
Submitted:
30 September 2023
Posted:
02 October 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
SARS-CoV-2 can trigger autoimmune central nervous system (CNS) diseases in genetically susceptible individuals, a mechanism poorly understood. Molecular mimicry (MM) has been identified in other viral diseases as potential triggers of autoimmune CNS events. This study investigated if MM is the process through which SARS-CoV-2 induce the breakdown of immune tolerance. The frequency of autoimmune CNS disorders was evaluated in a prospective cohort with patients admitted in the COVID-19 Intense Care Unity (ICU) in Rio de Janeiro. Then, an in silico analysis was performed to identify the conserved regions which share high identity between SARS-Cov-2 anti-gens and human proteins. The sequences with significant identity and antigenic properties were then assessed for their binding capacity to HLA subtypes. Of the 112 patients included, 3 were classified as having an autoimmune disorder. A total of eleven combinations had significant linear and three-dimensional overlap. NMDAR1, MOG and MPO were the self-antigens with more significant combinations, followed by GAD65. All sequences presented at least one epitope with strong or intermediate binding capacity to the HLA subtypes selected. This study underscores the possibility that CNS autoimmune attacks observed in COVID-19 patients, including those in our population, could be driven by MM in genetically predisposed individuals.
Keywords:
COVID-19
; SARS-CoV-2
; Molecular mimicry
; immune tolerance
; autoimmune disorders
; central nervous system.
1. Introduction
The recent emergence of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) virus, responsible for the disease caused by the novel coronavirus (COVID19), has already affected nearly seven hundred million individuals worldwide, with 6,919,573 deaths until September 2023 [1]. As evidenced in recent years, the occurrence of viral epidemics characterized by unpredictable clinical outcomes has been highly frequent on a global scale [2]. Therefore, the end of the public health emergency of international concern declared on May 5, 2023, by the World Health Organization (WHO) does not mean that COVID-19 has ceased to be a serious public health problem. Like in other viral infections [3]–[5], extensive studies have provided strong evidence of the frequent occurrence of neurological manifestations that resemble clinical patterns seen in autoimmune para- or post-infectious diseases triggered by SARS-CoV-2 [6], [7]. Moreover, the hypothesis that viral infections can act as triggers for the development of autoimmune diseases is not a new one [8]. Due to the association between infection with other coronaviruses and autoimmunity, it is reasonable to assume that there is a connection between SARS-CoV-2 infection and certain autoimmune diseases that will be diagnosed later [9]–[11].
It is already well discussed in the literature that viral diseases have been identified as potential triggers of inflammatory demyelinating diseases (IDD) and autoimmune encephalitis (AE) [12], [13]. Numerous demyelinating disorders such as multiple sclerosis (MS), Neuromyelitis Optica Spectrum Diseases (NMOSD), Acute Disseminated Encephalomyelitis (ADEM), Myelitis and Myelin Oligodendrocyte Glycoprotein Antibody-associated Disease (MOGAD) have been described as post and para-infectious complications of COVID-19 [14], as well as the most common forms of AE following SARS-CoV-2 infection are limbic encephalitis and NMDAR receptor antibody-mediated (anti-NMDAR) encephalitis[15]. Important studies have identified the presence of autoantibodies associated with-glutamic acid decarboxylase 65-kilodalton isoform (anti-GAD65) [16], myelin oligodendrocyte glycoprotein (anti-MOG) and others [17]–[20].
Several hypotheses have been proposed to explain the molecular basis of the loss of immune tolerance and induction of autoimmune mechanisms, including hyperinflammation syndrome caused by SARS-CoV-2, molecular mimicry (MM) by viral proteins, immune cell activation through bystander effect, the release of autoantigens from virus-damaged tissues, lymphocyte activation mediated by superantigens, and epitope spreading [21]–[24]. Some recent studies on MM have shown similarities between SARS-CoV-2 protein sequences and human proteins found in multiple organs/tissues (neurological, vascular, and cardiac), indicating the potential for cross-reactive immune recognition of these regions by T cells and antibodies produced by B cells [22], [25]–[27]. However, the true spectrum of autoimmune conditions, their pathophysiology, prevalence, and the risk of their development in individuals after SARS-CoV-2 infection remain unknown, representing just the tip of the iceberg.
In this study, we intend to investigate the potential role of MM between SARS-CoV-2 antigens and human autoantigens of CNS autoimmune diseases.
2. Materials and Methods
2.1. Study population
A prospective cohort study was performed with patients admitted in the COVID-19 Intense Care Unity (ICU) of Clementino Fraga Filho University Hospital in Rio de Janeiro, RJ, Brazil. This work was approved by the National Council for Ethics in Research (CAAE 33659620.1.1001.5258) and accompanied afterward on the post-COVID ambulatory of the same hospital. All subjects signed an informed consent agreeing to participate in this research. From 2020 to 2022, COVID-19 patients were evaluated with complete physical and neurological examinations and searched for CNS autoimmune diseases. They were also actively asked for long COVID symptoms, with focus on cognitive impairment. Besides, all patients were assessed for objective alterations with the neurocognitive battery tests: Symbol Digit Modalities Test (SDMT) and Montreal Cognitive Assessment (MoCA). Patients with previously known CNS autoimmune diseases or neurocognitive disturbances were excluded from the study.
2.2. Linear sequence analysis
Peptide sharing between SARS-CoV-2 antigens and autoantigens was analyzed in accordance with França et al (2023). Briefly, a viral polyprotein library was constructed using the major viral antigens reported in the literature and protein sequences available in NCBI Protein Reference Sequences (https://www.ncbi.nlm.nih.gov/protein) (Table 1). An extended research was conducted to build an autoantigen library from UniProtKB Database (www.uniprot.org/ ), based on results from PubMed of autoantigen related to demyelinating brain diseases and autoimmune encephalitis (Table 2).
The sequence alignment was done using EMBOSS WATER (https://www.ebi.ac.uk/Tools/psa/emboss_ water/), an online server that uses the Smith-Waterman algorithm (modified for speed enhancements) to calculate the local alignment of two sequences, narrowing down the regions with more identity [28]. The chosen regions were analyzed for linear homology between the identified human proteins to SARS-COV-2 proteins using BLASTp (https://blast.ncbi.nlm.nih.gov/Blast.cgi) [29]. We used the default BLASTp algorithm parameters to consider a significant result [30].
2.3. Three-dimensional comparative modelling
The combinations that shared significant linear identity according to BLASTp, were them investigated for three-dimensional similarities. The three-dimensional models were built using the Swiss-Model, an online modeling server (https://swissmodel.expasy.org/). The template modeling scores (TM-scores), and root mean square deviation (RMSD) of the SARS-CoV-2 antigens and autoantigens three-dimensional overlap were calculated using TM-Align (https://seq2fun.dcmb.med.umich.edu//TM-align/), an algorithm for sequence-independent protein structure comparisons. TM-align first generates optimized residue-to-residue alignment based on structural identity using heuristic dynamic programming iterations. The TM-score value scales the structural identity varying from 0.0 to 1.0, where scores below 0.3 correspond to randomly chosen unrelated proteins, while those higher than 0.5 assume generally the same fold between two structures, based on the Protein Data Bank (PDB) [31]. The RMSD considers the root-mean-square distance between corresponding residues and is calculated after an optimal rotation of one structure to another.
2.4. Antigenic prediction
To confirm whether the SARS-CoV-2 sequences studied have antigenic properties, VaxiJen version 2.0 (http://www.ddgpharmfac.net/vaxijen/VaxiJen/VaxiJen.html) was used. A threshold antigenic score of 0.4 was defined to filter probable non-antigenic sequences. Vaxijen server performs alignment-independent prediction, which is based on auto cross-covariance (ACC) transformation of protein sequences into uniform vectors of principal amino acid properties.
2.5. Search for potential T cell epitopes
The sequences with significant TM-Score and antigenic properties were used as the input in a neural network–based algorithm to predict T cell epitopes showing binding capacity to HLA subtypes using the Immune Epitope Database and Analysis (IEDB) major histocompatibility complex-I (MHC-I) Binding Predictions (http://tools.iedb.org/mhci/) and major histocompatibility complex-II (MHC-II) Binding Predictions (http://tools.iedb.org/mhcii/) resource. This approach enabled the distinction of T cell epitopes recognized by HLA. As HLAs exhibit high polymorphism, we choose HLAs variants from MHC-I and MHC-II, with known associations with CNS autoimmune diseases. The representatives used were HLA-I A*31:01, HLA-I B*07:02, HLA-II DRB1*1501, HLA-II DQA1*0102-HLA-II DQB1*0602 and HLA-II DRB1∗03:01.
The prediction method used was SMM-align. The predicted output is given in units of Half maximal inhibitory concentration (IC50nM). Therefore, a lower number indicates higher affinity. As a rough guideline, peptides with IC50values <50 nM are considered high affinity, <500 nM intermediate affinity and <5000 nM low affinity. Since most known epitopes have high or intermediate affinity, we only considered a noteworthy result the epitopes with <500 nM [32].
3. Results
3.1. Study population
A total of 112 patients were evaluated. Mean age was 65.95 (17-95) and 58 (51.78%) were female. Among them, 3 were classified as having IDD. As our population consisted mainly of patient from the first and second wave of the COVID-19 pandemic and all of them hospitalized on the ICU, 51 (45.53%) patients progressed to death during or nearly after the hospitalization. The patients with IDD survived and remain with controlled disease one year after the episodes. Among the survivors, 32 patients (52.45%) presented cognitive complains during long COVID. Regarding the neurocognitive battery, 15 (24.59%) patients had cognitive impairment according to the two test results, while 20 had alteration only in SDMT and 33 had alteration only in MoCA.
3.1.1. Patient 1
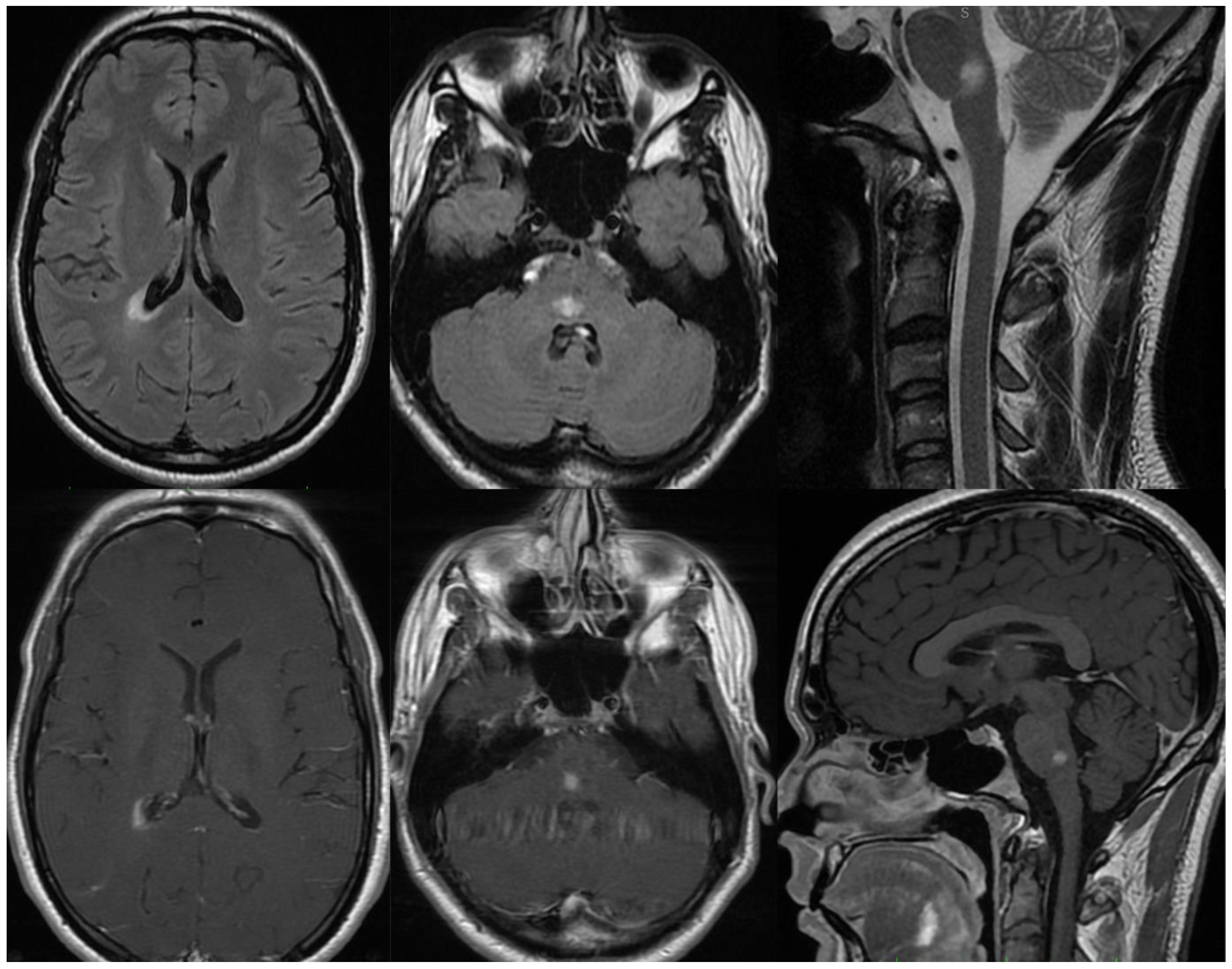
Patient 1, male, 24 years-old, with history of IDD in the family (Mother diagnosed with MS), was admitted to the hospital presenting paresthesia in upper and lower limb on the left side. The Magnetic Resonance Imaging (MRI) showed two white-matter lesions hyperintense in T2/FLAIR, with gadolinium enhancement, one on the periventricular region and one on the medullary bulb transition (Figure 1). Cerebrospinal fluid (CSF) exam was negative for infections, including SARS-CoV-2, and oligoclonal bands, with normal cell and protein count. Serum research was positive for anti-MOG abs through flow cytometry and negative for anti-AQP4 abs. Screening for metabolic and other autoimmune diseases was negative. Even asymptomatic, as part of the hospital protocol, he was tested for COVID-19 with Polymerase Chain Reaction (PCR) test, that was positive. He was treated with intravenous glucocorticoids, evolving with complete recovery. On the long COVID assessment, he presented new symptoms and new lesions on the MRI, being diagnosed with MOGAD.
Figure 1.
MRI of Patient 1. Two lesions hyperintense in T2/FLAIR, one on the periventricular region and one on the medullary bulb transition.
Figure 1.
MRI of Patient 1. Two lesions hyperintense in T2/FLAIR, one on the periventricular region and one on the medullary bulb transition.
3.1.2. Patient 2
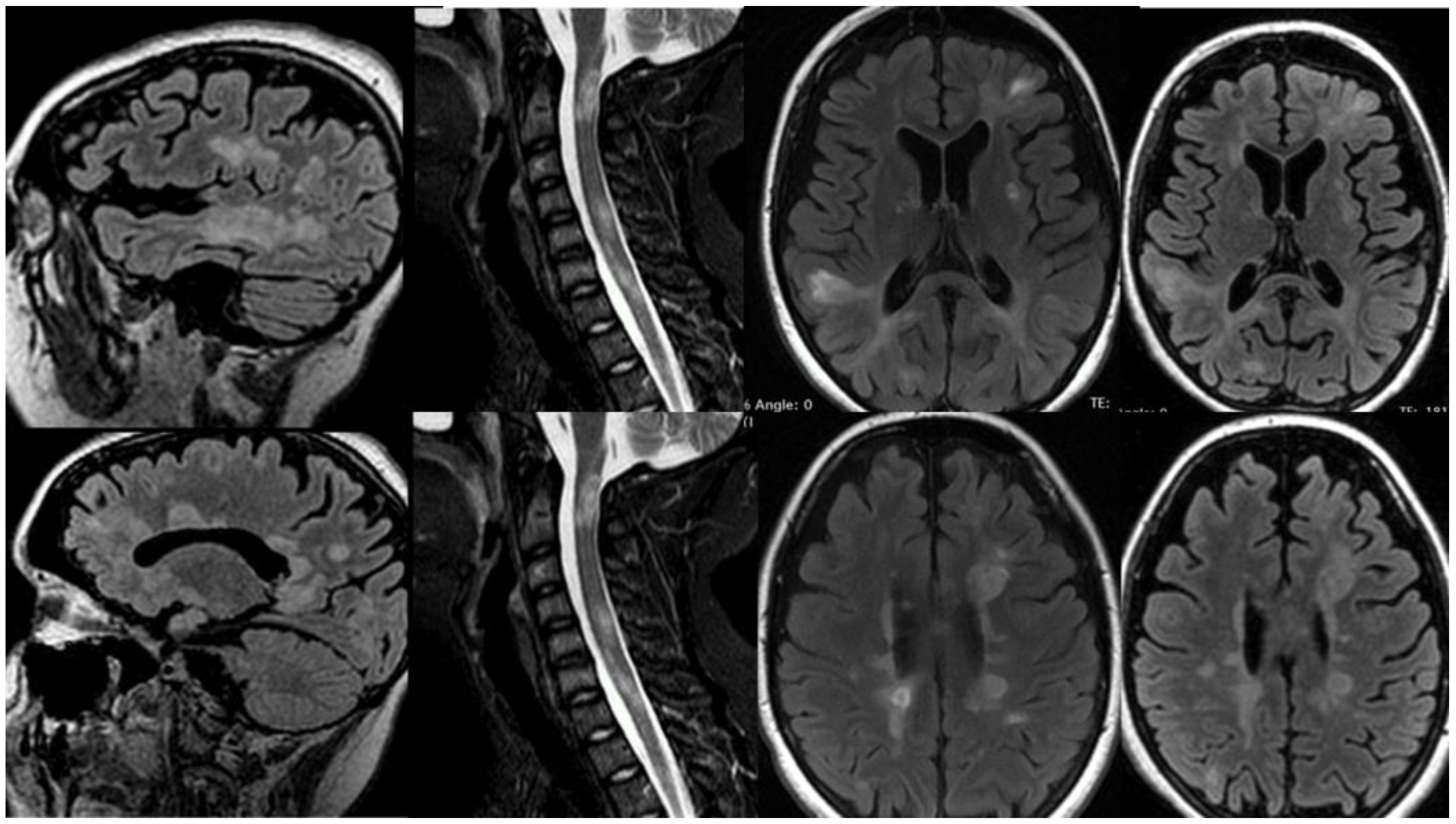
Patient 2, female, 19 years-old, evolved twenty days after SARS-CoV-2 m-RNA vaccination with subacute paresthesia in hands and feet, followed by urinary incontinence, visual disturbance, mental confusion, appendicular ataxia, progressive tetraparesis, and coma within days. Her Expanded Disability Status Scale (EDSS) during the acute state was 9.5. The MRI showed countless white-matter lesions hyperintense in T2/FLAIR, several with gadolinium enhancement (Figure 2). CSF exam was positive for oligoclonal bands, and negative for infections, including SARS-CoV-2. Screening for autoimmune diseases, including MOG and AQP4 abs, and metabolic diseases were negative. First, she received a diagnosis of ADEM following COVID-19 vaccination and was treated with pulse therapy with glucocorticoids, partially recovering from the attack. Nonetheless, after 6 months she once again evolved with a new aggressive demyelinating event, being diagnosed with MS and treated with Natalizumab. The patient stabilized with the treatment, and, after a year, she recovered nearly a hundred percent (EDSS 3.0).
Figure 2.
MRI of Patient 2. Countless white-matter lesions hyperintense on brain and spinal cord in T2/FLAIR, several with gadolinium enhancement.
Figure 2.
MRI of Patient 2. Countless white-matter lesions hyperintense on brain and spinal cord in T2/FLAIR, several with gadolinium enhancement.
3.1.3. Patient 3
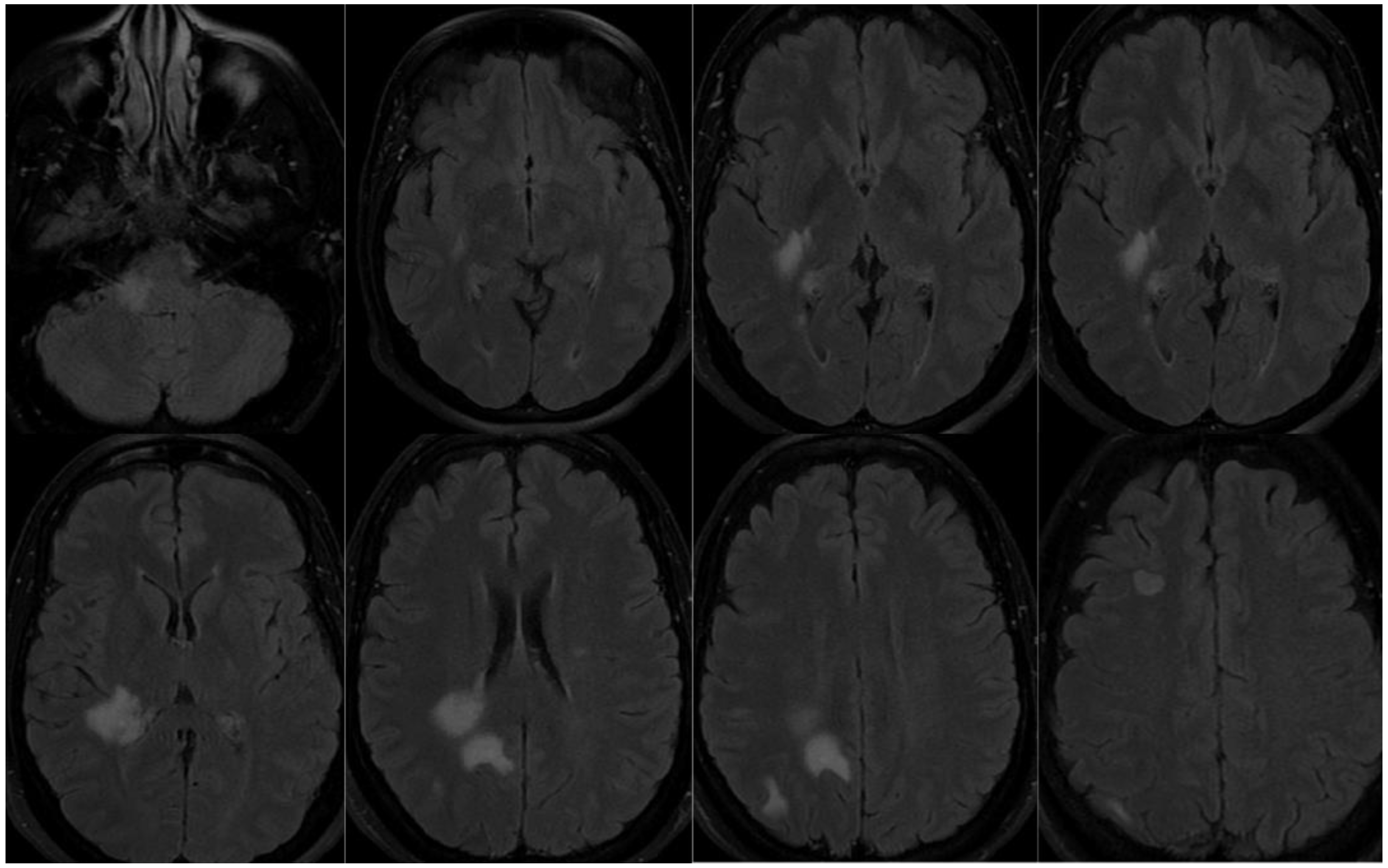
Patient 3, female, 39 years-old, with history of psoriasis, spontaneous miscarriages and reducing gastroplasty is admitted due to paresthesia in the right hemiface, and left lower limb, as well as deviation of the labial commissure, right auricular fullness, vertigo and headache. Diagnostic investigation with complementary tests was initiated. Eighteen days before she had COVID-19 confirmed with PCR, with headache, cough and sore throat. MRI showed oval white-matter lesions on the cortex, periventricular and brainstem, on the right, hyperintense in T2/FLAIR (Figure 3). CSF exam was positive for oligoclonal bands, and negative for infections, including SARS-CoV-2, and autoantibodies. Screening for autoimmune diseases, including MOG and AQP4 abs, and metabolic diseases were negative. At first, she was diagnosed with ADEM following SARS-CoV-2 infection. Nonetheless, three months after, she evolved with new focal symptoms and new lesions on the MRI, with gadolinium enhancement. Therefore, she was diagnosed with MS and treated with dimethyl fumarate. The disease stabilized and she stayed asymptomatic since then.
Figure 3.
MRI patient 3. Oval white-matter lesions on the cortex, periventricular and brainstem, on the right, hyperintense in T2/FLAIR.
Figure 3.
MRI patient 3. Oval white-matter lesions on the cortex, periventricular and brainstem, on the right, hyperintense in T2/FLAIR.
3.2. Sequence identification
The extended literature research led to the selection of eight viral proteins and ten self-proteins associated with CNS demyelinating diseases and auto-immunes encephalitis, listed as follow:
Sars-CoV-2 proteins: Spike protein (S), envelope protein (E), leader protein/non-structural protein 1 (Nsp1), non-structural protein 2 (Nsp2), non-structural protein 3 (Nsp3), non-structural protein 13/helicase (Nsp13), ORF7a and nucleocapsid (N) (Table 1).

Table 1.
SARS-CoV-2 proteins selected.
| Protein | Number of aminoacids | Gene | NCBI Reference Sequence | Uniprot ID |
|---|---|---|---|---|
| E | 75aa | E | YP_009724392 | P0DTC4 |
| Nsp1 | 180aa | ORF1a | YP_009742608.1 | P0DTD1 |
| M | 222aa | M | YP_009724393.1 | P0DTC5 |
| Nsp2 | 638aa | ORF1a | YP_009742609.1 | P0DTD1 |
| Nsp3 | 1945aa | ORF1a | YP_009742610.1 | P0DTD1 |
| Nsp13 | 601aa | ORF1a | NP_828870.1 | P0DTD1 |
| ORF7a | 121aa | ORF7a | YP_009724395.1 | P0DTC7 |
| S | 1273 aa | S | YP_009724390.1 | P0DTC2 |
Self-proteins: Glutamic acid decarboxylase 65-kilodalton isoform (GAD65), Myelin proteolipid protein (PLP), Myelin basic protein (MBP), Myelin-oligodendrocyte glycoprotein (MOG), Myelin-associated glycoprotein (MAG), Myelin-associated oligodendrocyte basic protein (MOBP), Transaldolase, 2’,3’-Cyclic-nucleotide 3’-phosphodiesterase (CNP), Aquaporin-4 (AQP4), N-methyl-D-aspartate receptor 1 (NMDAR1), and Myeloperoxidase (MPO) (Table 2).
Table 2.
Self-proteins associated with IDDs and autoimmune encephalitis.
| Protein | Number of aminoacids | Gene | NCBI Reference Sequence | Uniprot ID |
|---|---|---|---|---|
| 2’,3’-Cyclic-nucleotide 3’-phosphodiesterase (CNP) | 421aa | CNP | NP_149124.3 | P09543 |
| Aquaporin-4 (AQP4) | 323aa | AQP4 | NP_001641.1 | P55087 |
| Glutamic acid decarboxylase 65-kilodalton isoform (GAD65) | 585aa | GAD2 | NP_001127838.1 | Q05329 |
| Myelin associated glycoprotein (MAG) | 626aa | MAG | NP_002352.1 | P20916 |
| Myelin basic protein (MBP) | 304aa | MBP | NP_001020272.1 | P02686 |
| Myelin oligodendrocyte glycoprotein (MOG) | 247aa | MOG | NP_996532.2 | Q16653 |
| Myelin-associated oligodendrocytic basic protein (MOBP) | 183aa | MOBP | NP_001380633.1 | Q13875 |
| Myeloperoxidase (MPO) | 745aa | MPO | NP_000241.1 | P05164 |
| N-methyl-D-aspartate receptor 1 (NMDAR1) | 938aa | GRIN1 | NP_015566.1 | Q05586 |
| Transaldolase | 337aa | TALDO1 | NP_006746.1 | P37837 |
3.3. Linear and three-dimensional analysis
A total of 80 possible arrangements were made through the bioinformatics approach, to identify the sequences that shared linear and three-dimensional identity with human autoantigens of demyelinating brain diseases and autoimmune encephalitis.
The resulting arranges were ranked based on the highest TM-scores, excluding randomly arranged, unrelated proteins (TM-score < 0.3), leaving 29 arranges (Table 3). It is noteworthy that we only considered meaningful the SARS-CoV-2 sequences with reported linear identity on BLASTp, varying from 62.50% to 100% of identity, with significant E-score and antigenic properties according to VaxiJen (threshold antigenic score of 0.4).
Table 3.
Linear and three-dimensional identity between SARS-CoV-2 antigens and self-antigens, and antigenic properties of each combination.
Table 3.
Linear and three-dimensional identity between SARS-CoV-2 antigens and self-antigens, and antigenic properties of each combination.
| Sars-Cov-2 antigens | Autoantigens | Region of the SARS-Cov-2 antigen with more identity | Region of the autoantigen with more identity | % Identity | E-value | SWISS MODEL SARS-CoV-2 antigen | SWISS MODEL autoantigen | TM-Score | RMSD | Overall prediction Vaxijen linear | Overall prediction VaxiJen three-dimensional model |
|---|---|---|---|---|---|---|---|---|---|---|---|
| M | NMDAR1 | 54-70 | 562-577 | 100 | 3,00E-04 | 74-106 | 626-658 | 0.89 | 0.55 | 0.9306 | 0.5324 |
| M | MPO | 134-162 | 57-85 | 100 | 1.0 | 76-105 | 67-96 | 0.73 | 1.21 | 0.4740 | 0.5177 |
| Nsp2 | NMDAR1 | 448-465 | 325-344 | 71.43 | 0.038 | 549-584 | 621-650 | 0.69 | 1.35 | 0.5983 | 0.4174 |
| S | MOG | 249-278 | 83-102 | 83.33 | 0.069 | 944-974 | 150-180 | 0.63 | 2.06 | 0.4706 | 0.4059 |
| ORF7a | MOG | 25-32 | 95-102 | 75 | 2,00E-04 | 17-81 | Complete structure | 0.62 | 2.83 | 0.4846 | 0.6598 |
| N | MPO | 227-236 | 149-157 | 70 | 0.002 | 388-419 | 67-98 | 0.59 | 1.91 | 0.4117 | 0.4124 |
| Nsp13 | GAD65 | 466-472 | 439-445 | 66.67 | 3,00E-04 | 290-349 | 312-389 | 0.52 | 3.22 | 0.6555 | 0.4695 |
| Nsp1 | GAD65 | 131-138 | 137-139 | 100 | 0.005 | 32-61 | 302-331 | 0.52 | 1.57 | -0.3299 | 0.6325 |
| Nsp1 | MOG | 103-114 | 210-221 | 71.43 | 0.003 | 33-62 | 204-232 | 0.50 | 1.59 | 0.6013 | 0.6757 |
| Nsp3 | MPO | 903-908 | 613-618 | 83.33 | 2,00E-05 | 180-209 | 71-112 | 0.50 | 2.79 | 1.8236 | 0.8823 |
| S | NMDAR1 | 1020-1027 | 223-230 | 100 | 0.017 | 1020-1050 | 221-250 | 0.50 | 1.47 | 0.8726 | 0.7359 |
| Nsp3 | NMDAR1 | 1800-1809 | 766-774 | 100 | 5,00E-04 | 399-535 | 153-278 | 0.49 | 3.86 | 0.4238 | 0.4044 |
| Nsp13 | PLP | 88-94 | 99-105 | 71.43 | 1,00E-04 | 310-342 | 175-210 | 0.49 | 2.82 | -0.0624 | 0.4685 |
| Nsp1 | PLP | 34-62 | 196-211 | 100 | 0.006 | 33-64 | 240-272 | 0.47 | 1.69 | 0.7645 | 0.5854 |
| Nsp1 | Transaldolase | 84-99 | 139-154 | 100 | 0.001 | 33-62 | 145-176 | 0.46 | 2.18 | 0.6898 | 0.6757 |
| Nsp2 | MPO | 419-425 | 159-165 | 100 | 0.15 | 672-707 | 67-95 | 0.46 | 1.16 | 0.5813 | 0.5890 |
| Nsp3 | PLP | 88-94 | 99-105 | 71.43 | 1,00E-04 | 180-209 | 34-63 | 0.45 | 2.07 | -0.0624 | 0.8823 |
| M | MAG | 132-138 | 369-375 | 100 | 2,00E-04 | 156-186 | 295-325 | 0.43 | 2.59 | 0.4549 | 0.6527 |
| S | Transaldolase | 1110-1117 | 37-44 | 62.50 | 0.002 | 276-305 | 125-156 | 0.42 | 2.10 | 0.8734 | 0.6476 |
| Nsp13 | Transaldolase | 146-151 | 307-312 | 83.33 | 0.001 | 367-396 | 135-164 | 0.41 | 2.60 | 1.0726 | 1.1104 |
| N | CNP | 243-252 | 191 200 | 100 | 0.001 | 195-239 | 322-363 | 0.41 | 2.72 | 0.7056 | 0.6601 |
| Nsp13 | NMDAR1 | 301-306 | 628-636 | 100 | 3,00E-04 | 503-532 | 683-712 | 0.41 | 2.90 | 0.9060 | 0.4818 |
| Nsp2 | Transaldolase | 389-403 | 95-109 | 66.67 | 0.19 | 249-280 | 134-163 | 0.40 | 1.90 | 0.5146 | 0.7524 |
| Nsp2 | CNP | 270-278 | 348-356 | 77.78 | 4,00E-07 | 395-432 | 349-382 | 0.39 | 2.38 | 0.9295 | 0.4387 |
| N | MOBP | 198-209 | 168-178 | 77.78 | 5,00E-06 | 202-233 | 29-74 | 0.38 | 2.38 | 0.4291 | 0.6320 |
| N | PLP | 170-184 | 120-134 | 75 | 0.0003 | 212-241 | 75-98 | 0.37 | 1.44 | 0.4459 | 0.7169 |
| Nsp1 | MBP | 76-102 | 198-116 | 100 | 0.010 | 32-67 | 211-240 | 0.36 | 3.22 | 0.6583 | 0.4770 |
| Nsp3 | AQP4 | 63-77 | 51-65 | 100 | 0.27 | 2667-2697 | 197-227 | 0.35 | 2.36 | 0.4168 | 0.5551 |
| Nsp2 | MBP | 61-73 | 107-119 | 100 | 8,00E-04 | 280-309 | 200-229 | 0.33 | 2.79 | 0.6625 | 0.5624 |
Among these arranges, eleven three-dimensional models had a significant linear and three-dimensional overlap of autoimmune CNS proteins and SARS CoV-2 proteins (TM-score ≥ 0.5). The most similar structures were M and NMDAR1 (TM-score: 0.89), M and MPO (TM-Score = 0.73), nsp2 and NMDAR1 (TM-score = 0.69), S and MOG (TM-score = 0.63), ORF7a and MOG (TM-score = 0.62), N and MPO (TM-score = 0.59), nsp13 and GAD65 (TM-score = 0.52), nsp1 and GAD65 (TM-score = 0.52), nsp1 and MOG (TM-score = 0.50), nsp3 and MPO (TM-score = 0.50), S and NMDAR1 (TM-score = 0.50) (Figure 1).
Figure 4.
three-dimensional models had a significant linear and three-dimensional overlap of autoimmune CNS proteins and SARS CoV-2 proteins according to TM-align. a) M and NMDAR1 (TM-score: 0.89). b) M and MPO (TM-Score = 0.73). c) nsp2 and NMDAR1 (TM-score = 0.69). d) S and MOG (TM-score = 0.63). e) ORF7a and MOG (TM-score = 0.62). f) N and MPO (TM-score = 0.59). g) nsp13 and GAD65 (TM-score = 0.52). h) nsp1 and GAD65 (TM-score = 0.52). i) nsp1 and MOG (TM-score = 0.50). j) nsp3 and MPO (TM-score = 0.50). k) S and NMDAR1 (TM-score = 0.50).
Figure 4.
three-dimensional models had a significant linear and three-dimensional overlap of autoimmune CNS proteins and SARS CoV-2 proteins according to TM-align. a) M and NMDAR1 (TM-score: 0.89). b) M and MPO (TM-Score = 0.73). c) nsp2 and NMDAR1 (TM-score = 0.69). d) S and MOG (TM-score = 0.63). e) ORF7a and MOG (TM-score = 0.62). f) N and MPO (TM-score = 0.59). g) nsp13 and GAD65 (TM-score = 0.52). h) nsp1 and GAD65 (TM-score = 0.52). i) nsp1 and MOG (TM-score = 0.50). j) nsp3 and MPO (TM-score = 0.50). k) S and NMDAR1 (TM-score = 0.50).
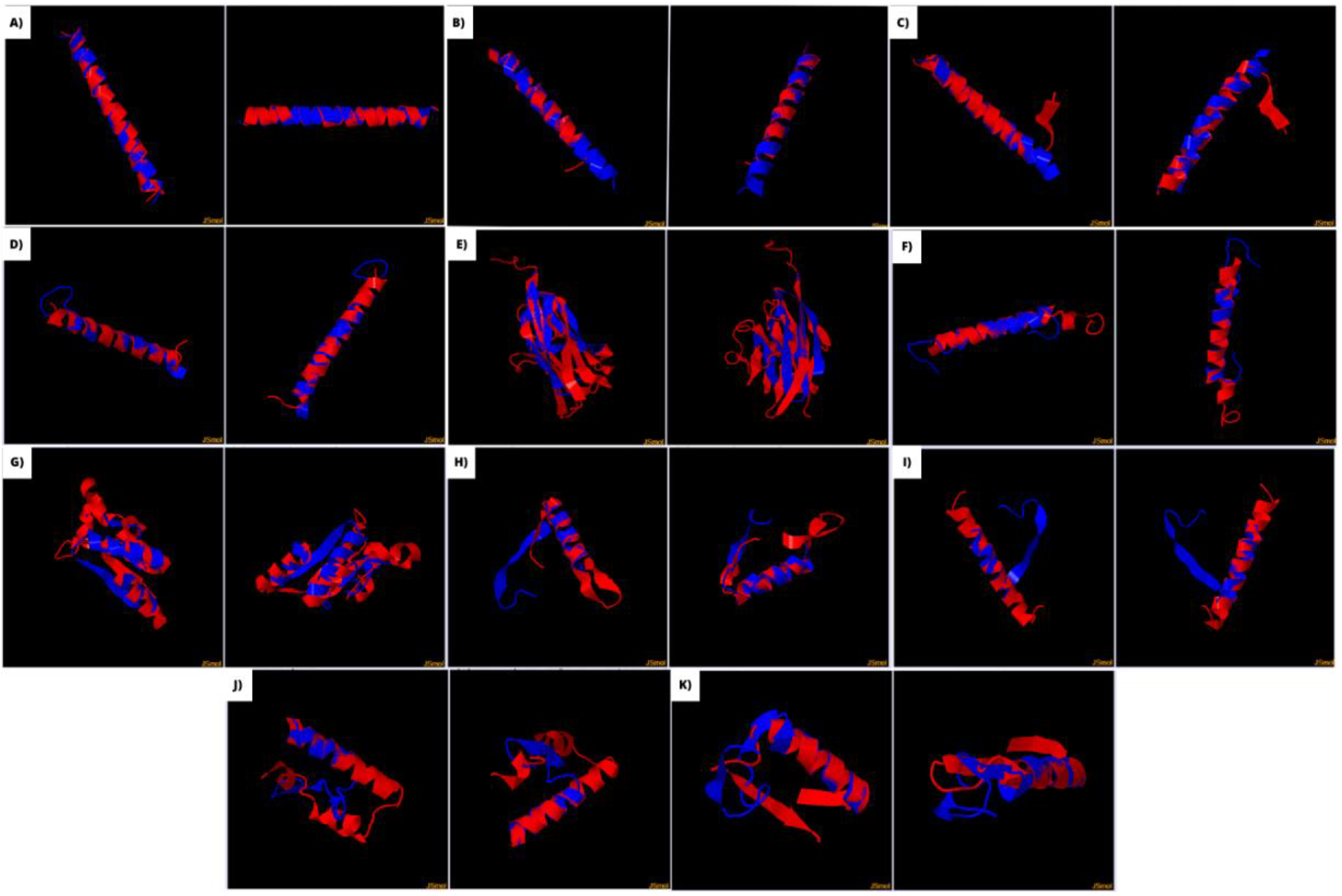
NMDAR1, MOG and MPO were the self-antigens with more significant identity with SARS-CoV-2 antigens, each one with 3 different proteins. GAD65 had also significant identity with 2 virus antigens. PLP, MBP, MOBP, MAG, AQP4 and transaldolase had demonstrated significant linear identity with at least one virus protein, along with three-dimensional overlap not considered random. Nonetheless, their TM-score were below 0.5, meaning they are not on the same fold. This means their MM is possible, yet less achievable in practice.
3.3. Search for potential T cell epitopes
The sequences of the eleven combinations were used as the input in a neural network–based algorithm to predict their binding capacity to HLA subtypes related to autoimmune CNS diseases. All sequences presented at least one epitope with strong or intermediate binding capacity to the chosen HLA subtypes (Table 4). The arranges with strongest binding capacities were seen with nsp1 and GAD65 epitopes of HLA-A*31:01 (Ic50 31.44 and 27.31, respectively) and with nsp13 and GAD65 epitopes of HLA-A*31:01 (Ic50 18.72 and 35.44, respectively).
Table 4.
The viral epitopes for MHC-I and MHC-II and the corresponding homologous human proteins.
| SARS-CoV-2 antigen | Autoantigens | Allele | Potencial SARS-CoV-2 epitope | Corresponding human epitope | IC50 virus peptide | IC50 human peptide |
|---|---|---|---|---|---|---|
| M | NMDAR1 | HLA-DQA1*01:02/DQB1*06:02 | NWITGGIAIAMACLV | VWAGFAMIIVASYTA | 57.00 | 60.00 |
| HLA-DRB1*15:01 | LMWLSYFIASFRLFA | GFAMIIVASYTANLA | 68.00 | 49.00 | ||
| HLA-A*31:01 | LSYFIASFR | LGMVWAGFAM | 12.20 | 262.06 | ||
| M | MPO | HLA-A*31:01 | LSYFIASFR | KQLVDKAYK | 12.20 | 68.78 |
| HLA-B*07:02 | GGIAIAMACLV | RLRSGSASPM | 321.23 | 159.89 | ||
| Nsp2 | NMDAR1 | HLA-DQA1*01:02/DQB1*06:02 | RVLQKAAITILDGIS | VWAGFAMIIVASYTA | 93.00 | 60.00 |
| HLA-DRB1*15:01 | ITILDGISQYSLRLI | VWAGFAMIIVASYTA | 146.00 | 60.00 | ||
| HLA-A*31:01 | RTLETAQNSVR | GAPRSFSAR | 309.40 | 129.86 | ||
| HLA-B*07:02 | SVRVLQKAAI | APRSFSARIL | 357.95 | 29.30 | ||
| S | MOG | HLA-DRB1*15:01 | LNTLVKQLSSNFGAI | GVLVLLAVLPVLLLQ | 89.00 | 58.00 |
| HLA-DQA1*01:02/ DQB1*06:02 | QNAQALNTLVKQLSS | WVSPGVLVLLAVLPV | 319.00 | 229.00 | ||
| ORF7a | MOG | HLA-DQA1*01:02/DQB1*06:02 | YQECVRGTTVLLKEP | YWVSPGVLVLLAVLP | 261.00 | 210.00 |
| N | MPO | HLA-A*31:01 | FSKQLQQSMSS | KQLVDKAYK | 46.72 | 68.78 |
| Nsp13 | GAD65 | HLA-DRB1*15:01 | AIGLALYYPSARIVY | AKQKGFVPFLVSATA | 71.00 | 196.00 |
| HLA-DQA1*01:02/DQB1*06:02 | IVYTACSHAAVDALC | LVSATAGTTVYGAFD | 347.00 | 52.00 | ||
| HLA-A*31:01 | KYLPIDKCSR | KHKWKLSGVER | 18.72 | 35.44 | ||
| HLA-B*07:02 | LPIDKCSR | VPFLVSAT | 187.50 | 69.82 | ||
| Nsp1 | GAD65 | HLA-DRB1*03:01 | SVEEVLSEARQHLKD | RGKMIPSDLERRILE | 489.00 | 448.00 |
| HLA-A*31:01 | HLKDGTCGLVE | KMIPSDLERR | 31.44 | 27.31 | ||
| Nsp1 | MOG | HLA-A*31:01 | HLKDGTCGLVE | CWKITLFVIVP | 31.44 | 284.79 |
| Nsp3 | MPO | HLA-A*31:01 | SYKDWSYSGQS | RLRSGSASPME | 38.77 | 111.56 |
| S | NMDAR1 | HLA-DQA1*01:02/ DQB1*06:02 | ASANLAATKMSECVL | ASEDDAATVYRAAAM | 213.00 | 121.00 |
| HLA-A*31:01 | KMSECVLGQSKR | LSASEDDAATVYR | 297.9 | 495.7 | ||
| HLA-DQA1*01:02/ DQB1*06:02 | AQYTSALLAGTITSG | SRRVLLLAGRLAAQS | 122.00 | 199.00 | ||
| HLA-B*07:02 | MIAQYTSAL | MAAESRRVL | 61.88 | 54.29 |
NMDAR1 combinations had the highest number of epitopes with strong or intermediate binding capacity with two combinations binding to all four selected HLAs subtypes (nsp2 and NMDAR1, and S and NMDAR1), and the same epitope had identity with different virus antigens (M and nsp2).
Interestingly, different combinations shared the same viral epitope, such as M with NMDAR1 and MPO, and nsp1 with GAD65 and MOG (Table 4). This reinforces the potential of such region to possibly trigger an unwanted autoimmune response, by mimicking distinct self-proteins.
4. Discussion
SARS-CoV-2 is widely studied for the generation of multi-system autoimmune reactions [33]. In that sense, the triggering of CNS autoimmune diseases seems to be a consequence of an imprecise adaptive immune system response to the presence of viral antigens. It is well known that some viruses demonstrated neurotropic features [34]–[37], and replication within the brain tissue, as shown by our team with Zika virus (ZIKV) [34]. However, even ZIKV nervous system manifestation is not always associated with acute infection and MM seems to justify these events [38], [39]. Similarly, in COVID-19, viral load or severe acute infection doesn’t seem to be the only mechanism to justify CNS involvement [40], [41]. The occurrence of IDD phenotypes and encephalitis as para or postinfectious event, seems to be an immune-mediated response induced by SARS-CoV-2 [42].
A large study from various global health organizations found that the incidence of autoimmune diseases was significantly higher in the COVID-19 cohort compared to the non-COVID-19 group after a 6-month follow-up period [43]. Another similar study identified a 43% higher likelihood of developing an autoimmune disease between 3 to 15 months after infection compared to a non-COVID-19 cohort [44]. Despite the progress made, cases of CNS autoimmunity after COVID-19 are rare and mainly consist of isolated case reports or case series, which provide limited information regarding clinical outcomes [45].
Although the target of such supposed autoimmune mechanisms, precisely regarding the CNS manifestations, are still not fully understood, our findings suggest that cross-reaction with selected CNS proteins associated with autoimmune brain diseases is possible to occur secondary to the immune response to SARS CoV-2 infection. However, the risk of developing these diseases or experiencing relapses in the setting of COVID-19 remains relatively low [46]. In our cohort, 3 patients developed IDD following SARS-CoV-2 infection or vaccination. Since the CNS autoimmune manifestations after COVID-19 are rare, it is expected that genetic predisposition plays an essential role in the disease mechanism [47]. Despite the low frequency, the identification of IDD in such circumstances is primordial, taking in consideration the high prevalence of SARS-CoV-2 infection and the possible critical state in which the patients may encounter it. For example, our first patient evolved with EDSS 9.5, a near-death experience in a young individual with no previous comorbidity. Moreover, is important to consider SARS-CoV-2 infection as a possible trigger of IDDs, because some patients present demyelinating events as the only symptom of COVID-19, as happened with our second patient.
Molecular mimicry has been described as an essential immune mechanism involved in autoimmune reactions, especially from viruses [48]. The sharing of a linear amino acid sequence or a three-dimensional conformation fit between an antigen of the virus and a host self-protein can trigger a cross-reaction from the adaptive immune system and, therefore, have a major role in initiating an autoimmune response in genetically susceptible individuals [49]. Several researchers have recognized molecular mimicry as a component of COVID-19 pathophysiology [50]–[52]. As example, Lucchese et al. observed that molecular mimicry between SARS-CoV-2 antigens and respiratory pacemaker neurons may contribute to understanding respiratory failure [53]. Hence, MM may be a key component of the immune system dysregulated response in the CNS.
In this study, 80 possible arranges of identity among SARS-CoV-2 antigens and self-antigens related to autoimmune CNS diseases were made. Among these arranges, eleven models had a significant linear and three-dimensional overlap of autoimmune CNS proteins and SARS CoV-2 proteins (TM-score ≥ 0.5). NMDAR1, MOG and MPO were the self-antigens with more significant identity with SARS-CoV-2 antigens, followed by GAD65. Notably, dysregulated serum levels of autoantibodies NMDAR, GAD65 and MOG were detected in patients with severe COVID-19 compared with healthy controls and mild COVID-19 patients [54].
MS is a classic example of an autoimmune CNS disease characterized by chronic inflammation and demyelination [55]. SARS-CoV-2 must likely act as a precipitating factor rather than being a direct cause of MS, triggering autoimmunity in genetically predisposed individuals. In our cohort, two patients had SARS-CoV-2 related events (vaccination and infection) as trigger for MS. Both patients were female, however their ages, comorbidities, symptoms and MRI lesions were considerably different, highlighting the importance of genetic predisposal and other environmental factors on the course of the disease [56].
MBP, MOBP, PLP and MAG are myelin proteins known to be critical autoantigens in causing demyelination in CNS leading to MS [57], [58]. In our study, the TM-scores among these proteins and SARS-CoV-2 antigens were low, but not considered randomly arranged, unrelated proteins. This can mean that MM among these MS autoantigens is feasible, however, the evidence is not strong. Nevertheless, MPO, a pro-oxidative enzyme associated with immune-inflammatory, oxidative stress pathways, and cortical demyelination [59], has been MPO gaining acceptance as an important modulator of MS activity [60]. Higher expressing MPO genotype is overrepresented in early-onset MS in females [61] and immunohistochemical analysis shows that MPO is present in microglia in and around MS lesions [61]. This study found a significant overlap of MPO among 3 different SARS-CoV-2 antigens (M, N, nsp3). SARS-CoV-2 ability to mimic MPO seem to provide a greater threat for triggering new onset MS or worsening of MS symptoms in genetically predisposed patients, as seen during the COVID-19 pandemic [62], [63].
MOGAD is an emerging subset of CNS demyelinating disease [64], [65], and has also been related to COVID-19 [14]. In our study, one patient evolved with MOGAD during an asymptomatic SARS-CoV-2 infection. Besides, our in silico analysis showed that MOG shared significant linear and three-dimensional identity with 3 different virus antigens (S, ORF7a and nsp1), being the most prominent overlap with Spike protein. This goes accordingly to recent literature, that identified anti-MOG antibodies in the acute and post-infectious phase of SARS-CoV-2 infection and COVID-19 vaccination [66], [67]. Moreover, the literature has shown that diseases associated with anti-MOG almost tripled during the COVID-19 pandemic [68].
Our study highlights that NMDAR1 has 3 domains with significant linear and three-dimensional identity with SARS-CoV-2 antigens, including Spike protein, and all of them have binding capacity to T-cells that would consider them epitopes. It has been proposed that SARS-CoV-2’s molecular mimicry may induce anti-NMDAR encephalitis after COVID-19 [69]. This may be a key mechanism beneath CNS manifestations of COVID-19 disease and vaccination associated with anti-NMDAR antibodies [15], [70].
Moreover, GAD65 had the strongest binding capacity to HLA in this work, with two different combinations of mimicry. Cases of autoimmune encephalitis associated with GAD65 have been described following SARS-CoV-2 infection [71], [72]. This finding reinforces the association of SARS-CoV-2 MM and the clinical findings related to anti-GAD65 antibody.
A growing body of evidence has demonstrated the relationship between ADEM and SARS-CoV-2 infection [73]. ADEM following SARS-CoV-2 infection and vaccination have been associated to MOG [74]–[77] and NMDAR [78] antibodies. Interestingly, in our study, both MOG and NMDAR are associated with Spike protein, the most common antigen presented in SARS-CoV-2 vaccination, which may indicate the relevance of MM in post-COVID ADEM manifestations.
In this cohort, 52.45% of the COVID-19 ICU patients presented cognitive impairment during the post-acute phase of COVID-19. Notably, NMDAR, GAD65 and MPO may be involved not only in acute encephalitis or demyelinating events but also in neurocognitive and psychiatric manifestations, frequently seen in long COVID patients [69], [79]–[81]. Pathological results in cognitive screening were associated with the presence of antibodies against NMDAR and GAD65 in CSF of long COVID patients [82]. The MM between these autoantibodies and SARS-CoV-2 antigens may be a prominent asset to understanding the pathogenesis of long COVID cognitive and psychiatric symptoms.
Genetic susceptibility seems to explain the heterogeneity of response to immune tolerance breakdown and molecular mimicry between autoantigens and viral proteins [49]. Due to the limited knowledge about genetic susceptibility to explain mechanisms involved in the pathophysiology of AE, we choose HLA variants with known associations with CNS autoimmune diseases [58], [83]–[87]. Interestingly enough, all eleven combinations with significant linear and three-dimensional identity presented at least one epitope with strong or intermediate binding capacity to the chosen HLA subtypes. In this manner, the investigation of the connection between HLA alleles related to CNS autoimmune diseases and the MM found in this paper can strengthen the results and possibly help to elucidate the pathophysiology of these manifestations.
It is worth to highlight the results regarding the S protein. Spike or its fragments, has the ability to cross the blood-brain barrier (BBB), irrespective of the presence of the viral RNA [88]. Furthermore, some cases have reported an association of CNS demyelination events with the use of vaccines with S protein as the main antigen for the generation of immunological memory, which has become a major concern for health authorities worldwide [89]. Thus, the MM regarding S may be more common than the others described in this article. Indeed, both MOG and NMDAR1, which presented significant linear and three-dimensional overlap with Spike, have been associated with COVID-19 in a more expressive way than the other autoimmune affection [15], [68], [69], and have been related to COVID-19 vaccination [70], [90]–[92]. One of our patients triggered IDD following COVID-19 vaccination. Although is not possible to affirm causality, MM must be considered as a possible mechanism for this phenomenon.
As limitations of this study, is important to mention that is a theoretical work, however it is based on our cohort findings and recent literature studies regarding SARS-CoV-2. Besides, it uses validated software’s to give results as close as possible to reality. Additionally, the study used a limited number of HLA alleles in the prediction of T-cell binding capacity, only the most common HLA alleles on the literature associated with CNS autoimmune diseases, in order to increase the specificity of the results. Thereby, is possible that some epitopes of rarer HLA weren’t included in this study. Further studies are needed to validate the in silico work described here, as well as to understand probable genetic susceptibility some individuals must develop such manifestations.
5. Conclusions
The presented study proposes a demonstration of possible molecular mimicry between SARS-CoV-2 antigens and CNS autoimmune self-antigens, especially MOG, NMDAR1, GAD65 and MPO in genetically susceptible individuals. This goes accordingly to our cohort, with three cases of IDD, and to the most recent literature. Therefore, advancing our understanding of the key mechanisms of SARS-CoV-2-mediated autoimmunity is urgent.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, EGG. SVAL, MMS and FLFD.; methodology, EGG, FLFD, JVRV, RAF; software, EGG and FLFD; validation, ALS, RAF, JVRV, CFT, VGCC, VCSRP, MMS and FLFD.; formal analysis, EGG.; investigation, EGG, ALS, LAD, SVAL, CFT, VGCC, VCSRP, MMS and JVRV; resources, SVAL; data curation, ALS, EGG, RAF, LAD, FLFD, JVRV, CFT, VGCC, VCSRP, MMS and SVAL; writing—original draft preparation, EGG.; writing—review and editing, ASL, RAF, SVAL, FLFD and JVRV.; visualization, EGG, LAD, ASL, RAF, FLFD, JVRV, CFT, VGCC, VCSRP, MMS and SVAL; supervision, FLFD, MMS and SVAL; project administration, SVAL; funding acquisition, SVAL. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) grant numbers: E26/210.254/2020, E-26/210.657/2021, E-26/210.273/2018, E-26/201.040/2021, E-26/200.733/2021, E26/2006.022/2022 and E-26/201.406/2021; by Financiadora de Estudos e Projetos (FINEP), and by FIOCRUZ Ministry of Health/INOVA Program, grant number: VPPCB-005-FIO-20-2-56.
Data Availability Statement
The results presented in this article are supported by data in other articles published in MDPI journals. The clinical profile and risk factors for severe COVID-19 in our cohort of hospitalized patients, comparing the First and Second Pandemic Waves was published on the Journal of Clinical Medicine in 2023, under the doi: 10.3390/jcm12072568. Disease severity was associated with older age, pre-existing neurological comorbidities, higher viral load, and dyspnea. Laboratory biomarkers related to white blood cells, coagulation, cellular injury, inflammation, renal, and liver injuries were significantly associated with severe COVID-19. During the second wave of the pandemic, the necessity of invasive respiratory support was higher, and more individuals with COVID-19 developed acute hepatitis, suggesting that the progression of the second wave resulted in an increase in severe cases.
We used transcriptome analysis of these patients to understand key genes and cellular mechanisms that are most affected by the severe outcome of COVID-19. Transcriptomic analysis revealed 1009 up-regulated and 501 down-regulated genes in the SARS group, with 10% of both being composed of long non-coding RNA. Ribosome and cell cycle pathways were enriched among down-regulated genes. The most connected proteins among the differentially expressed genes involved transport dysregulation, proteasome degradation, interferon response, cytokinesis failure, and host translation inhibition. Furthermore, interactome analysis showed Fibrillarin to be one of the key genes affected by SARS-CoV-2. This protein interacts directly with the N protein and long non-coding RNAs affecting transcription, translation, and ribosomal processes. This work was published on the International Journal of Molecular Sciences in 2022, under the doi: 10.3390/ijms232113588.
We also published a study that aimed to establish a relationship between miRNA and neurological manifestations in our cohort of COVID-19 patients co-infected with HHV-6 and evaluate miRNAs as potential biomarkers. miRNA analysis by real-time polymerase chain reaction (qPCR) revealed miRNAs associated with neuroinflammation were highly expressed in patients with neurological disorders and HHV-6 detection. When compared with the group of patients without detection of HHVs DNA and without neurological alterations, the group with detection of HHV-6 DNA and neurological alteration, displayed significant differences in the expression of mir-21, mir-146a, miR-155 and miR-let-7b (p < 0.01). This work was published on the International Journal of Molecular Sciences in 2023, under the doi: 10.3390/ijms241311201.
One of our authors (Salvio, AL) also analyzed the Effectiveness of Household Disinfection Techniques to Remove SARS-CoV-2 from Cloth Masks. The study showed that all biocidal treatments successfully disinfected the tissue tested. This work was published on Pathogens in 2022, under the doi: 10.3390/pathogens11080916.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- worldometers.info/coronavirus/, “COVID-19 CORONAVIRUS PANDEMIC.”.
- N. I. Nii-Trebi, “Emerging and Neglected Infectious Diseases: Insights, Advances, and Challenges,” Biomed Res Int, vol. 2017, pp. 1–15, 2017, doi: 10.1155/2017/5245021. [CrossRef]
- S. V. Alves-Leon et al., “Zika virus found in brain tissue of a multiple sclerosis patient undergoing an acute disseminated encephalomyelitis-like episode,” Multiple Sclerosis Journal, vol. 25, no. 3, pp. 427–430, Mar. 2019, doi: 10.1177/1352458518781992. [CrossRef]
- F. C. Rueda-Lopes et al., “Clinical and magnetic resonance imaging patterns of extensive Chikungunya virus–associated myelitis,” J Neurovirol, vol. 27, no. 4, pp. 616–625, Aug. 2021, doi: 10.1007/s13365-021-00962-4. [CrossRef]
- S. V. Alves-Leon et al., “Exome-Wide Search for Genes Associated With Central Nervous System Inflammatory Demyelinating Diseases Following CHIKV Infection: The Tip of the Iceberg,” Front Genet, vol. 12, Mar. 2021, doi: 10.3389/fgene.2021.639364. [CrossRef]
- P. Feizi et al., “Central nervous system (CNS) inflammatory demyelinating diseases (IDDs) associated with COVID-19: A case series and review,” J Neuroimmunol, vol. 371, p. 577939, Oct. 2022, doi: 10.1016/j.jneuroim.2022.577939. [CrossRef]
- G. Toscano et al., “Guillain–Barré Syndrome Associated with SARS-CoV-2,” New England Journal of Medicine, vol. 382, no. 26, pp. 2574–2576, Jun. 2020, doi: 10.1056/NEJMc2009191. [CrossRef]
- M. B. A. Oldstone, “Molecular mimicry and immune-mediated diseases,” The FASEB Journal, vol. 12, no. 13, pp. 1255–1265, Oct. 1998, doi: 10.1096/fasebj.12.13.1255. [CrossRef]
- M. Lima et al., “Coronaviruses and their relationship with multiple sclerosis: is the prevalence of multiple sclerosis going to increase after the Covid-19 pandemia?,” Rev Neurosci, vol. 33, no. 7, pp. 703–720, Oct. 2022, doi: 10.1515/revneuro-2021-0148. [CrossRef]
- J. C. Gerdes, I. Klein, B. L. DeVald, and J. S. Burks, “Coronavirus isolates SK and SD from multiple sclerosis patients are serologically related to murine coronaviruses A59 and JHM and human coronavirus OC43, but not to human coronavirus 229E,” J Virol, vol. 38, no. 1, pp. 231–238, Apr. 1981, doi: 10.1128/jvi.38.1.231-238.1981. [CrossRef]
- R. Moody, K. Wilson, K. L. Flanagan, A. Jaworowski, and M. Plebanski, “Adaptive Immunity and the Risk of Autoreactivity in COVID-19,” Int J Mol Sci, vol. 22, no. 16, p. 8965, Aug. 2021, doi: 10.3390/ijms22168965. [CrossRef]
- C. Hammer et al., “Neuropsychiatric disease relevance of circulating anti-NMDA receptor autoantibodies depends on blood–brain barrier integrity,” Mol Psychiatry, vol. 19, no. 10, pp. 1143–1149, Oct. 2014, doi: 10.1038/mp.2013.110. [CrossRef]
- H. Prüss, “Autoantibodies in neurological disease,” Nature Reviews Immunology, vol. 21, no. 12. Nature Research, pp. 798–813, Dec. 01, 2021. doi: 10.1038/s41577-021-00543-w. [CrossRef]
- P. Feizi et al., “Central nervous system (CNS) inflammatory demyelinating diseases (IDDs) associated with COVID-19: A case series and review,” J Neuroimmunol, vol. 371, p. 577939, Oct. 2022, doi: 10.1016/j.jneuroim.2022.577939. [CrossRef]
- H. Wang, “COVID−19, Anti-NMDA Receptor Encephalitis and MicroRNA,” Front Immunol, vol. 13, Mar. 2022, doi: 10.3389/fimmu.2022.825103. [CrossRef]
- M. Salari, B. Zaker Harofteh, and M. Etemadifar, “Autoimmune meningoencephalitis associated with anti-glutamic acid decarboxylase antibody following COVID-19 infection: A case report,” Clin Case Rep, vol. 10, no. 12, Dec. 2022, doi: 10.1002/ccr3.6597. [CrossRef]
- L. Llorente Ayuso, P. Torres Rubio, R. F. Beijinho do Rosário, M. L. Giganto Arroyo, and F. Sierra-Hidalgo, “Bickerstaff encephalitis after COVID-19,” J Neurol, vol. 268, no. 6, pp. 2035–2037, Jun. 2021, doi: 10.1007/s00415-020-10201-1. [CrossRef]
- M. Gaughan, S. Connolly, S. O’Riordan, N. Tubridy, C. McGuigan, and J. A. Kinsella, “Pediatric Parainfectious Encephalitis Associated With COVID-19,” Neurology, vol. 96, no. 11, pp. 541–544, Mar. 2021, doi: 10.1212/WNL.0000000000011476. [CrossRef]
- N. Ahsan, S. Jafarpour, and J. D. Santoro, “Myelin oligodendrocyte glycoprotein antibody encephalitis following severe acute respiratory syndrome coronavirus 2 in a pediatric patient,” Clin Exp Pediatr, vol. 64, no. 6, pp. 310–312, Jun. 2021, doi: 10.3345/cep.2020.01963. [CrossRef]
- A. Guilmot et al., “Immune-mediated neurological syndromes in SARS-CoV-2-infected patients,” J Neurol, vol. 268, no. 3, pp. 751–757, Mar. 2021, doi: 10.1007/s00415-020-10108-x. [CrossRef]
- Marino Gammazza et al., “Human molecular chaperones share with SARS-CoV-2 antigenic epitopes potentially capable of eliciting autoimmunity against endothelial cells: possible role of molecular mimicry in COVID-19,” Cell Stress Chaperones, vol. 25, no. 5, pp. 737–741, Sep. 2020, doi: 10.1007/s12192-020-01148-3. [CrossRef]
- D. Kanduc, “From Anti-SARS-CoV-2 Immune Responses to COVID-19 via Molecular Mimicry,” Antibodies, vol. 9, no. 3, p. 33, Jul. 2020, doi: 10.3390/antib9030033. [CrossRef]
- E. Gusev, A. Sarapultsev, L. Solomatina, and V. Chereshnev, “SARS-CoV-2-Specific Immune Response and the Pathogenesis of COVID-19,” Int J Mol Sci, vol. 23, no. 3, p. 1716, Feb. 2022, doi: 10.3390/ijms23031716. [CrossRef]
- Sharma and J. Bayry, “High risk of autoimmune diseases after COVID-19,” Nat Rev Rheumatol, vol. 19, no. 7, pp. 399–400, Jul. 2023, doi: 10.1038/s41584-023-00964-y. [CrossRef]
- A. Marino Gammazza et al., “Human molecular chaperones share with SARS-CoV-2 antigenic epitopes potentially capable of eliciting autoimmunity against endothelial cells: possible role of molecular mimicry in COVID-19,” Cell Stress Chaperones, vol. 25, no. 5, pp. 737–741, Sep. 2020, doi: 10.1007/s12192-020-01148-3. [CrossRef]
- G. Lucchese and A. Flöel, “SARS-CoV-2 and Guillain-Barré syndrome: molecular mimicry with human heat shock proteins as potential pathogenic mechanism,” Cell Stress Chaperones, vol. 25, no. 5, pp. 731–735, Sep. 2020, doi: 10.1007/s12192-020-01145-6. [CrossRef]
- A. Felipe Cuspoca, P. Isaac Estrada, and A. Velez-van-Meerbeke, “Molecular Mimicry of SARS-CoV-2 Spike Protein in the Nervous System: A Bioinformatics Approach,” Comput Struct Biotechnol J, vol. 20, pp. 6041–6054, 2022, doi: 10.1016/j.csbj.2022.10.022. [CrossRef]
- F. Madeira et al., “Search and sequence analysis tools services from EMBL-EBI in 2022,” Nucleic Acids Res, vol. 50, no. W1, pp. W276–W279, Jul. 2022, doi: 10.1093/nar/gkac240. [CrossRef]
- S. F. Altschul, W. Gish, W. Miller, E. W. Myers, and D. J. Lipman, “Basic local alignment search tool,” J Mol Biol, vol. 215, no. 3, pp. 403–410, Oct. 1990, doi: 10.1016/S0022-2836(05)80360-2. [CrossRef]
- S. F. Altschul et al., “Protein database searches using compositionally adjusted substitution matrices,” FEBS Journal, vol. 272, no. 20, pp. 5101–5109, Oct. 2005, doi: 10.1111/j.1742-4658.2005.04945.x. [CrossRef]
- Y. Zhang, “TM-align: a protein structure alignment algorithm based on the TM-score,” Nucleic Acids Res, vol. 33, no. 7, pp. 2302–2309, Apr. 2005, doi: 10.1093/nar/gki524. [CrossRef]
- M. Nielsen, C. Lundegaard, and O. Lund, “Prediction of MHC class II binding affinity using SMM-align, a novel stabilization matrix alignment method,” BMC Bioinformatics, vol. 8, no. 1, p. 238, Dec. 2007, doi: 10.1186/1471-2105-8-238. [CrossRef]
- M. Mohkhedkar, S. S. K. Venigalla, and V. Janakiraman, “Untangling COVID-19 and autoimmunity: Identification of plausible targets suggests multi organ involvement,” Mol Immunol, vol. 137, pp. 105–113, Sep. 2021, doi: 10.1016/j.molimm.2021.06.021. [CrossRef]
- C. P. Figueiredo et al., “Zika virus replicates in adult human brain tissue and impairs synapses and memory in mice,” Nat Commun, vol. 10, no. 1, Dec. 2019, doi: 10.1038/s41467-019-11866-7. [CrossRef]
- U. K. Ranawaka, “Emerging tropical neurological infections,” Clinical Medicine, vol. 22, no. 1, pp. 18–20, Jan. 2022, doi: 10.7861/clinmed.2021-0799. [CrossRef]
- L. Zerboni, M. Reichelt, and A. Arvin, “Varicella-Zoster Virus Neurotropism in SCID Mouse–Human Dorsal Root Ganglia Xenografts,” 2010, pp. 255–276. doi: 10.1007/82_2009_8. [CrossRef]
- M. K. Das, A. Sarma, and T. Chakraborty, “Nano-ART and NeuroAIDS,” Drug Deliv Transl Res, vol. 6, no. 5, pp. 452–472, Oct. 2016, doi: 10.1007/s13346-016-0293-z. [CrossRef]
- V. da Silva, F. L. Fontes-Dantas, T. V. Dantas, A. Dutra, O. J. M. Nascimento, and S. V. Alves-Leon, “Shared Molecular Signatures Across Zika Virus Infection and Multiple Sclerosis Highlight AP-1 Transcription Factor as a Potential Player in Post-ZIKV MS-Like Phenotypes,” Mol Neurobiol, vol. 60, no. 8, pp. 4184–4205, Aug. 2023, doi: 10.1007/s12035-023-03305-y. [CrossRef]
- L. C. França et al., “Mimetismo molecular entre o vírus Zika e os distúrbios inflamatórios desmielinizantes do sistema nervoso central: o papel do epítopo NS5 do vírus Zika e dos autoantígenos PLP,” Arq Neuropsiquiatr, vol. 81, no. 4, pp. 357–368, Apr. 2023, doi: 10.1055/s-0043-1768698. [CrossRef]
- L. T. de F. Cavalcante et al., “Buffy Coat Transcriptomic Analysis Reveals Alterations in Host Cell Protein Synthesis and Cell Cycle in Severe COVID-19 Patients,” Int J Mol Sci, vol. 23, no. 21, p. 13588, Nov. 2022, doi: 10.3390/ijms232113588. [CrossRef]
- L. Yan et al., “Cryo-EM Structure of an Extended SARS-CoV-2 Replication and Transcription Complex Reveals an Intermediate State in Cap Synthesis,” Cell, vol. 184, no. 1, pp. 184-193.e10, Jan. 2021, doi: 10.1016/j.cell.2020.11.016. [CrossRef]
- J. Almqvist et al., “Neurological manifestations of coronavirus infections – a systematic review,” Ann Clin Transl Neurol, vol. 7, no. 10, pp. 2057–2071, Oct. 2020, doi: 10.1002/acn3.51166. [CrossRef]
- R. Chang, T. Yen-Ting Chen, S.-I. Wang, Y.-M. Hung, H.-Y. Chen, and C.-C. J. Wei, “Risk of autoimmune diseases in patients with COVID-19: a retrospective cohort study,” EClinicalMedicine, vol. 56, p. 101783, Feb. 2023, doi: 10.1016/j.eclinm.2022.101783. [CrossRef]
- Tesch et al., “Incident autoimmune diseases in association with a SARS-CoV-2 infection: A matched cohort study”, doi: 10.1101/2023.01.25.23285014. [CrossRef]
- D. G. Corrêa, F. C. de Souza Lima, D. da Cruz Bezerra, A. C. Coutinho, and L. C. Hygino da Cruz, “COVID-19 associated with encephalomyeloradiculitis and positive anti-aquaporin-4 antibodies: Cause or coincidence?,” Multiple Sclerosis Journal, vol. 27, no. 6, pp. 973–976, May 2021, doi: 10.1177/1352458520949988. [CrossRef]
- Lotan, S. Nishiyama, G. S. Manzano, M. Lydston, and M. Levy, “COVID-19 and the risk of CNS demyelinating diseases: A systematic review,” Front Neurol, vol. 13, Sep. 2022, doi: 10.3389/fneur.2022.970383. [CrossRef]
- C. Galeotti and J. Bayry, “Autoimmune and inflammatory diseases following COVID-19,” Nat Rev Rheumatol, vol. 16, no. 8, pp. 413–414, Aug. 2020, doi: 10.1038/s41584-020-0448-7. [CrossRef]
- A. Schattner and B. Rager-Zisman, “Virus-Induced Autoimmunity,” Clinical Infectious Diseases, vol. 12, no. 2, pp. 204–222, Mar. 1990, doi: 10.1093/clinids/12.2.204. [CrossRef]
- M. B. A. Oldstone, “Molecular mimicry and immune-mediated diseases,” The FASEB Journal, vol. 12, no. 13, pp. 1255–1265, Oct. 1998, doi: 10.1096/fasebj.12.13.1255. [CrossRef]
- S. Morsy and A. Morsy, “Epitope mimicry analysis of SARS-COV-2 surface proteins and human lung proteins,” J Mol Graph Model, vol. 105, Jun. 2021, doi: 10.1016/j.jmgm.2021.107836. [CrossRef]
- V. A. Petrenko, J. W. Gillespie, L. M. De Plano, and M. A. Shokhen, “Phage-Displayed Mimotopes of SARS-CoV-2 Spike Protein Targeted to Authentic and Alternative Cellular Receptors,” Viruses, vol. 14, no. 2, Feb. 2022, doi: 10.3390/v14020384. [CrossRef]
- K. Karagöz, M. R. Munk, M. Kaya, R. Rückert, M. Yıldırım, and L. Karabaş, “Using bioinformatic protein sequence similarity to investigate if SARS CoV-2 infection could cause an ocular autoimmune inflammatory reactions?,” Exp Eye Res, vol. 203, Feb. 2021, doi: 10.1016/j.exer.2020.108433. [CrossRef]
- Lucchese and A. Flöel, “Molecular mimicry between SARS-CoV-2 and respiratory pacemaker neurons,” Autoimmun Rev, vol. 19, no. 7, p. 102556, Jul. 2020, doi: 10.1016/j.autrev.2020.102556. [CrossRef]
- Y. Lavi et al., “Dysregulated Levels of Circulating Autoantibodies against Neuronal and Nervous System Autoantigens in COVID-19 Patients,” Diagnostics, vol. 13, no. 4, p. 687, Feb. 2023, doi: 10.3390/diagnostics13040687. [CrossRef]
- Giovannoni and G. Ebers, “Multiple sclerosis: the environment and causation,” Curr Opin Neurol, vol. 20, no. 3, pp. 261–268, Jun. 2007, doi: 10.1097/WCO.0b013e32815610c2. [CrossRef]
- T. Olsson, L. F. Barcellos, and L. Alfredsson, “Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis,” Nat Rev Neurol, vol. 13, no. 1, pp. 25–36, Jan. 2017, doi: 10.1038/nrneurol.2016.187. [CrossRef]
- N. Kaushansky, M. Eisenstein, R. Zilkha-Falb, and A. Ben-Nun, “The myelin-associated oligodendrocytic basic protein (MOBP) as a relevant primary target autoantigen in multiple sclerosis,” Autoimmun Rev, vol. 9, no. 4, pp. 233–236, Feb. 2010, doi: 10.1016/j.autrev.2009.08.002. [CrossRef]
- F. Mohammadi-Milasi, K. Mahnam, and M. Shakhsi-Niaei, “In silico study of the association of the HLA-A*31:01 allele (human leucocyte antigen allele 31:01) with neuroantigenic epitopes of PLP (proteolipid protein), MBP (myelin basic protein) and MOG proteins (myelin oligodendrocyte glycoprotein) for studying the multiple sclerosis disease pathogenesis,” J Biomol Struct Dyn, vol. 39, no. 7, pp. 2526–2542, May 2021, doi: 10.1080/07391102.2020.1751291. [CrossRef]
- M. Minohara et al., “Upregulation of myeloperoxidase in patients with opticospinal multiple sclerosis: Positive correlation with disease severity,” J Neuroimmunol, vol. 178, no. 1–2, pp. 156–160, Sep. 2006, doi: 10.1016/j.jneuroim.2006.05.026. [CrossRef]
- E. Gray, T. L. Thomas, S. Betmouni, N. Scolding, and S. Love, “Elevated Activity and Microglial Expression of Myeloperoxidase in Demyelinated Cerebral Cortex in Multiple Sclerosis,” Brain Pathology, vol. 18, no. 1, pp. 86–95, Jan. 2008, doi: 10.1111/j.1750-3639.2007.00110.x. [CrossRef]
- R. M. Nagra et al., “Immunohistochemical and genetic evidence of myeloperoxidase involvement in multiple sclerosis,” J Neuroimmunol, vol. 78, no. 1–2, pp. 97–107, Sep. 1997, doi: 10.1016/S0165-5728(97)00089-1. [CrossRef]
- Lotan, S. Nishiyama, G. S. Manzano, M. Lydston, and M. Levy, “COVID-19 and the risk of CNS demyelinating diseases: A systematic review,” Front Neurol, vol. 13, Sep. 2022, doi: 10.3389/fneur.2022.970383. [CrossRef]
- S. Sedighi et al., “Comprehensive Investigations Relationship Between Viral Infections and Multiple Sclerosis Pathogenesis,” Curr Microbiol, vol. 80, no. 1, p. 15, Jan. 2023, doi: 10.1007/s00284-022-03112-z. [CrossRef]
- S. Mariotto et al., “Clinical spectrum and IgG subclass analysis of anti-myelin oligodendrocyte glycoprotein antibody-associated syndromes: a multicenter study,” J Neurol, vol. 264, no. 12, pp. 2420–2430, Dec. 2017, doi: 10.1007/s00415-017-8635-4. [CrossRef]
- A. Cobo-Calvo et al., “Clinical spectrum and prognostic value of CNS MOG autoimmunity in adults,” Neurology, vol. 90, no. 21, pp. e1858–e1869, May 2018, doi: 10.1212/WNL.0000000000005560. [CrossRef]
- V. Sehgal, P. Bansal, S. Arora, S. Kapila, and G. S. Bedi, “Myelin Oligodendrocyte Glycoprotein Antibody Disease After COVID-19 Vaccination - Causal or Incidental?,” Cureus, Jul. 2022, doi: 10.7759/cureus.27024. [CrossRef]
- M. Netravathi et al., “COVID-19 vaccine associated demyelination & its association with MOG antibody,” Mult Scler Relat Disord, vol. 60, p. 103739, Apr. 2022, doi: 10.1016/j.msard.2022.103739. [CrossRef]
- Lambe et al., “Myelin oligodendrocyte glycoprotein-IgG associated disorders (MOGAD) following SARS-CoV-2 infection: A case series,” J Neuroimmunol, vol. 370, p. 577933, Sep. 2022, doi: 10.1016/j.jneuroim.2022.577933. [CrossRef]
- V. Vasilevska, P. C. Guest, H. G. Bernstein, M. L. Schroeter, C. Geis, and J. Steiner, “Molecular mimicry of NMDA receptors may contribute to neuropsychiatric symptoms in severe COVID-19 cases,” Journal of Neuroinflammation, vol. 18, no. 1. BioMed Central Ltd, Dec. 01, 2021. doi: 10.1186/s12974-021-02293-x. [CrossRef]
- M. Etemadifar, H. Nouri, M. Salari, and N. Sedaghat, “Detection of anti-NMDA receptor antibodies following BBIBP-CorV COVID-19 vaccination in a rituximab-treated person with multiple sclerosis presenting with manifestations of an acute relapse,” Hum Vaccin Immunother, vol. 18, no. 1, Jan. 2022, doi: 10.1080/21645515.2022.2033540. [CrossRef]
- Valadez-Calderon, A. Ordinola Navarro, E. Rodriguez-Chavez, and O. Vera-Lastra, “Co-expression of anti-NMDAR and anti-GAD65 antibodies. A case of autoimmune encephalitis in a post-COVID-19 patient,” Neurología, vol. 37, no. 6, pp. 503–504, Jul. 2022, doi: 10.1016/j.nrl.2021.09.003. [CrossRef]
- M. Salari, B. Zaker Harofteh, and M. Etemadifar, “Autoimmune meningoencephalitis associated with anti-glutamic acid decarboxylase antibody following COVID-19 infection: A case report,” Clin Case Rep, vol. 10, no. 12, Dec. 2022, doi: 10.1002/ccr3.6597. [CrossRef]
- F. Assi, R. Abdallah, A. Mecheik, H. H. Rahhal, and J. Wazne, “Acute Disseminated Encephalomyelitis Following COVID-19 Infection,” Cureus, Jan. 2023, doi: 10.7759/cureus.33365. [CrossRef]
- Mumoli, V. Vescio, D. Pirritano, E. Russo, and D. Bosco, “ADEM anti-MOG antibody-positive after SARS-CoV2 vaccination,” Neurological Sciences, vol. 43, no. 2, pp. 763–766, Feb. 2022, doi: 10.1007/s10072-021-05761-7. [CrossRef]
- Y. Wang, Y. Wang, L. Huo, Q. Li, J. Chen, and H. Wang, “SARS-CoV-2-associated acute disseminated encephalomyelitis: a systematic review of the literature,” J Neurol, vol. 269, no. 3, pp. 1071–1092, Mar. 2022, doi: 10.1007/s00415-021-10771-8. [CrossRef]
- Miyamoto, J. Koh, M. Takahashi, M. Niwa, and H. Ito, “A case of anti-MOG antibody-positive ADEM following COVID-19 mRNA vaccination,” Neurological Sciences, vol. 43, no. 6, pp. 3513–3514, Jun. 2022, doi: 10.1007/s10072-022-06019-6. [CrossRef]
- A. A. Pinto, L. S. Carroll, V. Nar, A. Varatharaj, and I. Galea, “CNS inflammatory vasculopathy with antimyelin oligodendrocyte glycoprotein antibodies in COVID-19,” Neurology - Neuroimmunology Neuroinflammation, vol. 7, no. 5, p. e813, Sep. 2020, doi: 10.1212/NXI.0000000000000813. [CrossRef]
- Naidu and R. Tayler, “Anti N-Methyl-D-Aspartate receptor antibody associated Acute Demyelinating Encephalomyelitis in a patient with COVID-19: a case report,” J Med Case Rep, vol. 17, no. 1, p. 247, Jun. 2023, doi: 10.1186/s13256-023-03979-x. [CrossRef]
- H. K. Al-Hakeim, H. T. Al-Rubaye, D. S. Al-Hadrawi, A. F. Almulla, and M. Maes, “Long-COVID post-viral chronic fatigue and affective symptoms are associated with oxidative damage, lowered antioxidant defenses and inflammation: a proof of concept and mechanism study,” Mol Psychiatry, vol. 28, no. 2, pp. 564–578, Feb. 2023, doi: 10.1038/s41380-022-01836-9. [CrossRef]
- H. K. Al-Hakeim, H. T. Al-Rubaye, A. F. Almulla, D. S. Al-Hadrawi, and M. Maes, “Chronic Fatigue, Depression and Anxiety Symptoms in Long COVID Are Strongly Predicted by Neuroimmune and Neuro-Oxidative Pathways Which Are Caused by the Inflammation during Acute Infection,” J Clin Med, vol. 12, no. 2, p. 511, Jan. 2023, doi: 10.3390/jcm12020511. [CrossRef]
- F. Li and J. Z. Tsien, “Memory and the NMDA Receptors,” New England Journal of Medicine, vol. 361, no. 3, pp. 302–303, Jul. 2009, doi: 10.1056/NEJMcibr0902052. [CrossRef]
- C. Franke et al., “Association of cerebrospinal fluid brain-binding autoantibodies with cognitive impairment in post-COVID-19 syndrome,” Brain Behav Immun, vol. 109, pp. 139–143, Mar. 2023, doi: 10.1016/j.bbi.2023.01.006. [CrossRef]
- S. H. Mueller et al., “Genetic predisposition in anti-LGI1 and anti-NMDA receptor encephalitis,” Ann Neurol, vol. 83, no. 4, pp. 863–869, Apr. 2018, doi: 10.1002/ana.25216. [CrossRef]
- “Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis,” Nature, vol. 476, no. 7359, pp. 214–219, Aug. 2011, doi: 10.1038/nature10251. [CrossRef]
- S. Vieira Alves-Leon et al., “ACUTE DISSEMINATED ENCEPHALOMYELITIS Clinical features, HLA DRB1*1501, HLA DRB1*1503, HLA DQA1*0102, HLA DQB1*0602, and HLA DPA1*0301 allelic association study,” 2009.
- A. De Silvestri et al., “The Involvement of HLA Class II Alleles in Multiple Sclerosis: A Systematic Review with Meta-analysis,” Dis Markers, vol. 2019, pp. 1–7, Nov. 2019, doi: 10.1155/2019/1409069. [CrossRef]
- F. Barcellos et al., “Heterogeneity at the HLA-DRB1 locus and risk for multiple sclerosis,” Hum Mol Genet, vol. 15, no. 18, pp. 2813–2824, Sep. 2006, doi: 10.1093/hmg/ddl223. [CrossRef]
- F. L. Fontes-Dantas et al., “SARS-CoV-2 spike protein induces TLR4-mediated long-term cognitive dysfunction recapitulating post-COVID syndrome in mice,” Cell Rep, p. 112189, Feb. 2023, doi: 10.1016/j.celrep.2023.112189. [CrossRef]
- I. Ismail and S. Salama, “A systematic review of cases of CNS demyelination following COVID-19 vaccination,” J Neuroimmunol, vol. 362, p. 577765, Jan. 2022, doi: 10.1016/j.jneuroim.2021.577765. [CrossRef]
- P. Flannery, I. Yang, M. Keyvani, and G. Sakoulas, “Acute Psychosis Due to Anti-N-Methyl D-Aspartate Receptor Encephalitis Following COVID-19 Vaccination: A Case Report,” Front Neurol, vol. 12, Nov. 2021, doi: 10.3389/fneur.2021.764197. [CrossRef]
- H. C. Lee et al., “Anti-N-Methyl-D-Aspartate Receptor Encephalitis after BNT162b2 COVID-19 Vaccination,” J Epilepsy Res, vol. 12, no. 2, pp. 71–73, Dec. 2022, doi: 10.14581/jer.22013. [CrossRef]
- S. Jarius, N. Bieber, J. Haas, and B. Wildemann, “MOG encephalomyelitis after vaccination against severe acute respiratory syndrome coronavirus type 2 (SARS-CoV-2): case report and comprehensive review of the literature,” J Neurol, vol. 269, no. 10, pp. 5198–5212, Oct. 2022, doi: 10.1007/s00415-022-11194-9. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



