
Submitted:
30 September 2023
Posted:
02 October 2023
You are already at the latest version
Abstract
Angiogenesis is a process whereby new blood vessels are born from preexistent ones, which has a role in transporting oxygen and nutrients. On the other hand, inhibiting this process is a strategy used to treat some diseases, including cancers, diabetic retinopathy, and rheumatoid arthritis. This study investigates the effect of a new nutraceutical (Terasen®) on angiogenesis. Terasen® is made from a standardized extract of Euterpe oleraceae (Açaí), Myrciaria dubia (Camu-Camu), and purified oil of Bixa orellana (Annatto) and Astrocaryum aculeatum (Tucumã). The in vivo angiogenesis model used was the chorioallantoic membrane (CAM) from chicken embryos. The embryos were divided into seven groups: a vehicle control treated with ethanol solution (ETH), a naïve control treated only with saline solution (SOR), a positive control treated with dexamethasone (DEX), and four groups treated with Terasen® at different concentrations (0.5, 1, 2, and 3 mg/ml). The parameters assessed were the vessels’ area, length, and amount. The results show a statistically significant and concentration-dependent inhibition of angiogenesis caused by Terasen®. Our results suggest the nutraceutical has a high antiangiogenesis potential, possibly due to its bioactive compounds, including anthocyanins, quercetin, and tocotrienols.
Keywords:
angiogenesis
; nutraceuticals
; chorioallantoic membrane
; anthocyanins
; tocotrienols.
1. Introduction
Angiogenesis is defined as the formation of new blood vessels from the preexistent ones [1], it is a continuous process that begins during embryo development and lasts throughout the entire organism’s life [2]. Angiogenesis is crucial to cells’ growth, differentiation, and tissue formation since blood vessels are necessary to distribute oxygen and nutrients to cells [3]. It is critical during development and other conditions, including wound healing [2]. In addition, inducing the growth of arteries in patients with severe limb ischemia using growth factors can prevent limb amputations [1].
Under typical healthy conditions, stimulating and inhibitory factors of angiogenesis are in balance. Nevertheless, this balance is impaired under certain pathological conditions, and the stimulating factors induce angiogenesis, favoring some pathologies that rely on it [4]. Hence, inhibiting angiogenesis can have a beneficial role against such diseases, including cancers, macular degeneration, diabetic retinopathy, rheumatoid arthritis, and psoriasis [5].
A popular model to evaluate angiogenesis in vivo uses the chicken embryos’ chorioallantoic membrane (CAM) [1]. The CAM is a highly vascularized extra-embryonic membrane responsible for the embryo’s respiration [6]. It is formed from the fourth day after incubation through the fusion of the external mesodermal allantoid layer with the mesodermal layer of the chorion. From the 8th to the 10th day, there is a fast vascular network growth with capillary differentiation and complete development of the CAM central area. During this period, it is possible to analyze and quantify the CAM’s angiogenesis through the vessel network density, which helps test potential pro- or anti-angiogenic drugs [1,7].
Terasen® is a nutraceutical made from encapsulated granules from standardized extracts of the association of standardized extracts of Euterpe oleracea, Myrciaria dubia, and purified oil of Bixa orellana and Astrocaryum aculeatum. Some important phytochemical markers from this product are anthocyanins, quercetin, ellagic acid, gallic acid, carotenoids, geranylgeraniol, and tocotrienols. These compounds have been extensively studied for their biological activities, including anti-angiogenic potential [8,9,10,11,12,13]. Hence, this study aimed to evaluate the potential anti-angiogenic activity of Terasen® using the CAM model.
2. Materials and Methods
2.1. Product Obtention and Scanning Electron Microscopy (SEM) Analysis
The nutraceutical granules from Terasen® were kindly provided by Ages Bioactive Compounds (São Paulo, SP, Brazil), lot URU201101. The products’ granules were analyzed through SEM through photomicrographs using a Hitachi T3030 Plus Tabletop equipment (15 kV).
2.2. Phytochemical Analysis
2.2.1. Anthocyanins
The total monomeric anthocyanins were quantified through the differential pH method and expressed in cyanidin-3-O-glucoside (C3OG) equivalents using a technique based on the AOAC method [9]. First, a buffer solution (pH 1.0, KCl 0.025 M, Sigma, USA) was added to a 10 mL volumetric flask with 50 mg of the test product. Another 10 mL volumetric flask was filled with another buffer solution (pH 4.5, sodium acetate 0.4 M, Sigma, USA). Both flasks were gently stirred and left to rest for 15 minutes. Next, the solutions were filtered using filter paper (Whatman n° 1), and the absorbance of the samples was registered using a UV-Vis spectrophotometer (Shimadzu, Japan) at 520 and 700 nm. The total anthocyanin content (TAC) was calculated as cyanidin-3-O-glucoside equivalents (mg/mL) using the following equation 1:
Where:
- A
- = (A 520nm -A700nm) pH1.0 - (A520nm - A700nm) pH 4.5;
- A
- ε = cyanidin-3-O-glucoside extinction coefficient;
- A
- MM = cyanidin-3-O-glucoside molecular mass;
- A
- DF = dilution factor;
- V
- = solution volume;
- m
- = mass of the extract.
2.2.2. Total Flavonoids
The total flavonoids were quantified as quercetin equivalents using a calibration curve with concentrations within an interval from 5 µg/ml to 50 µg/ml based on a method published before [14]. First, Terasen® was solubilized in methanol at 4 mg/ml. Next, 1 mL of aluminum chloride (Sigma Aldrich, Germany) at 5% (m/v) in methanol was added to the standard and sample solutions, stirred using a vortex for 30 seconds, and left to rest for 30 minutes away from heat and light. After 30 minutes, the samples were assessed using a UV-Vis spectrophotometer at 420 nm with a glass cuvette with a 1 cm optical path.
2.2.3. Geranylgeraniol
Geranylgeraniol was quantified using a calibration curve between 0.1 and 2 mg/ml. The sample solution was prepared using 10 mg/ml of Terasen® in hexane. For the quantification, was used GC-MS with Shimadzu equipment (GC2010, Kyoto, Japan) with an autoinjector (AOC 5000), mass detector (MS2010, 70 eV), and fused silica column (DB-5MS, 30 m x 0.025 mm x 0.25 m).
The running parameters were set as follows: 120 °C of initial temperature with 3 °C of heating per minute until 280 °C; from this moment, the temperature was kept constant for 3 minutes, totaling 56.3 of analysis. The injection temperature was at 270 °C with 80.8 kPa and a 10.0 split ratio. The ion source and interface temperature were set at 260 °C.
2.2.4. Δ-tocotrienol
To quantify δ-tocotrienol from Terasen® was used a calibration curve between 0.2 and 1 mg/ml. Terasen® was used at 10 mg/ml in hexane for the analysis. This analysis was performed through GC-MS using the same equipment described for geranylgeraniol.
The parameters were set as follows: 180 °C of initial temperature for five minutes, followed by 3 °C of heating per minute until 300 °C; from this moment, the temperature was kept constant for 15 minutes totalizing 40 of analysis. The injection temperature was 210 °C with 80.1 kPa and a 5.0 split ratio. The ion source and interface temperature were set at 250 °C.
2.3. In Vivo Anti-Angiogenic CAM Assay
The eggs used in this experiment were from standard chicken (Gallus gallus), bought from Agro Cam (Macapá, Amapá, Brazil). The procedure used followed the method described by (West et al., 2001).
First, the eggs were washed under ambient temperature with sodium hypochlorite at 2% (v/v) and incubated horizontally to ensure embryo growth away from the CAM. The incubation occurred at 37 – 37.5 °C with 60 – 70% humidity, and the first day of incubation was defined as day zero (Figure 1).
On the third day of incubation, a small hole was made at the pointy side of the eggs using a 5 mL syringe, and 2 mL of albumin was removed. The eggs were put back into the incubator horizontally with the hole facing upwards.
On the fourth day was used a cutting machine with a rotary clamp drill and cutting blade (Dremel 3000, Bosch, Ciudad de Mexico, Mexico) to make rectangular-shaped cuts (3 x 2.5 cm) at the eggs’ side. The cut eggshell was gently removed with a clamp, exposing the CAM, and the open hole was covered with micropore tape (Sanfarma, São Paulo, Brazil). After this procedure, the eggs were returned to the incubator horizontally, with the rectangular role facing upwards.
On the eighth day of incubation, the eggs received their treatment according to their group; there were seven groups with five eggs each: a naïve control group treated only with saline solution 0.9% (SOR, Farmax Lab S.A., lot 0873, Divinópolis, Minas Gerais, Brazil), a vehicle control group treated with 90% ethanol (ETH, Santa Cruz), a positive control group treated with dexamethasone 4 mg/ml (DEX, Teuto Lab, lot 94570172, Anápolis, Goiás, Brazil), and four groups treated with Terasen® (solubilized in 90% ethanol) at different concentrations – 0.5 mg/ml (TERA 0.5), 1 mg/ml (TERA 1), 2 mg/ml (TERA 2), and 3 mg/ml (TERA 3). Before the treatment, the eggs’ CAM were photographed to record the vessels before treatments with high-quality pictures (4,000 x 3,000 px), using a camera coupled to a stereomicroscope (Estereotereo Luxeo 4D, Labomed, CA, EUA).
Finally, on the tenth day, the CAM was examined and photographed again to evaluate the effect of the treatments. All the process is summarized in Figure 1.
The blood vessels from the pictures were analyzed using ImageJ (v1.53u, Massachusetts EUA) with the Angiogenesis Analyzer plugin. The parameters evaluated were the number of blood vessels, area, and the average vessels’ length.
2.4. Statistical Analysis
All results were expressed as a mean ± SEM, and the groups were compared through One-way ANOVA followed by Tukey posthoc test in case of statistical difference (p < 0.05). All analyses were performed using GraphPad Prism v5.03.
3. Results
3.1. Granules Analysis
The SEM pictures of Terasen® are shown in Figure 2. It is observed that the formulation has irregular-shaped particles of aggregated granules. Such particles are formed due to the production method that propitiates the formation of lecithin clusters containing Chronic® [16] inside, adsorbed in other excipients.
3.2. Phytochemical Analysis
The concentration of total monomeric anthocyanins in Terasen® was 0.43417 ± 0.1525 mg/g C3OG equivalents (0.04341 ± 0.0152 %). The quantification was performed through the differential pH method of AOAC (2006); hence there was no calibration curve.
The calibration curve of the quercetin-equivalent total flavonoids is shown in Figure 3A. The line equation was calculated as y =39.808x-0.007629 with R² = 0.991. It was calculated 2.4501 ± 0.7341 mg/g of total flavonoids in the sample (2.4501 ± 0.0734%).
3.3. Effect of Terasen® on Angiogenesis
4. Discussion
The initiation of new blood vessel formation occurs through angiogenic stimulation, which can be triggered by factors such as oxygen reduction (hypoxia), decreased blood flow (ischemia), or the release of signaling molecules and growth factors. This process sets off a complex cascade of cellular and molecular events that culminate in the generation of new vessels, fulfilling the increased oxygen and nutrient demands in tissues requiring enhanced vascularization [17]. Angiogenesis activation promotes the activation of matrix metalloproteinases (MMPs), responsible for the degradation and reorganization of the extracellular matrix during new blood vessel formation [18,19].
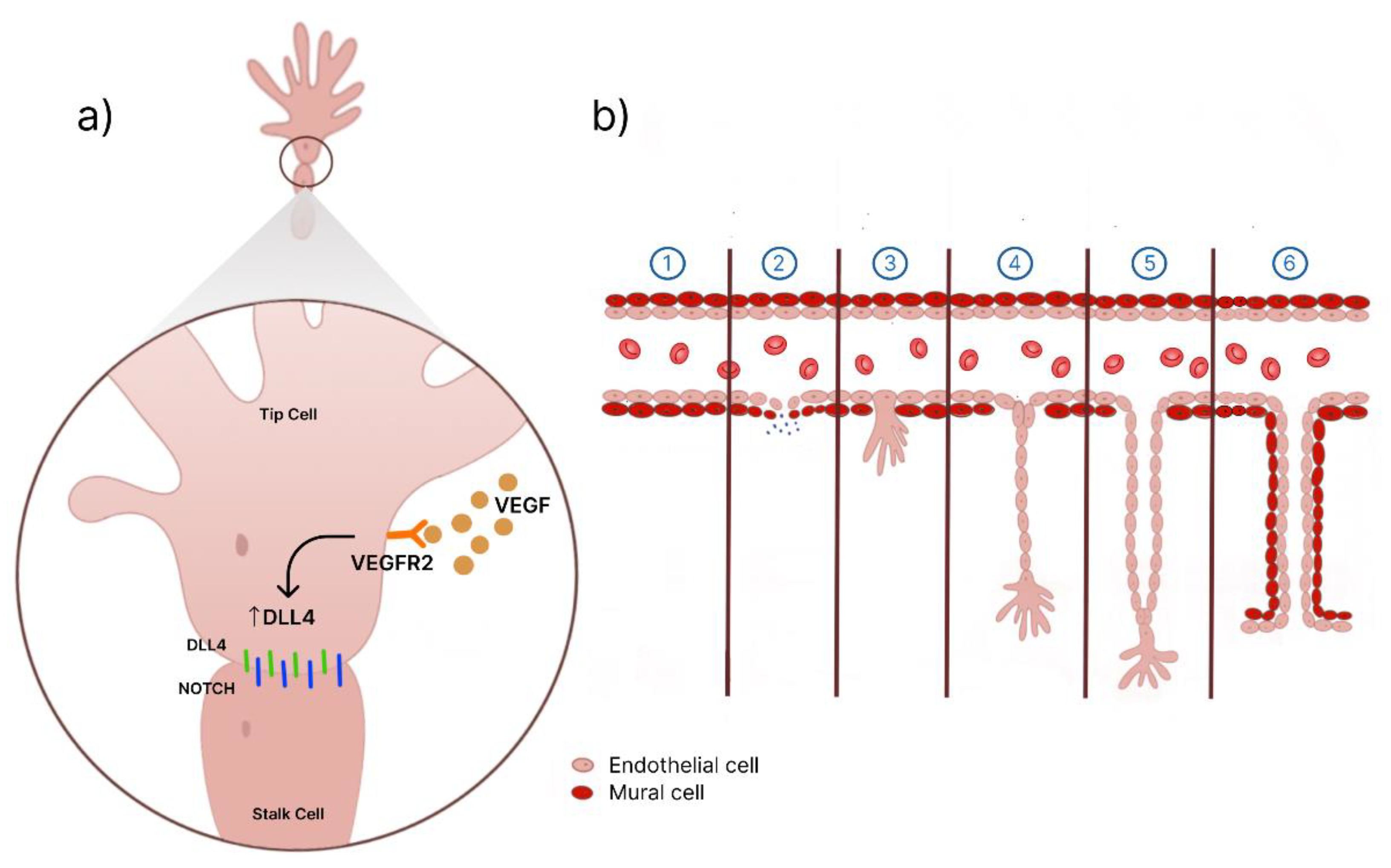
Subsequently, proliferation and migration occur, with new branches formed by tip cells. Tip cells are proliferative endothelial cells that respond to the angiogenic factor VEGF, triggering filopodia production that guides the formation of new blood vessels toward the VEGF gradient [18]. Furthermore, VEGF plays a pivotal role in the differentiation of tip and stalk cells [20]. Stimulation by VEGF-VEGFR2 in tip cells leads to the expression of the DLL4 protein, which binds to Notch receptors in stalk cells [20]. Activation mediated by DLL4-Notch results in downregulating VEGFR-2 expression, whereas inhibition of the Dll4-Notch signaling pathway leads to an upregulation of VEGFR-2 [21]. Thus, blockade of the Dll4-Notch pathway and positive regulation of VEGFR-2 in the VEGF signaling pathway can promote angiogenesis.
Tip cells lead the process of new blood vessel formation, accompanied by stalk cells, working together to facilitate lumen formation and the development of the tubular structure. Fibroblasts are recruited to the site as a new sprout emerges and differentiate into smooth muscle cells, lining the vessel wall [17,18]. Following this stage, anastomosis occurs, which involves the connection of the new vessels with pre-existing ones [18]. A simplified diagram is shown in Figure 6.
Despite the unquestionable significance of angiogenesis, it is involved in several pathological conditions. Therefore, the inhibition of angiogenesis has become a focus in treating various diseases. In this context, we have developed a nutraceutical derived from herbal products as a potential strategy to inhibit angiogenesis and aid in treating these conditions. Following the formulation development, the initial stage involved the characterization of its morphology.
SEM analysis allows for characterizing the formulation’s structure, surface morphology, drug adsorption homogeneity in the excipient, and droplet scattering [22,23]. The SEM analysis of Terasen® reveals the presence of agglomerated particles with irregular surfaces and pores. The rotation speed employed during the formulation process plays a role in determining the formulation’s droplet size distribution and mechanical stability [24]. In this study, the granules were formed by adding sunflower lecithin at 3,600 rpm. According to Liu et al. [24], high rotation speeds (> 170 rpm) can result in the formation of irregularly shaped granules, promote impact with liquid droplets during pulverization, and increase the yield of larger granules. The uneven granular surface also enhances the penetration of the dissolution medium [25].
The added lecithin in the formulation serves as an adjuvant and is composed of a mixture of phosphatidic acids and phospholipids. Following international regulations, lecithin should exhibit insolubility of > 60% in acetone and < 0.3% in hexane [26,27]. The increasing interest in lecithin as an ingredient stems from its unique chemical properties, offering pharmaceutical, cosmetic, and nutraceutical value [28]. The chemical structure of its compounds, particularly phospholipids, can influence its molecular geometry and functional properties, such as self-assembly and emulsification. Lecithin can form lamellar structures with single or double layers depending on the proportion of phosphatidylcholine and phosphatidylinositol. This property can enhance the stability and hydration of agglomerated structures [26,28,29].
After conducting a morphological analysis of the formulation, we proceeded to evaluate the concentration of its main bioactive compounds, including total monomeric anthocyanins, total quercetin-equivalent flavonoids, geranylgeraniol, and δ-tocotrienol. These compounds were selected as they represent the major bioactive constituents present in the ingredients of Terasen®, the name of the nutraceutical composed of standardized extracts of E. oleracea, M. dubia, and purified oil of B. orellana and A. aculeatum.
The anthocyanins present in Terasen® originate from the dry extract of E. oleracea. [30] quantified the total monomeric anthocyanins in eight samples of E. oleracea extract and reported an average concentration of 3.6 mg/g of C3OG equivalents, with values ranging from 1 to 7.3 mg/g. In our study, the results obtained were lower, which is expected since other ingredients were added to the nutraceutical formulation, not solely the E. oleracea extract. The primary anthocyanins found in E. oleracea include Cyanidin-3-O-rutinoside, Cyanidin-3-O-glucoside, Pelargonidin-3-O-rutinoside, and Peonidin 3-O-rutinoside [31].
Most of the total flavonoids in Terasen® are derived from the dry extracts of E. oleracea and M. dubia. [30] analyzed the quercetin-equivalent flavonoid content in eight samples of E. oleracea extract and reported an average concentration of 2.1 mg/g, with values ranging from 1.3 to 3 mg/g. According to Kang et al. [32], the main flavonoids found in açaí are dihydrokaempferol, isovitexin, and velutin. As for M. dubia, Fidelis et al. [33] reported quercetin concentrations of 0.007 mg/g in the aqueous extract and 0.0085 mg/g in the hydroethanolic extract, as well as rutin concentrations of 0.035 mg/g in the aqueous extract and 0.14 mg/g in the hydroethanolic extract. Given that our product contains 2.4 mg/g of quercetin-equivalent flavonoids, the concentration in the extracts is expected to be high. Additionally, M. dubia is known for its high content of ascorbic acid, polyphenols, flavonols, and carotenoids. Interestingly, Souza et al. [34] also detected anthocyanins in ripe and semi-ripe M. dubia fruits.
The oil derived from B. orellana, which is included in the formulation of Terasen®, contains δ-tocotrienol and geranylgeraniol as its main compounds. Previous studies have reported the presence of δ-tocotrienol in the seeds of B. orellana [35,36,37] and geranylgeraniol [37,38].
Following the phytochemical analysis, we conducted the chicken CAM (chorioallantoic membrane) test to evaluate the effect of the product on angiogenesis. This test is widely recognized as a robust technique for studying the process of angiogenesis [39]. The product was tested at different concentrations, with dexamethasone used as a positive control. Additionally, a vehicle control group was included to assess the impact of the vehicle used in the formulation of Terasen®, and a naïve control group was treated with saline solution alone.
The results obtained from the study demonstrated that Terasen® exhibited remarkable anti-angiogenic activity. This was evident from the significant and concentration-dependent reduction in the vascular area, quantity, and length.
The anti-angiogenic activity of Terasen® can be attributed to its composition. For example, anthocyanins have been shown to suppress angiogenesis through various mechanisms, including inhibiting H2O2 and reducing VEGF receptor expression in endothelial cells [13]. Other studies have also supported the anti-angiogenic activity of anthocyanins, including those from black carrot (Daucus carota) extract [12] and crowberry (Empetrum nigrum) [40].
The impact of gallic acid on the suppression of new blood vessel growth in tumors can be linked to its effects on the communication between CXCR4/SDF-1 molecules. This communication plays a crucial role in the formation of blood vessels, and gallic acid increases VEGF expression to inhibit this process [41].
Liang et al. [42] showed the impact of gallic acid on osteosarcoma cells and mouse tumor models. The findings demonstrated that gallic acid effectively suppressed cell proliferation, triggered apoptosis, and curbed tumor growth in a manner that was dependent on the dosage. Additionally, the acid was observed to impede the expression of proliferation and angiogenesis markers in tumor tissue.
Ellagic acid has been investigated for its impact on VEGF and PDGF (platelet-derived growth factor) receptor signaling and endothelial and smooth muscle cell migration. The results revealed that ellagic acid inhibited the phosphorylation of VEGFR-2 and PDGFR receptors, blocking subsequent signaling [43].
Fikry et al. [44] conducted a comparative analysis of the efficacy of caffeic acid and ellagic acid versus celecoxib for treating rheumatoid arthritis. The findings revealed that both acids effectively mitigated the severity of arthritis by reducing edema, inflammation, cartilage and bone destruction, and pannus formation. Additionally, they successfully decreased pro-inflammatory molecules and factors related to angiogenesis, including VEGF.
In a study conducted by Metibemu et al. [45], the effects of carotenoid phytochemicals derived from Spondias mombin on the breast carcinoma model in Wistar rats were evaluated. The study focused on the impact of these phytochemicals on angiogenesis and the expression of related genes. The results indicated that treatment with carotenoids led to a significant reduction in the expression of pro-angiogenic genes, including VEGF, VEGFR, EGFR, HIF-1, and MMP-2. Additionally, the carotenoids were found to inhibit the VEGFR-2 kinase domain, suggesting their potential as an anti-angiogenic and anti-proliferative agent.
Quercetin, one of the flavonoids in M. dubia, is associated with reduced blood vessel formation by inhibiting endothelial cell growth, which is thought to involve the inhibition of VEGFR2 [46]. Studies using the CAM model have shown that quercetin can inhibit angiogenic activity and VEGF transcription induced by cancerous cells [47]. Additionally, quercetin conjugated with titanium dioxide nanotubes has significantly decreased blood vessel formation in the CAM model [48]. In mice bearing B16F10 tumors, treatment with this preparation led to decreased levels of phosphorylated STAT3, which is involved in the induction of angiogenesis.
Tocotrienols have also been reported to inhibit angiogenesis. γ-tocotrienol inhibited angiogenesis in the CAM induced by hepatocellular carcinoma by suppressing VEGF [49]. Furthermore, δ-tocotrienol has decreased angiogenesis in mouse dorsal air sacs and the CAM assay [50]. Although the role of geranylgeraniol in angiogenesis has not been extensively explored, it possesses health-promoting properties such as anti-inflammatory and lipid-lowering effects, thereby increasing the overall nutraceutical value of the product [16,51].
Overall, the observed anti-angiogenic activity of Terasen® can be attributed to the combined effects of its bioactive compounds, including anthocyanins, quercetin, tocotrienols, and geranylgeraniol, which exert inhibitory effects on various angiogenesis-related processes and signaling pathways.
5. Conclusions
In this study, we investigated the potential anti-angiogenic properties of a newly developed nutraceutical comprising the dry extracts of E. oleracea and M. dubia, as well as the purified oil of B. orellana and A. aculeatum. The morphological analysis revealed that the nutraceutical is composed of granules. To further characterize the nutraceutical, a phytochemical analysis was conducted to quantify the major compounds present in the formulation. The analysis revealed significant amounts of anthocyanins, flavonoids, δ-tocotrienol, and geranylgeraniol. The results demonstrated a significant and concentration-dependent decrease in angiogenesis in the CAM model.
Author Contributions
Conceptualization, J.C.T.C. and E.L.M.; methodology, E.L.M.; validation, E.L.M. and B.A.M.T.; formal analysis, H.O.C.; investigation, A.V.T.L.T., B.A.M.T. and N.N.D.C.; resources, J.C.T.C. and A.M.F.; writing—original draft preparation, E.L.M.; writing—review and editing, E.L.M. and A.C.M.P.; visualization, L.F.M. and A.N.C.; supervision, J.C.T.C.; project administration, E.L.M. and J.C.T.C.; funding acquisition, J.C.T.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received National Council for Scientific and Technological Development (CNPq) external funding - CNPq Call - No. 68/2022 - Master's and Doctoral Program for Innovation MAI/DAI.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data will be made available upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dai, X.; Cui, S.; Wang, T.; Liu, Q.; Song, H.; Wang, R. Endogenous Opioid Peptides, Endomorphin-1 and -2 and Deltorphin I, Stimulate Angiogenesis in the CAM Assay. Eur J Pharmacol 2008, 579, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.; Belinha, J.; Mangir, N.; MacNeil, S.; Natal Jorge, R. Simulation of the Process of Angiogenesis: Quantification and Assessment of Vascular Patterning in the Chicken Chorioallantoic Membrane. Comput Biol Med 2021, 136, 104647. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Bai, L.; Zhou, J.; Chen, H.; Zhang, L. Sequential Delivery of VEGF, FGF-2 and PDGF from the Polymeric System Enhance HUVECs Angiogenesis in Vitro and CAM Angiogenesis. Cell Immunol 2018, 323, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, H.; Deng, R. Angiogenesis as a Potential Treatment Strategy for Rheumatoid Arthritis. Eur J Pharmacol 2021, 910, 174500. [Google Scholar] [CrossRef] [PubMed]
- Gatne, D.P.; Mungekar, S.; Addepalli, V.; Mohanraj, K.; Ghone, S.A.; Rege, N.N. Development of Collateral Vessels: A New Paradigm in CAM Angiogenesis Model. Microvasc Res 2016, 103, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Two New Applications in the Study of Angiogenesis the CAM Assay: Acellular Scaffolds and Organoids. Microvasc Res 2022, 140. [Google Scholar] [CrossRef]
- Watchararot, T.; Prasongchean, W.; Thongnuek, P. Angiogenic Property of Silk Fibroin Scaffolds with Adipose-Derived Stem Cells on Chick Chorioallantoic Membrane. R Soc Open Sci 2021, 8. [Google Scholar] [CrossRef]
- Gulla, S.; Reddy, V.C.; Araveti, P.B.; Lomada, D.; Srivastava, A.; Reddy, M.C.; Reddy, K.R. Synthesis of Titanium Dioxide Nanotubes (TNT) Conjugated with Quercetin and Its in Vivo Antitumor Activity against Skin Cancer. J Mol Struct 2022, 1249, 131556. [Google Scholar] [CrossRef]
- Nakagawa, K.; Shibata, A.; Yamashita, S.; Tsuzuki, T.; Kariya, J.; Oikawa, S.; Miyazawa, T. In Vivo Angiogenesis Is Suppressed by Unsaturated Vitamin E, Tocotrienol. J Nutr 2007, 137, 1938–1943. [Google Scholar] [CrossRef]
- Oh, S.J.; Kim, O.; Lee, J.S.; Kim, J.A.; Kim, M.R.; Choi, H.S.; Shim, J.H.; Kang, K.W.; Kim, Y.C. Inhibition of Angiogenesis by Quercetin in Tamoxifen-Resistant Breast Cancer Cells. Food Chem Toxicol 2010, 48, 3227–3234. [Google Scholar] [CrossRef]
- Siveen, K.S.; Ahn, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Yap, W.N.; Kumar, A.P.; Fong, C.W.; Tergaonkar, V.; Hui, K.M.; et al. Y-Tocotrienol Inhibits Angiogenesis-Dependent Growth of Human Hepatocellular Carcinoma through Abrogation of AKT/MTOR Pathway in an Orthotopic Mouse Model. Oncotarget 2014, 5, 1897–1911. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Denaro, M.; Barreca, D.; D’Angelo, V.; Germanò, M.P.; Trombetta, D. Polyphenolic Profile and Biological Activities of Black Carrot Crude Extract (Daucus Carota L. Ssp. Sativus Var. Atrorubens Alef.). Fitoterapia 2018, 124, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Stoner, G.D. Anthocyanins and Their Role in Cancer Prevention. Cancer Lett 2008, 269, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.; Bezerra, I.; Ferreira, M.; Soares, L. Spectrophotometric Quantification of Flavonoids in Herbal Material, Crude Extract, and Fractions from Leaves of Eugenia Uniflora Linn. Pharmacognosy Res 2017, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- West, D.C.; Thompson, W.D.; Sells, P.G.; Burbridge, M.F. Angiogenesis Assays Using Chick Chorioallantoic Membrane. Angiogenesis Protocols 2001, 107–129. [Google Scholar] [CrossRef]
- Matias Pereira, A.C.; de Oliveira Carvalho, H.; Gonçalves, D.E.S.; Picanço, K.R.T.; de Lima Teixeira dos Santos, A.V.T.; da Silva, H.R.; Braga, F.S.; Bezerra, R.M.; de Sousa Nunes, A.; Nazima, M.T.S.T.; et al. Co-treatment of Purified Annatto Oil (Bixa Orellana l.) and Its Granules (Chronic®) Improves the Blood Lipid Profile and Bone Protective Effects of Testosterone in the Orchiectomy-induced Osteoporosis in Wistar Rats. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Aires, M. de M. Fisiologia; 5. ed.; Guanabara Koogan: Rio de Janeiro, 2018.
- Šalandová, M.; Hengel, I.A.J.; Apachitei, I.; Zadpoor, A.A.; Eerden, B.C.J.; Fratila-Apachitei, L.E. Inorganic Agents for Enhanced Angiogenesis of Orthopedic Biomaterials. Adv Healthc Mater 2021, 10, 2002254. [Google Scholar] [CrossRef]
- Golan, D.E. Princípios de Farmacologia - A Base Fisiopatológica Da Farmacoterapia ; 2. ed.; Guanabara Koogan, 2009.
- Cha, S.; Kim, H.-G.; Jang, H.; Lee, J.; Chao, T.; Baek, N.-I.; Song, I.-S.; Lee, Y.M. Steppogenin Suppresses Tumor Growth and Sprouting Angiogenesis through Inhibition of HIF-1α in Tumors and DLL4 Activity in the Endothelium. Phytomedicine 2023, 108, 154513. [Google Scholar] [CrossRef]
- Zhu, T.; Xie, W.; Wang, L.; Jin, X.; Meng, X.; Sun, G.; Sun, X. Notoginsenoside R1 Activates the NAMPT-NAD+-SIRT1 Cascade to Promote Postischemic Angiogenesis by Modulating Notch Signaling. Biomedicine & Pharmacotherapy 2021, 140, 111693. [Google Scholar] [CrossRef]
- Klang, V.; Valenta, C.; Matsko, N.B. Electron Microscopy of Pharmaceutical Systems. Micron 2013, 44, 45–74. [Google Scholar] [CrossRef]
- Szulc, K.; Lenart, A. Surface Modification of Dairy Powders: Effects of Fluid-Bed Agglomeration and Coating. Int Dairy J 2013, 33, 55–61. [Google Scholar] [CrossRef]
- Liu, Y.; Scharf, D.; Graule, T.; Clemens, F.J. Granulation Processing Parameters on the Mechanical Properties of Diatomite-Based Porous Granulates. Powder Technol 2014, 263, 159–167. [Google Scholar] [CrossRef]
- Kharb, V.; Saharan, V.A.; Kharb, V.; Jadhav, H.; Purohit, S. Formulation and Evaluation of Lipid Based Taste Masked Granules of Ondansetron HCl. European Journal of Pharmaceutical Sciences 2014, 62, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Bot, F.; Cossuta, D.; O’Mahony, J.A. Inter-Relationships between Composition, Physicochemical Properties and Functionality of Lecithin Ingredients. Trends Food Sci Technol 2021, 111, 261–270. [Google Scholar] [CrossRef]
- Van Nieuwenhuyzen, W.; Tomás, M.C. Update on Vegetable Lecithin and Phospholipid Technologies. European Journal of Lipid Science and Technology 2008, 110, 472–486. [Google Scholar] [CrossRef]
- Bueschelberger, H.-G. Lecithins. Emulsifiers in Food Technology 2007, 1–39. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Zhang, T.; Wang, C.; Huang, Z.; Luo, X.; Deng, Y. A Review on Phospholipids and Their Main Applications in Drug Delivery Systems. Asian J Pharm Sci 2015, 10, 81–98. [Google Scholar] [CrossRef]
- Lisboa, C.R.; Oliveira, M. do S.P. de; Chisté, R.C.; Carvalho, A.V. Compostos Bioativos e Potencial Antioxidante de Diferentes Acessos de Euterpe Oleracea e Euterpe Precatoria Do Banco Ativo de Germoplasma de Açaí. Research, Society and Development 2022, 11, e428111234824. [CrossRef]
- Costa, R.; Azevedo, D.; Barata, P.; Soares, R.; Guido, L.F.; Carvalho, D.O. Antiangiogenic and Antioxidant In Vitro Properties of Hydroethanolic Extract from Açaí (Euterpe Oleracea) Dietary Powder Supplement. Molecules 2021, 26, 2011. [Google Scholar] [CrossRef]
- Kang, J.; Xie, C.; Li, Z.; Nagarajan, S.; Schauss, A.G.; Wu, T.; Wu, X. Flavonoids from Acai (Euterpe Oleracea Mart.) Pulp and Their Antioxidant and Anti-Inflammatory Activities. Food Chem 2011, 128, 152–157. [Google Scholar] [CrossRef]
- Fidelis, M.; Santos, J.S.; Escher, G.B.; Vieira do Carmo, M.; Azevedo, L.; Cristina da Silva, M.; Putnik, P.; Granato, D. In Vitro Antioxidant and Antihypertensive Compounds from Camu-Camu (Myrciaria Dubia McVaugh, Myrtaceae) Seed Coat: A Multivariate Structure-Activity Study. Food and Chemical Toxicology 2018, 120, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.; Oliveira, T.; Mattietto, R.; Nascimento, W.; Lopes, A. Bioactive Compounds in the Peel of Camu Camu Genotypes from Embrapa’s Active Germplasm Bank. Food Science and Technology (Brazil) 2018, 38, 67–71. [Google Scholar] [CrossRef]
- Frega, N.; Mozzon, M.; Bocci, F. Identification and Estimation of Tocotrienols in the Annatto Lipid Fraction by Gas Chromatography-Mass Spectrometry. JAOCS, Journal of the American Oil Chemists’ Society 1998, 75, 1723–1727. [Google Scholar] [CrossRef]
- Costa, C.K.; Silva, C.B.; Lordello, A.L.L.; Zanin, S.M.W.; Dias, J.F.G.; Miguel, M.D.; Miguel, O.G. Identificação de δ Tocotrienol e de Ácidos Graxos No Óleo Fixo de Urucum (Bixa Orellana Linné). Revista Brasileira de Plantas Medicinais 2013, 15, 508–512. [Google Scholar] [CrossRef]
- de Oliveira Carvalho, H.; Sauma, A.L.R.; do Nascimento, A.L.; de Lima Teixeira, A.V.T.; Gonçalves, D.E.S.; Gomes, L.; da Costa Furtado, G.; da Silva, H.R.; de Souza, G.C.; Pereira, A.C.M.; et al. Intramuscular Compatibility of an Injectable Anti-Inflammatory Nanodispersion from a Standardized Bixa Orellana Oil (Chronic®): A Toxicological Study in Wistar Rats. Inflammopharmacology 2023. [Google Scholar] [CrossRef]
- Vardanega, R.; Nogueira, G.C.; Nascimento, C.D.O.; Faria-Machado, A.F.; Meireles, M.A.A. Selective Extraction of Bioactive Compounds from Annatto Seeds by Sequential Supercritical CO2 Process. J Supercrit Fluids 2019, 150, 122–127. [Google Scholar] [CrossRef]
- Wan, X.; Zhang, Z.; Rui, L.; Wilson, R.S.; Wu, X.; Liu, X. An Improved In Vivo Angiogenesis Model of Chicken Chorioallantoic Membranes in Surrogate Shells Revealed the Pro-Angiogenesis Effects of Chylomicrons. Vasc Cell 2019, 11, 1. [Google Scholar] [CrossRef]
- Bae, H.S.; Kim, H.J.; Jeong, D.H.; Hosoya, T.; Kumazawa, S.; Jun, M.; Kim, O.Y.; Kim, S.W.; Ahn, M.R. In Vitro and In Vivo Antiangiogenic Activity of Crowberry (Empetrum Nigrum Var. Japonicum). 2016, 11, 503–506. [CrossRef]
- Lu, Y.; Jiang, F.; Jiang, H.; Wu, K.; Zheng, X.; Cai, Y.; Katakowski, M.; Chopp, M.; To, S.-S.T. Gallic Acid Suppresses Cell Viability, Proliferation, Invasion and Angiogenesis in Human Glioma Cells. Eur J Pharmacol 2010, 641, 102–107. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, X.; Li, H.; Tao, Y.; Tao, L.; Yang, Z.; Zhou, X.; Shi, Z.; Tao, H. Gallic Acid Induces the Apoptosis of Human Osteosarcoma Cells In Vitro and In Vivo via the Regulation of Mitogen-Activated Protein Kinase Pathways. Cancer Biother Radiopharm 2012, 27, 701–710. [Google Scholar] [CrossRef]
- Labrecque, L.; Lamy, S.; Chapus, A.; Mihoubi, S.; Durocher, Y.; Cass, B.; Bojanowski, M.W.; Gingras, D.; Béliveau, R. Combined Inhibition of PDGF and VEGF Receptors by Ellagic Acid, a Dietary-Derived Phenolic Compound. Carcinogenesis 2005, 26, 821–826. [Google Scholar] [CrossRef]
- Fikry, E.M.; Gad, A.M.; Eid, A.H.; Arab, H.H. Caffeic Acid and Ellagic Acid Ameliorate Adjuvant-Induced Arthritis in Rats via Targeting Inflammatory Signals, Chitinase-3-like Protein-1 and Angiogenesis. Biomedicine & Pharmacotherapy 2019, 110, 878–886. [Google Scholar] [CrossRef]
- Metibemu, D.S.; Akinloye, O.A.; Akamo, A.J.; Okoye, J.O.; Ojo, D.A.; Morifi, E.; Omotuyi, I.O. VEGFR-2 Kinase Domain Inhibition as a Scaffold for Anti-Angiogenesis: Validation of the Anti-Angiogenic Effects of Carotenoids from Spondias Mombin in DMBA Model of Breast Carcinoma in Wistar Rats. Toxicol Rep 2021, 8, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Morbidelli, L. Polyphenol-Based Nutraceuticals for the Control of Angiogenesis: Analysis of the Critical Issues for Human Use. Pharmacol Res 2016, 111, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Kim, O.; Lee, J.S.; Kim, J.-A.; Kim, M.R.; Choi, H.S.; Shim, J.-H.; Kang, K.W.; Kim, Y.C. Inhibition of Angiogenesis by Quercetin in Tamoxifen-Resistant Breast Cancer Cells. Food and Chemical Toxicology 2010, 48, 3227–3234. [Google Scholar] [CrossRef]
- Gulla, S.; Reddy, V.C.; Araveti, P.B.; Lomada, D.; Srivastava, A.; Reddy, M.C.; Reddy, K.R. Synthesis of Titanium Dioxide Nanotubes (TNT) Conjugated with Quercetin and Its in Vivo Antitumor Activity against Skin Cancer. J Mol Struct 2022, 1249, 131556. [Google Scholar] [CrossRef]
- Siveen, K.S.; Ahn, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Yap, W.N.; Kumar, A.P.; Fong, C.W.; Tergaonkar, V.; Hui, K.M.; et al. γ-Tocotrienol Inhibits Angiogenesis-Dependent Growth of Human Hepatocellular Carcinoma through Abrogation of AKT/MTOR Pathway in an Orthotopic Mouse Model. Oncotarget 2014, 5, 1897–1911. [Google Scholar] [CrossRef]
- Nakagawa, K.; Shibata, A.; Yamashita, S.; Tsuzuki, T.; Kariya, J.; Oikawa, S.; Miyazawa, T. In Vivo Angiogenesis Is Suppressed by Unsaturated Vitamin E, Tocotrienol. J Nutr 2007, 137, 1938–1943. [Google Scholar] [CrossRef]
- Batista, M.A.; de Lima Teixeira dos Santos, A.V.T.; do Nascimento, A.L.; Moreira, L.F.; Souza, I.R.S.; da Silva, H.R.; Pereira, A.C.M.; da Silva Hage-Melim, L.I.; Carvalho, J.C.T. Potential of the Compounds from Bixa Orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols. Molecules 2022, 27, 1584. [Google Scholar] [CrossRef]
Figure 1.
Schematics showing the process to evaluate the embryos’ CAM.
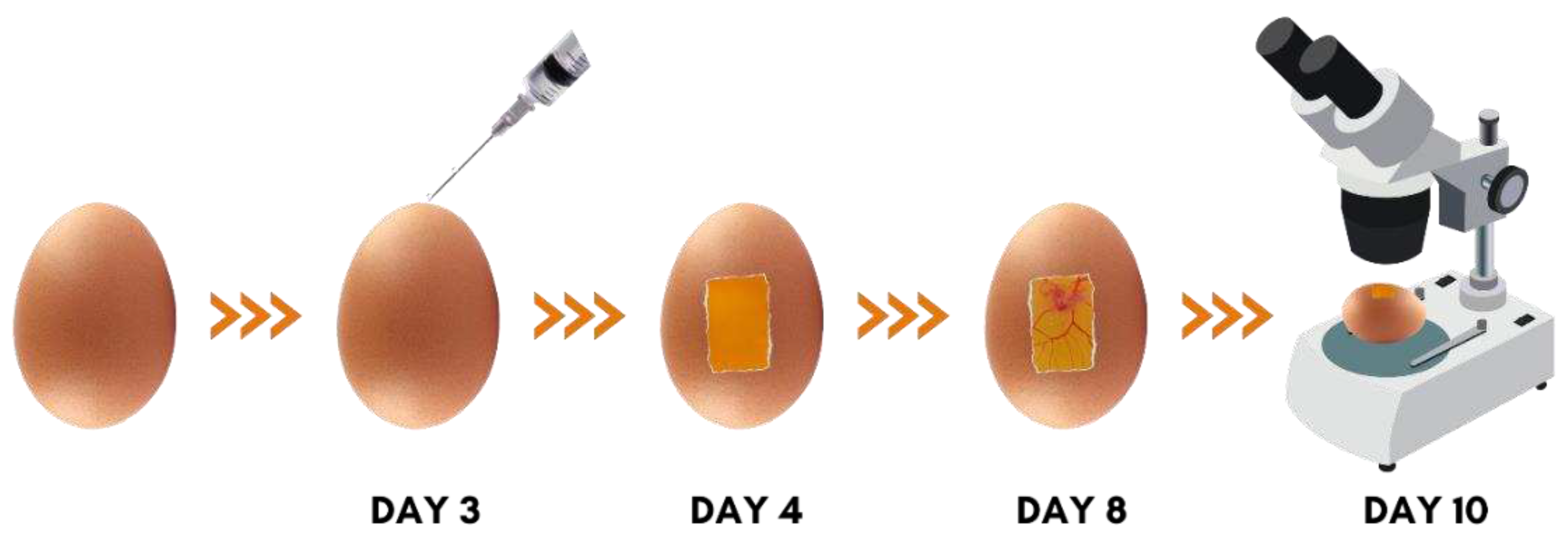
Figure 2.
Scanning Electron Microscopy (SEM) analysis from Terasen® granules.

Figure 3.
Phytochemical analysis of Terasen®. A: Calibration curves of total flavonoids; B: Calibration curves of geranylgeraniol; C: Calibration curves of δ-tocotrienol.
Figure 3.
Phytochemical analysis of Terasen®. A: Calibration curves of total flavonoids; B: Calibration curves of geranylgeraniol; C: Calibration curves of δ-tocotrienol.
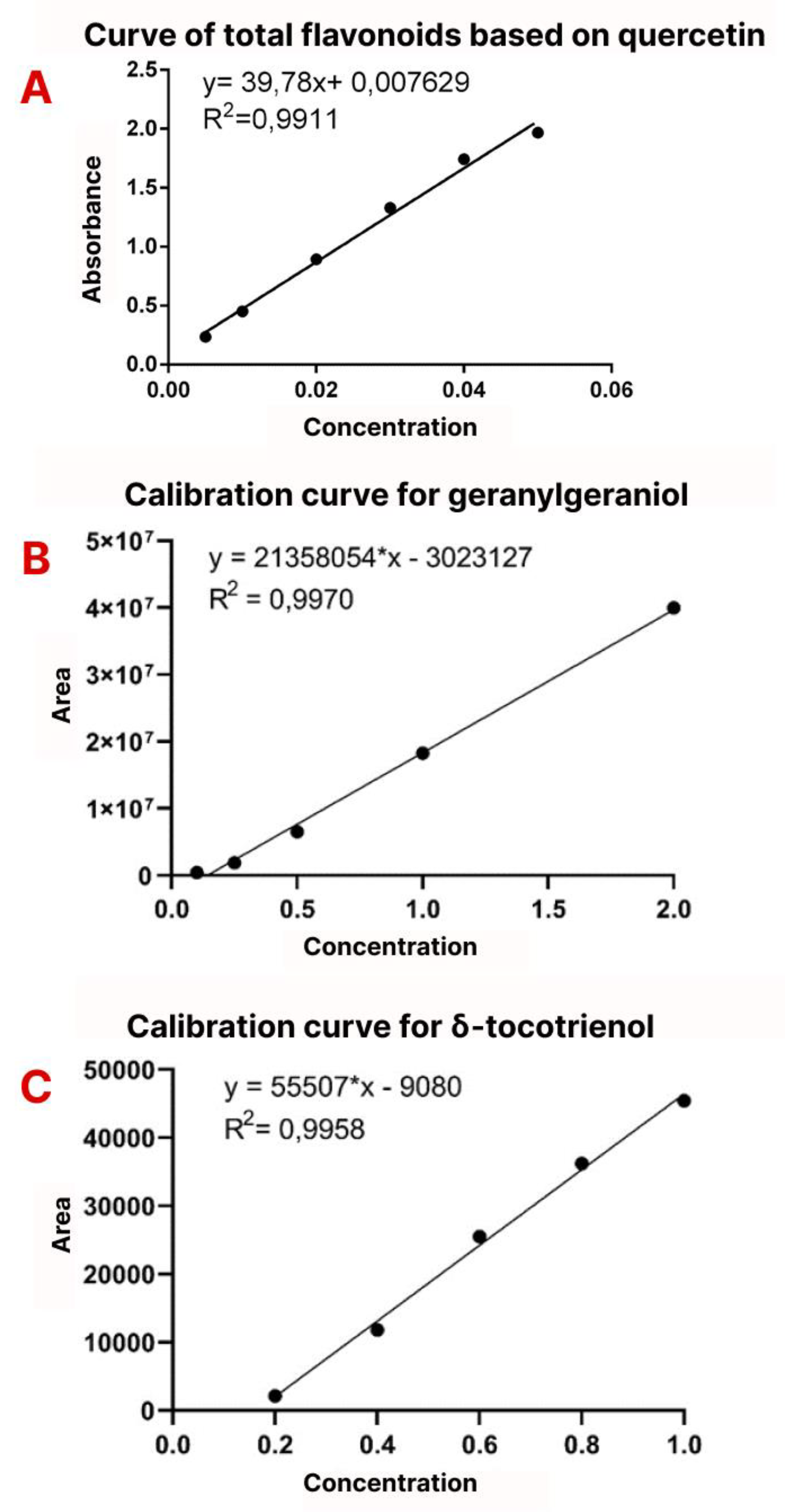
Figure 4.
Phytochemical analysis of Terasen®. A: GC chromatogram of geranylgeraniol. B: GC chromatogram of δ-tocotrienol.
Figure 4.
Phytochemical analysis of Terasen®. A: GC chromatogram of geranylgeraniol. B: GC chromatogram of δ-tocotrienol.
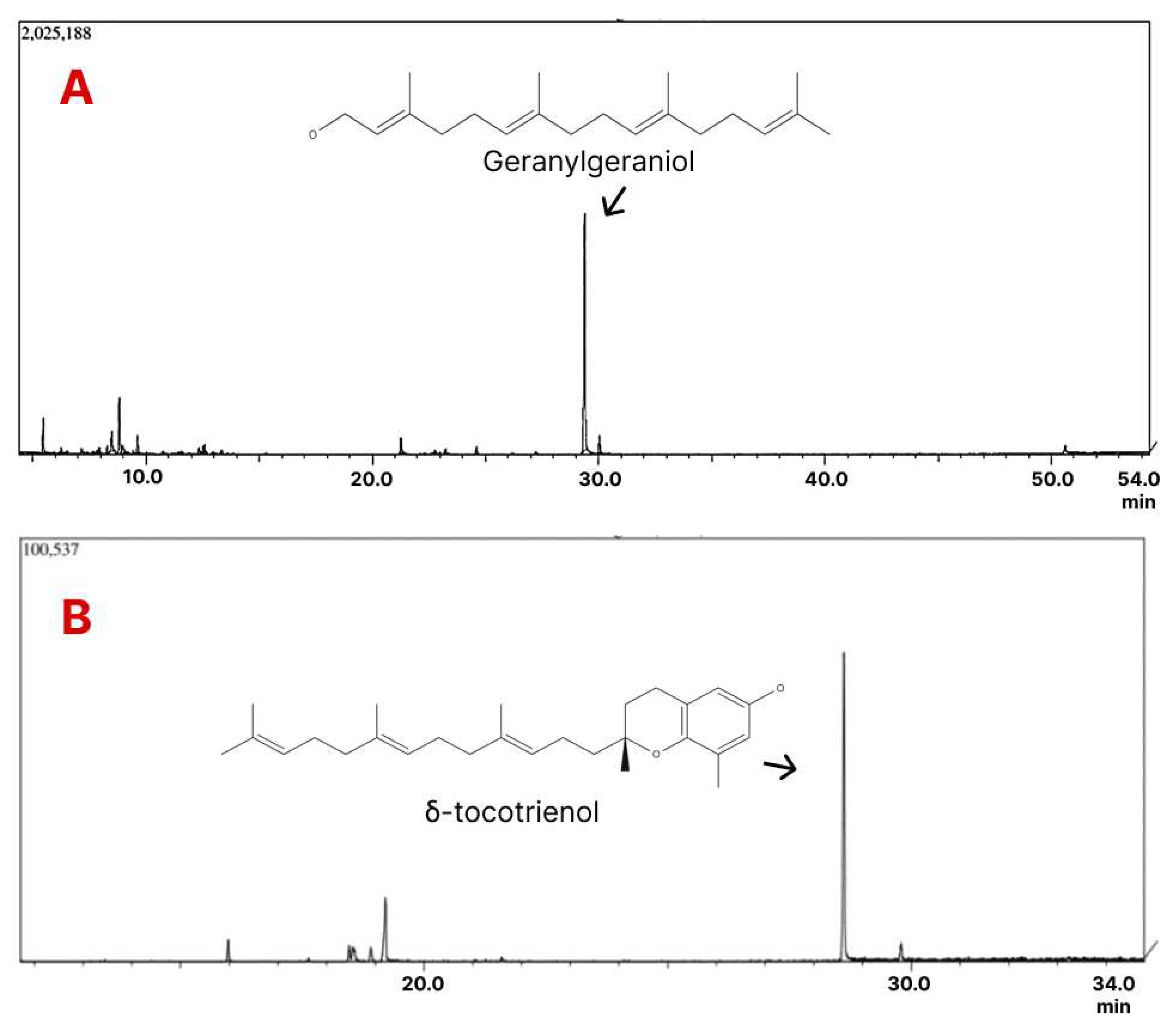
Figure 5.
CAM’s blood vessels network before and after treatments with saline solution 0.9% (CON), 90% ethanol (ETH), dexamethasone 4 mg/ml (DEX), and Terasen® (solubilized in 90% ethanol) at different concentrations – 0.5 mg/ml (TERA 0.5), 1 mg/ml (TERA 1), 2 mg/ml (TERA 2), and 3 mg/ml (TERA 3).
Figure 5.
CAM’s blood vessels network before and after treatments with saline solution 0.9% (CON), 90% ethanol (ETH), dexamethasone 4 mg/ml (DEX), and Terasen® (solubilized in 90% ethanol) at different concentrations – 0.5 mg/ml (TERA 0.5), 1 mg/ml (TERA 1), 2 mg/ml (TERA 2), and 3 mg/ml (TERA 3).
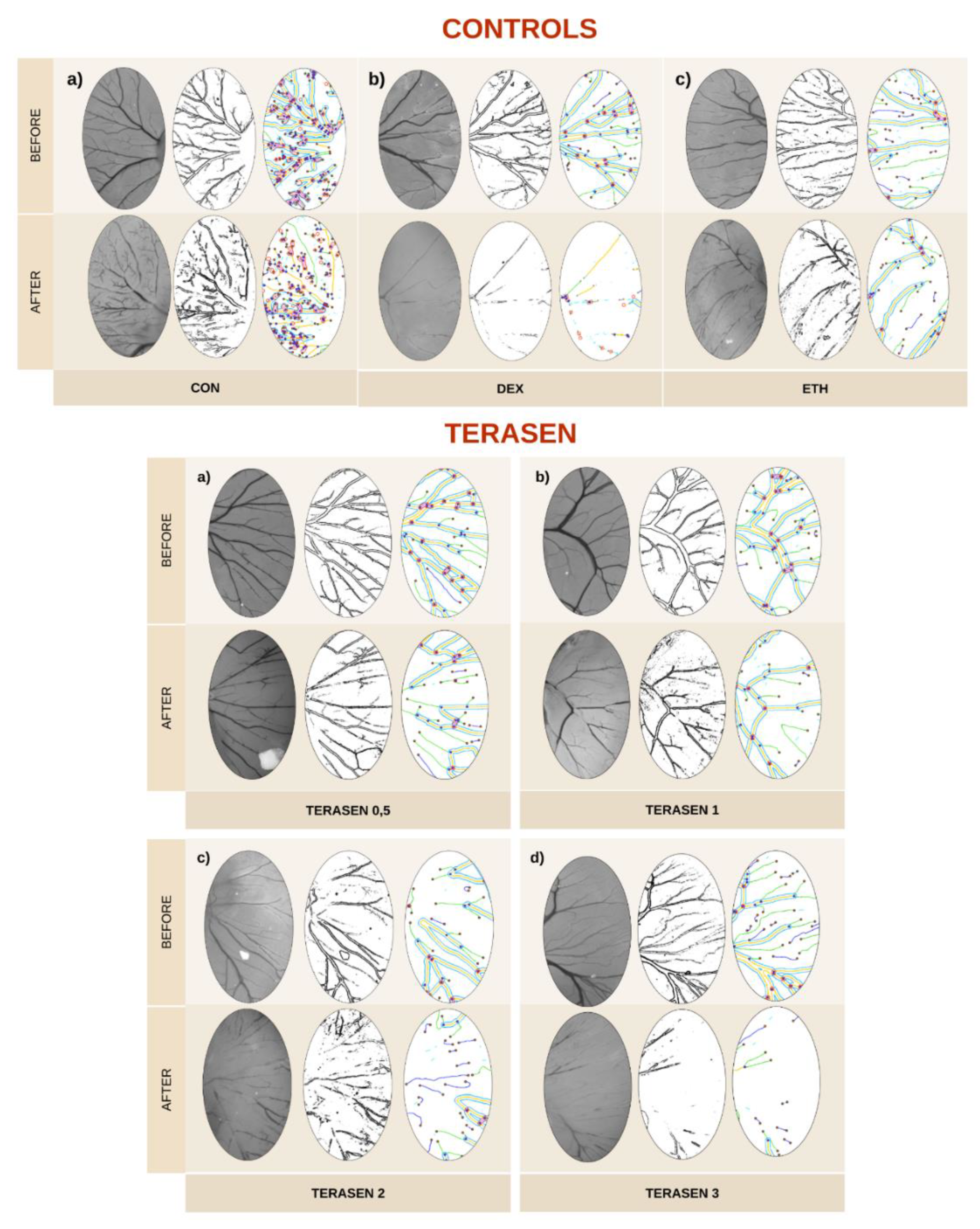
Figure 6.
Effects of the treatments with saline solution 0.9% (CON), 90% ethanol (ETH), dexamethasone 4 mg/ml (DEX), and Terasen® (solubilized in 90% ethanol) at different concentrations – 0.5 mg/ml (TERA 0.5), 1 mg/ml (TERA 1), 2 mg/ml (TERA 2), and 3 mg/ml (TERA 3) on angiogenic parameters assessed: Vascular area, vessel length variation, and vessel number variation (from top to bottom). *: p < 0.05 vs naïve control (ANOVA followed by Tukey posthoc test).
Figure 6.
Effects of the treatments with saline solution 0.9% (CON), 90% ethanol (ETH), dexamethasone 4 mg/ml (DEX), and Terasen® (solubilized in 90% ethanol) at different concentrations – 0.5 mg/ml (TERA 0.5), 1 mg/ml (TERA 1), 2 mg/ml (TERA 2), and 3 mg/ml (TERA 3) on angiogenic parameters assessed: Vascular area, vessel length variation, and vessel number variation (from top to bottom). *: p < 0.05 vs naïve control (ANOVA followed by Tukey posthoc test).
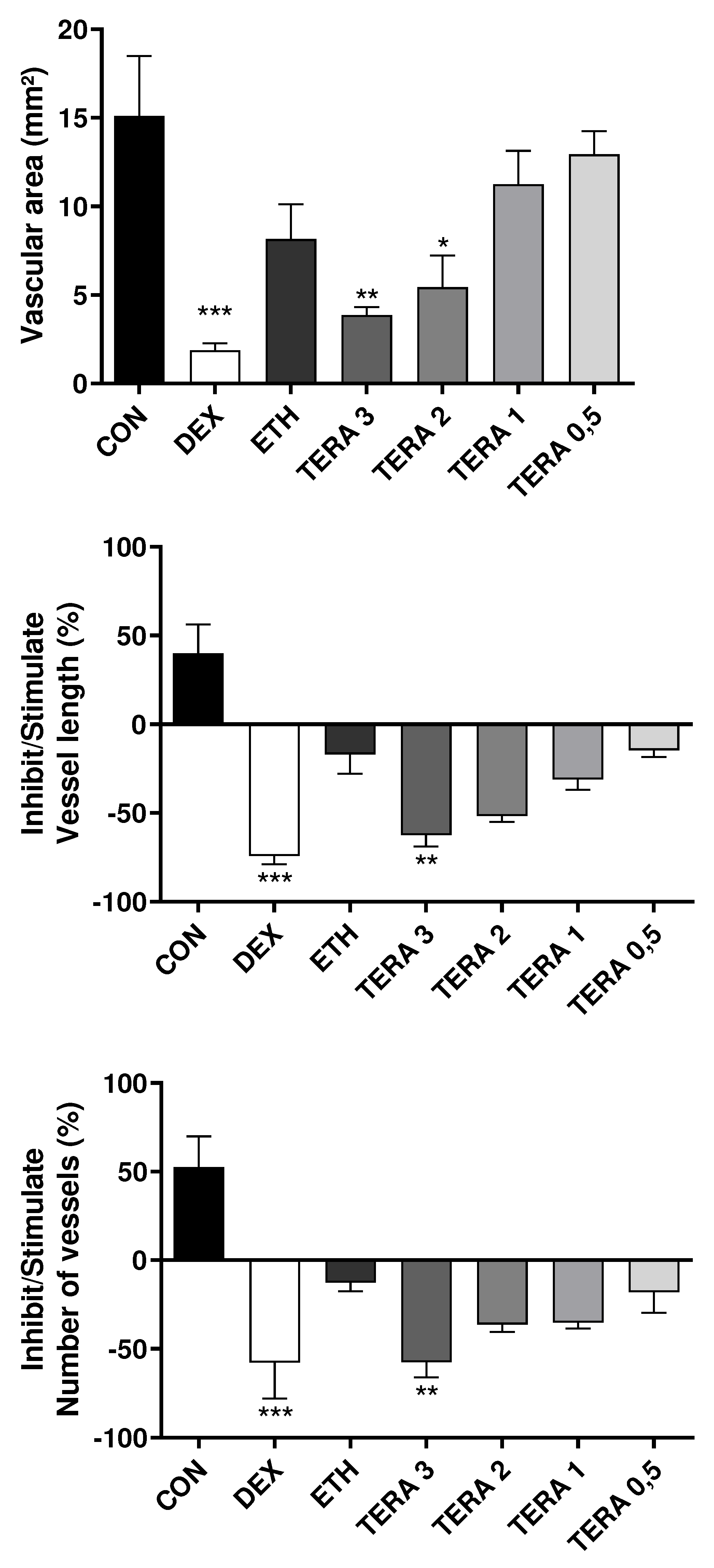
Figure 7.
Schematics of the angiogenic process. A: tip cells and stalk cells interaction. B: Formation of the new vessel. 1. Angiogenic stimulus. 2. The activation of matrix metalloproteinases (MMPs) leads to the degradation of the extracellular matrix. 3. Formation of tip cells. 4. Proliferation and migration. 5. Lumen formation. 6. Maturation and blood flow.
Figure 7.
Schematics of the angiogenic process. A: tip cells and stalk cells interaction. B: Formation of the new vessel. 1. Angiogenic stimulus. 2. The activation of matrix metalloproteinases (MMPs) leads to the degradation of the extracellular matrix. 3. Formation of tip cells. 4. Proliferation and migration. 5. Lumen formation. 6. Maturation and blood flow.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



