
Submitted:
02 October 2023
Posted:
03 October 2023
You are already at the latest version
Abstract
CXCL14 is one of the most evolutionarily conserved members of the chemokine family and is constitutionally expressed in multiple organs, suggesting that it is involved in the homeostasis maintenance of the system. CXCL14 is highly expressed in colon epithelial cells and shows obvious gene silencing in clinical colon cancer samples, suggesting that its silencing is related to the immune escape of cancer cells. In this paper, we analyzed the expression profiles of multiple human clinical colon cancer datasets and mouse colon cancer models to reveal the variation trend of CXCL14 expression during colitis, colon polyps, primary colon cancer, and liver metastases. The relationship between CXCL14 gene silencing and promoter hypermethylation was revealed through the colorectal carcinoma methylation database. The results suggest that CXCL14 is a tumor suppressor gene in colorectal carcinoma, which is activated first and then silenced during the process of tumor occurrence and deterioration. Promoter hypermethylation is the main cause of CXCL14 silencing. The methylation level of CXCL14 is correlated with the anatomic site of tumor occurrence and positively correlated with patient age, and associated with prognosis. Reversing the hypermethylation of CXCL14 may be an epigenetic therapy for colon cancer.
Keywords:
colorectal cancer
; CXCL14
; gene silence
; metastasis
; DNA methylation
Introduction
Chemokines are cytokines that appeared with the emergence of chordates. There are 41 ligands and 21 receptor genes in the human genome[1]. The chemotactic action of chemokines induces the effector cells to move against the gradient through the concentration gradient formed by the local tissue expression and secretion. This system plays an important role in embryonic development and immune cell homing. In addition to initial chemotaxis, chemokines are also involved in metabolic regulation[2], angiogenesis[3], and tissue development [4].
CXCL14 is one of the earliest members appeared in the chemokine family [5]. Compared with other members, it evolves extremely slowly and nonsense SNPs are rare, indicating its significant function. In healthy people, CXCL14 is usually highly expressed in tissues with a rapid renewal rate, such as epithelial cells of small intestine, colorectal, gastric mucosa, endometrium, mammary gland, skin, and other tissues. and it is low in resting state cells, such as the liver, muscle, connective tissue, brain, and other parenchymal cells. CXCL14 has shown abnormal expression levels in some immune diseases and a large number of tumor samples [6-8]. CXCL14 gene knockout has effects on the survival rate and metabolism of sugar and lipids in mouse embryos [9].
Contrary to its important function, the receptor for CXCL14 has not yet been discovered. Ariadni Kouzeli et al [10] systematically studied the interaction between CXCL14 and all genes of the chemokine receptor family and found that CXCL14 could not activate anyone. However, it is interesting that CXCL14 can synergistically promote the physiological activity of several other chemokines. Philipp von Hundelshausen et al. [11] systematically studied the pattern of heterodimers formed by chemokines and found that a large number of heterodimers existed in the chemokine family. It is suggested that CXCL14 functions by promoting or inhibiting the activity of other chemokines. This may explain why the effects of CXCL14 may be opposite in different tissues and tumor types [12], because the chemokines that partner with CXCL14 in different tissues can vary greatly in both type and concentration. CXCL14 appears as a tumor suppressor in colon cancer.
Tumor development is the result of activation of oncogenes and/or inactivation of tumor suppressor genes. It was previously thought that these genetic variations were all due to genetic mutations [13]. However, it is now known that epigenetic variation is another pathway that leads to the silencing of tumor suppressor genes, and abnormal expression of genes caused by abnormal DNA methylation is more frequent than changes caused by gene copy number [14]. We investigated the methylation level variation and gene mutation frequency of colon cancer tumors and found that the CXCL14 promoter was particularly hypermethylated in colon cancer samples.
The relationship between CXCL14 hypermethylation and gene silencing was also found in some tumor types. Tessema et al. found that CXCL14 was epigenetically silenced in lung cancer and considered this gene to be an important epigenetic therapeutic site [7]. Cao et al. observed that the epigenetic silencing of CXCL14 in colon cancer is a mechanism leading to tumor metastasis and invasion [15]. Hu et al. also found that abnormal methylation of promoters in gastric tumors resulted in inhibition of CXCL14 expression [16].
To further reveal the changes of CXCL14 in colon cancer, this paper studied the changes of CXCL14 expression from normal tissue to colitis, to the primary tumor, and eventually to metastatic cancer by mining multiple datasets. We explored the changes of CXCL14 methylation in colon cancer and its correlation with CXCL14 gene silencing, prognosis, and important clinical markers, and revealed the silencing mechanism of CXCL14 and its important significance in the development of colon cancer.
Methods
mRNA Expression Data
The four groups of human colon cancer expression profiles analyzed in this study are as follows. (1) Memorial Sloan-Kettering Cancer Center, a total of 390 microarray data (E-GEOD-41258), Includes primary colon adenocarcinomas, adenomas, metastasis, and corresponding normal mucosae[17]. (2) Different stage colorectal carcinomas (CRC) and inflammatory bowel diseases (IBD), (E-GEOD-4183), including biopsies of 15 patients with CRC, 15 with adenoma, 15 with IBD, and 8 healthy normal controls[18]. (3) 36 CRC tissues and 24 non-cancerous colorectal tissue. (E-GEOD-23878). (4)Pairs of primary tumors, hepatic metastases, and normal controls [19].
Two mouse primary colorectal cancer microarray databet was analyzed. They are about AOM/DSS induced colitis-associated cancer (CAC) model[20,21] and ApcMin/+/J transgenic mice-based spontaneous tumor model[21].
DNA Methylation Data
We used the mexpress database (http://mexpress.be) to study the methylation status of the CXCL14 gene in the TCGA-COAD dataset [22]. The methylation of sites of probe,6, 7, and 8, that was significantly hypermethylated in clinical colorectal carcinoma, were used to analyze associations with clinical indexes in Table 2.
Survival Analysis
Cases were separated into groups of higher or lower CXCL14 mRNA expression by the best expression cut-off of the default setting of the Human Protein Atlas (https://www.proteinatlas.org/) and Gepia2 (http://gepia2.cancer-pku.cn) platform. For survival analysis of CXCL14 methylation, the cases were split into groups of hypermethylation and hypomethylation by the maxstat algorithm. Overall survival curves were achieved using the Kaplan-Meier assessment and the log-rank test.
Cell Culture
293T, HCT15 and HT29 cells were cultured in RPMI1640 medium containing 10% FBS, and HCT116 cells were cultured in McCoy’s 5A containing 10% FBS. The cell culture conditions were 37℃, 5% carbon dioxide, and 70% humidity. Decitabine (DAC) was purchased from MCE (HY-A0004, shanghai, China) and formulated with DMSO for 10mM storage. When DAC is treated with cells, DAC is diluted to the desired concentration in the medium and added to the cell culture dish.
Real-Time Quantitative PCR
Cell total RNA was extracted using the TaKaRa MiniBEST Universal RNA Extraction Kit (Cat 9767, TAKARA, Japan). Reverse transcription was performed using HiScript II 1st Strand cDNA Synthesis Kit (Cat R212-01, Vazyme, China). Quantitative PCR was performed using the TAKARA SybrGreen kit (RR820Q, TAKARA, Japan). The quantitative PCR primer for human CXCL14 was 5’-AAGCCAAAGTACCCGCACTG (Forward), 5’-GACCTCGGTACCTGGACACG (Reverse), the reference gene β-actin primers are: 5’-GTGAAGGTGACAGCAGTCGGTT(Forward), 5’-GAAGTGGGGTGGCTTTTAGGAT(Reverse). The relative expression of target genes was calculated by the ΔΔCt method.
Promoter Assay
Plasmid pGL4.11 [Luc2p] was used as vector for promoter assay, in which truncated CXCL14 promoter fragments were cloned between KpnI and HindIII sites (Figure 6D). When studying the effect of methylation on promoters, CpG methyltransferase M. SSSI (EM0821, Thermo Scientific, Shanghai, China) was used to methylate PCR products, and PCR recovery kit was used to recover reaction products. The methylated fragments were then ligated into the vector using Gibson Assembly cloning kit (C115, Vezyme, Nanjing, China), and the product concentration was measured by spectrophotometry. The 293T cells in the 24-well plate were transfected with 0.9 ug promoter plasmid and 0.1 ug pEGFP-N1 plasmid using Lipofectamin2000. After 48 hours, the cells were digested with trypsin and loaded into a white wall transparent bottom 96-well plate. The green fluorescence (EGFP) was measured with a Tecan M200 pro microplate reader, and then the chemiluminescence intensity was measured immediately after adding 150 μg/mL D-luciferin. The Luc/EGFP value is used as the promoter activity.
Statistical Analysis
SPSS software was used for statistical analysis in this study. The student T-test is used to assess the significance between two sets of data. Paired T-tests are used to test the significance of differences between paired data from the same individual. Chi-square tests were used to examine the association between CXCL14 methylation and different clinical indexes. Spearman’s correlation approach was examined in methylation β-value and patient ages. *: p<0.05, **: p<0.01, ***: p<0.001.
Result
CXCL14 Silences as the Colorectal Cancer Progresses
We mined four groups of human colon cancer expression profile data sets and found that CXCL14 expression levels had obvious changes in different stages of colon cancer development. Figure 1A is RNA microarray data from 259 colon cancer samples and 12 human cell lines collected at Memorial Sloan-Kettering Cancer Center between 1992 and 2004[18]. From the data, we can clearly see the expression changes of CXCL14 in different cancer stages. Firstly, CXCL14 expression levels were normally distributed in healthy tissues, but after the occurrence of intestinal polyps, the expression level dispersion increased, indicating a slight dysregulation in the expression level. After further development into primary tumors, the dispersion of expression levels increased further and the median decreased, indicating that the dysregulation of the expression level was intensified, and the selection of cancer cells in vivo was progressing in the direction of CXCL14 silencing. The phenomenon of gene silencing became more significant after liver metastasis or lung metastasis, indicating that CXCL14 silencing has certain advantages for tumor metastasis or tumor formation after metastasis. CXCL14 silencing occurred completely in 12 tumor cell lines.
In the dataset, E-GEOD-4183 (Figure 1B), Orsolya Galamb, et al. collected 15 patients with CRC, 15 with adenoma, 15 with IBD, and 8 healthy normal controls[18]. It was also found that in inflammatory bowel disease, although the mean expression of CXCL14 did not change significantly, the dispersion increased, and the CV% increased from 24% in healthy tissues to 83% in colitis tissues, indicating dysregulation of CXCL14 expression. CXCL14 levels decreased significantly with adenoma and colorectal cancer (CRC) (p<0.001).
In another set of expression profile data of colon cancer and adjacent tissues (Figure 1C), it was also found that the expression of CXCL14 in colon cancer tissues decreased and was accompanied by an increase in dispersion.
In the last set of data, which was about a clinical follow-up investigation, Pierre Martineau et al. tracked and collected primary cancer tissue, metastatic tissue and healthy colon tissues from 19 tumor patients, and performed RNA microarray analysis[17]. We analyzed the expression of CXCL14 and found that CXCL14 expression continued to decline during tumor progression (Figure 1D-G).
From the above four sets of expression profile data, it can be seen that CXCL14 silencing becomes more and more significant with the increase of colon cancer malignancy.
After that, we investigated the expression profiles of two independent mouse colon cancer models. In the first experiment (Figure 1H), two modeling methods were used to establish the primary model of colon cancer in mice [20]. In the first, the CAC model was induced by AOM/DSS, and the second was spontaneous colon cancer modeling by C57BL/6-ApcMinC/Nju transgenic mice (SporCRC). The expression profiles of cancer tissues and adjacent tissues in the CAC group and SporCRC group were detected respectively, and it was found that the CXCL14 expression level in tumor tissues was higher than that in corresponding healthy control tissues. In the second model (Figure 1I) [21], the authors used a single AOM/DSS model, and it was observed that the expression level of CXCL14 increased.
Therefore, the expression changes of CXCL14 in human colon cancer and in mouse colon cancer development seem to be opposite, whether they are contradictory, we will discuss in the following.
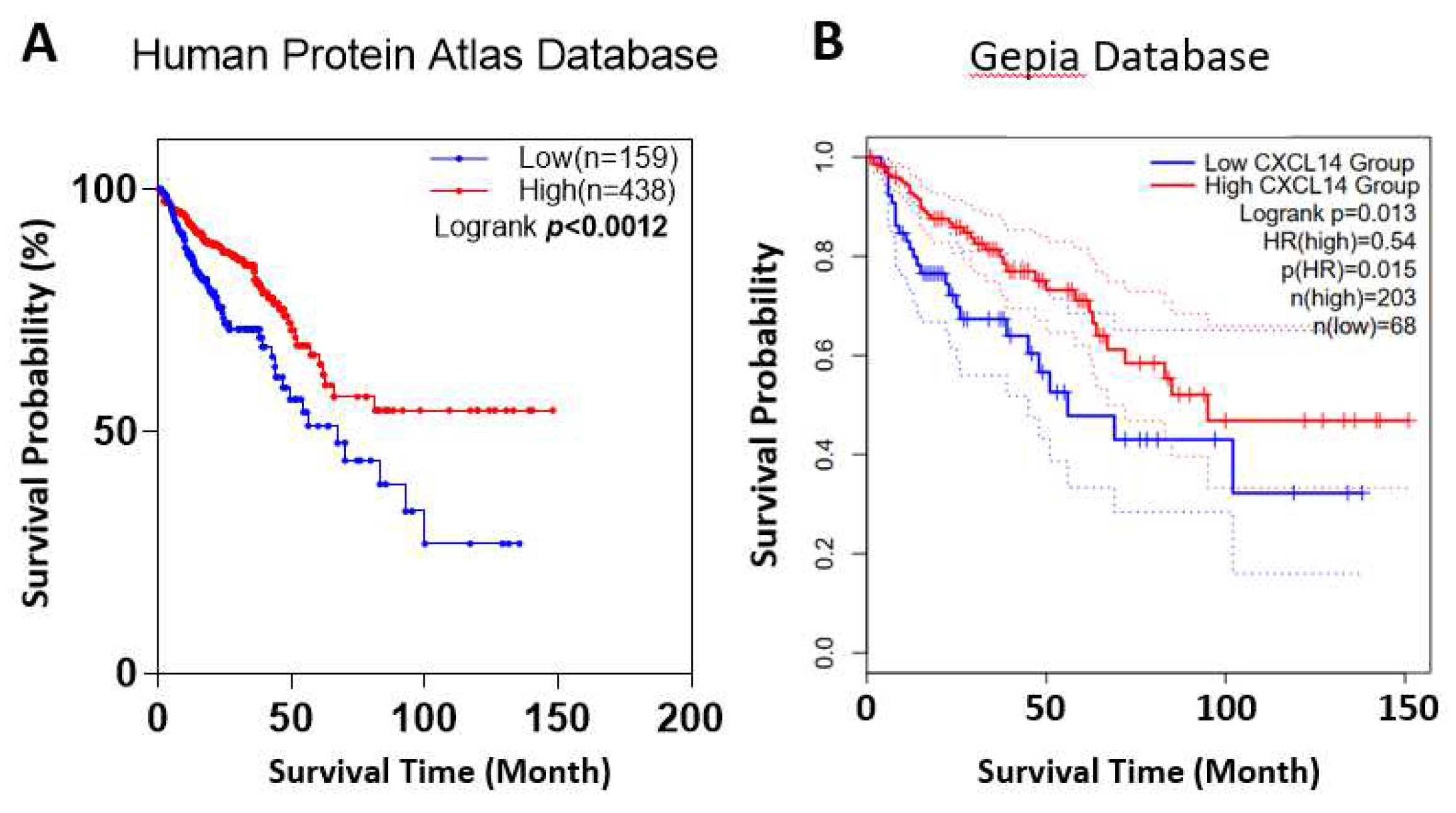
Clinical data show that patients with higher CXCL14 expression have a better prognosis. Two colon cancer datasets from two databases, Protein Atlas and Gepia, respectively, showed that higher CXCL14 expression was associated with longer survival.
CXCL14 Silencing is Associated with the Promoter CpG Methylation
CXCL14 rarely has gene mutations in colon cancer. We analyzed the colorectal adenocarcinoma dataset (TCGA, PanCancer Atlas) and found that in clinical samples of colon cancer, only 0.6% of total 526 samples had genetic variation, in which 1 sample had gene deletion and 2 samples had nonsense mutations.
We then analyzed the colon cancer methylation dataset[22] and found that CXCL14 methylation in colon cancer tissues showed significant hypermethylation across the entire promoter region as well as the 5′ untranslated region. Especially in the range -103~90 (probe 6, 7, 8), the hypermethylation was extremely significant (Figure 3A). A comparison of cancer/paracancer tissue pairs showed that methylation levels in cancer tissue were significantly higher in more than 75% of samples than in paracancer control tissue (more than three-fold increase, Figure 3C-E). At probe 6, 7, and 8, samples were divided into hypermethylation and hypomethylation group, the student t-test result denoted that the mRNA level in the two groups is significantly different (Figure 3F~G). In the following analysis, the average β-value of probes 6, 7, and 8 in each sample was used.
Figure 2.
Survival curves of colon cancer with different CXCL14 expression levels.fi(A)Kaplan-Meier survival curve of TCGA database Colon Adenocarcinoma (TCGA-COAD). (B) survival curve of the Gepia COAD dataset.
Figure 2.
Survival curves of colon cancer with different CXCL14 expression levels.fi(A)Kaplan-Meier survival curve of TCGA database Colon Adenocarcinoma (TCGA-COAD). (B) survival curve of the Gepia COAD dataset.
The Methylation of CXCL14 in Colon Cancer is Closely Related to Anatomical Site and Patient Age
We further analyzed the association between CXCL14 hypermethylation and clinical parameters (Table 1). Based on the upper limit of the methylation level of CXCL14 in healthy tissues 0.1, cancer tissues were divided into CXCL14 unmethylated group (β-value<0.1) and the hypermethylated group (β-value>0.1). A comparison between the non-methylated group and the hypermethylated group showed that CXCL14 hypermethylation was associated with the anatomical site (p<10-6), age at diagnosis (p<0.0001), and clinical stage of the tumor (p=0.019). Comparing the tumor cells from different colonic sites, it was found that the methylation level of the tumor cells originating from the anterior segment of the colon (cecum, ascending colon, hepatic flexure, and transverse colon) was significantly higher than that of the control paracancer tissues. However, the methylation of tumor cells from the posterior segment of the colon (descending colon, sigmoid colon) was significantly lower than that of the first half. Accordingly, CXCL14 expression decreased significantly in the anterior segment of the colon compared to the posterior segment (Figure 4B).
CXCL14 Methylation and Senescence
By correlation analysis, it was found that CXCL14 methylation increased slowly with age in normal colon tissue, and the correlation was very strong (r=0.58, p<0.0001). However, the methylation trend of CXCL14 in tumor samples showed a distinct differentiation. We used the upper limit of CXCL14 methylation level in normal colon samples, β-value=0.1, as the boundary to divide tumor samples into hypermethylated and hypomethylated groups. The hypermethylated group accounted for 72% of the total samples. Although elevated methylation was still linearly correlated with age in this sample, the degree of correlation was significantly reduced (r=0.194, p=0.0047). On the contrary, in the hypomethylated group (28% of total tumor samples), the methylation accumulation process tended to stall and did not increase with age, which was a different mode from both normal tissue and hypermethylated tumor samples.
CXCL14 Methylation was Associated with Survival
We analyzed the correlation between methylation levels and survival at different probe sites and found that higher methylation level at probe 3, 9, 11, and 12 was associated with worse prognosis, while high methylation levels at probe 6,7, and 8, in contrast, associated with longer survival (Figure 5D-E). Since we had observed abnormally low methylation in some cases in Figure 4D, we excluded these cases and reanalyzed the survival time, and found that high methylation level was associated with poor prognosis (Figure 5I~J), although the correlation was not significant.
The Methylation and Demethylation of CXCL14 Promoter Directly Regulate CXCL14 Expression
We treated human colon cancer cells with DAC and found that CXCL14 expression was positively correlated with DAC dose and treatment time (Figure 6A-C). We cloned the sequence of the human CXCL14 promoter from upstream -1000bp to downstream 100bp from the transcription start site (TSS), and inserted the upstream of pGL4.11 plasmid Luc2p gene. Then, through promoter truncation, key transcription factor binding sites were found, located in the -170bp~100bp region (Figure 6D). After the -170~100 region was methylated with M. SSSI enzyme, the promoter activity was significantly inhibited, indicating that the methylation of this region has an inhibitory effect on CXCL14 promoter activity. (Figure 6E).
Discussion
In this study, we found significant differences in the regulation of CXCL14 expression between human colon cancer tumors and mouse colon cancer models. CXCL14 is generally silenced in clinical tumor tissues, while it is upregulated in primary mouse tumors. Whether this contradiction is based on the inherent differences between humans and mice or some other reason deserves further discussion. First, we know that human tumor tissues progress for years to decades before being discovered, so the clinical tumor samples we observe are the result of many mutations and clonal differentiation, and at the same time, many years of selection in vivo. While, the mouse tumor samples were all primary tumors that developed within a few months, with limited time for mutation and selection. So, there’s a difference in the stage between the human and mouse tumors. Secondly, it should be noted that although the expression of CXCL14 decreased with the increase of cancer cell malignancy in a general trend, the expression of CXCL14 increased in some tumors (Figure 1A~D).
Taking the above two points together, we propose that the up-regulation of CXCL14 in the primary tumor of mice is not inconsistent with its silencing in clinical samples, but with its expression at different stages of tumor development. The mouse tumor model revealed the gene regulation pattern of CXCL14 in the early stage of tumor formation. Because of some currently unknown mechanisms, CXCL14 is activated during tumor formation, possibly by recruiting immune cells to participate in the tumor immune response. The clinical tumor samples reflect the CXCL14 status in the middle and late stages of tumor development. It is the result of genetic and/or epigenetic variation and has undergone long-term evolution and subclonal selection. However, this hypothesis is difficult to confirm in clinical samples, because early latent tumors are difficult to obtain clinically, so it is difficult to prove the hypothesis that CXCL14 is activated and then silenced during tumorigenesis. This hypothesis may be proved by long-term systematic animal tumor research.
The silencing of CXCL14 in colon cancer is mainly due to promoter hypermethylation. Since the database shows that the mutation rate of CXCL14 gene is very low in various tumor samples (not shown), the gene mutation is not the cause of CXCL14 silencing. Why in the tumor genome, some genes are more prone to change their expression levels through gene mutations, such as the well-known p53, while others, such as CXCL14, are more prone to epigenetic silencing? We believe that compared with simple deletion of genes, gene missense mutation can produce new phenotypes, one of the important aspects of which is to promote tumor metastasis. For example, some p53 mutants not only lose the cancer-suppressing function of the wild-type gene but also interfere with other signaling pathways, thus obtains survival advantages through the gain-of-function mechanism [23]. With chemokines like CXCL14, tumor cells could gain a survival advantage by simply reducing immune surveillance through gene silencing. Because epigenetic modification is less stable than genetic information, in the process of tumor progression, epigenetic silencing becomes the fate of the genes that are more sensitive to epigenetic regulation in tumor cells.
CXCL14 methylation levels were correlated with tumor anatomical location. Firstly, we report that the level of CXCL14 promoter methylation in colon cancer is correlated with the anatomical site of the primary tumor. The data showed that tumors from the anterior segment of the colon had a greater methylation increase. Although the methylation level from the posterior segment of colon cancer is still elevated compared with normal paracancer tissue, the extent of the methylation level increase is significantly decreased compared with the tumor from the anterior segment. This result is interesting, but we can’t yet explain why this phenomenon occurs.
We also found a relationship between the degree of CXCL14 methylation and age. In healthy colon tissue, CXCL14 methylation was significantly positively correlated with age (r=0.58), the regression line almost passes through the origin of coordinates. This fits well with the epigenetic clock model [24]. For tumor tissue, the samples can be divided into two subgroups, one group showed hypermethylation, while the other one showed hypomethylation. Hypermethylated groups exhibit faster methylation accumulation than healthy tissues, which is consistent with what has been reported in the literature [25]. However in the hypomethylated group, the epigenetic clock stopped completely, and methylation levels did not accumulate with age. Whether this group of tumors shows a reversal of epigenetic aging, and which kind of tumors have better prognosis compared with those that accelerate epigenetic aging, is a question worthy of attention.
We investigated the correlation between methylation level and survival, and found that methylation level was significantly negatively correlated with overall survival at probe positions 3-5, 9-12, but was opposite at probe positions 6, 7, and 8 (Figure 5C-E). To explain this contradiction, we specifically excluded the abnormal hypomethylated samples at 6, 7, and 8 probe locations and re-performed correlation analysis. We found that after such treatment, methylation levels at these three probe locations were negatively correlated with survival, which was consistent with the trend in the other locations. Therefore, we believe that the hypomethylated cases at the 6,7,8 probe location show different characteristics from the hypermethylated cases, and they have a worse prognosis than the CXCL14 promoter hypermethylated cases, and the mechanism needs to be further studied.
In summary, this paper describes the dynamic profile of CXCL14 expression with tumor development from the time axis through bioinformatics mining. The results revealed a dynamic process in which CXCL14 expression is up-regulated in early cancerous tissues and goes to silencing in metastatic tumors, suggesting that colon cancer cells gain proliferation and metastasis advantages through CXCL14 silencing that caused by its promoter methylation. Survival analysis confirmed that the gene silencing of CXCL14 in clinical colon cancer samples was associated with poor prognosis and revealed its anti-colon cancer function. In vitro experiments have shown that CXCL14 promoter methylation is an important cause of gene silencing. However, the mechanism of CXCL14 silencing promoting the progression of colon cancer is still unclear, which may be related to the immune surveillance of CXCL14[3,26-28] or/and antiangiogenic function [29].
Funding& Acknowledgements
B. Ma thanks support from Natural Science Foundation of China (Grant No. 32171246) and Shanghai Municipal Government Science Innovation grant 21JC1403700. Y. Wang thanks support from the grants from the National Natural Science Foundation of China (No.32200531), the Joint Research Funds for Medical and Engineering and Scientific Research at Shanghai Jiao Tong University (YG2022QN114 and YG2022QN082), and Startup Fund for Young Faculty at SJTU (SFYF at SJTU).
References
- Shields, D.C. Molecular evolution of CXC chemokines and receptors. Trends Immunol 2003, 24, 355. [Google Scholar] [CrossRef] [PubMed]
- Nara, N.; Nakayama, Y.; Okamoto, S.; Tamura, H.; Kiyono, M.; Muraoka, M.; Tanaka, K.; Taya, C.; Shitara, H.; Ishii, R.; et al. Disruption of CXC motif chemokine ligand-14 in mice ameliorates obesity-induced insulin resistance. J Biol Chem 2007, 282, 30794–30803. [Google Scholar] [CrossRef]
- Shellenberger, T.D.; Wang, M.; Gujrati, M.; Jayakumar, A.; Strieter, R.M.; Burdick, M.D.; Ioannides, C.G.; Efferson, C.L.; El-Naggar, A.K.; Roberts, D.; et al. BRAK/CXCL14 is a potent inhibitor of angiogenesis and a chemotactic factor for immature dendritic cells. Cancer Res 2004, 64, 8262–8270. [Google Scholar] [CrossRef]
- Kuang, H.; Chen, Q.; Zhang, Y.; Zhang, L.; Peng, H.; Ning, L.; Cao, Y.; Duan, E. The cytokine gene CXCL14 restricts human trophoblast cell invasion by suppressing gelatinase activity. Endocrinology 2009, 150, 5596–5605. [Google Scholar] [CrossRef] [PubMed]
- Huising, M.O.; Stet, R.J.; Kruiswijk, C.P.; Savelkoul, H.F.; Lidy Verburg-van Kemenade, B.M. Molecular evolution of CXC chemokines: extant CXC chemokines originate from the CNS. Trends Immunol 2003, 24, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Guo, L.; Tian, J.; He, H.; Marinova, E.; Zhang, P.; Zheng, B.; Han, S. Overexpression of CXC chemokine ligand 14 exacerbates collagen-induced arthritis. J Immunol 2010, 184, 4455–4459. [Google Scholar] [CrossRef]
- Tessema, M.; Klinge, D.M.; Yingling, C.M.; Do, K.; Van Neste, L.; Belinsky, S.A. Re-expression of CXCL14, a common target for epigenetic silencing in lung cancer, induces tumor necrosis. Oncogene 2010, 29, 5159–5170. [Google Scholar] [CrossRef]
- Song, E.Y.; Shurin, M.R.; Tourkova, I.L.; Gutkin, D.W.; Shurin, G.V. Epigenetic mechanisms of promigratory chemokine CXCL14 regulation in human prostate cancer cells. Cancer Res 2010, 70, 4394–4401. [Google Scholar] [CrossRef]
- Hara, T.; Nakayama, Y. CXCL14 and insulin action. Vitam Horm 2009, 80, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Kouzeli, A.; Collins, P.J.; Metzemaekers, M.; Meyrath, M.; Szpakowska, M.; Artinger, M.; Struyf, S.; Proost, P.; Chevigne, A.; Legler, D.F.; et al. CXCL14 Preferentially Synergizes With Homeostatic Chemokine Receptor Systems. Frontiers in immunology 2020, 11, 561404. [Google Scholar] [CrossRef]
- von Hundelshausen, P.; Agten, S.M.; Eckardt, V.; Blanchet, X.; Schmitt, M.M.; Ippel, H.; Neideck, C.; Bidzhekov, K.; Leberzammer, J.; Wichapong, K.; et al. Chemokine interactome mapping enables tailored intervention in acute and chronic inflammation. Sci Transl Med 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Tanegashima, K. Pleiotropic functions of the CXC-type chemokine CXCL14 in mammals. J Biochem 2012, 151, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Kinzler, K.W. Cancer genes and the pathways they control. Nat Med 2004, 10, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Zardo, G.; Tiirikainen, M.I.; Hong, C.; Misra, A.; Feuerstein, B.G.; Volik, S.; Collins, C.C.; Lamborn, K.R.; Bollen, A.; Pinkel, D.; et al. Integrated genomic and epigenomic analyses pinpoint biallelic gene inactivation in tumors. Nat Genet 2002, 32, 453–458. [Google Scholar] [CrossRef]
- Cao, B.; Yang, Y.; Pan, Y.; Jia, Y.; Brock, M.V.; Herman, J.G.; Guo, M. Epigenetic silencing of CXCL14 induced colorectal cancer migration and invasion. Discov Med 2013, 16, 137–147. [Google Scholar]
- Hu, C.; Lin, F.; Zhu, G.; Xue, X.; Ding, Y.; Zhao, Z.; Zhang, L.; Shen, X. Abnormal hypermethylation of promoter region downregulates chemokine CXC ligand 14 expression in gastric cancer. Int J Oncol 2013, 43, 1487–1494. [Google Scholar] [CrossRef] [PubMed]
- Sheffer, M.; Bacolod, M.D.; Zuk, O.; Giardina, S.F.; Pincas, H.; Barany, F.; Paty, P.B.; Gerald, W.L.; Notterman, D.A.; Domany, E. Association of survival and disease progression with chromosomal instability: a genomic exploration of colorectal cancer. Proc Natl Acad Sci U S A 2009, 106, 7131–7136. [Google Scholar] [CrossRef]
- Galamb, O.; Gyorffy, B.; Sipos, F.; Spisak, S.; Nemeth, A.M.; Miheller, P.; Tulassay, Z.; Dinya, E.; Molnar, B. Inflammation, adenoma and cancer: objective classification of colon biopsy specimens with gene expression signature. Dis Markers 2008, 25, 1–16. [Google Scholar] [CrossRef]
- Del Rio, M.; Mollevi, C.; Vezzio-Vie, N.; Bibeau, F.; Ychou, M.; Martineau, P. Specific extracellular matrix remodeling signature of colon hepatic metastases. PLoS One 2013, 8, e74599. [Google Scholar] [CrossRef]
- Neufert, C.; Becker, C.; Tureci, O.; Waldner, M.J.; Backert, I.; Floh, K.; Atreya, I.; Leppkes, M.; Jefremow, A.; Vieth, M.; et al. Tumor fibroblast-derived epiregulin promotes growth of colitis-associated neoplasms through ERK. J Clin Invest 2013, 123, 1428–1443. [Google Scholar] [CrossRef]
- Tang, A.; Li, N.; Li, X.; Yang, H.; Wang, W.; Zhang, L.; Li, G.; Xiong, W.; Ma, J.; Shen, S. Dynamic activation of the key pathways: linking colitis to colorectal cancer in a mouse model. Carcinogenesis 2012, 33, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Hoadley, K.A.; Yau, C.; Hinoue, T.; Wolf, D.M.; Lazar, A.J.; Drill, E.; Shen, R.; Taylor, A.M.; Cherniack, A.D.; Thorsson, V.; et al. Cell-of-Origin Patterns Dominate the Molecular Classification of 10,000 Tumors from 33 Types of Cancer. Cell 2018, 173, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Kon, N.; Yi, J.; Zhao, H.; Zhang, W.; Tang, Q.; Li, H.; Kobayashi, H.; Li, Z.; Duan, S.; et al. Specific regulation of BACH1 by the hotspot mutant p53(R175H) reveals a distinct gain-of-function mechanism. Nature cancer 2023, 4, 564–581. [Google Scholar] [CrossRef]
- Seale, K.; Horvath, S.; Teschendorff, A.; Eynon, N.; Voisin, S. Making sense of the ageing methylome. Nat Rev Genet 2022, 23, 585–605. [Google Scholar] [CrossRef] [PubMed]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell 2013, 49, 359–367. [Google Scholar] [CrossRef]
- Starnes, T.; Rasila, K.K.; Robertson, M.J.; Brahmi, Z.; Dahl, R.; Christopherson, K.; Hromas, R. The chemokine CXCL14 (BRAK) stimulates activated NK cell migration: implications for the downregulation of CXCL14 in malignancy. Exp Hematol 2006, 34, 1101–1105. [Google Scholar] [CrossRef]
- Ozawa, S.; Kato, Y.; Kubota, E.; Hata, R. BRAK/CXCL14 expression in oral carcinoma cells completely suppresses tumor cell xenografts in SCID mouse. Biomed Res 2009, 30, 315–318. [Google Scholar] [CrossRef]
- Shurin, G.V.; Ferris, R.L.; Tourkova, I.L.; Perez, L.; Lokshin, A.; Balkir, L.; Collins, B.; Chatta, G.S.; Shurin, M.R. Loss of new chemokine CXCL14 in tumor tissue is associated with low infiltration by dendritic cells (DC), while restoration of human CXCL14 expression in tumor cells causes attraction of DC both in vitro and in vivo. J Immunol 2005, 174, 5490–5498. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, Q.; Wu, X.; Yu, Y.; Zhang, H. Effect of chemokine CXCL14 on in vitro angiogenesis of human hepatocellular carcinoma cells. Arch Physiol Biochem 2022, 128, 1316–1322. [Google Scholar] [CrossRef]
Figure 1.
CXCL14 was downregulated in clinical colorectal carcinoma. (A) CXCL14 expression level in 259 samples of different stages of tumorigenesis and ten colorectal carcinoma cell lines. (B) CXCL14 expression level of E-GEOD-4183 dataset, including 15 patients with CRC, 15 with adenoma, 15 with IBD, and 8 healthy normal controls. (C) CXCL14 expression profile in E-GEOD-23878 dataset with 35 CRC and 24 normal samples. (D) Overview of CXCL14 expression in E-GEOD-49355 dataset of normal colon, primary tumor, and liver metastasis samples of 13 patients. (E) Line chart of CXCL14 expression of different sample types. (F-G) Paired comparisons of Normal-Primary tumor and Primary-Metastasis tumor. (H) Expression profile in mouse Primary colon cancer model. (I) Expression profile in AOM/DSS induced CRC model.
Figure 1.
CXCL14 was downregulated in clinical colorectal carcinoma. (A) CXCL14 expression level in 259 samples of different stages of tumorigenesis and ten colorectal carcinoma cell lines. (B) CXCL14 expression level of E-GEOD-4183 dataset, including 15 patients with CRC, 15 with adenoma, 15 with IBD, and 8 healthy normal controls. (C) CXCL14 expression profile in E-GEOD-23878 dataset with 35 CRC and 24 normal samples. (D) Overview of CXCL14 expression in E-GEOD-49355 dataset of normal colon, primary tumor, and liver metastasis samples of 13 patients. (E) Line chart of CXCL14 expression of different sample types. (F-G) Paired comparisons of Normal-Primary tumor and Primary-Metastasis tumor. (H) Expression profile in mouse Primary colon cancer model. (I) Expression profile in AOM/DSS induced CRC model.
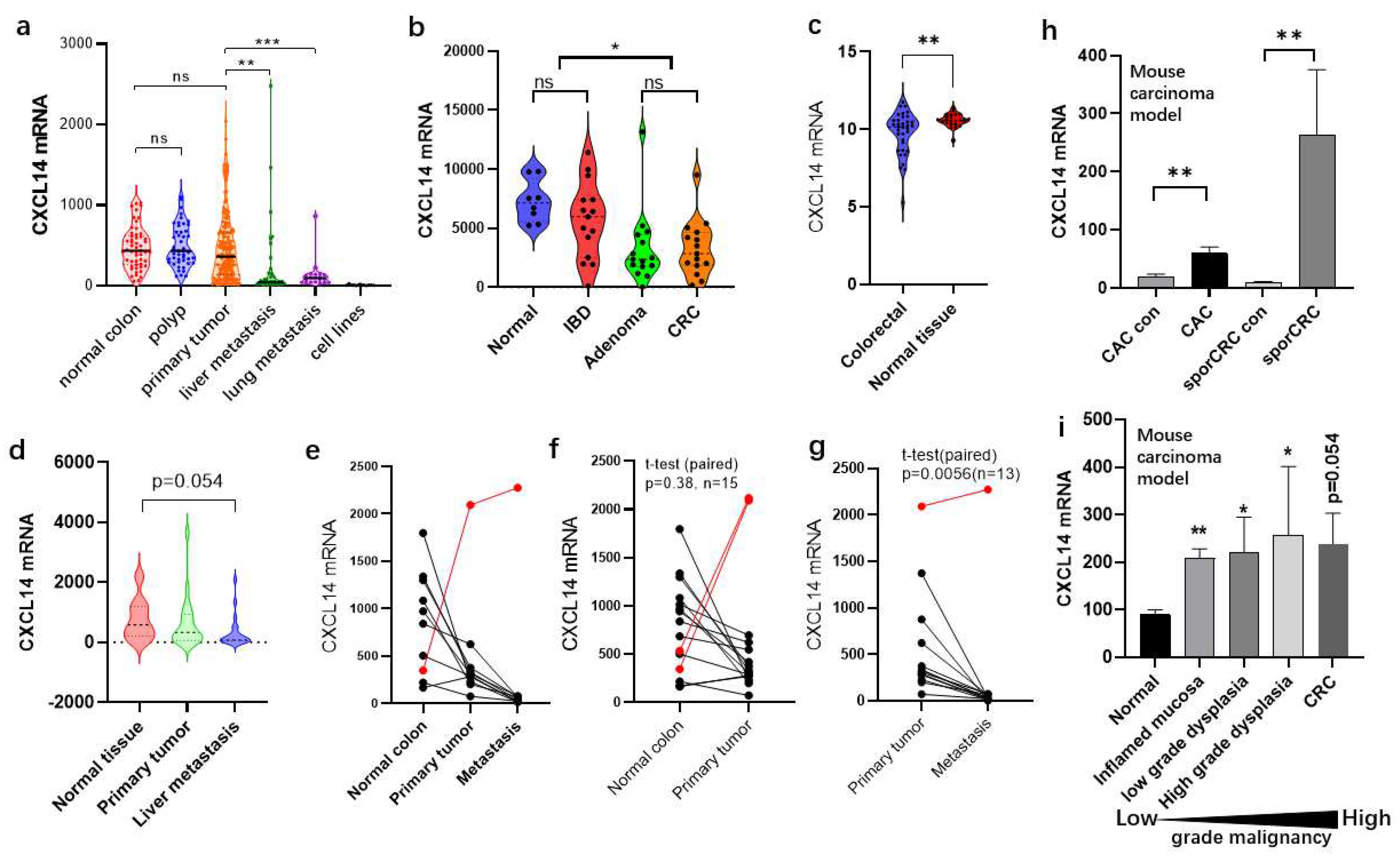
Figure 3.
CXCL14 promoter was hypermethylated in the primary tumor. (A) Tumor sample and normal control of the TCGA dataset, 5mC was sequenced by bisulfite sequencing. (B) Probe number in Fig A. (C-E) Paired samples of primary tumor and normal control were compared in #6~8 locus. (F~H) The correlation between RNA expression and methylation level was assayed by a correlation study.
Figure 3.
CXCL14 promoter was hypermethylated in the primary tumor. (A) Tumor sample and normal control of the TCGA dataset, 5mC was sequenced by bisulfite sequencing. (B) Probe number in Fig A. (C-E) Paired samples of primary tumor and normal control were compared in #6~8 locus. (F~H) The correlation between RNA expression and methylation level was assayed by a correlation study.
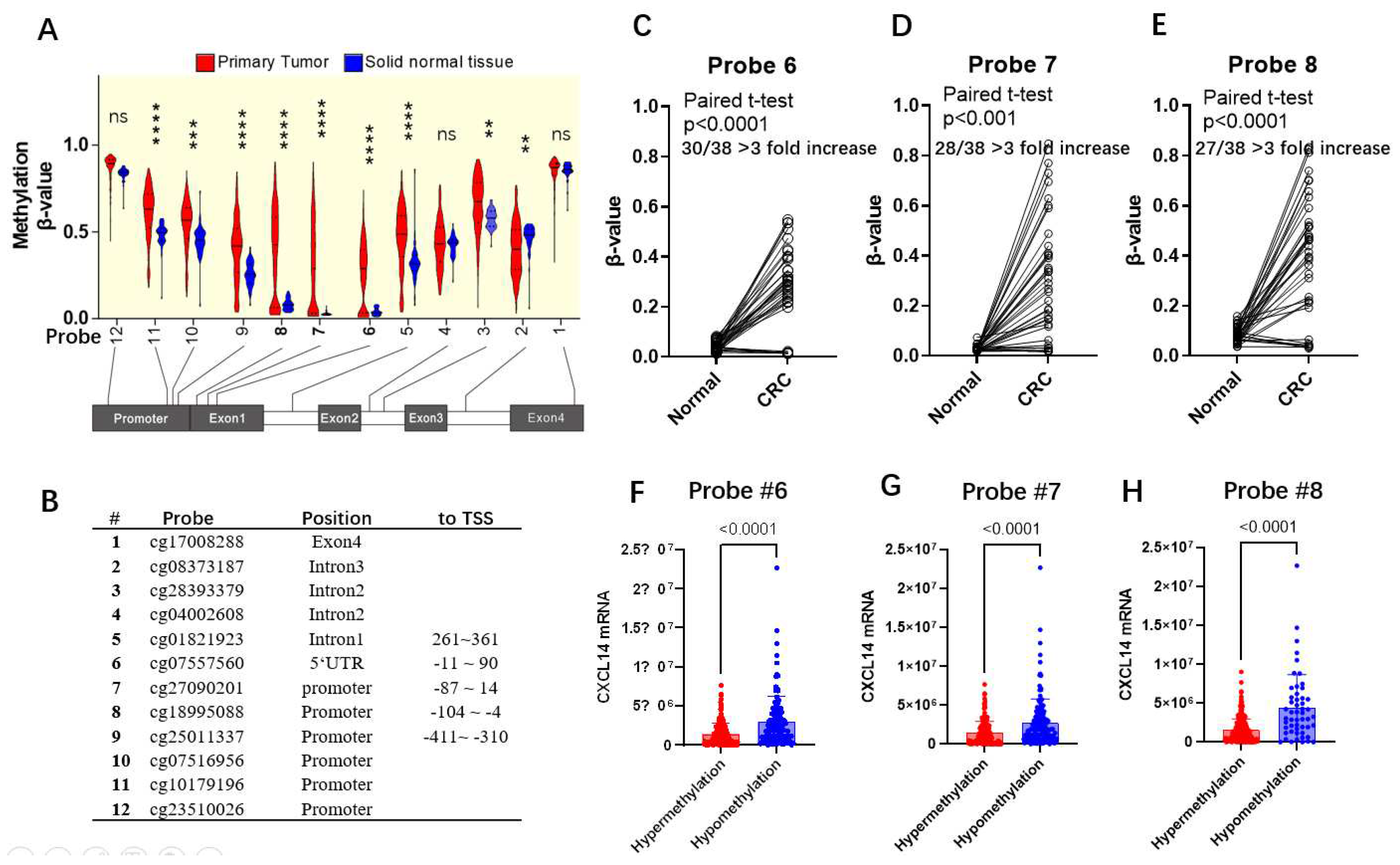
Figure 4.
Methylation of CXCL14 promoter was correlated with anatomic neoplasm subdivision of the primary tumor and age at diagnosis. (A) The CXCL14 promoter methylation level in the different anatomic subdivisions of the colon. (B) CXCL14 mRNA level in different anatomic subdivisions of the colon. (C) The correlation analysis of CXCL14 methylation level of normal colon tissue and donor’s age. (D) Tumor samples were separated into hyper- and hypo-methylation groups by 0.1 of β-value. (E) The group of hypermethylation showed a mild correlation between β-value and age at diagnosis. (F) There is no correlation in the group of hypomethylation. *** p<0.001.
Figure 4.
Methylation of CXCL14 promoter was correlated with anatomic neoplasm subdivision of the primary tumor and age at diagnosis. (A) The CXCL14 promoter methylation level in the different anatomic subdivisions of the colon. (B) CXCL14 mRNA level in different anatomic subdivisions of the colon. (C) The correlation analysis of CXCL14 methylation level of normal colon tissue and donor’s age. (D) Tumor samples were separated into hyper- and hypo-methylation groups by 0.1 of β-value. (E) The group of hypermethylation showed a mild correlation between β-value and age at diagnosis. (F) There is no correlation in the group of hypomethylation. *** p<0.001.
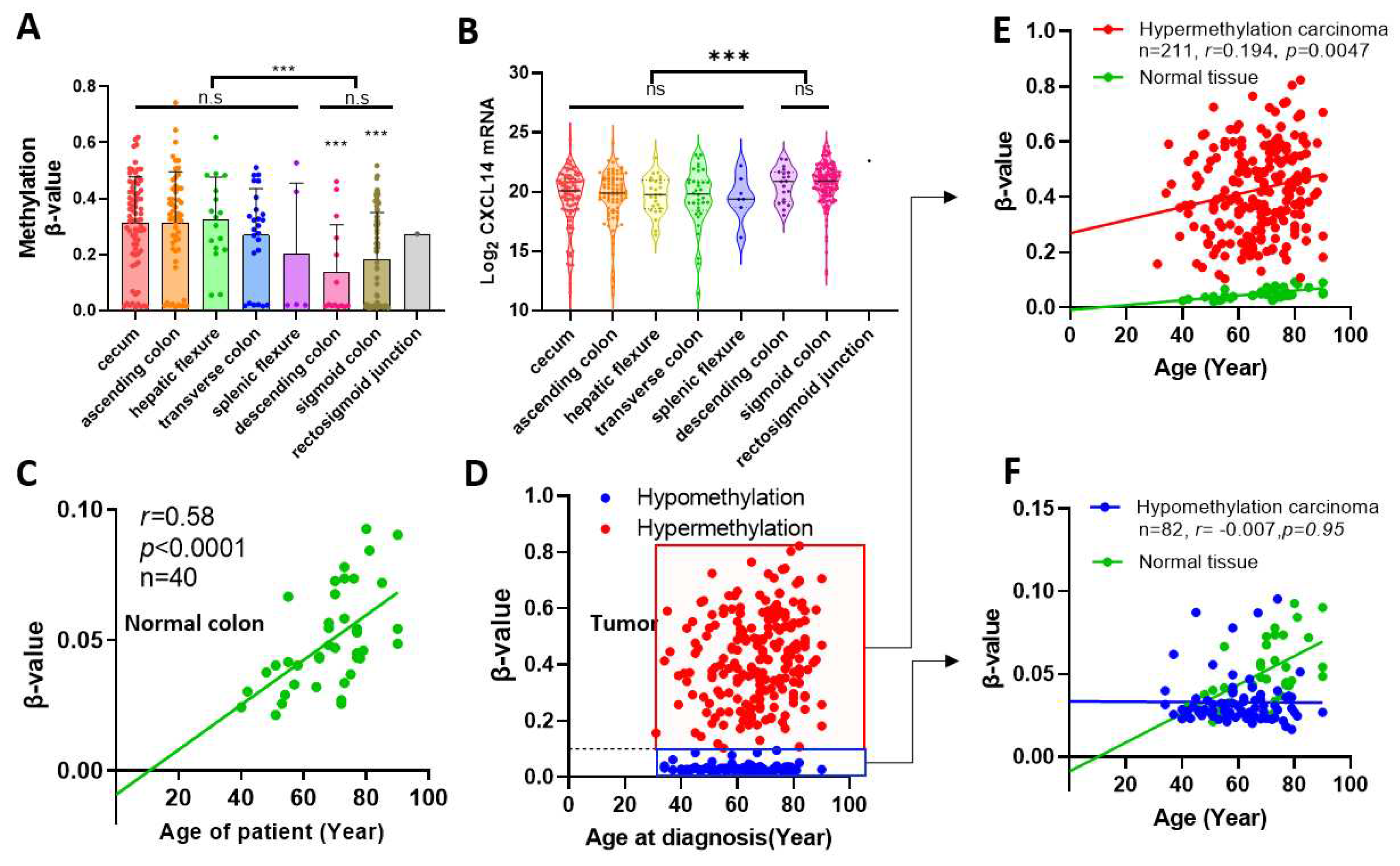
Figure 5.
Methylation of CXCL14 promoter is associated with prognosis. (A)~(H) Kaplan-Meier survival curve of higher and lower CXCL14 promoter methylation group for different probes in TCGA colorectal database. (I)~(K) Kaplan-Meier survival curve examination of overall survival in subjects with high and low methylation levels in the hypermethylation group as mentioned in Figure 4D.
Figure 5.
Methylation of CXCL14 promoter is associated with prognosis. (A)~(H) Kaplan-Meier survival curve of higher and lower CXCL14 promoter methylation group for different probes in TCGA colorectal database. (I)~(K) Kaplan-Meier survival curve examination of overall survival in subjects with high and low methylation levels in the hypermethylation group as mentioned in Figure 4D.
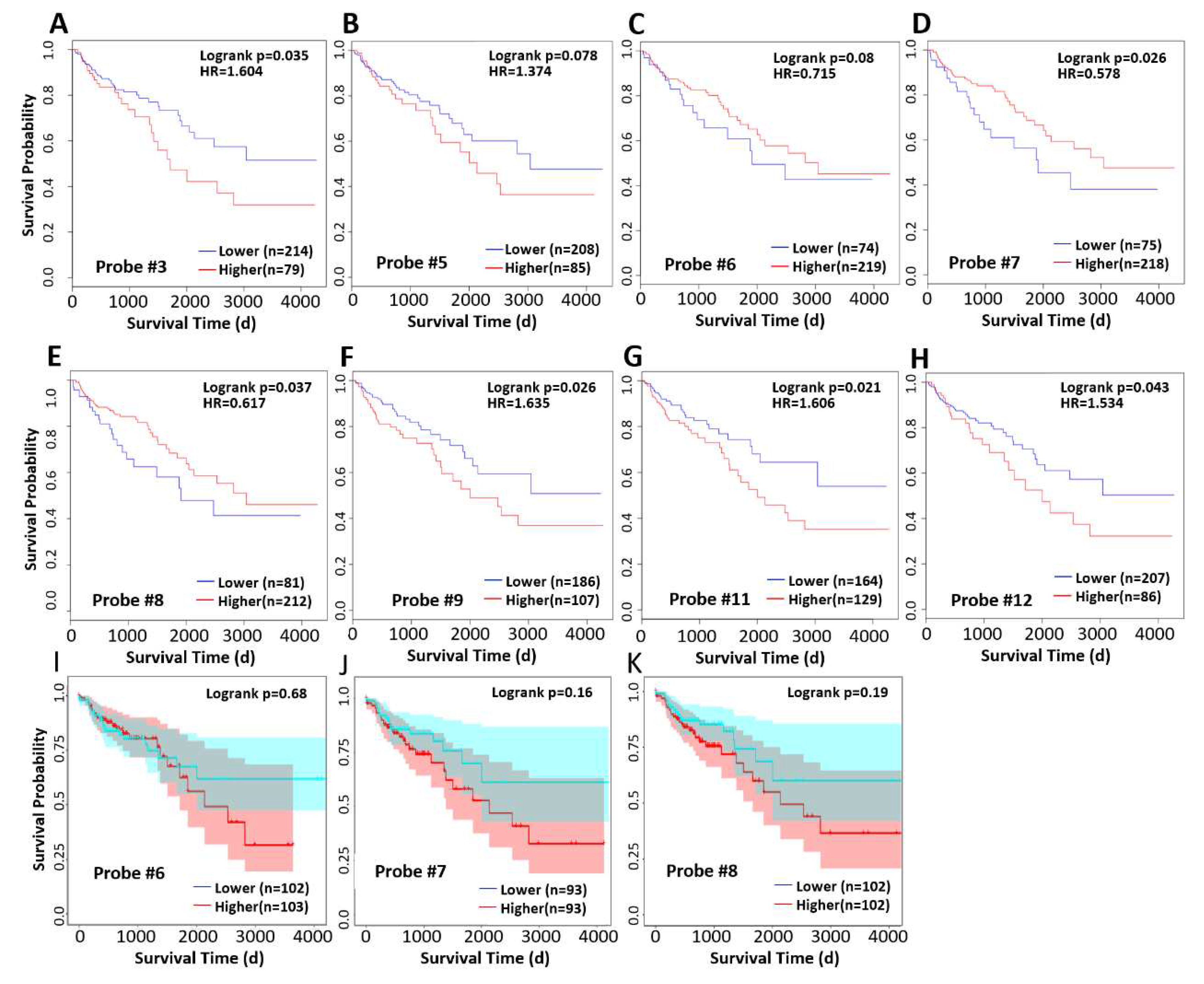
Figure 6.
CXCL14 promoter methylation is negatively correlated with transcription level. (A) HT-29 and (B) HCT116 were treated with DAC with gradient concentration, and CXCL14 silencing was reversed. (C) CXCL14 transcription in HT29 was activated by DAC treatment and decreased after drug withdrawal. (D) Plasmids containing different lengths of truncated CXCL14 promoter were evaluated by luciferase assay. (E) Methylation of the -170~100 region of CXCL14 promoter by M.SssI treatment decreases CXCL14 expression.
Figure 6.
CXCL14 promoter methylation is negatively correlated with transcription level. (A) HT-29 and (B) HCT116 were treated with DAC with gradient concentration, and CXCL14 silencing was reversed. (C) CXCL14 transcription in HT29 was activated by DAC treatment and decreased after drug withdrawal. (D) Plasmids containing different lengths of truncated CXCL14 promoter were evaluated by luciferase assay. (E) Methylation of the -170~100 region of CXCL14 promoter by M.SssI treatment decreases CXCL14 expression.
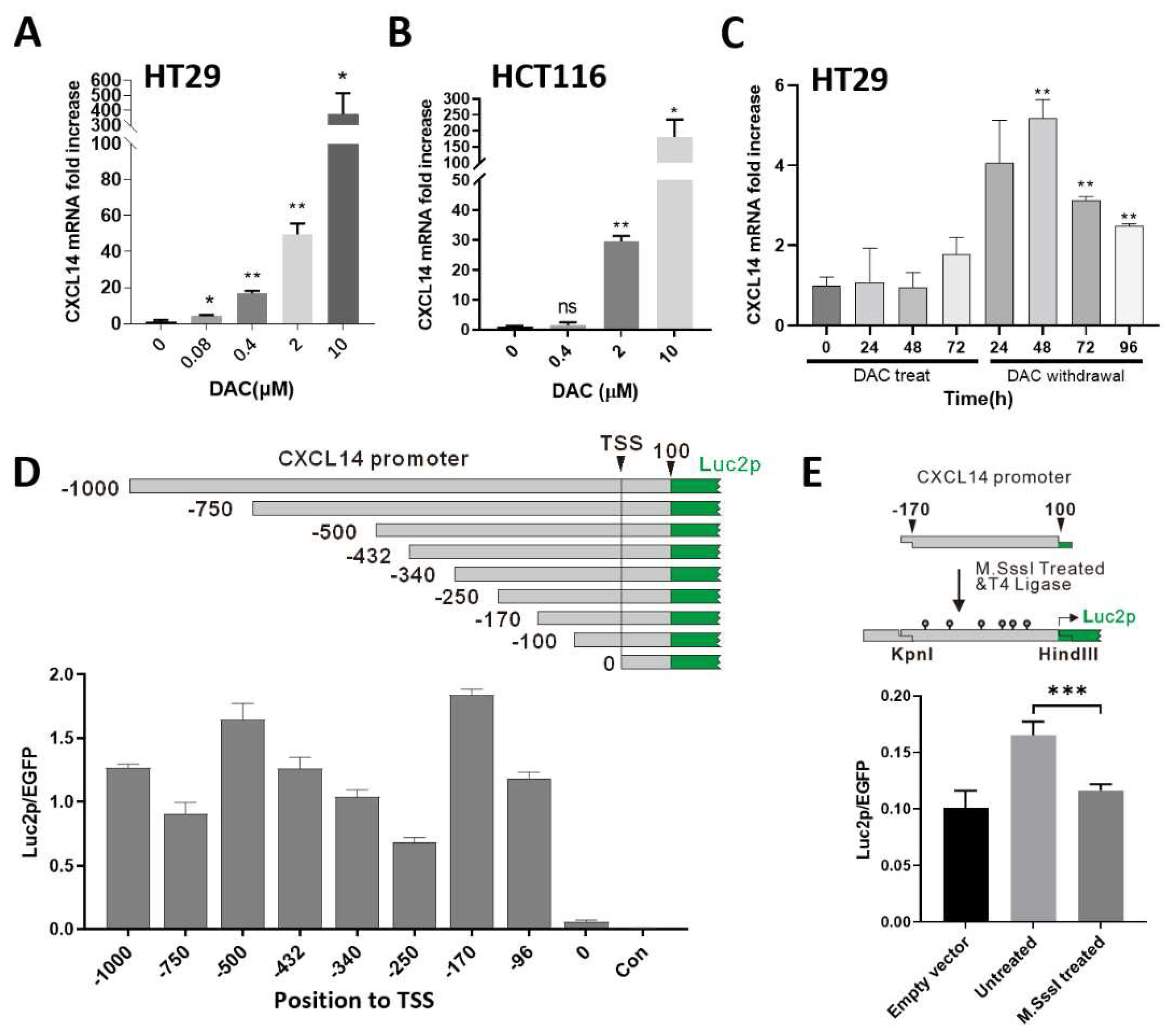

Table 2.
Correlation of CXCL14 methylation and clinical indexes.
| Clinical Parameters | Number | CXCL14 Methylation Status | ||||||
|---|---|---|---|---|---|---|---|---|
| Unmethylated n=84(28%) |
Methylated n=212(72%) |
p-value (χ2) |
||||||
| Gender | Female | 136 | (55%) | 36 | (44%) | 100 | (47%) | 0.614 |
| Male | 112 | (45%) | 46 | (56%) | 112 | (53%) | ||
| Race | Asian | 12 | (4%) | 3 | (4%) | 9 | (5%) | 0.33 |
| Black | 58 | (21%) | 21 | (27%) | 37 | (19%) | ||
| White | 206 | (75%) | 54 | (69%) | 152 | (77%) | ||
| Age | <60 | 98 | (33%) | 43 | (52%) | 55 | (26%) | <0.0001 |
| ≥60 | 196 | (67%) | 39 | (48%) | 157 | (74%) | ||
|
anatomic neoplasm subdivision |
cecum | 75 | (27%) | 13 | (16%) | 62 | (31%) | <10^6 |
| ascending colon | 55 | (20%) | 11 | (14%) | 44 | (22%) | ||
| hepatic flexure | 22 | (8%) | 0 | (0%) | 22 | (11%) | ||
| transverse colon | 25 | (9%) | 6 | (8%) | 19 | (10%) | ||
| splenic flexure | 5 | (2%) | 3 | (4%) | 2 | (1%) | ||
| descending colon | 14 | (5%) | 9 | (11%) | 5 | (3%) | ||
| sigmoid colon | 83 | (30%) | 38 | (48%) | 45 | (23%) | ||
| Poly | Yes | 77 | (35%) | 24 | (39%) | 53 | (34%) | 0.457 |
| No | 140 | (65%) | 37 | (61%) | 103 | (66%) | ||
|
Kras gene mutation |
Yes | 21 | (48%) | 11 | (61%) | 10 | (38%) | 0.139 |
| No | 23 | (52%) | 7 | (39%) | 16 | (62%) | ||
|
lymphatic invasion |
Yes | 79 | (31%) | 25 | (36%) | 54 | (29%) | 0.268 |
| No | 180 | (69%) | 45 | (64%) | 135 | (71%) | ||
|
microsatellite instability |
Yes | 11 | (13%) | 10 | (17%) | 1 | (3%) | 0.126 |
| No | 77 | (87%) | 48 | (82%) | 29 | (96%) | ||
|
New tumor event |
Yes | 62 | (23%) | 17 | (23%) | 45 | (23%) | 0.998 |
| No | 208 | (77%) | 57 | (77%) | 151 | (77%) | ||
|
Perineural invasion present |
Yes | 45 | (25%) | 14 | (30%) | 31 | (24%) | 0.423 |
| No | 132 | (75%) | 33 | (70%) | 99 | (76%) | ||
|
neoplasm cancer status |
Yes | 62 | (25%) | 23 | (33%) | 39 | (22%) | 0.069 |
| No | 187 | (75%) | 47 | (67%) | 140 | (78%) | ||
|
Tumor stage |
I | 44 | (15%) | 6 | (8%) | 38 | (18%) | 0.019 |
| II | 114 | (40%) | 30 | (39%) | 84 | (41%) | ||
| III | 85 | (30%) | 23 | (30%) | 62 | (30%) | ||
| IV | 41 | (14%) | 18 | (23%) | 23 | (11%) | ||
Table 1.
Dataset used in the study.
| Species | Sample type | Sample number | Accession No. | reference | |
|---|---|---|---|---|---|
| 1 | Human | Primary colon adenocarcinomas, adenomas, metastasis and corresponding normal mucosae. | 390 | E-GEOD-41258 | [17] |
| 2 | Human | 15 CRC, 15 with adenoma, 15 IBD and 8 healthy normal controls | 53 | E-GEOD-4183 | [18] |
| 3 | Human | 36 CRC tissues and 24 non-cancerous colorectal tissue | 60 | E-GEOD-23878 | |
| 4 | Human | Paired primary tumors and hepatic metastases and normal controls | 57 | E-GEOD-49355 | [19] |
| 5 | Human | Colorectal adenomas, adenocarcinomas, and normal control | 336†/556 | TCGA-COAD PanCancer Atlas |
[22] |
| 6 | Mouse | AOM/DSS induced CAC, ApcMin/+/J spontaneous tumor and para-carcinoma tissue | 14 | E-GEOD-43338 | [20] |
| 7 | Mouse | Ulcerative CAC | 18 | E-GEOD-31106 | [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



