Submitted:
01 October 2023
Posted:
04 October 2023
You are already at the latest version
Abstract
Amaranth has the potential to support Malawi's food and nutrition security, income generation and livelihoods, and climate change resilience efforts. Due to the high genetic variability of Ama-ranth, there is a need to develop stable and high-yielding genotypes for sustainable production. To determine the degree of genetic stability in different environments, five Amaranth accessions were subjected to stability analysis. The experiment was carried out at three sites (Bunda, Bembeke, and Chipoka) for two seasons in 2020-2021 in the central region of Malawi. It was laid out in Ran-domized Complete Block Design (RCBD) with four replicates. Eberhart and Russell linear regres-sion model was used for stability analysis and Pearson correlation was used to test the relationship between variables. Environmental variance + (genotype x environment) was significant for four of the parameters studied, namely grain yield, plant height, leaf length, and leaf width, indicating the presence of a remarkable interaction between genotypes and environment. The results of a pooled analysis of variance showed significant differences at a 5% significance level among the Amaranth accessions, indicating inherent genetic variability. Using the linear regression model of Eberhart and Russell, accessions PE-LO-BH -01 and LL-BH -04 were identified as the highest yielding stable genotypes for leaf and grain yield, respectively. In addition, Bembeke site was the most favourable environment for all the accessions. Thus, to enhance the production of amaranth in Malawi, LL-BH-04 and PE-LO-BH-01 were put forward for release as varieties for grain and leaf respectively. These results will also guide and support for future breeding programs.
Keywords:
amaranth
; environmental index
; linear regression
; stability
1. Introduction
Malawi is endowed with a diversity of Amaranth (Amaranthus spp) species [1]. The plant belongs to the Amaranthaceae family [1]. It is a "pseudo-cereal" plant due to its taste and culinary similarity to cereals, as well as its high nutritional and medicinal value. The genus Amaranth includes about 50-60 species grown as both leaves and grains [2]. Some of the species include A. cruentus L., A. hypochondriacus L., and A. caudatus. Amaranth is widely cultivated as a green leafy vegetable in most tropical countries. The colour of the leaves ranges from green, purple, red, and gold [3]. The crop has good nutrient density and is rich in proteins, carbohydrates, vitamins, magnesium, phosphorus, potassium, and antioxidants [4]. As such, it provides health benefits when included in the diet. The vegetable improves palatability and adds variety to diets, especially those of the poor. It is a particularly valuable source of food in times of hardship such as floods, famines, droughts, and wars [1]. In addition, cultivation leads to an increase in income. Socioeconomic surveys in different parts of Africa have shown that Amaranth provides employment opportunities and income to rural people [5]. As demand for this vegetable is currently gradually increasing, a potential market for it is emerging.
Although Amaranth is a beneficial crop, the crop is underutilized and under-researched. Nevertheless, the crop is gaining increasing consumer attention because of its high nutritional quality [1]. Over the years, much work has been done on the nutritional composition of amaranth [6,7], the phytochemical composition of indigenous vegetables, which include amaranth [8], and finally on the agromorphological characterization of amaranth accessions in Malawi [5]. Consequently, there are no commercial varieties on the market in Malawi. There is therefore a need to select the best accessions as potential candidates for cultivar development. However, the Amaranth plant is more sensitive to environmental fluctuations, and yields vary in different agroecological areas. The stability of yields and related traits is greatly affected by the constant fluctuations in climatic conditions, so it is important to identify genotypes that have stable performances in different environments. Hence, the interaction between genotypes and the environment is an important factor in deciding the suitability of a crop [9].
The adaptability of varieties to environmental variation is critical to stabilizing crop production in different environments and seasons. Therefore, the best way to identify stable genotypes is to test the selected promising genotypes in different environments. [10], reported that adaptability must be measured by phenotypic stability. Information on the nature and extent of GXE interactions with yield and its components obtained through stability analysis can help identify site-specific and broadly adaptable genotypes. However, there are very few previous studies evaluating genotype-environment interactions and identifying broadly adaptive genotypes in Amaranth. Therefore, it is important to quantify the GXE interaction to develop stable cultivars that increase the leaf yield and grain yield of Amaranth. Moreover, such information is critical for the expansion of this crop in urban and rural areas. The presence of substantial genetic variability in Amaranth combined with very high phenotypic flexibility requires the development of stable genotypes for sustainable crop production.
The main objective of this study was to conduct stability analysis and identify high yielding amaranth accessions for variety development. The amaranth accessions were tested in three agroecologies of Malawi. Stable and high yielding amaranth accessions were identified. The main hypothesis of this study is whether there are stable and high yielding amaranth accessions that can contribute to amaranth variety development. The identification of stable and high-yielding amaranth accessions will facilitate the development of amaranth varieties.
2. Materials and Methods
2.1. Plant material and experimental site
Five selected Amaranth accessions were used for stability studies. Seeds of the Amaranth accessions were from five districts in Malawi (Table 1). The amaranth accessions from five districts were collected in accordance with the local laws without any conflict of interest. The Amaranth accessions were studied in three different agroecological areas: Bunda, Bembeke, and Chipoka. These sites represent three different agroecological areas in Malawi (mid-elevation plateau, mid-elevation plateau and highlands, and lakeshore and upper Shire Valley) [11]. Bembeke is a horticultural research site for the Department of Agricultural Research Services (DARS), Chipoka is a field trial site for the Department of Agricultural Extension Services (DAES), while Bunda College is a site that has long been used by DARS and LUANAR as a field site for variety trials. The average temperature, rainfall, soil type, coordinates, and agroecological zones of the sites are shown in Table 2.
2.2. General agronomic practices
Land preparation was done in all three sites. Ridges were spaced at 75 cm. Plot size was 2 m x 4.2 m with a net plot area of 3 m2 containing 4 ridges per net plot. To maintain genetic purity, plots, and replicates were separated by 75 cm. Seeds were stored in tightly sealed glass containers. Seeds were sown directly 30 cm apart between planting stations for both the hot dry season and the hot wet season. Thinning was done two weeks after germination, leaving one plant per planting station. A total of 56 plants per plot were kept in each plot. Fertilizer was applied one week after seeding with 200 kg/ha of 23:21:0+4s as base fertilizer and 120 kg/ha of calcium ammonium nitrate (CAN) as a top dressing. Acephate was used for pest control. Weed control was done by hand hoeing at 2-week intervals immediately after germination. During the hot dry season, furrow irrigation was used three times a week.
2.3. Data collection
At the time of flowering, fifteen (15) randomly selected plants were chosen within a net plot of three square meters (3 m2) in each plot and the data were measured. Plant height was measured from the bottom to the base of the inflorescence. Leaf length was measured from the leaf tip to the leaf base while leaf width was measured at the longest extension on the blade edge perpendicular to the length axis of the leaf using a 30 cm ruler. Stem girth (mm) was measured (between 3rd and 4th node) using a caliper. When each plot was harvested, the marginal rows were excluded and grain yield and leaf yield were measured. Grain yield and leaf yield were converted to tons ha-1 using the following formula.
Yield ton ha-1 (Grain/leaf) = Weight per plot (kg) x 10,000 m2 x 1 ton
Plot area (m2) x 1,000 kg
Days to 50% flowering and days to 80% maturity were determined by counting the days from planting to the days when 50% of the plants in the net plot flowered and 80% of the plants in the net plot reached maturity.
2.4. Data analysis
The collected data was firstly subjected to a normality test across location analysis using the following linear model;
Yij = µ + di + (1 + βi) ej + δij + ɛij
Normalized data were then subjected to pooled analysis of variance, and stability analysis was performed according to the linear regression model described by Eberhart and Russell using R Studio version 4.2.2 with the R package metan. Linear regression is considered a measure of genotypic response to varying environments. This model is widely used because of its simplicity and reliability. It states that a genotype is stable if the regression coefficient is equal to one (bi=1) and there is a non-significant deviation from the regression coefficient close to zero (S2 di =0) along with a higher mean power. Linear regression coefficient equal to one (bi=1) indicates average sensitivity to environmental variation; regression coefficient less than one (bi < 1), indicates above average sensitivity to environmental variation and superior adaption to unfavorable environments; regression coefficient greater than one (bi>1), indicates higher sensitivity to environmental changes, but is specifically adapted to favourable environments. Deviation from regression if non-significant (S2 di =0), the performance of genotypes for a given environment can be accurately predicted [12,13]. Pearson correlation was also performed using R studio version 4.2.2 and the R package corrplot
3. Results
3.1. Genotype x Environmental effects
Genotypes were significant for leaf yield, grain yield, plant height, stem girth, leaf length, leaf width, days to 50% flowering, and days to 80% maturity, while they were not significant for inflorescence length (Table 3). Genotype x environment were significant only for grain yield, plant height, stem girth, leaf length, leaf width and inflorescence length (Table 3). The significant G × E interaction indicated that the genotypes responded differently to the different environments.
3.2. Yield variations of Amaranth accessions across the locations
Leaf and grain yields of the five Amaranth accessions were evaluated across the three sites. The yield variations of the two parameters are presented in table 4. In terms of leaf yield, all the Amaranth accession were not statistically different across the locations except for accessions LL-BH-04 and PE-UP-BH-01. PE-UP-BH-01 accession recorded the highest leaf yield at the Bembeke site (3.250±0.2398 tons ha-1), while at Bunda and Chipoka the yields were not statistically different at 5% level of significance (Table 4). LL-BH-04 Amaranth accession recorded the least leaf yield at the Bunda site (0.925±0.2097 tons ha-1) while at Chipoka and Bembeke the yields were statistically the same. In terms of grain yield, LL-BH-04 accession was the highest at each of the studied sites (Table 4). The results also indicate that the best leaf-yielding Amaranth accession was not the same for grain yield across the sites. The highest leaf yield accession was obtained in PE-UP-BH-01 Amaranth accession, while LL-BH-04 accession was the least at each site (Table 4). In terms of grain yield, it was indicated that at each site, LL-BH-04 Amaranth accession recorded the highest. The results from table 4 also indicate that the Bembeke site was the best for almost all the accessions studied for both leaf and grain yield.
3.3. Stability analyses for yield traits
Leaf yield and grain yield were examined using Eberhart and Russell model to test the response and stability of Amaranth cultivars in the three different environments representing the three agroecologies of Malawi. Mean, linear regression (bi), and deviations from regression (S2di) were the parameters used. From the results presented in Table 5, the magnitude of the regression coefficient and deviations from regression varied across genotypes, as indicated by mean power, bi, and S2di. The analysis in Table 5 reveals that accession PE-UP-BH-01 (mean = 2.69, bi = 1.658, S2di = 0.0659) is stable for leaf yield across the zones while accession LL-BH-04 was the most stable and high-yielding grain accession in all environments with an overall mean of 6.79 tonnes per hectare, uniform regression coefficient (bi=1.53) and non-significant regression coefficient deviation from zero (S2di =0.37) (Table 6).
3.4. Stability analyses for yield attributes
The mean values for yield attributes, regression coefficient (bi), and deviation from regression (s2di) for five Amaranth accessions over three environments are presented in Table 7. The stability analysis presented in Table 7 shows that the deviation from the regression line (s2di) was non-significant among the Amaranth accessions in all yield attribute traits studied across the three different environments. However, the accessions varied in regression coefficient (bi) in all the traits. In terms of plant height, accession NU-BH-01 was the stable and widely adapted genotype with a high mean performance of 174.3 cm, regression coefficient (bi=1.35), and deviation from regression (s2di=0.0). However, Amaranth accessions MN-BH-01 (202.5 cm), PE-LO-BH-01 (167.9 cm), and PE-UP-BH-01 (164.9 cm) were not stable even though they recorded high mean value than the overall mean. The results indicate that Amaranth accession MN-BH-01 had a regression coefficient of 2.38 which reveals that the genotype was sensitive to fluctuating environments and adapted to favourable environments. While accessions PE-LO-BH -01 and PE-UP-BH -01 had a regression coefficient of less than 1. In terms of leaf length, accession PE-UP-BH -01 was the stable and widely adapted genotype with a high mean performance of 18.95 cm, a regression coefficient of 1.1, and deviation from the regression line of 0.0 while for leaf width accessions PE-LO-BH -01 (9.61 cm), MN-BH -01(9.78 cm) and PE-UP-BH -01 (9.34 cm) were stable. In terms of days to 80% maturity, accessions LL-BH-04, PE-LO-BH-01, and PE-UP-BH-01 were stable, however, their mean performance was not above the average. While in terms of inflorescence length, only accession LL-BH-04 was stable as its regression coefficient was close to one while the deviation from regression line was non-significant.
3.5. Phenotypic correlation between different characters in Amaranth
Correlations among different quantitative traits of amaranths with grain yield and leaf yield is presented in Figure 1. The Leaf yield was significantly positively correlated with days to 80% maturity (0.94*), inflorescence length (0.93*), grain yield (0.98**), and days to 50% flowering (0.99**). The results also shows that grain yield was significantly positively correlated with days to 50% flowering (0.96*), leaf yield (0.98*), days to 80% maturity (0.94*), and inflorescence length (0.93*) (Figure 1).
3.6. Environmental index of three environments for leaf and grain yield in Amaranth
Table 8 presents the environmental index and mean performance for leaf yield and grain yield across the three different environments studied. Overall, The Bembeke site recorded a high mean value of 2.34 and 5.96 tons/hectare for leaf yield and grain yield respectively. Furthermore, the Bembeke site had a positive environmental index of 0.323 and 2.36 for leaf and grain yield respectively. The results also depict that the Bembeke site was favourable environment for both traits. While the other two sites were unfavourable for both parameters as shown in Table 8.
4. Discussion
The pooled analysis of variance presented in Table 3 showed that accessions in different environments perform differently for the traits that interact with the environment. The results might be more genetic and not environmental influence. From the results, it was clearly shown that deviance from linear regression is also an extreme contributor to differences in genotype stability. This was evident by the significant variance observed from a pooled analysis of variance. Thus, both linear and nonlinear components contributed significantly to genotype-environment interactions. Hence overall, the results from a pooled analysis of variance revealed significant differences among the Amaranth accessions indicating the existence of inherent genetic variability and suggesting the need for stability analysis to identify stable accessions from the lot. Similar findings were also reported by Lakshmidevamma et al., 2022; Mbwambo et al., 2015.
The significant differences in leaf and grain yield across the three sites might be due to the influence of genotype and environmental conditions. The sites varied with mean temperature, soil type, and rainfall (table 2). Bembeke site was characterized by having low temperature, and sandy loam soils. Low temperature and sandy loam soils seem to be optimum for leaf and grain yield for all the Amaranth accessions studied. Even though the Bembeke site was the best site for both leaf and grain, the performance of the accessions varied for both leaf and grain yield. A similar trend was also observed in the other sites (Table 4). This indicates that the results might be due to genotype and not the environment. Additionally, it was also noted that no Amaranth accession was best for both leaf and grain yield. Amaranth accession PE-UP-BH-01 was the best for leaf yield across the sites while LL-BH-04 was the best for grain yield across the sites. Hence none of the Amaranth accessions studied was identified as a dual purpose. However, dual-purpose Amaranth is important as it allows smallholder farmers to exploit the full nutritional and income benefits of Amaranth [14]. As such the results suggest that the results were due to genotype since at each site they varied in terms of leaf and grain yield. However, the test was not powerful enough to conclude that the high-performing accession was stable across the studied sites. Hence, the study further employed stability analysis using Eberhart and Russell model to identify high-yielding stable genotypes for leaf and grain yield across fluctuating environments.
The present study was commissioned to identify stable high yielding Amaranth accessions for cultivar development. The analysis in Table 5 and Table 6 reveals that accessions PE-UP-BH-01 (mean = 2.69 tons/ha, bi = 1.658, S2di = 0.059) and LL-BH -04 (mean= 6.79 tons/ha, bi =1.53, S2di = 0.37) were stable accessions for leaf yield and grain yield respectively. These results indicate that the accessions had average environmental sensitivity, showing their adaptability and stability across different environments. These results are likely due to genetic differences among the Amaranth accessions. Hence the accessions PE-UP-BH-01 and LL-BH-01 could be utilized in all the environments to achieve higher and more stable leaf yield and grain yield respectively. The results support the findings of Lakshmidevamma et al., 2022; Mbwambo et al., 2015; Nyasulu et al., 2021; Stetter et al., 2015, who reported that Amaranth genotypes have different stabilities for leaf and grain yield.
The stability of genotypes in terms of yield is the consequence of the stability of their component traits [13]. Therefore, it is important to analyze the stability of these yield traits. The stability analyses for the component traits are presented in Table 7. Stable accessions for plant height were NU-BH -01 (174.3 cm), for leaf length PE-UP-BH -01 (18.95 cm), for leaf width PE-LO-BH -01 (9.61 cm), MN-BH -01(9.78 cm) and PE-UP-BH -01 (9.34 cm), for days to 80% maturity LL-BH-04, PE-LO-BH-01, and PE-UP-BH-01, while for inflorescence length was LL-BH-04. This shows that the accessions had higher mean values with uniform regression coefficients and insignificant deviation of regression coefficient from zero.
Although some accessions had higher means, they were not recommended as stable because their unit regression coefficient was greater than one and some were much less than one. The accessions PE-UP-BH -01 and PE-LO-BH -01 had bi=0.31 and 0.39, respectively, which is far less than one, indicating that they are adapted to unfavourable environments in terms of plant height. In MN-BH -01, the regression coefficient for leaf length was greater than 1, while in NU-BH -01 it was less than 1. This also means that MN-BH -01 was adapted to favourable environments, while NU-BH -01 was adapted to unfavourable environments. However, the studied parameters of yield attributes were not stable in LL-BH -04 accession but did not affect the stability of grain yield.
Many efforts have been made to establish the relationship between yield and yield components. [9], reported that yield traits do not occur independently, but as a result of interaction with other factors, resulting in a complex interaction that ultimately affects yield. Therefore, we conducted a Pearson correlation analysis to determine the relationship between yield and yield components for the current study (Figure 1). The results showed that leaf yield was significantly positively correlated with days to 80% maturity, inflorescence length, and days to 50% flowering. Pearson correlation analysis also showed that grain yield was positively correlated with days to 80% maturity, days to 50% flowering, inflorescence length and leaf yield. Therefore, the results suggest that increasing the value of the significant yield attributes would ultimately increase grain yield and leaf yield. Similar results were reported by [15].
The present study also examined the environmental indices listed in Table 8. The objective was to determine the most suitable environment that would have a positive effect on leaf and grain yield. It was found that the mean leaf and grain yields at the Bembeke site were significantly higher (2.34 and 5.93 tons ha-1, respectively) than the overall mean for both traits, indicating that the Bembeke site was a favourable environment compared to the other sites in the study. In addition, the positive environmental index for both leaf (0.323) and grain (2.36) yields reflected that it was the most favourable environment that resulted in the superior performance of the Amaranth accessions studied. As indicated in Table 2, the sites differed concerning temperature. The Bembeke site generally had a much lower temperature (17.3 0C) than the other two sites. The lower temperature at the Bembeke site proved to be favourable for these Amaranth accessions. However, the results do not agree with the findings of [16]. [16], reported that low air temperatures reduce the development of Amaranth plants. However, the results of this study showed the opposite. This might be the fact that low temperatures increases sucrose hydrolysis and therefore decreases its concentration in the flower bud which increases the uptake of carbon, hence a delay in flower induction and increase in flower size, hence an increase in leaf yield and grain yield [17]. This could also reveal Amaranth’s adaptability to various environments.
5. Conclusions
In the current study, among the five Amaranth accessions studied, Amaranth accession PE-UP-BH -01 was identified as a high-yielding stable genotype for leaf yield, and Amaranth accession LL-BH -04 for grain yield as they had higher mean yields than the overall mean with significant regression coefficient and non-significant deviation from regression close to zero. In addition, the accessions PE-LO-BH -01, PE-UP-BH -0, and MN-BH -01 showed high adaptability in leaf width, the accession PE-UP-BH-01 in leaf length, and the accession NU-BH -01 in plant height. Furthermore, the study revealed that the Bembeke site is a favourable environment for leaf and grain yield. Therefore, in this study, it was recommended that the PE-UP-BH-01 and LL-BH -04 accessions be used for possible release for leaf and grain yield, respectively, because they are more widely adaptable in Malawi.
Author Contributions
Conceptualization: MN and AS. Data collection and cleaning: MN. Data analysis: MN. Experiments layout: MN and AS. Funding acquisitions: AS. Supervision: AS, SZC, and MM. Original manuscript draft: MN. Writing review, editing, and final approval: MN, RMK, SZC, MM, RGM, and AS.
Funding
The study was supported by Capacity Building and Management of Climate Change (CABMACC) project number: CABMACC 001-2015, through the Horticulture Department of the Lilongwe University of Agriculture and Natural Resources (LUANAR), Bunda College Campus.
Data Availability Statement
Derived data supporting the results of this study are available from the corresponding authors (Mvuyeni Nyasulu and Abel Sefasi) upon request.
Acknowledgments
The study was conducted with financial support from Capacity Building and Management of Climate Change (CABMACC) through the Horticulture Department of Lilongwe University of Agriculture and Natural Resources (LUANAR), Bunda College Campus. The authors are grateful for the support of the administrators of CABMACC, the technical staff of the student farm of the Horticulture Department, Chipoka and Bembeke sites.
Conflicts of Interest
We the authors of this paper hereby declare that there are no competing interests in this publication.
References
- Mng'Omba, S.; Kwapata, M.; Bokosi, J. Performance of grain amaranth varieties under drought stressed conditions in two contrasting agro-ecological areas in malawi. Acta Hortic. 2003, 618, 313–319. [Google Scholar] [CrossRef]
- Akaneme, F.I.; Ani, G.O. Morphological assessment of genetic variability among accessions of Amaranthus hybridus World Appl. Sci. J. 2013, 28, 568–577, 2013. [Google Scholar] [CrossRef]
- Stetter, M.G.; Müller, T.; Schmid, K. Incomplete domestication of South American grain amaranth (Amaranthus caudatus) from its wild relatives. bioRxiv 2015. [CrossRef]
- Alegbejo, J.O. Nutritional Value and Utilization of Amaranthus (Amaranthus spp.)–A Review. Bayero J. Pure Appl. Sci. 2014, 6, 136–143. [Google Scholar] [CrossRef]
- Nyasulu, M.; Sefasi, A.; Chimzinga, S.; Maliro, M. Agromophological Characterisation of Amaranth Accessions from Malawi. Am. J. Plant Sci. 2021, 12, 1528–1542. [Google Scholar] [CrossRef]
- Issa, J.Y.; Onyango, A.; Makokha, A.; Okoth, J. Nutrients Content and Antioxidant Potential of Selected Traditional Vegetables Grown in Malawi. J. Agric. Stud. 2022, 9, 406–420. [Google Scholar] [CrossRef]
- Kachiguma, N.A.; Mwase, W.; Maliro, M.; Damaliphetsa, A. Chemical and Mineral Composition of Amaranth ( Amaranthus L.) Species Chemical and Mineral Composition of Amaranth ( Amaranthus L.) Species Collected From Central Malawi. J. Food Res. 2015, 4, 95–100. [Google Scholar] [CrossRef]
- Lesten, E.C.C.; Kingsley, G.M. Proximate and phytochemical composition of selected indigenous leafy vegetables consumed in Malawi. Afr. J. Food Sci. 2020, 14, 265–273. [Google Scholar] [CrossRef]
- Khan, M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Mamun, A. Path-coefficient and correlation analysis in Bambara groundnut (Vigna subterranea [L.] Verdc.) accessions over environments. Sci. Rep. 2022, 12, 1–12. [Google Scholar] [CrossRef]
- Begna, T. The Role of Genotype by Environmental Interaction in Plant Breeding. Int. J. Agric. Biosci. 2020, 11, 9–18. [Google Scholar] [CrossRef]
- P. C. Nalivata, M. W. Munthali, W. I. Makumba, and E. Mbewe, Optimizing Fertilizer Use within the Context of Integrated Soil Fertility Management in Malawi. Malawi: CABI, 2017.
- Eberhart, S.A.; Russell, W.A. Stability Parameters for Comparing Varieties. Crop. Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Lakshmidevamma, T.N.; Deshpande, S.K.; Jagadeesha, R.C.; Patil, B.R.; Patil, R.V.; Mirajkar, K. Assessment of Promising Diverse Germplasm Accessions for Stability with Respect to Yield and Its Attributing Traits in Vegetable Amaranth. Int. J. Environ. Clim. Chang. 2022, 12, 870–881. [Google Scholar] [CrossRef]
- Mbwambo, O.; Onyango, M.O.A.; Dinssa, F.F.; Ojiewo, C. Performances of elite amaranth genotypes in grain and leaf yields in Northern Tanzania. J. Hortic. For. 2015, 7, 16–23. [Google Scholar] [CrossRef]
- Kandel, M.; Rijal, T.R.; Kandel, B.P. Evaluation and identification of stable and high yielding genotypes for varietal development in amaranthus (Amaranthus hypochondriacus L.) under hilly region of Nepal. J. Agric. Food Res. 2021, 5, 100158. [Google Scholar] [CrossRef]
- Khandaker, L.; Akond, A.S.M.G.M.; Oba, S. Air temperature and sunlight intensity of different growing period affects the biomass, leaf color and betacyanin pigment accumulations in red amaranth ( amaranthus tricolor l.). J. Cent. Eur. Agric. 2010, 10, 439–448. [Google Scholar]
- Vaid, T.M.; Runkle, E.S.; Room, A.; Lansing, E.; Frantz, J.M.; Stop, M. Mean Daily Temperature Regulates Plant Quality Attributes of Annual Ornamental Plants. Hortscience 2014, 49, 574–580. [Google Scholar] [CrossRef]
Figure 1.
Pearson’s correlation for the quantitative variables. Abbreviations represent; leaf length = LL, leaf width = LW, plant height = PH, stem girth = SG, leaf yield = LY, grain yield = GY, inflorescence length=IF, days to 50% flowering=D50, Days to 80% maturity, ns=not significant.
Figure 1.
Pearson’s correlation for the quantitative variables. Abbreviations represent; leaf length = LL, leaf width = LW, plant height = PH, stem girth = SG, leaf yield = LY, grain yield = GY, inflorescence length=IF, days to 50% flowering=D50, Days to 80% maturity, ns=not significant.
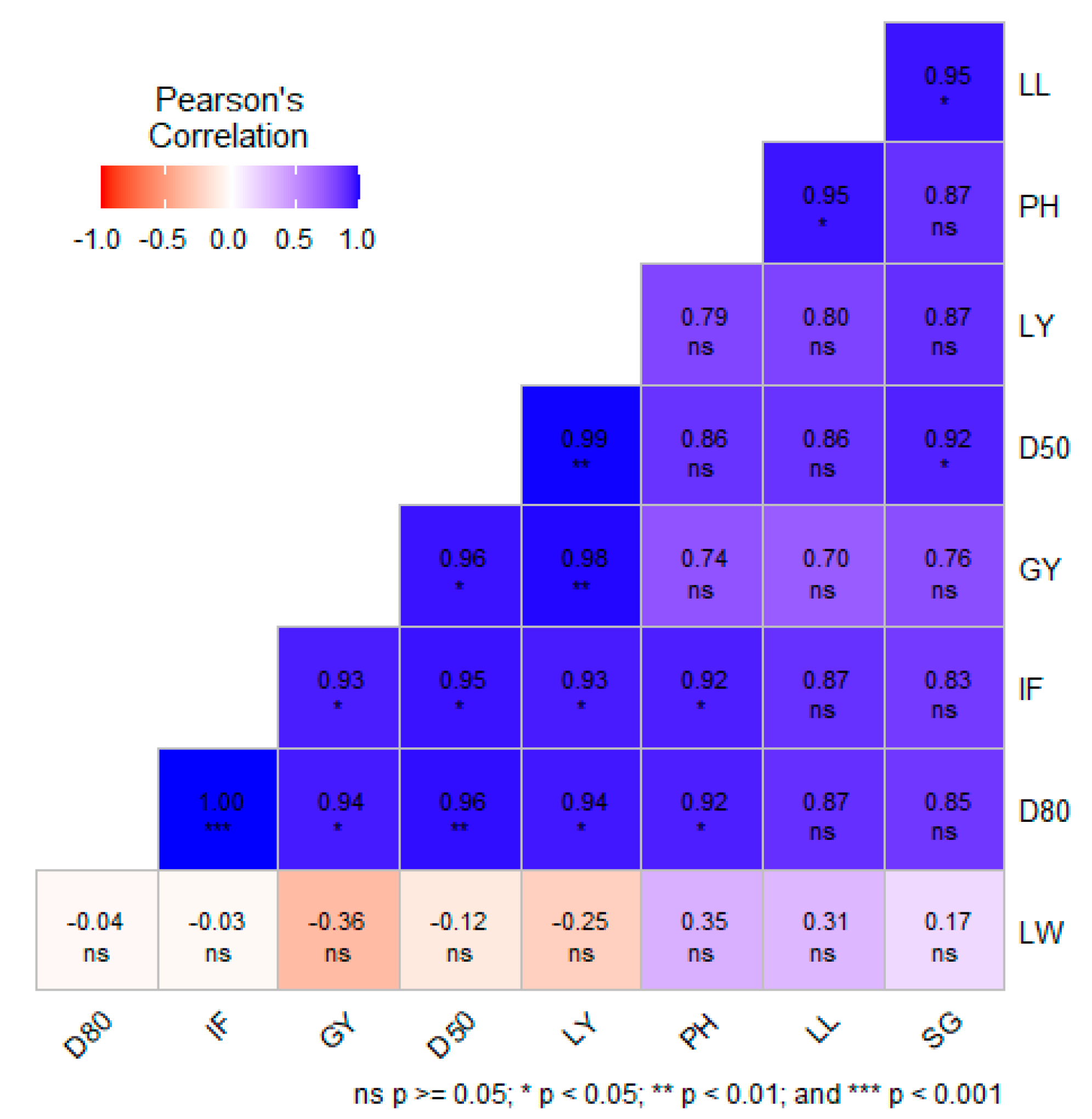

Table 1.
Source of Amaranth accessions used in the study.
| Accession | Origin district | Elevation (masl) | Latitude (south) | Longitude (east) | Agroecology |
|---|---|---|---|---|---|
| PE-UP-BH-01 | Phalombe upper | 803.75 | 15.77 | 36.66 | Mid-altitude plateau |
| PE-LO-BH-01 | Phalombe lower | 728.77 | 15.65 | 35.63 | Mid-altitude plateau |
| NU-BH-01 | Ntcheu | 1422.5 | 14.67 | 34.53 | Mid-altitude plateau |
| MN-BH-01 | Mwanza | 2529 | 15.37 | 34.29 | Lakeshore and upper shire valley |
| LL-BH-04 | Lilongwe | 1100 | 14.11 | 33.46 | Mid-altitude plateau |
masl= Meters above sea level.
Table 2.
Study sites characteristics.
| Location | Agro-ecological zone | Coordinates | Elevation (masl) | Average Temperature (0C) | Total rainfall (mm) | Soil type | |
|---|---|---|---|---|---|---|---|
| Latitude (South) | Longitude (East) | ||||||
| Bunda | Mid-elevation, upland plateau | 14° 11’ | 33° 46’ | 1,100 | 27.6 | 834.9 | Sandy-clay-loam |
| Bembeke | Highlands | 14°23.67’ | 34° 22.9´ | 1,474.3 | 17.3 | 667.5 | Sandy-loam |
| Chipoka | Lakeshore, middle and upper shire | 13° 57.07’ | 34°27.42 | 531.88 | 30.1 | 976.2 | Sandy-loam |
Table 3.
A pooled analysis of variance for stability parameters of Amaranth traits yield attributes.
Table 3.
A pooled analysis of variance for stability parameters of Amaranth traits yield attributes.
| Mean squares for different traits | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Source | DF | LY | GY | PH | SG | LL | LW | IF | D50 | D80 |
| ENV | 2 | 1.573 | 97.523* | 81642* | 13.601* | 1452* | 412* | 1994 | 6.17e+03* | 6771.3* |
| REP(ENV) | 9 | 0.908 | 1.035 | 8232* | 0.216 | 32.5* | 11.6 | 116 | 1.16e+01 | 95.4* |
| GEN | 4 | 3.098* | 27.404* | 185317* | 13.987* | 563.1* | 899* | 2133 | 3.08e+03* | 2682.5* |
| GEN x ENV (LINEAR) | 8 | 0.517 | 8.115* | 30621* | 5.772* | 91.7* | 64.8* | 4119* | 3.70e-28 | 16.9 |
| Pooled error | 36 | 0.817 | 0.532 | 4298 | 0.616 | 45.0 | 11 | 1035 | 3.47e+01 | 45.6 |
* - Significant at 5 % level of significance, GEN=Genotype, ENV=Environment, GY= Grain yield (tons ha-1), LY= Leaf yield (tons ha-1), PH= plant height (cm), SG= stem girth (mm), LL= leaf length (cm), LW= leaf width (cm), IF= inflorescence length (cm), D50= Days to 50% flowering, D80= Days to 80% Maturity.
Table 4.
Yield variations of Amaranth accessions across the sites.
| Accession | Leaf yield (tons/ha) | Grain yield (tons/ha) | |
|---|---|---|---|
| Bunda | LL-BH-04 | 0.925±0.2097a | 5.725±1.1131de |
| NU-BH-01 | 1.850±0.7837ab | 1.375±0.0479a | |
| MN-BH-01 | 1.600±0.1472ab | 2.750±0.0645abc | |
| PE-LO-BH-01 | 2.075±0.8440ab | 1.825±0.0854a | |
| PE-UP-BH-01 | 2.750±0.2398ab | 4.400±0.2345cd | |
| Mean | 1.840±0.2540 | 3.215±0.4250b | |
| Chipoka | LL-BH-04 | 1.525±0.3301ab | 2.250±0.2500ab |
| NU-BH-01 | 1.450±0.2661ab | 1.125±0.0479a | |
| MN-BH-01 | 2.350±0.1555ab | 2.125±0.2428 a | |
| PE-LO-BH-01 | 1.950±0.1500ab | 1.150±0.0645 a | |
| PE-UP-BH-01 | 2.075±0.0854ab | 1.100±0.0408 a | |
| Mean | 1.870±0.1159 | 1.550±0.1358 a | |
| Bembeke | LL-BH-04 | 1.425±0.2097ab | 9.075±0.7432 f |
| NU-BH-01 | 2.350±0.7837ab | 2.350±0.4992 ab | |
| MN-BH-01 | 2.100±0.1472 ab | 4.025±0.2358 bcd | |
| PE-LO-BH-01 | 2.575±0.8440 ab | 7.000±0.0408 e | |
| PE-UP-BH-01 | 3.250±0.2398b | 7.175±0.27803 e | |
| Mean | 2.340±0.2540 | 5.925±0.5789 c |
Table 5.
Stability parameters of Amaranth accessions for leaf yield per hectare.
| SI.NO | Accession | Leaf yield per hectare (Tons ha-1) | bi | S2di | |||
|---|---|---|---|---|---|---|---|
| Bunda | Chipoka | Bembeke | Grand mean | ||||
| 1 | LL-BH-04 | 0.93 | 1.53 | 1.43 | 1.30 | 0.468 | -0.032 |
| 2 | PE-LO-BH-01 | 2.08 | 1.95 | 2.76 | 2.26 | 1.145 | -0.192 |
| 3 | MN-BH-01 | 1.60 | 2.35 | 2.10 | 2.02 | 0.329 | 0.070 |
| 4 | NU-BH-01 | 1.85 | 1.45 | 2.35 | 1.88 | 1.401 | -0.106 |
| 5 | PE-UP-BH-01 | 2.75 | 2.08 | 3.25 | 2.69 | 1.658 | 0.059 |
| C.V. (%) | 16.0 | 15.9 | 16.4 | - | |||
| Mean | 1.84 | 1.71 | 2.34 | 1.98 | |||
| S.E(mean) | 0.25 | 0.25 | 0.12 | ||||
| Mean of bi | 1.00 | ||||||
Table 6.
Stability parameters of Amaranth accessions for grain yield per hectare.
| SI.NO | Accession | Grain yield per hectare (Tons ha-1) | bi | S2di | |||
|---|---|---|---|---|---|---|---|
| Bunda | Chipoka | Bembeke | Grand mean | ||||
| 1 | LL-BH-04 | 5.73 | 5.55 | 9.08 | 6.79 | 1.53 | 0.37 |
| 2 | PE-LO-BH-01 | 1.83 | 1.15 | 7.00 | 3.33 | 1.39 | 1.44 |
| 3 | MN-BH-01 | 2.75 | 2.13 | 4.03 | 2.97 | 0.44 | -0.13 |
| 4 | NU-BH-01 | 1.38 | 1.23 | 2.35 | 1.62 | 0.29 | -0.10 |
| 5 | PE-UP-BH-01 | 4.40 | 1.10 | 7.18 | 4.23 | 1.35 | 0.51 |
| C.V. (%) | 9.30 | 8.11 | 10.6 | - | |||
| Mean | 3.22 | 2.23 | 5.93 | 3.79 | |||
| S.E(mean) | 0.25 | 0.12 | 0.25 | ||||
| Mean of bi | 1.00 | ||||||
Table 7.
Stability analyses of yield traits of Amaranth accessions.
| Accession | Plant height(cm) | Leaf length (cm) | Leaf width (cm) | D80 | IF (cm) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | bi | S2di | Mean | bi | S2di | Mean | bi | S2di | Mean | bi | S2di | Mean | bi | S2di | ||
| LL-BH-04 | 98.4 | 0.58 | 0.0 | 13.59 | 1.1 | 0.0 | 6.41 | 0.85 | 0.02 | 105 | 1.01 | 0.04 | 28.9 | 0.83 | 0.0 | |
| PE-LO-BH-01 | 167.9 | 0.39 | 0.0 | 17.41 | 0.3 | 0.0 | 9.61 | 1.01 | 0.02 | 125 | 1.04 | 0.09 | 45.0 | 3.26 | 0.01 | |
| MN-BH-01 | 202.5 | 2.38 | 0.0 | 19.20 | 2.8 | 0.0 | 9.78 | 1.51 | 0.04 | 141 | 1.04 | 1.60 | 62.8 | -0.81 | 0.01 | |
| NU-BH-01 | 174.3 | 1.35 | 0.0 | 18.25 | 0.2 | 0.0 | 3.88 | 0.82 | 0.19 | 142 | 0.89 | 0.14 | 41.3 | -1.78 | 0.02 | |
| PE-UP-BH-01 | 164.9 | 0.31 | 0.0 | 18.95 | 1.1 | 0.0 | 9.34 | 0.82 | 0.00 | 125 | 1.02 | 0.16 | 46.9 | 3.5 | 0.02 | |
| Mean | 161.6 | 17.48 | 7.80 | 128 | 44.98 | |||||||||||
| S.E(mean) ± | 2.63 | 0.23 | 0.15 | 6.72 | 2.67 | |||||||||||
Table 8.
Favourable and unfavourable environments for grain yield and leaf yield per hectare.
| SI.NO | Location | Leaf yield (Tons ha-1) | Grain yield (Tons ha-1) | ||||
|---|---|---|---|---|---|---|---|
| Mean | Index | Status | mean | Index | Status | ||
| 1 | Bunda | 1.84 | -0.177 | Unfavourable | 3.22 | -0.348 | Unfavourable |
| 3 | Chipoka | 1.71 | -0.147 | Unfavourable | 2.23 | -2.01 | Unfavourable |
| 3 | Bembeke | 2.34 | 0.323 | Favourable | 5.93 | 2.36 | Favourable |
| Grand mean | 1.98 | 3.79 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



