
Submitted:
29 September 2023
Posted:
30 September 2023
You are already at the latest version
Abstract
The subtropical forest soil plays a pivotal role in terrestrial carbon (C) cycling. Although global warming is expected to influence subtropical soil C cycling, a consensus on its impact on soil C dynamics remains elusive. Given the significant response of soil microbial respiration to temperature in subtropical soils, understanding its behavior under varying temperature regimes is critical for predicting soil organic C (SOC) responses to climate warming. We conducted a short-term laboratory soil warming incubation experiment, sampling both warmed and un-warmed soils from a subtropical plantation in southeastern China, incubating them at 20 °C, 30 °C, and 40 °C. Our aim was to study the SOC mineralization response to increasing temperatures. Our findings revealed that the temperature sensitivity (Q10) of SOC mineralization to short-term experimental warming varied between the warmed soil and the un-warmed soil. The Q10 of the un-warmed soil escalated with the temperature treatment (20-30 ℃: 1.31, 30-40℃: 1.63). Conversely, the Q10 of the warmed soil decreased (20-30 ℃:1.57, 30-40 ℃:1.41). Increasing temperature treatments enhanced hydrolytic enzyme activity but decreased soil substrate availability in both un-warmed and warmed soil. The Q10 of un-warmed soil was positively correlated with the response ratio of DOC. Additionally, soil microbial biomass exhibited a significant decline, leading to a reduced total amount of PLFAs and a decreased abundance of bacteria, fungi, and Gram-negative bacteria (GN). The changes in temperature also considerably altered the composition of the warmed soil microbial community. A drop in the microbial quotient coupled with a rise in the metabolic quotient indicated that warming amplified microbial respiration over microbial growth. The differential Q10 of SOC mineralization, in response to temperature across varying soil, can largely be attributed to shifts in soil nutrients, C-degrading enzyme activities, and microbial communities (the ratio of fungal to bacterial PLFAs).
Keywords:
Warming
; Soil organic carbon mineralization
; Temperature sensitivity
; Substrate availability
; Enzyme activity
; Microbial community structure
1. Introduction
Soil plays a central regulatory role in the global carbon (C) cycle [1]. Being the largest terrestrial C pool, soil organic carbon (SOC) contains three times as much C as that stored in the atmosphere and plants [2]. Given this vast pool size, even a slight change in SOC dynamic could significantly affect the global C cycle. While warming experiments in temperate and boreal regions have enriched our understanding of soil C cycling in response to climate warming [3,4], our knowledge regarding the SOC response in tropical and subtropical (sub/tropical) forests remains limited [5,6]. It’s noteworthy that sub/tropical forest ecosystems store 46 % and 11 % of the world’s living terrestrial C and soil C, respectively [7]. This lack of studies in the region underscores a knowledge gap in our understanding of the responses of sub/tropical forests to climate warming.
The temperature sensitivity (Q10) is a key metric that gauges the response of SOC decomposition to temperature change. It’s been observed that Q10 profoundly influenced by biotic factors like soil microbial biomass and community composition, and abiotic ones such as incubation temperature and nutrient availability [8,9,10]. However, variations in incubation conditions, like different temperatures and durations among experiments, have led to inconsistent predictions about the Q10 of SOC mineralization [11]. Q10 also mirrors the differences in thermal acclimation of microbial communities between long- and short-term responses to temperature [12]. Such community thermal acclimation can result from direct shifts in the extracellular enzyme pool towards distinct activation energies and temperature optima or indirect alterations in resources composition and availability [13,14,15]. Furthermore, since Q10 is temperature-dependent, the estimation of Q10 based on non-site-specific incubation temperatures might not accurately capture in-situ temperature sensitivity [11]. Therefore, analyzing samples from the same site under different warming treatments can provide more accurate insights into the differences in Q10 of SOC decomposition due to climate warming, and can also help determine if soils act as sources or sinks.
The quality and quantity of substrates influence microbial use of organic substances and the Q10 values of SOC decomposition [16,17]. For instance, as the temperature rises, the increase of dissolved organic carbon content and soil bacterial abundance can enhance SOC mineralization [18]. Regardless of warming, a richer soil C boosts the extracellular enzymatic pool and its temperature sensitivity [12]. Contrarily, Jiang et al. [19] found that Q10 had a positively correlation with the changes in soil dissolved organic carbon, soil ammonium nitrogen contents, and β-glucosidase activities, but a negatively one with fungi: bacteria and urease activities post a 5℃ of warming. Clearly, soil substrate quality and quantity play pivotal roles in SOC decomposition by microbes [20]. Temperature-induced changes in nutrient demand and supply can impact the available C to microbes [21,22]. Additionally, microbial biomass and community composition have a pronounced effect Q10 values [23,24]. For example, temperature can accelerate SOC decomposition by altering soil enzymes activity and SOC turnover rate [25]. On the other hand, soil microorganisms can adapt to sustained temperature increments by modifying their community composition [26]. However, not all substrates are created equal in fulfilling microbial demands. Factors such as enzyme investment, activation energy for decomposition, and energetic richness of compounds determine their usability by microbes [27,28,29]. Moreover, the various components of soil organic matter (SOM) differ in their decomposition temperature sensitivity, leading to significant shifts in SOM chemistry with chronic warming [21,30]. This means that long-term warming-induced changes in SOM chemistry could alter the potential resource pool available for organisms when they encounter short-term temperature fluctuations [31]. Given this complex interplay, there’s a pressing need for further research on the effects of soil microbial communities and their relationship with soil substrate on SOC mineralization.
This study was conducted on a field soil warming experiment in a subtropical plantation in southeast China. Previous studies on this experiment revealed that warming can reshape microbial community structure and enzyme activity, leading to a significant imbalance between soil N and C decomposition, and suggesting that heterotrophic respiration could be more sensitive to climate warming [32,33]. Therefore, we designed a laboratory-based soil incubation experiment that investigates the changes in Q10 of SOC mineralization between un-warmed and warmed soils across three temperature regimes (20, 30, and 40℃). This investigation is rooted in a field soil warming experiment conducted in a subtropical plantation in southeastern China. In this study, we hypothesis that the difference in Q10 of SOC mineralization in warmed and un-warmed soils to warming hinges on alterations in microbial and enzyme activities.
2. Materials and Methods
2.1. Study Site and Soil Sampling
The experiment was located at the Chenda research site (300 m above sea level) of Sanming Forest Ecosystem and Global Change National Observation and Research Station, Fujian Province (26°19N, 117°36E), southeastern China. This region is characterized by a subtropical monsoonal climate with a mean annual precipitation of 1670 mm (from 1959-2015), with 77% occurring from March to August, and a mean annual temperature of 19.1 °C [33].
In October 2013, ten experimental plots were established, following a randomized block design. Each block consisted of a warmed and un-warmed plot, with each plot measuring 2 m × 2 m. Heating cables (TXLP/1, Nexans, Norway) were buried in both un-warmed and warmed plots at a depth of 10 cm with a horizontal interval of 20 cm. However, the cables in the un-warmed plots remained unheated. In the warming plots, soil temperature was continuously maintained at 5 °C above that in the un-warmed plots. The warming experiment was carried out in a young Chinese fir plantation established in 2013. In June 2016, nine 0-10 cm depth soil cores were collected each of 5 plots per treatment using a 3.5 cm diameter soil corer. These samples were immediately transported to the laboratory, stored at 4 °C before analyses and subsequently incubated. For determining basic soil properties (Table 1), fresh soil was sieved through a 2 mm mesh and soil moisture was measured by drying the soil (2.0 g) at 105 °C for 24 h.
2.2. Laboratory Incubation Experiment
The incubation experiment encompassed three temperature levels (20, 30, and 40 °C) and two types soil (warmed soil and un-warmed soil). In total, 18 soil samples were placed into a 500-mL incubation jar, maintaining 60% of the field capacity moisture content. All samples were pre-incubated at 20 °C for two weeks, following the procedure by Hamdi et al. [34], to mitigate the burst of respiration due to wetting the dry soils. Throughout the incubation, three samples of each treatment were used for the determination of soil CO2 concentration, The CO2 concentration determination was carried out on days 1, 7, 14, 24, 34, 49, and 63. During each gas sampling, compressed air was used to flush the headspace for 60 s to standardize the starting atmospheric CO2 concentration of each incubation jar [35]. Two hours later, the gas sample of each jar was collected again. The gas sample was injected into an evacuated 20 mL glass vial with a syringe to measure its CO2 concentration using a thermal conductivity detector at 400 °C on gas chromatography (GC-2014, Shimadzu, Kyoto, Japan) within 24 h. Three blank jars without soil were used to determine the background CO2 concentrations. To minimize the effect of different levels of SOC on the amount of CO2 produced, the cumulative CO2-C was expressed as a proportion of the SOC (μg CO2-C g-1 SOC h-1).
2.3. Soil Chemical Analyses
Total soil nitrogen (TN) and SOC were determined using a Vario MAX CN elemental analyzer (Elementar Vario EL III, Germany). The Dissolved organic carbon (DOC) and nitrogen (DON) were extracted from 10 g of incubation soil with 40 mL Milli Q water at 20 °C and shaking for 30 min [36]. The mixture solution was then centrifuged at 11.5×g for 20 min and then filtered through a 0.45-μm glass fabric filter paper. The DOC concentration was determined using a TOC analyzer (Shimadzu Corporation, Japan), the DON concentration was determined using a Continuous Flow Analyzer (SKALAR San++, Holland). Soil ammonium and nitrate determinations, 5 g of freshly collected soil was extracted with a 2 mol L-1 KCl solution [37]. The solution was shaken for 20 min and then filtered, and the concentrations of soil ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3--N) in the supernatant were determined using a Continuous Flow Analyzer (SKALAR San++, Holland).
2.4. Microbial Biomass C and N
Soil microbial biomass carbon (MBC) and nitrogen (MBN) were determined using the chloroform (CHCl3) fumigation and potassium sulfate (K2SO4) extraction techniques as described by Vance et al. [38] and Xu et al. [39]. In summary, CHCl3 was used to fumigate 5-g fresh soil for 24 h in dark plastic bags paired with unfumigated ambient. A 20-mL solution of 0.5 mol L-1 K2SO4 was used and the samples were fumigated and then shaken for 30 min at a rate of 250 r min-1. Samples were centrifuged at 13.1×g for 10 min and then passed through a 0.45-μm glass fabric filter paper. The filtrates were analyzed for organic C using a TOC analyzer (Shimadzu VCPH/TNM-1, Japan). The difference in organic C in fumigated and unfumigated samples was assumed to originate from MBC. MBC concentrations were corrected for unrecovered biomass using a conversion factor of 0.45. The difference in total N between fumigated and non-fumigated samples was assumed to originate from MBN. The filtrates were used to determine total N (TN) using a continuous flow analytic system analyzer (Skalar San++, Holland. Fitted with a TN unit), and TN was converted to MBN using a factor of 0.54. The ratio of MBC to SOC represents the microbial quotient (qMBC), and the ratio of basal respiration to total MBC represents the metabolic quotient (qCO2) [40].
2.5. Enzyme Analysis
The Enzyme analysis was conducted following a procedure described in Saiya-Cork et al. [41] and Sinsabaugh et al. [42]. Enzymes assayed in this study included β-1, 4-glucosidase (βG), Cellobiohydrolase (CBH); β-1,4-N-acetylglucosaminidase (NAG); Acid phosphatase (AP); Phenol Oxidase (PHO); Peroxidase (PEO). Suspensions of 1 g soil to 125 mL of acetate buffer at a concentration of 50 mol L-1 were prepared for each sample and agitated for 1 min using a Brinkmann Polytron PT 3000 homogenizer. The sample suspensions were continuously mixed with a magnetic stir plate during which 200 mL of the suspensions was portioned into 96-well microplates at 16 replicate wells per sample per assay. Fluorimetric assays were conducted for all enzymes except phenol oxidase and peroxidase. Fluorescence was measured using a microplate fluorometer with 365 nm excitation and 450 nm emission filter. After a correction for quenching and negative ambients, all activity units were presented as nmol h-1 g-1, and 4-dihydroxyphenylalanine (DOPA) was used as the substrate for spectrophotometrical analysis for phenol oxidase and peroxidase activity. Each sample well had the 50-μL solution of 25 mM DOPA and peroxidase assays had 10-μL of 0.3 % H2O2. Blank wells had 200-μL of the sample suspension and 50 μL of acetate buffer. A 50-μL solution of DOPA and 200 μL of acetate buffer were added to negative ambient wells for phenol oxidase. Negative ambient and blank wells for peroxidase had 10 μL of H2O2. This produced eight replicate wells for blanks and ambients, and 16 replicate sample wells for each enzyme assay. Microplates were incubated at 20 °C for 18 h in the dark. The activity was considered as a measure of absorbance at 450 nm by using a microplate spectrophotometer.
2.6. Phospholipid Fatty Acids (PLFAs) Analysis
At the end of the incubation, the microbial community structure was determined using the phospholipid fatty acids (PLFAs) analysis as described by Wan et al. [43]. In summary, an extracted mixture of chloroform, methanol, and citrate buffer with a volume ratio of 1:2:0.8 from a 10-g dry sieved soil was used for PLFA analysis. The extraction process included two phases: the chloroform phase and the citrate buffer phase. The lipid materials were recovered and evaporated under nitrogen gas during the chloroform phase. These lipids were then resuspended in chloroform and then fractionated on silicic chromatography acid columns (CNWBOND, 500 mg, 3 mL). Neutral lipids, glycolipids, and phospholipids were eluted with 5.0 mL of chloroform, acetone, and methanol, respectively. Nitrogen gas was used to dry the phospholipid fraction before a mild alkaline methanolysis was conducted to prepare for fatty acid methyl esters. For alkaline methanolysis, one mL of methyl alcohol and methylbenzene with a ratio of 1:1 in volume and then 1.0 mL of methanolic KOH were added to each sample. After swirling to mix, the samples were sealed and placed in a bath at 37 °C for 30 min. Afterward, 2.0 mL of hexane was added to each sample and after swirling 0.2 mL of 1.0 M acetic acid was added to each sample. 2.0 mL of deionized H2O was added to each sample to break phase and vortex samples for 30 s and then the samples were centrifuged for 2 min. The top phase was transferred to clean labeled 10-mL vials with a short Pasteur pipette. Afterward, 2.0 mL of hexane was added to each sample and then swirled. Nitrogen gas was also used to evaporate these methyl esters and stored at -20 °C until running gas chromatography analysis (GC). For the CG process, 200-μL HPLC-grade ethyl acetate was used to dissolve and separate the individual methyl esters. They were then run on GC with SGE 25QC3 BP-5 25 m × 0.32 μm film in thickness. A chromatographic retention time comparison to bacterial methyl esters was used to identify and quantify the separated fatty acids (Supeloc Bacterial Acid Methyl Esters CP Mix 47080-U). The abundance of individual fatty acids was expressed as nanomoles per gram of dry soil and using standard nomenclature. The following PLFAs were used as biomarkers: i14:0, i15:0, a15:0, i16:0, i17:0, and a17:0 for Gram-positive bacteria (GP), 16:1ω7c, cy17:0, 18:1ω7c, 18:1ω5c and cy19:0 for Gram-negative bacteria (GN), 18:1ω6c and 18:2ω9c for fungi, 10Me16:0, 10Me17:0 and 10Me18:0 for actinomycetes (ACT), and 16:1ω5c for arbuscular mycorrhizal fungi (AMF) [44,45]. Total bacterial PLFAs were calculated as the sum of Gram-negative and Gram-positive bacteria. The ratio of fungal to bacterial PLFAs (F:B) was used to estimate the relative importance of the bacterial and fungal metabolic presence in the community.
2.7. Calculation of Rate, Cumulative Mineralization, and Temperature Sensitivity
The CO2-C mineralization rate was determined using the following equation:
where R is mineralization rate (μg CO2-C g-1 SOC h-1), k is the coefficient of conversion to standard units of CO2 (1.964 g m-3), v is the jar volume (m3), m is the soil weight (g) and C is the SOC content (g kg-1), Δc/Δt is the concentration of CO2 per unit time (mg kg-1 h-1), and T is incubation temperature (°C) [46].
The cumulative CO2-C mineralization was determined using the following equation:
where Cm is cumulative mineralization (mg CO2-C g-1 SOC), R is the average mineralization rate between Ti and Ti+1 (μg CO2- C g-1 SOC h-1), T is incubation time between Ti+1 and Ti (Day), i is gas sampling time.
The short-term temperature sensitivity (i.e., based on differences in respiration between warmed and un-warmed soils) of soil decomposition for each incubation temperature was determined using the following equation:
where C is cumulative respiration, T is the temperature (°C), and the subscripts low and high indicate incubation at 20, 30, and 40 °C, respectively [47,48].
2.8. Statistical Analyses
Student’s-t test was used to analyze the differences in the measured items (i.e., cumulative mineralization, NH4+-N, NO3--N, DON, DOC, MBC, MBN, qMBC, qCO2, soil enzyme activity, and microbial community) between incubation for the different temperature treatments in un-warmed and warmed soil. Two-way ANOVA was performed to examine the effects of soil types and incubation temperatures on the cumulative SOC mineralization. The concentrations of the individual PLFAs (expressed as mol %) were subjected to principal component analysis (PCA) to determine the differences in soil microbial community composition between treatments.
We used a structural equation modeling (SEM) approach to examine the relationships among temperature, soil property, enzyme activity and microbial community and the Q10 of warmed and un-warmed SOC mineralization. All the data in the SEMs were scaling by one standard deviation [49]. We used composite variables to explain the collective effects of temperature (20, 30, and 40 ℃), soil property (NH4+-N, NO3--N, DON, DOC, MBC, MBN), enzyme activity (βG, CBH, NAG, AP, PHO, PEO) and microbial community (GP, GN, ACT, AMF, Fungi, Bacteria and F:B) on the Q10 of warmed and un-warmed SOC mineralization. Each of the composite variables was selected based on the multiple regression for mass loss rate and Akaike’s Information Criterion (AIC). Model fit was assessed using Fisher’s C statistic, where good-fitting models yield small C statistics and P values > 0.05 indicate that the data is well represented by the model. Piecewise SEM was based on linear mixed-effects models using the R package piecewise SEM. All the statistical analyses were performed in R v. 4.2.2 and with a significance level of 0.05.
3. Results
3.1. Soil Organic Carbon Mineralization and Its Temperature Sensitivity
Throughout the incubation period, the SOC mineralization rates for all treatments consistently declined (Figure 1a). Both soil types and incubation temperatures significantly influenced the mineralization of SOC, and their interaction was also notable (all p < 0.001, Table 2). As the incubation temperature rose, a marked increase in the SOC mineralization rates was observed (p < 0.05). By the end of the incubation, the cumulative SOC mineralization of un-warmed soil was highest at 40 ℃ (663.79 ± 6.70 μg CO2-C g-1), followed by 30 ℃ (420.72 ± 12.80 μg CO2-C g-1 soil) and 20 ℃ (322.14 ± 11.75 μg CO2-C g-1 soil). Similarly, the warmed soil exhibited a trend where cumulative SOC mineralization increased with temperature. However, for every temperature level, the un-warmed soil always displayed higher mineralization than the warmed soil (Figure 1b).
Regarding temperature sensitivity, variations in Q10 were observed for QT20 and QT30 between the two soils (Figure 2). The Q10 of 20-30 ℃ (QT20) and 30-40 ℃ (QT30) of the two soils varied significantly (Figure 2). The QT20 of the warmed soil (1.31 ± 0.03) was higher than that of the un-warmed soil (1.63 ± 0.07), but the QT30 of the warmed soil (1.58± 0.04) was lower than that of the un-warmed soil (1.41 ± 0.03) (Figure 2).
3.2. Soil Nutrients, MBC, Microbial Metabolic Quotients
With an increase in incubation temperature, Soil NO3--N levels rose significantly in both soil types. For the 40 ℃-incubation temperature, significant differences between the two soil types were observed for NH4+-N and DON levels (Table 3). Meanwhile, as temperatures rose, there was a decline in DOC and MBC, with un-warmed soils consistently recording higher levels than the warmed soils. For MBN, levels were notably higher at 40 ℃ for both soil types, with the un-warmed soil showing a more pronounced increase. Variations in soil qMBC and qCO2 were also influenced by temperature changes, with distinct patterns observed between the two soil types at certain temperatures (Table 3).
3.3. Soil Enzyme Activities and Microbial PLFAs
Increasing the incubation temperature from 20 ℃ to 30 ℃ boosted the activity of several soil enzymes, except AP (Figure 3). When the temperature was further increased, certain enzyme activities, like CBH and NAG, showed a decline. However, oxidase activity in warmed soil was consistently higher than in un-warmed soil across all temperatures (Figure 3).
The microbial community composition was also significantly impacted by temperature changes (Figure 4). All the PLFA biomarkers showed a decreasing trend (except for GP in the un-warmed soil). The PLFA of total microbes, total bacteria, GN, AMF, ACT, and fungus of the warmed soil significantly (p < 0.05) decreased compared to those of the un-warmed soil (Figure 4). 40 ℃ incubation significantly increased the ratio of gram-positive bacteria to gram-negative bacteria (GP:GN) for both soils and the ratio was significantly higher in the warmed soil than in the un-warmed soil (Figure 4). Increasing the incubation temperature significantly decreased F:B for both soils and the ration was also significantly higher in the warmed soil than the un-warmed soil for each incubation temperature (Figure 4). The first principal component of the PLFA pattern explained 82.35 % of the variation in the data, while PC2 explained another 11.21 % (Figure 5). According to the results of anosim analysis, there were significant differences in microbial community results between soil treatment and temperature treatment (p < 0.05) (Figure 5).
3.4. Factors Affecting the Temperature Sensitivity of SOC Mineralization
For the un-warmed soil, the Q10 of SOC mineralization showed several distinct relationships. It showed a significant positive correlation with the response ratios (RRs) of NH4+-N (Figure 6a) and DOC (Figure S1c). On the other hand, it had a significant negative correlation with the response ratios of NO3--N (Figure 6b), MBN (Figure S1b), and enzyme activities such as βG (Figure 6c), NAG, APC (Figure S1f,h), PHO, PEO (Figure 6d, e), and microbial communities such as GN, Fungi, F:B and GP:GN (Figure 6f,g,h,i).
For the warmed soil, it showed a significant negative correlation with the response ratios of NH4+-N (Figure 6a) and DON (Figure S1e), as well as with PHO and PEO (Figure 6d,e) and GP:GN (Figure 6i). Conversely, a significant positive correlation was observed with the RRs of NO3--N (Figure 6b), βG (Figure 6c) and CBH (Figure S1d), GN, Fungi, F:B (Figure 6f,g,h), AMF, ACT, Total PLFAs and Bacteria (Figure S1i,j,k,l).
The structural equation model explained 97 % and 98 % of the variance in the Q10 of un-warmed and warmed soil, respectively (Figure 7). This model depicted that incubation temperature, enzyme activity, microbial community, and soil properties directly and indirectly affected the Q10 of both soil types. Notably, while the effects of soil properties on Q10 were similar for both soil types, other factors, such as incubation temperature, enzyme activity, and microbial community, showed contrasting impacts on the two soils. For instance, in un-warmed soil, while the incubation temperature exhibited a positive effect on Q10, the soil properties, enzyme activity, and microbial community seemed to diminish Q10 (Figure 7a). On the other hand, in warmed soil, both incubation temperature and soil properties had a detrimental effect on Q10, while the enzyme activity and microbial community enhanced its value (Figure 7b).
4. Discussion
4.1. Response of SOC Mineralization of Un-Warmed and Warmed Soil to Warming
The enhancement of SOC mineralization rate by the increased incubation temperatures was evident for both un-warmed and warmed soil (Figure 1a). Additionally, the cumulative SOC mineralization from un-warmed soil was consistently higher than that from warmed soil at the same incubation temperature (Figure 1b). This aligns with many studies suggesting that higher temperature can intensify C mineralization and subsequently increase soil CO2 emission [22,48,50]. For instance, Gudasz et al. [51] also found that cumulative SOC mineralization escalated with increasing temperatures. Furthermore, our study indicated that greater SOC mineralization in un-warmed soil could be linked to the higher DOC and MBC levels in un-warmed soil than in warmed soil. Specifically, labile SOC contents, including DOC and MBC, were found to be 39-66 % and 20-31 % greater in the un-warmed soil (Table 3). For example, in the Great Hing’an Mountains in temperate northeast China, between 1.3-2.1 % of SOC was mineralized over a 42-day period at 15 °C without 13C-glucose [19]. Similarly, research from Northeastern China’s permafrost peatlands demonstrated 0.6-11.1 % decreases in SOC after 90-day incubation at 15 °C [18]. Contrastingly, in tropical and subtropical forests, SOC mineralization recorded between 2-7 % [52,53]. The positive relationship we observed between incubation temperature and soil NO3--N, NH4+-N, and DON (Table 3) mirrors previous findings. Warming has been shown to enhanced net N mineralization and nitrification rate, leading to N loss from ecosystems [54]. Additionally, Dawes et al. [55] and Melillo et al. [56] reported that experimental soil warming in temperate forests stimulated soil organic N mineralization. Lastly, temperature also impacts SOC mineralization by modulating soil microbial activity and community composition [57]. At the same temperature, the cumulative SOC mineralization of warmed soil was lower than un-warmed soil, because the PLFA of total microbes, total bacteria, fungi, and F:B significantly (p < 0.05) decreased (Figure 4). The greater SOC in the temperate region than the tropical and subtropical regions but the cumulative mineralization is not much higher, which imply that there was likely more sensitive to global warming than temperate and boreal forests in the tropical and subtropical region [58].
4.2. Response of the Q10 of SOC Mineralization of Un-Warmed and Warmed Soil to Rising Incubation Temperature
Q10 is strongly affected by substrate availability and soil microbial characteristics [23,59]. In our observations, the QT20 treatment displayed a peculiar trend where the Q10 of warmed soil exceeded that of un-warmed soil. This suggests that at this range, the sensitivity of the un-warmed soil changes might be somewhat reduced. This behavior aligns with Bradford et al. [60], who pointed out potential reductions in labile C and thermal adaptation of microbial decomposers as factors reducing temperature sensitivity of SOC mineralization. Furthering this line of thought, Domeignoz-Horta et al. [12] proposed that metrics closely associated with microbial biomass-such as total organic carbon and size of the extracellular enzymatic pool- alongside the temperature sensitivity of extracellular enzyme activity, played pivotal roles in dictating respiration temperature sensitivity. Delving deeper into enzyme activity, we noticed a significant uptick in PEO and PHO enzyme activities for un-warmed soil as incubation temperature rose from 20 ℃ to 30 ℃, whereas warmed soil showcased no such pronounced change. Both the un-warmed soil and warmed soil displayed a negatively correlation with the response ratio of oxidases enzymes activity. Interestingly, in the QT20 treatment, the Q10 of the warmed soil surpassed that of the un-warmed soil, hinting at the former’s heightened sensitivity within this temperature range. This observation contrasts with findings from numerous studies that have reported a higher Q10 at reduced temperatures [61]. Another intriguing point is that warmed soil SOC mineralization Q10 was not correlated with the RRs of MBC and DOC (Figure S1a,c). This is different from many studies, which reported substrate availability had a significant effect on temperature sensitivity [22,62,63]. The decrease in microbial quotient combined with the increase in metabolic quotient indicated that warming increased microbial respiration rather than microbial growth, and the higher qCO2 indicated a lower assimilation rate and higher maintenance C demand [64]. According to the metabolic theory [65], if microbes are active for a long period, cumulative maintenance respiration would be large but the population size of microbes would be small. This might explain the higher qCO2 yet reduced MBC at the highest incubation temperature.
In the QT30 treatment, the Q10 of warmed soil was significantly lower than that of un-warmed soil. Our results indicated that both DOC and MBC decreased with increasing incubation temperature; moreover, the levels of DOC and MBC in un-warmed soil were significantly higher than those in warmed soil at the same incubation temperature (Table 3). With the increase of incubation temperature (30 ℃increased to 40 ℃), the PEO and PHO of warmed soil did not increase significantly, but un-warmed soil was no significantly. Our study showed that increased incubation temperature stimulated the activity of C-degrading enzymes (βG and CBH) and the acid-resistant C-degrading enzymes (PHO and PEO) (Figure 3). The Q10 also was significantly related to the response ratios of βG and CBH enzymes activity, as well as PHO and PEO activities. The N-degrading enzyme activity increased with increased incubation temperature, but the P-degrading enzyme activity showed a downward trend. The Q10 of un-warmed soil was significantly correlated with the RRs of NAG and AP. The result on enzyme activity in the present study indicates that incubation temperature can differentially affect the extractable C- and acid-resistant C-degrading enzymes and consequently change the SOC decomposition, and the decrease of labile SOC of warmed soil will also accelerate the decomposition rate of non-labile SOC, because oxidases enzymes are responsible for decomposing recalcitrant C fractions, such as lignin and humus [66]. Compared with warmed soil, the Q10 of organic carbon decomposition of un-warmed soil is higher at 40 ℃, which may be because its microorganisms still have higher nutrient acquisition as the temperature increases, since nutrient availability also has a significant effect on the Q10 values [67].
The profound of global warming on the microbial mineralization of soil organic matter are well-documented [68]. The important role microbes play in modulating the temperature sensitivity of SOM decomposition has seen increased interest [23,69]. Our research corroborates this, highlighting that a surge in incubation temperature drastically reduces the total PLFAs, bacteria, fungi, GN, AMF, and ACT. Interestingly, warmed soil consistently higher than un-warmed soil under identical temperature treatment (Figure 4), likely because warming amplifies environmental stressors affecting soil microorganisms. SOC Mineralization is predominantly by bacteria and fungi [2]. In our study, soil microbial community composition shifted after incubation, there were significant differences in microbial community results between soil treatment and temperature treatment (p < 0.05) (Figure 5). Fungi are associated with a slow energy channel (slow turnover of more acid resistant and N-poor substrates, leading to high soil-C accumulation). In contrast, bacteria are associated with a fast energy channel (fast turnover of extractable and N-rich substrates), leading to low soil-C accumulation [70]. Our result showing that the higher GP:GN at 40 ℃ (Figure 4) and F:B decreased with increasing incubation temperature, which implies possible bacterial-dominated microbial communities. The shift in microbial communities could be the underlying reason for disparities in SOC mineralization across different incubation temperatures. Furthermore, Li et al. [8] show that the Q10 increased with the decrease of F:B. In our study, the Q10 of warmed soil SOC mineralization was negatively correlated with GP:GN (Figure 6i), while demonstrating a positively correlation with GN, Fungi, F:B (Figure 6f,g,h), AMF, ACT, Total PLFAs and Bacteria (Figure S1i,j,k,l). The structural equation model showed that incubation temperature exerts both direct and indirect effects on the Q10 of un-warmed soil. In contrast, for warmed soil, these effects manifest predominantly through microbial community shift (Figure 7). Therefore, it is plausible that the divergent temperature sensitivity between un-warmed soil and warmed soil across varied incubation temperatures are primarily driven by microbial community structure.
5. Conclusions
Our comprehensive study underscores the multifaceted impacts of experimental warming on SOC mineralization dynamics. The research distinctly illuminates the intensified SOC mineralization under elevated temperatures, concomitant with a marked reduction in soil DOC concentrations. Intriguingly, the escalated incubation temperatures induced discernible shifts in soil microbial communities, characterized by a decline in total microbial biomass, PLFAs, and the abundance of key microbial taxa, including bacteria, fungi, and GN. The qMBC and qCO2 patterns further attest to the dominance of microbial respiration processes in the warmed conditions, as opposed to microbial growth. Central to our findings is the revelation that the Q10 of SOC mineralization is intricately tethered to alterations in soil nutrients, enzymatic activities, and microbial community structures, especially the fungi to bacteria ratio. The nuanced disparities in Q10 values between un-warmed and warmed soils across varying incubation temperatures accentuate the evolving nature of the warming effects on SOC mineralization. As global temperatures continue their upward trajectory, these insights become paramount, urging the scientific community and policymakers to factor in the dynamic responses of SOC mineralization when forecasting the soil C feedbacks to ongoing climatic changes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This work was supported by the State Key program of the National Natural Science Foundation of China (No. 31930071) and the Major program of the National Natural Science Foundation of China (No. 32192433).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We thank Dr. Teng-Chiu Lin for his guidance in writing and comments on the manuscript. We also thank Xiaojie Li, Xianfeng Li and Chao Li for their help in laboratory analysis.
References
- Davidson, EA.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.W.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; Nannipieri, P.; Rasse, D.P.; Weiner, S.; Trumbore, S.E. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Cavaleri, M.A.; Reed, S.C.; Smith, W.K.; Wood, T.E. Urgent need for warming experiments in tropical forests. Global Change Biology 2015, 21, 2111–2121. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Liang, N.S.; Sha, Q.; Xu, X.L.; Zhang, Y.P.; Lu, H.Z.; Song, L.; Song, Q.H.; Xie, Y.N. Heterotrophic respiration does not acclimate to continuous warming in a subtropical forest. Scientific Reports 2016, 6, 21561. [Google Scholar] [CrossRef] [PubMed]
- Litton, C.M.; Giardina, C.P.; Albano, J.K.; Long, M.S.; Asner, G.P. The magnitude and variability of soil-surface CO2 efflux increase with mean annual temperature in Hawaiian tropical montane wet forests. Biochemistry 2011, 43, 2315–2323. [Google Scholar] [CrossRef]
- Giardina, C.P.; Litton, C.M.; Crow, S.E.; Asner, G.P. Warming-related increases in soil CO2 efflux are explained by increased below-ground carbon flux. Nature Climate Change 2014, 4, 822–827. [Google Scholar] [CrossRef]
- Brown, S.; Lugo, A.E. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 1982, 14, 161–187. [Google Scholar] [CrossRef]
- Li, H.; Yang, S.; Semenov, M. V.; Yao, F.; Ye, J.I.; Bu, R.; Ma, R.; Lin, J.; Kurganova, I.; Wang, X.; Deng, Y.E.; Kravchenko, I.; Jiang, Y.; Kuzyakov, Y. Temperature sensitivity of SOM decomposition is linked with a K-selected microbial community. Global Change Biology 2021, 27, 2763–2779. [Google Scholar] [CrossRef]
- Qin, S.; Kou, D.; Mao, C.; Chen, Y.; Chen, L.; Yang, Y. Temperature sensitivity of permafrost carbon release mediated by mineral and microbial properties. Science Advances 2021, 7, eabe3596. [Google Scholar] [CrossRef]
- Liu, X.F.; Yang, Z.J.; Lin, C.F.; Giardina, C.P.; Xiong, D.C.; Lin, W.S.; Chen, S.D.; Xu, C.; Chen, G.S.; Xie, J.S.; Li, Y.Q.; Yang, Y.S. Will nitrogen deposition mitigate warming-increased soil respiration in a young subtropical plantation? Agricultural and Forest Meteorology 2017, 246, 78–85. [Google Scholar] [CrossRef]
- Wang, Q.K.; Liu, S.; Tian, P. Carbon quality and soil microbial property control the latitudinal pattern in temperature sensitivity of soil microbial respiration across Chinese forest ecosystems. Global change biology 2018, 24, 2841–2849. [Google Scholar] [CrossRef] [PubMed]
- Domeignoz-Horta, L.A.; Pold, G.; Erb, H.; Sebag, D.; Verrecchia, E.; Northen, T.; Louie, K.; Eloe-Fadrosh, E.; Pennacchio, C.; Knorr, M.A.; Frey, S.D.; Melillo, J.M.; DeAngelis, K.M. Substrate availability and not thermal acclimation controls microbial temperature sensitivity response to long-term warming. Global change biology 2023, 29, 1574–1590. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.M.; O’Neill, T.A.; Ryburn, J.; Liang, L.L.; Arcus, V.L.; Schipper, L.A. Rapid laboratory measurement of the temperature dependence of soil respiration and application to changes in three diverse soils through the year. Biogeochemistry 2017, 133, 101–112. [Google Scholar] [CrossRef]
- Moinet, G.Y.; Dhami, M.K.; Hunt, J.E.; Podolyan, A.; Liáng, L.L.; Schipper, L.A.; Whitehead, D.; Nuñez, J.; Nascente, A.; Millard, P. Soil microbial sensitivity to temperature remains unchanged despite community compositional shifts along geothermal gradients. Global Change Biology 2021, 27, 6217–6231. [Google Scholar] [CrossRef] [PubMed]
- Pold, G.; Billings, A.F.; Blanchard, J.L.; Burkhardt, D.B.; Frey, S.D.; Melillo, J.M.; Schnabel, J.; van Diepen, L.T.A.; DeAngelis, K. Long-term warming alters carbohydrate degradation potential in temperate forest soils. Applied and environmental microbiology 2016, 82, 6518–6530. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, M.; Kuzyakov, Y.; Sanaullah, M.; Heitkamp, F.; Zelenev, V.; Kumar, A.; Blagodatskaya, E. Microbial decomposition of soil organic matter is mediated by quality and quantity of crop residues: mechanisms and thresholds. Biology and Fertility of Soils 2017, 53, 287–301. [Google Scholar] [CrossRef]
- Zhran, M.; Ge, T.; Tong, Y.; Deng, Y.; Wei, X.; Lynn, T.M.; Zhu, Z.; Wu, J.; Gunina, A. Assessment of depth-dependent microbial carbon use efficiency in long-term fertilized paddy soil using an 18O-H2O approach. Land Degradation & Development 2021, 32, 199-207.
- Song, Y.Y.; Liu, C.; Song, C.C.; Wang, X.W.; Ma, X.Y.; Gao, J.L.; Gao, S.Q.; Wang, L.L. Linking soil organic carbon mineralization with soil microbial and substrate properties under warming in permafrost peatlands of Northeastern China. Catena 2021, 203, 105348. [Google Scholar] [CrossRef]
- Jiang, L.; Ma, X.; Song, Y.Y.; Gao, S.Q.; Ren, J.S.; Zhang, H.; Wang, X.W. Warming-Induced Labile Carbon Change Soil Organic Carbon Mineralization and Microbial Abundance in a Northern Peatland. Microorganisms 2022, 10, 1329. [Google Scholar] [CrossRef]
- Jagadamma, S.; Mayes, M.; Steinweg, J.M.; Schaeffer, S.M. Substrate quality alters the microbial mineralization of added substrate and soil organic carbon. Biogeosciences 2014, 11, 4665–4678. [Google Scholar] [CrossRef]
- Pold, G.; Grandy, A.S.; Melillo, J.M.; Deangelis, K.M. Changes in substrate availability drive carbon cycle response to chronic warming. Soil Biology and Biochemistry 2017, 110, 68–78. [Google Scholar] [CrossRef]
- Li, X.J.; Xie, J.S.; Zhang, Q.F.; Lyu, M.K.; Xiong, X.L.; Liu, X.F.; Lin, T.C.; Yang, Y.S. Substrate availability and soil microbes drive temperature sensitivity of soil organic carbon mineralization to warming along an elevation gradient in subtropical Asia. Geoderma 2020, 364, 114198. [Google Scholar] [CrossRef]
- Karhu, K.; Auffret, M.D.; Dungait, J.A.J.; Hopkins, D.W.; Prosser, J.I.; Singh, B.K.; Subke, J.-A.; Wookey, P.A.; Ågren, G.I.; Sebastià, M.- T.; Gouriveau, F.; Bergkvist, G.; Meir, P.; Nottingham, A.T.; Salinas, N.; Hartley, I. P. Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 2014, 513, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Morrissey, E.M.; Mau, R.L.; Hayer, M.; Piñeiro, J.; Mack, M.C.; Marks, J.C.; Bell, S.L.; Miller, S.N.; Schwartz, E.; Dijkstra, P.; Koch, B.J.; Stone, B.W.; Purcell, A.M.; Blazewicz, S.J.; Hofmockel, K.S.; Pett-Ridge, J.; Hungate, B.A. The temperature sensitivity of soil: Microbial biodiversity, growth, and carbon mineralization. The ISME Journal 2021, 15, 2738–2747. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R.P. Soil enzyme activity: a brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biology and Fertility of Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Xu, X.Y.; Liu, X.R.; Li, Y.; Ran, Y.; Liu, Y.P.; Zhang, Q.C.; Li, Z.; He, Y.; Xu, J.H.; Di, H.J. High temperatures inhibited the growth of soil bacteria and archaea but not that of fungi and altered nitrous oxide production mechanisms from different nitrogen sources in an acidic soil. Soil Biology and Biochemistry 2017, 107, 168–179. [Google Scholar] [CrossRef]
- Gommers, P.; Van Schie, B.; Van Dijken, J.; Kuenen, J. Biochemical limits to microbial growth yields: An analysis of mixed substrate utilization. Biotechnology and bioengineering 1988, 32, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Gunina, A., & Kuzyakov, Y. From energy to (soil organic) matter. Global change biology 2022, 28, 2169-2182.
- LaRowe, D.E.; Van Cappellen, P. Degradation of natural organic matter: A thermodynamic analysis. Geochimica et Cosmochimica Acta 2011, 75, 2030–2042. [Google Scholar] [CrossRef]
- VandenEnden, L.; Anthony, M.A.; Frey, S.D.; Simpson, M.J. Biogeochemical evolution of soil organic matter composition after a decade of warming and nitrogen addition. Biogeochemistry 2021, 156, 161–175. [Google Scholar] [CrossRef]
- Liu, X.J.A.; Pold, G.; Domeignoz-Horta, L.A.; Geyer, K.M.; Caris, H.; Nicolson, H.; Kemner, K.M.; Frey, S.D.; Melillo, J.M.; DeAngelis, K.M. Soil aggregate-mediated microbial responses to long-term warming. Soil Biology and Biochemistry 2021, 152, 108055. [Google Scholar] [CrossRef]
- Li, Y.Q.; Qing, Y.X.; Lyu, M.K.; Chen, S.D.; Yang, Z.J.; Lin, C.F.; Yang, Y.S. Effects of artificial warming on different soil organic carbon and nitrogen pools in a subtropical plantation. Soil Biology and Biochemistry 2018, 124, 161–167. [Google Scholar] [CrossRef]
- Liu, X.F.; Chen, S.D.; Yang, Z.J.; Lin, C.F.; Xiong, D.C.; Lin, W.S.; Xu, C.; Chen, G.S.; Xie, J.S.; Li, Y.Q.; Yang, Y.S. Will heterotrophic soil respiration be more sensitive to warming than autotrophic respiration in subtropical forests?: Warming effect on soil respiration. European Journal of Soil Science 2019, 70, 655–663. [Google Scholar] [CrossRef]
- Hamdi, S.; Chevallier, T.; Aïssa, N.B.; Hammouda, M.B.; Gallali, T.; Chotte, J.; Bernoux, M. Short-term temperature dependence of heterotrophic soil respiration after one-month of pre-incubation at different temperatures. Soil Biology and Biochemistry 2011, 43.9, 1752–1758. [Google Scholar] [CrossRef]
- Huang, J.X.; Lin, T.C.; Xiong, D.C.; Yang, Z.J.; Liu, X.F.; Chen, G.S.; Xie, J.S.; Li, Y.Q.; Yang, Y.S. Organic carbon mineralization in soils of a natural forest and a forest plantation of southeastern China. Geoderma 2019, 344, 119–126. [Google Scholar] [CrossRef]
- Jones, D.; Willett, V. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biology and Biochemistry 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Carter, M.R.; Gregorich, E.G. Soil sampling and methods of analysis. Soil sampling and methods of analysis 2007, Chapter 19, 207–214. [Google Scholar]
- Vance, E.; Brookes, P.; Jenkinson, D. An extraction method for measuring soil microbial biomass C. Soil biology and Biochemistry 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Xu, G.; Chen, J.; Berninger, F.; Pumpanen, J.; Bai, J.; Yu, L.; Duan, B. Labile, recalcitrant, microbial carbon and nitrogen and the microbial community composition at two Abies faxoniana forest elevations under elevated temperatures. Soil Biology and Biochemistry 2015, 91, 1–13. [Google Scholar] [CrossRef]
- Kaschuk, G.; Alberton, O.; Hungria, M. Three decades of soil microbial biomass studies in Brazilian ecosystems: lessons learned about soil quality and indications for improving sustainability. Soil Biology and Biochemistry 2010, 42, 1–13. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. Effects of long-term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biology and Biochemistry 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Antibus, R.K.; Linkins, A.E.; McClaugherty, C.A.; Rayburn, L.; Repert, D.A.; Weiland, T. Wood decomposition over a first-order watershed: mass loss as a function of lignocellulase activity. Soil biology and biochemistry 1992, 24, 743–749. [Google Scholar] [CrossRef]
- Wan, X.H.; Huang, Z.Q.; He, Z.M.; Yu, Z.P.; Wang, M.H.; Davis, M.R.; Yang, Y.S. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant and soil 2015, 387, 103–116. [Google Scholar] [CrossRef]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biology and Biochemistry 2011, 43, 1621–1625. [Google Scholar] [CrossRef]
- McKinley, V.L.; Peacock, A.D.; White, D.C. Microbial community PLFA and PHB responses to ecosystem restoration in tallgrass prairie soils. Soil Biology and Biochemistry 2005, 37, 1946–1958. [Google Scholar] [CrossRef]
- Hu, R.G.; Hatano, R.; Kusa, K.; Sawamoto, T. Soil respiration and net ecosystem production in an onion field in Central Hokkaido, Japan. Soil science and plant nutrition 2004, 50, 27–33. [Google Scholar] [CrossRef]
- Creamer, C.A.; De Menezes, A.B.; Krull, E.S.; Sanderman, J.; Newton-Walters, R.; Farrell, M. Microbial community structure mediates response of soil C decomposition to litter addition and warming. Soil Biology and Biochemistry 2015, 80, 175–188. [Google Scholar] [CrossRef]
- Tian, Q.X.; Wang, X.G.; Wang, D.Y.; Wang, M.; Liao, C.; Yang, X.L.; Liu, F. Decoupled linkage between soil carbon and nitrogen mineralization among soil depths in a subtropical mixed forest. Soil Biology and Biochemistry 2017, 109, 135–144. [Google Scholar] [CrossRef]
- Lefcheck, J.S. Piecewise SEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods in Ecology and Evolution 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Wang, X.W.; Li, X.Z.; Hu, Y.M.; Lv, J.J.; Sun, J.; Li, Z.M.; Wu, Z.F. Effect of temperature and moisture on soil organic carbon mineralization of predominantly permafrost peatland in the Great Hing’an Mountains, Northeastern China. Journal of Environmental Sciences 2010, 22, 1057–1066. [Google Scholar] [CrossRef]
- Gudasz, C.; Bastviken, D.; Steger, K.; Premke, K.; Sobek, S.; Tranvik, L.J. Temperature-controlled organic carbon mineralization in lake sediments. Nature 2010, 466, 478–481. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L.; He, T.X.; Liu, L.; Wu, J.W. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biology and Biochemistry 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Wang, Q.K.; Zeng, Z.Q.; Zhong, M.C. Soil moisture alters the response of soil organic carbon mineralization to litter addition. Ecosystems 2016, 19, 450–460. [Google Scholar] [CrossRef]
- Bai, E.; Li, S.L.; Xu, W.H.; Li, W.; Dai, W.W.; Jiang, P. A meta-analysis of experimental warming effects on terrestrial nitrogen pools and dynamics. New Phytologist 2013, 199, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Dawes, M.A.; Schleppi, P.; Hättenschwiler, S.; Rixen, C.; Hagedorn, F. Soil warming opens the nitrogen cycle at the alpine treeline. Global Change Biology 2017, 23, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Melillo, J.M.; Butler, S.; Johnson, J.; Mohan, J.; Steudler, P.; Lux, H.; Burrows, E.; Bowles, F.; Smith, R.; Scott, L.; Vario, C.; Hill, T.; Burton, A.; Zhou, Y.M.; Tang, J. Soil warming, carbon–nitrogen interactions, and forest carbon budgets. Proceedings of the National Academy of Sciences 2011, 108, 9508–9512. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.G.; Valentine, D.W.; Ruess, R.W. Soil and root respiration in mature Alaskan black spruce forests that vary in soil organic matter decomposition rates. Canadian Journal of Forest Research 2005, 35, 161–174. [Google Scholar] [CrossRef]
- Zhou, P.; Li, Y.; Ren, X.E.; Xiao, H.A.; Tong, C.L.; Ge, T.; Brookes, P.; Shen, J.L.; Wu, J.S. Organic carbon mineralization responses to temperature increases in subtropical paddy soils. Journal of soils and sediments 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Eberwein, J.R.; Oikawa, P.Y.; Allsman, L.A.; Jenerette, G.D. Carbon availability regulates soil respiration response to nitrogen and temperature. Soil Biology and Biochemistry 2015, 88, 158–164. [Google Scholar] [CrossRef]
- Bradford, M.A.; Davies, C.A.; Frey, S.D.; Maddox, T.R.; Mohan, J.E.; Reynolds, J.F.; Treseder, K.K.; Wallenstein, M.D. Thermal adaptation of soil microbial respiration to elevated temperature. Ecology letters 2008, 11, 1316–1327. [Google Scholar] [CrossRef]
- Hamdi, S.; Moyano, F.; Sall, S.; Bernoux, M.; Chevallier, T. Synthesis analysis of the temperature sensitivity of soil respiration from laboratory studies in relation to incubation methods and soil conditions. Soil Biology and Biochemistry 2013, 58, 115–126. [Google Scholar] [CrossRef]
- Gershenson, A.; Bader, N.E.; Cheng, W.C. Effects of substrate availability on the temperature sensitivity of soil organic matter decomposition. Global Change Biology 2009, 15, 176–183. [Google Scholar] [CrossRef]
- Su, J.; Zhang, H.Y.; Han, X.G.; Penuelas, J.; Filimonenko, E.; Jiang, Y.; Kuzyakov, Y.; Wei, C.Z. Low carbon availability in paleosols nonlinearly attenuates temperature sensitivity of SOM decomposition. Global Change Biology 2022, 28, 4180–4193. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.H.; Domsch, K.H. Soil microbial biomass: the eco-physiological approach. Soil Biology and Biochemistry 2010, 42, 2039–2043. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Follstad Shah, J.J. Ecoenzymatic Stoichiometry and Ecological Theory. Annual Review of Ecology, Evolution, and Systematics 2012, 43, 313-343.
- Ding, F.; Sun, W.J.; Huang, Y.; Hu, X.Y. Larger Q10 of carbon decomposition in finer soil particles does not bring long-lasting dependence of Q10 on soil texture. European Journal of Soil Science 2018, 69, 336–347. [Google Scholar] [CrossRef]
- Xu, X.; Luo, Y.Q.; Zhou, J.Z. Carbon quality and the temperature sensitivity of soil organic carbon decomposition in a tallgrass prairie. Soil Biology and Biochemistry 2012, 50, 142–148. [Google Scholar] [CrossRef]
- Bai, Z.; Ma, Q.; Wu, X.; Zhang, Y.L.; Yu, W.T. Temperature sensitivity of a PLFA-distinguishable microbial community differs between varying and constant temperature regimes. Geoderma 2017, 308, 54–59. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef]
Figure 1.
Temporal patterns of SOC mineralization rates (a) and cumulative SOC mineralization during the incubation period (b) at different incubation temperatures (20 ℃, 30 ℃, 40 ℃) in un-warmed soil, and warmed soil. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
Figure 1.
Temporal patterns of SOC mineralization rates (a) and cumulative SOC mineralization during the incubation period (b) at different incubation temperatures (20 ℃, 30 ℃, 40 ℃) in un-warmed soil, and warmed soil. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
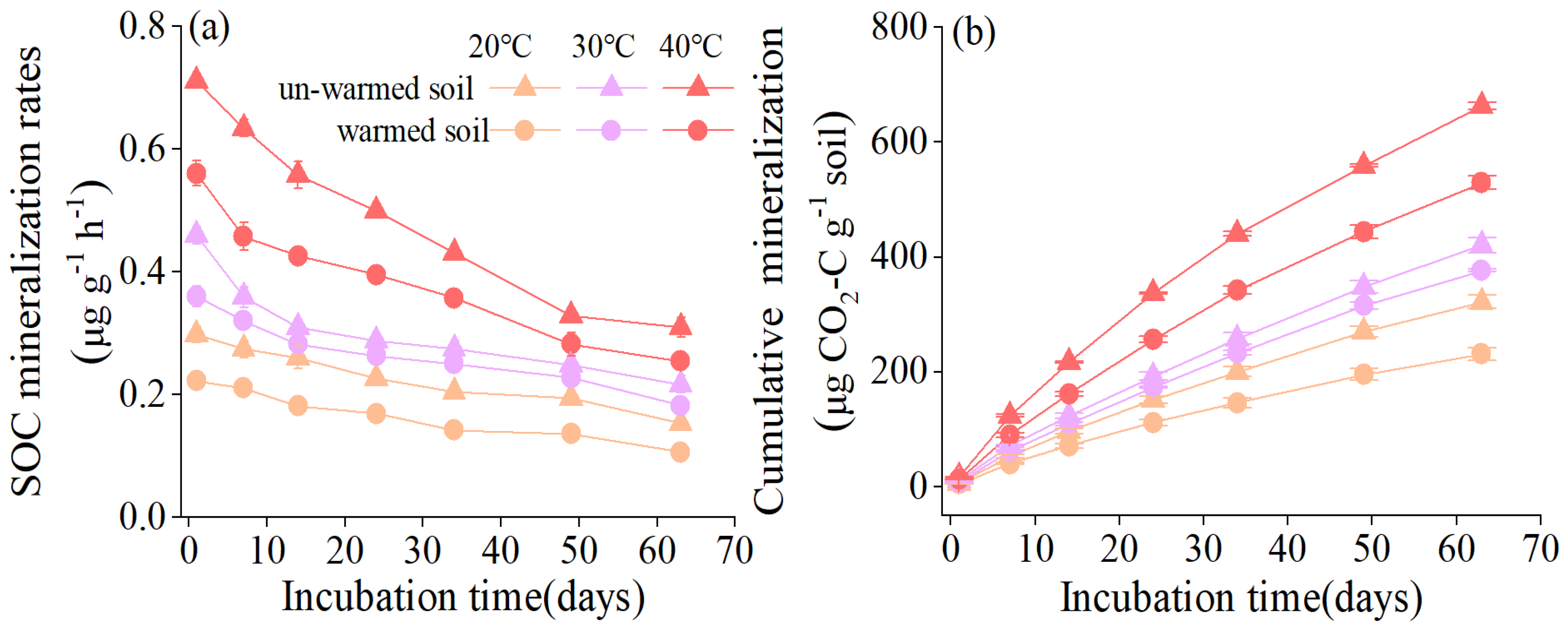
Figure 2.
The Q10 values of soil organic carbon mineralization of un-warmed soil and warmed soil over two temperature ranges (20-30 ℃ (QT20) and 30-40 ℃ (QT30)), respectively. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
Figure 2.
The Q10 values of soil organic carbon mineralization of un-warmed soil and warmed soil over two temperature ranges (20-30 ℃ (QT20) and 30-40 ℃ (QT30)), respectively. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
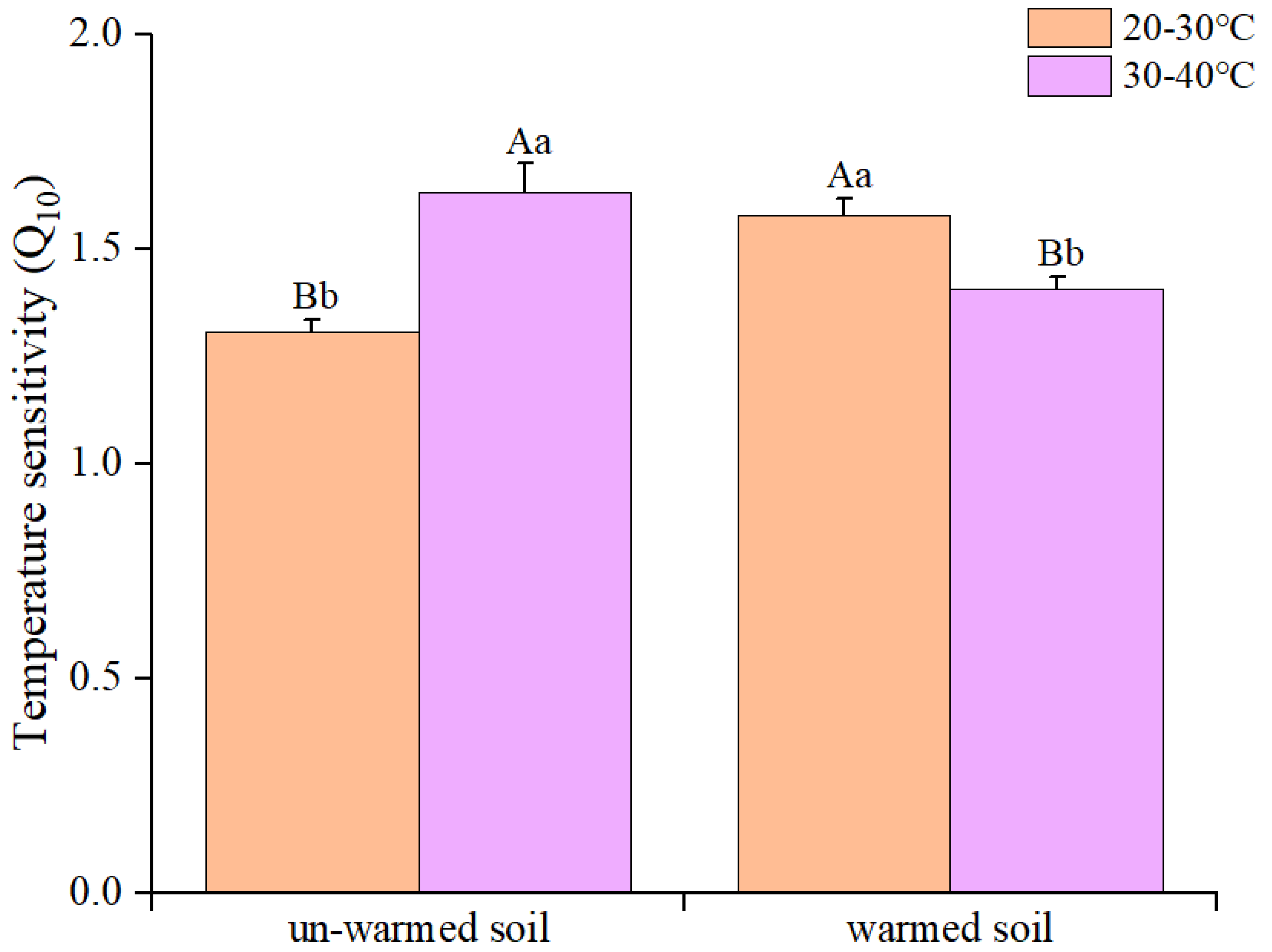
Figure 3.
Effects of incubation temperature on soil enzyme activity between un-warmed soil and warmed soil. βG: β-1, 4-glucosidase, CBH: Cellobiohydrolase; NAG: β-1, 4-N-acetylglucosaminidase; AP: Acid phosphatase. PHO: Phenol Oxidase; PEO: Peroxidase. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
Figure 3.
Effects of incubation temperature on soil enzyme activity between un-warmed soil and warmed soil. βG: β-1, 4-glucosidase, CBH: Cellobiohydrolase; NAG: β-1, 4-N-acetylglucosaminidase; AP: Acid phosphatase. PHO: Phenol Oxidase; PEO: Peroxidase. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
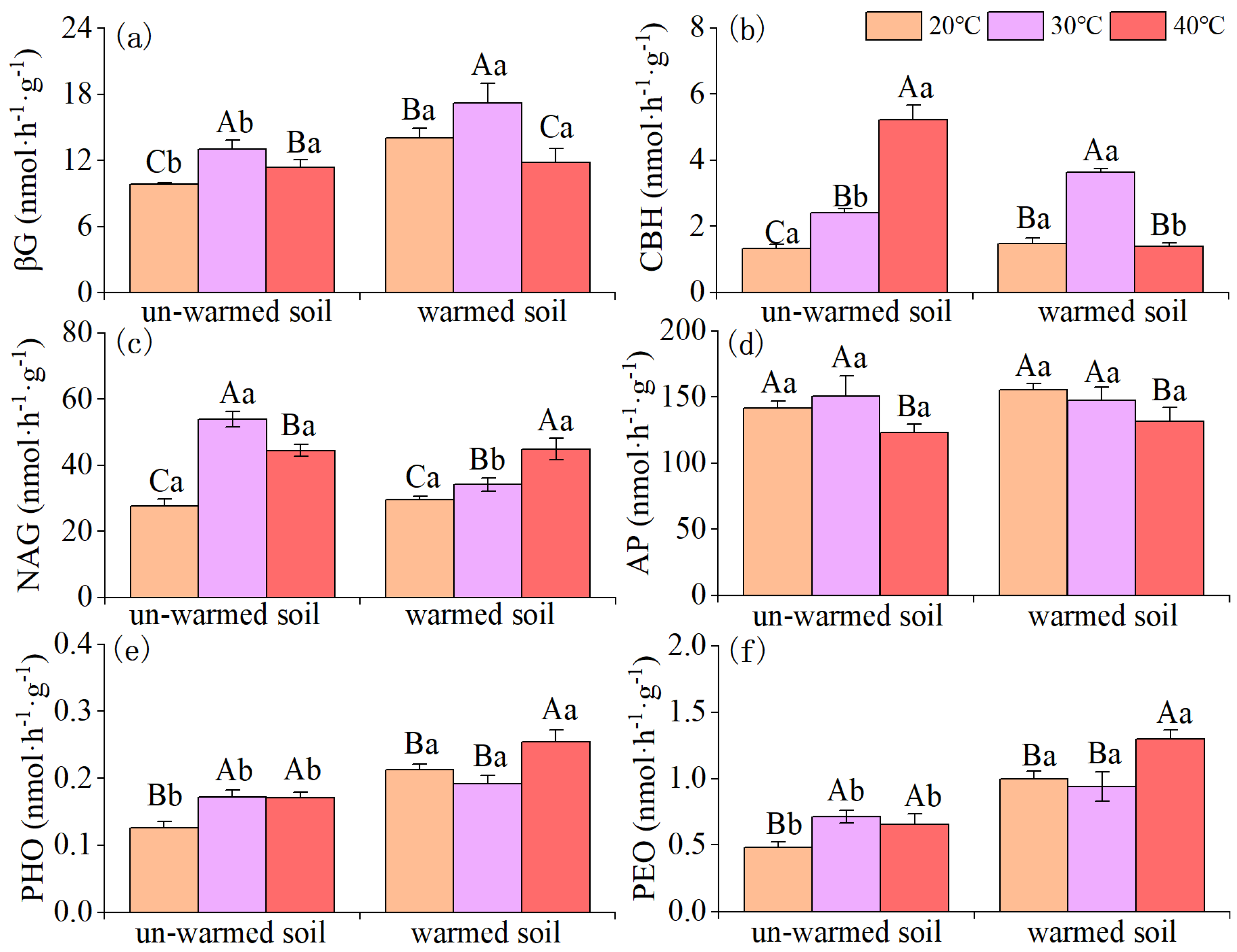
Figure 4.
Effects of incubation temperature on the phospholipid fatty acid biomarker contents (in nmol g−1 soil) between un-warmed soil and warmed soil. Total, total microbial PLFAs; fungi, total fungi PLFAs; total bacteria PLFAs, the sum of GP, GN and unspecific bacteria; AMF, Arbuscular mycorrhiza fungi; ACT: actinomycetes; F:B ratio, the ratio of total fungi to total bacteria PLFAs; GP, total gram-positive bacteria PLFAs; GN, total gram-negative bacteria PLFAs; GP:GN, the ratio of total gram-positive bacteria to gram-negative bacteria PLFAs. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
Figure 4.
Effects of incubation temperature on the phospholipid fatty acid biomarker contents (in nmol g−1 soil) between un-warmed soil and warmed soil. Total, total microbial PLFAs; fungi, total fungi PLFAs; total bacteria PLFAs, the sum of GP, GN and unspecific bacteria; AMF, Arbuscular mycorrhiza fungi; ACT: actinomycetes; F:B ratio, the ratio of total fungi to total bacteria PLFAs; GP, total gram-positive bacteria PLFAs; GN, total gram-negative bacteria PLFAs; GP:GN, the ratio of total gram-positive bacteria to gram-negative bacteria PLFAs. Bars are standard deviation (n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
Figure 5.
Principal components analysis (PCA) of microbial communities in the soils with three incubation temperatures: 20, 30, and 40 ℃. Blue circle: un-warmed soil, red circle: un-warmed soil. The higher the incubation temperature, the darker the color.
Figure 5.
Principal components analysis (PCA) of microbial communities in the soils with three incubation temperatures: 20, 30, and 40 ℃. Blue circle: un-warmed soil, red circle: un-warmed soil. The higher the incubation temperature, the darker the color.
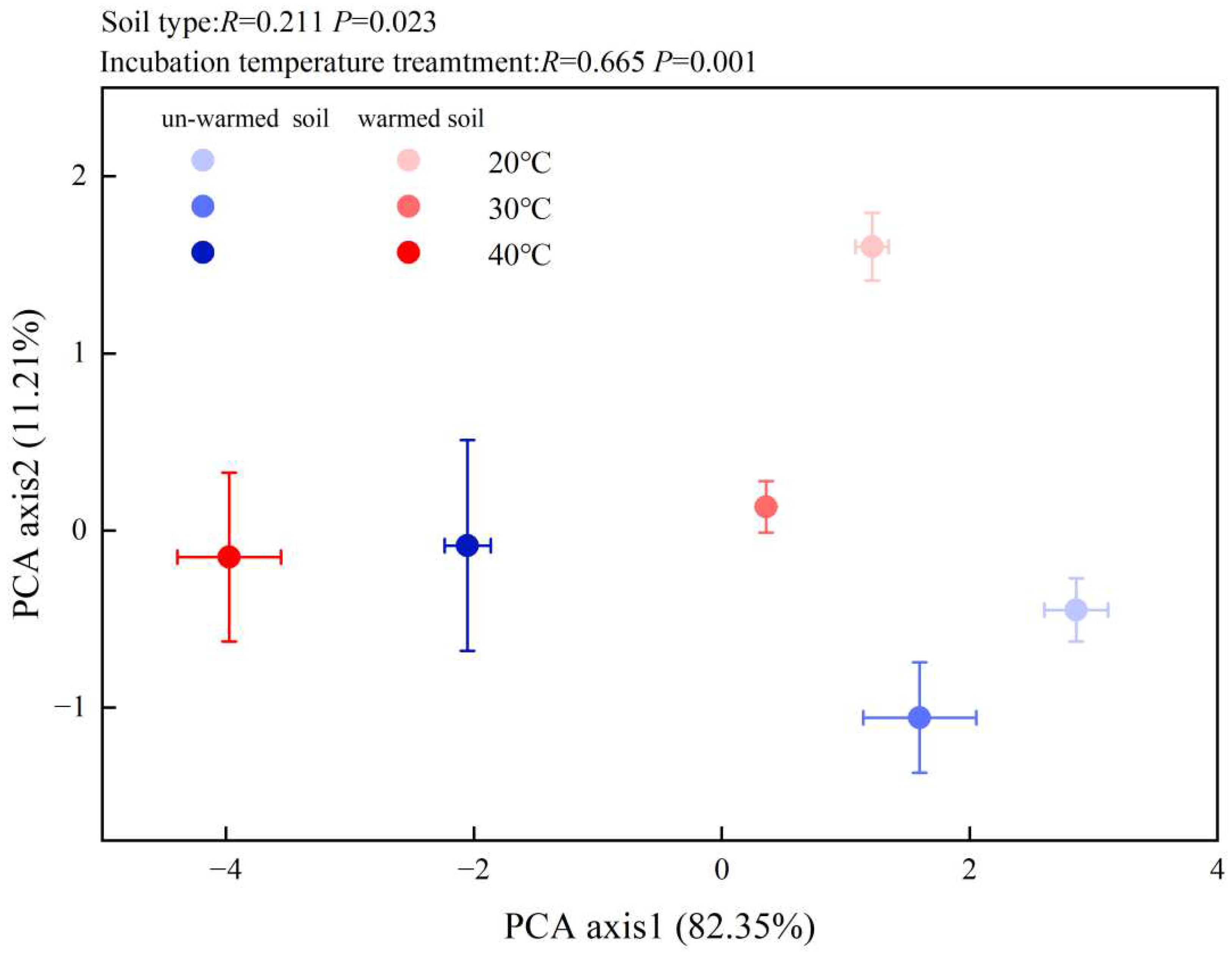
Figure 6.
Relationships between temperature sensitivity (Q10) over two temperature ranges (20-30 ℃ (QT20) and 30-40 ℃ (QT30)) and the response ratios (RRs) of NH4+-N (a), NO3--N (b), βG (c), PHO (d), PEO (e), GN (f), Fungi (g), F:B (h) and GP:GN (i). Black square and red point represent correlations in un-warmed and warmed soil, respectively. Blue and red lines represent relationships in un-warmed soil and warmed soil, respectively. Orange and red ranges represent 95% confidence interval in un-warmed soil and warmed soil, respectively.
Figure 6.
Relationships between temperature sensitivity (Q10) over two temperature ranges (20-30 ℃ (QT20) and 30-40 ℃ (QT30)) and the response ratios (RRs) of NH4+-N (a), NO3--N (b), βG (c), PHO (d), PEO (e), GN (f), Fungi (g), F:B (h) and GP:GN (i). Black square and red point represent correlations in un-warmed and warmed soil, respectively. Blue and red lines represent relationships in un-warmed soil and warmed soil, respectively. Orange and red ranges represent 95% confidence interval in un-warmed soil and warmed soil, respectively.
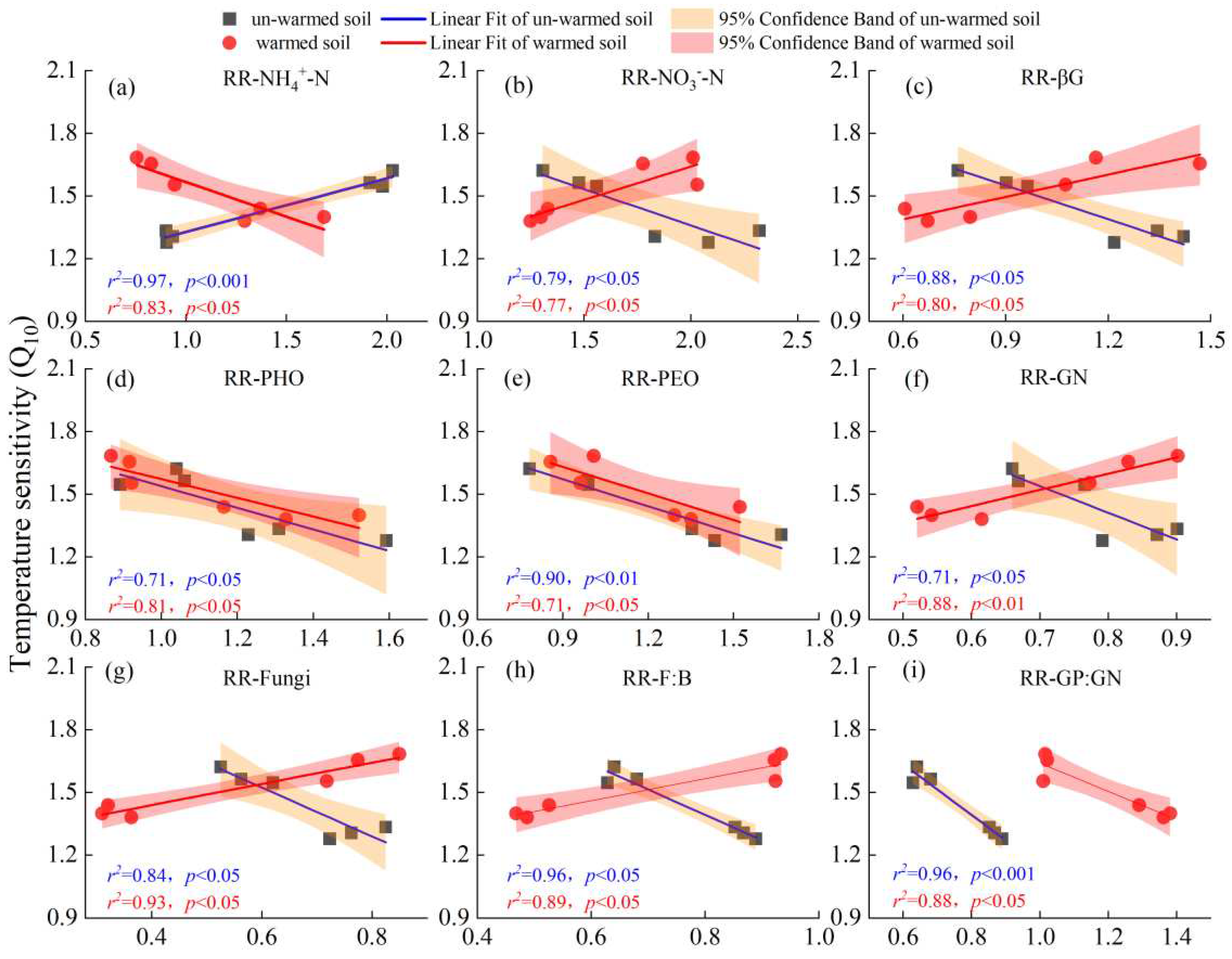
Figure 7.
Structural equation model exploring the direct and indirect effects of incubation temperature, soil propertiy, enzyme activity and microbial communitiy on the Q10 of soil organic carbon mineralization (a: un-warmed soil: The final results of model fitting were: C = 7.60, df = 1, P =0.006, AIC = 35.60, BIC=32.68, b: warmed soil: The final results of model fitting were: C = 0.50, df = 1, P =0.48, AIC =28.50, BIC=25.58). The soil properties are contents of NH4+-N, NO3−-N, MBC, MBN, DON and DOC. The enzyme activities are contents of βG, CBH, APC, NAG, PHO and PEO. The microbial communities are contents of GP, GN, ACT, AMF, Fungi, Bacteria and F:B. The blue boxes indicate the incubation temperature; the green boxes indicate the target factor (e.g. the Q10 of soil organic carbon mineralization of un-warmed soil and warmed soil, respectively); the orange boxes indicate the Biological and abiotic factors (e.g. soil properties, microbial communities and enzyme activities). The numbers adjacent to the arrows are standardised path coefficients. The solid lines indicate a positive effect and the dashed lines indicate a negative effect. The thickness of the lines indicates the size of the coefficients. The proportion of variance explained (R2) appears alongside each response variable in the model.
Figure 7.
Structural equation model exploring the direct and indirect effects of incubation temperature, soil propertiy, enzyme activity and microbial communitiy on the Q10 of soil organic carbon mineralization (a: un-warmed soil: The final results of model fitting were: C = 7.60, df = 1, P =0.006, AIC = 35.60, BIC=32.68, b: warmed soil: The final results of model fitting were: C = 0.50, df = 1, P =0.48, AIC =28.50, BIC=25.58). The soil properties are contents of NH4+-N, NO3−-N, MBC, MBN, DON and DOC. The enzyme activities are contents of βG, CBH, APC, NAG, PHO and PEO. The microbial communities are contents of GP, GN, ACT, AMF, Fungi, Bacteria and F:B. The blue boxes indicate the incubation temperature; the green boxes indicate the target factor (e.g. the Q10 of soil organic carbon mineralization of un-warmed soil and warmed soil, respectively); the orange boxes indicate the Biological and abiotic factors (e.g. soil properties, microbial communities and enzyme activities). The numbers adjacent to the arrows are standardised path coefficients. The solid lines indicate a positive effect and the dashed lines indicate a negative effect. The thickness of the lines indicates the size of the coefficients. The proportion of variance explained (R2) appears alongside each response variable in the model.
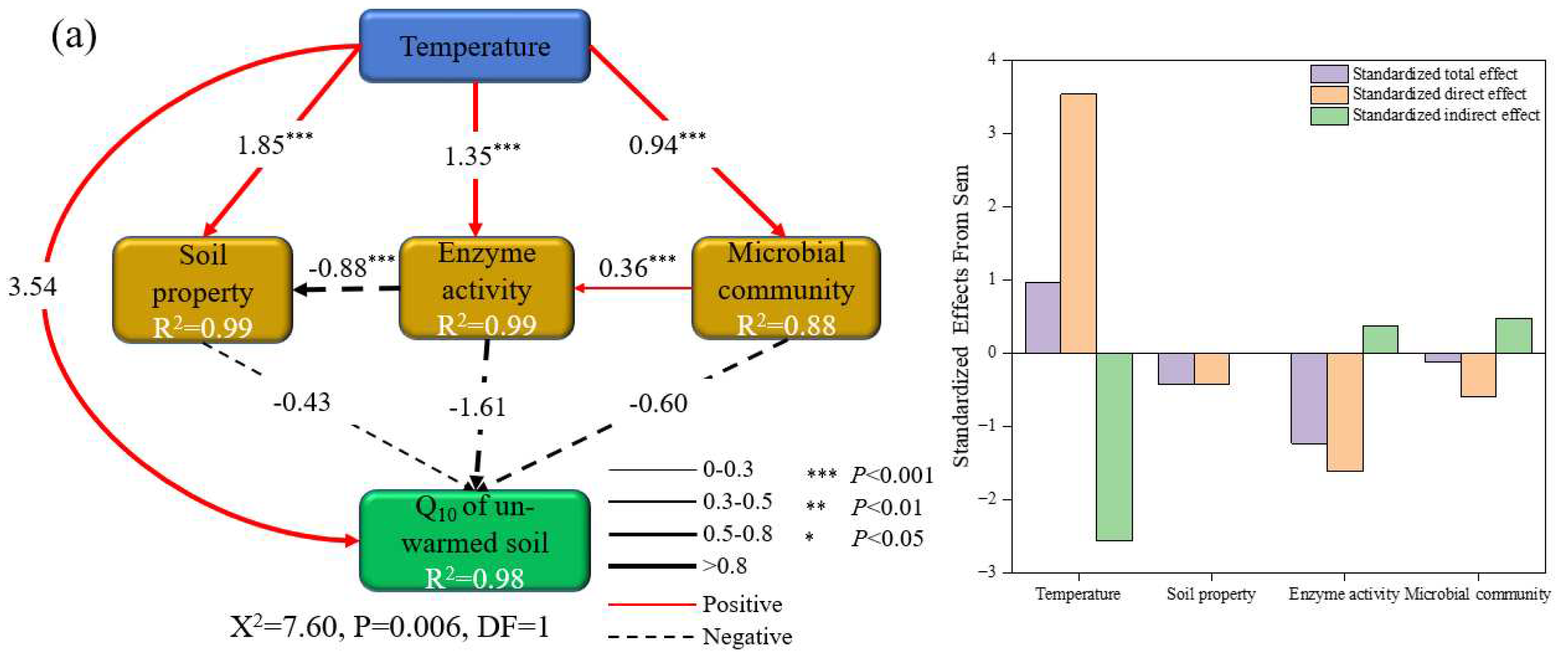
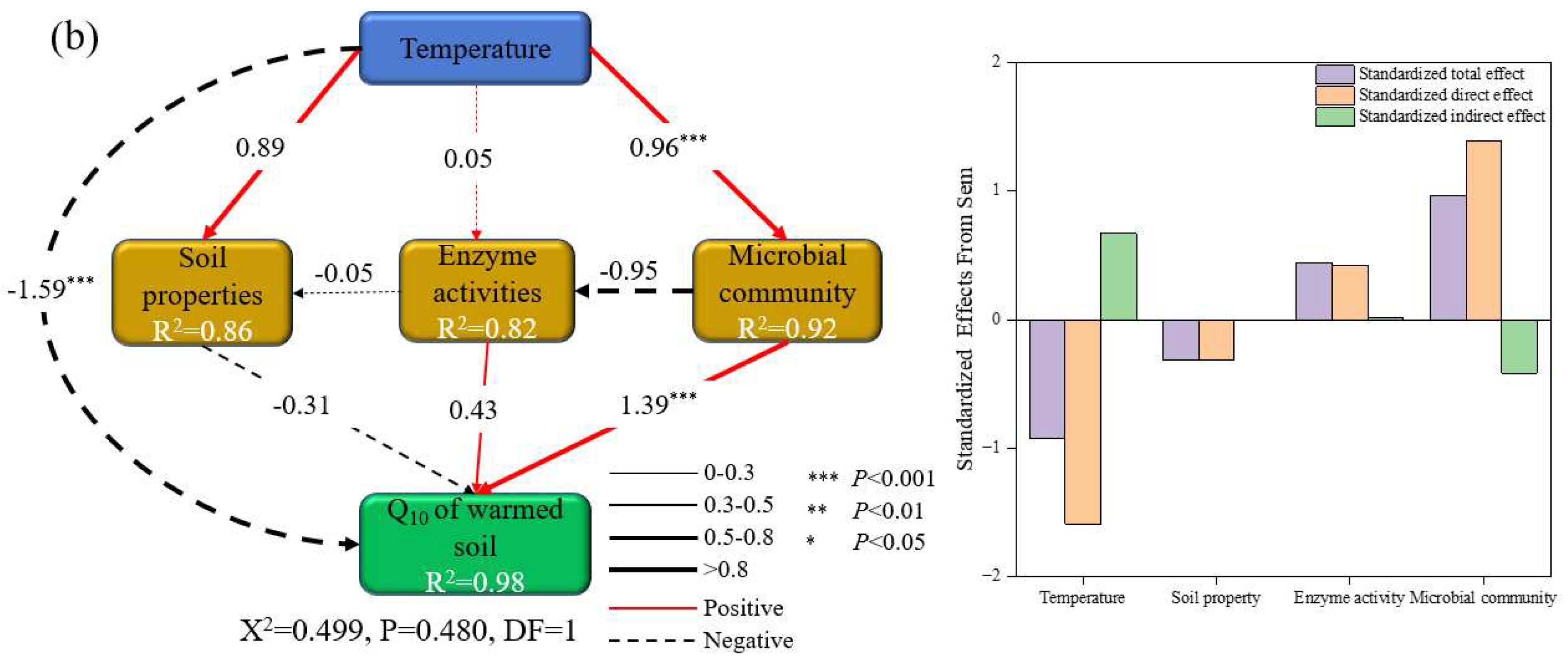

Table 1.
Mean soil organic carbon (SOC), total N (TN), post-incubation soil ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3--N), dissolved organic N (DON), dissolved organic C (DOC), microbial biomass carbon (MBC), nitrogen (MBN), microbial quotient, and metabolic quotient at different incubation temperatures in un-warmed and warmed soil. Values are expressed as (mean ± standard deviation; n = 3). Different capital letters denote significant difference between situ un-warmed soil and warmed soil (p < 0.05).
Table 1.
Mean soil organic carbon (SOC), total N (TN), post-incubation soil ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3--N), dissolved organic N (DON), dissolved organic C (DOC), microbial biomass carbon (MBC), nitrogen (MBN), microbial quotient, and metabolic quotient at different incubation temperatures in un-warmed and warmed soil. Values are expressed as (mean ± standard deviation; n = 3). Different capital letters denote significant difference between situ un-warmed soil and warmed soil (p < 0.05).
| Treatment | SOC | TN | NH4+-N | NO3--N | DON | DOC | MBC | MBN |
| (mg·g-1) | (mg·g-1) | (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | |
| un-warmed soil | 13.02±1.14A | 1.12±0.08A | 4.48±0.60A | 2.04±0.54A | 1.66±0.21A | 13.58±1.46A | 285.50±20.74A | 24.60±2.20A |
| warmed soil | 11.60±1.38A | 0.98±0.09A | 4.08±1.38A | 2.32±0.45A | 1.17±0.25A | 9.86±2.24B | 203.55±28.75B | 19.07±2.43B |
Table 2.
Results of two-way ANOVA for responses of the cumulative SOC mineralization to soil type and temperature (20, 30, and 40 °C). ***: p < 0.001.
Table 2.
Results of two-way ANOVA for responses of the cumulative SOC mineralization to soil type and temperature (20, 30, and 40 °C). ***: p < 0.001.
| Treatment | Cumulative SOC mineralization | ||||||
| 1 d | 7 d | 14d | 24d | 34 d | 49d | 63d | |
| Temperature | 1010.31*** | 807.56*** | 1130.97*** | 1718.71*** | 1449.05*** | 1380.53*** | 1515.73*** |
| Soil type | 249.58*** | 215.57*** | 228.79*** | 386.28*** | 307.45*** | 308.32*** | 349.39*** |
| Temperature × Soil type | 10.36*** | 37.93*** | 45.45*** | 58.25*** | 39.60*** | 31.91*** | 29.24*** |
Table 3.
Mean post-incubation soil ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3--N), dissolved organic N (DON), dissolved organic C (DOC), microbial biomass carbon (MBC), nitrogen (MBN), microbial quotient, and metabolic quotient at different incubation temperatures in un-warmed and warmed soil. Values are expressed as (mean ± standard deviation; n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
Table 3.
Mean post-incubation soil ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3--N), dissolved organic N (DON), dissolved organic C (DOC), microbial biomass carbon (MBC), nitrogen (MBN), microbial quotient, and metabolic quotient at different incubation temperatures in un-warmed and warmed soil. Values are expressed as (mean ± standard deviation; n = 3). Different capital letters denote significant difference among incubation temperatures and different lower-case letters denote significant difference between un-warmed soil and warmed soil at the same incubation temperature (p < 0.05).
| Treatment | NH4+-N | NO3--N | DON | DOC | MBC | MBN | Microbial quotient | Metabolic quotient | |
| (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | (mg·kg-1) | (%) | (mg CO2-C g−1 MBC h−1) | ||
| un-warmed soil | 20 ℃ | 15.50±0.76Ba | 7.73±0.67Ca | 1.42±0.27Ba | 26.03±1.99Aa | 233.18±14.38Aa | 23.31±3.11Aa | 1.80±0.13Aa | 0.66±0.06Ca |
| 30 ℃ | 14.14±0.55Ba | 11.08±0.76Bb | 2.57±0.8Ba | 18.67±1.68Ba | 167.65±9.02Ba | 25.82±4.54Aa | 1.34±0.22Ba | 1.30±0.09Ba | |
| 40 ℃ | 27.88±0.72Aa | 15.96±0.51Ab | 5.68±0.18Aa | 15.88±0.43Ca | 94.26±6.12Ca | 17.54±2.28Ba | 0.78±0.07Ca | 3.30±0.38Aa | |
| warmed soil | 20 ℃ | 15.95±1.47Ba | 6.88±0.28Ca | 1.48±0.37Ba | 15.71±1.05Ab | 178.58±12.37Ab | 18.03±3.31Aa | 1.44±0.15Ab | 0.60±0.08Ca |
| 30 ℃ | 13.46±2.09Ba | 13.32±0.66Ba | 1.71±0.15Ba | 13.45±0.74Bb | 136.89±9.21Bb | 20.69±2.27Aa | 1.15±0.08Ba | 1.34±0.10Ba | |
| 40 ℃ | 19.22±0.63Ab | 17.20±0.30Aa | 4.54±0.41Ab | 10.96±1.08Cb | 79.23±7.18Cb | 13.32±1.72Bb | 0.67±0.06Ca | 3.23±0.25Aa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



