
Submitted:
02 October 2023
Posted:
02 October 2023
You are already at the latest version
Abstract
Material and methods. The trial was carried out over two years in Southern Italy. Two grapevine rootstocks, 110R and SO4, were compared to evaluate the water extraction ability from the soil and the effect on the yield and quality of the Cardinal grapevine table cultivar. Therefore, a new approach to plant water consumption based on sap flow was adopted.
Results. The earlier and faster water refilling of the xylem in Cardinal onto 110R (C/110R) appears responsible for the earlier evolution of the phenological phases than Cardinal onto SO4 (C/SO4). The maximum length of the principal shoot was reached in Cardinal/110R compared to C/SO4, while a higher number of lateral shoots with lower internode has changed canopy architecture and light distribution in C/SO4. The 110R used more water compared to SO4; it was possible to quantify the real transpired flux of the plant per day: sap flow was 12.3 L.plant-1.d-1 and 11.7 L.plant-1d-1 in C/110R in the first and second year, respectively, while it was 14% lower in alternative graft combination.
Conclusions. However, C/SO4 was able to sustain leaf water status and physiological mecha-nisms, although with lesser performance than C/110R, but that did not cause negative effects on production parameters.
Keywords:
drought
; phenology
; quality
; sap flow
; viticulture
1. Introduction
Grafting is a propagation method that has been used for millennia [1] and is currently widely used in grapevine culture. The graft of Vitis vinifera scion onto a rootstock of Vitis spp. americana (Vitis berlandieri Planch, Vitis riparia Michx., and Vitis rupestris Scheele) or their interspecific hybrids, was a necessary propagation method to contrast phylloxera damage on grapevines at the end of the 19th century [2]. This insect damaged the root system of Vitis vinifera, but not that of Vitis americana, which is poorly suited to the production of quality grapes. Therefore, the graft on rootstock become an essential practice for planting new vineyards [3], and subsequently, it has allowed it to overcome other biotic and abiotic adversities, such as drought [4]. In the Mediterranean environment, water is the most limiting resource for viticulture; indeed, grapevine trees are often exposed to water deficit conditions during the growth period and this phenomenon has been exacerbated in the last years such to the effect of climate change [5]. Drought conditions affect optimal water transport through the soil-plant-air continuum, which is interconnected by a continuous film of water [6]. However, the uptake of water could be increased in grafted plants using drought-tolerant rootstocks [7,8,9,10] such as V. Rupestris or its hybrid. Previous studies indicated that 110R hybrid rootstock had a strong drought tolerance with its deep-growing and well-branched root system in the field [9,11]; the root hydraulic conducibility is also very important [10] to overcome soil drought conditions. Furthermore, is to be considered that a cyclic correlation is generated between the shoot and the root system, whereby the behaviour of the shoot depends on the substances absorbed and synthesised by the root (hormones) and the behaviour of the root system depends by the substance synthesised in the shoot. This correlation is modified in graft plants. Indeed, the graft of scion onto a rootstock generates a new individual with two different genotypes whose behaviour will depend on the mutual interaction between them. Therefore, the same scion genotype may have a different result when grafted onto two different rootstocks in the same cultivation conditions. Climate change is upsetting the established relationships between plants and the environment and the use of rootstock should be re-evaluated using the new technology available. Indeed, evaluations of drought tolerance induced by rootstocks were based primarily on vegetative vigour (trunk circumference), fruit attributes (berry size, berry colour estimate, total soluble solids and total acids) and yield [12]. More recent studies have incorporated physiological indicators, such as stomatal conductance [13,14], leaf water potential [15,16,17,18], root hydraulic conductivity [10] and, recently, sap flow measurements [19]. The sap flow with HFD (Heat Field Deformation) technology allowed the study of water pathways in the xylem. During the day water moves from the roots to the canopy and subsequent loss to air [day-time transpiration (Ed)]. Instead, during the night, the water pathways can be two: from the roots to the canopy and subsequent loss to air [night-time transpiration (En)] and movement from one part of the root system to another and subsequent loss to the soil [Hydraulic Redistribution (HR)] [20]. In this trial, two hybrid rootstocks [(V. berlandieri Planch x V. rupestris Scheele) and (V. berlandieri Planch x V. riparia L.)] were compared to evaluate the water extraction ability from the soil and the effect on yield and grape quality of a table cultivar in a Mediterranean environment where temperatures in recent years have increased. Furthermore, a new approach based on sap flow measurement integrated with a tomograph image was proposed to evaluate plant water consumption to goal water saving used in the climate change context.
2. Materials and Methods
2.1. Experimental site, plant material and experimental design
The experiment was conducted over two years (2021–2022) on Maiorana Farm, in Curinga (CZ) Italy (38°51ʹ42.8” N, 16°15ʹ45.8” E). The table grapevine trees “Cardinal” cultivar had been grafted onto 110R (Vitis berlandieri Planch x Vitis rupestris Scheele) and SO4 (Selection OPPENHEIM 4) (Vitis berlandieri Planch x Vitis riparia L.) rootstock and has been planted in the spring of 2006 in subacidic (pH 6) sandy soil, containing 2.12% organic matter and 1.6 g·kg−1 nitrogen content. The plants had been spaced 2 m × 2m apart (2500 plants ha−1), and the trees had been trained to “Tendone” system with 4 cordons. The water was administered every two days with 4 drippers per plant to 4 L.h-1 per 1 hour (16 L.day-1) every three days. The experiment consisted of three blocks with four trees per block and per graft combination.
2.2. Phenological surveys
The main phenophases were monitored every week, using the BBCH scale (Biologische Bundesanstalt, Bundessortenamt and Chemical industry) on spurs pruned two buds, distributed along two of four cordons per plant. The observations were started in the first days of March (03 Mar 2021; 04 Mar 2022) (BBCH 00) and concluded in the third ten days of July (22 Jul 2021; 22 Jul 22) (BBCH 89).
Shoot growth was monitored every week on alternating spurs distributed on two cordons oriented East and West. For each shoot measured, the number of nodes and the number of anticipated shoots (lateral shoot) were detected before harvesting; at this time a sample of 24 leaves (2 leaves per plant randomly selected) was taken from each graft combination and the leaf area was determined in the laboratory using an area meter (Li-Cor 3100, LI-COR, Lincoln, Nebraska, USA). Finally, the leaf area per shoot, per plant, and LAI (leaf Area Index) were calculated.
2.3. Gas exchange and radiometric measurements
The gas exchange and PPFD (Photosynthetic Photon Flux Density) measurements were carried out during a clear sunny summer day (from 11:00 a.m. to 01:00 p.m.), on four clean days during the last week of the summer months (June, July, and August) in both years. Gas exchange measurements [net assimilation of CO2, (An), stomatal conductance (gs), and internal CO2 concentration (Ci)] were performed on leaves completely developed located in the central layer of the canopy (six leaves × four plants × three blocks) using a portable photosynthesis system (Li-Cor 6400XT; LI-COR Biosciences, Lincoln, Nebraska, USA). Subsequently, on the same leaves the SPAD index was measured using a SPAD 502 instrument (Spectrum Technologies Inc., Aurora, Illinois, USA). It was possible to measure the PPFD inside the canopy as well as the radiation that reaches the ground. The measurements were conducted at noon on the last day of July during the days with clean sky using a SunScan Probe (Mod. SS1, Delta-T Devices Ltd, Burwell, Cambridge, UK).
2.4. Tomography measurements
The distribution of the conductive and resistive areas of the xylem was obtained using a (Picus TreeTronic3®, Argus, Berlin, Germany, UE) in the trees of the two grafted combinations. The electrical resistance of the wood is influenced by the moisture content of the wood tissue. In the course of the scans, the instrument transmits electrical voltages (Ω Ohm) to the wood up to 100 volts at all measuring points (PM) (up to 24) positioned around the circumference of the sample tested. The tomograph software calculated the apparent electrical resistance of the xylem and outputs it as a tomogram, a 2D colour map.
2.5. Leaf water potential
The leaf water potential (ΨL) was determined on the same leaves on which gas exchange and fluorescence measurements were conducted. The leaf water potential was expressed in units of pressure (Bar) using the Schölander pressure chamber (PMS Instrument Company, Oregon, USA).
2.6. Sap Flow–HFD
Sap Flow Meter HFD8-100 (ICT International, Armidale, NSW, Australia) were installed for xylem flow measurement. The HFD technique combines the most advantageous features of the previously developed methods, measuring asymmetric and symmetric temperature gradients (dTsym and dTas), around a heat source, and avoids the limitations associated with the arrangement of previous sensors. In addition, the sensor configuration using the HFD method involves placing the upper thermometer of the asymmetrical thermocouple next to a heater, which allows it to track heat dissipation and deformation in both axial and tangential directions around a linear heater. The Sap Flow Meter HFD8-100 can measure high sap flow rates as well as low to zero and reverse sap flow. The needles have sensors distributed on all lengths spacing 5 mm between them.
2.7. Total sap flow measurement
The measurement of the xylem flow detected by the Sap Flow Meter HFD8-100 was expressed in cm3.cm-2.h-1. The average value detected by the various sensors distributed every 0.5 cm along the probe (needle) of the Sap-Flow HFD was determined, which in turn was averaged over the entire day. This last parameter multiplied by the conductive area (detected and determined with the tomograph) was used to calculate the Daily Total Sap Flow.
2.8. Harvest
The yield per plant (weight and grapes number) was determined at harvest. Two grape bunches per tree (8 grape bunches per block per graft combination = 36 grape bunches per graft combination) were randomly collected and taken to the “Colture Arboree” Laboratory, of AGRARIA Department, at the Mediterranean University of Reggio Calabria.
2.9. Winter pruned
Each year at the end of July, the pruning material was weighed per plant.
2.10. Biometric measurements, maturation indexes, and nutraceutical parameters
The fresh weight (FW) of clusters and the rasp were determined using an analytical balance (BJ 610C, Precisa Instruments AG, Dietikon, Switzerland). For each grape bunch, the berries were detached and the fresh weight of the rasp was measured. Furthermore, 100 berries randomly taken per plant were used to determine the height (H), diameter (D), and fresh weight using an electronic balance (Mettler-Toledo, Grelfensee, Switzerland). For each berry the veraison surface was determined, for which purpose a scale of values was prepared: 0 (0% of the surface area covered), 1 (<50% of the surface area covered), 2 (>50% of the surface area covered) and 3 (100% of the surface area covered). Five groups of ten berries were used to juice extract. The juice was used to measure the total soluble solids (TSS), using a handheld digital refractometer (PR-1; Atago, Tokyo, Japan), and titratable acidity (TA), using 10 mL of juice diluted with distilled water (1:1) and titrated to pH 8.2 with 0.1 N NaOH (%). The TSS/TA ratio was calculated. On five other groups of 10 berries, the analyses of total polyphenol content (TPC) and total antioxidant capacity (TAC) were performed. The pulp portion (20 g), removed before juice extraction, was homogenized using an Ultraturrax blender (20.000 rpm, T 25 Basic; IKA, Werke, Germany, UE). The TPC and TAC were analyzed separately using a Lambda 35 spectrophotometer (PerkinElmer Corporation, Waltham, Massachusetts, USA). Before measuring the TPC and TAC, standard curves were prepared for each test. The TPC (mg gallic acid equivalents g−1 FW) was determined using the Folin–Ciocalteu method [25]. The TAC was determined using the modified TEAC assay and expressed as mmol Trolox, equivalent to g−1 FW [26,27].
2.11. Statistical Analysis
All data were subjected to two-way ANOVA tests (treatments and years), and the means were compared by using Tukey’s test when the ANOVA indicated significant (P < .05) variable effects. All data analyses were performed using IBM® SPSS® Statistic version 22 (SPSS Inc. IBM Company, Armonk, New York, USA).
3. Results
An earlier water refiling of the xylem (more bluish area compared to the reddish area in the tomographs) (Figure 1), was observed in the Cardinal grafted on 110R than in those grafted on SO4 in the first ten days of March in both years.
In fact, the Cardinal onto 110R showed a conductive xylem area (CXA) of 66% in 2021 and 61% in 2022, whereas CXA was significantly lower in the plants grafted on SO4, 49% and 44%, respectively in 2021 and 2022 (Table 1). No difference was observed in the phenological stage between the two graft combinations during the first ten days of March month in both years. Indeed, the buds were at BBCH 0 'winter bud' in the two grafting combinations (Table 2).
At the beginning of the second ten days of March 2021, the tomograph shows an increase in the conductive area in C/SO4 compared to the previous survey reaching 54% of the total xylem area (Figure 1; Table 1). At this time, the CXA value in Cardinal onto 110R was similar to the previous survey and it was still significantly higher than the C/SO4 graft combination (Table 1); this trend was observed in the next year too. The differences in phenophases evolution between the two graft combinations were clear at the beginning of the second ten days of March in the first year, whereas it appeared at the end of the second ten days of March in the next year. The fast water refilling of the xylem in Cardinal onto 110R appears responsible for the earlier evolution of the phenological phases than Cardinal onto SO4. In fact, at the end of the second ten days of March, the buds in BBCH 5 phase were 15% and 27% in C/SO4, in 2021 and 2022 respectively (Table 2 and Table 3), whereas the buds including from the BBCH 5 phase to BBCH10 phase in C/110 R were 45% in 2021 and 36 % in 2022. At the end of March, the buds in BBCH 0 stage in C/SO4 were 64% and 70% in the first and second observation years, respectively (Table 2 and Table 3), whereas the percentage was lower for the alternative graft combination in both years: 18% during 2021, and 42% in 2022 (Table 2 and Table 3). It is a clear faster bud-break with 110R than SO4 until this time. At the end of April, the conductive area was significantly higher in C/110R than in C/SO4 (Figure 1, Table 1). However, the percentage of buds unbroken (BBCH 0) was similar between the two graft combinations; whereas the percentage of buds in the BBCH 55 stage was 20% higher in C/110R than in C/SO4 (58.97%), in the first year (Table 2). In the next year, the BBCH 55 stage was always higher in C/110R (27.06%) than in C/SO4 (8.33%) (Table 3). During the first ten days of May, the percentage of buds in the BBCH 57 stage reached 56% onto C/SO4 and it was 12% higher onto C/110R in 2021 (Table 2). In the next year, the slow phenophases evolution of Cardinal onto SO4 was observed; indeed the buds percentage at BBCH 57 phase were 26% and 48% in C/SO4 and C/110 R, respectively (Table 3). In the second ten days of May, the buds in the phase (BBCH 65) were 64% on C/SO4 and 78% on 110R in the first observation year (Table 2), whereas in the second year, the percentages were 32% and 58%, for C/SO4 and C/110R, respectively (Table 3). Therefore, the delay in the evolution of phenophases was confirmed for both graft combinations. The percentage of buds in the BBCH 71 stage was 71% in C/SO4 during the first days of June in the first observation year (Table 2), whereas the alternative graft combination had a high percentage of buds in the more advanced stage (BBCH 75) (82%) in the same date (Table 2). In the next year, always during the first days of June, the two graft combinations were near with regard phenophases; indeed 55% and 61% of buds were in the BBCH 71 stage onto SO4 and 110 R graft combinations, respectively (Table 3). At the end of July, during the first year, a high percentage (58 %) of the buds were in the BBCH 77 stage in the C/SO4 combination, while 75 % of the buds were in a more advanced stage, BBCH 81, in the C/110R combination (Table 2). In the following year, the percentage of buds in stage BBCH 81 was 20% higher in C/110R than in C/SO4 (Table 3). Therefore, the different evolution of the phenophases continues to be evident between the two grafting combinations; it was always delayed in C/SO4 compared to C/110R, but the percentage of sprouting was not different in both years. In the third ten days of July, the conductive xylem area was not different between the two graft combinations (Table 2). The shoot growth trend, recorded weekly from the third ten days of April to the third ten days of July, was linear and showed a clear significant difference between the two grafting combinations in both years (Figure 2a,b).
The maximum length was reached in the shoots of the Cardinal plants grafted on 110R. Indeed the growth stopped around 180 cm (2021) and 210 cm (2022) onto C/110 R (Figure 2a,b), whereas in the alternative graft combination, the shoot stopped its growth at a length of 145 cm in 2021 and at 160 cm in 2022 (Figure 2a,b). The percentage difference in shoot length between the two graft combinations was similar between two years, about 35 %. The final internode length was significantly higher in C/110R than in alternative graft combinations (17% and 13%, in the first and second year, respectively) (Table 4).
The overall number of leaves overall leaf area per shoot (principal shoot and lateral shoot/s), and leaf area per plant were not statistically different, in both years (Table 4). The number of leaves and leaf area per principal shoot was significantly higher in C/110 R than in C/SO4. Instead, the leaf number of the lateral shoot and his leaf area were higher in C/SO4 than in C/110R (Table 4). Furthermore, the branching of the principal shoot was higher in C/SO4 than in C/110R (Table 4). Then, the principal shoots of the Cardinal/SO4 were characterised by a significantly higher number of lateral shoots per unit measure of the primary shoot, in both years. This aspect is very important because it causes a different architecture inside the canopy of the "Tendone" system between two grafted combinations and different light distributions within the canopy. However, the total pruned material was significantly different between the two graft combinations; it was higher on C/SO4 in both years (Table 4). Indeed, the PPFD measurement had shown values of 53 % and 61% higher in the middle layer of the C/110R canopy, than those found in the plants grafted onto SO4, respectively, in the first year and second year. The light that reaches the soil was also 37% higher in C/110 R (Figure 3) in both years.
Furthermore, the temperature measured on the berry surface showed a significant increase of approximately 1.3 °C on the bunch of C/110R than C/SO4 as an effect of higher solar exposition of the berry. This effect was recorded in both years. (29.60±0.29 C/110 R, 2021- 28.27±0.164 C/110R in 2022 year; 28.45±0.33 C/SO4 in 2021; 27.15±0.14 C/SO4 in 2022 year). The Pn was statistically higher in the leaf of C/110R than C/SO4 in both years (15% and 30%, respectively in 2021 and 2022 year) Table 5. Stomatal conductance was also statistically higher in the C/110R graft combination (0.086 mol H2O m-2 leaf area s-1, 2021; 0.090 mol H2O m-2 leaf area s-1; 2022) than in the C/SO4 (0.073 mol H2O m-2 leaf area s-1, 2021; 0.072 mol H2O m-2 leaf area s-1, 2022), Table 5. In addition, plants grafted on SO4, showed lower leaf water potential (-5.62±0.15, 2021; -5.37±0.20, 2022) than on 110R, respectively (-4.56±0.11, 2021, -4.64±0.10, 2022); furthermore, no differences were observed in the Spad Index; indeed, the values were similar between the two graft combinations over two observation years (Table 5). At the end of July, no differences were observed in the xylem conductive area (Figure 1; Table 1), but differences appeared in sap flow measurements along the needles of the sap flow meter (Figure 4).
The higher water uptake of C/110R than C/SO4 was confirmed by the data obtained from plant-based sap flow sensors (Table 5, Figure 4). In C/SO4, the flow through the sap flow meter needles was only higher at 3.5 cm from the outside to the inside of the xylem, compared to the alternative grafted combination; at other needle points, the flux was always higher in C/110 R. With 110R rootstock, the xylem of the Cardinal was active from 0.5 cm up to a xylem depth of 5.5 cm, whereas with the alternative rootstock, the flux was detected up to a xylem depth of 3.5 cm (Figure 4). Therefore, the overall flux was higher in C/110 R than in C/SO4. Considering the average sap flow measurement, the xylem flow was 20/25% (2021-2022) higher on the Cardinal grafted on 110R (25.34 cm3 cm-2 h-1), compared to the Cardinal grafted on SO4 (20.33 cm3 cm-2 h-1) (Table 5). Then, it was possible to quantify the real transpired flux of the plant per day: sap flow was 12.33 l.plant-1.d-1 and 11.74 l.plant-1.d-1 in C/110R, respectively in the first and second year, whereas it was 14% lower in alternative graft combination in both years (Table 5). A nocturnal redistribution of xylem flux from the epigeal to the hypogean portion is most evident in C/SO4, with a negative peak at dawn. No statistically significant differences were observed in production parameters (yield, number of bunches per plant, bunch weight) between the two graft combinations; the number of bunches per plant averaged close to 60 in the first year and around 50 in the second year, whereas the production was 21% lower in the second year than first year (Table 6).
The values were statistically higher in the first year (Table 6). The sprouting bud percentage was not influenced by rootstock (Table 2 and Table 3), and the real fertility was also similar between the two graft combinations, but it was lower in the second year (Table 6). As regard the biometric parameters, at harvest (BBCH 89), the bunch weight was not significantly different between the two graft combination, but it was 16% lower in the second year (Table 6). This lower weight can be attributed to the lower berry number per bunch during the second year (Table 6). From the first to second sample time, the biometric measurements of the berry showed no significant statistical differences between the two graft combinations (Table 7); indeed, the berry reached the highest weight and size during the end of July onto 110R whereas the berry must wait for the beginning august to reach the highest biometric parameters onto SO4 (Table 7). The lower pigmentation of peel was observed during the first and second weeks of July on berries of Cardinal grafted onto 110 R (Table 7). However, the colour peel did not show significant differences between the two graft combinations during the third ten days of July and at harvest (2.36±0.11 in the C/110R and 2.50±0.12 in the C/SO4). The behaviour of biometric parameters was similar in both two years, whereas was different for degree ripening; however, no interaction was recorded for all parameters reported in Table 8.
The pattern of Total Soluble Solids t and pH in the juice of the berry was similar between graft combinations and between years and increased from the first week of July to the first ten days of August in both years (Table 8). At the beginning of August, the TSS was 14.74 °Brix (±0.72) in the C/SO4 and 14.48 °Brix (±0.30) in the 110R, whereas pH was 3.50 in both graft combinations. Furthermore, the evolution during the samplings of titratable acidity and TSS/TA did not change between years but between the two graft combinations. Indeed, titratable acidity reached an average lower concentration in C/110R than in C/SO4, already at the end of July (Table 8). As a consequence of this, the sugar/acid ratio reached a value significantly higher in the 110R (23.66±0.71) than in C/SO4 (19.18±0.75) at the end of July without differences in both two years. That indicates that the Cardinal cultivar, grafted onto 110R, had reached the organoleptic characteristics appreciated by consumers well in advance. Indeed, at the last sampling, the SSR/TA ratio was 31.68 (±0.27) and 28.84 (±0.53) in the 110R and SO4 grafted combination, respectively (Table 8). No interaction year x treatment was observed. As regards nutraceutical analyses, the Total Polyphenol Content recorded in the pulp and of the berry, values were overlapping between two graft combinations (Table 9).
No differences were observed between the two years and no interaction was recorded. Instead, the value of TPC was significantly higher in the C/110 R than C/SO4 at the end of July and the TAC of peel was also significantly higher in berries of C/110 R than C/SO4. At the beginning of August, the TPC decreased in the berry of both graft combinations; the TAC decreased also in the berry of C/110 R whereas increased in the berry of C/SO4 reaching values similar to C/110 R.
4. Discussion
Growing buds require a carbohydrate supply, mostly mono- or disaccharides (i.e., glucose, fructose, and sucrose). Furthermore, in the weeks before the bud-burst, the xylem fills with water and the pressure in the xylem builds to high levels [28]. In grapevine, it is shown that there is low starch variation near the bud during bud-break, whereas there is an increase in starch degrading enzyme activity and significant decreases of starch reserves at distant locations [29,30,31,32] and xylem sap becomes enriched as well as sugars also phytohormones. The cytokinin is mainly produced in root tips and moves from root tips to the bud through the xylem: it controls growth and shoot development [33]. A higher cytokinin content was found in 110R rootstocks compared to, a progenitor of SO4 used in our experiment, the Kober 5BB [34]; a delay of bud–break was observed with this last rootstock; however, no differences in final bud-break percentage between the two graft combinations. Tomograph analyses effected can explain this. Indeed, the tomograph has shown that SO4 has a slowdown in xylem filling in spring compared to 110R and this can be one cause of the delayed bud break Cardinal onto SO4. Following this hypothesis, it is possible to assume in combination with 110R, cytokinins and carbohydrate supply by reserves at distant locations reach the buds earlier than in the alternative combination causing earlier sprouting. Many workers have reported that the size of the root system is an important factor in determining shoot growth. In our experiment, rootstock extensions the growth of the principal shoot with a leaf-number increase; therefore, the primary effect of 110R rootstocks was on the cell extension of the shoot: higher length of principal shoot and his internodes than to C/SO4. However, no differences were observed in the overall leaf area of the principal shoot and lateral shoot between the two graft combinations, Indeed, the total leaf area of lateral shoots was higher on SO4 rootstock than on 110 R, whereas the leaf of the principal shoot was higher in C/110 R than C/SO4. However, the shorter internode and higher number of short lateral induced on Cardinal by SO4 have caused a change of canopy architecture with lower gaps in C/SO4 than in C/110R grafted vines without change in final biomass removed with pruned weight. Therefore, the increase of solar radiation and the temperature inside the canopy in Cardinal canopy grafted onto 110R did not adversely affect photosynthetic activity. Indeed, light exposure generally stimulates anthocyanin accumulation in grape berries, but, conversely, high temperatures inhibit color formation [35,36]. Ref. [35] reported that the optimal temperature range for anthocyanin accumulation was between 17 and 26 °C. Therefore, during the ripening Cardinal, in July month, the slower coloration on the berries of Cardinal/1110R was probably a consequence of the higher temperature (29.60 °C±0.29) of the clusters more exposed to light than in the Cardinal/SO4 combination (28.27±0.16), as confirmed by previous studies [35]. Furthermore, it has been reported that more shaded bunches have a higher acidity [37]. Then, the lower acidity recorded on berry of C/110R can be also attributed to the greater exposure of the grapes to the higher light and increasing temperature, compared to the grapes of plants grafted on SO4. Furthermore, the berries of C/110R reached higher sweetness early compared to C/SO4, but with lower peel coloration compared to the alternative graft combination. Leaf water potential and gas exchange parameters were always better on the 110R graft combination than with SO4 rootstock, supporting the higher drought tolerance of the 110R rootstock [7]. Indeed, the overall xylem flux transpired by the plant was higher in the graft combination on 110R than on SO4; this showed a higher root water extraction from the rhizosphere by the V. berlandieri x V. rupestris rootstocks. In addition, the prevailing flow in the outermost part of the xylem might depend on the more superficial roots, as detected in C/SO4, while, the inner one, on the deeper roots, as detected in C/110R. The An/gs and An/Ci detected in the leaf of Cardinal grafted onto 110R were similar to that recorded in the alternative graft combination. The biomass production was higher in C/SO4 than in C/110 R as shown by the pruned material weight. The higher pruned weight is attributed to weight lateral shoot axes because the Cardinal onto SO4 produce more lateral shoots than onto 110 R, but no differences were recorded about the overall leaf area per plant. Because no difference was recorded to yield, it is hypothesised that the Cardinal/onto 110R has a greater accumulation of dry matter in the reserve tissues, which contributed to the earlier spring vegetative resumption. Interesting was the behaviour of sap flow change in the Cardinal by changing the rootstock. It is evident that the conductive part of the xylem detected by the sap flow needle is deeper in the Cardinal grafted on 110R than in the combination with SO4, resulting in a higher daily water flow potential. In the C/SO4 combination, however, a flux perturbation is evident at sunrise caused by downward and upward flow, resulting in a negative peak. As regards the production parameters, the 110R rootstock has influenced the ripening and sweetness that was anticipated, but the rootstock did not influence the other maturation indexes, yield, and nutraceutical parameters.
5. Conclusions
Under the drought conditions of southern Italy, the 110 R rootstock uses, also, the water present in the deeper layers, accumulated during the rainy season due to its structure characterized by deep roots, improving photosynthesis, elongation of the principal shoot with a lower number of lateral shoots. In the berry of C/110 R, the sweetness was reached early, but with lower peel coloration compared to the berry of C/SO4. The SO4 rootstock changed the pattern of shoot growth and micro-environmental condition in the canopy compared to 110R, favouring the colouring of the pulp, but delaying the increase in sweetness. No other variations were found for the other ripening indices, nutraceutical parameters and production per plant. The higher biomass produced in C/SO4 could be one of the factors responsible for the slower sprouting of C/SO4 because the plant uses more photosynthesis products in the construction of lateral shoots. However, SO4 has shown to be a good user of irrigation water and, therefore, a viable rootstock in surface soils with little water reserve. A study on the root synthesis of shoot-promoting hormones could make a further contribution to the results of the experiments conducted. The use of the two rootstocks could be exploited to have an earlier production with the 110 R rootstock and more pigmented with the SO4.
References
- Mudge, K.; Janick, J.; Scofield, S.; Goldschmidt, E.E. A history of grafting. Hortic Rev. 2009, 35, 437–493. [Google Scholar]
- Gautier, A.T.; Chambaud, C.; Brocard, L.; Ollat, N.; Gambetta, G.A.; Delrot, S.; et al. 2019. Merging genotypes: graft union formation and scion–rootstock interactions. J. Exp. Bot. 2019, 70, 747–755. [Google Scholar] [CrossRef]
- Granett, J.; Walker, A.M.; Kocsis, L.; Omer, A.D. Biology and management of grape Phylloxera. Annu. Rev. Entomol. 2001, 46, 387–412. [Google Scholar] [CrossRef] [PubMed]
- Galet, P.; Smith, J. (1998) Grape varieties and rootstock varieties. Collection Avenir oenologie (Oenoplurimédia: Chaintré, France).
- Jones, G.V. Climate, grapes, and wine: Structure and suitability in a changing climate. Acta Hortic. 2012, 931, 19–28. [Google Scholar] [CrossRef]
- Janott, M.; Gayler, S.; Gessler, A.; Javaux, M.; Klier, C.; Priesacket, E. A one-dimensional model of water flow in soil–plant systems based on plant architecture. Plant Soil 2011, 341, 233–256. [Google Scholar] [CrossRef]
- Pongrácz, D.P. Rootstocks for grapevines. David Philip Publisher: Cape Town, South Africa, 1983.
- Martínez-Ballesta, M.C.; Alcaraz-Lopez, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological aspects of rootstock-scion interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Serra, I.; Strever, A.; Myburgh, P.; Deloire, A. The interaction between rootstocks and cultivars (Vitis vinifera L.) to enhance drought tolerance in grapevine. Aust. J. Grape Wine Res. 2014, 20, 1–14. [Google Scholar] [CrossRef]
- Gullo, G.; Dattola, A.; Vonella, V.; Zappia, R. Evaluation of water relation parameters in vitis rootstocks with different drought tolerance and their effects on growth of a grafted cultivar. J. Plant Physiol. 2018, 226, 172–178. [Google Scholar] [CrossRef]
- Sucu, S.; Yagcı, A. Determination of sapling yield and quality features in some rootstocks and Sultanı Çekırdeksız (Vitis Vinifera L.) variety grafted on these rootstocks. Ege Üniv. Ziraat Fak. Dergisi 2017, 54, 53–59. [Google Scholar] [CrossRef]
- Lider, L.L. A grape rootstock trial in the Napa Valley: a nine-year progress report of three stocks on bale loam soil. Am. J. Enol. Vitic. 1957, 8, 121–126. [Google Scholar] [CrossRef]
- Carbonneau, A. The early selection of grapevine rootstocks for resistance to drought conditions. Am. J. Enol. Vitic. 1985, 36, 195–198. [Google Scholar] [CrossRef]
- Iacono, F.; Peterlunger, E. Rootstock-scion interaction may affect drought tolerance in Vitis vinifera cultivars. Implications in selection programs. Acta Hortic. 2000, 528, 543–554. [Google Scholar] [CrossRef]
- Ezzahouani, A; Williams, L. E. The influence of rootstock on leaf water potential, yield, and berry composition of Ruby Seedless grapevines. Am. J. Enol. Vitic. 1995, 46, 559–563. [Google Scholar] [CrossRef]
- Choné, X.; Van Leeuwen, C.; Dubourdieu, D.; Gaudillère, J. Stem water potential is a sensitive indicator of grapevine water status. Ann. Bot. 2001, 87, 477–483. [Google Scholar] [CrossRef]
- Deloire, A.; Carbonneau, A.; Wang, Z.; Ojeda, H. Vine and water: a short review. J. Int. Sci. Vigne Vin 2004, 38, 1–13. [Google Scholar] [CrossRef]
- Williams, L.E. Interaction of rootstock and applied water amounts at various fractions of estimated evapotranspiration (ETc) on productivity of Cabernet Sauvignon. Aust. J. Grape Wine Res. 2010, 16, 434–444. [Google Scholar] [CrossRef]
- Nadezhdina, N. A simplified equation for sap flow calculation based on the Heat-Field Deformation (HFD) measurements. Acta Hortic. 2012, 951, 11. [Google Scholar] [CrossRef]
- Howard, A.R.; Van Iersel, M.W.; Richards, J. H.; Donovan, L. Night-time transpiration can decrease hydraulic redistribution. Plant, Cell Environ. 2009, 32, 1060–1070. [Google Scholar] [CrossRef]
- Nadezhdina, N. Temperature gradients around a linear heater due to moving sap. In: Proceedings of the “4th International Workshop on Measuring Sap Flow in Intact Plants” (Cermák J, Nadezhdina N eds). Zidlochovice (Czech republic). Publishing House of Mendel University, Brno, Czech Republic, 1998, pp. 65–71.
- Nadezhdina, N.; Tributsch, H.; Èermák, J. Infra-red images of heat filed around a linear heater and sap flow in stems of lime trees under natural and experimental conditions. Ann. Sci. For. 2004, 61, 203–213. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Èermák, J.; Neruda, J.; Prax, A.; Ulrich, R.; Nadezhdin, V.J.; Gašpárek, J.; Pokorny, E. Roots under the load of heavy machinery in spruce trees. Eur. J. For. Res. 2006, 125, 111–128. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Vandegehuchte, M.; Steppe, K. Sap flux density measurements based on the heat field deformation method. Trees – Structure and Function, 2012.
- Slinkard, K.; Singleton, V.L. Total phenol analysis: automation and comparison with manual methods. Am. J. Enol. Vitic. 1997, 28, 49–55. [Google Scholar] [CrossRef]
- Pellegrini, N.; Re, R.; Yang, M.; Rice, Evans C.
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Sperry, J.S.; Holbrook, N.M.; Zimmermann, M.H.; Tyree, M.T. Spring filling of xylem vessels in wild grapevine. Plant Physiol. 1987, 83, 414–417. [Google Scholar] [CrossRef]
- Maurel, K.; Leite, G.B.; Bonhomme, M.; Guilliot, A.; Rageau, R.; Pétel, G.; Sakr, S. Trophic control of bud break in peach (Prunus persica) trees: a possible role of hexoses. Tree Physiol. 2004, 24, 579–588. [Google Scholar] [CrossRef]
- Bonhomme, M.; Peuch, M.; Ameglio, T.; Rageau, R.; Guilliot, A.; Decourteix, M.; et al. Carbohydrate uptake from xylem vessels and its distribution among stem tissues and buds in walnut (Juglans regia L.). Tree Physiol. 2010, 30, 89–102. [Google Scholar] [CrossRef]
- Keller, M.; Zhang, Y.; Shrestha, P.M.; Biondi, M.; Bondada, B.R. Sugar demand of ripening grape berries leads to recycling of surplus phloem water via the xylem. Plant, Cell Environ. 2015, 38, 1048–1059. [Google Scholar] [CrossRef]
- Rubio, S.; Noriega, X.; Pérez, F.J. ABA promotes starch synthesis and storage metabolism in dormant grapevine buds. J. Plant Physiol. 2019, 234–235, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Van Staden, J.; Davey, J.G. The synthesis, transport and metabolism of endogenous cytokinins. Plant, Cell and Environ. 1979, 2, 93–106. [Google Scholar] [CrossRef]
- Nikolaou, G.; Neocleous, D.; Christou, A.; Kitta, E.; Katsoulas, N. Implementing sustainable irrigation in water-scarce regions under the impact of climate change. Agronomy 2020, 10, 1120. [Google Scholar] [CrossRef]
- Pirie, A.; Mullins, M.G. Changes in anthocyanin and phenolics content of grapevine leaf and fruit tissues treated with sucrose, nitrate, and abscisic acid. Plant Physiol. 1976, 58, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Dokoozlian, N.K. Grape berry growth and development. In: Christensen LP (ed) Raisin production manual. Oakland, California, 2000, pp. 30–37.
- Jogaiah, S.; Striegler, K.R.; Bergmeier, E.; Harris, J. Influence of cluster exposure to sun on fruit composition of ‘Norton'Grapes (Vitis aestivalis Michx) in Missouri. Int. J. Fruit Sci. 2012, 12, 410–426. [Google Scholar] [CrossRef]
Figure 1.
Representative tomograms of the different periods, years and combinations of grafting. (a) 2021; (b) (2022).
Figure 1.
Representative tomograms of the different periods, years and combinations of grafting. (a) 2021; (b) (2022).
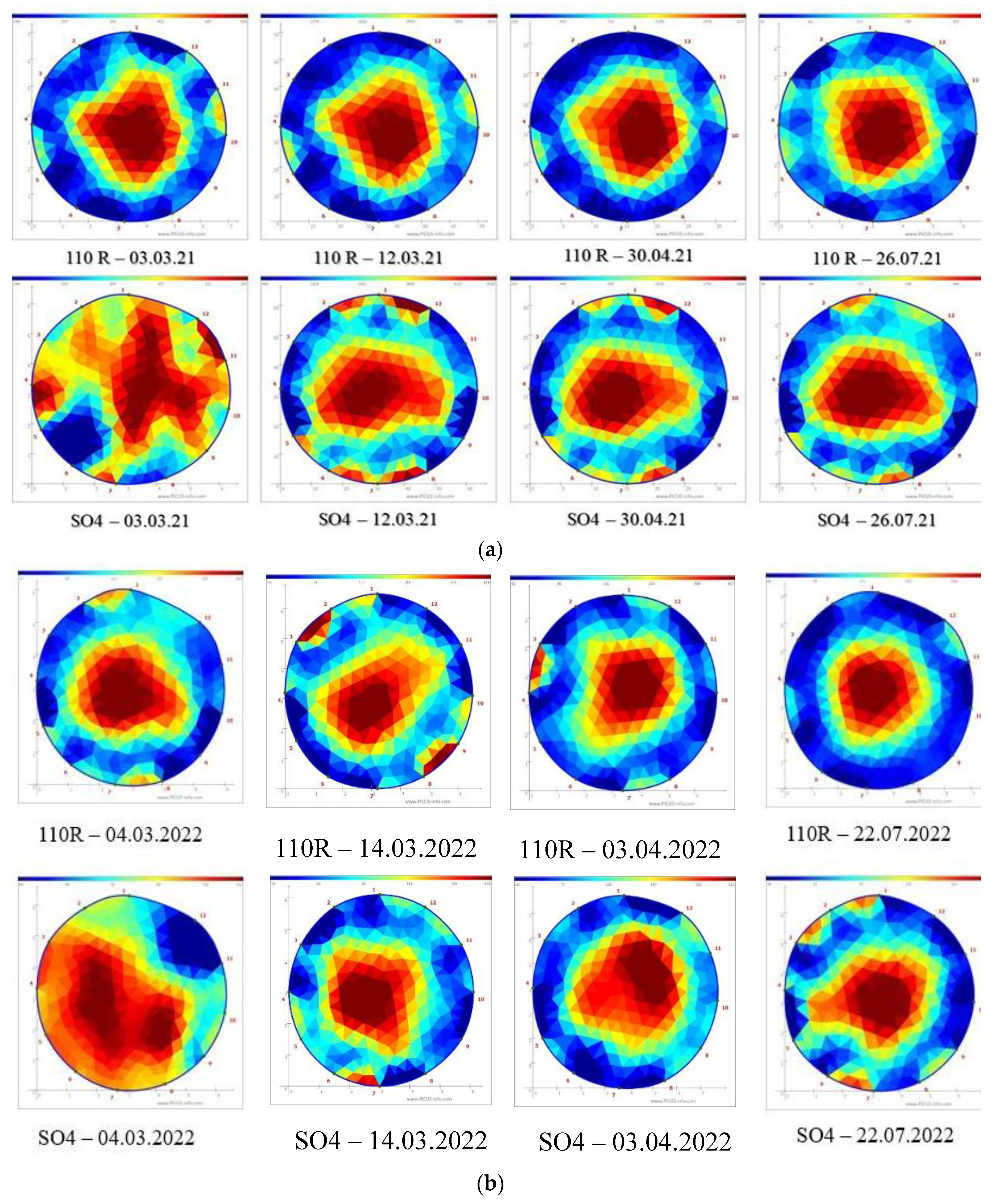
Figure 2.
Vegetative growth measured during 2021 (a) and 2022 (b) in both graft combinations.
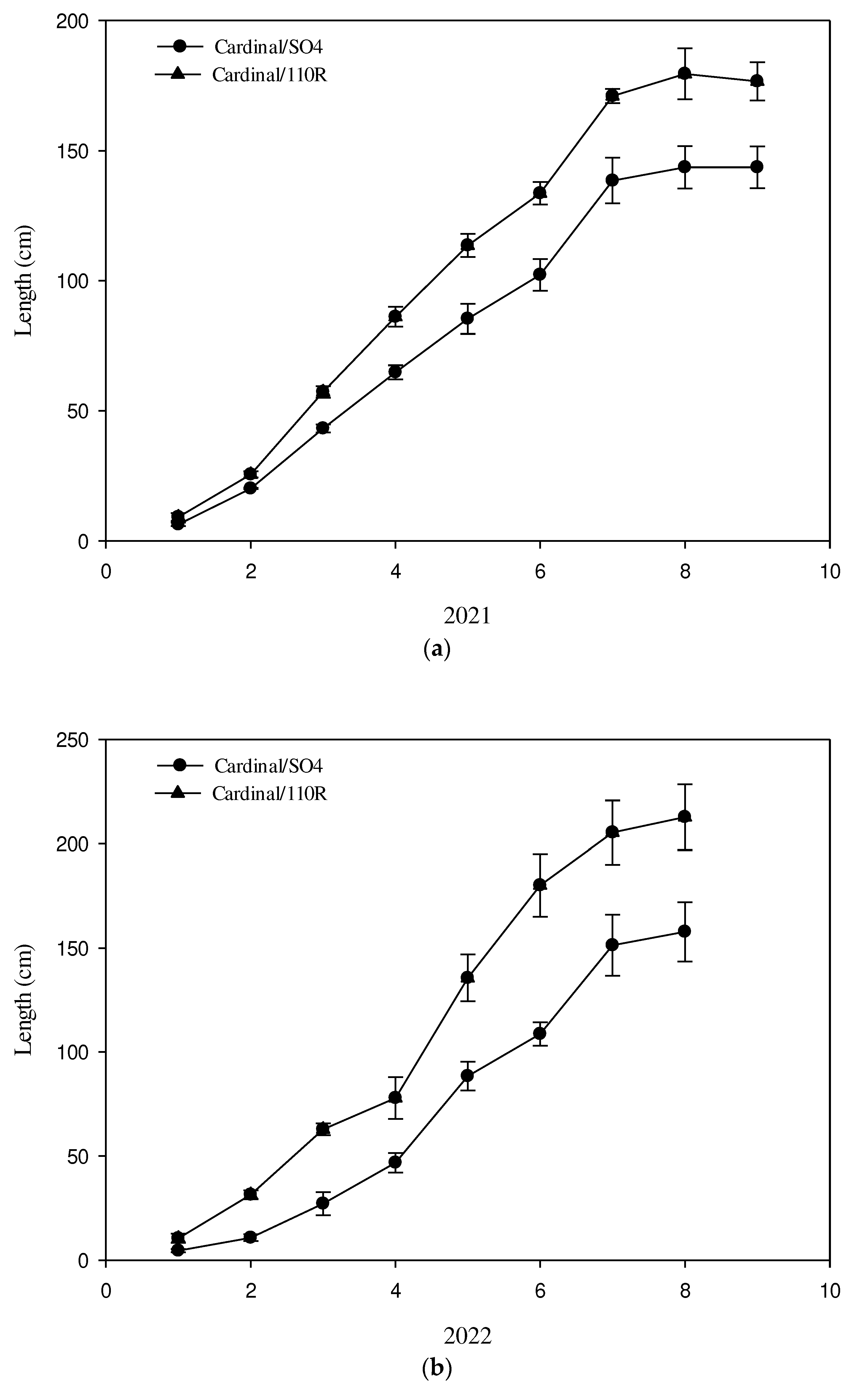
Figure 3.
Distribution, intracanopy and ground, of PAR, expressed in µmoles m2 s-1, in the two graft combinations.
Figure 3.
Distribution, intracanopy and ground, of PAR, expressed in µmoles m2 s-1, in the two graft combinations.

Figure 4.
Sap flow measurements in stem of Cardinal grapevine cultivar grafted onto two rootstocks 110 R (a) and SO4 (b) during five summer days.
Figure 4.
Sap flow measurements in stem of Cardinal grapevine cultivar grafted onto two rootstocks 110 R (a) and SO4 (b) during five summer days.
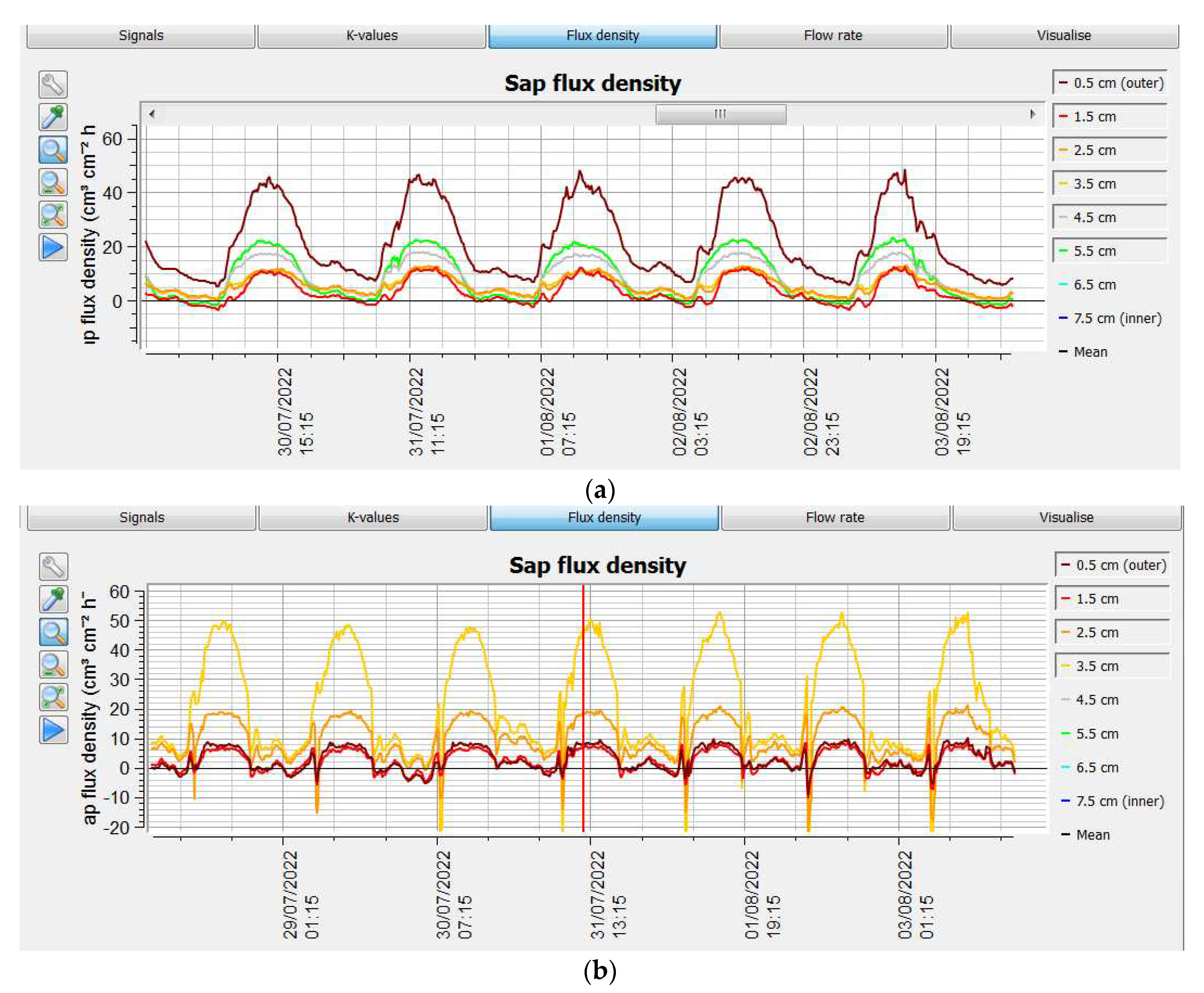

Table 1.
Evolution of conductive xylem areas (CXA) at different measurement times for the different graft combinations on two observation years.
Table 1.
Evolution of conductive xylem areas (CXA) at different measurement times for the different graft combinations on two observation years.
| Graft combination | CXA | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| First ten days March | Second ten days March | Third ten days April | Third ten days July | ||||||
| 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | ||
| C/110R | % | 66.47a (±1.71) |
60.73a (±0.44) |
68.61a (±2.29) |
61.22a (±0.47) |
67.72a (±1.05) |
65.33a (±1.56) |
65.03n.s. (±2.60) |
70.88 n.s. (±1.26) |
| C/SO4 | 49.39b (±2.33) |
44.12b (±1.32) |
53.91b (±2.17) |
56.15b (±0.82) |
61.56b (±2.80) |
62.33b (±2.80) |
62.97 (±1.91) |
68.54 (±0.66) |
|
Table 2.
Evolution of phenological stages (%) in grapevine plants of Cardinal cultivar grafted on two rootstocks: SO4 and 110 R (2021).
Table 2.
Evolution of phenological stages (%) in grapevine plants of Cardinal cultivar grafted on two rootstocks: SO4 and 110 R (2021).
| CARDINAL/SO4 | |||||||||||||
| Period | 00 | 05 | 09 | 10 | 12 | 53 | 55 | 57 | 65 | 71 | 75 | 77 | 81 |
| 03 Mar 2021 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 12 Mar 2021 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 19 Mar 2021 | 84,61 | 15,38 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 26 Mar 2021 | 64,1 | 33,33 | 0 | 2,564 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 02 Apr 2021 | 12,82 | 33,33 | 35,89 | 17,94 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 08 Apr 2021 | 5,12 | 15,38 | 23,07 | 48,71 | 7,69 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 Apr 2021 | 5,12 | 12,82 | 5,12 | 30,76 | 25,64 | 20,51 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 30 Apr 2021 | 5,12 | 10,25 | 2,56 | 5,12 | 12,82 | 5,12 | 58,97 | 0 | 0 | 0 | 0 | 0 | 0 |
| 10 May 2021 | 5,12 | 10,25 | 0 | 0 | 15,38 | 2,56 | 10,25 | 56,41 | 0 | 0 | 0 | 0 | 0 |
| 20 May 2021 | 5,12 | 10,25 | 0 | 0 | 15,38 | 2,56 | 2,56 | 64,1 | 0 | 0 | 0 | 0 | 0 |
| 28 May 2021 | 5,12 | 10,25 | 0 | 0 | 15,38 | 0 | 0 | 2,56 | 66,66 | 0 | 0 | 0 | 0 |
| 04 Jun 2021 | 5,12 | 10,25 | 0 | 0 | 12,82 | 0 | 0 | 0 | 0 | 71,79 | 0 | 0 | 0 |
| 21 Jun 2021 | 5,12 | 10,25 | 0 | 0 | 12,82 | 0 | 0 | 0 | 0 | 71,79 | 0 | 0 | 0 |
| 09 Jul 2021 | 5,12 | 10,25 | 0 | 0 | 12,82 | 0 | 0 | 0 | 0 | 0 | 71,79 | 0 | 0 |
| 22 Jul 2021 | 5,12 | 7,69 | 0 | 0 | 12,82 | 0 | 0 | 0 | 0 | 0 | 0 | 58,97 | 15,38 |
| CARDINAL/110R | |||||||||||||
| Period | 00 | 05 | 09 | 10 | 12 | 53 | 55 | 57 | 65 | 71 | 75 | 77 | 81 |
| 03 Mar 2021 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 12 Mar 2021 | 78,57 | 10,71 | 3,57 | 7,14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 19 Mar 2021 | 53,57 | 28,57 | 14,28 | 3,57 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 26 Mar 2021 | 17,85 | 39,28 | 25 | 17,85 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 02 Apr 2021 | 10,71 | 3,57 | 42,85 | 32,14 | 10,71 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 08 Apr 2021 | 10,71 | 0 | 3,57 | 28,57 | 50 | 7,14 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 14 Apr 2021 | 10,71 | 0 | 0 | 3,57 | 32,14 | 42,85 | 10,71 | 0 | 0 | 0 | 0 | 0 | 0 |
| 30 Apr 2021 | 10,71 | 0 | 0 | 0 | 10,71 | 0 | 78,57 | 0 | 0 | 0 | 0 | 0 | 0 |
| 10 May 2021 | 10,71 | 0 | 0 | 0 | 10,71 | 0 | 10,71 | 67,85 | 0 | 0 | 0 | 0 | 0 |
| 20 May 2021 | 10,71 | 0 | 0 | 0 | 10,71 | 0 | 0 | 78,57 | 0 | 0 | 0 | 0 | 0 |
| 28 May 2021 | 10,71 | 0 | 0 | 0 | 10,71 | 0 | 0 | 0 | 78,57 | 0 | 0 | 0 | 0 |
| 04 Jun 2021 | 10,71 | 0 | 0 | 0 | 10,71 | 0 | 0 | 0 | 78,57 | 0 | 0 | 0 | 0 |
| 21 Jun 2021 | 10,71 | 0 | 0 | 0 | 7,142 | 0 | 0 | 0 | 82,14 | 0 | 0 | 0 | 0 |
| 09 Jul 2021 | 10,71 | 0 | 0 | 0 | 7,142 | 0 | 0 | 0 | 0 | 0 | 82,14 | 0 | 0 |
Table 3.
Evolution of phenological stages (%) in grapevine plants of Cardinal cultivar grafted on two rootstocks: SO4 and 110 R (2022).
Table 3.
Evolution of phenological stages (%) in grapevine plants of Cardinal cultivar grafted on two rootstocks: SO4 and 110 R (2022).
| CARDINAL/110R | ||||||||||||
| Period | 00 | 05 | 09 | 10 | 12 | 53 | 55 | 57 | 65 | 71 | 75 | 77 |
| 04 Mar 2022 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 11 Mar 2022 | 98,85 | 1,15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 18 Mar 2022 | 64,37 | 32,18 | 3,45 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 25 Mar 2022 | 42,35 | 52,94 | 4,71 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 01 Apr 2022 | 30,59 | 12,94 | 35,29 | 18,82 | 2,35 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 08 Apr 2022 | 15,29 | 4,71 | 22,35 | 43,53 | 12,94 | 1,18 | 0 | 0 | 0 | 0 | 0 | 0 |
| 15 Apr 2022 | 10,59 | 0 | 4,71 | 20 | 29,41 | 35,29 | 0 | 0 | 0 | 0 | 0 | 0 |
| 22 Apr 2022 | 9,41 | 0 | 0 | 0 | 27,06 | 36,47 | 27,06 | 0 | 0 | 0 | 0 | 0 |
| 02 May 2022 | 9,52 | 0 | 0 | 0 | 3,57 | 20,24 | 30,95 | 35,71 | 0 | 0 | 0 | 0 |
| 13 May 2022 | 8,54 | 0 | 0 | 1,22 | 2,44 | 15,85 | 24,39 | 47,56 | 0 | 0 | 0 | 0 |
| 20 May 2022 | 8,54 | 0 | 0 | 0 | 2,44 | 17,07 | 19,51 | 52,44 | 0 | 0 | 0 | 0 |
| 03 Jun 2022 | 8,54 | 0 | 0 | 0 | 2,44 | 17,07 | 0 | 3,66 | 7,32 | 60,98 | 0 | 0 |
| 17 Jun 2022 | 8,64 | 0 | 0 | 0 | 0 | 16,05 | 0 | 0 | 0 | 2,47 | 54,32 | 18,52 |
| 01 Jul 2022 | 8,64 | 0 | 0 | 0 | 0 | 16,05 | 0 | 0 | 0 | 0 | 71,60 | 3,70 |
| 22 Jul 2022 | 8,64 | 0 | 0 | 0 | 0 | 16,05 | 0 | 0 | 0 | 0 | 2,47 | 72,84 |
| CARDINAL/SO4 | ||||||||||||
| Period | 00 | 05 | 09 | 10 | 12 | 53 | 55 | 57 | 65 | 71 | 75 | 77 |
| 04 Mar 2022 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 11 Mar 2022 | 96,67 | 3,33 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 18 Mar 2022 | 71,67 | 26,67 | 1,67 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 25 Mar 2022 | 70 | 26,67 | 1,67 | 1,67 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 01 Apr 2022 | 40 | 28,33 | 21,67 | 8,33 | 1,67 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 08 Apr 2022 | 18,33 | 10 | 28,33 | 36,67 | 5,00 | 1,67 | 0 | 0 | 0 | 0 | 0 | 0 |
| 15 Apr 2022 | 10 | 0 | 8,33 | 46,67 | 28,33 | 6,67 | 0 | 0 | 0 | 0 | 0 | 0 |
| 22 Apr 2022 | 10 | 0 | 0 | 1,67 | 31,67 | 48,33 | 8,33 | 0 | 0 | 0 | 0 | 0 |
| 02 May 2022 | 8,33 | 1,67 | 0 | 0 | 6,67 | 35,00 | 40 | 8,33 | 0 | 0 | 0 | 0 |
| 13 May 2022 | 8,33 | 0 | 0 | 1,67 | 0 | 30 | 31,67 | 26,67 | 0 | 1,67 | 0 | 0 |
| 20 May 2022 | 8,47 | 0 | 0 | 0 | 1,69 | 30,51 | 20,34 | 37,29 | 1,69 | 0 | 0 | 0 |
| 03 Jun 2022 | 8,62 | 0 | 0 | 0 | 0 | 29,31 | 1,72 | 0 | 5,17 | 55,17 | 0 | 0 |
| 17 Jun 2022 | 8,62 | 0 | 0 | 0 | 0 | 29,31 | 0 | 0 | 0 | 0 | 22,41 | 39,66 |
| 01 Jul 2022 | 8,62 | 0 | 0 | 0 | 0 | 31,03 | 0 | 0 | 0 | 0 | 53,45 | 6,90 |
| 22 Jul 2022 | 8,62 | 0 | 0 | 0 | 0 | 31,03 | 0 | 0 | 0 | 0 | 8,62 | 51,72 |
Table 4.
Vegetative parameters of shoot and plant of Cardinal cultivar grafted onto 110 R and SO4.
| Number of leaves per principal shoot (PS) n° |
Total Leaf area per Principal shoot m2 |
Number of leaves per Lateral Shoot n° | Leaf area per lateral shoot (cm2) | Total number of leaves per shoot (PS+LS) n° |
Total Leaf area per shoot (PS+LS) m2 |
Leaf area cm2 |
Leaf area per plant m2 |
Lateral shoot % |
LAI | Internode length (cm) |
Winter pruning residue (Kg pianta-1) |
|
| C/SO4 | 16.30±1.9d | 0.12±0.02d | 27.53b | 0.217267b | 43.83±10.83ns | 0.33±0.05ns | 78.92±6.47ns | 26.66.26±4.1b | 20.1±1.5a | 6.66±0.7b | 7.33±0.12b | 3.10±0.30a |
| C/110R | 19.18±0.4c | 0.15±0.015c | 17.48c | 0.137358d | 36.66±6.90 | 0.28±0.05 | 78.58±3.87 | 22.20±2.8c | 14.53±1.8b | 5.50±0.4c | 8.59±0.15a | 2.50±0.20b |
| C/SO4 | 21.61±2.2b | 0.21±0.01b | 29.73a | 0.293376a | 51.34±10.83 | 0.40±0.06 | 98.68±3.13ns | 36.26±4.1a | 19±1.1a | 9.26±0.7a | 7.47±0.14b | 3.80±0.40a |
| C/110R | 25.02±1.5a | 0.24±0.01a | 16.26d | 0.158194c | 41.28±6.90 | 0.32±0.04 | 97.29±2.17 | 26.22±4.8b | 15±1.2b | 6.59±0.4b | 8.48±0.18a | 2.30±0.30b |
| year | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| I=YxT | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
Different letters indicate significant differences (P ≤ 0.05); n.s.=non-significant. I= Interaction Treatment x Year.
Table 5.
Main physiological parameters in Cardinal cultivar grafted onto 110 R and SO4 rootstocks ( 2021 and 2022).
Table 5.
Main physiological parameters in Cardinal cultivar grafted onto 110 R and SO4 rootstocks ( 2021 and 2022).
| Year | Graft combination | Pn µmol CO2 m-2leaf area s-1 |
gs mol H2O m-2leaf area s-1 |
Ci ppm |
Tr | Pn/Tr | Pn/Ci | SPAD | ΨLeaf Bar |
Sap Flow cm3 cm-2 h-1 |
Daily Xylem Flow L cm-2 d-1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | C/SO4 | 12.21±0.87b | 0.073±0.004b | 118.1±17.80b | 3.102±0.322ns | 3.93±0.23ns | 0.10±0.05ns | 36.88±0.71ns | -5.62±0.15b | 25.74±1.12b | 10.60±0.4b |
| C/110R | 13.83±0.85a | 0.086±0.008a | 128.45±20.43b | 3.008±0.212 | 4.59±0.23 | 0.10±0.02 | 36.14±0.62 | -4.56±0.11a | 30.89±1.35a | 12.33±0.15a | |
| 2022 | C/SO4 | 10.79±1.13c | 0.072±0.01b | 157.21±33.40a | 3.296±0.588ns | 3.60±0.78ns | 0.10±0.09ns | 37.20±0.23 | -5.37±0.20b | 20.33±1.24c | 10.21±0.8b |
| C/110 R | 14.04±1.97a | 0.090±0.01a | 140.70±14.28a | 3.239±0.708 | 5.05±0.28 | 0.07±0.01 | 36.84±0.38 | -4.64±0.10a | 25.34±1.01b | 11.74±0.5a | |
| Year | ns | ns | * | ns | ns | ns | ns | ns | * | ns | |
| I = T x Y | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |
Table 6.
Yield per plant, number of clusters and berries, budding percentage and real fertility in vine plants of the Cardinal cultivar grafted onto 110 R and SO4 (2021 and 2022).
Table 6.
Yield per plant, number of clusters and berries, budding percentage and real fertility in vine plants of the Cardinal cultivar grafted onto 110 R and SO4 (2021 and 2022).
| Year | Graft combination | Number of clusters per plant | Yield per plant Kg. plant-1 |
Bunch weight g |
N° of berries per bunch | Bud-break % |
Real Fertility R.F. |
| 2021 | C/SO4 | 59.18±4.37a | 21.3±1.9a | 362±25 n.s. | 51±5 n.s. | 95.60±2.23n.s. | 0.77±0.12a |
| C/110R | 63.75±3.3a | 25.9±2.4a | 393±22 | 56±4 | 89.70±2.06 | 0.87±0.11a | |
| 2022 | C/SO4 | 48.00±4.19b | 18.44±1.5b | 326.05±18.64 | 54±2 | 92.38±3.15 | 0.66±0.06b |
| C/110R | 53.83±5.90b | 18.59±1.76b | 305.59±21.64 | 59±4 | 92.36±2.89 | 0.73±0.04b | |
| Year | * | * | * | n.s. | n.s. | n.s. | |
| I = T x Y | ns | ns | ns | ns | ns | ns | |
Different letters indicate significant differences (P ≤ 0.05); n.s.=non-significant. I= Interaction Treatment x Year.
Table 7.
Biometric parameters and degree of berry ripening in table grape Cardinal Cultivar grafted on two different rootstocks: 110R and SO4 (2021 and 2022).
Table 7.
Biometric parameters and degree of berry ripening in table grape Cardinal Cultivar grafted on two different rootstocks: 110R and SO4 (2021 and 2022).
| Time of sampling | Graft combination | Average weight g |
Polar diameter mm |
Equatorial diameter mm |
Degree of ripening % |
| First week July (12-July-2021 13-July-2022) |
110R | 5.16±0.21d | 19.68±0.25d | 20.64±0.30c | 0.76±0.14f |
| SO4 | 4.91±0.19d | 19.56±0.21d | 20.04±0.27c | 1.06±0.14e | |
| Second week July 19-July-2021 20-July-2022) |
110R | 5.77±0.26c | 20.17±0.27c | 20.74±0.33c | 1.68±0.14d |
| SO4 | 5.87±0.22c | 20.33±0.26c | 20.62±0.2c | 1.98±0.14c | |
| Third ten days July (26-July-2021 26-July-2022) |
110R | 7.24±0.27a | 22.24±0.25b | 22.54±0.29b | 2.26±0.10b |
| SO4 | 6.84±0.22b | 22.20±0.24b | 22.14±0.27b | 2.22±0.12b | |
| First ten day August (02-Aug-2021 3-Aug-2022) |
110R | 7.58±0.23a | 23.05±0.22a | 22.83±0.27a | 2.36±0.11a |
| SO4 | 7.40±0.24a | 23.84±0.23a | 23.34±0.28a | 2.50±0.12a | |
| Year | ns | ns | ns | * | |
| I = Year x Treatment | ns | ns | ns | ns | |
Different letters indicate significant differences (P ≤ 0.05); n.s.=non-significant. I= Interaction Treatment x Year.
Table 8.
Main ripening indices, in table grape plants of Cardinal cultivar grafted on two different rootstocks: 110R and SO4 (2021 and 2022).
Table 8.
Main ripening indices, in table grape plants of Cardinal cultivar grafted on two different rootstocks: 110R and SO4 (2021 and 2022).
| Time of sampling | Graft combination | TSS (°Brix) |
pH | TA (%) |
TSS/TA |
|---|---|---|---|---|---|
| First week July (12-JUL-2021 13 _Julyy 2022) |
110R | 10.10±0.31d | 2.94±0.03d | 1.21±0.05a | 8.43±0.68c |
| SO4 | 10.40±0.11d | 2.94±0.02d | 1.28±0.03a | 8.14±0.20c | |
| Second week July (19-JUL-2021 20 –July 2022) |
110R | 11.60±0.14c | 3.13±0.02c | 0.79±0.06b | 14.97±1.17d |
| SO4 | 11.74±0.19c | 3.10±0.02c | 0.85±0.03b | 13.91±0.60d | |
| Third ten days July (26-JUL-2021 26 July 2022) |
110R | 13.56±0.31b | 3.60±0.33a | 0.57±0.01d | 23.66±0.71b |
| SO4 | 13.42±0.18b | 3.28±0.02b | 0.670.02c | 19.18±0.75c | |
| First ten day August (02-AUG-2021 3-Aug-2022) |
110R | 14.48±0.30a | 3.50±0.02a | 0.46±0.01d | 31.68±0.27a |
| SO4 | 14.74±0.31a | 3.51±0.02a | 0.53±0.04d | 28.84±0.53a | |
| Year | ns | ns | ns | ns | |
| I = T x Y | ns | ns | ns | ns | |
Different letters indicate significant differences (P ≤ 0.05); n.s.=non-significant. I = Interaction Treatment x Year.
Table 9.
Main nutraceutical parameters in table grape plants of Cardinal cv grafted on two different rootstocks: 110R and SO4 (2021 and 2022).
Table 9.
Main nutraceutical parameters in table grape plants of Cardinal cv grafted on two different rootstocks: 110R and SO4 (2021 and 2022).
| Time of sampling | Graft combination | TPC pulp mg gallic acid/gr. FW |
TAC pulp µmoles trolox/gr. FW |
TPC peel mg gallic acid/gr. FW |
TAC peel µmoles trolox/gr. FW |
| First week July (12-July-2021 13-July-2022) |
C/110R | 0.569±0.049ns | 1.204±0.054ns | 0.994±0.086ns | 26.691±0.494a |
| C/SO4 | 0.650±0.085 | 1.137±0.207 | 1.311±0.172 | 23.841±0.041b | |
| Second week July 19-July-2021 20-July-2022) |
C/110R | 0.678±0.036ns | 1.114±0.093ns | 1.249±0.067ns | 25.859±0.121ns |
| C/SO4 | 0.615±0.038 | 1.589±0.150 | 1.172±0.073 | 25.680±0.013 | |
| Third ten days July (26-July-2021 26-July-2022) |
C/110R | 0.590±0.039ns | 1.146±0.068ns | 1.427±0.095ns | 31.337±0.887a |
| C/SO4 | 0.659±0.040 | 1.469±0.138 | 1.239±0.075 | 25.703±0.529b | |
| First ten day August (02-Aug-2021 3-Aug-2022) |
C/110R | 0.592±0.048ns | 1.074±0.052ns | 1.232±0.099ns | 28.244±0.450ns |
| C/SO4 | 0.524±0.010 | 1.066±0.062 | 1.072±0.022 | 27.407±0.353 | |
| Year | ns | ns | ns | ns | |
| I = T x Y | ns | ns | ns | ns | |
Different letters indicate significant differences (P ≤ 0.05); n.s. = non-significant. I = (Interaction Treatment x Year).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



