
Submitted:
27 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
Avipoxvirus (APV), a linear dsDNA virus belonging to the subfamily Chordopoxvirinae of the family Poxviridae, infects more than 278 species of domestic and wild birds. It is responsible for causing the avian pox disease, which is characterized by its cutaneous and diphtheric forms. With a high transmission capacity, it can cause high economic losses and damage to the ecosys-tem. Several diagnostic methods are available and vaccination of birds can be an effective pre-ventive measure. To update the molecular characterization and phylogenetic analysis of viruses isolated in Portugal between 2017 and 2023, ten APV-positive samples were analyzed. A P4b gene fragment was amplified by PCR and the nucleotide sequence of the amplicons was deter-mined by Sanger sequencing. The sequences obtained were aligned using ClustalW, and a max-imum likelihood phylogenetic tree was constructed. With this study, it was possible to verify that the analyzed sequences are distributed in subclades A1, A2, B1, and B3. Since some of them are quite similar to others from different countries and obtained in different years, it is possible to conclude that there have been several viral introductions in Portugal. Finally, it was possible to successfully update the data on avipoxviruses in Portugal.
Keywords:
Avipoxvirus
; molecular characterization
; phylogenetic analysis
; viral introduction
; Portugal
1. Introduction
The family Poxviridae is divided into subfamilies Chordopoxvirinae and Entomopoxvirinae, responsible for infecting vertebrates and insects, respectively [1]. The genus Avipoxvirus is included within the subfamily Chordopoxvirinae, together with other seventeen genera [2]. Considering the different avian species that the virus infects, there are twelve species of avipoxviruses (APVs) [3].
According to Manarolla et al. [4], the nomenclature given to APVs is based on the type of species present in each clade, having already been identified five clades, A to E. Clade A is associated with fowlpox, clade B with canarypox, and clade C with psittacinepox viruses. To date, seven subclades within clade A and four subclades within clade B were described. Clade D includes a unique strain, QP-241, which was isolated in Italy from a quail [4]. Clade E contains sequences isolated from APVs from chickens in Brazil [5] and Mozambique [6], and also from a turkey in Hungary [7].
The avipoxviruses are very large viruses [8] that can be either oval or brick-shaped [9]. The dsDNA genome is in a linear configuration [9] and has a very low GC content [10]. Among all the poxviruses, avipoxviruses have the largest genome, with about 180 kbp [11].
APV infection can cause the development of avian pox disease, which can be presented by its cutaneous or diphtheric forms. These are caused by different routes of infection. The cutaneous form is originated by mechanical trauma [12], causes nodular lesions in feather-free areas and is associated with very low mortality rates [9]. The diphtheric form, on the other hand, occurs after inhalation or ingestion of the virus [12] and is characterized by the formation of proliferative nodular lesions in the mucous membranes of the digestive and respiratory tracts [9]. Since the nodular lesions can cause difficulty in breathing and eating, the mortality rate of this form of the disease is higher [13]. Transmission may occur through mechanical vectors like arthropods, aerosols released by sick birds, direct contact with lesions, or ingestion of contaminated food or drink [14]. Wild birds’ behavior, such as bird migration, introduction of new species, and habitat change, also lead to transmission [3]. However, the disease can be prevented by implementing sanitary measures to avoid mechanical vectors and contaminated sources [5] or through birds’ vaccination [15].
Infection leads to a variety of negative consequences, both economical and in the ecosystem. A decrease in egg production and immunity is commonly seen, resulting in a decreased ability to survive to secondary infections. Reduced mating success, retarded growth, blindness, feeding difficulties, and death can also occur [3,16]. Therefore, poultry farmers suffer significant economic losses due to the need to replace livestock, sales losses, and sanitation costs [5]. At the ecosystem level, the birds are more vulnerable, leading to increased predation, and a decrease in mating, thus resulting in a decline in population [17].
In this paper, molecular characterization and phylogenetic analysis of avipoxviruses isolated from ten samples detected in Portugal since 2017 are performed, in order to infer about the possible origin of the viruses circulating in the country.
2. Materials and Methods
2.1. Samples
Ten avipoxvirus positive samples were detected at INIAV since 2017. Those positive samples were preserved at -80°C.

Table 1.
Avipoxvirus positive samples diagnosed at INIAV since 2017 and used in this study.
| Host | Sequence ID | Accesion number | Country | Collection date | Isolation source | |
| Common name | Scientific name | |||||
| Flamingo | Phoenicopterus ruber | 24569-17 | OQ615872 | Portugal: Lisboa | 26/Sep/17 | Pool of organs |
| Chicken | Gallus gallus | 23049-18 | OQ615873 | Portugal: Porto Santo, Madeira | 24/Jul/18 | Pool of organs |
| Puffin | Fratercula | 11612-19 | OQ615874 | Portugal: Lisboa | 16/Apr/19 | Cutaneous lesion |
| Canary | Serinus canaria | 37026-19 | OQ615875 | Portugal: Freixianda, Ourém | 22/Nov/19 | Pool of organs |
| Canary | Serinus canaria | 03779-20 | OQ615876 | Portugal | 04/Feb/20 | Pool of organs |
| Chicken | Gallus gallus | 04482-20 | OQ615877 | Portugal | 11/Feb/20 | Cutaneous lesion |
| Blackbird | Turdus merula | 16735-20 | OQ615878 | Portugal | 09/Jun/20 | Cutaneous lesion |
| Chicken | Gallus gallus | P-08508-21 | OQ615879 | Portugal: Maia, Porto | 22/Sep/21 | Pool of organs |
| Penguin | Spheniscidae | P-09292-22 | OQ615880 | Portugal: Avintes, Porto | 17/Oct/22 | Pool of organs |
| Chicken | Gallus gallus | 00917-23 | OQ615881 | Portugal: Évora | 16/Jan/23 | Pool of organs |
2.2. Nucleic acids extraction
The viral DNA extraction was performed in a nucleic acid extraction workstation Kingfisher Flex (Thermo Fisher Scientific, Massachusetts, USA) using IndiMag Kit (Indical Bioscience, Germany), following the manufacturer’s instructions. After extraction, the DNA was stored at 4°C.
2.3. PCR amplification of P4b gene fragment
The amplification of the P4b gene fragment was performed by conventional PCR with primers described by Huw Lee and Hwa Lee [18], using NZYTaq II 2x Green Master Mix (0.2 U/μL) (NZYTech, Portugal). The reaction contained 0.5 μL of each primer (50 pmol/μL) (Table 2) and 5 μL of DNA, in a total of 25 μL. The PCR program was executed in an UNO II thermocycler thermoblock (Biometra, Germany) and consisted of an initial denaturation at 95°C for 2 min, followed by 50 cycles of denaturation at 95°C for 30 s, annealing at 60°C for 40 s, and extension at 72°C for 1 min. A final extension was performed at 72°C for 10 min. The PCR product was separated by a 1% agarose gel electrophoresis stained with GreenSafe Premium (NZYTech, Portugal). The expected fragments of 578 bp, corresponding to the P4b gene amplified fragment were excised and purified using NZYGelpure Kit (NZYTech, Portugal), according to the manufacturer’s instructions.
Table 2.
Sequences of P4b primers used in PCR reactions, described by Huw Lee and Hwa Lee [18].
Table 2.
Sequences of P4b primers used in PCR reactions, described by Huw Lee and Hwa Lee [18].
| Primer | Sequence | bp | Tm (°C) | % GC |
| Pox-VP1 | 5’ – CAGCAGGTGCTAAACAACAA – 3’ | 20 | 62 | 45 |
| Pox-VP2 | 5’ – CGGTAGCTTAACGCCGAATA – 3’ | 20 | 64 | 50 |
2.4. P4b gene fragment Sanger sequencing
The P4b gene fragments were sequenced by the Sanger method, using BigDye Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific, Massachusetts, USA), following the manufacturer’s guidelines. The primers used in the sequencing reaction are described in Table 2. In order to improve the sequencing quality, different volumes of sample were used in each reaction depending on the DNA concentration. The samples were sequenced in a 3130 Genetic Analyzer (Applied Biosystems, Massachusetts, USA). The sequence alignment was performed using SeqScape Software from Applied Biosystems. With the use of Clustal Omega by EMBL-EBI, percent identity matrices were also created between the sequences of subclades A1, A2, B1, and B3 (data not shown) and the sequences isolated and sequenced in this work. The GenBank accession numbers given to sequences obtained in this study are represented in Table 1.
2.5. Phylogenetic analysis
To study the phylogenetic origin of the avipoxviruses detected and their phylogenetic relationship with the sequences published in GenBank (Table 3), a multiple alignment between them was performed using ClustalW method. Sequences from different countries, years, and clades were chosen, in order to get a reliable analysis. To generate the phylogenetic trees, maximum likelihood, neighbor-joining, and bayesian methods were used. Each tree was produced using MEGA11 with the Tamura 3-parameter G+I, determined as the best fit model also by MEGA11, and 1000 bootstrap replicates.
Table 3.
Details of APVs sequences published in GenBank and used in this study.
| Host | Accesion number | Country | Collection date | Clade | |
| Common name | Scientific name | ||||
| Turkey | Meleagris gallopavo | AY530304 | Germany | 2001 | A1 |
| Silver pheasant | Lophura nycthemera | HM481406 | India | 2008 | A1 |
| Chicken | Gallus gallus | KF722860 | Tanzania | 2012 | A1 |
| Chicken | Gallus gallus | KM974727 | Portugal | 2013 | A1 |
| Chicken | Gallus gallus | KP987214 | Nigeria | 2013 | A1 |
| Backyard turkey | Meleagris gallopavo | KU522210 | Iran | 2015 | A1 |
| Quail | Coturnix coturnix | DQ873809 | India | - | A2 |
| Indian little brown dove | Spilopelia senegalensis | HM481408 | India | 2009 | A2 |
| Eurasian stone-curlew | Burhinus oedicnemus | HM627224 | Spain | 1980 | A2 |
| Rock dove | Columba livia | KC017966 | USA | 1995 | A2 |
| Indian peafowl | Pavo cristatus | KC017975 | Hungary | 2003 | A2 |
| Booted eagle | Hieraaetus pennatus | KC017976 | Spain | 2000 | A2 |
| Pigeon | Columbidae | KJ913659 | Tanzania | 2013 | A2 |
| Wood-pigeon | Columba palumbus | EU798994 | Czech Republic | 2008 | A3 |
| Pelagic cormorant | Phalacrocorax pelagicus | KC017982 | USA | 1989 | A3 |
| Eurasian eagle owl | Bubo bubo | KC017983 | South Korea | - | A3 |
| Common murre | Uria aalge | KC017985 | USA | 1991 | A3 |
| Laysan albatross | Phoebastria immutabilis | KC017986 | USA | 1983 | A3 |
| Magellanic penguin | Spheniscus magellanicus | KC017987 | Argentina | 2007 | A3 |
| Falcon | Falco sp. | AY530306 | United Arab Emirates | 2002 | A4 |
| Red-footed falcon | Falco vespertinus | KC017989 | Hungary | 2007 | A4 |
| Trumpeter swan | Cygnus buccinator | KC017990 | USA | 1991 | A5 |
| Mottled duck | Anas fulvigula | KC017991 | USA | 2005 | A5 |
| Redhead duck | Aythya americana | KC017993 | USA | 1991 | A5 |
| Trumpeter swan | Cygnus buccinator | KC017995 | USA | 1989 | A5 |
| Wood duck | Aix sponsa | KC017996 | USA | 1991 | A5 |
| Domestic mallard duck | Anas platyrhynchos | KJ192189 | China | 2013 | A5 |
| Mourning dove | Zenaida macroura | KC018000 | USA | 1987 | A6 |
| Canada goose | Branta canadensis | KC018002 | USA | 1992 | A6 |
| Common buzzard | Buteo buteo | KC018009 | Hungary | 2000 | A7 |
| Stone curlew | Burhinidae | AY530310 | United Arab Emirates | 1998 | B1 |
| Palila | Loxioides bailleui | EF568381 | USA | - | B1 |
| Amakihi | Hemignathus virens | EF568401 | USA | - | B1 |
| Blue jay | Cyanocitta cristata | GQ487567 | Canada | 1998 | B1 |
| Canary | Serinus canaria | GU108510 | Austria | 2009 | B1 |
| Red crossbill | Loxia curvirostra | HM627227 | Spain | 1930 | B1 |
| Golden eagle | Aquila chrysaetos | KC018058 | Spain | 2000 | B1 |
| House sparrow | Passer domesticus | HM627220 | Marocco | 2009 | B2 |
| Flamingo | Phoenicopterus ruber | HQ875129 | Portugal | 2010 | B2 |
| Great bustard | Otis tarda | KC018066 | Hungary | 2005 | B2 |
| American crow | Corvus brachyrhynchos | DQ131891 | USA | 2003 | B3 |
| House finch | Haemorhous mexicanus | DQ131896 | USA | 2003 | B3 |
| Great blue heron | Ardea herodias | DQ131898 | USA | 2004 | B3 |
| Northern cardinal | Cardinalis cardinalis | DQ131899 | USA | 2003 | B3 |
| Red-tailed hawk | Buteo jamaicensis | DQ131901 | USA | 2003 | B3 |
| Chicken | Gallus gallus | AM050382 | United Kingdom | 1986 | C |
| Parrot | Psittaciformes | AM050383 | United Kingdom | 1989 | C |
| Lovebird | Agapornis | AY530311 | Germany | - | C |
| Yellow-crowned amazon | Amazona ochrocephala | KC018069 | USA | 1980 | C |
| Quail | Coturnix coturnix | GQ180200 | Italy | - | D |
| Chicken | Gallus gallus | MW349699 | Brazil | 2019 | E |
| Chicken | Gallus gallus | MW349701 | Brazil | 2019 | E |
| Domestic mallard duck | Anas platyrhynchos | KJ192189 | China | 2013 | A5 |
| Mourning dove | Zenaida macroura | KC018000 | USA | 1987 | A6 |
3. Results and Discussion
3.1. PCR amplification of P4b gene fragment
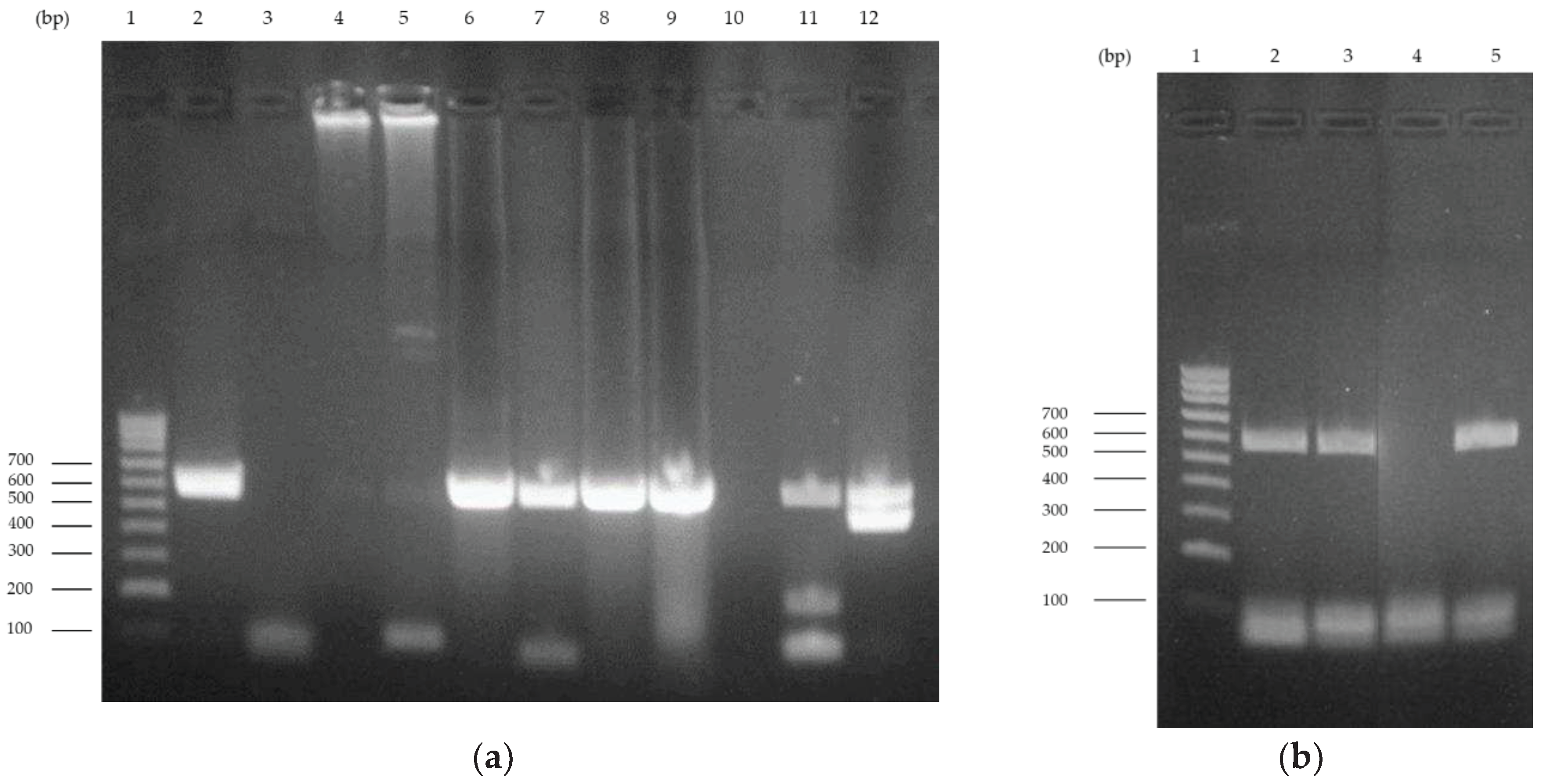
The amplification of the 578 bp P4b gene fragment was confirmed by 1% agarose gel electrophoresis. The results obtained are shown in Figure 1. It is possible to observe the presence of the expected fragment in all lanes except lanes 3, 4, and 5, which correspond to samples 11612-19 (puffin), 37026-19 (canary), and 03779-20 (canary), respectively. Amplification of DNA from samples 37026-19 and 03779-20 obtained in a previous extraction process was achieved (lanes 11 and 12) (Figure 1a). A new PCR reaction led to the amplification of the sample 11612-19 (Figure 1b).
Figure 1.
1% agarose gel electrophoresis of the 578 bp P4b gene fragment PCR amplification. (a) Lane 1 corresponds to the molecular weight marker, NZYDNA Ladder V (NZYTech, Portugal). Lanes 2 to 9 correspond to the amplification of samples 24569-17 (flamingo), 11612-19 (puffin), 37026-19 (canary), 03779-20 (canary), 04482-20 (chicken), 16735-20 (blackbird), P-08508-21 (chicken), and 23049-18 (chicken), respectively. Lane 10 is the negative control. Lanes 11 and 12 correspond to samples 37026-19 (canary) and 03779-20 (canary) obtained in a previous extraction. (b) Lane 1 corresponds to the molecular weight marker, NZYDNA Ladder V. Lanes 2 and 3 correspond to sample 11612-19 (puffin) from different extraction reactions. Lanes 4 and 5 are the negative and positive controls, respectively.
Figure 1.
1% agarose gel electrophoresis of the 578 bp P4b gene fragment PCR amplification. (a) Lane 1 corresponds to the molecular weight marker, NZYDNA Ladder V (NZYTech, Portugal). Lanes 2 to 9 correspond to the amplification of samples 24569-17 (flamingo), 11612-19 (puffin), 37026-19 (canary), 03779-20 (canary), 04482-20 (chicken), 16735-20 (blackbird), P-08508-21 (chicken), and 23049-18 (chicken), respectively. Lane 10 is the negative control. Lanes 11 and 12 correspond to samples 37026-19 (canary) and 03779-20 (canary) obtained in a previous extraction. (b) Lane 1 corresponds to the molecular weight marker, NZYDNA Ladder V. Lanes 2 and 3 correspond to sample 11612-19 (puffin) from different extraction reactions. Lanes 4 and 5 are the negative and positive controls, respectively.
3.2. P4b gene fragment sequencing
The alignment of forward and reverse nucleotide sequences obtained, originated a sequence of 538 bp for all samples, except for sample 03779-20 (canary), after removing the primers sequences. Due to incomplete sequencing of the 3' end, only 513 bp were obtained for this sample. This result can be explained by DNA damage, incorrect amplification, or inadequate purification.
The representation of the alignment between the studied sequences is present in Figure 2, and its analysis is done in the following section, together with the phylogenetic analysis.
Figure 2.
Alignment of the studied sequences.

3.3. Phylogenetic analysis
The avipoxvirus nucleotide sequences obtained were phylogenetically analyzed using different algorithms, specifically maximum likelihood, neighbor-joining, and bayesian analysis. The three algorithms originated phylogenetic trees with similar topologies. However, the maximum likelihood tree originated a better resolution of the sequences, and therefore this was the chosen method (Figure 3). The phylogenetic tree obtained, including representatives from all subclades (except B4), shows that clades C and D have a common most recent ancestor, as observed in previous studies [5,14].
The phylogenetic analysis reveals that the samples from Portugal are quite distributed, with five samples belonging to subclade A1, one to subclade A2, three to subclade B1, and one to subclade B3. Moreover, their distribution is quite consistent with the nomenclature given, since samples from Gallus gallus belong to the fowlpox clade (clade A), while samples from Turdus merula and Serinus canaria, both passeriformes, belong to the canarypox clade (clade B). Phoenicopterus ruber, Fratercula, and Spheniscidae are not fowl or Passeriformes representatives. The viruses detected in the first two hosts were included in the fowlpox clade, while the penguinpox virus belongs to the canarypox clade.
When comparing the alignment (Figure 2) and the percent identity matrix of the sequences obtained in this study (Table 4), several conclusions can be reached. Sample 16735-20 (blackbird) is the most distinct of all, as it has several different nucleotides along the entire sequence. Therefore, it has a very low identity percentage relative to all the other sequences, with the highest identity percentage value being only 81.60%. The nucleotide sequence of sample 24569-17 (flamingo) is the second most distinct sequence, showing a maximum identity percentage of 90.52%. In addition, it contains an extra valine codon at position 23.
By comparing the alignment of all the sequences present in the phylogenetic tree, it is observable that sample 16735-20 (blackbird) has two unique mutations. It contains a cytosine in position nt 63 instead of a thymine, and a thymine at position nt 148, instead of a guanine or an adenine.
Interestingly, when analyzing the percentage identity matrix between the sequences of the A1 subclade, it can be observed that the puffin (Fratercula) isolate sequence is not completely identical with any other sequence, but the identity percentages are quite high, showing that the sequences are very similar.
On the other hand, all the chicken (Gallus gallus) isolates have an identity percentage of 100%, except sample sequence 04482-20. Yet the identity percentage is still very high, 99.63%, with only two nucleotides differing. It can also be observed that the sequences of the samples isolated from turkey (Meleagris, Germany, 2001) and wild turkey (Meleagris gallopavo, Iran, 2015) are also similar to those isolated from chickens. By analyzing the position of the remaining sequences in the phylogenetic tree, it is possible to verify the presence of samples isolated from chickens also in subclades C and E. When comparing those of subclade A1 with these, it is found that the percentage of identity is quite low, approximately 74%, confirming the classification in different clades.
Regarding the sequences of A2 subclade, it is possible to observe that the sequence of the sample isolated from the flamingo (Phoenicopterus ruber) is different from all the other sequences. However, the percentages are quite high, varying between 92.15% (pigeon) (Columbia livia, USA, 1995) and 99.63% (pigeon) (Columbidae, Tanzania, 2013). Additionally, it can be seen that both APVs isolated from flamingos in Portugal belong to different clades, the one from 2010 belongs to B2 and the one from 2017 to A2. Indeed, the identity percentage between them is only 75.84%. When they are aligned, it can be noticed that the 2010 sequence has an extra isoleucine codon at position nt 24, while the 2017 has an extra phenylalanine codon at location nt 60.
The analysis of the sequences of subclade B1 shows that both isolates of canaries (Serinus canaria) from Portugal share the same nucleotide sequence. The canary’s isolate from Austria is not 100% identical with these canaries’ sequences, however it is very similar, having an identity percentage of 99.81%, with only one nucleotide differing. The sequence from the penguin (Spheniscidae) isolate is 100% identical with the sequences from three other isolates, the golden eagle isolate (Aquila chrysaetos, Spain, 2000), the stone-curlew isolate (Burhinidae, United Arab Emirates, 1998) and the crossbill isolate (Loxia curvirostra, Spain, 1930). Comparing the sequence of the avipoxvirus isolated from the penguin in Portugal with the one from Argentina, it is found that they are classified in distinct subclades, B1 and A3, respectively. These two sequences show several mutations between them, showing a percentage of identity of only 72.68%.
Comparing the sequences of the B3 subclade, it is possible to verify that the blackbird (Turdus merula) isolate sequence is not completely identical with any other sequence. However, the percentage of identity is quite high among all of the sequences (97.03% to 97.58%).
The results of this study indicate that the APVs circulating in Portugal in chickens and canaries are quite similar, since the nucleotide sequences of the isolates did not undergo considerable changes. However, the isolates from flamingos show the opposite situation, since the virus that circulated in 2010 is very different from that circulating in 2017. This suggests the occurrence of a second viral introduction in Portugal. Finally, as the nucleotide sequence of the isolate from penguin is very similar to that isolated from golden eagle, stone-curlew, and crossbill, it is possible to hypothesize that there was a third viral introduction in Portugal.
Figure 3.
Evolutionary analysis by Maximum Likelihood method. The evolutionary history was inferred by using the Maximum Likelihood method and Tamura 3-parameter model [19]. The tree with the highest log likelihood (-3505.45) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. A discrete Gamma distribution was used to model evolutionary rate differences among sites (5 categories (+G, parameter = 1.4400)). The rate variation model allowed for some sites to be evolutionarily invariable ([+I], 36.73% sites). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. This analysis involved 62 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. There were a total of 485 positions in the final dataset. Evolutionary analyses were conducted in MEGA X [20].
Figure 3.
Evolutionary analysis by Maximum Likelihood method. The evolutionary history was inferred by using the Maximum Likelihood method and Tamura 3-parameter model [19]. The tree with the highest log likelihood (-3505.45) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. A discrete Gamma distribution was used to model evolutionary rate differences among sites (5 categories (+G, parameter = 1.4400)). The rate variation model allowed for some sites to be evolutionarily invariable ([+I], 36.73% sites). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. This analysis involved 62 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. There were a total of 485 positions in the final dataset. Evolutionary analyses were conducted in MEGA X [20].

Table 4.
Percent identity matrix between APVs sequences isolated in this study.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| 1 | Phoenicopterus ruber (24569-17) | 90.52 | 89.78 | 89.78 | 89.78 | 90.15 | 75.46 | 72.68 | 73.05 | 72.71 | |
| 2 |
Fratercula (11612-19) |
90.52 | 99.26 | 99.26 | 99.26 | 98.88 | 76.39 | 74.54 | 74.91 | 74.66 | |
| 3 |
Gallus gallus (23049-18) |
89.78 | 99.26 | 100.00 | 100.00 | 99.63 | 76.21 | 74.35 | 74.72 | 74.46 | |
| 4 |
Gallus gallus (P-08508-21) |
89.78 | 99.26 | 100.00 | 100.00 | 99.63 | 76.21 | 74.35 | 74.72 | 74.46 | |
| 5 |
Gallus gallus (00917-23) |
89.78 | 99.26 | 100.00 | 100.00 | 99.63 | 76.21 | 74.35 | 74.72 | 74.46 | |
| 6 |
Gallus gallus (04482-20) |
90.15 | 98.88 | 99.63 | 99.63 | 99.63 | 76.39 | 74.72 | 75.09 | 74.85 | |
| 7 |
Turdus merula (16735-20) |
75.46 | 76.39 | 76.21 | 76.21 | 76.21 | 76.39 | 81.60 | 81.60 | 81.48 | |
| 8 |
Spheniscidae (P-09292-22) |
72.68 | 74.54 | 74.35 | 74.35 | 74.35 | 74.72 | 81.60 | 99.63 | 99.81 | |
| 9 |
Serinus canaria (37026-19) |
73.05 | 74.91 | 74.72 | 74.72 | 74.72 | 75.09 | 81.60 | 99.63 | 100.00 | |
| 10 |
Serinus canaria (03779-20) |
72.71 | 74.66 | 74.46 | 74.46 | 74.46 | 74.85 | 81.48 | 99.81 | 100.00 |
4. Conclusions
In conclusion, it was possible to verify that the samples studied are widely distributed in the phylogenetic tree, with representatives in subclades A1, A2, B1, and B3. In addition, it was possible to observe that some of the isolates, such as those from chickens and canaries, suggest that the APV in circulation in Portugal originated from the same virus. Other isolates, such as flamingo and penguin isolates, suggest new viral introductions, since they are similar to other strains from different years and geographic origins. These data suggest that the viruses circulating in Portugal have distinct origins. Interestingly, the sequence from the blackbird isolate presents two unique mutations, when compared with all the sequences used in the phylogenetic analysis. With this study, it was possible to update the molecular and taxonomic data of avipoxviruses in Portugal.
Acknowledgments
The authors thank Maria João Teixeira, Fátima Cordeiro, Rosário Ferreira and Sandra Nunes for technical assistance.
References
- Murphy, F.A.; Gibbs, E.P.J.; Horzinek, M.C.; Studdert, M.J. Veterinary Virology; 3rd ed.; Academic Press: San Diego, 1999; ISBN 978-0-12-511340-3.
- Brennan, G.; Stoian, A.M.M.; Yu, H.; Rahman, M.J.; Banerjee, S.; Stroup, J.N.; Park, C.; Tazi, L.; Rothenburg, S. Molecular Mechanisms of Poxvirus Evolution. mBio 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.; Athukorala, A.; Raidal, S.R. Molecular Characterisation of a Novel Pathogenic Avipoxvirus from an Australian Passerine Bird, Mudlark (Grallina Cyanoleuca). Virology 2021, 554, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Manarolla, G.; Pisoni, G.; Sironi, G.; Rampin, T. Molecular Biological Characterization of Avian Poxvirus Strains Isolated from Different Avian Species. Vet. Microbiol. 2010, 140, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chacón, R.D.; Astolfi-Ferreira, C.S.; Pereira, P.C.; Assayag, M.S.; Campos-Salazar, A.B.; De la Torre, D.; de Sá, L.R.M.; de Almeida, S.R.Y.; Rici, R.E.G.; Piantino Ferreira, A.J. Outbreaks of Avipoxvirus Clade E in Vaccinated Broiler Breeders with Exacerbated Beak Injuries and Sex Differences in Severity. Viruses 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Mapaco, L.P.; Lacerda, Z.; Monjane, I.V.A.; Gelaye, E.; Sussuro, A.H.; Viljoen, G.J.; Dundon, W.G.; Achá, S.J. Identification of Clade E Avipoxvirus, Mozambique, 2016. Emerg. Infect. Dis. 2017, 23, 1602–1604. [Google Scholar] [CrossRef] [PubMed]
- Bányai, K.; Palya, V.; Dénes, B.; Glávits, R.; Ivanics, É.; Horváth, B.; Farkas, S.L.; Marton, S.; Bálint, Á.; Gyuranecz, M.; et al. Unique Genomic Organization of a Novel Avipoxvirus Detected in Turkey (Meleagris Gallopavo). Infect. Genet. Evol. 2015, 35, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Recent Advances in Animal Virology; Malik, Y.S., Singh, R.K., Yadav, M.P., Eds.; Springer Singapore: Singapore, 2019; ISBN 9789811390722.
- Bolte, A.L.; Meurer, J.; Kaleta, E.F. Avian Host Spectrum of Avipoxviruses. Avian Pathol. 1999, 28, 415–432. [Google Scholar] [CrossRef] [PubMed]
- Henriques, A.M.; Fagulha, T.; Duarte, M.; Ramos, F.; Barros, S.C.; Luís, T.; Bernardino, R.; Fernandes, T.L.; Lapão, N.; Da Silva, J.F.; et al. Avian Poxvirus Infection in a Flamingo (Phoenicopterus Ruber) of the Lisbon Zoo. J. Zoo Wildl. Med. 2016, 47, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Wittek, R. Organization and Expression of the Poxvirus Genome. Experientia 1982, 38, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Jarmin, S.; Manvell, R.; Gough, R.E.; Laidlaw, S.M.; Skinner, M.A. Avipoxvirus Phylogenetics: Identification of a PCR Length Polymorphism That Discriminates between the Two Major Clades. J. Gen. Virol. 2006, 87, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Kirubaharan, J.J.; Rajasekaran, R.; Shilpa, P.; Vidhya, M.; Rajalakshmi, S. Isolation, Molecular Detection and Phylogenetic Analysis of Avipox Virus Obtained from Pigeon. Indian J. Vet. Sci. Biotechnol. 2018, 14. [Google Scholar] [CrossRef]
- Van Der Meer, C.S.; Paulino, P.G.; Jardim, T.H.A.; Senne, N.A.; Araujo, T.R.; Dos Santos Juliano, D.; Massard, C.L.; Peixoto, M.P.; Da Costa Angelo, I.; Santos, H.A. Detection and Molecular Characterization of Avipoxvirus in Culex Spp. (Culicidae) Captured in Domestic Areas in Rio de Janeiro, Brazil. Sci. Rep. 2022, 12, 13496. [Google Scholar] [CrossRef] [PubMed]
- Boyle, D.B. Genus Avipoxvirus. In Poxviruses; Mercer, A.A., Schmidt, A., Weber, O., Eds.; Birkhäuser Basel: Basel, 2007; pp. 217–251. ISBN 978-3-7643-7556-0. [Google Scholar]
- Ghalyanchilangeroudi, A.; Hosseini, H.; Morshed, R. Molecular Characterization and Phylogenetic Analysis of Avian Pox Virus Isolated from Pet Birds and Commercial Flocks, in Iran. Slov. Vet. Res. 2018, 55. [Google Scholar] [CrossRef]
- Loc, G.L.; Bertagnoli, S.; Ducatez, M.; Camus-Bouclainville, C.; Guérin, J.-L. Diversity of Avipoxviruses in Captive-Bred Houbara Bustard. Vet. Res. 2014, 45. [Google Scholar] [CrossRef]
- Lee, L.H.; Lee, K.H. Application of the Polymerase Chain Reaction for the Diagnosis of Fowl Poxvirus Infection. J. Virol. Methods 1997, 63, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the Number of Nucleotide Substitutions When There Are Strong Transition-Transversion and G+C-Content Biases. Mol. Biol. Evol. 1992. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



