Submitted:
01 October 2023
Posted:
01 October 2023
You are already at the latest version
Abstract
Modern research achievements support health-promoting effects of natural products and diets rich in polyphenols. Pomegranate (PG) (Punica granatum L.) contain a considerable number of bioactive compounds that exert a broad spectrum of beneficial biological activities, including antimicrobial, antidiabetic, antiobesity, and atheroprotective properties. In this view, the reviewed literature shows that PG intake might reduce insulin resistance, cytokine levels, redox gene expression, blood pressure elevation, vascular injuries and lipoprotein oxidative modifications. Lipid parameters corrective capabilities of PG-ellagitannins are also sufficiently reported to be significantly effective in reducing hyperlipidemia (TC, LDL-C, VLDL-C, and TAGs), plasma HDL-C concentrations, TC/HDL-C and LDL-C/HDL-C ratio. The health benevolent effects of pomegranate consumption appear to be produced through the amelioration of adipose tissue endocrine function, fatty acid utilization, GLUT receptor expression, paraoxonase activity enhancement, PPAR and NFκB modulation. Although the results from animal experiments are encouraging, human findings published in this field are inconsistent and still limited in many points of view. The present review discusses and provides a critical analysis of PG's bioeffects on metabolic syndrome components, type-2 diabetes, obesity, and dyslipidemia, as well as on some cardiovascular-related diseases. A brief overview of the pharmacokinetic properties, safety, and bioavailability of PG-ellagitannins is also included.
Keywords:
Punica granatum L.
; pharmacokinetic
; metabolic disorders
; dyslipidaemia
1. Introduction
The major advances made in the history of humanity, particularly in the fields of transportation, food security and telecommunication, have greatly facilitated the life of the 21st-century humans. As a result of this materialistic evolution, humans have become more and more sedentary and increasingly under stressful situations. Simultaneously, other non-healthy behaviours, such as sleep deprivation, smoking, alcoholism, and non-nutritious dietary habits, have worsened the situation. These lifestyle changes have largely impacted human health capital and are potentially incriminated in the occurrence of numerous chronic illnesses, including cardio-metabolic and vascular diseases.
The above analysis explains, at least in part, the following worrying prevalence rates: in 2016, nearly 1.9 billion adults are already diagnosed as overweight worldwide [1] and 650 million of them are obese. In June 2021, alarming percentages concerning overweight and obesity rates (39% and 13% of adults, respectively) have been communicated by the same organization. A simple comparison of these rates with those registered in the middle of the 1970s, that were three times lower than today’s [1], highlighted how much the world is at real risk of a global epidemic, especially as future projections continue to increase in an exponential way. Another factor that complicates the situation is childhood obesity, which is becoming more and more noticeable. This means more chances to develop cardiovascular diseases, insulin resistance, psychological disorders, and ultimately premature mortality.
According to the International Diabetes Federation [2], 415 million diabetics have been positively diagnosed in 2015. Future projections are alarming and the federation predicts approximately 642 million diabetics by the year 2040 [3]. From an economic point of view, the global financial burden of diabetes illnesses was estimated at approximately US $1.31 trillion in 2015 [4]. The estimations have included all types of diabetes, 184 countries, adults aged between 20 and 79 years, direct and indirect expenditures. This burden is expected to increase as the incidence of obesity continues to elevate.
Pharmacological treatments, including anti-obesity drugs, insulin-sensitizing molecules and insulin-secretagogue agents, have provided solutions to these illnesses. However, pharmacovigilance reports and case series studies [5,6] have signalled serious adverse effects related to the use of some of these agents. In this context, troglitazone has been removed from the US market in March 2000 [5], and the prescription of others is being questioned and largely restricted [7], particularly for reasons related to hypoglycaemia or coma risk [8,9,10], [8,9,10] weight gain [11,12], cardiac events [13,14,15], hepatic dysfunction [5,16], and nephrotoxicity [17].
A large body of evidence accumulated in the last decades supports the safety of natural nutrients and their strong correlation with cellular homeostasis, chronic disease prevention, cognitive function, and life quality improvement. Therefore, various research teams have focused their research interest in identification and characterization bioactivities from natural compounds. Aromatic and medicinal plants, such as pomegranate (PG), have been reported in the ethnomedicine literature to treat microbial infections, diabetes mellitus, obesity and its-related metabolic abnormalities. Currently, empirical arguments confirm these folk remedy applications and attribute to PG consumption strong normoglycemic and anti-dyslipidemia properties.
The present review discusses the most relevant published in vitro and in vivo data related to the anti-hyperglycaemic and anti-hyperlipidaemic regulatory effects of PG consumption. Relevant clinical and pharmacokinetic studies performed in this order are also reviewed.
2. Pharmacokinetic and safety of ellagitannin constituents
Human investigations on the bioavailability of ellagitannins (ETs) and ETs-derivative nutrients are relatively recent, since the first paper on ETs from PG was published in 2004 [18]. Related animal studies are also recent and have been performed using mainly rats as an experimental model to investigate ET biotransformation, absorption, and clearance [19].
3. Catabolism of ellagitannins
It is proven that under physiological conditions, ET polyphenols are subject to non-enzymatic hydrolysis, involving the acidic degradation and the lysis by the intestinal bacteria. In colon area, the resulting ellagic acid (EA) molecules are subjected to biochemical modifications, namely dihydroxylation, decarboxylation, and lactone-ring cleavage to produce dibenzopyran-6-one derivatives known as urolithins [20]. In this regard, analysis of human fecal culture has identified urolithin A, thought to be produced from EA and related polyphenols [20]. However, glucuronide or sulfate conjugates of this metabolite are not detected [20]. In the same context, a study from Cerdá et al., [19] demonstrated that rat intestinal microflora was able to transform ET-punicalagin to the urolithin metabolites.
Studies on jejunum and colon tissues [21] of Iberian pigs fed acorn ETs have demonstrated the presence of EA compounds and their bioconversion products. These studies also revealed inter-individual variability in terms of the rate and profile of metabolites produced, primarily because catabolic physiological capacities are dependent on the composition and efficiency of gut microbial organisms, which vary greatly between individuals.
4. Absorption and bioavailability of ellagitannins
Scientific experiments conducted to investigate the ability of PG compounds to penetrate the intestinal lumen and to achieve human blood, suggest the bioavailability of EA at the following concentrations : 31.9 ng/mL [18] and 33 ng/mL [22]. These values are achieved in a post-ingestion time of 1 h and the reported quantities are eliminated at T-max of 4 h. Additionally, ET-punicalagin molecules were detected at a concentration of 30 µg/mL, in a rat animal model [19]. However, polyphenols or related hydrolysis products from other species like raspberries were not able to achieve the plasma circulation of healthy volunteers [23]. The production capacity of urolithins and their absorption capabilities, present a great variability and many factors have been proposed to explain this variability; such as molecule physicochemical characteristics (the chemical structure, degree of lipophilicity, solubility, etc.), and the individual specificities (microbiota composition, intestinal pH, etc.).
5. Biodistribution and clearance of ellagitannins
Urolithins tissue accumulation outcomes are in favor of an important deposition in the colon and prostate organs [24]. However, the capabilities of ET metabolites including punicalagin, to achieve and to accumulate in other organs such as the liver and kidney are very limited [19]. Moreover, others investigators did not report any detection [21]. This last publication did not suggest any deposition of these molecules in the brain, adipose, and muscle tissues. The distribution and/or deposition abilities of PG-ETs may be influenced by organ cells biochemical characteristics, such as the selective properties and/or permeability (e.g. in the brain, the blood-brain barrier could limit the access of these molecules to the intraneuronal compartment), the general structure of the ET-molecule (which gives it or not a susceptibility to resist the hepatic catabolism reactions), and personal blood transport performance.
In terms of clearance, punicalagin metabolites are detected in urine under 6-H-dibenzo[b,d]pyran-6-one derivative forms [19]. Indeed, feces and urine investigations were able to quantify around 3–6 % of the total polyphenols ingested (0.6 to 1.2 g, daily). The authors hypothesized the quasi-biotransformation of these compounds into undetectable molecules or a possible accumulation in non-investigated tissues [19]. Furthermore, clearance of ETs-polyphenols from other sources have been also studied, and the post-ingestion of 35 g of walnuts excretion analysis, showed the presence of urolithin A, without any detection of its sulfate or glucuronide derivatives [20]. However, feces elimination of urolithins glucuronide forms is confirmed in another study [21], thing which is in agreement with the discussed inter-individual variability in terms of absorption rat, gut-microbiota composition and ETs-microflora degradation abilities.
6. Safety of pomegranate and pomegranate products
Although traditional remedy literature supports PG utilization and its capacities to ameliorate health performances, some toxicological evaluations indicated cellular components alteration and nuclear damages after PG administration. According to Tripathi and Singh [25], the concentration required to kill 50% of snail Lymnaea acuminate (LC50) using PG bark was fixed at 22.42 mg/l and vary in a dose- and time-dependent manner. In addition, recombinogenic, mutagenic, and clastogenic effects, have been observed in mice after whole PG fruit consumption [26], suggesting that PG may contain toxic substances, most likely, the alkaloids ingredients [25].
Conversely, in vivo data [27], have been recently published attributing an antigenotoxic actions of PG fruit administered at different concentrations (87.5, 175, 350, and 700 mg/kg of b.w). Additionally, 21 days of subacute toxicity assays, also did not reveal any toxicity markers. Moreover, an embryo toxicity study [28] showed that the hydroalcoholic extract of PG fruit was safe at a dose less than 0.1 mg per embryo, and its intraperitoneal administration lethal dose 50 (LD50), is reached only in high dose (731.1 mg/kg). Furthermore, Jahromi et al., [29], suggest the fact that the administration of the following doses : 0.5, 1.9 and 7.5 mg/kg of PG peel extract, did not provoke any toxic signs or behavioral disorder symptoms. Also, data from this paper, do not suggest any cholesterol, glucose, or hepatic enzyme disturbances.
PG pure dietary-compounds and metabolites, such as punicalagin and EA have been also investigated for their toxicity and safety properties [30,31]. In a 90-day subchronic experiment [30], 1.25, 2.5, and 5% doses of EA, have been given to F344 rats as a powdered basal diet to evaluate its bio-alteration potential. Outcomes from this study did not show any toxic effects or supplementation-related clinical signs. Moreover, 37 days repeated oral administration of a 6% ETs-punicalagin did not produce any liver or kidney toxicity [31]. Furthermore, this compound is not only safe but also can protect the hippocampal HT22 cells and H9c2 cardiomyocytes from glutamate and doxorubicin-induced toxicity [32,33]. In vitro and in vivo toxicological examinations of PG-seed oil, a rich source of punicic acid concluded that this treatment was neither mutagenic nor clastogenic, and the post-mortem analysis did not reveal any cellular abnormalities [34]. In this experiment, the no observed adverse effect concentration was fixed at 4.3 g/ kg/ day.
In all, PG or PG pure chemical intake seems to be safe, and the doses expected to have toxic effects, are much higher than those reported in ethnomedicine remedies and than those currently used for healing purposes.
7. Pomegranate consumption and obesity
This pathology has a multifactorial etiology, which includes genetic, epigenetic, and environmental roots. Indeed, the unlimited access to a diet often rich in fat, and poor in fibres, with a considerable reduction of physical activity, in addition to the in utero epigenetic modifications, represent the major elements underlying the obesity epidemic.
Physiopathological aspects of this energetic imbalance are illustrated in the hormono-metabolic change that occurs in the adipose tissue, due to its exposure to nutritional affluence that exceeds the organism’s spending. In fact, the abdominal hypertrophy is accompanied by a qualitative and quantitative change in hormonal products produced from the endocrine organ. This is exemplified in the increase of pro-inflammatory genes activation (TNF-α, IL-6, etc.), and the reduction of the mRNA and protein insulin-sensitizing molecules (adiponectin) expression. This resulting simultaneous chronic inflammatory and the hypo-adiponectinemia state, are associated with a sustainable decrease in myocyte glucose uptake, low fatty acid β-oxidation, and excessive glucose production. The activated form of the fatty acids, the acyl-COA, can activate a family of kinases (PKC), and through this pathway, participate in the enzymatic phosphorylation process of serine/threonine residues of insulin receptor substrate. This event alters the insulin signaling transduction (PI3 kinase and MAP kinase pathways) and thereby contributes to the development of the insulin resistance phenomena. The metabolic repercussions of such pathological situation exceed the adipocyte territory, to affect the energetic functioning of the myocyte, hepatocyte, and the cellular homeostasis of all peripheral insulin-dependent tissues.
The reversible nature of obesity permits to the hygienic-dietary measures to restore metabolic homeostasis and prevents, at least, in some circumstances, the onset of other obesity-related pathologies. The nutritional component of this approach may be reinforced by the improvement of the organism antioxidant and anti-inflammatory status. This can be achieved through the consumption of vegetables and fruits with a low glycemic index, hydrogen atom transfers abilities, electron donor capacities, and chelate transition metals capabilities. The fruit of PG was proposed as a candidate that responds to these kinds of dietary requirements.
Experimental evidence attributed to PG intake anti-obesity and anti-diabetic capacities (Table 1 and Table 2). According to Vroegrijk et al., [35], diet supplementation with PG-seed oil proved its potent to reduce body weight, body fat mass, and to increase peripheral insulin sensitivity, in C57Bl/J6 mice. These findings reinforce the previous observations made by Lei et al., [36], which found a significant lowering effect on the main lipid parameters, including plasma total cholesterol (TC) concentration, triglycerides (TG) content, and TC/HDL-C ratio. Moreover, enzymes involved in lipid metabolism, such as acyl-CoA oxidase, carnitine palmitoyltransferase-1, as well as nuclear receptors such PPAR-α, have been positively influenced by PG flower constituents [37]. Furthermore, data from Oliveira et al., [38], showed the decrease in body weight after 30 days of feeding with PG extract, without any effect on plasma glucose concentration. However, opposite findings concerning feed intake and weight gain tendencies, have been published by Shabtay and colleagues [39]. Similarly, PG peel extract did not modify body weight gain and did not alleviate inflammation in an animal model of obesity (Balb/c mice) [40]. Nutrigenomic variability, in which individual genetic differences can influence the biological response to phytochemical nutrients, may explain the reported inconsistency.

Table 1.
Summary of the most relevant human and animal findings related to the anti-hyperlipidemic effects of PG consumption.
Table 1.
Summary of the most relevant human and animal findings related to the anti-hyperlipidemic effects of PG consumption.
| Group | Population / disease induction | PG part or product | PG dose and duration | HDL | LDL | TC | VLDL | HDL-C | LDL-C | VLDL-C | TG | Ref |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clinical studies ( the values are expressed as mean ± SD) | ||||||||||||
| Treated group | Hypertensive subjects | PG juice | 150 ml/day, for 2 weeks | ni | ni | 218.73±42.81 a | ni | 49.27±8.06 a | 127.27±24.22 a | ni | 171.18±78.92 a | [41] |
| Placebo group | _ | _ | ni | ni | 187.00±30.27 a | ni | 40.40±6.91 a | 109.40±25.82 a | ni | 165.60±124.32 a | ||
| Treated group | Dyslipidemic patients | PG-seed oil | 800 mg twice daily, for 4 weeks | ni | ni | ni | ni | 1.38±0.44 b | ni | ni | 2.75±1.40 b | [42] |
| Placebo group | _ | _ | ni | ni | ni | ni | 1.25±0.26 b | ni | ni | 3.12±1.59 b | ||
| Treated group | COPD |
PG PG juice |
400 ml daily, for 5 weeks |
55.05 ±12.01 a | 130.48±32.29 a | 209.68±39.10 a | ni | ni | ni | ni | 170.68±187.10 a | [43] |
| Placebo group | _ | _ | 56.75±20.81 a | 116.06±29.14 a | 201.34±32.64 a | ni | ni | ni | ni | 137.91±778.22 a | ||
| Treated group |
Obese patients obesity |
PG juice | 120 ml, for 1 month | ni | ni | 4.7±0.7 b | ni | 1.1±0.1 b | 2.9±0.8 b | ni | 1.3±0.3 b | [44] |
| Placebo group | _ | - | ni | ni | 4.8±0.6 b | ni | 1.2±0.2 b | 2.9±0.5 b | ni | 1.2±0.5 b | ||
| Treated group |
Hyper-cholesterolemic patients |
PGE + Simvastatin |
PGE: 1g/day Simvastatin :20mg/day, for 2 months |
ni | ni | 202±29 a | ni | 45 ± 11 a | 129±15 a | ni | 187±138 a | [45] |
| Placebo group | - | - | i | ni | 192±35 a | ni | 46±12 a | 123±27 a | ni | 110±19 a | ||
| Treated group |
PGE + Simvastatin |
PGE: 1g/day Simvastatin : 20 mg/day, for 1 month |
ni | ni | 208 ± 40 a | ni | 47±13 a | 135±37 a | ni | 129±43 a | ||
| Placebo group | _ | _ | ni | ni | 198 ±55 a | ni | 46 ±19 a | 123±38 a | ni | 144±71 a | ||
| Treated group | Hemodialysis patients | PG Juice |
0.7 mM of polyphenols, 3 times/week, for 1 year |
36.8 ± 10.8 a | 100 ± 33.1 a | 167.3 ± 43.5 a | ni | ni | ni | ni | 167.3 ± 86.3 a | [46] |
| Placebo group | _ | _ | 34.3 ± 15.4 a | 94.3 ± 27.2 a | 165.1 ± 35.8 a | ni | ni | ni | ni | 206.1 ± 109.4 a | ||
| Treated group | CHD patients | PG juice |
240 ml/day, for 3 months |
48±11 a | 91±33 a | 170± 42 a | ni | ni | ni | ni | 149±107 a | [47] |
| Placebo group | _ | _ | 46 ± 12 a | 80±35 a | 157±32 a | ni | ni | ni | ni | 155±102 a | ||
| Treated group | Volunteers at high CVD risk | PG Juice | 500 ml/day, for 4 weeks | ni | ni | 5.45±1.0 b | ni | 1.52±0.44 b | 3.31±0.73 b | ni | 1.147±0.39 b | [48] |
| Placebo group | _ | _ | ni | ni | 4.51±0.51 b | ni | 1.46±0.56 b | 2.54±0.79 b | ni | 1.14±0.51 b | ||
| Animal studies (data is reported as mean ± SEM/SE/SD) | ||||||||||||
| Treated group | by adding 10% of lipid in the basal diet | Hydroethanolic extract of PG | 50 mg/kg/day, for 23 days | 89 ± 11 a | 209 ± 23 a | 87 ± 9 a | ni | ni | ni | ni | 381 ± 23 a | [49] |
| 100 mg/kg/day, for 23 days | 128 ± 5 a | 145 ± 29 a | 82 ± 5 a | ni | ni | ni | ni | 325 ± 43 a | ||||
| 200 mg/kg/day, for 23 days | 179 ± 18 a | 79 ± 8 a | 80 ± 9 a | ni | ni | ni | ni | 302 ± 31 a | ||||
| 300 mg/kg/day, for 23 days | 185 ± 20 a | 61 ± 7 a | 81 ± 7 a | ni | ni | ni | ni | 210 ± 27 a | ||||
| Control + | _ | _ | 98 ±9 a | 92 ±6 a | 73 ±8 a | ni | ni | ni | ni | 146 ± 21 a | ||
| Treated group | Hyper cholestrolemic diet | PG peel powder | (5%) | 38.40±5.18 a | 40.73±1.85 a | 92.33±4.76 a | 13.20±0.69 a | ni | ni | ni | 66±3.46 a | [50] |
| (10%) | 36.93±5.53 a | 46.67±1.97 a | 96.00±4.11 a | 12.40±0.84 a | ni | ni | ni | 62± 3.69 a | ||||
| (15%) | 41.50±5.98 a | 45.77±2.13 a | 97.67±4.52a | 10.40±0.59 a | ni | ni | ni | 52±3.99 a | ||||
|
PG peel extract |
(1%) | 41.40±5.18 a | 16.33±1.85 a | 75.00± 3.66 a | 17.27±0.79 a | ni | ni | ni | 86.33±4.78 a | |||
| (2%) | 42.93±5.53 a | 9.67±1.97 a | 69.00± 3.89 a | 16.40±0.67 a | ni | ni | ni | 82±4.11 a | ||||
| (3%) | 40.50±5.98 a | 12.77±2.13 a | 70.00± 3.92 a | 16.73±0.64 a | ni | ni | ni | 83.67±5.12 a | ||||
| Control + | _ | - | 41.93±5.53 a | 87.53±1.97 a | 154.33±5.13 a | 24.87±0.77 a | ni | ni | ni | 124.33± 3.70 a | ||
| Treated group | High fat diet | PG peel extract | 200 mg/kg, for 56 days | ni | ni | 172.3 ± 3.94 | ni | 40.03 ± 1.03 | 93.84 ± 3.69 | 38.49 ± 0.62 | 192.4 ± 3 | [51] |
| Control + | _ | - | ni | ni | 271.8 ± 3.94 | ni | 29.30 ± 1.03 | 185.3 ± 3.69 | 57.26 ± 0.62 | 285.5 ± 3 | ||
| Treated group |
Intraperitoneal injection of STZ (60 mg/kg) |
PG flowers extract |
250 mg/kg, for 21 dyas | ni | ni | 124.50 ± 8.62 a | ni | 45.17 ± 4.84 a | 61.67 ± 6.12 a | 17.67 ± 4.50 a | 88.17 ± 7.05 a | [52] |
| 500 mg/kg, for 21 days | ni | ni | 118.67 ± 9.60 a | ni | 48.67 ± 5.16 a | 54.67 ± 4.89 a | 14.34 ± 2.95 a | 72.83 ± 6.52 a | ||||
| Control+ | _ | 0 mg/kg, for 21 days | ni | ni | 292.33 ± 4.64 a | ni | 32.83 ± 4.22 a | 234.34 ± 6.12 a | 25.26 ± 0.93 a | 126.33 ± 4.64 a | ||
| Treated group | Intraperitonial injection of STZ (65 mg/kg) | PG leaves | 50 mg/kg, for 28 days | ni | ni | 162.25 ± 5.28 c | ni | 37.79 ± 1.92 c | 106.22 ± 6.14 c | 18.76 ±0.73 c | 93.845 ± 3.66 c | [53] |
| 100 mg/kg, for 28 days | ni | ni | 142.38 ± 2.70 c | ni | 63.32 ± 3.11 c | 80.36 ± 2.08 c | 15.99 ± 1.57 c | 84.53 ± 4.49 c | ||||
| 200 mg/kg, for28 days | ni | ni | 139.45 ± 1.98 c | ni | 44.54 ± 2.97 c | 60.41 ± 3.57 c | 15.71 ± 1.83 c | 76.25 ± 9.96 c | ||||
| Control+ | - | 0 mg/kg, for 28 days | ni | ni | 229.08 ± 7.51 c | ni | 20.47 ± 1.31 c | 179.50 ± 6.68 c | 29.09 ± 0.70 c | 145.46 ± 3.53 c | ||
| Treated group | Intraperitonial injection of poloxamer 407 | PG flowers | 500 mg/Kg, after 15 h | 6.23 ± 0.39 b | 8.56 ± 0.62 b | 16.9 ± 0.60 b | 2.11 ±0.23 b | ni | ni | ni | 10.57 ± 1.17 b | [54] |
| Control+ | _ | _ | 5.04 ± 0.20 b | 9.9 ± 0.67 b | 18.39 ± 0.63 b | 3.38 ±0.08 b | ni | ni | ni | 16.93 ± 0.75 b | ||
| Treated group | PG flowers | 500 mg/Kg, after 24 h | 6.06 ± 0.29 b | 10.74 ± 0.95 b | 19.72 ± 0.67 b | 2.91 ±0.09 b | ni | ni | ni | 14.56 ± 0.46 b | ||
| Control+ | _ | 0 mg/kg | 5.05 ± 0.17 b | 15.7 ± 0.80 b | 24.28 ± 0.89 b | 3.52 ±0.09 b | ni | ni | ni | 17.66 ± 0.46 b | ||
| Treated group | High cholesterol diet | PG juice |
0.2 mL/animal, for 30 days |
78.58±4.79 a | 19.38±10.34 a | 135.83 ± 13.9 b | 37.87±5.36 a | ni | ni | ni | 189.33 ± 26.81 a | [55] |
| Control + | _ | 67.70±2.34 a | 169.93± 31.90 a | 267 ±31.78 a | 29.37±1.18 a | ni | ni | ni | 146.83 ± 5.88 a | |||
| Treated group | High cholesterol diet | PGME | 200 mg, for 30 day | 1.54±0.208 | 0.58 ± 0.118 | 1.93 ± 0.191 | 0.23 ± 0.06 | ni | ni | ni | 1.05 ± 0.17 | [56] |
| 300 mg, for 30 day | 1.29 ± 0.68 | 0.21 ± 0.057 | 1.63 ± 0.125 | 0.10 ±0.028 | ni | ni | ni | 0.91 ± 0.12 | ||||
| 400 mg, for 30 day | 0.91±0.117 | 0.17 ± 0.049 | 1.04 ±0.159 | 0.07 ±0.026 | ni | ni | ni | 0.46 ± 0.15 | ||||
| Control + | _ | 0 mg | 2.16 ±0.150 | 0.68 ± 0.050 | 2.55 ± 0.211 | 0.27 ±0.072 | ni | ni | ni | 1.17 ± 0.13 | ||
Abbreviations. TC: total cholesterol; TG: triglycerides; HDL: high-density lipoprotein; HDL-C: high-density lipoprotein-cholesterol; LDL: low-density lipoprotein; LDL-C: low-density lipoprotein-cholesterol; VLDL: very low-density lipoprotein; VLDL-C: very low-density lipoprotein-cholesterol; ni: not investigated; a: mg/dl; b: mmol/l; COPD: chronic obstructive pulmonary disorder; PGME: PG mesocarp extract. CHD: ischemic coronary heart disease; c: (mg %); CVD: cardiovascular diseases; COPD: chronic obstructive pulmonary disease; STZ: streptozotocin.
8. Type 2 diabetes and pomegranate consumption
Various innate factors (genetic), such as some nucleotide gene polymorphisms or acquired factors (environmental) like food misbehavior, in addition to the in utero epigenome changes (methylation, acetylation, etc.), play a determining role in the pathophysiology of insulin resistance. The interconnectivity of metabolic pathways allows this pathological mechanism to influence, not only the incidence of type 2 diabetes but also the cardiovascular and hepatic diseases (atherosclerosis and non-alcoholic fatty liver disease, respectively). The increase in circulating free fatty acids and serum pro-inflammatory cytokines (TNF-α, IL-6, etc.) which accompanies the abdominal hypertrophy, interfere with insulin-substrate-receptor signaling cascade. Such event disrupts insulin signal transduction, and thereby its anabolic effects. This is reflected on the one hand, by the decline in peripheral glucose uptake, and glycogen synthesis. On the other hand, by the enhancement of free fatty acids releases and hepatic glucose production improvement. Faced with the resulting hyperglycemia, the islets of Langerhans try to compensate the growing insulin demand through the hyperinsulinism phenomena. This temptation restores and maintains, at least temporarily, the cellular energetic equilibrium. However, the persistence of the insulin resistance process leads progressively to the exhaustion of the β-insular function. Then, the resulting glucolipotoxicity state pushes the transition from a non-pathological situation of glucose intolerance to a potentially deleterious type 2 diabetes.
To prevent, or at least, to attenuate such fatal consequences, natural molecules with an aromatic nucleus and hydroxyl groups, should be envisaged, particularly in light of associated anti-diabetic medication toxic effects (as discussed earlier). PG-polyphenols have been well documented in the folk medicine literature to have anti-diabetic effects. In fact, this plant may act through numerous mechanisms, including PPAR-γ activity modulation [37,57,58,59,60], resistin protein degradation [61], adiponectin gene expression [62], α-glucosidase enzymatic activities inhibition [63,64], Glut-4 mRNA expression [59], and β-mass regeneration [65].
In this context, Parmar et al., [63], reported that short-term treatment with PG peel extract, may reduce α-amylase activity, serum glucose concentration, and lipid peroxidation content. The achieved hypoglycemic effects appear to be associated with the increase in insulin secretion capacities, as this hormone was found in plasma at high levels compared to the control group. These results are consistent with those published by Li et al., [64]. This paper demonstrated the ability of PG flowers to normalize postprandial hyperglycemia and to inhibit the catalytic activity of the α-amylase in a dose-dependent manner. Additionally, Vroegrijk and coworkers [35], proved that PG-seed oil treatment can improve peripheral insulin sensitivity in a C57Bl/J6 mice. Furthermore, α-glucosidase and α-amylase assays show the capacity of PG leaves extract to inhibit these enzymes. Enzymatic inhibition of pancreatic lipase enzyme was also reported [66]. However, contradictory findings related to the anti-hyperglycemic potential of PG have been also published [40,67]. These papers did not suggest any significant change after PG treatment. Also, fasting blood glucose and pancreatic β-mass were not improved.
PG preparations and pure PG-related constituents have been also investigated for their metabolic effects. These include, juice [68], punic acid [69], EA [70], punicalagin [71] and catalpic acid [72]. Outcomes from these studies indicate that these molecules exert numerous antidiabetic and biological effects.
PPAR-γ is a subtype nuclear receptor which is predominantly expressed in the adipose tissue [73,74]. Its capability to exert physiological functions, such as adipocyte differentiation [74], lipid accumulation [75], lipoprotein lipase mRNA expression [76], proinflammatory gene repression and resistin gene downregulation [77], have made this receptor a privileged target for anti-diabetic drugs. The genes under PPAR-γ control are thought to be incriminated in the development of insulin resistance and type 2 diabetes progression.
Phenolic molecules from PG may modulate PPAR-γ activities [37,57,58,59,60]. The observed antidiabetic effects PPAR-γ related activation is believed to be produced in two different ways depending on the ligand molecule. The first one is through PPAR-γ selective agonist actions (without any activation of the associated PPAR-γ adipogenic factors (DRIP205/TRAP220)), which terminate by an improvement of insulin sensitivity [57]. This proved a solution to weight gain frequently observed with the full activator’s antidiabetic pharmacological drugs. The second pathway implies a different mechanism and consists of inhibiting PPAR-γ activation to prevent the associated undesirable adipogenesis effects. Authors, attributed this activity to quercetin, a flavonoid found in PG [78,79]. Consequently, PG appears to exert a dual function, in which the agonist PPAR-γ activities appear to be weak, in comparison to the PPAR-γ antagonistic actions [57]. In this view, Mueller and Jungbauer [57], showed that PG fruit extract (standardized on 40% of EA) was the highest PPAR-γ antagonist among fifty extracts positively screened for their PPAR-γ antagonism activity. The findings related to this kind of modulation are in agreement with the reported in vitro [37,58,59] and in vivo [37,59,60] studies. Therefore, natural selective PPAR-γ agonists or in contrast PPAR-γ antagonists should be targeted to replace the thiazolidinedione’s utilization and to reduce adipose tissue hypertrophy.
Clinical studies performed in this field are mainly carried out using PG juice and their outcomes are largely inconsistent. In this regard, although Parsaeyan et al., [80], have demonstrated that daily supplementation of 200 ml of PG juice for six weeks, can reduce fasting blood sugar, unexpected findings suggest that PG juice did not modify the insulin secretory performances or hormone sensitivity [44]. This suggests the fact that the supplement may contain insulin-like compounds and can act through non-insulin mechanisms, especially as it was able to alleviate inflammatory response. Furthermore, arguments with statistical power comes from a systematic literature review and meta-analysis of seven trials to indicate the fact that PG intake did not show any significant effects on glycemic markers [81].
In all, data from pure compounds seems to be consistent, while outcomes from extracts or products both on animal and human studies are conflicting. This can be explained by the probable interactions that could occur between different PG-bioactive substances that contain each extract/product, and which may potentiate and/or attenuate the cellular effect and the biological response.
Figure 1.
The Anti-hyperlipidemic and normoglycaemic molecular mechanisms of PG phytochemicals active compounds. As summarized, PG nutrients are able to reduce resistin protein, α-glucosidase, and redox-sensitive gene expression, as well as, to increase adiponectinemia and to ameliorate cellular glucose uptake. Inhibitory effects against enzymes involved in lipid metabolism, including carnitine palmitoyltransferase I, acyl-coenzyme A oxidase, and lipoprotein lipase, are also proposed. Furthermore, secondary metabolites from PG organisms could improve blood pressure, inflammatory state, and the antioxidant enzymatic capabilities as well as to modulate PPAR-γ activities. Abbreviations. GILM: genes involved in lipid metabolism; CPT1: carnitine palmitoyltransferase 1; ACO: acyl-coenzyme A oxidase; LPL: lipoprotein lipase; redox SG: redox-sensitive genes; GAA: the gene that codes for α-glucosidase; AOX genes: antioxidant genes; CAT: catalase; GSH: glutathione; GLUT-4: glucose transporter-4; ADIPOQ: adiponectin gene; NF-κB: nuclear factor-kappa B; PPAR-γ : peroxisome proliferator-activated receptor gamma.
Figure 1.
The Anti-hyperlipidemic and normoglycaemic molecular mechanisms of PG phytochemicals active compounds. As summarized, PG nutrients are able to reduce resistin protein, α-glucosidase, and redox-sensitive gene expression, as well as, to increase adiponectinemia and to ameliorate cellular glucose uptake. Inhibitory effects against enzymes involved in lipid metabolism, including carnitine palmitoyltransferase I, acyl-coenzyme A oxidase, and lipoprotein lipase, are also proposed. Furthermore, secondary metabolites from PG organisms could improve blood pressure, inflammatory state, and the antioxidant enzymatic capabilities as well as to modulate PPAR-γ activities. Abbreviations. GILM: genes involved in lipid metabolism; CPT1: carnitine palmitoyltransferase 1; ACO: acyl-coenzyme A oxidase; LPL: lipoprotein lipase; redox SG: redox-sensitive genes; GAA: the gene that codes for α-glucosidase; AOX genes: antioxidant genes; CAT: catalase; GSH: glutathione; GLUT-4: glucose transporter-4; ADIPOQ: adiponectin gene; NF-κB: nuclear factor-kappa B; PPAR-γ : peroxisome proliferator-activated receptor gamma.
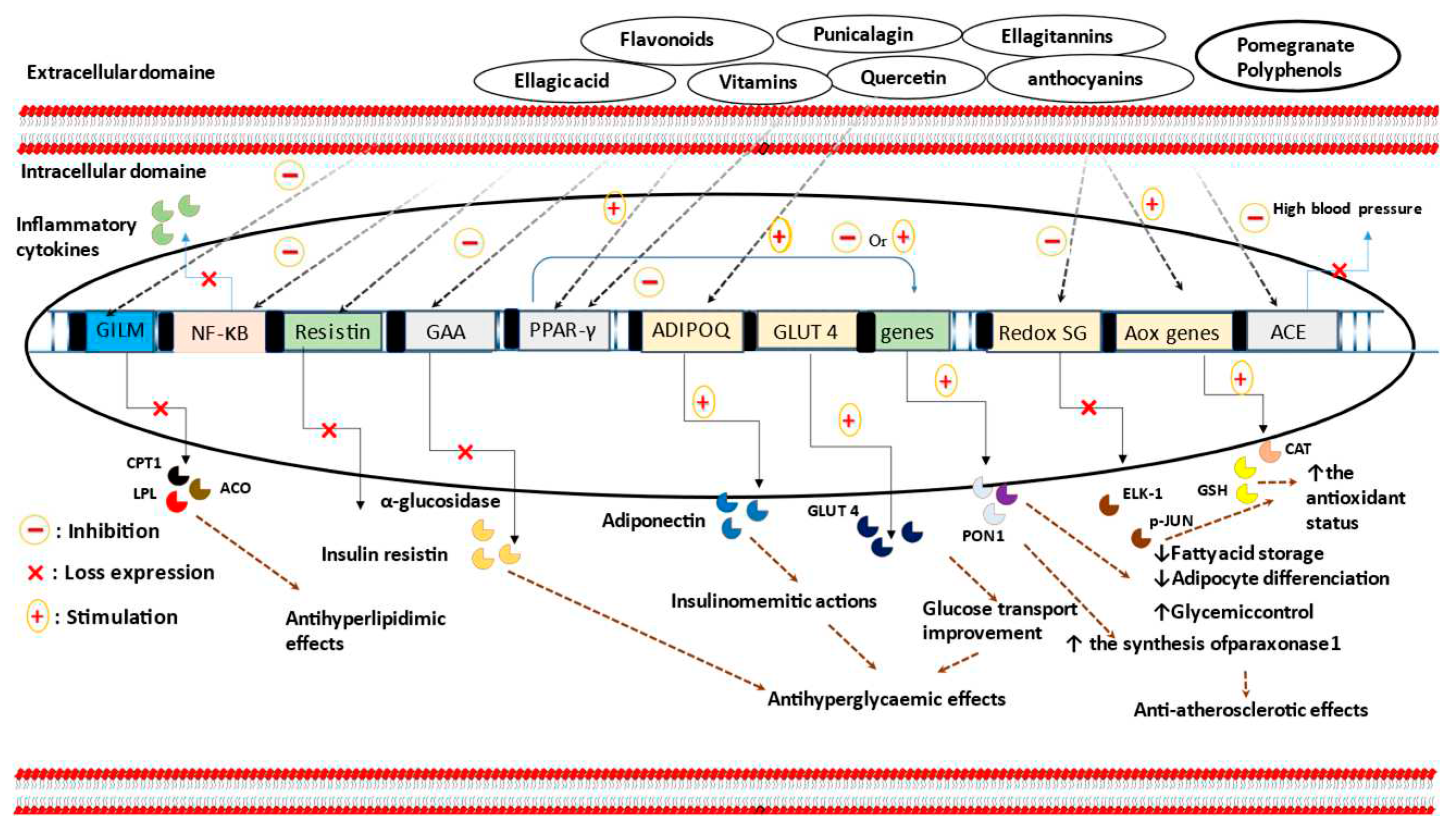
Table 2.
Summary of the main effects of PG intake on type 2 diabetes, obesity and cardiovascular diseases.
Table 2.
Summary of the main effects of PG intake on type 2 diabetes, obesity and cardiovascular diseases.
| Animal model/ population/ cell line |
Disease or induction of the disease | PG part or product | Dose and period of treatment | Findings | Ref |
|---|---|---|---|---|---|
| PG intake effect on obesity and diabetes mellitus | |||||
| Male C57Bl/J6 mice | High-fat diet | PG-seed oil | 1%, for 12 weeks | ↓ body weight; ↓ body fat mass; ↔ liver insulin sensitivity; ↑ peripheral insulin sensitivity; ↔ food intake; ↔ energy expenditure. | [35] |
| Male CD-1 mice | High-fat diet | PG-seed oil | 61.79 mg/day, for 14 weeks | ↓ weight gain; ↓ body weight; ↓ absolute weight gain; ↓ percentage of weight gain; ↔ lean mass; ↔ cholesterol profile; ↓ leptin; ↑ adiponectin. | [62] |
| Zucker diabetic fatty rats | Genetic manipulation |
PG flower extract |
500 mg/kg/day, for 6-weeks | ↓ plasma glucose; ↔ fasting serum glucose; ↑ cardiac PPAR-γ mRNA expression; ↑ GLUT-4 mRNA; ↑ mRNA expression of inhibitor-kBα. |
[59] |
| Zucker lean rats | - | ↔ plasma glucose; ↔ fasting plasma glucose. | |||
| Human THP-1-derived macrophage cells | - | 50µg/ml, for 48 h | ↑ PPAR-γ gene expression; ↑ PPAR-γ-dependent mRNA expression. ↑ lipoprotein lipase activity. |
||
| Swiss albino male mice | Alloxan injection | PG peel extract | 200 mg/kg/day, for 10 days | ↓ plasma glucose; ↓ α-amylase activity; ↓ water consumption; ↓ lipid peroxidation; ↑ plasma insulin. | [63] |
| Male albino rats | Alloxan injection | PG peel aqueous extract | 0.43g/kg BW, for 4-weeks | ↓ blood glucose; ↑ insulin level; ↑ β-cells regeneration. | [65] |
| Male Sprague Dawley rats | Alloxan injection | PG seed | 60 g/ kg/day, for 15 days | ↔ serum glucose; ↔ fasting blood glucose; ↔ size of islets; ↔ islets density; ↔ percent of β-cells in each islet; ↔ number of islets. |
[67] |
| albino rats | Alloxan injection | PG flower extract | 300 or 400or 500 mg/kg. Sampling time: at 1 and 2 h. |
↓ blood glucose. |
[82] |
| Zucker diabetic fatty rats | Genetic manipulation |
PG flower extract |
250,500, and 1000 mg/kg/day, for 2 weeks. 200 µl, for 5 min for the in vitro assay. |
↓ postprandial hyperglycemia; ↓ α-glucosidase activity (IC50: 1.8 µg/ml); ↓ plasma glucose levels after sucrose loading. |
[64] |
| Adult albino rats | Streptozotocin treatment | PG seed extract | 150, 300 and 600 mg/kg, for 2, 4, 8 and 12 h |
↓ blood glucose in time and dose-dependent manner. | [83] |
| 3T3-L1 pre-adipocytes |
- |
Punicic acid | 1.25, 2.5, 5 and 10 µM | ↑ PPAR-α and γ activity; ↓ fasting plasma glucose; ↑ glucose normalizing capabilities; ↓ NF-κB activation; ↓ TNF-α expression. | [84] |
| ovariectomized mice | Surgical intervention |
PG fruit extract |
30 mg/kg/day, for 12 weeks | ↓ serum resistin concentrations. |
[61] |
| 3T3-L1 adipocytes | - | 50 and 100 µg/mL, for 9 and 12 h | ↓ resistin protein secretion; ↔ resistin mRNA expression; ↑ intracellular resistin degradation; ↔ adiponectin secretion. | ||
| Ellagic acid | 20, 40, and 70 µM, for 12h | ↓ resistin protein secretion; ↔ adiponectin secretion; ↓ intracellular resistin time-dependently. | |||
| Punicic acid | 5 and 10 µM, for 9h | ↔ resistin molecule secretion. | |||
| C57BL/6J obese mice | High-fat diet | Catalpic acid | 1g/100g, for 78 days | ↓ insulin; ↓ fasting blood glucose; ↑ glucose normalizing ability; ↓ abdominal white adipose tissue storage; ↑ PPAR-α expression; ↑ HDL-C; ↓ TG. |
[72] |
| male and female ICR mice | High-fat diet | PG leaf extract | 400 or 800 mg/kg daily, for 5 weeks | ↓ body weight; ↓ energy intake; ↓ TC; ↓ TG; ↓ TC/HDL-C ratio; ↓ glucose; ↓ fat absorption; ↓ appetite. |
[36] |
| Zucker diabetic fatty rats | Genetic manipulation | PG flower extract | 500 mg/ kg daily, for 6 weeks | ↓ TG; ↓ TC; ↓ fatty acids; ↓ fatty acids transport proteins; ↓ PPAR-; ↓ acyl-CoA oxidase; ↓ 5 -AMP-activated protein kinase-α-2; ↓ carnitine palmitoyltransferase-1; ↓ acetyl-CoA carboxylase mRNA. |
[85] |
| Zucker diabetic fatty rats |
Genetic manipulation |
PG flower extract |
500 mg/kg/day, for 6 weeks | ↓ TG; ↓ lipid droplets; ↑ PPAR-α; ↑ acyl-CoA oxidase; ↑ carnitine palmitoyltransferase-1; ↓ gene expression of stearoyl-CoA desaturase-1; ↔ fatty acids and TG synthesis; ↔ fatty acids and TG hydrolysis; ↔ fatty acids and TG uptake. |
[37] |
| HepG2 cell line | - | 10, 50 and 100 µg/ml, for 48h | ↑ PPAR-α; ↑ Acyl-CoA oxidase mRNA. | ||
| Type 2 diabetic and hyperlipidemic patients | Diabetes mellitus and hyperlipidemia | PG juice | 40 g/day of concentrated PG juice, for 8 weeks | ↓ TC; ↓ LDL-C; ↓ LDL-C/HDL-C; ↓ TC/HDL-C; ↔TG; ↔ HDL-C. |
[86] |
|
Calves |
- |
Polyphenols PG extract | 5 or 10 g/day (0,15, and 30 mg of gallic acid equiv/kg/day), for 70 days | ↔ on body weight or intake, during the first 30 postnatal days, but are ↓ after this period; ↔ glucose concentration; ↔ 3-hydroxybutyrate; ↓ fat digestion; ↔ dry matter; ↔ starch digestibility; ↔ organic matter. |
[38] |
| Calves | - | PG peel | Ad libitum, for 2 months | ↑ feed intake; ↑ weight gain tendency. | [39] |
| Balb/c mice | High-fat feeding | PG peel extract | 0.2% (6 mg/day/mouse), for 4 weeks | ↔ weight gain; ↔ glycaemia; ↔ glucose tolerance; ↓ TC; ↓ LDL-C; ↔ IL-1β, IL-6 and COX-2 in the liver; ↓ IL-1β, IL-6 and COX-2 both in in the gastrointestinal tract and visceral adipose tissue. |
[40] |
|
Male wistar rats |
High lipid diet | PG peel extract | 50, 100, 200, and 300mg/kg, for 23 days | ↓ body weight; ↓ TC; ↓ LDL-C; ↓ alkaline phosphatase; ↓ TG; ↑ HDL-C; ↓ AST; ↓ ALT. |
[49] |
| Pomegranate intake and cardiovascular diseases | |||||
| J774.A1 macrophages |
- |
PG juice | 75 mmol/L, for 90 min | ↑ Ox-LDL degradation by 40%; ↔ on macrophage degradation of native LDL; ↔ macrophage cholesterol efflux capacities; ↓ macrophage cholesterol biosynthesis (by 50%). | [87] |
|
Human coronary artery endothelial cells |
High shear stress | PG juice | 7–14 µl of PG juice, for 24h. | ↓ the activation of redox-sensitive genes (ELK-1 and p-JUN); ↑ eNOS expression. |
[88] |
| Low-density-lipoprotein receptor-deficient mice (LDLR_/_ mice) | Genetic manipulation and high-cholesterol diet |
PG juice |
31 µl/day (0.875 µmol of total polyphenols), for 24 weeks | ↓ the activation of redox-sensitive genes (ELK-1 and p-JUN); ↑ eNOS expression; ↓ the progression of atherosclerosis lesions in mice. | [88] |
| Carotid artery stenosis subjects | carotid artery stenosis |
PG juice |
50 ml, for 1 or 3 years |
↓ carotid intima-media thickness; ↑ PON 1 activity; ↓ LDL basal oxidative state; ↓ LDL susceptibility to oxidation; ↓ antibodies against ox-LDL; ↑ total antioxidant status; ↓ antibodies against oxidized LDL; ↓ systolic blood pressure. |
[89] |
| Apolipoprotein E-deficient mice | Genetic manipulation | PG juice | 31 µL of PJ/d (0.875 mmol of total polyphenols/d ), for 2 months | ↑ PON1 activity; ↓ MPM lipid peroxide; ↓ Ox-LDL MPM uptake; ↓ MPM cholesterol esterification; ↑ macrophage cholesterol efflux; ↓ macrophage Ox-LDL uptake; ↓ cholesterol esterification; ↓ atherosclerosis lesions size. |
[90] |
| Apolipoprotein E-deficient mice | Genetic manipulation |
PG byproduct |
17 or 51.5 µg of gallic acid equiv/kg/day, for 3 months | ↓ atherosclerotic lesion size; ↓ cellular lipid peroxide; ↓ glutathione levels; ↑ POM-2 lactonase activity; ↓ Ox-LDL uptake. |
[91] |
| J774A.1 macrophage | - | 10 or 50 µmol/L of total polyphenols, for 18 h | ↓ cellular total peroxide; ↓ Ox-LDL uptake. | ||
| J774A.1 macrophage | - | PG juice | 10–50 µM of total polyphenols, for 18 h | ↑ expression and enzymatic activity of PON-2; ↑ PPAR-γ and AP-1 activation; ↓ macrophage oxidative status; ↓ Ox-LDL uptake. | [58] |
| Wistar albino rats |
High-fat diet |
PG peel extract | 50 or 100 mg/kg, for 6 weeks |
↓ TC; ↓ LDL-C; ↓ VLDL-C; ↓ TAGs; ↑ HDL-C ; ↑ GR; ↑ SOD; ↑ CAT; ↑ GSH; ↓ MDA; ↑ PON-1 activities; ↓ LDH activity; ↑ TNF-α; ↑ CD36. |
[92] |
| Ellagic acid | 1mg/kg, for 6 weeks | ||||
| Punicalagin | 7mg/kg, for 6 weeks | ||||
| Zucker diabetic fatty rats | Genetic manipulation |
PGF extract |
500 mg/kg, for 6 weeks |
↓ cardiac fibronectin expression; ↓ collagen I and III mRNAs; ↓ expression of endothelin -1; ↓ endothelin receptor a; ↓ c-jun and inhibitor-kBβ expression; ↑ inhibitor-kBα; ↓ LPS-induced NF-kB activation. | [93] |
Abbreviations. ↔: no effect; ↓: decrease; ↑: increase; TC: total cholesterol; HDL-C: high-density lipoprotein cholesterol; LDL-C: low-density lipoprotein cholesterol; TG: triglycerides; VLDL: very low-density lipoprotein; VLDL-C: very low-density lipoprotein cholesterol; TAGs: triacylglycerols; ox-LDL: oxidized LDL; LDLR_/_: low-density-lipoprotein receptor-deficient; CAS: atherosclerotic patients with carotid artery stenosis; MPM: mice peritoneal macrophages; COX-2: cyclooxygenase-2; TGCOX-2: transgenic mice COX-2; PPAR: peroxisome proliferator-activated receptor; NF-κB: nuclear factor-kappa B; LPS: lipopolysaccharide; PON: paraoxonase; eNOS: endothelial nitric oxide synthase; TNF-α: tumor necrosis factor α; ALT: alanine transaminase; AST: aspartate transaminase; GLUT: glucose transporter; GSH: glutathione; SOD: superoxyde dismutase; GR: glutathione reductase; CAT: catalase; CD36; cluster of differentiation 36; MDA: malondialdehyde; AP1: activator protein 1.
9. Atheroprotective activities and antidyslipidemic effects of pomegranate consumption
Vascular cell dysfunction and arterial endothelial tissue activation are thought to be the first events in the atherogenic process. The increase in endothelial permeability observed in such circumstances permits to LDL and LDL-C to migrate from the vascular compartment to the intima domain. In this area, these lipoproteins establish molecular interactions with matrix proteoglycans using their Apo-B100, and then will be subjected to post-translational biochemical modifications, in particular oxidative modifications. The local endothelial hypertrophy that occurs because of this non-physiological accumulation stimulates through chemotactic signalling pathways the recruitment of macrophages; cells specialized in the clearance of cellular debris. In parallel, complex phenomena, involving a decrease of reverse cholesterol efflux capacity, migration and proliferation of smooth muscle cells, activation of metalloproteases, and installation of a local pro-inflammatory secretory profile, act in a concomitant manner towards the fragilization of the arterial wall and conditioning the anatomical development and clinical expression of atherosclerosis. These series of deleterious events is thought to be associated with dyslipidemia, visceral obesity, and diabetes mellitus.
These medical conditions are considered majors modifiable risk factors for atherosclerosis. Therefore, safe strategies aimed at correcting atherosclerosis-related metabolic abnormalities, are of paramount importance as they could restore vascular homeostasis. This may be achieved through polymedication and lifestyle amelioration. To ensure this goal, functional food containing vitamins, phenols, and polyphenols compounds, may be added to this global approach. In this regard, an extensive amount of data has been raised to support the anti-atherogenicity potent of PG-diet intake.
Poly-molecular substances from PG exhibit a range of beneficial biological activities in the vascular microenvironment and may act on a series of risk factors that impair the endothelial anatomy and function. Nutrients isolated from PG, such as ellagic acid can suppress pro-matrix metalloproteinases and prevent their interaction with the imprisoned arterial intima LDL molecules [94]. This fact, contribute to inhibit one of the first events involved in the pathogenesis of atherosclerosis. Moreover, punicalagin compound, gallic acid and its derivatives are able to stimulate and to promoter paraoxonase-1 activity in a HuH7 hepatocyte cells [95]. The preventive atherosclerosis effects related to PG consumption, may be also achieved through eNOS expression improvement [88], and mRNA paraoxonase-1 transcription [89,90,95].
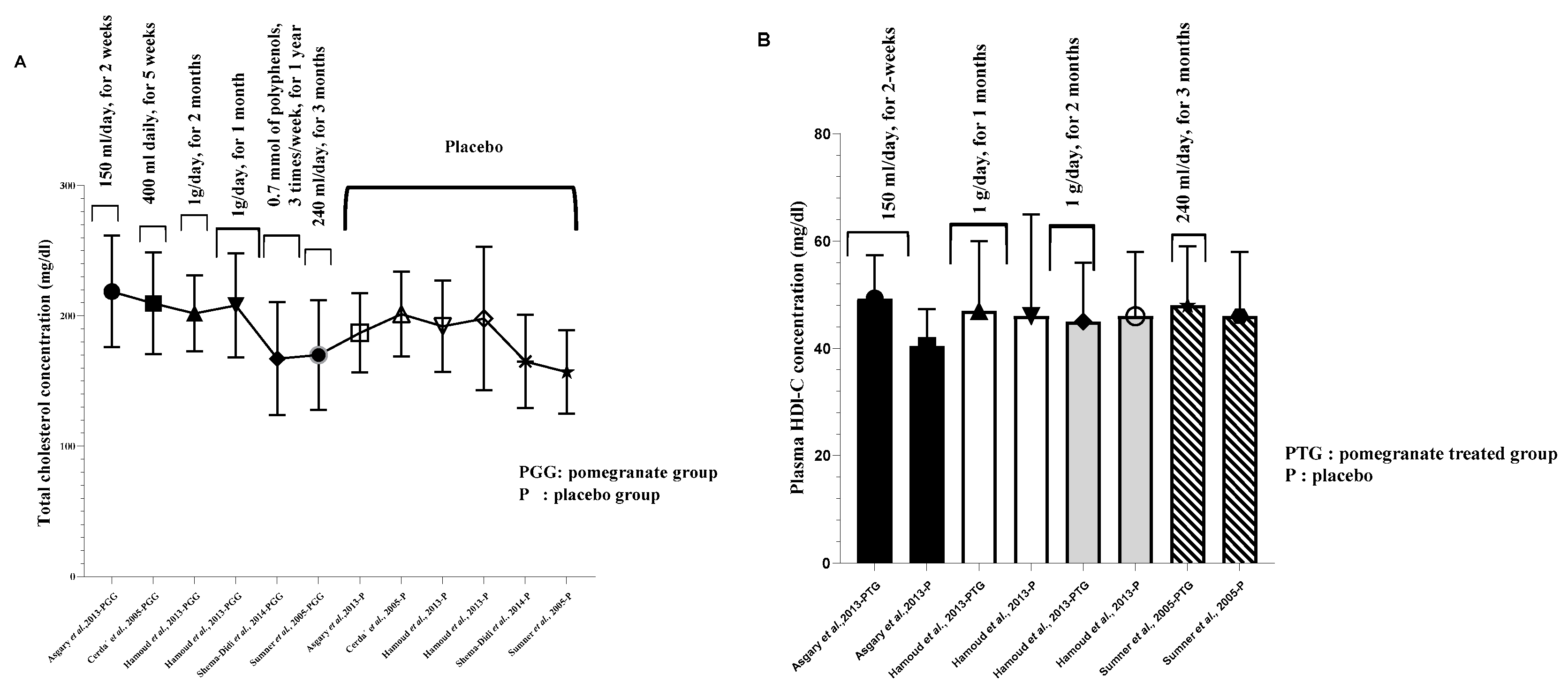
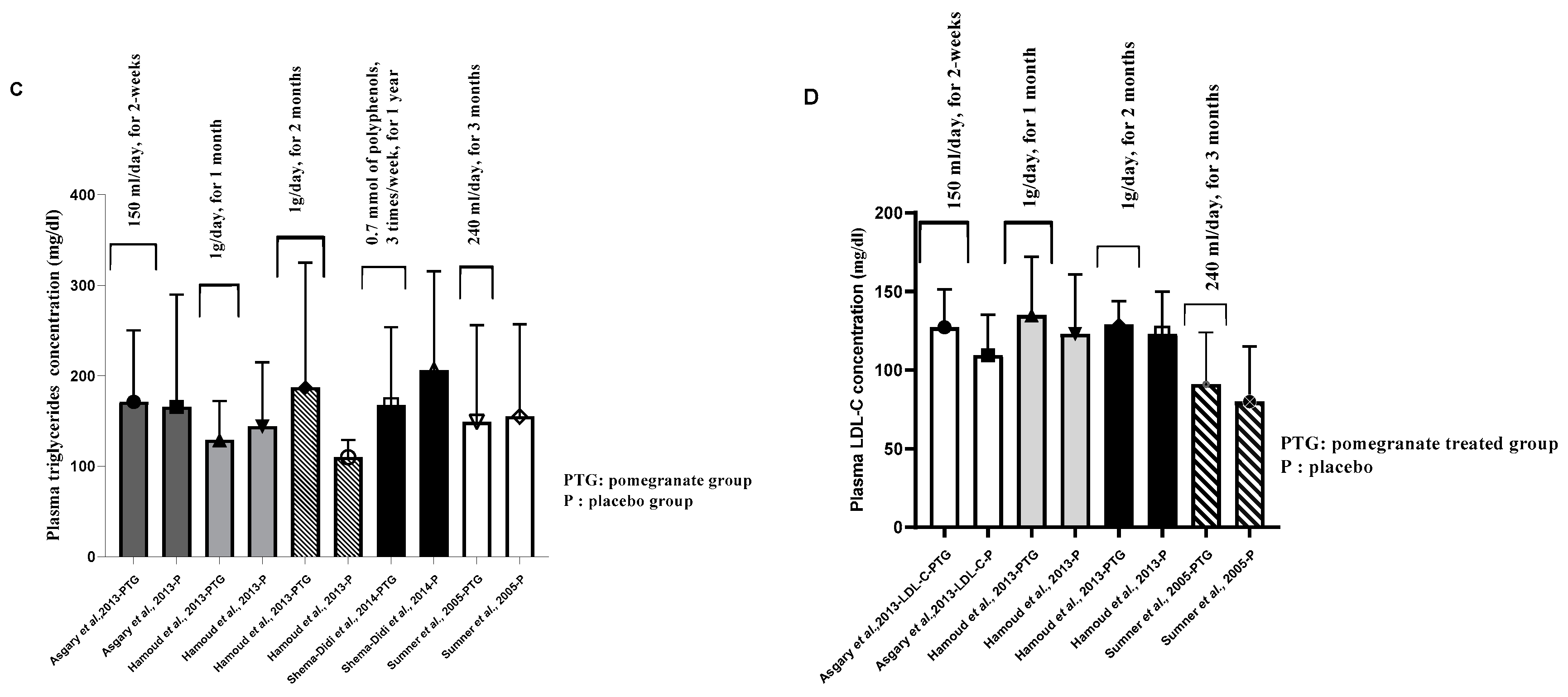
Figure 2.
Pomegranate bioeffects on human lipid parameters. A. Pomegranate consumption effect on total cholesterol. B. Pomegranate impac on plasma HDL-C concentration. C. Pomegranate intake and plasma triglycerides concentration. D. The modulatory effect of pomegranate supplementation on plasma LDL-cholesterol level. All values are reported as mean ± SD.
Figure 2.
Pomegranate bioeffects on human lipid parameters. A. Pomegranate consumption effect on total cholesterol. B. Pomegranate impac on plasma HDL-C concentration. C. Pomegranate intake and plasma triglycerides concentration. D. The modulatory effect of pomegranate supplementation on plasma LDL-cholesterol level. All values are reported as mean ± SD.
Pomegranate juice supplementation was also tested for possible atheroprotective effects. According to Burgermeister et al., PG juice products may modulate paraoxonase-2 (PON 2) activities in a PPAR-γ depending pathways [96], and one year of supplementation, may be effective in increasing plasma paraoxonase activity and serum total antioxidant status by 83% and 130%, respectively [89]. PG juice treatment could also improve lactonase activity and paraoxonase enzyme stability, as well as HDL-associated paraoxonase arylesterase form, in comparison to the unsupplemented group [80,97]. This last claim is of fundamental importance because enzymatic stability and biological activities of PON 1 improve when this enzyme is bound to HDL. Additionally, this preparation modulates paraoxonase-2 activity in a PPAR-γ-related signaling cascades [96]. In in vivo experiment using apolipoprotein E-deficient mice, showed the fact that this product when it is added to diet, may increase macrophage cholesterol efflux and protect LDL against oxidative alterations [90], an initiating event and a crucial step in the atherosclerosis development. Bioprotective effects illustrated in attenuating LDL oxidation, macrophage cholesterol biosynthesis reduction [87], and redox-sensitive gene inactivation [88].
Arterial protection mediated by PG substances may also occur through non-enzymatic mechanisms, involving a significant reduction of VCAM-1, ICAM-1, and E-selectin molecules [41]. Furthermore, TG/HDL-C ratio, a predictor element for cardiovascular diseases [98], has been observed to be significantly reduced after PG-seed oil treatment [99]. These observations and others suggest cardiomyocyte and coronary artery protection of PG supplementation. However, effect on reverse cholesterol efflux are improbable, as investigators [41,48,80,86,100] did not find any significant amelioration of serum HDL-C concentration (Figure 3) and apolipoprotein-A level, compared to the placebo group.
10. Antidyslipidemic effects of pomegranate and pomegranate products
Hypercholesterolemia and hypertriglyceridemia states appear to be independent risk factors for chronic diseases, like diabetes mellitus and atherosclerosis [101]. According to Soliman et al., [92], PG peel and PG-ETs polyphenols were significantly effective in reducing hyperlipidemia (TC, LDL-C, VLDL-C, and TAGs), as well as in improving plasma HDL-C concentrations in a Balb/c mice. Investigators have evaluated this potent in human subjects [48,80,86]. In fact, Esmaillzadeh and colleagues [86], demonstrated that daily consumption of concentrated PG juice for 8 weeks, may regulate lipid parameters in type 2 diabetes and dyslipidemic population. This regulation is exemplified mainly in the decrease of serum TC and LDL-C concentrations, as well as TC/HDL-C and LDL-C/HDL-C ratio. Similar results have been reported in Parsaeyan’s study [80], in which short-term treatment (6 weeks) with 200 ml of PG juice, resulted in a diminution of plasma TC and serum LDL-C levels. Identical results have been obtained in a medium-term supplementation (eight weeks), exemplified in a significant reduction in plasma TC, LDL-C, LDL-C/HDL-C, and TC/HDL-C ratio [100]. However, contradictory outcomes are also published, and did not suggest any changes in lipid profile after PG intervention [48].
The lack of long-term supplementation, and the low number of participants, are the main limitations of the reported clinical data. The last issue has been overcome by a fixed-effect meta-analysis, which included 12 studies (a total of 545 individuals) [102]. Unfortunately, it did not suggest any significant effect of PG intake or PG-derived products on serum lipid profile.
11. Insulin resistance, blood pressure, and pomegrnate consumption
The reduction of nitric oxide molecules, insulin resistance, oxidative stress, obesity, type 2 diabetes and its vascular complications, are thought to be the major leading causes of vascular dysfunction and cardiovascular abnormalities development. The vasodilator effect and tubular sodium reabsorption performances mediated by insulin hormones are found to be altered in such pathological conditions. This is aggravated by the vasoconstrictive effect exercised by misused free fatty acids, contributing to blood pressure abnormalities.
In vitro and in vivo evidence [103,104,105,106,107], have been sufficiently accumulated to push many research teams to investigate the antihypertensive properties of PG in human conditions [41,46,47,48,89,108,109]. Outcomes from these clinical interventions are in favour of an improvement of vascular function, lipid metabolism homeostasis maintaining, and blood pressure normalization. A systematic review and meta-analysis performed by Sahebkar and coworkers [110], come to a conclusion confirming these health-promoting capacities. Authors suggest, a significant beneficial impact of PG juice supplementation on both diastolic and systolic blood pressure, for doses ranging from 150 ml to 500 ml per day, and for periods between 4 weeks and 18 months. From a mechanistic point of view, inhibition of angiotensin-converting enzymatic activities [104,111], and tubular degeneration protection [105], are among the proposed operative actions of PG biochemical components. However, the included studies have limitations, regarding the number of subjects recruited [47,89], lack of certain anthropometric measurements (age, gender, BMI) [46,47,89], and non-investigation of the main lipid parameters (LDL-C, HDL-C, TC, and TG) [47,89,108]. Concerns which could provide a more or less broad view of the health status of subjects enrolled, and thereby help to make a reliable interpretation of the obtained results.
12. Conclusion
The expansion of obesity prevalence influences the incidence of obesity-related pathologies, such as non-alcoholic fatty liver disease, diabetes mellitus, and cardiovascular abnormalities. Therefore, taking measures aimed at reducing and/or limiting the progression of obesity and its pathological consequences, is of crucial importance and should be a public health priority. As we reviewed, a considerable amount of research has been published and are largely conflicting. The reported inconsistency is associated, on the one hand, with the variability of the plant part used (as there is an asymmetric distribution of the bioactive compounds), cultivar,
geographical area and its related bioclimatic and edaphic characteristics. On the other hand, even if we achieve to standardize the fruit, results are sensitive to other factors related to the in vivo conditions, mainly plasma bioavailability, organ accessibility and nutrigenomics considerations. All in all, there are substantial arguments that support the medicinal value of PG and PG-active substances against metabolic syndrome components. Effective management of this group of metabolic abnormalities implies the adoption of global therapeutics strategies, in which hygienic-dietary recommendations, and lifestyle modification, should be take a central place.
Author Contributions
Conceptualization: M.A. and H.B.; methodology: M.A., H.B., and S.B.; software: M.A., M.R., and H.B.; writing—original draft preparation: M.A., K.B, H.B., and SB; writing—review and editing: M.A., H.B, A.K and T.F.; visualization: A.K, M.R and T.F.; supervision: H.B and S.B.; project administration: H.B.; funding acquisition: H.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by (Agence Nationale des Plantes Médicinales et Aromatiques; Centre National de Recherche Scientifique et Technique and Université Sultan Moulay Slimane), (project number: 3/2021-348/20. Morocco (H.B.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization: “Obesity and overweight,” 2021.
- S. Webber, International Diabetes Federation, vol. 102, no. 2. 2013.
- F. Meuleneire, Management of diabetic foot ulcers using dressings with Safetac®: A review of case studies, vol. 4, no. 4. 2008.
- C. Bommer et al., “The global economic burden of diabetes in adults aged 20–79 years: a cost-of-illness study,” Lancet Diabetes Endocrinol., vol. 5, no. 6, pp. 423–430, 2017. [CrossRef]
- H. Dysfunction, “Associated with Troglitazone,” New Engl. J. Med. Corresp., vol. 338, no. 13, pp. 916–917, 1998.
- A. Muthukumar, M. Sridevi, R. Gayathri, S. Muthukumar, S. R. M. S, and A. Muthukumar, “A Pharmacovigilance Study of antidiabetic Drugs in Hypertensive Patients in a Territory Hospital in Coimbatore Zone,” Sch. Reasearch Libr., vol. 9, no. 8, pp. 70–76, 2017.
- C.J. Rosen, “Revisiting the Rosiglitazone Story — Lessons Learned,” N. Engl. J. Med., vol. 363, no. 9, pp. 803–806, 2010. [CrossRef]
- G. Viberti et al., “A Diabetes Outcome Progression Trial (ADOPT),” Health Care (Don. Mills)., vol. 25, no. 10, pp. 1737–1743, 2002.
- S. E. Kahn et al., “Glycemic Durability of Rosiglitazone, Metformin, or Glyburide Monotherapy,” N. Engl. J. Med., vol. 355, no. 23, pp. 2427–2443, 2006. [CrossRef]
- A.D. Wright, C. A. Cull, K. M. Macleod, and R. R. Holman, “Hypoglycemia in Type 2 diabetic patients randomized to and maintained on monotherapy with diet, sulfonylurea, metformin, or insulin for 6 years from diagnosis: UKPDS73,” J. Diabetes Complications, vol. 20, no. 6, pp. 395–401, 2006. [CrossRef]
- A.N. Jacob, K. Salinas, B. Adams-Huet, and P. Raskin, “Weight gain in type 2 diabetes mellitus,” Diabetes, Obes. Metab., vol. 9, no. 3, pp. 386–393, 2007. [CrossRef]
- P. Raskin, M. RENDELL, M. C. RIDDLE, and J. F. DOLE, “A Randomized Trial of Rosiglitazone,” 2001.
- S. R. et al Mehta, “Effect of Rosiglitazone on the Risk of Myocardial Infarction and Death from Cardiovascular Causes.,” N. Engl. J. Med., pp. 687–696, 2015.
- J. Ludvigsson, “Novel therapies in the management of type I diabetes mellitus,” Panminerva Med., vol. 54, no. 4, pp. 257–270, 2012.
- F.-Y. Hsiao, W.-F. Huang, Y.-W. Wen, P.-F. Chen, K. N. Kuo, and Y.-W. Tsai, “Thiazolidinediones and Cardiovascular Events in Patients with Type 2 Diabetes Mellitus,” Drug Saf., vol. 32, no. 8, pp. 675–690, 2009. [CrossRef]
- J. S. Floyd, E. Barbehenn, P. Lurie, M. Sidney, and Wolfe, “Case series of liver failure associated with rosiglitazone and pioglitazone,” Pharmacoepidemiol. Drug Saf., vol. 18, pp. 1238–1243, 2009. [CrossRef]
- A. Singh, S. R. Sarkar, L. W. Gaber, and M. A. Perazella, “Acute Oxalate Nephropathy Associated With Orlistat, a Gastrointestinal Lipase Inhibitor,” Am. J. Kidney Dis., vol. 49, no. 1, pp. 153–157, 2007. [CrossRef]
- N. P. Seeram, R. Lee, and D. Heber, “Bioavailability of ellagic acid in human plasma after consumption of ellagitannins from pomegranate (Punica granatum L.) juice,” Clin. Chim. Acta, vol. 348, no. 1–2, pp. 63–68, 2004. [CrossRef]
- B. Cerdá, R. Llorach, J. J. Cerón, J. C. Espín, and F. A. Tomás-Barberán, “Evaluation of the bioavailability and metabolism in the rat of punicalagin, an antioxidant polyphenol from pomegranate juice,” Eur. J. Nutr., vol. 42, no. 1, pp. 18–28, 2003. [CrossRef]
- B. Cerdá, P. Periago, J. C. Espín, and F. A. Tomás-Barberán, “Identification of urolithin A as a metabolite produced by human colon microflora from ellagic acid and related compounds,” J. Agric. Food Chem., vol. 53, no. 14, pp. 5571–5576, 2005. [CrossRef]
- J. C. Espín, R. González-Barrio, B. Cerdá, C. López-Bote, A. I. Rey, and F. A. Tomás-Barberán, “Iberian pig as a model to clarify obscure points in the bioavailability and metabolism of ellagitannins in humans,” J. Agric. Food Chem., vol. 55, no. 25, pp. 10476–10485, 2007. [CrossRef]
- L. A. L. H. Ingorani and H. A. D. Erendorf, “Absorption , Metabolism , and Antioxidant Effects of Pomegranate ( Punica granatum L .) Polyphenols after Ingestion of a Standardized Extract in Healthy Human Volunteers,” pp. 8956–8961, 2006. [CrossRef]
- R. González-Barrio, G. Borges, W. Mullen, and A. Crozier, “Bioavailability of anthocyanins and ellagitannins following consumption of raspberries by healthy humans and subjects with an ileostomy,” J. Agric. Food Chem., vol. 58, no. 7, pp. 3933–3939, 2010. [CrossRef]
- N. P. Seeram et al., “Pomegranate ellagitannin-derived metabolites inhibit prostate cancer growth and localize to the mouse prostate gland,” J. Agric. Food Chem., vol. 55, no. 19, pp. 7732–7737, 2007. [CrossRef]
- S. M. Tripathi and D. K. Singh, “Molluscicidal activity of Punica granatum bark and Canna indica root,” Brazilian J. Med. Biol. Res., vol. 33, no. 11, pp. 1351–1355, 2000. [CrossRef]
- A. Sánchez-Lamar et al., “Assessment of the genotoxic risk of Punica granatum L. (Punicaceae) whole fruit extracts,” J. Ethnopharmacol., vol. 115, no. 3, pp. 416–422, 2007. [CrossRef]
- E. P. Vale, L. R. do Rego, D. D. N. Pureza, P. G. de B. Silva, F. F. O. de Sousa, and M. de A. B. Monteiro Neto, “Cytogenetic and toxicological effects of Punica granatum Linnaeus fruit peel hydroethanolic extract in mice,” South African J. Bot., vol. 130, pp. 465–470, 2020. [CrossRef]
- A. Vidal et al., “Studies on the toxicity of Punica granatum L. (Punicaceae) whole fruit extracts,” J. Ethnopharmacol., vol. 89, no. 2–3, pp. 295–300, 2003. [CrossRef]
- S. B. Jahromi et al., “Punica granatum peel extract toxicity in mice,” Jundishapur J. Nat. Pharm. Prod., vol. 10, no. 4, pp. 4–9, 2015. [CrossRef]
- M. Tasaki et al., “Safety assessment of ellagic acid, a food additive, in a subchronic toxicity study using F344 rats,” Food Chem. Toxicol., vol. 46, no. 3, pp. 1119–1124, 2008. [CrossRef]
- B. Cerdá, J. J. Cerón, F. A. Tomás-Barberán, and J. C. Espín, “Repeated oral administration of high doses of the pomegranate ellagitannin punicalagin to rats for 37 days is not toxic,” J. Agric. Food Chem., vol. 51, no. 11, pp. 3493–3501, 2003. [CrossRef]
- K. Pathakoti, L. Goodla, M. Manubolu, and T. Tencomnao, “Metabolic Alterations and the Protective Effect of Punicalagin Against Glutamate-Induced Oxidative Toxicity in HT22 Cells,” Neurotox. Res., vol. 31, no. 4, pp. 521–531, 2017. [CrossRef]
- M. Ye, L. Zhang, Y. Yan, and H. Lin, “Punicalagin protects H9c2 cardiomyocytes from doxorubicin-induced toxicity through activation of Nrf2/HO-1 signaling,” Biosci. Rep., vol. 39, no. 5, pp. 1–8, 2019. [CrossRef]
- I. A. T. M. Meerts et al., “Toxicological evaluation of pomegranate seed oil,” Food Chem. Toxicol., vol. 47, no. 6, pp. 1085–1092, 2009. [CrossRef]
- I. O. C. M. Vroegrijk et al., “Pomegranate seed oil, a rich source of punicic acid, prevents diet-induced obesity and insulin resistance in mice,” Food Chem. Toxicol., vol. 49, no. 6, pp. 1426–1430, 2011. [CrossRef]
- F. Lei et al., “Evidence of anti-obesity effects of the pomegranate leaf extract in high-fat diet induced obese mice,” Int. J. Obes., vol. 31, no. 6, pp. 1023–1029, 2007. [CrossRef]
- K. Z. Y. Xu, C. Zhu, M. S. Kim, J. Yamahara, and Y. Li, “Pomegranate flower ameliorates fatty liver in an animal model of type 2 diabetes and obesity,” J. Ethnopharmacol., vol. 123, no. 2, pp. 280–287, 2009. [CrossRef]
- R. A. Oliveira et al., “Effects of feeding polyphenols from pomegranate extract on health, growth, nutrient digestion, and immunocompetence of calves,” J. Dairy Sci., vol. 93, no. 9, pp. 4280–4291, 2010. [CrossRef]
- A. Shabtay et al., “Nutritive and antioxidative potential of fresh and stored pomegranate industrial byproduct as a novel beef cattle feed,” J. Agric. Food Chem., vol. 56, no. 21, pp. 10063–10070, 2008. [CrossRef]
- A. M. Neyrinck, V. F. Van Hée, L. B. Bindels, F. De Backer, P. D. Cani, and N. M. Delzenne, “Polyphenol-rich extract of pomegranate peel alleviates tissue inflammation and hypercholesterolaemia in high-fat diet-induced obese mice: Potential implication of the gut microbiota,” Br. J. Nutr., vol. 109, no. 5, pp. 802–809, 2013. [CrossRef]
- S. Asgary, A. Sahebkar, M. R. Afshani, M. Keshvari, S. Haghjooyjavanmard, and M. Rafieian-Kopaei, “Clinical evaluation of blood pressure lowering, endothelial function improving, hypolipidemic and anti-inflammatory effects of pomegranate juice in hypertensive subjects,” Phyther. Res., vol. 28, no. 2, pp. 193–199, 2014. [CrossRef]
- G. Asghari et al., “Effect of pomegranate seed oil on serum TNF-α level in dyslipidemic patients,” Int. J. Food Sci. Nutr., vol. 63, no. 3, pp. 368–371, 2012. [CrossRef]
- B. Cerdá et al., “Pomegranate juice supplementation in chronic obstructive pulmonary disease: A 5-week randomized, double-blind, placebo-controlled trial,” Eur. J. Clin. Nutr., vol. 60, no. 2, pp. 245–253, 2006. [CrossRef]
- M. González-Ortiz, E. Martínez-Abundis, M. C. Espinel-Bermúdez, and K. G. Pérez-Rubio, “Effect of pomegranate juice on insulin secretion and sensitivity in patients with obesity,” Ann. Nutr. Metab., vol. 58, no. 3, pp. 220–223, 2011. [CrossRef]
- S. Hamoud et al., “Pomegranate extract (POMx) decreases the atherogenicity of serum and of human monocyte-derived macrophages (HMDM) in simvastatin-treated hypercholesterolemic patients: A double-blinded, placebo-controlled, randomized, prospective pilot study,” Atherosclerosis, vol. 232, no. 1, pp. 204–210, 2014. [CrossRef]
- L. Shema-Didi, B. Kristal, S. Sela, R. Geron, and L. Ore, “Does Pomegranate intake attenuate cardiovascular risk factors in hemodialysis patients?,” Nutr. J., vol. 13, no. 1, pp. 2–9, 2014. [CrossRef]
- M. D. Sumner et al., “Effects of pomegranate juice consumption on myocardial perfusion in patients with coronary heart disease,” Am. J. Cardiol., vol. 96, no. 6, pp. 810–814, 2005. [CrossRef]
- C. Tsang, N. F. Smail, S. Almoosawi, I. Davidson, and E. A. S. Al-Dujaili, “Intake of polyphenol-rich pomegranate pure juice influences urinary glucocorticoids, blood pressure and homeostasis model assessment of insulin resistance in human volunteers,” J. Nutr. Sci., vol. 1, pp. 1–9, 2012. [CrossRef]
- A. Sadeghipour, M. Eidi, A. Ilchizadeh Kavgani, R. Ghahramani, S. Shahabzadeh, and A. Anissian, “Lipid lowering effect of punica granatum L. Peel in high lipid diet fed male rats,” Evidence-based Complement. Altern. Med., vol. 2014, 2014. [CrossRef]
- F. L. A. Hossin, “Effect of pomegranate (Punica granatum) peels and it’s extract on obese hypercholesterolemic rats,” Pakistan J. Nutr., vol. 8, no. 8, pp. 1251–1257, 2009. [CrossRef]
- A. E. El-Hadary and M. F. Ramadan, “Phenolic profiles, antihyperglycemic, antihyperlipidemic, and antioxidant properties of pomegranate (Punica granatum) peel extract,” J. Food Biochem., vol. 43, no. 4, pp. 1–9, 2019. [CrossRef]
- P. Bagri, M. Ali, V. Aeri, M. Bhowmik, and S. Sultana, “Antidiabetic effect of Punica granatum flowers: Effect on hyperlipidemia, pancreatic cells lipid peroxidation and antioxidant enzymes in experimental diabetes,” Food Chem. Toxicol., vol. 47, no. 1, pp. 50–54, 2009. [CrossRef]
- A. N. Patel, D. D. Bandawane, and N. K. Mhetre, “Pomegranate (Punica granatum Linn.) leaves attenuate disturbed glucose homeostasis and hyperglycemia mediated hyperlipidemia and oxidative stress in streptozotocin induced diabetic rats,” Eur. J. Integr. Med., vol. 6, no. 3, pp. 307–321, 2014. [CrossRef]
- M. Sarker, Z. A. Mahmud, S. K. Saha, N. S. Tithi, M. S. Ali, and S. C. Bachar, “Antihyperlipidemic activity of flowers of punica granatum in poloxamer-407 induced hyperlipidemic mice model,” Pharmacogn. J., vol. 4, no. 27, pp. 66–70, 2012. [CrossRef]
- K. G. Al-Fartosi, R. J. Tuama, A. B. Roomi, & Saad, and H. Jasim, “Effect of Pomegranate (Punica Granatum L) Juice on Lipid Profile of Hyperlipidemic Female Mice,” Int. J. Res. Applied, Nat. Soc. Sci., vol. 3, no. 10, pp. 2321–8851, 2015.
- M. F. Saleh and F. H. Mohammed, “ANTI HYPERLIPIDEMIC EFFECT OF ETHANOLIC EXTRACT OF PUNICA GRANATUM L . MESOCARP IN HYPERLIPIDEMIC RATS,” vol. 20, no. 2, pp. 8989–8994, 2020.
- M. Mueller and A. Jungbauer, “Culinary plants, herbs and spices - A rich source of PPARγ ligands,” Food Chem., vol. 117, no. 4, pp. 660–667, 2009. [CrossRef]
- M. Shiner, B. Fuhrman, and M. Aviram, “Macrophage paraoxonase 2 (PON2) expression is up-regulated by pomegranate juice phenolic anti-oxidants via PPARγ and AP-1 pathway activation,” Atherosclerosis, vol. 195, no. 2, pp. 313–321, 2007. [CrossRef]
- T. H. W. Huang et al., “Anti-diabetic action of Punica granatum flower extract: Activation of PPAR-γ and identification of an active component,” Toxicol. Appl. Pharmacol., vol. 207, no. 2, pp. 160–169, 2005. [CrossRef]
- H. Kohno, R. Suzuki, Y. Yasui, M. Hosokawa, K. Miyashita, and T. Tanaka, “Pomegranate seed oil rich in conjugated linolenic acid suppresses chemically induced colon carcinogenesis in rats,” Cancer Sci., vol. 95, no. 6, pp. 481–486, 2004. [CrossRef]
- Y. Makino-Wakagi, Y. Yoshimura, Y. Uzawa, N. Zaima, T. Moriyama, and Y. Kawamura, “Ellagic acid in pomegranate suppresses resistin secretion by a novel regulatory mechanism involving the degradation of intracellular resistin protein in adipocytes,” Biochem. Biophys. Res. Commun., vol. 417, no. 2, pp. 880–885, 2012. [CrossRef]
- B. K. McFarlin, K. A. Strohacker, and M. L. Kueht, “Pomegranate seed oil consumption during a period of high-fat feeding reduces weight gain and reduces type 2 diabetes risk in CD-1 mice,” Br. J. Nutr., vol. 102, no. 1, pp. 54–59, 2009. [CrossRef]
- H. S. Parmar and A. Kar, “Antidiabetic potential of Citrus sinensis and Punica granatum peel extracts in alloxan treated male mice,” BioFactors, vol. 31, no. 1, pp. 17–24, 2007. [CrossRef]
- Y. Li et al., “Punica granatum flower extract, a potent α-glucosidase inhibitor, improves postprandial hyperglycemia in Zucker diabetic fatty rats,” J. Ethnopharmacol., vol. 99, no. 2, pp. 239–244, 2005. [CrossRef]
- E. A. M. Khalil, “Antidiabetic effect of an aqueous extract of Pomegranate (Punica granatum L.) peels in normal and alloxan diabetic rats,” Egypt. J. Hosp. Med., vol. 16, no. 1, pp. 92–99, 2004. [CrossRef]
- Y. A., S. E.Y., F. I., S. F., and K. A. I., “Antiobesity, antidiabetic and antioxidant activities of senna (senna alexandrina mill.) and pomegranate (punica granatum l.) leaves extracts and its fractions,” Int. J. Pharm. Phytopharm. Res., vol. 8, no. 3, pp. 18–24, 2018.
- G. Jelodar, M. Maleki, and S. Sirus, “EFFECT OF WALNUT LEAF, CORIANDER AND POMEGRANATE ON BLOOD GLUCOSE AND HISTOPATHOLOGY OF PANCREAS OF ALLOXAN INDUCED DIABETIC RATS,” Afr. J. Trad., vol. 31, no. 1, pp. 299–305, 2007. [CrossRef]
- B. Hosseini, A. Saedisomeolia, L. G. Wood, M. Yaseri, and S. Tavasoli, “Effects of pomegranate extract supplementation on inflammation in overweight and obese individuals: A randomized controlled clinical trial,” Complement. Ther. Clin. Pract., vol. 22, pp. 44–50, 2016. [CrossRef]
- Y. Khajebishak, L. Payahoo, M. Alivand, and B. Alipour, “Punicic acid: A potential compound of pomegranate seed oil in Type 2 diabetes mellitus management,” J. Cell. Physiol., vol. 234, no. 3, pp. 2112–2120, 2019. [CrossRef]
- S. Ö. Akarca Dizakar, G. S. Saribas, and A. Tekcan, “Effects of ellagic acid in the testes of streptozotocin induced diabetic rats,” Drug Chem. Toxicol., vol. 0, no. 0, pp. 1–8, 2021. [CrossRef]
- Y. Zhang, Y. Cao, J. Chen, H. Qin, and L. Yang, “A New Possible Mechanism by Which Punicalagin Protects against Liver Injury Induced by Type 2 Diabetes Mellitus: Upregulation of Autophagy via the Akt/FoxO3a Signaling Pathway,” J. Agric. Food Chem., vol. 67, no. 50, pp. 13948–13959, 2019. [CrossRef]
- R. Hontecillas, M. Diguardo, E. Duran, M. Orpi, and J. Bassaganya-Riera, “Catalpic acid decreases abdominal fat deposition, improves glucose homeostasis and upregulates PPAR α expression in adipose tissue,” Clin. Nutr., vol. 27, no. 5, pp. 764–772, 2008. [CrossRef]
- L. Fajas et al., “The organization, promoter analysis, and expression of the human PPARγ gene,” J. Biol. Chem., vol. 272, no. 30, pp. 18779–18789, 1997. [CrossRef]
- A. Chawla and A. Lazar, “Peroxisome Expression In Adipocyte Differentiation Receptor ( WAR ) y : And Induction,” Endocrinology, vol. 135, no. 2, pp. 798–800, 1994.
- P. Tontonoz, E. Hu, and B. M. Spiegelman, “Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor,” Cell, vol. 79, no. 7, pp. 1147–1156, 1994. [CrossRef]
- J. Berger and D. E. Moller, “THE MECHANISMS OF ACTION OF PPARs.,” Annu. Rev. Med, vol. 53, pp. 409–435, 2002. [CrossRef]
- S. M. Rangwala and M. A. Lazar, “Peroxisome proliferator-activated receptor γ in diabetes and metabolism,” Trends Pharmacol. Sci., vol. 25, no. 6, pp. 331–336, 2004. [CrossRef]
- R. S. Farag, M. S. Abdel-Latif, S. Emam, and S. Tawfeek, “Phytochemical screening and polyphenol constituents of pomegranate peels and leave juices,” Agric. Soil Sci., vol. 1, no. 6, pp. 86–93, 2014.
- J.-H. EokLee, S.-D. Kim, J.-Y. Lee, K.-N. Kim, and H.-S. Kim, “Analysis of Flavonoids in Concentrated Pomegranate extract by HPLC with Diod Array Detection.,” Food Sci. Biotechnol, vol. 14, no. 1, pp. 171–174, 2005.
- N. Parsaeyan, H. Mozaffari-Khosravi, and M. R. Mozayan, “Effect of pomegranate juice on paraoxonase enzyme activity in patients with type 2 diabetes,” J. Diabetes Metab. Disord., vol. 11, no. 1, pp. 2–5, 2012. [CrossRef]
- S. Jandari, E. Hatami, R. Ziaei, A. Ghavami, and A. M. Yamchi, “The effect of pomegranate (Punica granatum) supplementation on metabolic status in patients with type 2 diabetes: A systematic review and meta-analysis,” Complement. Ther. Med., vol. 52, p. 102478, 2020. [CrossRef]
- M. A. Jafri, M. Aslam, K. Javed, and S. Singh, “Effect of punica granatum Linn. (flowers) on blood glucose level in normal and alloxan- induced diabetic rats,” J. Ethnopharmacol., pp. 309–314, 2000. [CrossRef]
- A. K. Das, S. C. Mandal, S. K. Banerjee, S. Sinha, B. P. Saha, and M. Pal, “Studies on the hypoglycaemic activity of Punica granatum seed in streptozotocin induced diabetic rats,” Phyther. Res., vol. 15, no. 7, pp. 628–629, 2001. [CrossRef]
- R. Hontecillas, M. O’Shea, A. Einerhand, M. Diguardo, and J. Bassaganya-Riera, “Activation of ppar γ and α by punicic acid ameliorates glucose tolerance and suppresses obesity-related inflammation,” J. Am. Coll. Nutr., vol. 28, no. 2, pp. 184–195, 2009. [CrossRef]
- T. H. W. Huang et al., “Pomegranate flower improves cardiac lipid metabolism in a diabetic rat model: Role of lowering circulating lipids,” Br. J. Pharmacol., vol. 145, no. 6, pp. 767–774, 2005. [CrossRef]
- A. Esmaillzadeh, F. Tahbaz, I. Gaieni, H. Alavi-Majd, and L. Azadbakht, “Concentrated pomegranate juice improves lipid profiles in diabetic patients with hyperlipidemia,” J. Med. Food, vol. 7, no. 3, pp. 305–308, 2004. [CrossRef]
- B. Fuhrman, N. Volkova, and M. Aviram, “Pomegranate juice inhibits oxidized LDL uptake and cholesterol biosynthesis in macrophages,” J. Nutr. Biochem., vol. 16, no. 9, pp. 570–576, 2005. [CrossRef]
- F. De Nigris et al., “Beneficial effects of pomegranate juice on oxidation-sensitive genes and endothelial nitric oxide synthase activity at sites of perturbed shear stress,” Proc. Natl. Acad. Sci. U. S. A., vol. 102, no. 13, pp. 4896–4901, 2005. [CrossRef]
- M. Aviram et al., “Pomegranate juice consumption for 3 years by patients with carotid artery stenosis reduces common carotid intima-media thickness, blood pressure and LDL oxidation,” Clin. Nutr., vol. 23, no. 3, pp. 423–433, 2004. [CrossRef]
- M. Kaplan et al., “Pomegranate juice supplementation to atherosclerotic mice reduces macrophage lipid peroxidation, cellular cholesterol accumulation and development of atherosclerosis,” J. Nutr., vol. 131, no. 8, pp. 2082–2089, 2001. [CrossRef]
- M. Rosenblat, N. Volkova, R. Coleman, and M. Aviram, “Pomegranate byproduct administration to apolipoprotein E-deficient mice attenuates atherosclerosis development as a result of decreased macrophage oxidative stress and reduced cellular uptake of oxidized low-density lipoprotein,” J. Agric. Food Chem., vol. 54, no. 5, pp. 1928–1935, 2006. [CrossRef]
- E. ElHussieny, R. Soliman, and N. ElBeih, “Ameliorative Effects of Pomegranate Peel Extract and Some of its Bioactive Components against Hyperlipidemia-Induced Atherosclerosis in Male Rats,” Egypt. J. Exp. Biol., vol. 14, no. 1, p. 85, 2018. [CrossRef]
- T. H. W. Huang et al., “Pomegranate flower extract diminishes cardiac fibrosis in zucker diabetic fatty rats: Modulation of cardiac endothelin-1 and nuclear factor-kappaB pathways,” J. Cardiovasc. Pharmacol., vol. 46, no. 6, pp. 856–862, 2005. [CrossRef]
- J. N. Losso, R. R. Bansode, A. Trappey, H. A. Bawadi, and R. Truax, “In vitro anti-proliferative activities of ellagic acid,” J. Nutr. Biochem., vol. 15, no. 11, pp. 672–678, 2004. [CrossRef]
- J. Khateeb, A. Gantman, A. J. Kreitenberg, M. Aviram, and B. Fuhrman, “Paraoxonase 1 (PON1) expression in hepatocytes is upregulated by pomegranate polyphenols: A role for PPAR-γ pathway,” Atherosclerosis, vol. 208, no. 1, pp. 119–125, 2010. [CrossRef]
- E. Burgermeister, D. Chuderland, T. Hanoch, M. Meyer, M. Liscovitch, and R. Seger, “Interaction with MEK Causes Nuclear Export and Downregulation of Peroxisome Proliferator-Activated Receptor γ,” Mol. Cell. Biol., vol. 27, no. 3, pp. 803–817, 2007. [CrossRef]
- W. Rock, M. Rosenblat, R. Miller-Lotan, A. P. Levy, M. Elias, and M. Aviram, “Consumption of Wonderful variety pomegranate juice and extract by diabetic patients increases paraoxonase 1 association with high-density lipoprotein and stimulates its catalytic activities,” J. Agric. Food Chem., vol. 56, no. 18, pp. 8704–8713, 2008. [CrossRef]
- F. Hadaegh, D. Khalili, A. Ghasemi, M. Tohidi, F. Sheikholeslami, and F. Azizi, “Triglyceride/HDL-cholesterol ratio is an independent predictor for coronary heart disease in a population of Iranian men,” Nutr. Metab. Cardiovasc. Dis., vol. 19, no. 6, pp. 401–408, 2009. [CrossRef]
- P. Mirmiran, M. R. Fazeli, G. Asghari, A. Shafiee, and F. Azizi, “Effect of pomegranate seed oil on hyperlipidaemic subjects: A double-blind placebo-controlled clinical trial,” Br. J. Nutr., vol. 104, no. 3, pp. 402–406, 2010. [CrossRef]
- A. Esmaillzadeh, F. Tahbaz, I. Gaieni, H. Alavi-Majd, and L. Azadbakht, “Cholesterol-lowering effect of concentrated pomegranate juice consumption in type II diabetic patients with hyperlipidemia,” Int. J. Vitam. Nutr. Res., vol. 76, no. 3, pp. 147–151, 2006. [CrossRef]
- J. M. McKenney, “Pharmacotherapy of dyslipidemia,” Cardiovasc. Drugs Ther., vol. 15, no. 5, pp. 413–422, 2001. [CrossRef]
- A. Sahebkar, L. E. Simental-Mendía, P. Giorgini, C. Ferri, and D. Grassi, “Lipid profile changes after pomegranate consumption: A systematic review and meta-analysis of randomized controlled trials,” Phytomedicine, vol. 23, no. 11, pp. 1103–1112, 2016. [CrossRef]
- F. de Nigris et al., “The influence of pomegranate fruit extract in comparison to regular pomegranate juice and seed oil on nitric oxide and arterial function in obese Zucker rats,” Nitric Oxide - Biol. Chem., vol. 17, no. 1, pp. 50–54, 2007. [CrossRef]
- K. B. Arun, P. Jayamurthy, C. V. Anusha, S. K. Mahesh, and P. Nisha, “Studies on Activity Guided Fractionation of Pomegranate Peel Extracts and Its Effect on Antidiabetic and Cardiovascular Protection Properties,” J. Food Process. Preserv., vol. 41, no. 1, pp. 1–12, 2017. [CrossRef]
- M. Mohan, H. Waghulde, and S. Kasture, “Effect of pomegranate juice on angiotensin II-induced hypertension in diabetic wistar rats,” Phyther. Res., vol. 24, no. SUPPL. 2, pp. 196–203, 2010. [CrossRef]
- R. L. dos Santos et al., “Pomegranate peel extract attenuates oxidative stress by decreasing coronary angiotensin-converting enzyme (ACE) activity in hypertensive female rats,” J. Toxicol. Environ. Heal. - Part A Curr. Issues, vol. 79, no. 21, pp. 998–1007, 2016. [CrossRef]
- M. H. Davidson et al., “Effects of Consumption of Pomegranate Juice on Carotid Intima-Media Thickness in Men and Women at Moderate Risk for Coronary Heart Disease,” Am. J. Cardiol., vol. 104, no. 7, pp. 936–942, 2009. [CrossRef]
- A. Lynn, H. Hamadeh, W. C. Leung, J. M. Russell, and M. E. Barker, “Effects of Pomegranate Juice Supplementation on Pulse Wave Velocity and Blood Pressure in Healthy Young and Middle-aged Men and Women,” Plant Foods Hum. Nutr., vol. 67, no. 3, pp. 309–314, 2012. [CrossRef]
- G. Sohrab et al., “Effects of pomegranate juice consumption on oxidative stress in patients with type 2 diabetes: a single-blind, randomized clinical trial,” Int. J. Food Sci. Nutr., vol. 68, no. 2, pp. 249–255, 2017. [CrossRef]
- A. Sahebkar, C. Ferri, P. Giorgini, S. Bo, P. Nachtigal, and D. Grassi, “Effects of pomegranate juice on blood pressure: A systematic review and meta-analysis of randomized controlled trials,” Pharmacol. Res., vol. 115, pp. 149–161, 2017. [CrossRef]
- M. Aviram and L. Dornfeld, “Pomegranate juice consumption inhibits serum angiotensin converting enzyme activity and reduces systolic blood pressure,” Atherosclerosis, vol. 158, no. 1, pp. 195–198, 2001. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



