
Submitted:
03 October 2023
Posted:
04 October 2023
You are already at the latest version
Abstract
We have used Daphnia cucullata as a model organism for the first time in the four deepest Latvian lakes from the Boreal biogeographical region lakes in order to find the genetic diversity of Daphnia cucullata populations. During the research, the most appropriate microsatellite markers for future genetic studies of Daphnia cucullata populations in Boreal biogeographical region lakes. It was the loci Dgm105 and Dgm101, in which the maximum number of alleles and the maximum number of private alleles were found. The Dgm105 locus had five private alleles (62% of all detected alleles), while the Dgm101 locus had four private alleles (57% of all detected alleles) in these loci. We was determined the observed heterozygosity (Hobs) and the expected heterozygosity (Hexp) level (by Hardy-Weinberg), the number of polymorphic loci, the number of detected alleles in each analyzed microsatellite locus, the average number of alleles at the locus (Na), the average effective number of alleles at the locus (Ne), FST of the population genetic differentiation, the genetic distance (D) (by Nei) and significance (χ2- test) of differences between the levels of observed and expected heterozygosity.
It was shown that Daphnia cucullata populations from lakes with a low number of zooplankton taxa (Riča and Geraņimovas-Ilzas) have a higher genetic diversity compared to lakes with a high number of zooplankton taxa (Dridzis and Svente) in lakes. It was found, that Daphnia cucullata populations from lakes Dridzis and Svente have the least genetic distance and these populations form a single genetic group, as confirmed by clustering
Keywords:
Cladocera
; Daphnia cucullata
; genetic diversity
; microsatellite-PCR
; Boreal biogeographical region lakes
1. Introduction
Zooplankton (e.g. Cladocera) plays an essential role in the transformation of substances and energy in water bodies and are an important stage in the food chain. They regulate bacterial and detrital quantity and they are an important component of the feed for juvenile fish, plankton-feeding fish and many other aquatic animals [1,2,3,4]. Some Cladocera species such as Daphnia magna, Daphnia pulex, Daphnia cucullata are often used as bioindicators of water pollution and a good model organism of freshwater ecology [5,6,7,8,9,10]. Daphnia has been considered to be a control organism in the freshwater as a kind of convergence model with the adaptive features in radically different habitats [11,12]. For understanding the adaptive processes in Daphnia populations under the changing environmental conditions the investigations of genetic structure of population are necessary [13,14,15,16,17].
In lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas Daphnia cucullata species are one of dominating species in the species composition [18,19,20,21,22,23].
Microsatellite markers are useful for population genetics studies because they typically have high allelic variation within and between populations, thus increasing the probability of distinguishing between populations and detecting changes over time [24]. Also, microsatellites are neutral markers (in the non-coding regions of the genome) and do not affect the fitness of the organism. This provides an opportunity to understand gene flow and to analyze the degree of connection among contemporary of Daphnia cucullata populations [25] or of Daphnia cucullata populations through time from different waterbodies [26,27,28,29]. Microsatellite analysis method can be used to determine the genetic variability and genetic monitoring of populations of Cladoceran different species specimen as too [13,26,30,31,32,33]. Microsatellite loci are frequently used in studies of the genetic structure of Daphnia genus different species (Daphnia pulex, Daphnia magna, Daphnia longispina, Daphnia galeata, Daphnia hyalina, Daphnia rosea, Daphnia curvirostris) [13,26,29,30,34,35,36,37,38], there are relatively few studies investigating the genetic structure of the Daphnia cucullata species [13].
Therefore, the aim of this study at first time was determined the genetic structure of Daphnia cucullata native population in four deepest Latvian lakes from Boreal biogeographical region lakes by microsatellite markers.
2. Material and methods
2.1. Sampling sites and material collection of Daphnia cucullata
Material for the study of genetic diversity in Daphnia cucullata population was collected in four deep lakes of Eastern Latvia, which belong to deep, well-transparent mesotrophic and mesoeutrophic Latvian lakes and belongs Daugava River basin. Lakes Dridzis, Svente and Riča are mesotrophic, but Lake Geraņimovas-Ilzas is mesoeutrophic [39]. Samples were taken in the deepest places in each lake, namely Dridzis, Svente, Riča and Geraņimovas-Ilzas.
This lakes are relatively similar in terms of their morphometric characteristics. Lake Dridzis is the deepest Lake in Latvia. Moreover, it is the deepest Lake in Baltia [40]. Lake Geraņimovas-Ilzas is the fifth deepest, but Lake Svente is the tenth deepest Lake in Latvia [40,41,42]. In turn, Lake Riča is the ninth deepest Lake in Latvia [43]. Investigated lakes characteristics and location are presented in Figure 1 and Table 1. Geographical data of collected material localities were obtained by echo sounders with GPS receiver LOWRANCE LMS-522C and TRIMBLE Juno SB.
Zooplankton samples were taken with Hydro-bios Apstein type plankton net with an opening-closing mechanism (mesh size 64 µm) to filter the water column, which was taken from the deep water to the surface. The collection of the zooplankton samples was performed using the APHA standard methods procedure [44,45]. The collected water sample material was preserved immediately after collecting by adding 98% ethanol to water sample hence the final concentration in the sample is ± 70%. Samples tissues were stored at -200C. After the splitting collected material into the species, the species resulting material was stored in 96% ethanol. The samples had to be preserved immediately after harvesting to prevent individuals from biochemical and molecular degradation [16].
2.2. Genetic analysis
2.2.1. DNA extraction
Genomic DNA extraction from adult Daphnia cucullata individuals (20–30 specimens from each population) was performed using slightly modified ‘salting out’ extraction methodology earlier described by Fitzsimmons and Innes [46]. The method consists of the following steps: zooplankton samples were transferred to 1.5 ml reaction tubes, containing 100 μl of buffer A (100 mM Tris–HCl (pH 7.5), 100 mM ethylenediaminetetraacetic acid (EDTA), 100 mM NaCl and 0.5% SDS) was added. Tubes were incubated at 700C for 35 min. Two hundred microliters of LiCl–KAc solution (one part 5 M KAc by volume with 2.5 parts 6 M LiCl) was added before tubes were incubated on ice for 15–20 min. Samples were spun at 13 700 g for 15 min. Supernatant was transferred into new tubes. One hundred and sixty microliters of cold (-20 0C) isopropanol was added, and the sample was mixed and then spun for 15 min. We aspirated away the supernatant by vacuum, spun, and then aspirated the remaining liquid. Samples were washed twice with cold (40C) 70% ethanol, being spun for 2 min before supernatant was aspirated away each time. DNA was resuspended in 35 μl of double-distilled water and left at 4 0C overnight [46,47].
2.2.2. Determination of the quantity and quality of isolated DNA
The concentration of DNA samples and quantity, quality and suitability for PCR were determined using spectrophotometer BioSpec- Nano (Shimadzu, Japan). The dry DNA samples were dissolved in dd H2O for quantifying DNA. The ratio of absorbance at 260 and 280 nm (A260/280>1.8) and A260/230 were used to assess the purity of nucleic acids. The quality and suitability of the isolated DNA samples for PCR were checked on 1.5% agarose gel [16,47] with ethidium bromide.
2.2.3. Microsatellites analysis
We used nine primers for microsatellite loci of nuclear DNA (DaB10/15; Dp512; Dp519; DaB17/16; DaB17/17; SwiD1; Dgm101; Dgm105; Dgm109) for the Latvian Daphnia cucullata population genetic research, but six microsatellite primers with good representativity (SwiD1; Dgm105; Dgm101; DaB17/17; Dgm109; Dp519) were selected for the analysis. There were three of them were dinucleotide microsatellite primers (SwiD1; Dgm101; Dp519) and two trinucleotide microsatellite primers (Dgm105; Dgm109).
Microsatellite amplification was performed using Eppendorf Mastercycler® pro (Eppendorf) automated polymerase chain reaction (PCR) system. PCR was performed in 10 µl for 0.2 ml PCR tubes. PCR mixture components were 2.3 µl of genomic DNA sample (20 ng) in Dilution buffer (Thermo Scientific), 5 µl 2x Phire Animal Tissue PCR Buffer (with dNTPs and MgCl2) (Thermo Scientific); 0.2 µl - Phire Hot Start II DNA polimerase (Thermo Scientific); 1.25 µl–8 µmol/ µl primers F; 1.25 µl–8 µmol/ µl primers R. Primers were obtained with fluorescently-4 labeled TMR, HEX, FAM. PCR were performed using the thermal cycling programme, following amplification cycle: denaturation 98 °C 5 min, 40 cycles: 98 °C 5 s (denaturation), X °C or 55 °C 10 s (solicitation or primer annealing), 72 °C 20 s (synthesis), 72 °C 1 min, 4 °C (cooling). Amplification was repeated three times with each primer, including a positive and negative control. After the PCR amplification the products were maintained at 4°C until the analytical separation using automated sequencer GeneScan®Analysis ABI PRISM 3100 (Applied Biosystems) as international size standard.
Based on obtained results for future genetic diversity study of Daphnia cucullata populations were selected only primers that provide high levels of good amplifications and informative DNA fragments.
2.2.4. Statistical processing and analysis of the obtained data
The obtained data were processed and analysed using computer softwares GeneAlex 6.41 [48] and POPGENE 1.32 [49]. Allele number per locus, frequency, private alleles in each population (Nei, 1987), observed (Hobs) and expected (Hexp) heterozygosity level in polymorphic loci (according to Hardy-Weinberg) [50] were measured, and their differences among Daphnia cucullata individuals from different sampling places were calculated using and significance with χ2 criteria were calculated using GeneAlex 6.41 and POPGENE 1.32. The genetic relatedness of Daphnia cucullata populations was estimated with genetic distance (D) [51]. Genetic differentation among the Daphnia cucullata populations was estimated by principal component analysis (PCA) and pairwise FST values [52]. To estimate and visualise the genetic structure and differentation of the studied Daphnia cucullata populations STRUCTURE 2.3.4 [53] and STRUCTURE HARVESTER [54] were used.
3. Results
The size of the scored polymorphic DNA fragments ranged from 122 bp to 303 bp (Table 2). The highest number of base pairs were found in loci Dgm109 (250–303 bp) and Dgm105 (165-240 bp), but the lowest in loci DaB17/17 (100–106 bp) and SwiD1 (122-127 bp) (Table 2).
According to the results obtained in the study, it can be seen (Table 3) that the average level of polymorphism in all studied Daphnia cucullata populations was the same and amounted to 100%, because all six analyzed microsatellite loci were polymorphic in all studied Daphnia cucullata populations. Number of polymorphic loci of Daphnia cucullata population in the lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas ranged from 33% to 100%. The lowest number of polymorphic microsatellite loci of Daphnia cucullata population was found in the Lake Dridzis (33%), while the highest number of polymorphic microsatellite loci was found in the lakes Riča (100%) and Geraņimovas-Ilzas (83%) (Figure 2).
- * GERANIM- Lake Geraņimovas-Ilzas
Analyzing the general parameters of the abundance of obtained alleles (Table 3), it can be seen that the detected number of alleles in a investigated locus differs in each population. Also, the number of detected alleles in each analyzed microsatellite locus was different. The maximum number of alleles was found in the loci Dgm105 (eight alleles) and Dgm101 (seven alleles), moreover, it should be noted that the maximum number of private alleles was also found in these loci, where the Dgm105 locus had five private alleles (62% of the all detected alleles), while the Dgm101 locus had four private alleles (57% of all detected alleles) in these loci (Table 3). On the other hand, the lowest number of alleles was found in locus Dp519 (two alleles). It is also characteristic that private allele was not detected in this locus at all (Table 3).
Analyzing the occurrence of alleles in the investigated Daphnia cucullata populations (Figure 3) shows that the alleles abundance were different, but this difference is not statistically significant (p> 0.05). The largest number of detected alleles per locus were for the Daphnia cucullata population of Lake Geraņimovas-Ilzas (3.17), followed by the Daphnia cucullata population of Lake Riča (3.00). The number of detected alleles per locus is relatively lower for the Daphnia cucullata populations of lakes Dridzis (1.67) and Svente (1.83) (Figure 3).
The average number of alleles per locus with a frequency of more than 5% is equal to the average number of alleles per locus in all studied Daphnia cucullata populations (Figure 3). The average number of rare alleles per loci of the Daphnia cucullata specimens under research, which found to be less than 50%, is the same for populations of lakes Geraņimovas-Ilzas and Riča and respectively it is 1, while for the Daphnia cucullata populations of lakes Dridzis and Svente and respectively it is 0.5, but overall these differences are not significant (p> 0.05) (Figure 3).
The number of average effective alleles per locus differs significantly from the average observed number of alleles in Daphnia cucullata populations of lakes Riča and Svente (p< 0.05), while these differences are not significant for Daphnia cucullata populations of lakes Dridzis and Geraņimovas-Ilzas (p> 0.05) (Figure 3).
The average level of the observed heterozygosity (Ho) was highh in all studied Daphnia cucullata populations, ranging from 1.67 to 3.17. The minimum value of Ho was 1.67 in Lake Dridzis, and the maximum was 3.17 in Lake Geraņimovas-Ilzas. While the average level of the expected heterozygosity (He) ranged from 1.15 to 2.43. The minimum value of He was 1.15 in Lake Svente, and the maximum was 2.43 in Lake Geraņimovas-Ilzas. In general, in all Daphnia cucullata populations under research the average observed level and the average expected level of heterozygosity (according to Hardy-Weinberg) was different, but these differences were insignificant (p <0.001) (Figure 3).
We investigated the polymorphism of microsatellites lake sites to examine the variability of microsatellites in Lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas, which are deep lakes of Eastern Latvia. The analysed loci (SwiD1; Dgm105; Dgm101; DaB17/17; Dgm109 and Dp519) were polymorphic in the investigated Daphnia cucullata populations and the level of polymorphism was very high. Genetic diversity across the studied Daphnia cucullata samples found in each studied loci and each each location are presented in Table 4. The greatest number of alleles (19) were found at loci DaB17/17 and Dp519 in the population of Lake Riča, and the minimum number of alleles (1) at locus Dgm101 was found in the population of Lake Dridzis. It should be noted that no alleles were not detected at loci Dgm101 and Dgm109 in the population of Lake Svente. Private alleles were found at the loci SwiD1, Dgm105, Dgm101, DaB17/17 and Dgm109.
A significant homozygote excess was observed in Daphnia cucullata population from Lake Dridzis at one locus DaB17/17 p< 0.001, from Lake Riča at five loci (SwiD1, Dgm101, DaB17/17, Dgm109, Dp519 p< 0.001), from Lake Svente at four loci (SwiD1, Dgm105 p< 0.05, DaB17/17 and Dp519 p< 0.001) and from Lake Geraņimovas-Ilzas at three loci (SwiD1, Dgm101 p< 0.01 and DaB17/17 p< 0.001) (Table 5). Microsatellite locus DaB17/17 has maximal differentiation (p< 0.001) between the level of observed and expected heterozygosity in all investigated lakes. In addition, microsatellite loci SwiD1, Dgm101, Dgm109, Dp519 and DaB17/17 has maximal differentiation (p< 0.001) in Lake Riča (Table 5). It should be noted that microsatellite loci Dgm105, Dgm101, Dgm109 and Dp519 were monomorphic in Lake Dridzis. Whereas microsatellite loci Dgm101 and Dgm109 were monomorphic in Lake Svente, but microsatellite locus Dp519 also was monomorphic in Lake Geraņimovas-Ilzas (Table 5).
The smallest genetic distance (D) [55] in the Daphnia cucullata populations under research was observed between lakes Riča and Geraņimovas-Ilzas (0.16), while the greatest genetic distance was found between lakes Dridzis and Geraņimovas-Ilzas (0.70) and between lakes Geraņimovas-Ilzas and Svente (1.35) (Table 6).
By contrast, FST values for different Daphnia cucullata populations under research ranged from 0.08 to 0.50. The highest values were between Daphnia cucullata populations of lakes Riča and Svente (0.50) and lakes Svente and Geraņimovas-Ilzas (0.49) (Table 6). The lowest FST values were between Daphnia cucullata populations of lakes Riča and Geraņimovas-Ilzas (0.08) (Table 6).
Principal component analysis (PCA), a graph of genetic structuring among four Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas, clearly showed the genetic structuring into different genetic groups (Figure 4). Stable groups of Daphnia cucullata populations were formed between lakes Dridzis and Svente and between lakes Riča and Geraņimovas-Ilzas. In the principal component analysis plot, PC 1 and PC 2 explained 77.43% and 15.71% of the total genetic diversity. A similar result was obtained using Bayesian clustering analysis (STRUCTURE 2.3.4) [53] (Figure 5) and number of clusters of individuals using Evano et al. clustering approach (Figure 6).
4. Discussion
Daphnia cucullata species are one of the most common specimens in the species composition of Lakeland at Boreal biogeographical regions in Europe [18,19,21,23,47,57].The analysis of the genetic structure of a typical representative of the zooplankton i.e Daphnia is important in terms of predicting the impact of anthropogenic and climatic factors both on zooplankton and aquatic ecosystems in general species [13,26,30,31,32,33].
For study of genetic structure of Daphnia cucullata we used the microsatellite loci, which are frequently used in studies of the genetic structure of others Daphnia genus different species [13,26]. Six of the microsatellites markers have good representation in populations. However, some primers that have been successfully used in the study of European populations have not been amplified in Latvian populations. For example, three of the microsatellite loci (DaB10/17; Dp512; DaB17/16), which were presented for European Daphnia i.e Daphnia cucullata population from Switzerland and the Netherlands lakes analyse, did not appear in Daphnia cucullata population from Latvian lakes. This may be indicated a significant difference between the genotypes of Daphnia cucullata from Continental and Boreal biogeographical regions in Europe. The sizes of amplified loci of Daphnia cucullata populations from Continental and Boreal (Latvia) regions were similar (Table 1).
Few microsatellite loci of Daphnia cucullata populations in the Boreal region are generally slightly longer than those in the Continental region (Table 1). For example, microsatellites loci Dgm105 (165–240 bp) and Dgm 109 (250–303 bp) were longer in the Boreal region Lakeland (Latvia). Compare with Continental region data microsatellites loci Dgm105 size was 172-197 bp and Dgm 109 was 247-266 bp (Table 1). It was can be explained that Daphnia cucullata specimens from Continental region were taken from cultivated material in the laboratory, which was not exposed to the influence of various anthropogenic factors [13], but in our study the nature Daphnia populations was used directly from lakes and they have been regularly exposed to various anthropogenic factors, mainly to the impact of agriculture.
Relatively some differences between allele lengths among the Daphnia cucullata populations in Boreal and Continental biogeographical regions in Europe most probably are the result of an accidental genetic drift, but not that of mutation [58].
The highest number of alleles using microsatellite markers analysis was found in loci Dgm105 and Dgm101 (8 and 7), these loci also had the highest number of private alleles (62% and 57%) of all detected alleles in these loci. However, the number of alleles in loci Dgm105 and Dgm101 was much lower in Daphnia cucullata populations in Switzerland and Netherlands lakes (2 and 3). The number of alleles in loci DaB17/17 and Dgm109 was the same (4 and 5), but the number of alleles in locus Dp519 was lower compared to Switzerland and Netherlands data [13,26]. The observed small differences between the allele lengths in the Daphnia cucullata populations in the lakes we studied with those found in Switzerland and the Netherlands are most likely the result of random genetic drift and not mutations [58]. It is possible that the increase in allelic diversity is influenced by various chemical compounds in the water, and as one of the main influencing factors, we should mention different changes in temperature conditions in the lakes we studied with those found in Switzerland and the Netherlands [4,58,59,60,61].
In our research, the highest level of polymorphism of Daphnia cucullata populations by microsatellites in Boreal biogeographical region Lakeland during the summer season was observed in lakes Riča (100%) and Geraņimovas-Ilzas (83%), and the lowest in Lake Dridzis (33%). The polymorphism levels by microsatellites in lakes Svente and Riča were from 26% to 29%. The highest level of genetic polymorphism of Daphnia cucullata populations using RAPD analysis was observed in lakes Dridzis (50%) and Geraņimovas-Ilzas (33%). In lakes Svente and Riča it was between 26% and 29% [47]. The obtained rather different results of the level of genetic polymorphism can probably be explained by the specificity of the selected nuclear DNA markers (RAPD and microsatellites).
Haag et al. [62] showed that older populations have a higher genetic diversity and that genetic differentiation among pools decreases with population age. They assumed that the bottleneck effect may be twofold: namely decreasing genetic diversity and population-wide inbreeding. Subsequent immigration may not only introduce new genetic material, but also lead to the selection of noninbred hybrids, which may cause immigrant alleles to increase in frequency, thus leading to an increase in the genetic diversity in the older population. Consequently, it is possible that in addition to the clones we found there are various other clones of Daphnia cucullata in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas. The genetic structure of metapopulations offers insights into the genetic differentiation and it shows consequences of local extinction and recolonization. Research of rock pools metapopulation by allozymes showed that the genetic differentiation among pools of metapopulation is strong [62]. It is assumed that the genetic structure of a population in the metapopulation is largely explained by three consequences of the founder events: strong drift during colonization, subsequent immigration, and effects of selection through hitching of neutral genes with linked loci under selection [47,62].
Heterozygosity serves as an indicator of evolutionary potential and is important in determining population dynamics as well as population viability. A decrease in heterozygosity may lead to a decrease in adaptation in the population Populations which showed extremely high levels of heterozygosity were comprised largely of hybrids [5,34]. The average level of heterozygosity in the studied Daphnia cucullata populations by microsatellites in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas is relatively high and ranges from 1.67 to 3.17. The average level of heterozygosity, based on RAPD analysis, in the studied Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas ranges from 0.18 to 0.20 [47]. A substantial difference between the observed and expected level of heterozygosity was found only in the locus SwiD1 in the Daphnia cucullata population from Lake Geraņimovas-Ilzas (p<0.01). A substantial difference between the observed and expected level of heterozygosity was found in Daphnia cucullata populations of Lake Riča in the loci SwiD1, Dgm101, DaB17/17, Dgm109, Dp519 (p<0.001), and Geraņimovas-Ilzas in the locus DaB17/17. A decline in heterozygosity can lead to a lower adaptation of the population. In our study, populations of Daphnia cucullata that showed extremely high levels of heterozygosity were mostly hybrids [5,63,64]. χ2- test is identify whether a disparity between actual and predicted data is due to chance or to a link between the variables under consideration. The obtained data of genetic distance and genetic differentiation of Daphnia cucullata populations by microsatellites in our studied lakes confirm our previous data about genetic distance and genetic differentiation of Daphnia cucullata populations by RAPD [47] and show that the studied populations of Daphnia cucullata are different among themselves.
In our study, the Bayesian and Evano clustering approaches show that the populations of Daphnia cucullata (Riča and Geraņimovas-Ilzas), which are relatively far from each other, form a separate genetic group. It is difficult to explain the fact that populations that are geographically distant from each other and whose lakes are not connected to each other are the most similar. One of the factors that can affect the transfer of Daphnia cucullata individuals or their gills from one water body to another are water birds, which during migration could transfer these individuals or their gills from one water body to another [65,66,67]. It has been shown that migrating waterfowl can carry zooplankton ephippia up to 50 kilometers per day when flying between feeding or roosting sites. The literature mentions that the maximum distance that waterfowl can fly from one water body to another is 1500 kilometers [65]. Studies using mtDNA have shown that, for example, the distribution of taxa Daphnia ambigua and Daphnia laevis coincides with the flight directions of migratory waterfowl [65]. In addition, Daphnia lumholtzi, for example, was shown to spread faster than Bythotrephes longimanus and Bythotrephes cederstroemi (Cladocera), as their ephippia are less viable in the intestinal tract of birds than Daphnia lumholtzi [65]. Ephippia can also be carried by wind [68,69,70,71]. Ephippia can withstand harsh environmental conditions (freezing, desiccation), and in spring, under favorable conditions, young parthenogenetic females hatch from winter eggs [4,59,60,61,72,73,74].
Differences in the genetic structure of Daphnia cucullata populations can also be explained by a large role of cyclic parthenogenesis and the biotope size [4,58,60,61,71,75,76,77]. In cyclic parthenogenetic zooplankton, larger biotopes have a larger pool of ephippia than smaller biotopes, and thus the number of eggs from the pool of ephippia will increase at the beginning of the growing season. Ephippia accumulate annually in lake sediments, and under favorable conditions up to a century old ephippia can hatch into diploid individuals [25,35,78,79,80]. Using of these eggs from the pool of ephippia increases genetic diversity and thus significantly affects the genetic structure of cyclic parthenogenetic Daphnia populations [71,77,80,81].
In our previous study [23] was found that Daphnia cucullata populations from lakes Riča and Geraņimovas-Ilzas have a low number of zooplankton taxa (47 and 43 respectively), bet Daphnia cucullata populations from the lakes Dridzis and Svente have a large number of zooplankton taxa (72 and 69 respectively) [23]. In the present study we found that Daphnia cucullata populations from lakes Riča and Geraņimovas-Ilzas Daphnia cucullata have a higher genetic diversity compared to populations from lakes Dridzis and Svente with a large number of zooplankton taxa.
A small number of taxa affects the ecological existence niche of Daphnia cucullata taxa, because in this case, Daphnia cucullata has less interaction and competition with other taxa, it adapts more to different conditions, interbreeds, eats more, etc., therefore it has more genetic diversity. Wetzel [4], De Meester et al. [58], Kalff [60], Lampert & Sommer [61], Vanoverbeke & De Meester [71], Hebert [75], De Meester [76], Vanoverbeke et al. [77], discuss extensively that there are several explanations for the positive correlation between genetic diversity and habitat size. For example, it is a well-known fact that ecological diversity increases with increasing size of the biotope. Furthermore, the population size tends to be positively correlated with the biotope size. The length of the clonal phase and the frequency of sexual reproduction are thought to be related to the biotope size and persistence [75,76,77]. In large water bodies, where the biotope conditions are more stable, parthenogenesis is maintained for a longer time. Since larger biotopes often contain more ecological niches than smaller ones, this may favor the coexistence of ecologically distinct genotypes [76].
This can probably be explained by the similarity of the studied lakes in terms of average depth and catchment area, similar stable environmental conditions, therefore it is possible that parthenogenesis in these lakes is maintained for a longer period of time and the coexistence of different genotypes is possible here. In parthenogenetic populations of large water bodies, the survival of single dominant highly heterozygous clones is observed, as it is evidenced by the inevitable cessation of sexual genetic recombination, and heterozygous excess as a characteristic feature of long-lived Daphnia populations [78,80]. Smaller biotopes have smaller populations than larger ones, making a population with the same number of clones more vulnerable to random clonal extinction [77,82,83].
Haag et al. [62] suggest that older populations have greater genetic diversity and that genetic differentiation between populations decreases with population age. Daphnia cucullata is a cyclic parthenogenic organism whose life cycle consists of a unisexual, apomictic phase that dominates during the favorable growing season when females produce diploid parthenogenetic offspring. Parthenogenetic reproduction continues until adverse weather conditions occur, when some eggs develop into males and others into haploid eggs that require fertilization [58,74]. Parthenogenetic reproduction throughout the summer does not suppress the amount of genetic variation. Males appear when there is a high population density or a rapid depletion of nutrients. In this case, diploids produce winter eggs, or ephippia [25,35,78,79,80]. Depending on the relative importance of recombination and parthenogenetic reproduction, populations of Daphnia cucullata will have different local diversity and genetic population structure [58,73]. When populations have sufficient food (e.g., algae and bacteria) and favorable living conditions (e.g., temperature, dissolved oxygen), they exhibit high numbers of parthenogenetically or bisexually reproducing females throughout the reproductive season [4,84]. Haag et al. [62] assumed that the "bottleneck effect" can be two-fold: namely, a decrease in genetic diversity and close relatedness of population individuals.
The immigration of new individuals may not only introduce new genetic material but also lead to the selection of closely related hybrids, which may lead to an increase in the frequency of “immigrant” alleles, thereby leading to an increase in genetic diversity in older populations. The genetic structure of metapopulations provides insight into genetic differentiation and shows the consequences of local extinction and recolonization. As an example, studies of allozyme metapopulations of upland water bodies have shown that there is marked genetic differentiation between metapopulation pools [62]. Population genetic structure in a metapopulation is assumed to be largely explained by three consequences of the bottleneck effect: strong drift during colonization, subsequent immigration, and the effect of selection by bringing neutral genes with linked loci into selection [62].
Many authors widely discussed that there exist several explanations for a positive correlation between genetic diversity and habitat size. In addition, population size tends to be positively correlated with habitat size. The length of the clonal phase and the frequency of sexual reproduction appear to be related to the size and permanency of the habitat [75,76,77]. As larger habitats often harbor more ecological niches than smaller ones, this may contribute to the coexistence of ecologically different genotypes [76]. Moreover, in cyclic parthenogenetic zooplankton, large habitats will have a larger stock of resting eggs than smaller habitats and hence a higher recruitment of sexual eggs from the resting egg bank at the beginning of the growing season. The recruitment of sexual eggs from the resting egg bank will increase genetic diversity and thus have a profound impact on the genetic structure of cyclical parthenogenetic Daphnia populations [71,77,80,81]. Smaller habitats harbor smaller populations than larger ones, making a population with an equal number of clones more vulnerable to chance extinctions of clones [77].
Author Contributions
Conceptualization, A.B. (Aija Brakovska) and N.Š. (Nataļja Škute); methodology, A.B. and N.Š.; formal analysis, A.B. and N.Š.; investigation, A.B.; resources, A.B. and N.Š.; writing—original draft preparation, A.B. and N.Š.; writing—review and editing, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partly supported by the projects ESF Project No. 8.2.2.0/20/I/003 “Strengthening of Professional Competence of Daugavpils University Academic Personnel of Strategic Specialization Branches 3rd Call” and ESF Project No. 2009/0214/1DP/1.1.1.2.0/09/APIA/VIAA/089 “Formation of Interdisciplinarity Research Group for Securing the Sustainibility of Salmonid Lakes in Latvia”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data on zooplankton species diversity are publicly available in my publications in internationally peer-reviewed scientific journals.
Acknowledgments
We thank Jana Paidere for their help in zooplankton samples collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hebert, P. D. N. Competition in zooplankton communities. Annales Zoologici Fennici 1982, 19, 349–356. [Google Scholar]
- Malone, B.J.; McQueen, D.J. Horizontal patchiness in zooplankton populations in two Ontario kettle lakes. Hydrobiologia 1983, 99, 101–124. [Google Scholar] [CrossRef]
- Pinel-Alloul, B. Spatial heterogeneity as a multiscale characteristic of zooplankton community. Hydrobiologia 1995, 300/301, 17–42. [Google Scholar] [CrossRef]
- Wetzel, R.G. Limnology. Lake and River Ecosystems. Third Edition. Academic Press 2001, 1006. [Google Scholar]
- Adamowicz, S.J.; Hebert, P.D.N.; Marinone, M.C. Species diversity and endemism in the Daphnia of Argentina: a genetic investigation. Zoological Journal of the Linnean Society of London 2004, 140, 171–205. [Google Scholar] [CrossRef]
- Colbourne, J.K.; Hebert, P.D.N. The systematics of North American Daphnia (Crustacea: Anomopoda): a molecularphylogenetic approach. Philosophical Transactions of the Royal Society of London. Series B 1996, 351, 349–360. [Google Scholar]
- Colbourne, J.K.; Wilson, C.C.; Hebert, P.D.N. The systematics of Australian Daphnia and Daphniopsis (Crustacea: Cladocera): A shared phylogenetic history transformed by habitat-specific rates of evolution. Biological Journal of the Linnean Society 2006, 89, 469–488. [Google Scholar] [CrossRef]
- Dussart, B.H.; Defaye, D. Introduction to the Copepoda (2nd edition revised and enlarged). Guides to the Identification of the Microinvertebrates of the Continetal Waters of the World. 16 Backhuys Publishers, Leiden. 2001, 344.
- Harris, K. D. M.; Bartlett, N. J.; Lloyd, V.K. Daphnia as an emerging epigenetic model organism. Genetic Research International. ID 147892 2012, 1–8. [Google Scholar] [CrossRef]
- Lampert, W. Daphnia: model herbivore, predator and prey. Polish Journal of Ecology 2006, 54, 607–620. [Google Scholar]
- Colbourne, J.K.; Hebert, P.D.N.; Taylor, D.J. Evolutionary origins of phenotypic diversity in Daphnia. In Molecular Evolution and Adaptive Radiation. (Givnish, T. J. & Sytsma, K. J. eds), Cambridge University Press, London. 1997, 163 –188.
- Dodson, S.I.; Hanazato, T. Commentary on effects of anthropogenic and natural organic-chemicals on development, swimming behavior and reproduction of Daphnia, a key member of aquatic ecosystems. Environmental Health Perspectives 1995, 103, 7–11. [Google Scholar]
- Brede, N.; Thielsch, A.; Sandrock, C.; Spaak, P.; Keller, B.; Streit, B.; Schwenk, K. Microsatellite markers for European Daphnia. Molecular Ecology Notes 2006, 6, 536–539. [Google Scholar] [CrossRef]
- Cousyn, C.; De Meester, L.; Colbourne, J.K.; Brendonck, L.; Verschuren, D.; Volckaert, F. Rapid, local adaptation of zooplankton behavior to changes in predation pressure in the absence of neutral genetic changes. Proceedings of the National Academy of Sciences of the United States of America 2001, 98, 6256–6260. [Google Scholar] [CrossRef] [PubMed]
- De Meester, L.; Weider, L.J.; Tollrian, R. Alternative antipredator defences and genetic polymorphism in a pelagic predator-prey system. Nature 1995, 378, 483–485. [Google Scholar] [CrossRef]
- Harris, R.P.; Wiebe, P.H.; Lenz, J.; Skjoldal, H.R.; Huntley, M. Zooplankton Methodology Manual. Elsevier Academic Press 2005, 533–570. [Google Scholar]
- Hellsten, M.E.; Sundberg, P. Genetic variation in two sympatric European populations of Bosmina spp. (Cladocera) tested with RAPD markers. Hydrobiologia 2000, 421, 157–164. [Google Scholar] [CrossRef]
- Brakovska, A. Daphnia cucullata Sars, 1862 (CRUSTACEA: CLADOCERA) distribution and location in composition of zooplankton cenosis in Lake Dridzis. Acta Biologica Universitatis Daugavpiliensis 2014, 14, 1–19. [Google Scholar]
- Brakovska, A.; Škute, R. Ecological characterization of zooplankton groups in the deepest lakes of East Latvia. Acta Biologica Universitatis Daugavpiliensis 2007, 7, 165–174. [Google Scholar]
- Brakovska, A.; Škute, R. Ecological evaluation of zooplankton groups in Lake Geranimovas-Ilzas and Lake Garais. Proceedings of the 7th International Scientific and Practical Conference Environment. Technology. Resources 2009, 2, 43–50. [Google Scholar] [CrossRef]
- Brakovska, A.; Škute, R.; Škute, A. Heterogeneity of distribution and community composition of zooplankton in upper layers of Lake Svente. Zoology and Ecology 2012, 22, 172–180. [Google Scholar] [CrossRef]
- Brakovska, A.; Škute, N. Optimisation of DNA extraction and RAPD-PCR amplification for population genetic analysis of Daphnia cucullata Sars, 1862 (Crustacea: Cladocera). Acta Biologica Universitatis Daugavpiliensis 2013, 13, 11–20. [Google Scholar]
- Brakovska, A.; Paidere, J.; Škute, A. Dynamics and factors influencing zooplankton in the lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas (Eastern Latvia). Acta Biologica Universitatis Daugavpiliensis 2020, 20, 71–94. [Google Scholar]
- Selkoe, K. A.; Toonen, R. J. Microsatellites for ecologists: a practical guide to using and evaluating microsatellite markers. Ecology Letters 2006, 9, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Haag, T.; Santos, A. S.; Sana, D. A.; Morato, R. G.; Cullen, L. Jr.; Crawshaw, P. G. Jr.; De Angelo, C.; Di Bitetti, M. S.; Salzano, F. M.; Eizirik, E. The effect of habitat fragmentation on the genetic structure of a top predator: loss of diversity and high differentiation among remnant populations of Atlantic Forest jaguars (Panthera onca). Molecular Ecology 2010, 22, 4906–4921. [Google Scholar] [CrossRef] [PubMed]
- Colbourne, J.K.; Robison, B.; Bogart, K.; Lynch, M. Five hundred and twenty-eight microsatellite markers for ecological genomic investigations using Daphnia. Molecular Ecology Notes 2004, 4, 485–490. [Google Scholar] [CrossRef]
- Frisch, D.; Morton, P. K.; Roy Chowdhury, P.; Culver, B. W.; Colbourne, J. K.; Weider, L. J.; Jeyasingh, P. D. A millennial-scale chronicle of evolutionary responses to cultural eutrophication in Daphnia. Ecology Letters 2014, 17, 360–368. [Google Scholar] [CrossRef]
- Forest, F.; Chase, M. W.; Persson, C.; Crane, P. R.; Hawkins, J. A. The role of biotic and abiotic factors in evolution of ant dispersal in the milkwort family (polygalaceae). Evolution 2007, 61, 1675–1694. [Google Scholar] [CrossRef]
- Pálsson, S. Microsatellite variation in Daphnia pulex from both sides of the Baltic Sea. Molecular Ecology 2000, 9, 1075–1088. [Google Scholar] [CrossRef]
- Ender, A.; Schwenk, K.; Stadler, T.; Streit, B.; Schierwater, B. RAPD identification of microsatellites in Daphnia. Molecular Ecology 1996, 5, 437–441. [Google Scholar] [CrossRef]
- Fox, J.A. New microsatellite primers for Daphnia galeata mendotae. Molecular Ecology Notes 2004, 4, 544–546. [Google Scholar] [CrossRef]
- Lynch, M.; Spitze, K. Evolutionary genetics of Daphnia. In: Ecological Genetics (ed. Real L) Princeton University Press, Princeton, NJ. 1994, 109–128.
- Schwenk, K.; Spaak, P. Evolutionary and ecological consequences of interspecific hybridization in cladocerans. Experientia 1995, 51, 465–481. [Google Scholar] [CrossRef]
- Cousyn, C.; De Meester, L.; Colbourne, J.K.; Brendonck, L.; Verschuren, D.; Volckaert, F. Rapid, local adaptation of zooplankton behavior to changes in predation pressure in the absence of neutral genetic changes. Proceedings of the National Academy of Sciences 2001, 98, 6256–6260. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Pfrender, M.; Spitze, K.; Lehman, N.; Hicks, J.; Allen, D.; Latta, L.; Ottene, M.; Bogue, F.; Colbourne, J. The quantitative and molecular genetic architecture of a subdivided species Evolution 1999, 53, 100–110. 53.
- Mergeay, J.; Verschuren, D.; De Meester, L. Cryptic invasion and dispersal of an American Daphnia in East Africa. Limnol. Oceanogr 2005, 50, 1278–1283. [Google Scholar] [CrossRef]
- Silva R., M.; Pereira, F.; Carneiro, J.; Sobral, O.; Ribeiro, R.; Amorim, A.; Soares A. M. V., M.; Lopes, I. Microevolution in a Natural Population of Daphnia longispina Exposed to Acid Mine Drainage. In Interdisciplinary Studies on Environmental Chemistry — Biological Responses to Contaminants, Eds., N. Hamamura, S. Suzuki, S. Mendo, C. M. Barroso, H. Iwata and S. Tanabe. Terrapub, 2010, 213–218.
- Reid, V. A.; Carvalho, G. R.; George, D. G. Molecular genetic analysis of Daphnia in the English Lake District: species identity, hybridisation and resting egg banks. Freshwater Biology 2020, 44, 247–253. [Google Scholar] [CrossRef]
- Urtāne, L. Cladocera as types and trophic status indicators of Latvian Lakes. Thesis 1998, 168. Not published (In Latvian).
- www.ezeri.lv database. (accessed 24.01.2023).
- Eipurs, I. Lake Geraņimovas-Ilzas. The Nature of Latvia. Riga, Encyclopedia of Latvia. 1995, 2, 106. (In Latvian) [Google Scholar]
- Tidriķis. A. Lake Svente. The Nature of Latvia. Riga, Publishing house. 1998, 5, 180. (In Latvian)
- Tidriķis, A. Lake Riču. The Nature of Latvia. Riga, Publishing house. 1997, 4, 243–244. (In Latvian) [Google Scholar]
- APHA. Standart methods for the examination of waters and wastewater. 21<sup>st</sup> edition. Washington, D.C. APHA. Standart methods for the examination of waters and wastewater. 21st edition. Washington, D.C., American Public Health Association. 2005.
- Wetzel, R.G.; Likens, G.E. Limnological Analyses. New York, Springer Science, Business Media 2000, 429.
- Fitzsimmons, J.M.; Innes, D.M. No evidence of Wolbachia among Great Lakes area populations of Daphnia pulex (Crustacea: Cladocera). Journal of Plankton Research 2005, 27, 121–124. [Google Scholar] [CrossRef]
- Brakovska, A.; Paidere, J.; Škute, R.; Škute, N.; Škute, A. Occurrence of Cladocera and genetic diversity of Daphnia cucullata in pelagic zone of the Latvian salmonid lakes. Estonian Journal of Ecology 2013, 62, 244–264. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Yeh, F. C.; Yang, R. C.; Boyle, T. POPGENE 32-version 1. 31. Population Genetics Software. 1999.
- Nei, M. Analysis of gene diversity in subdivided populations. Proc.Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Tajima, F.; Tateno, Y. Accurancy of estimated phylogenetic trees from molecular data. 2 Genefrequency data. Journal of Molecular Evolution 1983, 19, 153–170. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics. Columbia University Press, New York. 1987, 512.
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, A.J. Interring weak population structure with the assistance of sample group information. Molecular Ecology Resources, 2009, 9, 1322–1332. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conservation Genetics Resources 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Nei, M. Estimation of Average Heterozygosity and Genetic Distance from a small number of Individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Molecular Ecology 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Biogeographical regions in Europe — European Environment Agency (europa.eu). (accessed 24.01.2023).
- De Meester, L.; Vanoverbeke, J.; De Gelas, K.; Ortells, R.; Spaak, P. Genetic structure of cyclic parthenogenetic zooplankton populations — a conceptual framework. Archiv für Hydrobiologie 2006, 167, 217–244. [Google Scholar] [CrossRef]
- De Meester, L.; Gómez, A.; Simon, J.C. Evolutionary and ecological genetics of cyclical parthenogens. In Moya A, Font E (eds) Evolution from molecules to ecosystems. Oxford University Press, Oxford, 2004, 122–134.
- Kalff, J. Limnology: Inland water ecosystems. Prentice- Hall, Upper Saddle River, NJ. 2002, 592.
- Lampert, W.; Sommer, U. Limnoecology: The Ecology of Lakes and Streams. Oxford University Press, New York Oxford. 1997, 382.
- Haag, C.R.; Riek, M.; Hottinger, J.W.; Pajunen, V.I.; Ebert, D. Genetic diversity and genetic differentiation in Daphnia metapopulations with subpopulations of known age. Genetics 2005, 170, 1809–1820. [Google Scholar] [CrossRef]
- Taylor, D.J; Hebert, P.D.N. Habitat dependent hybrid parentage and differential introgression between neighboringly sympatric Daphnia species. Proceedings of the National Academy of Sciences USA 1993a, 90, 7079–7083. [Google Scholar] [CrossRef]
- Taylor, D.J.; Hebert, P.D.N. A reappraisal of phenotypic variation in Daphnia galeata mendotae: the role of interspecific hybridization. Canadian Journal of Fisheries and Aquatic Sciences 1993b, 50, 2137–2146. [Google Scholar] [CrossRef]
- Green, A.J.; Figuerola, J. Recent advances in the study of longdistance dispersal of aquatic invertebrates via birds. Diversity and Distributions 2005, 11, 149–156. [Google Scholar] [CrossRef]
- Figuerola, J.; Green, A.J.; Santamaria, L. Passive internal transport of aquatic organisms by waterfowl in Doñana, south-west Spain. Global Ecology and Biogeography 2003, 12, 427–436. [Google Scholar] [CrossRef]
- Figuerola, J.; Green, A.J.; Michot, T.C. Invertebrate eggs can fly: evidence of waterfowl-mediated gene flow in aquatic invertebrates. American Naturalist 2005, 165, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Moran, C. Enzyme variability in natural populations of Daphnia carinata King. Heredity 1980, 45, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Crease, T. J.; Lynch, M.; Spitze, K. Hierarchical analysis of population genetic variation in mitochondrial and nuclear genes of Daphnia pulex. Molecular Biology and Evolution 1990, 7, 444–458. [Google Scholar]
- Gomez, A.; Carvalho, G.R. Sex, parthenogenesis and genetic structure of rotifers: microsatellite analysis of contemporary and resting egg bank populations. Molecular Ecology 2000, 9, 203–214. [Google Scholar] [CrossRef]
- Vanoverbeke, J.; De Meester, L. Among-populational genetic differentiation in the cyclical parthenogen Daphnia magna (Crustacea: Anomopoda) and its relation to geographic distance and clonal diversity. Hydrobiologia 1997, 126, 135–142. [Google Scholar] [CrossRef]
- Decaestecker, E.; De Meester, L.; Mergeay, J. Cyclical Parthenogenesis in Daphnia: Sexual Versus Asexual Reproduction. Lost Sex 2009, 295–316. [Google Scholar]
- Hughes, R.N. A Functional Biology of Clonal Animals. Chapman & Hall, London, New York. 1989.
- Hobæk, A.; Larsson, P. Sex determination in Daphnia magna. Ecology 1990, 71, 2255–2268. [Google Scholar] [CrossRef]
- Hebert, P.D.N. Genetics of Daphnia. In: Peters RH, De Bernardi R (eds) Daphnia. MemInst Ital Idrobiol 1987, 45, 439–460. [Google Scholar]
- De Meester, L. Local genetic differentiation and adaptation in freshwater zooplankton populations: patterns and processes. Ecoscience 1996, 3, 385–399. [Google Scholar] [CrossRef]
- Vanoverbeke, J.; De Gelas, K.; De Meester, L. Habitat size and the genetic structure of a cyclical parthenogen, Daphnia magna. Heredity 2007, 98, 419–426. [Google Scholar] [CrossRef]
- Deng, H.W.; Lynch, M. Change of genetic architecture in response to sex. Genetics 1996, 143, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Hairston, N.G.; Holtmeier, C.L.; Lampert, W.; Weider, L. J.; Post, D. M.; Fischer, J. M.; Cáceres, C. E.; Fox, J. A.; Gaedke, U. Natural selection for grazer resistance to toxic cyanobacteria: Evolution of phenotypic plasticity? Evolution 2001, 55, 2203–2214. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Deng, H.W. Genetic slippage in response to sex. American Naturalist 1994, 144, 242–261. [Google Scholar] [CrossRef]
- Michels, E.; Audenaert, E.; Ortells, R.; De Meester, L. Population genetic structure of three pond-inhabiting Daphnia species on a regional scale (Flanders, Belgium). Freshwater Biology 2003, 48, 1825–1839. [Google Scholar] [CrossRef]
- Spaak, P.; Denk, A.; Boersma, M.; Weider, L. J. Spatial and temporal patterns of sexual reproduction in a hybrid Daphnia species complex. Journal of Plankton Research 2004, 26, 625–635. [Google Scholar] [CrossRef]
- Wolf, H. G. Interspecific hybridization between Daphnia hyalina, D. galeata and D. cucullata and seasonal abundances of these species and their hybrids. Hydrobiologia 1987, 145, 213–217. [Google Scholar] [CrossRef]
- Gliwicz, M. Z.; Ślusarczyk, A.; Ślusarczyk, M. Life history synchronization in a long-lifespan single-cohort Daphnia population in a fishless alpine lake. Oecologia, 2001, 128, 368–378. [Google Scholar] [CrossRef]
Figure 1.
Location of lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas (map author: E.Iliško).
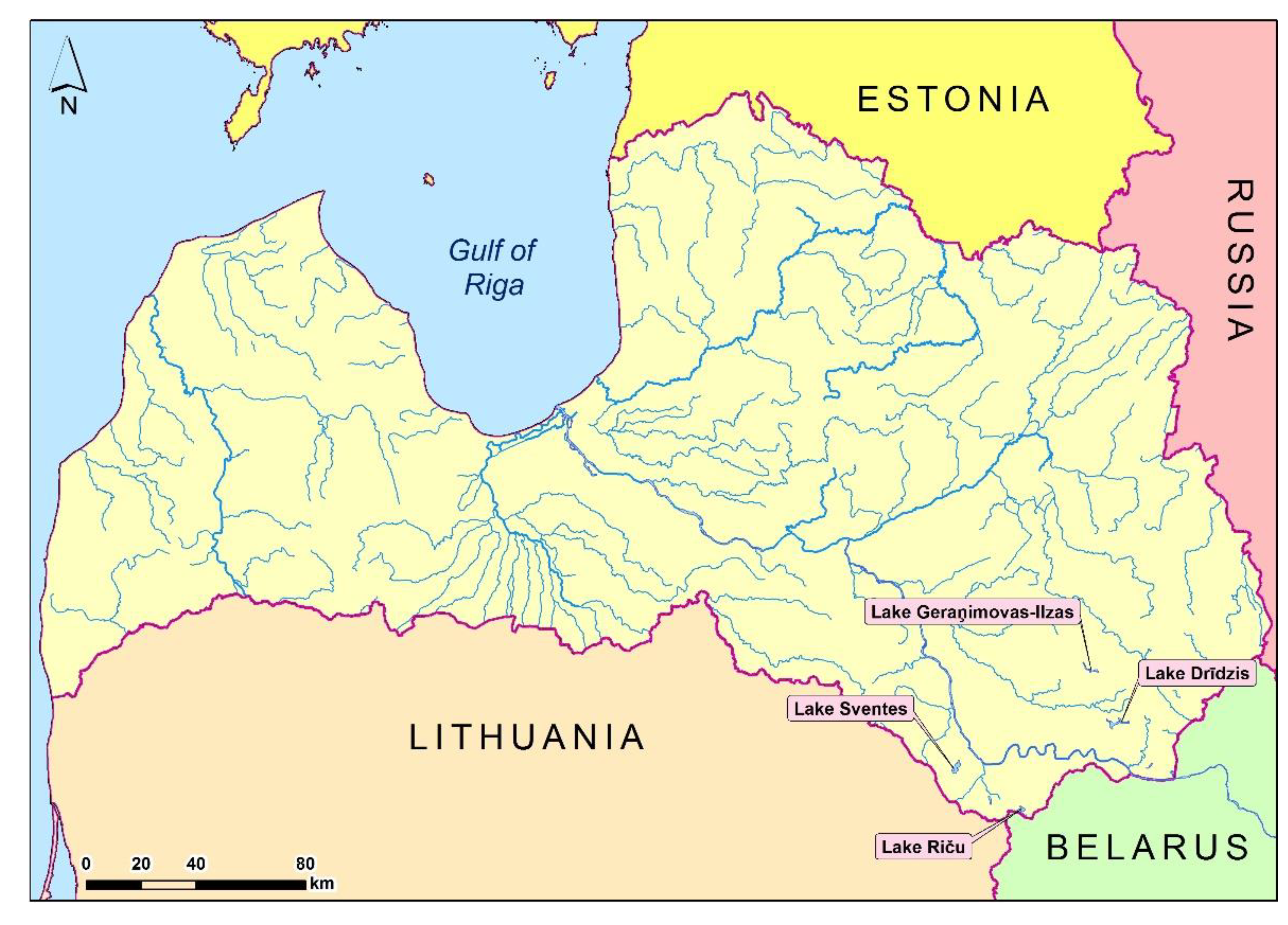
Figure 2.
Percentage of polymorphic loci of Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
Figure 2.
Percentage of polymorphic loci of Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
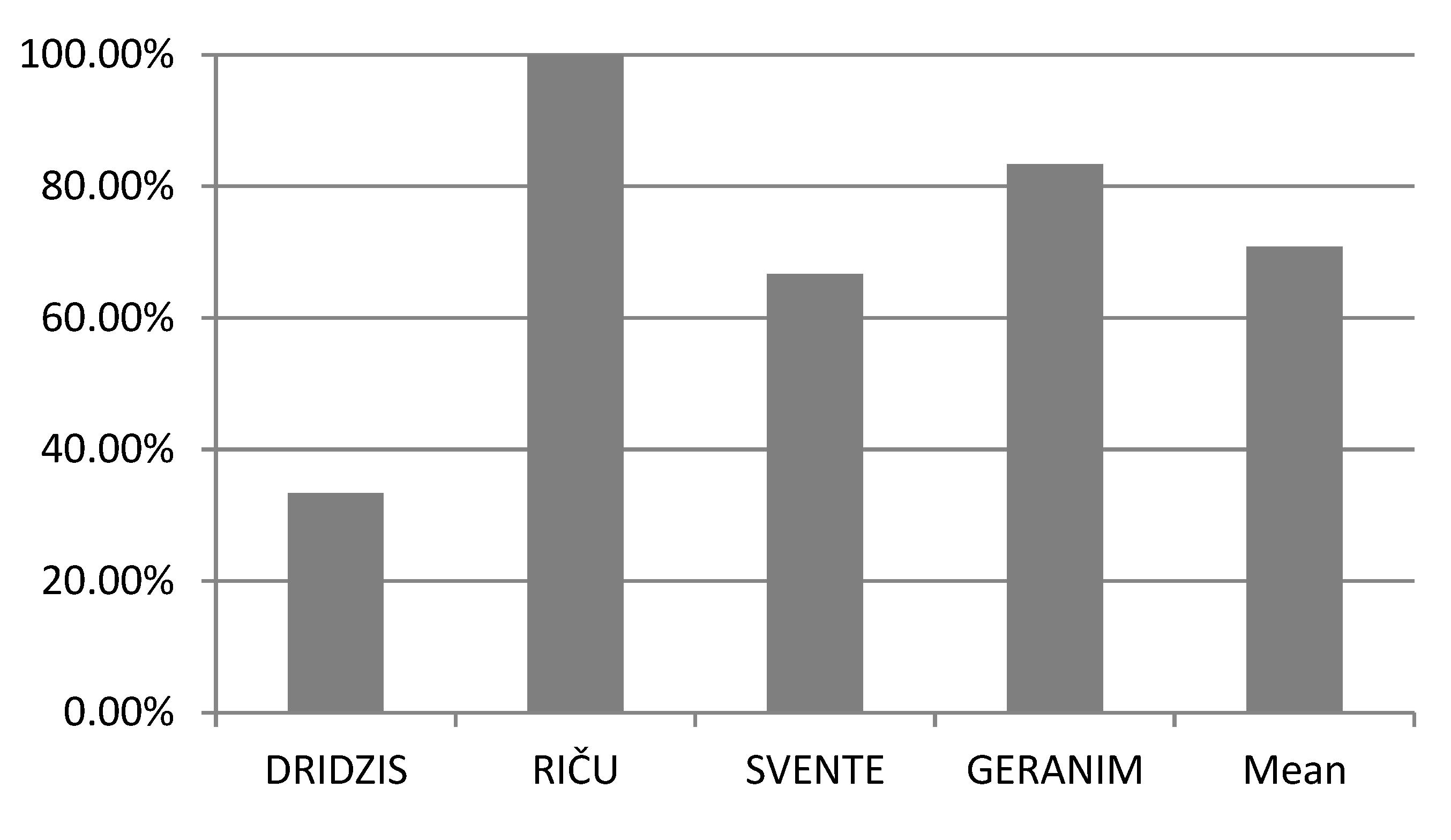
Figure 3.
Allelic patterns across Daphnia cucullata populations between lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas using microsatellites-PCR analysis (Na- the average number of alleles at the locus; Ne- the average effective number of alleles at the locus; Na ≥ 5%- average number of alleles with an incidence of more than 5%; No≤ 50%- average number of alleles with an incidence of less than 50%; No- average number of private alleles; ± standard deviation).
Figure 3.
Allelic patterns across Daphnia cucullata populations between lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas using microsatellites-PCR analysis (Na- the average number of alleles at the locus; Ne- the average effective number of alleles at the locus; Na ≥ 5%- average number of alleles with an incidence of more than 5%; No≤ 50%- average number of alleles with an incidence of less than 50%; No- average number of private alleles; ± standard deviation).
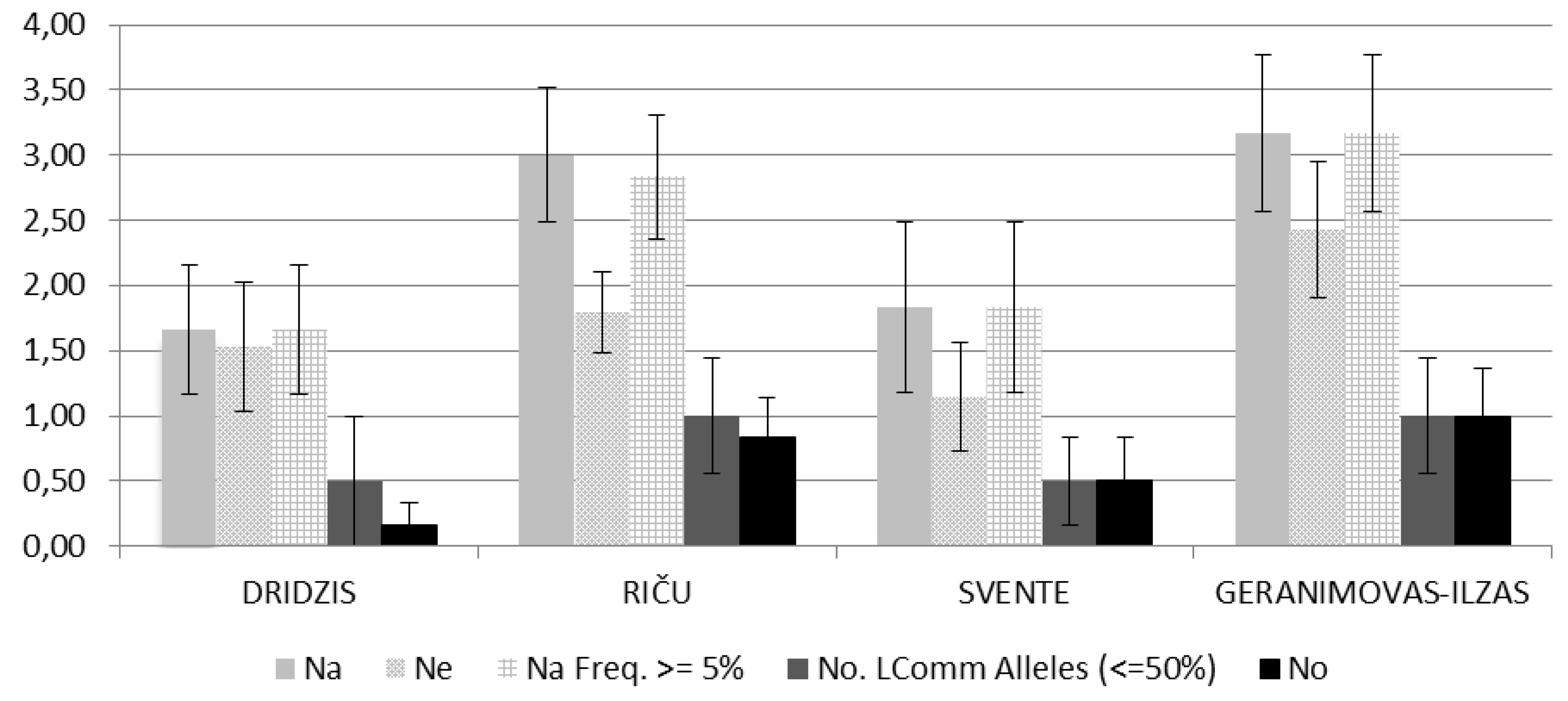
Figure 4.
Principal component analysis (PCA). Plot of genetic structuring after Nei genetic distance data among Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
Figure 4.
Principal component analysis (PCA). Plot of genetic structuring after Nei genetic distance data among Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
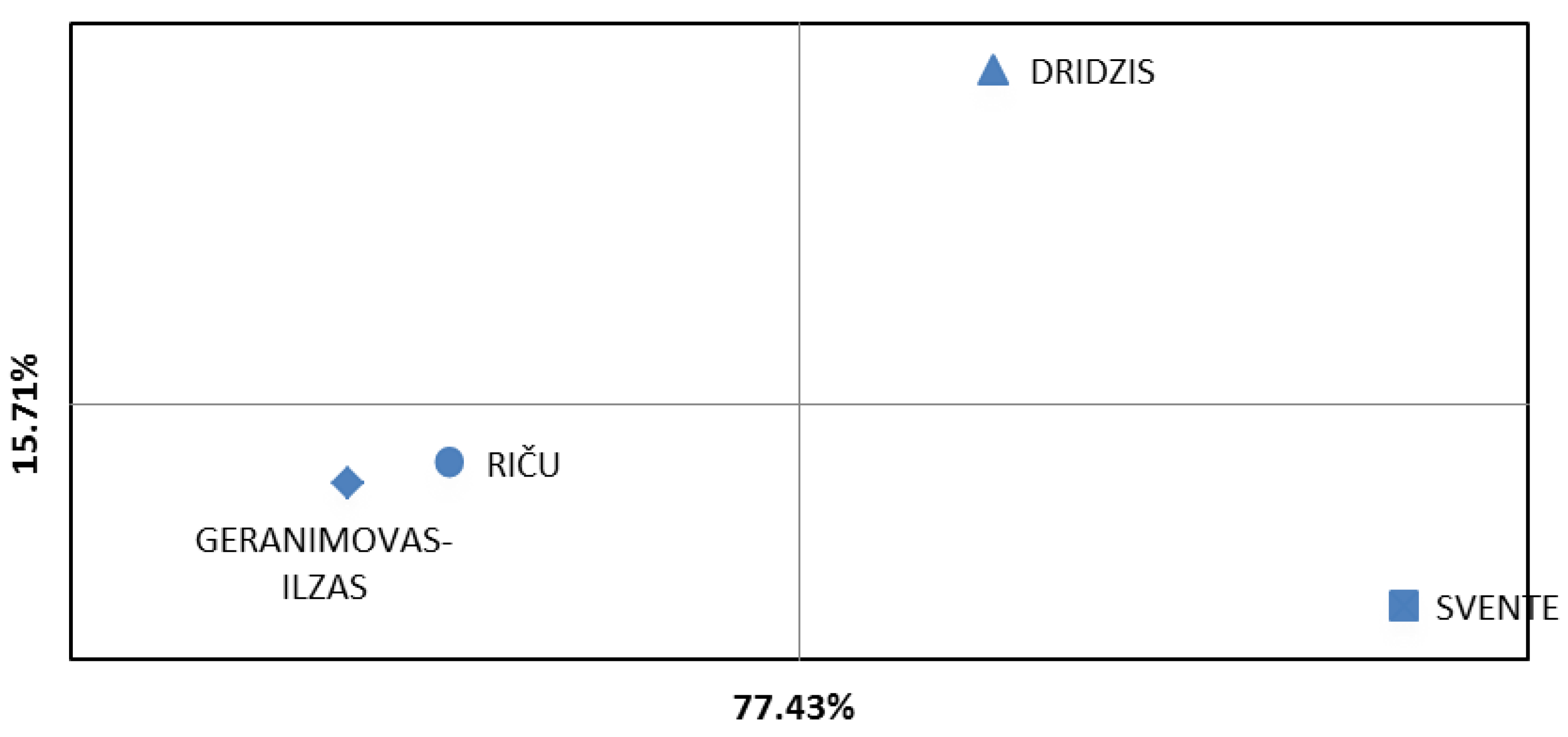
Figure 5.
Bayesian clustering of individuals using STRUCTURE 2.3.4 [53] In the STRUCTURE analysis color lines separate individuals from different sampling sites and each individual is represented by a vertical line, which is partitioned into K-coloured segments representing an individual's estimated membership in K clusters (1- Dridzis, 2- Riča, 3- Svente, 4- Geraņimovas-Ilzas).
Figure 5.
Bayesian clustering of individuals using STRUCTURE 2.3.4 [53] In the STRUCTURE analysis color lines separate individuals from different sampling sites and each individual is represented by a vertical line, which is partitioned into K-coloured segments representing an individual's estimated membership in K clusters (1- Dridzis, 2- Riča, 3- Svente, 4- Geraņimovas-Ilzas).
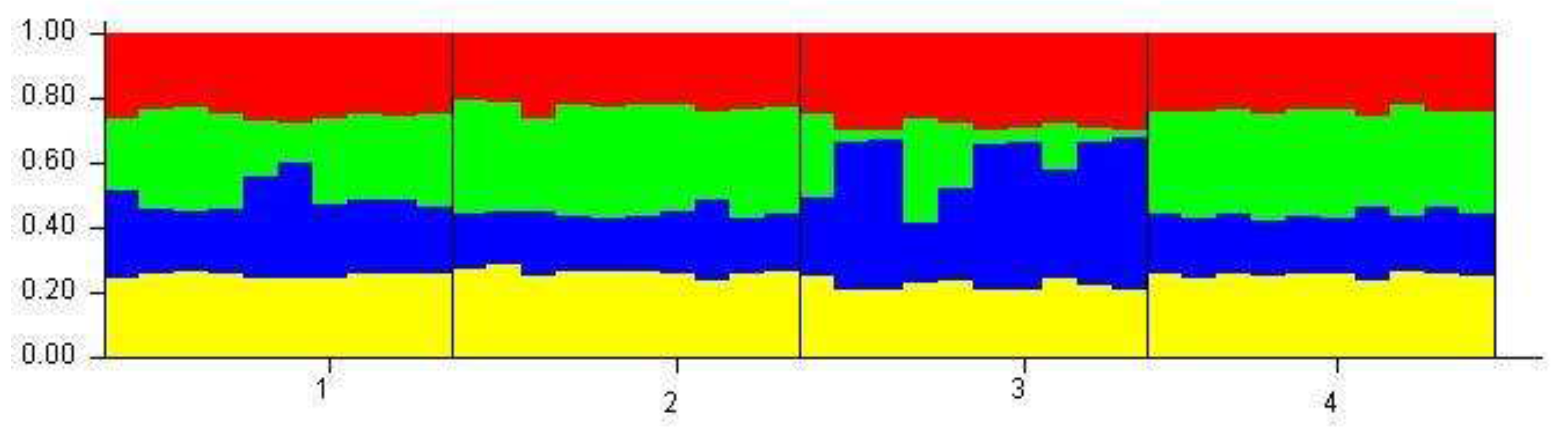
Figure 6.
The number of clusters of individuals using Evano et al. clustering approach [56] assuming two genetic clusters (K=2; ∆K=1.59; InP(K) ± SD = -298.76 ± 78.73).
Figure 6.
The number of clusters of individuals using Evano et al. clustering approach [56] assuming two genetic clusters (K=2; ∆K=1.59; InP(K) ± SD = -298.76 ± 78.73).
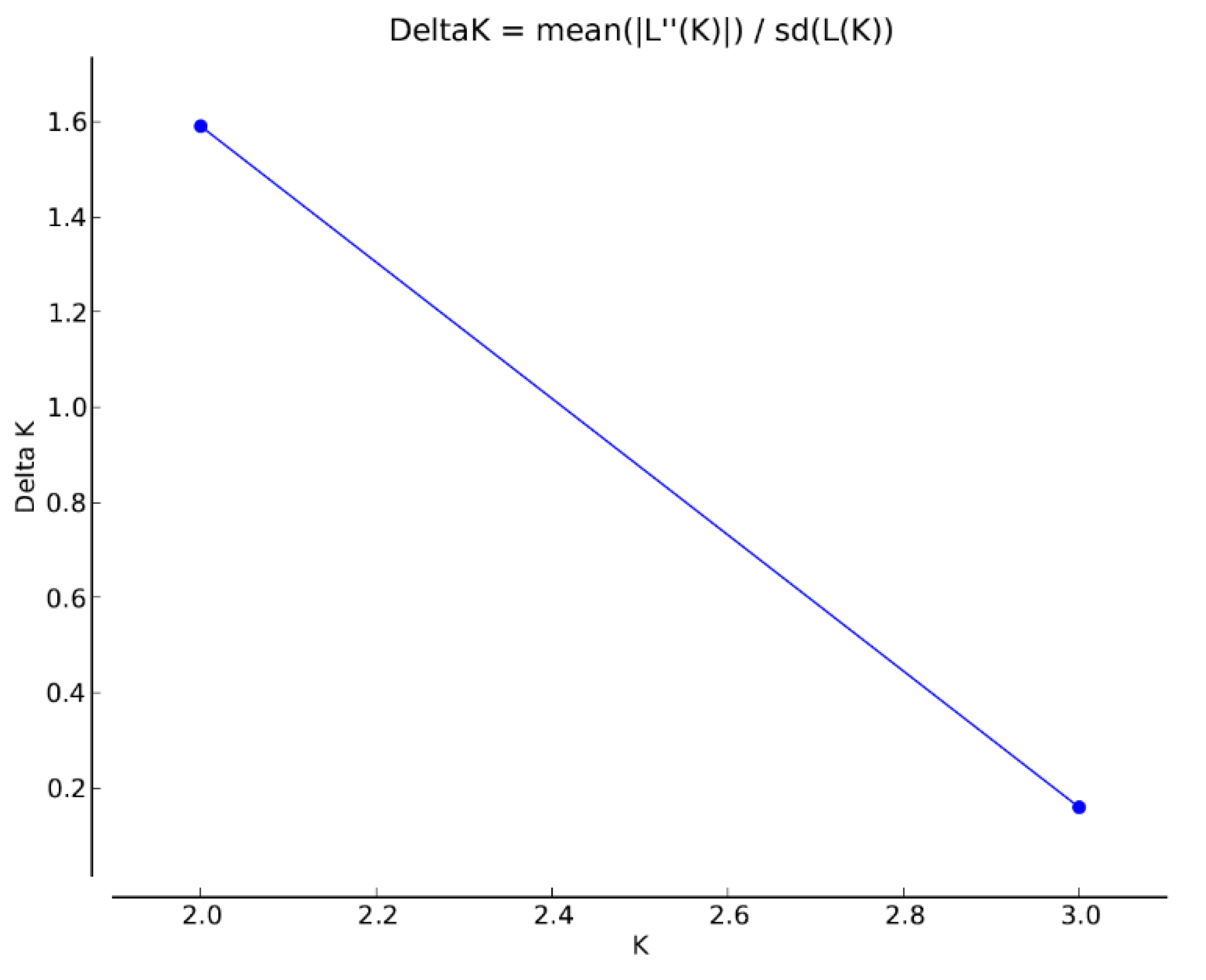

Table 1.
Hydrogeographical, hydrological and morphological parameters of investigated lakes.
| Lakes | Coordinates X/Y |
Elevation of lakes above sea level,m | Surface area with island, km2 | Surface area without island, km2 | Max. depth, m* | Mean depth, m* | Catchment basin, km2 | Shore length, km |
|---|---|---|---|---|---|---|---|---|
| Dridzis | 705390.852/ 208462.077 |
159.8 | 7.72 | 7.56 | 64 | 12.8 | 46 | 42 |
| Riča | 670715.594/ 175721.067 |
145.8 | 13.12 | 13.07 | 39 | 9.7 | 123*/130** | 34 |
| Svente | 647412.511/ 192388.091 |
136.9 | 7.06 | 7.03 | 38 | 7.8 | 20 | 26 |
| Geraņimovas-Ilzas | 696251.015/ 228167.042 |
150.7 | 3.17 | 3.17 | 46 | 9.8 | 66 | 24 |
* Catchment basin in the territory of Latvia. ** Catchment basin in the territory of Belarus.
Table 2.
Characteristics of the 9 microsatellites: locus name, primer sequences, repeat motif, modification, fragment size range, annealing temperature (Ta).
Table 2.
Characteristics of the 9 microsatellites: locus name, primer sequences, repeat motif, modification, fragment size range, annealing temperature (Ta).
|
Locus |
Primer sequences (5′−3′) |
Repeat unit |
Label dye |
Size range (bp) (our data) |
Size range (bp) (data after Brede et al.) |
Ta(°C) |
|---|---|---|---|---|---|---|
| SwiD1 | F:GCCGTGTTCGAAAGCTAGTC R: AGCCGAACGAAAAACATGC |
(TG)18 | 5'TAM | 122–127 | 116-142 | 59.4 |
| Dgm105 | F:ATGTGAGCGCGCGAGCATTT R:GTCCAGCCGGCCCATTTCAGTT |
(CAG)8AG | 5'FAM | 165–240 | 172-197 | 59.4 |
| Dgm101 | F: TCTTGCTCGAATTCTCTCC R: CCTGTCTCACACGGAGC |
(GA)10AGA | 5'HEX | 165–180 | 162-177 | 54.5 |
| DaB17/17 | F:GAGAACCTTTTATCAGCTTCG R:ACTCATCTGGTGAGATGGATC |
T9 | 5'TAM | 100–106 | 100-109 | 55.9 |
| Dgm109 | F: CCAGCTGTTGACCACCTG R: TGCGCGAGGATTTCCAACAC |
(ACC)7AC | 5'FAM | 250–303 | 247-266 | 58.2 |
| Dp519 | F:AGTCGCGACGACATAAAGC R:GTGGTAGTTGTGGAATCCG |
(TG)6(GA)7 | 5'HEX | 140–142 | 144-160 | 56.7 |
| DaB10/15 | F:AGAGAAGTGTTTGCGTTTC R:TGTTTCCTATATCCCTCGG |
TC6 | 5'TAM | No result | 75–89 | 52.4 |
| Dp512 | F:TTTCGTTCTACCCAGGGAAG R:TTTGCTCGTCTGTGATAGGC |
(TG)4…(GT)8 | 5'HEX | No result | 125-141 | 57.3 |
| DaB17/16 | F: AGGGAACGAGCGGCGATAAG R:TCTTTGGCAGGCCACTGCCAAGG |
GA10 | 5'FAM | No result | 189-195 | 61.4 |
Table 3.
Abundance of alleles in the studied microsatellite loci.
| Locus | Total number of alleles in the locus | Number of private alleles in the locus | Proportion of private alleles (%) | Number of populations in which private alleles have been detected |
|---|---|---|---|---|
| SwiD1 | 5 | 1 | 20 | 1 |
| Dgm105 | 8 | 5 | 62 | 3 |
| Dgm101 | 7 | 4 | 57 | 3 |
| DaB17/17 | 4 | 2 | 50 | 2 |
| Dgm109 | 5 | 3 | 60 | 2 |
| Dp519 | 2 | 0 | 0 | 0 |
Table 4.
Genetic diversity across studied Daphnia cucullata samples found in each studied loci and each studied lakes.
Table 4.
Genetic diversity across studied Daphnia cucullata samples found in each studied loci and each studied lakes.
| Sample | SwiD1 | Dgm105 | Dgm101 | DaB17/17 | Dgm109 | Dp519 | |
|---|---|---|---|---|---|---|---|
| Dridzis | N | 4 | 4 | 1 | 14 | 4 | 14 |
| Na | 4 | 1 | 1 | 2 | 1 | 1 | |
| No | 0 | 0 | 1 | 0 | 0 | 0 | |
| Ho | 0 | 0 | 0 | 0 | 0 | 0 | |
| He | 0.75 | 0 | 0 | 0.13 | 0 | 0 | |
| Riča | N | 13 | 12 | 12 | 19 | 11 | 19 |
| Na | 2 | 4 | 5 | 2 | 3 | 2 | |
| No | 1 | 1 | 2 | 0 | 1 | 0 | |
| Ho | 0 | 0.25 | 0 | 0 | 0 | 0 | |
| He | 0.14 | 0.51 | 0.68 | 0.46 | 0.31 | 0.1 | |
| Svente | N | 4 | 8 | 0 | 15 | 0 | 16 |
| Na | 3 | 4 | 0 | 2 | 0 | 2 | |
| No | 0 | 2 | 0 | 1 | 0 | 0 | |
| Ho | 0 | 0.25 | 0 | 0 | 0 | 0 | |
| He | 0.62 | 0.33 | 0 | 0.12 | 0 | 0.37 | |
| Geranimovas-Ilzas | N | 7 | 6 | 7 | 14 | 6 | 8 |
| Na | 2 | 5 | 4 | 3 | 4 | 1 | |
| No | 0 | 2 | 1 | 1 | 2 | 0 | |
| Ho | 0 | 0.17 | 0 | 0 | 0.17 | 0 | |
| He | 0.24 | 0.74 | 0.73 | 0.36 | 0.68 | 0 |
* Na, the average number in a locus; Ne, the average effective number of alleles in a locus; No, the average number of private alleles; Ho, observed heterozygosity; He, expected heterozygosity.
Table 5.
Significance (χ2- test) of differences between the levels of observed and expected heterozygosity in studied Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
Table 5.
Significance (χ2- test) of differences between the levels of observed and expected heterozygosity in studied Daphnia cucullata populations in lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
| Population/ Microsatellite loci |
SwiD1 | Dgm105 | Dgm101 | DaB17/17 | Dgm109 | Dp519 |
|---|---|---|---|---|---|---|
| Dridzis | ns | M | M | *** | M | M |
| Riča | *** | ns | *** | *** | *** | *** |
| Svente | * | * | M | *** | M | *** |
| Geraņimovas-Ilzas | ** | ns | ** | *** | ns | M |
- ns- not significant, M- monomorphic loci, *p< 0.05, ** p< 0.01, *** p< 0.001.
Table 6.
Genetic distance (D) [55] and genetic differentiation (after FST values) among Daphnia cucullata populations between lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
Table 6.
Genetic distance (D) [55] and genetic differentiation (after FST values) among Daphnia cucullata populations between lakes Svente, Riča, Dridzis and Geraņimovas-Ilzas.
| Population | Dridzis | Riča | Svente | Geraņimovas-Ilzas |
|---|---|---|---|---|
| Dridzis | 0.29 | 0.45 | 0.37 | |
| Riča | 0.56 | 0.50 | 0.08 | |
| Svente | 0.50 | 1.14 | 0.49 | |
| Geraņimovas-Ilzas | 0.70 | 0.16 | 1.35 |
* genetic distance (D) values below diagonal; ** genetic differentiation (FST values) over diagonal.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



