
Submitted:
06 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
Lactic acid bacteria are traditionally applied in a variety of fermented food products and have the capability to produce a wide range of bioactive ingredients during fermentation including vitamins, bacteriocins, bioactive peptides and bioactive compounds. The bioactivity and health benefits associated with these ingredients have gained interest in applications in the functional dairy market and have relevance both as components produced in situ and as functional additives. This review provides a brief description of the regulations regarding the functional food market in the European Union as well as an overview of some of the functional dairy products currently available in the Irish and European markets. A better understanding of the production of these ingredients excreted by lactic acid bacteria can further drive the development and innovation of the continuously growing functional food market.
Keywords:
Lactic acid bacteria
; functional food
; dairy products
; health benefits
; vitamins
; bacteriocins
; bioactive peptides
; bioactive compounds.
1. Introduction
Lactic acid bacteria (LAB) are a diverse group of microorganisms that have an essential role in the food and pharmaceutical industries [1]. The bacteria in this group can generally be characterised as gram-positive, non-spore-forming with a cocci or rod shape with their main metabolic product being lactic acid. Organisms belonging to either of the genera Lactobacillus, Leuconostoc, Pediococcus and Streptococcus are considered LAB, but it has been suggested that also Aerococcus, Alloiococcus, Carnobacterium, Dolosigranulum, Enterococcus, Globicatella, Lactococcus, Oenicoccus, Tetragenococcus, Vagococcus and Weissella should be included [2,3].
Historically LAB has been cultured and applied in the transformation of raw food material into fermented products such as yoghurt, cheese, bread and a wide range of fermented meat and vegetables (Figure 1) [4]. The fermentation process aids in the conservation of food material, enhances the texture and flavour profiles of products and increases the health benefits that can be derived upon consumption [2]. Their traditional applications in food production have given LAB GRAS (Generally Regarded as Safe) status [5].
This review explores the diverse range of functional ingredients produced by lactic acid bacteria including vitamins, bacteriocins, bioactive peptides and bioactive compounds. Exopolysaccharides are another metabolite excreted by LAB and its production, purification and health benefits have previously been reviewed extensively, and will therefore not be discussed in this review [6]. Additionally, lactic acid itself is a viable food ingredient produced by LAB and has been discussed at length in several publications [7,8,9] and is therefore not the target of this review.
1.1. Food product regulations
The European Union (EU) has a common framework of regulations for food, that can be intricate and thorough to navigate. The objective of this framework is to guarantee that a level of food safety standards are met and that the marketing of products is not misleading or confusing for consumers with unsubstantiated claims, thereby safeguarding consumer interests [10]. The European Food Safety Authority (EFSA) provides this framework of scientific advice and risk assessments, and aids in the decision making. EFSA furthermore evaluates the scientific claims made by companies and determines the validity and substantiation of suggested health claims [11].
Introducing innovative products with novel food ingredients and health claims on the European market the products need to comply with The Nutrition and Health Claims Regulations (NHCR) (Regulation EC No. 1924/2006) [12]. To receive approval for sale in the EU, the product must therefore adhere to the regulations set forth by the (EFSA). Specifically, the following points must be met [13]:
- The food/constituent is defined and characterised.
- The claimed effect is based on a nutrient essentiality OR the claimed effect is defined and a beneficial physiological effect can be measured in vivo in humans.
- The food/constituent is required for bodily function OR a cause-and-effect relationship between consumption and human health has been established.
- The quantity of the food/constituent can be consumed as part of a balanced diet to obtain the claimed effect.
Additional guidance for the determination of benefits to human health can be found in documents describing the following health claims related to [14,15]:
- Functions of the nervous system
- Physical performance.
- Bone, joints, skin and oral health.
- Appetite ratings, weight management and blood glucose concentrations.
- The immune system, the gastrointestinal tract and defence against pathogenic organisms.
This extensive framework aims to promote trust in the functional food market among consumers but can also be a challenge for food innovation in the food industry. The high standard of the NHCR regulations can be discouraging for producers of functional products, with “wording of claims” and “missing transparency being reported by the industry as the major challenges [10].
Health claims stating that microorganisms act as probiotics have not yet been authorised by the European Commission, however, some member states do allow the use of the term “probiotic effect” [14].
1.2. Current functional dairy products on the European market
The growth of the functional food market worldwide has been forecasted to continue its increase throughout the next decade with an expected global market increase from 177.4 USD billion in 2021 to 219.5 USD billion in 2026 [6]. With the increased focus on health and nutrition after the COVID-19 pandemic, consumers are actively seeking out products with added health benefits such as dairy products that can aid in the support of nutritional requirements or generally improved health [16]. Additionally, the dairy market is dynamic and undergoing constant development and innovation. The total amount of dairy products sold in Europe is steadily increasing according to MarketLine and will reach 155 billion euros in 2023 (Figure 2). This, in combination with the growing interest in functionality in foods among customers, renders dairy products an optimal choice for the enhancement of fortification.
The majority of food products on the European market with health claims deal with the addition of vitamins, minerals, protein and/or fibre [17]. Regarding microorganisms like LAB, NHCR regulations prevent the term probiotic from being applied as this claim has not yet been authorized by the European Commission, however, some member states do allow the use of the term “probiotic effect” [14].
Table 1 provides a summary of dairy products available within the Irish and European markets and includes kefir, yoghurt and drinking yoghurt, milk, butter and milk powders.
A current trend among consumers is the tailoring of functional products to specific needs and population groups, in which the fortified milk range by Avonmore (Tirlán) is a good example as it includes Avonmore Super Milk for improved nutrition and bone health, Avonmore Fibre Plus Milk for improved immune function and gut health and Avonmore Slimline Milk targeted population groups with higher iron requirements such as pregnant women (Table 1).
Milk powders with functionality are currently mostly aimed at infant and child nutrition but are starting to see their introduction in adult nutrition as well with the launch of Aerabo skim milk powders from Dairygold, which is being sold on the Chinese market. Like the Avonmore range of functional milk, the Dairygold line of functional milk powders also targets its application towards different groups: Aerabo Active Vitality targeted the middle-aged and senior population ([18], Aerabo Active Boost for improved immune support [19] and Aerabo Active Light for improved nutrition and wellbeing with a lower calorie content [20].

Table 1.
summary of dairy products available in the Irish and European markets and includes kefir, yoghurt and drinking yoghurt, milk, butter and milk powders.
Table 1.
summary of dairy products available in the Irish and European markets and includes kefir, yoghurt and drinking yoghurt, milk, butter and milk powders.
| Product | Product name | Functional ingredient | Health benefit | Company | Ref |
|---|---|---|---|---|---|
| Kefir | Kefir Smoothie | Probiotics Calcium |
Gut health | Müller | [21] |
| Kefir | Kefir Yoghurt | Probiotics | Gut health | Glenillen Farm | [22] |
| Kefir | Spoonable Kefir | Probiotics Calcium |
Gut health | Irish Yogurts, Clonakilty | [23] |
| Kefir | Kefir Drink Original | Probiotics Vitamin B12 and B2 Calcium |
Immune health Gut health |
Biotiful Gut Health | [24] |
| Kefir | Kefir Yoghurt Original | Probiotics Vitamin B12 and B2 Calcium |
Digestion Immune health Gut health |
Biotiful Gut Health | [25] |
| Spreads | Flora 100% natural ingredients | Vitamin A Omega 3 and 6 |
Nutrition | Flora | [26] |
| Spreads | Flora ProActiv | Plant sterols | Cholesterol-lowering | Flora | [27] |
| Spreads | Benecol© Spread | Plant sterols | Cholesterol-lowering | Raisio Lpc. | [28] |
| Milk | Avonmore Super Milk | Vitamins | Bone health Nutrition |
Tirlán | [29] |
| Milk | Avonmore Fibre Plus Milk | Vitamins Fibre |
Gut health Immune health Nutrition |
Tirlán | [30] |
| Milk | Avonmore Slimline Milk | Vitamins Iron |
Nutrition |
Tirlán | [31] |
| Milk powder | Nido© | Vitamins Minerals |
Nutrition | Nestlé | [32] |
| Milk powder | Baby&Me© Organic | Vitamins Minerals Oligosaccharides |
Nutrition | Arla | [33] |
| Milk powder | Aerabo Active Vitality | Amino acids Antioxidants Glucosamine Minerals Vitamins |
Bone health Nutrition |
Dairygold | [18] |
| Milk powder | Aerabo Active Boost | Amino acids Antioxidants Beta carotene Vitamins |
Immune support Nutrition |
Dairygold | [19] |
| Milk powder | Aerabo Active Light | Amino acids Antioxidants Beta carotene Minerals Vitamins |
Nutrition | Dairygold | [20] |
| Milk drink | Goede Morgen! | Vitamins | Nutrition | Friesland Campina | [34] |
| Yoghurt | Activia | Probiotics | Gut health | Danone | [35] |
| Yoghurt | Danonino | Minerals Vitamins |
Nutrition | Danone | [36] |
| Yoghurt | Benecol© Yoghurt | Plant sterols | Cholesterol-lowering | Raisio Lpc | [37] |
| Yoghurt | Natural Yoghurt | Probiotics | Gut health | Glenillen farm | [38] |
| Yoghurt | Greek style natural | Probiotics | Gut health | Irish Yoghurts, Clonakilty | [39] |
| Yoghurt drink |
Actimel | Probiotics Vitamins |
Immune health | Danone | [40] |
| Yoghurt drink | Benecol© Yoghurt Drink | Plant sterols | Cholesterol-lowering | Raisio Lpc. | [37] |
2. Vitamins
Vitamins are organic compounds that constitute an essential part of the human diet. Humans are for the most part unable to synthesise vitamins themselves and rely on supplementation through a balanced diet. Certain LAB have the capability to synthesize some of these essential vitamins that humans are incapable of producing including folate (vitamin B9), cobalamin (vitamin B12), riboflavin (vitamin B2), menaquinone (vitamin K2) and thiamine (vitamin B1) [41,42].
The ingestion of vitamins is widely known to be associated with a range of health benefits (Figure 3).
2.1. Folate
Folate is essential for a wide range of biological functions including amino acid metabolism and DNA repair and is essential for cell division [43].
LAB, Bifidobacteria and PAB are all capable of producing folate while also being food-grade microorganisms (Table 2). Folate yields are however generally reported to be very low, even in genetically engineered strains [44,45,46]. Folate is therefore primarily produced synthetically as folic acid, but the human dihydrofolate reductase enzyme has an extremely low rate of folic acid conversion into bioactive vitamins and a high concentration of this synthetic form is therefore needed [47].
Table 2.
Overview of folate-producing lactic acid bacteria.
| Microorganism(s) | Fermentation | Medium | Yield | Ref |
|---|---|---|---|---|
| L. acidophilus | Flask | MRS | 37.2 µg/l | [48] |
| L. amylovorus | Flask | MRS | 81.2 µg/l | [48] |
|
L. amylovorus S. thermophilus L. delbrueckii subsp. bulgaricus |
Flask | Milk | 263 µg/l | [48] |
| L. brevis | Flask | Supplemented whey permeate | 131 µg/l | [49] |
| L. casei | Flask | MRS | 1.5 µg/l | [48] |
| L. coryniformis | Flask | MRS | 81 µg/l | [50] |
| L. delbrueckii subsp. bulgaricus | Flask | MRS | 54 µg/l | [51] |
|
L. delbrueckii subsp. bulgaricus S. thermophilus |
Flask | Non-fat milk | 180 µg/l | [52] |
| L. fermentum | Flask | MRS | 6.9 µg/l | [48] |
| L. fermentum | Flask | Supplemented whey permeate | 84 µg/l | [49] |
| L. helveticus | Flask | MRS | 89 µg/l | [51] |
| L. lactis | Flask | MRS | 45 µg/l | [51] |
| L. lactis subsp. cremoris | 5 L batch bioreactor | Skim milk powder | 187 µg/l | [53] |
| L. lactis subsp. lactis | Flask | M17 | 291 µg/l | [51] |
| L. lactis subsp. lactis biovar diacetylactis | Flask | M17 | 100 µg/l | [51] |
| L. paracasei subsp. paracasei | Flask | MRS | 38.7 µg/l | [48] |
| L. plantarum | Flask | MRS | 57.2 µg/l | [48] |
| L. plantarum | Flask | Supplemented whey permeate | 397 µg/l | [49] |
| L. plantarum | Flask | MRS | 108 µg/l | [50] |
|
L. plantarum P. freudenreichii |
1 L bioreactor | Supplemented whey permeate | 8399 µg/l | [54] |
| L. reuteri | Flask | Supplemented whey permeate | 125 µg/l | [49] |
| L. sakei | Flask | MRS | 107 µg/l | [50] |
| S. thermophilus | Flask | M17 | 202 µg/l | [51] |
| S. thermophilus | 2 L batch bioreactor | Modified M17 | 54.53 µg/l | [55] |
2.1.1. Production of folate
Hugenschmidt et. al. (2010), screened 151 LAB strains and 100 PAB strains for their ability to produce folate in a supplemented whey permeate media. Strains belonging to Lactobacillus showed the highest extracellular folate production, with L. plantarum SM39 being the highest producer with 3.97 ng/ml [49]. This strain was subsequently used in a follow-up study examining the production of folate and cobalamin in co-culture with P. freudenreichii, in a supplemented whey media. The addition of para-aminobenzoic acid (pABA) to the fermentation media increased the folate yield more than 10-fold [54]. This stimulating effect of pABA has also been observed in other studies on the strains S. thermophilus and L. lactis, where the production yield of folate was dependent on pABA in the media. This can be explained by pABA being one of the precursors of folate production. The level of pABA needed for optimal folate production is not fully described, with one study reporting that production yields did not increase with a concentration of pABA above 1 µM [56] while another reported this number to be above 100 µM [51].
Mousavi et. al. (2013) tested the effect of different carbon sources (glucose, maltose, sucrose, lactose) and nitrogen sources (meat extract, yeast extract, peptone/casein, peptone/meat, (NH4)2 SO4- and NH4NO32-) was tested to find optimal cultivation conditions for folate production of S. thermophilus. Here, it was concluded that lactose and yeast extract were the optimal carbon source and nitrogen source [55].
Another study further supplemented media with the prebiotics mannitol and sorbitol increased the yield of folate due to the stimulatory effect of prebiotics on cell growth. Folate molecules are highly sensitive to oxidation, and the reducing agents sodium thioglycolate, sodium ascorbate and cysteine hydrochloride have been added with a positive effect on the final yield of folate produced by a strain of L. lactis [41].
Two studies carried out fermentation of S. thermophilus in bioreactors and observed a high increase in folate production when pH was controlled, but where one study observed 7.3-9.3 as the optimal pH range [51], another determined pH 7 to be the optimal pH [56].
The production of folate in situ has been studied as well. Yoghurt is an example of a product well known for its potential of being a source of folate, although the concentrations of folate are however highly variable [42,52]. By fermenting milk with high folate-producing strains of the known yoghurt-producing species S. thermophilus and L. delbrueckii subsp. bulgaricus it was possible to increase folate content by 125% compared to commercial yoghurts [52]. Currently, commercial yoghurts can contribute to 10-20% of daily levels of folate but with both optimum strain selection and growth factors this number has great potential for future increase [51].
2.1.2. Health benefits of folate
Folate is an essential nutritional component, that cannot be synthesised by mammalian cells, and thus it must be supplemented through diet. Some population groups exhibit a higher risk of folate deficiency which is especially prevalent in the elderly population due to lower food consumption but also in pregnant women as higher levels of folate are required to assist in neural tube development [57]. Obtaining the right amount of folate is essential as excessive intake can lead to the accumulation of folic acid in the bloodstream and mask a potential cobalamin deficiency, as symptoms are comparable [58].
Low levels of folate in the diet during pregnancy have been associated with neural tube defects which can lead to severe consequences for the developing foetus including anencephaly (absence of major parts of the brain), spina bifida (spinal cord not formed properly), encephalocele (sacks containing brain tissue protrudes through the skull) and stillbirth [59]. Due to this, most countries are recommending folate supplementation of at least 400µg/day for pregnant women [59,60].
Folate can lower the levels of homocysteine, an amino acid that has been associated with neurodegeneration and Alzheimer’s disease [61,62]. Large studies in the UK and Sweden have indeed found an association between folate levels and cognitive function and Alzheimer’s disease [61,63]. A correlation between folate deficiency and depression and schizophrenia has also been described [64]. This was investigated in a study where patients suffering from depression and schizophrenia were supplemented with 15 mg of methyl folate per day for 6 months in addition to standard psychotropic treatment and a significant improvement of clinal and social recovery was observed [65].
The ability of folate to lower homocysteine levels can furthermore reduce the risk of cardiovascular diseases and studies have shown an inverse relationship between folate and cardiovascular disease [66,67]. Oral administration of folate has shown the ability to reduce homocysteine levels by 25% in subjects that are not folate deficient [68].
Folate levels might also modulate the risk of cancer, as a higher intake of folate has been associated with a lowered risk of colon cancer [69]. Balancing the intake of folate is also a necessity as it has been demonstrated how a modest supplementation of folate can reduce carcinogenesis while an excessive amount might increase the growth of tumours [69].
2.2. Cobalamin
Cobalamin is an important co-factor in the metabolism of amino acids, carbohydrates, fatty acids and nucleic acids [42]
Chemical production of cobalamin is far too expensive to be commercially viable and involves more than different 70 steps, so industrial preparation takes place through bacterial fermentation [70]. In industrial fermentations, species of P. freudenreichii, P. denitrificans and B. megaterium are highly used. Of these, only P. freudenreichii has GRAS status and is therefore the only of these microorganisms that can currently be utilised directly in food production (Table 3) [45,71].
2.2.1. Production of cobalamin
The production of cobalamin from different strains of Propionibacterium in a whey-based fermentation medium has also been optimized in several studies [77,78,79,80]. The production of cobalamin by P. shermanii was optimised by investigating optimal levels of whey and yeast extract in the growth medium. It was determined that with a given level of whey, a certain amount of yeast extract was needed, with 10% whey solids and 1.5% yeast extract resulting in maximum yield [77].
The addition of the amino acids betaine and choline as well as diammonium hydrogen phosphate ((NH4)2HPO4) to a whey-based media showed an increase in cobalamin production. It was determined that the addition higher than 0.5% w/v of either of the amino acids did not increase cobalamin production further. In addition, the effect of betaine was observed to be more stimulating than choline [79,80]. This stimulating effect of betaine concurs with studies on cobalamin production of P. denitrificans [73,81] and is explained by betaine acting as a methyl group donor for the highly complicated cobalamin structure that contains up to eight methyl groups [73].
PAB are microaerophilic and produce cobalamin in high yield only when the concentration of oxygen is kept low [70]. One step in the cobalamin synthesis does however require oxygen, and the fermentation process producing cobalamin from P. freudenreichii is therefore often divided into two stages, as the optimal yield is dependent on both an anaerobic and aerobic phase [82]. A periodic fermentation with P. freudenreichii was developed with a fluctuation of oxygen supply and a cyclic operation switching between anaerobic and aerobic conditions. This operation mode resulted in a low concentration of propionic acid together with a high level of cobalamin [83].
One of the challenges of producing cobalamin from Propionibacterium is the accumulation of metabolites such as propionic acid and acetic acid which harbours growth inhibitory effects [84]. In a study by Miyano et al (2000) three different strategies were applied to circumvent this in P. freudenreichii fermentations: periodic changing of DO concentration between 0 and 1 ppm, a cell recycle system and mixed culture with R. eutropha that are capable of assimilating propionic acid. From these three methods, the cell recycle system gave the highest cobalamin productivity, but the mixed cell system resulted in the highest cobalamin produced per unit volume of medium [76].
2.2.2. Health risks associated with cobalamin deficiency
Cobalamin deficiency can occur in the elderly population, children, pregnant women and people on a plant-based diet that excluded natural cobalamin-high foods such as meat and dairy [85]. This deficiency is usually caused either by a lack of cobalamin in the diet or by malabsorption. It is estimated that 6% of the UK population under 60 and 20% of the population over 60 years exhibit a cobalamin deficiency. [85]. The deficiency of cobalamin typically appears with symptoms such as fatigue and anaemia, but can also result in bone marrow suppression and risk of cardiomyopathy in more serious cases of deficiency [85]. Patients with a cobalamin deficiency can be ordinated oral treatments of the vitamins, while fortification of foods to make them more functional could be the solution in countries with high numbers of deficiencies in the population [86].
In addition to low folate levels inducing the risk of neural tube development, there is also evidence that low cobalamin levels might induce the same risk [87,88]. This area of research is however still lacking in data on both need and dosage, but the benefits of cobalamin supplementation outweigh any potential consequences [89].
As mentioned, the levels of homocysteine appear to be related to cardiovascular disease. A study that found a reduction of homocysteine levels by supplementation with 5 mg folate, found a small, but further decrease by also supplementing the diet with 0.4 mg/day cobalamin with 7% [90]. Most of the research on cardiovascular disease is however done on folate with or without the addition of cobalamin, making the information on the specific effect of cobalamin limited [91].
2.3. Riboflavin
Traditionally riboflavin has been produced chemically, but the biotechnological production through strains such as B. subtilis, A.gossypii, C. famata and L. lactis has gained more interest, with only L. lactic belonging to the group of LAB (Table 4) [95]. Although some strains of bacteria and yeast are considered good producers of riboflavin, the ascomycete fungi A. gossypii is considered the best as it can produce 40000 times more riboflavin than it needs for its growth [96].
2.3.1. Production of riboflavin
L. acidophilus and L. lactis have been applied in studies examining their riboflavin production [95,97]. L. lactis and L. acidophilus were grown on both milk and whey. Whey appeared to be a more optimal growth medium for riboflavin production, and L. acidophilus was the optimal producer of riboflavin with a final yield of 2.93 mg/L [103]. Although this yield is significantly lower than those that are possible to obtain with A. gossypii the interest in producing riboflavin from LAB is due to the potential of producing fermented foods with high levels of riboflavin produced in situ [104]. Mohedano et al (2019) investigated Five strains of L. plantarum as a probiotic by examining their ability to produce riboflavin as well as their capability to survive under digestive tract stresses. One strain of L plantarum 3.33 mg/L survived well under gastric stress conditions, alluding to its potential as a probiotic strain for functional foods [100].
High yields of 5.72 mg/L riboflavin have been obtained for another strain of L. plantarum by optimising its growth medium. Glucose and sucrose were evaluated as carbon sources at both 30°C and 37°C, and the highest yields of riboflavin were observed when growing in a sucrose-based medium at 30°C [102].
2.3.2. Health benefits of riboflavin
A balanced diet will in most cases supply the necessary amount of riboflavin in healthy humans. Nevertheless, different population groups can be at risk of insufficient riboflavin supply such as the elderly, children and pregnant women [105].
Riboflavin has been shown to exert an antioxidant effect in two ways: by prevention of lipid peroxidation and by the attenuation of reperfusion of oxidative injury [106]. The research conducted on this antioxidant activity in humans is however limited [107]. This antioxidant activity is attributed to the role riboflavin plays in the activity of several antioxidant enzymes such as superoxide dismutase, catalase and glutathione peroxidase [106].
Some studies have also shown that intake of adequate amounts of riboflavin could lead to a decreased risk of breast cancer [108], lung cancer [109], colorectal cancer [110], gastric cancer [111] and ovarian cancer [112], emphasizing its importance in a healthy lifestyle.
It has been hypothesized that riboflavin can have a neuroprotective effect on diseases such as Parkinson’s disease, migraine and multiple sclerosis [107]. A low level of riboflavin has been reported in patients with Parkinson’s disease. By adding a riboflavin supplement to the diet and eliminating red meat, it was possible to increase motor functions [113]. This same effect has however not been observed in other studies [114,115].
2.4. Menaquinone
There are two available forms of vitamin K: phylloquinone produced by plants and menaquinone primarily produced by bacteria [70]. It is an essential cofactor involved in the post-transitional carboxylation found in proteins related to blood clotting, cardiovascular disease and bone health [42]. B. subtilis is the most well-studied strain for menaquinone production, but some LAB is also capable of synthesising menaquinone (Table 5) [121].
2.4.1. Production of menaquinone
The potential of synthesis of menaquinone by LAB has been researched in a study that screened 21 strains. Of the screened species of LAB, five strains were selected due to their high production of menaquinone: three strains of L. lactis subsp. Cremoris, one strain of L. lactis subsp. lactis and one strain of L. lactis. All five of these strains were able to produce a meaningful amount of menaquinone when grown in non-fat dry milk [123].
P. freudenreichii has been investigated both for its ability to produce menaquinone and its precursor 1,4-Dihydroxy2-Naphthoic Acid (DHNA). When grown in a skim milk-based media in a 3L bioreactor, relatively high yields of menaquinone of 0.12 mM were obtained [124].
2.4.2. The role of menaquinone in cardiovascular disease and bone health
Menaquinone has long been recognised for its role in blood coagulation but has shown a potential to improve bone health and inhibit the growth of cancer [125,126].
Studies have shown that the intake of specifically vitamin K in the form of menaquinone is associated with reduced coronary calcification and a reduced risk of cardiovascular disease [127,128,129]. The effects of vitamin K in the form of phylloquinone and menaquinone over 10 years in 4807 Dutch men and women over the age of 55 were investigated [127]. Interestingly, this study found that the intake of menaquinone but not phylloquinone reduced the risk of coronary heart disease and coronary calcification [127]. This comparison of phylloquinone to menaquinone was also done in another study that found a similar effect of only menaquinone reducing the risk of coronary calcification [130].
Menaquinone can play an important role in bone health as three different menaquinone-dependent proteins have been isolated from bone [128]. The effect of menaquinone on bone mineral density in human intervention studies is inconclusive, with some showing a beneficial effect [131,132] while another study has concluded no beneficial effect [133]. One major limitation of the study by Emaus et al (2010) is however the short follow-up period, of 1 year with another study stating that effects might only become significant after 2 years of intervention [132]. In addition to improving bone mineral density, menaquinone has also shown indications of improving bone strength in a human intervention study [132].
3. Bacteriocins
Bacteriocins are antimicrobial peptides produced by certain bacteria that can kill or inhibit the growth of both gram-negative and gram-positive bacteria (Table 6) [134]. Several applications of bacteriocins have been reported in the literature including, control of microflora in fermentation products [135] and extension of shelf life [136]. Bacteriocins are proving worthy of commercial interest, due to their preservation qualities that can aid in fulfilling consumer demand for foods that are safe and long-lasting without the use of chemical preservatives [137,138].
Currently, the only commercially available bacteriocins are nisin produced by L. lactis and pediocin PA-1 produced by Pediococcus species [139]. These are both approved for safe use in food by the Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA). In addition to nisin and pediocin, several other bacteriocins are highly studied, but not yet approved for safe use in foods including lactococcins, enterocins and aureocins [140].
Table 6.
Overview of nisin and pediocin produced by LAB.
| Microorganism(s) | Product | Fermentation | Medium | Yield | Ref |
|---|---|---|---|---|---|
| L. lactis | Nisin | Flask | Skim milk-based media | 75 IU/ml | [141] |
| L. lactis | Nisin | Shake flask | Whey based media | 92.9 mg/L | [142] |
| L. lactis | Nisin | Shake flask | Whey based media | 1167 AU/ml | [143] |
| L. lactis | Nisin | Shake flask | Whey based media | 2618.7 IU/ml | [144] |
| L. lactis | Nisin | Single batch – free cells | Whey based media | 32800 & 41000 BU/ml (16400 & 20500 IU/ml) |
[145] |
| L. lactis | Nisin | Single batch – free cells | MRS | 160 AU/ml | [146] |
| L. lactis | Nisin | Repeated cycle batch - ICT | Whey based media | 20480 IU/ml | [147] |
| L. lactis | Nisin | Fed-batch | Complex | 4185 IU/ml | [148] |
| L. lactis | Nisin | Fed-batch | Whey | 60.3 BU/ml | [149] |
| L. lactis | Nisin | Fed-batch | Whey + glucose | 124 BU/ml | [150] |
| L. lactis | Nisin | Fed-batch | Whey + MRS nutrients | 258.47 BU/ml | [151] |
| L. lactis | Nisin | Fed-batch | Defined | 2594 · 106 IU/ml | [152] |
|
L. delbrueckii subsp. bulgaricus S. thermophilus |
Nisin | Continuous fermentation | Skim milk-based media | 4500 BU/ml | [153] |
|
L. lactis K. marxianus |
Nisin | Single batch – free cells | Defined | 98 mg/L (3920 IU) | [154] |
|
L. lactis S. cerevisiae |
Nisin | Shake flask | Defined | 150.3 mg/L | [155] |
|
L. lactis P. acidilactici |
Nisin Pediocin |
Shake flask | Whey based media | 74 BU/ml 195 BU/ml |
[156] |
|
L. lactis P. acidilactici |
Nisin Pediocin |
Shake flask | Whey based media | 9 BU/ml 45 BU/ml |
[157] |
|
L. lactis P. acidilactici |
Nisin Pediocin |
Shake flask | MRS Whey based media |
50 and 22.9 BU/ml 493.2 and 57.9 BU/ml |
[158] |
|
L. lactis P. acidilactici |
Nisin Pediocin |
Single batch – free cells | Whey based media | 3000 AU/ml (18h) 1359 AU/ml (16h) |
[159] |
| P. acidilactici | Pediocin | Flask | Whey based media | 12800 AU/ml | [160] |
| P. acidilactici | Pediocin | Flask | MRS | 12800 AU/ml | [161] |
| P. acidilactici | Pediocin | Flask | Whey based media | 150000 AU/ml | [162] |
| P. acidilactici | Pediocin | Shake flask | Whey based media | 189 BU/ml | [163] |
| P. acidilactici | Pediocin | Shake flask | MRS Whey based media |
493.2 BU/ml 167.3 BU/ml |
[164] |
| P. acidilactici | Pediocin | Single batch – free cells | TGE broth | 40000 AU/ml | [165] |
| P. acidilactici | Pediocin | Repeated cycle batch - ICT | MRS Whey based media |
4096 AU/ml ( 0.75h) 4096 AU/ml (2h) |
[166] |
| P. acidilactici | Pediocin | Fed-batch | Defined | 712 BU/ml | [163] |
| P. acidilactici | Pediocin | Fed-batch | Whey based media | 517 BU/ml | [164] |
|
P. acidilactici S. thermophilus L. delbrueckii subsp. bulgaricus |
Pediocin | Flask | Skim milk-based media | 6400 AU/ml | [161] |
|
P. acidilactici P. pentosaceus |
Pediocin | Flask | Whey based media | 3220 AU/ml 26100 AU/ml |
[167] |
|
P. pentosaceus L. plantarum |
Pediocin | Flask | Whey based media | 51200 AU/ml | [168] |
3.1. Production of nisin
Nisin is commercially used in over 48 countries and has FDA and EFSA approval [139]. It was first isolated from milk in 1928 and gained approval as a safe food additive in 1969 by the Joint Food and Agriculture Organization/World Health Organization (FAO/WHO) [169]. Nisin is usually manufactured via fermentation of milk or whey using L. lactis. The fermentation broth is subsequently harvested, concentrated, separated and spray-dried [134]. To reach the optimal nisin production more complex media including MRS are usually required together with control over parameters such as pH and temperature [148].
Several studies have compared the nisin production yields from L. lactis grown on whey-based feedstocks with MRS media [149,158]. A selection of studies has reported that the production yields using MRS medium can reach yields of 50 and 55 AU/ml [149,158], whereas typical yields of 22 and 22,5 AU/ml are achieved using diluted whey medium
[149,158]. Malvido et. al (2019) supplemented a base whey media with MRS nutrients. Nutrients were supplemented in concentrations corresponding to MRS medium of 25%, 50%, 75%, 100% and 125% w/v. The addition of MRS nutrients increased the nisin yield from 22.67 bacteriocin units (BU) for diluted whey to 57.26 BU for whey supplemented with 100 v/w of MRS nutrients. There was an observed increase in yield with MRS nutrient addition, however only up until 100% where the yield stagnated [151].
It has been reported that to further optimise the production of nisin, it is necessary to provide additional protein in the form of e.g. peptone, tryptone or yeast extract [148].
To optimise nisin production, a fed-batch fermentation on a whey media with a feed composition of concentrated glucose (400g/L), concentrated whey and 4% yeast extract, and a feeding volume addition corresponding to the amount needed to restore initial total sugar concentration was studied. Nisin production was increased to 50.6 and 60.3 BU/ml when compared to yield obtained in batch cultures grown in whey of 22.5 BU/ml [158] and 50 BU/ml when compared to growth batch culture growth in MRS [149].
The production efficiency and costs of production in fed-batch fermentation of nisin were compared in supplemented and unsupplemented whey. They had a feed consisting of concentrated whey and concentrated glucose and a feed profile adding volumes to the fermentation to restore the initial total sugar composition. The cost of nisin in a fed-batch fermentation supplemented with MRS nutrients was 30% lower than the cost of nisin in fed-batch fermentation with unsupplemented media. Here, the highest yield obtained on a whey-based fed-batch fermentation with MRS nutrients was 258 BU/ml compared to 124.66 BU/ml in a whey-based fed-batch with no added nutrients [151].
3.2. Production of pediocin
Pediocin is a bacteriocin with a broad spectrum, meaning that it is capable of inhibiting several different species of gram-positive bacteria [170]. It is however particularly known for its ability to inhibit the growth of L. monocytogenes and S. aureus which are both common food pathogens [171]. Both P. acidilactici and P. pentosaceus are capable of pediocin production (Table 6) and are well-known from food fermentations, and are either naturally present or added as a starter culture in the fermentation of vegetables and sausages [170,172].
The influence of the different salts ammonium phosphate, calcium chloride, potassium dihydrogen phosphate and manganese(II)sulphate monohydrate on pediocin production was tested in a direct plate bioassay for rapid assessment. Manganese is an essential growth factor for lactic acid bacteria, and the addition of MgSO4 also resulted in a significantly increased pediocin yield. Each of ammonium phosphate, calcium chloride and potassium dihydrogen phosphate resulted in a suppression of pediocin production [146]. A whey and yeast extract-based media could therefore possibly be further optimized by the addition of manganese. The growth and pediocin production of pediococcus has also been tested in both a repeated batch and a fed-batch setup with whey and yeast extract as the batch media, with reports of increased yield [173,174,175].
Pediococcus species more readily metabolise glucose than other carbon sources [146,159]. Excessive glucose in the fermentation broth can however have an inhibitory effect, and its addition in a simple batch resulted in decreased pediocin, suggesting substrate inhibition. The addition of glucose in the feed of a fed-batch fermentation instead resulted in great improvements in the pediocin yields, showing at least a 2-fold increase of 517 BU/ml in a fed-batch compared to 167 BU/ml in a simple batch in one study [174]. Pediocin has furthermore been determined to be a pH-dependent metabolite, and that pH should be maintained at levels lower than 5 to achieve maximum production [157,174,176]. This is due to a requirement of low pH for the post-translational processing of pre-pediocin to active pediocin [165].
3.3. Mixed culture induction
The presence of other bacterial strains can enhance bacteriocin production either by the production or consumption of metabolites or by acting as a stress signal which in some cases increases bacteriocin production. Pediocin production by P. pentosaceus was increased by 250% in the presence of L. plantarum [168], while another study observed that the co-culturing of P. acidilactici with S. thermophilus and L. delbrueckii enhanced pediocin production in fermented milk [161].
S. cerevisiae improved nisin production in a study by Shimizu et. al. (1999) from L. lactis by 85% to a final yield of 150.3 mg/L. This was attributed to the lactic- and acetic acid assimilation by S. cerevisiae which normally acts as a limiting factor for nisin production [155]. A similar enhancement of 70% was observed when L. lactis was co-cultured with another yeast, K. marxianus, where it was also concluded that the increased nisin production was caused by lactic acid assimilation and the resulting control of pH levels. [154].
3.4. Health benefits of bacteriocins
Bacteriocins produced by LAB possess several health benefits. Their antimicrobial activity can be applied in the treatment of harmful bacteria, as they can selectively target pathogenic organisms, while not affecting commensal bacteria. This makes them a valuable alternative to antibiotics that are less target-specific [177].
The antimicrobial nature of bacteriocins makes them Additionally, research is suggesting that bacteriocins can have applications in the treatment of cancer as well as immunomodulatory effects (Figure 3).
3.4.1. Treatment of pathogens and alternatives to antibiotics
The anti-listerial effect of both purified pediocin and its producer strain P. acidilactici was investigated in vivo in mice infected with L. monocytogenes. The effect of oral administration of P. acidilactici did not result in a decrease of L. monocytogenes in the intestine, liver or spleen, while the administration of purified pediocin resulted in a 2-log decrease of L. monocytogenes. None of the treatments altered the composition of the gut microbiota, making purified pediocin a promising agent against L. monocytogenes [178]. In this study, however, administration of P. acidilactici was only given as a single dose and purified pediocin was administered over 3 days. Another study administered both a pediocin and non-pediocin strain of P. acidilactici and a nisin-producing strain of L. lactis for 16 days in mice infected with vancomycin-resistant enterococci (VRE). Results showed, that on day 6 none of the infected mice administered either pediocin-producing P. acidilactici or nisin-producing L. lactis had detectable levels of VRE [179].
S. aureus is one of the most common pathogens in the upper respiratory tract and nisin has been assessed for its ability to combat S. aureus infections in vivo in immunocompromised rats. Rats were provided nisin intranasally, and growth of S. aureus was inhibited [180]. These results suggest that nisin and pediocin can indeed be used to fight pathogens and could have importance against pathogens with resistance to antibiotics. Another advantage of bacteriocins over antibiotics is their specific activity that does not affect the commensal bacteria in the gut, as antibiotics do [181]. A study investigated the sensitivity of 21 common intestinal bacteria to pediocin and two types of nisin in vitro. Both types of nisin inhibition of all the gram-positive bacteria and only one of the gram-negative bacteria. Pediocin on the other hand did not inhibit any of the 21 intestinal bacteria assayed, which further strengthens its potential as a non-invasive alternative to pathogenic infections [181].
3.4.2. Anti-cancer treatment
There is research that suggests the cytotoxicity of bacteriocins against cancer cells [182].
Nisin has been reported to have cytotoxic and antitumour effects against head and neck squamous cell carcinoma in vivo in mice [183]. Mice were fed 200 mg/kg of nisin for three weeks which resulted in a significant reduction of tumour size compared to control. These effects of nisin were also tested in vitro and were found to be due to induced apoptosis, cell cycle arrest and reduction in cell proliferation [183]. This effect of nisin on head and neck squamous cell carcinoma was also observed in vivo in another study in mice [184]. Here, it was also found that when getting dosages of nisin, the tumour size would reduce and would prolong survival.
Nisin has also shown possible application as an adjunct together with the chemotherapeutic agent doxorubicin in the treatment of skin cancer [185]. Carcinogenic mice were treated with either nisin, doxorubicin or a combination of the two. The tumour size was reduced by 14% with nisin, 51% with doxorubicin and 66.82% when mice were treated with both agents, as compared to untreated groups.
3.4.3. Immunomodulatory role of bacteriocins
Host-defence peptides are ubiquitous and play an important role in the innate immune system. Despite being smaller and having a different structure, bacteriocins share similar physiochemical properties to host-defence peptides, which could indicate that they might have similar immunomodulatory properties [188,189]. This immunomodulatory ability has been demonstrated in vivo in mice [190] and turbot [191]. By feeding mice nisin in the commercial form Nisaplin an increase in the T-lymphocytes CD4 and CD8 and a decrease in B-lymphocyte levels was observed. Long-term administration resulted in a return to normal levels of B- and T-lymphocytes and an increase in the macrophage/monocyte population [190].
An in vitro study found that nisin could induce the stimulation of the chemokines monocyte chemoattractant protein- 1 (MCP-1), Gro-α and IL-8 while significantly reducing TNF-α in response to bacterial lipopolysaccharide in human peripheral blood mononuclear cells [189].
Lastly, a study has found that nisin was able to activate neutrophils and suggested that nisin might be capable of influencing multiple subsets of host immune cells [192].
The research conducted on nisin’s ability to alter the host immune response is limited, so further studies on the area are needed.
4. Bioactive peptides
Proteins that occur naturally in food can in addition to providing nutritional benefits, also contain sequences called bioactive peptides that can exert health various health benefits (Figure 3) [193]. When present in the parental protein these sequences are inactive but can be released in 3 ways: through hydrolysis by gastrointestinal enzymes, through the action of proteolytic enzymes derived from plants or microbes or by microbial fermentation with microbes exerting proteolytic activity [194]. The focus in this section will solely be on the bioactive peptides released by microbial fermentation. During the fermentation of milk, LAB is capable of hydrolysing milk proteins to make nitrogen sources like peptides and amino acids available [195].
4.1. Angiotensin-converting enzyme inhibitory peptides
Several members of the species lactobacillus, bifidobacterium and pediococcus are described in the literature as angiotensin-converting enzyme inhibitory peptides (ACE inhibitory peptides) producers [196,197,198], Table 7. Functional dairy products containing ACE-inhibitory peptides such as fermented milk [199,200,201,202] and cheese [203,204] have been developed. These products are characterized by a high degree of proteolysis, whereas products with a lower proteolytic activity such as yoghurt, fresh cheese and quark have a lower ACE-inhibitory activity [205].
Un-fermented whey already contains a certain amount of ACE-inhibitory peptides, but by fermenting it with the microbiota naturally present in cheese whey, the ACE-inhibitory activity increased from 22% in unfermented whey to 60-70% in fermented whey [210]. Another study by Ahn et. al. (2009) showed that a further supplementation of glucose and yeast extract to whey led to an increase in the biomass, thereby increasing the proteolytic activity and in turn a higher yield of peptides [197]. Here, L. brevis, L. helveticus and L. paracasei were used for peptide production, with the peptides derived from whey fermentation of L. helveticus showing high inhibitory activity with IC50 values of 5,3 and 7,8 µg/ml.
P. acidilactici is another species that has also been shown to exhibit good ACE inhibitory activity and was found by screening 34 different strains of LAB grown in a whey-based media [196]. P. acidilactici had an ACE-inhibitory activity of 84.7% with an IC50 of 19.78 µg/ml.
Sodium caseinate was used to produce a fermentate with ACE inhibitory activity by L. animalis [211]. The fermentate had an 85.51% ACE-inhibitory activity with an IC50 value of 8 µg/ml, which is only a slightly higher dose needed than the commercially available captopril that demonstrates inhibitory activity at a dose of 5 µg/ml [211]. Raveschot et. al. ( 2020) compared the production of ACE inhibitory peptide productivity of L. helveticus in 3 different fermentation setups: a simple batch, a continuous bioreactor and a continuous membrane bioreactor (MBR). The mean productivity of the MBR setup was 0,27 g/L/h which was comparable to that of the simple batch of 0,33 g/L/h but higher than the continuous bioreactor which had a mean productivity of 0,103 g/L/h. The specific productivity was also calculated as a function of bacterial biomass, where the MBR setup was superior to the other setups with a rate of 15.8 g/g compared to 9.99 g/g and 7.13 g/g in batch and continuous batch respectively. In this study, a fermentate with an IC50 value of 0.47 mg/L for L. helveticus was obtained [212].
4.1.1. Antihypertensive effect and cardiovascular diseases
Drugs that inhibit ACE are common for the treatment of hypertension. ACE is needed for the conversion of angiotensin 1 to angiotensin 2 which narrows the blood vessels and can cause higher blood pressure [213].
The blood-pressure-lowering effects of ACE inhibitory peptides have been demonstrated in several studies with subjects suffering from hypertension [214,215,216]. Ingestion of tablets of milk fermented with L. helveticus was administered for 4 weeks in subjects with either high-normal blood pressure or mild hypertension. For both groups, a lowered blood pressure was observed for subjects ingesting the tablets compared to a placebo group. The placebo tablets had a similar composition to the test-group tablets, but had no ACE inhibitory peptides, indicating the blood pressure-lowering effect was due to the presence of the peptides [214].
Another proposed effect of milk fermented with L. helveticus is the alleviation of arterial stiffness. Long-term administration of the milk-containing peptides showed a significant reduction in arterial stiffness compared to the placebo group [217]. This study also investigated the improvement of endothelial function but found no effects from the peptide-containing milk. Another study did however observe an improved vascular endothelial function when administering peptides to hypertensive subjects [218]. This mechanism was determined to be related to the enhanced production of vasodilating substances.
4.2. Peptides with antioxidant activity
Some LAB possess the ability to excrete peptides with antioxidant activity (Table 8). These peptides can decrease the risk of accumulation of reactive oxygen species (ROS) as well as degrading superoxide anions and hydrogen peroxide [219,220].
Virtanen et. al. (2007) screened 25 LAB strains for their ability to exhibit antioxidant activity in milk media [223]. While all strains had antioxidant activity to some degree, 6 strains had a higher inhibition rate: 2 Lc. cremoris, L. lactis, L. jensenii, L. acidophilus and L. helveticus. A correlation between the degree of hydrolysis and radical scavenging activity was found for all strains, with an additional correlation of bacterial growth to radical scavenging activity found for L. acidophilus and L. helveticus. Lipid peroxidation inhibitory activity was found to be more related to bacterial growth, than to proteolysis. When two or three of the strains were cultivated together, this increased antioxidant activity [223]. This mixed-strain approach to a higher antioxidant activity was used in the other two studies either co-cultivating yoghurt bacteria with lactobacilli [221].
The traditional yoghurt fermenting strains S. thermophilus and L. delbrueckii subsp. bulgaricus was used to produce yoghurt and test for its potential antioxidant activity. This study concluded that yoghurt can indeed have the potential as a natural antioxidant [220].
Another study also used the yoghurt strains but supplemented them with additions of L. casei, L. paracasei and/or L. acidophilus to enhance the antioxidant activity of yoghurt. The yoghurt fermenting strains alone demonstrated good antioxidant activity but the addition of the lactobacillus strains resulted in a lowering of the IC50 value. The highest antioxidant activity observed was when all 3 strains of lactobacillus were added [221]. and in another study by Lin and Yen (1999),
L. casei was used in a study to investigate the influence of several nutrients on the production of antioxidant peptides in goat milk fermentation [222]. Here, it was found that the addition of casein peptone, glucose and calcium lactate had significant positive effects on the antioxidant activity. The effect of glucose and calcium lactate are most likely caused by stimulation of L. casei growth. Casein peptone could increase the release of peptide, which has also been observed in another study that has shown for caseinate [224].
4.2.1. Health benefits of antioxidant peptides produced by LAB
Some of the antioxidant potential in fermented dairy products is considered to be caused by the high release of antioxidant peptides by the proteolytic systems in many LAB [225]. In vivo studies in either animal or human models demonstrating the specific effect of antioxidant peptides are scarce [225]. One study fed ageing rats with either unfermented milk or milk fermented with L. fermentum and observed that after 2 months, the numbers of the antioxidant enzymes superoxide dismutase, catalase and glutathione peroxidase were higher in the liver cells of rats fed the fermented milk compared to the unfermented milk [226]. The influence of fermented and unfermented milk on a weanling rat model was also compared in another study [227]. Here, the ability to reduce lipid peroxidative stress was evaluated, but no difference between the two milk types was observed. There was still observed an antiperoxidative effect when compared to the control, indicating the milk protein rather than the lactobacillus to be responsible for this effect. Both of the above-mentioned results were also demonstrated in a third study on rats fed either fermented or unfermented whey. Here, an observed increase in antioxidant enzymes and antiperoxidative activity was observed for both fermented and unfermented whey, although this study did observe peroxidative changes to be more pronounced in fermented whey [228].
5. Bioactive compounds
Some species of LAB have gained attention for their ability to produce the bioactive compounds gamma-aminobutyric acid (GABA) and carotenoids [229,230]. GABA is a neurotransmitter with known health benefits such as its ability to calm the nervous system while carotenoids are pigments with known antioxidant activity (Figure 3) [231,232].
5.1. Gamma-aminobutyric acid
γ-aminobutyric acid (GABA) is a bioactive amine that has been shown to confer health benefits. It acts as a neurotransmitter in the nervous system and can lower blood pressure in mildly hypertensive patients [233] and has also been suggested to have anti-tumour effects [234]. It is naturally present in several foods such as tomatoes, teas, soybeans and fermented foods and can therefore be obtained naturally through diet, but much higher concentrations are obtainable through LAB fermentations (Table 9) [235].
5.1.1. Production of GABA
The pH value during the fermentation plays an important role both in the cell growth of LAB and in their production of GABA. The synthesis of GABA is dependent on the enzyme glutamic acid decarboxylase (GAD) which acts as a catalysator, and pH is therefore an important factor when considering yields of GABA [264,267]. The importance of maintaining a pH value of between 4.5-5 has been reported in several studies that have documented optimal GABA synthesis when pH was kept at this level [236,237,238,248,251,264]. A pH value of 6 did result in higher biomass in a study cultivating L. brevis, but even with the higher biomass, the yield of GABA was still higher at pH 5 [238].
GABA production itself is not affected by lower temperatures of around 30°C but can lead to lower yields of biomass which in turn leads to less total yield of GABA [236,238]. Higher temperatures above 40°C did support growth but were inhibitory to GABA production [236,237,238,253]. The optimal temperature for both cell growth and GABA synthesis is therefore within the range of 35-40°C.
Due to the different optima for cell growth and GABA synthesis, a two-stage fermentation was developed for L. brevis. Here, cultures were maintained at 35°C and a pH value of 5 for the first 32 hours to stimulate cell growth and was subsequently adjusted to 40°C and a pH of 4.5. This resulted in an increase of GABA from 398 mmol/L in the one-stage fermentation to 474.70 mmol/L in the two-stage fermentation [239].
Another factor that can increase GAD activity is the addition of pyridoxal-5’-phosphate (PLP), which acts as a coenzyme. Studies have reported an increase in the GABA yield of L. paracasei from 200mM without PLP to 300 mM with PLP [251] and in S. thermophilus fermentation from 6 g/L without PLP to 8 g/L with PLP addition after 24 hours [264]. Other studies did however not observe any change in yield in L. brevis fermentations, but it was suggested to be due to sufficient synthesis in the fermentation broth [238,253].
Monosodium glutamate (MSG) is a stimulant of GABA production, but if it is present at too high levels, it has been shown to have inhibitory effects [240,264]. In L. plantarum cultivations, GABA synthesis was significantly influenced by the addition of 100 mM of MSG in the fermentation broth, as it induced an increase in production by 7.7 to a value of 721.35 mM [268]. This stimulatory effect has similarly been observed for L. brevis within a range of 2.25% and 4-6% v/w [237,254], for S. thermophilus within a range of 10-20 g/L [264] and L. lactis and L. plantarum at 1% v/w [253].
A strategy to overcome the inhibitory effects of high MSG concentrations is to supplement it to the feed in fed-batch fermentations [239,238,239]. In a study by Peng et. al. (2013) an initial concentration of 100mmol/L MSG was added to a defined medium consisting of glucose, yeast extract, peptone, sodium acetate and ions. The feed consisted of 106 mmol of MSG and was added at 32h and 56h. By the end of fermentation, the final yield of GABA was 526.33 mmol/L. The yield was only slightly higher than without the feed (474.79 mmol/L) but due to the additional increase in volume, a much higher total GABA productivity was obtained [239].
To a fermentation broth consisting of glucose, peptone, tween80 and MnSO4, 400mM of glutamate was added. It was found that concentrations exceeding 500mM strongly inhibited cell growth. The feed was added at 12h and 24h and consisted of 280.8g and 224.56g of glutamate. This feeding strategy was detrimental to the cell growth, but highly effective for a high yield of GABA in a short fermentation time, as it yielded 1095 mM [238].
A very high yield of 205.8 g/L GABA by L. brevis has also been found in a medium with 295 g/L of L-glutamic acid [241]. The issue with L-glutamic acid or glutamate is however the high production cost, making it less feasible for a cost-effective production [254].
Co-culturing microbes is another efficient approach to increase the yield of GABA and has been successfully applied in many studies. Fermentation of milk with a combination of a strain with high proteolytic activity together with a GABA-producing strain has led to efficient GABA production together with additional reports of a lowered fermentation time [247,261,266]. The proteolytic strains are capable of breaking down milk proteins into peptides which can then be used as substrates by the GABA-producing strain.
The addition of a GABA-producing LAB strain to fermentation with S. thermophilus and L. delbrueckii sups. bulgaricus enhanced the level of GABA in fermented milk with MSG in two studies [257,266].
Co-culturing of LAB and fungi has been reported with combinations of L. plantarum and Ceriporia lacerata leading to the production of 15.53 g/L while the combination of Lactobacillus futsaii and Candida rogusa lead to a GABA productivity of 135 mg/L/h [269,270].
Whey could be a suitable and low-cost option for GABA production by LAB and has been investigated as a substrate [245,261]. A GABA yield by cultivation of L. brevis of 553 mg/L by utilizing a media consisting of 14.95% of whey and 4.95% of MSG [245]. A higher GABA yield of 3.65 g/L was obtained in whey-fermentations of a co-culture of L. plantarum and L. lactis subsp lactis. In this study, soy protein hydrolysate was used as a source of glutamic acid and when added to the whey the yield increased from 1 g/L to 3.65 g/L [261]. Both of these studies utilize very simple media for GABA production on whey and do not use pH control, the addition of MSG, PLP, and tween80. It is possible that these additions and a potential fed-batch fermentation setup could further increase the yield of GABA.
5.1.2. Health benefits of GABA
GABA have a wide variety of different health benefits including hypotensive effect, it can act as a neurotransmitter and can aid diabetes, cancer and asthma [271].
The hypotensive effect of GABA is similar to that of ACE-inhibitory peptides, by inhibiting noradrenalin release which inhibits perivascular nerve stimulation and thereby facilitates a hypotensive effect [272]. Spontaneously hypertensive rats (SHR) and normotensive Wistar-Kyoto rats (WKY) were given GABA to investigate its blood pressure-lowering capabilities. This was given as a single dose of 0.5mg/kg and showed a significant lowering of the systolic blood pressure in SHR but not in WKY rats [272]. This was confirmed in the same two rat models in a later study dosing GABA either pure or in an enriched fermented milk product [273]. Here, a long-term dosage was also investigated and a significantly slower increase in blood pressure of both rat models was observed for those administered either GABA or GABA-enriched fermented milk, compared to controls [273]. Milk fermented with strain L. casei shirota and L. lactis product containing GABA was developed and tested in mildly hypertensive patients [233]. The blood pressure was significantly decreased in patients after 2 to 4 weeks and remained decreased through all 12 weeks [233].
Disruption in the GABA receptor expression has been associated with anxiety spectrum disorders and has also been shown to have a role in mood disorders like depression [274]. Administration of L. rhamnosus to mice modulated the GABA receptors, and mice showed a behaviour that was less anxious and reduced stress-related, and depressive behaviour compared to the control group [275]. This stress-reducing effect of GABA has also been demonstrated in humans through drinking GABA-enhanced water [276] and by ingestion of GABA-enriched chocolate [277].
By consuming a powdered preparation of GABA, an improvement in sleep was observed by reduced sleep latency as well as an increase in non-rapid eye movement sleep in healthy adults [278]. For a group of patients suffering from chronic fatigue, the ingestion of a GABA-enhanced drink before solving an arithmetic task resulted in reduced occupational fatigue afterwards [279]
A study has suggested GABA as a tool to prevent obesity induced by a high-fat diet. It was demonstrated that GABA improved oxidative stress and thyroid function and the treatment with GABA on high-fat diet mouse models prevented weight gain [280].
GABA also has potential as a therapeutic against diabetes and has shown an inducing effect on the secretion of insulin [281] and can even reverse established diabetes in a diabetic mouse model [282]. Based on the strong in vivo data of GABA on diabetes, the administration of a food product with GABA present serves as a promising tool in the treatment and prevention of diabetes.
Lastly, GABA could be used as an anti-cancer agent by having tumour-suppressing activity in lung adenocarcinoma cells [283] and inhibiting cell proliferation in colon cancer cells [284]. Mouse and human leukaemia cells as well as human cervical cancer cells were treated with germinated rice enhanced with GABA. Here, GABA had an inhibitory action on leukaemia cell proliferation and also showed a stimulatory action on cancer cell apoptosis [285].
5.2. Carotenoids
Carotenoids are bioactive molecules that are naturally occurring in most fruits and vegetables. They can be produced by a wide range of organisms such as plants, algae, fungi, yeast and bacteria. Humans, however, are unable to synthesise carotenoids and depend on uptake through the diet [230]. Intake of carotenoids is associated with many health benefits, primarily by exerting antioxidant activity [286]. Several carotenoids are produced industrially and can be used both as a source for fortification of foods and as colourants [287]. The majority of studies on the biotechnological production of carotenoids are from yeasts, with fewer reports of synthesis through bacterial production [288,289].
5.2.1. Production of carotenoids
The research conducted on the production of carotenoids by LAB is sparse as yeasts are preferred due to higher yields, however, some research has been conducted investigating this trait in LAB (Table 10). 158 different strains of LAB were screened for their ability to produce carotenoids and out of these only 36 produced the pigment. Specifically, strains belonging to L. plantarum, L. fermentum and P. acidilactici had this ability. All these strains did however produce carotenoids in low amounts, with a strain of L. fermentum yielding the highest amount of 765 µg/kg CDW [290].
Garrido-Fernández et. al. (2010) cultivated L plantarum in four different brands of MRS as well as two defined media and were able to obtain 54.55 mg/kg of cell dry weight in a defined media. The effect of length of cultivation was tested by sampling at 24H, 48H, 72H and 96H, but a decrease of carotenoid concentration in the broth was observed over time, and 24H was selected as the optimum cultivation time [291]. L. plantarum was also examined in another study that used a Plackett-Burman approach to media optimisation using date syrup as a carbon source which had a final maximum yield of 54.89 mg/kg of cell dry weight [292].
It has been found that the cultivation of LAB under aerobic conditions is essential for optimal carotenoid production as it increases the expression of genes in carotenoid biosynthesis [293].
A research group has investigated the co-culturing of LAB with the yeast R. rubra [294,295,296,297]. These studies however used R. rubra as the main carotenoid producer, while the LAB are added to the culture to create an optimal growth environment for R. rubra.
Table 10.
Summary of carotenoid-producing strains and yields.
| Microorganism(s) | Fermentation | Medium | Yield | Ref |
|---|---|---|---|---|
| L. fermentum | Flask | MRS | 765 µg/kg CDW | [290] |
| L. plantarum | Flask | Supplemented MRS | 54.89 mg/kg CDW | [292] |
| L. plantarum | Flask | Defined medium | 54.55 mg/kg CDW | [290] |
|
L. casei R.rubra |
Flask | Supplemented whey ultrafiltrate | 12.35 mg/ml | [294] |
|
L. delbrueckii. subsp. bulgaricus S. thermophilus R. rubra |
Flask | Supplemented whey ultrafiltrate | 13.09 mg/ml | [295] |
|
L. delbrueckii. subsp. bulgaricus S. thermophilus R. rubra |
Flask | Supplemented whey ultrafiltrate | 13.37 mg/L | [296] |
|
L. helveticus R. glutinis |
Batch | Supplemented whey ultrafiltrate | 8.3 mg/L | [297] |
5.2.2. Health benefits of carotenoids
Carotenoids play an important role in human nutrition as they can act as a precursor for the synthesis of vitamin A [298]. Although vitamin A is naturally present in many food products such as dairy, fish, orange and yellow vegetables and leafy greens, deficiencies of vitamin A are still a prominent issue in some developing countries [299].
Low-density lipoprotein (LDL) is linked to the pathogenesis of atherosclerosis. Healthy adults consumed a carotenoid supplement in various concentrations, and it was found that a daily concentration over 3.6 mg of a carotenoid led to an inhibition of LDL oxidation, thereby having potential as a parameter for the prevention of atherosclerosis [300]. Carotenoid supplementation has also shown an effect on the improvement of lipid profiles. Non-obese subjects with mild hyperlipidaemia were given a dose of either 0, 6, 12 or 18 mg of carotenoids per day for 12 weeks. BMI and LDL levels remained unaffected, but triglyceride levels decreased significantly with doses of 12 and 18 mg/day and levels of high-density lipoprotein (HDL) increased significantly in doses of 6 and 12 mg/day [301]. Long-term supplementation of a carotenoid did however not show to impose any effects on the development of cardiovascular disease or mortality associated with cardiovascular disease [302,303].
In vitro and in vivo studies have indicated that carotenoids stimulate bone formation and bone cell proliferation while also having an inhibitory effect on bone resorption [304,305,306,307]. In human trials, the intake of carotenoids over a 4 year period was also associated with a beneficial effect on bone health and results showed a protective role of carotenoids for bone mineral density [308].
Human trials have demonstrated a positive effect of dietary supplementation of carotenoids in weight management and obesity [309]. Canas et al (2017) could report a decrease in the BMI z-score, waist-to-height ratio and subcutaneous adipose tissue in children with mild obesity over a period of 6 months of supplementation of a mixture of carotenoids. These results were concluded to be directly correlated with an increased concentration of serum β-carotene [310]. Similarly, in another study, healthy overweight adults were given a capsule containing carotenoids daily for 12 weeks. They observed a significant reduction in the BMI and the abdominal fat area [311].
A long-term study over 18 years with 4052 healthy participants receiving 50mg of beta carotene on alternate days or a placebo showed promising effects on cognitive function. A significant difference was observed for the global cognitive score as well as verbal memory scores for the group receiving beta-carotene. In addition, it was tested if there was an effect of shorter-term administration of 1 year by recruiting new subjects to either a treatment or placebo group, but here no significant difference between the two groups was observable [312]. It has been suggested that the antioxidant effect of carotenoids could have a preventative effect and act as a treatment for Alzheimer’s Disease [313], but a study has found no effect of carotenoids on the decreased risk of Alzheimer’s Disease and dementia [314].
6. Conclusions
The introduction of novel functional products on the European Market can be a challenging process as claims related to health need to be scientifically substantiated and evaluated in a process that can be complicated for producers to navigate. This extensive framework of regulations does, however, simultaneously ensure trust from consumers in new food products.
LAB are an essential constituent of a wide range of fermented products which are already an integral part of the human diet. Here, they enhance the nutritional, organoleptic and functional properties of the products. Understanding the different bioactive ingredients such as vitamins, bacteriocins, bioactive peptides and bioactive compounds excreted during fermentation by LAB can aid in the targeted development of products with desired health claims. This includes the selection of appropriate strains, optimised media composition as well as selection of suitable bioprocess parameters.
Consumers are increasingly seeking products that offer additional health benefits to nutrition. Exploring the potential and understanding the production processes of the bioactive ingredients produced by LAB is therefore important as it can further drive forward the innovation and development of the functional food industry market.
Author Contributions
Conceptualization, H.M.S. and B.F.; writing—original draft preparation, H.M.S.; writing—review and editing, H.M.S., K.D.R., B.F., S.M., C.L., D.B. and G.M.; project administration, B.F., C.L. and D.B.; funding acquisition, B.F., C.L., and D.B. All authors have read and agreed to the published version of the manuscript.
Funding
This publication has emanated from research conducted with the financial support of Science Foundation Ireland (SFI), under grant number [20/FIP/PL/8940].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Papagianni, M. Metabolic Engineering of Lactic Acid Bacteria for the Production of Industrially Important Compounds. Comput. Struct. Biotechnol. 2012, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Lactic Acid Bacteria : Their Applications in Foods. J. Bacteriol. Mycol. 2018, 6, 89–94. [Google Scholar] [CrossRef]
- Khalid, K. An Overview of Lactic Acid Bacteria. Int. J. Biosci. 2011, 1, 1–13. [Google Scholar]
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva, R.C. da; Ibrahim, S.A. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 202–232. [Google Scholar] [CrossRef]
- Feord, J. Lactic Acid Bacteria in a Changing Legislative Environment. Antonie Van Leeuwenhoek 2002, 82, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, H.M.; Rochfort, K.D.; Maye, S.; MacLeod, G.; Brabazon, D.; Loscher, C.; Freeland, B. Exopolysaccharides of Lactic Acid Bacteria: Production, Purification and Health Benefits towards Functional Food. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, J. The Lactic Acid Bacterium as a Cell Factory for Food Ingredient Production. Int. Dairy J. 2008, 18, 466–475. [Google Scholar] [CrossRef]
- Taskila, S.; Ojamo, H. The Current Status and Future Expectations in Industrial Production of Lactic Acid by Lactic Acid Bacteria. Lact. Acid Bact. - R D Food, Heal. Livest. Purp. 2013. [Google Scholar] [CrossRef]
- Vijayakumar, J.; Aravindan, R.; Viruthagiri, T. Recent Trends in the Production, Purification and Application of Lactic Acid. Chem. Biochem. Eng. Q. 2008, 22, 245–264. [Google Scholar]
- Khedkar, S.; Bröring, S.; Ciliberti, S. Exploring the Nutrition and Health Claims Regulation ( EC ) No. 1924 / 2006 : What Is the Impact on Innovation in the EU Food Sector ? Int. J. Food Sci. Nutr. 2016, 0, 000. [Google Scholar] [CrossRef]
- Verhagen, H.; Verloo, D.; Bronzwaer, S.; Kass, G.; Robinson, T.; Vrbos, D.; Hugas, M. Food Safety Regulatory Research Needs 2030. EFSA J. 2019, 17. [Google Scholar] [CrossRef]
- (Institution), E.C. Regulation (EC) No. 1924/2006 of the European Parliament and of the Council of 20th December 2006 on Nutrition and Health Claims Made on Foods; 2006; 20 December.
- Scientific and Technical Guidance for the Preparation and Presentation of a Health Claim Application (Revision 3)1; 2021; Vol. 19;
- Collins, N. ; Hans Verhagen Nutrition and Health Claims in the European Union in 2022. Available online: https://www.raps.org/news-and-articles/news-articles/2022/9/nutrition-and-health-claims-in-the-european-union (accessed on 10 July 2023).
- EFSA Nutrition Applications: Regulations and Guidance. Available online: https://www.efsa.europa.eu/en/applications/nutrition/regulationsandguidance (accessed on 10 July 2023).
- Schnettler, B.; Bou, L.; García, M.; Mercedes, S. Consumers ’ Willingness to Pay for Health Claims during the COVID-19 Pandemic : A Moderated Mediation Analysis. J. Agric. Food Res. 2023, 11. [Google Scholar] [CrossRef]
- Boer, A. De Fifteen Years of Regulating Nutrition and Health Claims in Europe: The Past, the Present and the Future. Nutrients 2021, 13. [Google Scholar]
- Dairygold Food Ingredients Ltd. Aerabo Active Vitality. Available online: https://aerabo.ie/ (accessed on 10 July 2023).
- Dairygold Food Ingredients Ltd. Aerabo Active Boost. Available online: https://aerabo.ie/ (accessed on 7 October 2023).
- Dairygold Food Ingredients Ltd. Aerabo Active Light. Available online: https://aerabo.ie/ (accessed on 10 July 2023).
- Müller Kefir Smoothie. Available online: https://www.muller.co.uk/our-brands/muller-kefir-smoothie/muller-kefir-smoothie (accessed on 7 July 2023).
- Glenillen Farm Gut Health Kefir. Available online: https://glenilenfarm.com/gut-health-kefir/?_gl=1*872m4o*_up*MQ..*_ga*MzI2MTQ0NTAzLjE2ODg3MzU0Mzc.*_ga_DQMB2TF4BR*MTY4ODczNTQzNi4xLjAuMTY4ODczNTQzNi4wLjAuMA.. (accessed on 7 July 2023).
- Irish Yogurts Clonakilty (Institution) Spoonable Kefir.
- Biotiful Gut Health (institution) Kefir Drink Original. Available online: https://biotifulguthealth.com/products/kefir/ (accessed on 7 July 2023).
- Biotiful Gut Health (institution) Kefir Yoghurt Original. Available online: https://biotifulguthealth.com/products/kefir-yogurt-original/ (accessed on 7 July 2023).
- Upfield Flora 100% Natural Ingredients Dairy Free & Rich In Omega 3. Available online: https://www.flora.com/en-gb/flora/products/flora-original (accessed on 7 July 2023).
- Upfield Flora ProActiv Spreads. Available online: https://www.pro-activ.com/en-ie/produ (accessed on 7 July 2023).
- Raisio Plc Benecol Spreads.
- Tirlán Avonmore Super Milk. Available online: https://www.avonmore.ie/products/whole-super-milk (accessed on 7 July 2023).
- Tirlán Avonmore Fibre Plus Milk. Available online: Avonmore Fibre Plus Milk (accessed on 7 July 2023).
- Tirlán Avonmore Slimline Milk. Available online: https://www.avonmore.ie/products/slimline-milk (accessed on 7 July 2023).
- Nestle Nido. Available online: https://www.nestle.com/brands/baby-foods/nido (accessed on 7 July 2023).
- Arla Baby&Me Organic. Available online: https://www.arla.com/our-brands/babyme-organic/ (accessed on 7 July 2023).
- Campina, F. Goede Morgen. Available online: https://www.vifit.nl/producten (accessed on 7 July 2023).
- Danone Activia. Available online: https://www.danone.com/brands/dairy-plant-based-products/activia.html (accessed on 7 July 2023).
- Danone Danonino. Available online: https://www.danone.com/brands/dairy-plant-based-products/danonino.html (accessed on 7 July 2023).
- Raisio Plc Benecol Yoghurt. Available online: https://benecol.ie/products/#/feed&categories=yogurts (accessed on 7 July 2023).
- Farm, G. Natural Yoghurt. Available online: https://glenilenfarm.com/natural-yoghurt/?_gl=1*1k6o5yn*_up*MQ..*_ga*MTk3NzM2Mzc5OS4xNjg4NzM1OTY2*_ga_DQMB2TF4BR*MTY4ODczNTk2NS4xLjAuMTY4ODczNTk2NS4wLjAuMA.. (accessed on 7 July 2023).
- Irish Yogurts Clonakilty Greek Style Natural. Available online: https://www.irish-yogurts.ie/products/?gclid=Cj0KCQjw756lBhDMARIsAEI0AgkFfC8BwPWXFKmOU8J4UcStFaReW7gm2CKQBogmHBWcMsi1PuDHvHIaAo2sEALw_wcB (accessed on 7 July 2023).
- Danone Actimel. Available online: https://www.danone.com/brands/dairy-plant-based-products/actimel.html (accessed on 7 July 2023).
- Gangadharan, D.; Nampoothiri, K.M. Folate Production Using Lactococcus Lactis Ssp Cremoris with Implications for Fortification of Skim Milk and Fruit Juices. LWT - Food Sci. Technol. 2011, 44, 1859–1864. [Google Scholar] [CrossRef]
- Linares, D.M.; Fitzgerald, G.; Hill, C.; Stanton, C.; Ross, P. Production of Vitamins, Exopolysaccharides and Bacteriocins by Probiotic Bacteria. In Probiotic Dairy Products; 2017; pp. 359–388 ISBN 9781119214106.
- Saubade, F.; Hemery, Y.M.; Guyot, J.P.; Humblot, C. Lactic Acid Fermentation as a Tool for Increasing the Folate Content of Foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 3894–3910. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Wegkamp, A.; De Vos, W.M.; Smid, E.J.; Hugenholtz, J. High-Level Folate Production in Fermented Foods by the B12 Producer Lactobacillus Reuteri JCM1112. Appl. Environ. Microbiol. 2008, 74, 3291–3294. [Google Scholar] [CrossRef] [PubMed]
- Hugenholtz, J.; Smid, E.J. Nutraceutical Production with Food-Grade Microorganisms. Curr. Opin. Biotechnol. 2002, 13, 497–507. [Google Scholar] [CrossRef]
- Sybesma, W.; Starrenburg, M.; Kleerebezem, M.; Mierau, I.; De Vos, W.M.; Hugenholtz, J. Increased Production of Folate by Metabolic Engineering of Lactococcus Lactis. Appl. Environ. Microbiol. 2003, 69, 3069–3076. [Google Scholar] [CrossRef]
- Revuelta, J.L.; Serrano-Amatriain, C.; Ledesma-Amaro, R.; Jiménez, A. Formation of Folates by Microorganisms: Towards the Biotechnological Production of This Vitamin. Appl. Microbiol. Biotechnol. 2018, 102, 8613–8620. [Google Scholar] [CrossRef]
- Laiño, J.E.; Juarez del Valle, M.; Savoy de Giori, G.; LeBlanc, J.G.J. Applicability of a Lactobacillus Amylovorus Strain as Co-Culture for Natural Folate Bio-Enrichment of Fermented Milk. Int. J. Food Microbiol. 2014, 191, 10–16. [Google Scholar] [CrossRef]
- Hugenschmidt, S.; Schwenninger, S.M.; Gnehm, N.; Lacroix, C. Screening of a Natural Biodiversity of Lactic and Propionic Acid Bacteria for Folate and Vitamin B12 Production in Supplemented Whey Permeate. Int. Dairy J. 2010, 20, 852–857. [Google Scholar] [CrossRef]
- Masuda, M.; Ide, M.; Utsumi, H.; Niiro, T.; Shimamura, Y.; Murata, M. Production Potency of Folate, Vitamin B12, and Thiamine by Lactic Acid Bacteria Isolated from Japanese Pickles. Biosci. Biotechnol. Biochem. 2012, 76, 2061–2067. [Google Scholar] [CrossRef] [PubMed]
- Sybesma, W.; Starrenburg, M.; Tijsseling, L.; Hoefnagel, M.H.N.; Hugenholtz, J. Effects of Cultivation Conditions on Folate Production by Lactic Acid Bacteria. Appl. Environ. Microbiol. 2003, 69, 4542–4548. [Google Scholar] [CrossRef]
- Laiño, J.E.; Juarez del Valle, M.; Savoy de Giori, G.; LeBlanc, J.G.J. Development of a High Folate Concentration Yogurt Naturally Bio-Enriched Using Selected Lactic Acid Bacteria. LWT - Food Sci. Technol. 2013, 54, 1–5. [Google Scholar] [CrossRef]
- Gangadharan, D.; Nampoothiri, K.M. Folate Production Using Lactococcus Lactis Ssp Cremoris with Implications for Fortification of Skim Milk and Fruit Juices. LWT - Food Sci. Technol. 2011, 44, 1859–1864. [Google Scholar] [CrossRef]
- Hugenschmidt, S.; Schwenninger, S.M.; Lacroix, C. Concurrent High Production of Natural Folate and Vitamin B12 Using a Co-Culture Process with Lactobacillus Plantarum SM39 and Propionibacterium Freudenreichii DF13. Process Biochem. 2011, 46, 1063–1070. [Google Scholar] [CrossRef]
- Mousavi, S.S.; Moeini, H.; Mohamad, R.; Dinarvand, M.; Ariff, A.; Ling, F.H.; Raha, A.R. Effects of Medium and Culture Conditions on Folate Production by Streptococcus Thermophilus BAA-250. Res. Biotechnol. 2013, 4, 21–29. [Google Scholar]
- Mousavi, S.S.; Moeini, H.; Mohamad, R.; Dinarvand, M.; Ariff, A.; Ling, F.H.; Raha, A.R. Effects of Medium and Culture Conditions on Folate Production by Streptococcus Thermophilus BAA-250; 2013; Vol. 4;
- Rad, A.H.; Yari Khosroushahi, A.; Khalili, M.; Jafarzadeh, S. Folate Bio-Fortification of Yoghurt and Fermented Milk: A Review. Dairy Sci. Technol. 2016, 96, 427–441. [Google Scholar] [CrossRef]
- Choi, J.H.; Yates, Z.; Veysey, M.; Heo, Y.R.; Lucock, M. Contemporary Issues Surrounding Folic Acid Fortification Initiatives. Prev. Nutr. Food Sci. 2014, 19, 247–260. [Google Scholar] [CrossRef]
- Blencowe, H.; Cousens, S.; Modell, B.; Lawn, J. Folic Acid to Reduce Neonatal Mortality from Neural Tube Disorders. Int. J. Epidemiol. 2010, 39, 110–121. [Google Scholar] [CrossRef]
- Yates, A.A.; Schlicker, S.A.; Suitor, C.W. Dietary Reference Intakes: The New Basis for Recommendations for Calcium and Related Nutrients, B Vitamins, and Choline. J. Am. Diet. Assoc. 1998, 98, 699–706. [Google Scholar] [CrossRef]
- Wang, H.X.; Wahlin, Å.; Basun, H.; Fastbom, J.; Winblad, B.; Fratiglioni, L. Vitamin B12 and Folate in Relation to the Development of Alzheimer’s Disease. Neurology 2001, 56, 1188–1194. [Google Scholar] [CrossRef]
- Ho, P.I.; Ashline, D.; Dhitavat, S.; Ortiz, D.; Collins, S.C.; Shea, T.B.; Rogers, E. Folate Deprivation Induces Neurodegeneration: Roles of Oxidative Stress and Increased Homocysteine. Neurobiol. Dis. 2003, 14, 32–42. [Google Scholar] [CrossRef]
- Duthie, S.J.; Whalley, L.J.; Collins, A.R.; Leaper, S.; Berger, K.; Deary, I.J. Homocysteine, B Vitamin Status, and Cognitive Function in the Elderly. Am. J. Clin. Nutr. 2002, 75, 908–913. [Google Scholar] [CrossRef]
- Morris, M.S. Folate, Homocysteine, and Neurological Function. Nutr. Clin. Care 2007, 44, 113–122. [Google Scholar] [CrossRef]
- Godfrey, P.S.A.; Toone, B.K.; Carney, M.W.P.; Flynn, T.G.; Bottiglien, T.; Laundy, M.; Chanarin, I.; Reynolds, E.H. Enhancement of Recovery from Psychiatric Illness by Methylfolate. Psychosomatics 1990, 336, 392–395. [Google Scholar]
- Iyer, R.; Tomar, S.K. Folate: A Functional Food Constituent. J. Food Sci. 2009, 74. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, Y.; Wang, Y.; Li, L.; Liao, Y.; Zhang, Y.; Yu, D.; Omboni, S. The Effect of Folic Acid in Patients with Cardiovascular Disease: A Systematic Review and Meta-Analysis. Med. (United States) 2019, 98. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Brattström, L.; Landgren, F.; Israelsson, B.; Lindgren, A.; Hultberg, B.; Andersson, A.; Cuskelly, G.; McNulty, H.; Strain, S.S.; et al. Lowering Blood Homocysteine with Folic Acid Based Supplements: Meta-Analysis of Randomised Trials. Br. Med. J. 1998, 316, 894–898. [Google Scholar] [CrossRef]
- Ulrich, C.M. Folate and Cancer Prevention: A Closer Look at a Complex Picture. Am. J. Clin. Nutr. 2007, 86, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Survase, S.A.; Bajaj, I.B.; Singhal, R.S. Biotechnological Production of Vitamins. Food Technol. Biotechnol. 2006, 44, 381–396. [Google Scholar]
- Martens, J.H.; Barg, H.; Warren, M.; Jahn, D. Microbial Production of Vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Chanto, A.; Afschar, A.S.; Wagner, F. Optimization of a Propionibacterium Acidipropionici Continuous Culture Utilizing Sucrose. Appl. Microbiol. Biotechnol. 1994, 3, 16–21. [Google Scholar] [CrossRef]
- Li, K.T.; Liu, D.H.; Li, Y.L.; Chu, J.; Wang, Y.H.; Zhuang, Y.P.; Zhang, S.L. Improved Large-Scale Production of Vitamin B12 by Pseudomonas Denitrificans with Betaine Feeding. Bioresour. Technol. 2008, 99, 8516–8520. [Google Scholar] [CrossRef]
- Li, K.-T.; Liu, D.-H.; Chu, J.; Wang, Y.-H.; Zhuang, Y.-P.; Zhang, S.-L. An Effective and Simplified PH-Stat Control Strategy for the Industrial Fermentation of Vitamin B12 by Pseudomonas Denitrificans. Bioprocess Biosyst. Eng. 2008, 31, 605–610. [Google Scholar] [CrossRef]
- Ye, K.; Shijo, M.; Jin, S.; Shimizu, K. Efficient Production of Vitamin B12 from Propionic Acid Bacteria under Periodic Variation of Dissolved Oxygen Concentration. J. Ferment. Bioeng. 1996, 82, 484–491. [Google Scholar] [CrossRef]
- Miyano, K.I.; Ye, K.; Shimizu, K. Improvement of Vitamin B12 Fermentation by Reducing the Inhibitory Metabolites by Cell Recycle System and a Mixed Culture. Biochem. Eng. J. 2000, 6, 207–214. [Google Scholar] [CrossRef]
- Bullerman, L. .; Berry, E.C. Use of Cheese Whey for Vitamin B12 Production: I. Whey Solids and Yeast Extract Levels. Appl. Microbiol. 1966, 14, 353–355. [Google Scholar] [CrossRef]
- Marwaha, S.S.; Sethi, R.P. Utilization of Dairy Waste for Vitamin B12 Fermentation. Agric. Wastes 1984, 9, 111–130. [Google Scholar] [CrossRef]
- Marwaha, S.S.; Sethi, R.P.; Kennedy, J.F.; Kumar, R. Simulation of Fermentation Conditions for Vitamin B12 Biosynthesis from Whey. Enzyme Microb. Technol. 1983, 5, 449–453. [Google Scholar] [CrossRef]
- Marwaha, S.S.; Sethi, R.P.; Kennedy, J.F. Role of Amino Acids, Betaine and Choline in Vitamin B12 Biosynthesis by Strains of Propionibacterium. Enzyme Microb. Technol. 1983, 5, 454–456. [Google Scholar] [CrossRef]
- Li, K.T.; Liu, D.H.; Chu, J.; Wang, Y.H.; Zhuang, Y.P.; Zhang, S.L. An Effective and Simplified PH-Stat Control Strategy for the Industrial Fermentation of Vitamin B12 by Pseudomonas Denitrificans. Bioprocess Biosyst. Eng. 2008, 31, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Hunik, J.H. Process for the Production of Vitamin B12 2002, 1.
- Ye, K.; Shijo, M.; Jin, S.; Shimizu, K. Efficient Production of Vitamin from Propionic Acid Bacteria under Periodic Variation of Dissolved Oxygen Concentration. J. Ferment. Bioeng. 1996, 82, 484–491. [Google Scholar] [CrossRef]
- Hsu, S.-T.; Yang, S.-T. Propionic Acid Fermentation of Lactose by Propionibacterium Acidipropionici: Effects of PH. J. Biol. Chem. 1991, 56, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Hunt, A.; Harrington, D.; Robinson, S. Vitamin B12 Deficiency. BMJ 2014, 349. [Google Scholar] [CrossRef]
- Allen, L.H. How Common Is Vitamin B-12 Deficiency? Am. J. Clin. Nutr. 2009, 89, 693–696. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.G.; Wyatt, P.R.; Thompson, M.D.; Vermeulen, M.J.; Meier, C.; Wong, P.Y.; Farrell, S.A.; Cole, D.E.C. Vitamin B12 and the Risk of Neural Tube Defects in a Folic-Acid-Fortified Population. Epidemiology 2007, 18, 362–366. [Google Scholar] [CrossRef]
- Afman, L.A.; Van Der Put, N.M.J.; Thomas, C.M.G.; Trijbels, J.M.F.; Blom, H.J. Reduced Vitamin B12 Binding by Transcobalamin II Increases the Risk of Neural Tube Defects. QJM - Mon. J. Assoc. Physicians 2001, 94, 159–166. [Google Scholar] [CrossRef]
- Molloy, A.M. Should Vitamin B12 Status Be Considered in Assessing Risk of Neural Tube Defects? Ann. N. Y. Acad. Sci. 2018, 1414, 109–125. [Google Scholar] [CrossRef]
- Clarke, R.; Frost, C.; Sherliker, P.; Lewington, S.; Collins, R.; Brattstrom, L.; Brouwer, I.; van Dusseldorp, M.; Steegers-Theunissen, R.P.M.; Cuskelly, G.; et al. Dose-Dependent Effects of Folic Acid on Blood Concentrations of Homocysteine: A Meta-Analysis of the Randomized Trials. Am. J. Clin. Nutr. 2005, 82, 806–812. [Google Scholar] [CrossRef]
- O’Leary, F.; Samman, S. Vitamin B12 in Health and Disease. Nutrients 2010, 2, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Dhonukshe-Rutten, R.A.M.; Van Dusseldorp, M.; Schneede, J.; De Groot, L.C.P.G.M.; Van Staveren, W.A. Low Bone Mineral Density and Bone Mineral Content Are Associated with Low Cobalamin Status in Adolescents. Eur. J. Nutr. 2005, 44, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Muthayya, S.; Kurpad, A. V.; Duggan, C.P.; Bosch, R.J.; Dwarkanath, P.; Mhaskar, A.; Mhaskar, R.; Thomas, A.; Vaz, M.; Bhat, S.; et al. Low Maternal Vitamin B12 Status Is Associated with Intrauterine Growth Retardation in Urban South Indians. Eur. J. Clin. Nutr. 2006, 60, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Roman-Garcia, P.; Quiros-Gonzalez, I.; Mottram, L.; Lieben, L.; Sharan, K.; Wangwiwatsin, A.; Tubio, J.; Lewis, K.; Wilkinson, D.; Santhanam, B.; et al. Vitamin B12-Dependent Taurine Synthesis Regulates Growth and Bone Mass. J. Clin. Invest. 2014, 124, 2988–3002. [Google Scholar] [CrossRef] [PubMed]
- Sybesma, W.; Burgess, C.; Starrenburg, M.; Sinderen, D. van; Hugenholtz, J. Multivitamin Production in Lactococcus Lactis Using Metabolic Engineering. Metab. Eng. 2004, 6, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Stahmann, K.P.; Kupp, C.; Feldmann, S.D.; sahm, H. Formation and Degradation of Lipid Bodies Found in the Riboflavin-Producing Fungus Ashbya Gossypii. Appl. Microbiol. Biotechnol. 1994, 42, 121–127. [Google Scholar] [CrossRef]
- Guru, V.; Viswanathan, K. Riboflavin Production in Milk Whey Using Probiotic Bacteria-Lactobacillus Acidophilus and Lactococcus Lactis. Indian J. Fundam. Appl. Life Sci. 2013, 3, 169–176. [Google Scholar]
- Russo, P.; Capozzi, V.; Arena, M.P.; Spadaccino, G.; Dueñas, M.T.; López, P.; Fiocco, D.; Spano, G. Riboflavin-Overproducing Strains of Lactobacillus Fermentum for Riboflavin-Enriched Bread. Appl. Microbiol. Biotechnol. 2014, 98, 3691–3700. [Google Scholar] [CrossRef]
- Jayashree, S.; Rajendhran, J.; Jayaraman, K.; Kalaichelvan, G.; Gunasekaran, P. Improvement of Riboflavin Production by Lactobacillus Fermentum Isolated from Yogurt. Food Biotechnol. 2011, 25, 240–251. [Google Scholar] [CrossRef]
- Mohedano, M.L.; Hernández-Recio, S.; Yépez, A.; Requena, T.; Martínez-Cuesta, M.C.; Peláez, C.; Russo, P.; LeBlanc, J.G.; Spano, G.; Aznar, R.; et al. Real-Time Detection of Riboflavin Production by Lactobacillus Plantarum Strains and Tracking of Their Gastrointestinal Survival and Functionality in Vitro and in Vivo Using MCherry Labeling. Front. Microbiol. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Arena, M.P.; Russo, P.; Capozzi, V.; López, P.; Fiocco, D.; Spano, G. Probiotic Abilities of Riboflavin-Overproducing Lactobacillus Strains: A Novel Promising Application of Probiotics. Appl. Microbiol. Biotechnol. 2014, 98, 7569–7581. [Google Scholar] [CrossRef] [PubMed]
- Juarez del Valle, M.; Laiño, J.E.; Savoy de Giori, G.; LeBlanc, J.G. Factors Stimulating Riboflavin Produced by Lactobacillus Plantarum CRL 725 Grown in a Semi-Defined Medium. J. Basic Microbiol. 2017, 57, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Guru V; Viswanathan K Riboflavin Production in Milk Whey Using Probiotic Bacteria-Lactobacillus Acidophilus and Lactococcus Lactis. Indian J. Fundam. Appl. Life Sci. 2013, 3, 169–176.
- Burgess, C.; O’Connell-Motherway, M.; Sybesma, W.; Hugenholtz, J.; Van Sinderen, D. Riboflavin Production in Lactococcus Lactis: Potential for in Situ Production of Vitamin-Enriched Foods. Appl. Environ. Microbiol. 2004, 70, 5769–5777. [Google Scholar] [CrossRef] [PubMed]
- Powers, H.J. Riboflavin (Vitamin B-2) and Health. Am. J. Clin. Nutr. 2003, 77, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Ashoori, M.; Saedisomeolia, A. Riboflavin (Vitamin B2) and Oxidative Stress: A Review. Br. J. Nutr. 2014, 111, 1985–1991. [Google Scholar] [CrossRef] [PubMed]
- Saedisomeolia, A.; Ashoori, M. Riboflavin in Human Health: A Review of Current Evidences. In Advances in Food and Nutrition Research; Elsevier Inc., 2018; Vol. 83, pp. 57–81.
- Shrubsole, M.J.; Shu, X.O.; Li, H.L.; Cai, H.; Yang, G.; Gao, Y.T.; Gao, J.; Zheng, W. Dietary B Vitamin and Methionine Intakes and Breast Cancer Risk among Chinese Women. Am. J. Epidemiol. 2011, 173, 1171–1182. [Google Scholar] [CrossRef]
- Takata, Y.; Cai, Q.; Beeghly-Fadiel, A.; Li, H.; Shrubsole, M.J.; Ji, B.T.; Yang, G.; Chow, W.H.; Gao, Y.T.; Zheng, W.; et al. Dietary B Vitamin and Methionine Intakes and Lung Cancer Risk among Female Never Smokers in China. Cancer Causes Control 2012, 23, 1965–1975. [Google Scholar] [CrossRef]
- Van Den Donk, M.; Buijsse, B.; Van Den Berg, S.W.; Ocké, M.C.; Harryvan, J.L.; Nagengast, F.M.; Kok, F.J.; Kampman, E. Dietary Intake of Folate and Riboflavin, MTHFR C677T Genotype, and Colorectal Adenoma Risk: A Dutch Case-Control Study. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 1562–1566. [Google Scholar] [CrossRef]
- Eussen, S.J.P.M.; Vollset, S.E.; Hustad, S.; Midttun, Ø.; Meyer, K.; Fredriksen, Å.; Ueland, P.M.; Jenab, M.; Slimani, N.; Ferrari, P.; et al. Vitamins B2 and B6 and Genetic Polymorphisms Related to One-Carbon Metabolism as Risk Factors for Gastric Adenocarcinoma in the European Prospective Investigation into Cancer and Nutrition. Cancer Epidemiol. Biomarkers Prev. 2010, 19, 28–38. [Google Scholar] [CrossRef]
- Kabat, G.C.; Miller, A.B.; Jain, M.; Rohan, T.E. Dietary Intake of Selected B Vitamins in Relation to Risk of Major Cancers in Women. Br. J. Cancer 2008, 99, 816–821. [Google Scholar] [CrossRef]
- Coimbra, C.G.; Junqueira, V.B.C. High Doses of Riboflavin and the Elimination of Dietary Red Meat Promote the Recovery of Some Motor Functions in Parkinson’s Disease Patients. Brazilian J. Med. Biol. Res. 2003, 36, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.C.; Gorell, J.M.; Rybicki, B.A.; Sanders, K.; Peterson, E.L. Adult Nutrient Intake as a Risk Factor for Parkinson’s Disease. Int. J. Epidemiol. 1999, 28, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Håglin, L.; Johansson, I.; Forsgren, L.; Bäckman, L. Intake of Vitamin B before Onset of Parkinson’s Disease and Atypical Parkinsonism and Olfactory Function at the Time of Diagnosis. Eur. J. Clin. Nutr. 2017, 71, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Naghashpour, M.; Amani, R.; Sarkaki, A.; Ghadiri, A.; Samarbafzadeh, A.; Jafarirad, S.; Saki Malehi, A. Brain-Derived Neurotrophic and Immunologic Factors: Beneficial Effects of Riboflavin on Motor Disability in Murine Model of Multiple Sclerosis. Iran. J. Basic Med. Sci. 2016, 19, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Naghashpour, M.; Majdinasab, N.; Shakerinejad, G.; Kouchak, M.; Haghighizadeh, M.H.; Jarvandi, F.; Hajinajaf, S. Riboflavin Supplementation to Patients with Multiple Sclerosis Does Not Improve Disability Status nor Is Riboflavin Supplementation Correlated to Homocysteine. Int. J. Vitam. Nutr. Res. 2013, 83, 281–290. [Google Scholar] [CrossRef]
- Lewis, D.; Ashwal, S.; Hershey, A.; Hirtz, D.; Yonker, M.; Silberstein, S. Practice Parameter: Pharmacological Treatment of Migraine Headache in Children and Adolescents: Report of the American Academy of Neurology Quality Standards Subcommittee and the Practice Committee of the Child Neurology Society. Neurology 2004, 63, 2215–2224. [Google Scholar] [CrossRef] [PubMed]
- Schoenen, J.; Jacquy, J.; Lenaerts, M. Effectiveness of High-Dose Riboflavin in Migraine Prophylaxis: A Randomized Controlled Trial. Neurology 1998, 50, 466–470. [Google Scholar] [CrossRef]
- Boehnke, C.; Reuter, U.; Flach, U.; Schuh-Hofer, S.; Einhäupl, K.M.; Arnold, G. High-Dose Riboflavin Treatment Is Efficacious in Migraine Prophylaxis: An Open Study in a Tertiary Care Centre. Eur. J. Neurol. 2004, 11, 475–477. [Google Scholar] [CrossRef]
- Mahdinia, E.; Demirci, A.; Berenjian, A. Production and Application of Menaquinone-7 (Vitamin K2): A New Perspective. World J. Microbiol. Biotechnol. 2017, 33, 1–7. [Google Scholar] [CrossRef]
- Lim, S.D.; Kim, K.S.; Do, J.R. Physiological Characteristics and Production of Vitamin K2 by Lactobacillus Fermentum LC272 Isolated from Raw Milk. Korean J. Food Sci. Anim. Resour. 2011, 31, 513–520. [Google Scholar] [CrossRef]
- Morishita, T.; Tamura, N.; Makino, T.; Kudo, S. Production of Menaquinones by Lactic Acid Bacteria. J. Dairy Sci. 1999, 82, 1897–1903. [Google Scholar] [CrossRef]
- Furuichi, K.; Hojo, K. ichi; Katakura, Y.; Ninomiya, K.; Shioya, S. Aerobic Culture of Propionibacterium Freudenreichii ET-3 Can Increase Production Ratio of 1,4-Dihydroxy-2-Naphthoic Acid to Menaquinone. J. Biosci. Bioeng. 2006, 101, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Berenjian, A.; Mahanama, R.; Kavanagh, J.; Dehghani, F. Vitamin K Series: Current Status and Future Prospects. Crit. Rev. Biotechnol. 2015, 35, 199–208. [Google Scholar] [CrossRef] [PubMed]
- DiNicolantonio, J.J.; Bhutani, J.; O’Keefe, J.H. The Health Benefits of Vitamin K. Open Hear. 2015, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Geleijnse, J.M.; Vermeer, C.; Grobbee, D.E.; Schurgers, L.J.; Knapen, M.H.J.; Van Der Meer, I.M.; Hofman, A.; Witteman, J.C.M. Dietary Intake of Menaquinone Is Associated with a Reduced Risk of Coronary Heart Disease: The Rotterdam Study. J. Nutr. 2004, 134, 3100–3105. [Google Scholar] [CrossRef] [PubMed]
- Beulens, J.W.J.; Booth, S.L.; Van Den Heuvel, E.G.H.M.; Stoecklin, E.; Baka, A.; Vermeer, C. The Role of Menaquinones (Vitamin K2) in Human Health. Br. J. Nutr. 2013, 110, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Gast, G.C.M.; de Roos, N.M.; Sluijs, I.; Bots, M.L.; Beulens, J.W.J.; Geleijnse, J.M.; Witteman, J.C.; Grobbee, D.E.; Peeters, P.H.M.; van der Schouw, Y.T. A High Menaquinone Intake Reduces the Incidence of Coronary Heart Disease. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Beulens, J.W.J.; Bots, M.L.; Atsma, F.; Bartelink, M.L.E.L.; Prokop, M.; Geleijnse, J.M.; Witteman, J.C.M.; Grobbee, D.E.; van der Schouw, Y.T. High Dietary Menaquinone Intake Is Associated with Reduced Coronary Calcification. Atherosclerosis 2009, 203, 489–493. [Google Scholar] [CrossRef]
- Kanellakis, S.; Moschonis, G.; Tenta, R.; Schaafsma, A.; Van Den Heuvel, E.G.H.M.; Papaioannou, N.; Lyritis, G.; Manios, Y. Changes in Parameters of Bone Metabolism in Postmenopausal Women Following a 12-Month Intervention Period Using Dairy Products Enriched with Calcium, Vitamin D, and Phylloquinone (Vitamin K 1) or Menaquinone-7 (Vitamin K 2): The Postmenopausal Health Stud. Calcif. Tissue Int. 2012, 90, 251–262. [Google Scholar] [CrossRef]
- Knapen, M.H.J.; Drummen, N.E.; Smit, E.; Vermeer, C.; Theuwissen, E. Three-Year Low-Dose Menaquinone-7 Supplementation Helps Decrease Bone Loss in Healthy Postmenopausal Women. Osteoporos. Int. 2013, 24, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- Emaus, N.; Gjesdal, C.G.; Almås, B.; Christensen, M.; Grimsgaard, A.S.; Berntsen, G.K.R.; Salomonsen, L.; Fønnebø, V. Vitamin K2 Supplementation Does Not Influence Bone Loss in Early Menopausal Women: A Randomised Double-Blind Placebo-Controlled Trial. Osteoporos. Int. 2010, 21, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- de Arauz, L.J.; Jozala, A.F.; Mazzola, P.G.; Vessoni Penna, T.C. Nisin Biotechnological Production and Application: A Review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Gálvez, A.; López, R.L.; Abriouel, H.; Valdivia, E.; Omar, N. Ben Application of Bacteriocins in the Control of Foodborne Pathogenic and Spoilage Bacteria. Crit. Rev. Biotechnol. 2008, 28, 125–152. [Google Scholar] [CrossRef] [PubMed]
- Balciunas, E.M.; Castillo Martinez, F.A.; Todorov, S.D.; Franco, B.D.G. de M.; Converti, A.; Oliveira, R.P. de S. Novel Biotechnological Applications of Bacteriocins: A Review. Food Control 2013, 32, 134–142. [Google Scholar] [CrossRef]
- Amiali, M.N.; Lacroix, C.; Simard, R.E.; Anastasiadou, S.; Papagianni, M.; Ambrosiadis, I.; Koidis, P.; Balciunas, E.M.; Castillo Martinez, F.A.; Todorov, S.D.; et al. High Nisin Z Production by Lactococcus Lactis UL719 in Whey Permeate with Aeration. Int. Dairy J. 2008, 9, 17. [Google Scholar] [CrossRef]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, Natural Antimicrobials for Food Preservation. Int. J. Food Microbiol. 2011, 2, 90. [Google Scholar] [CrossRef]
- Deegan, L.H.; Cotter, P.D.; Hill, C.; Ross, P. Bacteriocins : Biological Tools for Bio-Preservation and Shelf-Life Extension. Int. Dairy J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- García-Parra, M.D.; García-Almendárez, B.E.; Guevara-Olvera, L.; Guevara-González, R.G.; Rodríguez, A.; Martínez, B.; Domínguez-Domínguez, J.; Regalado, C. Effect of Sub-Inhibitory Amounts of Nisin and Mineral Salts on Nisin Production by Lactococcus Lactis UQ2 in Skim Milk. Food Bioprocess Technol. 2011, 4, 646–654. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Liao, W.; Wen, Z.; Chen, S. Simultaneous Production of Nisin and Lactic Acid from Cheese Whey: Optimization of Fermentation Conditions through Statistically Based Experimental Designs. Appl. Biochem. Biotechnol. - Part A Enzym. Eng. Biotechnol. 2004, 114, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Jozala, A.F.; Silva, D.P.; Vicente, A.A.; Teixeira, J.A.; Júnior, P.; Penna, T.C. V Processing of Byproducts to Improve Nisin Production by Lactococcus Lactis. Int. Dairy J. 2011, 10, 14920–14925. [Google Scholar] [CrossRef]
- Senan, S.; El-aal, H.A.; Dave, R.; Hassan, A. Production and Stability of Nisin in Whey Protein Concentrate. LWT - Food Sci. Technol. 2016, 71, 125–129. [Google Scholar] [CrossRef]
- Amiali, M.N.; Lacroix, C.; Simard, R.E. High Nisin Z Production by Lactococcus Lactis UL719 in Whey Permeate with Aeration. World J. Microbiol. Biotechnol. 1998, 14, 887–894. [Google Scholar] [CrossRef]
- Anastasiadou, S.; Papagianni, M.; Ambrosiadis, I.; Koidis, P. Rapid Quantifiable Assessment of Nutritional Parameters Influencing Pediocin Production by Pediococcus Acidilactici NRRL B5627. Bioresour. Technol. 2008, 99, 6646–6650. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, N.; Fliss, I.; Lacroix, C. High Nisin-Z Production during Repeated-Cycle Batch Cultures in Supplemented Whey Permeate Using Immobilized Lactococcus Lactis UL719. Int. Dairy J. 2001, 11, 953–960. [Google Scholar] [CrossRef]
- Lv, W.; Zhang, X.; Cong, W. Modelling the Production of Nisin by Lactococcus Lactis in Fed-Batch Culture. Appl. Microbiol. Biotechnol. 2005, 68, 322–326. [Google Scholar] [CrossRef]
- Malvido, M.C.; González, E.A.; Guerra, N.P. Nisin Production in Realkalized Fed-Batch Cultures in Whey with Feeding with Lactose- or Glucose-Containing Substrates. Appl. Microbiol. Biotechnol. 2016, 100, 7899–7908. [Google Scholar] [CrossRef]
- Malvido, M.C.; Alonso González, E.; Outeiriño, D.; Fajardo Bernárdez, P.; Pérez Guerra, N. Combination of Food Wastes for an Efficient Production of Nisin in Realkalized Fed-Batch Cultures. Biochem. Eng. J. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Malvido, M.C.; González, E.A.; Bazán Tantaleán, D.L.; Bendaña Jácome, R.J.; Guerra, N.P. Batch and Fed-Batch Production of Probiotic Biomass and Nisin in Nutrient-Supplemented Whey Media. Brazilian J. Microbiol. 2019, 50, 915–925. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, L.; Jing, Y.; Li, X.; Zhao, Y. Variable Volume Fed-Batch Fermentation for Nisin Production by Lactococcus Lactis Subsp. Lactis W28. Appl. Biochem. Biotechnol. 2009, 152, 372–382. [Google Scholar] [CrossRef]
- Simova, E.D.; Beshkova, D.M.; Angelov, M.P.; Dimitrov, Z.P. Bacteriocin Production by Strain Lactobacillus Delbrueckii Ssp. Bulgaricus BB18 during Continuous Prefermentation of Yogurt Starter Culture and Subsequent Batch Coagulation of Milk. J. Ind. Microbiol. Biotechnol. 2008, 35, 559–567. [Google Scholar] [CrossRef]
- Shimizu, H.; Mizuguchi, T.; Tanaka, E.; Shioya, S. Nisin Production by a Mixed-Culture System Consisting of Lactococcus Lactis and Kluyveromyces Marxianus. Appl. Environ. Microbiol. 1999, 65, 3134–3141. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hu, B.; Liu, Y.; Chen, S. Stimulation of Nisin Production from Whey by a Mixed Culture of Lactococcus Lactis and Saccharomyces Cerevisiae. Appl. Biochem. Biotechnol. 2006, 131, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Guerra, N.P.; Pastrana, L. Enhanced Nisin and Pediocin Production on Whey Supplemented with Different Nitrogen Sources. Biotechnol. Lett. 2001, 23, 609–612. [Google Scholar] [CrossRef]
- Guerra, N.P.; Pastrana, L. Influence of PH Drop on Both Nisin and Pediocin Production by Lactococcus Lactis and Pediococcus Acidilactici. Lett. Appl. Microbiol. 2003, 37, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Guerra, N.P.; Rua, M.L.; Pastrana, L. Nutritional Factors Affecting the Production of Two Bacteriocins from Lactic Acid Bacteria on Whey. Int. J. Food Microbiol. 2001, 70, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Goulhen, F.; Meghrous, J.; Lacroix, C. Production of a Nisin Z/Pediocin Mixture by PH-Controlled Mixed-Strain Batch Cultures in Supplemented Whey Permeate. J. Appl. Microbiol. 1999, 86, 399–406. [Google Scholar] [CrossRef]
- Liao, C.; Yousef, A.E.; Richter, E.R.; Chism, G.W. Pediococcus Acidilactici PO2 Bacteriocin Production in Whey Permeate and Inhibition of Listeria Monocyfogenes in Foods. J. Food Sci. 1993, 58, 430–434. [Google Scholar] [CrossRef]
- Somkuti, G.A.; Steinberg, D.H. Pediocin Production in Milk by Pediococcus Acidilactici in Co-Culture with Streptococcus Thermophilus and Lactobacillus Delbrueckii Subsp. Bulgaricus. J. Ind. Microbiol. Biotechnol. 2010, 37, 65–69. [Google Scholar] [CrossRef]
- Halami, P.M.; Chandrashekar, A. Enhanced Production of Pediocin C20 by a Native Strain of Pediococcus Acidilactici C20 in an Optimized Food-Grade Medium. Process Biochem. 2005, 40, 1835–1840. [Google Scholar] [CrossRef]
- Guerra, N.P.; Bernárdez, P.F.; Castro, L.P. Fed-Batch Pediocin Production on Whey Using Different Feeding Media. Enzyme Microb. Technol. 2007, 41, 397–406. [Google Scholar] [CrossRef]
- Guerra, N.P.; Bernárdez, P.F.; Agrasar, A.T.; López Macías, C.; Castro, L.P. Fed-Batch Pediocin Production by Pediococcus Acidilactici NRRL B-5627 on Whey. Biotechnol. Appl. Biochem. 2005, 42, 17. [Google Scholar] [CrossRef]
- Biswas, S.R.; Ray, P.; Johnson, M.C.; Ray, B. Influence of Growth Conditions on the Production of a Bacteriocin, Pediocin AcH, by Pediococcus Acidilactici H. Appl. Environ. Microbiol. 1991, 57, 1265–1267. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Fliss, I.; Drider, D.; Lacroix, C. Pediocin PA-1 Production during Repeated-Cycle Batch Culture of Immobilized Pediococcus Acidilactici UL5 Cells. J. Biosci. Bioeng. 2008, 105, 513–517. [Google Scholar] [CrossRef]
- Garsa, A.K.; Kumariya, R.; Kumar, A.; Lather, P.; Kapila, S.; Sood, S.K. Industrial Cheese Whey Utilization for Enhanced Production of Purified Pediocin PA-1. LWT - Food Sci. Technol. 2014, 59, 656–665. [Google Scholar] [CrossRef]
- Gutiérrez-Cortés, C.; Suarez, H.; Buitrago, G.; Nero, L.A.; Todorov, S.D. Enhanced Bacteriocin Production by Pediococcus Pentosaceus 147 in Co-Culture With Lactobacillus Plantarum LE27 on Cheese Whey Broth. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Shin, J.M.; Gwak, J.W.; Kamarajan, Pachiyappan J. Fenno, C.; Rickard, A.H.; Kapila, L.Y. Biomedical Applications of Nisin. J. Appl. Microbiol. 2016, 120, 145–166. [Google Scholar] [CrossRef]
- Porto, M.C.W.; Kuniyoshi, T.M.; Azevedo, P.O.S.; Vitolo, M.; Oliveira, R.P.S. Pediococcus Spp.: An Important Genus of Lactic Acid Bacteria and Pediocin Producers. Biotechnol. Adv. 2017, 35, 361–374. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Martínez, M.I.; Kok, J. Pediocin PA-1, a Wide-Spectrum Bacteriocin from Lactic Acid Bacteria. Crit. Rev. Food Sci. Nutr. 2002, 42, 91–121. [Google Scholar] [CrossRef]
- Papagianni, M.; Anastasiadou, S. Pediocins: The Bacteriocins of Pediococci. Sources, Production, Properties and Applications. Microb. Cell Fact. 2009, 8, 1–16. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Fliss, I.; Drider, D.; Lacroix, C. Pediocin PA-1 Production during Repeated-Cycle Batch Culture of Immobilized Pediococcus Acidilactici UL5 Cells. J. Biosci. Bioeng. 2008, 105, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Pérez Guerra, N.; Bernárdez, P.F.; Agrasar, A.T.; López Macías, C.; Castro, L.P. Fed-Batch Pediocin Production by Pediococcus Acidilactici NRRL B-5627 on Whey. Biotechnol. Appl. Biochem. 2005, 42, 17. [Google Scholar] [CrossRef] [PubMed]
- Guerra, N.P.; Bernárdez, P.F.; Castro, L.P. Fed-Batch Pediocin Production on Whey Using Different Feeding Media. Enzyme Microb. Technol. 2007, 41, 397–406. [Google Scholar] [CrossRef]
- Yang, R.; Ray, B. Factors Influencing Production of Bacteriocins by Lactic Acid Bacteria. Food Microbiol. 1994, 11, 281–291. [Google Scholar] [CrossRef]
- Dobson, A.; Cotter, P.D.; Paul Ross, R.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dabour, N.; Zihler, A.; Kheadr, E.; Lacroix, C.; Fliss, I. In Vivo Study on the Effectiveness of Pediocin PA-1 and Pediococcus Acidilactici UL5 at Inhibiting Listeria Monocytogenes. Int. J. Food Microbiol. 2009, 133, 225–233. [Google Scholar] [CrossRef]
- Millette, M.; Cornut, G.; Dupont, C.; Shareck, F.; Archambault, D.; Lacroix, M. Capacity of Human Nisin- and Pediocin-Producing Lactic Acid Bacteria to Reduce Intestinal Colonization by Vancomycin-Resistant Enterococci. Appl. Environ. Microbiol. 2008, 74, 1997–2003. [Google Scholar] [CrossRef]
- De Kwaadsteniet, M.; Doeschate, K.T.; Dicks, L.M.T. Nisin F in the Treatment of Respiratory Tract Infections Caused by Staphylococcus Aureus. Lett. Appl. Microbiol. 2009, 48, 65–70. [Google Scholar] [CrossRef]
- Blay, G. Le; Lacroix, C.; Zihler, A.; Fliss, I. In Vitro Inhibition Activity of Nisin A, Nisin Z, Pediocin PA-1 and Antibiotics against Common Intestinal Bacteria. Lett. Appl. Microbiol. 2007, 45, 252–257. [Google Scholar] [CrossRef]
- Kaur, S.; Kaur, S. Bacteriocins as Potential Anticancer Agents. Front. Pharmacol. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.E.; Ritchie, K.; Kamarajan, P.; Miao, D.; Kapila, Y.L. Nisin, an Apoptogenic Bacteriocin and Food Preservative, Attenuates HNSCC Tumorigenesis via CHAC1. Cancer Med. 2012, 1, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Kamarajan, P.; Hayami, T.; Matte, B.; Liu, Y.; Danciu, T.; Ramamoorthy, A.; Worden, F.; Kapila, S.; Kapila, Y. Nisin ZP, a Bacteriocin and Food Preservative, Inhibits Head and Neck Cancer Tumorigenesis and Prolongs Survival. PLoS One 2015, 10, 1–20. [Google Scholar] [CrossRef]
- Preet, S.; Bharati, S.; Panjeta, A.; Tewari, R.; Rishi, P. Effect of Nisin and Doxorubicin on DMBA-Induced Skin Carcinogenesis—a Possible Adjunct Therapy. Tumor Biol. 2015, 36, 8301–8308. [Google Scholar] [CrossRef] [PubMed]
- Villarante, K.I.; Elegado, F.B.; Iwatani, S.; Zendo, T.; Sonomoto, K.; de Guzman, E.E. Purification, Characterization and in Vitro Cytotoxicity of the Bacteriocin from Pediococcus Acidilactici K2a2-3 against Human Colon Adenocarcinoma (HT29) and Human Cervical Carcinoma (HeLa) Cells. World J. Microbiol. Biotechnol. 2011, 27, 975–980. [Google Scholar] [CrossRef]
- Kumar, B.; Balgir, P.P.; Kaur, B.; Mittu, B.; Chauhan, A. In Vitro Cytotoxicity of Native and Rec-Pediocin CP2 Against Cancer Cell Lines: A Comparative Study. Pharm. Anal. Acta 2012, 03, 2–6. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Nijnik, A.; Magrangeas-Janot, L.; Pasupuleti, M.; Thorson, L.; Ma, S.; Easton, D.M.; Bains, M.; et al. Manipulation of Innate Immunity by a Bacterial Secreted Peptide: Lantibiotic Nisin Z Is Selectively Immunomodulatory. Innate Immun. 2013, 19, 315–327. [Google Scholar] [CrossRef]
- De Pablo, M.A.; Gaforio, J.J.; Gallego, A.M.; Ortega, E.; Gálvez, A.M.; Alvarez De Cienfuegos López, G. Evaluation of Immunomodulatory Effects of Nisin-Containing Diets on Mice. FEMS Immunol. Med. Microbiol. 1999, 24, 35–42. [Google Scholar] [CrossRef]
- Villamil, L.; Figueras, A.; Novoa, B. Immunomodulatory Effects of Nisin in Turbot (Scophthalmus Maximus L.). Fish Shellfish Immunol. 2003, 14, 157–169. [Google Scholar] [CrossRef]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory Efficacy of Nisin-a Bacterial Lantibiotic Peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food Protein-Derived Bioactive Peptides: Production, Processing, and Potential Health Benefits. J. Food Sci. 2012, 77. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Bioactive Peptides: Production and Functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Del Mar Contreras, M.; Recio, I. Antihypertensive Peptides: Production, Bioavailability and Incorporation into Foods. Adv. Colloid Interface Sci. 2011, 165, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Daliri, E.B.; Lee, B.H.; Park, B.; Kim, S.; Oh, D.-H. Antihypertensive Peptides from Whey Proteins Fermented by Lactic Acid Bacteria. Food Sci. Biotechnol. 2018, 27, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.E.; Park, S.Y.; Atwal, A.; Gibbs, B.F.; Lee, B.H. Angiotensin I-converting Enzyme (Ace) Inhibitory Peptides from Whey Fermented by Lactobacillus Species. J. Food Biochem. 2009, 33, 587–602. [Google Scholar] [CrossRef]
- Fuglsang, A.; Rattray, F.P.; Nilsson, D.; Nyborg, N.C.B. Lactic Acid Bacteria: Inhibition of Angiotensin Converting Enzyme in Vitro and in Vivo. Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 2003, 83, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Miralles, B.; Amigo, L.; Ramos, M.; Recio, I. Identification of Antioxidant and ACE-Inhibitory Peptides in Fermented Milk. J. Sci. Food Agric. 2005, 85, 1041–1048. [Google Scholar] [CrossRef]
- Yamamoto, N.; Akino, A.; Takano, T. Antihypertensive Effects of Different Kinds of Fermented Milk in Spontaneously Hypertensive Rats. Biosci. Biotechnol. Biochem. 1994, 58, 776–778. [Google Scholar] [CrossRef]
- Seppo, L.; Jauhiainen, T.; Poussa, T.; Korpela, R. A Fermented Milk High in Bioactive Peptides Has a Blood Pressure-Lowering Effect in Hypertensive Subjects. Am. J. Clin. Nutr. 2003, 77, 326–330. [Google Scholar] [CrossRef]
- Nejati, F.; Rizzello, C.G.; Di Cagno, R.; Sheikh-Zeinoddin, M.; Diviccaro, A.; Minervini, F.; Gobbetti, M. Manufacture of a Functional Fermented Milk Enriched of Angiotensin-I Converting Enzyme (ACE)-Inhibitory Peptides and γ-Amino Butyric Acid (GABA). LWT - Food Sci. Technol. 2013, 51, 183–189. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.Á.; Ramos, M.; Recio, I. Angiotensin-Converting Enzyme-Inhibitory Peptides in Manchego Cheeses Manufactured with Different Starter Cultures. Int. Dairy J. 2002, 12, 697–706. [Google Scholar] [CrossRef]
- Ryhänen, E.L.; Pihlanto-Leppälä, A.; Pahkala, E. A New Type of Ripened, Low-Fat Cheese with Bioactive Properties. Int. Dairy J. 2001, 11, 441–447. [Google Scholar] [CrossRef]
- Korhonen, H. Milk-Derived Bioactive Peptides: From Science to Applications. J. Funct. Foods 2009, 1, 177–187. [Google Scholar] [CrossRef]
- Ahn, J.E.; Park, S.Y.; Atwal, A.; Gibbs, B.F.; Lee, B.H. Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides from Whey Fermented by Lactobacillus Species. J. Food Biochem. 2009, 33, 587–602. [Google Scholar] [CrossRef]
- Wang, H.K.; Dong, C.; Chen, Y.F.; Cui, L.M.; Zhang, H.P. A New Probiotic Cheddar Cheese with High ACE-Inhibitory Activity and γ-Aminobutyric Acid Content Produced with Koumiss-Derived Lactobacillus Casei Zhang. Food Technol. Biotechnol. 2010, 48, 62–70. [Google Scholar]
- Minervini, F.; Bilancia, M.T.; Siragusa, S.; Gobbetti, M.; Caponio, F. Fermented Goats’ Milk Produced with Selected Multiple Starters as a Potentially Functional Food. Food Microbiol. 2009, 26, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhao, S.; Wang, H.; Cai, C.; Chen, Y.; Zhang, H. ACE-Inhibitory Activity and Gamma-Aminobutyric Acid Content of Fermented Skim Milk by Lactobacillus Helveticus Isolated from Xinjiang Koumiss in China. Eur. Food Res. Technol. 2009, 228, 607–612. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; González-Velázquez, D.A.; Torres-Llanez, M.J.; Martínez-Porchas, M.; García-Sifuentes, C.O.; González-Córdova, A.F.; Vallejo-Córdoba, B. Cheese Whey Fermentation by Its Native Microbiota: Proteolysis and Bioactive Peptides Release with ACE-Inhibitory Activity. Fermentation 2020, 6, 1–12. [Google Scholar] [CrossRef]
- Hayes, M.; Stanton, C.; Slattery, H.; O’Sullivan, O.; Hill, C.; Fitzgerald, G.F.; Ross, R.P. Casein Fermentate of Lactobacillus Animalis DPC6134 Contains a Range of Novel Propeptide Angiotensin-Converting Enzyme Inhibitors. Appl. Environ. Microbiol. 2007, 73, 4658–4667. [Google Scholar] [CrossRef]
- Raveschot, C.; Deracinois, B.; Bertrand, E.; Flahaut, C.; Frémont, M.; Drider, D.; Dhulster, P.; Cudennec, B.; Coutte, F. Integrated Continuous Bioprocess Development for ACE-Inhibitory Peptide Production by Lactobacillus Helveticus Strains in Membrane Bioreactor. Front. Bioeng. Biotechnol. 2020, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pessione, E.; Cirrincione, S. Bioactive Molecules Released in Food by Lactic Acid Bacteria: Encrypted Peptides and Biogenic Amines. Front. Microbiol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Aihara, K.; Nakamura, Y.; Kajimoto, O.; Hirata, H.; Takahashi, R. Effect of Powdered Fermented Milk with Lactobacillus Helveticus on Subjects with High-Normal Blood Pressure or Mild Hypertension. J. Am. Coll. Nutr. 2005, 24, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Jiang, Y.; Yang, W.; Du, F.; Yao, Y.; Shi, C.; Wang, C. Effective Treatment of Hypertension by Recombinant Lactobacillus Plantarum Expressing Angiotensin Converting Enzyme Inhibitory Peptide. Microb. Cell Fact. 2015, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hata, Y.; Yamamoto, M.; Ohni, M.; Nakajima, K.; Nakamura, Y.; Takano, T. A Placebo-Controlled Study of the Effect of Sour Milk on Blood Pressure in Hypertensive Subjects. Am. J. Clin. Nutr. 1996, 64, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Jauhiainen, T.; Rönnback, M.; Vapaatalo, H.; Wuolle, K.; Kautiainen, H.; Groop, P.H.; Korpela, R. Long-Term Intervention with Lactobacillus Helveticus Fermented Milk Reduces Augmentation Index in Hypertensive Subjects. Eur. J. Clin. Nutr. 2010, 64, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; Nonaka, A.; Matsushita, A.; Uchida, N.; Ohki, K.; Asakura, M.; Kitakaze, M. Milk Casein-Derived Tripeptides, VPP and IPP Induced NO Production in Cultured Endothelial Cells and Endothelium-Dependent Relaxation of Isolated Aortic Rings. Heart Vessels 2011, 26, 549–556. [Google Scholar] [CrossRef]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two Antioxidative Lactobacilli Strains as Promising Probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef]
- Lin, M.Y.; Yen, C.L. Reactive Oxygen Species and Lipid Peroxidation Product-Scavenging Ability of Yogurt Organisms. J. Dairy Sci. 1999, 82, 1629–1634. [Google Scholar] [CrossRef]
- Sah, B.N.P.; Vasiljevic, T.; McKechnie, S.; Donkor, O.N. Effect of Probiotics on Antioxidant and Antimutagenic Activities of Crude Peptide Extract from Yogurt. Food Chem. 2014, 156, 264–270. [Google Scholar] [CrossRef]
- Shu, G.; Shi, X.; Chen, L.; Kou, J.; Meng, J.; Chen, H. Antioxidant Peptides from Goat Milk Fermented by Lactobacillus Casei L61: Preparation, Optimization, and Stability Evaluation in Simulated Gastrointestinal Fluid. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, T.; Pihlanto, A.; Akkanen, S.; Korhonen, H. Development of Antioxidant Activity in Milk Whey during Fermentation with Lactic Acid Bacteria. 2007, 102, 106–115. [CrossRef]
- Leclerc, P.L.; Gauthier, S.F.; Bachelard, H.; Santure, M.; Roy, D. Antihypertensive Activity of Casein-Enriched Milk Fermented by Lactobacillus Helveticus. Int. Dairy J. 2002, 12, 995–1004. [Google Scholar] [CrossRef]
- Fardet, A.; Rock, E. In Vitro and in Vivo Antioxidant Potential of Milks, Yoghurts, Fermented Milks and Cheeses: A Narrative Review of Evidence. Nutr. Res. Rev. 2018, 31, 52–70. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Kapila, R.; Kapasiya, M.; Saliganti, V.; Dass, G.; Kapila, S. Dietary Supplementation of Milk Fermented with Probiotic Lactobacillus Fermentum Enhances Systemic Immune Response and Antioxidant Capacity in Aging Mice. Nutr. Res. 2014, 34, 968–981. [Google Scholar] [CrossRef]
- Bay, B.H.; Lee, Y.K.; Tan, B.K.H.; Ling, E.A. Lipid Peroxidative Stress and Antioxidative Enzymes in Brains of Milk-Supplemented Rats. Neurosci. Lett. 1999, 277, 127–130. [Google Scholar] [CrossRef]
- Zommara, M.; Tachibana, N.; Sakono, M.; Suzuki, Y.; Oda, T.; Hashiba, H.; Imaizumi, K. Whey from Cultured Skim Milk Decreases Serum Cholesterol and Increases Antioxidant Enzymes in Liver and Red Blood Cells in Rats. Nutr. Res. 1996, 16, 293–302. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Ram, S.; Mitra, M.; Shah, F.; Rani, S.; Mishra, S. Bacteria as an Alternate Biofactory for Carotenoid Production: A Review of Its Applications, Opportunities and Challenges. J. Funct. Foods 2020, 67, 103867. [Google Scholar] [CrossRef]
- Yoto, A.; Murao, S.; Motoki, M.; Yokoyama, Y.; Horie, N.; Takeshima, K.; Masuda, K.; Kim, M.; Yokogoshi, H. Oral Intake of γ-Aminobutyric Acid Affects Mood and Activities of Central Nervous System during Stressed Condition Induced by Mental Tasks. Amino Acids 2012, 43, 1331–1337. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Inoue, K.; Shirai, T.; Ochiai, H.; Kasao, M.; Hayakawa, K.; Kimura, M.; Sansawa, H. Blood-Pressure-Lowering Effect of a Novel Fermented Milk Containing γ-Aminobutyric Acid (GABA) in Mild Hypertensives. Eur. J. Clin. Nutr. 2003, 57, 490–495. [Google Scholar] [CrossRef]
- Thaker, P.H.; Yokoi, K.; Jennings, N.B.; Li, Y.; Rebhun, R.B.; Rousseau, D.L.; Fan, D.; Sood, A.K. Inhibition of Experimental Colon Cancer Metastasis by the GABA-Receptor Agonist Nembutal. Cancer Biol. Ther. 2005, 4, 753–758. [Google Scholar] [CrossRef]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of Oral Gamma-Aminobutyric Acid (GABA) Administration on Stress and Sleep in Humans: A Systematic Review. Front. Neurosci. 2020, 14. [Google Scholar] [CrossRef]
- Huang, J.; Mei, L.H.; Wu, H.; Lin, D.Q. Biosynthesis of γ-Aminobutyric Acid (GABA) Using Immobilized Whole Cells of Lactobacillus Brevis. World J. Microbiol. Biotechnol. 2007, 23, 865–871. [Google Scholar] [CrossRef]
- Binh, T.T.T.; Ju, W.T.; Jung, W.J.; Park, R.D. Optimization of γ-Amino Butyric Acid Production in a Newly Isolated Lactobacillus Brevis. Biotechnol. Lett. 2014, 36, 93–98. [Google Scholar] [CrossRef]
- Li, H.; Qiu, T.; Huang, G.; Cao, Y. Production of Gamma-Aminobutyric Acid by Lactobacillus Brevis NCL912 Using Fed-Batch Fermentation. Microb. Cell Fact. 2010, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Huang, J.; Hu, S.; Zhao, W.; Yao, S.; Mei, L. A Two-Stage PH and Temperature Control with Substrate Feeding Strategy for Production of Gamma-Aminobutyric Acid by Lactobacillus Brevis CGMCC 1306. Chinese J. Chem. Eng. 2013, 21, 1190–1194. [Google Scholar] [CrossRef]
- Villegas, J.M.; Brown, L.; Savoy de Giori, G.; Hebert, E.M. Optimization of Batch Culture Conditions for GABA Production by Lactobacillus Brevis CRL 1942, Isolated from Quinoa Sourdough. LWT - Food Sci. Technol. 2016, 67, 22–26. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, X.; Fu, J.; Wang, S.; Chen, Y.; Chang, K.; Li, H. Substrate Sustained Release-Based High Efficacy Biosynthesis of GABA by Lactobacillus Brevis NCL912. Microb. Cell Fact. 2018, 17, 1–8. [Google Scholar] [CrossRef]
- Wu, C.; Hsuesh, Y.-H.; Kuo, J.-M.; Liu, S.-J. Characterization of a Potential Probiotic Lactobacillus Brevis RK03 and Efficient Production of γ -Aminobutyric Acid in Batch Fermentation. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Cataldo, P.G.; Villegas, M.; Savoy, G.; Giori, D.; Saavedra, L.; Hebert, E.M. Enhancement of γ -Aminobutyric Acid (GABA ) Production by Lactobacillus Brevis CRL 2013 Based on Carbohydrate Fermentation. Int. J. Food Microbiol. 2020, 333. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-I.; Lee, J.-W.; Park, S.; Lee, M.-Y.; Ji, G.-E.; Park, M.-S.; Heo, T.-R. Improvement of Gamma-Aminobutyric Acid Production Using Cell Entrapment of Lactobacillus Brevis GABA 057. J. Microbiol. Biotechnol. 2005, 16, 562–568. [Google Scholar]
- Alizadeh Behbahani, B.; Jooyandeh, H.; Falah, F.; Vasiee, A. Gamma-Aminobutyric Acid Production by Lactobacillus Brevis A3: Optimization of Production, Antioxidant Potential, Cell Toxicity, and Antimicrobial Activity. Food Sci. Nutr. 2020, 8, 5330–5339. [Google Scholar] [CrossRef] [PubMed]
- Park, K.B.; Oh, S.H. Production of Yogurt with Enhanced Levels of Gamma-Aminobutyric Acid and Valuable Nutrients Using Lactic Acid Bacteria and Germinated Soybean Extract. Bioresour. Technol. 2007, 98, 1675–1679. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.J.; Nam, Y. Do; Park, S.L.; Lee, S.Y.; Yi, S.H.; Lim, S. Il γ-Aminobutyric Acid Production in Skim Milk Co-Fermented with Lactobacillus Brevis 877G and Lactobacillus Sakei 795. Food Sci. Biotechnol. 2013, 22, 751–755. [Google Scholar] [CrossRef]
- Cho, Y.R.; Chang, J.Y.; Chang, H.C. Production of Gamma Amino-Bytyric Acid by Lactobacillus Buchneri Isolated from Kimchi and Its Neuroprotective Effect on Neuronal Cells. J. Microbiol. Biotechnol. 2007, 17, 104–109. [Google Scholar] [PubMed]
- Park, K.-B.; Oh, S.-H. Isolation and Characterization of Lactobacillus Buchneri Strains with High GABA Producing Capacity from Naturally Aged Cheese.Pdf. Food Sci. Biotechnol. 2006, 15, 86–90. [Google Scholar]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-Aminobutyric Acid by Lactic Acid Bacteria Isolated from a Variety of Italian Cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef]
- Komatsuzaki, N.; Shima, J.; Kawamoto, S.; Momose, H.; Kimura, T. Production of γ-Aminobutyric Acid (GABA) by Lactobacillus Paracasei Isolated from Traditional Fermented Foods. Food Microbiol. 2005, 22, 497–504. [Google Scholar] [CrossRef]
- Servili, M.; Rizzello, C.G.; Taticchi, A.; Esposto, S.; Urbani, S.; Mazzacane, F.; Di Maio, I.; Selvaggini, R.; Gobbetti, M.; Di Cagno, R. Functional Milk Beverage Fortified with Phenolic Compounds Extracted from Olive Vegetation Water, and Fermented with Functional Lactic Acid Bacteria. Int. J. Food Microbiol. 2011, 147, 45–52. [Google Scholar] [CrossRef]
- Tung, Y.T.; Lee, B.H.; Liu, C.F.; Pan, T.M. Optimization of Culture Condition for ACEI and GABA Production by Lactic Acid Bacteria. J. Food Sci. 2011, 76, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, D.H.; Kang, H.J.; Shin, M.; Yang, S.Y.; Yang, J.; Jung, Y.H. Enhanced Production of γ-Aminobutyric Acid (GABA) Using Lactobacillus Plantarum EJ2014 with Simple Medium Composition. Lwt 2021, 137, 110443. [Google Scholar] [CrossRef]
- Ratanaburee, A.; Kantachote, D. Enhancement of γ -Aminobutyric Acid in a Fermented Red Seaweed Beverage by Starter Culture Lactobacillus Plantarum DW12. Electron. J. Biotechnol. 2011, 14. [Google Scholar] [CrossRef]
- Park, S.; Kim, K.; Lee, M.; Lim, S. Physiological Characteristics and GABA Production of Lactobacillus Plantarum K255 Isolated from Kimchi. Korean J. Food Sci. Anim. Resour. 2013, 33, 595–602. [Google Scholar] [CrossRef]
- Shan, Y.; Man, C.X.; Han, X.; Li, L.; Guo, Y.; Deng, Y.; Li, T.; Zhang, L.W.; Jiang, Y.J. Evaluation of Improved γ-Aminobutyric Acid Production in Yogurt Using Lactobacillus Plantarum NDC75017. J. Dairy Sci. 2015, 98, 2138–2149. [Google Scholar] [CrossRef] [PubMed]
- Tajabadi, N.; Ebrahimpour, A.; Baradaran, A.; Rahim, R.A.; Mahyudin, N.A.; Yazid, M.; Manap, A.; Bakar, F.A.; Saari, N. Optimization of γ-Aminobutyric Acid Production by Lactobacillus Plantarum Taj-Apis362 from Honeybees. Molecules 2015, 20, 6654–6669. [Google Scholar] [CrossRef] [PubMed]
- Cagno, R. Di; Mazzacane, F.; Rizzello, C.G.; Angelis, M. De; Giuliani, G.; Meloni, M.; Servi, B. De; Gobbetti, M. Synthesis of γ -Aminobutyric Acid ( GABA ) by Lactobacillus Plantarum DSM19463 : Functional Grape Must Beverage and Dermatological Applications. Appl. Microb. cell Physiol. 2010, 86, 731–741. [Google Scholar] [CrossRef]
- Park, S.; Lee, J.; Lim, S. The Probiotic Characteristics and GABA Production of Lactobacillus Plantarum K154 Isolated from Kimchi. Food Sci. Biotechnol. 2014, 23, 1951–1957. [Google Scholar] [CrossRef]
- Karimian, E.; Moayedi, A.; Khomeiri, M.; Aalami, M.; Mahoonak, A.S. Application of High-GABA Producing Lactobacillus Plantarum Isolated from Traditional Cabbage Pickle in the Production of Functional Fermented Whey-Based Formulate. J. Food Meas. Charact. 2020, 14, 3408–3416. [Google Scholar] [CrossRef]
- Lin, Q. Submerged Fermentation of Lactobacillus Rhamnosus YS9 for γ-Aminobutyric Acid (GABA) Production. Brazilian J. Microbiol. 2013, 44, 183–187. [Google Scholar] [CrossRef]
- Kook, M.C.; Seo, M.J.; Cheigh, C.I.; Lee, S.J.; Pyun, Y.R.; Park, H. Enhancement of γ-Amminobutyric Acid Production by Lactobacillus Sakei B2-16 Expressing Glutamate Decarboxylase from Lactobacillus Plantarum ATCC 14917. J. Appl. Biol. Chem. 2010, 53, 816–820. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lü, F.X.; Lu, Z.X.; Bie, X.M.; Jiao, Y.; Sun, L.J.; Yu, B. Production of γ-Aminobutyric Acid by Streptococcus Salivarius Subsp. Thermophilus Y2 under Submerged Fermentation. Amino Acids 2008, 34, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.M.; O’Callaghan, T.F.; O’Connor, P.M.; Ross, R.P.; Stanton, C. Streptococcus Thermophilus APC151 Strain Is Suitable for the Manufacture of Naturally Gaba-Enriched Bioactive Yogurt. Front. Microbiol. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Wu, Q.; Law, Y.S.; Shah, N.P. Dairy Streptococcus Thermophilus Improves Cell Viability of Lactobacillus Brevis NPS-QW-145 and Its γ-Aminobutyric Acid Biosynthesis Ability in Milk. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kook, M.C.; Cho, S.C. Production of GABA (Gamma Amino Butyric Acid) by Lactic Acid Bacteria. Korean J. Food Sci. Anim. Resour. 2013, 33, 377–389. [Google Scholar] [CrossRef]
- Zhuang, K.; Jiang, Y.; Feng, X.; Li, L.; Dang, F.; Zhang, W.; Man, C. Transcriptomic Response to GABA-Producing Lactobacillus Plantarum CGMCC 1.2437T Induced by L-MSG. PLoS One 2018, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sanchart, C.; Watthanasakphuban, N.; Boonseng, O.; Nguyen, T.H.; Haltrich, D.; Maneerat, S. Tuna Condensate as a Promising Low-Cost Substrate for Glutamic Acid and GABA Formation Using Candida Rugosa and Lactobacillus Futsaii. Process Biochem. 2018, 70, 29–35. [Google Scholar] [CrossRef]
- Lee, E.J.; Lee, S.P. Novel Bioconversion of Sodium Glutamate to γ-Amino Butyric Acid by Co-Culture of Lactobacillus Plantarum K154 in Ceriporia Lacerata Culture Broth. Food Sci. Biotechnol. 2014, 23, 1997–2005. [Google Scholar] [CrossRef]
- Diez-Gutiérrez, L.; San Vicente, L.; Luis, L.J.; Villarán, M. del C.; Chávarri, M. Gamma-Aminobutyric Acid and Probiotics: Multiple Health Benefits and Their Future in the Global Functional Food and Nutraceuticals Market. J. Funct. Foods 2020, 64, 103669. [Google Scholar] [CrossRef]
- Hayakawa, K.; Kimura, M.; Kamata, K. Mechanism Underlying γ-Aminobutyric Acid-Induced Antihypertensive Effect in Spontaneously Hypertensive Rats. Eur. J. Pharmacol. 2002, 438, 107–113. [Google Scholar] [CrossRef]
- Hayakawa, K.; Kimura, M.; Kasaha, K.; Matsumoto, K.; Sansawa, H.; Yamori, Y. Effect of a γ-Aminobutyric Acid-Enriched Dairy Product on the Blood Pressure of Spontaneously Hypertensive and Normotensive Wistar–Kyoto Rats. Br. J. Nutr. 2004, 92, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A. V.; Nutt, D.J. Role of GABA in Anxiety and Depression. Depress. Anxiety 2007, 24, 495–517. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M. V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus Strain Regulates Emotional Behavior and Central GABA Receptor Expression in a Mouse via the Vagus Nerve. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and Immunity Enhancement Effects of γ-Aminobutyric Acid (GABA) Administration in Humans. BioFactors 2006, 26, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Takishima, T.; Kometani, T.; Yokogoshi, H. Psychological Stress-Reducing Effect of Chocolate Enriched with γ-Aminobutyric Acid (GABA) in Humans: Assessment of Stress Using Heart Rate Variability and Salivary Chromogranin A. Int. J. Food Sci. Nutr. 2009, 60, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Yamatsu, A.; Yamashita, Y.; Pandharipande, T.; Maru, I.; Kim, M. Effect of Oral γ-Aminobutyric Acid (GABA) Administration on Sleep and Its Absorption in Humans. Food Sci. Biotechnol. 2016, 25, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Kanehira, T.; Nakamura, Y.; Nakamura, K.; Horie, K.; Horie, N.; Furugori, K.; Sauchi, Y.; Yokogoshi, H. Relieving Occupational Fatigue by Consumption of a Beverage Containing γ-Amino Butyric Acid. J. Nutr. Sci. Vitaminol. (Tokyo). 2011, 57, 9–15. [Google Scholar] [CrossRef]
- Xie, Z.; Xia, S.; Le, G. Gamma-Aminobutyric Acid Improves Oxidative Stress and Function of the Thyroid in High-Fat Diet. J. Funct. Foods 2014, 8, 76–86. [Google Scholar] [CrossRef]
- Adeghate, E.; Ponery, A.S. GABA in the Endocrine Pancreas : Cellular Localization and Function in Normal and Diabetic Rats. Tissue Cell 2002, 34, 1–6. [Google Scholar] [CrossRef]
- Soltani, N.; Qiu, H.; Aleksic, M.; Glinka, Y.; Zhao, F.; Liu, R.; Li, Y.; Zhang, N.; Chakrabarti, R.; Ng, T.; et al. GABA Exerts Protective and Regenerative Effects on Islet Beta Cells and Reverses Diabetes. PNAS 2011, 108, 4–9. [Google Scholar] [CrossRef]
- Schuller, H.M.; Al-wadei, H.A.N.; Majidi, M. Gamma-Aminobutyric Acid, a Potential Tumor Suppressor for Small Airway-Derived Lung Adenocarcinoma. Carcinogenesis 2008, 29, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Du, A.; Xiong, Y.; Jiang, J.; Zhang, Y.; Tian, Z. γ -Aminobutyric Acid Inhibits the Proliferation and Increases Oxaliplatin Sensitivity in Human Colon Cancer Cells. Tumor Biol. 2016, 37, 14885–14894. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.; Oh, S. Effects of Germinated Brown Rice Extracts with Enhanced Levels of GABA on Cancer Cell Proliferation and Apoptosis. J. Med. Food 2004, 7, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Eggersdorfer, M.; Wyss, A. Carotenoids in Human Nutrition and Health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Sowmya, R.; Sachindra, N.M. Carotenoid Production by Formosa Sp. KMW, a Marine Bacteria of Flavobacteriaceae Family: Influence of Culture Conditions and Nutrient Composition. Biocatal. Agric. Biotechnol. 2015, 4, 559–567. [Google Scholar] [CrossRef]
- Giuffrida, D.; Sutthiwong, N.; Dugo, P.; Donato, P.; Cacciola, F.; Girard-Valenciennes, E.; Le Mao, Y.; Monnet, C.; Fouillaud, M.; Caro, Y.; et al. Characterisation of the C50 Carotenoids Produced by Strains of the Cheese-Ripening Bacterium Arthrobacter Arilaitensis. Int. Dairy J. 2016, 55, 10–16. [Google Scholar] [CrossRef]
- Turpin, W.; Renaud, C.; Avallone, S.; Hammoumi, A.; Guyot, J.P.; Humblot, C. PCR of CrtNM Combined with Analytical Biochemistry: An Efficient Way to Identify Carotenoid Producing Lactic Acid Bacteria. Syst. Appl. Microbiol. 2016, 39, 115–121. [Google Scholar] [CrossRef]
- Garrido-Fernández, J.; Maldonado-Barragán, A.; Caballero-Guerrero, B.; Hornero-Méndez, D.; Ruiz-Barba, J.L. Carotenoid Production in Lactobacillus Plantarum. Int. J. Food Microbiol. 2010, 140, 34–39. [Google Scholar] [CrossRef]
- Elsanhoty, R.M.; Al-Turki, I.A.; Ramadan, M.F. Screening of Medium Components by Plackett-Burman Design for Carotenoid Production Using Date (Phoenix Dactylifera) Wastes. Ind. Crops Prod. 2012, 36, 313–320. [Google Scholar] [CrossRef]
- Hagi, T.; Kobayashi, M.; Nomura, M. Aerobic Conditions Increase Isoprenoid Biosynthesis Pathway Gene Expression Levels for Carotenoid Production in Enterococcus Gilvus. FEMS Microbiol. Lett. 2015, 362, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Simova, E.D.; Frengova, G.I.; Beshkova, D.M. Effect of Aeration on the Production of Carotenoid Pigments by Rhodotorula Rubra-Lactobacillus Casei Subsp. Casei Co-Cultures in Whey Ultrafiltrate. Zeitschrift für Naturforsch. C 2003, 58, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Simova, E.D.; Frengova, G.I.; Beshkova, D.M. Synthesis of Carotenoids by Rhodotorula Rubra GED8 Co-Cultured with Yogurt Starter Cultures in Whey Ultrafiltrate. J. Ind. Microbiol. Biotechnol. 2004, 31, 115–121. [Google Scholar] [CrossRef]
- Frengova, G.I.; Simova, E.D.; Beshkova, D.M. β-Carotene-Rich Carotenoid-Protein Preparation and Exopolysaccharide Production by Rhodotorula Rubra GED8 Grown with a Yogurt Starter Culture. Zeitschrift für Naturforsch. C 2006, 61, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Frengova, G.I.; Simova, E.D.; Beshkova, D.M. Effect of Temperature Changes in the Production of Yeast Pigments Co-Cultivated with Lacto-Acid Bacteria in Whey Ultrafiltrate. Biotechnol. Lett. 1995, 17, 1001–1006. [Google Scholar] [CrossRef]
- Rodriguez-concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-gomez, L.; Hornero-mendez, D.; Limon, M.C.; Meléndez-martínez, A.J.; Olmedilla-alonso, B.; Palou, A.; et al. A Global Perspective on Carotenoids: Metabolism, Biotechnology, and Benefits for Nutrition and Health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [PubMed]
- WHO The World Health Report 2002: Reducing Risks, Promoting Healthy Life; 2002; 2002.
- Hirano, R.; Iwamoto, T.; Hosodad, K.; Kurata, H.; Matsumoto, A.; Miki, W.; Kamiyama, M.; Itakura, H.; Yamamoto, S.; Kondo, K. Inhibition of Low-Density Lipoprotein Oxidation by Astaxanthin Tamami of Medicine Introduction Oxidative Modification of Low-Density Lipoprotein ( LDL ) Has Been Implicated in the Pathogenesis of Atheroscle- Rosis ( 1, 2 ). The Rapid Uptake of Oxidative. J. Atheroscler. Thromb. 2000, 7, 216–222. [Google Scholar]
- Yoshida, H.; Yanai, H.; Ito, K.; Tomono, Y.; Koikeda, T.; Tsukahara, H.; Tada, N. Administration of Natural Astaxanthin Increases Serum HDL-Cholesterol and Adiponectin in Subjects with Mild Hyperlipidemia. Atherosclerosis 2010, 209, 520–523. [Google Scholar] [CrossRef]
- Hennekens, C.H.; Buring, J.E.; Manson, J.E.; Stampfer, M.; Rosner, B.; Cook, N.R.; Belanger, C.; LaMotte, F.; Gaziano, J.M.; Ridker, P.M.; et al. Lack of Effect of Long-Term Supplementation with Beta Carotene on the Incidence of Malignant Neoplasms and Cardiovascular Disease. N. Engl. J. Med. 1996, 334, 1145–1149. [Google Scholar] [CrossRef]
- Greenberg, E.R.; Baron, J.A.; Karagas, M.R.; Stukel, T.A.; Nierenberg, D.W.; Stevens, M.M.; Mandel, J.S.; Haile, R.W. Mortality Associated with Low Plasma Concentration of Beta Carotene and the Effect of Oral Supplementation. J. Am. Med. Assoc. 1996, 275, 699–703. [Google Scholar] [CrossRef]
- Yamaguchi, M. β-Cryptoxanthin Stimulates Bone Formation and Inhibits Bone Resorption in Tissue Culture in Vitro. Mol. Cell. Biochem. 2004, 258, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Uchiyama, S. Effect of Carotenoid on Calcium Content and Alkaline Phosphatase Activity in Rat Femoral Tissues in Vitro: The Unique Anabolic Effect of β-Cryptoxanthin. Biol. Pharm. Bull. 2003, 26, 1188–1191. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, S.; Sumida, T.; Yamaguchi, M. Oral Administration of β-Cryptoxanthin Induces Anabolic Effects on Bone Components in the Femoral Tissues of Rats in Vivo. Biol. Pharm. Bull. 2004, 27, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; Rao, A.V.; Rao, L.G. Lycopene II - Effect on Osteoblasts: The Carotenoid Lycopene Stimulates Cell Proliferation and Alkaline Phosphatase Activity of SaOS-2 Cells. J. Med. Food 2003, 6, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Hannan, M.T.; Blumberg, J.; Cupples, L.A.; Kiel, D.P.; Tucker, K.L. Inverse Association of Carotenoid Intakes with 4-y Change in Bone Mineral Density in Elderly Men and Women: The Framingham Osteoporosis Study. Am. J. Clin. Nutr. 2009, 89, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Mounien, L.; Tourniaire, F. Anti-Obesity Effect of Carotenoids: Direct Impact on Adipose Tissue and Adipose Tissue-Driven Indirect Effect. Nutrients 2019, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Canas, J.A.; Lochrie, A.; McGowan, A.G.; Hossain, J.; Schettino, C.; Balagopal, P.B. Effects of Mixed Carotenoids on Adipokines and Abdominal Adiposity in Children: A Pilot Study. J. Clin. Endocrinol. Metab. 2017, 102, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Kakutani, R.; Hokari, S.; Nishino, A.; Ichihara, T.; Sugimoto, K.; Takaha, T.; Kuriki, T.; Maoka, T. Effect of Oral Paprika Xanthophyll Intake on Abdominal Fat in Healthy Overweight Humans: A Randomized, Double-Blind, Placebo-Controlled Study. J. Oleo Sci. 2018, 67, 1149–1162. [Google Scholar] [CrossRef]
- Grodstein, F.; Kang, J.H.; Glynn, R.J.; Cook, N.R.; Gaziano, J.M. A Randomized Trial of Beta Carotene Supplementation and Cognitive Function in Men: The Physicians’ Health Study II. Arch. Intern. Med. 2007, 167, 2184–2190. [Google Scholar] [CrossRef]
- Otaegui-Arrazola, A.; Amiano, P.; Elbusto, A.; Urdaneta, E.; Martínez-Lage, P. Diet, Cognition, and Alzheimer’s Disease: Food for Thought. Eur. J. Nutr. 2014, 53, 1–23. [Google Scholar] [CrossRef]
- Devore, E.E.; Grodstein, F.; Van Rooij, F.J.A.; Hofman, A.; Stampfer, M.J.; Witteman, J.C.M.; Breteler, M.M.B. Dietary Antioxidants and Long-Term Risk of Dementia. Arch. Neurol. 2010, 67, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.L.; Dawsey, S.M.; Kamangar, F.; Fan, J.H.; Abnet, C.C.; Sun, X. Di; Johnson, L.L.; Gail, M.H.; Dong, Z.W.; Yu, B.; et al. Total and Cancer Mortality after Supplementation with Vitamins and Minerals: Follow-up of the Linxian General Population Nutrition Intervention Trial. J. Natl. Cancer Inst. 2009, 101, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.R.; Lee, I.; Manson, J.E.; Buring, J.E.; Hennekens, C.H. Effects of Beta-Carotene Supplementation on Cancer Incidence by Baseline Characteristics in the Physicians ’ Health Study (United States). Cancer Causes Control 2000, 11, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Albanes, D.; Heinonen, O.P.; Huttunen, J.K.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Haapakoski, J.; Rautalahti, M.; Hartman, A.M.; Palmgren, J.; et al. Effects Cancer Cancer of A-Tocopherol and n-Carotene Incidence in the Alpha-Tocopherol Prevention Study13 Supplements On. Am. Soc. Clin. Nutr. 1995, 62, 1–4. [Google Scholar]
- Li, J.Y.; Taylor, P.R.; Li, B.; Dawsey, S.; Wang, G.Q.; Ershow, A.G.; Guo, W.; Liu, S.F.; Yang, C.S.; Shen, Q.; et al. Nutrition Intervention Trials in Linxian, China: Multiple Vitamin/Mineral Supplementation, Cancer Incidence, and Disease-Specific Mortality among Adults with Esophageal Dysplasia. J. Natl. Cancer Inst. 1993, 85, 1492–1498. [Google Scholar] [CrossRef]
Figure 1.
Examples of functional dairy products including milk, yoghurt, cheese and milk powder (created with Bing Image Creator).
Figure 1.
Examples of functional dairy products including milk, yoghurt, cheese and milk powder (created with Bing Image Creator).

Figure 2.
Total amount of dairy products sold in Europe in euros billions (€) from 2014-2023. The data include milk, yoghurt, dairy-based and soy-based desserts, cream, butter, and spreadable fats. The figure was made with data obtained from MarkeLine.
Figure 2.
Total amount of dairy products sold in Europe in euros billions (€) from 2014-2023. The data include milk, yoghurt, dairy-based and soy-based desserts, cream, butter, and spreadable fats. The figure was made with data obtained from MarkeLine.
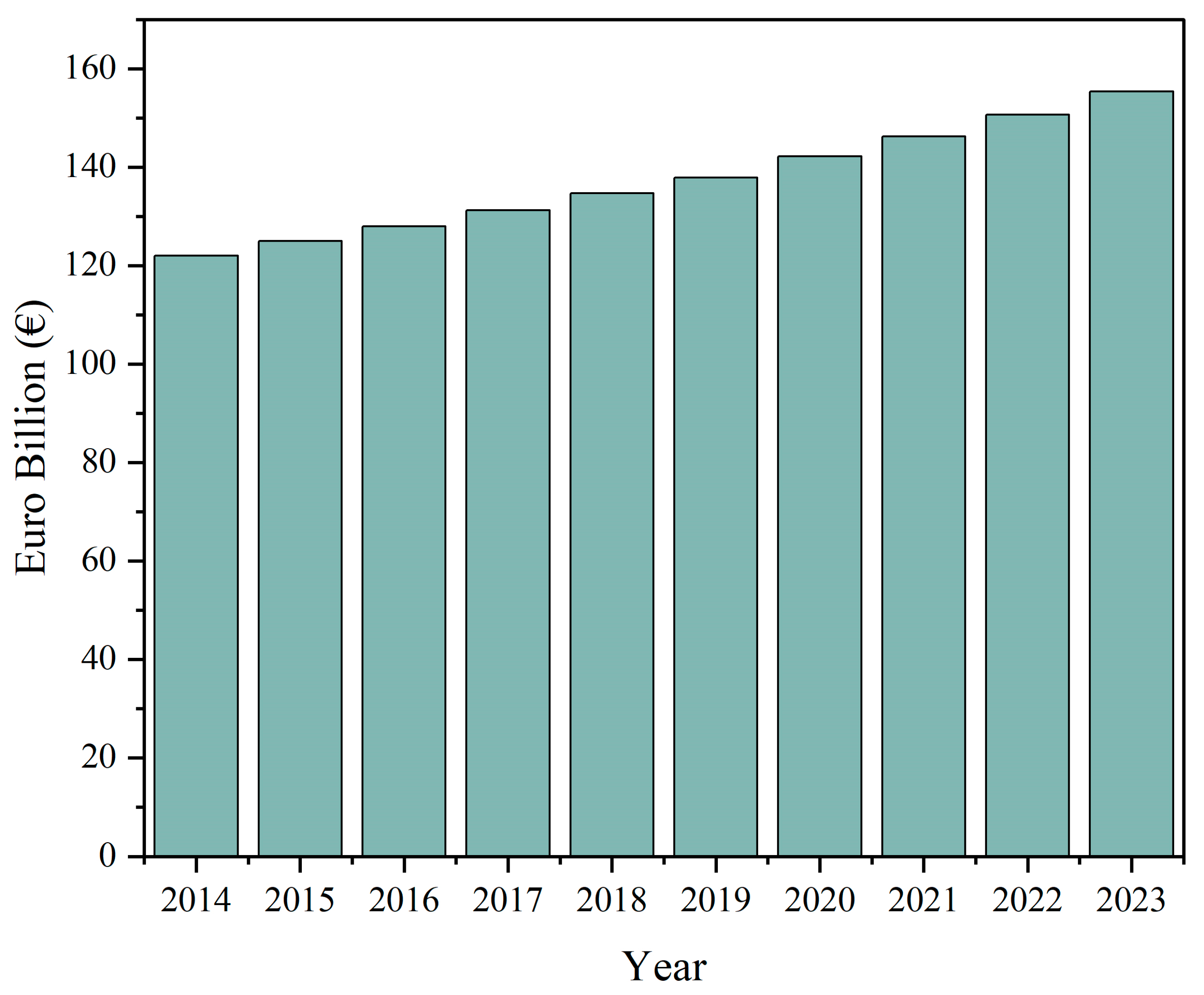
Figure 3.
Overview of health benefits related to consumption or usage of vitamins, bacteriocins, bioactive peptides and bioactive compounds.
Figure 3.
Overview of health benefits related to consumption or usage of vitamins, bacteriocins, bioactive peptides and bioactive compounds.
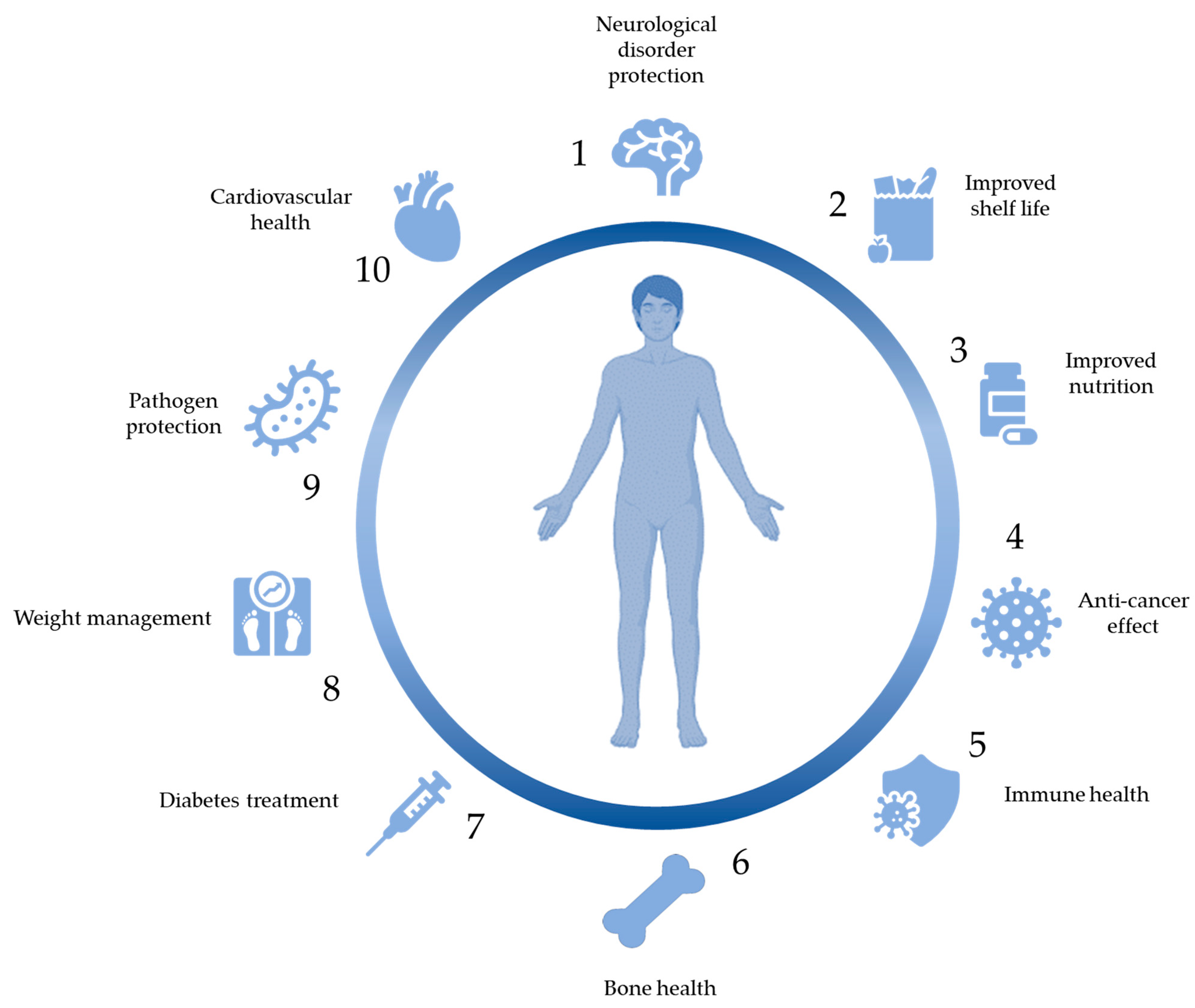
Table 3.
Overview of cobalamin producing Propionibacteria.
| Microorganism(s) | Fermentation | Medium | Yield | Ref |
|---|---|---|---|---|
| P. acidipropionici | 1 L fermentor | Complex media | 3.3 mg/L | [72] |
| P. denitrificans | 120 L Batch | Complex media | 177.49 µg/l | [73] |
| P. denitrificans | 120 L Batch | Complex media | 214.3 µg/mL | [74] |
| P. freudenreichii | 5 L fermentor | Defined media | 9.45 µg/mL | [75] |
| P. freudenreichii | Cell recycle system | Complex media | 24.93 µg/mL | [76] |
| P. shermanii | Flask | Whey based medium | 8.43 µg/l | [77] |
| P. shermanii | Flask | Whey based medium | 2.97 µg/l | [78] |
| P. shermanii | Flask | Whey based medium | 4.51 µg/l | [79] |
Table 4.
Overview of riboflavin-producing microorganisms.
| Microorganism(s) | Fermentation | Medium | Yield | Ref |
|---|---|---|---|---|
| L. acidophilus | Flask | Whey based medium | 2.93 mg/L | [97] |
| L. fermentum | Flask | CDM | 1.2 mg/L | [98] |
| L. fermentum | Flask | CDM | 3.49 mg/L | [99] |
| L. lactis | Flask | Whey based medium | 2.61 mg/L | [97] |
| L. plantarum | Flask | MRS | 3.33 mg/L | [100] |
| L. plantarum | Flask | Supplemented MRS | 3.33 mg/L | [101] |
| L. plantarum | Flask | CDM | 5.72 mg/L | [102] |
Table 5.
Overview of menaquinone-producing LAB.
| Microorganism(s) | Fermentation | Medium | Yield | Ref |
|---|---|---|---|---|
| L. fermentum | Flask | Rogosa medium Skim milk |
184 µg/L 63.93 µg/L |
[122] |
| L. lactis subsp. cremoris | Flask | Milk-based media | 534 nmol/g of cells | [123] |
| L. lactis subsp. lactis | Flask | Milk-based media | 717 nmol/g of cells | [123] |
| Leu. lactis | Flask | Milk-based media | 173 nmol/g of cells | [123] |
| P. freudenreichii | 3 L fermentor | Milk-based media | 0.3 mM | [124] |
Table 7.
Summary of ACEI peptide-producing LAB and their yields.
| Microorganism(s) | Fermentation | Medium | ACEI ability | IC50 | Ref |
|---|---|---|---|---|---|
| L. acidophilus | Flask | Skim milk-based media | - | 730 µg/mL | [202] |
| L. brevis | Flask | MRS | 79.03 | 1280 µg/mL | [196] |
| L. brevis | Flask | Whey based media | 64.7 | 1130 µg/mL | [206] |
| L. casei | Flask | Skim milk-based media | - | 250 µg/mL | [202] |
| L. casei | Flask | Skim milk-based media | 100 | [207] | |
| L. casei | Flask | Goats milk-based media | 34.3 | - | [208] |
| L. delbruckii subsp. bulgaricus | Flask | Skim milk-based media | - | 780 µg/mL | [202] |
| L. helveticus | Flask | Whey based media | 84.2 | 860 µg/mL | |
| L. helveticus | Flask | Skim milk-based media | - | 1460 µg/mL | [202] |
| L. helveticus | Flask | Skim milk-based media | 67.18 | - | [209] |
| L. helveticus | Flask | Goats milk-based media | 51.3 | - | [208] |
| L. paracasei | Flask | Whey based media | 63.9 | 1130 µg/mL | [206] |
| L. plantarum | Flask | MRS | 84.0 | 65.53 µg/mL | [196] |
| L. plantarum | Flask | Skim milk-based media | - | 910 µg/mL | [202] |
| L. plantarum | Flask | Goats milk-based media | 37.7 | - | [208] |
| L. rhamnosus | Flask | Skim milk-based media | - | 700 µg/mL | [202] |
| L. rhamnosus | Flask | MRS | 52.4 | 2130 µg/mL | [196] |
| L. sakei | Flask | Skim milk-based media | - | 1220 µg/mL | [202] |
| Lc. lactis | Flask | Skim milk-based media | - | 220 µg/mL | [202] |
| E. durans | Flask | Skim milk-based media | - | 450 µg/mL | [202] |
| E. faecium | Flask | MRS | 55.4 | 70.5 µg/mL | [196] |
| P. acidilactici | Flask | MRS | 84.7 | 19.78 µg/mL | [196] |
| P. pentosaceus | Flask | MRS | 72.9 | 2070 µg/mL | [196] |
| P. pentosaceus | Flask | Skim milk-based media | - | 780 µg/mL | [202] |
| S. thermophilus | Flask | Skim milk-based media | - | 820 µg/mL | [202] |
| Streptococcus thermophilus | Flask | Goats milk-based media | 40 | - | [208] |
|
Streptococcus thermophilus, Lactobacillus casei, Lactobacillus helveticus |
Flask | Goats milk-based media | 82 | - | [208] |
|
S. thermophilus, L. casei, Lactobacillus plantarum |
Flask | Goats milk-based media | 43.3 | - | [208] |
Table 8.
Summary of antioxidant peptide-producing LAB and their reported activity.
| Microorganism(s) | Fermentation | Medium | DPPH (mg/ml) or % |
ABTS (mg/ml) |
HFRSR (%) |
Antimutagenic activity (% inhibition) | Ref |
|---|---|---|---|---|---|---|---|
|
S. thermophilus L. bulgaricus |
Cups | Skim milk-based media | 2.23 mg/ml | 2.43 mg/ml | - | 15.87 | [221] |
|
S. thermophilus L. bulgaricus L. acidophilus |
Cups | Skim milk-based media | 2.05 mg/ml | 2.28 mg/ml |
- | 18.35 | [221] |
|
S. thermophilus L. bulgaricus L. casei |
Cups | Skim milk-based media | 1.83 mg/ml |
1.91 mg/ml | - | 18.83 | [221] |
|
S. thermophilus L. bulgaricus L. paracasei |
Cups | Skim milk-based media | 1.82 mg/ml |
1.98 mg/ml | - | 18.48 | [221] |
|
S. thermophilus L. bulgaricus L. acidophilus L. casei |
Cups | Skim milk-based media | 1.8 mg/ml |
1.73 mg/ml | - | 20.25 | [221] |
| Cups | Skim milk-based media | 1.77 mg/ml |
1.8 mg/ml | - | 23.06 | [221] |
|
| L. casei | Flask | Goat milk-based media | 63.48 % | - | 88.01 | - | [222] |
| L. mesenteroides subsp. cremoris | Flask | Skim milk-based media | - | 0.7 nmol-1/mmol-1 | - | - | [223] |
| L. lactis subsp. lactis | Flask | Skim milk-based media | 0.15 nmol-1/mmol-1 | - | - | [223] | |
| L. acidophilus | Flask | Skim milk-based media | - | 0.6nmol-1/mmol-1 | [223] | ||
| L. jensenii | Flask | Skim milk-based media | - | 0.6 nmol-1/mmol-1 | - | - | [223] |
| L. helveticus | Flask | Skim milk-based media | - | 0.4 nmol-1/mmol-1 | - | - | [223] |
Table 9.
Summary of GABA-producing strains and yields.
| Microorganism(s) | Fermentation | Medium | Yield | Ref |
|---|---|---|---|---|
| L. acidophilus | Flask | Goats milk-based media | 1.92 mg/kg | [208] |
| L. brevis | Packed bed reactor | Defined media | 55 mM | [236] |
| L. brevis | Batch | Modified MRS | 44.4 mg/ml | [237] |
| L. brevis | Fed-batch | Defined media | 1005.81 mM | [238] |
| L. brevis | Fed-batch | Complex | 526.33 mmol/L | [239] |
| L. brevis | Flask | MRS + MSG | 255 mM | [240] |
| L. brevis | Batch | Defined media | 205.8 g/L | [241] |
| L. brevis | Batch | Defined media | 62.5 g/L | [242] |
| L. brevis | Flask | Supplemented MRS | 265 mM | [243] |
| L. brevis | Entrapped cell | Defined media + MSG, | 223 mM | [244] |
| L. brevis | Flask | Whey permeate-based media + MSG | 553.5 mg/L | [245] |
| L. brevis | Flask | MRS + MSG | 2.5 g/L | [246] |
|
L. brevis L. sakei |
Flask | Skim milk-based media + MSG | 22.41 mM | [247] |
| L. buchneri | Flask | Modified MRS | 251mM | [248] |
| L. buchneri | Flask | MRS + MSG | 4.4 g/L | [249] |
| L. casei | Flask | Skim milk-based media | 677.35 mg/kg | [207] |
| L. casei | Flask | Goats milk-based media | 1.65 mg/kg | [208] |
| L. delbrueckii subsp. bulgaricus | Flask | Skim milk-based media | 9 mg/kg | [250] |
| L. helveticus | Flask | Skim milk-based media | 165.11 mg/L | [209] |
| L. helveticus | Flask | Goats milk-based media | 0.06 mg/kg | [208] |
| L. paracasei | Flask | Modified MRS | 302 mM | [251] |
| L. paracasei | Flask | Skim milk-based media | 20 mg/kg | [250] |
| L. paracasei and L. plantarum | Flask | Skim milk base dmedia | 6.7 mg/100ml | [252] |
| L. plantarum | Flask | Skim milk + MSG | 629 mg/ml | [253] |
| L. plantarum | Flask | Defined media + MSG | 19.8 g/L | [254] |
| L. plantarum | Flask | MRS | 4.156 mg/L | [255] |
| L. plantarum | Flask | MRS + MSG | 821.24 mg/L | [256] |
| L. plantarum | Flask | Skim milk basd media + MSG | 314 mg/100g | [257] |
| L. plantarum | Flask | MRS + MSG | 7.15 mM | [258] |
| L. plantarum | Flask | Grape must | 4.85 mM | [259] |
| L. plantarum | Flask | MRS + MSG | 201.78 mg/L | [260] |
| L. plantarum | Flask | Skim milk-based media | 6 mg/kg | [250] |
| L. plantarum | Flask | Skim milk + yeast extract | 77.4 mg/kg | [202] |
| L. plantarum | Flask | Goats milk | 12.84 mg/kg | [208] |
| L. plantarum and L. sakei | Flask | Whey based media | 365.6 mg/100 ml | [261] |
| L. rhamnosus | Flask | Complex media + MSG + PLP | 187 mM | [262] |
| L. sakei | Batch | MRS + MSG | 265.3 mM | [263] |
| L. sakei | Batch | MRS + MSG | 217 mM | [263] |
| S. salivarius subsp. thermophilus | Flask | Complex media | 7984.75 mg/L | [264] |
| S. thermophilus | Flask | Skim milk-based media + MSG | 2.2 mg/ml | [265] |
| S. thermophilus | Flask | Goats milk-based media | 1.59 mg/kg | [208] |
|
S. thermophilus L. brevis |
Flask | Skim milk + MSG | 314.97 mg/kg | [266] |
|
S. thermophilus, L. casei, L. helveticus |
Flask | Goats milk-based media | 5.79 mg/kg | [208] |
|
S. thermophilus, L. casei, L. plantarum |
Flask | Goats milk-based media | 30.86 mg/kg | [208] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



