
Submitted:
27 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
Fusarium wilt diseases severely influence the growth and productivity of numerous crop plants. The consortium of antagonistic rhizospheric Bacillus strains and quercetin were evaluated imperatively as a possible remedy to effectively manage the Fusarium wilt disease of tomato plants. The selection of Bacillus strains was made based on in-vitro antagonistic bioassays against Fusarium oxysporum f.sp. lycoprsici (FOL). Whereas, quercetin was selected after screening a library of phytochemicals during in-silico molecular docking analysis using tomato LysM receptor Kinases “SILKY12” based on its dual role in symbiosis and plant defense responses. After the selection of test materials, pot trials were conducted where tomato plants were provided consortium of Bacillus strains as soil drenching and quercetin as a foliar spray in different concentrations. The combined application of consortium () and quercetin (1.0 mM) reduced the Fusarium wilt disease index up to 69% also resulting in increased plant growth attributes. Likewise, the imperative application of the Bacillus consortium and quercetin (1.0 mM) significantly increased total phenolic contents and activities of the enzymes of the phenylpropanoid pathway. Non-targeted metabolomics analysis was performed to investigate the perturbation in metabolites. FOL pathogen negatively affected a range of metabolites including carbohydrates, amino acids, phenylpropanoids, and organic acids. Thereinto, combined treatment of Bacillus consortium and quercetin (1.0 mM) ameliorated the production of different metabolites in tomato plants. These findings prove the imperative use of Bacillus consortium and quercetin as an effective and sustainable remedy to manage Fusarium wilt disease of tomato plants and to promote the growth of tomato plants under pathogen stress conditions.
Keywords:
fusarium wilt
; bacillus
; quercetin
; antagonism
; induced resistance
1. Introduction
Tomatoes are the second most commercially significant vegetable produced globally [1]. According to estimates, over 160 million tons of tomatoes were produced worldwide in 2017. The Eurotomaton Union, China, Turkey, the United States, and India are the prominent tomato-producing nations. Out of the 160 million tons of tomatoes produced overall, about 40 million tons are processed [2]. Plant diseases can occur across the whole life cycle of crop plants and are one of the biggest threats to human welfare. Plant diseases result in a 13-22% yield loss and billions of dollars are lost in economic terms [3].
Nonetheless, the major constraints in tomato cultivation and reduced yield are due to the phytopathogens. The tomato plants fall susceptible to a variety of biotic and abiotic stresses from the seedling to the maturity stage [4]. The Fusarium oxysporum species are extremely harmful and widely distributed around the world. The fusarium wilt disease of tomato caused by Fusarium oxysporum f.sp. lycopersici is a major pathological constraint affecting tomato crops worldwide. Fusarium wilt is one of the most common and destructive diseases as the pathogen is soil-borne and can be transmitted through water and contaminated soil [5]. The pathogen can persist as chlamydospores in soil and agricultural leftovers for up to six years in the absence of a sensitive host [6]. Any stage of the crop can be easily infected by the F. oxysporum. The pathogen damages the vascular system of the plant and the noticeable symptoms can appear very late until the plant begins bearing fruits. This disease is still a major problem in tomato-growing areas due to the highly variable nature of the pathogen [7].
Beneficial endophytic microbiomes significantly influence plant responses under stressed conditions by mediating the functioning of the plant micro-ecosystem [8]. The biological management strategy for wilt diseases is a durable and economical approach without damaging the environment. The effectiveness of biological control agents can be sometimes comparable to the toxic chemical fungicides used to control wilt disease. Biological control agents have been used for more than a century to ensure the long-term management of several important pathogens of field crops [9]. Different studies reported the use of Bacillus bacteria to manage plant diseases [10,11,12]. The ability to produce resistant endospores and antibiotics makes Bacillus an attractive biological control agent. These microbes are equipped with the production of antibiotics, lytic enzymes, and phytohormones, and can diminish the pathogen infection pressure by competitive exclusion and reducing the growth of pathogens without killing them [13,14]. Secondly, they cause resistance in plants against invading pathogens by the activation of induced systemic resistance [15].
To initiate these beneficial responses, specific signaling molecules are secreted by the microbes which are recognized by the plants [16,17]. Plants are equipped with pattern-recognition receptors (PRRs) used to sense elicitor chemicals (microbial-/pathogen-associated molecular patterns) produced by microbes. PRRs are mainly cell surface recognition proteins playing a vital role in the signaling process that initiates the plant’s immune responses [18]. LysM receptor kinase proteins are members of the PRRs family, responsible for the perception of signal molecules secreted by microbes [19,20].
Secondly, LysM domain proteins are responsible for the reception of the pathogen-associated molecular patterns (PAMPs) that trigger plant immune responses to avoid or limit the invasion of pathogens [21]. In Arabidopsis and rice plants, PAMPs secreted by the pathogens are perceived by LysM proteins [19,22]. LysM and chitin molecules form a heterotetramer complex activating the downstream immune responses [23,24]. Different members of LysM-type receptor kinases have been identified in tomato plants regarding their roles in signaling and symbiosis. Among them, SlLYK12 in tomato plants is mainly involved in microbial colonization [25]. This dual function of SlLYK12 receptor kinases in beneficial symbiosis and pathogen-triggered innate immunity makes them the target of choice in immuno- and bioinformatic studies.
Due to the use of diverse mechanisms by Bacillus bacterial to manage plant disease and the importance of receptor kinases in symbiosis and plant defense responses against invading pathogens, the present study aimed at the imperative use of the consortia of beneficial Bacillus microbes and synthetic chemical agonist capable to dock with LysM receptor kinases SlLYK12 of tomato to manage Fusarium wilt disease. Employing this integrated approach based on the bioassay-guided selection of biocontrol agents and searching synthetic agonists/chemicals by in-silico studies to facilitate symbiosis and trigger receptor kinases-mediated immune responses is likely to be a novel approach for combating plant disease.
2. Materials and Methods
2.1. Isolation and selection of antagonistic rhizospheric bacterial strains
The heat shock method was used for the selective isolation of bacteria belonging to the Bacillus genera [18]. The rhizosphere soil samples were collected from healthy-looking vegetable crop plants in agricultural fields situated in different areas of the Punjab Province, Pakistan. For isolation purposes, one gram soil sample was mixed in distilled sterilized water (10mL) and incubated at 80°C for 10 minutes in the hot water bath [26]. The whole mixture was serially diluted up to 10-7 dilutions using distilled autoclaved water. Finally, 100 µL of the mixture from the last three dilutions was spread on Lysogeny Broth (LB) media plates. The plates were incubated at 30±2°C overnight. All single colonies were cultured on LB plates using the streaking inoculation method.
Antagonistic bacteria were screened in-vitro on PDA medium plates in a dual-culture assay as suggested by Anith, et al. [27]. The 5 mm diameter plugs from F. oxysporum cultured one-week-old culture were transferred to one side of the media plates. The bacterial isolates were inoculated on the other side of the media plate in a straight-line manner. Each treatment was repeated three times. All plates were then kept at 28°C for 5 days. Bacteria were characterized into three categories e.g., bacteria showing zone of growth inhibition; bacteria showing contact inhibition; bacteria showing no inhibition [28]. The bacteria showing a clear zone of growth inhibition were selected for further studies.
2.2. Molecular identification of best-performing bacterial strains
To identify the best-performing strains (IS1, IS6, and IS7), their 16S rDNA motif were amplified and obtained sequences were analyzed. Briefly, a pure culture of bacteria was grown in LB broth medium at 30±2°C, under continuous shaking (180 rpm) in a shaking incubator overnight. Total genomic DNA was extracted with the help of Rapid Bacterial Genomic DNA Isolation Kit (Sangon Biotech). Afterward, routine PCR was performed to amplify the 16S rDNA sequence using the universal primers 27F (5’-AGAGTTTGATCCTGGCTCAG-3´) and 1492R (5´-GGTTACCTTGTTACGACTT-3´) [29]. After successful PCR, the obtained PCR products were sequenced by the Sanger dideoxy method at Macrogen Incorporation (Korea). The Phylogenetic trees were constructed based on the 16S rRNA sequences with the MEGA 6.0 software. The maximum likelihood method was adopted to construct the tree with 1000 bootstrap replications [30].
2.3. Screening of potential agonist/s of tomato receptor-like kinases SlLYK12
2.3.1. Preparation of protein structure and quality analysis
Tomato receptor-like kinase SlLYK12 was chosen as the target protein because of its dual role in plant symbiosis and defense response (). Homology modeling of SlLYK12 was performed using Alphafold to generate a 3D structure from the protein sequence retrieved from the public database (NCBI Accession No. NP_001234725). The quality of generated models was validated as described by Karunarathna, et al. [31]. The Ramachandran plot was generated and VERIFY3D, ERRAT, PROVE, and PROCHECK, criteria were used to analyze the overall quality of the model [32].
2.3.2. Model refinement and validation
The selected protein model was repaired and refined by running the YASRARA md_refine macro provided in the YASARA Structure software [33]. The macro runs a 500 ps simulation and saves snapshots every 25 ps. All parameters were kept at the values defined by the macro. This macro is responsible for energy minimization with combined steepest descent, fixing the backbone atoms to avoid potential damage to the model, and full unrestrained all-atom simulated annealing minimization. The refined model was further subjected to molecular dynamics simulations for >50 ns using the md_run.mcr macro of YASARA as described in Choudhary, et al. [34].
Afterward, the protein model was analyzed and validated using the ProtParam tool of ExPASy Proteomics Server for various parameters such as estimated half-life, theoretical pI, instability index, aliphatic index, and grand average of hydropathicity (GRAVY) [35].
2.3.3. Virtual screening by molecular docking
The database of phytochemicals was downloaded from the Plant Secondary Compound Database http://pscdb.appsbio.utalca.cl/viewIndex/index.php. These included alkaloids, flavonoids, phenylpropanoids, and phytohormones. Before docking, ligands were prepared to perform the following: conversions of 2D to 3D, energy minimization, the addition of hydrogen atoms, and necessary structural optimizations using the Openbebal program. Similarly, receptor molecule was prepared by adding polar hydrogen atoms and removing water molecules on Discovery Studio software. The molecular docking studies of active compounds were performed using AutoDock Vina by PyRx virtual screening software. A blind docking strategy was adopted because a completely new receptor molecule was used with no previous knowledge of potential binding sites for ligands [36,37,38]. Preliminary docking was performed to screen the whole legend library and separate the legend/s fulfilling the selection criteria. The results for receptor-ligand complexes were arranged in ascending energy order to determine the interaction profile. Ligand/s showing maximum vina score and commercial availability were chosen. Afterward, re-docking of the selected legend was performed. The selected conformation of the top order ligand/s associated with SlLYK12 receptor kinases was analyzed to dissect the interaction profiles by Discovery Studio software and Pylip online server. PyMOL was used as molecular visualization software.
2.4. In-vivo effect of the synthetic elicitor and consortium of Bacillus strains on Fusarium wilt disease development and growth of tomato plants
The combined application of biotic and synthetic elicitor was performed by soil drenching and foliar application, respectively. Before seed priming, antagonistic bacteria were analyzed for consortia compatibility by growing them in a single petri plate as streaked as the cross pattern. For soil drenching, an aqueous suspension of the consortia bacterial strains was made. All the bacteria strains were raised together in an LB broth medium overnight and cells were collected by centrifugation. Bacterial suspension was prepared in distilled autoclaved water and the concentration was adjusted to 106-7 c.f.u/mL by taking O.D of 1 at 600nm. Plastic pots were filled with sterilized potting mix and fifty mL of aqueous suspension of bacteria was added in the allotted pots. The next day, surface sterilized tomato seeds were cultivated in the pots. After emergence, thinning was performed to ensure uniformity of seedlings, and one healthy seedling was left in the pot. Additionally, a chemical elicitor (quercetin) was applied as a foliar application at varying concentrations (1.0, 0.1, 0.01 mM) twenty days post-emergence according to the experimental design as described in Table 1. Ten days after quercetin application, conidial suspension of FOL was prepared as mentioned in [39] and 50 mL of that was added to the rhizosphere of the tomato seedlings. Plants receiving FOL served as pathogen control. Whereas, plants raised from only distilled autoclaved water-primed seeds and sprayed with distilled autoclaved water served as non-treated control. The fungicide control consisted of the soil drenching of Carbendazim fungicide (1.5 g/L) as suggested by [40].
Five biological replicate pots were included in each treatment and the whole experiment was repeated twice. Plants were irrigated with distilled sterilized water and kept at green house under natural daylight conditions. After twenty days of pathogen application, the disease scoring was performed and the disease index was determined as described by Hua, et al. [41] and Wei, et al. [42] respectively. Additionally, growth attributes of tomato plants like shoot length, root length, shoot biomass, and root biomass were also determined at harvest. Total chlorophyll contents were analyzed by spectrophotometer as described by Arnon [43].
2.5. Analysis of the biochemical basis of induced defense responses in tomato plants against Fusarium wilt disease
Another independent pot experiment was performed using the same strategy as mentioned in the previous section using selected treatments but with a shorter duration. Hereafter five days of pathogen application the harvest was taken. This experiment was performed for the elucidation of mechanisms behind disease suppression. The treatments were as following: Con= Untreated control, PC= Pathogen control, T1= Consortia + Quercetin (1.0 mM) + FOL, T2= Consortia + FOL, and T3= Quercetin (1.0 mM) + FOL.
2.5.1. Analysis of total phenolics and plant defense-related enzymes
The quantification of total phenolic compounds and enzymes involved in phenylpropanoid pathways was performed five days after pathogen application. Leaf samples were used for extraction and quantification purposes. The total phenolic compounds were analyzed by the Folin ciaocaltue method (Singleton and Rossi, 1965). The leave samples were ground into a fine powder using liquid nitrogen and enzyme extraction was performed in 0.1 M sodium phosphate buffer (pH 7.5) as described by Sundaramoorthy, et al. [44]. The peroxidase, polyphenol oxidase, and phenylalanine ammonia-lyase (PAL) activities were estimated as described by Dalisay and Kuć [45]; Mozzetti, et al. [46] and Cahill and McComb [47] respectively.
2.5.2. Analysis of the metabolomic profile of tomato plants
A non-targeted metabolomic analysis was performed on the UHPLC triple quadrupole MS/MS apparatus (Agilent, CA, USA) to observe the changes in the metabolomic profile of the tomato plants. For that purpose, leaves from the plants of different treatments were ground to powder material in the pestle mortar using liquid nitrogen. The mixture of (methanol/water, 80/20, v/v) containing 1 ng/µL of the internal standard was used for extraction as described by Jo, et al. [48]. The extract was centrifuged and passed by a microfilter assembly. Chromatographic separation was performed on an Agilent 1200 ultra-performance liquid chromatography system (Agilent, USA) fitted with a C18 analytical column (Agilent). The data for the identification and quantification of compounds was obtained from a Triple Quad tandem mass spectrometer (6470) system equipped with an Electrospray Ionization Source (ESI). QC sample was made after pooling the samples of all treatments in a single vial in equal quantities. The chromatographic conditions were adopted as mentioned in Rossouw et al. (2019). The MS scan range was 50−1500 m/z with a 100 ms scan time. The acquired mass data was converted into an mzxml format. Afterward, the data was loaded onto MZmine 2.53 software and further processed as described in Li, et al. [49]. Identification of compounds was performed using the NIST MS/MS library and previously published literature.
2.6. Statistical analysis
All the experiments were repeated twice and mean values are provided. The data were analyzed statistically. The analysis of ANOVA and DNMRT was performed by using the Excel addon “DSAASTAT” developed by Onofri (Italy) [50].
3. Results
3.1. Selection of antagonistic rhizospheric bacterial strains
Altogether, 11 different bacterial isolates were isolated from the rhizosphere of tomato plants using the heat shock method aimed at Bacillus genera. The purified bacterial isolates were screened for the presence of antagonistic activity against FOL using a dual culture assay. The analysis showed that out of the 11 isolates 05 could successfully inhibit the growth of fusarium wilt pathogen (Table 2). In these cases, three bacterial isolates (BS6, BS7, and BS9) showing a clear zone of inhibition were separated (Figure 1; Table 2). Two of the bacterial isolates could inhibit the growth of the pathogen upon contact with a fungal pathogen colony. These were denoted as bacterial isolates having contact inhibition activity. The rest of the bacterial strains showed no inhibition activity (Table 2). Bacteria showing a clear zone of inhibition were selected for further experimentation.
3.2. Molecular identification of bacterial strains
Three bacterial strains with antagonistic activity against FOL were identified by the amplification and sequencing of the 16rRNA region. The BLAST analysis showed that the bacterial isolates BS6, BS7, and BS9 shared >99% homology with Bacillus velezensis (NCBI Accession No. LC516497), Bacillus thuringiensis (NCBI Accession No. LT838123) and Bacillus fortis (NCBI Accession No. MG563939), respectively (Figure 2). The obtained sequences were aligned with the reference sequences downloaded from the NCBI database using the Clustal W multiple alignment program. Whereas, MEGA version 10 software was utilized to produce a maximum likelihood phylogenetic tree (Figure 2).
3.3. Screening of potential agonist/s of tomato receptor-like kinases SlLYK12
Before the molecular docking, the SlLYK12 model was refined and the quality analysis of the refined SlLYK12 protein was performed by online quality structure assessment tools (Table ). Here no residue of the SlLYK12 was in the disallowed region, whereas >90% of residues were found in the favored region (Table 3). The quality analysis using VERIFY3D, ERRAT, and PROCHECK servers indicated a good overall quality of the SlLYK12 protein model. The refined SlLYK12 model was further analyzed for physicochemical parameters such as theoretical isoelectric point, estimated half-life, instability index, aliphatic index, and the grand average hydropathicity (Table S1). The half-life of SlLYK12 (30 h), the instability index (<40), the higher aliphatic index (87.62) and the lower GRAVY value (-0.05) of our refined SlLYK12 model present the good quality of the receptor representing the stable receptor model (Table S1).
The molecular docking analysis was performed using a library of phytochemicals carefully developed keeping in view the commercial availability and economic feasibility. The top score legends are mentioned in Table 4 with binding affinity >8 after performing a preliminary docking analysis. Quercetin (8.7 binding affinity) was selected based on the top highest affinity scores and devised screening criteria to avoid a lengthy study. The SlLYK12 + quercetin complex comprised three hydrogen bonds and the rest of the intermolecular bond e.g., van der Waals force, pi alkyl (Figure 3) as depicted by the Discovery Studio.
3.4. In-vivo effect of the synthetic elicitor and consortium of Bacillus strains on Fusarium wilt disease development and growth of tomato plants
In pot trials, different treatments consisting of synthetic elicitor (quercetin) and bacterial consortia in different combinations were assessed for their efficiency in suppressing fungal wilt disease. A varied degree of protection was observed ranging 17–69% under greenhouse conditions. The highest level of disease index was observed in the pathogen control treatment. The valuation of fusarium wilt protection showed that the combination of both synthetic elicitor (quercetin) and bacterial consortia was significantly higher than disease protection provided by the quercetin or bacterial consortia individually (Figure 4). A combination of quercetin (1.0mM) and bacterial consortia reduced the disease index up to 69.63% as compared to the pathogen control (Figure 4). Similarly, another treatment consisting of the combined applications of quercetin (0.1 mM) and bacterial consortia reduced the disease index up to 51.36% compared to the pathogen control. The foliar application of quercetin alone significantly reduced disease index up to 29.27- (0.01 mM) and 37.61% (0.1 mM) over pathogen control. Treatment with bacterial consortia alone reduced disease by>35% compared to the pathogen control (Figure 4).
Regarding growth parameters, the minimum shoot length, root length, shoot biomass, root biomass, and chlorophyll contents were related to the pathogen control plants, followed by carbendazim fungicide control (Table 5). The rest of the treatments significantly increased the aforementioned growth attributes of tomato plants compared to the pathogen control (Table 5). The combined application of quercetin (1.0 mM) and bacterial consortia increased shoot and root- biomasses nearly 2.0-fold compared to pathogen control plants respectively. These outcomes made it evident that the combined treatment of quercetin (1.0 mM) and bacterial consortia effectively rescued the growth of tomato plants attacked by the fusarium wilt pathogen and made it comparable to the non-treated control plants in some instances (Table 5). Hence, based on the findings of pot trials the treatment consisting of the combined application of quercetin (1.0mM) and bacterial consortia was selected to elucidate the possible mechanisms behind disease suppression. It is to be mentioned here that the rest of the treatments including consortia alone also significantly increased the growth attributes of tomato plants but the application of quercetin combined with the consortia showed a more pronounced increase in the growth attributes of tomato plants.
3.5. Analysis of the biochemical basis of induced defense responses in tomato plants against Fusarium wilt disease
3.5.1. Analysis of total phenolics and defense-related enzymes
Now the efforts were diverted to determine the role of quercetin (1.0mM) and bacterial consortia in the induction of tomato systemic resistance through the comparative quantification of enzymes involved in phenylpropanoid pathways and phenolic compounds.
The results found that a combination of quercetin (1.0mM) and bacterial consortia had a higher significant effect on the inducible production of total phenolics, and activities of PO, PPO, and PAL enzymes than those of single elicitor treatments (Table 6). The combined applications have increased to 1.42-, 1.79-, 2.36- and 1.34-fold increase in the quantities of total phenolics, PO, PPO, and PAL enzymes compared to the pathogen control (Table 6). The outcomes further confirm that the treatment of quercetin and consortia alone a potential inducers in defense responses but to a lesser extent compared to the combined application of quercetin and bacterial consortia (Table 6).
3.5.2. Non-targeted metabolomic analysis
The up-regulated defense system in tomato plants was further analyzed by performing non-targeted metabolomic analysis. The metabolomic data was obtained from UHPLC-triple quadrupole -MS/MS analysis. The representative chromatograms indicate varying levels of different metabolites among different treatments (Figure 5). The quantities of phenylpropanoids including ferulic acid, cinnamic acid, caffeic acid, and gallic acid,) positively responded to the quercetin (1.0mM) and bacterial consortia in either combination (Figure 6 and Figure 7). Most of the phenylpropanoid metabolites showed higher values of fold change in combined treatment (quercetin 1.0mM + bacterial consortia) compared to the rest of the treatments (Figure 6).
The rest of the metabolites interfering with plant physiology belonged to carbohydrates, amino acids, alkaloids, and organic acids, etc. (Figure 6). Given that tomato plants exhibited a more evident physiological response to the combined application of quercetin 1.0mM + bacterial consortia followed by the fusarium wilt pathogen. The metabolic response was also significant for the rest of the treatments (Figure 6). For pathogens alone, the effect on major classes of compounds was clear compared to that of non-treated control plants. Similarly, the metabolomics responses varied clearly in tomato plants treated with quercetin and bacterial consortia alone compared to the non-treated control plants (Figure 6).
4. Discussion
Tomato is a valuable vegetable crop in the Solanaceae family. Tomatoes are grown both in open fields and in greenhouses. This crop generates a high return for farmers and provides plenty of job opportunities for rural residents [51]. Rhizospheric beneficial bacteria have several strategies, both direct and indirect, to manage plant diseases [52]. In this study, an imperative approach was used based on the consortia of antagonistic Bacillus microbes and synthetic elicitors capable of triggering symbiosis and plant immunity, to manage Fusarium wilt disease of tomato plants.
The plant disease suppression mediated by the Bacillus spp. rely on the direct antagonistic effects of antimicrobial metabolites produced by bacteria against pathogens or by activation of defense responses in the host plant [53]. In the first phase of the present study, we isolated Bacillus isolates using the heat shock method from the rhizospheric soil and demonstrated their antagonistic capabilities in in-vitro studies. The strains were differentiated into non-antagonists, antagonists with contact inhibition, and antagonists with the clear zone of inhibition. The three Bacillus isolates (BS6, BS7, and BS9) showed a clear zone of inhibition against the radial growth of F. oxysporum. The isolates were identified as Bacillus velezensis BS6, B. thuringiensis BS7, and B. fortis BS9 based on 16s rRNA gene sequencing. These findings are in line with some previous studies that Bacillus species, including Bacillus velezensis, B. thuringiensis, and B. fortis strains have antagonistic activities against different plant pathogens [54,55,56,57,58,59]. These antagonistic properties can be attributed to the production of antifungal lipopeptides by Bacillus species with antifungal activities for different phytopathogenic fungi including F. oxysporum [60]. The findings of this preliminary study provide us with a basis to develop consortia of these antagonistic Bacillus isolates for the management of Fusarium wilt during subsequent experiments.
Receptor-like kinases play important roles in plant immunity. Plants used a large number of receptor kinases as pattern recognition receptors for the detection of microbial-derived molecular patterns to initiate inducible defense [61]. Next, in-silico studies were performed to find synthetic agonists capable of binding with the receptor-like kinases “SILKY12” of tomato plants. The homology modeling is used to obtain a 3D representation of the target receptor whenever experimental structures are not available for docking. The use of DeepMind’s artificial intelligence model, AlphaFold (AF), set a milestone within the field of homology modeling [62]. The SlLYK12 model was created with the Alphafold, online server. The model showed the superior quality/criteria scores devised by the modeling servers [35]. Further quality analysis of the selected models from each modeling server was performed by online quality structure assessment tools available on the Saves server. The quality analysis using VERIFY3D, ERRAT, and PROCHECK servers also indicated a good overall quality of the trRosetta model. Henceforth, validation scores suggest that trRosetta-modelled SlLYK12 can be used for further molecular docking analysis. The refined trRosetta model was further analyzed for physicochemical parameters such as theoretical isoelectric point, estimated half-life, instability index, aliphatic index, and the grand average hydropathicity. Protein-ligand blind docking strategy was adopted as it is considered a powerful method to explore the best binding sites of receptor molecules and the binding conformation of ligands. Among five top score legends, quercetin was selected based on the top highest affinity scores and screening criteria to avoid a lengthy study. The SlLYK12 + quercetin complex was stabilized by the presence of three hydrogen bonds and the rest of the intermolecular bond e.g., van der Waals force, and hydrophobic contacts. The presence and number of hydrogen bonding indicated that the agonist could trigger the SlLYK12 receptor kinase to modulate further downstream processes [63]. Similarly, the presence of hydrophobic contacts indicates stable protein folding, biological activeness, and reduced undesirable interactions [64].
Ultimately, the treatments for pot trials were devised based on the findings of these preliminary studies. The Consortia capability of three strains was analyzed before the pot trial. Afterward, consortia were applied as soil drenching and quercetin as a foliar amendment. Here lower concentrations of quercetin were used (1.0, 0.1, and 0.01) to ensure the economical feasibility of the whole process. The results of pot trials showed that the combined use of consortia of Bacillus microbes in association with the quercetin (1.0 mM) proved to be an effective strategy to manage Fusarium wilt disease of tomato as well as increased plant growth. The combined treatment based on consortium and quercetin reduced the disease index by>60% and significantly improved plant growth attributes including shoot length, root length, shoot biomass, root biomass and total chlorophyll contents in tomato plants. The findings of the pot trial were found consistent with some previous studies, that showed the combined use of beneficial microbes along with the synthetic elicitor can provide better protection against both biotic and abiotic stress conditions [65,66]. The application of Bacillus consortia can produce antibiotic substances, which can play a critical role in the suppression of the pathogen population inside plant rhizosphere. In addition, the application of quercetin can enhance the symbiotic beneficial impact of consortia, and can further rescue plant growth under disease stress conditions. As observed in the pot trial, the integration of Bacillus consortium (soil drenching) alongside quercetin (foliar spray) greatly improved the growth-related attributes compared to the treatment where both were applied alone, possibly due to the collaboration with phytohormones synthesis, the increased cell division and better performance of photosynthesis machinery.
A thorough understanding of the interactions between tomato plants, Bacillus consortium, and quercetin having an effective impact on plant disease suppression and development is required. To this end, another independent experiment was performed to elucidate the mechanisms behind disease suppression mediated by the combined application of a consortium of Bacillus strains and quercetin. Consequently, it was observed that the combined approach was more stimulatory to increase the production of total phenolic compounds and enzymes involved in the phenylpropanoid pathway like peroxidase, polyphenol oxidase, and phenylalanine ammonia-lyase than the Bacillus consortium or quercetin alone. Peroxidase is an important defense protein that contributes to the biosynthesis of lignin [67] and antimicrobial phytoalexins inside plants. Similarly, polyphenol oxidase and phenylalanine ammonia-lyase play an important role in the biosynthesis of toxic substances via the phenylpropanoid pathway to hinder the growth of pathogens inside plants [68]. In a previous study, the induced resistance through increased production of defense-related enzymes in tomato plants treated with rhizospheric microbes and chemical elicitor salicylic acid [69]. Their findings proved that upon pathogen inoculation, relatively higher activities of defense-related enzymes were seen treated with both bacterial and chemical elicitors compared to the plants only receiving pathogen inoculum.
A non-targeted metabolomic analysis was performed to further improve our understanding to this end. The findings were comparable as mentioned in the previous section. A large number of perturbations were seen in the metabolic profile of tomato plants under different treatments. It could be due to the reason that Bacillus consortium in combination with quercetin greatly affected the physiology of plants as the varying abundance of a range of metabolites including carbohydrates, amino acids, organic acids, phenylpropanoids, polyols, flavonoids, etc., was seen across different treatments. Surprisingly, the metabolic reprogramming in tomato plants exposed to the pathogen and Bacillus consortium+ quercetin was more extensive compared to the other treatments. The representative chromatograms and heatmap showed vividly that combined treatment influenced the metabolites in tomato plants differently under the attack of a pathogen from the treatments where Bacillus consortium and quercetin were applied alone. The pathogen attack negatively affected the production of a range of metabolites with few exceptions. Whereas, the application of Bacillus consortium and quercetin either alone and/or in combination seemed to restore the production of several metabolites in tomato plants. The application of the Bacillus consortium and quercetin increased the production of different phenolic acids in tomato plants subsequently challenged with the pathogen. These changes in the levels of phenylpropanoids may affect the ability of the tomato plant to restrict the growth and colonization of pathogens, reflected in a lowered disease index in pot trials [70]. Similarly, increased production of carbohydrates amino acids, sugars, and polyols could be attributed to the higher availability of raw material for building blocks in tomato plants leading to increases in growth-related parameters [71]. These findings are of great importance to understanding a complex mechanism governing disease suppression and growth enhancement under the influence of beneficial microbes and synthetic elicitors. Additional omics can also be used to further understand the response of plants to non-toxic chemicals and beneficial microbes in the presence of a nasty pathogen.
5. Conclusions
We have demonstrated that the imperative use of Bacillus consortium and quercetin application suppresses the Fusarium wilt disease of tomato and significantly enhances tomato defense responses and growth parameters. In the presence of beneficial microbes, quercetin can act as a cofactor to ensure symbiosis, elicit plant defense responses, and regulate physiological traits. The quercetin can be used as an additive in the beneficial microbes-based formulation to facilitate symbiosis and trigger receptor kinases-mediated immune responses. In conclusion, our findings provide practical information for the imperative use of beneficial microbes and chemical elicitors to manage plant disease.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, W.A., investigation, A. H. and S. H., methodology, A. H. and W. A.; resources, T. A. and Z. A.; supervision, W. A. and T. A.; validation, W. A., writing—original draft preparation, A. H, and W. A., project administration, H. R.; funding acquisition, M. H. A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Researchers Supporting Project number (RSPD2023R1048), King Saud University, Saudi Arabia.
Data Availability Statement
The data supporting the findings of this study are available within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hussain, I.; Alam, S.S.; Khan, I.; Shah, B.; Naeem, A.; Khan, N.; Ullah, W.; Iqbal, B.; Adnan, M.; Junaid, K. Study on the biological control of fusarium wilt of tomato. J Entomol Zool Studies 2016, 4, 525–528. [Google Scholar]
- Pathak, T.B.; Stoddard, C.S. Climate change effects on the processing tomato growing season in California using growing degree day model. Modeling Earth Systems and Environment 2018, 4, 765–775. [Google Scholar] [CrossRef]
- Venbrux, M.; Crauwels, S.; Rediers, H. Current and emerging trends in techniques for plant pathogen detection. Frontiers in Plant Science 2023, 14, 1120968. [Google Scholar] [CrossRef]
- Brahimi, M.; Boukhalfa, K.; Moussaoui, A. Deep learning for tomato diseases: classification and symptoms visualization. Applied Artificial Intelligence 2017, 31, 299–315. [Google Scholar] [CrossRef]
- Chacón, C.; Bojórquez-Quintal, E.; Caamal-Chan, G.; Ruíz-Valdiviezo, V.M.; Montes-Molina, J.A.; Garrido-Ramírez, E.R.; Rojas-Abarca, L.M.; Ruiz-Lau, N. In vitro antifungal activity and chemical composition of Piper auritum Kunth essential oil against Fusarium oxysporum and Fusarium equiseti. Agronomy 2021, 11, 1098. [Google Scholar] [CrossRef]
- Asif, M.; Haider, M.S.; Akhter, A. Impact of Biochar on Fusarium Wilt of Cotton and the Dynamics of Soil Microbial Community. Sustainability 2023, 15, 12936. [Google Scholar] [CrossRef]
- Chitwood-Brown, J.; Vallad, G.E.; Lee, T.G.; Hutton, S.F. Breeding for resistance to Fusarium wilt of tomato: A review. Genes 2021, 12, 1673. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.; Liu, L.; Jin, X.; Duan, C.; Cui, Y.; Wang, J.; Ma, D.; Zhao, W.; Wang, Y.; Fang, L. Co-inoculation effect of plant-growth-promoting rhizobacteria and rhizobium on EDDS assisted phytoremediation of Cu contaminated soils. Chemosphere 2020, 254, 126724. [Google Scholar] [CrossRef] [PubMed]
- Heimpel, G.E.; Mills, N.J. Biological control; Cambridge University Press: 2017.
- Anckaert, A.; Arguelles Arias, A.; Hoff, G.; Calonne-Salmon, M.; Declerck, S.; Ongena, M. The use of Bacillus spp. as bacterial biocontrol agents to control plant diseases. 2021. [Google Scholar]
- Wang, X.; Zhao, D.; Shen, L.; Jing, C.; Zhang, C. Application and mechanisms of Bacillus subtilis in biological control of plant disease. Role of rhizospheric microbes in soil: Volume 1: Stress management and agricultural sustainability 2018, 225–250. [Google Scholar] [CrossRef]
- Cawoy, H.; Bettiol, W.; Fickers, P.; Ongena, M. Bacillus-based biological control of plant diseases. Pesticides in the modern world-pesticides use and management 2011, 273–302. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, Y.; Liu, Y.; Zhang, R. Research progress in biocontrol of Bacillus spp. against plant diseases. Jiangsu Journal of Agricultural Sciences 2012, 28, 999–1006. [Google Scholar]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: a review. Biotechnology & Biotechnological Equipment 2017, 31, 446–459. [Google Scholar] [CrossRef]
- Patel, S.; Saraf, M. Interaction of root colonizing biocontrol agents demonstrates the antagonistic effect against Fusarium oxysporum f. sp. lycopersici on tomato. European Journal of plant pathology 2017, 149, 425–433. [Google Scholar] [CrossRef]
- Smith, D.L.; Praslickova, D.; Ilangumaran, G. Inter-organismal signaling and management of the phytomicrobiome. Frontiers in plant science 2015, 6, 722. [Google Scholar] [CrossRef]
- Bukhat, S.; Imran, A.; Javaid, S.; Shahid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defense signaling. Microbiological research 2020, 238, 126486. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; He, S.Y. Innate immunity in plants: an arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–744. [Google Scholar] [CrossRef] [PubMed]
- Gimenez-Ibanez, S.; Ntoukakis, V.; Rathjen, J.P. The LysM receptor kinase CERK1 mediates bacterial perception in Arabidopsis. Plant signaling & behavior 2009, 4, 539–541. [Google Scholar] [CrossRef]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proceedings of the National Academy of Sciences 2007, 104, 19613–19618. [Google Scholar] [CrossRef] [PubMed]
- Shirron, N.; Yaron, S. Active suppression of early immune response in tobacco by the human pathogen Salmonella Typhimurium. PLoS One 2011, 6, e18855. [Google Scholar] [CrossRef] [PubMed]
- Gimenez-Ibanez, S.; Hann, D.R.; Ntoukakis, V.; Petutschnig, E.; Lipka, V.; Rathjen, J.P. AvrPtoB targets the LysM receptor kinase CERK1 to promote bacterial virulence on plants. Current biology 2009, 19, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. elife 2014, 3, e03766. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant pattern-recognition receptors. Trends in immunology 2014, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Sun, X.; Wang, N.; Song, F.; Liang, Y. Tomato LysM receptor-like kinase SlLYK12 is involved in arbuscular mycorrhizal symbiosis. Frontiers in Plant Science 2018, 9, 1004. [Google Scholar] [CrossRef]
- Sulistiyani, T.R.; Kusmiati, M.; Putri, G.A. The 16S rRNA analysis and enzyme screening of Bacillus from rhizosphere soil of Lombok Island. Jurnal Ilmu Pertanian Indonesia 2021, 26, 582–590. [Google Scholar] [CrossRef]
- Anith, K.; Radhakrishnan, N.; Manomohandas, T. Screening of antagonistic bacteria for biological control of nursery wilt of black pepper (Piper nigrum). Microbiological Research 2003, 158, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Palys, S.; Di Falco, M.; Tsang, A.; Périnet, P.; Ramanan, U.S.; Dayanandan, S. Isolation and Characterization of Bacillus velezensis EB14, an Endophytic Bacterial Strain Antagonistic to Poplar Stem Canker Pathogen Sphaerulina musiva and Its Interactions with the Endophytic Fungal Microbiome in Poplar. PhytoFrontiers™ 2021, 1, 229–238. [Google Scholar] [CrossRef]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Applied and environmental microbiology 1997, 63, 3233–3241. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Molecular biology and evolution 2018, 35, 1547. [Google Scholar] [CrossRef]
- Karunarathna, K.H.T.; Senathilake, N.; Mewan, K.M.; Weerasena, O.; Perera, S. In silico structural homology modelling of EST073 motif coding protein of tea Camellia sinensis (L). Journal of Genetic Engineering and Biotechnology 2020, 18, 1–10. [Google Scholar] [CrossRef]
- Vriend, G. WHAT IF: a molecular modeling and drug design program. Journal of molecular graphics 1990, 8, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Vollan, H.S.; Caugant, D.A.; Eldholm, V.; Alfsnes, K.; Debech, N.; Brynildsrud, O. Naturally occurring Neisseria gonorrhoeae can have large deletions in housekeeping gene abcZ, making them untypable with multilocus sequence typing. Microbial Genomics 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Janjanam, J.; Kumar, S.; Kaushik, J.K.; Mohanty, A.K. Structural and functional characterization of buffalo oviduct-specific glycoprotein (OVGP1) expressed during estrous cycle. Bioscience Reports 2019, 39. [Google Scholar] [CrossRef]
- Mora Lagares, L.; Minovski, N.; Caballero Alfonso, A.Y.; Benfenati, E.; Wellens, S.; Culot, M.; Gosselet, F.; Novič, M. Homology modeling of the human p-glycoprotein (Abcb1) and insights into ligand binding through molecular docking studies. International journal of molecular sciences 2020, 21, 4058. [Google Scholar] [CrossRef] [PubMed]
- Forli, S.; Olson, A.J. A force field with discrete displaceable waters and desolvation entropy for hydrated ligand docking. Journal of medicinal chemistry 2012, 55, 623–638. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.-X.; Cao, Y. CB-Dock2: Improved protein–ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Research 2022, 50, W159–W164. [Google Scholar] [CrossRef]
- Meng, X. Y, Zhang H-X, Mezei M, Cui M. Molecular docking: a powerful approach for structure-based drug discovery. Curr. Comp. Aided Drug Design 2011, 7, 146–157. [Google Scholar] [CrossRef] [PubMed]
- PURWATI, R.D.; Hidayah, N. Inoculation methods and conidial densities of Fusarium oxysporum f. sp. cubense in Abaca. HAYATI Journal of Biosciences 2008, 15, 1–7. [Google Scholar] [CrossRef]
- Abbasi, S.; Safaie, N.; Sadeghi, A.; Shamsbakhsh, M. Streptomyces strains induce resistance to Fusarium oxysporum f. sp. lycopersici race 3 in tomato through different molecular mechanisms. Frontiers in Microbiology 2019, 10, 1505. [Google Scholar] [CrossRef] [PubMed]
- Hua, G.K.H.; Timper, P.; Ji, P. Meloidogyne incognita intensifies the severity of Fusarium wilt on watermelon caused by Fusarium oxysporum f. sp. niveum. Canadian Journal of Plant Pathology 2019, 41, 261–269. [Google Scholar] [CrossRef]
- Wei, L.; Jiangchun, H.; Shujin, W. Growth-promotion and biocontrol of cucumber fusarium wilt by marine Bacillus subtilis 3512A. Journal of Shenyang Agricultural University 2008. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant physiology 1949, 24, 1. [Google Scholar] [CrossRef]
- Sundaramoorthy, S.; Raguchander, T.; Ragupathi, N.; Samiyappan, R. Combinatorial effect of endophytic and plant growth promoting rhizobacteria against wilt disease of Capsicum annum L. caused by Fusarium solani. Biological Control 2012, 60, 59–67. [Google Scholar] [CrossRef]
- Dalisay, R.; Kuć, J. Persistence of reduced penetration by Colletotrichum lagenarium into cucumber leaves with induced systemic resistance and its relation to enhanced peroxidase and chitinase activities. Physiological and Molecular Plant Pathology 1995, 47, 329–338. [Google Scholar] [CrossRef]
- Mozzetti, C.; Ferraris, L.; Tamietti, G.; Matta, A. Variation in enzyme activities in leaves and cell suspensions as markers of incompatibility in different Phytophthora-pepper interactions. Physiological and Molecular Plant Pathology 1995, 46, 95–107. [Google Scholar] [CrossRef]
- Cahill, D.M.; McComb, J.A. A comparison of changes in phenylalanine ammonia-lyase activity, lignin and phenolic synthesis in the roots of Eucalyptus calophylla (field resistant) and E. marginata (susceptible) when infected with Phytophthora cinnamomi. Physiological and Molecular Plant Pathology 1992, 40, 315–332. [Google Scholar] [CrossRef]
- Jo, H.E.; Song, K.; Kim, J.-G.; Lee, C.H. Non-targeted metabolomic analysis for the comparative evaluation of volatile organic compounds in 20 globally representative cucumber lines. Frontiers in Plant Science 2022, 13, 1028735. [Google Scholar] [CrossRef]
- Li, X.; Wang, P.; Tong, Y.; Liu, J.; Shu, G. UHPLC-Q-Exactive orbitrap MS/MS-Based untargeted metabolomics and molecular networking reveal the differential chemical constituents of the bulbs and flowers of Fritillaria thunbergii. Molecules 2022, 27, 6944. [Google Scholar] [CrossRef]
- Onofri, A. Routine statistical analyses of field experiments by using an Excel extension. In Proceedings of the Proceedings 6th National Conference Italian Biometric Society:“La statistica nelle scienze della vita e dell’ambiente”, Pisa, 2007; pp. 93–96. [Google Scholar]
- Mari, F.; Memon, R.; Lohano, H. Measuring returns to scale for onion, tomato and chillies production in Sindh province of Pakistan. International Journal of Agriculture and Biology (Pakistan) 2007. [Google Scholar]
- Deng, X.; Zhang, N.; Shen, Z.; Zhu, C.; Li, R.; Salles, J.F.; Shen, Q. Rhizosphere bacteria assembly derived from fumigation and organic amendment triggers the direct and indirect suppression of tomato bacterial wilt disease. Applied soil ecology 2020, 147, 103364. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P.; Touré, Y.; Destain, J.; Jabrane, A.; Thonart, P. Involvement of fengycin-type lipopeptides in the multifaceted biocontrol potential of Bacillus subtilis. Applied microbiology and biotechnology 2005, 69, 29–38. [Google Scholar] [CrossRef]
- Adrees, H.; Haider, M.S.; Anjum, T.; Akram, W. Inducing systemic resistance in cotton plants against charcoal root rot pathogen using indigenous rhizospheric bacterial strains and chemical elicitors. Crop protection 2019, 115, 75–83. [Google Scholar] [CrossRef]
- Mojica-Marín, V.; Luna-Olvera, H.A.; Sandoval-Coronado, C.F.; Pereyra-Alférez, B.; Morales-Ramos, L.H.; Hernández-Luna, C.E.; Alvarado-Gomez, O.G. Antagonistic activity of selected strains of Bacillus thuringiensis against Rhizoctonia solani of chili pepper. African Journal of Biotechnology 2008, 7. [Google Scholar]
- Yan, H.; Qiu, Y.; Yang, S.; Wang, Y.; Wang, K.; Jiang, L.; Wang, H. Antagonistic activity of Bacillus velezensis SDTB038 against Phytophthora infestans in Potato. Plant Disease 2021, 105, 1738–1747. [Google Scholar] [CrossRef] [PubMed]
- Rayavarapu, V.B.; Padmavathi, T. Bacillus sp. as potential plant growth promoting rhizobacteria. Int J Adv Life Sci 2016, 9, 29–36. [Google Scholar]
- Bisen, K.; Singh, V.; Keswani, C.; Ray, S.; Sarma, B.K.; Singh, H. Use of Biocontrol Agents for the Management of Seed-Borne Diseases. Seed-Borne Diseases of Agricultural Crops: Detection, Diagnosis & Management 2020, 651–663. [Google Scholar]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annual review of phytopathology 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Gond, S.K.; Bergen, M.S.; Torres, M.S.; White Jr, J.F. Endophytic Bacillus spp. produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiological research 2015, 172, 79–87. [Google Scholar] [CrossRef]
- Sun, Y.; Qiao, Z.; Muchero, W.; Chen, J.-G. Lectin receptor-like kinases: the sensor and mediator at the plant cell surface. Frontiers in plant science 2020, 11, 596301. [Google Scholar] [CrossRef] [PubMed]
- Scardino, V.; Di Filippo, J.I.; Cavasotto, C.N. How good are AlphaFold models for docking-based virtual screening? Iscience 2023, 26. [Google Scholar] [CrossRef] [PubMed]
- Oltrogge, L.M.; Boxer, S.G. Short hydrogen bonds and proton delocalization in green fluorescent protein (GFP). ACS Central Science 2015, 1, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Fu, H.; Fryar, K.L.; Landua, J.; Trevino, S.R.; Shirley, B.A.; Hendricks, M.M.; Iimura, S.; Gajiwala, K.; Scholtz, J.M.; et al. Contribution of Hydrophobic Interactions to Protein Stability. Journal of Molecular Biology 2011, 408, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.; Omara, A.E.D.; Ahmed, A. The coupling effects of plant growth promoting rhizobacteria and salicylic acid on physiological modifications, yield traits, and productivity of wheat under water deficient conditions. Agronomy 2019, 9, 524. [Google Scholar] [CrossRef]
- Mufti, R.; Bano, A.; Munis, M.F.H.; Andleeb, T.; Quraishi, U.M.; Khan, N. Integrated Application of Salicylic Acid and PGPRs to Control Fusarium Wilt of Chickpea. Frontiers in Bioscience-Landmark 2023, 28, 20. [Google Scholar] [CrossRef]
- Chittoor, J.M.; Leach, J.E.; White, F.F. Induction of peroxidase during defense against pathogens. Pathogenesis-related proteins in plants 1999, 171–193. [Google Scholar]
- Vanitha, S.C.; Niranjana, S.R.; Umesha, S. Role of phenylalanine ammonia lyase and polyphenol oxidase in host resistance to bacterial wilt of tomato. Journal of Phytopathology 2009, 157, 552–557. [Google Scholar] [CrossRef]
- Nikoo, F.S.; Sahebani, N.; Aminian, H.; Mokhtarnejad, L.; Ghaderi, R. Induction of systemic resistance and defense-related enzymes in tomato plants using Pseudomonas fluorescens CHAO and salicylic acid against root-knot nematode Meloidogyne javanica. Journal of Plant Protection Research 2014. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Labuschagne, N.; Dubery, I.A. Metabolomic evaluation of tissue-specific defense responses in tomato plants modulated by PGPR-priming against Phytophthora capsici infection. Plants 2021, 10, 1530. [Google Scholar] [CrossRef] [PubMed]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Labuschagne, N.; Dubery, I.A. Metabolic profiling of PGPR-treated tomato plants reveal priming-related adaptations of secondary metabolites and aromatic amino acids. Metabolites 2020, 10, 210. [Google Scholar] [CrossRef]
Figure 1.
In-vitro analysis of the antagonistic properties of Bacillus isolates. Clear zones of growth inhibition were developed by strains BS6 (A), BS7 (B) and BS9 (C). Contact growth inhibition as displayed by strain BS2 (B).
Figure 1.
In-vitro analysis of the antagonistic properties of Bacillus isolates. Clear zones of growth inhibition were developed by strains BS6 (A), BS7 (B) and BS9 (C). Contact growth inhibition as displayed by strain BS2 (B).
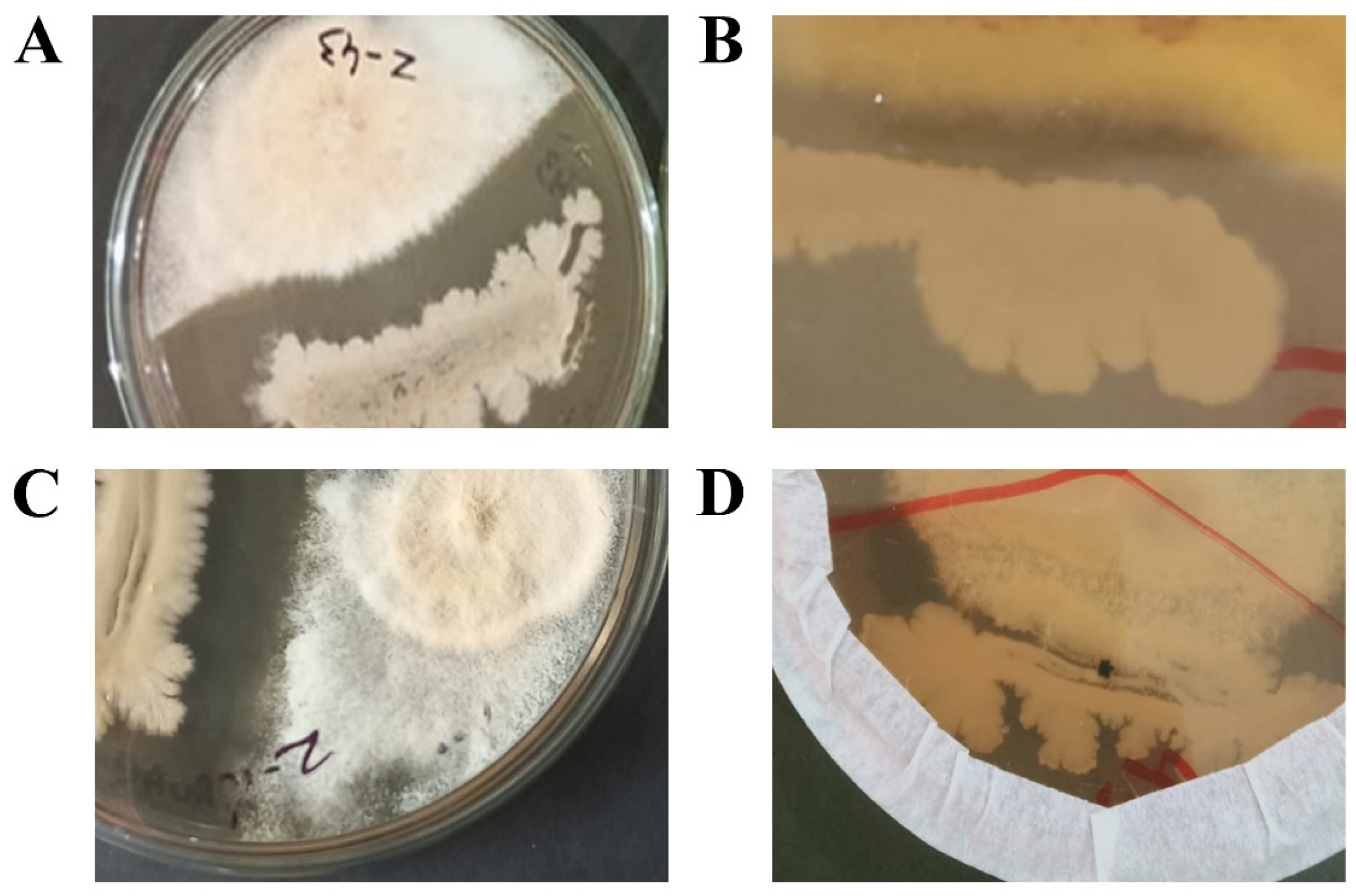
Figure 2.
Maximum likelihood tree of sequences of selected bacterial strains and retrieved from NCBI database.
Figure 2.
Maximum likelihood tree of sequences of selected bacterial strains and retrieved from NCBI database.
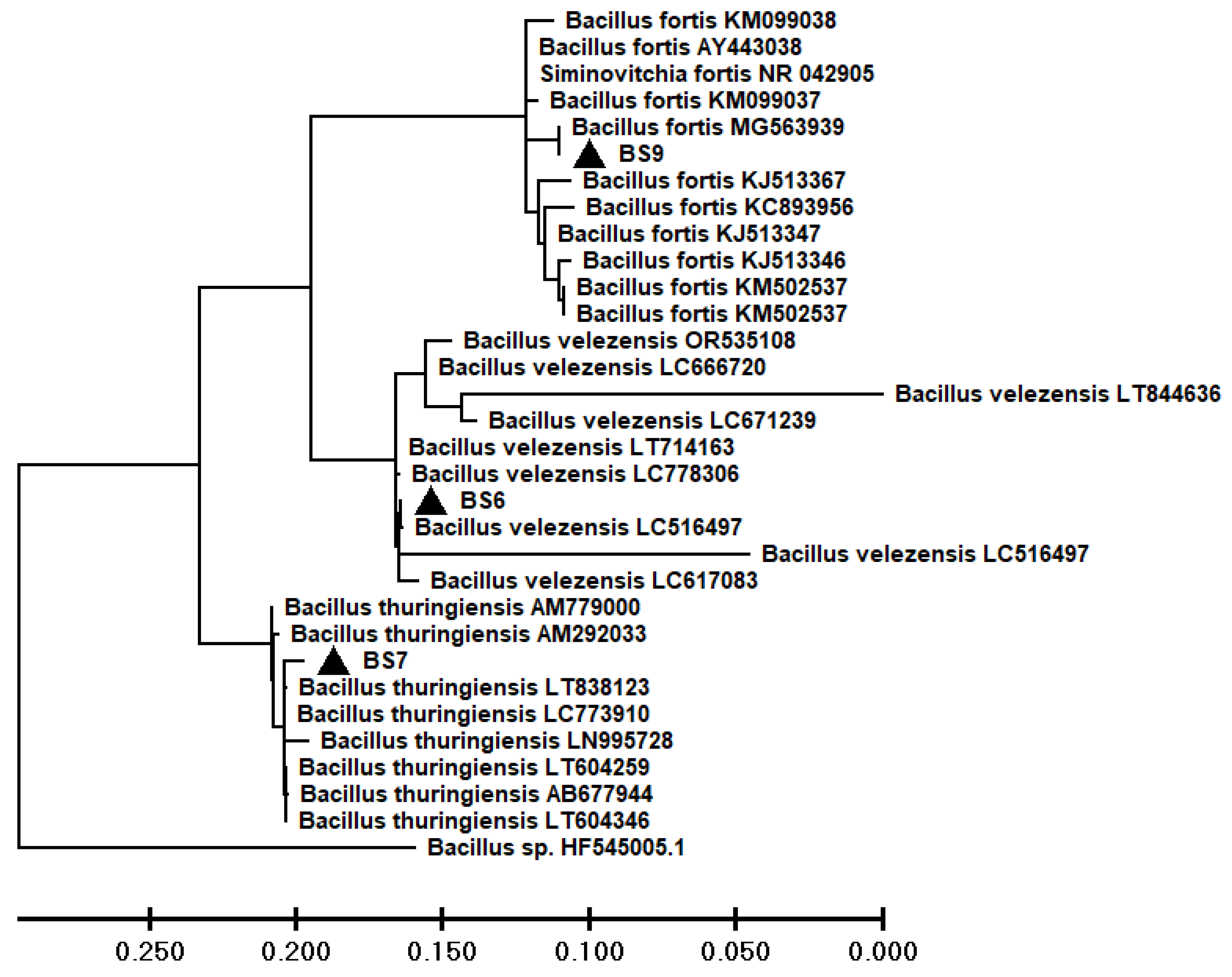
Figure 3.
The snapshots of quercetin ligand docked within the SILKY12 receptor (A) and their interactions (B).
Figure 3.
The snapshots of quercetin ligand docked within the SILKY12 receptor (A) and their interactions (B).
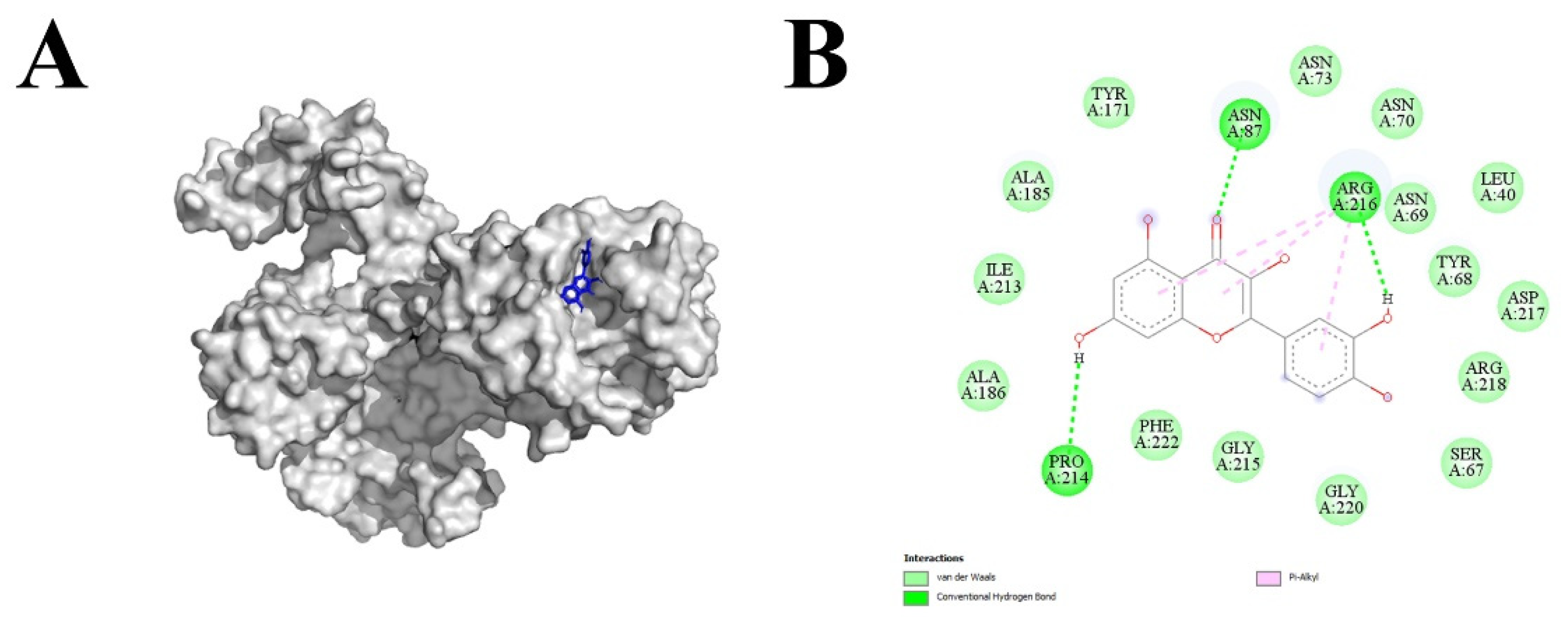
Figure 4.
The potential of bacterial consortia and synthetic elicitor to suppress Fusarium wilt disease. Vertical bars represent standard error. Capital letters present the level of significance by ANOVA and DNMRT at p= 0.05. details of treatments are provided in Table 1.
Figure 4.
The potential of bacterial consortia and synthetic elicitor to suppress Fusarium wilt disease. Vertical bars represent standard error. Capital letters present the level of significance by ANOVA and DNMRT at p= 0.05. details of treatments are provided in Table 1.
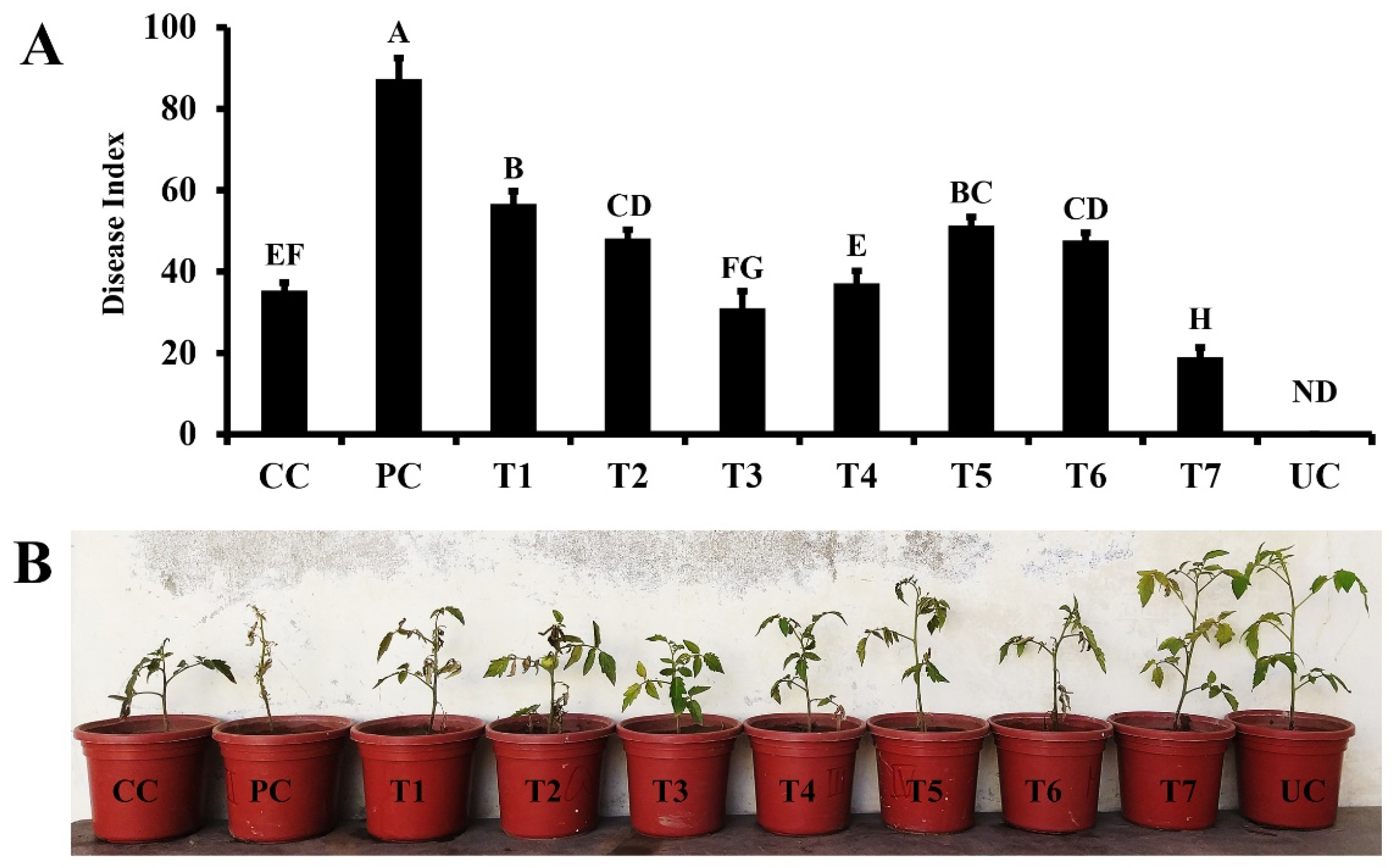
Figure 5.
Representative chromatograms of non-targeted metabolomic analysis of tomato plants under different treatments. A= Pathogen control, B= Quercetin (1.0 mM) + FOL, C= Consortia + FOL, and D= Consortia + Quercetin (1.0 mM) + FOL.
Figure 5.
Representative chromatograms of non-targeted metabolomic analysis of tomato plants under different treatments. A= Pathogen control, B= Quercetin (1.0 mM) + FOL, C= Consortia + FOL, and D= Consortia + Quercetin (1.0 mM) + FOL.
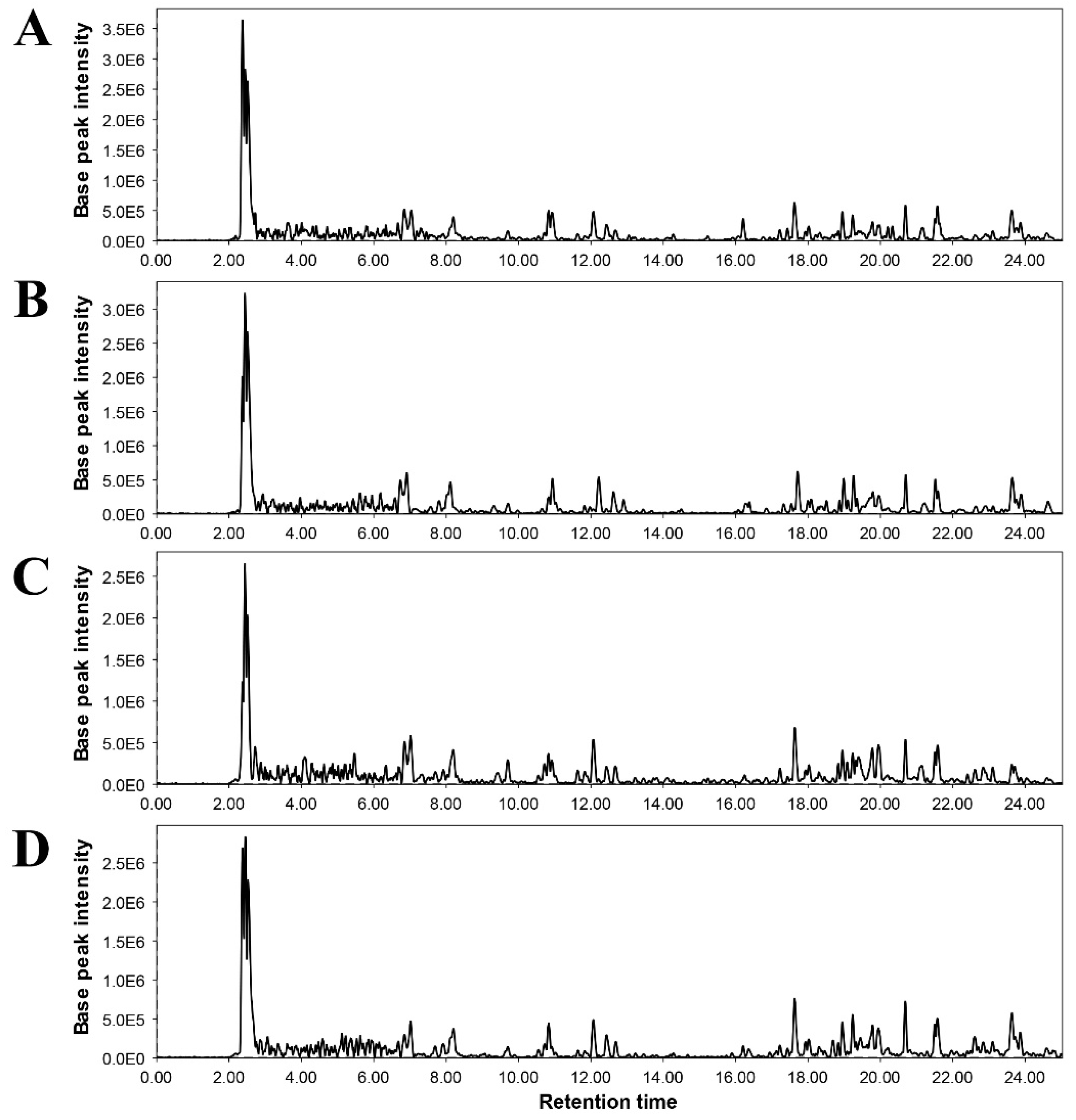
Figure 6.
Heat map showing changes in the levels of different metabolites. Data is presented as fold change as compared to the untreated control treatment.
Figure 6.
Heat map showing changes in the levels of different metabolites. Data is presented as fold change as compared to the untreated control treatment.
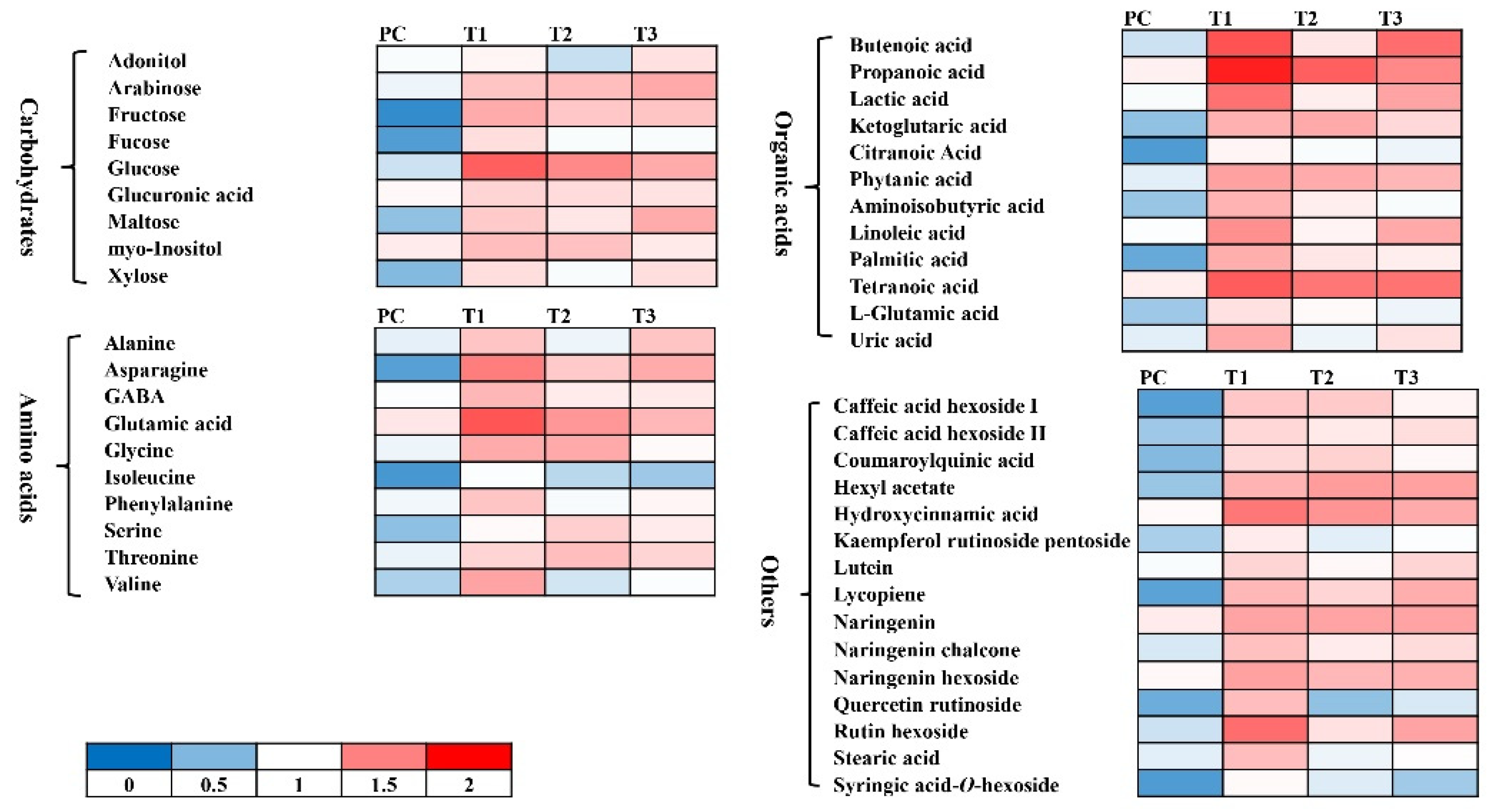
Figure 7.
Heat map showing changes in the levels of different phenolic acids. Data is presented as fold change as compared to the untreated control treatment. * Represent significant differences among treatments as governed by ANOVA at p> 0.05.
Figure 7.
Heat map showing changes in the levels of different phenolic acids. Data is presented as fold change as compared to the untreated control treatment. * Represent significant differences among treatments as governed by ANOVA at p> 0.05.
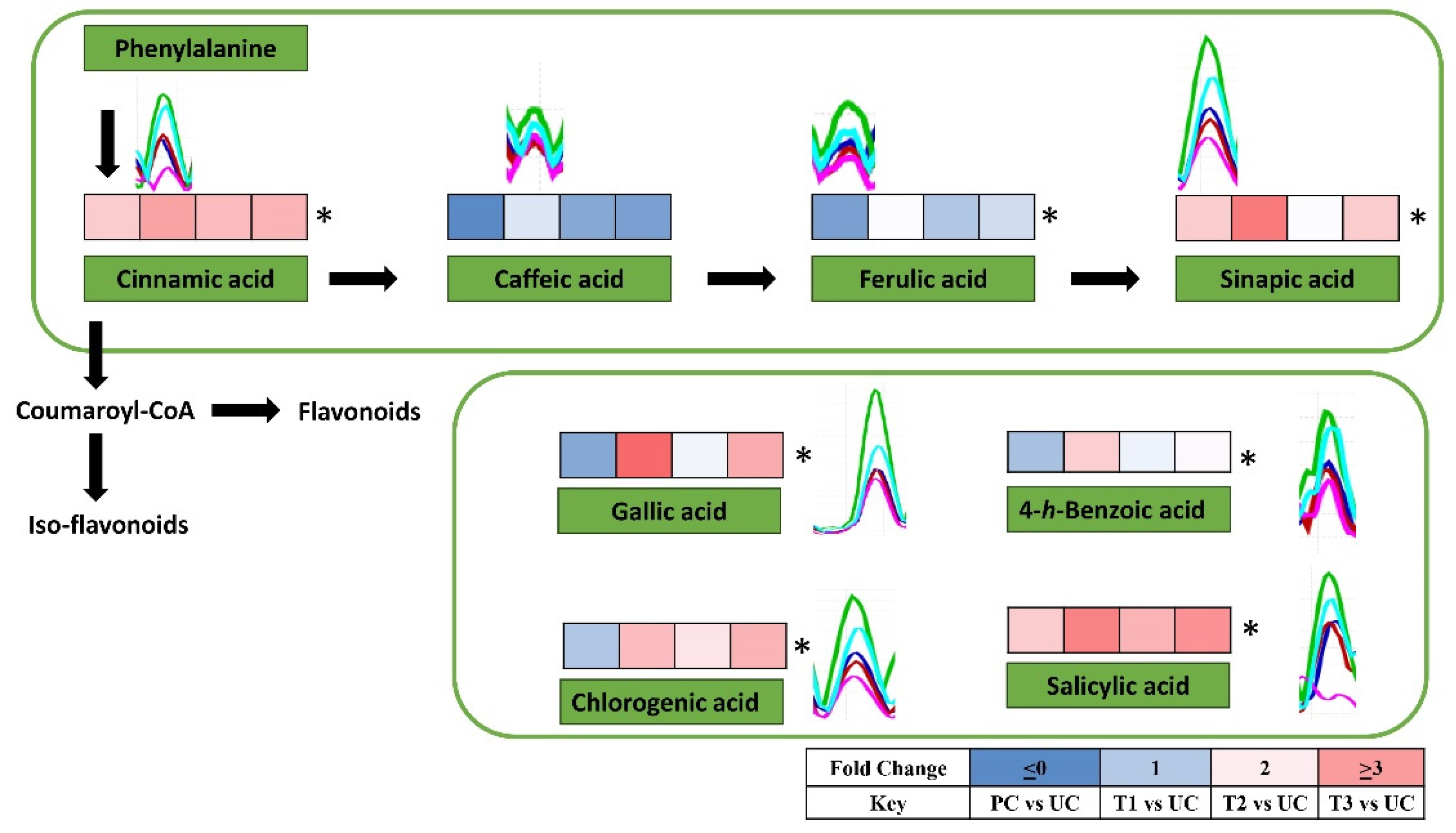

Table 1.
Details of treatments.
| Treatment | Description |
|---|---|
| CC | Carbendazim control |
| PC | Pathogen control (FOL) |
| T1 | Quercetin (1.0 mM) + FOL |
| T2 | Quercetin (0.1 mM) + FOL |
| T3 | Quercetin (0.01 mM) + FOL |
| T4 | Consortium + FOL |
| T5 | Quercetin (1.0 mM) + Consortium + FOL |
| T6 | Quercetin (0.1 mM) + Consortium + FOL |
| T7 | Quercetin (0.01 mM) + Consortium + FOL |
| UC | Untreated control |
Table 2.
Characterization of bacterial strains for antagonism properties.
| No | Code | Gram staining | Antagonistic phenotype |
|---|---|---|---|
| 1 | BS1 | Positive | NA |
| 2 | BS2 | Positive | CGI |
| 3 | BS3 | Positive | NA |
| 4 | BS4 | Positive | NA |
| 5 | BS5 | Positive | NA |
| 6 | BS6 | Positive | ZGI |
| 7 | BS7 | Positive | ZGI |
| 8 | BS8 | Positive | CGI |
| 9 | BS9 | Positive | ZGI |
| 10 | BS10 | Positive | NA |
| 11 | BS11 | Positive | NA |
NA No Activity, CGI= Contact growth inhibition, ZGI= Zone of growth inhibition.
Table 3.
Quality assessment of the predicted 3D models.
| Parameter | Value |
|---|---|
| PROCHECK | |
| Errors | 5 |
| Warning | 2 |
| Pass | 2 |
| ERRAT | 97.03 |
| Ramachandran plot | |
| FR | 91.8 |
| AR | 8.1 |
| GR | 0.2 |
| DR | 0.00 |
| Verify3D | 62.4 |
FR= most favored regions, AR= additional allowed regions, GR= generously allowed regions, DR= disallowed regions.
Table 4.
Estimate of the overall binding free energies of the top five legends.
| Compound | Formula | Molecular weight | Binding Affinity (kcal/mol) |
|---|---|---|---|
| Quercetin | C15H10O7 | 302.2 | -8.7 |
| Galbacin | C20H20O5 | 340.4 | -8.7 |
| Traumatin | C12H20O3 | 212.2 | -8.3 |
| 7-Oxotyphasterol | C28H48O5 | 464.7 | -8.2 |
Table 5.
Effect of the application of consortium of antagonistic bacterial strains and quercetin on growth parameters of tomato plants.
Table 5.
Effect of the application of consortium of antagonistic bacterial strains and quercetin on growth parameters of tomato plants.
| Treatment | Shoot length (cm) | Root length (cm) | Shoot biomass (g) | Root Biomass (g) | Total Chlorophyll (mg/g fw) |
|---|---|---|---|---|---|
| CC | 08.01±0.5e-g | 08.62±0.5ef | 1.82±0.07de | 0.31±0.02gh | 11.23±0.76e-g |
| PC | 09.28±0.8ef | 07.23±0.5e-g | 1.58±0.09ef | 0.24±0.02i | 07.17±0.04h |
| T1 | 11.12±0.9de | 09.26±0.5de | 2.06±0.08b-d | 0.43±0.03ef | 10.87±0.76fg |
| T2 | 10.38±1.4de | 10.71±0.7cd | 1.93±0.12cd | 0.51±0.04c-e | 15.91±1.27cd |
| T3 | 13.66±1.5cd | 11.96±0.7b-d | 2.24±0.13bc | 0.57±0.02cd | 16.08±1.27cd |
| T4 | 14.26±1.3b-d | 12.81±0.9bc | 2.59±0.14bc | 0.65±0.03c | 16.53±0.98c |
| T5 | 17.05±1.0bc | 12.54±1.1bc | 2.66±0.24ab | 0.59±0.04cd | 19.58±1.13a |
| T6 | 19.17±1.5ab | 15.97±1.4ab | 2.89±0.10a | 0.81±0.05ab | 18.32±1.64ab |
| T7 | 21.09±1.1a | 17.26±1.2a | 3.07±0.18a | 0.87±0.06a | 19.53±1.25a |
| Con | 23.18±1.6a | 15.62±1.1ab | 3.24±0.21a | 0.92±0.05a | 21.35±1.57a |
Values are mentioned as mean ± standard error. Capital letters present a level of significance as governed by ANOVA and DNMRT at p= 0.05. Details of treatments are provided in Table 1.
Table 6.
Effect of combined application of rhizobacteria and synthetic elicitors on defense-related activities of tomato plants against Fusarium wilt disease.
Table 6.
Effect of combined application of rhizobacteria and synthetic elicitors on defense-related activities of tomato plants against Fusarium wilt disease.
| Treatment | Total Phenolics (mg g-1FW) | PO (ΔOD min-1 g-1 FW) | PPO (ΔOD min-1 g-1 FW) | PAL (ΔOD min-1 g-1 FW) |
|---|---|---|---|---|
| Con | 2.03 ± 0.14e | 0.09±0.00e | 0.72±0.03c | 0.18 ± 0.01d |
| PC | 4.93 ± 0.51b-d | 0.68±0.04c | 1.12±0.25b | 0.49 ± 0.02bc |
| T1 | 7.18 ± 0.37a | 1.03±0.25a | 2.68±0.09a | 0.66 ± 0.05a |
| T2 | 5.39±0.20bc | 0.87±0.03b | 2.17±0.15a | 0.52±0.03ab |
| T3 | 6.28±0.37ab | 0.56±0.02cd | 1.32±0.07b | 0.63±0.04a |
Values are mentioned as mean ± standard error. Capital letters present a level of significance as governed by ANOVA and DNMRT at p= 0.05. Con= Untreated control, PC= Pathogen control, T1= Consortia + Quercetin (1.0 mM) + FOL, T2= Consortia + FOL, and T3= Quercetin (1.0 mM) + FOL.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



