
Submitted:
18 October 2023
Posted:
20 October 2023
You are already at the latest version
Abstract
Studies in the field of experimental hypnosis highlighted the role of hypnotizability in the physiological variability of the general population. It is associated, in fact, with a few differences which are observable in the ordinary state of consciousness and in the absence of suggestions. The aim of the present scoping review is summarizing them, and indicate their relevance to the neural mechanisms of hypnosis and to the prognosis and treatment of a few medical conditions. Individuals with high, medium and low hypnotizability scores display different cerebral functional differences – i.e., functional equivalence between imagery and perception/action, excitability of the motor cortex, interoceptive accuracy - possibly related to brain structural and functional characteristics -, and different control of blood supply at peripheral and cerebral level, likely due to different availability of endothelial nitric oxide. These differences are reviewed to support the idea of their participation in hypnotic behaviour and to indicate their prognostic and therapeutic usefulness in a few medical conditions.
Keywords:
motor imagery
; functional equivalence
; interoception
; cerebral blood flow
; endothelial function
; hypnosis.
1. Introduction
Hypnotizability, an individual trait substantially stable through life (Piccione et al., 1989), is associated with the proneness to enter hypnosis and/or to experience alteration of perception, memory and behavior following the administration of specific suggestions (Acunzo and Terhune, 2021). The observation that suggestions are effective in both the ordinary state of consciousness and after hypnotic induction (Santarcangelo et al., 2005; Carli et al., 2006; Derbyshire et al., 2009; Green and Lynn, 2011; Parris and Dienes, 2013; Zhang et al., 2018) has highlighted the role the trait of hypnotizability has, together with other individual traits and with contextual factors, in hypnotic behavior. Hypnotizability, in fact, just roughly predicts the response to suggestions, as placebo and hypnotizability- related mechanisms can cooperate, for instance, in the cognitive control of pain (Santarcangelo and Carli, 2021).
The contribution of several factors to hypnotic behavior is in line with the bio-psycho-social model of hypnosis (Jensen et al., 2015), rather than with the relevance of the induction of the hypnotic state.
Standard hypnotizability scales classify the general population into highly (highs), medium (mediums) and low hypnotizable persons (lows) according to the scales total score or on the basis of the specific scales items the subjects pass (Terhune et al., 2011).
The hypnotizability-related physiological correlates are physiological differences associated with different levels of hypnotizability which can be observed in the ordinary state of consciousness and in the absence of suggestions. Among the others, differences have been found in brain morpho-functional characteristics (Landry et al., 2017; Picerni et al., 2019), in the functional equivalence between imagery and perception/action, within groups topological homogeneity and modes of information processing (Ibanez-Marcelo et al., 2019), excitability of the motor cortex (Spina et al., 2020; Cesari et al., 2020), vascular peripheral (Jambrik et al., 2004; 2005) and cerebral control of blood flow (Rashid et al., 2022a,b), postural and visuomotor control (Santarcangelo et al., 2008; Menzocchi et al., 2014), interoception (Callara et al., 2023; Rosati et al., 2021; Diolaiuti et al., 2020), polymorphism of µ1 receptors (Presciuttini et al., 2018).
The object of this review is the description of the physiological correlates of hypnotizability which can account for a few hypnotic behaviors, or exhibit a prognostic role in medical conditions, or allow personalized pharmacological treatments. From this perspective, we describe facts and the hypotheses which can be suggested on the basis of current evidence.
1.1. Cerebral Morpho-Functional and Vascular Correlates of Hypnotizability
The earliest neuroimaging study conducted in individuals with different ability to accept suggestions revealed a larger anterior part of the corpus callosum (Horton et al., 2004), but recent investigation has not confirmed this difference between hypnotizability groups (Bastos et al., 2021). Reduction in the entire brain volume (including white and grey matter) has been observed in individuals able to experience deeper hypnotic trance (McGeown et al., 2015), i.e., the highs. This suggests than not only possible genetic markers, usually associated with localized variations - as occurs, for instance, in schizophrenia (Ahmed et al., 2021) and Parkinson disease (Ohira et al., 2022) -, but also maturation processes related to different availability of maturation factors, i.e., endothelial nitric oxide (NO), could be involved in the observed hypnotizability-related brain volume variations. In highs other studies revealed reduced grey matter volume (GMV) in the insula (Figure 1A), larger GMV in the mid-temporal and mid-occipital cortices, stronger functional connectivity between the anterior cingulate and the prefrontal dorsolateral cortex (Landry et al., 2017). Cerebellar morpho-functional differences have also been reported (Figure 1A). They consist of reduced GMV in the highs’ left lobules IV-VI compared to lows (Picerni et al., 2019).
The morphological variants of the cerebellar and insula GMV do not indicate impaired functions. Classical cerebellar tasks, such as postural and visuomotor control, in fact, are appropriately although less precisely performed by highs (Santarcangelo et al., 2008; Menzocchi et al., 2014). The highs’ postural control is less close than lows’, in fact, the set point for postural control which integrates peripheral sensory reafferents when the body centre of pressure are farther from the origin of sway compared to lows (Santarcangelo et al., 2008). This means that highs exhibit larger and faster body sway compared to lows, although reporting the same perception of body sway. Nonetheless, they do not fall down and, when they stand up on a very unstable platform, the differences disappear, likely due to attentional effort (Mecacci et al., 2013). Also visuomotor control, which is modulated by the direction of gaze (during prisms application and after their removal), is less precise in highs than in lows and both the error and the variability of the error are larger than in lows (Menzocchi et al., 2014). In both cases - postural and visuomotor control - the absence of learning across trials characterizes highs. Nonetheless, the typical cerebellar operation – changing the direction of launches toward a target as a function of prisms application and removal- is appropriately performed. Moreover, the performance of another typical cerebellar test, mental rotation, does not differ between highs, mediums and lows (Incognito et al., 2019).
The hypnotizability-related differences in cerebral blood flow were studied through near-infared spectroscopy (NIRS). It was shown that only highs exhibit a significant increase in blood supply during cognitive tasks (Figure 1B), suggesting that they can better adjust brain oxigenation to metabolic demands (Rashid et al., 2022a), which largely depends on both endothelial and neuronal nitric oxide release (Hoiland, et al., 2020). The metabolic demand, however, could be lower in medium-to-high than in low-to-medium hypnotizables, as suggested by the negative correlation between hypnotizability and cerebrovascular reactivity observed during visual stimulation (Rashid et al. 2022b) and could be due to the highs’ peculiar mode of information processing showing small and distributed changes in the brain asset (Ibanez-Marcelo et al., 2019). The highs’ greater increase in cerebral blood oxygenation during cognitive tasks could at least partially account for their greater attentional stability depending on the brain dopamine content (for review, Presciuttini et al., 2014) and, maybe, on cerebellar function (see 1.4).
1.2. Functional Equivalence between Real and Imagined Perception/Action
The functional equivalence (FE) between actual and imagined perception/action is indicated by the degree of superimposition between the cortical activations observed during these conditions (Jeannerod and Frak, 1999; Guillot et al., 2012; Hurst and Boe, 2022; Yao, et al., 2023; Henschke et al., 2023). It has been studied in highs and lows through topological analysis of the EEG, which revealed stronger FE between actual and imagined sensori-motor conditions in highs than in lows (Ibanez-Marcelo et al., 2019). In contrast, significant hypnotizability-related differences in the vividness of imagery have not been unanimously reported (Srzich et al., 2016). EEG topological analysis confirmed the hypothesis based on a behavioral experiment in which the earliest component of the vestibulo-spinal reflex (VR) was elicited by galvanic stimulation of the labyrinth, which is not under volitional control (Santarcangelo et al., 2010). VR develops in the frontal plane when the head is directed forward and in the sagittal plane when the head is rotated toward one side owing to the interaction between vestibular and neck proprioceptive information controlled by the cerebellum. Highs exhibited the same amplitude of the VR earlier component during both the actual and imagined rotated posture of the head (Santarcangelo et al., 2010). In line with this behavioural finding, topological EEG studies revealed similar topological asset during actual and imagined rotated posture of the head in highs (Ibanez-Marcelo et al., 2019, 2020). The same studies suggested differences in the modes of the cortical elaboration of sensory and imaginative stimuli. During imagery tasks, in fact, highs showed slight, distributed cortical topological changes almost not detectable through spectral analysis, whereas lows exhibited task-related localized changes detectable also through spectral analysis (Ibanez-Marcelo et al., 2019a, b; Cavallaro et al., 2010).
The highs’ stronger FE between imagined and actual action (Ibanez-Marcelo et al., 2019) together with the greater excitability of their motor cortex (Spina et al., 202; Cesari et al., 2020) can increase the likelihood of ideomotor responses, thus reducing the perception of effort and agency. The experience of involuntariness in suggested action, in fact, is one of the most important characteristics of hypnotic behaviour (Dell, 2010) and has been interpreted according to both dissociative (Kirsch and Lynn, 1998) and socio-cognitive views (Kirsch and Lynn, 1997). These two main theories can be theoretically reconciled, however, based on the complex nature of movement, which is often automatic and perceived as involuntary also in the ordinary state of consciousness (Lynn and Green, 2011).
The same EEG study (Ibanez-Marcelo et al., 2019) revealed greater topological homogeneity among highs than among lows, during all conditions. Work in progress (Lucas et al., personal communication) confirms this finding during baseline and extends this finding to hypnosis. This may seem to contrast with the observation of different types of highs according to the quality of the scales items they pass rather than according to the scales total score (Terhune et al., 2011). A possible interpretation is that the processes less general than those reflected by topology at mesoscopic level are not detected by the performed topological measures.
1.3. Motor Cortex Excitability
Studies of the motor cortex excitability have been performed through transcranial magnetic stimulation (TMS) of the motor cortex and recording of the evoked muscle activity in one hand. They have shown greater excitability of the right motor cortex in highs than in lows in resting conditions and during imagery of movement of the left hand, with mediums exhibiting intermediate excitability (Spina et al., 2020). The electromyographic activity at rest and during imagery of movement of the left hand, in fact, showed lower thresholds and higher amplitudes in highs than in lows. In contrast, TMS of the left motor cortex increases the excitability of the motor cortex and decreases the motor threshold in the right hand only during imagery. The high dopaminergic tone (Acunzo et al., 2021) of the highs’ cerebral cortex cannot account for the difference in the excitability of the right motor cortex by itself. In contrast to the left motor cortex, which is influenced only by dopamine content, the right cortex is influenced, in fact, also by the reduced cerebellar inhibition possibly due to the reduced volume of the left cerebellar lobules IV and V. The higher excitability of the right motor cortex might take part in the greater proneness of highs to respond to ideomotor suggestions by the left hand. For instance, the larger lowering of the left arm with respect to the right arm during suggestions of arm heaviness could be at least partially accounted for by greater excitability of the right motor cortex (Santarcangelo et al., 2005).
1.4. Attention and Emotion
The highs’ attention is greatly stable, in that highs are scarcely distractible from their current focus of attention. In the general population low distractibility is associated with high cortical dopamine content (Colzato et al., 2010), and the same occurs in highs (Raz and Posner, 2006). The highs’ higher dopaminergic tone, however, is suggested by neuropsychological tests, such as Stroop test, Vigilance task, Letter fluency task, Choice reaction times (Kallio et al., 2001), but the genetic argument widely cited to support it - the polymorphism of the Catechol-O- Methil- Transferase (COMT), responsible for reduced dopamine catabolism – is somehow weak. Genetic results on hypnotizability-related difference in COMT polymorphism, in fact, are inconsistent among each other, as COMT differences between highs and lows have been found present (Szekely et al., 2010), absent (Presciuttini et al., 2014; Bryant et al., 2013), present only among males (Lichtenberg et al., 2000), only in participants with peculiar attentional capabilities independently from hypnotizability (Rominger et al., 2014). The latter finding suggests that the reported hypnotizability-related COMT differences might not be the only factor of the highs’ attentional stability. Thus, it may be worthwile to note that the cerebellum is involved in both motor and non-motor functions (Strick et al., 2009) and, specifically, it allows the quick changes in the focus of attention. The same cerebellar peculiarities influencing the highs’ sensorimotor behavior can affect also their cognitive performance (Stric et al., 2009).
In addition, since the hypnotizability-related cerebellar peculiarities involve also the left lobule VI, which is associated with cognitive-emotional functions (Laricchiuta et al., 2022), we may hypothesize that the highs’ greater proneness to experience emotions (Crowson et al., 1991; Kirenskaya et al., 2011, Facco et al., 2017) and also their lower tendency to avoid possibly unpleasant situations compared to mediums (Diolaiuti et al., 2020) can be at least partially accounted for by cerebellar morpho-functional characteristics.
1.5. Paradoxical Pain Control by Cerebellar Stimulation
The reduced grey matter volume in the cerebellar lobules IV-V could be responsible for the highs’ less close postural and visuomotor control and absence of learning across successive trials of postural and visuomotor tasks compared to lows (Santarcangelo and Scattina, 2016). Moreover, reduced inhibition from these lobules onto left motor areas can be responsible for the greater excitability of the highs’ right motor cortex, which accounts for their lower motor threshold in the left (Spina et al., 2020) but not right hand (Cesari et al., 2020) in resting conditions. The highs’ higher excitability of both sides motor areas observed during motor imagery could be an effect of their stronger FE between the imagined and actual sensorimotor context.
The increase in the reported pain intensity and in the amplitude of cortically evoked nociceptive potentials observed after bilateral cerebellar anodal transcranial direct current stimulation (tDCS) in medium-to high hypnotizable participants (Bocci et al., 2017) contrasts with the findings obtained in the general population and in low-to-medium hypnotizables. Together with the highs’ higher emotional intensity (Kirenskaia et al., 2011; Facco et al., 2017), it could be due to reduced inhibition of regions of the pain matrix involved in cognition and emotion by the cerebellar left lobule VI (Laricchiuta et al., 2022). Nonetheless, it contrasts with the highs’ greater excitability of the motor cortex (Spina et al., 2020), whose activation is involved in pain reduction (Gan et al., 2021), and should be attributed to the cerebellar projections to other regions of the pain matrix. The prefrontal cortex, with its connections to the insula, is a good candidate (Ong et al., 2019; Seminowicz and Moayedi, 2017).
1.6. Interoception
Interoception is the perception of the bodily state (Craig, 2002). It is sustained by afferent signals, central integration and mental representation of visceral signals (Critchley and Garfinkel, 2017) and is extremely important, being linked to phenomenal consciousness, body awareness, cognition and affect (Tsakiris and Critchley, 2016). The insula is the structure most involved in interoception. Specifically, its anterior and posterior division are mainly connected to the prefrontal and orbitofrontal cortex. Three dimensions of interoception are usually considered: accuracy (IA) – the ability to detect interoceptive signals-, sensitivity (IS) – the interpretation of signals, and awareness, which represents the correspondence between interoceptive accuracy and sensitivity. IA is measured by behavioral tasks, for instance the heartbeats count and its comparison with the ECG recorded heartbeats. IS is measured by questionnaires such as the Multisensory assessment of Interoceptive Awareness (MAIA, Mehling et al., 2018), whose subscales indicate the awareness of body sensations (noticing), and that certain body sensations are the sensory aspect of emotional state; (emotional awareness), the tendency to ignore/distract oneself from sensations of pain/discomfort (not distracting), to not experience emotional distress or worry with sensations of pain or discomfort (not worrying), the ability to sustain and control attention to body sensation (attention regulation), to regulate psychological distress by attention to body sensations (self-regulation), to actively listen to the body for insight (body listening) and to experience of one’s body as safe and trustworthy (trusting). The Body Perception Questionnaire (Porges, 1993) refers to the awareness of bodily signals and to the detection of supra and subdiaphragmatic information, thus showing a closer relation with the activity of the autonomic system.
The difference in the insula grey matter volume and in its connections can be involved in hypnotizability-related differences in interoception. Highs display lower interoceptive accuracy – the ability to detect visceral signals measured by the heartbeat count test - than lows, with mediums exhibiting intermediate values (Rosati et al., 2021). Accordingly, their heartbeat-evoked cortical potential is smaller than lows’ in the centro-parietal regions, which are reached by projections from the anterior insula (Callara et al., 2023).
The highs’ ability to modify the experience of their body, usually indicated as an effect of dissociation, can be sustained by their low interoceptive accuracy possibly depending on the insula morpho-functional characteristics, which could allow them to feel a body condition different from the real one (and facilitate dissociative experiences).
1.7. Hypnotizability and Brain Injuries
A few hypnotizability-related brain functions allow to hypothesize that highs could be less vulnerable to brain injuries and more resilient to them compared to lows.
The topologically different cortical elaboration of sensory and cognitive information – distributed in highs, localized in lows (Ibanez-Marcelo et al., 2019, 2020) - suggests, in fact, that brain lesions could be less impairing in highs than in lows. Clinical studies, however, are required to ascertain whether brain lesions produce less deficits in highs than in lows.
The highs’ better cerebrovascular reactivity may buffer transient alteration of blood flow and, theoretically, their stronger FE between imagery and perception/action (Ibanez-Marcelo et al., 2019), together with the higher excitability of the motor cortex (Spina et al., 2020; Cesari et al., 2020), makes highs more prone than lows to take advantage from mental training after brain lesions of any origin (Fontanelli et al., 2022). The latter finding suggests that hypnotic assessment could predict the outcome of mental training and Brain Computer Interface interventions, which display large variability in the outcome and is ineffective in part of the general population (Peters et al., 2022). Studies in progress are aimed at assessing whether training to mental imagery can increase FE in mediums/lows, thus extending the utilization of mental training to larger part of the population. Preliminary findings show that motor imagey training improves the velocity and accuracy of movement and that the improvement lasts at least two weeks after 5 days training (Terreni, 2023). TMS and anodal tDCS aimed at improving motor responses are more expensive and time consuming than mental training, and the duration of their effects has not been consistently reported (Han et al., 2023).
Figure 1.
Association between hypnotizability-related morpho-functional differences and behavior. A, at brain level; B) at vascular level.
Figure 1.
Association between hypnotizability-related morpho-functional differences and behavior. A, at brain level; B) at vascular level.

1.8. Cardiovascular Control
The most important vascular difference between highs and lows is in the post-occlusion flow-mediated endothelial function (FMD, Figure 1B). It is defined as the difference in an artery diameter measured after and before the artery occlusion and is usually tested in the brachial artery. In the general healthy population and in lows, after occlusion the flow-mediated dilation is larger than before it, as the swirling blood flow following dis-occlusion promotes the release of nitric oxide from endothelial cells. In highs, the brachial artery post-occlusion flow-mediated dilation is significantly less reduced than in lows during tonic nociceptive stimulation (Jambrik et al., 2005) and not reduced at all during mental computation (Jambrik et al., 2004, 2005). Since FMD is considered a reliable index of cardiovascular health (Matsukawa et al., 2015), in the absence of risk factors, high hypnotizability should predict better cardiovascular health.
Endothelial nitric oxide controls vascular dilation, thus is relevant to the function of all organs and systems (Grześk et al., 2023). Larger post occlusion flow-mediated dilation, in fact, is considered a predictor of less vulnerability to cardiovascular events (Mućka et al., 2022), and drugs containing nitric oxide donors are administered in clinical trials (Escobar Cervantes et al., 2023). Moreover, nitric oxide exerts a central inhibition of the sympathetic activity (Kishi, 2013), which is increased in patients with heart failure (Quarti-Trevano et al., 2023). In this respect, high hypnotizability may have a favorable prognostic role in case of cardiovascular events. In fact, nitric oxide inhalation positively influences the course of cardio-cerebrovascular diseases (Huerta de la Cruz et al., 2022). We can also hypothesize that highs are less vulnerable to vascular-based cognitive decline owing to their cerebrovascular reactivity (Rajeev et al., 2023; Uemura et al., 2020). Nonetheless, eccessive NO has been associated with Alzheimer earlier degeneration (Malinski, 2007). Thus, it is important to remark that hypnotizability-related differences in basal blood flow have not been observed.
During long lasting relaxation, highs increase their parasympathetic tone more than lows (Santarcangelo et al., 2012). The parasympathetic tone is indicated by the High Frequency component of the tacogram power spectrum (a signal obtained by reporting the sequence of the distances between consecutive R waves of the ECG). Also, at variance with lows, in standing position the highs’ increase in the Low Frequency component of heart rate variability – related to the sympathetic activity - is not significant (Santarcangelo et al., 2008). Both findings could be accounted for by higher release of nitric oxide in the bulbar regions responsible for sympathetic inhibition in the general population (Kishi, 2013). Thus, the highs’ greater proneness to induce relaxation responses (Hoffman et al., 1982) could work as a natural protection against stress.
Low sympathetic activity could also induce more efficient activity of the immune system, with useful effects on autoimmune conditions (Ruzyla-Smith et al., 1995). Hypnotic treatments, which induces relaxation responses, influence the immune system by modulation of the autonomic activity and consequent greater decreases in highs than in lows in the activity of Natural Killers lymphocytes and lymphocyte proliferative response. In highs, hypnotic suggestions of relaxation and wellbeing buffer the decline in NKP, CD8, and CD8/CD4 ratio occurring during examination-related stress in students, and upregulate the expression of immune-related genes. In cancer patients and geriatric patients positive immune effects of hypnotic relaxation have also been reported (Fontanelli et al., 2021). Finally, the highs’ ability to modulate their autonomic activity (Sebastiani et al., 2005) could positively influence their microbiota (De Benedittis, 2022), whose alteration is also involved in cognitive decline owing to the cerebral effects of locally produced cytokines and the activation af afferent vagal fibers (Weber et al., 2023).
2. Limitations and Conclusions
A limitation of the reported studies is that mediums, who represent 70% of the general population, have been seldom enrolled. Thus, at the moment, only part of the hypnotizability-related findings can be extended to the general population (Jensen et al., 2017). In the studies of FE (Ibanez-Marcelo et al., 2019; Ruggirello et al., 2020) and FMD (Jambrik et al., 2004, 2005 a, b) only highs and lows have been recruited. In a few studies – motor cortex excitability (Spina et al., 2020), interoceptive accuracy (Rosati et al., 2021) - mediums exhibit intermediate values, not always significantly different from highs and lows. In other experiments – cerebellar tDCS stimulation before nociceptive stimulation (Bocci et al., 2018), and cerebral blood flow (Rashid et al., 2022 a,b), interoception accuracy (Rosati et al., 2021) - the participants have been divided in low-to-medium hypnotizables (score 0-5 on the Stanford Hypnotic Susceptibility Scale, form A) and medium-to-high hypnotizables (score 7-12 on SHSS, A), thus reducing the sensitivity of the study to hypnotizability.
Finally, some of the suggested mechanisms of the highs’ higher parasympathetic tone, i.e., the sympathetic inhibition by NO at bulbar level should be experimentally confirmed in humans.
A question arising from the present approach to hypnotizability is when and how we will be able to perform hypnotic assessment instrumentally. Attempts have been done through EEG studies, but they are not satisfying because the suggested indices have been obtained during sessions including suggestions and/or hypnosis (Baghdadi and Nasrabadi, 2012; Yargholi and Nasrabadi, 2015). A possible discriminant index obtained in resting conditions is the Determinism of the EEG Recurrence Plot, which approximates a good separation between highs and lows (Madeo et al., 2013; Chiarucci et al., 2014).
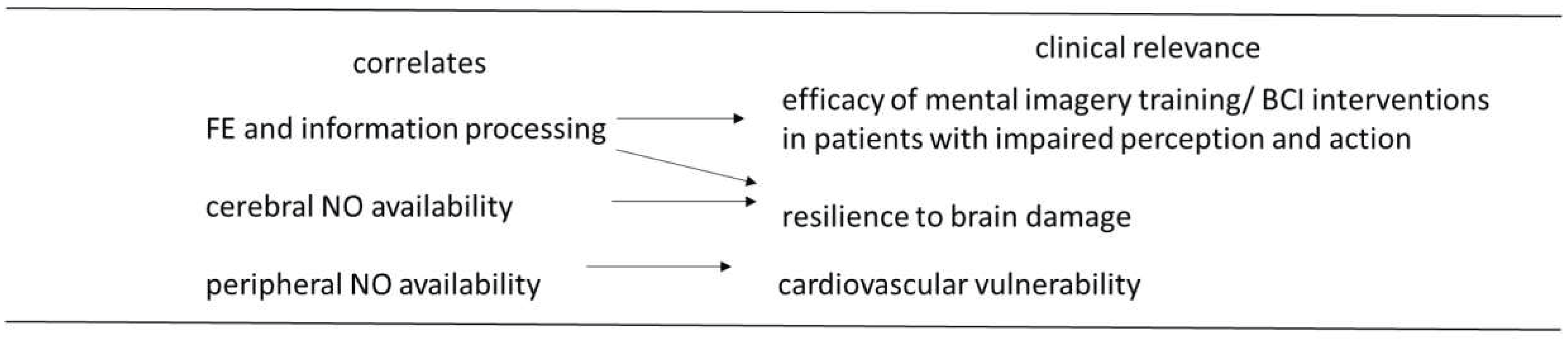
In conclusion, the present review is an attempt to translate psychological characteristics into physiological functions. It describes hypnotizability-related physiological characteristics (Figure 2) possibly accounting for a) a few hypnotic behaviours (the response to ideomotor suggestions owing to stronger FE between imagery and perception/action and greater excitability of the motor cortex), b) the stability of attention due not only to higher dopamine content but also to cerebellar peculiarities. Other correlates of hypnotizability predict c) greater resilience to brain injuries and efficacy of mental training and better cardio- and cerebrovascular functions (availability and sensitivity to endothelial nitric oxide) and allow d) to personalize pharmacological pain therapies (owing to different sensitivity of highs and lows’ µ1 receptors). Finally, this review is a call to medical doctors to consider the relevance of hypnotic assessment to their clinical practice.
Acknowledgments
This article was produced by Eleonora Malloggi while attending the PhD program in Space Science and Technology at the University of Trento, Cycle XXXVIII, with the support of a scholarship financed by the Ministerial Decree no. 352 of 9th April 2022, based on the NRRP - funded by the European Union - NextGenerationEU - Mission 4 “Education and Research”, Component 1 “Enhancement of the offer of educational services: from nurseries to universities” - Investment 4.1 “Extension of the number of research doctorates and innovative doctorates for public administration and cultural heritage”.
References
- Acunzo, D. J., & Terhune, D. B. (2021). A Critical Review of Standardized Measures of Hypnotic Suggestibility. The International journal of clinical and experimental hypnosis, 69(1), 50–71. [CrossRef]
- Acunzo, D.J., Oakley, D.A., & Terhune, D.B. (2021). The neurochemistry of hypnotic suggestion. American Journal of Clinical hypnosis, 63(4), 355-371. [CrossRef]
- Ahmad, B., Gill, I., Liblik, K., Uppal, J. S., & El-Diasty, M. (2023). The Role of Hypnotherapy in Postoperative Cardiac Surgical Patients, A Scoping Review of Current Literature. Current problems in cardiology, 48(9), 101787. Advance online publication. [CrossRef]
- Ahmed, A.O., Kramer, S., Hofman, N., Flynn, J., Hansen, M., Martin, V., Pillai, A., Buckley, P.F. (2021). A Meta-Analysis of Brain-Derived Neurotrophic Factor Effects on Brain Volume in Schizophrenia: Genotype and Serum Levels. Neuropsychobiology, 80(5), 411-424. [CrossRef]
- Baghdadi, G., Nasrabadi, A.M. (2021a). Comparison of different EEG features in estimation of hypnosis susceptibility level. Comput Biol Med, 42(5), 590-7. [CrossRef]
- Baghdadi, G. & Nasrabadi, A.M. (2012b). EEG phase synchronization during hypnosis induction. J Med Eng Technol, 36(4), 222-9. [CrossRef]
- Bastos, M.A.V. Jr, Oliveira Bastos, P.R.H., Foscaches Filho, G.B., Conde, R.B., Ozaki, J.G.O., Portella, R.B., Iandoli, D. Jr, Lucchetti, G. (2022). Corpus callosum size, hypnotic susceptibility and empathy in women with alleged mediumship: a controlled study. Explore (NY), 18(2), 217-225. [CrossRef]
- Bocci, T., Barloscio, D., Parenti, L., Sartucci, F., Carli, G., & Santarcangelo, E.L. (2017). High Hypnotizability Impairs the Cerebellar Control of Pain. Cerebellum (London, England), 16(1), 55–61. [CrossRef]
- Callara, A. L., Fontanelli, L., Belcari, I., Rho, G., Greco, A., Zelič, Ž., Sebastiani, L., & Santarcangelo, E.L. (2023). Modulation of the heartbeat evoked cortical potential by hypnotizability and hypnosis. Psychophysiology, e14309. Advance online publication. [CrossRef]
- Carli, G., Rendo, C., Sebastiani, L., Santarcangelo, E.L. (2006). Suggestions of altered balance: Possible equivalence of imagery and perception. Int J Clin Exp Hypn, 54(2), 206-23. [CrossRef]
- Cavallaro, F.I., Cacace, I., Del Testa, M., Andre, P., Carli, G., De Pascalis, V., Rocchi, R., & Santarcangelo, E. L. (2010). Hypnotizability-related EEG alpha and theta activities during visual and somesthetic imageries. Neuroscience letters, 470(1), 13–18. [CrossRef]
- Cesari, P., Modenese, M., Benedetti, S., Emadi Andani, M., & Fiorio, M. (2020). Hypnosis-induced modulation of corticospinal excitability during motor imagery. Scientific reports, 10(1), 16882. [CrossRef]
- Chiarucci, R., Madeo, D., Loffredo, M.I., Castellani, E., Santarcangelo, E.L., Mocenni, C. (2014). Cross-evidence for hypnotic susceptibility through nonlinear measures on EEGs of non-hypnotized subjects. Sci Rep, 8, 4-5610. [CrossRef]
- Colzato L. S., Waszak F., Nieuwenhuis S., Posthuma D., Hommel B. (2010). The flexible mind is associated with the catechol-O-methyltransferase (COMT) Val158Met polymorphism: evidence for a role of dopamine in the control of task-switching. Neuropsychologia, 48, 2764–2768.
- Craig, A.D. (2002). How do you feel? Interoception: the sense of the physiological condition of the body. Nat Rev Neurosci, 3(8), 655-66. [CrossRef]
- Critchley, H.D., Garfinkel, S.N. (2017). Interoception and emotion. Curr Opin Psychol, 17:7-14. [CrossRef]
- Crowson, J.J. Jr, Conroy, A.M., Chester, T.D. (1991). Hypnotizability as related to visually induced affective reactivity: a brief communication. Int J Clin Exp Hypn, 39(3), 140-4. [CrossRef]
- Császár-Nag, N., & Bókkon, I. (2022). Hypnotherapy and IBS: Implicit, long-term stress memory in the ENS. Heliyon, 9(1), e12751. [CrossRef]
- De Benedittis, G. (2022). Hypnobiome: A New, Potential Frontier of Hypnotherapy in the Treatment of Irritable Bowel Syndrome-A Narrative Review of the Literature. The International journal of clinical and experimental hypnosis, 70(3), 286–299. [CrossRef]
- Dell P. F. (2010). Involuntariness in hypnotic responding and dissociative symptoms. Journal of trauma & dissociation: the official journal of the International Society for the Study of Dissociation (ISSD), 11(1), 1–18. [CrossRef]
- Derbyshire, S. W., Whalley, M. G., & Oakley, D. A. (2009). Fibromyalgia pain and its modulation by hypnotic and non-hypnotic suggestion: an fMRI analysis. European journal of pain (London, England), 13(5), 542–550. [CrossRef]
- Elkins G. (2022). From Research to Clinical Practice. The International journal of clinical and experimental hypnosis, 70(3), 209–211. [CrossRef]
- Escobar Cervantes, C., Esteban Fernández, A., Recio Mayoral, A., Mirabet, S., González Costello, J., Rubio Gracia, J., Núñez Villota, J., González Franco, Á., & Bonilla Palomas, J. L. (2023). Identifying the patient with heart failure to be treated with vericiguat. Current medical research and opinion, 39(5), 661–669. [CrossRef]
- Facco, E., Testoni, I., Ronconi, L., Casiglia, E., Zanette, G., & Spiegel, D. (2017). Psychological Features of Hypnotizability: A First Step Towards Its Empirical Definition. The International journal of clinical and experimental hypnosis, 65(1), 98–119. [CrossRef]
- Fiorio, M., Modenese, M., & Cesari, P. (2020). The rubber hand illusion in hypnosis provides new insights into the sense of body ownership. Scientific reports, 10(1), 5706. [CrossRef]
- Fontanelli, L., Spina, V., Chisari, C., Siciliano, G., & Santarcangelo, E. L. (2022). Is hypnotic assessment relevant to neurology?. Neurological sciences: official journal of the Italian Neurological Society and of the Italian Society of Clinical Neurophysiology, 43(8), 4655–4661. [CrossRef]
- Gan, Z., Li, H., Naser, P.V., Oswald, M.J., Kuner, R. (2021). Suppression of Neuropathic Pain and Comorbidities by Recurrent Cycles of Repetitive Transcranial Direct Current Motor Cortex Stimulation in Mice. Sci. Rep., 11, 9735.
- Green, J.P., Lynn, S.J. (2011). Hypnotic responsiveness: expectancy, attitudes, fantasy proneness, absorption, and gender. Int J Clin Exp Hypn, 59(1), 103-21. [CrossRef]
- Grześk, G., Witczyńska, A., Węglarz, M., Wołowiec, Ł., Nowaczyk, J., Grześk, E., & Nowaczyk, A. (2023). Soluble Guanylyl Cyclase Activators-Promising Therapeutic Option in the Pharmacotherapy of Heart Failure and Pulmonary Hypertension. Molecules (Basel, Switzerland), 28(2), 861. [CrossRef]
- Guillot, A., Di Rienzo, F., Macintyre, T., Moran, A., & Collet, C. (2012). Imagining is Not Doing but Involves Specific Motor Commands: A Review of Experimental Data Related to Motor Inhibition. Frontiers in human neuroscience, 6, 247. [CrossRef]
- Han, K., Liu, J., Tang, Z., Su, W., Liu, Y., Lu, H., Zhang, H. (2023). Effects of excitatory transcranial magnetic stimulation over the different cerebral hemispheres dorsolateral prefrontal cortex for post-stroke cognitive impairment: a systematic review and meta-analysis. Front Neurosci, 17, 1102311. PMID: 37260845; PMCID: PMC10228699. [CrossRef]
- Henschke, J. U., & Pakan, J. M. P. (2023). Engaging distributed cortical and cerebellar networks through motor execution, observation, and imagery. Frontiers in systems neuroscience, 17, 1165307. [CrossRef]
- Hoffman, J.W., Benson, H., Arns, P.A., Stainbrook, G.L., Landsberg, G.L., Young, J.B., Gill, A. (1982). Reduced sympathetic nervous system responsivity associated with the relaxation response. Science, 215(4529), 190-2. [CrossRef]
- Hoiland, R. L., Caldwell, H. G., Howe, C. A., Nowak-Flück, D., Stacey, B. S., Bailey, D. M., Paton, J. F. R., Green, D. J., Sekhon, M. S., Macleod, D. B., & Ainslie, P. N. (2020). Nitric oxide is fundamental to neurovascular coupling in humans. The Journal of physiology, 598(21), 4927–4939. [CrossRef]
- Horton, J.E., Crawford, H.J., Harrington, G., Downs, J.H. 3rd. (2004). Increased anterior corpus callosum size associated positively with hypnotizability and the ability to control pain. Brain, 127(Pt 8), 1741-7. [CrossRef]
- Huerta de la Cruz, S., Santiago-Castañeda, C. L., Rodríguez-Palma, E. J., Medina-Terol, G. J., López-Preza, F. I., Rocha, L., Sánchez-López, A., Freeman, K., & Centurión, D. (2022). Targeting hydrogen sulfide and nitric oxide to repair cardiovascular injury after trauma. Nitric oxide: biology and chemistry, 129, 82–101. [CrossRef]
- Hurst, A. J., & Boe, S. G. (2022). Imagining the way forward: A review of contemporary motor imagery theory. Frontiers in human neuroscience, 16, 1033493. [CrossRef]
- Ibáñez-Marcelo, E., Campioni, L., Manzoni, D., Santarcangelo, E. L., & Petri, G. (2019). Spectral and topological analyses of the cortical representation of the head position: Does hypnotizability matter?. Brain and behavior, 9(6), e01277. [CrossRef]
- Ibáñez-Marcelo, E., Campioni, L., Phinyomark, A., Petri, G., & Santarcangelo, E. L. (2019a). Topology highlights mesoscopic functional equivalence between imagery and perception: The case of hypnotizability. NeuroImage, 200, 437–449. [CrossRef]
- Incognito, O., Menardo, E., Di Gruttola, F., Tomaiuolo, F., Sebastiani, L., Santarcangelo, E.L. (2019). Visuospatial imagery in healthy individuals with different hypnotizability levels. Neurosci Lett, 690, 158-161. [CrossRef]
- ambrik, Z., Santarcangelo, E. L., Ghelarducci, B., Picano, E., & Sebastiani, L. (2004). Does hypnotizability modulate the stress-related endothelial dysfunction?. Brain research bulletin, 63(3), 213–216. [CrossRef]
- Jambrik, Z., Santarcangelo, E. L., Rudisch, T., Varga, A., Forster, T., & Carli, G. (2005). Modulation of pain-induced endothelial dysfunction by hypnotisability. Pain, 116(3), 181–186. [CrossRef]
- Jeannerod, M., & Frak, V. (1999). Mental imaging of motor activity in humans. Current opinion in neurobiology, 9(6), 735–739. [CrossRef]
- Jensen, M.P., Adachi, T., Tomé-Pires, C., Lee, J., Osman, Z.J., Miró, J. (2015). Mechanisms of hypnosis: toward the development of a biopsychosocial model. Int J Clin Exp Hypn, 63(1), 34-75. Erratum in: Int J Clin Exp Hypn, 63(2), 247. [CrossRef]
- Kallio, S., Revonsuo, A., Hämäläinen, H., Markela, J., & Gruzelier, J. (2001). Anterior brain functions and hypnosis: a test of the frontal hypothesis. The International journal of clinical and experimental hypnosis, 49(2), 95–108. [CrossRef]
- Kirenskaya, A. V., Novototsky-Vlasov, V. Y., Chistyakov, A. N., & Zvonikov, V. M. (2011). The relationship between hypnotizability, internal imagery, and efficiency of neurolinguistic programming. The International journal of clinical and experimental hypnosis, 59(2), 225–241. [CrossRef]
- Kirsch, I., & Lynn, S. J. (1997). Hypnotic involuntariness and the automaticity of everyday life. The American journal of clinical hypnosis, 40(1), 329–348. [CrossRef]
- Kirsch, I., & Lynn, S. J. (1998). Dissociation theories of hypnosis. Psychological bulletin, 123(1), 100–115. [CrossRef]
- Kishi, T. (2013). Regulation of the sympathetic nervous system by nitric oxide and oxidative stress in the rostral ventrolateral medulla: 2012 Academic Conference Award from the Japanese Society of Hypertension. Hypertension research : official journal of the Japanese Society of Hypertension, 36(10), 845–851. [CrossRef]
- Landry, M., Lifshitz, M., & Raz, A. (2017). Brain correlates of hypnosis: A systematic review and meta-analytic exploration. Neuroscience and biobehavioral reviews, 81(Pt A), 75–98. [CrossRef]
- Laricchiuta, D., Picerni, E., Cutuli, D., & Petrosini, L. (2022). Cerebellum, Embodied Emotions, and Psychological Traits. Advances in experimental medicine and biology, 1378, 255–269. [CrossRef]
- Lichtenberg, P., Bachner-Melman, R., Gritsenko, I., Ebstein, R.P. (2000). Exploratory association study between catechol-O-methyltransferase (COMT) high/low enzyme activity polymorphism and hypnotizability. Am J Med Genet, 96(6), 771-4. [CrossRef]
- Lifshitz, M., Howells, C., & Raz, A. (2012). Can expectation enhance response to suggestion? De-automatization illuminates a conundrum. Consciousness and cognition, 21(2), 1001–1008. [CrossRef]
- Lynn, S. J., & Green, J. P. (2011). The sociocognitive and dissociation theories of hypnosis: toward a rapprochement. The International journal of clinical and experimental hypnosis, 59(3), 277–293. [CrossRef]
- Lynn, S. J., Green, J. P., Zahedi, A., & Apelian, C. (2023). The response set theory of hypnosis reconsidered: toward an integrative model. The American journal of clinical hypnosis, 65(3), 186–210. [CrossRef]
- Ludwig, V. U., Seitz, J., Schönfeldt-Lecuona, C., Höse, A., Abler, B., Hole, G., Goebel, R., & Walter, H. (2015). The neural correlates of movement intentions: A pilot study comparing hypnotic and simulated paralysis. Consciousness and cognition, 35, 158–170. [CrossRef]
- Madeo, D., Castellani, E., Santarcangelo, E.L., Mocenni, C. (2013). Hypnotic assessment based on the recurrence quantification analysis of EEG recorded in the ordinary state of consciousness. Brain Cogn, 83(2), 227-33. [CrossRef]
- Malinski T. (2007). Nitric oxide and nitroxidative stress in Alzheimer’s disease. Journal of Alzheimer’s disease : JAD, 11(2), 207–218. [CrossRef]
- Matsuzawa, Y., Kwon, T.G., Lennon, R.J., Lerman, L.O., Lerman, A. (2015). Prognostic Value of Flow-Mediated Vasodilation in Brachial Artery and Fingertip Artery for Cardiovascular Events: A Systematic Review and Meta-Analysis. J Am Heart Assoc, 4(11), e002270. [CrossRef]
- McGeown, W.J., Mazzoni, G., Vannucci, M., & Venneri, A. (2015). Structural and functional correlates of hypnotic depth and suggestibility. Psychiatry Reseach, 28, 231(2), 151-9. [CrossRef]
- Mecacci, G., Menzocchi, M., Zeppi, A., Carli, G., & Santarcangelo, E.L(2013). Body sway modulation by hypnotizability and gender during low and high demanding postural conditions. Archives Italilennes de Biologie, 151(3), 99-105. [CrossRef]
- Mehling, W.E., Acree, M., Stewart, A., Silas, J., Jones, A. (2018). The Multidimensional Assessment of Interoceptive Awareness, Version 2 (MAIA-2). PLoS One, 13(12), e0208034. [CrossRef]
- Mućka, S., Miodońska, M., Jakubiak, G. K., Starzak, M., Cieślar, G., & Stanek, A. (2022). Endothelial Function Assessment by Flow-Mediated Dilation Method: A Valuable Tool in the Evaluation of the Cardiovascular System. International journal of environmental research and public health, 19(18), 11242. [CrossRef]
- Ohira, K., Yokota, H., Hirano, S., Nishimura, M., Mukai, H., Horikoshi, T., Sawai, S., Yamanaka, Y., Yamamoto, T., Kakeda, S., Kuwabara, S., Tanaka, T., Uno, T. (2022). DRD2 Taq1A Polymorphism-Related Brain Volume Changes in Parkinson’s Disease: Voxel-Based Morphometry. Parkinsons Dis, 2022, 8649195. [CrossRef]
- Ong, W.Y., Stohler, C.S., Herr, D.R. (2019). Role of the Prefrontal Cortex in Pain Processing. Mol Neurobiol, 56(2), 1137-1166. [CrossRef]
- Parris, B.A. & Dienes, Z. (2013). Hypnotic suggestibility predicts the magnitude of the imaginative word blindness suggestion effect in a non-hypnotic context. Conscious Cogn, 22(3), 868-74. [CrossRef]
- Parris, B. A., Hasshim, N., & Dienes, Z. (2021). Look into my eyes: Pupillometry reveals that a post-hypnotic suggestion for word blindness reduces Stroop interference by marshalling greater effortful control. The European journal of neuroscience, 53(8), 2819–2834. [CrossRef]
- Peters, B., Eddy, B., Galvin-McLaughlin, D., Betz, G., Oken, B., & Fried-Oken, M. (2022). A systematic review of research on augmentative and alternative communication brain-computer interface systems for individuals with disabilities. Frontiers in human neuroscience, 16, 952380. [CrossRef]
- Piccione, C., Hilgard, E. R., & Zimbardo, P. G. (1989). On the degree of stability of measured hypnotizability over a 25-year period. Journal of personality and social psychology, 56(2), 289–295. [CrossRef]
- Picerni, E., Santarcangelo, E. L., Laricchiuta, D., Cutuli, D., Petrosini, L., Spalletta, G., & Piras, F. (2019). Cerebellar Structural Variations in Participants with Different Hypnotizability. Cerebellum (London, England), 18(1), 109–118. [CrossRef]
- Porges, S.W. Body Perception Questionnaire. Laboratory of Developmental Assessment, University of Maryland; College Park, MD, USA: 1993.
- Presciuttini, S., Gialluisi, A., Barbuti, S., Curcio, M., Scatena, F., Carli, G., & Santarcangelo, E. L. (2014). Hypnotizability and Catechol-O-Methyltransferase (COMT) polymorphysms in Italians. Frontiers in human neuroscience, 7, 929. [CrossRef]
- Quarti-Trevano, F., Dell’Oro, R., Cuspidi, C., Ambrosino, P., & Grassi, G. (2023). Endothelial, Vascular and Sympathetic Alterations as Therapeutic Targets in Chronic Heart Failure. Biomedicines, 11(3), 803. [CrossRef]
- Rajeev, V., Chai, Y. L., Poh, L., Selvaraji, S., Fann, D. Y., Jo, D. G., De Silva, T. M., Drummond, G. R., Sobey, C. G., Arumugam, T. V., Chen, C. P., & Lai, M. K. P. (2023). Chronic cerebral hypoperfusion: a critical feature in unravelling the etiology of vascular cognitive impairment. Acta neuropathologica communications, 11(1), 93. [CrossRef]
- Rashid, A., Santarcangelo, E. L., & Roatta, S. (2022a). Does hypnotizability affect neurovascular coupling during cognitive tasks?. Physiology & behavior, 257, 113915. [CrossRef]
- Rashid, A., Santarcangelo, E. L., & Roatta, S. (2022b). Cerebrovascular reactivity during visual stimulation: Does hypnotizability matter?. Brain research, 1794, 148059. [CrossRef]
- Raz A., Fan J., & Posner M. I. (2006). Neuroimaging and genetic associations of attentional and hypnotic processes. Journal of Physiology (Paris), 99, 483–491. [CrossRef]
- Rominger, C., Weiss, E.M., Nagl, S., Niederstätter, H., Parson, W., Papousek, I. (2014). Carriers of the COMT Met/Met allele have higher degrees of hypnotizability, provided that they have good attentional control: a case of gene-trait interaction. Int J Clin Exp Hypn, 62(4), 455-82. [CrossRef]
- Rosati, A., Belcari, I., Santarcangelo, E. L., & Sebastiani, L. (2021). Interoceptive Accuracy as a Function of Hypnotizability. The International journal of clinical and experimental hypnosis, 69(4), 441–452. [CrossRef]
- Ruggirello, S., Campioni, L., Piermanni, S., Sebastiani, L., & Santarcangelo, E. L. (2019). Does hypnotic assessment predict the functional equivalence between motor imagery and action?. Brain and cognition, 136, 103598. [CrossRef]
- Ruzyla-Smith, P., Barabasz, A., Barabasz, M. & Warner, D. (1995). Effects of hypnosis on the immune response: B-cells, T-cells, helper and suppressor cells. American Journal of Clinical Hypnosis, 38, 71-79.
- Santarcangelo, E. L., & Carli, G. (2021). Individual Traits and Pain Treatment: The Case of Hypnotizability. Frontiers in neuroscience, 15, 683045. [CrossRef]
- Santarcangelo, E.L., Cavallaro, E., Mazzoleni, S., Marano, E., Ghelarducci, B., Dario, P., Micera, S., & Sebastiani, L. (2005). Kinematic strategies for lowering of upper limbs during suggestions of heaviness: a real-simulator design. Experimental Brain Research, 162(1),35-45. [CrossRef]
- Santarcangelo, E. L., Scattina, E., Carli, G., Ghelarducci, B., Orsini, P., & Manzoni, D. (2010). Can imagery become reality?. Experimental Brain Research, 206(3), 329–335. [CrossRef]
- Santarcangelo, E. L., & Scattina, E. (2016). Complementing the Latest APA Definition of Hypnosis: Sensory-Motor and Vascular Peculiarities Involved in Hypnotizability. The International journal of clinical and experimental hypnosis, 64(3), 318–330. [CrossRef]
- Santarcangelo, E. L., Balocchi, R., Scattina, E., Manzoni, D., Bruschini, L., Ghelarducci, B., & Varanini, M. (2008). Hypnotizability-dependent modulation of the changes in heart rate control induced by upright stance. Brain research bulletin, 75(5), 692–697. [CrossRef]
- Santarcangelo, E. L., Paoletti, G., Balocchi, R., Carli, G., Morizzo, C., Palombo, C., & Varanini, M. (2012). Hypnotizability modulates the cardiovascular correlates of participantive relaxation. The International journal of clinical and experimental hypnosis, 60(4), 383–396. [CrossRef]
- Scacchia, P., & De Pascalis, V. (2020). Effects of Prehypnotic Instructions on Hypnotizability and Relationships Between Hypnotizability, Absorption, and Empathy. The American journal of clinical hypnosis, 62(3), 231–266. [CrossRef]
- Sebastiani, L., D’Alessandro, L., Menicucci, D., Ghelarducci, B., & Santarcangelo, E. L. (2007). Role of relaxation and specific suggestions in hypnotic emotional numbing. International journal of psychophysiology, 63(1), 125–132. [CrossRef]
- Seminowicz, D.A., & Moayedi, M. (2017). The Dorsolateral Prefrontal Cortex in Acute and Chronic Pain. J Pain, 18(9), 1027-1035. [CrossRef]
- Spina, V., Chisari, C., & Santarcangelo, E. L. (2020). High Motor Cortex Excitability in Highly Hypnotizable Individuals: A Favourable Factor for Neuroplasticity?. Neuroscience, 430, 125–130. [CrossRef]
- Srzich, A. J., Byblow, W. D., Stinear, J. W., Cirillo, J., & Anson, J. G. (2016). Can motor imagery and hypnotic susceptibility explain Conversion Disorder with motor symptoms?. Neuropsychologia, 89, 287–298. [CrossRef]
- Strick, P.L., Dum, R.P., & Fiez, J.A. (2009). Cerebellum and nonmotor function. Annual Reviews of Neuroscience, 32, 413-34. [CrossRef]
- Szekely, A., Kovacs-Nagy, R., Bányai, E.I., Gosi-Greguss, A.C., Varga, K., Halmai, Z., Ronai, Z., Sasvari-Szekely, M. (2010). Association between hypnotizability and the catechol-O-methyltransferase (COMT) polymorphism. Int J Clin Exp Hypn, 58(3), 301-15. [CrossRef]
- Terhune, D.B., Cardeña, E., Lindgren, M. (2011). Dissociated control as a signature of typological variability in high hypnotic suggestibility. Conscious Cogn, 20(3), 727-36. [CrossRef]
- Terreni, C. Efficacia di un training immaginativo sul movimento: studio sperimentale. Master thesis, Pisa University, 2023.
- Tsakiris, M., & Critchley, H. (2016). Interoception beyond homeostasis: affect, cognition and mental health. Philos Trans R Soc Lond B Biol Sci, 371(1708), 20160002. [CrossRef]
- Uemura, M. T., Maki, T., Ihara, M., Lee, V. M. Y., & Trojanowski, J. Q. (2020). Brain Microvascular Pericytes in Vascular Cognitive Impairment and Dementia. Frontiers in aging neuroscience, 12, 80. [CrossRef]
- Weber, C., Dilthey, A., & Finzer, P. (2023). The role of microbiome-host interactions in the development of Alzheimer’s disease. Frontiers in cellular and infection microbiology, 13, 1151021. [CrossRef]
- Wieder, L., Brown, R., Thompson, T., & Terhune, D. B. (2021). Suggestibility in functional neurological disorder: a meta-analysis. Journal of neurology, neurosurgery, and psychiatry, 92(2), 150–157. [CrossRef]
- Wolf, T. G., Faerber, K. A., Rummel, C., Halsband, U., & Campus, G. (2022). Functional Changes in Brain Activity Using Hypnosis: A Systematic Review. Brain sciences, 12(1), 108. [CrossRef]
- Yargholi, E., & Nasrabadi, A.M. (2015). Chaos-chaos transition of left hemisphere EEGs during standard tasks of Waterloo-Stanford Group Scale of hypnotic susceptibility. J Med Eng Technol, 39(5), 281-5. [CrossRef]
- Yao, W. X., Ge, S., Zhang, J. Q., Hemmat, P., Jiang, B. Y., Liu, X. J., Lu, X., Yaghi, Z., & Yue, G. H. (2023). Bilateral transfer of motor performance as a function of motor imagery training: a systematic review and meta-analysis. Frontiers in psychology, 14, 1187175. [CrossRef]
- Zambach, S.A., Cai, C., Helms, H.C.C., Hald, B.O., Dong, Y., Fordsmann, J.C., Nielsen, R.M., Hu, J., Lønstrup, M., Brodin, B., et al. (2021). Precapillary sphincters and pericytes at first-order capillaries as key regulators for brain capillary perfusion. Proc. Natl. Acad. Sci, 118, e2023749118.
- Zeng, J., Wang, L., Cai, Q., Wu, J., & Zhou, C. (2022). Effect of hypnosis before general anesthesia on postoperative outcomes in patients undergoing minor surgery for breast cancer: a systematic review and meta-analysis. Gland surgery, 11(3), 588–598. [CrossRef]
- Zhang, Y., Wang, Y., Ku, Y. (2018). Hypnotic and non-hypnotic suggestion to ignore pre-cues decreases space-valence congruency effects in highly hypnotizable individuals. Conscious Cogn, 65, 293-303. [CrossRef]
Figure 2.
Hypnotizability-related correlates relevant to prognosis and treatment. FE, functional equivalence between actual and imagined perception/action; NO, nitiric oxide.
Figure 2.
Hypnotizability-related correlates relevant to prognosis and treatment. FE, functional equivalence between actual and imagined perception/action; NO, nitiric oxide.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



