
Submitted:
08 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
This review provides information on acne transcriptomics allowing deeper insights into acne pathogenesis and isotretinoin´s mode of action. Puberty-induced insulin-like growth factor 1 (IGF-1), insulin and androgen signaling activate the kinase AKT and mechanistic target of rapamycin complex 1 (mTORC1). Western diet (hyperglycemic carbohydrates, milk/dairy products) as well co-stimulate AKT/mTORC1 signaling. AKT-mediated phosphorylation of nuclear FoxO1 and FoxO3 results in their extrusion into the cytoplasm, a critical switch, which enhances the transactivation of lipogenic and proinflammatory transcription factors including androgen receptor (AR), sterol regulatory element-binding transcription factor 1 (SREBF1), peroxisome proliferator-activated receptor γ (PPARγ), and signal transducer and activator of transcription 3 (STAT3) but reduces FoxO1-dependent expression of GATA binding protein 6 (GATA6), the key transcription factor of infundibular keratinocyte homeostasis. AKT-mediated phosphorylation of the p53-binding protein MDM2 promotes the degradation of p53. In contrast, isotretinoin enhances the expression of p53, FoxO1 and FoxO3 in sebaceous glands of acne patients. Overexpression of these proapoptotic transcription factors explains isotretinoin´s desired sebum-suppressive effect via induction of sebocyte apoptosis but also its adverse effects including teratogenicity (neural crest cell apoptosis), reduced ovarian reserve (granulosa cell apoptosis), risk of depression (apoptosis of hypothalamic neurons), VLDL hyperlipidemia, intracranial hypertension and dry skin.
Keywords:
acne
; apoptosis
; FoxO1
; FoxO3
; isotretinoin
; mTORC1
; p53
; pathogenesis
; therapy
; transcriptomics
1. Introduction
In recent years, tremendous scientific progress has been made in unraveling the transcriptional regulation in acne vulgaris. This review provides updated information on acne transcriptomics allowing deeper insights into the pathogenesis and treatment of acne. Acne vulgaris is a very common chronic inflammatory skin disorder with a complex pathogenesis. Four factors play vital roles in acne pathophysiology: hyperseborrhea and dysseborrhea, altered keratinization of the pilosebaceous duct (comedogenesis), effects mediated by Cutibacterium acnes (C. acnes) and Th17 cell-driven inflammation [1,2].
Isotretinoin is the most efficient anti-acne drug improving all major factors of acne pathogenesis. This review focuses on three key transcription factors, p53, FoxO1 and FoxO3, whose suppression plays important roles in acne pathogenesis, whereas their upregulation by isotretinoin explains isotretinoin´s beneficial as well as its adverse effects in the treatment of acne.
2. Growth Factor Signaling in Acne Activating the Kinase AKT
Various growth factors induce acne pathogenesis. Their input signals converge in the activation of the kinase AKT (protein kinase B), which modifies the activity and expression of important transcription factors promoting the disease.
2.1. Insulin-Like Growth Factor 1
Insulin-like growth factor 1 (IGF-1) is a potent mitogen [3]. IGFs bind specifically to the IGF-1 receptor (IGF1R) on the cell surface of targeted tissues [4]. During puberty increasing production of growth hormone (GH) leads to the activation of the GH/IGF-1 axis [5]. IGF-1 is the key hormone of puberty [6] and promotes sexual differentiation and linear bone growth [7]. Acne becomes manifest at a relatively late stage of puberty, at or shortly after peak height velocity in boys and menarche in girls [8]. Deplewski and Rosenfield [9] emphasized the important role of IGF-1 in the development of the pilosebaceous unit. Increased serum IGF-1 levels correlate with acne lesion counts in females [10] and increased sebum secretion rates in male acne patients [11]. IGF-1 is overexpressed in epidermis and pilosebaceous units of acne patients compared to controls [12]. Western diet, especially hyperglycemic carbohydrates [13], milk and yogurt consumption [14,15] have been associated with IGF-1-mediated acne pathogenesis [16,17]. Individuals with Laron syndrome, who exhibit a congenital IGF-1 deficiency, are of short stature and do never develop acne [18], unless therapeutically substituted with recombinant IGF-1 [19].
2.2. IGF-1-PI3K-AKT-Mediated Downregulation of FoxO1 and FoxO3
After binding of IGF-1 to its receptor, the phosphoinositide 3-kinase (PI3K) gets activated resulting in the activation of the kinase AKT (also known as protein kinase B), a key checkpoint of growth network regulation [20]. Among many regulatory effectors, AKT phosphorylates and thereby inactivates the transcription factors FoxO1 and FoxO3, predicted to play a key role in acne pathogenesis [21]. After AKT-mediated phosphorylation, nuclear FoxO1 is extruded into the cytoplasm, confirmed in IGF-1-stimulated SZ95 sebocytes in vitro [22]. FoxO1 acts as a negative nuclear coregulator of lipogenic transcription factors including androgen receptor (AR) [23], sterol regulatory element-binding transcription factor 1 (SREBF1) [24], peroxisome proliferator-activated receptor-γ (PPARγ) [25] and of the pro-inflammatory signal transducer and activator of transcription 3 (STAT3), respectively [26].
FoxO1 binds to STAT3 and prevents STAT3 from interacting with the SP1.POMC promoter complex, and consequently, inhibits STAT3-mediated leptin action. As shown in SZ95 sebocytes, leptin promotes a proinflammatory lipid profile and induces inflammatory pathways in human SZ95 sebocytes [27].
Thus, increased IGF-1 signaling via AKT-mediated suppression of FoxO1 enhances the transcriptional activity of AR, SREBF1, PPARγ and STAT3, crucial transcription factors promoting sebaceous lipogenesis [28,29,30]. In fact, IGF-1 has been shown to induce SREBF1 expression and lipogenesis in SEB-1 sebocytes via activation of the PI3K/AKT pathway [30], whereas a low glycemic diet, which decreases IGF-1 serum levels [12], reduces SREBF1 expression in sebaceous glands (SGs) of acne patients [31] (Figure 1).
2.3. IGF-1-IGF1R-PI3K-AKT-MDM2-Mediated Downregulation of P53
IGF-1/IGF1R/PI3K/AKT activation inhibits the activity of p53 [32]. AKT activated by exogenous IGF-1 promotes the phosphorylation of the p53-binding protein mouse double minute 2 (MDM2). This phosphorylation increases the ability of MDM2 to degrade p53 [32]. MDM2 promotes cell survival and cell cycle progression by inhibiting p53. To regulate p53, MDM2 must gain nuclear entry, thereby diminishes cellular levels of p53, and decreases p53 transcriptional activity [32]. Phosphorylated MDM2 finally promotes the proteasomal degradation of p53 [33,34]. Thus, IGF-1-AKT signaling lowers the level of nuclear p53, recently suggested to play a key role in acne pathogenesis and treatment [2,35] (Figure 1).
2.4. IGF-1-IGF1R-PI3K-AKT-Mediated Activation of mTORC1
Mechanistic target of rapamycin complex 1 (mTORC1) is a key growth factor-and nutrient-dependent regulatory kinase orchestrating cell proliferation and anabolism and inhibiting autophagy [36,37]. Activated AKT phosphorylates tuberin (TSC2). TSC2 is inactivated by AKT-dependent phosphorylation, which destabilizes TSC2 and disrupts its interaction with hamartin (TSC1) [38]. The TSC1-TSC2 (hamartin-tuberin) complex, through its GAP (GTPase-activating protein) activity towards the small G-protein RHEB (Ras homolog protein enriched in brain) is a critical negative regulator of mTORC1 [39]. Thus, AKT-mediated phosphorylation of TSC2 plays a key role for growth factor-stimulated mTORC1 activation, predicted to be of fundamental importance in acne pathogenesis [40,41]. Indeed, increased expression of mTOR and mTORC1 activation has been detected in the epidermis and SGs of acne patients [42,43,44]. Western diet over-stimulates insulin/IGF-1/AKT/mTORC1 signaling [45]. Recent evidence indicates that milk is a highly specialized signaling system of mammals promoting mTORC1-dependent translation for postnatal growth [46]. mTORC1 has a special impact in regulating protein translation of specific transcription factors involved in the pathogenesis of acne including SREBFs, PPARγ, STAT3 and hypoxia-inducible factor-1α (HIF-1α) [47]. Further evidence obtained in adipocytes underlines the role of mTORC1 in leptin biosynthesis at the level of translation [48] (Figure 1).
2.5. Insulin-INSR-PI3K-AKT-Mediated Activation of mTORC1
Acne has been associated with syndromes associated with insulin resistance, especially polycystic ovary syndrome (PCOS) [49], which results in hyperinsulinemia [50]. Insulin and IGF-1 are sister growth hormones and insulin has been shown to bind and activate both, insulin receptor (INSR) and IGF1R. Both INSR and IGF1R also elicit common downstream signaling with phosphorylation of a family of INSR substrates and activation of the PI3K-AKT-mTORC1 pathway [51]. Milk exhibits a high insulinemic index [52]. High intakes of milk, but not meat, increase serum insulin and insulin resistance as shown in 8-year-old boys [53]. Thus, milk consumption stimulates insulin and IGF-1 signaling [45], a meaningful mechanism to stimulate mTORC1-driven postnatal growth and anabolism [46]. In contrast, metformin therapy lowers insulin resistance [54] and thus reduces insulin signaling towards SGs. Mirdamadi et al. [22] convincingly demonstrated that both insulin and IGF-1 enhance the PI3K/AKT pathway in SZ95 sebocytes in vitro.
2.6. FGFR2-PI3K-AKT-Mediated Activation of mTORC1
Patients with congenital fibroblast growth factor receptor 2 (FGFR2) gain-of-function mutations (S252W or P253R FGFR2) - either germline (Apert syndrome) [55,56,57,58] or segmental somatic (acne nevus of Munro) [59,60,61] develop acne during puberty and respond to isotretinoin treatment [62,63,64]. FGFR2 is a tyrosine kinase receptor that like INSR or IGF1R activates multiple pathways including PI3K/AKT signaling [55]. Gain-of-function FGFR2 signaling thus adds on IGF1R and INSR signaling of puberty enhancing the AKT-mTORC1 pathway featuring the acne in Apert syndrome and acne nevus of Munro (Figure 1).
2.7. Androgen Receptor Signaling Converges with PI3K-Mediated Activation of AKT
Decades ago, acne was believed to be primarily an androgen-driven disease. Whereas the impact of androgens on acne pathogenesis is well accepted [2,28,65], recent observations underline a complex synergistic crosstalk between IGF-1, insulin and androgens in the activation of AKT and mTORC1. Androgens operate via ligand binding to AR. IGF-1 plays a critical role in the synthesis of adrenal and gonadal androgens [2,9,66,67,68,69,70,71] and promotes 5-reductase-mediated conversion of testosterone to dihydro-testosterone (DHT) [72], the high affinity ligand of AR [73,74]. Androgens induce sebaceous differentiation in sebocytes expressing a stable functional AR [75]. Notably, two androgen response elements (AREs) have been identified in the upstream promoter of the IGF1 gene [76]. It has been shown in prostate cancer cells that androgens also upregulate IGF1R expression [77]. DHT increases the association of mTOR with RICTOR instead of RAPTOR consistent with selective activation of mechanistic target of rapamycin complex 2 (mTORC2) by DHT [78]. Like PI3K, the growth promoting kinase mTORC2 phosphorylates and activates AKT and thereby contributes to nuclear extrusion of FoxO1 [78]. Thus, the insulin/IGF-1-PI3K signaling pathway and the androgen-mTORC2 signaling pathway converge in the activation of AKT (Figure 1). AKT-mediated nuclear extrusion of FoxO1, which is a negative coregulator of AR, further enhances the transcriptional activity of AR [23]. In addition, AKT phosphorylates AR enhancing AR transactivation and AR nuclear stability [79]. AKT-MDM2-mediated suppression of p53 [32,33,34] will further enhance AR signaling because AR expression is negatively regulated by p53 [80]. Notably, androgen-insensitive subjects who lack functional ARs do not produce sebum and do not develop acne [81] underlining the important contribution of AR signaling in the pathogenesis of acne.
3. Hypoxia-Inducible Factor-1α and Leptin
In 2014, Danby [82] postulated that ductal hypoxia may link comedogenesis and inflammation in acne. It has been hypothesized that ductal hypoxia as well as mTORC1 activation via upregulation of HIF-1 may enhance sebocyte-derived leptin secretion into the ascending sebum that may subsequently stimulate the proliferation of adjacent infundibular keratinocytes promoting comedogenesis [83]. The key transcription factor of hypoxia is HIF-1 [84]. HIF-1 translation is upregulated by activated mTORC1 [47].
Remarkably, HIF-1 transactivates the human leptin (LEP) gene promoter [85]. It has been shown in human keratinocytes that leptin mediates mitogenic stimuli promoting keratinocyte proliferation [86]. Primary keratinocytes under hypoxic conditions induce filaggrin (FLG) gene expression in a HIF-1α- and HIF-2α-dependent manner [87]. In fact, increased filaggrin expression has been observed in the sebaceous duct and infundibulum of patients with acne vulgaris [88]. The upregulation of the leptin-induced proinflammatory cytokines in normal human keratinocytes is mainly regulated by STAT3 signaling [89], which is also stimulated by overactivated mTORC1 [47].
HIF-1α acts via the retinoic acid receptor-related orphan receptor-γt (RORγt) to drive Th17 cell differentiation and is thus a key reprogrammer of metabolism in inflammatory cells promoting inflammatory gene expression [90]. HIF-1α controls the balance of Th17 cells and regulatory T cells [91]. Under conditions of hypoxia, most eukaryotic cells can shift their primary metabolic strategy from predominantly mitochondrial respiration towards increased glycolysis to maintain ATP levels [92]. At the transcriptional level, this metabolic switch is critically dependent on HIF-1α, which induces the expression of glycolytic enzymes. Glycolysis generates indispensable metabolic intermediates [93] that are required for the rapid proliferation of keratinocytes [94,95], sebocytes [96] and Th17 cells [97], thus linking comedogenesis and Th17 cell-mediated inflammation in acne [82,98]. A pilot study indicated that acne lesion extraction reduces HIF-1α expression [99]. Choi et al. [100] recently reported increased levels of SREBF1 and perilipin 2 (PLIN2) that were upregulated by HIF-1α in SZ95 sebocytes under hypoxia indicating that a hypoxic microenvironment can increase lipogenesis and provides a link between seborrhea and inflammation. However, we have to keep in mind that not only local hypoxia, but also increased mTORC1 signaling generally enhances the expression of HIF-1α [47] and leptin [48], respectively.
Surprisingly, there are no data on the effect of benzoyl peroxide, the most commonly used topical anti-acne agent, on ductal HIF-1α expression in acne skin. Li et al. [101] reported that minocycline, a commonly used oral antibiotic for the treatment of inflammatory acne [102,103], induces HIF-1α proteasomal degradation under hypoxia by increasing the expression prolyl hydroxylase-2 and HIF-1α/von Hippel-Lindau protein interaction, thereby overcoming hypoxia-induced HIF-1α stabilization. Azelaic acid, another topically used anti-acne agent, might also interfere with HIF-1α because it is known to inhibit anaerobic glycolysis [104]. An intensive crosstalk between p53 and HIF-1α as mediators of molecular responses to physiological and genotoxic stresses has been reported [105]. Activated p53 decreases HIF-1α protein levels by accelerated proteasome-dependent degradation [106]. In contrast, AKT activation inhibits p53-mediated HIF-1α degradation [107]. Remarkably, PI3K-AKT activation is required for the hypoxic stabilization of HIF-1α and hypoxia alone is not sufficient to render HIF-1α resistant to proteasomal cleavage and degradation [108]. Of note, we recently detected increased expression of HIF-1α in lesional skin of patients with acne inversa (hidradenitis suppurativa) [109].
4. Infundibular GATA-Binding Protein 6
Recent studies reported that the infundibulum and sebaceous ducts are lined by molecularly distinct differentiated cells expressing markers including keratin 79 and the transcription factor GATA-binding protein 6 (GATA6) [110,111,112]. Loss of GATA6 causes dilation of the hair follicle canal and sebaceous duct [111]. Importantly, Oulès et al. [112] observed reduced expression of GATA6 in the upper pilosebaceous unit in acne patients. GATA6 controls keratinocyte proliferation and differentiation to prevent hyper-keratinization of the infundibulum, which is the primary pathological event in acne vulgaris (comedogenesis). Okabe et al. [113] found the presence of retinoic acid response elements (RAREs) in the putative regulatory region of the GATA6 gene and identified retinoic acid as a critical signal inducing GATA6 transcription. Topical treatment of acne with all-trans retinoic acid (ATRA) may thus normalize disturbed follicular keratinization promoting comedolysis via the induction of GATA6 expression. Importantly, the expression of GATA6 is also upregulated by FoxO1 [112]. GATA6 exhibits three putative FoxO1 binding sites. Inhibition of AKT reduces phosphorylated FoxO1 and increases nuclear localization of FoxO1 subsequently enhancing GATA6 expression [114]. In contrast, overexpression of IGF-1/AKT signaling may thus reduce infundibular GATA6 expression promoting comedogenesis via suppression of nuclear FoxO1 and GATA6. mTORC2 signaling, the androgen-driven pathway to activate AKT, also negatively regulates GATA6 expression in a FoxO1-dependent manner [114]. Thus, increased insulin/IGF-1-AKT signaling as well as androgen-mTORC2-AKT signaling converge in the suppression of GATA6. Intriguingly, overexpression of GATA6 suppresses AR expression [110], pointing to enhanced infundibular AR signaling in reduced states of GATA6. Notably, inhibition of mTORC1 stabilizes GATA6 and promotes the nuclear accumulation of GATA6 [115]. In accordance with Oulès et al. [112], GATA6 expression contributes to the therapeutic effect of ATRA, the main topical treatment for comedonal acne. Isotretinoin/ATRA induced overexpression of p53 and FoxO1 with associated attenuation of PI3K-AKT-mTORC1 signaling may stabilize infundibular GATA6 in acne, a potential key mechanism of comedolysis. Isotretinoin treatment of human SEB-1 sebocytes induces cell cycle arrest associated with increased expression of p21 (cyclin-dependent kinase inhibitor 1A; CDKN1A) [116]. The CDKN1A gene is transcriptionally activated by p53 [117]. Remarkably, transient transfection with a GATA6 expression vector inhibited S-phase entry in vascular smooth muscle cells and in mouse embryonic fibroblasts lacking both p53 alleles. The GATA6-induced growth arrest correlated with a marked increase in the expression of p21 [118]. Thus, there appear to be a close interactions between isotretinoin-induced upregulation of p53-FoxO1, p53-p21 and FoxO1-GATA6, respectively. In contrast, hypoxia-induced microRNA-181b has been shown to target GATA6 [119], thus eventually destabilizing GATA6-controlled infundibular keratinocyte homeostasis linking ductal hypoxia to ductal GATA6 deficiency in acne.
5. Transforming Growth Factor β
Genome-wide associations studies identified gene loci associated with impaired transforming growth factor β (TGFβ) signaling including the genes TGFB2, OVOL1 and FST [120,121]. In fact, experimental evidence supports the view that TGFβ signaling is reduced in SGs of acne patients [112]. Activation of the TGFβ signaling pathway is necessary and sufficient for maintaining sebocytes in an undifferentiated state. The presence of TGFβ suppresses genes required for the production of sebaceous lipids and for sebocyte differentiation such as fatty acid desaturase 2 (FADS2) and PPARγ, thereby decreasing lipid accumulation through a TGFβR2-SMAD2 dependent pathway [122]. There is an important molecular crosstalk between GATA6 and TGFβ. GATA6 is a critical signal for the activation of TGFβ [113]. In a sebaceous organoid model, Oulès et al. [112] obtained evidence that GATA6-mediated TGFβ activation is a key process controlling the repression of the interfollicular fate of keratinocytes and promoting junctional zone/sebaceous duct differentiation and downregulation of the infundibular differentiation program [112]. Furthermore, it has been shown in chondrocytes that TGFβ stimulates FoxO1 expression [123] pointing to a crosstalk of GATA6, TGFβ and FoxO1, respectively.
6. Cutibacterium acnes
Obstruction of the pilosebaceous unit leads to hypoxia, favoring the development of Cutibacterium acnes (C. acnes), formerly designated as Propionibacterium acnes (P. acnes). Multiple studies suggest that dysbiosis of the skin microbiota significantly contributes to acne development. C. acnes is the dominant resident bacterial species in SG-enriched areas of the skin implicated in acne pathogenesis and disease progression [124]. Acne has been linked to dysregulated innate immunity in response to C. acnes promoting increased Th17 cell differentiation with increased IL-17 levels [125], thus contributing to sebocyte-mediated polarization of cutaneous T cells towards the Th17 phenotype [126]. To mimic the effect of C. acnes on sebocytes, Oulès et al. [112] treated control or GATA6-expressing sebocytes with peptidoglycan (PGN), which is the main component of the Gram+ bacterial cell wall, which is recognized by toll-like receptor 2 (TLR2). GATA6 expression led to a decrease in PGN-induced IL-17 expression. When they explored the effect of live C. acnes bacteria on control and GATA6 expressing sebocytes, GATA6 expression showed a trend towards deceased IL-17 expression [112]. It has been reported that C. acnes membrane fractions increase IGF-1 and IGF1R expression in the epidermis of explants [127], a constellation that may reduce FoxO1-mediated GATA6 expression. C. acnes also induces TLR2 and TLR4 on keratinocytes, a mechanism that could play an essential role in acne-linked inflammation [128], as TLR2 activation drives an inflammatory transcriptional program in Th17 cells [129]. Biofilm C. acnes in contrast to planktonic bacteria are characterized by upregulated stress-induced genes and upregulation of genes coding for the potential virulence-associated CAMP factors [130] including exogeneous triacylglycerol lipase (gehA) [131] enhancing the release of free fatty acids like palmitic and oleic acid [132,133]. Lipase-mediated triacylglycerol hydrolysis thus generates saturated free fatty acids that bind and activate TLR2-triggered proinflammatory signaling [134]. Gene expression of interleukin 8 (IL-8) and TLR2 was enhanced by cell-free extracts of C. acnes in SZ95 sebocytes [135]. Palmitic acid can stimulate IL-6, TNF-α, IL-1β productions in HaCaT keratinocytes promoting cell proliferation, potentially contributing to inflammation and pilosebaceous duct hyperkeratinization in acne [136].
Taken together, C. acnes-derived signaling augments a variety of transcriptomic deviations observed in acne vulgaris.
7. Transcriptomic Effects of Isotretinoin Treatment
Isotretinoin counteracts critical pathways of growth factor and androgen signaling that are overstimulated in acne vulgaris.
7.1. P53/FoxO1-Mediated Suppression of IGF-1/IGF1R/PI3K/AKT/mTORC1 Signaling
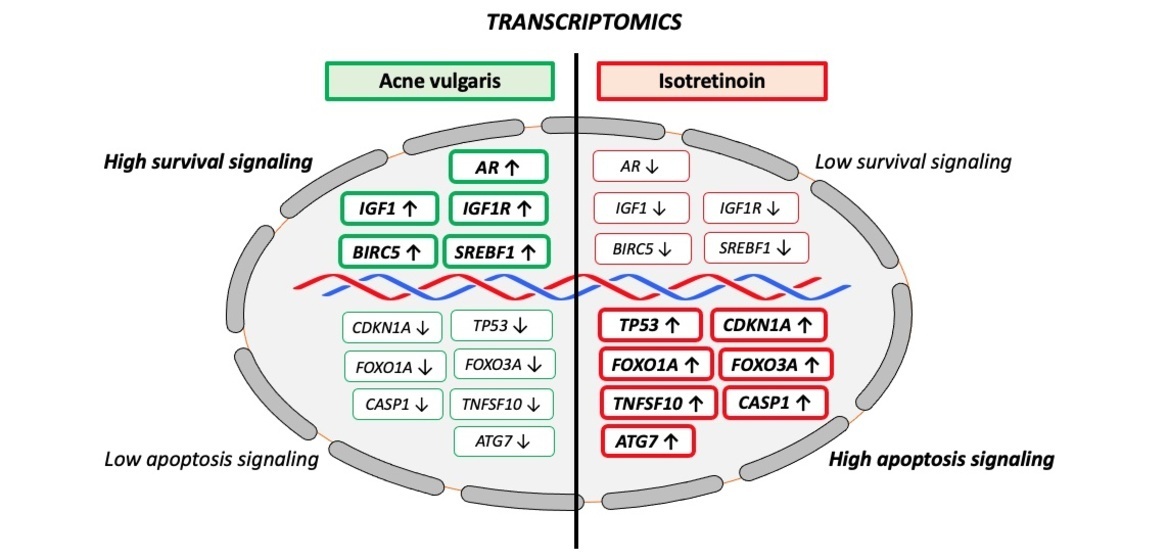
Oral isotretinoin is the most effective treatment of recalcitrant and severe acne improving all major aspects of acne pathogenesis [137,138]. It has been predicted that isotretinoin upregulates the expression of FoxO1 and FoxO3 [139,140,141]. In the meantime, experimental evidence confirmed that isotretinoin enhances the expression of FoxO1 in primary human keratinocytes [142], SZ95 immortalized human sebocytes [143] and SGs of isotretinoin-treated acne patients [144]. We could demonstrate by immunohistochemistry of SGs that isotretinoin treatment increased nuclear accumulation of FoxO1 and FoxO3 proteins [144] pointing to changes of FoxO-mediated gene expression or FoxO translocation. Furthermore, it has been hypothesized that isotretinoin upregulates p53 in the pilosebaceous unit [35]. Notably, p53 maintains baseline expression of FOXO1A [145]. Importantly, FOXO3A has also been confirmed to be target gene of p53 [146]. Experimental evidence confirmed that isotretinoin treatment of primary human keratinocytes increased the expression of p53 [142]. Recently, we could demonstrate that isotretinoin treatment enhanced nuclear accumulation of p53 in SGs of acne patients [147]. Of note, prior to isotretinoin treatment, acne patients exhibited lower cutaneous expression levels of p53 protein compared to acne-free controls [147].
p53, FoxO1 and FoxO3 are fundamental negative regulators of cell cycle progression and cell proliferation. These proapoptotic transcription factors exert synergistic effects in promoting catabolism, autophagy and apoptosis, respectively [148,149,150,151,152,153,154] (Figure 2).
p53 suppresses IGF-1/PI3K/AKT/mTORC1 activity at multiple regulatory checkpoints [155,156,157,158,159,160,161,162,163,164]. The IGF1 promoter is negatively regulated by p53 [159]. In fact, Karadag et al. [160] observed decreased serum levels of IGF-1 in isotretinoin-treated acne patients. In addition, IGF1R expression is also negatively regulated by p53 [161]. Disruption of endogenous IGF1R led to inhibition of insulin receptor (INSR) promoter activity by p53 [162]. Of note, the expression of negative regulators of the PI3K-AKT-mTORC1 signaling pathway including phosphatase and tensin homolog (PTEN) (a negative regulator of PI3K), TSC2 (negative regulator of RHEB), AMP-activated protein kinase β1 (PRKAB1) (activator of TSC2) are all upregulated by p53 [163]. DEP-domain containing mTOR-interacting protein (DEPTOR) is a natural inhibitor of mTORC1 and mTORC2. DEPTOR as well is a downstream target p53, whose activity positively correlates with DEPTOR expression [165].
Taken together, isotretinoin-mediated upregulation of p53 counteracts multiple regulatory checkpoints of exaggerated IGF-1/IGF1R/PI3K/AKT-mTORC1 and potentially mTORC2 signaling pathways in acne patients (Figure 3).
7.2. P53/FoxO1 Upregulation Suppresses AR Signaling
7.3. GATA6 Upregulation Suppresses Comedogenesis
Reduced infundibular GATA6 expression has been related to disturbed follicular keratinization (comedo formation) in acne patients [112]. Systemic isotretinoin treatment has a two principal impacts on the upregulation of infundibular GATA6 expression: 1) After isomerization of isotretinoin (13-cis retinoic acid) to ATRA [170], ATRA activates RAR [171] enhancing GATA6 expression via RAR binding to RARE on the GATA6 promoter [113]. In addition, p53-mediated upregulation of nuclear FoxO1 may activate three FoxO1 binding sites on the GATA6 promoter [112,114] further promoting its expression. Upregulated GATA6 represses AR activation [110], a synergistic interplay normalizing disturbed infundibular keratinization (comedogenesis) in acne patients.
7.4. Sebum Suppression, Sebocyte Autophagy and Apoptosis
It is well accepted that binding of ATRA initiates changes in interactions of RAR/RXRs with co-repressor and co-activator proteins, activating transcription of primary target genes, alters interactions with proteins that induce epigenetic changes and induces transcription of genes encoding transcription factors and signaling proteins that further modify gene expression [172]. Isotretinoin-induced upregulation of p53 explains sebocyte apoptosis and sebum suppression as the major desired pharmacological effects in the treatment of acne [35,173]. Nelson et al. [174] demonstrated that tumor necrosis factor-related apoptosis inducing ligand (TRAIL) contributes to the apoptotic effect of isotretinoin in human sebocytes. Importantly, p53 upregulates the expression of the proapoptotic proteins TRAIL (TNFSF10) [175], FoxO1 (FOXO1A)[144,145] and FoxO3 (FOXO3A)[144], respectively. In contrast, survivin (BIRC5), a member of the inhibitors of the apoptosis gene family, which is overexpressed in serum of acne patients [176,177], is suppressed by p53 [178]. Thus, isotretinoin-mediated upregulation of nuclear p53 in SGs [147] represents most likely the desired sebum-suppressive effect in promoting sebocyte apoptosis. In SGs, isotretinoin (13-cis retinoic acid) is converted to all-trans retinoic acid (ATRA) [170]. Via binding to cellular retinoic acid binding protein 2 (CRABP2), ATRA is transported into the nucleus, where in a RAR-dependent fashion the transcription of p53 is upregulated [179]. ATRA/RAR-mediated upregulation of p53 expression has also been reported in other benign and malignant cells [180,181,182,183,184,185]. Preferential CRABP2 expression in basal and suprabasal layers of SGs is mandatory for ATRA-induced transcriptomic changes resulting in sufficient sebum suppression [186]. It is of critical functional importance that partitioning of ATRA between the nuclear receptors RAR and PPARβ/δ is regulated by the intracellular lipid binding proteins CRABP2 and fatty acid binding protein 5 (FABP5) [187]. In cells with a high CRABP2/FABP5 ratio, ATRA functions through RAR and is a proapoptotic agent, but in cells that highly express FABP5 it signals through PPARβ/δ and promotes cell survival [187]. Increased expression of CRABP2 in isotretinoin-treated SGs of patients with acne has been observed after weeks of oral isotretinoin exposure [186], whereas short-term (6 h, 24 h) isotretinoin exposure of immortalized SZ95 sebocytes did neither exhibit increased CRABP2 nor upregulated p53, FoxO1 or FoxO3 expression [188,189].
Recently, Seo et al. [143] reported that autophagy is constitutively active in maturing sebocytes of human SGs, whereas autophagy-related protein expression is repressed in SGs of acne patients. The authors found that isotretinoin activates autophagy in immortalized SZ95 sebocytes and induces sebosuppression [143]. Blocking autophagy using siRNA targeting autophagy-related 7 (ATG7) resulted in a significant loss of the sebosuppressive activity of isotretinoin in SZ95 sebocytes stimulated with testosterone and linoleic acid. Unfortunately, the investigators overlooked the reported intimate interaction between ATG7 and p53. ATG7 binding to p53 enhances the transcription of the cell cycle inhibitor p21 [190]. Among the genes directly activated by p53 are several autophagy genes, including ATG7 [191]. It is well established that there is an important relationship between autophagy and p53. Autophagy suppresses p53, whereas p53 also activates autophagy [192]. In this regard, studying autophagy in immortalized SZ95 sebocytes is critical because Simian virus transfection used for sebocyte immortalization is mediated by inactivation of p53 by direct binding of SV40 large T antigen to p53 [193,194]. Proapoptotic p53 signaling may thus be compromised in p53-inactivated immortalized sebocytes, which may thus be unable to execute to complete program of p53-induced apoptosis but instead remain in a preliminary state of autophagy. In contrast, human sebocytes in vivo are not artificially p53-inactivated and the clinical period to obtain an histological “involution” of SGs in acne patients takes weeks of isotretinoin treatment compared to short term cell culture studies [195].
7.5. Teratogenicity and Neural Crest Cell Apoptosis
Neural crest cells (NCCs) and neural crest (NC)-derived neuroblastoma cells are very susceptible to isotretinoin-induced apoptosis [173,196,197]. Neural precursor cells possess multiple p53-dependent apoptotic pathways [198]. Translational evidence suggests that isotretinoin-induced upregulation of p53 promotes NCC apoptosis suggested to operate as the major pathogenic mechanism of isotretinoin´s teratogenicity [199]. Increased p53 signaling is also associated with Treacher Collins-, CHARGE- and fetal alcohol syndrome, which all exhibit dysmorphic craniofacial features resembling retinoid embryopathy [199]. It has been demonstrated in mouse embryonic fibroblasts that p53 controls the NC/EMT gene network [200]. Upregulation or stabilization of p53 in the cranial neural tube reduces cranial NC delamination and promotes neural tube defects in chick embryos [200]. p53 was shown to play a major role in Treacher Collins syndrome, a congenital haploinsufficiency disorder in humans that arises from mutations in the TCOF1 gene. In the absence of one Tcof1 allele in the mouse, upregulation of p53-related apoptotic genes in NC progenitors results in severe craniofacial defects [201]. Keeping in mind that pathogenic mechanisms involved in Treacher Collins syndrome are dependent on the p53 pathway, recent in vivo ATRA-treatment results showed similar processes where ATRA-exposure led to an increase of apoptosis processes at late time points and thus could participate in producing developmental birth defects [202]. Using a proteomic approach, a new ATRA target, EFTUD2 (elongation factor Tu GTP-binding domain-containing 2) has been identified whose dysregulation leads to craniofacial defects [202]. Homozygous deletion of Eftud2 causes brain and craniofacial malformations, affecting the same precursors as in mandibulofacial dysostosis with microcephaly patients [203]. Remarkably, increased p53 activity and NCC death are responsible for craniofacial malformations in the Eftud2; Wnt1-Cre2 mutant mouse model [203]. Overactivation of the p53 pathway in Eftud2 knockdown cells was attenuated by overexpression of non-spliced MDM2, and craniofacial development was improved when Eftud2-mutant embryos were treated with pifithrin-α, an inhibitor of p53 [203].
Thus, accumulating evidence underlines the role of overactivated p53 signaling in NCC apoptosis linking syndromes with dysmorphic craniofacial features with the pathogenesis of isotretinoin-induced embryopathy (teratogenicity).
7.6. Depression and Impaired Hippocampal Neurogenesis
The question whether isotretinoin causes depression and anxiety in acne patients is still a matter of debate [204,205,206,207,208,209]. However, a subgroup of patients may be at risk of developing depression or suicidal ideation with isotretinoin treatment [205,208]. The hippocampus is one of the brain regions where new neurons are constantly formed in a process called hippocampal neurogenesis [210,211,212,213]. Reduced hippocampal volume and low numbers of hippocampal neural progenitors have been reported in depressed humans [213]. The commonly used anti-depressive drug lithium has been shown to increase hippocampal neurogenesis in rodents and humans [214,215,216,217]. Long-term lithium therapy has been shown to suppress neuronal p53 levels [218,219] and reduces FoxO3 transcriptional activity by decreasing its intracellular content [220]. Intriguingly, acne is a possible adverse effect of lithium therapy [221,222,223], a potential result of attenuated p53/FoxO signaling. Remarkably, isotretinoin treatment of mice results in both decreased hippocampal neurogenesis and reduction in hippocampal volume [224,225]. Treatment of hypothalamic cells with 10 μM isotretinoin for 48 h decreased cell growth to 45.6 + 13% of control [225]. Griffin et al. [225] hypothesized that the ability of isotretinoin to decrease hypothalamic cell numbers may contribute to the increased depression-related behaviors observed in mice. Intracerebroventricularly applied ATRA to adult rats increased RARα protein expression in the hippocampus suggesting an activation of ATRA/RARα-induced signaling mechanisms [226]. In these rats, ATRA-induced impairments in hippocampal neurogenesis correlated with depression-like symptoms [226]. Remarkably, retinoic acid-inducible gene 1 (RAI1) was found to be significantly upregulated in brains from patients with schizophrenia, bipolar disorder, or major depression [227]. Gene expression profiling reveals the role of RAI1 like receptor signaling in p53 dependent apoptosis induced by psoralen + UV-A (PUVA) in keratinocytes [228]. A direct relationship between p53 expression and loss of viability in CNS neurons [229] and neuronal cell death is well established [230,231,232]. In fact, adenovirus-mediated delivery of the p53 gene causes cortical and hippocampal neuronal cell death with some features typical of apoptosis [233].
Recent studies have shown that FoxOs are as well implicated in the pathophysiology of depression [234,235]. They play an essential role for neural stem cell homeostasis [236]. In gerbil and mouse brains dephosphorylation of FoxO1 following transient forebrain ischemia resulted in FoxO1 translocation into the nucleus in neurons [237]. The activation of FoxO1 preceded delayed neuronal death in the vulnerable hippocampal regions following ischemic brain injury. Notably, FoxO1 activation was accompanied by an increase in DNA binding activity for FoxO1-responsive element on the Fas ligand promoter (FASLG) [237]. In accordance to AKT-mediated regulation of FoxO1, AKT-induced phosphorylation of FoxO3 promotes cell survival via extrusion of FoxO3 from the nucleus into the cytoplasm [238,239], whereas nuclear non-phosphorylated FoxO3 triggers apoptosis inducing Fas ligand (FASLG) expression [239]. Of note, FOXO1A is a target of FoxO3, which induces the expression of FoxO1 [240]. IGF-1-induced AKT-mediated phosphorylation of FoxO3 has been observed in NC-derived PC12 cells, where IGF-1 inhibits apoptosis of neuronal cells [241]. Studies in murine models confirmed that FoxO1 and FoxO3 influence behavioral processes linked to anxiety and depression [242]. In fact, IGF-1-PI3K-AKT-mediated phosphorylation of FoxO3 induces survival of cultured hippocampal neurons [243]. In contrast, overexpression of FoxO3 induces apoptosis of cultured hippocampal neurons [243]. In this regard, it is of critical concern that we observed nuclear overexpression of both FoxO1 and FoxO3 in SGs of acne patients treated with isotretinoin [144], a pharmacologically constellation that may negatively affect the homeostasis hypothalamic neurons. In accordance, Kim et al. [244] showed that overexpression of FoxO1 suppresses neuronal differentiation, whereas in the absence of FoxO-dependent homeostatic processes, there was significant decrease in the neuronal stem cell pool and accompanying neurogenesis in the adult mouse brain.
Thus, isotretinoin-mediated upregulation of p53, FoxO1 and FoxO3 might negatively affect hypothalamic neurogenesis eventually increasing the risk of depression in predisposed individuals.
7.7. Reduced Ovarian Reserve and Granulosa Cell Apoptosis
Systemic isotretinoin also modifies the pituitary-ovarian axis causing mild suppression of pituitary hormone levels including growth hormone (GH), thyroid stimulating hormone (TSH), prolactin, adrenocorticotropic hormone (ACTH), and luteinizing hormone (LH) [245,246]. Follicle-stimulating hormone (FSH) and LH are required for the maturation of ovarian follicles. FoxO1 is critically involved in the regulation of gonadotropin expression [247,248,249,250]. Increased nuclear localization of FoxO1 decreases FSH β-polypeptide (FSHB) mRNA levels in murine primary pituitary cells [248]. FoxO1 overexpression in pituitary gonadotrope cells also inhibits the transcription of the β-subunit of LH (LHB) [249].
In rats, isotretinoin reduced ovarian reserve, a process associated with granulosa cell (GC) apoptosis [251]. In women treated with isotretinoin, anti-Müllerian hormone, antral follicle count and ovarian volume were significantly reduced compared to pretreatment values pointing to negative effects on ovarian reserve [252]. FoxO1 and FoxO3 are highly expressed in GCs of ovarian follicles. They interact with activin to regulate genes controlling follicle growth or with bone morphogenic protein 2 (BMP2) to control genes associated with metabolic stress and apoptosis leading to follicle death [253]. Overexpression of FoxO1 inhibits the viability of GCs in the mouse [254]. Constitutive activation of FoxO1 in murine GCs not only abolishes the protection from FSH, but activates autophagic gene expression [255]. Further experimental evidence in mice and chicken supports a critical role of FoxO1 and FoxO3 in GC apoptosis [256,257]. In addition, FSH and FoxO1 regulate genes in the sterol/steroid and lipid biosynthetic pathways in GCs [258]. It has been shown in porcine GCs that p53 promotes apoptosis and suppresses cell proliferation [259].
The impact of isotretinoin action on the pituitary-ovarian axis and GCs might as well be caused by overexpressed p53/FoxO1/FoxO3 signaling resulting in gonadotropin suppression and GC apoptosis, respectively [260].
7.8. Hypertriglyceridemia
Isotretinoin-mediated upregulation of p53 and FoxO1 also provides a reasonable explanation for isotretinoin-induced hyperlipidemia, which results primarily from hepatic oversecretion of apolipoprotein B100-containing very low density-lipoproteins (VLDL) [261,262]. Gustafson et al. [263] showed that rats respond with a prompt increase (+ 250%) in plasma triacylglycerol levels within the first few days after starting the administration of isotretinoin, which was exclusively due to a marked increase in VLDL. We observed a significant increase in VLDL apolipoprotein B (apoB) levels during isotretinoin treatment of acne patients [262]. The hepatic synthesis of apo B (B-100) is absolutely required for VLDL assembly and secretion. It contains several very hydrophobic areas that serve as strong lipid-binding domains [264,265]. Intriguingly, the APOB gene has been identified as a target gene of p53 [266]. Loading of hepatic triglycerides onto apoB100 is mediated by microsomal triglyceride transfer protein (MTTP), which is also essential for VLDL assembly and secretion and is induced by FoxO1 [267]. In addition, FoxO1 stimulates hepatic apolipoprotein C-III (APOC3) expression. FoxO1 binds to its consensus site in the APOC3 promoter [268,269]. In men, isotretinoin treatment (80 mg/d; 5 d) resulted in elevated plasma apoC-III concentrations at the transcriptional levels [270]. It has been shown that apoC-III strongly inhibits hepatic uptake of VLDL and intermediate density lipoproteins (IDL) [271] and inhibits the activity of lipoprotein lipase [272,273].
Thus, isotretinoin-mediated changes with transcriptional upregulation of p53 and FoxO1 explain isotretinoin-induced hypertriglyceridemia by increased hepatic VLDL apo B synthesis, triglyceride loading onto VLDL, heapticVLDL secretion but impaired VLDL triglyceride hydrolysis and hepatic VLDL and IDL uptake.
7.9. Increased Transepidermal Water Loss and Dry Skin
Dry skin is the most common mucocutaneous side effect of oral isotretinoin treatment [274]. Increased transepidermal water loss induced by oral isotretinoin treatment has been related to upregulated expression of aquaporin 3 (AQP3) [275]. AQP3 controls a water-, glycerol-, and hydrogen peroxide-transporting channel that plays a key role in various processes involved in keratinocyte function [276]. AQP3 is another p53 target gene [277].
Isotretinoin-induced p53 overexpression in epidermal keratinocytes [142] thus explains increased AQP3-mediated transepidermal water loss resulting in dry skin.
7.10. Intracranial Hypertension
Intracranial hypertension (pseudotumor cerebri) is a potential adverse effect of oral isotretinoin treatment [278,279]. An association between increased choroid plexus (ChP) aquaporin 1 (AQP1) and intracranial pressure has also been observed in obese Zucker rats [280]. AQP1 plays an essential role in water movement through ChP epithelial cells. Of note, CRABP2 is specifically expressed in ChP [281]. AQP1 is primarily located in the apical membrane of the ChP epithelium and has been implicated in playing a pivotal role in cerebrospinal fluid secretion [282]. AQP1 is widely distributed in the human brain and is associated with water secretion into the subarachnoid space and has been suggested to play a key role in idiopathic and drug-induced intracranial hypertension [283]. A close correlation between upregulated expression of p53, p21 and AQP1 has been reported in rat kidney, heart, lung and small intestine [284]. In fact, a recent bioinformatic analysis of the AQP1 promoter revealed the presence of DNA binding sites for p53 [285].
Thus, isotretinoin-induced overexpression of p53 in ChP epithelial cells may explain the occurrence of intracranial hypertension in predisposed individuals.
7.11. Inflammatory Flare at Initiation of Isotretinoin Treatment
Interleukin 1β (IL1B) mRNA and the active processed form of IL-1β are abundant in inflammatory acne lesions [286,287,288]. Caspase 1, also known as IL-1 converting enzyme, is a protease responsible for the processing of the key proinflammatory cytokine IL-1β from the inactive precursor pro-IL-1β to the active, secreted IL-1β [289,290]. In addition to the processing of IL-1β, caspase-1 plays an important and a conserved role as a cell death protease [288]. Remarkably, CASP1 is a target gene of p53 [291]. Thus, initiation of isotretinoin therapy with upregulation of p53 might explain the inflammatory acne flare at the beginning of isotretinoin treatment [292] and the occurrence of acne fulminans after initiation of isotretinoin therapy [293,294]. Growing evidence suggests that many of the signaling molecules known to regulate programmed cell death can also modulate inflammasome activation in a cell-intrinsic manner [295].
8. Limitations of Immortalized Human Sebocytes
In 1979, p53 was first identified in complex with the SV40 tumor-virus oncoprotein [296,297,298,299]. The immortalized human sebocyte cell lines SZ95 [300] and SEB-1 [301] were generated in 1999 and 2001, respectively. They are widely used tools for acne research and the exploration of pharmaceutical effects of potential anti-acne agents. Both immortalized sebocyte cell lines are generated by Simian virus (SV) transfection [300,301]. Importantly, SV40 large T antigen binds and inactivates p53 [193,194]. Furthermore, it has been shown that SV40 T antigen inhibits binding of p53 to cellular DNA [302]. As p53, called the guardian of the genome [303], plays key roles in the regulation of lipid metabolism [304,305,306,307,308,309], cell cycle control [310,311], autophagy [192] and apoptosis [312,313,314], viral inactivation of p53 by SV40 transfection will certainly cause deviations from normal physiological and pharmacological cellular responses compared to primary human non-immortalized sebocytes or in vivo SG specimens obtained from human subjects, in whom p53 is not artificially inactivated. Although an immortalized cell should generally be resistant against cell death, Wróbel et al. [315] reported apoptosis in human immortalized SZ95 sebocytes after 6 h of SZ95 sebocyte treatment with high staurosporine concentrations (10-6-10-5 M). However, treatment with 13-cis retinoic acid (10-8-10-5 M) did not affect externalized phosphatidylserine levels, DNA fragmentation, and lactate dehydrogenase cell release, despite increased caspase 3 levels leading to the conclusion that isotretinoin (13-cis retinoic acid) did not execute programmed death in human SZ95 sebocytes [315].
Under today´s perception, these former experiments require a critical re-evaluation: p53-inactivated SZ95 sebocytes apparently fail to induce a sufficient isotretinoin-mediated upregulation of p53 resulting in the incomplete execution of apoptosis. In addition, immortalized sebocytes treated with isotretinoin exhibit peculiar aberrations in the regulation of lipid metabolism. Short-term exposure of human SEB-1 sebocytes to 13-cis retinoic acid (10-7 M, 10-6 M and 10-5 M) for 48 hours did not exhibit apoptosis, but paradoxically induced a dose-dependent increase in total lipid synthesis, SREBF1 and 5-lipoxigenase (ALOX5) expression [188]. Of note, it has been reported that SV40 large T antigen/p53 complex activates the promoter of the IGF1 gene [316]. The isotretinoin-induced upregulation of p53 in SEB-1 cells may thus have increased the formation of SV40 large T antigen/p53 complex enhancing IGF-1-AKT-SREBF1 signaling enhancing lipogenesis [317]. A partial increase in isotretinoin-mediated upregulation of p53 in SEB-1 sebocytes may also explain the observed upregulation of ALOX5, a further target gene of p53 [318]. SREBF1 expression is under physiological conditions negatively regulated by p53 [309] pointing to insufficient isotretinoin/p53-mediated SREBF1 suppression in immortalized SEB-1 sebocytes. Also, the statement that “isotretinoin is indirectly effective in sebocytes” [189], requires a re-evaluation. Observed metabolic effects in immortalized p53-inactived sebocytes should not be transferred to responses of primary human sebocytes, which have never been used as physiological controls in the majority of experiments with immortalized sebocytes. However, SGs of patients treated with isotretinoin in vivo exhibit significant nuclear upregulation of p53 [147], marked sebum reduction [319] and SG involution [320], most likely driven by p53-mediated apoptosis [117,173,174]. Immortalized sebocytes apparently mostly respond reasonably when studied under proliferative stimuli like the exposure of IGF-1 and insulin, but have an overlooked handicap with proapoptotic signaling, especially isotretinoin-regulated transcription.
The final physiological fate of sebocytes in SGs is holocrine secretion [321], a unique DNase2-dependent mode of programmed cell death [322]. Notably, during Drosophila embryonic development, cell death eliminates 30% of the primordial germ cells. Therefore, an intrinsic alternative cell death pathway is operative and mediated by DNase2 release from lysosomes, leading to nuclear translocation and subsequent DNA double-strand breaks [323,324]. Notably, nuclear accumulation of DNase2 requires binding to the mitochondrial protein apoptosis inducing factor (AIFM1), which is upregulated by p53 [325]. This sheds a new light on the mode of isotretinoin-treatment, which via upregulation of p53 may accelerate AIFM1/DNase2-dependend programmed cell death, a phenomenon, which cannot be followed in p53-inactivated immortalized sebocytes. It is thus important for future research, to focus on the transcriptomics of death pathways to get deeper insights for the understanding of sebocyte biology as well as therapeutic responses [326].Table 1 and Table 2 summarize p53- and FoxO1-controlled target genes, which are critically involved in acne pathogenesis and isotretinoin´s mode of action.
9. Conclusions and Future Perspectives
The common inflammatory skin disease acne vulgaris, a highly prevalent disease in Western countries [327,328], is related to hereditary and multiple environmental factors [329,330], especially Western style diet (hyperglycemic carbohydrates, milk and dairy products) promoting insulin/IGF-1-driven nutrigenomic aberrations [331,332] (Figure 1). The observed changes in transcriptional regulation are primarily linked to downstream IGF-1/insulin/PI3K/AKT/mTORC1 signaling and androgen/AR/mTORC2/AKT signaling, which compromise the nuclear activities of p53, FoxO1 and FoxO3. Nuclear downregulation of FoxO1 results in transactivation of AR, SREBF1, PPARγ, STAT3 and attenuation of GATA6, critical transcription factors promoting sebocyte lipogenesis (increased sebum production), inflammation and infundibular hyperkeratinization (comedogenesis), respectively. AKT-MDM2-mediated suppression of p53 modifies the expression of critical p53 downstream target genes.
In contrast, the most effective anti-acne agent, isotretinoin, attenuates all major pathogenic mechanisms of the disease. Since the introduction of isotretinoin for the treatment of cystic and conglobate acne in 1979 by Peck et al. [333], isotretinoin´s inhibitory mode of action on the pilosebaceous unit remained a miracle. In the same year p53 was detected [296].
After four decades, evidence summarized in this review allows the conclusion that isotretinoin via upregulation of p53, FoxO1 and FoxO3 counteracts all major pathways in acne pathogenesis. Isotretinoin-induced sebocyte apoptosis with sebum suppression appears to be a result of upregulation of the proapoptotic transcription factors p53 [147], FoxO1 [144] and FoxO3 [146]. However, forced p53/FoxO1/FoxO3 signaling also explains the adverse effects of isotretinoin treatment including its teratogenicity (NCC apoptosis), reduction of ovarian reserve (granulosa cell apoptosis), risk of depression (reduced hypothalamic neurogenesis), dry skin (upregulation of AQP3), hypertriglyceridemia (upregulation of APOB) and risk of intracranial hypertension (upregulation of AQP1) among other adverse effects.
p53-inactivation by SV40 large T antigen in immortalized sebocytes (SZ95; SEB-1) is an overlooked critical limitation of these in vitro sebocyte models for acne research and certain pharmacological studies that depend on adequate death signaling pathways. After a period of enthusiasm, it´s apparently time to go back to primary sebocytes and in vivo studies. After four decades since the introduction of isotretinoin for acne therapy, we are at the beginning to understand its mode of action on the level of transcriptomic regulation. In the future, the role of STAT3 in acne pathogenesis deserves closer investigation, as STAT3 inhibits the expression of p53 [334] and interacts with FoxO1 [26]. Another highly interesting research question should be answered: Does the upregulation of p53 by isotretinoin accelerate the process of programmed sebocyte death by holocrine secretion?
Finally, it should be realized that the reported transcriptomic deviations exhibit a significant overlap with the transcriptomic landscape observed in common cancers of Western civilization, especially prostate and breast cancer [335,336,337]. It is thus not surprising that epidemiological studies reported an association between severe acne during adolescence and later risk of prostate cancer [338,339] and breast cancer [340], respectively. In contrast, the transcriptional changes induced by isotretinoin are useful in the prevention and therapy of various cancers [341,342,343,344,345].
Author Contributions
Conceptualization, methodology, formal analysis, writing, original draft preparation, all B.C.M.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Major parts of this review have been presented on at the Scientific Symposium at University Medical Center Minden, Germany, on the occasion of the 70th birthday of Prof. Dr. Rudolf Stadler on May 12, 2023. The author likes to thank Naglaa F. Agamia (Alexandria), Pedro Carrera-Bastos (Lisbon), WenChieh Chen (Zurich), Loren Cordain (Colorado), Swen Malte John (Osnabrück), Ayse Karadag (Istanbul), Claus Leitzmann (Giessen), Gerd Plewig (Munich), Gerd Schmitz (Regensburg), Ralf Weiskirchen (Aachen) and Christos Zouboulis (Dessau) for successful scientific cooperations and academic input and support over many years.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
| ACTH, adrenocorticotropic hormone |
| AIFM1, apoptosis-inducing factor, mitochondria associated |
| AKT, Akt kinase (protein kinase B) |
| ALOX5, arachidonate 5-lipoxigenate |
| APOB, apolipoprotein B |
| APOCIII, apolipoprotein CIII |
| AQP1, aquaporin 1 |
| AQP3, aquaporin 3 |
| AR, androgen receptor |
| ARE, androgen response element |
| ATG7, autophagy-related 7 |
| ATP, adenosine triphosphate |
| ATRA, all-trans retinoic acid |
| BIRC5, baculoviral IAP repeat-containing protein |
| BMP2, bone morphogenic protein 2 |
| C. acnes, Cutibacterium acnes |
| CASP1, caspase 1 |
| CDKN1A, cyclin-dependent kinase inhibitor 1A (p21) |
| ChP, choroid plexus |
| CRABP2, cellular retinoic acid binding protein 2 |
| DEPTOR, DEP-domain containing mTOR-interacting protein |
| EFTUD2, elongation factor Tu GTP-binding domain-containing 2 |
| FABP5, fatty acid binding protein 5 |
| FADS2, fatty acid desaturase 2 |
| FASLG, Fas ligand |
| FGFR2, fibroblast growth factor receptor 2 |
| FLG, filaggrin |
| FoxO1, forkhead box O1 |
| FoxO3, forkheadbox O3 |
| FSH, follicle stimulating hormone |
| FSHB, FSH β-polypeptide downregulation |
| FST, follistatin |
| GAP, GTPase-activating protein |
| GATA6, GATA binding protein 6 |
| GC, granulosa cell |
| GH, growth hormone |
| HIF-1, hypoxia-inducible factor 1 |
| HIF-2α, hypoxia-inducible factor 2α |
| IGF-1, insulin-like growth factor 1 |
| IGF1R, insulin-like growth factor receptor |
| IL-1β, interleukin-1β |
| IL-6, interleukin 6 |
| IL-8, interleukin 8 |
| IL-17, interleukin 17 |
| INSR, insulin receptor |
| Isotretinoin, 13-cis retinoic acid |
| LEP, leptin |
| LH, luteinizing hormone |
| LHB, β-subunit of LH downregulation |
| MDM2, MDM2 protooncogene |
| mTORC1, mechanistic target of rapamycin complex 1 |
| mTORC2, mechanistic target of rapamycin complex 2 |
| MTTP, microsomal triglyceride transfer protein |
| NC, neural crest |
| NCC, neural crest cell |
| OVOL1, ovo-like 1 |
| PGN, peptidoglycan |
| PI3K, phosphoinositide 3-kinase |
| PLIN2, perilipin 2 |
| POMC, proopiomelanocortin |
| PRKAB1, protein kinase, AMP-activated, noncatalytic, β-1 |
| PPAR/, peroxisome proliferator-activated receptor / |
| PPAR, peroxisome proliferator-activated receptor |
| PTEN, phosphatase and tensin homolog |
| RAI1, retinoic acid-inducible gene 1 |
| RAPTOR, regulatory associated protein of mTOR; |
| RAR, retinoic acid receptor |
| RARE, retinoic acid response element |
| RICTOR, rapamycin-insensitive companion of mTOR |
| RHEB, Ras protein homolog enriched in brain |
| RORγt, retinoic acid receptor-related orphan receptor-γt |
| SG, sebaceous gland |
| SMAD2, SMAD family member 2 |
| SREBF1, sterol regulatory element binding transcription factor 1 |
| STAT3, signal transducer and activator of transcription 3 |
| SV, Simian virus |
| TCOF1, treacle ribosome biogenesis factor 1 |
| TGFβ, transforming growth factor beta |
| TGFB2, transforming growth factor beta 2 |
| TLR2, toll-like receptor 2 |
| TLR4, toll-like receptor 4 |
| TNF-α, tumor necrosis factor-α |
| TNFSF10, tumor necrosis factor ligand superfamily, member 10 |
| TP53, tumor protein |
| TRAIL, tumor necrosis factor-related apoptosis inducing ligand (TNFSF10) |
| TSC1, TSC complex subunit 1 (hamartin) |
| TSC2, TSC complex subunit 2 (tuberin) |
| TSH, thyroid stimulating hormone |
| VLDL, very low density lipoprotein |
References
- Cong, T.X.; Hao, D.; Wen, X.; Li, X.H.; He, G. , Jiang, X. From pathogenesis of acne vulgaris to anti-acne agents. Arch. Dermatol. Res. 2019, 311, 337–349. [Google Scholar] [CrossRef]
- Acne Pathogenesis. In Plewig and Kligman´s Acne and Rosacea; Plewig, G., Melnik, B., Chen, W., Eds.; Springer: Cham, Germany, 2019; pp. 45–61. [Google Scholar] [CrossRef]
- Benito, M.; Valverde, A.M.; Lorenzo, M. IGF-I: a mitogen also involved in differentiation processes in mammalian cells. Int. J. Biochem. Cell Biol. 1996, 28, 499–510. [Google Scholar] [CrossRef]
- Hakuno, F.; Takahashi, S.I. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef]
- Christoforidis, A.; Maniadaki, I.; Stanhope, R. Growth hormone / insulin-like growth factor-1 axis during puberty. Pediatr. Endocrinol. Rev. 2005, 3, 5–10. [Google Scholar] [PubMed]
- Upners, E.N.; Busch, A.S.; Almstrup, K.; Petersen, J.H.; Assens, M.; Main, K.M.; Jensen, R.B.; Juul, A. Does height and IGF-I determine pubertal timing in girls? Pediatr. Res. 2021, 90, 176–183. [Google Scholar] [CrossRef]
- Racine, H.L.; Serrat, M.A. The actions of IGF-1 in the growth plate and its role in postnatal bone elongation. Curr. Osteoporos. Rep. 2020, 18, 210–227. [Google Scholar] [CrossRef] [PubMed]
- Fellowes, H.M.; Billewicz, W.Z.; Thomson, A.M. Is acne a sign of normal puberty? A longitudinal study. J. Biosoc. Sci. 1981, 13, 401–407. [Google Scholar] [CrossRef]
- Deplewski, D.; Rosenfield, R.L. Role of hormones in pilosebaceous unit development. Endocr. Rev. 2000, 21, 363–392. [Google Scholar] [CrossRef]
- Cappel, M.; Mauger, D.; Thiboutot, D. Correlation between serum levels of insulin-like growth factor 1, dehydroepiandrosterone sulfate, and dihydrotestosterone and acne lesion counts in adult women. Arch. Dermatol. 2005, 143, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Vora, S.; Ovhal, A; Jerajani, H. ; Nair, N.; Chakrabortty A. Correlation of facial sebum to serum insulin-like growth factor-1 in patients with acne. Br. J. Dermatol. 2008, 159, 990–991. [Google Scholar] [CrossRef]
- Seleit, I.; Bakry, O.A.; Abdou, A.G.; Hashim, A. Body mass index, selected dietary factors, and acne severity: are they related to in situ expression of insulin-like growth factor-1? Anal. Quant. Cytopathol. Histpathol. 2014, 36, 267–278. [Google Scholar]
- Burris, J.; Shikany, J.M.; Rietkerk, W.; Woolf, K. A low glycemic index and glycemic load diet decreases insulin-like growth factor-1 among adults with moderate and severe acne: A short-duration, 2-week randomized controlled trial. J. Acad. Nutr. Diet. 2018, 118, 1874–1885. [Google Scholar] [CrossRef]
- Qin, L.Q.; He, K.; Xu, J.Y. Milk consumption and circulating insulin-like growth factor-I level: a systematic literature review. Int. J. Food Sci. Nutr. 2009, 60 Suppl. 7, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Watling, C.Z.; Kelly, R.K.; Tong, T.Y.N.; Piernas, C.; Watts, E.L.; Tin Tin, S.; Knuppel, A.; Schmidt, J.A.; Travis, R.C.; Key, T.J.; et al. Associations of circulating insulin-like growth factor-I with intake of dietary proteins and other macronutrients. Clin. Nutr. 2021, 40, 4685–4693. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C.; Schmitz, G. Role of insulin, insulin-like growth factor-1, hyperglycaemic food and milk consumption in the pathogenesis of acne vulgaris. Exp. Dermatol. 2009, 18, 833–841. [Google Scholar] [CrossRef]
- Meixiong, J.; Ricco, C.; Vasavda, C.; Ho, B.K. Diet and acne: A systematic review. JAAD Int. 2022, 7, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Ben-Amitai, D.; Laron, Z. Effect of insulin-like growth factor-1 deficiency or administration on the occurrence of acne. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 950–954. [Google Scholar] [CrossRef]
- Klinger, B.; Anin, S.; Silbergeld, A.; Eshet, R.; Laron, Z. Development of hyperandrogenism during treatment with insulin-like growth factor-I (IGF-I) in female patients with Laron syndrome. Clin. Endocrinol. (Oxf.) 1998, 48, 81–87. [Google Scholar] [CrossRef]
- Manning, B.D.; Toker, A. AKT/PKB signaling: Navigating the network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef]
- Melnik, B.C. FoxO1 - the key for the pathogenesis and therapy of acne? J. Dtsch. Dermatol. Ges. 2010, 8, 105–114. [Google Scholar] [CrossRef]
- Mirdamadi, Y.; Thielitz, A.; Wiede, A.; Goihl, A.; Papakonstantinou, E.; Hartig, R.; Zouboulis, C.C. : Reinhold, D.; Simeoni, L.; Bommhardt, U.; et al. Insulin and insulin-like growth factor-1 can modulate the phosphoinositide-3-kinase/Akt/FoxO1 pathway in SZ95 sebocytes in vitro. Mol. Cell Endocrinol. 2015, 415, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Yanase, T.; Morinaga, H.; Okabe, T.; Nomura, M.; Daitoku, H.; Fukamizu, A.; Kato, S.; Takayanagi, R.; Nawata, H. Insulin-like growth factor 1/insulin signaling activates androgen signaling through direct interactions of Foxo1 with androgen receptor. J. Biol. Chem. 2007, 282, 7329–7338. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, W.; O-Sullivan, I.; Williams, J.B.; Dong, Q.; Park, E.A.; Raghow, R.; Unterman, T.G.; Elam, M.B. FoxO1 inhibits sterol regulatory element-binding protein-1c (SREBP-1c) gene expression via transcription factors Sp1 and SREBP-1c. J. Biol. Chem. 2012, 287, 20132–20143. [Google Scholar] [CrossRef]
- Fan, W.; Imamura, T.; Sonoda, N.; Sears, D.D.; Patsouris, D.; Kim, J.J.; Olefsky, J.M. FOXO1 transrepresses peroxisome proliferator-activated receptor gamma transactivation, coordinating an insulin-induced feed-forward response in adipocytes. J. Biol. Chem. 2009, 284, 12188–12197. [Google Scholar] [CrossRef]
- Yang, G.; Lim, C.Y.; Li, C.; Xiao, X.; Radda, G.K.; Li, C.; Cao, X.; Han, W. FoxO1 inhibits leptin regulation of pro-opiomelanocortin promoter activity by blocking STAT3 interaction with specificity protein 1. J. Biol. Chem. 2009, 284, 3719–3727. [Google Scholar] [CrossRef]
- Törőcsik, D.; Kovács, D.; Camera, E.; Lovászi, M.; Cseri, K.; Nagy, G.G.; Molinaro, R.; Rühl, R.; Tax, G.; Szabó, K.; et al. Leptin promotes a proinflammatory lipid profile and induces inflammatory pathways in human SZ95 sebocytes. Br. J. Dermatol. 2014, 171, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Zouboulis, C.C. Hormones and the pilosebaceous unit. Dermatoendocrinol. 2009, 1, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.M.; Gilliland, K.; Clawson, G.A.; Thiboutot, D. IGF-1 induces SREBP-1 expression and lipogenesis in SEB-1 sebocytes via activation of the phosphoinositide 3-kinase/Akt pathway. J. Invest. Dermatol. 2008, 128, 1286–1293. [Google Scholar] [CrossRef]
- Mastrofrancesco, A.; Ottaviani, M.; Cardinali, G.; Flori, E.; Briganti, S.; Ludovici, M.; Zouboulis, C.C.; Lora, V.; Camera, E.; Picardo, M. Pharmacological PPARγ modulation regulates sebogenesis and inflammation in SZ95 human sebocytes. Biochem. Pharmacol. 2017, 138, 96–106. [Google Scholar] [CrossRef]
- Kwon, H.H.; Yoon, J.Y.; Hong, J.S.; Jung, J.Y.; Park, M.S.; Suh, D.H. Clinical and histological effect of a low glycaemic load diet in treatment of acne vulgaris in Korean patients: a randomized, controlled trial. Acta Derm. Venereol. 2012, 92, 241–246. [Google Scholar] [CrossRef]
- Mayo, L.D.; Donner, D.B. A phosphatidylinositol 3-kinase/Akt pathway promotes translocation of Mdm2 from the cytoplasm to the nucleus. Proc. Natl. Acad. Sci. U S A 2001, 98, 11598–11603. [Google Scholar] [CrossRef] [PubMed]
- Haupt, Y.; Maya, R.; Kazaz, A.; Oren, M. Mdm2 promotes the rapid degradation of p53. Nature 1997, 387, 296–299. [Google Scholar] [CrossRef]
- Ogawara, Y.; Kishishita, S.; Obata, T.; Isazawa, Y.; Suzuki, T.; Tanaka, K.; Masuyama, N.; Gotoh, Y. Akt enhances Mdm2-mediated ubiquitination and degradation of p53. J. Biol. Chem. 2002, 277, 21843–21850. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. p53: key conductor of all anti-acne therapies. J. Transl. Med. 2017, 15, 195. [Google Scholar] [CrossRef] [PubMed]
- Dibble, C.C.; Manning, B.D. Signal integration by mTORC1 coordinates nutrient input with biosynthetic output. Nat. Cell Biol. 2013, 15, 555–564. [Google Scholar] [CrossRef]
- Ben-Sahra, I.; Manning, B.D. mTORC1 signaling and the metabolic control of cell growth. Curr. Opin. Cell Biol. 2017, 45, 72–82. [Google Scholar] [CrossRef]
- Inoki, K.; Li, Y.; Zhu, T.; Wu, J.; Guan, K.L. TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat. Cell Biol. 2002, 4, 648–657. [Google Scholar] [CrossRef]
- Huang, J.; Manning, B.D. The TSC1-TSC2 complex: a molecular switchboard controlling cell growth. Biochem. J. 2008, 412, 179–190. [Google Scholar] [CrossRef]
- Melnik, B. Dietary intervention in acne: Attenuation of increased mTORC1 signaling promoted by Western diet. Dermatoendocrinol. 2012, 4, 20–32. [Google Scholar] [CrossRef]
- Melnik, B.C.; Zouboulis, C.C. Potential role of FoxO1 and mTORC1 in the pathogenesis of Western diet-induced acne. Exp. Dermatol. 2013, 22, 311–315. [Google Scholar] [CrossRef]
- Monfrecola, G.; Lembo, S.; Caiazzo, G.; De Vita, V.; Di Caprio, R.; Balato, A. , Fabbrocini, G. Mechanistic target of rapamycin (mTOR) expression is increased in acne patients' skin. Exp. Dermatol. 2016, 25, 153–155. [Google Scholar] [CrossRef]
- Agamia, N.F.; Abdallah, D.M.; Sorour, O.; Mourad, B.; Younan, D.N. Skin expression of mammalian target of rapamycin and forkhead box transcription factor O1, and serum insulin-like growth factor-1 in patients with acne vulgaris and their relationship with diet. Br. J. Dermatol. 2016, 174, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Lembo, S.; Di Caprio, R.; Balato, A.; Caiazzo, G.; Fabbrocini, G.; Skroza, N.; Tolino, E.; Potenza, C. The increase of mTOR expression is consistent with FoxO1 decrease at gene level in acne but not in psoriasis. Arch. Dermatol. Res. 2020, 312, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C.; John, S.M.; Schmitz, G. Over-stimulation of insulin/IGF-1 signaling by western diet may promote diseases of civilization: lessons learnt from Laron syndrome. Nutr. Metab. (Lond.) 2011, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Milk--a nutrient system of mammalian evolution promoting mTORC1-dependent translation. Int. J. Mol. Sci. 2015, 16, 17048–17087. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. Regulation of mTORC1 and its impact on gene expression at a glance. J. Cell Sci. 2013, 126, 1713–1719. [Google Scholar] [CrossRef]
- Chakrabarti, P.; Anno, T.; Manning, B.D.; Luo, Z.; Kandror, K.V. The mammalian target of rapamycin complex 1 regulates leptin biosynthesis in adipocytes at the level of translation: the role of the 5'-untranslated region in the expression of leptin messenger ribonucleic acid. Mol. Endocrinol. 2008, 22, 2260–2267. [Google Scholar] [CrossRef]
- Chen, W.; Obermayer-Pietsch, B.; Hong, J.B.; Melnik, B.C.; Yamasaki, O.; Dessinioti, C.; Ju, Q.; Liakou, A.I.; Al-Khuzaei, S.; Katsambas, A.; et al. Acne-associated syndromes: models for better understanding of acne pathogenesis. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 637–646. [Google Scholar] [CrossRef]
- Shanik, M.H.; Xu, Y.; Skrha, J.; Dankner, R.; Zick, Y.; Roth, J. Insulin resistance and hyperinsulinemia: is hyperinsulinemia the cart or the horse? Diabetes Care 2008, 31, S262–S268. [Google Scholar] [CrossRef]
- Cai, W.; Sakaguchi, M.; Kleinridders, A.; Gonzalez-Del Pino, G.; Dreyfuss, J.M.; O'Neill, B.T.; Ramirez, A.K.; Pan, H.; Winnay, J.N.; Boucher, J.; et al. Domain-dependent effects of insulin and IGF-1 receptors on signalling and gene expression. Nat. Commun. 2017, 8, 14892. [Google Scholar] [CrossRef]
- Hoyt, G.; Hickey, M.S.; Cordain, L. Dissociation of the glycaemic and insulinaemic responses to whole and skimmed milk. Br. J. Nutr. 2005, 93, 175–177. [Google Scholar] [CrossRef]
- Hoppe, C.; Mølgaard, C.; Vaag, A.; Barkholt, V.; Michaelsen, K.F. High intakes of milk, but not meat, increase s-insulin and insulin resistance in 8-year-old boys. Eur. J. Clin. Nutr. 2005, 59, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Macut, D.; Bjekić-Macut, J.; Rahelić, D.; Doknić, M. Insulin and the polycystic ovary syndrome. Diabetes Res. Clin. Pract. 2017, 130, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Park, W.J.; Theda, C.; Maestri, N.E.; Meyers, G.A.; Fryburg, J.S.; Dufresne, C.; Cohen, M.M. Jr.; Jabs, E.W. Analysis of phenotypic features and FGFR2 mutations in Apert syndrome. Am. J. Hum. Genet. 1995, 57, 321–328. [Google Scholar] [PubMed]
- Xie, Y.; Su, N.; Yang, J.; Tan, Q.; Huang, S.; Jin, M.; Ni, Z.; Zhang, B.; Zhang, D.; Luo, F.; et al. FGF/FGFR signaling in health and disease. Signal Transduct. Target Ther. 2020, 5, 181. [Google Scholar] [CrossRef]
- Solomon, L.M.; Fretzin, D.; Pruzansky, S. Pilosebaceous abnormalities in Apert's syndrome. Arch. Dermatol. 1970, 102, 381–385. [Google Scholar] [CrossRef]
- Krafchik, B. Acne in Apert syndrome. Clin. Plast. Surg. 1991, 18, 407. [Google Scholar] [CrossRef]
- Munro, C.S.; Wilkie, A.O. Epidermal mosaicism producing localised acne: somatic mutation in FGFR2. Lancet 1998, 352, 704–705. [Google Scholar] [CrossRef]
- Melnik, B.C.; Vakilzadeh, F.; Aslanidis, C.; Schmitz, G. Unilateral segmental acneiform naevus: a model disorder towards understanding fibroblast growth factor receptor 2 function in acne? Br. J. Dermatol. 2008, 158, 1397–1399. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, B.; Wu, Z. Segmental hypopigmented acneiform nevus with FGFR2 gene mutation. An. Bras. Dermatol. 2023, 98, 710–712. [Google Scholar] [CrossRef]
- Melnik, B.; Schmitz, G. FGFR2 signaling and the pathogenesis of acne. J. Dtsch. Dermatol. Ges. 2008, 6, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Role of FGFR2-signaling in the pathogenesis of acne. Dermatoendocrinol. 2009, 1, 141–156. [Google Scholar] [CrossRef]
- Melnik, B.C.; Schmitz, G.; Zouboulis, C.C. Anti-acne agents attenuate FGFR2 signal transduction in acne. J. Invest. Dermatol. 2009, 129, 1868–1877. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Wei, Z.; Ju, Q.; Chen, W. Sex hormones and acne: State of the art. J. Dtsch. Dermatol. Ges. 2021, 19, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Belgorosky, A.; Baquedano, M.S.; Guercio, G.; Rivarola, M.A. Expression of the IGF and the aromatase/estrogen receptor systems in human adrenal tissues from early infancy to late puberty: implications for the development of adrenarche. Rev. Endocr. Metab. Disord. 2009, 10, 51–61. [Google Scholar] [CrossRef] [PubMed]
- De Mellow, J.S.; Handelsman, D.J.; Baxter, R.C. Short-term exposure to insulin-like growth factors stimulates testosterone production by testicular interstitial cells. Acta Endocrinol. (Copenh.) 1987, 115, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Haskell, J.; Vinson, N.; Terracio, L. Direct stimulatory effects of insulin-like growth factor-I on Leydig cell steroidogenesis in primary culture. Biochem. Biophys. Res. Commun. 1986, 137, 950–956. [Google Scholar] [CrossRef]
- Kasson, B.G.; Hsueh, A.J. Insulin-like growth factor-I augments gonadotropin-stimulated androgen biosynthesis by cultured rat testicular cells. Mol. Cell Endocrinol. 1987, 52, 27–34. [Google Scholar] [CrossRef]
- Cara, J.F.; Rosenfield, R.L. Insulin-like growth factor I and insulin potentiate luteinizing hormone-induced androgen synthesis by rat ovarian thecal-interstitial cells. Endocrinology 1988, 123, 733–739. [Google Scholar] [CrossRef]
- Cara, J.F. Insulin-like growth factors, insulin-like growth factor binding proteins and ovarian androgen production. Horm. Res. 1994, 42, 49–54. [Google Scholar] [CrossRef]
- Horton, R.; Pasupuletti, V.; Antonipillai, I. Androgen induction of steroid 5 alpha-reductase may be mediated via insulin-like growth factor-I. Endocrinology 1993, 133, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Grino, P.B. , Griffin, J.E., Wilson, J.D. Testosterone at high concentrations interacts with the human androgen receptor similarly to dihydrotestosterone. Endocrinology 1990, 126, 1165–1172. [Google Scholar] [CrossRef]
- Davey, R.A.; Grossmann, M. Androgen receptor structure, function and biology: From bench to bedside. Clin. Biochem. Rev. 2016, 37, 3–15. [Google Scholar] [PubMed]
- Barrault, C.; Garnier, J.; Pedretti, N.; Cordier-Dirikoc, S.; Ratineau, E.; Deguercy, A.; Bernard, F.X. Androgens induce sebaceous differentiation in sebocyte cells expressing a stable functional androgen receptor. J. Steroid. Biochem. Mol. Biol. 2015, 152, 34–44. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, W.; Zhao, J.; Pan, J.; Wu, Q.; Zhang, Y.; Bauman, W.A.; Cardozo, C.P. Identification of androgen response elements in the insulin-like growth factor I upstream promoter. Endocrinology 2007, 148, 2984–2993. [Google Scholar] [CrossRef]
- Pandini, G.; Mineo, R.; Frasca, F.; Roberts, C.T., Jr.; Marcelli, M.; Vigneri, R.; Belfiore, A. Androgens up-regulate the insulin-like growth factor-I receptor in prostate cancer cells. Cancer Res. 2005, 65, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Zhang, T.; Dizeyi, N.; Chen, S.; Wang, H.; Swanson, K.D.; Cai, C.; Balk, S.P.; Yuan, X. Androgen receptor enhances p27 degradation in prostate cancer cells through rapid and selective TORC2 activation. J. Biol. Chem. 2012, 287, 2090–2098. [Google Scholar] [CrossRef]
- Koryakina, Y.; Ta, H.Q.; Gioeli, D. Androgen receptor phosphorylation: biological context and functional consequences. Endocr. Relat. Cancer 2014, 21, T131–T145. [Google Scholar] [CrossRef]
- Alimirah, F.; Panchanathan, R.; Chen, J.; Zhang, X.; Ho, S.M.; Choubey, D. Expression of androgen receptor is negatively regulated by p53. Neoplasia 2007, 9, 1152–1159. [Google Scholar] [CrossRef]
- Imperato-McGinley, J.; Gautier, T.; Cai, L.Q.; Yee, B.; Epstein, J.; Pochi, P. The androgen control of sebum production. Studies of subjects with dihydrotestosterone deficiency and complete androgen insensitivity. J. Clin. Endocrinol. Metab. 1993, 76, 524–528. [Google Scholar] [CrossRef]
- Danby, F.W. Ductal hypoxia in acne: is it the missing link between comedogenesis and inflammation? J. Am. Acad. Dermatol. 2014, 70, 948–949. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Is sebocyte-derived leptin the missing link between hyperseborrhea, ductal hypoxia, inflammation and comedogenesis in acne vulgaris? Exp. Dermatol. 2016, 25, 181–182. [Google Scholar] [CrossRef]
- Corcoran, S.E.; O'Neill, L.A. HIF1α and metabolic reprogramming in inflammation. J. Clin. Invest. 2016, 126, 3699–3707. [Google Scholar] [CrossRef] [PubMed]
- Grosfeld, A.; Andre, J.; Hauguel-De Mouzon, S.; Berra, E. , Pouyssegur, J.; Guerre-Millo, M. Hypoxia-inducible factor 1 transactivates the human leptin gene promoter. J. Biol. Chem. 2002, 277, 42953–42957. [Google Scholar] [CrossRef]
- Frank, S.; Stallmeyer, B.; Kämpfer, H.; Kolb, N.; Pfeilschifter, J. Leptin enhances wound re-epithelialization and constitutes a direct function of leptin in skin repair. J. Clin. Invest. 2000, 106, 501–509. [Google Scholar] [CrossRef]
- Wong, W.J.; Richardson, T.; Seykora, J.T.; Cotsarelis, G.; Simon, M.C. Hypoxia-inducible factors regulate filaggrin expression and epidermal barrier function. J. Invest. Dermatol. 2015, 135, 454–461. [Google Scholar] [CrossRef]
- Kurokawa, I.; Mayer-da-Silva, A.; Gollnick, H.; Orfanos, C.E. Monoclonal antibody labeling for cytokeratins and filaggrin in the human pilosebaceous unit of normal, seborrhoeic and acne skin. J. Invest. Dermatol. 1988, 91, 566–571. [Google Scholar] [CrossRef]
- Lee, M.; Lee, E.; Jin, S.H.; Ahn, S.; Kim, S.O.; Kim, J.; Choi, D.; Lim, K.M.; Lee, S.T.; Noh, M. Leptin regulates the pro-inflammatory response in human epidermal keratinocytes. Arch. Dermatol. Res. 2018, 310, 351–362. [Google Scholar] [CrossRef]
- Shi, L.Z.; Wang, R.; Huang, G.; Vogel, P.; Neale, G.; Green, D.R.; Chi, H. HIF1alpha-dependent glycolytic pathway orchestrates a metabolic checkpoint for the differentiation of TH17 and Treg cells. J. Exp. Med. 2011, 208, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Dang, E.V.; Barbi, J.; Yang, H.Y.; Jinasena, D.; Yu, H.; Zheng, Y.; Bordman, Z.; Fu, J.; Kim, Y.; Yen, H.R.; et al. Control of T(H)17/T(reg) balance by hypoxia-inducible factor 1. Cell 2011, 146, 772–784. [Google Scholar] [CrossRef]
- Kierans, S.J.; Taylor, C.T. Regulation of glycolysis by the hypoxia-inducible factor (HIF): implications for cellular physiology. J. Physiol. 2021, 599, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic glycolysis: meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Khin, P.P.; Lim, O.K.; Jun, H.S. LPA/LPAR1 signaling induces PGAM1 expression via AKT/mTOR/HIF-1α pathway and increases aerobic glycolysis, contributing to keratinocyte proliferation. Life Sci. 2022, 311, 121201. [Google Scholar] [CrossRef]
- Shou, Y.; Zhu, R.; Tang, Z.; Man, X.Y. A prediction model identifying glycolysis signature as therapeutic target for psoriasis. Front. Immunol. 2023, 14, 1188745. [Google Scholar] [CrossRef] [PubMed]
- Downie, M.M.; Kealey, T. Human sebaceous glands engage in aerobic glycolysis and glutaminolysis. Br. J. Dermatol. 2004, 151, 320–327. [Google Scholar] [CrossRef]
- Soto-Heredero, G.; Gómez de Las Heras, M.M.; Gabandé-Rodríguez, E.; Oller, J.; Mittelbrunn, M. Glycolysis - a key player in the inflammatory response. FEBS J. 2020, 287, 3350–3369. [Google Scholar] [CrossRef]
- Kelhälä, H.L.; Palatsi, R.; Fyhrquist, N.; Lehtimäki, S.; Väyrynen, J.P.; Kallioinen, M.; Kubin, M.E.; Greco, D.; Tasanen, K.; Alenius, H.; et al. IL-17/Th17 pathway is activated in acne lesions. PLoS One 2014, 9, e105238. [Google Scholar] [CrossRef]
- Sitohang, I.B.S.; Soebaryo, R.W.; Kanoko, M. Acne lesion extraction versus oral doxycycline for moderate acne vulgaris: A randomized clinical trial. J. Clin. Aesthet. Dermatol. 2021, 14, E61–E65. [Google Scholar]
- Choi, K.; Jin, M.; Zouboulis, C.C.; Lee, Y. Increased lipid accumulation under hypoxia in SZ95 human sebocytes. Dermatology 2021, 237, 131–141. [Google Scholar] [CrossRef]
- Li, C.H.; Liao, P.L.; Yang, Y.T.; Huang, S.H. , Lin, C.H.; Cheng, Y.W.; Kang, J.J. Minocycline accelerates hypoxia-inducible factor-1 alpha degradation and inhibits hypoxia-induced neovasculogenesis through prolyl hydroxylase, von Hippel-Lindau-dependent pathway. Arch. Toxicol. 2014, 88, 659–671. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Hekmatjah, J.; Kircik, L.H. Oral tetracyclines and acne: A systematic review for dermatologists. J. Drugs Dermatol. 2020, 19, s6–s13. [Google Scholar]
- Eichenfield, D.Z. : Sprague, J.; Eichenfield, L.F. Management of acne vulgaris: A review. JAMA 2021, 326, 2055–2067. [Google Scholar] [CrossRef]
- Nazzaro-Porro, M. Azelaic acid. J. Am. Acad. Dermatol. 1987, 17, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Obacz, J.; Pastorekova, S.; Vojtesek, B.; Hrstka, R. Cross-talk between HIF and p53 as mediators of molecular responses to physiological and genotoxic stresses. Mol. Cancer 2013, 12, 93. [Google Scholar] [CrossRef]
- Kaluzová, M.; Kaluz, S.; Lerman, M.I.; Stanbridge, E.J. DNA damage is a prerequisite for p53-mediated proteasomal degradation of HIF-1alpha in hypoxic cells and downregulation of the hypoxia marker carbonic anhydrase IX. Mol. Cell Biol. 2004, 24, 5757–5766. [Google Scholar] [CrossRef]
- Choy, M.K.; Movassagh, M.; Bennett, M.R.; Foo, R.S. PKB/Akt activation inhibits p53-mediated HIF1A degradation that is independent of MDM2. J. Cell Physiol. 2010, 222, 635–639. [Google Scholar] [CrossRef]
- Joshi, S.; Singh, A.R.; Durden, D.L. MDM2 regulates hypoxic hypoxia-inducible factor 1α stability in an E3 ligase, proteasome, and PTEN-phosphatidylinositol 3-kinase-AKT-dependent manner. J. Biol. Chem. 2014, 289, 22785–22797. [Google Scholar] [CrossRef] [PubMed]
- Agamia, N.F.; Sorror, O.A.; Sayed, N.M.; Ghazala, R.A.; Echy, S.M.; Moussa, D.H.; Melnik, B.C. Overexpression of hypoxia-inducible factor-1α in hidradenitis suppurativa: the link between deviated immunity and metabolism. Arch. Dermatol. Res. 2023, 315, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Donati, G.; Rognoni, E.; Hiratsuka, T.; Liakath-Ali, K.; Hoste, E.; Kar, G.; Kayikci, M.; Russell, R.; Kretzschmar, K.; Mulder, K.W.; et al. Wounding induces dedifferentiation of epidermal Gata6+ cells and acquisition of stem cell properties. Nat. Cell Biol. 2017, 19, 603–613. [Google Scholar] [CrossRef]
- Swanson, J.B.; Vagnozzi, A.N.; Veniaminova, N.A.; Wong, S.Y. Loss of Gata6 causes dilation of the hair follicle canal and sebaceous duct. Exp. Dermatol. 2019, 28, 345–349. [Google Scholar] [CrossRef]
- Oulès, B.; Philippeos, C.; Segal, J.; Tihy, M.; Vietri Rudan, M.; Cujba, A.M.; Grange, P.A.; Quist, S.; Natsuga, K.; Deschamps, L.; et al. Contribution of GATA6 to homeostasis of the human upper pilosebaceous unit and acne pathogenesis. Nat. Commun. 2020, 11, 5067. [Google Scholar] [CrossRef]
- Okabe, Y.; Medzhitov, R. Tissue-specific signals control reversible program of localization and functional polarization of macrophages. Cell 2014, 157, 832–844. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.H.; Collins, S.L.; Sun, I.H.; Tam, A.J.; Patel, C.H.; Arwood, M.L.; Chan-Li, Y.; Powell, J.D.; Horton, M.R. mTORC2 signaling selectively regulates the generation and function of tissue-resident peritoneal macrophages. Cell Rep. 2017, 20, 2439–2454. [Google Scholar] [CrossRef]
- Xie, Y.; Jin, Y.; Merenick, B.L.; Ding, M.; Fetalvero, K.M.; Wagner, R.J.; Mai, A.; Gleim, S.; Tucker, D.F.; Birnbaum, M.J.; et al. Phosphorylation of GATA-6 is required for vascular smooth muscle cell differentiation after mTORC1 inhibition. Sci. Signal. 2015, 8, ra44. [Google Scholar] [CrossRef]
- Nelson, A.M.; Gilliland, K.L.; Cong, Z.; Thiboutot, D.M. 13-cis Retinoic acid induces apoptosis and cell cycle arrest in human SEB-1 sebocytes. J. Invest. Dermatol. 2006, 126, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- Engeland, K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022, 29, 946–960. [Google Scholar] [CrossRef]
- Perlman, H.; Suzuki, E.; Simonson, M.; Smith, R.C.; Walsh, K. GATA-6 induces p21(Cip1) expression and G1 cell cycle arrest. J. Biol. Chem. 1998, 273, 13713–13718. [Google Scholar] [CrossRef]
- Xu, X.; Ge, S.; Jia, R.; Zhou, Y.; Song, X.; Zhang, H.; Fan, X. Hypoxia-induced miR-181b enhances angiogenesis of retinoblastoma cells by targeting PDCD10 and GATA6. Oncol. Rep. 2015, 33, 2789–2796. [Google Scholar] [CrossRef] [PubMed]
- Navarini, A.A.; Simpson, M.A.; Weale, M.; Knight, J.; Carlavan, I.; Reiniche, P.; Burden, D.A.; Layton, A.; Bataille, V.; Allen, M.; et al. Genome-wide association study identifies three novel susceptibility loci for severe acne vulgaris. Nat. Commun. 2014, 5, 4020. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.L.; Saklatvala, J.R.; Dand, N.; Hagenbeek, F.A.; Li, X.; Min, J.L.; Thomas, L. , Bartels, M.; Jan Hottenga, J.; Lupton, M.K.; et al. Genome-wide association meta-analysis identifies 29 new acne susceptibility loci. Nat. Commun. 2022, 13, 702. [Google Scholar] [CrossRef]
- McNairn, A.J.; Doucet, Y.; Demaude, J.; Brusadelli, M.; Gordon, C.B.; Uribe-Rivera, A.; Lambert, P.F.; Bouez, C.; Breton, L.; Guasch, G. TGFβ signaling regulates lipogenesis in human sebaceous glands cells. BMC Dermatol. 2013, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shen, J.; Ying, J.; Xiao, D.; O'Keefe, R.J. FoxO1 is a crucial mediator of TGF-β/TAK1 signaling and protects against osteoarthritis by maintaining articular cartilage homeostasis. Proc. Natl. Acad. Sci. U S A 2020, 117, 30488–30497. [Google Scholar] [CrossRef]
- Huang, C.; Zhuo, F.; Han, B.; Li, W.; Jiang, B.; Zhang, K.; Jian, X.; Chen, Z.; Li, H.; Huang, H.; et al. . The updates and implications of cutaneous microbiota in acne. Cell Biosci. 2023, 13, 113. [Google Scholar] [CrossRef]
- Kistowska, M.; Meier, B.; Proust, T.; Feldmeyer, L.; Cozzio, A.; Kuendig, T.; Contassot, E.; French, L.E. Propionibacterium acnes promotes Th17 and Th17/Th1 responses in acne patients. J. Invest. Dermatol. 2015, 135, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Coenye, T.; He, L.; Kabashima, K.; Kobayashi, T.; Niemann, C.; Nomura, T.; Oláh, A.; Picardo, M.; Quist, S.R.; et al. Sebaceous immunobiology - skin homeostasis, pathophysiology, coordination of innate immunity and inflammatory response and disease associations. Front. Immunol. 2022, 13, 1029818. [Google Scholar] [CrossRef]
- Isard, O.; Knol, A.C.; Ariès, M.F.; Nguyen, J.M.; Khammari, A.; Castex-Rizzi, N.; Dréno, B. Propionibacterium acnes activates the IGF-1/IGF-1R system in the epidermis and induces keratinocyte proliferation. J. Invest. Dermatol. 2011, 131, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Jugeau, S.; Tenaud, I.; Knol, A.C.; Jarrousse, V.; Quereux, G.; Khammari, A.; Dreno, B. Induction of toll-like receptors by Propionibacterium acnes. Br. J. Dermatol. 2005, 153, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Marks, K.E.; Flaherty, S.; Patterson, K.M.; Stratton, M.; Martinez, G.J.; Reynolds, J.M. Toll-like receptor 2 induces pathogenicity in Th17 cells and reveals a role for IPCEF in regulating Th17 cell migration. Cell Rep. 2021, 35, 109303. [Google Scholar] [CrossRef]
- Jahns, A.C. , Eilers, H., Alexeyev, O.A. Transcriptomic analysis of Propionibacterium acnes biofilms in vitro. Anaerobe 2016, 42, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Mayslich, C.; Grange, P.A.; Dupin, N. Cutibacterium acnes as an opportunistic pathogen: An update of its virulence-associated factors. Microorganisms 2021, 9, 303. [Google Scholar] [CrossRef]
- Coenye, T.; Peeters, E.; Nelis, H.J. Biofilm formation by Propionibacterium acnes is associated with increased resistance to antimicrobial agents and increased production of putative virulence factors. Res. Microbiol. 2007, 158, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Spittaels. K.J.; Achermann, Y. The role of biofilm formation in the pathogenesis and antimicrobial susceptibility of Cutibacterium acnes. Biofilm 2021, 4, 100063. [Google Scholar] [CrossRef]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.G.; Ono-Moore, K.D.; Schneider, D.A.; Newman, J.W.; Adams, S.H.; Hwang, D.H. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef]
- Huang, Y.C.; Yang, C.H.; Li, T.T.; Zouboulis, C.C.; Hsu, H.C. Cell-free extracts of Propionibacterium acnes stimulate cytokine production through activation of p38 MAPK and Toll-like receptor in SZ95 sebocytes. Life Sci. 2015, 139, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.R.; Zhang, J.A.; Zhang, Q.; Permatasari, F.; Xu, Y.; Wu, D.; Yin, Z.Q.; Luo, D. Palmitic acid induces production of proinflammatory cytokines interleukin-6, interleukin-1β, and tumor necrosis factor-α via a NF-𝜅B-dependent mechanism in HaCaT keratinocytes. Mediators Inflammation 2013, 2013, 530429. [Google Scholar] [CrossRef]
- Vallerand, I.A.; Lewinson, R.T.; Farris, M.S.; Sibley, C.D.; Ramien, M.L.; Bulloch, A.G.M.; Patten, S.B. Efficacy and adverse events of oral isotretinoin for acne: A systematic review. Br. J. Dermatol. 2018, 178, 76–85. [Google Scholar] [CrossRef]
- Bagatin, E.; Costa, C.S. The use of isotretinoin for acne - an update on optimal dosing, surveillance, and adverse effects. Expert Rev. Clin. Pharmacol. 2020, 13, 885–897. [Google Scholar] [CrossRef]
- Melnik, B.C. Isotretinoin and FoxO1: A scientific hypothesis. Dermatoendocrinol. 2011, 3, 141–165. [Google Scholar] [CrossRef]
- Melnik, BC. Is nuclear deficiency of FoxO1 due to increased growth factor/PI3K/Akt-signalling in acne vulgaris reversed by isotretinoin treatment? Br. J. Dermatol. 2010, 162, 1398–1400. [Google Scholar] [CrossRef]
- Melnik, BC. The role of transcription factor FoxO1 in the pathogenesis of acne vulgaris and the mode of isotretinoin action. G. Ital. Dermatol. Venereol. 2010, 145, 559–571. [Google Scholar]
- Shi, G.; Liao, P.Y.; Cai, X.L.; Pi, X.X.; Zhang, M.F.; Li, S.J.; Quan, J.H.; Fan, Y.M. FoxO1 enhances differentiation and apoptosis in human primary keratinocytes. Exp. Dermatol. 2018, 27, 1254–1260. [Google Scholar] [CrossRef]
- Seo, S.H.; Jung, J.Y.; Park, K.; Hossini, A.M.; Zouboulis, CC.; Lee, SE. Autophagy regulates lipid production and contributes to the sebosuppressive effect of retinoic acid in human SZ95 sebocytes. J. Dermatol. Sci. 2020, 98, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Agamia, N.F.; Hussein, O.M.; Abdelmaksoud, R.E.; Abdalla, D.M.; Talaat, I.M.; Zaki, E.I.; El Tawdy, A.; Melnik, B.C. Effect of oral isotretinoin on the nucleo-cytoplasmic distribution of FoxO1 and FoxO3 proteins in sebaceous glands of patients with acne vulgaris. Exp. Dermatol. 2018, 27, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Pappas, K.; Xu, J.; Zairis, S.; Resnick-Silverman, L.; Abate, F.; Steinbach, N.; Ozturk, S.; Saal, L.H.; Su, T.; Cheung, P.; et al. p53 maintains baseline expression of multiple tumor suppressor genes. Mol. Cancer Res. 2017, 15, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Renault, V.M.; Thekkat, P.U.; Hoang, K.L.; White, J.L.; Brady, C.A.; Kenzelmann Broz, D.; Venturelli, O.S.; Johnson, T.M.; Oskoui, P.R.; Xuan, Z.; et al. The pro-longevity gene FoxO3 is a direct target of the p53 tumor suppressor. Oncogene 2011, 30, 3207–3221. [Google Scholar] [CrossRef]
- Agamia, N.F. , El Mulla, K.F.; Alsayed, N.M.; Ghazala, R.M.; El Maksoud, R.E.A.; Abdelmeniem, I.M.; Talaat, I.M.; Zaki, I.I.; Sabah, R.M.; Melnik, B.C. Isotretinoin treatment upregulates the expression of p53 in the skin and sebaceous glands of patients with acne vulgaris. Arch. Dermatol. Res. 2023, 315, 1355–1365. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, N.; Hadden, T.J.; Rishi, A.K. Akt, FoxO and regulation of apoptosis. Biochim. Biophys. Acta 2011, 1813, 1978–1986. [Google Scholar] [CrossRef]
- He, W.; Zhang, A.; Qi, L.; Na, C.; Jiang, R.; Fan, Z.; Chen, J. FOXO1, a potential therapeutic target, regulates autophagic flux, oxidative stress, mitochondrial dysfunction, and apoptosis in human cholangiocarcinoma QBC939 cells. Cell Physiol. Biochem. 2018, 45, 1506–1514. [Google Scholar] [CrossRef]
- Jiang, Y.; Luo, W.; Zhou, F.; Gong, P.; Xiong, Y. The role of FOXO1-mediated autophagy in the regulation of bone formation. Cell Cycle 2023, 22, 829–840. [Google Scholar] [CrossRef]
- Maiuri, M.C.; Galluzzi, L.; Morselli, E.; Kepp, O.; Malik, S.A.; Kroemer, G. Autophagy regulation by p53. Curr. Opin. Cell Biol. 2010, 22, 181–185. [Google Scholar] [CrossRef]
- Schuler, M.; Green, D.R. Mechanisms of p53-dependent apoptosis. Biochem. Soc. Trans. 2001, 29, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Di, J.; Cao, H.; Bai, J.; Zheng, J. p53-mediated autophagic regulation: A prospective strategy for cancer therapy. Cancer Lett. 2015, 363, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Maki, C.G. The IGF-1R/AKT pathway determines cell fate in response to p53. Transl. Cancer Res. 2016, 5, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Zhang, H.; Levine, A.J.; Jin, S. The coordinate regulation of the p53 and mTOR pathways in cells. Proc. Natl. Acad. Sci. U S A 2005, 102, 8204–8209. [Google Scholar] [CrossRef]
- Feng, Z.; Levine, A. .J. The regulation of energy metabolism and the IGF-1/mTOR pathways by the p53 protein. Trends Cell Biol. 2010, 20, 427–434. [Google Scholar] [CrossRef]
- Feng, Z. p53 regulation of the IGF-1/AKT/mTOR pathways and the endosomal compartment. Cold Spring Harb. Perspect. Biol. 2010, 2, a001057. [Google Scholar] [CrossRef]
- Goetz, E.M.; Shankar, B.; Zou, Y.; Morales, J.C.; Luo, X.; Araki, S.; Bachoo, R.; Mayo, L.D.; Boothman, D.A. ATM-dependent IGF-1 induction regulates secretory clusterin expression after DNA damage and in genetic instability. Oncogene 2011, 30, 3745–3754. [Google Scholar] [CrossRef] [PubMed]
- Karadag, A.S.; Ertugrul, D.T.; Tutal, E.; Akin, K.O. Short-term isotretinoin treatment decreases insulin-like growth factor-1 and insulin-like growth factor binding protein-3 levels: does isotretinoin affect growth hormone physiology? Br. J. Dermatol. 2010, 162, 798–802. [Google Scholar] [CrossRef]
- Werner, H.; Karnieli, E.; Rauscher, F.J.; LeRoith, D. Wild-type and mutant p53 differentially regulate transcription of the insulin-like growth factor I receptor gene. Proc. Natl. Acad. Sci. U S A 1996, 93, 8318–8323. [Google Scholar] [CrossRef]
- Sarfstein, R.; Werner, H. Tumor suppressor p53 regulates insulin receptor (INSR) gene expression via direct binding to the INSR promoter. Oncotarget 2020, 11, 2424–2437. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Hu, W.; de Stanchina, E.; Teresky, AK.; Jin, S.; Lowe, S.; Levine, A.J. The regulation of AMPK beta1, TSC2, and PTEN expression by p53: stress, cell and tissue specificity, and the role of these gene products in modulating the IGF-1-AKT-mTOR pathways. Cancer Res. 2007, 67, 3043–3053. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Qu, R.; Liu, D.; Xiong, X.; Liang, T.; Zhao, Y. The cross talk between p53 and mTOR pathways in response to physiological and genotoxic stresses. Front. Cell Dev. Biol. 2021, 9, 775507. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Dai, X.; Gong, L.; Chen, X.; Wang, L.; Xiong, X.; Zhao, Y. DEPTOR is a direct p53 target that suppresses cell growth and chemosensitivity. Cell Death Dis. 2020, 11, 976. [Google Scholar] [CrossRef]
- Cottle, D.L.; Kretzschmar, K.; Schweiger, P.J.; Quist, S.R.; Gollnick, H.P.; Natsuga, K. , Aoyagi, S.; Watt, F.M. c-MYC-induced sebaceous gland differentiation is controlled by an androgen receptor/p53 axis. Cell Rep. 2013, 3, 427–441. [Google Scholar] [CrossRef]
- Guseva, N.V.; Rokhlin, O.W.; Glover, R.A.; Cohen, M.B. P53 and the proteasome regulate androgen receptor activity. Cancer Biol Ther. 2012, 13, 553–558. [Google Scholar] [CrossRef]
- Boudou, P.; Soliman, H.; Chivot, M.; Villette, J.M.; Vexiau, P.; Belanger, A.; Fiet, J. Effect of oral isotretinoin treatment on skin androgen receptor levels in male acneic patients. J. Clin. Endocrinol. Metab. 1995, 80, 1158–1161. [Google Scholar] [CrossRef]
- Melnik, B.C. Acne vulgaris: The metabolic syndrome of the pilosebaceous follicle. Clin. Dermatol. 2018, 36, 29–40. [Google Scholar] [CrossRef]
- Tsukada, M.; Schröder, M.; Roos, T.C.; Chandraratna, RA.; Reichert, U.; Merk, H.F.; Orfanos, C.E.; Zouboulis, C.C. 13-cis retinoic acid exerts its specific activity on human sebocytes through selective intracellular isomerization to all-trans retinoic acid and binding to retinoid acid receptors. J. Invest. Dermatol. 2000, 115, 321–327. [Google Scholar] [CrossRef]
- Huang, P.; Chandra, V.; Rastinejad, F. Retinoic acid actions through mammalian nuclear receptors. Chem. Rev. 2014, 114, 233–254. [Google Scholar] [CrossRef]
- Gudas, L.J.; Wagner, J.A. Retinoids regulate stem cell differentiation. J. Cell Physiol. 2011, 226, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Apoptosis may explain the pharmacological mode of action and adverse effects of isotretinoin, including teratogenicity. Acta Derm. Venereol. 2017, 97, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.M.; Cong, Z.; Gilliland, K.L.; Thiboutot, D.M. TRAIL contributes to the apoptotic effect of 13-cis retinoic acid in human sebaceous gland cells. Br. J. Dermatol. 2011, 165, 526–533. [Google Scholar] [CrossRef]
- Kuribayashi, K.; Krigsfeld, G.; Wang, W.; Xu, J.; Mayes, P.A.; Dicker, DT.; Wu, G.S; El-Deiry, W.S. TNFSF10 (TRAIL), a p53 target gene that mediates p53-dependent cell death. Cancer Biol. Ther. 2008, 7, 2034–2038. [Google Scholar] [CrossRef] [PubMed]
- Assaf, H.A.; Abdel-Maged, W.M.; Elsadek, B.E.; Hassan, M.H.; Adly, M.A.; Ali, S.A. Survivin as a novel biomarker in the pathogenesis of acne vulgaris and its correlation to insulin-like growth factor-I. Dis. Markers 2016, 2016, 7040312. [Google Scholar] [CrossRef] [PubMed]
- Aksoy Saraç, G.; Kader, S.; Akdağ, T. Elevated survivin levels in patients with acne vulgaris. J. Cosmet. Dermatol. 2022, 21, 1744–1748. [Google Scholar] [CrossRef]
- Mirza, A.; McGuirk, M.; Hockenberry, T.N.; Wu, Q.; Ashar, H.; Black, S.; Wen, S.F.; Wang, L.; Kirschmeier, P.; Bishop, W.R.; et al. Human survivin is negatively regulated by wild-type p53 and participates in p53-dependent apoptotic pathway. Oncogene 2002, 21, 2613–2622. [Google Scholar] [CrossRef]
- Melnik, B.C. Mechanism of action of isotretinoin. In: Retinoids in Dermatology (Karadag, A.S.; Aksoy, B.; Parish, L.C. (eds.), chap. 4, 1st ed., CRC Press, Taylor & Francis Group, Boca Raton, USA, 2020.
- Ronca, F.; Yee, K.S.; Yu, V.C. Retinoic acid confers resistance to p53-dependent apoptosis in SH-SY5Y neuroblastoma cells by modulating nuclear import of p53. J. Biol. Chem. 1999, 274, 18128–18134. [Google Scholar] [CrossRef]
- Zheng, A.; Mäntymaa, P.; Säily, M.; Savolainen, E.; Vähäkangas, K.; Koistinen, P. p53 pathway in apoptosis induced by all-trans-retinoic acid in acute myeloblastic leukaemia cells. Acta Haematol. 2000, 103, 135–143. [Google Scholar] [CrossRef]
- Curtin, J.C.; Dragnev, K.H.; Sekula, D.; Christie, A.J.; Dmitrovsky, E.; Spinella, M.J. Retinoic acid activates p53 in human embryonal carcinoma through retinoid receptor-dependent stimulation of p53 transactivation function. Oncogene 2001, 20, 2559–2569. [Google Scholar] [CrossRef]
- Mrass, P.; Rendl, M.; Mildner, M.; Gruber, F.; Lengauer, B.; Ballaun, C.; Eckhart, L.; Tschachler, E. Retinoic acid increases the expression of p53 and proapoptotic caspases and sensitizes keratinocytes to apoptosis: a possible explanation for tumor preventive action of retinoids. Cancer Res. 2004, 64, 6542–6548. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, F.; Yuan, Y.; Ding, C.; Zhang, L.; Li, Q. All-trans retinoic acid upregulates the expression of p53 via Axin and inhibits the proliferation of glioma cells. Oncol. Rep. 2013, 29, 2269–2274. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Luquin, I.; Lázcoz, P.; Celay, J.; Castresana, J.S.; Encío, I.J. In vitro assessment of the role of p53 on chemotherapy treatments in neuroblastoma cell lines. Pharmaceuticals (Basel) 2021, 14, 1184. [Google Scholar] [CrossRef] [PubMed]
- Sitzmann, J.H.; Bauer, F.W.; Cunliffe, W.J.; Holland, B.; Lemotte, P.K. In situ hybridization analysis of CRABP II expression in sebaceous follicles from 13-cis retinoic acid-treated acne patients. Br. J. Dermatol. 1995, 133, 241–248. [Google Scholar] [CrossRef]
- Schug, T.; Berry, D.C.; Shaw, N.S.; Travis, S.N.; Noy, N. Opposing effects of retinoic acid on cell growth result from alternate activation of two different nuclear receptors. Cell 2007, 129, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Burney, W.; Bosanac, S.S.; Nguyen, C.; Isseroff, R.R.; Sivamani, R.K. Short-term exposure of human sebocytes to 13-cis-retinoic acid induces acnegenic changes. Br. J. Dermatol. 2018, 179, 1201–1202. [Google Scholar] [CrossRef]
- Kovács, D.; Hegyi, K.; Szegedi, A.; Deák, D.; Póliska, S.; Rühl, R.; Zouboulis, C.C.; Törőcsik, D. Isotretinoin is indirectly effective in sebocytes. Br. J. Dermatol. 2020, 182, 1052–1054. [Google Scholar] [CrossRef]
- Lee, I.H.; Kawai, Y.; Fergusson, M.M.; Rovira, I.I.; Bishop, AJ.; Motoyama, N.; Cao, L.; Finkel, T. Atg7 modulates p53 activity to regulate cell cycle and survival during metabolic stress. Science 2012, 336, 225–228. [Google Scholar] [CrossRef]
- Kenzelmann Broz, D.; Spano Mello, S.; Bieging, K.T.; Jiang, D.; Dusek, R.L.; Brady, C.A.; Sidow, A.; Attardi, L.D. Global genomic profiling reveals an extensive p53-regulated autophagy program contributing to key p53 responses. Genes Dev. 2013, 27, 1016–1031. [Google Scholar] [CrossRef]
- White, E. Autophagy and p53. Cold Spring Harb. Perspect. Med. 2016, 6, a026120. [Google Scholar] [CrossRef]
- McCormick, F.; Clark, R.; Harlow, E.; Tjian, R. SV40 T antigen binds specifically to a cellular 53 K protein in vitro. Nature 1981, 292, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Dobbelstein, M.; Roth, J. The large T antigen of simian virus 40 binds and inactivates p53 but not p73. J. Gen. Virol. 1998, 79, 3079–3083. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.A.; Comite, H.; Mescon, H.; Pochi, P.E. Isotretinoin in the treatment of acne: histologic changes, sebum production, and clinical observations. Arch. Dermatol. 1982, 118, 555–558. [Google Scholar] [CrossRef]
- Lammer, E.J.; Armstrong, D.L. Malformations in hindbrain structures among humans exposed to isotretinoin (13-cis-retinoic acid) during early embryogenesis. In: Morriss-Kay, G. (editor). Retinoids in Normal Development and Teratogenesis. New York: Oxford University Press, 1991; p. 281-295.
- Draghici, C.C.; Miulescu, R.G.; Petca, R.C.; Petca, A.; Dumitrașcu, MC.; Șandru, F. Teratogenic effect of isotretinoin in both fertile females and males (Review). Exp. Ther. Med. 2021, 21, 534. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, R.S.; Geng, Y.; Klocke, B.J.; Roth, K.A. Neural precursor cells possess multiple p53-dependent apoptotic pathways. Cell Death Differ. 2006, 13, 1727–1739. [Google Scholar] [CrossRef]
- Melnik, B.C. Overexpression of p53 explains isotretinoin's teratogenicity. Exp.Dermatol. 2018, 27, 91–93. [Google Scholar] [CrossRef]
- Rinon, A.; Molchadsky, A.; Nathan, E.; Yovel, G.; Rotter, V.; Sarig, R.; Tzahor, E. p53 coordinates cranial neural crest cell growth and epithelial-mesenchymal transition/delamination processes. Development 2011, 138, 1827–1838. [Google Scholar] [CrossRef]
- Jones, N.C.; Lynn, M.L.; Gaudenz, K.; Sakai, D.; Aoto, K.; Rey, J.P.; Glynn, E.F.; Ellington, L.; Du, C.; Dixon, J.; et al. Prevention of the neurocristopathy Treacher Collins syndrome through inhibition of p53 function. Nat. Med. 2008, 14, 125–133. [Google Scholar] [CrossRef]
- Berenguer, M.; Darnaudery, M.; Claverol, S.; Bonneu, M.; Lacombe, D.; Rooryck, C. Prenatal retinoic acid exposure reveals candidate genes for craniofacial disorders. Sci. Rep. 2018, 8, 17492. [Google Scholar] [CrossRef]
- Beauchamp, M.C.; Djedid, A.; Bareke, E.; Merkuri, F.; Aber, R.; Tam, A.S.; Lines, M.A.; Boycott, K.M.; Stirling, P.C.; Fish, J.L.; et al. Mutation in Eftud2 causes craniofacial defects in mice via mis-splicing of Mdm2 and increased p53. Hum. Mol. Genet. 2021, 30, 739–757. [Google Scholar] [CrossRef]
- Ludot, M. , Mouchabac, S., Ferreri, F. Inter-relationships between isotretinoin treatment and psychiatric disorders: Depression, bipolar disorder, anxiety, psychosis and suicide risks. World J. Psychiatry 2015, 5, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.M.; Sobreira, G.; Velosa, J.; Telles Correia, D.; Filipe, P. Association of isotretinoin with depression and suicide: A review of current literature. J. Cutan. Med. Surg. 2018, 22, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Metekoglu, S.; Oral, E.; Ucar, C.; Akalin, M. Does isotretinoin cause depression and anxiety in acne patients? Dermatol. Ther. 2019, 32, e12795. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, J.; Wang, W.; Ai, M.; Zhang, Q.; Kuang, L. Use of isotretinoin and risk of depression in patients with acne: A systematic review and meta-analysis. BMJ Open 2019, 9, e021549. [Google Scholar] [CrossRef] [PubMed]
- Bremner, J.D. Isotretinoin and neuropsychiatric side effects: Continued vigilance is needed. J. Affect. Disord. Rep. 2021, 6, 100230. [Google Scholar] [CrossRef]
- Kridin, K.; Ludwig, R.J. Isotretinoin and the risk of psychiatric disturbances: A global study shedding new light on a debatable story. J. Am. Acad. Dermatol. 2023, 88, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S. Depression: a case of neuronal life and death? Biol. Psychiatry 2004, 56, 140–145. [Google Scholar] [CrossRef]
- Sapolsky, R.M. Depression, antidepressants, and the shrinking hippocampus. Proc. Natl. Acad. Sci. U S A 2001, 98, 12320–12322. [Google Scholar] [CrossRef]
- Vaidya, V.A.; Fernandes, K.; Jha, S. Regulation of adult hippocampal neurogenesis: relevance to depression. Expert Rev. Neurother. 2007, 7, 853–864. [Google Scholar] [CrossRef]
- Apple, D.M.; Fonseca, R.S.; Kokovay, E. The role of adult neurogenesis in psychiatric and cognitive disorders. Brain Res. 2017, 1655, 270–276. [Google Scholar] [CrossRef]
- Gomes-Leal, W. Adult hippocampal neurogenesis and affective disorders: New neurons for psychic well-being. Front. Neurosci. 2021, 15, 594448. [Google Scholar] [CrossRef]
- Chen, G.; Rajkowska, G.; Du, F.; Seraji-Bozorgzad, N.; Manji, H.K. Enhancement of hippocampal neurogenesis by lithium. J. Neurochem. 2000, 75, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Chang, M.Y.; Yu, I.T.; Kim, J.H.; Lee, S.H.; Lee, Y.S.; Son, H. Lithium selectively increases neuronal differentiation of hippocampal neural progenitor cells both in vitro and in vivo. J. Neurochem. 2004, 89, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Palmos, A.B.; Duarte, R.R.R.; Smeeth, D.M.; Hedges, C.; Nixon, D.F.; Thuret, S.; Powell, T.R. Lithium treatment and human hippocampal neurogenesis. Transl. Psychiatry 2021, 11, 555. [Google Scholar] [CrossRef]
- Lu, R.; Song, L.; Jope, R.S. Lithium attenuates p53 levels in human neuroblastoma SH-SY5Y cells. Neuroreport 1999, 10, 1123–1125. [Google Scholar] [CrossRef]
- Chen, R.W.; Chuang, D.M. Long term lithium treatment suppresses p53 and Bax expression but increases Bcl-2 expression. A prominent role in neuroprotection against excitotoxicity. J. Biol. Chem. 1999, 274, 6039–6042. [Google Scholar] [CrossRef]
- Mao, Z.; Liu, L.; Zhang, R.; Li, X. Lithium reduces FoxO3a transcriptional activity by decreasing its intracellular content. Biol. Psychiatry 2007, 62, 1423–1430. [Google Scholar] [CrossRef]
- Okrasinski, H. Lithium acne. Dermatologica 1977, 154, 251–253. [Google Scholar] [CrossRef]
- Oztas, P.; Aksakal, A.B.; Oztas, M.O.; Onder, M. Severe acne with lithium. Ann. Pharmacother. 2001, 35, 961–962. [Google Scholar] [CrossRef] [PubMed]
- Scarfi, F.; Arunachalam, M. Lithium acne. CMAJ 2013, 85, 1525. [Google Scholar] [CrossRef]
- Crandall, J.; Sakai, Y.; Zhang, J.; Koul, O.; Mineur, Y.; Crusio, W.E.; McCaffery, P. 13-cis-retinoic acid suppresses hippocampal cell division and hippocampal-dependent learning in mice. Proc. Natl. Acad. Sci. U S A 2004, 101, 5111–5116. [Google Scholar] [CrossRef]
- Griffin, J.N.; Pinali, D.; Olds, K.; Lu, N.; Appleby, L.; Doan, L.; Lane, M.A. 13-Cis-retinoic acid decreases hypothalamic cell number in vitro. Neurosci. Res. 2010, 68, 185–190. [Google Scholar] [CrossRef]
- Hu, P.; Wang, Y.; Liu, J.; Meng, F.T.; Qi, X.R.; Chen, L.; van Dam, A.M.; Joëls, M.; Lucassen, P.J.; Zhou, J.N. Chronic retinoic acid treatment suppresses adult hippocampal neurogenesis, in close correlation with depressive-like behavior. Hippocampus 2016, 26, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Haybaeck, J.; Postruznik, M.; Miller, C.L.; Dulay, J.R.; Llenos, I.C.; Weis, S. Increased expression of retinoic acid-induced gene 1 in the dorsolateral prefrontal cortex in schizophrenia, bipolar disorder, and major depression. Neuropsychiatr. Dis. Treat. 2015, 11, 279–289. [Google Scholar] [CrossRef]
- Chowdhari, S.; Saini, N. Gene expression profiling reveals the role of RIG1 like receptor signaling in p53 dependent apoptosis induced by PUVA in keratinocytes. Cell Signal. 2016, 28, 25–33. [Google Scholar] [CrossRef]
- Xiang, H.; Hochman, D.W.; Saya, H.; Fujiwara, T.; Schwartzkroin, P.A.; Morrison, R.S. Evidence for p53-mediated modulation of neuronal viability. J. Neurosci. 1996, 16, 6753–6765. [Google Scholar] [CrossRef]
- Morrison, R.S.; Kinoshita, Y. The role of p53 in neuronal cell death. Cell Death Differ. 2000, 7, 868–879. [Google Scholar] [CrossRef]
- Morrison, R.S.; Kinoshita, Y.; Johnson, M.D.; Guo, W.; Garden, G.A. p53-dependent cell death signaling in neurons. Neurochem. Res. 2003, 28, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Huang, Z.; Xia, J.; Cui, Z.; Li, L.; Qi, Z.; Liu, W. Gene expression profile associated with Asmt knockout-induced depression-like behaviors and exercise effects in mouse hypothalamus. Biosci. Rep. 2022, 42, BSR20220800. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Kinoshita, Y.; Knudson, C.M.; Korsmeyer, S.J.; Schwartzkroin, P.A.; Morrison, R.S. Bax involvement in p53-mediated neuronal cell death. J. Neurosci. 1998, 18, 1363–1373. [Google Scholar] [CrossRef]
- Wang, H.; Quirion, R.; Little, P.J.; Cheng, Y.; Feng, Z.P.; Sun, H.S.; Xu, J.; Zheng, W. Forkhead box O transcription factors as possible mediators in the development of major depression. Neuropharmacology 2015, 99, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Rana, T.; Behl, T.; Sehgal, A.; Mehta, V.; Singh, S.; Sharma, N.; Bungau, S. Elucidating the possible role of FoxO in depression. Neurochem. Res. 2021, 46, 2761–2775. [Google Scholar] [CrossRef] [PubMed]
- Paik, J.H.; Ding, Z.; Narurkar, R.; Ramkissoon, S.; Muller, F.; Kamoun, W.S.; Chae, S.S.; Zheng, H.; Ying, H.; Mahoney, J.; et al. FoxOs cooperatively regulate diverse pathways governing neural stem cell homeostasis. Cell Stem Cell 2009, 5, 540–553. [Google Scholar] [CrossRef]
- Fukunaga, K.; Ishigami, T.; Kawano, T. Transcriptional regulation of neuronal genes and its effect on neural functions: expression and function of forkhead transcription factors in neurons. J. Pharmacol. Sci. 2005, 98, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Brunet, A.; Park, J.; Tran, H.; Hu, L.S.; Hemmings, B.A.; Greenberg, M.E. Protein kinase SGK mediates survival signals by phosphorylating the forkhead transcription factor FKHRL1 (FOXO3a). Mol. Cell Biol. 2001, 21, 952–965. [Google Scholar] [CrossRef]
- Essaghir, A.; Dif, N.; Marbehant, C.Y.; Coffer, P.J.; Demoulin, J.B. The transcription of FOXO genes is stimulated by FOXO3 and repressed by growth factors. J. Biol. Chem. 2009, 284, 10334–10342. [Google Scholar] [CrossRef]
- Zheng, W.H.; Kar, S.; Quirion, R. Insulin-like growth factor-1-induced phosphorylation of the forkhead family transcription factor FKHRL1 is mediated by Akt kinase in PC12 cells. J. Biol. Chem. 2000, 275, 39152–39158. [Google Scholar] [CrossRef]
- Polter, A.; Yang, S.; Zmijewska, A.A.; van Groen, T.; Paik, J.H.; Depinho, R.A.; Peng, S.L.; Jope, R.S.; Li, X. Forkhead box, class O transcription factors in brain: regulation and behavioral manifestation. Biol. Psychiatry 2009, 65, 150–159. [Google Scholar] [CrossRef]
- Zheng, W.H.; Kar, S.; Quirion, R. Insulin-like growth factor-1-induced phosphorylation of transcription factor FKHRL1 is mediated by phosphatidylinositol 3-kinase/Akt kinase and role of this pathway in insulin-like growth factor-1-induced survival of cultured hippocampal neurons. Mol. Pharmacol. 2002, 62, 225–233. [Google Scholar] [CrossRef]
- Kim, D.Y.; Hwang, I.; Muller, F.L.; Paik, J.H. Functional regulation of FoxO1 in neural stem cell differentiation. Cell Death Differ. 2015, 22, 2034–2045. [Google Scholar] [CrossRef] [PubMed]
- Karadag, A.S.; Ertugrul, D.T.; Tutal, E, Akin, K. O. Isotretinoin influences pituitary hormone levels in acne patients. Acta Derm. Venereol. 2011, 91, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Karadag, A.S.; Takci, Z.; Ertugrul, D.T.; Bilgili, S.G.; Balahoroglu, R.; Takir, M. The effect of different doses of isotretinoin on pituitary hormones. Dermatology 2015, 230, 354–359. [Google Scholar] [CrossRef]
- Richards, J.S. , Sharma, S.C., Falender, A.E., Lo, Y.H. Expression of FKHR, FKHRL1, and AFX genes in the rodent ovary: Evidence for regulation by IGF-I, estrogen, and the gonadotropins. Mol. Endocrinol. 2002, 16, 580–599. [Google Scholar] [CrossRef]
- Skarra, D.V.; Arriola, D.J.; Benson, C.A.; Thackray, V.G. Forkhead box O1 is a repressor of basal and GnRH-induced Fshb transcription in gonadotropes. Mol. Endocrinol. 2013, 27, 1825–1839. [Google Scholar] [CrossRef]
- Arriola, D.J.; Mayo, S.L.; Skarra, D.V.; Benson, C.A.; Thackray, V.G. FOXO1 transcription factor inhibits luteinizing hormone β gene expression in pituitary gonadotrope cells. J. Biol. Chem. 2012, 287, 33424–33435. [Google Scholar] [CrossRef]
- Thackray, V.G. Fox tales: regulation of gonadotropin gene expression by forkhead transcription factors. Mol. Cell Endocrinol. 2014, 385, 62–70. [Google Scholar] [CrossRef]
- Abali, R.; Yuksel, M.A.; Aktas, C.; Celik, C.; Guzel, S.; Erfan, G.; Sahin, O. Decreased ovarian reserve in female Sprague-Dawley rats induced by isotretinoin (retinoic acid) exposure. Reprod. Biomed. Online 2013, 27, 184–191. [Google Scholar] [CrossRef]
- Aksoy, H.; Cinar, L.; Acmaz, G.; Aksoy, U.; Aydin, T.; Vurdem, U.E.; Oz, L.; Idem Karadag, O. , Kartal D. The effect of isotretinoin on ovarian reserve based on hormonal parameters, ovarian volume, and antral follicle count in women with acne. Gynecol. Obstet. Invest. 2015, 79, 78–82. [Google Scholar] [CrossRef]
- Liu, Z.; Castrillon, DH.; Zhou, W.; Richards, J.S. FOXO1/3 depletion in granulosa cells alters follicle growth, death and regulation of pituitary FSH. Mol. Endocrinol. 2013, 27, 238–252. [Google Scholar] [CrossRef]
- Shen, M.; Liu, Z.; Li, B.; Teng, Y.; Zhang, J.; Tang, Y.; Sun, SC.; Liu, H. Involvement of FoxO1 in the effects of follicle-stimulating hormone on inhibition of apoptosis in mouse granulosa cells. Cell Death Dis. 2014, 5, e1475. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Jiang, Y.; Guan, Z.; Cao, Y.; Li, L.; Liu, H.; Sun, S.C. Protective mechanism of FSH against oxidative damage in mouse ovarian granulosa cells by repressing autophagy. Autophagy 2017, 13, 1364–1385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Q.; Gao, B.W.; Wang, J.; Ren, Q.L.; Chen, J.F.; Ma, Q.; Zhang, Z.J.; Xing, B.S. Critical role of FoxO1 in granulosa cell apoptosis caused by oxidative stress and protective effects of grape seed procyanidin B2. Oxid. Med. Cell Longev. 2016, 2016, 6147345. [Google Scholar] [CrossRef]
- Cui, C.; Han, S.; Yin, H.; Luo, B.; Shen, X.; Yang, F.; Liu, Z.; Zhu, Q.; Li, D.; Wang, Y. FOXO3 is expressed in ovarian tissues and acts as an apoptosis initiator in granulosa cells of chickens. Biomed. Res. Int. 2019, 2019, 6902906. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Rudd, M.D.; Hernandez-Gonzalez, I.; Gonzalez-Robayna, I.; Fan, H.Y.; Zeleznik, A.J.; Richards, J.S. FSH and FOXO1 regulate genes in the sterol/steroid and lipid biosynthetic pathways in granulosa cells. Mol. Endocrinol. 2009, 23, 649–661. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Benco, A.; Tandlmajerova, A.; Vasícek, D.; Kotwica, J.; Darlak, K.; Valenzuela, F. Transcription factor p53 can regulate proliferation, apoptosis and secretory activity of luteinizing porcine ovarian granulosa cell cultured with and without ghrelin and FSH. Reproduction 2008, 136, 611–618. [Google Scholar] [CrossRef]
- Abdelhamed, A.; Ezz El-Dawla, R.; Karadag, A.S.; Agamia, N.F.; Melnik, B.C. The impact of isotretinoin on the pituitary-ovarian axis: An interpretative review of the literature. Reprod. Toxicol. 2021, 104, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Marsden, J. Hyperlipidaemia due to isotretinoin and etretinate: possible mechanisms and consequences. Br. J. Dermatol. 1986, 114, 401–407. [Google Scholar] [CrossRef]
- Melnik, B.; Bros, U.; Plewig, G. Characterization of apoprotein metabolism and atherogenic lipoproteins during oral isotretinoin treatment. Dermatologica 1987, 175 Suppl 1, 158–168. [Google Scholar] [CrossRef]
- Gustafson, S.; Vahlquist, C.; Sjöblom, L.; Eklund, A.; Vahlquist, A. Metabolism of very low density lipoproteins in rats with isotretinoin (13-cis retinoic acid)-induced hyperlipidemia. J. Lipid Res. 1990, 31, 183–190. [Google Scholar] [CrossRef]
- Chen, Z.; Newberry, E.P.; Norris, J.Y.; Xie, Y.; Luo, J.; Kennedy, S.M.; Davidson, N.O. ApoB100 is required for increased VLDL-triglyceride secretion by microsomal triglyceride transfer protein in ob/ob mice. J. Lipid Res. 2008, 49, 2013–2022. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, S.O.; Borén, J. Apolipoprotein B secretory regulation by degradation. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1334–1338. [Google Scholar] [CrossRef]
- Ashur-Fabian, O.; Har-Zahav, A.; Shaish, A.; Wiener Amram, H.; Margalit, O.; Weizer-Stern, O.; Dominissini, D.; Harats, D.; Amariglio, N.; Rechavi, G. apoB and apobec1, two genes key to lipid metabolism, are transcriptionally regulated by p53. Cell Cycle 2010, 9, 3761–3770. [Google Scholar] [CrossRef]
- Kamagate, A.; Dong, H.H. FoxO1 integrates insulin signaling to VLDL production. Cell Cycle 2008, 7, 3162–3170. [Google Scholar] [CrossRef] [PubMed]
- Altomonte, J.; Cong, L.; Harbaran, S.; Richter, A.; Xu, J.; Meseck, M.; Dong, H.H. Foxo1 mediates insulin action on apoC-III and triglyceride metabolism. J. Clin. Invest. 2004, 114, 1493–1503. [Google Scholar] [CrossRef]
- Peng, S.; Li, W.; Hou, N.; Huang, N. A Review of FoxO1-regulated metabolic diseases and related drug discoveries. Cells 2020, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Vu-Dac, N.; Gervois, P.; Torra, I.P.; Fruchart, J.C.; Kosykh, V.; Kooistra, T.; Princen, H.M.; Dallongeville, J.; Staels, B. Retinoids increase human apo C-III expression at the transcriptional level via the retinoid X receptor. Contribution to the hypertriglyceridemic action of retinoids. J. Clin. Invest. 1998, 102, 625–632. [Google Scholar] [CrossRef]
- Mendivil, C.O.; Zheng, C.; Furtado, J.; Lel, J.; Sacks, F.M. Metabolism of very-low-density lipoprotein and low-density lipoprotein containing apolipoprotein C-III and not other small apolipoproteins. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 239–245, Erratum in: Arterioscler. Thromb. Vasc. Biol. 2010, 30, e171. [Google Scholar] [CrossRef]
- Larsson, M.; Vorrsjö, E.; Talmud, P.; Lookene, A.; Olivecrona, G. Apolipoproteins C-I and C-III inhibit lipoprotein lipase activity by displacement of the enzyme from lipid droplets. J. Biol. Chem. 2013, 288, 33997–34008. [Google Scholar] [CrossRef]
- Larsson, M.; Allan, C.M.; Jung, R.S.; Heizer, P.J.; Beigneux, A.P.; Young, S.G.; Fong, L.G. Apolipoprotein C-III inhibits triglyceride hydrolysis by GPIHBP1-bound LPL. J. Lipid Res. 2017, 58, 1893–1902. [Google Scholar] [CrossRef]
- Çölgeçen, E.; Özyurt, K.; Ferahbaş Kesikoğlu, A. The effect of systemic isotretinoin treatment on skin biophysical parameters among patients with acne vulgaris. Turk. J. Med. Sci. 2016, 46, 1641–1644. [Google Scholar] [CrossRef]
- Xing, F.; Liao, W.; Jiang, P.; Xu, W.; Jin, X. Effect of retinoic acid on aquaporin 3 expression in keratinocytes. Genet. Mol. Res. 2016, 15, 15016951. [Google Scholar] [CrossRef]
- Bollag, W.B.; Aitkens, L.; White, J.; Hyndman, K.A. Aquaporin-3 in the epidermis: more than skin deep. Am. J. Physiol. Cell Physiol. 2020, 318, C1144–C1153. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, V.; Olala, L.O.; Kagha, K.; Pan, Z.Q.; Chen, X.; Yang, R.; Cline, A.; Helwa, I.; Marshall, L.; Kaddour-Djebbar, I.; et al. Regulation of the glycerol transporter, aquaporin-3, by histone deacetylase-3 and p53 in keratinocytes. J. Invest. Dermatol. 2017, 137, 1935–1944. [Google Scholar] [CrossRef]
- Fraunfelder, F.W.; Fraunfelder, T.; Corbett, J.J. Isotretinoin-associated intracranial hypertension. Ophthalmology 2004, 111, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Reifenrath, J.; Rupprecht, C.; Gmeiner, V.; Haslinger, B. Intracranial hypertension after rosacea treatment with isotretinoin. Neurol. Sci. 2023. [Google Scholar] [CrossRef]
- Uldall, M.; Bhatt, D.K.; Kruuse, C.; Juhler, M.; Jansen-Olesen, I.; Jensen, R.H. Choroid plexus aquaporin 1 and intracranial pressure are increased in obese rats: towards an idiopathic intracranial hypertension model? Int. J. Obes (Lond.) 2017, 41, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Bushue, N.; Wan, Y.J. Retinoid pathway and cancer therapeutics. Adv. Drug Deliv. Rev. 2010, 62, 1285–1298. [Google Scholar] [CrossRef]
- González-Marrero, I.; Hernández-Abad, G.; González-Gómez, M.; Soto-Viera, M.; Carmona-Calero, E.M.; Castañeyra-Ruiz, L.; Castañeyra-Perdomo, A. Altered expression of AQP1 and AQP4 in brain barriers and cerebrospinal fluid may affect cerebral water balance during chronic hypertension. Int. J. Mol. Sci. 2022, 23, 12277. [Google Scholar] [CrossRef]
- Stiebel-Kalish, H.; Eyal, S.; Steiner, I. The role of aquaporin-1 in idiopathic and drug-induced intracranial hypertension. Med. Hypotheses 2013, 81, 1059–1062. [Google Scholar] [CrossRef]
- Lv, W.; Xue, L.; Liang, L.; Liu, D.; Li, C.; Liao, J.; Jin, Y. Endotoxin induced acute kidney injury modulates expression of AQP1, P53 and P21 in rat kidney, heart, lung and small intestine. PLoS One 2023, 18, e0288507. [Google Scholar] [CrossRef]
- Abreu-Rodríguez, I.; Sánchez Silva, R.; Martins, A.P.; Soveral, G.; Toledo-Aral, JJ.; López- Barneo, J.; Echevarría, M. Functional and transcriptional induction of aquaporin-1 gene by hypoxia; analysis of promoter and role of Hif-1α. PLoS One 2011, 6, e28385. [Google Scholar] [CrossRef]
- Kistowska, M.; Gehrke, S.; Jankovic, D.; Kerl, K.; Fettelschoss, A.; Feldmeyer, L.; Fenini, G.; Kolios, A.; Navarini, A.; Ganceviciene, R.; et al. IL-1β drives inflammatory responses to propionibacterium acnes in vitro and in vivo. J. Invest. Dermatol. 2014, 134, 677–685. [Google Scholar] [CrossRef] [PubMed]
- ElAttar, Y.; Mourad, B.; Alngomy, H.A.; Shams El Deen, A.; Ismail, M. Study of interleukin-1 beta expression in acne vulgaris and acne scars. J. Cosmet. Dermatol. 2022, 21, 4864–4870. [Google Scholar] [CrossRef] [PubMed]
- Thanh, L.T.V.; Anh, L.V.; Giang, T.H.; Hung, T.Q.; Trung, V.T.; Vuong, N.L. Immunohistochemical expression of interleukin 1 beta in papule biopsies from patients with acne vulgaris. Dermatol. Reports 2022, 14, 9444. [Google Scholar] [CrossRef]
- Denes, A.; Lopez-Castejon, G.; Brough, D. Caspase-1: is IL-1 just the tip of the ICEberg? Cell Death Dis. 2012, 3, e338. [Google Scholar] [CrossRef]
- Sutterwala, F.S.; Ogura, Y.; Szczepanik, M.; Lara-Tejero, M.; Lichtenberger, G.S.; Grant, E.P.; Bertin, J.; Coyle, A.J.; Galán, J.E.; Askenase, P.W.; et al. Critical role for NALP3/CIAS1/cryopyrin in innate and adaptive immunity through its regulation of caspase-1. Immunity 2006, 24, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Radha, V.; Furukawa, Y.; Swarup, G. Direct transcriptional activation of human caspase-1 by tumor suppressor p53. J. Biol. Chem. 2001, 276, 10585–10588. [Google Scholar] [CrossRef]
- Borghi, A.; Mantovani, L.; Minghetti, S.; Virgili, A.; Bettoli, V. Acute acne flare following isotretinoin administration: potential protective role of low starting dose. Dermatology 2009, 218, 178–180. [Google Scholar] [CrossRef]
- Fakih, A; Goens, J. ; Grozdev, I.; Dangoisse, C.; Richert, B. Acne fulminans induced by a low dose isotretinoin: case report and review of the literature. Dermatol. Online J. 2020, 26, 13030–qt14h2419w. [Google Scholar]
- Yorulmaz, A.; Hayran, Y. Isotretinoin-induced acne fulminans. J. Cutan. Med. Surg. 2022, 26, 435. [Google Scholar] [CrossRef]
- Lee, K.H.; Kang, T.B. The molecular links between cell death and inflammasome. Cells 2019, 8, 1057. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.P.; Crawford, L.V. T antigen is bound to a host protein in SV40-transformed cells. Nature 1979, 278, 261–263. [Google Scholar] [CrossRef]
- Linzer, D.I.; Levine, A.J. Characterization of a 54K dalton cellular SV40 tumor antigen present in SV40-transformed cells and uninfected embryonal carcinoma cells. Cell 1979, 17, 43–52. [Google Scholar] [CrossRef]
- Kress, M.; May, E.; Cassingena, R.; May, P. Simian virus 40-transformed cells express new species of proteins precipitable by anti-simian virus 40 tumor serum. J. Virol. 1979, 31, 472–483. [Google Scholar] [CrossRef]
- DeLeo, A.B.; Jay, G.; Appella, E.; Dubois, G.C.; Law, L.W.; Old, L.J. Detection of a transformation-related antigen in chemically induced sarcomas and other transformed cells of the mouse. Proc. Natl. Acad. Sci. U S A 1979, 76, 2420–2424. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Seltmann, H.; Neitzel, H.; Orfanos, C.E. Establishment and characterization of an immortalized human sebaceous gland cell line (SZ95). J. Invest. Dermatol. 1999, 113, 1011–1020. [Google Scholar] [CrossRef]
- Thiboutot, D.; Jabara, S.; McAllister, J.M.; Sivarajah, A.; Gilliland, K.; Cong, Z.; Clawson, G. Human skin is a steroidogenic tissue: steroidogenic enzymes and cofactors are expressed in epidermis, normal sebocytes, and an immortalized sebocyte cell line (SEB-1). J. Invest. Dermatol. 2003, 120, 905–914. [Google Scholar] [CrossRef]
- Bargonetti, J.; Reynisdóttir, I.; Friedman, P.N.; Prives, C. Site-specific binding of wild-type p53 to cellular DNA is inhibited by SV40 T antigen and mutant p53. Genes Dev. 1992, 6, 1886–1898. [Google Scholar] [CrossRef]
- Lane, D.P. Cancer. p53, guardian of the genome. Nature 1992, 358, 15–16. [Google Scholar] [CrossRef]
- Vousden, K.H.; Ryan, K.M. p53 and metabolism. Nat. Rev. Cancer. 2009, 9, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, I.; Ezra, O.; Rivlin, N.; Molchadsky, A.; Madar, S.; Goldfinger, N.; Rotter, V. p53, a novel regulator of lipid metabolism pathways. J. Hepatol. 2012, 56, 656–662. [Google Scholar] [CrossRef]
- Lacroix, M.; Linares, L.K.; Rueda-Rincon, N.; Bloch, K.; Di Michele, M.; De Blasio, C.; Fau, C.; Gayte, L.; Blanchet, E.; Mairal, A.; et al. The multifunctional protein E4F1 links P53 to lipid metabolism in adipocytes. Nat. Commun. 2021, 12, 7037. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, C.; Hu, W.; Feng, Z. Tumor suppressor p53 and metabolism. J. Mol. Cell Biol. 2019, 11, 284–292. [Google Scholar] [CrossRef]
- Lacroix, M.; Riscal, R.; Arena, G.; Linares, L.K.; Le Cam, L. Metabolic functions of the tumor suppressor p53: Implications in normal physiology, metabolic disorders, and cancer. Mol. Metab. 2020, 33, 2–22. [Google Scholar] [CrossRef]
- Yahagi, N.; Shimano, H.; Matsuzaka, T.; Najima, Y.; Sekiya, M.; Nakagawa, Y.; Ide, T.; Tomita, S.; Okazaki, H.; Tamura, Y.; et al. p53 Activation in adipocytes of obese mice. J. Biol. Chem. 2003, 278, 25395–25400. [Google Scholar] [CrossRef] [PubMed]
- Oren, M. Decision making by p53: life, death and cancer. Cell Death Differ. 2003, 10, 431–442. [Google Scholar] [CrossRef]
- Engeland, K. Cell cycle regulation: p53-p21-RB signaling. Cell Death Differ. 2022, 29, 946–960. [Google Scholar] [CrossRef]
- Shen, Y.; White, E. p53-dependent apoptosis pathways. Adv. Cancer Res. 2001, 82, 55–84. [Google Scholar] [CrossRef]
- Zamzami, N.; Kroemer, G. p53 in apoptosis control: an introduction. Biochem. Biophys. Res. Commun. 2005, 331, 685–687. [Google Scholar] [CrossRef]
- Meulmeester, E.; Jochemsen, A.G. p53: a guide to apoptosis. Curr. Cancer Drug Targets 2008, 8, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Wróbel, A.; Seltmann, H.; Fimmel, S.; Müller-Decker, K.; Tsukada, M.; Bogdanoff, B.; Mandt, N.; Blume-Peytavi, U.; Orfanos, C.E.; Zouboulis, C.C. Differentiation and apoptosis in human immortalized sebocytes. J. Invest. Dermatol. 2003, 120, 175–181. [Google Scholar] [CrossRef]
- Bocchetta, M.; Eliasz, S.; De Marco, M.A.; Rudzinski, J.; Zhang, L.; Carbone, M. The SV40 large T antigen-p53 complexes bind and activate the insulin-like growth factor-I promoter stimulating cell growth. Cancer Res. 2008, 68, 1022–1029. [Google Scholar] [CrossRef]
- Melnik, B.C.; John, S.M.; Agamia, N.F.; Chen, W.; De Vita, V.; Karadag, A.S.; Plewig, G.; Schmitz, G. Isotretinoin's paradoxical effects in immortalized sebocytes. Br. J. Dermatol. 2019, 180, 957–958. [Google Scholar] [CrossRef]
- Kumar, R.; Coronel, L.; Somalanka, B.; Raju, A.; Aning, O.A.; An, O.; Ho, Y.S.; Chen, S.; Mak. , SY.; Hor, P.Y.; et al. Mitochondrial uncoupling reveals a novel therapeutic opportunity for p53-defective cancers. Nat. Commun. 2018, 9, 3931. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.E.; Benoit, A.M.; Downing, D.T.; Strauss, J.S. Suppression of sebum secretion with 13-cis-retinoic acid: effect on individual skin surface lipids and implications for their anatomic origin. J. Invest. Dermatol. 1984, 82, 74–78. [Google Scholar] [CrossRef]
- Landthaler, M.; Kummermehr, J.; Wagner, A.; Plewig, G. Inhibitory effects of 13-cis-retinoic acid on human sebaceous glands. Arch. Dermatol. Res. 1980, 269, 297–309. [Google Scholar] [CrossRef]
- Hoover, E.; Aslam, S.; Krishnamurthy, K. Physiology, sebaceous glands. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023.
- Fischer, H.; Fumicz, J.; Rossiter, H.; Napirei, M.; Buchberger, M.; Tschachler, E.; Eckhart, L. Holocrine secretion of sebum is a unique DNase2-dependent mode of programmed cell death. J. Invest. Dermatol. 2017, 137, 587–594. [Google Scholar] [CrossRef]
- Tarayrah-Ibraheim, L.; Maurice, E.C.; Hadary, G.; Ben-Hur, S.; Kolpakova, A.; Braun, T.; Peleg, Y.; Yacobi-Sharon, K.; Arama, E. DNase II mediates a parthanatos-like developmental cell death pathway in Drosophila primordial germ cells. Nat. Commun. 2021, 12, 2285. [Google Scholar] [CrossRef]
- Xiong, C.; Ling, H.; Hao, Q.; Zhou, X. Cuproptosis: p53-regulated metabolic cell death? Cell Death Differ. 2023, 30, 876–884. [Google Scholar] [CrossRef]
- Stambolsky, P.; Weisz, L.; Shats, I.; Klein, Y.; Goldfinger, N.; Oren, M.; Rotter, V. Regulation of AIF expression by p53. Cell Death Differ. 2006, 13, 2140–2149. [Google Scholar] [CrossRef]
- Melnik, B.C. The TRAIL to acne pathogenesis: let's focus on death pathways. Exp. Dermatol. 2017, 26, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Cordain, L.; Lindeberg, S.; Hurtado, M.; Hill, K.; Eaton, S.B.; Brand-Miller, J. Acne vulgaris: a disease of Western civilization. Arch. Dermatol. 2002, 138, 1584–1590. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.K.; Bhate, K. A global perspective on the epidemiology of acne. Br. J. Dermatol. 2015, 172 Suppl 1, 3–12. [Google Scholar] [CrossRef]
- Moradi Tuchayi, S.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis. CC. Acne vulgaris. Nat. Rev. Dis. Primers 2015, 1, 15029. [Google Scholar] [CrossRef]
- Dréno, B.; Bettoli, V.; Araviiskaia, E.; Sanchez Viera, M.; Bouloc, A. The influence of exposome on acne. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 812–819. [Google Scholar] [CrossRef]
- Melnik, B.C. Diet in acne: further evidence for the role of nutrient signalling in acne pathogenesis. Acta Derm. Venereol. 2012, 92, 228–231. [Google Scholar] [CrossRef]
- Melnik, B.C. Linking diet to acne metabolomics, inflammation, and comedogenesis: an update. Clin. Cosmet. Investig. Dermatol. 2015, 8, 371–388. [Google Scholar] [CrossRef] [PubMed]
- Peck, G.L.; Olsen, T.G.; Yoder, F.W.; Strauss, J.S.; Downing, D.T.; Pandya, M.; Butkus, D.; Arnaud-Battandier, J. Prolonged remissions of cystic and conglobate acne with 13-cis-retinoic acid. N. Engl. J. Med. 1979, 300, 329–333. [Google Scholar] [CrossRef]
- Niu, G.; Wright, K.L.; Ma, Y.; Wright, G.M.; Huang, M.; Irby, R.; Briggs, J.; Karras, J.; Cress, W.D.; Pardoll, D.; et al. Role of Stat3 in regulating p53 expression and function. Mol. Cell Biol. 2005, 25, 7432–7440. [Google Scholar] [CrossRef]
- Melnik, B.C.; John, S.M.; Weiskirchen, R.; Schmitz, G. The endocrine and epigenetic impact of persistent cowmilk consumption on prostate carcinogenesis. J. Transl. Genet. Genom. 2022, 6, 1–45. [Google Scholar]
- Melnik, B.C.; John, S.M.; Carrera-Bastos, P.; Cordain, L.; Leitzmann, C.; Weiskirchen, R.; Schmitz, G. The role of cow's milk consumption in breast cancer initiation and progression. Curr. Nutr. Rep. 2023, 12, 122–140. [Google Scholar] [CrossRef]
- Melnik, B.C. Lifetime impact of cow's milk on overactivation of mTORC1: From fetal to childhood overgrowth, acne, diabetes, cancers, and neurodegeneration. Biomolecules 2021, 11, 404. [Google Scholar] [CrossRef]
- Sutcliffe, S.; Giovannucci, E.; Isaacs, W.B.; Willett, W.C.; Platz, E.A. Acne and risk of prostate cancer. Int. J. Cancer 2007, 121, 2688–2692. [Google Scholar] [CrossRef]
- Ugge, H.; Udumyan, R.; Carlsson, J.; Andrén, O.; Montgomery, S.; Davidsson, S.; Fall, K. Acne in late adolescence and risk of prostate cancer. Int. J. Cancer 2018, 142, 1580–1585. [Google Scholar] [CrossRef]
- Murphy, J.D.; Sandler, D.; White, A.J.; O'Brien, K.M. Severe acne and risk of breast cancer. Breast Cancer Res. Treat. 2019, 177, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Peck, G.L.; Gross, E.G.; Butkus, D.; DiGiovanna, J.J. Chemoprevention of basal cell carcinoma with isotretinoin. J. Am. Acad. Dermatol. 1982, 6, 815–823. [Google Scholar] [CrossRef]
- Bollag, W.; Holdener, E.E. Retinoids in cancer prevention and therapy. Ann. Oncol. 1992, 3, 513–526. [Google Scholar] [CrossRef]
- Bettoli, V.; Zauli, S.; Virgili, A. Retinoids in the chemoprevention of non-melanoma skin cancers: why, when and how. J. Dermatolog. Treat. 2013, 24, 235–237. [Google Scholar] [CrossRef]
- Veal, G.J.; Tweddle, D.A.; Visser, J.; Errington, J.; Buck, H.; Marange, J.; Moss, J.; Joseph, S.; Mulla, H. Pharmacokinetics and safety of a novel oral liquid formulation of 13-cis retinoic acid in children with neuroblastoma: A randomized crossover clinical trial. Cancers (Basel) 2021, 13, 1868. [Google Scholar] [CrossRef] [PubMed]
- Pile, H.D.; Sadiq, N.M. Isotretinoin. 2023 May 1. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023 Jan–. 1 May. [PubMed]
Figure 1.
Disturbed transcriptional regulation in acne vulgaris. Growth factors and androgens overstimulate the kinase AKT (protein kinase B) in acne vulgaris. Increased puberty-mediated insulin-like growth factor 1 (IGF-1) signaling increases the activity of phosphoinositide 3-kinase (PI3K), which phosphorylates and activates AKT. Increased IGF-1/insulin signaling by Western diet (hyperglycemic carbohydrates, milk and dairy products) as well as fibroblast growth factor receptor 2 (FGFR2) gain-of-function mutations (Apert syndrome, Munro acne nevus) further augment the activation of AKT. IGF-1 stimulates adrenal and gonadal androgen biosynthesis and activates 5α-reductase (5α-R) converting testosterone (T) to dihydrotestosterone (DHT), the high affinity ligand of androgen receptor (AR). Androgens activate mechanistic target of rapamycin complex 2 (mTORC2), which also phosphorylates and activates AKT. Thus, IGF-1 and androgens maintain a synergistic crosstalk resulting in the activation of AKT. AKT-mediated phosphorylation of AR mediates its nuclear transfer. In contrast, AKT-mediated phosphorylation of the transcription factors forkhead box O1 (FoxO1) and forkhead box O3 (FoxO3) promotes their export from the nucleus into the cytoplasm reducing their nuclear activity. FoxO1 is a suppressive nuclear coregulator of AR, peroxisome proliferator-activated receptor gamma γ (PPARγ), sterol regulatory element-binding transcription factor 1 (SREBF1) and signal transducer and activator of transcription 3 (STAT3) but promotes the expression of GATA-binding protein 6 (GATA6), the key regulatory transcription factor of infundibular keratinocytes. Thus, loss of nuclear FoxO1 activity enhances the expression of lipogenic genes (activated by AR, PPARγ, SREBF1) and proinflammatory genes (activated by STAT3) and attenuates the expression of GATA6. AKT-mediated phosphorylation of mouse-double minute 2 (MDM2) enhances the degradation of the transcription factor p53 resulting in reduced p53-mediated expression of FoxO1, FoxO3 and other p53 target genes. AKT-mediated phosphorylation of tuberin (TSC2) reduces its negative impact on Ras homolog protein enriched in brain (RHEB), the key activator of mechanistic target of rapamycin complex 1 (mTORC1). Activated mTORC1 stimulates protein translation of the transcription factors PPARγ, SREBF1, STAT3 and hypoxia-inducible factor 1α (HIF-1α). STAT3 is a negative regulator of TP53, whereas HIF-1α stimulates the expression of leptin (LEP) and interleukin 17A (IL17A) and Th17 cell differentiation.
Figure 1.
Disturbed transcriptional regulation in acne vulgaris. Growth factors and androgens overstimulate the kinase AKT (protein kinase B) in acne vulgaris. Increased puberty-mediated insulin-like growth factor 1 (IGF-1) signaling increases the activity of phosphoinositide 3-kinase (PI3K), which phosphorylates and activates AKT. Increased IGF-1/insulin signaling by Western diet (hyperglycemic carbohydrates, milk and dairy products) as well as fibroblast growth factor receptor 2 (FGFR2) gain-of-function mutations (Apert syndrome, Munro acne nevus) further augment the activation of AKT. IGF-1 stimulates adrenal and gonadal androgen biosynthesis and activates 5α-reductase (5α-R) converting testosterone (T) to dihydrotestosterone (DHT), the high affinity ligand of androgen receptor (AR). Androgens activate mechanistic target of rapamycin complex 2 (mTORC2), which also phosphorylates and activates AKT. Thus, IGF-1 and androgens maintain a synergistic crosstalk resulting in the activation of AKT. AKT-mediated phosphorylation of AR mediates its nuclear transfer. In contrast, AKT-mediated phosphorylation of the transcription factors forkhead box O1 (FoxO1) and forkhead box O3 (FoxO3) promotes their export from the nucleus into the cytoplasm reducing their nuclear activity. FoxO1 is a suppressive nuclear coregulator of AR, peroxisome proliferator-activated receptor gamma γ (PPARγ), sterol regulatory element-binding transcription factor 1 (SREBF1) and signal transducer and activator of transcription 3 (STAT3) but promotes the expression of GATA-binding protein 6 (GATA6), the key regulatory transcription factor of infundibular keratinocytes. Thus, loss of nuclear FoxO1 activity enhances the expression of lipogenic genes (activated by AR, PPARγ, SREBF1) and proinflammatory genes (activated by STAT3) and attenuates the expression of GATA6. AKT-mediated phosphorylation of mouse-double minute 2 (MDM2) enhances the degradation of the transcription factor p53 resulting in reduced p53-mediated expression of FoxO1, FoxO3 and other p53 target genes. AKT-mediated phosphorylation of tuberin (TSC2) reduces its negative impact on Ras homolog protein enriched in brain (RHEB), the key activator of mechanistic target of rapamycin complex 1 (mTORC1). Activated mTORC1 stimulates protein translation of the transcription factors PPARγ, SREBF1, STAT3 and hypoxia-inducible factor 1α (HIF-1α). STAT3 is a negative regulator of TP53, whereas HIF-1α stimulates the expression of leptin (LEP) and interleukin 17A (IL17A) and Th17 cell differentiation.
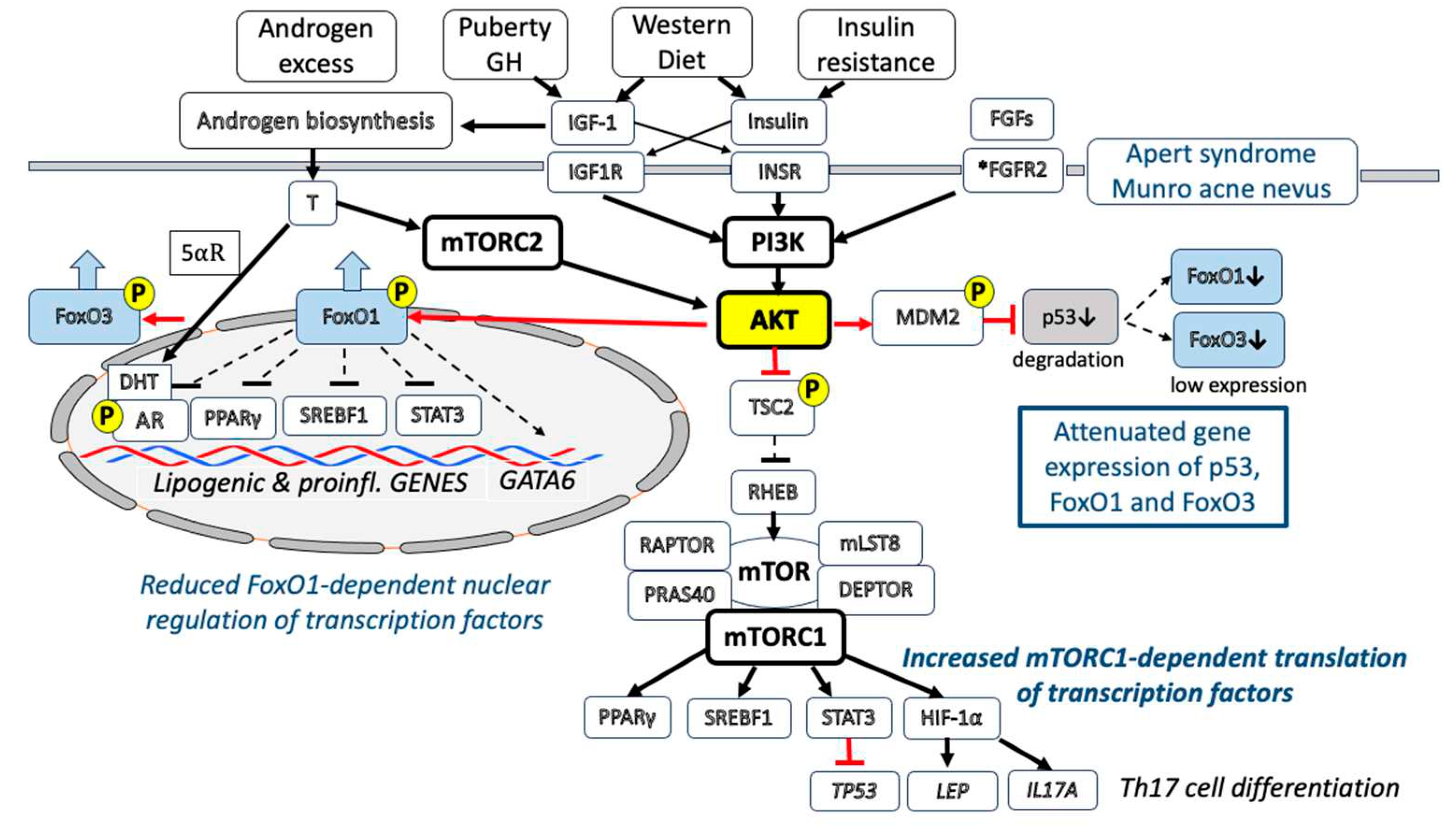
Figure 2.
Isotretinoin-induced upregulation of p53. After cellular uptake of isotretinoin, 13-cis retinoic acid is isomerized to all-trans retinoic acid (ATRA). The further transport of ATRA depends on the cellular abundance of cellular retinoic acid binding protein 2 (CRABP2), which in contrast to fatty acid binding protein 5 (FABP5) transfers ATRA to retinoic acid receptors (RAR) promoting the expression p53. p53 upregulates target genes promoting cell cycle arrest (CDKN1A), autophagy (ATG7) and apoptosis (FOXO1A, FOXO3A, CASP1, TNFSF10) but suppresses genes involved in growth factor signaling (IGF1, IGF1R), androgen signaling (AR), cell survival (BIRC5) and lipid biosynthesis (SREBF1), respectively.
Figure 2.
Isotretinoin-induced upregulation of p53. After cellular uptake of isotretinoin, 13-cis retinoic acid is isomerized to all-trans retinoic acid (ATRA). The further transport of ATRA depends on the cellular abundance of cellular retinoic acid binding protein 2 (CRABP2), which in contrast to fatty acid binding protein 5 (FABP5) transfers ATRA to retinoic acid receptors (RAR) promoting the expression p53. p53 upregulates target genes promoting cell cycle arrest (CDKN1A), autophagy (ATG7) and apoptosis (FOXO1A, FOXO3A, CASP1, TNFSF10) but suppresses genes involved in growth factor signaling (IGF1, IGF1R), androgen signaling (AR), cell survival (BIRC5) and lipid biosynthesis (SREBF1), respectively.
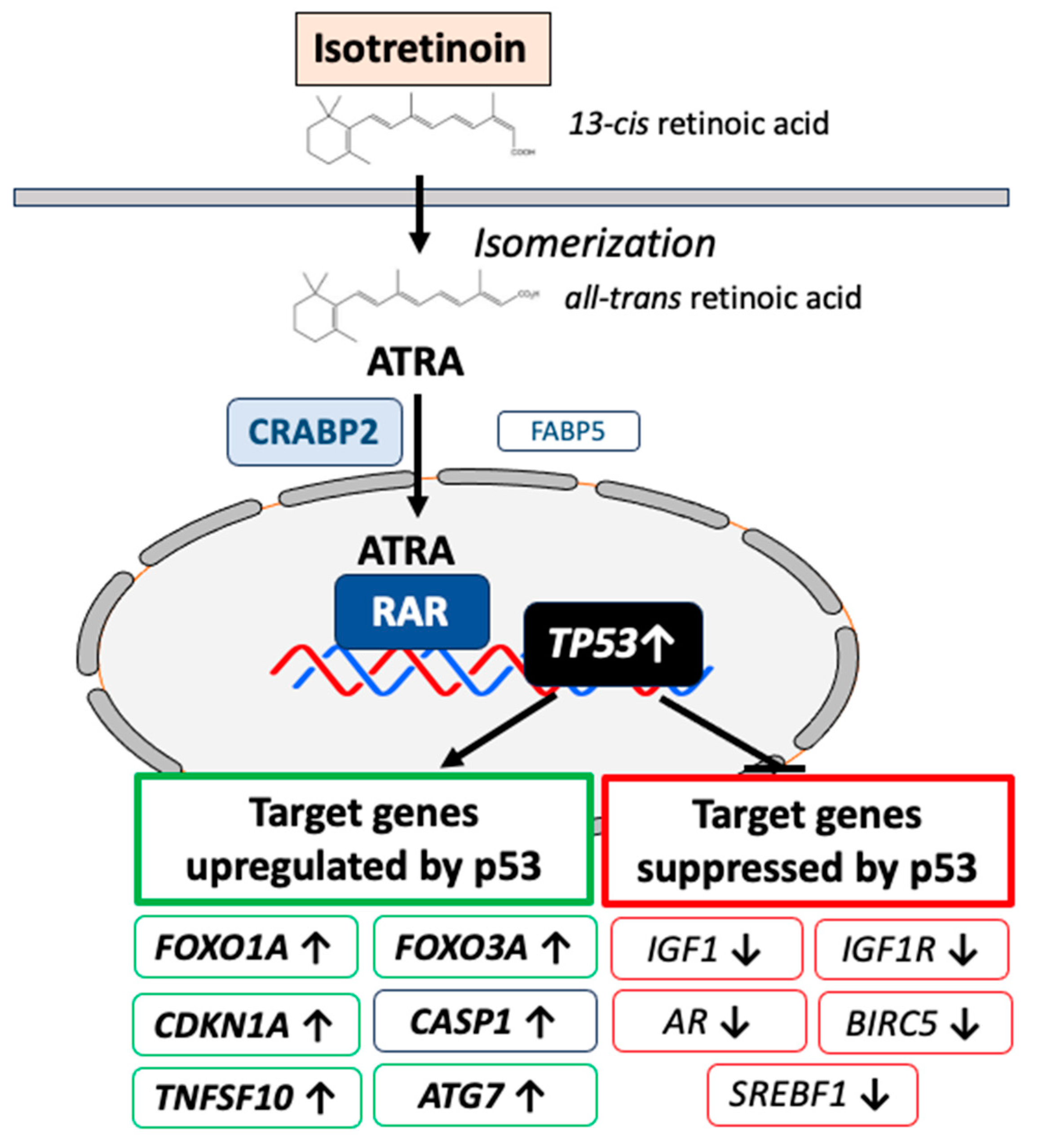
Figure 3.
Isotretinoin-induced overexpression of p53 counteracts dysregulated transcriptomics of acne vulgaris. Reduced gene expression of insulin-like growth factor 1 (IGF1) and insulin-like growth factor 1 receptor (IGF1R) reduces the activation of phosphoinositide 3-kinase (PI3K), which is further suppressed by upregulated expression of phosphatase and tensin homolog (PTEN). Reduced IGF-1/IGF1R signaling also attenuates androgen biosynthesis resulting in reduced mTORC2-mediated activation of AKT. Increased expression of tuberin (TSC2) suppresses Ras homolog protein enriched in brain (RHEB), thereby reduces the activity of mTORC1. mTORC1 and mTORC2 are further inhibited by induced expression of DEP-domain containing mTOR-interacting protein (DEPTOR), a natural inhibitor of both mTORC1 and mTORC2. Reduced mTORC1 results in impaired protein translation of peroxisome proliferator activated receptor γ (PPARγ), sterol regulatory element-binding transcription factor 1 (SREBF1), signal transducer and activator of transcription 3 (STAT3) and hypoxia-inducible factor 1α (HIF-1α). Forced nuclear expression of FoxO1 inhibits the transcriptional activity of androgen receptor (AR), PPARγ, SREBF1, STAT3 but induces the expression of GATA6. Thus, isotretinoin suppresses overactive growth factor and androgen signaling in acne patients but enhances GATA6 signaling, the key transcription factor controlling infundibular homeostasis. Forced overexpression of p53, FoxO1 and FoxO3 augments proapoptotic signaling explaining isotretinoin´s desired pharmacological mode of action (sebum suppression via sebocyte apoptosis) but also its major adverse effects, especially its teratogenicity (neural crest cell apoptosis).
Figure 3.
Isotretinoin-induced overexpression of p53 counteracts dysregulated transcriptomics of acne vulgaris. Reduced gene expression of insulin-like growth factor 1 (IGF1) and insulin-like growth factor 1 receptor (IGF1R) reduces the activation of phosphoinositide 3-kinase (PI3K), which is further suppressed by upregulated expression of phosphatase and tensin homolog (PTEN). Reduced IGF-1/IGF1R signaling also attenuates androgen biosynthesis resulting in reduced mTORC2-mediated activation of AKT. Increased expression of tuberin (TSC2) suppresses Ras homolog protein enriched in brain (RHEB), thereby reduces the activity of mTORC1. mTORC1 and mTORC2 are further inhibited by induced expression of DEP-domain containing mTOR-interacting protein (DEPTOR), a natural inhibitor of both mTORC1 and mTORC2. Reduced mTORC1 results in impaired protein translation of peroxisome proliferator activated receptor γ (PPARγ), sterol regulatory element-binding transcription factor 1 (SREBF1), signal transducer and activator of transcription 3 (STAT3) and hypoxia-inducible factor 1α (HIF-1α). Forced nuclear expression of FoxO1 inhibits the transcriptional activity of androgen receptor (AR), PPARγ, SREBF1, STAT3 but induces the expression of GATA6. Thus, isotretinoin suppresses overactive growth factor and androgen signaling in acne patients but enhances GATA6 signaling, the key transcription factor controlling infundibular homeostasis. Forced overexpression of p53, FoxO1 and FoxO3 augments proapoptotic signaling explaining isotretinoin´s desired pharmacological mode of action (sebum suppression via sebocyte apoptosis) but also its major adverse effects, especially its teratogenicity (neural crest cell apoptosis).
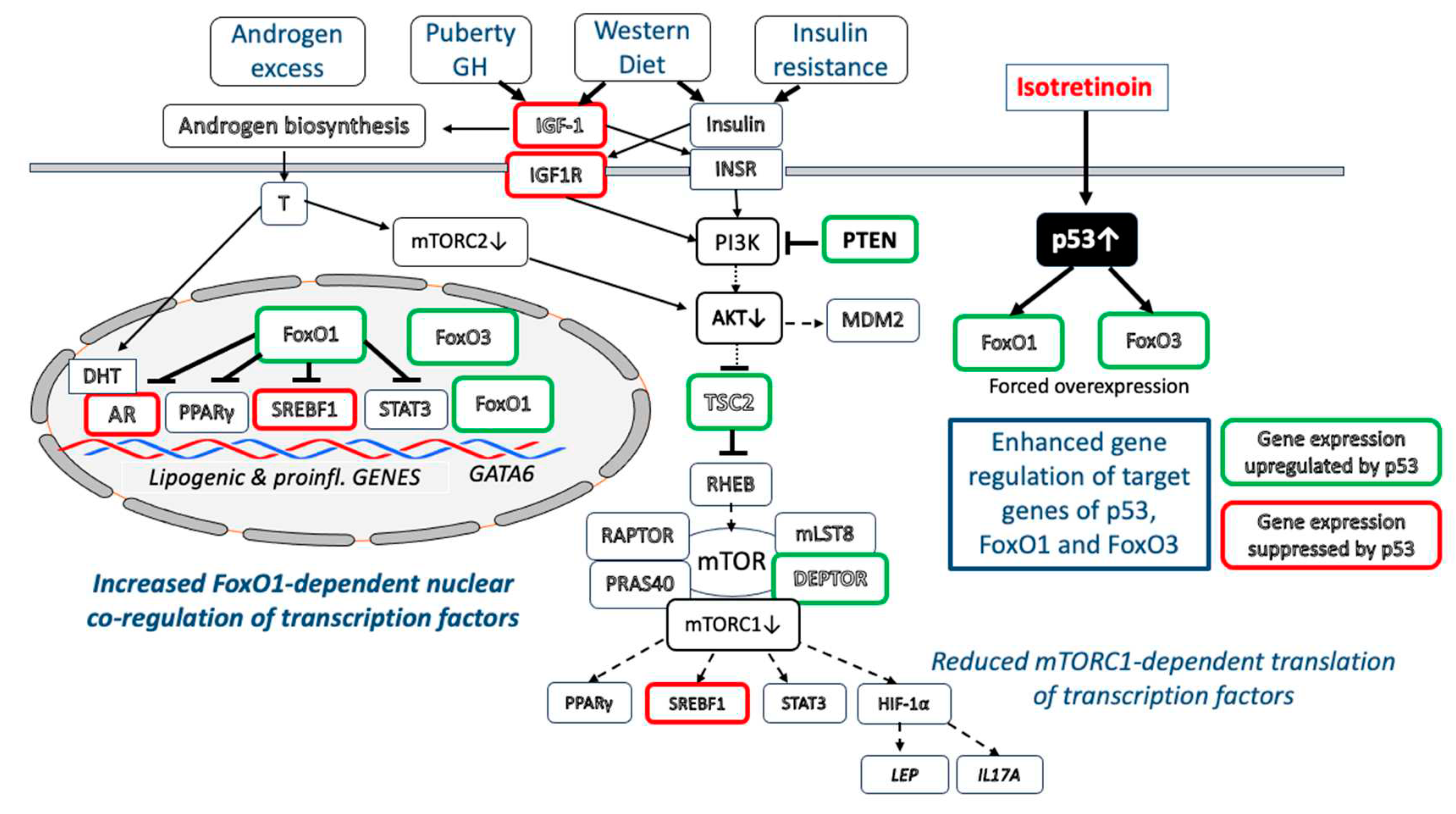

Table 1.
p53 target genes, whose expression is either down- or up-regulated by p53.
| Gene | Gene name | References |
|---|---|---|
| AR | Androgen receptor | [80] |
| IGF1 | Insulin-like growth factor 1 | [159] |
| IGF1R | Insulin-like growth factor I receptor | [161] |
| BIRC5 | Baculoviral IAP repeat-containing protein 5 | [178] |
| SREBF1 | Sterol regulatory element-binding transcription factor 1 | [309] |
| CDKN1A | Cyclin-dependent kinase inhibitor 1A (p21) | [117] |
| FOXO1A | Forkhead box O1A | [145] |
| FOXO3A | Forkhead box O3A | [146] |
| PTEN | Phosphatase and tensin homog | [163] |
| PRKAB1 | Protein kinase, AMP-activated, noncatalytic, -1 | [163] |
| TSC2 | TSC complex subunit 2 | [163] |
| DEPDC6 | DEP domain-containg protein 6 (DEPTOR) | [165] |
| TNFSF10 | Tumor necrosis factor ligand superfamily, member 10 | [175] |
| ATG7 | Autophagy-related 7 | [191] |
| APOB | Apolipoprotein B | [266] |
| AQP3 | Aquaporin 3 | [277] |
| AQP1 | Aquaporin 1 | [285] |
| CASP1 | Caspase 1 | [294] |
| ALOX5 | Arachidonate 5-lipoxigenate | [318] |
| AIFM1 | Apoptosis-inducing factor, mitochondria associated | [325] |
Table 2.
Genes, whose expression is either down-regulated or up-regulated by FoxO1.
| Gene | Gene name | References |
|---|---|---|
| AR | Androgen receptor | [23] |
| SREBF1 | Sterol regulatory element-binding transcription factor 1 | [24] |
| PPARG | Peroxisome proliferator-activated receptor gamma | [25] |
| STAT3 | Signal transducer and activator of transcription | [26] |
| FSHB | FSH -polypeptide downregulation | [248] |
| LHB | β-subunit of LH downregulation | [249] |
| GATA6 | GATA-binding protein 6 | [112,114] |
| FASLG | Fas ligand | [237] |
| MTTP | Microsomal triglyceride transfer protein | [267] |
| APOC3 | Apolipoprotein C-III | [268,269] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



