
Submitted:
05 October 2023
Posted:
05 October 2023
You are already at the latest version
Abstract
The relative proportions between nutrients affects the fate of them for metabolic interconversion, storage and turnover or, essentially for energy, in a process of partition modulated by hormonal and metabolic factors. A basic common chow model was used: ST or standard diet, HF high-fat and HP or high-protein had a common substrate. A CF cafeteria-type diet was added for comparison. Rats (female and male) received the diet for 30d. Nutrient intake and body composition were measured. Plasma glucose, lactate, testosterone (T) and estradiol (E2) were measured. Common groups energy intake, was largely based in carbohydrates CH, yielding 6C units, which were fully oxidized or stored as 2C fragments (fatty acids). Lactate (3C) was partially stored as glycerol of TAG. Accrual of 2C far exceeded about ten-fold that of 3C. Amino acid (AA) catabolism yielded 2C, 3C and 4C-5C fragments, the latter acting as Krebs cycle anaplerotic intermediates (KCAI), that facilitated the oxidation of 2C in detriment of TAG accumulation Lactate levels, were related to lactic dehydrogenase activities in muscle and liver. TAG accrual was maximal in CF, quite differently from HF with similar diet lipid proportion. Maximal rates of lipid storage were found in ST (and CF) groups. Females showed a higher ability to oxidize CH and to store less TAG. T increased N and protein deposition, whilst E2 decreased them favoring AA oxidation and the availability of KCAI. In the rat, an effective energy partition scheme requires that about half of the energy is provided by CH (polysaccharide), and needs enough protein to provide KCAI for a smooth 2C oxidation.
Keywords:
Energy partition
; Rat
; Estradiol
; Diets
; KCAI (Krebs cycle anaplerotic intermediates)
; Testosterone
; Energy balances.
1. Introduction
The relative composition of diets, i.e., the direct mass (but also energy content) relationships between the main classes of nutrients is critical for their function. This is largely related to the proportions of substrates, which render 3C (i.e., three-carbon intermediate metabolites related to glycolysis/ gluconeogenesis) or 2C (i.e., acetyl-CoA and 2-carbon intermediate metabolites related with it) [1]. Thus, energy partition constitutes and essential step in the distribution of energetic substrates. The role of liver in this process is essential [2], but its hormonal control, specially from steroid hormones must be specially taken into account.
Diet composition and energy partition
Experimental changes in the proportions of diet nutrients are commonly used to analyze the adaptability to the metabolic needs [3] and to understand (and find ways to prevent) the chronic energy-handling disorders affecting the maintenance of body weight homeostasis. Thus, we know that increases in dieraty protein tend to decrease food intake and induce a limited loss of weight [4], whereas high-energy diets (such as high-fat) are markedly obesogenic [5,6,7]: i.e., favor a higher lipid deposition but do not induce parallel effects on body protein. However, the question of direct translation of dietary energy into rapidly available energy sources is far from being a simple tallying of energy proportions and absolute loads, since these are not the only factors affecting energy partition and disposal: including the amount and nature of lipids, but also the proportions of other nutrients and the palatability of the food [8,9].
When trying to compare the metabolic effects of diet modifications, the main problem is the (necessary) use of standard diets as comparison or controls, compounded by the often quite different composition of these reference diets. The core of the question lies on the evolved and finely tuned mechanisms of interconversion of practically all foods ingested into a much shorter list of macronutrients and their intermediary catabolites (roughly equivalent to energy suppliers, direct or potential). These, are further broken down (hydrolyzed, oxidized) into smaller common metabolite fragments: 1C, 2C, 3C, and a few larger structures from amino acid hydrocarbon skeletons): 4C and 5C [10]. The 1C corresponding to one-carbon metabolism intermediates, 2C and 3C already defined above, and 4C – 5C being four-carbon or five-carbon compounds intermediates of the Krebs cycle [11] or metabolites related to them, as may be the case for propionate: a three-carbon compound with a metabolic 4C behavior and function. This short list accounts for practically all molecules usable by our cells after the digestive/ assimilation process of the food.
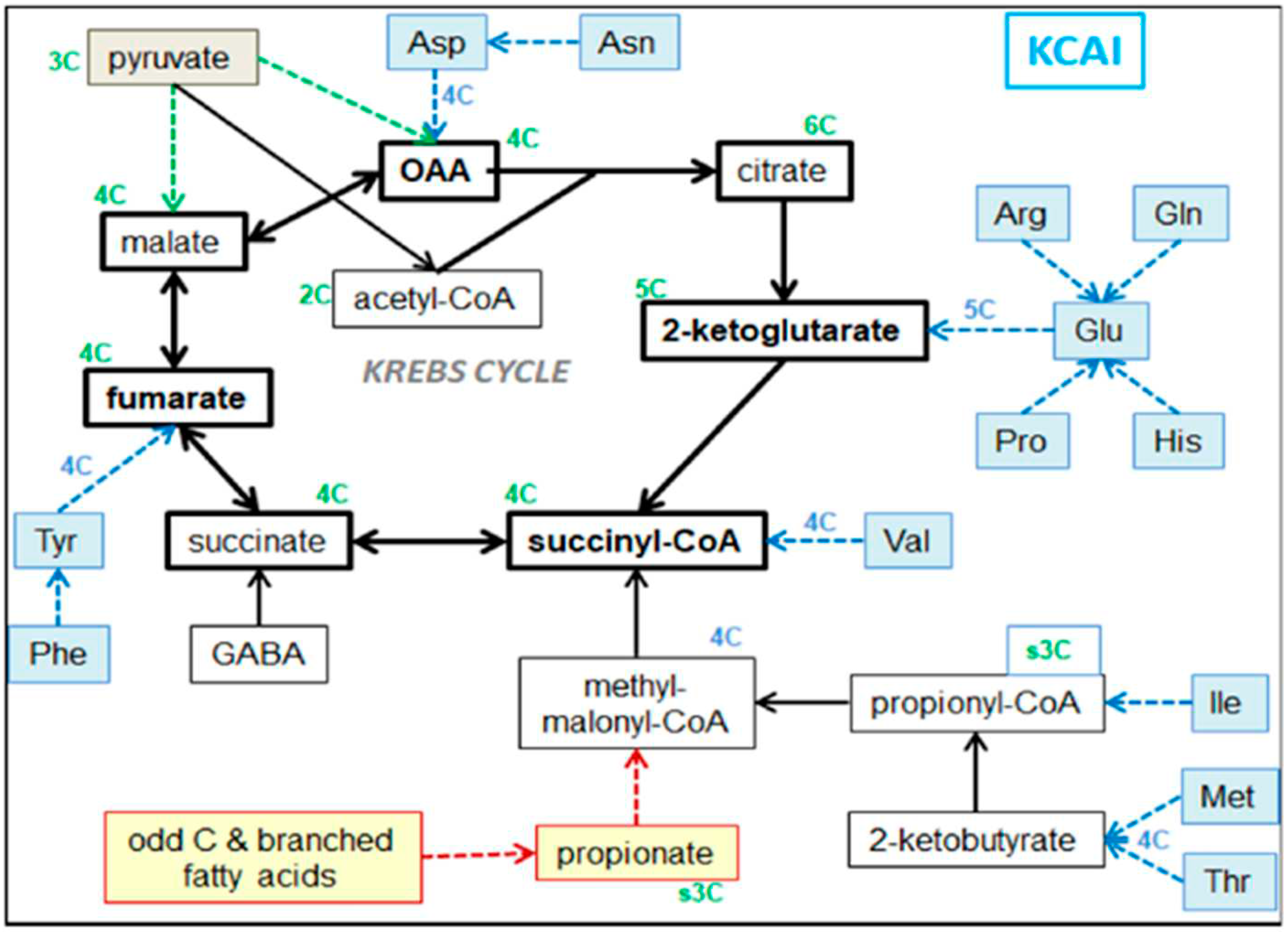
The large, combined mass of 3C and 2C (from carbohydrates and triacylglycerols –but also from many amino-acid after the loss of N– places the core of energy partition around 3C and 2C units, because of their mass, and the critical C3→C2 conversion step [12]. This reaction (pyruvate dehydrogenase) is a limiting factor in the oxidative processing of 3C, but the oxidation of 2C along the Krebs cycle is rate-limiting, depending in part on the anaplerotic effects (and, thus, availability) of 4C and 5C intermediates (i.e., oxaloacetate, malate, succinate and 2-ketoglutarate) to maintain the overall flow of 2C oxidation. The provision of these substrates from dietary sources (essentially amino acids) markedly widens the rate of oxidative potential of the mitochondria, and thus their ability to provide reducing power and ATP. This effect is more transcendent than the assumedly ‘minor relative energy’ contribution of these 4C-5C substrates from amino acid, since not only they are a majority of amino acid catabolite structures, but also include the propionate produced by the oxidation of odd- and some methyl-branched chain-fatty acids [13]. As an initial step in our study, we decided to group them (because of their shared metabolic function) as a distinct group of nutrients, which we defined as KCAI (Krebs cycle anaplerotic intermediates) when analyzing energy partition (Figure 1). The KCAI are eventually oxidized via 3C-2C.
Figure 1.
Biochemical (dietary) sources of KCAI: amino acids and some minoritary fatty acids, and their direct implication in the Krebs cycle.
Figure 1.
Biochemical (dietary) sources of KCAI: amino acids and some minoritary fatty acids, and their direct implication in the Krebs cycle.
Hormones and energy partition
Estrogens are a main regulatory factor in the complex mechanism that controls body weight [14,15,16] and justify many sex-related differences in metabolic handling of substrates. Estrogen receptor α (ERα) signaling in LH neurons [17] and VMH neurons plays a critical role for energy partition and thermogenesis [18]. Metabolic control by estrogen is directly related to the availability of testosterone (T) since its aromatization is the main path for 17β-estradiol (E2) synthesis [19]. The availability of E2 is, thus, linked to that of T and. In the metabolic syndrome (MS), one of the most generalized pathologic traits are the low levels –and availability– of T [20], paralleled by those of E2 [21]. The effects of this combined decrease deeply affect the handling of the eventual excess availability of 3C (and, especially, of 2C), with the consequence of 3C aggravating the problem of mounting 2C accumulation, which could not be oxidized in a timely way. This surplus of 2C is then mostly derived to the synthesis of fatty acids (and the ensuing accumulation of TAG) in the liver, blood and elsewhere; with ectopic depots in muscle and, especially in adipose tissue [22]. This is the biochemical basis for the excess of body fat resulting in obesity. The relative resistance to MS of pre-menopause women vs. mature men is a global reminder of the role of estrogens preventing diabetes hyperlipidemia and obesity by enhancing lipid oxidation [23]. The mechanisms explaining these effects of E2 are known: E2 elicits the mitochondrial oxidation of substrates [24], by regulating pyruvate dehydrogenase [12], which is the critical point in the partition of nutrients’ fate at the 3C to 2C point of no-return [25]. These effects are largely mediated by the modulation of insulin function [26], increasing the glucose uptake by muscle [27].
The direct relationship between dietary fatty acids and estrogen availability [28] and signaling is important for the regulation of lipid metabolism at the critical point of postprandial handling of the nutrients assimilated, helping to explain differential effects of diets directly related to their fatty acid nature and content. On the other hand, androgens contribute to maintain glycemic homeostasis, in both sexes through their insulin-like effects in muscle [29], but especially, in their implication on the maintenance of protein synthesis and ‒protection‒ of body protein against its use for energy [30]. The implication of steroid hormones in food intake regulation and glucose homeostasis, could be completed by the additional close interrelationships of androgens and glucocorticoids [19]. Glucocorticoids counter the E2 drive to oxidize 2C and favor lipogenesis and amino acid oxidation [31], including body protein (i.e., antagonizing the function of T), and inducing a marked ‘diabetogenic’ drive for gluconeogenesis. Thus, the balance between glucocorticoids, E2 and T (plus insulin) maintains a control system of liver energy partition which may help to cope with the eventual profound changes in diet supply of nutrients in type, proportion and amount. An additional –important– function of E2 is to regulate amino acid oxidation [2,16,19,32] (in part sharing the glucocorticoid drive, and countering the reverse action induced by insulin and, mainly, by T. This effect is not generalized to the whole body, an important difference with glucocorticoids, and is related to the disposal of excess ‘excess’ energy, elimination of unneeded 2-amino-N and, especially, the supply of amino acid hydrocarbon skeletons (i.e., KCAI) as additional thrust to 2C oxidation thanks to their anaplerotic action on the Krebs cycle [33].
The well-known effects of dietary lipid usually aggravating the excess of 2C induced by excess of energy intake stress the relatively known but unquantified implication of amino acids on energy partition, and on the general regulation of metabolism when considered not for their anabolic potential but as simple energy substrates [34,35]. The prevailing concept of amino acid nitrogen sparing for the preservation of hard to obtain amino acids, protein anabolic precursors [36] has been maintained for many years, but this concept needs additional conceptual correction. First, the differences in the critical sparing of amino acids are not the same for carnivores, herbivores (including ruminants) and the wide spectrum of omnivores [37]. And second, dietary protein is not only a source of amino acids for anabolism and synthesis of nitrogen-containing compounds, but also an important source of energy [38] for ‘omnivores’ such as men (and rats, despite not being the same type of omnivores), questions often not taken in consideration. Dietary protein is needed, also and precisely, for the expedite and effective oxidation of excess carbohydrate (and lipid: i.e., 2C sources) for energy, because in the diet most KCAI come from amino acids. Thus, a delicate equilibrium in dietary protein utilization is established between the need to spare N and essential amino acids for maintenance/ growth/ turnover of body protein, and their use as KCAI suppliers to aid on energy-staple substrate oxidation and, also, for their simple and direct oxidation for energy.
We know little about the transcendence of diet nutrients on the availability of 3C or 2C metabolites and how they affect energy partition. This limited knowledge extends, especially to the profound influence of E2 on glucose metabolism [16,39]. In the present study, we intended to group the wide experience of our group in the study of body weight regulation; and, especially, on the role of steroid hormones regulating the effects of diet composition on energy partition. We analyzed the relationships between energy partition and endogenous E2 and T levels in rats fed diets containing different carbohydrate, lipid and protein content, by using a highly controlled model. We expected that the different relation between 3C, 2C and KCAI availability (i.e., overall assimilation) of diet nutrients should condition energy partition, with E2 being a critical regulatory factor in the whole process.
2. Results
In all shared-origin diet groups, the rats increased their body weights along the 30-day study in a similar way, as previously published [2] (Figure S1), the only exception being the CF group, which showed higher increased rates for both sexes.
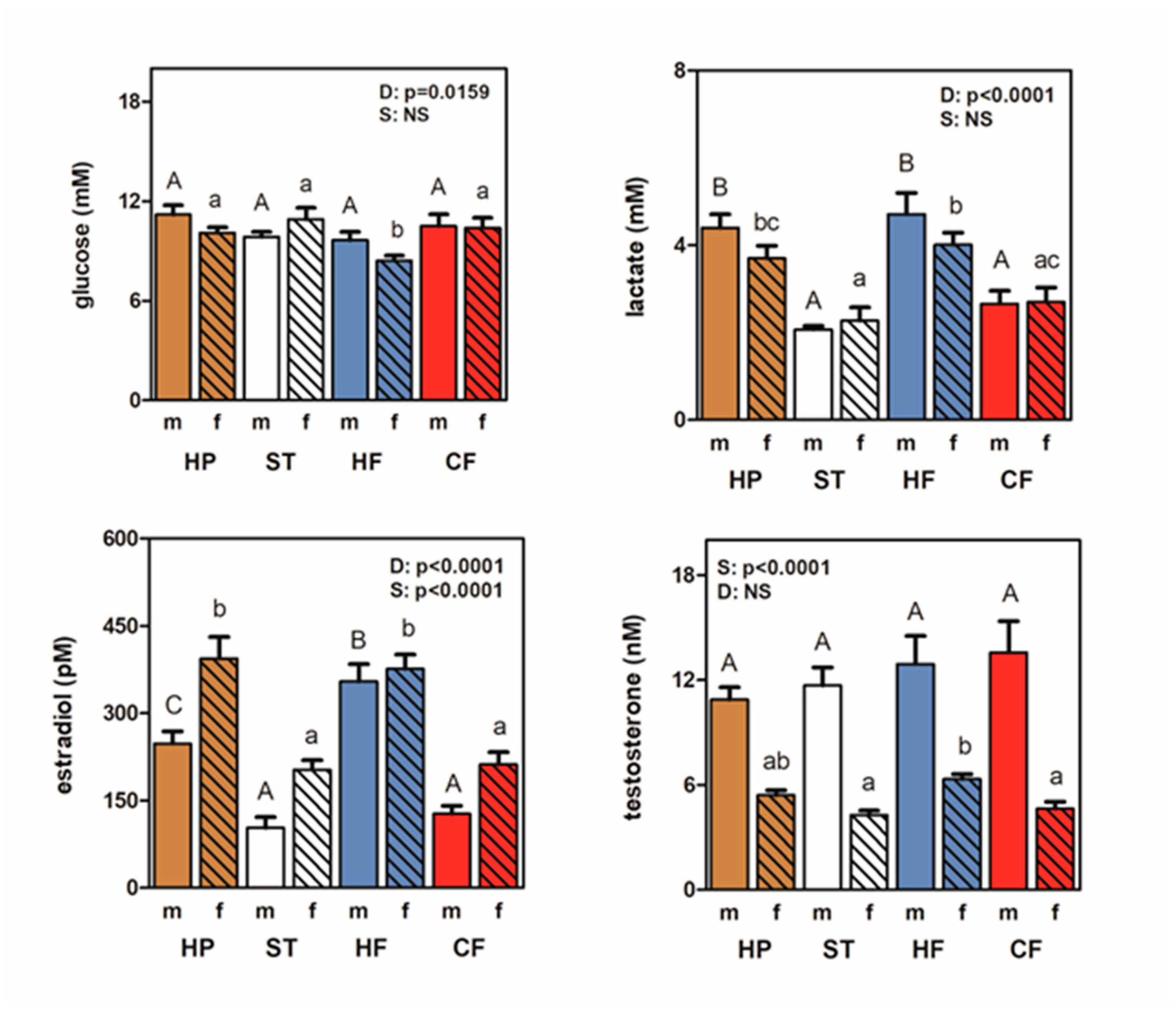
Figure 2 shows that in HF and HP groups, plasma E2 values increased, irrespective of sex; and total plasma glucose was well maintained along the whole study, except for HF females, which showed lower mean values than ST, in parallel with higher plasma testosterone. Plasma lactate was higher in HP and HF groups (but not in CF) compared with ST.
Figure 2.
Metabolite and hormone levels. Male (m, clear columns) and female (f, dashed) values for glucose, lactate, estradiol and testosterone are placed shown for HP (brown), ST (white), HF (blue) and CF (red) groups. Statistical differences between groups: two-way ANOVA (D, diet; S, sex). Bonferroni’s post hoc test: different letters represent statistically significant differences between groups of the same sex; NS: non-significant difference.
Figure 2.
Metabolite and hormone levels. Male (m, clear columns) and female (f, dashed) values for glucose, lactate, estradiol and testosterone are placed shown for HP (brown), ST (white), HF (blue) and CF (red) groups. Statistical differences between groups: two-way ANOVA (D, diet; S, sex). Bonferroni’s post hoc test: different letters represent statistically significant differences between groups of the same sex; NS: non-significant difference.
Plasma lactate was directly correlated with E2 (Figure 3), whereas no significant results were obtained for the correlation between lactate and testosterone levels in plasma (p=0.0994). The direct correlation between L-lactate dehydrogenase (EC 1.1.1.27) activity in liver (LDHa) or muscle (LDHb) vs. plasma lactate became significant when all the groups were considered as a combination of all, especially in muscle.
Figure 3.
Correlation between plasma lactate levels and lactate dehydrogenase in muscle and liver. Male (squares) and female (dots) values from HP (brown), ST (black), HF (green) and CF (red) groups. Black line indicate correlation for all groups.
Figure 3.
Correlation between plasma lactate levels and lactate dehydrogenase in muscle and liver. Male (squares) and female (dots) values from HP (brown), ST (black), HF (green) and CF (red) groups. Black line indicate correlation for all groups.
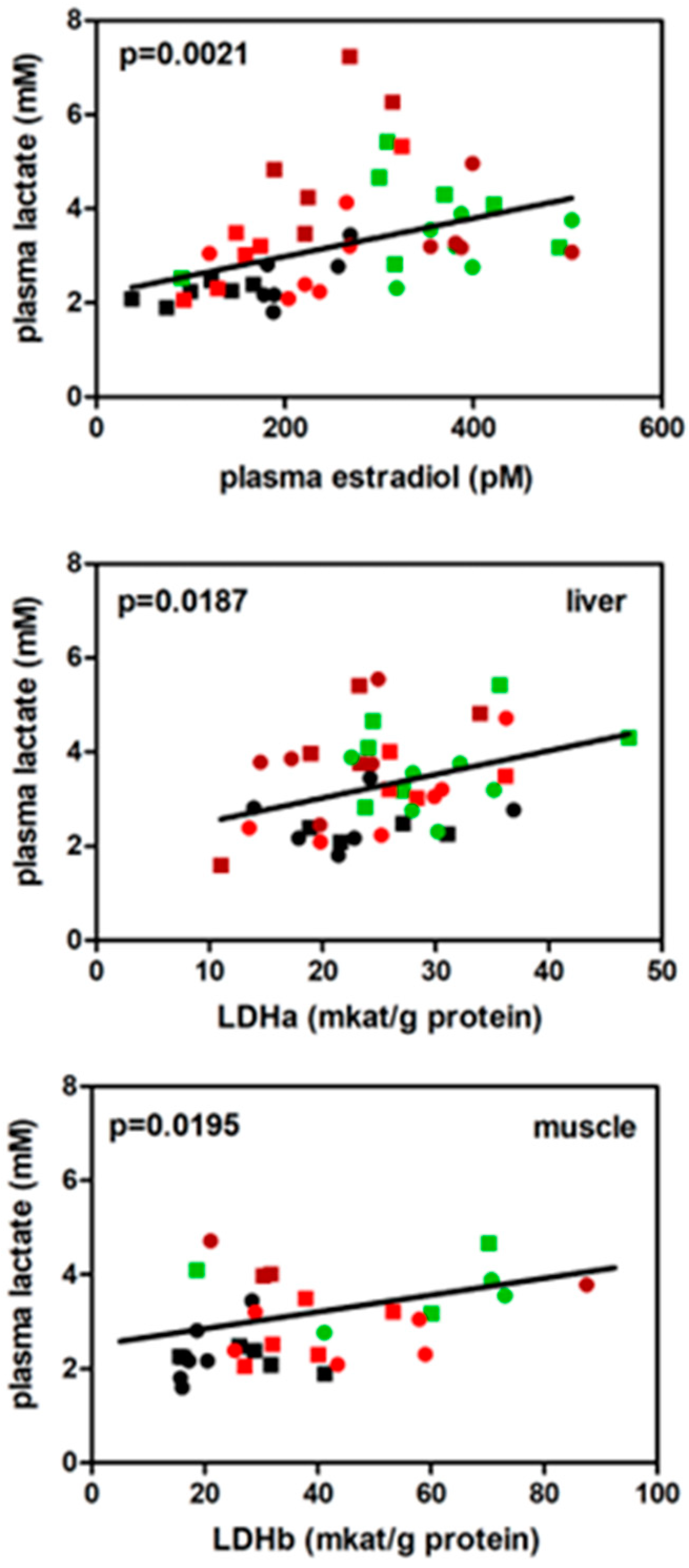
Figure 4A shows the energy ingested from lipids, proteins, and carbohydrates (shown in two separate groups: sugars –mono- and oligosaccharides– and polysaccharides), presented both in absolute and percentage values. All data were expressed as the ratio of nutrient ingested (in W, i.e., in energy units divided by time) with respect to the mean (for the 30-day treatment) body weight (BW) value adjusted with an allometric factor (BW0.75) [56,57] to allow comparisons between homologous parameters in groups of different body sizes. The CF groups again showed the highest absolute values, with relatively higher levels of food intake for males, except for polysaccharides, in which females took the lead. The same pattern was observed when comparing the relative intake, with protein ingested showing a similar percentage of total energy intake for both sexes.
Figure 4.
A. Energy substrate intake from protein (brown), lipid (yellow), sugars (green) and polysaccharides (white), either in stacked absolute values [MJ/kg mean weight0.75](left) and in stacked relative values [% of ingested](right). Values for two-way ANOVA and Bonferroni post-hoc test are indicated in the inserted tables. B. Energy derivable from substrates’ intake 1C (light green), 2C (yellow), 3C (red), 4C+5C (dark blue), 6C polysaccharide (white) and 6C sugars (green), either in stacked absolute values [MJ/kg mean weight0.75](left) and in stacked relative values [% of ingested](right]. Values for two-way ANOVA and Bonferroni post-hoc test are indicated in the tables.
Figure 4.
A. Energy substrate intake from protein (brown), lipid (yellow), sugars (green) and polysaccharides (white), either in stacked absolute values [MJ/kg mean weight0.75](left) and in stacked relative values [% of ingested](right). Values for two-way ANOVA and Bonferroni post-hoc test are indicated in the inserted tables. B. Energy derivable from substrates’ intake 1C (light green), 2C (yellow), 3C (red), 4C+5C (dark blue), 6C polysaccharide (white) and 6C sugars (green), either in stacked absolute values [MJ/kg mean weight0.75](left) and in stacked relative values [% of ingested](right]. Values for two-way ANOVA and Bonferroni post-hoc test are indicated in the tables.
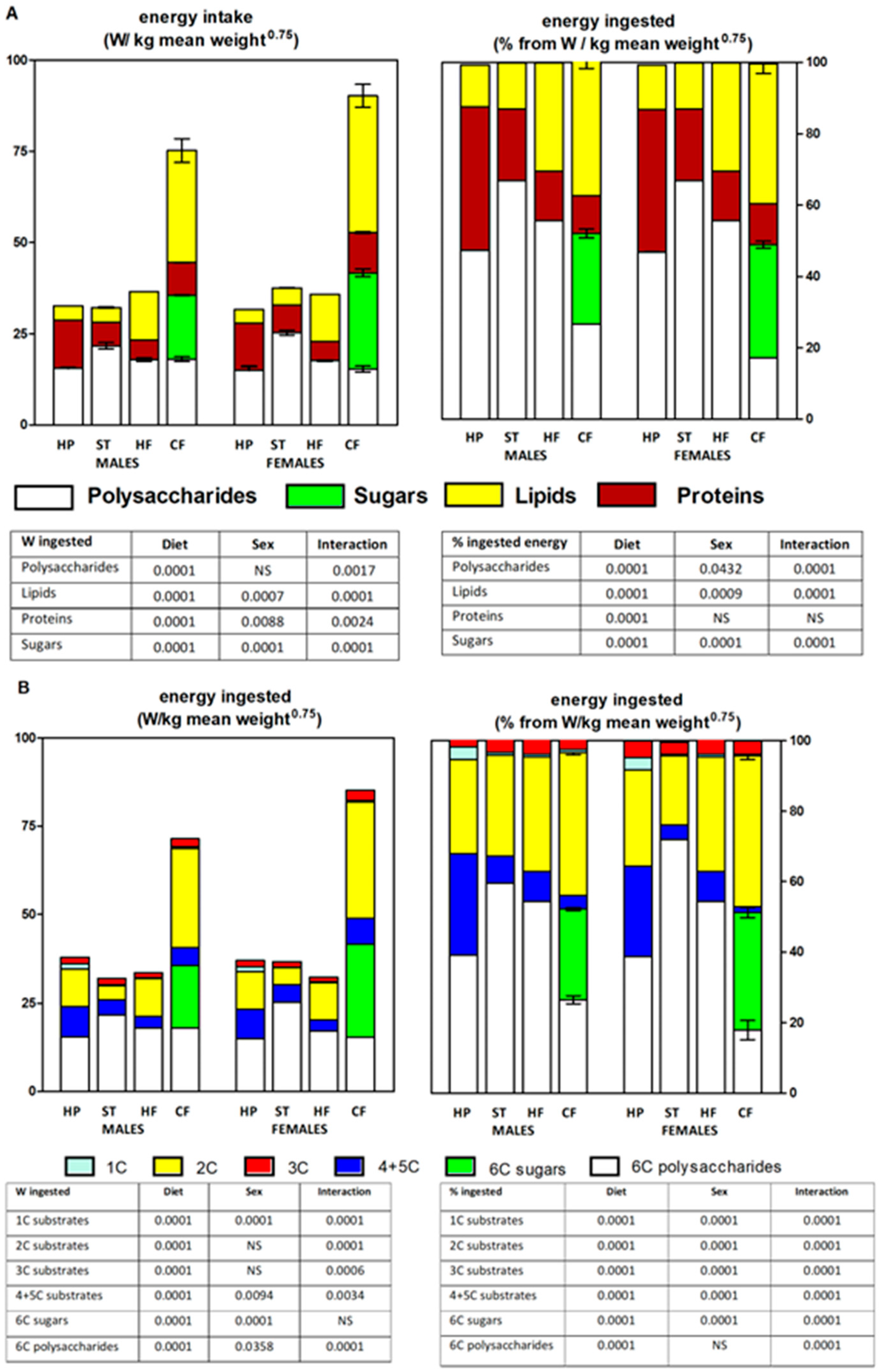
Figure 4B shows the comparative intake of foods yielding fragments of 1C, 2C, 3C, 4C+5C and 6 C, presented, as in Figure 3A, both in absolute and percentage values. The data were also expressed as the ratio of nutrient ingested (in W) with respect to the mean ‒of the 30-day treatment‒ body weight allometrically uniformized (BW0.75). The two CF groups showed the highest global 2C energy intakes, and, obviously, of 6C (sugars); the differences were significant for diet and sex. Fragments of 3C were produced in all groups with no differences between sexes. There was a sizeable production of 1C fragments (of no specified important energy value) in the two HP groups, which also showed a high generation of 4C+5C fragments (KCAI). The 6C units constituted, overall, the main energy nutrient ingested, in percentage of intake; the lowest 6C values were found in the HP groups, and the highest in the ST groups.
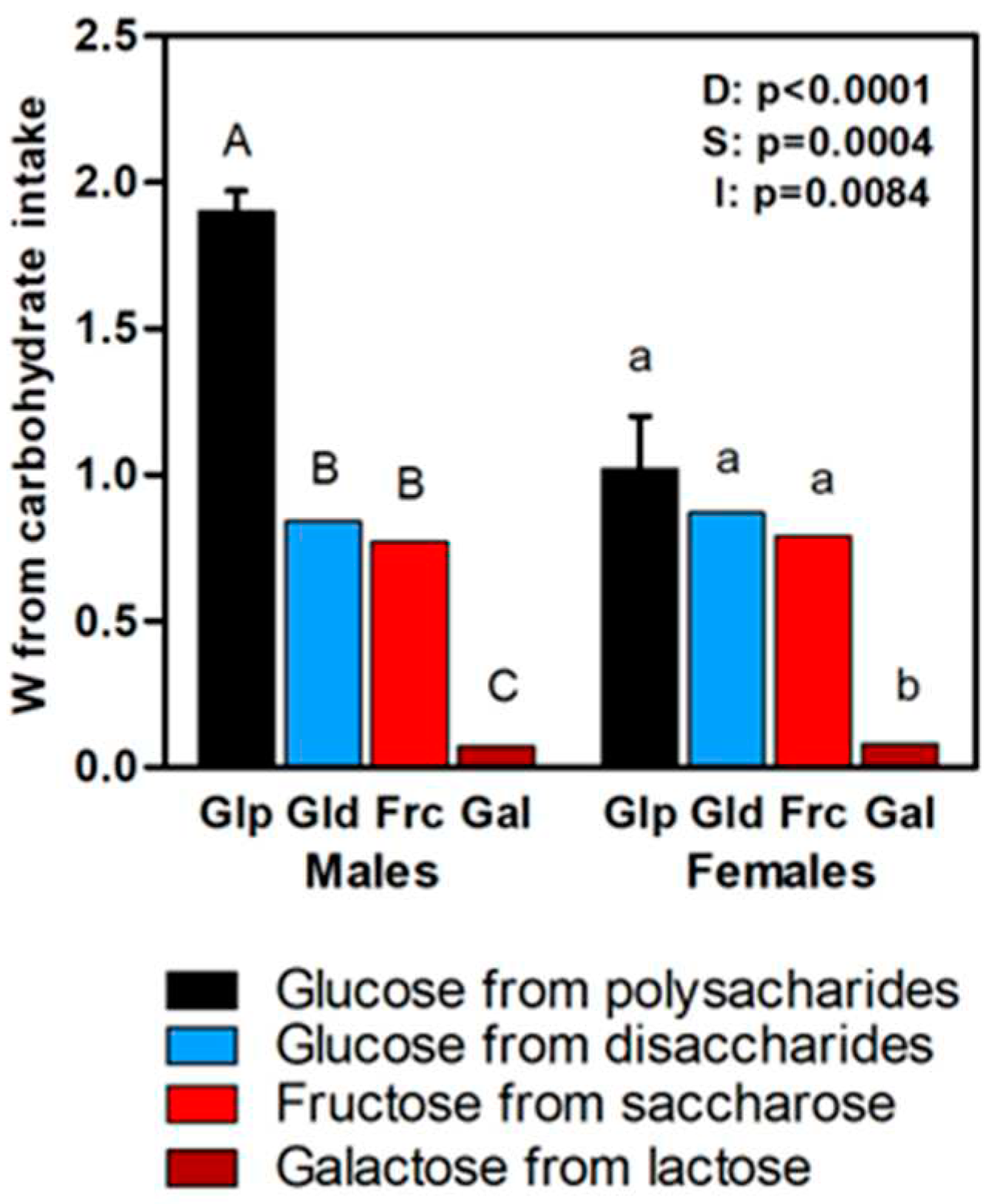
Figure 5 depicts the data for 6C intake (largely glucose) in the CF groups, and showed higher values for polysaccharide origin glucose intake in males, whereas in females these values were like those derived from disaccharides. Galactose (derived from lactose) values represented a small proportion of energy compared with those of poly- and disaccharides, but was a nutrient exclusive –in this study– of the CF diet. Fructose (derived from sucrose) represented a substantial part of the 6C fraction of the CF diet, especially in the females.
Figure 5.
Energy derived from carbohydrate intake in CF groups: black bar= glucose from polysaccharides; blue bar = glucose from disaccharides; red bar = fructose from disaccharides; brown bar= galactose from disaccharides. Values for two-way ANOVA: D (diet), S(sex) and I (interaction).
Figure 5.
Energy derived from carbohydrate intake in CF groups: black bar= glucose from polysaccharides; blue bar = glucose from disaccharides; red bar = fructose from disaccharides; brown bar= galactose from disaccharides. Values for two-way ANOVA: D (diet), S(sex) and I (interaction).
The type of diet clearly influenced the energy accrual in the body, in the form of molecules able of primarily yielding 3C or 2C. Figure 6 shows that in all cases, the mass of 2C fragments deposited in the body carcass was 10 to 20-fold higher than those corresponding to 3C; most of the 2C corresponding to fatty acids and the 3C to amino acids and glycerides-glycerol. The amount of energy deposited was rather uniform between groups when adjusted by the corrected mean body weight. Therefore, the efficiency of deposition was 30-80 times higher for 2C fragments as such than for the 3C. The highest rates of efficiency for 2C storage were detected in the groups receiving the ST diet, while the lowest values.were for HP and CF diets.. The lowest rates of 3C “efficiency” (i.e., deposition vs. intake) were those of HP groups.
Figure 6.
Energy deposited in form of 3C or 2C units in absolute values [W/kg mean weight0.75] (upper row) and as a percentage of efficiency (lower row). Male (m, clear columns) and female (f, dashed) values for HP (brown), ST (white), HF (blue) and CF (red) groups. Statistical differences between groups: two-way ANOVA (D, diet; S, sex). Bonferroni’s post hoc test: different letters represent statistically significant differences between groups of the same sex. A green horizontal dashed line indicates the 100 % of 2C accrued efficiency.
Figure 6.
Energy deposited in form of 3C or 2C units in absolute values [W/kg mean weight0.75] (upper row) and as a percentage of efficiency (lower row). Male (m, clear columns) and female (f, dashed) values for HP (brown), ST (white), HF (blue) and CF (red) groups. Statistical differences between groups: two-way ANOVA (D, diet; S, sex). Bonferroni’s post hoc test: different letters represent statistically significant differences between groups of the same sex. A green horizontal dashed line indicates the 100 % of 2C accrued efficiency.

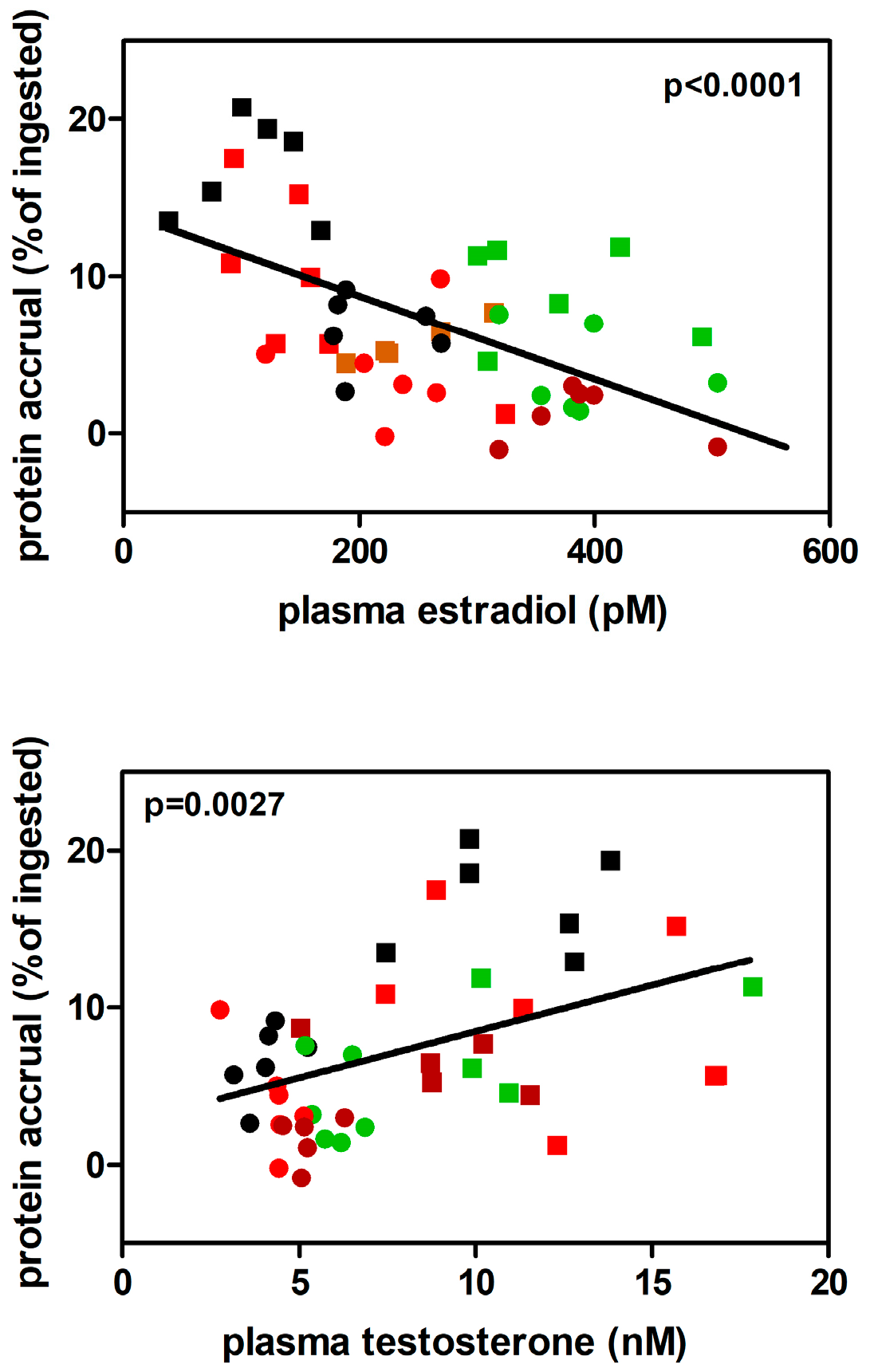
Nitrogen (massively protein) accrual correlates directly with T levels and inversely with E2 values, as shown in Figure 7 when all dietary treatments were included, suggesting that the correlations were rather independent of the nutrients-mix in the different groups. In short, T favored the accrual of protein, while E2 diminished it.
Figure 7.
Correlation between protein accrual (expressed as % of ingested protein) and plasma T or E2 levels. Males (squares) and females (dots) values takenfrom HP (brown), ST (black), HF (green) and CF (red) groups.
Figure 7.
Correlation between protein accrual (expressed as % of ingested protein) and plasma T or E2 levels. Males (squares) and females (dots) values takenfrom HP (brown), ST (black), HF (green) and CF (red) groups.
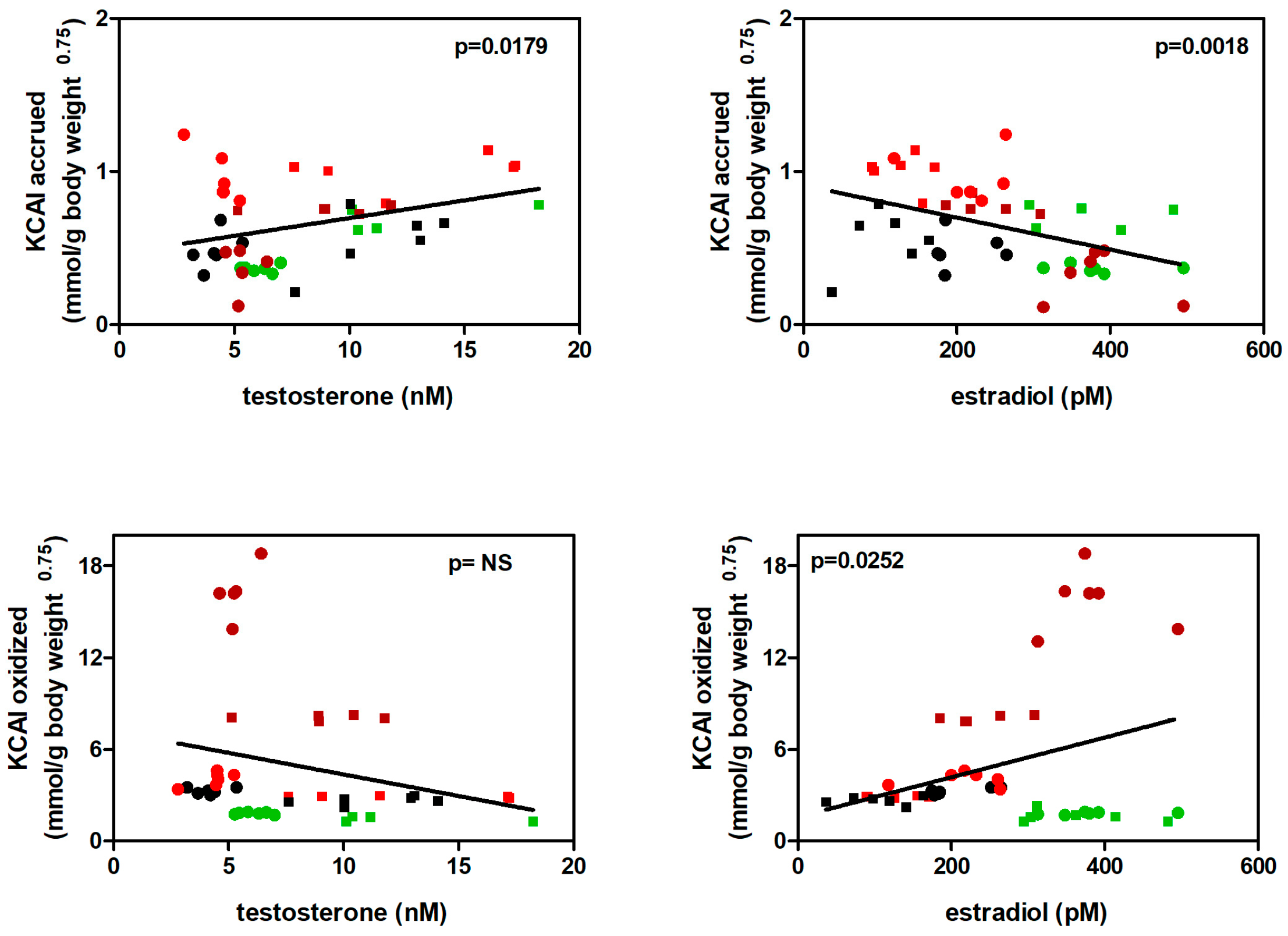
In parallel, the amount of KCAI accrued was directly correlated with circulating T levels, and inversely with those of E2 (Figure 8). Accrual of KCAI can be also equated (practically) to the deposition of amino acids (protein), and thus coincides with the data on accrual of nitrogen. Conversely, the KCAI oxidized tended to correlate inversely with T levels and directly with E2, in line with the information from accrual data, and showing that the fate of diet amino acids (at least the majority yielding KCAI) was unbalanced towards oxidation by E2 and to preservation/ accrual by T.
Figure 8.
Correlation between KCAI accrued or oxidized in front of plasma levels of T or E2. Males (squares) and females (dots); values taken from HP (brown), ST (black), HF (green) and CF (red) groups.
Figure 8.
Correlation between KCAI accrued or oxidized in front of plasma levels of T or E2. Males (squares) and females (dots); values taken from HP (brown), ST (black), HF (green) and CF (red) groups.
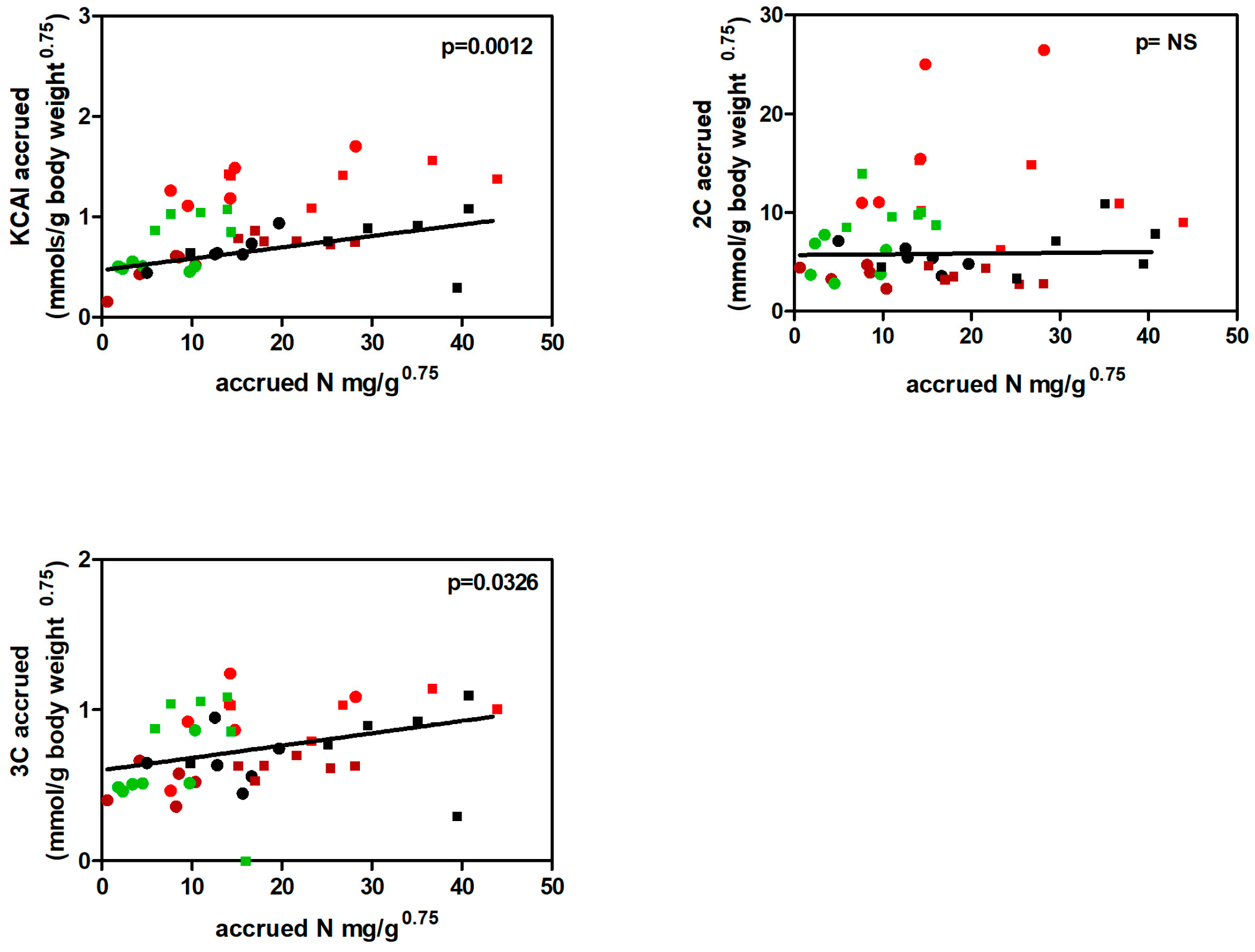
On the other hand, the data for 3C and KCAI fragments deposited were directly correlated with the amount of accrued N, whereas the correlation was lost with 2C fragments (Figure 9). The main difference between 3C and KCAI as to deposition being that the latter represented practically only amino acids, whereas the 3C was shared by amino acids and glycerides-derived glycerol.
Figure 9.
Correlation plots between protein (amino acid)-related indicators: KCAI or 3C or 2C accrued vs. accrued N. Males (squares) and females (dots); values taken from HP (brown), ST (black), HF (green) and CF (red) groups.
Figure 9.
Correlation plots between protein (amino acid)-related indicators: KCAI or 3C or 2C accrued vs. accrued N. Males (squares) and females (dots); values taken from HP (brown), ST (black), HF (green) and CF (red) groups.
3. Discussion
Studying the differential effects of diets of different food and nutrient composition has been (and remains) an almost impossible task due to the enormous variability of the foods consumed, both in humans and in most animal species in the wild. Every change represents a factor that needs to be either analyzed or adequately shown not to compromise the other results, which is even more difficult to achieve. The differences in protein or lipids, and the varying textures, palatability and, especially, nutrient composition makes difficult to isolate specific factors in order to establish their eventual overall (and specific) influence on health and disease [58]. In the present study, we used a previously tested model of standard-chow-derived high fat HF and high protein diets HP [2,42] compared to the original “standard” food ST from which they were derived, and also including a simplified cafeteria diet that we have used previously ([40,45] as a contrasting (and disrupting) factor, in order to be able to establish additional comparisons between the ‘shared-origin’ diets. Therefore, we used rats and diets as similar as possible to our long series of studies, with also comparable rat strain, age (size) and sex [7,40,46]. To establish a way to further compare animals of different sizes, and help ‘discount’ the allometric effect of body weight [57], we standardized the group data for body mass and diet intake, arbitrarily referred to the half-point of dietary treatment (day 15). This way we calculated mean day-15 values, for every group, of the rate of body composition, growth and the amount of each type of nutrient ingested per day. Since we introduced the factor of time in the comparisons we had, then, to express energy as energy rates (power units, W). These data were made comparable by correcting them by the allometrically adjusted data, as described previously [59,60] and widely used in comparative physiology when studying series with individuals of varying size.
As expected, the CF diet induced high intakes of energy (lipid and carbohydrate, including a large proportion of disaccharides), but total protein intake was comparable to that of ST, and the lipid ingestion was comparable to that of HF when the data were made homogeneous. Nevertheless, the carbohydrate proportion of energy intake was higher in CF than in all the other diets analyzed, although the sum of all carbohydrate-derivable energy (as a proportion of energy intake, not in comparable absolute terms) was similar for all groups.
In any case, the fact that the efficiency of lipid accrual (essentially as 2C) was higher for ST diet groups, and was in a comparable range for HP, CF and HF diets, clearly indicates that changes in the proportions of lipid and protein –taken irrespective of the rest of the diet– actually affect energy partition. Lipids being used in a significant proportion for energy, especially in liver [61], as previously proposed by our group [2] and confirmed again with the results presented here. The effect in HP and HF groups, however, can be attributed to higher circulating levels of E2.
Our data confirm that, irrespective of the type of diet used, testosterone tends to correlate with protein accrual, whereas E2 (mostly derived from testosterone via aromatization) correlated with oxidation of energy substrates, including the massive amino acid utilization by liver, especially in females [62]. Since amino acids are an important fraction of potential 3C substrates, the accrual of 3C was largely justified with the accrual of amino acids (but also by a smaller but significant part of 3C from glycerol in the deposition of TAG). The parallel deposition of KCAI was –again– related to amino acid deposition and both (3C and KCAI) again correlated, as expected, with total N accrual.
The diet comparison scheme we designed allows us to relativize the effects of both ingested lipid and protein as main inducers of the CF-increased food intake when compared with the three shared-origin diets, even when their lipid or protein proportions approaches those found in the CF groups. The remaining main bulk nutrient (making an absolute difference between CF and the other diets) being, essentially, the quite different intakes of 6C as [disaccharides and starch] in CF vs. [starch alone] in the ST-derived diets. The effects of comparable intakes of 6C containing (or not) a large proportion of readily assimilated disaccharides resulted in a higher energy intake (appetite) magnified in carbohydrates and lipids [63], and their derivation towards 2C storage (fat) in CF when compared with oxidation as main fate in the ST-related diets. Sucrose is known to increase food intake in humans and rats [64] and is a powerful energy partition destabilizer via disruption of glycolytic handling of glucose and its control by insulin [65].
Our data did not show deviations in food intake rates, within a same group along the 30-day period of study (Supplemental Figure 2). However, the differences between groups were patent. We can assume that the rapid absorption of disaccharides (already hydrolyzed to their constituting monosaccharides: glucose, fructose, and galactose) represent an immediate energy insult to the already high acute flow of glucose; fructose and galactose being also rapidly released into the intestine (and then absorbed). This bulk dumping of 6C induces a rise in glycaemia necessarily paralleled by a rise in insulin, which elicits hepatic insulin resistance [66]. The liver responds as best as possible to 6C overload, but it can do so more comfortably when the flow of 2C energy to CO2 may be enhanced by E2, and by the presence of KCAI [62], thus progressively lowering the load of excess 6C (finally 2C). The slower pace of release of glucose from the digestion of starch resulted in a higher proportion of their energy being used for oxidative energy rather than TAG deposition. This difference agrees with the critical role of postprandial energy partition in the liver [67] in addition to the disrupting effect of high glycemic index nutrients such as sucrose. Sex deeply influences this effect, with females being more resistant to the deleterious effect of excess fructose [68] than males, and they are –also– better adapted to carbohydrate (6C) loads than males. The results of the present study (Figure 5) agree with these postulates, since the load of 6C in CF male rats was (as % of total energy intake) 53% starch-derived glucose, 23% disaccharide-derived glucose, 22% fructose and 2.5% galactose. Whereas in female rats the intake of fructose and galactose was similar, disaccharide glucose almost represented the same amount than polysaccharide glucose; in all, the females proportionally accrued less energy than the males.
Nevertheless, the excess of glucose and the dumping of disaccharide-6C was not reflected into glycaemia a condition already described in the literature [69], which clearly indicates that the CF rats were able to dispose of all 6C substrates in real time without reverting to sustained hyperglycemia. The disposal of this huge amount of carbohydrate energy can occur only if the turnover/ metabolism of 6C (and its main intermediate metabolites, 3C) was highly increased with respect to the shared-origin diets. This anomalous ability to handling such excess energy cannot be (easily) justified by increased appetite (i.e., one of the effects of fructose), but may be –instead– a possible consequence of the huge intake of fructose and galactose. Fructose in excess does not follow the canonic glycolytic pathway [70] and their effects are more related to the complementary glycolysis integration pathways used. The case for galactose is quite different, since its oxidation is slow (in adults being mainly excreted unaltered or used by the microbiota), because the conditions of operation of the Leloir pathway are far from being optimal in a liver under high-load exposure to 6C [71]. In fact, galactose may become hepatotoxic [72], affecting the whole carbohydrate metabolism-regulation by insulin and intestinal peptides. This is a question that has not been discusses in depth, despite the presence of lactose in most cafeteria diets (and human Westernized diets [73]); the focus on dietary perils being mostly fixed on fructose, lipids and the overwhelming excess of energy. The distortion provoked by the ingestion of sugars yielding fructose and galactose in the CF groups, seems to alter the straightforward mechanism of dietary lipid (and carbohydrate) oxidation already observed in many HF diet rats. It must be stated, too, that the type of diet ‘biologically optimized’ for rat experimentation is quite different from that of rats in the wild and from those of humans, which have been more widely exposed to sugars different from polysaccharide-derived glucose in early periods of human evolution.
The fast rise of glucose (from sugars) necessarily spurns the conversion of increasing 3C to 2C thus limiting the near future maintenance of glycaemia, altering the rate of 3C oxidation and promoting the accumulation of lipid reserves. Nevertheless, using our model of shared-origin diets, we were able to present further evidence that rats fed the well-established chow pellets’ compositions use (need?) starch as the main energy staple [74]. Sugars as such being both uncommon and unnecessary in the adult diets, despite galactose being the only carbohydrate available in the diet during mammal suckling [71], defining the immediate post-natal period as a high-lipid high-protein high-oligosaccharide fairly ketogenic and markedly obesogenic diet (for a time) of unparalleled successful anabolic efficiency [75]. In addition, a sustained heavy load of dietary lipids must be combined with even larger sources of 3C to not only sustain energy needs, but to maintain glycemia and the synthesis of TAG (at least its glycerol moiety). Since the need to maintain glycaemia is critical for humans and rodents, we postulate that the maintenance of energy partition requires the inescapable ingestion of hexoses (or enough 3C). The best way to using them appears to be in the form of hydrolysable polysaccharides, since the slow pace of release of glucose from starchy foods (better with low glycemic index) does not elicit the inadequately massive metabolic (often pathogenic) responses such as those described for CF diets [69]. The effect of lipid as inducer of lipid accrual in rat diets probably can be more related to the additional presence of sugars or other factors (such as sodium) [9], than to the total energy availability [1]. And, even more probably to the different testosterone-E2 availability and equilibrium as main co-controllers, with insulin, of energy partition.
The extended use of diets low in carbohydrate (e.g., ketogenic) are supposed to favor the oxidation of lipids, but –instead– they accelerate the breakup of proteins and their amino acids for 3C (and 4C or 5C) fragments [76], eventually metabolized to 3C to provide substrates for gluconeogenesis or generate more 2C for energy. But this phenomenon of enhanced oxidation of 2C may have another (complementary?) interpretation: as postulated in this study, the accelerated catabolism of amino acids, potentiated by E2, may produce large amounts of KCAI, which act synergistically with E2 promoting the mitochondrial oxidation of 2C, instead of allowing the obesogenic effect provoked by the insufficient flow of carbon though the Krebs cycle. This interpretation may conflict with data that assume that estrogen control of liver amino acids [32] is constricted to of amino acid availability for protein synthesis. Our interpretation, stresses the need of a regular source of amino acids (or, alternatively, metabolize body proteins to obtain gluconeogenic substrates and/ or KCAI) in the diet further to its N-protein providing specific role. However, the main negative consequence is the need of disposing of the (unneeded, eventually toxic) N-waste, breaking the ‘need to spare N’ rule that has driven most of the nutritional analyses of N needs up to date, based on starvation and malnutrition [77], and far from the overeating and maladjustments of present day in large groups of population. The tight urea cycle constrictions are perfect for N conservation [78] as the use of the hydrocarbon skeleton of (scarce) amino acids requires a ready to access mechanism for controlled disposal of the excess N. For a rat (or human) it is metabolically more efficient (limits the need to excrete so much N) to ingest only the amino acids needed, and use the most common plant reserve, glucose, in the natural ‘second most common form’, starch. This type of polysaccharide has a principal place in our diet, being modulated and presented in different ways, but which is eventually translated into a slowly releasing source of 6C, but also provider of 3C (i.e., L-lactate, a critical energy substrate) used by most body tissues [79], which may also be easily converted into 2C for energy.
The data presented agree with this need to maintain an ‘adequate’ supply of polysaccharide-derived glucose for energy homeostasis, using fatty acids (as in HF) or protein (as in HP) as complementary energy substrates. The ‘rule of metabolic substrate oxidation priorities’ (essentially designed for the management of body reserves under starvation), compels first to oxidize fatty acids and exhaust the glycogen-glucose (if any remaining), and only then began using for energy our protein amino acids [80]. However, this set of priorities could not be directly applied to situations with excess energy available, quite different from the opposite and more common case of insufficient nutrient availability and neither to the management of external (dietary) sources. Thus, in many cases, protein becomes just another (main) source of energy [81] for humans, as in carnivores, to be used with the only limitations established by the tight protection of amino-N loss via the urea cycle, essentially designed for survival under low amino-N (and energy) availability [78]. However, the loss of excess N via an alternative pathway has been proven [40,82] and is related to the amount of energy available. It has been found that this N is lost as N2 [82], and the possible implication of mitochondria oxidative capabilities has been hinted at but not fully uncovered. This ‘alternative’ system has received little attention because of the intrinsic difficulties of measuring the emission of N2 by animals breathing air containing, already, more than 70% of this same gas. Nevertheless the measurement of a complete N balance (including urinary and fecal losses, and N accretion) repeatedly results in a N gap [40,83], process that has been also found to be related to arginine [84] and enhanced by E2 [2].
The increase of circulating E2 in HF and HP groups, is concurrent with maintained levels of testosterone (main precursor of E2) and occurs with practically unaltered basal glucose levels. Testosterone, differently from E2, seems not to respond to gross changes in the rat diet composition (at least during one-month treatment), as observed in this study. The decrease in the intake of 3C metabolite providers, irrespective from their original nutrient form, seems to be a main factor conditioning the observed increase of E2. This increase may reflect a situation opposed to the decrease in circulating estrogens described for rats fed high-fat diets [85] and also rich in salt and oligosaccharides [9]. In fact, we did not observe a decrease in E2 even in the CF rats, which suggests that the modulation of E2 levels by diet components needs further analyses, especially in cafeteria and Western-style diets.
In the liver, the excess of glycolysis-produced pyruvate is directed to produce lactate, when gluconeogenesis is not activated. When glucose availability coincides with excess lipids (as in the HF groups), most 3C fragments are derived to the peripheral use of lactate as substrate [79]. This process does not waste 3C energy in the liver since lactate is oxidized (to 2C and then to CO2), protractedly, in other tissues. This source of 3C may be –also—recycled to glucose, glycerol, alanine, etc. This distribution of 3C fragments for energy production elsewhere has the advantage of not directly affecting glycemia and circumventing the problems posed by the high reducing potential and isoforms of glucose, despite its effect on the blood pH equilibrium and the inter-organ handling of protons [86]. In any case, the 3C outlet of ‘predigested-glucose’ is efficient and functional. Under conditions of high 3C/2C availability ratio in the liver, oxidation of pyruvate (3C) to acetyl-CoA (2C) is activated for energy [87] or eventual lipogenesis [88], as obvious dump of excess energy accumulation.
The critical effects of E2 on the modulation of liver energy partition can be summarized in three processes:
Excess glucose may result, thus, in higher availability of 3C. However, pyruvate cannot be used for lipogenesis, since E2 downregulates hepatic lipogenesis [93], regulating the transcription of lipogenic genes through transcription factor XBP1 [94]; leaving open only the way to mitochondrial oxidation for energy and/or export as fatty acids (enhanced TAG turnover and transport), enhanced ketogenesis and activation of the mevalonate pathway. E2 indirect activation of β-oxidation and the concurrence of KCAI, higher oxidative mode and use of energy by the liver (processes in which E2 acts as enhancer or activator), allow the complete conversion of excess 2C carbon into energy (NADH+, ATP) and heat, mainly in the liver.
One of the consequences of higher plasma E2 in the HP and HF groups (compared with ST) is the higher circulating lactate levels, directly correlated with E2. In addition, lactate was correlated both with lactate dehydrogenase activities in liver and in the largest peripheral tissue, muscle, showing an intimate relationship with energy partition [1]. The existence of different isozymes that play towards the production or oxidation of lactate depending on its levels and supply, a reflection of the 3C energy availability, in liver or muscle [95] adds to its importance as dynamic marker of this same energy partition as a main form of carbohydrate-derived energy substrate. In any case, increased lactate levels are caused by an imbalance between its synthesis and uptake/ oxidation; but this is not only a function restricted to the liver, implying other potential massive lactate producers (from glucose) such as muscle and adipose tissue [96]. Circulating lactate levels respond not only to the liver-muscle relationships (as in the Cori cycle), but in some way, indicate that the pair 6C-3C (i.e glucose and mostly lactate) is sufficiently maintained, so part of the protected 3C can be converted to 2C and oxidized with the additional advantage of the ready to use metabolites for tissues circumventing the tight and often cumbersome mechanisms of control of glycemia. Evidently, E2 (in fact in cooperation with –and through— insulin) maintain the 3C homeostasis (and hence that of 6C) and allows its oxidation to 2C only when (and if) it is present in excess (as explained above) and the maintenance of basal glycemia is not threatened. This raised lactate levels are comparable to the described upregulation caused by E2 in humans with type 2 diabetes [97], although in this case, the diet, and not the glycemic status, is the main factor determining the lactate changes.
In our model, testosterone levels do not seem to downregulate lactate levels, as described in diabetic humans [97]. But high polyunsaturated fatty acid levels (mainly linolenic acid) in ST and HP diets did not affect –either— the levels of testosterone, which conflicts with reported data in humans [98], in which testosterone is a main factor in the maintenance of glycaemia [22].
In general terms, we have observed that 3C derived from non-carbohydrate ingested nutrients (as found in the HF and HP groups) promote the active use of other substrates for energy, including fatty acids and –especially-- amino acids in addition to the bulk of energy in the form of 6C carbohydrates. This was, in part, expected, but the twist of the complementary oxidation for energy of amino acids also facilitated the complete oxidation of 6C, circumventing the precarious solution of the lipogenic derivation of 2C, which eventually promotes obesity and other disorders in the context of the metabolic syndrome (MS).
Thus, in the HF group, fatty acids were actively oxidized to 2C to obtain energy (irrespective of sex), sparing the 3C providers, in shorter supply than in ST. In the HP group, the potentially ketogenic amino acids in excess (especially Leu and Lys) are probably used as source of 2C in significant amounts since these amino acids represent ca. 25% of total ingested amino acids [34]. At the same time, they become a main source of KCAI which help oxidize the excess 2C production from amino acids and 6C. Thus, both ‘supplemented’ diets can produce more 2C metabolites for energy than ST, sparing part of the 3C (lactate) for this purpose and nevertheless efficiently oxidizing most of dietary carbohydrate.
In sum, this study has given further support to the negative effect of large excesses of disaccharides (CF diet) on the tight control of food intake in the rats, favoring a disordered (and excessive) accrual of energy, mainly lipids. The data presented agree with the need (at least for rats, but the corresponding figures for humans could not be too far away) to provide a 50% of the diet energy needs as starch carbohydrates to maintain a well-preserved energy partition. An excessive proportion of carbohydrates in the diet may allow for a faster and more effective anabolic drive, but this includes an (usually) unsought for lipid deposition. The oxidation of protein as energy in the diet helps streamline and adequately use the main load of carbohydrate, a question long time known for which we offer here a plausible experimental analysis based on the transcendence of the KCAI as dietary component and the already explicited critical role of E2 as a main energy partition controller.
4. Materials and Methods
4.1. Animals and experimental setup
All animal-related procedures and the experimental setup were carried out in accordance with the animal handling guidelines of the Catalan, Spanish and European Authorities. The Committee on Animal Experimentation of the University of Barcelona specifically authorized the procedures used (# DAAM 6911).
Age-matched ten-week-old female (initial weight 233±8 g) and male (initial weight 364±12 g) Wistar rats (Janvier, Le-Genest-Saint-Isle, France) were used (N=52). The animals were randomly divided in four groups (n= 6-8 each) for each sex, and were fed ad libitum for 30 days:
1) ST group; fed the standard diet.
2) HF (high-fat) group; fed the basic standard diet containing added coconut oil.
3) HP (high-protein) group; fed the basic standard diet containing added proteins (gelatin + casein).
All animals had free access to water, and were housed (in same-sex pairs) in solid-bottom cages with wooden shards as bedding material. They were kept in a controlled environment (lights on from 08:00 to 20:00) temperature 21.5-22.5ºC, and 50-60% relative humidity). Body weight (for each rat) and food consumption (for each cage) were recorded daily. The calculation of ingested food in rats fed the cafeteria diet was done, as previously described, from the differences in the weight of the food offered and debris left (corrected for exposure dehydration) [41].
A detailed additional explanation of the procedures used (and complementary information) about methodology and the calculations’ model used is shown in the Appendix A data files.
4.2. Diets
The preparation and composition of the ‘shared origin’ diets (ST, HP, HL) used has been previously described in detail [42] (Table S1). The standard diet (ST), Teklad 2014 (Teklad diets, Madison WI, USA), essentially contained plant-derived foods: 20% of digestible energy derived from protein, 13% from lipids, and 67% from carbohydrates (largely starches, but including 10% oligosaccharides). The derived diets: high-fat (HF) and high-protein diets (HP) were prepared using the standard chow as base and main component, as previously described [43], constituting a group of three ‘shared origin’ diets, in which all components were given in the same relative proportion between them except for lipid in HF and protein in HP, which were at higher proportions than the rest. The HF diet contained 14% of digestible energy derived from protein, 37% from lipids, and 49% from carbohydrates, whereas in the HP diet, the energy derived from proteins was 41%, that from lipids 12% and 47% from carbohydrates.
The simplified cafeteria (CF) diet was quite different, formed by a free-access offer of the standard chow pellets, plain cookies spread with liver pâté, raw bacon, water and standard whole cow’s milk supplemented with 300 g/L sucrose and 30 g/L of a mineral and vitamin supplement [40]. All components were kept fresh (i.e., renewed daily). From the analysis of diet components and the ingested items, we calculated that for both sexes, about 40% of the energy ingested was derived from lipids, 12% from protein, and 48% from carbohydrates (including 20% from oligosaccharides, largely sucrose and lactose) [2]. More details related on energy determination are presented in Supplemental Material [44].
The ST, HF and HP diets were offered in the form of dry extruded pellets, and were well accepted (i.e., no aversion effects) by the animals [42]. The CF components were presented in a small accessible container (fortified milk was available in a drinking bottle hung in parallel to that of water). Diet’s fatty acid and amino acid composition and intake are described in Tables S2 to S5. The ST and HP diets contained 60% polyunsaturated fatty acids, whereas HL and CF diets contained 5% and 12%, respectively. Protein derived energy was 12% in CF, 19% in ST, and 40% in HP.
To facilitate comparisons between animal groups, substrates and mass/energy, the data for diet composition and dietary components eaten is presented in energy values (kJ/MJ), or power (W) when corrected by time (more details in the Appendix). Amino acid protein content and analysis [45,46,47,48], as well as lipid analyses and quantification [49,50] are described in Appendix.
Both, crude (cr) and digestible (dg) energy values were higher in the HF diet (cr 18.8 kJ/g; dg 14.6 kJ/g), since its energy density (kJ/g of food) was also higher than those of ST (cr 16.5 KJ/g; dg 12.1 KJ/g), HP (cr 17.4 KJ/g; dg 12.4 KJ/g) and CF (cr 12.4 KJ/g; dg 12 KJ/g) diets. The cafeteria diet had the lowest crude energy density value because of its low content of fiber and the abundant milk intake (i.e., including its zero-energy water content), although its digestible energy is akin to that of the standard diet.
4.3. Experimental procedure
After 30 days of exposure to the diet, at the beginning of light cycle, the rats were anesthetized with isoflurane and blood was withdrawn with dry-heparinized syringes through the exposed aorta, until euthanasi by exsanguination euthanasia. Plasma was obtained by centrifugation and kept at -20ºC until processed. Liver and hind leg muscle samples were extracted, immediately frozen in liquid nitrogen, and stored at -20ºC.
4.4. Analytical procedures
Plasma testosterone and 17β-estradiol were determined using ELISA kits EIA1559 and EIA2693 (DRG International, Marburg, Germany). Total plasma glucose (i.e., mixtures of α and β epimers) was measured with a glucose oxidase kit #11504 (Biosystems, Barcelona, Catalonia, Spain) supplemented with mutarotase (0.5 mkat/mL of reagent) (Calzyme, San Luis Obispo, CA, USA) [51,52]. L-Lactate was measured with kit #1001330 (Spinreact, Sant Esteve d'en Bas, Catalonia, Spain).
Samples of frozen liver and muscle (30-50mg) were powdered under liquid nitrogen. Total lactate dehydrogenase activity in liver and muscle was estimated from these samples as previously described [53].
4.5. Data homogenization. Calculations
Since daily food consumption was measured, the intake of the different nutrients was calculated from their contents in the diet. Glucose, non-ketogenic amino acids [54] and acylglycerol-derived glycerol were assumed to initially provide 3C fragments through major catabolic pathways, whereas the fatty acids from acylglycerols (which represent more than 98% of all diets’ lipid) and ketogenic amino acids were considered to yield 2C substrates. However, based in the final metabolic destination of each amino acid (Table S6), we were able to calculate the expected yield of 1C, 2C, 3C and 4+5C fragments derived from their catabolism and the data gathered for protein intake. We considered the sum of 4+5C fragments as the practical sum of KCAI, since the amount of odd-chain fatty acids (provider of propionate, a KCAI) ingested are negligible [55]; this was in agreement with our own fatty acid analyses. We have considered KCAI accrual as a representative value for total amino acid accrual, in spite of varied (albeit minor) disposal paths We have considered KCAI accrual as a representative value for total amino acid accrual, in spite that a tiny part of amino acid N may be derived in other pathways (Table S6).
Energy intake was calculated from the measured ingested food values, corrected by the respective kJ energy equivalents. A complete description of energy calculations is presented in Supplemental Material.
Statistical comparisons were performed using two-way ANOVA (sex and diet) and post hoc Bonferroni tests using the Prism 5.0 software (GraphPad Software Inc, La Jolla, CA, USA). The differences between groups were considered statistically significant when the p value was ≤ 0.05. Correlations between different parameters were determined by linear regression analysis using the Prism program and applying a 95% confidence interval.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
X.R. and M.A.: conceptualization; L.O.: animal handling and bench procedures; L.O. and X.R.: initial data curation; X.R. and J.-A.F.-L.: statistical validation; X.R.: graphics; X.R.: writing original draft; X.R. and M.A..: writing final version; X.R., M.A., L.O. and J.-A.F.-L.: final text revision and editing.
Funding
This research did not receive any specific funding.
Institutional Review Board Statement
The Committee on Animal Experimentation of the University of Barcelona authorized the specific procedures used (Catalonia Government attributed expedient number (# DAAM 6911).
Data Availability Statement
The raw data used in this study are available at the Universitat de Barcelona Repository. http://hdl.handle.net/2445/111074.
Conflicts of Interest
There are no conflicts to declare.
Appendix A
A question of definitions.
The foods constituting diets, contain nutrients in formerly biological structures, and are essentially made up of sugars and saccharides (mainly polysaccharides), proteins, and lipids (massively TAG) as energy substrates, plus materials we cannot digest (fibre). The carbohydrates are digested to produce 6C molecules (monosaccharides), and some 3C units (largely glycolytic derivatives), but also 5C (pentoses, that interconvert to 6C or 3C) and other compounds. Basically we obtain 6C and 3C with glycolysis as central path for their integration into energy metabolism. The 3C are oxidized by the pyruvate dehydrogenase complex to acetyl CoA, a 2C, which is then oxidized to CO2 in the Krebs cycle for reducing power and energy. Amino acids have a much more complex metabolism, with two definite parts, nitrogen (essentially the 2-amino group) and the hydrocarbon skeletons. The paths for N are the canonical urea cycle and a number of diverse ways of additional N excretion of limited entity in mammals, except for the mechanism of excretion of N2, which path has not yet been clarified, despite the abundant proof in favour of its operation. The hydrocarbon skeletons of amino acids, however render, essentially 3C and 2C units, plus a number of 4C and 5C intermediates that are directly related to the Krebs cycle (KCAI, as described in the text), which, eventually can be used as energy substrates via the conversion of 4C and 5C to 3C, the oxidation of 3C to 2C and the final conversion of 2C to CO2.
It must be considered that the simplification used above refers to substrates quite homogeneous as to their relationship to the central axis of glycolysis-Krebs cycle. A number of 2C (e.g., ethanol) or 3C (D-lactate, propionate) or even 4C (butyrate) could not be directly incorporated to this simplified narrative. Ethanol can be oxidized to acetate (a true 2C), D-lactate processing is slow and it may not be fully used (excretion, microbiota oxidation); propionate is a KCAI of 3C, but is metabolized as such, not via immediate oxidation to 2C. Butyrate, acetoacetate and 3-hydroxy-butyrate are metabolized as dual 2C providers. There are quite a number of exceptions, but the main line of nutrient-substrate to energy-production substrate follows majoritarian the path: 3C → 2C.
1. Rat weight changes
All animals were housed in same-sex pairs to limit the isolation stress in a controlled environment (21.5-22.5 ºC; 75-85% relative humidity; filtered air-renovation: 1 whole cycle/2 h; and artificial lights on from 08:00 to 20:00 GMT+2 --local time zone). Animal weight changes were measured and recorded (i.e., from day 0 to day 30 of the study. Furthermore, the daily intake of the diet components was registered for each cage for the same period.
2. Diet composition and analysis
The composition of diets used has been described in the Methods section. Here, we show a detailed composition of main components of the specific diets used, that in part was published in a previous paper [2] (Table S1).
Crude energy equivalences from the diet components were obtained using a bomb-calorimeter (Ika c7000), a procedure (and calibration/ adjustments) that was previously described [44].
Crude nitrogen content of the samples (and tissues) was measured using a semiautomatic Kjeldahl system as previously described [9]. Data from food components and biological samples were analyzed –in parallel—for humidity (dry weight, after 12 h at 40 ºC in an oven) in order to correct the crude weights for nutrient content.
2.1. Amino acid analyses and composition of diets
The analyses of amino acids from other biological samples (e.g., tissues, diet components) was done through initial careful three-step homogeneous sampling (i.e., powdering the liquid-nitrogen-frozen samples using mortar and pestle/ mechanical blender or homogenizer, the ground samples were further homogenized and solubilized in concentrated alkali and then centrifuged), then the extract was neutralized and the amino acids derivatized and their concentrations measured as described above. Amino acid analyses were done after deproteinization with cold acetone [9], and separation-quantification by amino acid analyser (ALPHA-PLUS (Pharmacia) using ninhydrin to derivatize amino acids. Norleucine or norvaline internal standards were used for first correction) and purified mixtures of standards were run with each batch for additional calibration. Part of this process (essentially the use of large instrumental systems) was carried out by the CCiT-UB (Scientific and Technological Services of support to Research) of the University of Barcelona.
The data for cafeteria diet components were previously published (in part) in [45]. The data for chow pellet were those given by the manufacturer (Diet 2014 Teklad).
2.2. Fatty acid composition of diets
Cafeteria diet fatty acid composition data (as well as rat biological samples’ composition) were obtained from dried samples, frozen, powdered and then extracted with trichloromethane:methanol [49], washed and then processed in GC-MS System (QP 2010 Shimadzu) at the CCiT-UB. Internal standard of bis-C17:0 diacylglycerol was added after the powdering process. The batch-correction was done using pre-prepared extended methylated fatty acid mixture (Supelco FAME mix C4–C24) of fatty acids as previously commented [50]. Most of the data used in the present study were taken from a previous publication of our group [48]. Coconut oil and Teklad chow fatty acid composition were analysed after extraction in chloromethane:methanol (2:1 v/v) as described above.
2.3. The model of energy used
The direct measure of the diet components consumed by each ‘cage’ (i.e., 2 rats) every day was corrected by the humidity content of the ‘pelleted’ diets (was checked but found to be sufficiently stable not to take this (negligible) factor into account). The weight of the solid components of the cafeteria diet, however, was corrected on a daily basis by using the weight decrement (water loss) of the samples exposed in the control cage (devoid of animals) set in paralle every day. The data for intake, i.e., the food offered minus the leftovers –corrected for humidity loss with the no-rat-cage controls—constituted the corrected amount of food ingested in each cage by the two resident rats.
The composite problem of different body change rates induced by diet consumption and the initial differences due to sex-related size and individual variability were corrected using an allometric factor that allows inter-species comparisons based on body size [56] but also same-species but different body size metabolic comparisons [57]. We used the BW0.75 allometric factor as the –probably—more adequate for this type of comparisons. In previous studies we already used this allometric factor to enable direct comparisons of heterogeneous-size experimental populations [59].
The power allometric correction, correlated with body energy expenditure and basal metabolic rate [56], however, is not sufficient to allow a direct comparison of the energy balances of the rats at different points of the one-month period studies. In this time, there are gross differences between groups, but also within the same group (case of BW). In order to homogenize the comparisons, we devised a process of standardization of the 30 days to a single dynamic point. Since all dietary-treatment groups started from the same rat population, strai, age and source, a group of rats (for each sex) was killed on day one, just at the time when the other rats were weighed and were offered the established diets. This 0 group weight allowed obtaining a zero-time body weight and composition. Since the BW curves showed a linear growth for each group along the study, we arbitrarily took day 15 as the point of comparison. Then for all parameter to be analysed we took the values for the regression lines’ cut at day 15. The data for this ‘means’ day as to body weight and composition, food consumption, and energy balances (i.e., energy intake, accrual and loss), were used for all comparisons. Then, we further completed the model by using the allometric exponent.
Consequently, to compare the energy intake or flow of energy for each group, the basic model described above of representing each group (diet, sex) as a compounded value of BW0.75 at day 15 was used; energy flow/ power values were compared as W/g0.75. The contribution of the different nutrients/ stores of energy were then compared using homogeneous unit data
2.4. Triacylglycerols
We assumed that most of the fatty acids in the diet were part of acylglycerols (largely TAG), thus, we considered essentially two components, fatty acids, and glycerol. Three mols of fatty acids esterifying one mol of glycerol. The mass of ingested lipids (discounting the proportion of glycerol) were corrected by the individual fatty composition, which gave the amount of individual fatty acids ingested. This value was used to calculate their final potential conversion into 2C fragments. These values initially expressed in molar units could be transformed in J and/or W.
2.5. Amino acids
As indicated above (Section 0), the catabolism of individual types of amino acids produces a variety of fragments of different number of C. The final fate for the hydrocarbon skeletons of amino acids is presented in Table S6. These data were collected and described previously [10]. The pathways and distribution between them for a given amino acid have been used to attribute their final catabolites to the groups of 1C, 2C, 3C and 4C+5C hydrocarbon intermediates derivable from the ingested amino acids. The data, initially in grams were converted to moles and finally distributed into the four primary hydrocarbon groups. Essentially, almost all amino acids produced fragments of 3 to 5 C, except for Leu and Lys, largely 2C direct producers.
When analyzing the deposition of amino acids, it must be taken into account that the complete synthesis of ‘essential’ amino acids is not biochemically feasible in most vertebrates’ tissues, but this problem can be in part circumvented via protection of their oxidation and the parallel synthesis of most amino acids derived from the central energy pathway of glycolysis and Krebs cycle intermediary metabolites. Thus, the accrual of essential amino acids could be done only via recycling and saving, but not newly synthesized contributions can be expected. Most of the newly formed amino acids are a product of 3C integration or recycling (KCIA-mediated) of other amino acids, incorporating newly provided 2-amino N since the initially excreted N is practically irretrievable.
2.6. Dietary starches and sugars
We assumed that most of the diet starches (easily digestible poly 6C (glucose) in long stretches of α1 →4 bond chains, with little ramification was completely degraded and converted in glucose which produced two 3C fragments each. Other polysaccharides resulted in similar effects. Variations in the main-chain link bonds, ramification, chemical modification of the chain, presence of pentoses and epimers of glucose, etc. may result in a myriad of (mainly) plant reserve polysaccharides which could not be degraded (i.e., become ‘fibre’ from a nutritional point of view) and/or have long digestions (i.e., have low glycemic indexes), but what has not been extracted and metabolized in a prudential amount of time is just excreted in the faeces.
The saccharides are a quite different question. We assumed that lactose produced one mol each of glucose and galactose; the last converted in glucose by the Leloir pathway. Probably this is not fully acceptable, since the utility of the Leloir path (critically at the epimerization in C4 of UDP-gal) as catabolic path has not been proven yet in adult mammals) and there is, simply, not enough information on the fate of this important nutrient for adults. The use of sucrose is much better known, since it is fully hydrolyzed to glucose and fructose, i.e., 2 6C, which are generally glycolysed to 3C and used as such or further oxidized to 2C.
The 2C fragments from the diet (circumventing the 3C step) were provided massively by fatty acids plus those obtained from the oxidation of amino acids and other minor sources (i.e., acetate).
The 3C fragments were the provided by the 6C from starch and disaccharides (sucrose and lactose) plus the 3C from amino acids plus glycerol. Since the diet did not contain measurable levels of odd-number or methy-branched fatty acids in the lipids, all KCAI in the diets used in this study were provided by amino acids, Table S6 shows their proportional contribution of 4C and 5C fragments to the overall diet.
3. Body composition
When the rats were euthanized, the gastrointestinal content was extracted and discarded and the corpse was used to obtain a “total rat” homogenate, as previously described [42]. The difference in weight from a rat anesthetized but alive and the corpse after euthanization using isoflurane and voiding of the gut contents was measured and used to determine true body composition. There was a small loss of other fluids, but –when possible—they were added to the digestion bag. Along this process, the control of weights is critical. Using these homogenates, we analysed the whole-body energy content (bomb-calorimeter), water content (differential drying), lipid content (as described above, using a two-phase extraction procedure) and protein content. The latter was estimated from the determination of total N content using a semi-automatic Kjeldahl system, as described above. The conversion of total N to protein was done using a N-content factor or 5.5 established previously for whole rat protein content [82].
Thus, we were able to determine the gross body composition of the rats at the end of the study (day 60). On the other hand, we already had sufficient data for untouched control animals that matched the present stud, previously published by us [5,45,83]. Using part of these data, and adjusting them to the parameters already available for the present group we established the body composition on day 0. These data allowed us to determine the net accrual of BW, water, lipid total N and protein.
The total amount of lipids and amino acids, in the form of individual fatty acids and individual amino acids was calculated from carcass composition (previous paragraph). Using data of days 0 and 30 (measured), the net overall balances of body fatty acid amino acid content of the rats along the one-month dietary exposure were calculated. These data, also allowed (including the whole dietary intake data) to calculate the oxidized (i.e., ingested minus accrued) individual fatty and amino acids. The data, initially expressed in molar units were converted to energy (W) units as explained above
Data of 2C or 3C or KCAI ingested, deposited, or oxidized were correlated with plasma hormone levels.
References
- Remesar, X.; Alemany, M. Dietary Energy Partition: The Central Role of Glucose. Int J Mol Sci 2020, 21, 7729. [Google Scholar] [CrossRef]
- Oliva, L.; Alemany, M.; Fernández-López, J.A.; Remesar, X. Circulating oestradiol determines liver lipid deposition in rats fed standard diets partially unbalanced with higher lipid or protein proportions. Br J Nutr 2022, 128, 1499–1508. [Google Scholar] [CrossRef]
- Chalvon-Demersay, T.; Blachier, F.; Tomé, D.; Blais, A. Animal models for the study of the relationships between diet and obesity: a focus on dietary protein and estrogen deficiency. Front Nutr 2017, 4, 5. [Google Scholar] [CrossRef]
- Zhou, J.; Keenan, M.J.; Losso, J.N.; Raggio, A.M.; Shen, L.; McCutcheon, K.L.; Tulley, R.T.; Blackman, M.R.; Martin, R.J. Dietary whey protein decreases food intake and body fat in rats. Obesity 2011, 19, 1568–1573. [Google Scholar] [CrossRef]
- Esteve, M.; Rafecas, I.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Dietary amino acid balances in young Wistar rats fed a cafeteria diet. Biochem Mol Biol Int 1993, 29, 1069–1081. [Google Scholar]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr Res Rev 2010, 23, 270–299. [Google Scholar] [CrossRef]
- Romero, M.M.; Roy, S.; Pouillot, K.; Feito, M.; Esteve, M.; Grasa, M.M.; Fernández-López, J.A.; Alemany, M.; Remesar, X. Treatment of rats with a self-selected hyperlipidic diet; increases the lipid content of the main adipose tissue sites in a proportion similar to that of the lipids in the rest of organs and tissues. PloSOne 2014, 9, e90995. [Google Scholar] [CrossRef]
- Davis, C.; Loxton, N.J. A psycho-genetic study of hedonic responsiveness in relation to “Food Addiction”. Nutrients 2014, 6, 4338–4353. [Google Scholar] [CrossRef]
- Oliva, L.; Aranda, T.; Caviola, G.; Fernández-Bernal, A.; Alemany, M.; Fernández-López, J.A.; Remesar, X. In rats fed high-energy diets; taste; rather than fat content; is the key factor increasing food intake: a comparison of a cafeteria and a lipid-supplemented standard diet. PeerJ 2017, 5, e3697. [Google Scholar] [CrossRef]
- Ferrer-Lorente, R.; Fernández-López, J.A.; Alemany, M. Estimation of the metabolizable energy equivalence of dietary proteins. Eur J Nutr 2007, 46, 1–11. [Google Scholar] [CrossRef]
- Owen, O.E.; Kalhan, S.C.; Hanson, R.W. The key role of anaplerosis and cataplerosis for citric acid cycle function. J Biol Chem 2002, 277, 30409–12. [Google Scholar] [CrossRef] [PubMed]
- Wieland, O.H. The mammalian pyruvate dehydrogenase complex: Structure and regulation. Rev Physiol Biochem Pharmacol 1983, 96, 123–170. [Google Scholar] [PubMed]
- Jenkins, B.; West, J.A.; Koulman, A. A review of odd-chain fatty acid metabolism and the role of pentadecanoic acid (C15:0) and heptadecanoic acid (C17:0) in health and disease. Molecules 2015, 20, 2425–2444. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.; Jones, M.; Misso, M.; Hewitt, K.; Hill, R.; Maffei, L.; Carani, C.; Boon, W.C. Estrogen; a fundamental player in energy homeostasis. J Steroid Biochem Mol Biol 2005, 95, 3–8. [Google Scholar] [CrossRef]
- Xu, Y.; Lopez, M. Central regulation of energy metabolism by estrogens. Mol Metab 2018, 15, 104–115. [Google Scholar] [CrossRef]
- Alemany, M. Estrogens and the regulation of glucose metabolism. World J Diabetes 2021. [Google Scholar] [CrossRef]
- Fukushima, A.; Hagiwara, H.; Fujioka, H.; Kimura, F.; Akema, T.; Funabashi, T. Sex differences in feeding behavior in rats: relationship with neuronal activation in the hypothalamus. Front Neurosci 2015, 9, 88. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Henever, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr Rev 2013, 34, 309–338. [Google Scholar] [CrossRef]
- Alemany, M. The roles of androgens in humans: Biology, Metabolic regulation and health. Int J Mol Sci 2022, 23, 11952. [Google Scholar] [CrossRef]
- Muraleedharan, V.; Jones, T.H. Testosterone and the metabolic syndrome. Therapy Adv Endocrinol Metab 2010, 1, 207–223. [Google Scholar] [CrossRef]
- Vermeulen, A.; Kaufman, J.M.; Goemaere, S.; van Pottelberg, I. Estradiol in elderly men. Aging Male 2002, 5, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.M.; Jones, T.H. Testosterone: a metabolic hormone in health and disease. J Endocrinol 2013, 217, R25–R45. [Google Scholar] [CrossRef] [PubMed]
- Guillaume, M.; Handgraaf, S.; Fabre, A.; Raymond-Letron, I.; Riant, E.; Montagner, A.; Vinel, A.; Buscato, M.; Smirnova, N.; Fontaine, C.; Guillou, H.; Arnal, J.F.; Gourdy, P. Selective activation of estrogen receptor α activation function-1 is sufficient to prevent obesity; steatosis; and insulin resistance in mouse. Am J Pathol 2017, 187, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. Estrogenic control of mitochondrial function. Redox Biol 2020, 31, 101435. [Google Scholar] [CrossRef] [PubMed]
- Bowler-Kinley, M.M.; Davis, W.I.; Wu, P.F.; Harris, R.A. Popov K.M. Evidence for existence of tissue-specific regulation of the mammalian pyruvate dehydrogenase complex. Biochem J 1998, 329, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Bian, C.; Bai, B.; Cao, C.; Li, S.; Zhao, Y. 17β-estradiol regulates glucose metabolism and insulin secretion in rat islet β cells through GPER and Akt/mTOR/GLUT2 pathway. Front Endocrinol 2019, 10, 511. [Google Scholar] [CrossRef]
- Gorres, B.K.; Bomhoff, G.L.; Morris, K.; Geiger, P.C. In vivo stimulation of estrogen receptor α increases insulin-stimulated skeletal muscle glucose uptake. J Physiol 2011, 598, 2041–2054. [Google Scholar] [CrossRef]
- Burning, P.F. : Bonfrèr JMG. Free Fatty Acid Concentrations Correlated with the Available Fraction of Estradiol in Human Plasma. Cancer Res 1986, 46, 2606–2609. [Google Scholar]
- Navarro, G.; Allard, C.; Xu, W.; Mauvais-Jarvis, F. The role of androgens in metabolism; obesity and diabetes in males and females. Obesity 2015, 23, 713–719. [Google Scholar] [CrossRef]
- Rossetti, M.L.; Steiner, J.L.; Gordon, B.S. Androgen-mediated regulation of skeletal muscle protein balance. Mol Cell Endocrinol 2017, 447, 35–44. [Google Scholar] [CrossRef]
- Kuo, T.; McQueen, A.; Chen, T.-C.; Wang, J.-C. Regulation of glucose homeostasiss by glucocorticoids. Adv Exp Med Biol 2015, 872, 99–126. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, M.; Shimomura, Y.; Nakai, N.; Jeoung, N.H.; Nahgasaki, M.; Murakami, T.; Sato, Y.; Harris, R.A. Estrogen controls branched-chain amino acid catabolism in female rats. J Nutr 2004, 134, 26-28-2633. [Google Scholar] [CrossRef]
- Arnold, P.K.; Finley, L.W.S. Regulation and function of the mammalian tricarboxylic acid cycle. J Biol Chem 2023, 299, 102838. [Google Scholar] [CrossRef] [PubMed]
- Kitabchi, A.E.; McDaniel, K.A.; Wan, J.Y.; Tylavsky, F.A.; Jacovino, C.A.; Sands, C.W.; Nyenwe, E.A.; Stentz, F.B. Effects of high-protein versus high-carbohydrate diets on markers of β-cell function; oxidative stress; lipid peroxidation; proinflammatory cytokines; and adipokines in obese; premenopausal women without diabetes. A randomized controlled trial. Diabetes Care 2013, 36, 1919–25. [Google Scholar] [CrossRef]
- Tang, M.H.; Armstrong, C.L.H.; Leidy, H.J.; Campbell, W.W. Normal vs. high-protein weight loss diets in men: effects on body composition and indices of metabolic syndrome. Obesity 2013, 21, E204–E210. [Google Scholar] [CrossRef]
- Cahill, G.F.; Aoki, T.T. Starvation and body nitrogen. Trans Am Clin Climatol Assoc 1971, 82, 43–51. [Google Scholar]
- Karasov, W.H.; Diamond, J.M. Interplay between Physiology and Ecology in digestion. BioScience 1988, 35, 602–611. [Google Scholar] [CrossRef]
- Lindsay, D.B. Amino acids as energy sources. Proc Nutr Soc 1980, 39, 53–59. [Google Scholar] [CrossRef]
- Liu, J.; Bisschop, P.H.; Eggels, L.; Foppen, E.; Ackermans, M.T.; Zhou, J.N.; Fliers, E.; Kalsbeek, A. Intrahypothalamic Estradiol regulates glucose metabolism via the sympathetic nervous system in female rats. Diabetes 2013, 62, 435–443. [Google Scholar] [CrossRef]
- Esteve, M.; Rafecas, I.; Remesar, X.; Alemany, M. Nitrogen balance of lean and obese Zucker rats subjected to a cafeteria diet. Int J Obes 1992, 16, 237–244. [Google Scholar]
- Prats, E.; Monfar, M.; Iglesias, R.; Castellà, J.; Alemany, M. Energy intake of rats fed a cafeteria diet. Physiol Behav 1989, 45, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Oliva, L.; Aranda, T.; Alemany, M.; Fernández-López, J.A.; Remesar, X. Unconnected body accrual of dietary lipid and protein in rats fed diets with different lipid and protein content. Mol Nutr Food Res 2020, 64, 2000265. [Google Scholar] [CrossRef] [PubMed]
- Oliva, L.; Alemany, M.; Remesar, X.; Fernández-López, J.A. The food energy/protein ratio regulates rat urea cycle operation but not total nitrogen losses. Nutrients 2019, 11, 316. [Google Scholar] [CrossRef]
- García-Peláez, B.; Vilà, R.; RemesarX. Maternal treatment with oleoyl-estrone induces resistance to lipid accrual in their descendants. Obesity 2008, 16, 2223–2231. [Google Scholar] [CrossRef] [PubMed]
- Esteve, M.; Rafecas, I.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Fatty acid utilization by young Wistar rats fed a cafeteria diet. Mol Cell Biochem 1992, 118, 67–74. [Google Scholar] [CrossRef]
- Eastoe, J.E. The amino acid composition of mammalian collagen and gelatin. Biochem J 1955, 61, 4, 589–600. [Google Scholar] [CrossRef]
- Hall, W.L.; Millward, D.J.; Long, S.J.; Morgan, L.M. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Br J Nutr 2003, 89, 239–248. [Google Scholar] [CrossRef]
- Rafecas, I.; Esteve, M.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Individual amino acid balances in young lean and obese Zucker rats fed a cafeteria diet. MolCell Biochem 1993, 121, 45–58. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane- Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J Biol Chem 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Remesar, X.; Antelo, A.; Llivina, C.; Albà, E.; Berdié, L.; Agnelli, S.; Arriarán, S.; Fernández-López, J.A.; Alemany, M. Influence of a hyperlipidic diet on the composition of the non-membrane lipid pool of red blood cells of male and female rats. Peer J 2015, 3, e1083. [Google Scholar] [CrossRef]
- Miwa, I.; Maeda, K.; Okuda, J. Anomeric compositions of D-glucose in tissues and blood of rat. Experientia 1978, 34, 167–169. [Google Scholar] [CrossRef]
- Oliva, L.; Barón, C.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Marked increase in rat red blood cell membrane protein glycosylation by one-month treatment with a cafeteria diet. PeerJ 2015, 3, e1101. [Google Scholar] [CrossRef] [PubMed]
- Arriarán, S.; Agnelli, S.; Sabater, D.; Remesar, X.; Fernández-López, J.A.; Alemany, M. Evidence of basal lactate production in the main white adipose tissue sites of rats. Effects of sex and a cafeteria diet. PlosOne 2015, 10, e0119572. [Google Scholar] [CrossRef] [PubMed]
- Bender, D.A. Amino acid metabolism, 2nd ed.; John Wiley & Sons: Chichester, United Kingdom, 1985; ISBN 9780470661512. [Google Scholar]
- Kalivianakis, M.; Minich, D.M.; Havinga, R.; Kuipers, F.; Stellaard, F.; Vonk, R.J.; Verkade, H, J. Detection of impaired intestinal absorption of long-chain fatty acids: validation studies of a novel test in a rat model of fat malabsorption. Am J Clin Nutr 2000, 72, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Kleiber, M. Body size and metabolic rate. Phys Rev 1947, 27, 511–541. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Energy metabolism; body size and problems of scaling. Fed Proc 1970, 29, 1524–1532. [Google Scholar]
- Roberts, B.; McCrory, M.A.; Saltzman, E. The Influence of Dietary Composition on Energy Intake and Body Weight. J Amer Coll Nutr 2002, 21, 140S–145S. [Google Scholar] [CrossRef]
- López-Martí, J.; Díaz-Silva, M.; Salas, A.; Grasa, M.M.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Oleoyl-estrone induces the massive loss of body weight in Zucker fa/fa rats fed a high-energy diet. J Nutr Biochem 2000, 11, 530–535. [Google Scholar] [CrossRef]
- Huang, Q.; Riviere, J.E. The application of allometric scaling principles to predict pharmacokinetic parameters across species. Expert Opin Drug Metab Toxicol 2014, 10, 1241–1253. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Estrogens in the regulation of liver lipid metabolism. Adv Exper Med Biol 2017, 1043, 227–256. [Google Scholar] [CrossRef]
- Della Torre, S.; Mitro, N.; Meda, C.; Lolli, F.; Pedretti, S.; Barcella, M.; Ottobrini, L.; Metzger, D.; Caruso, D.; Maggii, A. Short-term fasting reveals amino acid metabolism as a major sex- discriminating factor in the liver. Cell Metab 2018, 28, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, J.; Ferres, S.; Horgan, G. Energy Density of Foods: Effects on Energy Intake. Crit Rev Food Sci Nutr 2000, 6, 481–515. [Google Scholar] [CrossRef] [PubMed]
- Roncal-Jimenez, C.J.; Lanaspa, M.A.; Rivard, C.J. Nakagawa, T.; Sanchez-Lozada, G.; Jalal, D.; Andres-Hernando, A.; Tanabe, K.; Madero, M.; Li, N.; Cicerchi, C.; Mc Fann, K.; Sautin, Y.Y.; Johnson, R.J. Sucrose induces Fatty Liver and Pancreatic Inflammation in Male Breeder Rats Independent of Excess Energy Intake. Metabolism 2011, 60, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Basciano, H.; Federico, L.; Adeli, K. Fructose, insulin resistance, and metabolic dyslipemia. Nutr Metab 2005, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G. The metabolic syndrome or the insulin resistance syndrome? Different names; different concepts; and different goals. Endocrinol Metab Clin N Am 2004, 33, 283–303. [Google Scholar] [CrossRef]
- Hodson, L.; Gunn, P.J. The regulation of hepatic fatty acid synthesis and partitioning: The effect of nutritional state. Nat Rev End 2019, 15, 689–700. [Google Scholar] [CrossRef]
- Tran, C.; Jacot-Descombes, D.; Lecoultre, V.; Fielding, B.A.; Carrel, G.; Le, K.A.; Schneiter, P.; Bortolotti, M.; Frayn, K.N.; Tappy, l. Sex differences in lipid and glucose kinetics after ingestion of an acute oral fructose load. Br J Nutr 2010, 104, 1139–1147. [Google Scholar] [CrossRef]
- Sampey, B.P.; Vanhoose, A.M.; Winfield, H.M.; Freemerman, A.J.; Muehlbauer, M.J.; Fueger, P.T.; Newgard, C.B.; Makowski, l. Cafeteria diet is a robust model of human metabolic syndrome with liver and adipose inflammation: comparison to high-fat diet. Obesity 2011, 19, 1109–1117. [Google Scholar] [CrossRef]
- Muriel, P.; López-Sánchez, P.; Ramos-Tovar, E. Fructose and the liver. Int J Mol Sci 2021, 22, 6969. [Google Scholar] [CrossRef]
- Qi, X.; Tester, R.F. Fructose; galactose and glucose – In health and disease. Clin Nutr ESPEN 2019, 33, 18–28. [Google Scholar] [CrossRef]
- Lai, K.; Elsas, L.J.; Wierenga, K.J. Galactose Toxicity in Animals. IUBMB Life 2009, 61, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Lalanza, J.F.; Snoeren, E.M.S. The cafeteria diet: A standardized protocol and its effects on behavior. Neurosci. Biobehav. Rev. 2021, 122, 92–119. [Google Scholar] [CrossRef] [PubMed]
- Cahill, G.F. Fuel metabolism in starvation. Annu Rev Nutr 2006, 26, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Alemany, M. Different effects of hyperlipidic diets in human lactation and adulthood: growth versus the development of obesity. Reprod Biol Endocrinol 2011, 9, 101. [Google Scholar] [CrossRef]
- Paoli, A.; Bianco, A.; Grimaldi, K.A. The ketogenic diet and sport. A possible marriage? Exercise and Sports Sci Rev 2015, 43, 153–162. [Google Scholar] [CrossRef]
- Felig, P.; Owen, O.E.; Wahren, J.; Cahill, G.F. Amino acid metabolism during prolonged starvation. J Clin Invest 1969, 48, 584–94. [Google Scholar] [CrossRef]
- Dimski, D.S. Ammonia metabolism in the urea cycle: function and clinical implications. J Vet Int Med 1994, 8, 73–78. [Google Scholar] [CrossRef]
- Carrière, A.; Lagarde, D.; Jeanson, Y.; Portais, J.C.; Galinier, A.; Ader, I.; Casteilla, l. The emerging roles of lactate as a redox substrate and signaling molecule in adipose tissues. J Physiol Biochem, 2020, 76, 241–250. [Google Scholar] [CrossRef]
- Goodman, M.N.; McElaney, M.A.; Ruderman, N.B. Adaptation to prolonged starvation in the rat: curtailment of skeletal muscle proteolysis. Am J Physiol 1981, 241, E321–E327. [Google Scholar] [CrossRef]
- Giordano, M.; Castellino, P. Correlation between amino acid induced changes in energy expenditure and protein metabolism in humans. Nutrition 1997, 1, 309–312. [Google Scholar] [CrossRef]
- Costa, G.; Ullrich, L.; Kantor, F.; Holland, J.F. Production of elemental nitrogen by certain mammals including man. Nature 1918, 218, 546–55. [Google Scholar] [CrossRef]
- Rafecas, I.; Esteve, M.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Whole rat protein content estimation: applicability of the Nx6.25 factor. Br J Nutr 1994, 72, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Sabater, D.; Agnelli, S.; Arriarán, S.; Fernández-López, J.-A.; Romero, M. de M.; Alemany, M.; Remesar, X. Altered nitrogen balance and decreased urea excretion in male rats fed cafeteria diet are related to arginine availability. Biomed Res Int 2014, 959420. [Google Scholar] [CrossRef]
- Matute, M.L.; Kalkhoff, R.K. Sex steroids influence on hepatic gluconeogenesis and glycogen formation. Endocrinology 1973, 92, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Fernández-López, J.A.; Casado, J.; Argilés, J.M.; Alemany, M. Intestinal handling of a glucose gavage by the rat. Mol Cell Biochem 1992, 113, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Conte, A.; Damiano, F.; Siculella, L.; Zara, V. Differential effects of high-carbohydrate and high-fat diets on hepatic lipogenesis in rats. Eur J Nutr 2014, 53, 1103–1114. [Google Scholar] [CrossRef]
- Bertram, A.C.; Larsen, L.B.; Chen, X.; Jeppesen, B. Impact of high-fat and high-carbohydrate diets on liver metabolism studied in a rat model with a systems biology approach. J Agric Food Chem 2012, 60, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Imbert-Fernandez, Y.; Clem, B.F.; O’Neal, J.; Kerr, D.A.; Spaulding, R.; Lanceta, L.; Clem, A.L.; Telang, S.; Chesney, J. Estradiol stimulates glucose metabolism via 6-fosfofructo-2´kinase (PFKB3). J Biol Chem 2014, 289, 9440–9448. [Google Scholar] [CrossRef]
- Qiu, S.; Torrens-Vazquez, J.; Boulger, E.; Liu, H.; Xue, P.; Hussain, M.A.; Wolfe, A. Hepatic estrogen receptor α is critical for regulation of gluconeogenesis and lipid metabolism in males. PloSOne 2017, 7, 1661. [Google Scholar] [CrossRef]
- Yan, H.; Yang, W.; Zhou, F.; Li, X.; Pan, Q.; Shen, Z.; Han, G.; Newell-Fugate, A.; Tian, Y.; Majeti, R.; Liu, W.; Xu, Y.; Wu, C.; Allred, K.; Allred, C.; Sun, Y.; Guo, S. Estrogen Improves Insulin Sensitivity and Suppresses gluconeogenesis via the Transcription Factor Foxo. Diabetes 2019, 68, 291–304. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, K.; Sadana, P.; Chowdhury, F.; Gaillard, S.; Wang, F.; McDonnell, D.P.; Unterman, T.G.; Elam, M.B.; Park, E.A. Estrogen-related receptors. J Biol Chem 2006, 281, 39897–39906. [Google Scholar] [CrossRef] [PubMed]
- Maffei, L.; Murata, Y.; Rochira, V.; Tubert, G.; Aranda, C.; Vazquez, M.; Clyne, C.D.; Davis, S.; Simpson, E.R.; Carani, C. Dysmetabolic syndrome in a man with a novel mutation of the aromatase gene: effects of testosterone; alendronate; and estradiol treatment. J Clin Endocrinol Metab 2004, 89, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.N.; Della-Fera, M.A.; Baile, C.A. The mediation of hepatic lipogenesis through estrogens. PostDoc J 2013, 1, 27–3. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, J.; Tateyama, K.; Syuto, B.; Too, K. Lactate dehydrogenase and creatine phosphokinase isoenzymes in tissues of laboratory animals. Jpn J Vet Res 1990, 38, 19–29. [Google Scholar] [CrossRef]
- Rotondo, F.; Ho-Palma, A.C.; Romero, M.D.M. , Remesar, X.; Fernández-López, J.A.; Alemany, M. Higher lactate production from glucose in cultured adipose nucleated stromal cells than for rat adipocytes. Adipocyte 2019, 8, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Liu, F.; Li, Q.; Tang, J.; Zheng, T.; Lu, F.; Lu, H.; Jia, W. The gonadal hormone regulates the plasma lactate levels in type 2 diabetes treated with and without Metformin. Diabet Technol Therap 2012, 14, 469–474. [Google Scholar] [CrossRef]
- Mumford, S.L.; Chavarro, J.; Zhang, C.; Perkins, N.J.; Sjaarda, L.A.; Pollack, A.Z.; Schliep, K.C.; Michels, K.A.; Zarek, S.M.; Plowden, T.C.; Radin, R.G.; Messer, L.C.; Frankel, R.A.; Wactawski-Wende, J. Dietary fat intake and reproductive hormone concentrations andovulation in regularly menstruating. Am J Clin Nutr 2016, 103, 868–77. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



