Submitted:
02 October 2023
Posted:
03 October 2023
You are already at the latest version
Abstract
Crossostephium chinense is a wild species with strong salt tolerance, which has great potential to improve the salt tolerance of cultivated chrysanthemums. Conversely, the unique salt-tolerant molecular mechanisms of Cr. chinense are still unclear. This study performed a comparative physiological and transcriptome analysis of Cr. chinense, Chrysanthemum lavandulifolium and their hybrids. The physiological results showed that Cr. chinense maintained a higher superoxide dismutase (SOD) activity, to alleviate the oxidative damage to membrane. KEGG enrichment analysis showed that the plant hormone signaling transduction and the MAPK signaling pathway were mostly enriched in Cr. chinense and hybrids under salt stress. Further weighted gene co-expression network analysis (WGCNA) of DEGs suggested that the abscisic acid (ABA) signaling transduction may play a significant role in the salt-tolerant mechanisms of Cr. chinense and hybrids. The tissue-specific expression patterns of the candidate genes related to ABA signaling transduction and MAPK signaling pathway indicated that genes related to ABA signaling transduction performed significant expression levels under salt stress. This study provides a theoretical foundation for future research into the molecular mechanism of salt tolerance in chrysanthemums under salt stress.
Keywords:
salt stress
; Crossostephium chinense
; ABA signaling transduction
; molecular mechanism
1. Introduction
The accumulation of salts in soils is a problem that has limited plant growth and productivity [1]. More seriously, salt stress will bring out the appearance of wilt and death in plants [2]. For instance, salt stress has both osmotic and ionic effects on plant cells, including the secondary effects with oxidative stress and damage to cellular components [3].
Plants have the ability to defense varieties of abiotic stresses, activating various responses to maintain their normal growth [4]. Under salt stress, ion imbalance and water deficiency in the plant cell will cause osmotic stress, leading to the accumulation of osmolytes and antioxidants [5]. Correspondingly, plants can figure out these different stresses through protein kinases, which are involved in cellular regulation and metabolism. Recent studies suggest that protein kinases such as mitogen-activated protein kinase (MAPK) cascades, calcium-dependent protein kinases (CDPKs/CPKs) and sucrose nonfermenting 1 (SNF1)-related protein kinases (SnRKs) are vital to salt-stress-induced oxidative stress response [6,7,8]. Moreover, plants have developed phytohormone-mediated stress tolerant mechanisms [5]. Different mechanisms are exploited through regulation of stress-responsive gene expression to avoid and tolerate dehydration, involved in ABA and other signaling pathways [9]. Furthermore, MPKs are reported to be involved in ABA signaling and H2O2-mediated stomatal closure, indicating a potential link between MAPK cascades with ABA signaling [10].
Chrysanthemum is one of the most important ornamental plants in the world, while the planting areas of cultivated chrysanthemums are limited due to the susceptibility to salt stress. Therefore, improving salt tolerance is an important goal in chrysanthemum breeding. Previous studies have been found that some wild species in Artemisia and Chrysanthemum are more tolerant in salt stress [11,12,13]. Several studies have shown that the salt tolerance of wild chrysanthemums can be inherited to hybrids through distant hybridization [14,15]. The molecular mechanisms of chrysanthemum under salt stress are controlled by multiple genes involved in osmotic adjustment, ion transport and the regulation of ABA signaling [16,17,18]. Besides, transcription factor (TF) families in chrysanthemum including WRKYs, NACs, ZIPs, MBFs are reported to have important roles in salt stress [19,20,21,22].
Crossostephium chinense is a diploid species in the Asteraceae family, which is often distributed in the southeast of China with strong salt tolerance [23,24]. Our previous research has obtained several distant hybrids of Cr. chinense and C. lavandulifolium [25]. In this study, to deeply reveal the unique salt-tolerant mechanisms of Cr. chinense, hub genes were selected out through physiological, transcriptome and bioinformatics analysis. It was found that the genes involved in ABA signaling transduction showed dominantly expression levels. Our finding not only provided a better insight into the difference in salt tolerance between Cr. chinense and C. lavandulifolium, but also contributed potential pathways for breeding better salt-tolerant chrysanthemums.
2. Results
2.1. The activities of antioxidant-related enzymes
After treatment with 700 mM NaCl in seven days, the growth of Cr. chinense (CC) and its hybrids (CE, CT and CF) were significantly better than C. lavandulifolium (CL) (Figure 1A). The results showed that salt treatment aggravated the wilting of C. lavandulifolium with the accumulation of salinity while Cr. chinense and hybrids were slightly affected. To investigate the various response of plants under salt stress, activities of SOD and MDA were monitored at three time points. The SOD activity of all plants increased after 12 hours under salt stress (Figure 1B). Furthermore, the SOD activity of CC was substantially higher than other four plants at 12 h. The MDA activity of CC and three hybrids was obviously less than CL, and CC maintained at a minimum level (Figure 1C). The results indicated that Cr. chinense performed higher antioxidant enzyme activities, preventing oxidative damage to membrane. In conclusion, antioxidant enzyme activities of all plants began to change after 6 h under 700 mM NaCl treatment, suggesting that the genes related to salt stress were activated.
2.2. RNA-Sequencing and de novo Assembly
To further reveal the molecular mechanisms of Cr. chinense, leaf samples treated with 700mM for 6h were sequenced for analysis. After filtering the low-quality sequences and the adaptor sequences, a total of 1,200,999,186 clean reads, resulting in 168.16 Gb sequence data of 30 libraries. The statistics of the sequencing quality are provided in Table S2 and de novo assembly statistics are presented in Table S3. The unigene length distribution is shown in Figure 2A and the three biological replicates of each treatment were similar in the overall gene expression levels (Figure 2B). A total of 113700 unigenes were annotated after assembly. And the details of unigenes annotated by each database were shown in Table S4.
2.3. Comparative enrichment analyses of the DEGs under salt treatment
Comparing the salt treatment group with the control group, there were more downregulated DEGs in five samples under salt stress and the upregulated genes in Cr. chinense were less than hybrids and C. lavandulifolium (Figure 3A). In the comparison of salt treatment group to salt treatment group of CL, there were more downregulated genes in Cr. chinense (Figure 3B). DEGs were further analyzed to compare the different pathways of salt tolerance in plant materials. To validate the authenticity of the RNA-seq data, the expression levels of 10 randomly selected genes were detected by qRT-PCR. The qRT-PCR results were generally consistent with DEG analysis of the transcriptome (Figure S1).
The GO terms of C. lavandulifolium were compared with the salt-tolerant plants under salt stress. The DEGs in four groups were mostly enriched in the “molecular function (GO:0003674)” and “DNA binding transcription factor activity (GO:0003700)” categories. Additionally, the DEGs were specially enriched in “protein serine/threonine kinase activity (GO:0004674)” term of CT6h_vs_CL6h, “response to abscisic acid (GO:0009737)” and “response to salt stress (GO:0009651)” terms of CE6h_vs_CL6h, and “response to water deprivation (GO:0009414)” term of CF6h_vs_CL6h. The results indicated that the response of Cr. chinense and hybrids to salt stress involves multiple processes such as kinase activity and chloroplast activity (Figure 4). Besides, KEGG-based DEGs enrichment analysis showed that “plant hormone signal transduction” was mostly enriched in four groups under salt stress. Common categories of high enrichment included “photosynthesis” in CC6h_vs_CL6h, “MAPK signaling pathway”, “Phenylpropanoid biosynthesis” and “photosynthesis” in CT6h_vs_CL6h and “Galactose metabolism” in CE6h_vs_CL6h (Figure 5). According to the KEGG results, the “plant hormone signal transduction” and “MAPK signaling pathway” may be the potential biological pathways to resistant salt stress in Cr. chinense and hybrids.
2.4. Analysis of gene co-expression network
To further understand the gene expression of the CC, CL and three hybrids under salt stress, the hub genes related to salt tolerance were screened out through weighted gene co-expression network analysis (WGCNA). After filtering raw data in materials and methods, 34760 genes were retained for WGCNA analysis. The co-expression network was constructed based on the gene expression of 30 samples. In this study, we selected the weight value β = 22 to construct the scale-free networks, describing different modules with different colors. A total of 35 modules were identified after identifying the result of the system clustering tree by dynamic hybrid algorithm based on dissTOM (Figure 6). The blue module (5041) was with the maximum count, and the darkmagenta was with the minimum (40) (Table S5).
To screen out the modules that are significantly related to specific samples, the heatmap of the sample expression mode was conducted to select corresponding modules. As shown in the picture (Figure 7A), the yellow module had relatively high correlation with CC6h, indicating that the genes in yellow module may be related to salt stress response. Similarly, the green module was highly correlated with CE6h and CF6h, the red module was highly correlated with CT6h, and the brown module was highly related to CL6h. Therefore, this study identified 4 co-expression modules as key modules for further research. Analysis of the expression patterns of genes showed that genes in each module were differentially expressed in different samples (Figure 7B). Significantly, the genes in the green module were up-regulated in CE and CF, and the genes in the yellow module were specially up-regulated in CC.
KEGG Pathway enrichment analysis showed that the highly enriched pathways in Environmental Information Processing of four modules included the plant hormone signal transduction (map04075) and MAPK signaling pathway (map04016). The GO enrichment analysis showed that the highly enriched biological processes in green module included response to water deprivation (GO:0009414) and response to abscisic acid (GO:0009737). In yellow and red module, the biological processes were differentially enriched in protein phosphorylation (GO:0006468) and defense response (GO:0006952) (Figure S2). The results indicated that MAPK signaling pathway and the plant hormone signaling transduction especially abscisic acid signaling transduction played an important role in the response of Cr. chinense and its hybrids to salt stress.
The plant hormone signal transduction and MAPK signaling pathway were merged in each module and the genes enriched in each module were sorted according to the order of gene connectivity from high to low. The top 5 genes with gene connectivity were regarded as hub genes (Table S6). And the top 20 genes with weight values from high to low related to hub genes were considered as the associated genes (Table S7). In yellow module, 71 associated genes were screened, including TRINITY_DN84481_c0_g2, which encoded TATA-box binding protein-associated factor (TAF), TRINITY_DN65102_c0_g1, which encoded Trehalose-6-phosphate synthase (TPS) and genes encoding flavonoid biosynthesis, DNA binding and transporter activity. And most of associated genes were screened as unknown genes. In green module, 49 associated genes were screened, including TRINITY_DN83699_c2_g1, encoding 9-cis-epoxycarotenoid dioxygenase3 (NCED3), TRINITY_DN59565_c0_g1, encoding Calcineurin B-like calcium sensor interacting Protein Kinase (CIPK), TRINITY_DN70712_c0_g1, encoding MAPK kinases and genes encoding protein serine/threonine phosphatase activity and plasma membrane. In brown module, 64 associated genes were screened, including genes encoding protein binding and chlorophyll binding. In red module, 56 associated genes were screened, including genes encoding DNA binding and calcium ion binding (Table S8). The weighted gene co-expression network was drawn using Cytoscape_3.9.1 (Figure 8).
2.5. Tissue-specific expression pattern analysis of hub genes
To further explore the tissue-specific expression patterns of the hub genes under salt stress, qRT-PCR was performed on leaves, shoots and roots of five plant materials (Figure 9). Under salt stress, the gene TRINITY_DN66554_c0_g1 showed a significant downregulation trend, while the other seven genes were showed an upregulation trend. Remarkably, the gene TRINITY_DN77201_c0_g1 had higher expression levels in Cr. chinense, the gene TRINITY_DN61103_c1_g2 and the gene TRINITY_DN80388_c0_g1 showed higher expression levels in hybrids. The results of qRT-PCR analysis provided further evidence for revealing the salt-tolerant molecular mechanisms of Cr. chinense and hybrids.
3. Discussion
Soil salinization is one of the most severe environmental stresses, thus developing elite salt-tolerant germplasm resources through molecular breeding and traditional breeding approaches is available to use saline-alkali land [9]. Cr. chinense is a salt-tolerant cultivar but little is known about the salt-tolerant mechanisms. The intergeneric hybridization between Cr. chinense and C. lavandulifolium improves the salt tolerance of hybrids. Excessive salt induces oxidative stress, resulting in the increase of reactive oxygen species [26]. The excessive amounts of reactive oxygen will bring about membrane peroxidation reaction, producing harmful substances [16]. The physiological results showed that Cr. chinense maintained higher antioxidant enzyme activities, preventing oxidative damage to membrane. In the present study, we performed de novo transcriptome sequencing of Cr. chinense, C. lavandulifolium and their hybrids to better analyze the underlying molecular mechanisms of strong tolerance in Cr. chinense. To date, although a few transcriptome studies of salt stress in chrysanthemums have been published [27,28], but there is a lack of transcriptome analysis for exploring the differences between salt-tolerant cultivars and intergeneric hybrids. The results of GO and KEGG analysis of the whole transcriptome showed that the plant hormone signaling transduction and MAPK signaling pathway featured in the response of Cr. chinense and its hybrids to salt stress. The KEGG and GO pathway enrichment in WGCNA analysis further revealed that the ABA signaling transduction might play a crucial role in the salt-tolerant process. Therefore, the hub genes and associated genes related to the ABA signaling transduction and MAPK signaling pathway were selected to have an insight into the salt-tolerant functions.
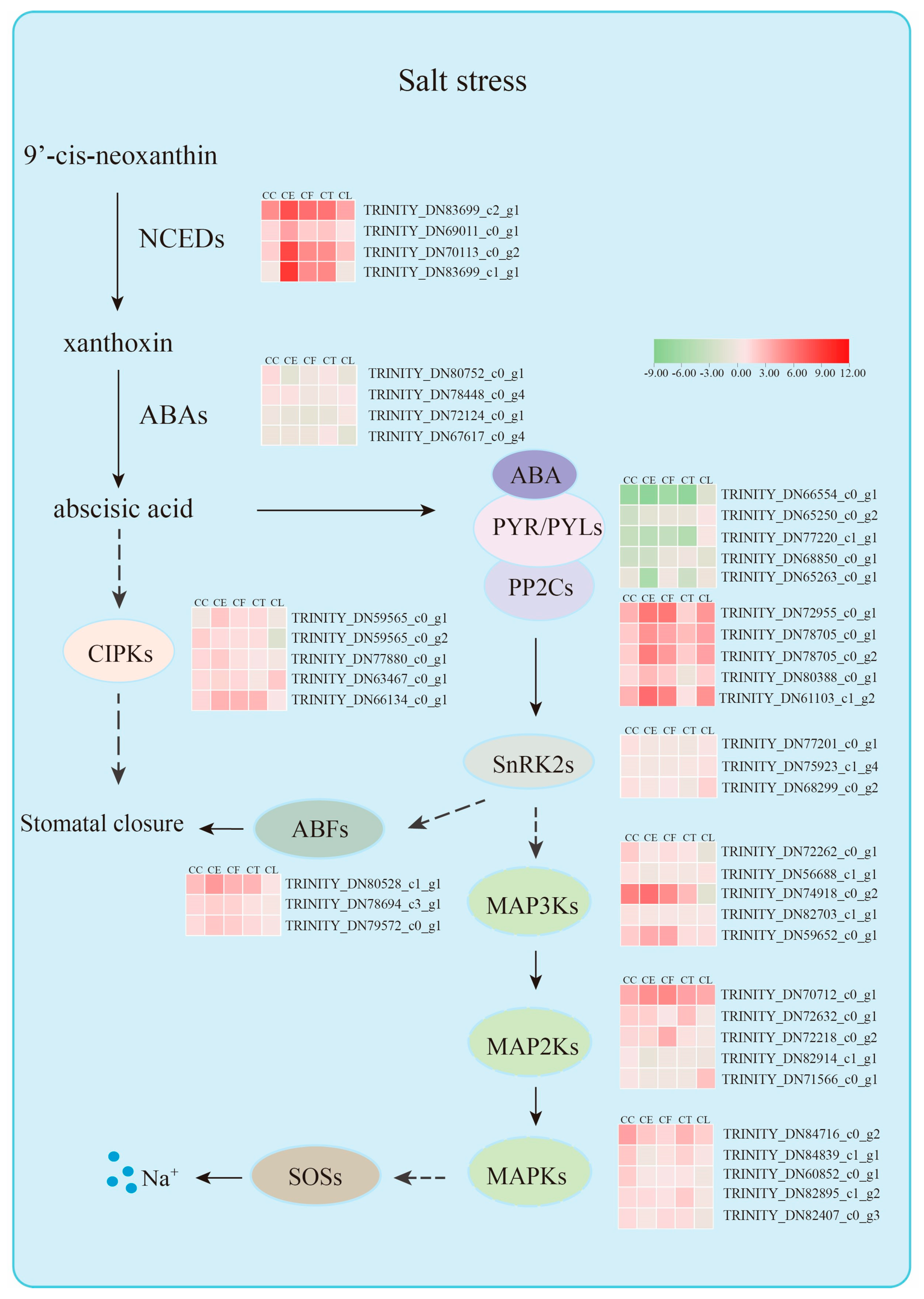
Plant hormones are significant signaling molecules that regulate growth, development, and defense in plants [29]. The 5 hub genes in green module, which was highly correlated with the sample CE and CF by salt stress, were all related to the hormone abscisic acid. ABA plays a crucial role in the closure of stomata by regulating guard cell ion fluxes towards varieties of stresses including salt, drought and water [30,31,32]. The ABA receptor-coupled core signaling pathway consists of three components containing the PYRABACTIN RESISTANCE (PYR)/PYR-LIKE (PYL)/REGULATORY COMPONENTS OF ABA RECEPTORS (RCAR) family proteins; the negative regulator clade A type 2 C protein phosphatases (PP2Cs); and the positive regulator SNF1-related protein kinase 2 s (SnRK2s) [33]. After ABA binding to PYR/PYL/RCARs, the ABA receptor complex inhibits the activity of PP2C phosphatases, releasing SnRK2s from PP2C-mediated inhibition [34]. SnRK2s are activated through autophosphorylation or other protein kinases, participating in varieties of physiological responses through phosphorylating target substrates including ion channels, transcription factors and transporters [35]. PP2CA and ABI1 are two branches of Protein phosphatases type 2C (PP2Cs) from group A [36]. And it is established that ABI1 is a negative regulator of SnRK2.4, and PP2CA interacts with and inhibits SnRK2.4, which belongs to SnRK2s activated by ABA [37]. However, the expression levels of PP2Cs are actually upregulated by abiotic stress and ABA, which is possibly induced by an ABA desensitization mechanism to adjust ABA signaling [38].
To prevent plants from losing water, ABA can mediate the closure of stomata pores by activating Ca2+ entry into the cytoplasm [39,40]. The evidence shows that Ca2+ signals can be decoded by several Ca2+ sensors including calcium dependent protein kinase CPK6, which can activate the downstream targets involved in stomatal closure [41]. In our study, we find that the gene TRINITY_DN59565_c0_g1 encoding CIPK2 had a correlation with the 5 hub genes in green module, which may be the potential conduction factors in ABA signaling pathway. In addition, the hub gene TRINITY_DN80528_c1_g1 encoding ABA-response element (ABRE)-binding factors (ABFs) in green module may be related to the hub gene TRINITY_DN77201_c0_g1 in yellow module. Recent research shows that PtrSnRK2.4 can interact with PtrABF2 by forming a heterodimer and PtrABF2 can be phosphorylated by PtrSnRK2.4 at Ser93 to modulate plants drought tolerance [42]. Besides, overexpressing DcABF3 in Arabidopsis increases the stomatal density and affects the water deficit tolerance, indicating that ABFs can mediate the stomatal development [43].
Moreover, the gene TRINITY_DN83699_c2_g1 encoding 9-cis-epoxycarotenoid dioxygenase3 (NCED3) as a candidate gene, was found to have a strong correlation with 5 hub genes in green module. Plant endogenous abscisic acid levels will increase dramatically, which is important to stomatal regulation and gene expression during periods of abiotic stress [44,45]. ABA is metabolized from xanthoxin, which is cleaved from carotenoids 9-cisneoxanthin and 9-cis-violoxanthin in the chloroplast [45]. And this carotenoid cleavage reaction is catalyzed by the 9-cisepoxycartenoid dioxygenases (NCEDs) [44]. Besides, both ABA-dependent and ABA-independent pathways govern the induction of NCED3 showing that NCED3 is NaCl-dependent [46]. In Arabidopsis, AtNCED3 was proved to be the major stress-induced NCED in leaves, and other four AtNCEDs (2, 5, 6 and 9) were found to differ in binding activity of the thylakoid membrane [47]. These evidences indicated that ABA played a significant role through varieties of ABA receptors and key enzyme genes. PP2Cs played a negative role in the process of plant endogenous ABA levels. Consistent with the previous conclusions, this study found that salt stress activated the expression of key elements in ABA signaling transduction, in which TRINITY_DN61103_c1_g2, TRINITY_DN80388_c0_g1, TRINITY_DN80528_c1_g1 and TRINITY_DN83699_c2_g1 were remained to be confirmed.
There has been a whole train of regulatory processes coordinating plant responses to abiotic stresses in plants, including chromatin modifications, transcriptional regulation, alternative splicing, protein phosphorylation and ubiquitnation/sumoylation [48]. Among them, protein phosphorylation is an important mode of signal transduction to plants, where protein kinases play a crucial role in transferring the phosphoryl group [49,50]. In this study, the MAPK signaling pathway was significantly enriched in red module, which were highly related to the sample CT by salt stress. Mitogen-activated protein kinase modulates plant tolerance to salt stress and the canonical MAPK is composed of three types of kinases: MAPK kinase kinases (MAPKKKs/MAP3Ks/MEKKs), MAPK kinases (MKKs/MAP2Ks/MEKs) and MAP kinases (MAPKs/MPKs) in a sequential phosphorelay starting from a MAPKKK [51,52]. Evidence shows that MAPK cascades like MAP3K17/18, MKK3 and MPK1/2/7/14 are activated by the PYR/PYL/RCAR-SnRK2-PP2C ABA core signaling module through protein synthesis of the MAP3Ks [53]. Furthermore, MPK1 and MPK2 can be activated by ABA in a SRK2D/E/I-dependent manner, connecting the ABA and MAPK modules [54]. In Arabidopsis, MPK6 can interact with and phosphorylate the C-terminal fragment of SOS1, which can extrude Na+ into the soil solution and load Na+ into the xylem [55]. Accordingly, we found the gene TRINITY_DN77201_c0_g1 in yellow module, which were highly associated with CC, and the gene TRINITY_DN75923_c1_g1 in brown module associated with CL were related to SnRK2s gene family. In conclusion, the MAPK signaling pathways can function in ABA signaling transduction through potential genes, which need to be unraveled in future researches.
Tissue-specific expression patterns of candidate genes could get a better knowledge of the gene functions in different plant materials. The significant expression levels of the gene TRINITY_DN66554_c0_g1 and TRINITY_DN83699_c2_g1 in leaves, shoots and roots indicated that the regulation of ABA played a key role in five materials response to salt stress. And the gene TRINITY_DN80388_c0_g1 and TRINITY_DN61103_c1_g2, which were related to green module, were highly expressed in hybrids. The gene TRINITY_DN77201_c0_g1, which was related to yellow module, was highly expressed in Cr. chinense. According to the hub genes and other potentially associated genes screened in each module, a hypothetical model was proposed to explain the mechanism of Cr. chinense under salt stress and analyze the differential expression of gene family in related pathways (Figure 10). However, further studies are required to validate the definite roles of genes and the potential connections between signaling pathways.
4. Materials and Methods
4.1. Plant materials and culture conditions
Cr. chinense (CC), C. lavandulifolium (CL) and their three hybrids (CE, CT, CF) were conserved in the breeding greenhouse of the National Flower Engineering Center, Beijing, China. Rooted seedlings at the 6-7 leaf stage were transplanted into mixture soil of peat, vermiculite and pearlite (3:1:1, v/v/v). The plants were cultured at 22 ± 2℃, 65 ± 5% relative humidity with a photoperiod of 16:8 (light: dark). The uniform plant seedlings with 8-10 leaves were selected for the following experiment.
4.2. Salt treatments and stress tolerance observation
To observe the salt-tolerant differences of Cr. chinense, C. lavandulifolium and its hybrids, the plants were irrigated with 700mM NaCl. All the plants were photographed on day 3 and day 7.
To ensure the best time point for transcriptome sampling, the seedlings were treated with 700 mM NaCl for 0, 6, 12 h separately. Fresh Leaves (about 0.1g) were harvested at three time points to determine the SOD and the MDA activity. The activities of SOD (U/g FW) and MDA (µmol/g FW) were measured according to the analysis kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China, www.njjcbio.com).
4.3. cDNA library construction and illumina sequencing
For transcriptome analysis, the plants were exposed to 700mM NaCl for 6 h. All the leaf samples of each set with three biological replicates were immediately immersed in liquid nitrogen and stored at -80℃ for RNA extraction. Total RNA was extracted from plant leaves using TRIzol reagent (Invitrogen, CA, USA) following the manufacturer’s procedure. To analyze the quantity and purity of the total RNA, the Bioanalyzer 2100 and RNA 1000 Nano LabChip Kit (Agilent, CA, USA) with RIN number >7.0 were used. Poly (A) RNA was purified from total RNA (5 µg) using poly-T oligo-attached magnetic beads and two rounds of purification. Following purification, the mRNA was fragmented into small pieces using divalent cations under elevated temperature. The cleaved RNA fragments were reverse-transcribed to create the final cDNA library in accordance with the protocol for the mRNASeq sample preparation kit (Illumina, San Diego, CA, USA). The average insert size for the paired-end libraries was 300 bp (±50 bp). The paired-end sequencing was performed on an Illumina NovaseqTM 6000 at the (LC Sciences, USA) following the vendor’s recommended protocol.
4.4. De novo assembly, unigene annotation and functional classification
Clean reads were obtained after using Cutadapt and perl scripts in house to remove the reads that contained adaptor contamination, low quality bases and undetermined bases [56]. Then sequence quality was verified using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/), based on Q20, Q30 and GC-content of the clean data. All downstream analyses were based on clean data of high quality. De novo assembly of the transcriptome was performed with Trinity 2.4.0 [57]. For shared sequence content, trinity groups transcripts clustered into unigenes. All assembled unigenes were aligned against the non-redundant (Nr) protein database (http://www.ncbi.nlm.nih.gov/), Gene ontology (GO) (http://www.geneontology.org), SwissProt (http://www.expasy.ch/sprot/), Kyoto Encyclopedia of Genes and Genomes (KEGG)(http://www.genome.jp/kegg/) and eggNOG (http://eggnogdb.embl.de/) databases using DIAMOND with a threshold of Evalue<0.00001 [58].
4.5. Differentially expressed unigene analysis
To perform expression level for Unigenes, salmon was used by calculating TPM [59,60]. The differentially expressed Unigenes were selected with log2 (fold change) >1 or log2 (fold change) <-1 and with statistical significance (p value < 0.05) by R package edgeR [61]. GO and KEGG enrichment analyses were performed on the differentially expressed unigenes using perl scripts in-house. And ten randomly selected DEGs were used for qRT-PCR analysis to validate the authenticity of the RNA-seq data.
4.6. Weighted gene co-expression network analysis
Co-expression-network analysis was constructed using the weighted gene correlation network analysis (WGCNA) package in R under the guidelines of the published tutorials. Genes with FPKM < 10 in 30 samples were filtered. Hierarchical clustering of the samples was conducted based on Euclidean distance computed on gene expression data, and integrated with the (clinical information of patients). Outlier samples were removed. The parameters were selected by default software parameters (soft threshold = 22 (estimate value); min module size = 30; merge cut height = 0.25). The core DEGs were further divided into 36 modules. For each module, the eigengene (the first component expression of genes in module) was determined, and the correlations of eigengenes were then subsequently calculated. Genes with high connectivity in the respective modules were considered hub genes. Network visualization for each module was performed using the Cytoscape software version 3.9.1 with a cut off of the weight parameter obtained from the WGCNA set at 0.3.
4.7. Tissue-specific expression patterns of the candidate genes
To analyze the gene expression of different tissues in plants, 8 candidate genes with high expression in key modules were selected for qRT-PCR analysis. The experimental plants were exposed to 700mM NaCl for 6h. The collecting methods of leaf, shoot and root samples were consistent with 2.3. The specific primers for these 8 genes and actin gene (an internal control) were designed by IDT (https://www.idtdna.com) (Table S1). 1 µg of total RNA were used to synthesize the cDNA according to the manufacture’s instruction of PrimeScriptTM RT reagent Kit with gDNA Eraser Perfect Real Time (Takara, Dalian, China). The qRT-PCR reaction (10 µL) was formulated using the TB Green Premix Ex Taq II (Takara, Dalian, China). All qRT-PCRs were carried out on a Piko® Thermal Cycler 96-well system (Thermo Scientific). The experimental methods of qRT-PCR were adapted to a previous study [62]. The average threshold cycle (CT) from three biological replicates was employed to calculate the gene expression fold change by the 2-△△Ct method. Data analysis was conducted using SPSS 20.0 (SPSS Inc., Chicago, IL, USA). The independent samples t-test was used to determine whether there were significant differences in transcript expression of salt treatment group relative to the control group.
5. Conclusions
Cr. chinense is a salt-tolerant wild species and its hybrids inherit this strong tolerance. The comparative transcriptome analyses indicated that the plant hormone signaling transduction and the MAPK signaling pathway were the key biological pathways. Further WGCNA analysis provided more insight into the ABA signaling pathway. The tissue-specific expression patterns of candidate genes showed that the ABA-related genes expressed significantly under salt stress. These candidate genes and pathways lay a foundation for further analyzing the salt-tolerant molecular mechanisms of Cr. chinense and improving the salt tolerance of cultivated chrysanthemums.
Author Contributions
Conceptualization, Y.W. and M.S.; methodology, Y.W. and M.L.; software, Y.W.; validation, Y.W., Z.G. and Y.L (Yufan Lu).; formal analysis, Y.W. and Y.X.; data curation, Y.W. and M.L.; writing-original draft preparation, Y.W.; writing-review and editing, Y.W.; visualization, Y.W. and Y.L. (Yilin Liang); supervision, M.S.; project administration, M.S.; funding acquisition, M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Science, Technology & Innovation Project of Xiongan New Area (2022XAGG0100) and National Natural Science Foundation of China (No. 32271947).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in Supplementary Materials.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, B. Protection of halophytes and their uses for cultivation of saline-alkali soil in China. Biology 2021, 10, 353. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. International Journal of Molecular Sciences 2021, 22, 4609. [Google Scholar] [CrossRef]
- Yu, Q.; An, L.; Li, W. The CBL–CIPK network mediates different signaling pathways in plants. Plant Cell Rep. 2014, 33, 203–214. [Google Scholar] [CrossRef]
- Feng, J.; Wang, L.; Wu, Y.; Luo, Q.; Zhang, Y.; Qiu, D.; Han, J.; Su, P.; Xiong, Z.; Chang, J. TaSnRK2.9, a sucrose non-fermenting 1-related protein kinase gene, positively regulates plant response to drought and salt stress in transgenic tobacco. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Z.; Chen, C.; Xu, M. The MKK2a gene involved in the MAPK signaling cascades enhances Populus salt tolerance. International Journal of Molecular Sciences 2022, 23, 10185. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef]
- Jammes, F.; Song, C.; Shin, D.; Munemasa, S.; Takeda, K.; Gu, D.; Cho, D.; Lee, S.; Giordo, R.; Sritubtim, S. MAP kinases MPK9 and MPK12 are preferentially expressed in guard cells and positively regulate ROS-mediated ABA signaling. Proceedings of the National Academy of Sciences - PNAS 2009, 106, 20520–20525. [Google Scholar] [CrossRef]
- Guan, Z.; Chen, F.; Teng, N.; Chen, S.; Liu, P. Study on the NaCl tolerance in five plant species from Dendrathema and its relatives. Scientia Agricultural Sinica 2010, 43, 787–794. [Google Scholar] [CrossRef]
- Guan, Z.; Chen, S.; Chen, F.; Liu, Z.; Fang, W.; Tang, J. Comparison of stress effect of NaCl, Na+ and Cl- on two Chrysanthemum species. Acta Horticulturae 2012, 937, 369–375. [Google Scholar] [CrossRef]
- Guan, Z.; Su, Y.; Teng, N.; Chen, S.; Sun, H.; Li, C.; Chen, F.; R, M.M.; S, D.Y.; T, T. Morphological, physiological, and structural responses of two species of Artemisia to NaCl Stress. The Scientific World 2013, 2013, 309808–309810. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Jiang, J.; Chen, S.; Teng, N.; Song, A.; Guan, Z.; Fang, W.; Chen, F.; Niedz, R.P. Combination of multiple resistance traits from wild relative species in chrysanthemum via trigeneric hybridization. PLoS One 2012, 7, e44337. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, J.; Chen, S.; Fang, W.; Guan, Z.; Liao, Y.; Chen, F. Rapid genomic and transcriptomic alterations induced by wide hybridization: Chrysanthemum nankingense × Tanacetum vulgare and C. crassum × Crossostephium chinense (Asteraceae). BMC Genomics 2013, 14, 902. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Xu, N.; Sun, Y.; Li, Q.; Yue, L.; Zhou, Y.; He, M. Chrysanthemum × grandiflora leaf and root transcript profiling in response to salinity stress. BMC Plant Biol. 2022, 22, 240. [Google Scholar] [CrossRef]
- Cheng, P.; Gao, J.; Feng, Y.; Zhang, Z.; Liu, Y.; Fang, W.; Chen, S.; Chen, F.; Jiang, J. The chrysanthemum leaf and root transcript profiling in response to salinity stress. Gene 2018, 674, 161–169. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, T.; Wang, K.; Liang, Q.; Bai, Z.; Liu, Q.; Pan, Y.; Jiang, B.; Zhang, L. Comparative analysis of the chrysanthemum leaf transcript profiling in response to salt stress. PLoS One 2016, 11, e159721. [Google Scholar] [CrossRef]
- Wang, K.; Wu, Y.; Tian, X.; Bai, Z.; Liang, Q.; Liu, Q.; Pan, Y.; Zhang, L.; Jiang, B. Overexpression of DgWRKY4 enhances salt tolerance in chrysanthemum seedlings. Front. Plant Sci. 2017, 8, 1592. [Google Scholar] [CrossRef]
- Wang, K.; Zhong, M.; Wu, Y.; Bai, Z.; Liang, Q.; Liu, Q.; Pan, Y.; Zhang, L.; Jiang, B.; Jia, Y. Overexpression of a chrysanthemum transcription factor gene DgNAC1 improves the salinity tolerance in chrysanthemum. Plant Cell Rep. 2017, 36, 571–581. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Liu, C.; Yin, H.; Liu, H.; Luo, H.; He, M.; Zhou, Y. CgbZIP1: A bZIP transcription factor from Chrysanthemum grandiflora confers plant tolerance to salinity and drought stress. Agronomy 2022, 12, 556. [Google Scholar] [CrossRef]
- Zhao, Q.; He, L.; Wang, B.; Liu, Q.; Pan, Y.; Zhang, F.; Jiang, B.; Zhang, L. Overexpression of a multiprotein bridging factor 1 gene DgMBF1 improves the salinity tolerance of chrysanthemum. International Journal of Molecular Sciences 2019, 20, 2453. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Sun, M.; Lin, S.; Guo, Y.; Yang, Y.; Zhang, T.; Zhang, J. Transcriptome analysis of Crossostephium chinensis provides insight into the molecular basis of salinity stress responses. PLoS One 2017, 12, e187124. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Sun, J.; Cao, P.; Ren, L.; Liu, C.; Chen, S.; Chen, F.; Jiang, J. Variation in tissue Na+ content and the activity of SOS1 genes among two species and two related genera of chrysanthemum. BMC Plant Biol. 2016, 16, 98. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Guo, Y.; Chen, J.; Cao, H.; Liu, M.; Guo, Z.; Zhang, Q.; Sun, M. Volatiles inheriting from Crossostephium chinense act as repellent weapons against aphids in Chrysanthemum lavandulifolium cultivars. Ind. Crop. Prod. 2021, 166, 113467. [Google Scholar] [CrossRef]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Liu, Z.; Wang, S.; Huang, B.; Hu, Z.; Liu, Y. Comparative transcriptome analysis of three chrysanthemums provides insights into flavonoid and terpenoid biosynthesis. J. Plant Biol. 2021, 64, 389–401. [Google Scholar] [CrossRef]
- Zhao, Q.; He, L.; Wang, B.; Liu, Q.; Pan, Y.; Zhang, F.; Jiang, B.; Zhang, L.; Liu, G.; Jia, Y. Transcriptome Comparative Analysis of Salt Stress Responsiveness in Chrysanthemum (Dendranthema grandiflorum) Roots by Illumina- and Single-Molecule Real-Time-Based RNA Sequencing. DNA. Cell. Biol. 2018, 37, 1016–1030. [Google Scholar] [CrossRef]
- Gharaghanipor, N.; Arzani, A.; Rahimmalek, M.; Ravash, R. Physiological and Transcriptome Indicators of Salt Tolerance in Wild and Cultivated Barley. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, W.; Yang, J.; Ismail, A.M. Role of ABA in integrating plant responses to drought and salt stresses. Field Crops Res. 2006, 97, 111–119. [Google Scholar] [CrossRef]
- Fujita, Y.; Nakashima, K.; Yoshida, T.; Katagiri, T.; Kidokoro, S.; Kanamori, N.; Umezawa, T.; Fujita, M.; Maruyama, K.; Ishiyama, K. Three SnRK2 Protein Kinases are the Main Positive Regulators of Abscisic Acid Signaling in Response to Water Stress in Arabidopsis. Plant Cell Physiol. 2009, 50, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Li, Y.; Wang, Y.; Liu, X.; Ma, L.; Zhang, Z.; Mu, C.; Zhang, Y.; Peng, L.; Xie, S. Initiation and amplification of SnRK2 activation in abscisic acid signaling. Nat. Commun. 2021, 12, 2456. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C Phosphatase Activity Function as Abscisic Acid Sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef]
- Nguyen, Q.; Lee, S.J.; Choi, S.W.; Na, Y.J.; Song, M.R.; Hoang, Q.; Sim, S.Y.; Kim, M.S.; Kim, J.I.; Soh, M.S. Arabidopsis Raf-Like Kinase Raf10 Is a Regulatory Component of Core ABA Signaling. Mol. Cells 2019, 42, 646–660. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; He, Z.; Zhao, X.; Wang, Q.; Zhou, B.; Yu, D.; Huang, X.; Tang, D.; Guo, X. Molecular character of a phosphatase 2C (PP2C) gene relation to stress tolerance in Arabidopsis thaliana. Mol. Biol. Rep. 2013, 40, 2633–2644. [Google Scholar] [CrossRef]
- Krzywinska, E.; Kulik, A.; Bucholc, M.; Fernandez, M.A.; Rodriguez, P.L.; Dobrowolska, G. Protein phosphatase type 2C PP2CA together with ABI1 inhibits SnRK2.4 activity and regulates plant responses to salinity. Plant Signal. Behav. 2016, 11, e1253647. [Google Scholar] [CrossRef]
- Wang, X.; Guo, C.; Peng, J.; Li, C.; Wan, F.; Zhang, S.; Zhou, Y.; Yan, Y.; Qi, L.; Sun, K. ABRE-BINDING FACTORS play a role in the feedback regulation of ABA signaling by mediating rapid ABA induction of ABA co-receptor genes. New Phytol. 2019, 221, 341–355. [Google Scholar] [CrossRef]
- Pei, Z.; Murata, Y.; Benning, G.; Thomine, S.; Klusener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signalling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef]
- Luan, S.; Wang, C. Calcium Signaling Mechanisms Across Kingdoms. Annu. Rev. Cell Dev.Biol. 2021, 37, 311–340. [Google Scholar] [CrossRef]
- Brandt, B.; Brodsky, D.E.; Xue, S.; Negi, J.; Iba, K.; Kangasjärvi, J.; Ghassemian, M.; Stephan, A.B.; Hu, H.; Schroeder, J.I. Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action. Proceedings of the National Academy of Sciences 2012, 109, 10593–10598. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Sun, P.; Kong, W.; Xie, Z.; Li, C.; Liu, J.H. SnRK2.4-mediated phosphorylation of ABF2 regulates ARGININE DECARBOXYLASE expression and putrescine accumulation under drought stress. New Phytol. 2023, 238, 216–236. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Que, F.; Li, T.; Zhang, R.; Khadr, A.; Xu, Z.; Tian, Y.; Xiong, A. DcABF3, an ABF transcription factor from carrot, alters stomatal density and reduces ABA sensitivity in transgenic Arabidopsis. Plant Sci. 2021, 302, 110699. [Google Scholar] [CrossRef] [PubMed]
- Kalladan, R.R.; Lasky, J.R.J.R.; Sharma, S.S.; Kumar, M.N.M.N.; Juenger, T.E.T.E.; Des Marais, D.L.D.L.; Verslues, P.E.P.E. Natural Variation in 9-Cis-Epoxycartenoid Dioxygenase 3 and ABA Accumulation1[OPEN]. Plant physiology (Bethesda) 2019, 179, 1620–1631. [Google Scholar] [CrossRef]
- Finkelstein, R. Abscisic Acid Synthesis and Response. The Arabidopsis Book 2013, 11, e0166. [Google Scholar] [CrossRef]
- Barrero, J.M.; Rodriguez, P.L.; Quesada, V.; Piqueras, P.; Ponce, M.R.; Micol, J.L. Both abscisic acid (ABA)-dependent and ABA-independent pathways govern the induction of NCED3, AAO3 and ABA1 in response to salt stress. Plant, Cell and Environment 2006, 29, 2000–2008. [Google Scholar] [CrossRef]
- Tan, B.; Joseph, L.M.; Deng, W.; Liu, L.; Li, Q.; Cline, K.; McCarty, D.R. Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. The Plant Journal 2003, 35, 44–56. [Google Scholar] [CrossRef]
- Haak, D.C.; Fukao, T.; Grene, R.; Hua, Z.; Ivanov, R.; Perrella, G.; Li, S. Multilevel Regulation of Abiotic Stress Responses in Plants. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Kumar, K.; Raina, S.K.; Sultan, S.M. Arabidopsis MAPK signaling pathways and their cross talks in abiotic stress response. J. Plant Biochem. Biotechnol. 2020, 29, 700–714. [Google Scholar] [CrossRef]
- Friso, G.; van Wijk, K.J. Update: Post-translational protein modifications in plant metabolism. Plant Physiol. 2015, 169, 1378–2015. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef] [PubMed]
- de Zelicourt, A.; Colcombet, J.; Hirt, H. The Role of MAPK Modules and ABA during Abiotic Stress Signaling. Trends Plant Sci. 2016, 21, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Danquah, A.; de Zélicourt, A.; Boudsocq, M.; Neubauer, J.; Frei Dit Frey, N.; Leonhardt, N.; Pateyron, S.; Gwinner, F.; Tamby, J.; Ortiz-Masia, D. Identification and characterization of an ABA-activated MAP kinase cascade inArabidopsis thaliana. The Plant Journal 2015, 82, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Sugiyama, N.; Takahashi, F.; Anderson, J.C.; Ishihama, Y.; Peck, S.C.; Shinozaki, K. Genetics and phosphoproteomics reveal a protein phosphorylation network in the abscisic acid signaling pathway in Arabidopsis thaliana. Sci. Signal. 2013, 6, rs8. [Google Scholar] [CrossRef]
- Yu, L.; Nie, J.; Cao, C.; Jin, Y.; Yan, M.; Wang, F.; Liu, J.; Xiao, Y.; Liang, Y.; Zhang, W. Phosphatidic acid mediates salt stress response by regulation of MPK6 in Arabidopsis thaliana. New Phytol. 2010, 188, 762–773. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet Journal. 2011, 17. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using diamond. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Zhong, J.; Wang, Y.; Lu, Y.; Ma, X.; Zhang, Q.; Wang, X.; Zhang, Q.; Sun, M. Identification and xpression Analysis of Chemosensory Genes in the Antennal Transcriptome of Chrysanthemum Aphid Macrosiphoniella sanborni. Insects 2022, 13, 597. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The effect of 700 mM NaCl exposure on the growth and enzymatic antioxidant activities of Cr. chinense (CC), C. lavandulifolium (CL) and three hybrids (CE, CT and CF): (A) phenotypic differences, (B) SOD, (C) MDA. Data are means of three replicates and follow normal distribution.
Figure 1.
The effect of 700 mM NaCl exposure on the growth and enzymatic antioxidant activities of Cr. chinense (CC), C. lavandulifolium (CL) and three hybrids (CE, CT and CF): (A) phenotypic differences, (B) SOD, (C) MDA. Data are means of three replicates and follow normal distribution.
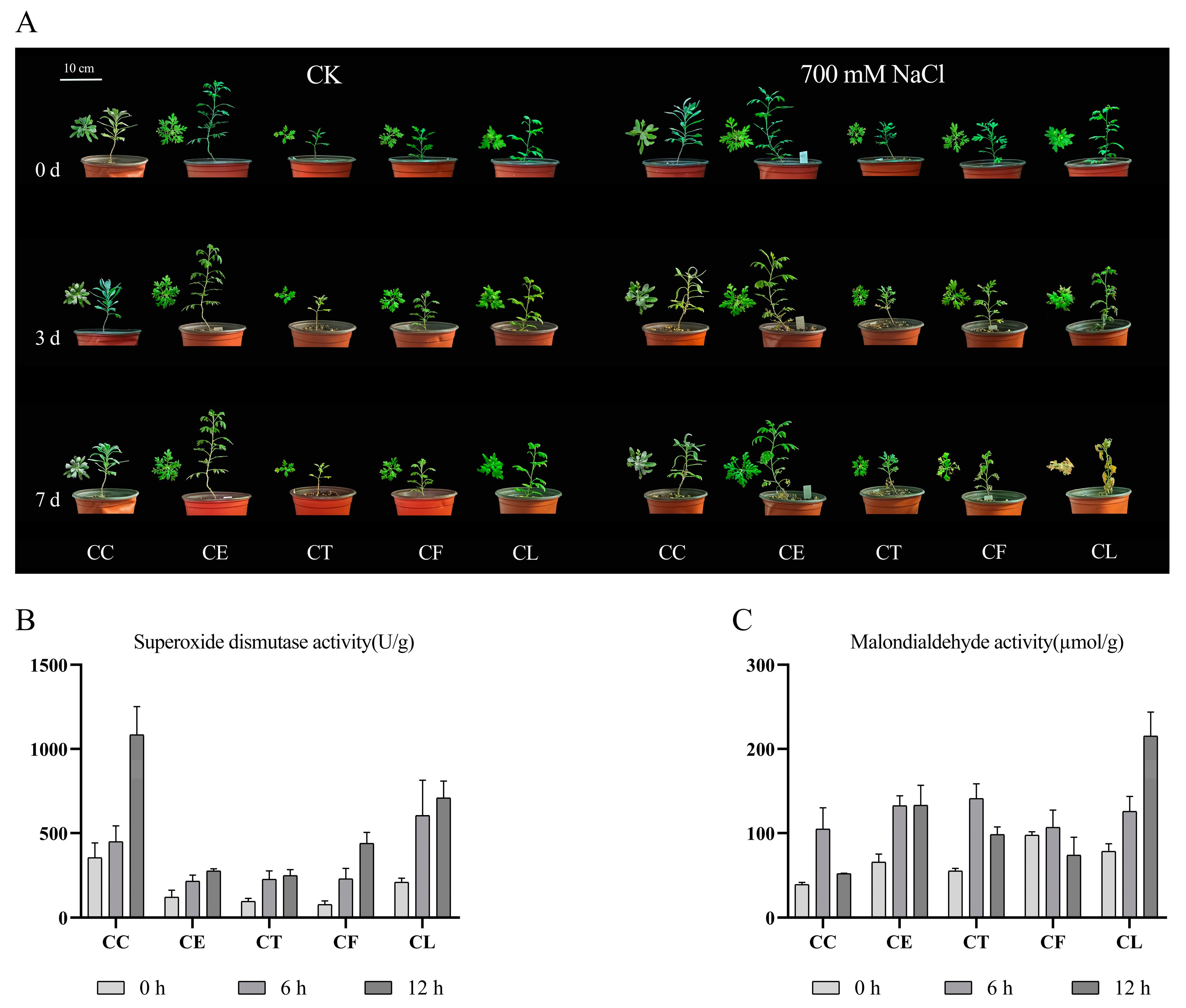
Figure 2.
Sequencing data and differentially expressed genes (DEGs) results: (A) length distribution of genes, (B) FPKM box diagram of all tested samples.
Figure 2.
Sequencing data and differentially expressed genes (DEGs) results: (A) length distribution of genes, (B) FPKM box diagram of all tested samples.
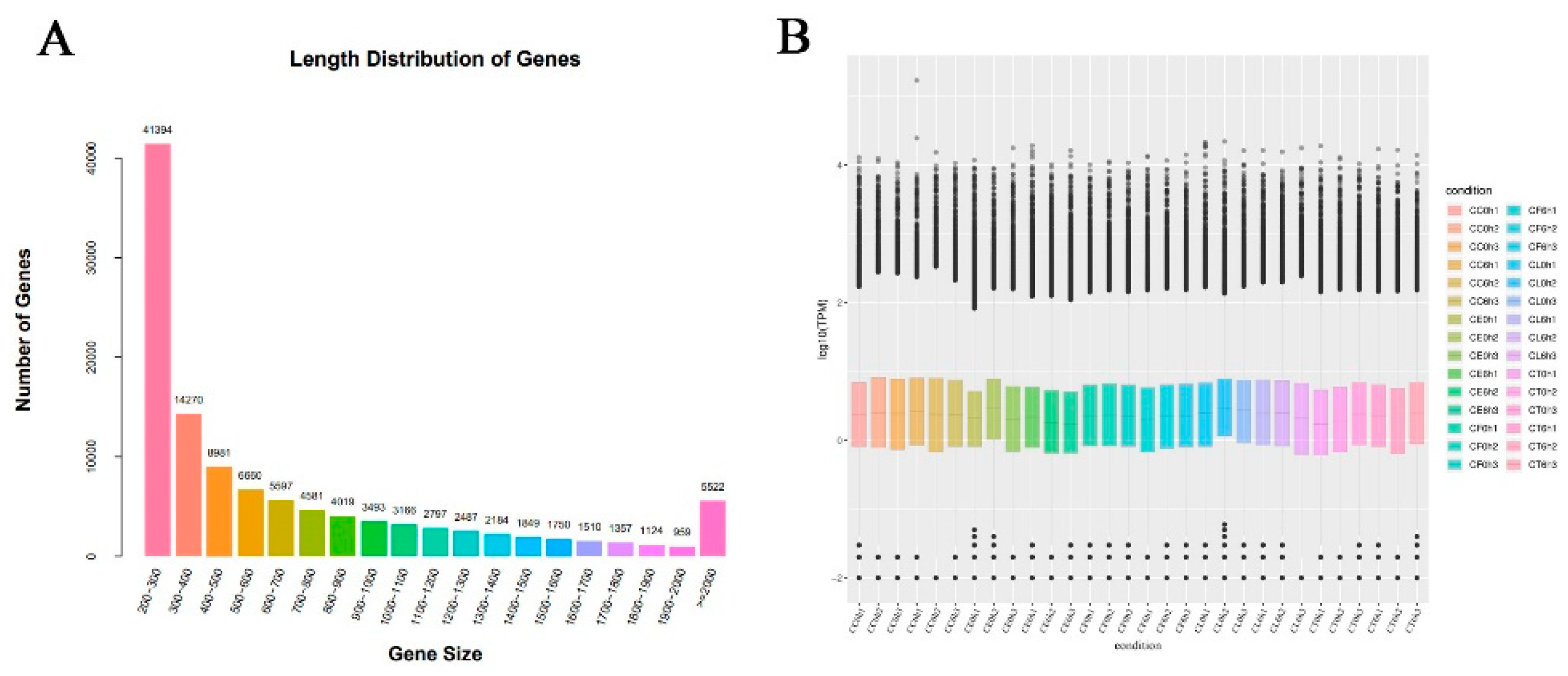
Figure 3.
The number of up-regulated and down-regulated DEGs in all comparison combinations.
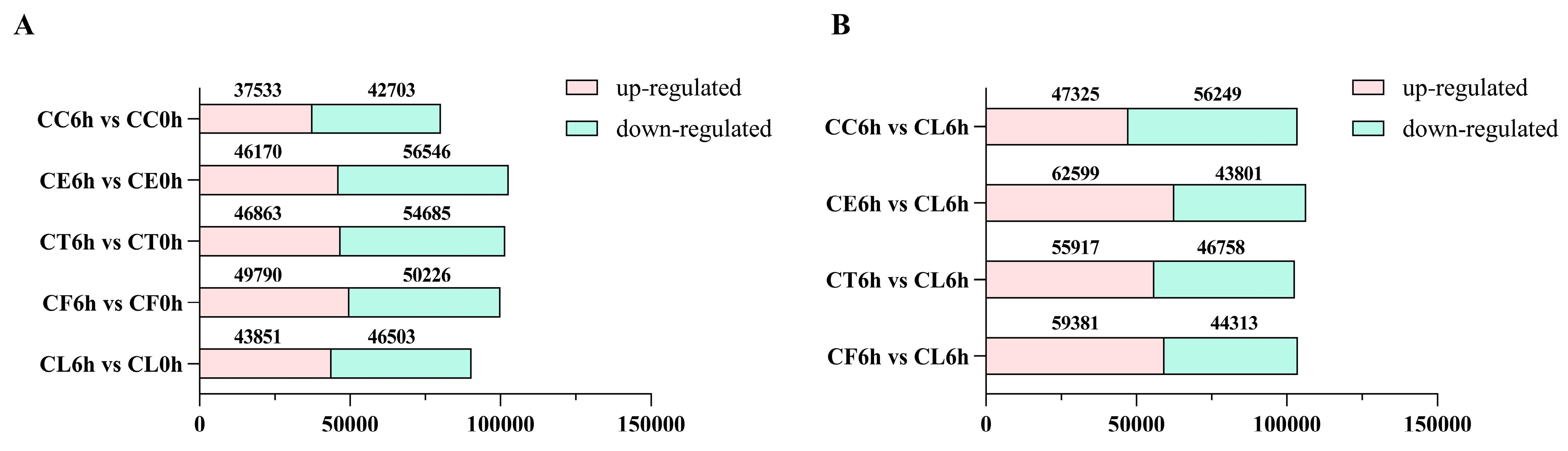
Figure 4.
Enrichment analysis of the GO annotation of DEGs in Cr.chinense, C.lavandulifolium and hybrids: (A) CC6h_vs_CL6h, (B) CE6h_vs_CL6h, (C) CT6h_vs_CL6h, (D) CF6h_vs_CL6h.
Figure 4.
Enrichment analysis of the GO annotation of DEGs in Cr.chinense, C.lavandulifolium and hybrids: (A) CC6h_vs_CL6h, (B) CE6h_vs_CL6h, (C) CT6h_vs_CL6h, (D) CF6h_vs_CL6h.
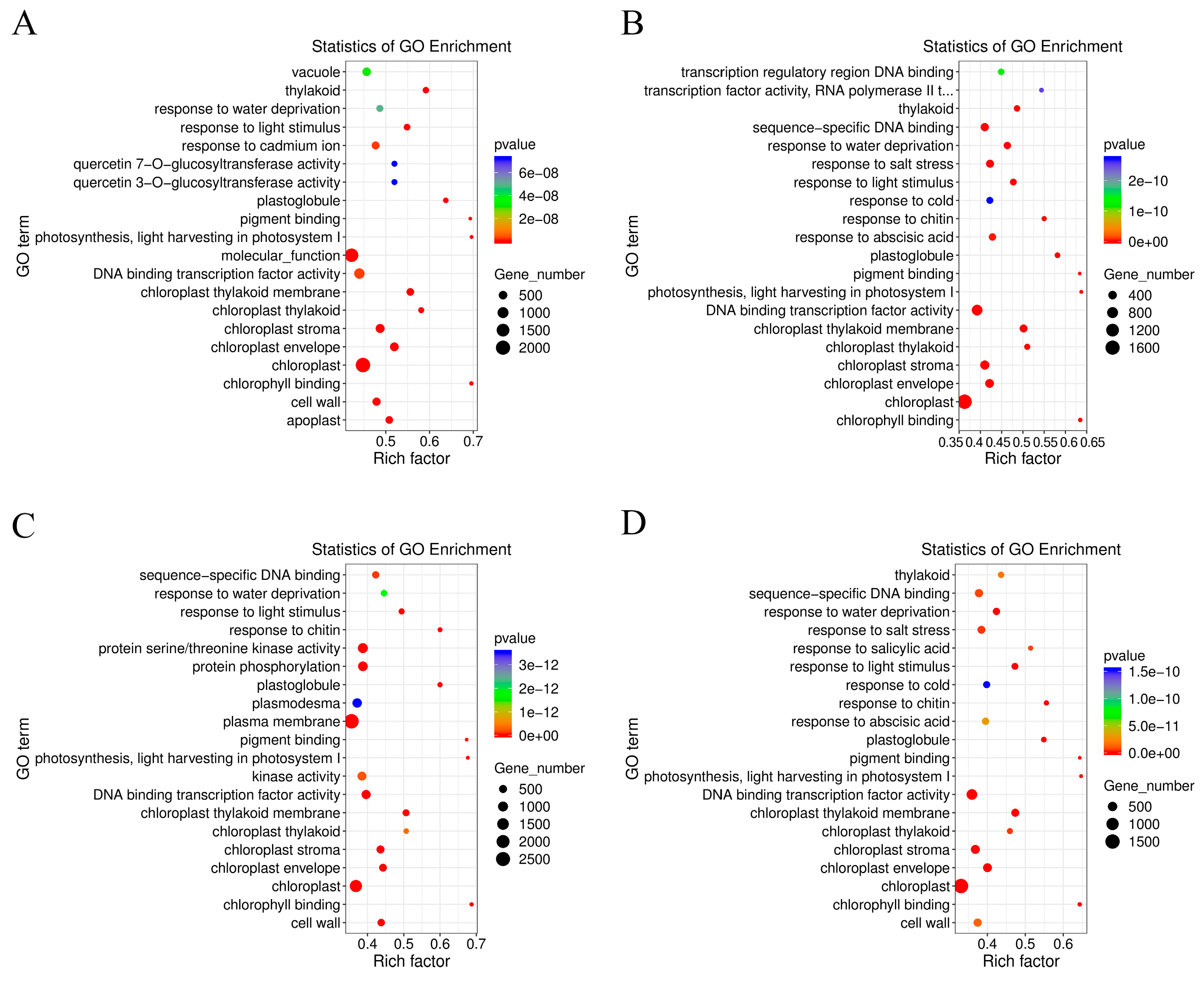
Figure 5.
Enrichment analysis of the KEGG pathway of DEGs in Cr.chinense, C.lavandulifolium and hybrids: (A) CC6h_vs_CL6h, (B) CE6h_vs_CL6h, (C) CT6h_vs_CL6h, (D) CF6h_vs_CL6h.
Figure 5.
Enrichment analysis of the KEGG pathway of DEGs in Cr.chinense, C.lavandulifolium and hybrids: (A) CC6h_vs_CL6h, (B) CE6h_vs_CL6h, (C) CT6h_vs_CL6h, (D) CF6h_vs_CL6h.
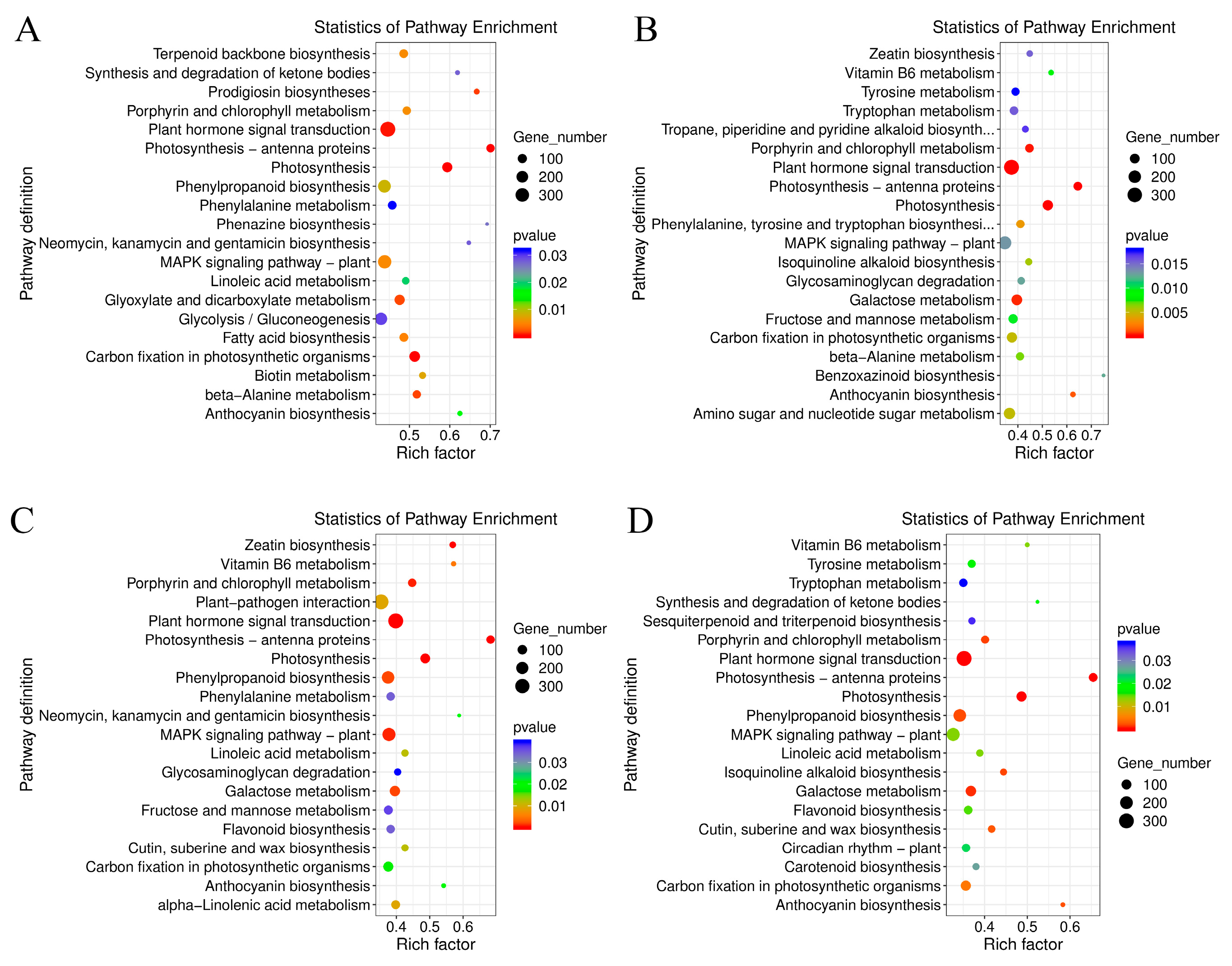
Figure 6.
Gene cluster dendrograms and module division.
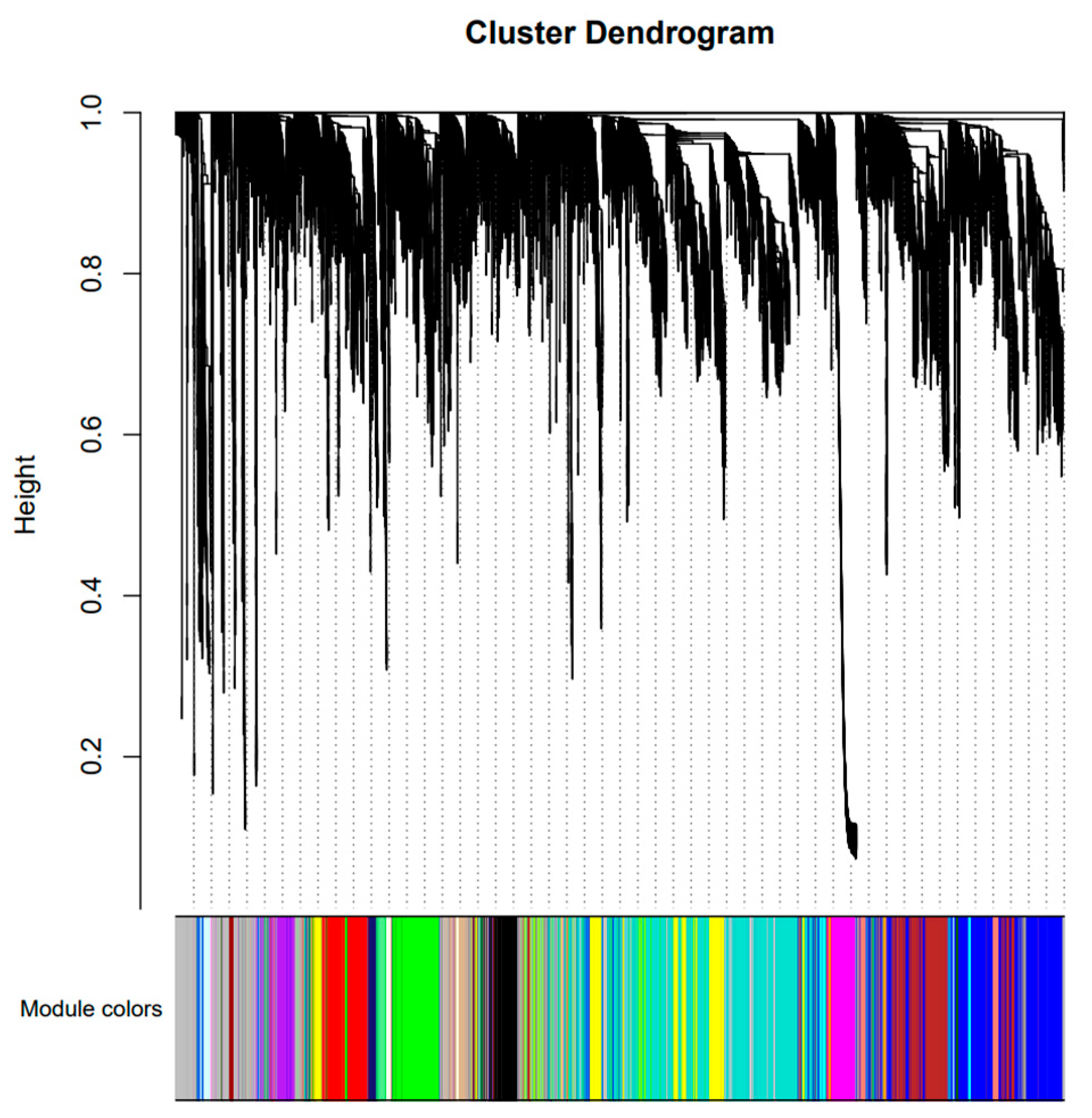
Figure 7.
Sample expression heatmap of co-expression modules. (A) the color in the heatmap indicates the characteristic value of the module, red represents high expression, and blue represents low expression, (B) the above figure is the expression heatmap of genes in the module in different samples, and the figure below shows the eigenvalues of modules in different samples, red indicates upregulation and green indicates downregulation.
Figure 7.
Sample expression heatmap of co-expression modules. (A) the color in the heatmap indicates the characteristic value of the module, red represents high expression, and blue represents low expression, (B) the above figure is the expression heatmap of genes in the module in different samples, and the figure below shows the eigenvalues of modules in different samples, red indicates upregulation and green indicates downregulation.
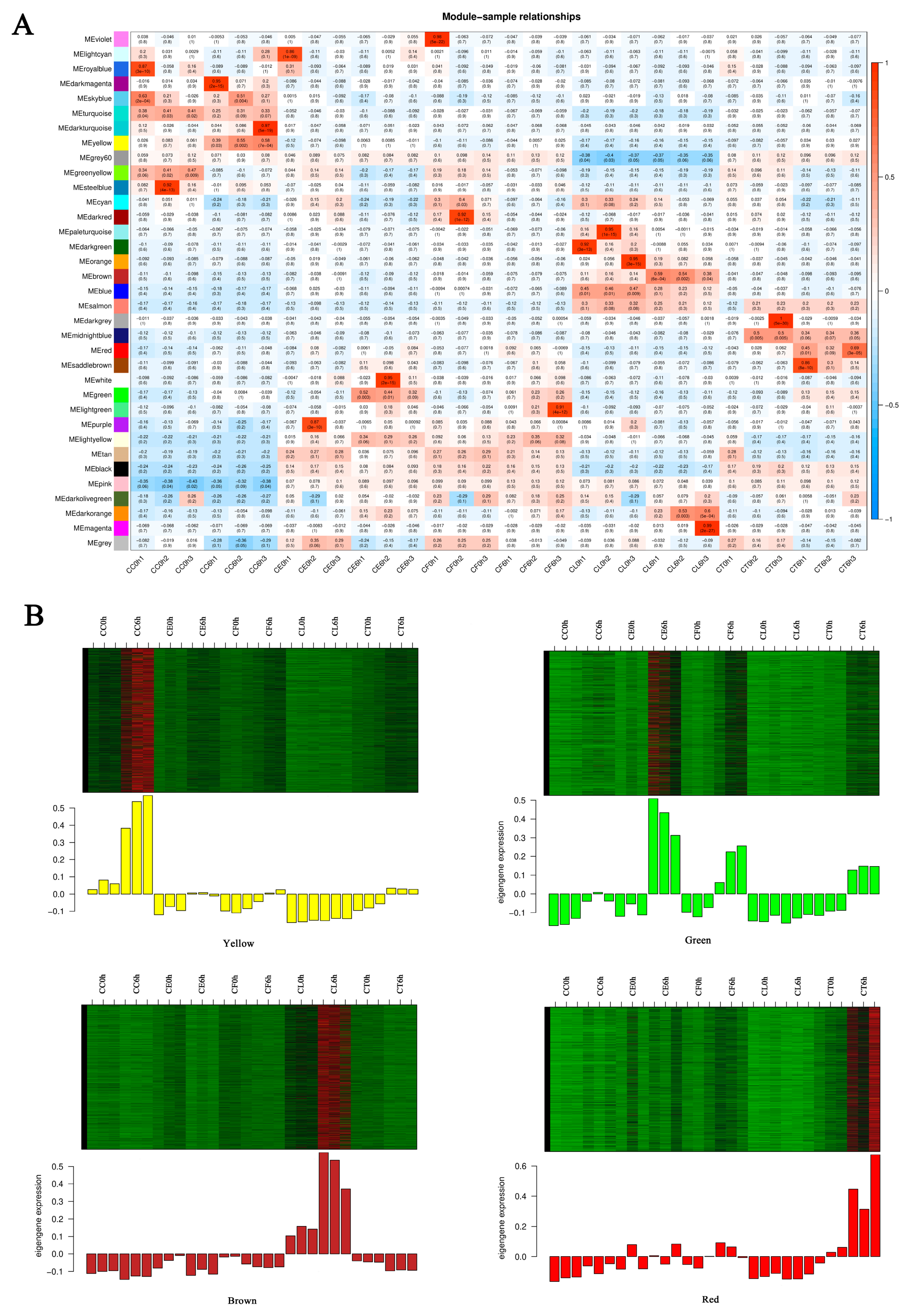
Figure 8.
Regulatory network diagram of hub genes and associated genes in key modules. Purple represents hub genes, green represents the top 20 genes associated with hub genes.
Figure 8.
Regulatory network diagram of hub genes and associated genes in key modules. Purple represents hub genes, green represents the top 20 genes associated with hub genes.
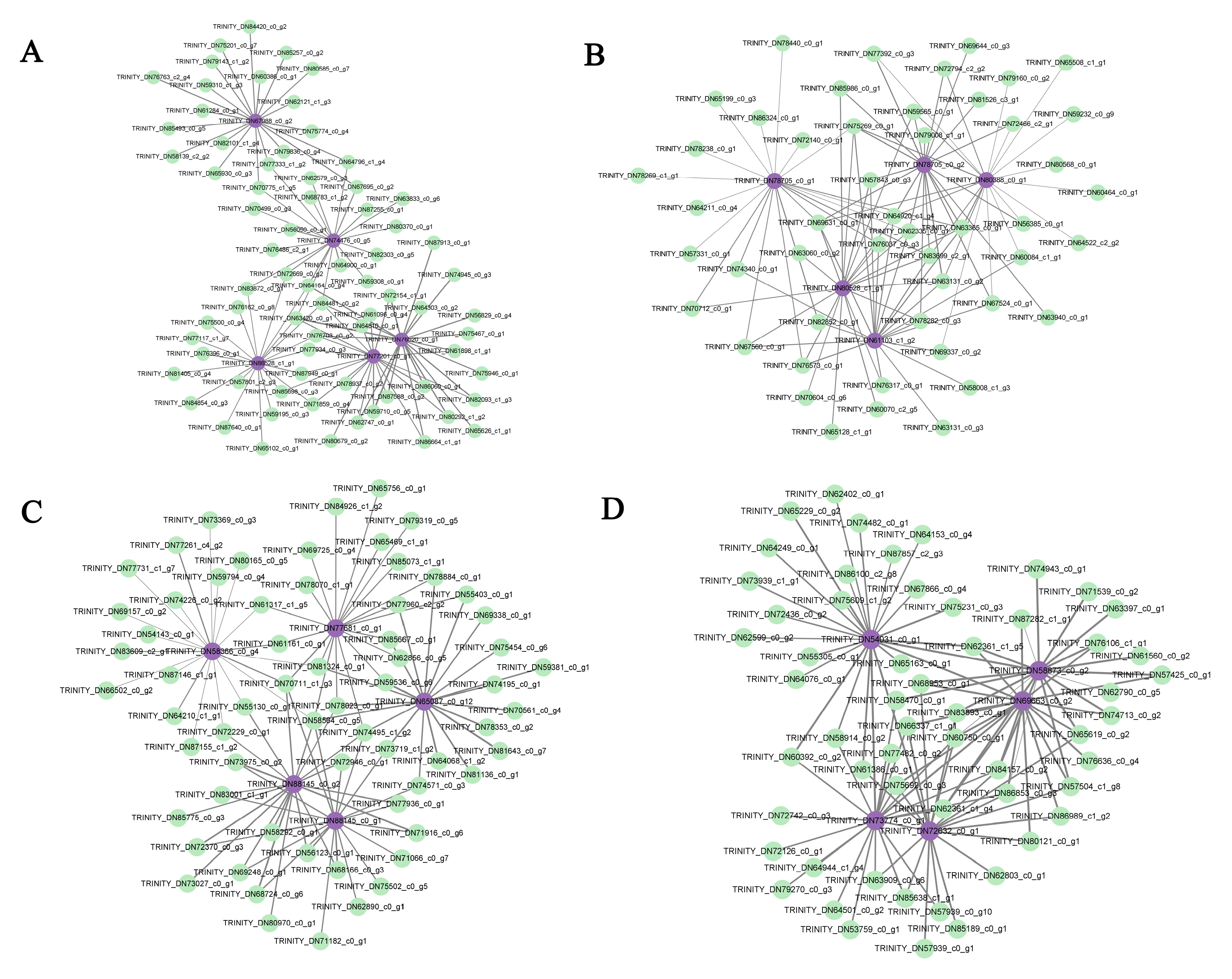
Figure 9.
The relative expression levels of hub genes under salt treatment in different plant tissues. Data are means of three replicates, ** (P<0.01) represents highly significant differences and * (P<0.05) represents significant differences based on the independent samples t-test.
Figure 9.
The relative expression levels of hub genes under salt treatment in different plant tissues. Data are means of three replicates, ** (P<0.01) represents highly significant differences and * (P<0.05) represents significant differences based on the independent samples t-test.
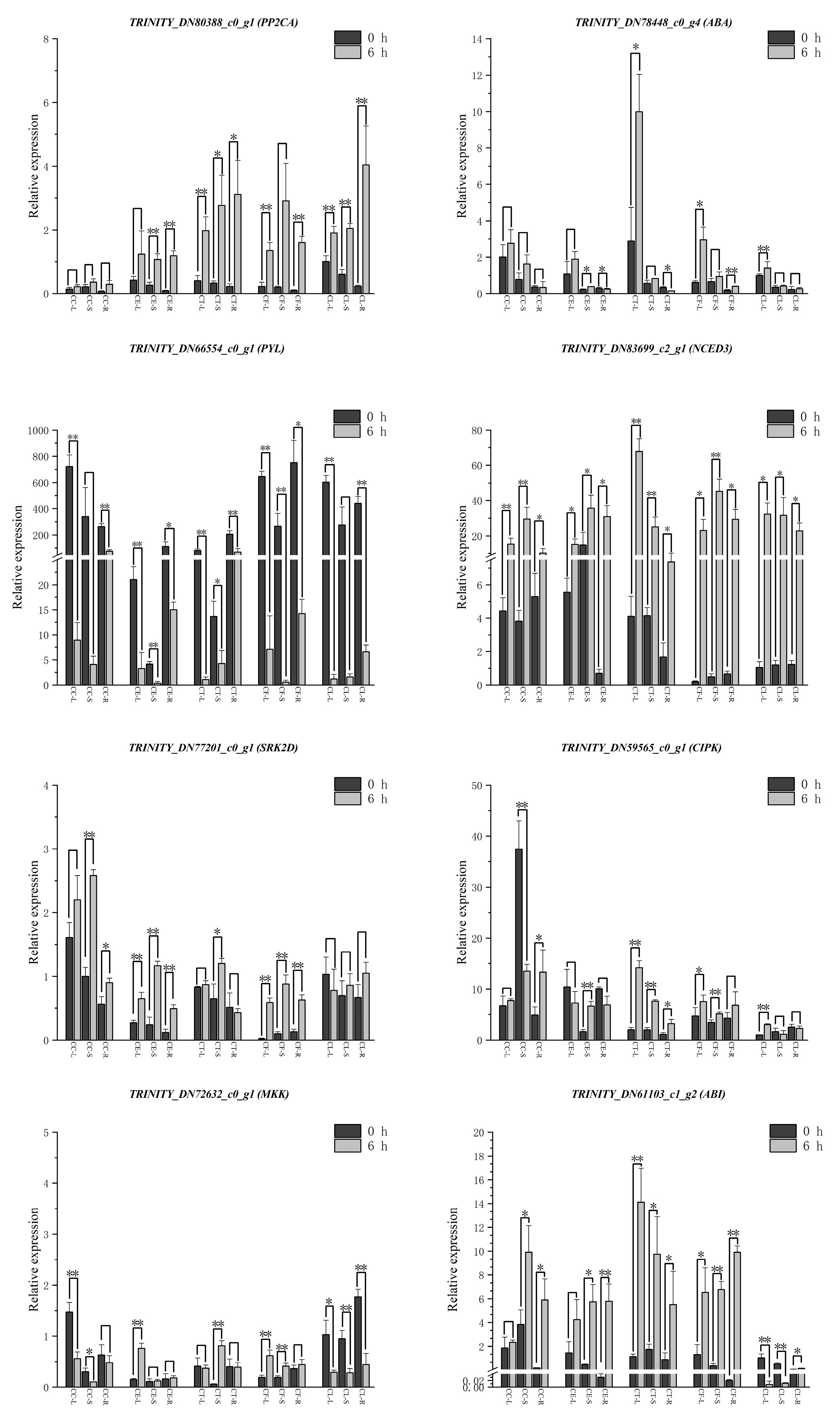
Figure 10.
Molecular mechanism diagram of salt stress resistance of Cr. Chinense.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



