Submitted:
06 October 2023
Posted:
08 October 2023
You are already at the latest version
Abstract
While it is well known that 98-99% of the human genome do not encode proteins, but are nevertheless transcriptionally active and give rise to a broad spectrum of noncoding RNAs (ncRNAs) with complex regulatory and structural functions, specific functions have so far been assigned to only a tiny fraction of all known transcripts. On the other hand, the striking observation of an overwhelmingly growing fraction of ncRNAs, in contrast to an only modest increase in the number of protein-coding genes, during evolution from simple organisms to humans, strongly suggests critical but so far essentially unexplored roles of the noncoding genome for human health and disease pathogenesis. Research into the vast realm of the noncoding genome during the past decades thus lead to a profoundly enhanced appreciation of the multi-level complexity of the human genome. Here, we address a few of the many huge remaining knowledge gaps and consider some newly emerging questions and concepts of research. We attempt to provide an up-to-date assessment of recent insights obtained by molecular and cell biological methods, and by the application of systems biology approaches. Specifically, we discuss current data regarding two questions: (1) By which mechanisms could evolutionary recent ncRNAs with critical regulatory functions in a broad spectrum of cell types (neural, immune, cardiovascular) constitute novel thera-peutic targets in human diseases ? (2) Since noncoding genome evolution is causally linked to brain evolution, and given the pro-found interactions between brain and immune system, could human-specific brain-expressed ncRNAs play a direct or indirect (immune-mediated) role in human diseases? Synergistic with the remarkable recent progress regarding delivery, efficacy, and safety of nu-cleic acid-based therapies, the ongoing large-scale exploration of the noncoding genome for human-specific therapeutic targets is encouraging to proceed with the development and clinical evaluation of novel therapeutic pathways suggested by these research fields.
Keywords:
primate evolution genetics
; noncoding genome
; immunogenetics
; neurogenetics
; neurobiology
; cell biology
; molecular biology
; neurological diseases
; cardiovascular diseases
; brain-immune system interactions
; nucleic acid therapeutics
; RNA interference
; AAV vectors
; blood-brain-barrier
1. Introduction
While it is well known that 98-99% of the human genome do not encode proteins, but are nevertheless transcriptionally active and give rise to a broad spectrum of noncoding RNAs [ncRNAs] with complex regulatory and structural functions, specific functions have so far been assigned to only a tiny fraction of all known transcripts [1,2,3,4,5,6,7,8,9].
On the other hand, the striking observation of an overwhelmingly growing fraction of ncRNAs, in contrast to an only modest increase in the number of protein-coding genes, during evolution from simple organisms to humans, strongly suggests critical but so far essentially unexplored roles of the noncoding genome for human health and disease pathogenesis [8,10,11,12].
Research into the vast realm of the noncoding genome during the past decades thus led to a profoundly enhanced appreciation of the multi-level complexity of the human genome. Here, we address a few of the many huge remaining knowledge gaps and consider some newly emerging questions and concepts of research. We provide a critical up-to-date assessment of recent insights obtained by molecular and cell biological methods, and by large-scale application of systems biology approaches. Specifically, we address two questions:
(1) How would evolutionary recent noncoding regions of the human genome with critical regulatory functions in a broad spectrum of cell types (neural, immune, cardiovascular) possibly constitute novel therapeutic targets in human diseases ?
(2) Since noncoding genome evolution is causally linked to brain evolution, and given the profound interactions between mind and brain and the immune system in humans, could human-specific brain-expressed genes play a direct or indirect (immune-mediated) role in human diseases ?
“Human-specific” in this context does not necessarily mean there are no paralogues or similar genes in other species, but that the respective gene has gained novel or particularly important functions in humans. This may simply occur, because an organ such as the brain has dramatically increased in size and complexity during evolution, requiring more sophisticated genome-level regulation and spatio-temporal coordination of cellular functions and cell-cell interactions.
2. Overwhelming Expansion of the Noncoding Genome in Higher Organisms
Landmark studies already decades ago documented intracellular presence of large amounts of RNAs that are transcribed, but do not encode proteins. Only part of these ncRNAs could later be linked to mRNA splicing (e.g., small nuclear RNAs [snRNA], or was specifically involved in the translation machinery and its regulation (e.g., transfer RNAs [tRNAs], ribosomal RNAs [rRNAs], small nucleolar RNAs [snoRNA], but the vast majority remained functionally cryptic for a long time. More recent studies then led to the discovery of entire new classes of small RNAs, generated not only by novel biosynthetic pathways and mediating gene expression post-transcriptionally (e.g., microRNA [miRNAs]), and also of unusually large long ncRNAs [lncRNAs] - in almost all cases – of still unknown functional significance. Broad evidence derived from multiple genetic, biochemical, and other experimental and clinical studies during the past decades clearly revealed a key role of ncRNAs in the genetic programming of complex organisms, during their development and in health and disease.
The steeply increasing fraction of ncRNAs in the genome during evolution, from simple to highly complex organisms, strongly contrast with the only modest increase in the number of protein-coding genes (Figure 1 revised) and is consistent with the assumption of an overwhelming role of the ncRNA species in higher organisms. Both Caenorhabditis elegans with ~1.000 somatic cells and humans with ~30 x 1012 somatic cells have ~20.000 protein-coding genes (‘g-value paradox’) [13]. Regarding disease pathogenesis, it is evident since the Encyclopedia of DNA Elements (ENCODE) project [3] that confinement to the analysis of protein-coding regions of the human genome is inadequate, because many noncoding variants are associated with important human diseases. Inclusion of the noncoding genomic elements in pathogenetic studies seems mandatory and one approach is comprehensive transcriptome mapping encompassing protein-coding genes, as well as diverse small and large ncRNAs.
Apparently ~5% of the human genome are functionally constrained [14]. Since only ~1.5% of the genome could possibly be assigned to protein-encoding genes, a major part of these constraints is necessarily associated with functionally important conserved noncoding elements (CNEs), preserved among organism through millions of years of evolution at the cellular and systemic level. As one example, there is evidence for higher-order genome organization functions of lncRNAs in diverse cell types and cell lineages, and during cell differentiation [15]. It was recently found that not only transcription, but also translation, is pervasive outside of protein-coding regions (e.g., lncRNAs, 3’-untranslated regions, introns). Although resulting polypeptides are generally nonfunctional, their translation is considered relevant for the emergence of novel functional genes [16].
One particular interesting but highly complex group of ncRNAs are those designated long noncoding RNAs (lncRNAs). The term 'lncRNAs' encompasses RNA polymerase I (Pol I), Pol II and Pol III transcribed RNAs, and RNAs from processed introns that are at least 200nt in length. More than 100.000 human lncRNAs have been identified many of which are primate-specific [17,18,19,20,21,22,23,24]. A recent consensus statement addressed definition, nomenclature, conservation, expression, phenotypic visibility, structure and functions of lncRNAs [25]. The paper emphasizes that many lncRNAs are cell lineage-specific, associated with developmental enhancers, and likely contribute to species diversity and evolution. Segments of lncRNAs may maintain sequence conservation comparable to protein-coding genes, exhibit conserved exon-intron structures and splice junctions, and retain orthologous functions despite rapid sequence evolution [26,27,28,29].
Importantly, noncoding genome research during the past decades not only revealed unanticipated multi-level complexity of the genome in higher organisms, but has also inspired fundamentally new therapeutic avenues as outlined in Figure 2 (revised).
3. Recent Searches for Evolutionary Relevant Primate/Human-Specific Noncoding Genes
Landmark studies: Nobel-price winning work by Pääbo et al. [30], based on experimentally revolutionary work ultimately enabling deep sequencing of the genomes of extinct primates and hominids, has revolutionized a field of research which was previously the exclusive realm of archaeologists and paleoanthropologists. The puzzle to mankind, when and where modern humans originated and how they differ from, and interacted with, other now-extinct forms of humans has come to be addressed now also by molecular geneticists [30,31,32,33,34,35,36,37,38,39] and protein biochemists [40]. These recent developments have enabled revolutionary novel approaches to diverse and so far enigmatic issues such as the evolutionary development of speech [41].
Screening for evolutionary relevant genetic changes during the diversion of humans from the other primate species naturally first focused on protein-coding genes with rather favourable signal/noise ratio and commonly more obvious functional impact as assessed by conventional molecular and cell biological approaches. Almost two decades ago, however, after the discovery that 98.5% of the human genome is transcribed but not translated into proteins, a first study already scanned for “human accelerated regions“ [HARs] with accelerated substitution rates in the human lineage [42]. That pioneering work identified one region (HAR1) expressed during human cortical development as the most significantly altered element. While the two transcripts from this region (HAR1F and HAR1R RNAs) lacked protein-coding potential, HAR1F transcript folded into a stable RNA structure as occurs with ncRNAs such as miRNAs and many lncRNAs [42]. Since that first study [42], further HARs were identified and investigated and as one important discovery their proximity to neuropsychiatric disease genes was revealed, as discussed in more detail further below.
Brain transcriptome: Pembroke et al. [43] recently discussed the evolutionary conservation and divergence of the human brain transcriptome in general. They emphasize that, although mouse models allow dissection of genetic effects on molecular, cellular, physiological, and behavioral brain phenotypes, the extent to which neurological or psychiatric traits are human- or primate-specific, and cannot be faithfully recapitulated in mouse models, is unknown. Pembroke et al. name multiple human neuropsychiatric and neurodegenerative disease risk genes (COMT, PSEN-1, LRRK2, SHANK3, SNCA) with grossly divergent expression pattern in mice vs. humans, for some of which functions at the cellular level have recently been assigned. We review similar possibly critical knowledge gaps in the cardiovascular field, which are likewise due to differences between the noncoding genome in humans vs. experimental animals (section 6 below).
Espinos et al. [44] discuss in-depth several genetic mechanism regulating the evolution of cortical neurogenesis. They address genes which emerged in the recent human and primate lineages and apparently promote cortical progenitor proliferation and increase neurogenesis. These include structurally conserved primate lncRNAs transiently expressed during human cortical differentiation, and modulating the expression of cell type-specific genes sequentially activated during cortical neurogenesis [45]. Several HARs [46,47] involved therein are predicted to act as regulatory enhancers [48] and are located in the vicinity of genes important for brain development [49].
4. Exploration of the Noncoding Genome for Human-Specific Therapeutic Targets – Three Levels of Increasingly Complex Genome - Disease Relationships
Obviously, any potential pathogenic role of “human-specific” protein-coding genes, as well as of noncoding RNAs, will remain hidden during studies of animal models (murine and diverse others) of human diseases. In principle, any such gene may play an important role in some human disease, however.
It is important to note that “human-specific” in this context does not necessarily mean there are no paralogues in other species, but that the respective gene has gained novel or particularly important functions in humans. This may occur, for instance, because an organ such as the brain has dramatically increased in size and complexity during evolution. Obviously, such genes are more likely to play a pathogenic role in diseases (e.g., neuropsychiatric) which are relatively unique to or display distinct phenotypes in humans.
From a technical standpoint, it is essential to exercise caution when designating a lncRNA as specific to humans or primates. This is because in numerous species, the non-protein coding regions are frequently incompletely annotated, and lncRNA databases remain in-adequately established. Additionally, since many lncRNAs are expected to play roles in the local chromatine remodeling (i.e., function in-cis), not only sequence conservation, but also positional conservation in the context of genomic location should be considered.
At a first level, human-specific proteins could directly alter known signal pathways or other canonical cellular functions. One example is interleukin 8 (IL8) which emerged as one of the most strongly deregulated genes in peripheral blood mononuclear cells (PBMCs) after human myocardial infarction [8,10,50], but does not exist in mice, the most commonly used animal model in cardiovascular research. Drury et al. [51], investigating the evolution and emergence of interferon regulatory factor 9 (IRF9), a key component of the ISGF3 complex and the cellular innate immune response, identified primate-specific IRF9 (PS-IRF9) isoforms unique to old world monkeys and great apes. Ellwanger et al. [52] analyzed the function of the primate-specific NLRP11 gene product, found it highly expressed in human immune cells (myeloid cells, B cells, lymphoma lines), and identified a novel role in the regulation of inflammatory responses in humans.
At a second level, human-specific noncoding RNAs non-existent in animal models may directly alter cellular functions and cell-cell interactions through non-canonical mechanism, and via complex interactions with other proteins and ncRNAs. An interesting recent study found a primate-specific lncRNA (CHROMR) to be induced by SARS-CoV-2 infection, coordinate expression of interferon-stimulated genes (ISGs), and restrict viral infection of macrophages [53]. Another study [54] found the primate-specific lncRNA (CHROME) elevated in plasma and atherosclerotic plaques of coronary artery disease (CAD) patients. Gain- and loss-of-function approaches showed that CHROME promotes cholesterol efflux and HDL biogenesis and constitutes a central component of the non-coding RNA circuitry controlling cholesterol homeostasis in humans.
Neuroscience has likewise identified brain-expressed lncRNA several of which are primate/human-specific and associated with brain development and neuropsychiatric diseases as outlined in section 7 below. Any of these could directly induce damage at their sites of expression in the brain.
At a third level, indirect impact of human-specific genes and ncRNAs upon pathogenesis may be exerted via peculiar interactions between human brain and mind and the immune system. Interactions between the immune system and the nervous system were initially described in the context of diseases. More recent studies have begun to reveal how immune cell-derived effectors can influence host behaviour even in the absence of infection [55]. Essentially, the immune system shapes nervous system function and controls manifestations of host behaviour. In the context of evolution, interactions between these two highly complex biological systems may have evolved to maximize an organism's ability to respond to environmental threats in order to survive [56].
5. Genetics of Immune System and Neuro-Immune Interactions Impact Upon a Large Spectrum of Human Diseases
From the discovery that about 98-99% of the human genome do not encode proteins, but instead generate a broad spectrum of ncRNAs many of whom are involved in the immune response [57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87], decades passed until finally successful clinical exploitation of ncRNAs and of novel drugs developed using them as blueprints was achieved [8,10] (Figure 2). Across the entire spectrum of medical disciplines it has been ascertained that the non-coding genome plays a key role in genetic programming and gene regulation during development as well as in health and disease (Figure 3 and Figure 4 both revised).
A particularly important biological network critically involved in multiple human diseases is the immune system. It is paramount to understanding disease pathogenesis, and to open new therapeutic avenues, from cardiovascular medicine to neurology and other clinical disciplines. For instance, in-depth molecular and genetic analyses of innate immunity have led to the identification of novel molecular players and therapeutic targets in cardiovascular diseases [88]. Immunity in general is deeply involved in many processes which are discussed more specifically in the following chapters. In this context Silverstein [56] has thoughtfully suggested that, in contrast to ”Darwinian” evolution involving adaptation to past challenges, evolution has “devised” two unique biological mechanisms permitting to anticipate future challenges: adaptive immune response and neural memory functions.
The study of immune systems evolution revealed differences, but also striking similarities of the immune mechanisms across different taxa in the context of evolution [89,90]. Major impact of the noncoding genome upon functions and stability of the immune response against diverse challenges has long been appreciated. There is a broad spectrum of broadly diverse non-coding RNAs (ncRNAs) involved in the human immune response [57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86]. Some of these have been identified as primate-specific, e.g. the lncRNA CHROMR [53]. These and related discoveries highlight diverse clinically relevant peculiarities of human immunology beyond the inbred mouse model [91].
6. Noncoding Genomic Regions Impact Upon Cardiovascular Pathogenesis in Humans
Within the field of cardiovascular medicine, a number of early experimental studies [92,93,94,95,96,97,98,99,100,101] revealed that certain ncRNAs (miRNAs) are regulators of cardiovascular homeostasis in animal models. This of course immediately suggested they might have potential to improve diagnostics and could possibly even be developed into novel therapeutics. The road to in-depth understanding of the molecular workings of at least a few of the numerous ncRNA classes, however, and beyond that the development of highly sophisticated bioengineered nucleic acid drugs [92,102,103,104,105,106,107,108,109,110,111,112] (Figure 2, Figure 5 and Figure 6) which are critically required to render them safe and efficacious for clinical applications, required critical input from several disciplines and two decades, counting from the early experimental work to the first clinically successful trials. In section 12 below we update the conceptual and methodological challenges on the road towards clinical exploitation of potential novel human-specific therapeutic targets.
Particularly advanced is the development of RNA interference [RNAi] drugs which use recently discovered pathways of endogenous short interfering RNAs (siRNAs) and have become highly versatile tools for the efficient silencing of any protein-coding or noncoding transcript and gene. A series of ground-breaking clinical trials [107,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134] has provided definite evidence of therapeutic potential of RNA interference [RNAi] and antisense drugs for cardiovascular disorders. For a more in-depth coverage of the enormously challenging bioengineering, safety and regulatory hurdles to be overcome towards clinical therapy during the past decades we may refer the reader to comprehensive recent reviews [8,10,135].
As examples of human-specific noncoding RNAs non-existent in animal models we invoke two primate-specific lncRNAs. One of these (CHROMR) [53] is induced by SARS-CoV-2 infection, coordinates expression of interferon-stimulated genes (ISGs), and restricts viral infection of macrophages [53]. Given the immense and diverse impact of SARS-CoV-2 upon human health, existence of this primate-specific lncRNA is of interest per se. Moreover, the human host’s innate immune response plays a critical role in COVID-19 pathogenesis [136,137,138,139]. Thus, several SARS-CoV-2 accessory proteins have been suggested to affect the innate immune response. Abnormal pro-inflammatory cytokine levels and immune cell infiltration are associated with tissue damage severity and morbidity. Overall, dysregulation of the host immune response and elevated cytokine release are crucial factors for the severity of COVID-19, and thus any peculiar human-specific immunoregulatory ncRNA deserves attention.
Another primate-specific lncRNA is CHROME [54]. It is elevated in plasma and atherosclerotic plaques of coronary artery disease (CAD) patients. Gain- and loss-of-function experiments revealed that CHROME promotes cholesterol efflux and HDL biogenesis and constitutes a key component of the ncRNA circuitry controlling cholesterol homeostasis in humans. Both publications illustrate how, after decades of traditional research in the respective fields, investigation of the noncoding genome still uncovers unexpected novel molecular players. These may significantly contribute to improved pathogenic understanding, beyond what could possibly be derived from animal models. Similar to these examples from the cardiovascular field, studies from the neurosciences and clinical neurology identified impact of species-specific transcripts upon human brain development and neurological disorders [45,140,141,142,143,144,145].
7. Primate/Human-Specific ncRNAs in Neural/Neuroimmune Cells and Their Impact Upon Brain Development and Neuropsychiatric Disorders
There is broad and growing interest in the identification of primate-specific genes involved in primate evolution and the evolution of humans. This includes not only coding genes, but also diverse types of noncoding transcripts, e.g. miRNAs [140] and lncRNAs [146]. One driver of this interest, beyond deeper understanding of human evolution [30], is a hope [140] to identify unique features of human brain development and function which may be critical towards the elucidation of higher cognitive functions, and of human-specific pathologies like neuropsychiatric and behavioral disorders.
After a first study had identified and partially characterized a “human accelerated region” (HAR) [42], further HARs were investigated and indeed - as one important discovery - their proximity to neuropsychiatric disease genes was revealed [39,147,148,149,150,151,152,153]. Since many HAR-associated genes are regulators and hubs in transcriptional networks, their differential expression would affect many other genes and cellular processes, suggesting outsized effects caused by noncoding HAR mutations. HAR enhancers may help to discover the genetic basis for disease, while medical genetics may reveal which HAR variants are pathogenic.
Further along this line, drug target data were employed to map neuropsychiatric disorders to HARs via nearby genes [154,155,156,157]. These genetic studies, involving large neuropsychiatric patient populations, suggest that human-specific noncoding transcripts from HARs as well as species-specific miRNAs are involved in human-specific pathologies like psychiatric and behavioral disorders. If so, sequence variants or altered expression pattern of these ncRNAs in the brain would directly lead to brain dysfunction and disease (second option in Figure 3).
Conveniently, more than two decades of research into the development of diverse types of RNA therapeutics towards clinical applications (as discussed above for cardiovascular and metabolic diseases), has set the stage for targeting/silencing of essentially any type of transcript considered relevant in other disease fields, too. In addition to RNA interference (RNAi) drugs, antisense therapeutics and CRISPR/Cas9-based approaches are being evaluated also with regard to their potential for the treatment of neuropsychiatric disorders [158,159]. Since many ncRNAs are specifically enriched in the central nervous system [CNS], and their dysregulation implicated in Alzheimer's disease and related dementias, Nguyen et al. [160] review conventional small molecule drugs targeting ncRNA, as possible therapeutics for ADRD.
Noncoding RNA targets: lncRNAs play important roles during normal brain development and in the pathogenesis of neurodegenerative disorders (ND) [161]. One target of particular therapeutic interest is Nuclear enriched abundant transcript 1 (NEAT1) (Figure 3), which plays a role in mediating nuclear retention of TAR DNA-binding protein 43 (TDP-43), and is potentially protective in certain proteinopathies [162,163,164,165,166,167] involving aberrant protein aggregates comprising tau, amyloid-β, and α-synuclein. TDP-43 is a highly conserved nuclear RNA/DNA-binding protein regulating RNA processing. Accumulation of TDP-43 aggregates in brain is common to ND such as amyotrophic lateral sclerosis, frontotemporal dementia, and Alzheimer’s disease (AD). Relevant with regard to possible therapeutic options, NEAT1 is upregulated in AD temporal cortex and hippocampus. Hippocampal knockdown of NEAT1 with siRNA improved memory in aged mice and vice versa [168]. Viral knockdown of NEAT1 rescued memory deficit in APP/PS1 mice [169].
Further lncRNA targets of interest include activity-dependent transported Adeptr [170], brain cytoplasmic BC200 [171,172,173,174], BACE1-AS (antisense relative to β-secretase 1 gene) [175,176,177], and Carip [178]. In addition, other types of ncRNAs (miRNAs, piRNAs, lncRNAs, circRNAs) are dysregulated in AD and related dementias with first evidence for therapeutic potential [160] (Figure 7). Thus, miR-132 is downregulated in AD hippocampus, prefrontal and temporal cortex. Viral overexpression and mimics rescued hippocampal cell death, tau homeostasis, hippocampal adult neurogenesis, and behavioral deficits in various AD mouse models [179,180,181]. miR-195 is downregulated in AD parietal cortex. Viral overexpression in mouse models decreased Aβ plaque, tau hyperphosphorylation, and rescued cognitive deficits in ApoE4+/+ mice [182].
These first experimental therapeutic studies hold promise for further developments with translational potential employing highly sophisticated and safe drug delivery systems. These should have well-established molecular and cellular mechanisms of action, including carrier ligand – cell surface receptor [183,184,185,186,187,188,189] interactions, as well as clearly characterized in vivo behaviour [190,191,192].
8. Multi-Level Functional Integration of Extended Noncoding Regions of the Human Genome – Critical Impact Upon Fundamental Cellular PROCESSES governing IMMUNE response and Oncogenesis
To illustrate the multi-level functional integration of major regions of the human genome, above and beyond individual ncRNAs, we invoke two examples addressing genome-level immunoregulation (NEAT1-MALAT1 cluster) (Figure 3 revised), and neoplastic transformation in glioblastoma (HOXD-embedded HOXD-AS2 and distant enhancer-associated LINC01116) (Figure 4 revised).
The evolutionary conserved NEAT1-MALAT1 cluster encounters interest in cardiovascular medicine [50,193,194], oncology [195,196,197,198,199,200], and neurosciences and clinical neurology [169,201,202,203]. While single studies from these fields identified pathogenic roles in specific disease settings, broad interdisciplinary interest apparently results from a deep-rooted complex stabilizing function of the cluster. Within the cardiovascular field, suppression of lncRNA NEAT1 was observed in circulating immune cells of post-myocardial infarction (MI) patients. Mice devoid of NEAT1 or MALAT1 displayed immune disturbances affecting monocyte-macrophage and T cell differentiation, and an immune system highly vulnerable to stress stimuli [204] and prone to the development of atherosclerosis. Uncontrolled inflammation is a key driver of multiple other diseases (see section 7 below), too, which may underly the current broad interdisciplinary interest.
The structure, organizational levels and functions of the noncoding genome are still largely unexplored [8,10,25]. One aspect of particular interest with regard to function(s) is the common unusual complexity of lncRNA interactions with other ncRNAs, proteins, and cellular and subcellular membrane components, as well as their posttranscriptional processing and intracellular kinetics. As an example, the human NEAT1-MALAT1 cluster generates lncRNA remaining nuclear, whereas tRNA-like transcripts (mascRNA, menRNA), enzymatically generated from these precursors, translocate to the cytosol. NEAT1-/- and MALAT1-/- mice display massive atherosclerosis and vascular inflammation [50,193,194,205]. A recent study found that these tRNA-like molecules are critical components of innate immunity and contribute to a balance response of immune cells to diverse challenges. They appear as prototypes of a new class of noncoding RNAs distinct from others (miRNAs, siRNAs) by biosynthetic pathway and intracellular kinetics.
For the long primary transcripts of NEAT1 a particularly interesting function of general cell-biological interest has been elucidated in much detail. These lncRNAs are critical for the formation of paraspeckles which are involved in multiple cellular functions [204]. Yamazaki et al. have put the phenomenon of paraspeckle formation into the much broader context of micellization and the formation of biomolecular condensates [206] which are essential for subcellular and nuclear compartmentalization. Obviously, molecules involved in these fundamental processes may have deep impact upon various cellular functions in a context-dependent manner (e.g., immune stress, infections, toxins). Association of such molecules with diverse diseases is therefore not entirely unexpected.
Other recent studies [207,208,209] have elucidated - in mechanistic detail – the complex multi-level functional integration of distinct regions of the human genome, expressing lncRNAs, in the molecular pathogenesis of glioblastoma (Figure 4 revised).
A remarkable molecular circuit involving ncRNAs plays a pivotal role in governing cell fate and transformation within the brain. This circuit comprises a miRNA, lncRNAs, and a small nuclear RNA (snRNA). The initial component of this intricate network, miR-10b, was originally identified as a unique miRNA that remains transcriptionally silenced in normal brain cells but becomes derepressed in low-grade gliomas and nearly all adult high-grade gliomas, including the most aggressive glioblastoma (GBM), overwise a highly heterogeneous class of brain tumors [210,211]. While activated in glial lineage cells, miR-10b functions through both classical and non-conventional pathways. It regulates the expression of multiple mRNAs associated with cell cycle, cell death, and invasion, such as CDKN1A, CDKN2A, BIM, as well as ncRNA targets [210,212,213]. Notably, an unbiased analysis of miR-10b targets has identified an essential ncRNA component of the spliceosomal machinery, U6 snRNA, as a principal direct target. By modulating the structure, modifications, and levels of U6 snRNA, miR-10b exerts influence over the splicing of numerous cancer-related genes [213].
Overall, glioma cells display a strong dependence on miR-10b, making this small tu-mor-promoting RNA an attractive target for the development of GBM therapies. Various miRNA-targeting modalities, formulations, and delivery approaches employing ASO, gene editing, and small molecules, are currently under development [214,215,216,217].
The intriguing phenomenon of miR-10b's silencing in neuroglial cells of the brain and its transcriptional activation during gliomagenesis has prompted investigations into the up-stream mechanisms responsible for this activation. Interestingly, the entire miR-10b locus, which also encodes 12 HOXD genes, becomes activated in gliomas through a mechanism involving lncRNA-mediated spatial chromatin reorganization (looping). Two interacting lncRNAs, HOXD-AS2 and LINC01116, associate with the HOXD3/HOXD4/miR-10b promoter and a distant enhancer, respectively, and both are necessary for the locus's derepression and gene expression ([207] and Figure 4 revised).
Furthermore, recent work suggests that the LINC01116 enhancer RNA (eRNA) also acts in trans and exhibits global genome wide-modifying activity. It directly binds to more than a thousand of gene promoters, including those of 44 glioma-specific transcription factors distributed across all chromosomes, derepressing them by removing the Polycomb re-pressive complex 2 (PRC2). Consequently, the activation of this single lncRNA in astro-cytes, which are cells of glioma origin, is sufficient to trigger the glioma transcriptional program. In addition to miR-10b and HOXD factors, this includes the activation of neuro-developmental regulators like OLIG2, SOX2, POU3F2, and SALL2, along with multiple oncogenes such as EGFR, PDGFR, TERT, BRAF, and miR-21, ultimately leading to malig-nant transformation [208]. Conversely, targeting LINC01116 with siRNA or ASO may hold substantial therapeutic potential for malignant gliomas, opening up excit-ing new avenues in neuro-oncology and, more broadly, neurology.
It's worth noting that this circuit encompasses various types of ncRNAs, both in terms of evolutionary conservation and functional diversity. These include the highly conserved and functionally essential in diverse eukaryotic species U6 snRNA, the relatively well-conserved miR-10b in vertebrates, and the largely primate-specific LINC01116. These observations prompt intriguing questions about the origin of human brain tumors, a phenomenon uncommon in other mammals, with only a few exceptions. This parallels the scenario seen in most human neurologic diseases.
The recent evolutionary emergence, and the species- and cell-type specificity of LINC01116 and many other lncRNAs invite further in-depth research, undoubtedly lead-ing to better understanding of human pathologies and the discovery of new therapeutic targets and biomarkers for various diseases. Furthermore, with regard to LINC01116, it's noteworthy that this transcript is expressed at high levels not only in the brain, but also in some normal and cancer cells outside the central nervous system. Additional studies into its functions beyond the CNS are needed. It is also imperative to integrate our under-standing of its chromatin-modifying activity with potential extra-nuclear functions [218,219].
9. Impact of Human-Specific Genes and ncRNAs Upon Pathogenesis via Peculiar Interactions between Human Brain and Mind and the Immune System
Considering three levels of increasingly complex genome - disease relationships as outlined in chapter 4, at the third level peculiar indirect pathways from the noncoding genome to neurological diseases may result from the well documented close and intense interactions between human brain and mind and the immune system. In the latter, immunoregulatory ncRNAs play important stabilizing roles and some of these, e.g. CHROMR, are primate-specific [53]. Sequence variants or altered expression pattern of these ncRNAs in immune cells may directly trigger brain dysfunction.
The grave adverse effect of psychological stress upon human diseases is well known since decades [220,221,222], and several stress-induced brain – immune system interactions have been elucidated at the molecular, cellular and systemic level [55,221,223]. In fact, the field of psychoneuroimmunology is one of the fastest-growing fields in the life sciences aiming to stepwise elucidate the highly complex interactions between nervous system and immune system at the molecular and cellular level [224,225,226,227,228,229].
Since psychological stress is certainly different in humans as compared to all other species, as a consequence of the peculiarity of the human brain and mind [157], any direct genetics-based immune dysfunction (section 5) may well synergize with any direct genetic predisposition towards neuropsychiatric disorders (section 7), indirectly resulting in grave brain dysfunction and disease once sufficient psychological stress peculiar to humans is imposed.
10. Current Status of Translational Research into Nucleic Acids-Based Therapeutics
Nucleic acid-based and nucleic acid-targeting therapeutics are currently developed at large scale for the prevention and management of multiple diseases for several reasons:
(1) Genetic studies have highlighted novel therapeutic RNA targets suggested to be causal for these diseases.
(2) There is a substantial recent progress in delivery, efficacy, and safety of nucleic acid-based therapies.
(3) They enable effective modulation of therapeutic targets that cannot be sufficiently or optimally addressed using traditional protein-targeting small molecule drugs or antibodies.
Nucleic acid-based therapeutics in development which are also being evaluated for the diseases outlined and discussed above include: mRNA-targeting drugs for gene silencing; miRNA inhibitors and mimics; gene augmentation therapies ; and genome-editing approaches:
(A) mRNA-targeting drugs for gene silencing: several large-scale clinical development programmes, using antisense oligonucleotides [ASO] or short interfering RNA [siRNA] therapeutics for prevention and management of cardiovascular disease have been initiated. These include ASO and/or siRNA molecules to lower apolipoprotein (a), proprotein convertase subtilisin/kexin type 9 (PCSK9), apoCIII, ANGPTL3 for the prevention and treatment of patients with atherosclerotic cardiovascular disease. In other fields of medicine including neurology, silencing of transthyretin (TTR) was evaluated for the treatment of TTR amyloidosis [10,132,230,231,232].
(B) miRNA mimics and inhibitors for miRNA modulation: Several types of mimetic drugs and inhibitors (e.g., antagomirs) have been developed and a few of them evaluated in clinical trials [134]. These drugs may be “classical” small molecule drugs [233,234,235] or engineered nucleic acids [105,217].
(C) Gene augmentation therapies: EMA/FDA have approved “classical” gene therapies, i.e. those involving vector-based transfer of the protein-coding cDNA sequence into the patient, for monogenic disorders. These include the haemophilias [236,237,238,239,240,241,242], homozygous hypercholesterolaemia [243], and others. Very recently, entirely different approaches towards gene augmentation are being developed which are based on highly specific modulation of RNA-based regulatory networks [244,245]. The molecular details of these approaches for RNA-targeted gene activation and their potential for clinical translation are discussed by Khorkova et al. [244].
(D) Genome editing approaches: these technologies, such as using CRISPR-Cas9, have proven powerful in stem cells, however, grave challenges are remaining, such as low rates of homology-directed repair in differentiated somatic cells (e.g., cardiomyocytes, neurons) and risk for off-target effects encompassing the germline.
Despite complex biotechnological challenges, current lack of efficient therapies for multiple severe and abundant diseases is clear evidence for the need to proceed beyond current options. Reassuringly, a remarkable number of pioneering clinical trials have proven technical and clinical feasibility of nucleic acid therapeutic approaches for important cardiovascular [107,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134] and haematological [236,237,238,239,240,241,242] diseases. These are truly fundamental achievements compared to the situation one decade ago.
The versatility of therapeutic non-coding RNA structures will certainly continue to expand the repertoire of our therapeutic tools, with majors leaps to be expected once critical technological issues are solved (Figure 2, Figure 5 and Figure 6). Fundamentally different from DNA, RNAs are carrying information not only in their linear sequences of nucleotides (primary structure), but local nucleotide pairing creates secondary structures, e.g. hairpins, and interactions among distantly located sequences create tertiary structures. In fact, this structural versatility needs to be considered for RNAs as therapeutic tools as well as targets. The plethora of RNA types, sequences, and structures created by evolution is a treasure trove of potential therapeutic tools and targets.
The strategies outlined above involve the use of "informational" drugs designed based on the sequences of specific targets. Additionally, extensive research in the field of RNA-targeted small molecules [234,235,244] complements these approaches and holds promise for enhancing drug delivery to traditionally challenging target tissues, such as the CNS. The current framework primarily employs small molecules as modulators of mRNA splicing and translation. However, it has the potential to expand to target various classes of ncRNAs and encompass both RNA inhibitors and activators. Initial strides have been taken to identify small molecule inhibitors of miRNAs that interfere with miRNA biogenesis [246,247]. Recent work has also out-lined a high-throughput screening strategy for identifying small molecule miRNA modulators using phenotypic expression-based profiling [160,233]. Furthermore, numerous opportunities exist for the targeting of highly structured lncRNAs once their functional domains become better investigated, as reviewed elsewhere [234].
11. State-of-the-Art in Nucleic Acid-Based Therapeutics and Their Molecular and Cell-Biological Foundations
It is important to note that nucleic acid-based approaches are currently the only therapeutic tools capable to address a multitude of therapeutic targets with proven key impact upon disease pathogenesis, but without any traditional pharmacological options (small molecule drugs, antibodies). Two decades of intense worldwide efforts to develop the novel approaches towards clinical utility have led to remarkable progress for a number of important diseases and in a few cases their entry into clinical practice. Whereas the liver and hepatocytes emerged as rather easily accessible target for RNAi or ASO strategies, once breakthrough molecular design and delivery discoveries (Figure 5) were made, efficient and selective targeting of many other organs and - in particular - proper targeting of specific cell population therein [248,249,250,251,252] has not yet been successfully applied in clinical trials.
While for many cell types and tissues there is currently still no clinically established targeting system available, this was also the case for liver/hepatocyte targeting just one decade ago. This problem is meanwhile solved through elegant molecular drug design and delivery systems. Considering the putative novel human-specific therapeutic targets discussed above, pathogenic involvement and possible molecular pathomechanisms have already been rather well documented in cell cultures, organoids, and animal models. Since several of these regard most severe and abundant diseases, this should be considered as strong incentive to solve the remaining methodological hurdles.
Promising progress has recently been made employing direct intrathecal delivery of chemically engineered siRNAs and ASOs to the CNS, enabling long-term target modulation [253,254]. These breakthrough molecular design studies may possibly – in the long run - have similar translational impact as those which have previously led to the successful liver-targeting trials reviewed above. One study engineered a peculiar divalent siRNA chemical scaffold which enabled potent modulation of gene expression throughout the CNS sustained over six months months upon a single intrathecal injection [253]. Another study achieved allele-specific gene silencing in Huntington's disease models when using chemically engineered siRNAs [254].
An alterative approach to nonviral, chemically synthesized, delivery systems for protein augmentation or RNAi-based target gene silencing is a broad spectrum of recombinant virus-based vectors (Figure 6). Due to the inherently grossly different organ and cell tropisms of the respective basic viruses, their broadly variable stability in the target cells, as well as their potential to induce systemic and local immune responses, selection of a suitable viral vector needs to consider multiple details of the attempted therapeutic strategy [92,93,248,255,256,257,258,259,260,261,262]. These include the desired duration of target modulation, the vector’s capacity to cross relevant anatomical, vascular and cellular barriers, and preferable even target cell-specific vector – cellular surface receptor interaction to achieve selectivity and avoid off-target side effects.
12. Unsolved Challenges and Novel Therapeutic Approaches Guided by Mechanistic Insights at the Molecular and Cell Biological Level
Molecular and cellular basis of liver targeting: Similar to the situation about a decade ago, when efficient and hepatocyte-selective in vivo delivery systems (Figure 3 and Figure 5) for experimentally already well established targets in the liver were still unavailable, brain or even brain cell-specific drug delivery is still in early infancy today. While meanwhile hepatocyte-specific genetic drug delivery is clinically applied, for none of a range of brain-targeting approaches based on synthetic nanoparticles [188,190,191,263,264,265,266,267,268,269,270,271], or involving rabies virus proteins [272,273] or vectors [274,275,276,277,278,279,280], the key therapeutic efficacy requirements (Figure 5) have been established so far. Nonetheless, they are most useful for experimental pathogenetic research purposes already.
Challenges of brain targeting: Targeted and safe delivery of any nucleic acid-based (siRNA, ASO) drug to specific regions of the brain appears as far greater challenge than for liver targeting or ex vivo blood stem cell modulation (Figure 7). A remarkable spectrum of brain-targeting approaches encompasses synthetic nanoparticles [188,190,191,263,264,265,266,267,268,269,270,271], rabies virus proteins [272,273] or vectors [274,275,276,277,278,279], yet none of these are established with respect to key efficacy requirements (Figure 5). Nonetheless, they are already most useful for pathogenetic research. Recombinant AAV vectors are currently encountering particular interest for brain-directed therapies due to favourable inherent or engineered properties as outlined below.
First experimental steps: A number of first experimental therapeutic studies in mice (section 3) holds promise for further developments with translational potential employing more sophisticated and safe drug delivery systems. These will need to have well-established molecular and cellular mechanisms of action, including carrier ligand – cell surface receptor [183,184,185,186,187,188,281] interactions, as well as clearly characterized in vivo behaviour [190,191,192]. The latter encompasses possible crossing of the blood-brain barrier via transcytosis [282,283,284,285,286], as well as local drug delivery through stereotactic approaches. Furthermore, non-invasive monitoring of therapeutic drug function in patients is highly desirable. For therapeutic proteins this may be enabled by a magnetic resonance imaging (MRI) approach [287,288,289] for real-time reporting of the gene therapy productin vivo by use of an MRI probe that is activated in the presence of therapeutic protein expression.
Advanced AAV-based vector systems: Currently, AAV-based vectors are encountering high interest for brain-targeted therapies [248,290,291,292]. In other fields, e.g. haematology and cardiovascular medicine, genetically engineered and surface-modified (pseudotyped) versions of this vector have been extensively studied previously for therapeutic gene augmentation [258,293] and RNA interference (RNAi) mediated gene silencing [92]. AAVs are considered as vectors of choice for many nervous system targets due to desirable safety profile, extensive basic science and clinical experience from other fields including clinical trials, stable transgene expression in post-mitotic cells, and neuronal tropism. Low immunogenicity of AAVs is a further critically important aspect, and the recently developed system of extracellular vesicle-encapsulated AAVs (EV-AAVs) enables efficient gene transfer even in the presence of pre-existing AAV-neutralizing antibodies in patients [258,294,295,296].
Anatomical barriers against nanoparticle or vector based therapeutics: One study [291] reported that a particular serotype, AAV9, is capable to cross the blood-brain barrier (BBB) [286,297], raising the possibility of intravascular administration as a non-invasive delivery route to achieve widespread CNS gene expression (Figure 7). Crossing of the blood-brain barrier appear to occur via transcytosis [282,283,284,285]. Notably, this same AAV serotype is also capable to enter the myocardium across the tight cardiovascular endothelium (impermeable for other AAV serotypes) and has previously been successfully employed for cardiac gene transfer [258,293] and the first demonstration of cardiac RNA interference (RNAi) therapy [92].
Differential tropism and cell type selectivity: So far, however, very little is known about differential tropism of the currently available AAV pseudotypes for different brain cell types of specific therapeutic interest [298,299]. From previous studies it is well known that genetic engineering of the vector surface i.e. pseudotyping has – in principle - the capacity to alter tropism in a desired direction [248,250,291,292,300,301]. However, recent high-throughput methods have identified the host proteins essential for vector attachment and internalization more comprehensively, and subsequent molecular studies including cryogenic electron microscopy (cryo-EM) have revealed unanticipated complexity and serotype specificity of the cellular vector entry process [302]. Theoretical predictability of the in vivo effects of vector modifications is therefore limited and extensive experimental validation essential [303].
Pillay et al. [304] used an unbiased genetic screen to identify proteins essential for AAV infection and identified a previously uncharacterized type I transmembrane protein, KIAA0319L, which they named AAV receptor (AAVR). They characterized AAVR as capable of rapid endocytosis and trafficking to the trans-Golgi network. AAVR was a critical host factor for all tested AAV serotypes and AAVR-/- mice were resistant to AAV infection, establishing AAVR as a universal receptor for involved in AAV infection. GPR108, a member of the G protein-coupled receptor superfamily, was subsequently identified as another highly conserved AAV entry factor [305]. Among 20 divergent AAVs across all AAV clades, only AAV5 transduction was unaffected in the GPR108 knockout (KO). Thus, this study identified the second of two AAV entry factors conserved between mice and humans and relevant in vitro and in vivo.
Starting from the AAV9 serotype holding promise for trans-BBB therapy, a recombinant AAV-PHP.eB was engineered by insertion of a 7-amino acid peptide and point mutations of neighboring residues into the AAV9 capsid, thereby enhancing potency in the central nervous system [306]. Consideration will be required for translation beyond mouse models, however, because the CNS transduction benefits of AAV-PHP.eB over AAV9 are dependent on administration route and mouse strain [307]. Specifically, AAV-PHP.eB produced higher CNS transduction than AAV9 after intravenous injection, but only in C57BL/6J and not in B6C3 mice. Another study [308] found mutation of certain tyrosine (Tyr) residues on the AAV2 capsid enhanced neuronal transduction in striatum and hippocampus, while analogous Tyr substitutions on AAV5 and AAV8 did not, emphasizing the unforeseeable impact of any modifications with regard to both targeting efficacy and specificity. Similar challenges with regard to clinical translation, generated by species differences, have been extensively investigated with regard to another “hard target” for nucleic acid therapeutics, i.e. the myocardium [8,92,258,294,309,310,311].
One approach towards selective targeting of specific brain cell types engineered AAVs deficient in heparan-sulfate proteoglycans (HSPGs) attachment, but instead recognizing the glutamate receptor 4 (GluA4) through a displayed GluA4-specific DARPin (designed ankyrin repeat protein). When injected into mouse brain, > 90% of transduced cells were interneurons. DARPin mediated selective attachment to GluA4-positive cells, while gene delivery still required expression of AAVR [312].
Another highly innovative strategy employs membrane protein-specific nanobodies inserted into a surface loop of the VP1 capsid protein of AAV2. Nanobodies are single immunoglobulin variable domains of heavy chain antibodies naturally occur in camelids [313]. Nanobodies specific for different membrane proteins dramatically enhanced the transduction of specific target cells by recombinant AAV2. Nanobody-VP1 fusion was incorporated into AAV1, AAV8, and AAV9 and effectively re-directed the target specificity of these AAV serotypes, too.
Transgene expression control: Beyond stability in the blood circulation and capability to cross the blood-brain barrier, it is also desirable to equip the vector with a promoter providing long-term transgene expression in the brain. Maturana et al. [314] have identified and characterized small alphaherpesvirus latency-associated promoters (LAPs) which enabled stable, pan-neuronal transgene transcription and translation from AAV-LAPs in the CNS for 6 months. Thus, these LAPs are suitable candidates for AAV-based CNS gene therapies requiring chronic transgene expression after one-time viral-vector administration.
13. Summary and Outlook
Figure 7 highlights the fact that liver-targeted genetic therapies (both for protein augmentation or gene silencing) are established for several important human metabolic and monogenic disease, since the key problem of proper targeting and stability upon systemic injection has been solved. In principle, genetic modulation of immune cells might expand the currently available arsenal of small molecule drugs, antibodies, and therapeutic cells. Except for ex vivo transduction of progenitor cells, however, immune cell subtype-specific transduction for the treatment of inborn haematological or immune diseases is still immature. The ultimate goal in neurology, of course, would be crossing of the intact blood-brain barrier (e.g. transcytosis), followed by spontaneous vascular egress and selective but efficient entry of the therapeutic nucleic acid into the target cell type where pathogenesis occurs.
It must be kept in mind that each new nucleic acid-based drug and therapeutic target may raise specific and previously unexpected issues, the most critical ones regarding safety (immune activation, off-target effects at the cellular level, systemic mis-targeting and accumulation, delayed-onset safe effects). It should be emphasized that each individual, e.g. siRNA or ASO drug formulation may have a specific side-effect, or particularly high efficacy, which is not ‘group-specific’ i.e. not common to the siRNA or ASO class of drugs in general. Seemingly minute molecular details of a siRNA, ASO, or gene therapy drug may determine whether e.g. immune reactions or thrombopenia will be triggered by this individual compound. High vigilance therefore needs to be focused upon each individual drug from the field of novel nucleic acid-based and epigenetic therapies.
14. Conclusions
There is an expanding spectrum of diseases for which nucleic acid-based therapeutics addressing fundamentally new therapeutic targets are envisaged or under development. Extensive genetic, experimental and clinical work has highlighted important new therapeutic targets possible causal or significantly contributing to development of these grave diseases.
Synergistic with the most remarkable recent progress regarding delivery, efficacy, and safety of nucleic acid-based therapies, past and ongoing large-scale exploration of the noncoding genome for human-specific therapeutic targets is encouraging to proceed with the development and clinical evaluation of such new therapeutic pathways.
Author Contributions
Based upon their extensive research in the respective fields WP and AK devised this review project and jointly drafted the manuscript. SS, RH and UL contributed critical revision of the manuscript.
Funding
The work in the Krichevsky Lab is supported by R01 CA215072 and R01 NS113929 NIH grants, and the Rainwater Foundation/ Tau Consortium award. W.P. received grants B19-006_SE (FKZ 81X2100257 and FKZ 81X2710170 ’Transcriptome analysis of circulating immune cells to improve the assessment of prognosis and the response to novel anti-inflammatory treatments after myocardial infarction’) and B18-004_SE (FKZ 81X2100253, ’Functional role and therapeutic potential of the long noncoding RNA NEAT1-MALAT1 genomic region in cardiovascular inflammation control and atheropathogenesis’) from the German Center for Cardiovascular Research (DZHK).
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
References
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. , Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [PubMed]
- International Human Genome Sequencing, C. , Finishing the euchromatic sequence of the human genome. Nature 2004, 431, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Consortium, E.P. , An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al; et al. , Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef]
- Gerstein, M.B.; Rozowsky, J.; Yan, K.K.; Wang, D.; Cheng, C.; Brown, J.B.; Davis, C.A.; Hillier, L.; Sisu, C.; Li, J.J.; et al. Comparative analysis of the transcriptome across distant species. Nature 2014, 512, 445–448. [Google Scholar] [CrossRef]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermuller, J.; Hofacker, I.L.; et al; et al. , RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. , GENCODE: The reference human genome annotation for The ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef]
- Poller, W.; Dimmeler, S.; Heymans, S.; Zeller, T.; Haas, J.; Karakas, M.; Leistner, D.M.; Jakob, P.; Nakagawa, S.; Blankenberg, S.; et al. , Non-coding RNAs in cardiovascular diseases: Diagnostic and therapeutic perspectives. Eur. Heart J. 2018, 39, 2704–2716. [Google Scholar] [CrossRef]
- Poller, W.; Tank, J.; Skurk, C.; Gast, M. , Cardiovascular RNA interference therapy: The broadening tool and target spectrum. Circ. Res. 2013, 113, 588–602. [Google Scholar] [CrossRef]
- Landmesser, U.; Poller, W.; Tsimikas, S.; Most, P.; Paneni, F.; Luscher, T.F. , From traditional pharmacological towards nucleic acid-based therapies for cardiovascular diseases. Eur. Heart J. 2020, 41, 3884–3899. [Google Scholar] [CrossRef]
- Poller, W.; Heidecker, B.; Ammirati, E.; Kuss, A.W.; Tzvetkova, A.; Poller, W.C.; Skurk, C.; Haghikia, A. Innate Immunity in Cardiovascular Diseases-Identification of Novel Molecular Players and Targets. J. Clin. Med. 2023, 12, 335. [Google Scholar] [CrossRef] [PubMed]
- Gast, M.; Nageswaran, V.; Kuss, A.W.; Tzvetkova, A.; Wang, X.; Mochmann, L.H.; Rad, P.R.; Weiss, S.; Simm, S.; Zeller, T.; et al. , tRNA-like Transcripts from the NEAT1-MALAT1 Genomic Region Critically Influence Human Innate Immunity and Macrophage Functions. Cells 2022, 11, 3970. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.W.; Wray, G.A. , The g-value paradox. Evol. Dev. 2002, 4, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Leypold, N.A.; Speicher, M.R. , Evolutionary conservation in noncoding genomic regions. Trends Genet. 2021, 37, 903–918. [Google Scholar] [CrossRef]
- Ramirez-Colmenero, A.; Oktaba, K.; Fernandez-Valverde, S.L. , Evolution of Genome-Organizing Long Non-coding RNAs in Metazoans. Front. Genet. 2020, 11, 589697. [Google Scholar] [CrossRef]
- Sandmann, C.L.; Schulz, J.F.; Ruiz-Orera, J.; Kirchner, M.; Ziehm, M.; Adami, E.; Marczenke, M.; Christ, A.; Liebe, N.; Greiner, J.; et al. , Evolutionary origins and interactomes of human, young microproteins and small peptides translated from short open reading frames. Mol. Cell 2023, 83, 994–1011. [Google Scholar] [CrossRef]
- Liu, S.J.; Horlbeck, M.A.; Cho, S.W.; Birk, H.S.; Malatesta, M.; He, D.; Attenello, F.J.; Villalta, J.E.; Cho, M.Y.; Chen, Y.; et al; et al. , CRISPRi-based genome-scale identification of functional long noncoding RNA loci in human cells. Science 2017, 355, aah7111. [Google Scholar] [CrossRef]
- Consortium, R.N. , RNAcentral 2021: Secondary structure integration, improved sequence search and new member databases. Nucleic Acids Res. 2021, 49, D212–D220. [Google Scholar] [CrossRef]
- Seifuddin, F.; Singh, K.; Suresh, A.; Judy, J.T.; Chen, Y.C.; Chaitankar, V.; Tunc, I.; Ruan, X.; Li, P.; Chen, Y.; et al. , lncRNAKB, a knowledgebase of tissue-specific functional annotation and trait association of long noncoding RNA. Sci. Data 2020, 7, 326. [Google Scholar] [CrossRef]
- Volders, P.J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. , LNCipedia 5: Towards a reference set of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef]
- Ma, L.; Cao, J.; Liu, L.; Du, Q.; Li, Z.; Zou, D.; Bajic, V.B.; Zhang, Z. , LncBook: A curated knowledgebase of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, 2699. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Zhang, L.; Guo, J.; Niu, Y.; Wu, Y.; Li, H.; Zhao, L.; Li, X.; Teng, X.; Sun, X.; et al; et al. , NONCODEV5: A comprehensive annotation database for long non-coding RNAs. Nucleic Acids Res. 2018, 46, D308–D314. [Google Scholar] [CrossRef] [PubMed]
- Mas-Ponte, D.; Carlevaro-Fita, J.; Palumbo, E.; Hermoso Pulido, T.; Guigo, R.; Johnson, R. , LncATLAS database for subcellular localization of long noncoding RNAs. RNA 2017, 23, 1080–1087. [Google Scholar] [CrossRef]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. , The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. , Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. , Conserved function of lincRNAs in vertebrate embryonic development despite rapid sequence evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef]
- Ross, C.J.; Ulitsky, I. , Discovering functional motifs in long noncoding RNAs. Wiley Interdiscip. Rev. RNA 2022, 13, e1708. [Google Scholar] [CrossRef]
- Ross, C.J.; Rom, A.; Spinrad, A.; Gelbard-Solodkin, D.; Degani, N.; Ulitsky, I. , Uncovering deeply conserved motif combinations in rapidly evolving noncoding sequences. Genome Biol. 2021, 22, 29. [Google Scholar] [CrossRef]
- Degani, N.; Lubelsky, Y.; Perry, R.B.; Ainbinder, E.; Ulitsky, I. , Highly conserved and cis-acting lncRNAs produced from paralogous regions in the center of HOXA and HOXB clusters in the endoderm lineage. PLoS Genet. 2021, 17, e1009681. [Google Scholar] [CrossRef]
- Paabo, S. , The human condition-a molecular approach. Cell 2014, 157, 216–226. [Google Scholar] [CrossRef]
- Essel, E.; Zavala, E.I.; Schulz-Kornas, E.; Kozlikin, M.B.; Fewlass, H.; Vernot, B.; Shunkov, M.V.; Derevianko, A.P.; Douka, K.; Barnes, I.; et al. , Ancient human DNA recovered from a Palaeolithic pendant. Nature 2023, 618, 328–332. [Google Scholar] [CrossRef]
- Bornschein, U.; Zeberg, H.; Enard, W.; Hevers, W.; Paabo, S. , Functional dissection of two amino acid substitutions unique to the human FOXP2 protein. Sci. Rep. 2023, 13, 3747. [Google Scholar] [CrossRef]
- Skov, L.; Peyregne, S.; Popli, D.; Iasi, L.N. M.; Deviese, T.; Slon, V.; Zavala, E.I.; Hajdinjak, M.; Sumer, A.P.; Grote, S.; et al. , Genetic insights into the social organization of Neanderthals. Nature 2022, 610, 519–525. [Google Scholar] [CrossRef]
- Pinson, A.; Xing, L.; Namba, T.; Kalebic, N.; Peters, J.; Oegema, C.E.; Traikov, S.; Reppe, K.; Riesenberg, S.; Maricic, T.; et al. , Human TKTL1 implies greater neurogenesis in frontal neocortex of modern humans than Neanderthals. Science 2022, 377, eabl6422. [Google Scholar] [CrossRef]
- Mora-Bermudez, F.; Kanis, P.; Macak, D.; Peters, J.; Naumann, R.; Xing, L.; Sarov, M.; Winkler, S.; Oegema, C.E.; Haffner, C.; et al. , Longer metaphase and fewer chromosome segregation errors in modern human than Neanderthal brain development. Sci. Adv. 2022, 8, eabn7702. [Google Scholar] [CrossRef] [PubMed]
- Haeggstrom, S.; Ingelman-Sundberg, M.; Paabo, S.; Zeberg, H. , The clinically relevant CYP2C8*3 and CYP2C9*2 haplotype is inherited from Neandertals. Pharmacogenomics J. 2022, 22, 247–249. [Google Scholar] [CrossRef]
- Brown, S.; Massilani, D.; Kozlikin, M.B.; Shunkov, M.V.; Derevianko, A.P.; Stoessel, A.; Jope-Street, B.; Meyer, M.; Kelso, J.; Paabo, S.; et al. , The earliest Denisovans and their cultural adaptation. Nat. Ecol. Evol. 2022, 6, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Hajdinjak, M.; Mafessoni, F.; Skov, L.; Vernot, B.; Hubner, A.; Fu, Q.; Essel, E.; Nagel, S.; Nickel, B.; Richter, J.; et al. , Initial Upper Palaeolithic humans in Europe had recent Neanderthal ancestry. Nature 2021, 592, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, S.; Noonan, J.P.; Paabo, S.; Rubin, E.M. , Accelerated evolution of conserved noncoding sequences in humans. Science 2006, 314, 786. [Google Scholar] [CrossRef] [PubMed]
- Madupe, P.P. Enamel proteins reveal biological sex and genetic variability within southern African Paranthropus. bioRxiv 2023. [Google Scholar] [CrossRef]
- Mortlock, D.P.; Fang, Z.M.; Chandler, K.J.; Hou, Y.; Bickford, L.R.; de Bock, C.E.; Eapen, V.; Clarke, R.A. , Transcriptional Interference Regulates the Evolutionary Development of Speech. Genes 2022, 13. [Google Scholar] [CrossRef]
- Pollard, K.S.; Salama, S.R.; Lambert, N.; Lambot, M.A.; Coppens, S.; Pedersen, J.S.; Katzman, S.; King, B.; Onodera, C.; Siepel, A.; et al. , An RNA gene expressed during cortical development evolved rapidly in humans. Nature 2006, 443, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Pembroke, W.G.; Hartl, C.L.; Geschwind, D.H. , Evolutionary conservation and divergence of the human brain transcriptome. Genome Biol. 2021, 22, 52. [Google Scholar] [CrossRef] [PubMed]
- Espinos, A.; Fernandez-Ortuno, E.; Negri, E.; Borrell, V. , Evolution of genetic mechanisms regulating cortical neurogenesis. Dev. Neurobiol. 2022, 82, 428–453. [Google Scholar] [CrossRef]
- Florio, M.; Heide, M.; Pinson, A.; Brandl, H.; Albert, M.; Winkler, S.; Wimberger, P.; Huttner, W.B.; Hiller, M. , Evolution and cell-type specificity of human-specific genes preferentially expressed in progenitors of fetal neocortex. Elife 2018, 7, e32332. [Google Scholar] [CrossRef]
- Hubisz, M.J.; Pollard, K.S. , Exploring the genesis and functions of Human Accelerated Regions sheds light on their role in human evolution. Curr. Opin. Genet. Dev. 2014, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lindblad-Toh, K.; Garber, M.; Zuk, O.; Lin, M.F.; Parker, B.J.; Washietl, S.; Kheradpour, P.; Ernst, J.; Jordan, G.; Mauceli, E.; et al. , A high-resolution map of human evolutionary constraint using 29 mammals. Nature 2011, 478, 476–482. [Google Scholar] [CrossRef]
- Uebbing, S.; Gockley, J.; Reilly, S.K.; Kocher, A.A.; Geller, E.; Gandotra, N.; Scharfe, C.; Cotney, J.; Noonan, J.P. , Massively parallel discovery of human-specific substitutions that alter enhancer activity. Proc. Natl. Acad. Sci. USA 2021, 118, e2007049118. [Google Scholar] [CrossRef]
- Won, H.; Huang, J.; Opland, C.K.; Hartl, C.L.; Geschwind, D.H. , Human evolved regulatory elements modulate genes involved in cortical expansion and neurodevelopmental disease susceptibility. Nat. Commun. 2019, 10, 2396. [Google Scholar] [CrossRef] [PubMed]
- Gast, M.; Rauch, B.H.; Haghikia, A.; Nakagawa, S.; Haas, J.; Stroux, A.; Schmidt, D.; Schumann, P.; Weiss, S.; Jensen, L.; et al. , Long noncoding RNA NEAT1 modulates immune cell functions and is suppressed in early onset myocardial infarction patients. Cardiovasc. Res. 2019, 115, 1886–1906. [Google Scholar] [CrossRef]
- Drury, S.; Claussen, G.; Zetterman, A.; Moriyama, H.; Moriyama, E.N.; Zhang, L. , Evolution and emergence of primate-specific interferon regulatory factor 9. J. Med. Virol. 2023, 95, e28521. [Google Scholar] [CrossRef]
- Ellwanger, K.; Becker, E.; Kienes, I.; Sowa, A.; Postma, Y.; Cardona Gloria, Y.; Weber, A.N. R.; Kufer, T.A. , The NLR family pyrin domain-containing 11 protein contributes to the regulation of inflammatory signaling. J. Biol. Chem. 2018, 293, 2701–2710. [Google Scholar] [CrossRef]
- van Solingen, C.; Cyr, Y.; Scacalossi, K.R.; de Vries, M.; Barrett, T.J.; de Jong, A.; Gourvest, M.; Zhang, T.; Peled, D.; Kher, R.; et al. , Long noncoding RNA CHROMR regulates antiviral immunity in humans. Proc. Natl. Acad. Sci. USA 2022, 119, e2210321119. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, E.J.; van Solingen, C.; Scacalossi, K.R.; Ouimet, M.; Afonso, M.S.; Prins, J.; Koelwyn, G.J.; Sharma, M.; Ramkhelawon, B.; Carpenter, S.; et al. , The long noncoding RNA CHROME regulates cholesterol homeostasis in primate. Nat. Metab. 2019, 1, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Salvador, A.F.; de Lima, K.A.; Kipnis, J. , Neuromodulation by the immune system: A focus on cytokines. Nat. Rev. Immunol. 2021, 21, 526–541. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, A.M. , Promethean Evolution: A Comparison of the Immune and Neural Systems. Perspect. Biol. Med. 2014, 57, 449–469. [Google Scholar] [CrossRef]
- Malekos, E.; Carpenter, S. , Short open reading frame genes in innate immunity: From discovery to characterization. Trends Immunol. 2022. [Google Scholar] [CrossRef]
- Michieletto, M.F.; Henao-Mejia, J. , Ontogeny and heterogeneity of innate lymphoid cells and the noncoding genome. Immunol. Rev. 2021, 300, 152–166. [Google Scholar] [CrossRef] [PubMed]
- Sadeq, S.; Al-Hashimi, S.; Cusack, C.M.; Werner, A. , Endogenous Double-Stranded RNA. Noncoding RNA 2021, 7, 15. [Google Scholar] [CrossRef]
- Imamura, K.; Imamachi, N.; Akizuki, G.; Kumakura, M.; Kawaguchi, A.; Nagata, K.; Kato, A.; Kawaguchi, Y.; Sato, H.; Yoneda, M.; et al. , Long noncoding RNA NEAT1-dependent SFPQ relocation from promoter region to paraspeckle mediates IL8 expression upon immune stimuli. Mol. Cell 2014, 53, 393–406. [Google Scholar] [CrossRef]
- Gomez, J.A.; Wapinski, O.L.; Yang, Y.W.; Bureau, J.F.; Gopinath, S.; Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. , The NeST long ncRNA controls microbial susceptibility and epigenetic activation of the interferon-gamma locus. Cell 2013, 152, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Collier, S.P.; Henderson, M.A.; Tossberg, J.T.; Aune, T.M. , Regulation of the Th1 genomic locus from Ifng through Tmevpg1 by T-bet. J. Immunol. 2014, 193, 3959–3965. [Google Scholar] [CrossRef]
- Ouyang, J.; Zhu, X.; Chen, Y.; Wei, H.; Chen, Q.; Chi, X.; Qi, B.; Zhang, L.; Zhao, Y.; Gao, G.F.; et al. , NRAV, a long noncoding RNA, modulates antiviral responses through suppression of interferon-stimulated gene transcription. Cell Host Microbe 2014, 16, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Hu, J.; Chen, J.L. , lncRNAs regulate the innate immune response to viral infection. Wiley Interdiscip. Rev. RNA 2016, 7, 129–143. [Google Scholar] [CrossRef]
- Li, Z.; Chao, T.C.; Chang, K.Y.; Lin, N.; Patil, V.S.; Shimizu, C.; Head, S.R.; Burns, J.C.; Rana, T.M. , The long noncoding RNA THRIL regulates TNFalpha expression through its interaction with hnRNPL. Proc. Natl. Acad. Sci. USA 2014, 111, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Plasek, L.M.; Valadkhan, S. , lncRNAs in T lymphocytes: RNA regulation at the heart of the immune response. Am. J. Physiol. Cell Physiol. 2021, 320, C415–C427. [Google Scholar] [CrossRef]
- Kambara, H.; Niazi, F.; Kostadinova, L.; Moonka, D.K.; Siegel, C.T.; Post, A.B.; Carnero, E.; Barriocanal, M.; Fortes, P.; Anthony, D.D.; et al. , Negative regulation of the interferon response by an interferon-induced long non-coding RNA. Nucleic Acids Res. 2014, 42, 10668–10680. [Google Scholar] [CrossRef]
- Winterling, C.; Koch, M.; Koeppel, M.; Garcia-Alcalde, F.; Karlas, A.; Meyer, T.F. , Evidence for a crucial role of a host non-coding RNA in influenza A virus replication. RNA Biol. 2014, 11, 66–75. [Google Scholar] [CrossRef]
- Spurlock, C.F., 3rd; Tossberg, J.T.; Guo, Y.; Collier, S.P.; Crooke, P.S., 3rd; Aune, T.M. , Expression and functions of long noncoding RNAs during human T helper cell differentiation. Nat. Commun. 2015, 6, 6932. [Google Scholar] [CrossRef]
- Aune, T.M.; Spurlock, C.F. , 3rd, Long non-coding RNAs in innate and adaptive immunity. Virus Res. 2016, 212, 146–160. [Google Scholar] [CrossRef]
- Koh, B.H.; Hwang, S.S.; Kim, J.Y.; Lee, W.; Kang, M.J.; Lee, C.G.; Park, J.W.; Flavell, R.A.; Lee, G.R. , Th2 LCR is essential for regulation of Th2 cytokine genes and for pathogenesis of allergic asthma. Proc. Natl. Acad. Sci. USA 2010, 107, 10614–10619. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xue, Y.; Han, Y.; Lin, L.; Wu, C.; Xu, S.; Jiang, Z.; Xu, J.; Liu, Q.; Cao, X. , The STAT3-binding long noncoding RNA lnc-DC controls human dendritic cell differentiation. Science 2014, 344, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Ranzani, V.; Rossetti, G.; Panzeri, I.; Arrigoni, A.; Bonnal, R.J.; Curti, S.; Gruarin, P.; Provasi, E.; Sugliano, E.; Marconi, M.; et al. , The long intergenic noncoding RNA landscape of human lymphocytes highlights the regulation of T cell differentiation by linc-MAF-4. Nat. Immunol. 2015, 16, 318–325. [Google Scholar] [CrossRef]
- Barriocanal, M.; Carnero, E.; Segura, V.; Fortes, P. , Long Non-Coding RNA BST2/BISPR is Induced by IFN and Regulates the Expression of the Antiviral Factor Tetherin. Front. Immunol. 2014, 5, 655. [Google Scholar] [CrossRef] [PubMed]
- Rapicavoli, N.A.; Qu, K.; Zhang, J.; Mikhail, M.; Laberge, R.M.; Chang, H.Y. , A mammalian pseudogene lncRNA at the interface of inflammation and anti-inflammatory therapeutics. Elife 2013, 2, e00762. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Yuan, J.; Zhang, C.; Zhu, Y.; Kuang, X.; Lan, L.; Wang, X. , The STAT3-regulated long non-coding RNA Lethe promote the HCV replication. Biomed. Pharmacother. 2015, 72, 165–171. [Google Scholar] [CrossRef]
- Hu, G.; Tang, Q.; Sharma, S.; Yu, F.; Escobar, T.M.; Muljo, S.A.; Zhu, J.; Zhao, K. , Expression and regulation of intergenic long noncoding RNAs during T cell development and differentiation. Nat. Immunol. 2013, 14, 1190–1198. [Google Scholar] [CrossRef]
- Cui, H.; Xie, N.; Tan, Z.; Banerjee, S.; Thannickal, V.J.; Abraham, E.; Liu, G. , The human long noncoding RNA lnc-IL7R regulates the inflammatory response. Eur. J. Immunol. 2014, 44, 2085–2095. [Google Scholar] [CrossRef]
- Dijkstra, J.M.; Alexander, D.B. , The " NF-k B interacting long noncoding RNA" ( NKILA) transcript is antisense to cancer-associated gene PMEPA1. F1000Res 2015, 4, 96. [Google Scholar] [CrossRef]
- Liu, B.; Sun, L.; Liu, Q.; Gong, C.; Yao, Y.; Lv, X.; Lin, L.; Yao, H.; Su, F.; Li, D.; et al. , A cytoplasmic NF-kappaB interacting long noncoding RNA blocks IkappaB phosphorylation and suppresses breast cancer metastasis. Cancer Cell 2015, 27, 370–381. [Google Scholar] [CrossRef]
- Imam, H.; Bano, A.S.; Patel, P.; Holla, P.; Jameel, S. , The lncRNA NRON modulates HIV-1 replication in a NFAT-dependent manner and is differentially regulated by early and late viral proteins. Sci. Rep. 2015, 5, 8639. [Google Scholar] [CrossRef]
- Krawczyk, M.; Emerson, B.M. , p50-associated COX-2 extragenic RNA (PACER) activates COX-2 gene expression by occluding repressive NF-kappaB complexes. Elife 2014, 3, e01776. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Masood, M.; Gaur, H.; Ahmad, S.; Syed, M.A. , Long non-coding RNA: An immune cells perspective. Life Sci. 2021, 271, 119152. [Google Scholar] [CrossRef] [PubMed]
- Henzinger, H.; Barth, D.A.; Klec, C.; Pichler, M. , Non-Coding RNAs and SARS-Related Coronaviruses. Viruses 2020, 12, 1374. [Google Scholar] [CrossRef]
- Zhu, L.; Lin, X.; Chen, M. , LncRNA NEAT1 correlates with Th17 cells and proinflammatory cytokines, also reflects stenosis degree and cholesterol level in coronary heart disease patients. J. Clin. Lab. Anal. 2022, 36, e23975. [Google Scholar] [CrossRef]
- Vlachogiannis, N.I.; Sachse, M.; Georgiopoulos, G.; Zormpas, E.; Bampatsias, D.; Delialis, D.; Bonini, F.; Galyfos, G.; Sigala, F.; Stamatelopoulos, K.; et al. , Adenosine-to-inosine Alu RNA editing controls the stability of the pro-inflammatory long noncoding RNA NEAT1 in atherosclerotic cardiovascular disease. J. Mol. Cell Cardiol. 2021, 160, 111–120. [Google Scholar] [CrossRef]
- Jing, D.; Zhu, F.; Xu, Z.; Zhang, G.; Zhou, G. , The role of long noncoding RNA (lncRNA) nuclear-enriched abundant transcript 1 (NEAT1) in immune diseases. Transpl. Immunol. 2022, 75, 101716. [Google Scholar] [CrossRef]
- Poller, W.; Heidecker, B.; Ammirati, E.; Kuss, A.W.; Tzvetkova, A.; Poller, W.C.; Skurk, C.; Haghikia, A. , Innate Immunity in Cardiovascular Diseases – Identification of Novel Molecular Players and Targets. Preprints 2022, 2022090487. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.M.; Lopez-Vazquez, A.; Lopez-Larrea, C. , Immune systems evolution. Adv. Exp. Med. Biol. 2012, 739, 237–251. [Google Scholar]
- Gourbal, B.; Pinaud, S.; Beckers, G.J. M.; Van Der Meer, J.W. M.; Conrath, U.; Netea, M.G. , Innate immune memory: An evolutionary perspective. Immunol. Rev. 2018, 283, 21–40. [Google Scholar] [CrossRef]
- Davis, M.M.; Brodin, P. , Rebooting Human Immunology. Annu. Rev. Immunol. 2018, 36, 843–864. [Google Scholar] [CrossRef] [PubMed]
- Suckau, L.; Fechner, H.; Chemaly, E.; Krohn, S.; Hadri, L.; Kockskamper, J.; Westermann, D.; Bisping, E.; Ly, H.; Wang, X.; et al. , Long-term cardiac-targeted RNA interference for the treatment of heart failure restores cardiac function and reduces pathological hypertrophy. Circulation 2009, 119, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Fechner, H.; Sipo, I.; Westermann, D.; Pinkert, S.; Wang, X.; Suckau, L.; Kurreck, J.; Zeichhardt, H.; Muller, O.; Vetter, R.; et al. , Cardiac-targeted RNA interference mediated by an AAV9 vector improves cardiac function in coxsackievirus B3 cardiomyopathy. J Mol Med (Berl) 2008, 86, 987–997. [Google Scholar] [CrossRef]
- Bonauer, A.; Carmona, G.; Iwasaki, M.; Mione, M.; Koyanagi, M.; Fischer, A.; Burchfield, J.; Fox, H.; Doebele, C.; Ohtani, K.; et al. , MicroRNA-92a controls angiogenesis and functional recovery of ischemic tissues in mice. Science 2009, 324, 1710–1713. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.M.; Penzkofer, D.; Teske, R.; Dutzmann, J.; Koch, A.; Bielenberg, W.; Bonauer, A.; Boon, R.A.; Fischer, A.; Bauersachs, J.; et al. , Inhibition of miR-92a improves re-endothelialization and prevents neointima formation following vascular injury. Cardiovasc. Res. 2014, 103, 564–572. [Google Scholar] [CrossRef]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. , MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Sassi, Y.; Avramopoulos, P.; Ramanujam, D.; Gruter, L.; Werfel, S.; Giosele, S.; Brunner, A.D.; Esfandyari, D.; Papadopoulou, A.S.; De Strooper, B.; et al. , Cardiac myocyte miR-29 promotes pathological remodeling of the heart by activating Wnt signaling. Nat. Commun. 2017, 8, 1614. [Google Scholar] [CrossRef]
- Ganesan, J.; Ramanujam, D.; Sassi, Y.; Ahles, A.; Jentzsch, C.; Werfel, S.; Leierseder, S.; Loyer, X.; Giacca, M.; Zentilin, L.; et al. , MiR-378 controls cardiac hypertrophy by combined repression of mitogen-activated protein kinase pathway factors. Circulation 2013, 127, 2097–2106. [Google Scholar] [CrossRef]
- Janssen, H.L.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. , Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef]
- Corsten, M.F.; Papageorgiou, A.; Verhesen, W.; Carai, P.; Lindow, M.; Obad, S.; Summer, G.; Coort, S.L.; Hazebroek, M.; van Leeuwen, R.; et al. , MicroRNA profiling identifies microRNA-155 as an adverse mediator of cardiac injury and dysfunction during acute viral myocarditis. Circ. Res. 2012, 111, 415–425. [Google Scholar] [CrossRef]
- Foinquinos, A.; Batkai, S.; Genschel, C.; Viereck, J.; Rump, S.; Gyongyosi, M.; Traxler, D.; Riesenhuber, M.; Spannbauer, A.; Lukovic, D.; et al. , Preclinical development of a miR-132 inhibitor for heart failure treatment. Nat. Commun. 2020, 11, 633. [Google Scholar] [CrossRef]
- Iwamoto, N.; Butler, D.C. D.; Svrzikapa, N.; Mohapatra, S.; Zlatev, I.; Sah, D.W. Y.; Meena; Standley, S. M.; Lu, G.; Apponi, L.H.; Frank-Kamenetsky, M.; et al., Control of phosphorothioate stereochemistry substantially increases the efficacy of antisense oligonucleotides. Nat. Biotechnol. 2017, 35, 845–851. [Google Scholar] [CrossRef]
- Hara, R.I.; Hisada, Y.; Maeda, Y.; Yokota, T.; Wada, T. , Artificial cationic oligosaccharides for heteroduplex oligonucleotide-type drugs. Sci. Rep. 2018, 8, 4323. [Google Scholar] [CrossRef] [PubMed]
- Farkas, N.; Scaria, P.V.; Woodle, M.C.; Dagata, J.A. , Physical-chemical measurement method development for self-assembled, core-shell nanoparticles. Sci. Rep. 2019, 9, 1655. [Google Scholar] [CrossRef]
- Smith, C.I. E.; Zain, R. , Therapeutic Oligonucleotides: State of the Art. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 605–630. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.M.; Choi, Y.H.; Tu, M.J. , RNA Drugs and RNA Targets for Small Molecules: Principles, Progress, and Challenges. Pharmacol. Rev. 2020, 72, 862–898. [Google Scholar] [CrossRef]
- Graham, M.J.; Lee, R.G.; Brandt, T.A.; Tai, L.J.; Fu, W.; Peralta, R.; Yu, R.; Hurh, E.; Paz, E.; McEvoy, B.W.; et al. , Cardiovascular and Metabolic Effects of ANGPTL3 Antisense Oligonucleotides. N. Engl. J. Med. 2017, 377, 222–232. [Google Scholar] [CrossRef]
- Zlatev, I.; Castoreno, A.; Brown, C.R.; Qin, J.; Waldron, S.; Schlegel, M.K.; Degaonkar, R.; Shulga-Morskaya, S.; Xu, H.; Gupta, S.; et al. , Reversal of siRNA-mediated gene silencing in vivo. Nat. Biotechnol. 2018, 36, 509–511. [Google Scholar] [CrossRef]
- Fechner, H.; Suckau, L.; Kurreck, J.; Sipo, I.; Wang, X.; Pinkert, S.; Loschen, S.; Rekittke, J.; Weger, S.; Dekkers, D.; et al. Highly efficient and specific modulation of cardiac calcium homeostasis by adenovector-derived short hairpin RNA targeting phospholamban. Gene Therapy 2007, 14, 211–218. [Google Scholar] [CrossRef]
- Le, B.T.; Paul, S.; Jastrzebska, K.; Langer, H.; Caruthers, M.H.; Veedu, R.N. , Thiomorpholino oligonucleotides as a robust class of next generation platforms for alternate mRNA splicing. Proc. Natl. Acad. Sci. USA 2022, 119, e2207956119. [Google Scholar] [CrossRef]
- Hwang, J.; Yokota, T. , Recent advancements in exon-skipping therapies using antisense oligonucleotides and genome editing for the treatment of various muscular dystrophies. Expert. Rev. Mol. Med. 2019, 21, e5. [Google Scholar] [CrossRef] [PubMed]
- Alexander, V.J.; Xia, S.; Hurh, E.; Hughes, S.G.; O'Dea, L.; Geary, R.S.; Witztum, J.L.; Tsimikas, S. , N-acetyl galactosamine-conjugated antisense drug to APOC3 mRNA, triglycerides and atherogenic lipoprotein levels. Eur. Heart J. 2019, 40, 2785–2796. [Google Scholar] [CrossRef]
- Ray, K.K.; Wright, R.S.; Kallend, D.; Koenig, W.; Leiter, L.A.; Raal, F.J.; Bisch, J.A.; Richardson, T.; Jaros, M.; Wijngaard, P.L. J.; et al. . Two Phase 3 Trials of Inclisiran in Patients with Elevated LDL Cholesterol. N. Engl. J. Med. 2020, 382, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- Raal, F.J.; Kallend, D.; Ray, K.K.; Turner, T.; Koenig, W.; Wright, R.S.; Wijngaard, P.L. J.; Curcio, D.; Jaros, M.J.; Leiter, L.A.; et al. Inclisiran for the Treatment of Heterozygous Familial Hypercholesterolemia. N. Engl. J. Med. 2020, 382, 1520–1530. [Google Scholar] [CrossRef]
- Benson, M.D.; Waddington-Cruz, M.; Berk, J.L.; Polydefkis, M.; Dyck, P.J.; Wang, A.K.; Plante-Bordeneuve, V.; Barroso, F.A.; Merlini, G.; Obici, L.; et al. , Inotersen Treatment for Patients with Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Gertz, M.A.; Scheinberg, M.; Waddington-Cruz, M.; Heitner, S.B.; Karam, C.; Drachman, B.; Khella, S.; Whelan, C.; Obici, L. , Inotersen for the treatment of adults with polyneuropathy caused by hereditary transthyretin-mediated amyloidosis. Expert. Rev. Clin. Pharmacol. 2019, 12, 701–711. [Google Scholar] [CrossRef]
- Wright, R.S.; Ray, K.K.; Raal, F.J.; Kallend, D.G.; Jaros, M.; Koenig, W.; Leiter, L.A.; Landmesser, U.; Schwartz, G.G.; Friedman, A.; et al. Pooled Patient-Level Analysis of Inclisiran Trials in Patients With Familial Hypercholesterolemia or Atherosclerosis. J. Am. Coll. Cardiol. 2021, 77, 1182–1193. [Google Scholar] [CrossRef]
- Ray, K.K.; Landmesser, U.; Leiter, L.A.; Kallend, D.; Dufour, R.; Karakas, M.; Hall, T.; Troquay, R.P.; Turner, T.; Visseren, F.L.; et al. Inclisiran in Patients at High Cardiovascular Risk with Elevated LDL Cholesterol. N. Engl. J. Med. 2017, 376, 1430–1440. [Google Scholar] [CrossRef]
- Ray, K.K.; Stoekenbroek, R.M.; Kallend, D.; Leiter, L.A.; Landmesser, U.; Wright, R.S.; Wijngaard, P.; Kastelein, J.J. P. , Effect of an siRNA Therapeutic Targeting PCSK9 on Atherogenic Lipoproteins. Circulation 2018, 138, 1304–1316. [Google Scholar] [CrossRef]
- Ray, K.K.; Stoekenbroek, R.M.; Kallend, D.; Nishikido, T.; Leiter, L.A.; Landmesser, U.; Wright, R.S.; Wijngaard, P.L. J.; Kastelein, J.J. P. , Effect of 1 or 2 Doses of Inclisiran on Low-Density Lipoprotein Cholesterol Levels: One-Year Follow-up of the ORION-1 Randomized Clinical Trial. JAMA Cardiol. 2019, 4, 1067–1075. [Google Scholar] [CrossRef]
- Witztum, J.L.; Gaudet, D.; Freedman, S.D.; Alexander, V.J.; Digenio, A.; Williams, K.R.; Yang, Q.; Hughes, S.G.; Geary, R.S.; Arca, M.; et al. Volanesorsen and Triglyceride Levels in Familial Chylomicronemia Syndrome. N. Engl. J. Med. 2019, 381, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Ramms, B.; Patel, S.; Nora, C.; Pessentheiner, A.R.; Chang, M.W.; Green, C.R.; Golden, G.J.; Secrest, P.; Krauss, R.M.; Metallo, C.M.; et al. ApoC-III ASO promotes tissue LPL activity in the absence of apoE-mediated TRL clearance. J. Lipid Res. 2019, 60, 1379–1395. [Google Scholar] [CrossRef]
- Tsimikas, S.; Karwatowska-Prokopczuk, E.; Gouni-Berthold, I.; Tardif, J.C.; Baum, S.J.; Steinhagen-Thiessen, E.; Shapiro, M.D.; Stroes, E.S.; Moriarty, P.M.; Nordestgaard, B.G.; et al. Lipoprotein(a) Reduction in Persons with Cardiovascular Disease. N. Engl. J. Med. 2020, 382, 244–255. [Google Scholar] [CrossRef]
- Viney, N.J.; van Capelleveen, J.C.; Geary, R.S.; Xia, S.; Tami, J.A.; Yu, R.Z.; Marcovina, S.M.; Hughes, S.G.; Graham, M.J.; Crooke, R.M.; et al. Antisense oligonucleotides targeting apolipoprotein(a) in people with raised lipoprotein(a): Two randomised, double-blind, placebo-controlled, dose-ranging trials. Lancet 2016, 388, 2239–2253. [Google Scholar] [CrossRef] [PubMed]
- Tsimikas, S.; Viney, N.J.; Hughes, S.G.; Singleton, W.; Graham, M.J.; Baker, B.F.; Burkey, J.L.; Yang, Q.; Marcovina, S.M.; Geary, R.S.; et al. Antisense therapy targeting apolipoprotein(a): A randomised, double-blind, placebo-controlled phase 1 study. Lancet 2015, 386, 1472–1483. [Google Scholar] [CrossRef]
- Alfano, L.N.; Charleston, J.S.; Connolly, A.M.; Cripe, L.; Donoghue, C.; Dracker, R.; Dworzak, J.; Eliopoulos, H.; Frank, D.E.; Lewis, S.; et al. Long-term treatment with eteplirsen in nonambulatory patients with Duchenne muscular dystrophy. Medicine (Baltimore) 2019, 98, e15858. [Google Scholar] [CrossRef] [PubMed]
- Minamisawa, M.; Claggett, B.; Adams, D.; Kristen, A.V.; Merlini, G.; Slama, M.S.; Dispenzieri, A.; Shah, A.M.; Falk, R.H.; Karsten, V.; et al. Association of Patisiran, an RNA Interference Therapeutic, With Regional Left Ventricular Myocardial Strain in Hereditary Transthyretin Amyloidosis: The APOLLO Study. JAMA Cardiol. 2019, 4, 466–472. [Google Scholar] [CrossRef]
- Ray, K.K.; Nicholls, S.J.; Buhr, K.A.; Ginsberg, H.N.; Johansson, J.O.; Kalantar-Zadeh, K.; Kulikowski, E.; Toth, P.P.; Wong, N.; Sweeney, M.; et al. Committees, Effect of Apabetalone Added to Standard Therapy on Major Adverse Cardiovascular Events in Patients With Recent Acute Coronary Syndrome and Type 2 Diabetes: A Randomized Clinical Trial. JAMA 2020, 323, 1565–1573. [Google Scholar] [CrossRef]
- Elman, L.; Youn, B.; Proud, C.M.; Frey, M.R.; Ajroud-Driss, S.; McCormick, M.E.; Michelson, D.; Cartwright, M.S.; Heiman-Patterson, T.; Choi, J.M.; et al. Real-world Adherence to Nusinersen in Adults with Spinal Muscular Atrophy in the US: A Multi-site Chart Review Study. J. Neuromuscul. Dis. 2022. [Google Scholar] [CrossRef]
- Qiu, J.; Wu, L.; Qu, R.; Jiang, T.; Bai, J.; Sheng, L.; Feng, P.; Sun, J. , History of development of the life-saving drug "Nusinersen" in spinal muscular atrophy. Front. Cell Neurosci. 2022, 16, 942976. [Google Scholar] [CrossRef]
- Adams, D.; Gonzalez-Duarte, A.; O'Riordan, W.D.; Yang, C.C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. , Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.D.; Adams, D.; Kristen, A.; Grogan, M.; Gonzalez-Duarte, A.; Maurer, M.S.; Merlini, G.; Damy, T.; Slama, M.S.; Brannagan, T.H., 3rd; Dispenzieri, A.; et al. , Effects of Patisiran, an RNA Interference Therapeutic, on Cardiac Parameters in Patients With Hereditary Transthyretin-Mediated Amyloidosis. Circulation 2019, 139, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.A. , Treating Disease at the RNA Level with Oligonucleotides. N. Engl. J. Med. 2019, 380, 57–70. [Google Scholar] [CrossRef]
- Taubel, J.; Hauke, W.; Rump, S.; Viereck, J.; Batkai, S.; Poetzsch, J.; Rode, L.; Weigt, H.; Genschel, C.; Lorch, U.; et al. , Novel antisense therapy targeting microRNA-132 in patients with heart failure: Results of a first-in-human Phase 1b randomized, double-blind, placebo-controlled study. Eur. Heart J. 2021, 42, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Deogharia, M.; Gurha, P. , The "guiding" principles of noncoding RNA function. Wiley Interdiscip. Rev. RNA 2022, 13, e1704. [Google Scholar] [CrossRef]
- Degenhardt, F.; Ellinghaus, D.; Juzenas, S.; Lerga-Jaso, J.; Wendorff, M.; Maya-Miles, D.; Uellendahl-Werth, F.; ElAbd, H.; Ruhlemann, M.C.; Arora, J.; et al. Detailed stratified GWAS analysis for severe COVID-19 in four European populations. Hum. Mol. Genet. 2022. [Google Scholar] [CrossRef]
- Dannebaum, R.; Suwalski, P.; Asgharian, H.; Du Zhipei, G.; Lin, H.; Weiner, J.; Holtgrewe, M.; Thibeault, C.; Muller, M.; Wang, X.; et al; et al. , Highly multiplexed immune repertoire sequencing links multiple lymphocyte classes with severity of response to COVID-19. EClinical Medicine 2022, 48, 101438. [Google Scholar] [CrossRef]
- Weiner, J.; Suwalski, P.; Holtgrewe, M.; Rakitko, A.; Thibeault, C.; Muller, M.; Patriki, D.; Quedenau, C.; Kruger, U.; Ilinsky, V.; et al. , Increased risk of severe clinical course of COVID-19 in carriers of HLA-C*04:01. EClinicalMedicine 2021, 40, 101099. [Google Scholar] [CrossRef]
- Schultheiss, H.P.; Baumeier, C.; Pietsch, H.; Bock, C.T.; Poller, W.; Escher, F. , Cardiovascular consequences of viral infections: From COVID to other viral diseases. Cardiovasc. Res. 2021, 117, 2610–2623. [Google Scholar] [CrossRef]
- Prodromidou, K.; Matsas, R. , Species-Specific miRNAs in Human Brain Development and Disease. Front. Cell Neurosci. 2019, 13, 559. [Google Scholar] [CrossRef]
- Prodromidou, K.; Vlachos, I.S.; Gaitanou, M.; Kouroupi, G.; Hatzigeorgiou, A.G.; Matsas, R. MicroRNA-934 is a novel primate-specific small non-coding RNA with neurogenic function during early development. Elife 2020, 9, e50561. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.H.; Park, S.J.; Cho, H.M.; Park, H.R.; Lee, J.R.; Kim, Y.H.; Huh, J.W. , A single mutation in the ACTR8 gene associated with lineage-specific expression in primates. BMC Evol. Biol. 2020, 20, 66. [Google Scholar] [CrossRef]
- Vaid, S.; Huttner, W.B. Transcriptional Regulators and Human-Specific/Primate-Specific Genes in Neocortical Neurogenesis. Int. J. Mol. Sci. 2020, 21, 4614. [Google Scholar] [CrossRef] [PubMed]
- Field, A.R.; Jacobs, F.M. J.; Fiddes, I.T.; Phillips, A.P. R.; Reyes-Ortiz, A.M.; LaMontagne, E.; Whitehead, L.; Meng, V.; Rosenkrantz, J.L.; Olsen, M.; et al. , Structurally Conserved Primate LncRNAs Are Transiently Expressed during Human Cortical Differentiation and Influence Cell-Type-Specific Genes. Stem Cell Reports 2019, 12, 245–257. [Google Scholar] [CrossRef]
- Shao, Y.; Chen, C.; Shen, H.; He, B.Z.; Yu, D.; Jiang, S.; Zhao, S.; Gao, Z.; Zhu, Z.; Chen, X.; et al. , GenTree, an integrated resource for analyzing the evolution and function of primate-specific coding genes. Genome Res. 2019, 29, 682–696. [Google Scholar] [CrossRef]
- Pruunsild, P.; Bengtson, C.P.; Loss, I.; Lohrer, B.; Bading, H. , Expression of the primate-specific LINC00473 RNA in mouse neurons promotes excitability and CREB-regulated transcription. J. Biol. Chem. 2023, 299, 104671. [Google Scholar] [CrossRef] [PubMed]
- Whalen, S.; Pollard, K.S. , Enhancer Function and Evolutionary Roles of Human Accelerated Regions. Annu. Rev. Genet. 2022, 56, 423–439. [Google Scholar] [CrossRef]
- Franchini, L.F.; Pollard, K.S. , Human evolution: The non-coding revolution. BMC Biol. 2017, 15, 89. [Google Scholar] [CrossRef]
- Waters, E.; Pucci, P.; Hirst, M.; Chapman, S.; Wang, Y.; Crea, F.; Heath, C.J. , HAR1: An insight into lncRNA genetic evolution. Epigenomics 2021, 13, 1831–1843. [Google Scholar] [CrossRef]
- Guardiola-Ripoll, M.; Fatjo-Vilas, M. , A Systematic Review of the Human Accelerated Regions in Schizophrenia and Related Disorders: Where the Evolutionary and Neurodevelopmental Hypotheses Converge. Int. J. Mol. Sci. 2023, 24, 3597. [Google Scholar] [CrossRef]
- Girskis, K.M.; Stergachis, A.B.; DeGennaro, E.M.; Doan, R.N.; Qian, X.; Johnson, M.B.; Wang, P.P.; Sejourne, G.M.; Nagy, M.A.; Pollina, E.A.; et al. , Rewiring of human neurodevelopmental gene regulatory programs by human accelerated regions. Neuron 2021, 109, 3239–3251. [Google Scholar] [CrossRef] [PubMed]
- Fulton, S.L.; Bendl, J.; Gameiro-Ros, I.; Fullard, J.F.; Al-Kachak, A.; Lepack, A.E.; Stewart, A.F.; Singh, S.; Poller, W.C.; Bastle, R.M.; et al. , ZBTB7A regulates MDD-specific chromatin signatures and astrocyte-mediated stress vulnerability in orbitofrontal cortex. bioRxiv 2023. [Google Scholar] [CrossRef]
- Luria, V.; Ma, S.; Shibata, M.; Pattabiraman, K.; Sestan, N. , Molecular and cellular mechanisms of human cortical connectivity. Curr. Opin. Neurobiol. 2023, 80, 102699. [Google Scholar] [CrossRef] [PubMed]
- Baird, D.A.; Liu, J.Z.; Zheng, J.; Sieberts, S.K.; Perumal, T.; Elsworth, B.; Richardson, T.G.; Chen, C.Y.; Carrasquillo, M.M.; Allen, M.; et al. , Identifying drug targets for neurological and psychiatric disease via genetics and the brain transcriptome. PLoS Genet. 2021, 17, e1009224. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.Y.; Quan, Y.; Zhang, H.Y. , Human accelerated genome regions with value in medical genetics and drug discovery. Drug Discov. Today 2020, 25, 821–827. [Google Scholar] [CrossRef]
- Xu, K.; Schadt, E.E.; Pollard, K.S.; Roussos, P.; Dudley, J.T. , Genomic and network patterns of schizophrenia genetic variation in human evolutionary accelerated regions. Mol. Biol. Evol. 2015, 32, 1148–1160. [Google Scholar] [CrossRef]
- Doan, R.N.; Shin, T.; Walsh, C.A. , Evolutionary Changes in Transcriptional Regulation: Insights into Human Behavior and Neurological Conditions. Annu. Rev. Neurosci. 2018, 41, 185–206. [Google Scholar] [CrossRef]
- Singh, D.D.; Verma, R.; Parimoo, P.; Sahu, A.; Kumar, V.; Upadhyay, E.; Yadav, D.K. , Potential Therapeutic Relevance of CRISPR/Cas9 Guided Epigenetic Regulations for Neuropsychiatric Disorders. Curr. Top. Med. Chem. 2021, 21, 878–894. [Google Scholar] [CrossRef]
- Preussner, M.; Smith, H.L.; Hughes, D.; Zhang, M.; Emmerichs, A.K.; Scalzitti, S.; Peretti, D.; Swinden, D.; Neumann, A.; Haltenhof, T.; et al. ASO targeting RBM3 temperature-controlled poison exon splicing prevents neurodegeneration in vivo. EMBO Mol. Med. 2023, 15, e17157. [Google Scholar] [CrossRef]
- Nguyen, L.D.; Chau, R.K.; Krichevsky, A.M. , Small Molecule Drugs Targeting Non-Coding RNAs as Treatments for Alzheimer's Disease and Related Dementias. Genes (Basel) 2021, 12, 2005. [Google Scholar] [CrossRef]
- Srinivas, T.; Mathias, C.; Oliveira-Mateos, C.; Guil, S. , Roles of lncRNAs in brain development and pathogenesis: Emerging therapeutic opportunities. Mol. Ther. 2023, 31, 1550–1561. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Duan, Y.; Duan, G.; Wang, Q.; Zhang, K.; Deng, X.; Qian, B.; Gu, J.; Ma, Z.; Zhang, S.; et al. Stress Induces Dynamic, Cytotoxicity-Antagonizing TDP-43 Nuclear Bodies via Paraspeckle LncRNA NEAT1-Mediated Liquid-Liquid Phase Separation. Mol. Cell 2020, 79, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Tollervey, J.R.; Curk, T.; Rogelj, B.; Briese, M.; Cereda, M.; Kayikci, M.; Konig, J.; Hortobagyi, T.; Nishimura, A.L.; Zupunski, V.; et al. Characterizing the RNA targets and position-dependent splicing regulation by TDP-43. Nat. Neurosci. 2011, 14, 452–458. [Google Scholar] [CrossRef]
- Tollervey, J.R.; Wang, Z.; Hortobagyi, T.; Witten, J.T.; Zarnack, K.; Kayikci, M.; Clark, T.A.; Schweitzer, A.C.; Rot, G.; Curk, T.; et al. Analysis of alternative splicing associated with aging and neurodegeneration in the human brain. Genome Res. 2011, 21, 1572–1582. [Google Scholar] [CrossRef]
- Sekar, D.; Tusubira, D.; Ross, K. , TDP-43 and NEAT long non-coding RNA: Roles in neurodegenerative disease. Front. Cell Neurosci. 2022, 16, 954912. [Google Scholar] [CrossRef] [PubMed]
- Kukharsky, M.S.; Ninkina, N.N.; An, H.; Telezhkin, V.; Wei, W.; Meritens, C.R.; Cooper-Knock, J.; Nakagawa, S.; Hirose, T.; Buchman, V.L.; et al. Long non-coding RNA Neat1 regulates adaptive behavioural response to stress in mice. Transl. Psychiatry 2020, 10, 171. [Google Scholar] [CrossRef] [PubMed]
- Oiwa, K.; Watanabe, S.; Onodera, K.; Iguchi, Y.; Kinoshita, Y.; Komine, O.; Sobue, A.; Okada, Y.; Katsuno, M.; Yamanaka, K. , Monomerization of TDP-43 is a key determinant for inducing TDP-43 pathology in amyotrophic lateral sclerosis. Sci. Adv. 2023, 9, eadf6895. [Google Scholar] [CrossRef]
- Butler, A.A.; Johnston, D.R.; Kaur, S.; Lubin, F.D. , Long noncoding RNA NEAT1 mediates neuronal histone methylation and age-related memory impairment. Sci. Signal 2019, 12, eaaw9277. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, J.; Wang, W.; Zhou, J.; Zhang, J. , Depletion of LncRNA NEAT1 Rescues Mitochondrial Dysfunction Through NEDD4L-Dependent PINK1 Degradation in Animal Models of Alzheimer's Disease. Front. Cell Neurosci. 2020, 14, 28. [Google Scholar] [CrossRef]
- Grinman, E.; Nakahata, Y.; Avchalumov, Y.; Espadas, I.; Swarnkar, S.; Yasuda, R.; Puthanveettil, S.V. , Activity-regulated synaptic targeting of lncRNA ADEPTR mediates structural plasticity by localizing Sptn1 and AnkB in dendrites. Sci. Adv. 2021, 7, eabf0605. [Google Scholar] [CrossRef]
- Kobayashi, S.; Takashima, A.; Anzai, K. , The dendritic translocation of translin protein in the form of BC1 RNA protein particles in developing rat hippocampal neurons in primary culture. Biochem. Biophys. Res. Commun. 1998, 253, 448–453. [Google Scholar] [CrossRef]
- Zalfa, F.; Giorgi, M.; Primerano, B.; Moro, A.; Di Penta, A.; Reis, S.; Oostra, B.; Bagni, C. , The fragile X syndrome protein FMRP associates with BC1 RNA and regulates the translation of specific mRNAs at synapses. Cell 2003, 112, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Briz, V.; Restivo, L.; Pasciuto, E.; Juczewski, K.; Mercaldo, V.; Lo, A.C.; Baatsen, P.; Gounko, N.V.; Borreca, A.; Girardi, T.; et al. The non-coding RNA BC1 regulates experience-dependent structural plasticity and learning. Nat. Commun. 2017, 8, 293. [Google Scholar] [CrossRef]
- Zhang, T.; Pang, P.; Fang, Z.; Guo, Y.; Li, H.; Li, X.; Tian, T.; Yang, X.; Chen, W.; Shu, S.; et al. Expression of BC1 Impairs Spatial Learning and Memory in Alzheimer's Disease Via APP Translation. Mol. Neurobiol. 2018, 55, 6007–6020. [Google Scholar] [CrossRef]
- Kamal, A.; Swellam, M.; N, M.S.; Darwish, M.K.; E, M.E.-N. , Long non-coding RNAs BACE1-AS and BC200 in multiple sclerosis and their relation to cognitive function: A gene expression analysis. Brain Res. 2023, 1814, 148424. [Google Scholar] [CrossRef]
- Bampatsias, D.; Mavroeidis, I.; Tual-Chalot, S.; Vlachogiannis, N.I.; Bonini, F.; Sachse, M.; Mavraganis, G.; Mareti, A.; Kritsioti, C.; Laina, A.; et al. Beta-Secretase-1 Antisense RNA Is Associated with Vascular Ageing and Atherosclerotic Cardiovascular Disease. Thromb. Haemost. 2022, 122, 1932–1942. [Google Scholar] [CrossRef] [PubMed]
- Sayad, A.; Najafi, S.; Hussen, B.M.; Abdullah, S.T.; Movahedpour, A.; Taheri, M.; Hajiesmaeili, M. , The Emerging Roles of the beta-Secretase BACE1 and the Long Non-coding RNA BACE1-AS in Human Diseases: A Focus on Neurodegenerative Diseases and Cancer. Front. Aging Neurosci. 2022, 14, 853180. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zhang, R.; Yang, Y.; Wu, E.; Tang, Y.; Zhao, Z.; Li, C.; Yang, L.; Teng, X.; Ye, Y.; et al. , Identification and characterization of long non-coding RNA Carip in modulating spatial learning and memory. Cell Rep. 2022, 38, 110398. [Google Scholar] [CrossRef]
- El Fatimy, R.; Li, S.; Chen, Z.; Mushannen, T.; Gongala, S.; Wei, Z.; Balu, D.T.; Rabinovsky, R.; Cantlon, A.; Elkhal, A.; et al. MicroRNA-132 provides neuroprotection for tauopathies via multiple signaling pathways. Acta Neuropathol. 2018, 136, 537–555. [Google Scholar] [CrossRef]
- Smith, P.Y.; Hernandez-Rapp, J.; Jolivette, F.; Lecours, C.; Bisht, K.; Goupil, C.; Dorval, V.; Parsi, S.; Morin, F.; Planel, E.; et al. miR-132/212 deficiency impairs tau metabolism and promotes pathological aggregation in vivo. Hum. Mol. Genet. 2015, 24, 6721–6735. [Google Scholar] [CrossRef]
- Walgrave, H.; Balusu, S.; Snoeck, S.; Vanden Eynden, E.; Craessaerts, K.; Thrupp, N.; Wolfs, L.; Horre, K.; Fourne, Y.; Ronisz, A.; et al. Restoring miR-132 expression rescues adult hippocampal neurogenesis and memory deficits in Alzheimer's disease. Cell Stem Cell 2021, 28, 1805–1821. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Huang, M.; Guo, L.; Zhu, L.; Hou, J.; Zhang, L.; Pero, A.; Ng, S.; El Gaamouch, F.; Elder, G.; et al. MicroRNA-195 rescues ApoE4-induced cognitive deficits and lysosomal defects in Alzheimer's disease pathogenesis. Mol. Psychiatry 2021, 26, 4687–4701. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Sonn, I.; Goto, M.; Murakami, R.; Sato, T.; Okano, H. , The original strain of SARS-CoV-2, the Delta variant, and the Omicron variant infect microglia efficiently, in contrast to their inability to infect neurons: Analysis using 2D and 3D cultures. Exp. Neurol. 2023, 363, 114379. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, R.; Hirai, Y.; Watanabe, S.; Harada, H.; Suzuki, T.; Hashiguchi, T.; Yanagi, Y.; Shirogane, Y. , Interaction of the Hemagglutinin Stalk Region with Cell Adhesion Molecule (CADM) 1 and CADM2 Mediates the Spread between Neurons and Neuropathogenicity of Measles Virus with a Hyperfusogenic Fusion Protein. J. Virol. 2023, 97, e0034023. [Google Scholar] [CrossRef] [PubMed]
- Dietert, K.; Mahesula, S.; Hegde, S.; Verschelde, J.; Reed, P.; Sprague, S.; Kokovay, E.; Sayre, N.L. , Loss of LRP1 in Adult Neural Stem Cells Impairs Migration to Ischemic Lesions. Stem Cells 2023, 41, 570–577. [Google Scholar] [CrossRef]
- Malik, J.R.; Acharya, A.; Avedissian, S.N.; Byrareddy, S.N.; Fletcher, C.V.; Podany, A.T.; Dyavar, S.R. , ACE-2, TMPRSS2, and Neuropilin-1 Receptor Expression on Human Brain Astrocytes and Pericytes and SARS-CoV-2 Infection Kinetics. Int. J. Mol. Sci. 2023, 24, 8622. [Google Scholar] [CrossRef]
- Bates, E.A.; Lovatt, C.; Plein, A.R.; Davies, J.A.; Siebzehnrubl, F.A.; Parker, A.L. , Engineering Adenoviral Vectors with Improved GBM Selectivity. Viruses 2023, 15, 1086. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, R.; Han, L.; Ke, W.; Shao, K.; Ye, L.; Lou, J.; Jiang, C. , Brain-targeting gene delivery and cellular internalization mechanisms for modified rabies virus glycoprotein RVG29 nanoparticles. Biomaterials 2009, 30, 4195–4202. [Google Scholar] [CrossRef]
- Malik, A.R.; Lips, J.; Gorniak-Walas, M.; Broekaart, D.W. M.; Asaro, A.; Kuffner, M.T. C.; Hoffmann, C.J.; Kikhia, M.; Dopatka, M.; Boehm-Sturm, P.; et al. SorCS2 facilitates release of endostatin from astrocytes and controls post-stroke angiogenesis. Glia 2020, 68, 1304–1316. [Google Scholar] [CrossRef]
- Rodgers, T.M.; Muzzio, N.; Valero, A.; Ahmad, I.; Ludtke, T.U.; Moya, S.E.; Romero, G. , Poly (beta-amino Ester) Nanoparticles Modified with a Rabies Virus-derived peptide for the Delivery of ASCL1 Across a 3D In Vitro Model of the Blood Brain Barrier. ACS Appl. Nano Mater. 2023, 6, 6299–6311. [Google Scholar] [CrossRef]
- Huang, R.; Ke, W.; Han, L.; Liu, Y.; Shao, K.; Ye, L.; Lou, J.; Jiang, C.; Pei, Y. , Brain-targeting mechanisms of lactoferrin-modified DNA-loaded nanoparticles. J. Cereb. Blood Flow. Metab. 2009, 29, 1914–1923. [Google Scholar] [CrossRef] [PubMed]
- Endo-Takahashi, Y.; Kurokawa, R.; Sato, K.; Takizawa, N.; Katagiri, F.; Hamano, N.; Suzuki, R.; Maruyama, K.; Nomizu, M.; Takagi, N.; et al. Ternary Complexes of pDNA, Neuron-Binding Peptide, and PEGylated Polyethyleneimine for Brain Delivery with Nano-Bubbles and Ultrasound. Pharmaceutics 2021, 13, 1003. [Google Scholar] [CrossRef] [PubMed]
- Gast, M.; Rauch, B.H.; Nakagawa, S.; Haghikia, A.; Jasina, A.; Haas, J.; Nath, N.; Jensen, L.; Stroux, A.; Bohm, A.; et al. Immune system-mediated atherosclerosis caused by deficiency of long non-coding RNA MALAT1 in ApoE-/-mice. Cardiovasc. Res. 2019, 115, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S.; Michalik, K.M.; Fischer, A.; Pfisterer, L.; Jae, N.; Winter, C.; Boon, R.A.; Muhly-Reinholz, M.; John, D.; Uchida, S.; et al. , Hematopoietic Deficiency of the Long Noncoding RNA MALAT1 Promotes Atherosclerosis and Plaque Inflammation. Circulation 2019, 139, 1320–1334. [Google Scholar] [CrossRef] [PubMed]
- Mamontova, V.; Trifault, B.; Burger, K. , Compartment-Specific Proximity Ligation Expands the Toolbox to Assess the Interactome of the Long Non-Coding RNA NEAT1. Int. J. Mol. Sci. 2022, 23, 4432. [Google Scholar] [CrossRef]
- Gu, J.; Zhang, B.; An, R.; Qian, W.; Han, L.; Duan, W.; Wang, Z.; Ma, Q. , Molecular Interactions of the Long Noncoding RNA NEAT1 in Cancer. Cancers (Basel) 2022, 14, 4009. [Google Scholar] [CrossRef] [PubMed]
- Park, M.K.; Zhang, L.; Min, K.W.; Cho, J.H.; Yeh, C.C.; Moon, H.; Hormaechea-Agulla, D.; Mun, H.; Ko, S.; Lee, J.W.; et al. , NEAT1 is essential for metabolic changes that promote breast cancer growth and metastasis. Cell Metab. 2021, 33, 2380–2397. [Google Scholar] [CrossRef]
- Naveed, A.; Cooper, J.A.; Li, R.; Hubbard, A.; Chen, J.; Liu, T.; Wilton, S.D.; Fletcher, S.; Fox, A.H. , NEAT1 polyA-modulating antisense oligonucleotides reveal opposing functions for both long non-coding RNA isoforms in neuroblastoma. Cell Mol. Life Sci. 2021, 78, 2213–2230. [Google Scholar] [CrossRef]
- Bhatt, U.; Kretzmann, A.L.; Guedin, A.; Ou, A.; Kobelke, S.; Bond, C.S.; Evans, C.W.; Hurley, L.H.; Mergny, J.L.; Iyer, K.S.; et al. The role of G-Quadruplex DNA in Paraspeckle formation in cancer. Biochimie 2021, 190, 124–131. [Google Scholar] [CrossRef]
- Knutsen, E.; Lellahi, S.M.; Aure, M.R.; Nord, S.; Fismen, S.; Larsen, K.B.; Gabriel, M.T.; Hedberg, A.; Bjorklund, S.S.; Oslo Breast Cancer Research, C.; et al. The expression of the long NEAT1_2 isoform is associated with human epidermal growth factor receptor 2-positive breast cancers. Sci. Rep. 2020, 10, 1277. [Google Scholar] [CrossRef]
- Sunwoo, J.S.; Lee, S.T.; Im, W.; Lee, M.; Byun, J.I.; Jung, K.H.; Park, K.I.; Jung, K.Y.; Lee, S.K.; Chu, K.; et al. Altered Expression of the Long Noncoding RNA NEAT1 in Huntington's Disease. Mol. Neurobiol. 2017, 54, 1577–1586. [Google Scholar] [CrossRef]
- Modic, M.; Grosch, M.; Rot, G.; Schirge, S.; Lepko, T.; Yamazaki, T.; Lee, F.C. Y.; Rusha, E.; Shaposhnikov, D.; Palo, M.; et al. Cross-Regulation between TDP-43 and Paraspeckles Promotes Pluripotency-Differentiation Transition. Mol. Cell 2019, 74, 951–965. [Google Scholar] [CrossRef]
- Boros, F.A.; Vecsei, L.; Klivenyi, P. , NEAT1 on the Field of Parkinson's Disease: Offense, Defense, or a Player on the Bench? J. Parkinsons Dis. 2021, 11, 123–138. [Google Scholar] [CrossRef]
- McCluggage, F.; Fox, A.H. , Paraspeckle nuclear condensates: Global sensors of cell stress? Bioessays 2021, 43, e2000245. [Google Scholar] [CrossRef] [PubMed]
- Gast, M.; Schroen, B.; Voigt, A.; Haas, J.; Kuehl, U.; Lassner, D.; Skurk, C.; Escher, F.; Wang, X.; Kratzer, A.; et al. Long noncoding RNA MALAT1-derived mascRNA is involved in cardiovascular innate immunity. J. Mol. Cell Biol. 2016, 8, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Yamamoto, T.; Hirose, T. , Micellization: A new principle in the formation of biomolecular condensates. Front. Mol. Biosci. 2022, 9, 974772. [Google Scholar] [CrossRef] [PubMed]
- Deforzh, E.; Uhlmann, E.J.; Das, E.; Galitsyna, A.; Arora, R.; Saravanan, H.; Rabinovsky, R.; Wirawan, A.D.; Teplyuk, N.M.; El Fatimy, R.; et al. Promoter and enhancer RNAs regulate chromatin reorganization and activation of miR-10b/HOXD locus, and neoplastic transformation in glioma. Mol. Cell 2022, 82, 1894–1908. [Google Scholar] [CrossRef] [PubMed]
- Deforzh, E.; Kharel, P.; Karelin, A.; Ivanov, P.; Krichevsky, A.M. HOXDeRNA activates a cancerous transcription program and super-enhancers genome-wide. bioRxiv 2023. [Google Scholar] [CrossRef]
- Zheng, C.; Wei, Y.; Zhang, Q.; Sun, M.; Wang, Y.; Hou, J.; Zhang, P.; Lv, X.; Su, D.; Jiang, Y.; et al. Multiomics analyses reveal DARS1-AS1/YBX1-controlled posttranscriptional circuits promoting glioblastoma tumorigenesis/radioresistance. Sci. Adv. 2023, 9, eadf3984. [Google Scholar] [CrossRef]
- Gabriely, G.; Yi, M.; Narayan, R.S.; Niers, J.M.; Wurdinger, T.; Imitola, J.; Ligon, K.L.; Kesari, S.; Esau, C.; Stephens, R.M.; et al. Human glioma growth is controlled by microRNA-10b. Cancer Res. 2011, 71, 3563–3572. [Google Scholar] [CrossRef]
- Lang, M.F.; Yang, S.; Zhao, C.; Sun, G.; Murai, K.; Wu, X.; Wang, J.; Gao, H.; Brown, C.E.; Liu, X.; et al. Genome-wide profiling identified a set of miRNAs that are differentially expressed in glioblastoma stem cells and normal neural stem cells. PLoS ONE 2012, 7, e36248. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Teo, S.; Lam, D.H.; Jeyaseelan, K.; Wang, S. , MicroRNA-10b pleiotropically regulates invasion, angiogenicity and apoptosis of tumor cells resembling mesenchymal subtype of glioblastoma multiforme. Cell Death Dis. 2012, 3, e398. [Google Scholar] [CrossRef] [PubMed]
- El Fatimy, R.; Zhang, Y.; Deforzh, E.; Ramadas, M.; Saravanan, H.; Wei, Z.; Rabinovsky, R.; Teplyuk, N.M.; Uhlmann, E.J.; Krichevsky, A.M. , A nuclear function for an oncogenic microRNA as a modulator of snRNA and splicing. Mol. Cancer 2022, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- Teplyuk, N.M.; Uhlmann, E.J.; Gabriely, G.; Volfovsky, N.; Wang, Y.; Teng, J.; Karmali, P.; Marcusson, E.; Peter, M.; Mohan, A.; et al. Therapeutic potential of targeting microRNA-10b in established intracranial glioblastoma: First steps toward the clinic. EMBO Mol. Med. 2016, 8, 268–287. [Google Scholar] [CrossRef]
- El Fatimy, R.; Subramanian, S.; Uhlmann, E.J.; Krichevsky, A.M. , Genome Editing Reveals Glioblastoma Addiction to MicroRNA-10b. Mol. Ther. 2017, 25, 368–378. [Google Scholar] [CrossRef]
- Zhang, Y.; Rabinovsky, R.; Wei, Z.; El Fatimy, R.; Deforzh, E.; Luan, B.; Peshkin, L.; Uhlmann, E.J.; Krichevsky, A.M. , Secreted PGK1 and IGFBP2 contribute to the bystander effect of miR-10b gene editing in glioma. Mol. Ther. Nucleic Acids 2023, 31, 265–275. [Google Scholar] [CrossRef]
- Wang, Y.; Malik, S.; Suh, H.W.; Xiao, Y.; Deng, Y.; Fan, R.; Huttner, A.; Bindra, R.S.; Singh, V.; Saltzman, W.M.; et al. Anti-seed PNAs targeting multiple oncomiRs for brain tumor therapy. Sci. Adv. 2023, 9, eabq7459. [Google Scholar] [CrossRef]
- Douka, K.; Birds, I.; Wang, D.; Kosteletos, A.; Clayton, S.; Byford, A.; Vasconcelos, E.J. R.; O'Connell, M.J.; Deuchars, J.; Whitehouse, A.; Aspden, J.L. , Cytoplasmic long noncoding RNAs are differentially regulated and translated during human neuronal differentiation. RNA 2021, 27, 1082–1101. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, X.; Zhang, M.; Zheng, Q.; Sun, Z.; He, Y.; Guo, W. , Promising Advances in LINC01116 Related to Cancer. Front. Cell Dev. Biol. 2021, 9, 736927. [Google Scholar] [CrossRef]
- Viswanathan, K.; Dhabhar, F.S. , Stress-induced enhancement of leukocyte trafficking into sites of surgery or immune activation. Proc. Natl. Acad. Sci. USA 2005, 102, 5808–5813. [Google Scholar] [CrossRef]
- Dhabhar, F.S.; Malarkey, W.B.; Neri, E.; McEwen, B.S. , Stress-induced redistribution of immune cells--from barracks to boulevards to battlefields: A tale of three hormones--Curt Richter Award winner. Psychoneuroendocrinology 2012, 37, 1345–1368. [Google Scholar] [CrossRef] [PubMed]
- Glaser, R.; Kiecolt-Glaser, J.K. , Stress-induced immune dysfunction: Implications for health. Nat. Rev. Immunol. 2005, 5, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Jensterle, M.; Herman, R.; Janez, A.; Mahmeed, W.A.; Al-Rasadi, K.; Al-Alawi, K.; Banach, M.; Banerjee, Y.; Ceriello, A.; Cesur, M.; et al. The Relationship between COVID-19 and Hypothalamic-Pituitary-Adrenal Axis: A Large Spectrum from Glucocorticoid Insufficiency to Excess-The CAPISCO International Expert Panel. Int. J. Mol. Sci. 2022, 23, 7326. [Google Scholar] [CrossRef] [PubMed]
- Schiller, M.; Ben-Shaanan, T.L.; Rolls, A. , Neuronal regulation of immunity: Why, how and where? Nat. Rev. Immunol. 2021, 21, 20–36. [Google Scholar] [CrossRef]
- Poller, W.C.; Downey, J.; Mooslechner, A.A.; Khan, N.; Li, L.; Chan, C.T.; McAlpine, C.S.; Xu, C.; Kahles, F.; He, S.; et al. Brain motor and fear circuits regulate leukocytes during acute stress. Nature 2022, 607, 578–584. [Google Scholar] [CrossRef]
- McAlpine, C.S.; Kiss, M.G.; Rattik, S.; He, S.; Vassalli, A.; Valet, C.; Anzai, A.; Chan, C.T.; Mindur, J.E.; Kahles, F.; et al. Sleep modulates haematopoiesis and protects against atherosclerosis. Nature 2019, 566, 383–387. [Google Scholar] [CrossRef]
- Poller, W.C.; Nahrendorf, M.; Swirski, F.K. , Hematopoiesis and Cardiovascular Disease. Circ. Res. 2020, 126, 1061–1085. [Google Scholar] [CrossRef]
- McAlpine, C.S.; Kiss, M.G.; Zuraikat, F.M.; Cheek, D.; Schiroli, G.; Amatullah, H.; Huynh, P.; Bhatti, M.Z.; Wong, L.P.; Yates, A.G.; et al. Sleep exerts lasting effects on hematopoietic stem cell function and diversity. J. Exp. Med. 2022, 219, e20220081. [Google Scholar] [CrossRef]
- Janssen, H.; Kahles, F.; Liu, D.; Downey, J.; Koekkoek, L.L.; Roudko, V.; D'Souza, D.; McAlpine, C.S.; Halle, L.; Poller, W.C.; et al. Monocytes re-enter the bone marrow during fasting and alter the host response to infection. Immunity 2023. [Google Scholar] [CrossRef]
- Vita, G.; Vita, G.L.; Stancanelli, C.; Gentile, L.; Russo, M.; Mazzeo, A. , Genetic neuromuscular disorders: Living the era of a therapeutic revolution. Part 1: Peripheral neuropathies. Neurol. Sci. 2019, 40, 661–669. [Google Scholar] [CrossRef]
- Mathew, V.; Wang, A.K. , Inotersen: New promise for the treatment of hereditary transthyretin amyloidosis. Drug Des. Devel Ther. 2019, 13, 1515–1525. [Google Scholar] [CrossRef]
- Kristen, A.V.; Ajroud-Driss, S.; Conceicao, I.; Gorevic, P.; Kyriakides, T.; Obici, L. , Patisiran, an RNAi therapeutic for the treatment of hereditary transthyretin-mediated amyloidosis. Neurodegener. Dis. Manag. 2019, 9, 5–23. [Google Scholar] [CrossRef]
- Krichevsky, A.; Nguyen, L.; Wei, Z.; Silva, M.; Barberan-Soler, S.; Rabinovsky, R.; Muratore, C.; Stricker, J.; Hortman, C.; Young-Pearse, T.; et al. Small Molecule Regulators of microRNAs Identified by High-Throughput Screen Coupled with High-Throughput Sequencing. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Umuhire Juru, A.; Hargrove, A.E. , Frameworks for targeting RNA with small molecules. J. Biol. Chem. 2021, 296, 100191. [Google Scholar] [CrossRef]
- Childs-Disney, J.L.; Yang, X.; Gibaut, Q.M. R.; Tong, Y.; Batey, R.T.; Disney, M.D. , Targeting RNA structures with small molecules. Nat. Rev. Drug Discov. 2022, 21, 736–762. [Google Scholar] [CrossRef]
- Pilunni, D.; Montrasio, L.; Navarra, P. , When innovation goes fast. The case of hemophilia. Curr. Opin. Pharmacol. 2019, 45, 95–101. [Google Scholar] [CrossRef]
- Majowicz, A.; Nijmeijer, B.; Lampen, M.H.; Spronck, L.; de Haan, M.; Petry, H.; van Deventer, S.J.; Meyer, C.; Tangelder, M.; Ferreira, V. , Therapeutic hFIX Activity Achieved after Single AAV5-hFIX Treatment in Hemophilia B Patients and NHPs with Pre-existing Anti-AAV5 NABs. Mol. Ther. Methods Clin. Dev. 2019, 14, 27–36. [Google Scholar] [CrossRef]
- Peyvandi, F.; Garagiola, I. , Clinical advances in gene therapy updates on clinical trials of gene therapy in haemophilia. Haemophilia 2019, 25, 738–746. [Google Scholar] [CrossRef]
- Samelson-Jones, B.J.; Finn, J.D.; George, L.A.; Camire, R.M.; Arruda, V.R. , Hyperactivity of factor IX Padua (R338L) depends on factor VIIIa cofactor activity. JCI Insight 2019, 5. [Google Scholar] [CrossRef]
- Hunt, B.J.; Corral, J.; Simioni, P. , Should we consider gene therapy in severe inherited thrombophilia?: Communication from the ISTH SSC subcommittee on Physiological Anticoagulants and Thrombophilia. J. Thromb. Haemost. 2023. [Google Scholar] [CrossRef]
- Valentino, L.A.; Kaczmarek, R.; Pierce, G.F.; Noone, D.; O'Mahony, B.; Page, D.; Rotellini, D.; Skinner, M.W. , Hemophilia Gene Therapy: First, Do No Harm. J. Thromb. Haemost. 2023. [Google Scholar] [CrossRef]
- Valentino, L.A.; Ozelo, M.C.; Herzog, R.W.; Key, N.S.; Pishko, A.M.; Ragni, M.V.; Samelson-Jones, B.J.; Lillicrap, D. , A Review of the Rationale for Gene Therapy for Hemophilia A With Inhibitors: One-Shot Tolerance and Treatment? J. Thromb. Haemost. 2023. [Google Scholar] [CrossRef]
- Hajighasemi, S.; Mahdavi Gorabi, A.; Bianconi, V.; Pirro, M.; Banach, M.; Ahmadi Tafti, H.; Reiner, Z.; Sahebkar, A. , A review of gene- and cell-based therapies for familial hypercholesterolemia. Pharmacol. Res. 2019, 143, 119–132. [Google Scholar] [CrossRef]
- Khorkova, O.; Stahl, J.; Joji, A.; Volmar, C.H.; Wahlestedt, C. , Amplifying gene expression with RNA-targeted therapeutics. Nat. Rev. Drug Discov. 2023, 22, 539–561. [Google Scholar] [CrossRef]
- Ray, R.M.; Hansen, A.H.; Slott, S.; Taskova, M.; Astakhova, K.; Morris, K.V. , Control of LDL Uptake in Human Cells by Targeting the LDLR Regulatory Long Non-coding RNA BM450697. Mol. Ther. Nucleic Acids 2019, 17, 264–276. [Google Scholar] [CrossRef]
- Costales, M.G.; Aikawa, H.; Li, Y.; Childs-Disney, J.L.; Abegg, D.; Hoch, D.G.; Pradeep Velagapudi, S.; Nakai, Y.; Khan, T.; Wang, K.W.; et al. Small-molecule targeted recruitment of a nuclease to cleave an oncogenic RNA in a mouse model of metastatic cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 2406–2411. [Google Scholar] [CrossRef]
- Benhamou, R.I.; Suresh, B.M.; Tong, Y.; Cochrane, W.G.; Cavett, V.; Vezina-Dawod, S.; Abegg, D.; Childs-Disney, J.L.; Adibekian, A.; Paegel, B.M.; et al. DNA-encoded library versus RNA-encoded library selection enables design of an oncogenic noncoding RNA inhibitor. Proc. Natl. Acad. Sci. USA 2022, 119. [Google Scholar] [CrossRef]
- Campos, L.J.; Arokiaraj, C.M.; Chuapoco, M.R.; Chen, X.; Goeden, N.; Gradinaru, V.; Fox, A.S. , Advances in AAV technology for delivering genetically encoded cargo to the nonhuman primate nervous system. Curr. Res. Neurobiol. 2023, 4, 100086. [Google Scholar] [CrossRef]
- Williams, J.A.; Paez, P.A. , Improving cell and gene therapy safety and performance using next-generation Nanoplasmid vectors. Mol. Ther. Nucleic Acids 2023, 32, 494–503. [Google Scholar] [CrossRef]
- Ail, D.; Malki, H.; Zin, E.A.; Dalkara, D. , Adeno-Associated Virus (AAV) - Based Gene Therapies for Retinal Diseases: Where are We? Appl. Clin. Genet. 2023, 16, 111–130. [Google Scholar] [CrossRef]
- Varshney, S.; Alam, A.; Kaur, A.; Dhoundiyal, S. , Niosomes: A Smart Drug Delivery System for Brain Targeting. Pharm. Nanotechnol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Narasipura, E.A.; VanKeulen-Miller, R.; Ma, Y.; Fenton, O.S. , Ongoing Clinical Trials of Nonviral siRNA Therapeutics. Bioconjug Chem. 2023. [Google Scholar] [CrossRef]
- Alterman, J.F.; Godinho, B.; Hassler, M.R.; Ferguson, C.M.; Echeverria, D.; Sapp, E.; Haraszti, R.A.; Coles, A.H.; Conroy, F.; Miller, R.; et al. A divalent siRNA chemical scaffold for potent and sustained modulation of gene expression throughout the central nervous system. Nat. Biotechnol. 2019, 37, 884–894. [Google Scholar] [CrossRef]
- Conroy, F.; Miller, R.; Alterman, J.F.; Hassler, M.R.; Echeverria, D.; Godinho, B.; Knox, E.G.; Sapp, E.; Sousa, J.; Yamada, K.; et al. Chemical engineering of therapeutic siRNAs for allele-specific gene silencing in Huntington's disease models. Nat. Commun. 2022, 13, 5802. [Google Scholar] [CrossRef]
- Fechner, H.; Haack, A.; Wang, H.; Wang, X.; Eizema, K.; Pauschinger, M.; Schoemaker, R.; vanVeghel, R.; Houtsmuller, A.; Schultheiss, H.-P.; Lamers, J.; Poller, W. , Expression of Coxsackie-adenovirus-receptor and av-integrin does not correlate with adenovector targeting in vivo indicating anatomical vector barriers. Gene Therapy 1999, 6, 1520–1535. [Google Scholar] [CrossRef]
- Noutsias, M.; Fechner, H.; Jonge, H. d.; Wang, X.; Dekkers, D.; Houtsmuller, A.; Pauschinger, M.; Bergelson, J.; Warraich, R.; Yacoub, M.; et al. Human Coxsackie-Adenovirus-Receptor is Co-Localized with Integrins avb3 and avb5 on the Cardiomyocyte Sarcolemma and Upregulated in Dilated Cardiomyopathy - Implications for Cardiotropic Viral Infections. Circulation 2001, 104, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Fechner, H.; Noutsias, M.; Tschoepe, C.; Hinze, K.; Wang, X.; Escher, F.; Pauschinger, M.; Dekkers, D.; Vetter, R.; Paul, M.; et al. Induction of coxsackievirus-adenovirus-receptor expression during myocardial tissue formation and remodeling: Identification of a cell-to-cell contact-dependent regulatory mechanism. Circulation 2003, 107, 876–882. [Google Scholar] [CrossRef]
- Ishikawa, K.; Weber, T.; Hajjar, R.J. , Human Cardiac Gene Therapy. Circ. Res. 2018, 123, 601–613. [Google Scholar] [CrossRef]
- O'Reilly, M.; Shipp, A.; Rosenthal, E.; Jambou, R.; Shih, T.; Montgomery, M.; Gargiulo, L.; Patterson, A.; Corrigan-Curay, J. , NIH oversight of human gene transfer research involving retroviral, lentiviral, and adeno-associated virus vectors and the role of the NIH recombinant DNA advisory committee. Methods Enzymol. 2012, 507, 313–335. [Google Scholar] [PubMed]
- Consiglieri, G.; Bernardo, M.E.; Brunetti-Pierri, N.; Aiuti, A. , Ex Vivo and In Vivo Gene Therapy for Mucopolysaccharidoses: State of the Art. Hematol. Oncol. Clin. North. Am. 2022, 36, 865–878. [Google Scholar] [CrossRef]
- Papaioannou, I.; Owen, J.S.; Yanez-Munoz, R.J. , Clinical applications of gene therapy for rare diseases: A review. Int. J. Exp. Pathol. 2023. [Google Scholar] [CrossRef]
- Grossen, P.; Skaripa Koukelli, I.; van Haasteren, J.; Machado, A.H. E.; Duerr, C. , The Ice Age - A Review on Formulation of Adeno-Associated Virus Therapeutics. Eur. J. Pharm. Biopharm. 2023. [Google Scholar] [CrossRef]
- Ke, W.; Shao, K.; Huang, R.; Han, L.; Liu, Y.; Li, J.; Kuang, Y.; Ye, L.; Lou, J.; Jiang, C. , Gene delivery targeted to the brain using an Angiopep-conjugated polyethyleneglycol-modified polyamidoamine dendrimer. Biomaterials 2009, 30, 6976–6985. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Singh, J. , In vitro and in vivo optimization of liposomal nanoparticles based brain targeted vgf gene therapy. Int. J. Pharm. 2021, 608, 121095. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, M.C. P.; Cronin, M.F.; Cryan, J.F.; O'Driscoll, C.M. , Modified cyclodextrin-based nanoparticles mediated delivery of siRNA for huntingtin gene silencing across an in vitro BBB model. Eur. J. Pharm. Biopharm. 2021, 169, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, T.; Shan, Y.; Li, G.; Ni, X.; Chen, X.; Hu, X.; Lin, L.; Li, Y.; Guan, Y.; et al. Therapeutic reversal of Huntington's disease by in vivo self-assembled siRNAs. Brain 2021, 144, 3421–3435. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, K.E.; Aouida, M.; Gupta, V.; Abdesselem, H.; El-Agnaf, O.M. A. , Development of non-viral vectors for neuronal-targeted delivery of CRISPR-Cas9 RNA-proteins as a therapeutic strategy for neurological disorders. Biomater. Sci. 2022, 10, 4959–4977. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Xie, R.; Burger, J.C.; Tong, Y.; Gong, S. , Overcoming the Blood-Brain Barrier for Gene Therapy via Systemic Administration of GSH-Responsive Silica Nanocapsules. Adv. Mater. 2023, 35, e2208018. [Google Scholar] [CrossRef] [PubMed]
- Wohlfart, S.; Gelperina, S.; Kreuter, J. , Transport of drugs across the blood-brain barrier by nanoparticles. J. Control Release 2012, 161, 264–273. [Google Scholar] [CrossRef]
- Andrieux, K.; Couvreur, P. , Polyalkylcyanoacrylate nanoparticles for delivery of drugs across the blood-brain barrier. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol 2009, 1, 463–474. [Google Scholar] [CrossRef]
- Huang, S.; Li, J.; Han, L.; Liu, S.; Ma, H.; Huang, R.; Jiang, C. , Dual targeting effect of Angiopep-2-modified, DNA-loaded nanoparticles for glioma. Biomaterials 2011, 32, 6832–6838. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Hu, S.; Wu, Z.; He, M. , Therapeutic effects of engineered exosome-based miR-25 and miR-181a treatment in spinocerebellar ataxia type 3 mice by silencing ATXN3. Mol. Med. 2023, 29, 96. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, R.G. R.; Granja, A.; Loureiro, J.A.; Pereira, M.C.; Pinheiro, M.; Neves, A.R.; Reis, S. , RVG29-Functionalized Lipid Nanoparticles for Quercetin Brain Delivery and Alzheimer's Disease. Pharm. Res. 2020, 37, 139. [Google Scholar] [CrossRef]
- Sumser, A.; Joesch, M.; Jonas, P.; Ben-Simon, Y. , Fast, high-throughput production of improved rabies viral vectors for specific, efficient and versatile transsynaptic retrograde labeling. Elife, 2022; 11. [Google Scholar] [CrossRef]
- Ghanem, A.; Conzelmann, K.K. , G gene-deficient single-round rabies viruses for neuronal circuit analysis. Virus Res. 2016, 216, 41–54. [Google Scholar] [CrossRef]
- Salin, P.; Blondel, D.; Kerkerian-Le Goff, L.; Coulon, P. , Golgi staining-like retrograde labeling of brain circuits using rabies virus: Focus onto the striatonigral neurons. J. Neurosci. Methods 2020, 344, 108872. [Google Scholar] [CrossRef]
- Sun, L.; Tang, Y.; Yan, K.; Yu, J.; Zou, Y.; Xu, W.; Xiao, K.; Zhang, Z.; Li, W.; Wu, B.; et al. Differences in neurotropism and neurotoxicity among retrograde viral tracers. Mol. Neurodegener. 2019, 14, 8. [Google Scholar] [CrossRef]
- Zhu, X.; Lin, K.; Liu, Q.; Yue, X.; Mi, H.; Huang, X.; He, X.; Wu, R.; Zheng, D.; Wei, D.; et al. Rabies Virus Pseudotyped with CVS-N2C Glycoprotein as a Powerful Tool for Retrograde Neuronal Network Tracing. Neurosci. Bull. 2020, 36, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.Z.; Li, L.; Ma, W.Y.; Yang, X.; Han, Z.P.; Luo, N.S.; Wang, J.; Xu, F.Q. , A rabies virus-based toolkit for efficient retrograde labeling and monosynaptic tracing. Neural Regen. Res. 2023, 18, 1827–1833. [Google Scholar] [CrossRef]
- Zhong, C.; Chen, Z.; Xia, Y.; Wu, J.; Zhang, F.; Cheng, C.; Wu, X.; Zhuang, Y.; Xiao, X. , Treatment of experimental autoimmune encephalomyelitis using AAV gene therapy by blocking T cell costimulatory pathways. Mol. Ther. Methods Clin. Dev. 2022, 25, 461–475. [Google Scholar] [CrossRef]
- Sandin, E.S.; Folberth, J.; Muller-Fielitz, H.; Pietrzik, C.U.; Herold, E.; Willnow, T.E.; Pfluger, P.T.; Nogueiras, R.; Prevot, V.; Krey, T.; et al. Is LRP2 Involved in Leptin Transport over the Blood-Brain Barrier and Development of Obesity? Int. J. Mol. Sci. 2021, 22, 4998. [Google Scholar] [CrossRef]
- Alnaqbi, N.; Mohammad, M.G.; Hamoudi, R.; Mabondzo, A.; Harati, R. , Molecular Heterogeneity of the Brain Endothelium. Curr. Issues Mol. Biol. 2023, 45, 3462–3478. [Google Scholar] [CrossRef]
- Mollgard, K.; Beinlich, F.R. M.; Kusk, P.; Miyakoshi, L.M.; Delle, C.; Pla, V.; Hauglund, N.L.; Esmail, T.; Rasmussen, M.K.; Gomolka, R.S.; et al. A mesothelium divides the subarachnoid space into functional compartments. Science 2023, 379, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, K.; Neyazi, B.; Stein, K.P.; Haghikia, A.; Sandalcioglu, I.E. , Is the central nervous system enclosed by a mesothel? Ther. Adv. Neurol. Disord. 2023, 16, 17562864231180335. [Google Scholar] [CrossRef]
- Hartmann, K.; Stein, K.P.; Neyazi, B.; Sandalcioglu, I.E. , First in vivo visualization of the human subarachnoid space and brain cortex via optical coherence tomography. Ther. Adv. Neurol. Disord. 2019, 12, 1756286419843040. [Google Scholar] [CrossRef] [PubMed]
- Andreone, B.J.; Chow, B.W.; Tata, A.; Lacoste, B.; Ben-Zvi, A.; Bullock, K.; Deik, A.A.; Ginty, D.D.; Clish, C.B.; Gu, C. , Blood-Brain Barrier Permeability Is Regulated by Lipid Transport-Dependent Suppression of Caveolae-Mediated Transcytosis. Neuron 2017, 94, 581–594. [Google Scholar] [CrossRef]
- Taghian, T.; Batista, A.R.; Kamper, S.; Caldwell, M.; Lilley, L.; Li, H.; Rodriguez, P.; Mesa, K.; Zheng, S.; King, R.M.; et al. Real-time MR tracking of AAV gene therapy with betagal-responsive MR probe in a murine model of GM1-gangliosidosis. Mol. Ther. Methods Clin. Dev. 2021, 23, 128–134. [Google Scholar] [PubMed]
- Gray-Edwards, H.L.; Maguire, A.S.; Salibi, N.; Ellis, L.E.; Voss, T.L.; Diffie, E.B.; Koehler, J.; Randle, A.N.; Taylor, A.R.; Brunson, B.L.; et al. 7T MRI Predicts Amelioration of Neurodegeneration in the Brain after AAV Gene Therapy. Mol. Ther. Methods Clin. Dev. 2020, 17, 258–270. [Google Scholar] [CrossRef]
- Gray-Edwards, H.L.; Jiang, X.; Randle, A.N.; Taylor, A.R.; Voss, T.L.; Johnson, A.K.; McCurdy, V.J.; Sena-Esteves, M.; Ory, D.S.; Martin, D.R. , Lipidomic Evaluation of Feline Neurologic Disease after AAV Gene Therapy. Mol. Ther. Methods Clin. Dev. 2017, 6, 135–142. [Google Scholar] [CrossRef]
- Hudry, E.; Vandenberghe, L.H. , Therapeutic AAV Gene Transfer to the Nervous System: A Clinical Reality. Neuron 2019, 101, 839–862. [Google Scholar] [CrossRef]
- Saraiva, J.; Nobre, R.J.; Pereira de Almeida, L. , Gene therapy for the CNS using AAVs: The impact of systemic delivery by AAV9. J. Control Release 2016, 241, 94–109. [Google Scholar] [CrossRef]
- Rosenberg, J.B.; Kaplitt, M.G.; De, B.P.; Chen, A.; Flagiello, T.; Salami, C.; Pey, E.; Zhao, L.; Ricart Arbona, R.J.; Monette, S.; et al. AAVrh.10-Mediated APOE2 Central Nervous System Gene Therapy for APOE4-Associated Alzheimer's Disease. Hum. Gene Ther. Clin. Dev. 2018, 29, 24–47. [Google Scholar] [CrossRef]
- Hammoudi, N.; Ishikawa, K.; Hajjar, R.J. , Adeno-associated virus-mediated gene therapy in cardiovascular disease. Curr. Opin. Cardiol. 2015, 30, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; La Salvia, S.; Liang, Y.; Adamiak, M.; Kohlbrenner, E.; Jeong, D.; Chepurko, E.; Ceholski, D.; Lopez-Gordo, E.; Yoon, S.; et al. Extracellular Vesicle-Encapsulated Adeno-Associated Viruses for Therapeutic Gene Delivery to the Heart. Circulation 2023, 148, 405–425. [Google Scholar] [CrossRef] [PubMed]
- Aronson, S.J.; Veron, P.; Collaud, F.; Hubert, A.; Delahais, V.; Honnet, G.; de Knegt, R.J.; Junge, N.; Baumann, U.; Di Giorgio, A.; et al. Prevalence and Relevance of Pre-Existing Anti-Adeno-Associated Virus Immunity in the Context of Gene Therapy for Crigler-Najjar Syndrome. Hum. Gene Ther. 2019, 30, 1297–1305. [Google Scholar] [CrossRef]
- Xiao, W.; Gao, G.; Ling, C.; Herzog, R.W.; Xiao, X.; Samulski, R.J. , Impact of neutralizing antibodies against AAV is a key consideration in gene transfer to nonhuman primates. Nat. Med. 2018, 24, 699. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Chiorini, J.A. , AAV transcytosis through barrier epithelia and endothelium. Mol. Ther. 2006, 13, 506–516. [Google Scholar] [PubMed]
- Dennys, C.N.; Sierra-Delgado, J.A.; Ray, S.S.; Hartlaub, A.M.; Roussel, F.S.; Rodriguez, Y.; Meyer, K. , In vitro Modeling for Neurological Diseases using Direct Conversion from Fibroblasts to Neuronal Progenitor Cells and Differentiation into Astrocytes. J. Vis. Exp. 2021. [Google Scholar] [CrossRef]
- Farfara, D.; Sooliman, M.; Avrahami, L.; Royal, T.G.; Amram, S.; Rozenstein-Tsalkovich, L.; Trudler, D.; Blanga-Kanfi, S.; Eldar-Finkelman, H.; Pahnke, J.; et al. Physiological expression of mutated TAU impaired astrocyte activity and exacerbates beta-amyloid pathology in 5xFAD mice. J. Neuroinflamm. 2023, 20, 174. [Google Scholar] [CrossRef]
- Zhang, L.; Rossi, A.; Lange, L.; Meumann, N.; Koitzsch, U.; Christie, K.; Nesbit, M.A.; Moore, C.B. T.; Hacker, U.T.; Morgan, M.; et al. Capsid Engineering Overcomes Barriers Toward Adeno-Associated Virus Vector-Mediated Transduction of Endothelial Cells. Hum. Gene Ther. 2019, 30, 1284–1296. [Google Scholar] [CrossRef]
- Greig, J.A.; Limberis, M.P.; Bell, P.; Chen, S.J.; Calcedo, R.; Rader, D.J.; Wilson, J.M. , Non-Clinical Study Examining AAV8.TBG.hLDLR Vector-Associated Toxicity in Chow-Fed Wild-Type and LDLR(+/-) Rhesus Macaques. Hum. Gene Ther. Clin. Dev. 2017, 28, 39–50. [Google Scholar] [CrossRef]
- Meyer, N.L.; Chapman, M.S. , Adeno-associated virus (AAV) cell entry: Structural insights. Trends Microbiol. 2022, 30, 432–451. [Google Scholar] [PubMed]
- Peters, C.W.; Maguire, C.A.; Hanlon, K.S. , Delivering AAV to the Central Nervous and Sensory Systems. Trends Pharmacol. Sci. 2021, 42, 461–474. [Google Scholar]
- Pillay, S.; Meyer, N.L.; Puschnik, A.S.; Davulcu, O.; Diep, J.; Ishikawa, Y.; Jae, L.T.; Wosen, J.E.; Nagamine, C.M.; Chapman, M.S.; et al. An essential receptor for adeno-associated virus infection. Nature 2016, 530, 108–112. [Google Scholar]
- Dudek, A.M.; Zabaleta, N.; Zinn, E.; Pillay, S.; Zengel, J.; Porter, C.; Franceschini, J.S.; Estelien, R.; Carette, J.E.; Zhou, G.L.; et al. GPR108 Is a Highly Conserved AAV Entry Factor. Mol. Ther. 2020, 28, 367–381. [Google Scholar]
- Jang, S.; Shen, H.K.; Ding, X.; Miles, T.F.; Gradinaru, V. , Structural basis of receptor usage by the engineered capsid AAV-PHP.eB. Mol. Ther. Methods Clin. Dev. 2022, 26, 343–354. [Google Scholar]
- Mathiesen, S.N.; Lock, J.L.; Schoderboeck, L.; Abraham, W.C.; Hughes, S.M. , CNS Transduction Benefits of AAV-PHP.eB over AAV9 Are Dependent on Administration Route and Mouse Strain. Mol. Ther. Methods Clin. Dev. 2020, 19, 447–458. [Google Scholar]
- Kanaan, N.M.; Sellnow, R.C.; Boye, S.L.; Coberly, B.; Bennett, A.; Agbandje-McKenna, M.; Sortwell, C.E.; Hauswirth, W.W.; Boye, S.E.; Manfredsson, F.P. , Rationally Engineered AAV Capsids Improve Transduction and Volumetric Spread in the CNS. Mol. Ther. Nucleic Acids 2017, 8, 184–197. [Google Scholar]
- Chamberlain, K.; Riyad, J.M.; Garnett, T.; Kohlbrenner, E.; Mookerjee, A.; Elmastour, F.; Benard, L.; Chen, J.; VandenDriessche, T.; Chuah, M.K.; et al. A Calsequestrin Cis-Regulatory Motif Coupled to a Cardiac Troponin T Promoter Improves Cardiac Adeno-Associated Virus Serotype 9 Transduction Specificity. Hum. Gene Ther. 2018, 29, 927–937. [Google Scholar] [CrossRef]
- Katz, M.G.; Fargnoli, A.S.; Kendle, A.P.; Hajjar, R.J.; Bridges, C.R. , Gene Therapy in Cardiac Surgery: Clinical Trials, Challenges, and Perspectives. Ann. Thorac. Surg. 2016, 101, 2407–2416. [Google Scholar] [PubMed]
- Hulot, J.S.; Ishikawa, K.; Hajjar, R.J. , Gene therapy for the treatment of heart failure: Promise postponed. Eur. Heart J. 2016, 37, 1651–1658. [Google Scholar]
- Hartmann, J.; Thalheimer, F.B.; Hopfner, F.; Kerzel, T.; Khodosevich, K.; Garcia-Gonzalez, D.; Monyer, H.; Diester, I.; Buning, H.; Carette, J.E.; et al. GluA4-Targeted AAV Vectors Deliver Genes Selectively to Interneurons while Relying on the AAV Receptor for Entry. Mol. Ther. Methods Clin. Dev. 2019, 14, 252–260. [Google Scholar] [PubMed]
- Eichhoff, A.M.; Borner, K.; Albrecht, B.; Schafer, W.; Baum, N.; Haag, F.; Korbelin, J.; Trepel, M.; Braren, I.; Grimm, D.; et al. Nanobody-Enhanced Targeting of AAV Gene Therapy Vectors. Mol. Ther. Methods Clin. Dev. 2019, 15, 211–220. [Google Scholar] [PubMed]
- Maturana, C.J.; Verpeut, J.L.; Pisano, T.J.; Dhanerawala, Z.M.; Esteves, A.; Enquist, L.W.; Engel, E.A. , Small Alphaherpesvirus Latency-Associated Promoters Drive Efficient and Long-Term Transgene Expression in the CNS. Mol. Ther. Methods Clin. Dev. 2020, 17, 843–857. [Google Scholar] [PubMed]
Figure 1.
- Evolutionary expansion of the noncoding genome in higher organisms. A, The basic molecules of life already found in the earliest and most simple organisms are increasingly supplemented, during the evolution to complex species, by molecules needed for correct embryonic development and homeostatic stability of their morphology and functions. B, Whereas the number of protein-coding genes remains similar from simple to complex species, it is the noncoding part of the genome that increases dramatically with morphological complexity to >98% in humans. C, Few types of noncoding RNAs arising from the noncoding genome have been phylogenetically mapped in depth. Thus, investigation of microRNA (miRNA) family evolution revealed impressive increases with the advent of vertebrates, and ancient miRNAs families can well be distinguished from those more recently arising. (Modified from Poller et al. 2013 by permission of Circ. Res.).
Figure 1.
- Evolutionary expansion of the noncoding genome in higher organisms. A, The basic molecules of life already found in the earliest and most simple organisms are increasingly supplemented, during the evolution to complex species, by molecules needed for correct embryonic development and homeostatic stability of their morphology and functions. B, Whereas the number of protein-coding genes remains similar from simple to complex species, it is the noncoding part of the genome that increases dramatically with morphological complexity to >98% in humans. C, Few types of noncoding RNAs arising from the noncoding genome have been phylogenetically mapped in depth. Thus, investigation of microRNA (miRNA) family evolution revealed impressive increases with the advent of vertebrates, and ancient miRNAs families can well be distinguished from those more recently arising. (Modified from Poller et al. 2013 by permission of Circ. Res.).
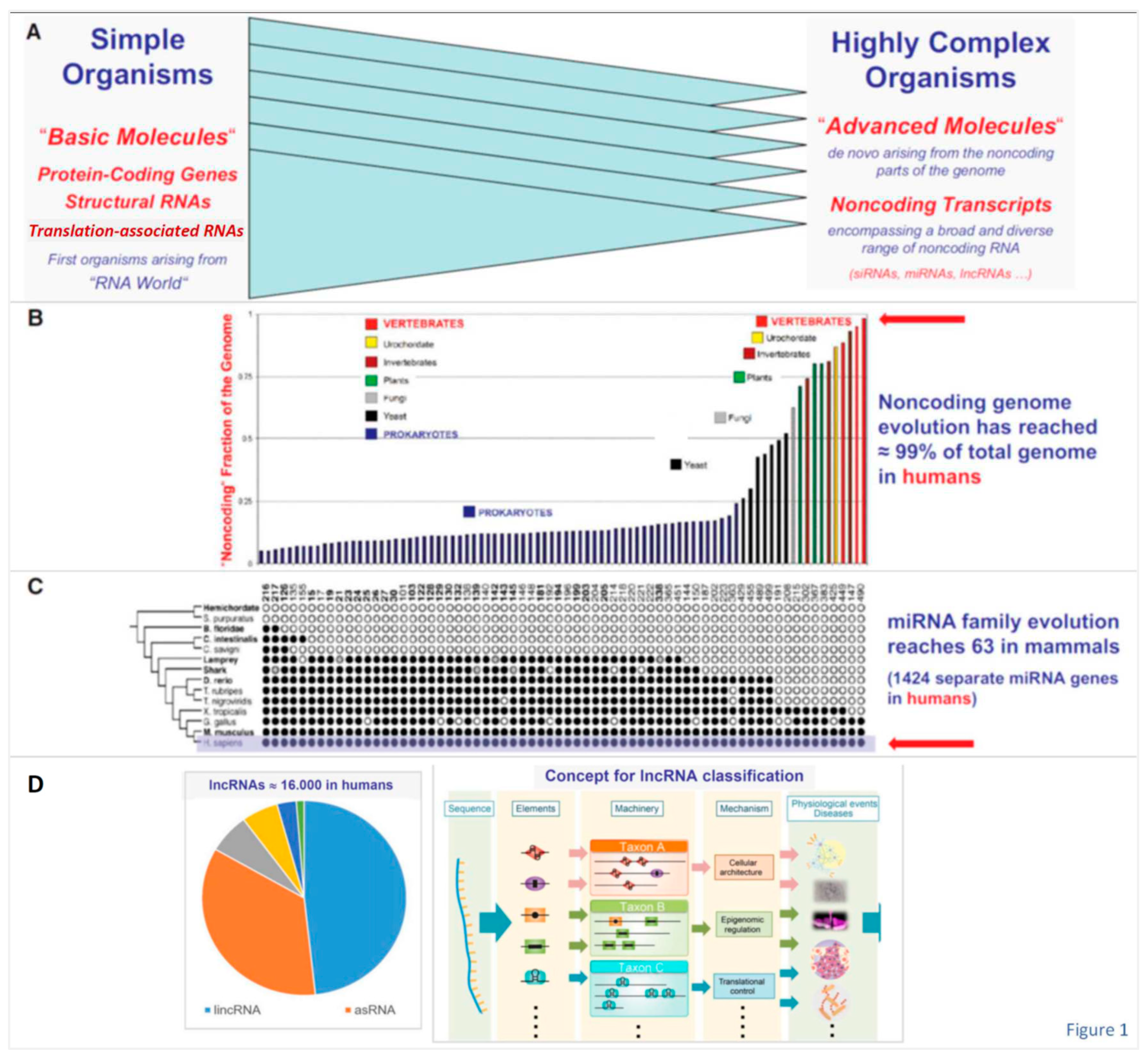
Figure 2.
– Endogenous non-coding RNAs as blueprints for RNA therapeutics. Non-coding RNAs may be addressed as therapeutic targets, but an increasing spectrum of endogenous ncRNAs (e.g., siRNAs) are also employed as blueprints for the development of novel therapeutic tools. The spectrum of possible therapeutic targets has vastly expanded beyond proteins, but likewise the therapeutic ‘toolbox’. One current topic is therapeutic RNA interference triggers (siRNAs) originally developed from endogenous siRNAs as blueprints, and made clinically applicable based on sophisticated chemical modifications and coupling to carriers/ligands for tissue targeting. Appreciation of the profound pathogenic impact of diverse small and long ncRNAs has inspired the development of multiple other therapeutic tools engaging these ncRNAs. The tools may be engineered nucleic acids themselves, acting through sequence homologies, or “classical” small molecule drugs designed to interact with e.g. conserved 3D structural motifs in lncRNAs which are not necessarily dependent on strict RNA sequence conservation.
Figure 2.
– Endogenous non-coding RNAs as blueprints for RNA therapeutics. Non-coding RNAs may be addressed as therapeutic targets, but an increasing spectrum of endogenous ncRNAs (e.g., siRNAs) are also employed as blueprints for the development of novel therapeutic tools. The spectrum of possible therapeutic targets has vastly expanded beyond proteins, but likewise the therapeutic ‘toolbox’. One current topic is therapeutic RNA interference triggers (siRNAs) originally developed from endogenous siRNAs as blueprints, and made clinically applicable based on sophisticated chemical modifications and coupling to carriers/ligands for tissue targeting. Appreciation of the profound pathogenic impact of diverse small and long ncRNAs has inspired the development of multiple other therapeutic tools engaging these ncRNAs. The tools may be engineered nucleic acids themselves, acting through sequence homologies, or “classical” small molecule drugs designed to interact with e.g. conserved 3D structural motifs in lncRNAs which are not necessarily dependent on strict RNA sequence conservation.
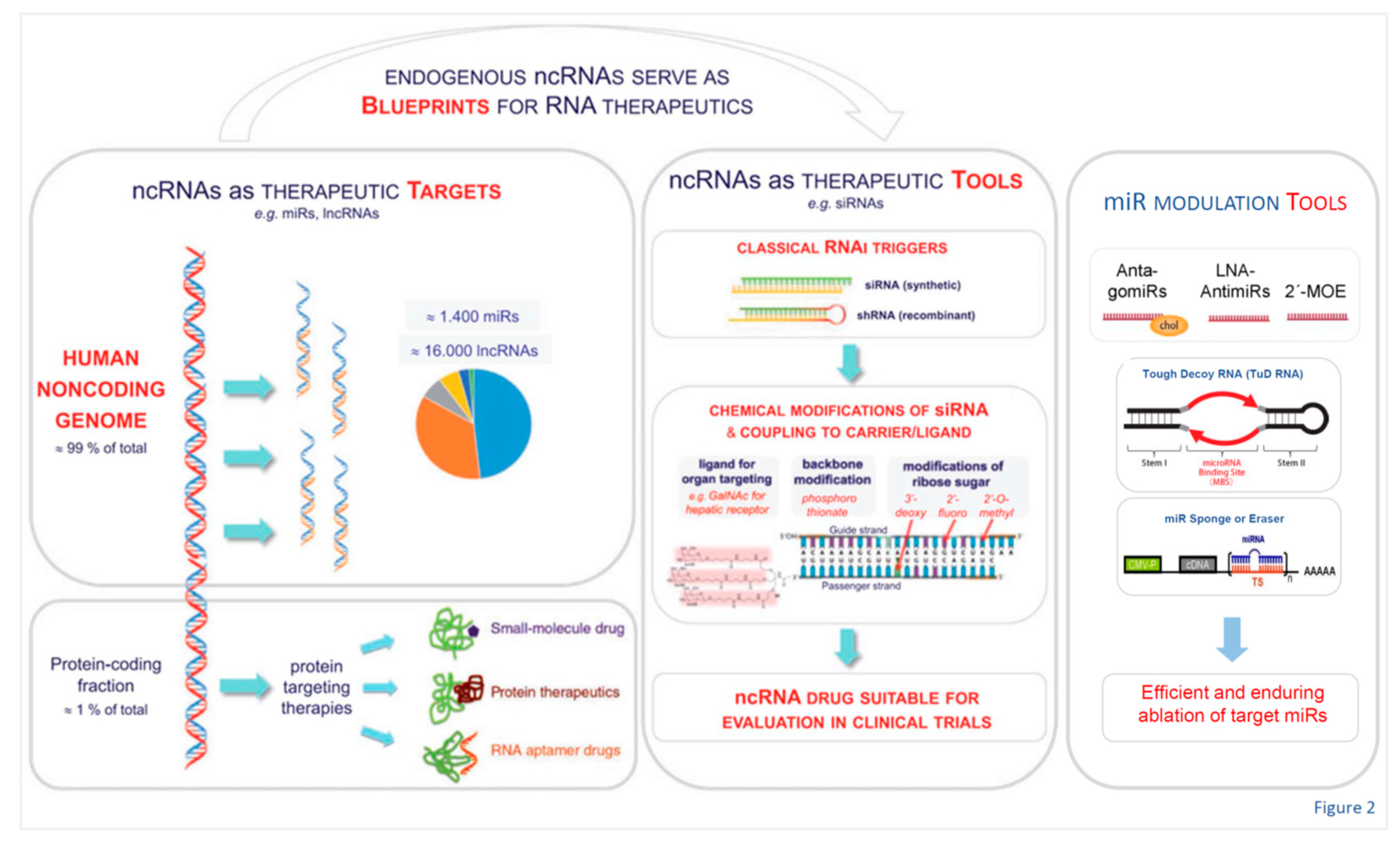
Figure 3.
– Multi-level functional integration of extended regions of the human genome – above and beyond individual noncoding RNAs. The NEAT1-MALAT1 genomic region encodes a biological integrated circuit controlling innate immune sensing and cell–cell interactions. From an evolutionary perspective, the NEAT1-MALAT1 genomic region appears as a highly integrated RNA processing circuitry critically contributing to immune homeostasis. Its components MEN-β, MEN-ε, menRNA, MALAT1, TALAM1, and mascRNA are obviously set for well-balanced interactions with each other. Genetic ablation of any element therefore leads to major dysfunction. Beyond prior work in NEAT1 and MALAT1 knockout mice, a recent cell biological study identified menRNA and mascRNA as novel components of innate immunity with deep impact upon cytokine regulation, immune cell–endothelium interactions, angiogenesis, and macrophage formation and functions. These tRNA-like transcripts appear to be prototypes of a class of ncRNAs distinct from other small transcripts (miRNAs, siRNAs) by biosynthetic pathway (enzymatic excision from lncRNAs) and intracellular kinetics, suggesting a novel link for the apparent relevance of the NEAT1-MALAT1 cluster in cardiovascular and neoplastic diseases. For the long primary transcripts of NEAT1 a function of general cell-biological interest has been identified. They are critical for the formation of paraspeckles which are involved in multiple cellular functions, and possibly also in the broader context of micellization and the formation of biomolecular condensates essential for proper subcellular and nuclear compartmentalization. Obviously, molecules involved in these fundamental processes may deeply impact upon various cellular functions in a context-dependent manner, so that their observed association with diverse diseases is therefore not entirely unexpected. Overall, the NEAT1-MALAT1 genomic region may serve as paradigm of a biological integrated circuit fine-tuning multiple cellular processes covering innate immune sensing and cell-cell interactions. (Modified from Poller et al. 2023 by permission from J. Clin. Med.).
Figure 3.
– Multi-level functional integration of extended regions of the human genome – above and beyond individual noncoding RNAs. The NEAT1-MALAT1 genomic region encodes a biological integrated circuit controlling innate immune sensing and cell–cell interactions. From an evolutionary perspective, the NEAT1-MALAT1 genomic region appears as a highly integrated RNA processing circuitry critically contributing to immune homeostasis. Its components MEN-β, MEN-ε, menRNA, MALAT1, TALAM1, and mascRNA are obviously set for well-balanced interactions with each other. Genetic ablation of any element therefore leads to major dysfunction. Beyond prior work in NEAT1 and MALAT1 knockout mice, a recent cell biological study identified menRNA and mascRNA as novel components of innate immunity with deep impact upon cytokine regulation, immune cell–endothelium interactions, angiogenesis, and macrophage formation and functions. These tRNA-like transcripts appear to be prototypes of a class of ncRNAs distinct from other small transcripts (miRNAs, siRNAs) by biosynthetic pathway (enzymatic excision from lncRNAs) and intracellular kinetics, suggesting a novel link for the apparent relevance of the NEAT1-MALAT1 cluster in cardiovascular and neoplastic diseases. For the long primary transcripts of NEAT1 a function of general cell-biological interest has been identified. They are critical for the formation of paraspeckles which are involved in multiple cellular functions, and possibly also in the broader context of micellization and the formation of biomolecular condensates essential for proper subcellular and nuclear compartmentalization. Obviously, molecules involved in these fundamental processes may deeply impact upon various cellular functions in a context-dependent manner, so that their observed association with diverse diseases is therefore not entirely unexpected. Overall, the NEAT1-MALAT1 genomic region may serve as paradigm of a biological integrated circuit fine-tuning multiple cellular processes covering innate immune sensing and cell-cell interactions. (Modified from Poller et al. 2023 by permission from J. Clin. Med.).
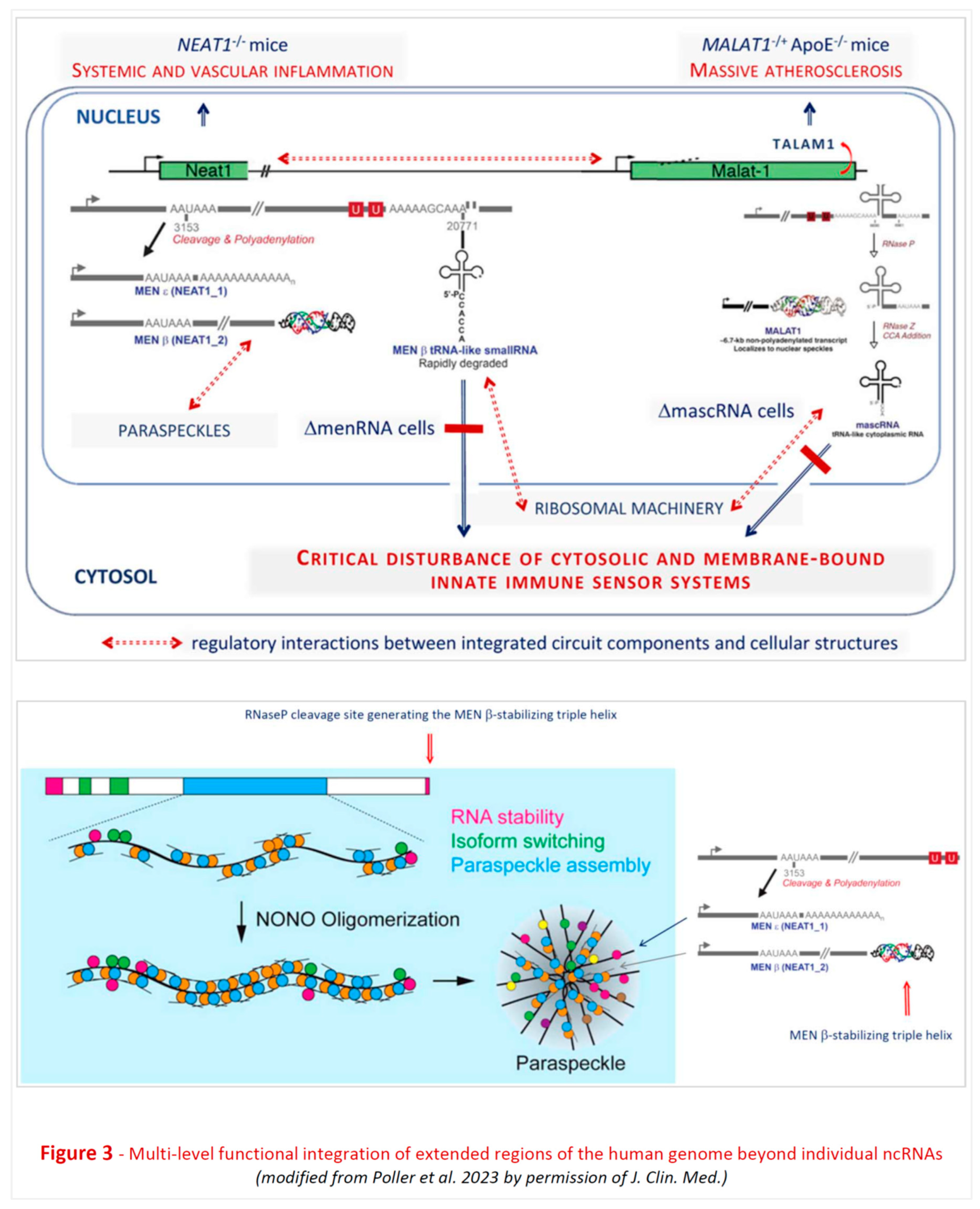
Figure 4.
–Promoter and enhancer RNAs regulate chromatin reorganization and activation of miR-10b/HOXD locus, and neoplastic transformation in glioma. miR-10b is silenced in normal neuroglial cells of the brain but commonly activated in glioma, where it assumes an essential tumor-promoting role. The entire miR-10b-hosting HOXD locus is activated in glioma via the cis-acting mechanism involving 3D chromatin reorganization and CTCF-cohesin-mediated looping. This mechanism requires two interacting lncRNAs, HOXD-AS2 and LINC01116, one associated with HOXD3/HOXD4/miR-10b promoter and another with the remote enhancer. Knockdown of either lncRNA in glioma cells alters CTCF and cohesin binding, abolishes chromatin looping, inhibits the expression of all genes within HOXD locus, and leads to glioma cell death. Conversely, in cortical astrocytes, enhancer activation is sufficient for HOXD/miR-10b locus reorganization, gene derepression, and neoplastic cell transformation. LINC01116 RNA is essential for this process. Our results demonstrate the interplay of two lncRNAs in the chromatin folding and concordant regulation of miR-10b and multiple HOXD genes normally silenced in astrocytes and triggering the neoplastic glial transformation. (Modified from Deforzh et al. 2022 by permission from Mol. Cell).
Figure 4.
–Promoter and enhancer RNAs regulate chromatin reorganization and activation of miR-10b/HOXD locus, and neoplastic transformation in glioma. miR-10b is silenced in normal neuroglial cells of the brain but commonly activated in glioma, where it assumes an essential tumor-promoting role. The entire miR-10b-hosting HOXD locus is activated in glioma via the cis-acting mechanism involving 3D chromatin reorganization and CTCF-cohesin-mediated looping. This mechanism requires two interacting lncRNAs, HOXD-AS2 and LINC01116, one associated with HOXD3/HOXD4/miR-10b promoter and another with the remote enhancer. Knockdown of either lncRNA in glioma cells alters CTCF and cohesin binding, abolishes chromatin looping, inhibits the expression of all genes within HOXD locus, and leads to glioma cell death. Conversely, in cortical astrocytes, enhancer activation is sufficient for HOXD/miR-10b locus reorganization, gene derepression, and neoplastic cell transformation. LINC01116 RNA is essential for this process. Our results demonstrate the interplay of two lncRNAs in the chromatin folding and concordant regulation of miR-10b and multiple HOXD genes normally silenced in astrocytes and triggering the neoplastic glial transformation. (Modified from Deforzh et al. 2022 by permission from Mol. Cell).
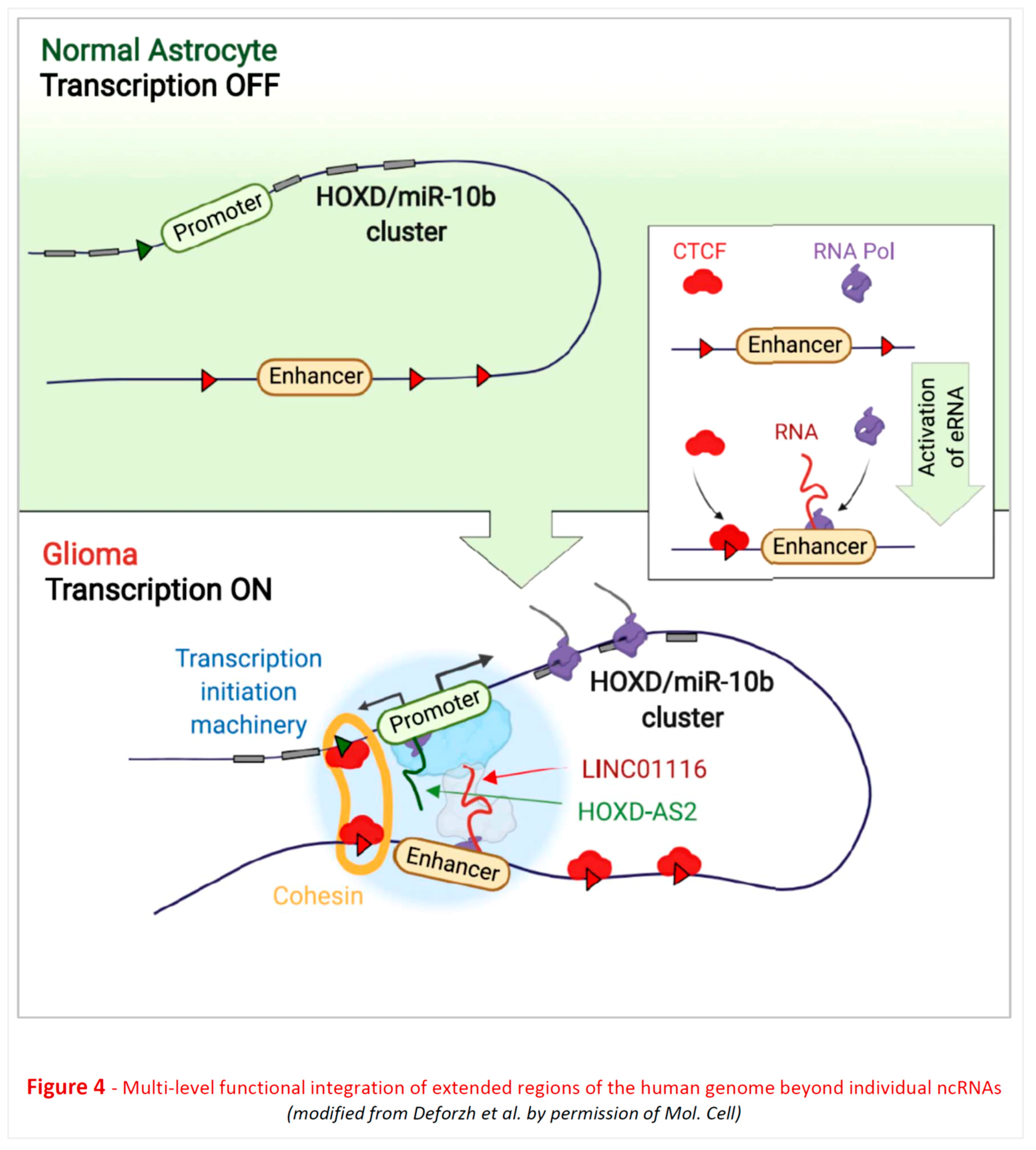
Figure 5.
– The fundamental clinical efficacy determinants of ASO and siRNA therapeutics. Despite broad diversity of the new nucleic acid-based therapeutic principles and tools, they share key common determinants of clinical efficacy which are critical for possible translational success and need to be closely monitored in any clinical trial.
Figure 5.
– The fundamental clinical efficacy determinants of ASO and siRNA therapeutics. Despite broad diversity of the new nucleic acid-based therapeutic principles and tools, they share key common determinants of clinical efficacy which are critical for possible translational success and need to be closely monitored in any clinical trial.
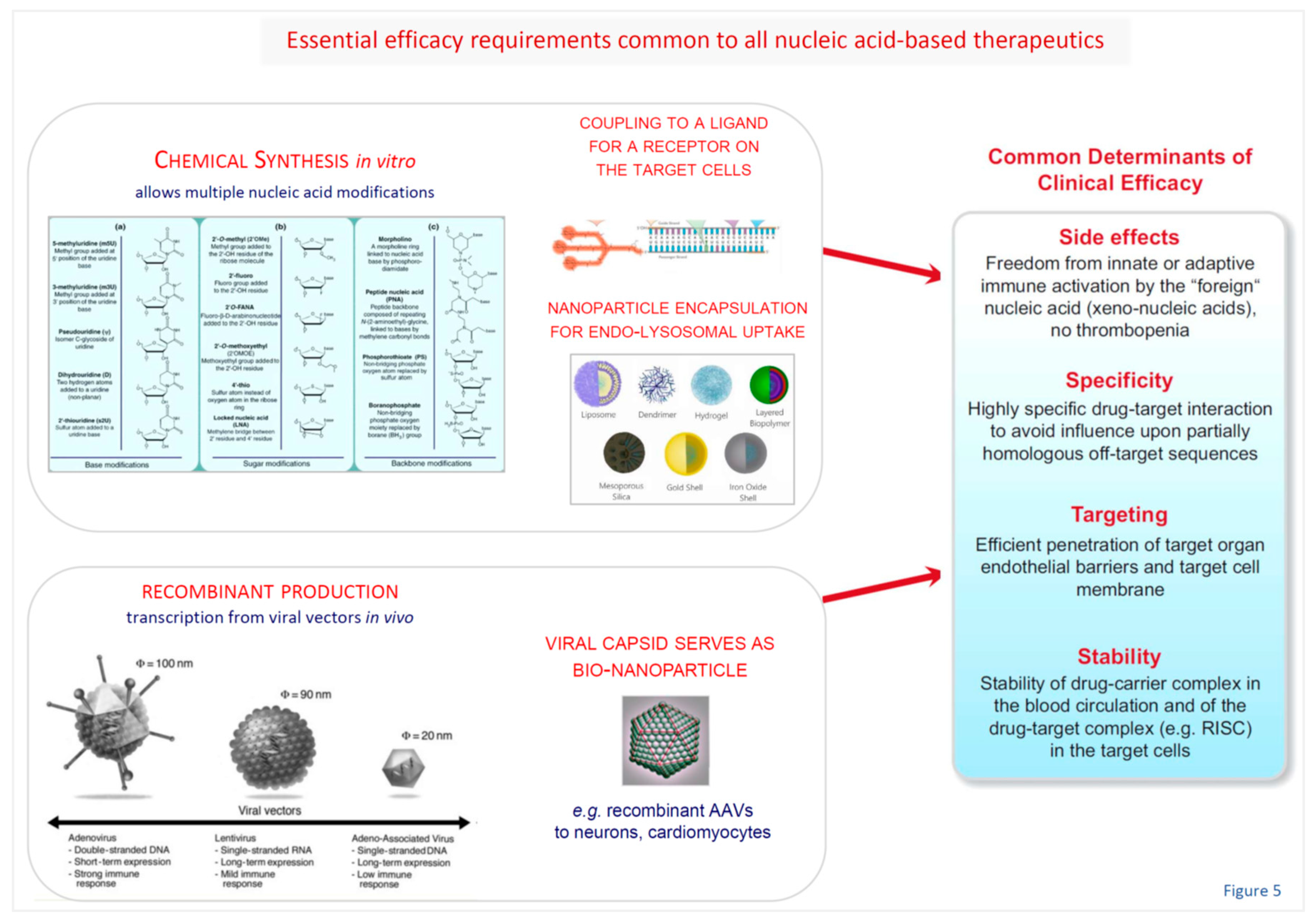
Figure 6.
– Vector-based genetic therapies for protein augmentation or RNAi-mediated target RNA depletion. The ‘classical’ approach of gene transfer for protein augmentation or (in the case of monogenic disorders) protein substitution recently gained clinical impact in the hemophilia field where the missing coagulation factor genes could be successfully and durably transferred to the liver using AAV vectors. In the cardiovascular field, cardiac-targeted gene augmentation (SERCA2a) or ablation (phospholamban) therapies were successful in animal models, but this could not yet be translated to the clinical arena due to still insufficient gene transfer efficacy in patients. The opposite approach is post-transcriptional silencing of genes involved in disease pathogenesis. Complementary to chemically synthesized base- and backbone-modified ASOs or siRNAs (Figure 6), silencing of any protein-coding or noncoding transcript may be achieved by viral vector-based RNA interference (RNAi). Two fundamentally distinct approaches (lower panel) use synthetic siRNAs, or recombinant shRNAs continuously produced from viral vectors. RNA is inherently unstable and must be modified to achieve sufficient biostability, and delivered via synthetic carriers, to become therapeutically useful. Viral vectors, which may be organ-targeted and regulatable, may circumvent targeting issues by their inherent biological properties, and the RNA stability problem by continuous synthesis in the host cells. Apart from these differences, the same characteristics will be considered when the therapeutic potential of synthetic or recombinant RNA drugs is assessed. AAV indicates adeno-associated virus; ASO, antisense oligonucleotide; LNA, locked nucleic acid; shRNA, short hairpin RNA; siRNA, short interfering RNA; and TS, target site. (Modified from Poller et al. 2013 by permission from Circ. Res.).
Figure 6.
– Vector-based genetic therapies for protein augmentation or RNAi-mediated target RNA depletion. The ‘classical’ approach of gene transfer for protein augmentation or (in the case of monogenic disorders) protein substitution recently gained clinical impact in the hemophilia field where the missing coagulation factor genes could be successfully and durably transferred to the liver using AAV vectors. In the cardiovascular field, cardiac-targeted gene augmentation (SERCA2a) or ablation (phospholamban) therapies were successful in animal models, but this could not yet be translated to the clinical arena due to still insufficient gene transfer efficacy in patients. The opposite approach is post-transcriptional silencing of genes involved in disease pathogenesis. Complementary to chemically synthesized base- and backbone-modified ASOs or siRNAs (Figure 6), silencing of any protein-coding or noncoding transcript may be achieved by viral vector-based RNA interference (RNAi). Two fundamentally distinct approaches (lower panel) use synthetic siRNAs, or recombinant shRNAs continuously produced from viral vectors. RNA is inherently unstable and must be modified to achieve sufficient biostability, and delivered via synthetic carriers, to become therapeutically useful. Viral vectors, which may be organ-targeted and regulatable, may circumvent targeting issues by their inherent biological properties, and the RNA stability problem by continuous synthesis in the host cells. Apart from these differences, the same characteristics will be considered when the therapeutic potential of synthetic or recombinant RNA drugs is assessed. AAV indicates adeno-associated virus; ASO, antisense oligonucleotide; LNA, locked nucleic acid; shRNA, short hairpin RNA; siRNA, short interfering RNA; and TS, target site. (Modified from Poller et al. 2013 by permission from Circ. Res.).
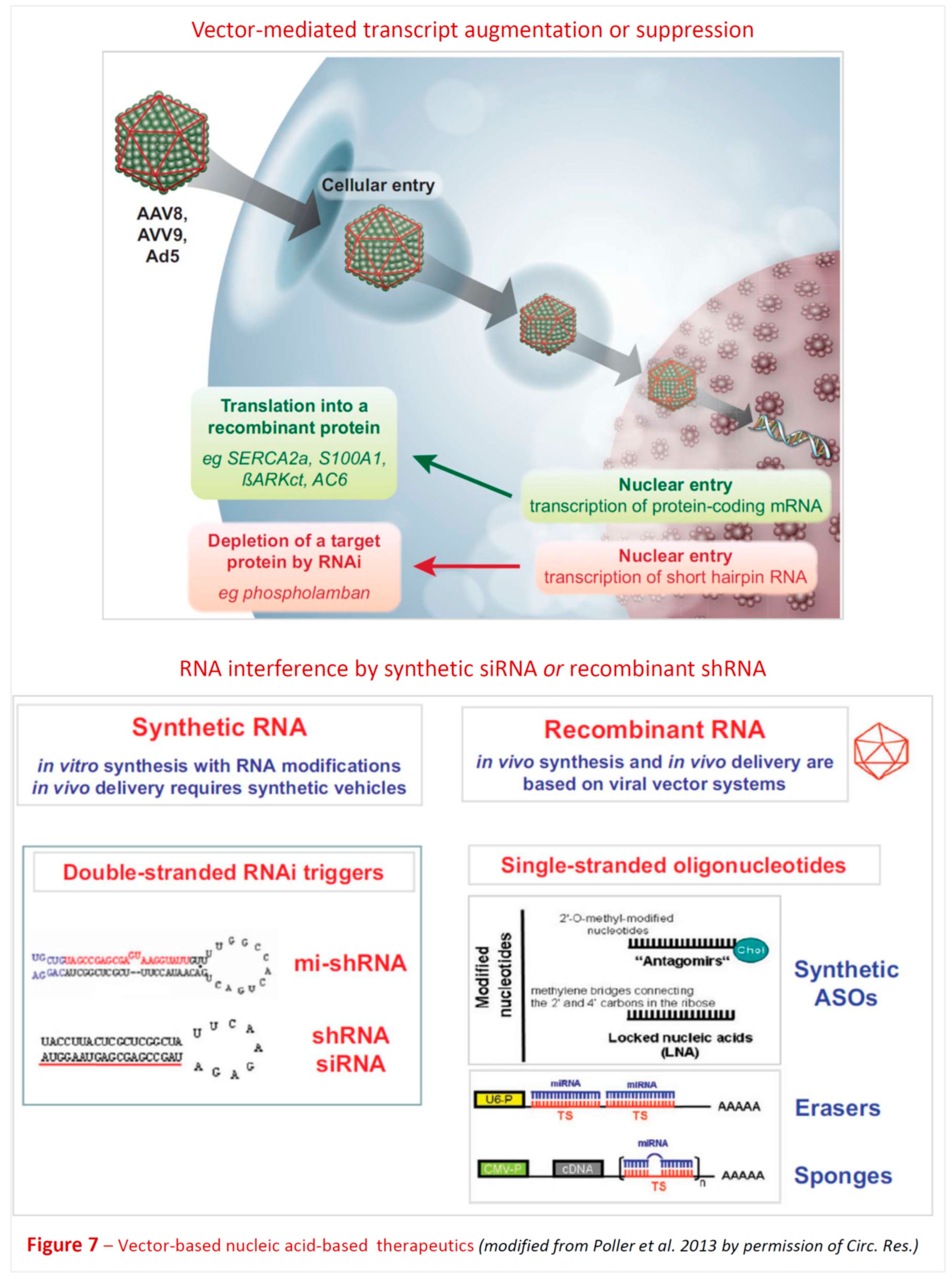
Figure 7.
– Summary and outlook. This figure aims to summarize – on the right side - the molecular and cellular basis of liver targeting of nucleic acid-based drugs, which is already being applied clinically for several disorders. Here, drug delivery is facilitated by the fenestrated endothelium of the liver, and the availability of a safe and efficient hepatocyte-specific ligand-receptor system. The left side outlines the multiple challenges arising once targeting to other organs is attempted, focusing upon brain delivery. Targeted and safe delivery of any nucleic acid-based (siRNA, ASO) drug to specific regions of the brain appears as far greater challenge than liver targeting or ex vivo blood stem cell modulation. A remarkable spectrum of brain-targeting approaches encompasses synthetic nanoparticles and viral vector, yet so far none of these are established with respect to key efficacy requirements (Figure 5). AAV vectors encounter high interest for brain-targeted therapies since genetically engineered and surface-modified (pseudotyped) versions of this vector have been extensively studied in other fields of medicine (e.g. haematology, cardiovascular medicine). The blood-brain-barrier (BBB) constitutes a particularly challenging anatomical barrier against nanoparticle or vector based drug delivery. Remarkably, the serotype AAV9 is capable to cross the BBB under certain conditions, raising the possibility of intravascular administration as a non-invasive delivery route of nucleic acid-based drugs to the CNS. Of note, this same AAV serotype is also able to enter the myocardium across the tight cardiovascular endothelium (impermeable for other AAV serotypes) and was previously employed for cardiac-targeted gene transfer and RNA interference therapy. Regarding the next step of delivery, little is known about differential tropism of currently available AAV variants for distinct brain cell types of specific therapeutic interest. Recent high-throughput screens have identified host proteins essential for AAV delivery in a comprehensive manner, and revealed unanticipated complexity and serotype specificity of the entry process. Theoretical predictability of any in vivo effects of vector modifications is therefore limited and experimental validation essential. The figure depicts recent experimental approaches to improve BBB passage and brain cell type-specific delivery. Starting from AAV9 holding promise for trans-BBB therapy, AAV-PHP.eB was engineered by insertion of a 7-amino acid peptide and point mutations of neighboring residues into the AAV9 capsid and enhanced CNS delivery in mice, however only under certain conditions. Similar challenges with regard to clinical translation, generated by species differences, have been extensively investigated before for another “hard target” i.e. the heart. AAVs deficient in HSPGs attachment, instead recognizing the glutamate receptor GluA4 through a displayed GluA4-specific DARPin, resulted in preferential >90% transduction of interneurons. Another highly innovative strategy employs membrane protein-specific nanobodies inserted into a surface loop of the VP1 capsid protein of AAVs. Nanobody-VP1 fusion was applied to AAV1, AAV2, AAV8, and AAV9 and effectively re-directed the target specificity of all these AAV serotypes. Beyond stability in the blood circulation and capability to cross the blood-brain barrier, transgene expression stability or even control is also desirable. Alphaherpesvirus latency-associated promoters (LAPs) enabled stable, pan-neuronal transgene transcription and translation from AAV-LAPs in the CNS for 6 months. Thus, these LAPs are suitable candidates for AAV-based CNS gene therapies requiring chronic transgene expression after one-time viral-vector administration.
Figure 7.
– Summary and outlook. This figure aims to summarize – on the right side - the molecular and cellular basis of liver targeting of nucleic acid-based drugs, which is already being applied clinically for several disorders. Here, drug delivery is facilitated by the fenestrated endothelium of the liver, and the availability of a safe and efficient hepatocyte-specific ligand-receptor system. The left side outlines the multiple challenges arising once targeting to other organs is attempted, focusing upon brain delivery. Targeted and safe delivery of any nucleic acid-based (siRNA, ASO) drug to specific regions of the brain appears as far greater challenge than liver targeting or ex vivo blood stem cell modulation. A remarkable spectrum of brain-targeting approaches encompasses synthetic nanoparticles and viral vector, yet so far none of these are established with respect to key efficacy requirements (Figure 5). AAV vectors encounter high interest for brain-targeted therapies since genetically engineered and surface-modified (pseudotyped) versions of this vector have been extensively studied in other fields of medicine (e.g. haematology, cardiovascular medicine). The blood-brain-barrier (BBB) constitutes a particularly challenging anatomical barrier against nanoparticle or vector based drug delivery. Remarkably, the serotype AAV9 is capable to cross the BBB under certain conditions, raising the possibility of intravascular administration as a non-invasive delivery route of nucleic acid-based drugs to the CNS. Of note, this same AAV serotype is also able to enter the myocardium across the tight cardiovascular endothelium (impermeable for other AAV serotypes) and was previously employed for cardiac-targeted gene transfer and RNA interference therapy. Regarding the next step of delivery, little is known about differential tropism of currently available AAV variants for distinct brain cell types of specific therapeutic interest. Recent high-throughput screens have identified host proteins essential for AAV delivery in a comprehensive manner, and revealed unanticipated complexity and serotype specificity of the entry process. Theoretical predictability of any in vivo effects of vector modifications is therefore limited and experimental validation essential. The figure depicts recent experimental approaches to improve BBB passage and brain cell type-specific delivery. Starting from AAV9 holding promise for trans-BBB therapy, AAV-PHP.eB was engineered by insertion of a 7-amino acid peptide and point mutations of neighboring residues into the AAV9 capsid and enhanced CNS delivery in mice, however only under certain conditions. Similar challenges with regard to clinical translation, generated by species differences, have been extensively investigated before for another “hard target” i.e. the heart. AAVs deficient in HSPGs attachment, instead recognizing the glutamate receptor GluA4 through a displayed GluA4-specific DARPin, resulted in preferential >90% transduction of interneurons. Another highly innovative strategy employs membrane protein-specific nanobodies inserted into a surface loop of the VP1 capsid protein of AAVs. Nanobody-VP1 fusion was applied to AAV1, AAV2, AAV8, and AAV9 and effectively re-directed the target specificity of all these AAV serotypes. Beyond stability in the blood circulation and capability to cross the blood-brain barrier, transgene expression stability or even control is also desirable. Alphaherpesvirus latency-associated promoters (LAPs) enabled stable, pan-neuronal transgene transcription and translation from AAV-LAPs in the CNS for 6 months. Thus, these LAPs are suitable candidates for AAV-based CNS gene therapies requiring chronic transgene expression after one-time viral-vector administration.
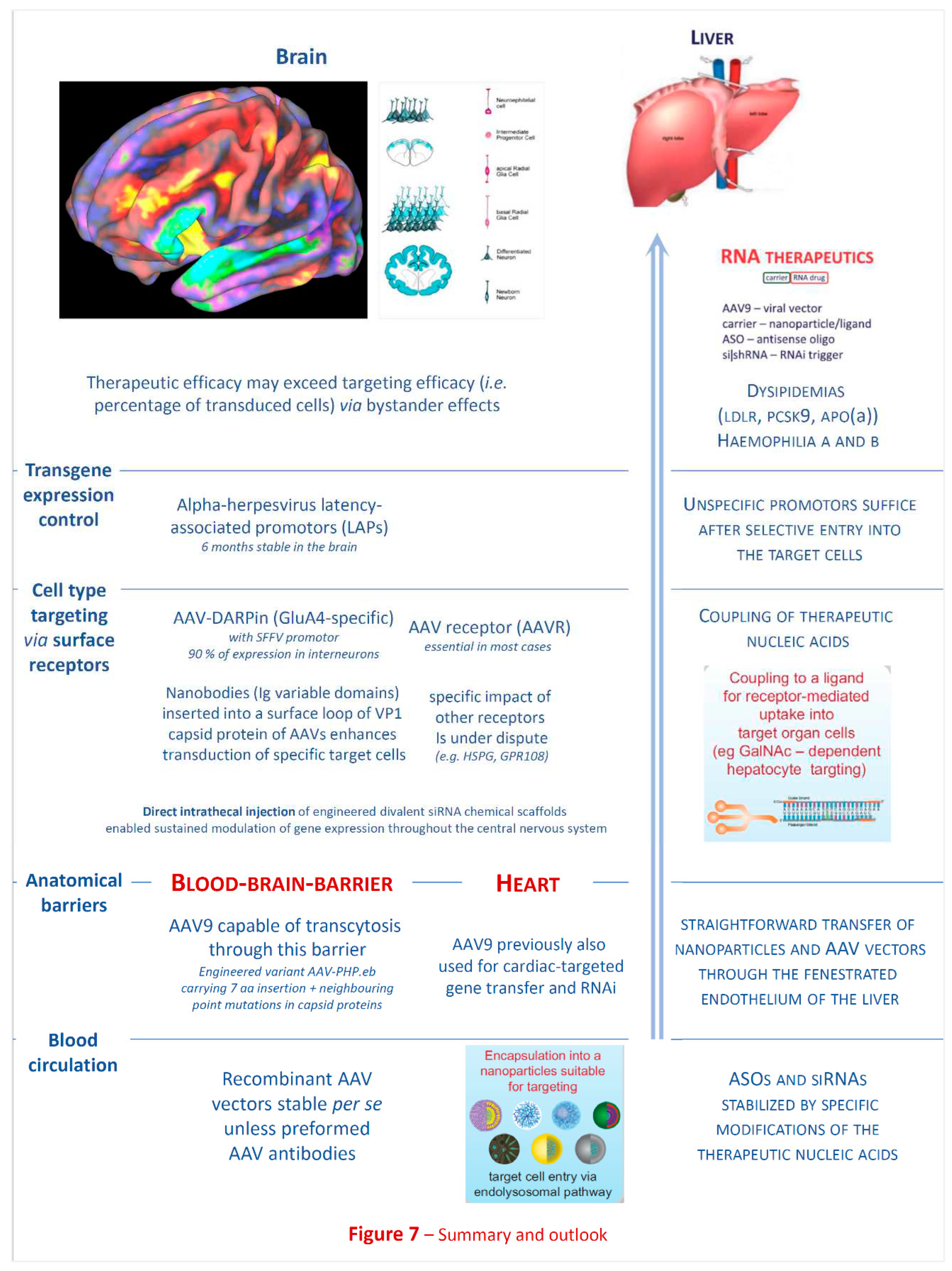
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



