
Submitted:
06 October 2023
Posted:
10 October 2023
You are already at the latest version
Abstract
It has been more than three decades since the discovery of multifunctional factors, Non-POU Domain-Containing Octamer-Binding Protein, NonO and Splicing Factor, Proline-and Glutamine-Rich, SFPQ. Some of their functions, including their participations in transcriptional and posttranscriptional regulation as well as their contribution to paraspeckle subnuclear body organization have been well-documented. In this review, we focus on the roles of NonO and SFPQ in cell cycle with particular attention to their participation in nonhomologous end joining (NHEJ) and homologous recombination (HR) of DNA damage responses induced by irradiation. In these contexts, absence or malfunction of either or both lead to genome instability and ultimately to apoptosis, cellular senescence, and tumor development. This review also will shed light on NonO and SFPQ function in DNA/RNA/protein binding and their association with Ku heterodimer of the NHEJ pathway required for repair of DNA double-strand breaks (DSBs).
Keywords:
NonO/p54nrb
; SFPQ/PSF
; DNA damage response
; ATR/ATM
; cell cycle
; checkpoint control
Introduction
NonO was first reported more than three decades ago [1,2,3] a single-strand octamer DNA binding protein and subsequently, as a DNA and RNA binding protein [2]. SFPQ was first purified with another RNA-binding protein, Polypyrimidine Tract Binding (PTB) [4]and defined as a splicing factor [5]. SFPQ participates in numerous mechanisms, including transcription (reviewed in [6,7]), post-transcription ([8,9]; reviewed in [10]), and RNA processing [11,12]. Early studies of NonO and SFPQ also focused on their direct regulation of transcription [13], post-transcription [14], and 3’ end processing [15]. Subsequently, NonO and SFPQ were found to be key components of nuclear paraspeckles, which control gene expression by the nuclear retention of RNA ([16,17]; and reviewed recently in [18]).
NonO and SFPQ (Figure 1) are members of the Drosophila Behavior/Human Splicing (DBHS) family of RNA/DNA binding cofactors liable for a range of cellular processes (reviewed in [19,20,21]). DBHS family also includes PSPC1, Hrp65 [22], NonO-1 [23], and NonA [24] (Figure 2A). All DBHS family members bind DNA and RNA via highly conserved RNA-binding domains (RRMs), a NOPS (NonA/ParaSpeckle) domain and coiled coil Regions (reviewed in [21]). These data suggested that the structures of NonO, SFPQ, and PSPC1 are similar, particularly in the regions required for the homo-/hetero- dimer interactions for their functions [25,26,27]. Accordingly, these observations were validated at the atomic level via Xray structure analyses [28,29,30].
Multifunctional roles of NonO and SFPQ
The studies above as well as others described below established NonO and SFPQ as “multifunctional” proteins (reviewed in [6]). For example, NonO is involved in cell proliferation ([26]; reviewed in [31]), whereas, SFPQ binds U5 small nuclear RNA and participate in splicing [32]. NonO, SFPQ and PSPC1 are key components of paraspeckles (reviewed in [21]). To mediate this function, NonO interact with CARM1 (the histone coactivator associated arginine methyltransferase 1) [33] to localize CARM1 within paraspeckles for its regulation in pre-implantation mouse embryo development [34]. NonO and SFPQ can act as RNA splicing regulators, and each has been implicated in neurogenesis (reviewed in [35]) in differentiation and development [36,37]. Loss of NonO or SFPQ results in apoptosis, and senescence ([38]; reviewed in [7]).
NonO and SFPQ can function individually, but often as heterodimers or as larger complexes [26,27]. For example, NonO, but not SFPQ, plays a significant role in circadian rhythm [39,40]. Disruption of NonO and SFPQ heterotetramers induces premature senescence [27]. Dysregulation of either NonO, or SFPQ, or both can lead to tumorigenesis [41,42]. For example, NonO upregulation can lead malignant breast cell proliferation [43], whereas NonO and SFPQ together promote castration-resistant prostate cancer progression [44,45]. Fusion of either NonO or SFPQ with the TFE3 transcriptional factor, has been observed in papillary renal cell carcinoma [46,47]. This same SFPQ-TFE3 oncoprotein inactivates TFE3 and p53 in papillary renal cell carcinoma [48] (Figure 2B).
Focus of this review
In addition to the multifunctional roles described above, NonO and SFPQ participate in double strand break repair in response to DNA damage [49,50]. Observations made several years ago showed that both NonO and SFPQ function in DNA unwinding and pairing [50]. In this review, we focus on these roles as well as how the interactions among NonO and/or SFPQ with other DNA damage repair factors illuminate their dual roles in double strand break (DSB) repair. We close with a more detailed consideration of NonO and SFPQ in cancer.
NonO and SFPQ in cell cycle and cell cycle arrest
Long ago, a dimeric complex of NonO:SFPQ complex was shown to be more highly proliferative in cancer cell lines than in normal cells [26]. NonO silencing induced reduction of the G1/S phase. Also, NonO was shown to be required for arrest of DNA replication (S phase) with ultraviolet-C (UVC) [51]. After UVC treatment, NonO localized with Rad9, an ortholog of 53BP1, foci [51]. These and other data indicated that NonO acts upstream of the ATR-mediated DNA-damage response cascade [51]. NonO is required for responding to UV irradiation-mediated checkpoint arrest. Tyrosine phosphorylation of SFPQ, on the other hand, has been reported to promote S-phase cell cycle arrest and its cytoplasmic localization [52].
NonO and SFPQ also are required for G2/M arrest [53,54]. In search of G2/M arrest-related proteins, Roberts et.al, [55] identified NonO as one of two protein targets phosphorylated by MKK 1/2 inhibitors [27,55]. Microtubule interfering agents and other agents, such as kinesin spindle protein (KSP) inhibitors, were required to induce NonO phosphorylation [56]. Similar approaches revealed that SFPQ is hyperphosphorylated in G2/M arrest [57]. Phosphorylation of NonO or SFPQ occurs when cell cycle arrests.
In summary, both NonO and SFPQ are required to arrest cell cycle and the phosphorylation of NonO and SFPQ might be involved for the triggering cell cycle or downstream regulation.
NonO and SFPQ participate in DNA damage repair
The initial indication that NonO was required for DNA damage repair came from observations by Roberts et al, [55]. Following purification during G2/M arrest, NonO was observed to be phosphorylated. Also SFPQ previously had been shown to be hyperphosphorylated during G2/M arrest [50]. These studies indicated that NonO and SFPQ phosphorylation are associated with cell cycle arrest. Further studies indicated that following cellular stress or UV-induced damage, NonO and SFPQ participate in double strand break (DSB) repair [50].
The two major pathways of DNA double-strand break (DSB) repair are Non-Homologous End Joining (NHEJ) and Homologous Recombination (HR) (Reviewed in [58]). NHEJ is the primary pathway for repairing DNA double-strand breaks (DSBs) (Reviewed in [59]) and one of two major pathways of double-strand break repair in mammalian cells. The Mre11-Rad50- NBS1 (MRN) complex plays a key role in NHEJ. NonO and SFPQ were isolated as a new ~200KDa fraction of MRN [49]. Subsequently, the complex was found to be devoid of a crucial ATP-dependent DNA ligase activity required to covalently join adjacent 5’ phosphate (5’P) and 3’ hydroxy (3’OH) termini of double-stranded [50]. Attenuation of NonO led to cell cycle arrest [60] and delayed DNA damage repair, whereas radiation-induced chromosomal aberrations were increased [60].
The mechanism employed by the NonO:SFPQ heterodimer is to promote binding of DNA substrates and stimulation of DNA end joining. This activity, in cooperation with Ku, leads to formation of the preligation complex [50].
NonO and SFPQ participate in Nonhomologous End Joining (NHEJ)
There are several key polypeptides required for NHEJ: Two subunits of Ku, which bind to DNA ends; DNA ligase IV (L4) and XRCC4 (X4), which form a complex that catalyzes strand ligation; and DNA-PKcs, the only active protein kinase described in the NHEJ pathway. Studies by Dynan et al. (2005) indicated that NonO and SFPQ also are two essential factors of NHEJ. The NonO:SFPQ heterodimer along with the Ku complex bind linear DNA fragments independently and to form a functional preligation complex [50].
Matrin 3, a nuclear matrix protein initially proposed to stabilize selective mRNA species, also is involved in this early stage of the DSB response [61]. Matrin 3:NonO:SFPQ:Ku can also bind substrate DNA in vitro [50]. However, NonO:SFPQ only cooperates with Ku in cis (i.e. on the same DNA strand). Generally, NonO:SFPQ:Ku bind cooperatively to DNA substrates in vitro to form functional preligation complexes [50].
Li et al. [60] demonstrated that attenuation of NonO expression increased the frequency of radio-induced chromosomal aberrations. Subsequently, the same group observed NonO knockdown produced a DSB repair-deficient, radiosensitive phenotype. These data suggest that loss of NonO may deter the cellular response to irradiation and DSB repair [60].
NonO:SFPQ and Ku also bind DNA substrates independently, but in different manners. Udayakumar et al, [62] observed that Ku promoted capture of radiolabeled DNA in a concentration-dependent manner. While NonO:SFPQ also promoted capture of radiolabeled DNA, but an ~4-fold higher concentrations was required for equivalent retention [62]. Furthermore, this group showed that, unlike Ku proteins, NonO:SFPQ can bind DNA substrates without free ends. This suggested that the NonO:SFPQ complex can capture DNA sequences and, in cooperation with Ku, stabilize a synaptic preligation complex. Also, NonO:SFPQ stimulates kinase activity and increases DNA-PKcs autophosphorylation [62].
PSPC1, which is highly similar to NonO and a DBHS member, has yet to be reported as a key component in DSB repair. However, knockdown of NonO can be rescued by overexpressing PSPC1 [63]. This suggested that PSPC1 can replace NonO in teaming up with SFPQ to participate in NHEJ.
NonO and SFPQ in Homologous Recombination (HR)
Even though NonO and SFPQ have been purified together and participate in DSB repair, NonO alone has also been found to stimulate NHEJ and suppress HR [64]. SFPQ, on the other hand, interacts directly with RAD51 to participate in HR [53]. RAD51 family members are key proteins involved in HR responses as well as in repair and genome stability [65,66]. RAD51 is an essential recombinase in both meiotic and mitotic homologous recombination [67]. NonO and SFPQ were co-purified, in a large-scale screen as RAD51 binding partners [68]. Subsequently, SFPQ was shown to directly interact with the RAD51D isoform and to mediate epistatic effects on cell viability. SFPQ also promotes sister chromatid cohesion and maintains chromosome integrity [53].
SFPQ also was shown to interact with RAD51 in a 1:1 ratio to modulate its HR activity [67]. Furthermore, SFPQ promotes strand exchange between ssDNA and DNA/RNA hybrids in a transcription-associated manner [67]. Finally, SFPQ stimulates RAD51-mediated homologous pairing and strand exchange under low RAD51 conditions, while it inhibits RAD51-mediated recombination when RAD51 concentrations are optimal [67].
The evidence is clear that SFPQ is necessary for HR repair during DSB damage as well as homology-directed repair and sister chromatid cohesion [53].
NonO and/or SFPQ are associated with DSB repair factors
Numerous experiments indicate that NonO and SFPQ associate with DSB repair factors. For example, NonO and SFPQ directly associate with topoisomerase I (TOPI), whose major function is to relax supercoiled DNA and to alleviate DNA helical constraints [69]. Cleavage and preligation half-reactions of TOPI are unaffected by NonO:SFPQ, whereas the propensity of the enzyme to “jumping” between separate DNA helices is stimulated [69]. The interaction was buttressed by the observation that a NonO:SFPQ:TOPI complex is pulled down by an antibody recognizing TOPII beta [70].
TOPI is bound with numerous cofactors, and one of them, TopBP1 has been reported to interact with NonO and SFPQ to repair laser-induced DNA damage sites [71]. NonO, possibly via its RRM domains, interacts with BRCT domains, 6-8, of TopBP1 as demonstrated in a yeast two-hybrid system [71]. Considering that TopBP1 expression comes earlier (5s, following DNA damage induction) than NonO and SFPQ (20s), TopBP1 might recruit NonO and SFPQ to DSB sites. The 3 proteins reach their maximal concentrations at around 60s. At a point, NonO and SFPQ have disengaged while TopBP1 remains bound [71].
The Ku DNA end-binding complex, which plays various roles with NonO:SFPQ as discussed previously, is involved with DSB at the earliest stage [61]. Ku proteins, associated with XRCC-DNA ligase IV and other NonO:SFPQ interacting proteins to form “scaffolds” that stabilize DNA pairing (reviewed in [72]). NonO:SFPQ stimulates autophosphorylation of DNA-PKs, but the mechanism of action remains to be investigated [62].
As with their roles in HR, RAD51 proteins perform key functions of homologous recombination in DNA repair and chromosomal integrity (reviewed in [72]). SFPQ and NonO were among four candidates pulled down in an attempt to identify interaction profiles of RAD51D, whereas SFPQ was pulled down by RAD51C [68]. SFPQ subsequently was shown to interact with RAD51D and participate in homology-directed repair of DSBs [53]. Depletion of both SFPQ and RAD51 lead to a lethal phenotype, whereas reduced expression of SFPQ and RAD51D interrupted the cell cycle progression leading to G2/M arrest and/or chromosomal aneuploidy. Thus, SFPQ directly participates in homologous repair in DSB DNA damage [53].
A series of parallel studies determined that SFPQ regulates RAD51-mediated homologous pairing between single-strand (ss) DNA and supercoil double-stranded (ds) DNA [67]. At low RAD51 concentration, SFPQ promotes homologous pairing, but inhibits such pairing at concentrations in which RAD51 can function alone [67]. In the same paper, it was observed that SFPQ modulated RAD51-mediated strand exchange in a concentration-dependent manner; i.e. promoting strand exchange at low RAD51 concentrations while inhibiting the reaction when RAD51 concentrations were optimal. SFPQ binding can compete with ssDNA binding of RAD51 when its concentration is high, culminating with disassembly of the RAD51-ssDNA interaction [67].
RAD9, the yeast ortholog of 53BP1, encodes an adaptor protein required for S. cerevisiae cell cycle checkpoint arrest in G1/S, intra-S, and G2/M [73]. RAD9 also plays a role in [73] post-replication repair (PRR) pathway via transmission of a checkpoint signal via phosphorylation of the RAD9-HUS1-RAD1 (9-1-1) clamp complex [74]. While there is no evidence indicating that NonO or SFPQ interacts with 53BP1, depletion of SFPQ delays DSB repair [61]. However, 53BP1 foci disappears when the expression levels of SFPQ are reduced [61].
Poly(ADP-ribose) polymerase-1 (PARP-1), one of several members of the PARP family, is a strong sensor of DNA damage (reviewed in [75]). PARP-1 rapidly produces Poly(ADP-Ribose) (PAR) [64], which appears to be involved in DNA damage repair (reviewed in [76]). NonO has been reported to be a PAR-binding protein [64], which colocalizes with PARP-1 and PAR at laser-IR-induced DNA damage sites immediately after the introduction of DNA lesions [64]. The recruitment of NonO to the DNA damage site is PARP-1 and PAR-dependent and it is mediated via RRM1 of NonO [64]. Moreover, either knockdown of NonO expression or PARP inhibition decreases NHEJ, while attenuation of NonO not only decreases NHEJ, but also facilitates repair by homologous recombination (HR) [64].
It has been speculated that NonO:SFPQ might substitute for XRCC-Like Factor (XLF) or for Non-Homologous End Joining Factor 1 (NHEJ1) to promote sequence-independent pairing of DNA substrates in vitro. But it is clear that the ability of NonO to bind RNA contributes to DSB repair [77].
Posttranscriptional modification of NonO facilitates DNA damage repair
Protein O-GlcNAcylation, catalyzed by O-GlcNAc transferase (OGT) has been linked with DNA damage (reviewed in [78]). The removal of OGlcNAcylation is catalyzed by O-GlcNAcase (OGA). OGA relocates to sites of DNA damage, where its C-terminal pseudo-Histone Acetyltransferase (HAT) domain plays a key role for its recruitment as well as its substrate recognition [79]. NonO and the Ku70/80 complex are O-GlcNAcylated by OGT [79]. Delay of OGlcNAcylation at DNA lesions delays NonO degradation and impairs NHEJ [79].
RING finger protein 8 (RNF8) is a major E3 ubiquitin ligase that, via its FHA domain, rapidly accumulates at sites of DNA damage via its FHA domain to facilitate phosphorylation of MDC1. Phosphorylation of MDC1 occurs in response to DNA damage and is mediated by phosphoinositol-3-kinase-related kinases, mainly by Ataxia Telangiectasia Mutated (ATM) or ATM and Rad3-related (ATR) [80,81]. NonO, but not SFPQ and PSPC1, is a substrate of RNF8 [81]. NonO is necessary for the loading of TopBP1 and ATR Interacting Proteins (ATRIP) to chromatin following UV irradiation [81]. RNF8 also mediates the ubiquitination and degradation of NonO − an event necessary to terminate ATR-CHK1 checkpoint signaling induced by UV-induced DNA Damage Repair (UV-DDR) and required for S phase progression [81]. Interfering with RNF-mediated degradation of NonO, via mutation of three key lysine residues prolongs S phase after UV exposure [81].
These data suggest that OGlcNAcylation and ubiquitination of NonO acts in the absence of SFPQ to stabilize and to retain UV-induced chromatin protein complexes. Conversely, the degradation of NonO is necessary for cell cycle progression.
NonO and SFPQ in telomere stability
Neither NonO nor SFPQ are directly involved in the activity of telomerase [27]. However, both are pulled down in a complex with TElomeric Repeat-containing RNA (TERRA) [82], a class of long noncoding RNAs transcribed at telomeres that actively participate in regulating telomere maintenance and chromosome end protection. While depletion of NonO or SFPQ in malignant cells does not impact total TERRA levels, it does increase TERRA foci per nucleus [82]. Further, depletion/loss of NonO or SFPQ increases the RNA:DNA-hybrid formation between TERRA and the C-rich telomeric strand [82].
Phosphorylation of ATR and serine 33 of the 32 kDa subunit of the Replication Protein A (RPA32pSer33) are both markers of replication stress [83]. Depletion of NonO and SFPQ results in the recruitment of RPA and this effect can be abolished by expression of RNaseH1 [82]. These observations indicate that NonO and SFPQ interact with TERRA to prevent the formation of RNA:DNA hybrid and R-loop replication defects at telomeric repeats [82].
TERRA are composed of DNA, RNA, and protein [82,84]. Perhaps, NonO, and/or SFPQ is involved in either DNA/RNA or protein interactions that recruit other members to stabilize the telomere complex. This hypothesis is buttressed by the observation that NonO and SFPQ have been recently discovered as regulators of telomere length homeostasis by suppressing telomere fragility and HR triggered by the TERRA-induced RNA:DNA hybrid. Loss of both NonO and SFPQ results in an increase in homologous recombination and altered telomere length homeostasis [82].
Summary and Cancer Implications
This review has addressed select roles of two multifunctional proteins—the Non-POU Domain-Containing Octamer-Binding Protein (NonO) and Splicing Factor, Proline-and Glutamine-Rich (SFPQ). We elected to focus on topics that have been less thoroughly reviewed, including NonO/SFPQ function in nonhomologous end joining (NHEJ), homologous recombination (HR) and repair of DNA double-strand breaks (DSBs). We attempted to summarize in these contexts how the absence or malfunction of NonO, SFPQ or both result in genome instability, apoptosis and cellular senescence. Finally, we assessed the roles of several NonO and SFPQ interacting proteins in these contexts, including Ku, PARP, TopBP1 and TERRA.
We find it interesting that NonO was initially isolated from a B cell leukemia [1]. Although not implicated clinically in B cell neoplasias, NonO is a key component in NHEJ and DSB repair—mechanisms critical for B- and T-cell receptor generation. During V(D)J recombination, DSBs induced by Recombination Activating Gene proteins (RAG1 and RAG2) are virtually all repaired by the NHEJ pathway for the benefit of antigen receptor gene diversity (reviewed in [85,86]).
While neither NonO nor SFPQ has been characterized further in lymphoma/leukemia, aberrant function of both has been observed in the etiology of colorectal, hepatocellular, renal, myeloid and prostate cancers ([87]; reviewed in [88]). Particularly penetrant is the connection of these two factors in neuroblastoma. Suitable treatments for low-risk patients exist, but high-risk neuroblastoma patients have exceedingly poor survival rates and lack therapeutic options. In particular, high overexpression of NonO is associated with poor survival [89]. This suggests that NonO might represent a potential therapeutic target. Unfortunately, mechanistic data implicating NonO in neuroblastoma, beyond its binding to a tumor-specific long non-coding RNA (LncMycnUS) remains unknown.
Recently Zhang and colleagues [90] proposed a model for a regulatory role for NonO in neuroblastoma. They contend that NonO binding to pre-mRNA of enhancer-regulated genes promotes the formation of RNA-processing “condensates” to allow efficient splicing. Such a model supports the growing body of evidence of NonO, and possibly SFPQ upregulation in different cancer cell types and clinical samples [31,45]. Although NonO is not a crucial component in spliceosome assembly, it interacts with critical spliceosomal proteins [44,91]. Such features suggest that NonO may be part of a “master” transcriptional regulator capable of assembling other transcription factors and regulators to drive significant overexpression in neuroblastoma.
As a clinical manifestation, NonO inhibitors might be good prospects for inhibiting growth via induction of apoptosis in neuroblastoma. A potential avenue of exploration should include small molecule inhibitors of NonO function. These observations may provide a new approach in the development of pharmaceutical drugs to manipulate aberrant RNA-binding capacity underlying cancer and other diseases.
Acknowledgments
We thank Tyson Phonesavanh for preparing the figures, Qinghua Xu for compiling References, and Lu Zhang for discussions of the manuscript. We thank members of the Tucker laboratory for discussions and reading of the manuscript. The paper is supported by NYU Shanghai Faculty Discretionary Funds. H.O.T. received support from NIH Grant R01CA31534, Cancer Prevention Research Institute of Texas (CPRIT) Grants RP100612, RP120348; and the Marie Betzner Morrow Centennial Endowment.
Conflicts of interest
These authors declare no conflict of interest.
References
- Hanke, J.H.; Landolfi, N.F.; Tucker, P.W.; Capra, J.D. Identification of Murine Nuclear Proteins That Bind to the Conserved Octamer Sequence of the Immunoglobulin Promoter Region. Proc Natl Acad Sci U S A 1988, 85, 3560–3564. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Hanke, J.H.; Carayannopoulos, L.; Craft, C.M.; Capra, J.D.; Tucker, P.W. NonO, a Non-POU-Domain-Containing, Octamer-Binding Protein, Is the Mammalian Homolog of Drosophila nonAdiss. Mol Cell Biol 1993, 13, 5593–5603. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Horowitz, D.S.; Kobayashi, R.; Krainer, A.R. Purification and cDNA Cloning of HeLa Cell P54nrb, a Nuclear Protein with Two RNA Recognition Motifs and Extensive Homology to Human Splicing Factor PSF and Drosophila NONA/BJ6. Nucleic Acids Res 1993, 21, 4085–4092. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.G.; Mayer, S.A.; Tempst, P.; Nadal-Ginard, B. Characterization and Molecular Cloning of Polypyrimidine Tract-Binding Protein: A Component of a Complex Necessary for Pre-mRNA Splicing. Genes Dev 1991, 5, 1237–1251. [Google Scholar] [CrossRef]
- Patton, J.G.; Porro, E.B.; Galceran, J.; Tempst, P.; Nadal-Ginard, B. Cloning and Characterization of PSF, a Novel Pre-mRNA Splicing Factor. Genes Dev 1993, 7, 393–406. [Google Scholar] [CrossRef]
- Shav-Tal, Y.; Zipori, D. PSF and P54(Nrb)/NonO--Multi-Functional Nuclear Proteins. FEBS Lett 2002, 531, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Yarosh, C.A.; Iacona, J.R.; Lutz, C.S.; Lynch, K.W. PSF: Nuclear Busy-Body or Nuclear Facilitator? Wiley Interdiscip Rev RNA 2015, 6, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Mitobe, Y.; Iino, K.; Takayama, K.-I.; Ikeda, K.; Suzuki, T.; Aogi, K.; Kawabata, H.; Suzuki, Y.; Horie-Inoue, K.; Inoue, S. PSF Promotes ER-Positive Breast Cancer Progression via Posttranscriptional Regulation of ESR1 and SCFD2. Cancer Res 2020, 80, 2230–2242. [Google Scholar] [CrossRef]
- Takayama, K.-I.; Honma, T.; Suzuki, T.; Kondoh, Y.; Osada, H.; Suzuki, Y.; Yoshida, M.; Inoue, S. Targeting Epigenetic and Posttranscriptional Gene Regulation by PSF Impairs Hormone Therapy-Refractory Cancer Growth. Cancer Res 2021, 81, 3495–3508. [Google Scholar] [CrossRef]
- Figueroa, A.; Fujita, Y.; Gorospe, M. Hacking RNA: Hakai Promotes Tumorigenesis by Enhancing the RNA-Binding Function of PSF. Cell Cycle 2009, 8, 3648–3651. [Google Scholar] [CrossRef]
- Stagsted, L.V.W.; O’Leary, E.T.; Ebbesen, K.K.; Hansen, T.B. The RNA-Binding Protein SFPQ Preserves Long-Intron Splicing and Regulates circRNA Biogenesis in Mammals. Elife 2021, 10, e63088. [Google Scholar] [CrossRef] [PubMed]
- Cosker, K.E.; Fenstermacher, S.J.; Pazyra-Murphy, M.F.; Elliott, H.L.; Segal, R.A. The RNA-Binding Protein SFPQ Orchestrates an RNA Regulon to Promote Axon Viability. Nat Neurosci 2016, 19, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, S.; Ikei, A.; Taguchi, Y.; Tabuchi, Y.; Fujimoto, N.; Obinata, M.; Uesugi, S.; Kurihara, Y. PSPC1, NONO, and SFPQ Are Expressed in Mouse Sertoli Cells and May Function as Coregulators of Androgen Receptor-Mediated Transcription. Biol Reprod 2006, 75, 352–359. [Google Scholar] [CrossRef] [PubMed]
- King, H.A.; Cobbold, L.C.; Pichon, X.; Pöyry, T.; Wilson, L.A.; Booden, H.; Jukes-Jones, R.; Cain, K.; Lilley, K.S.; Bushell, M.; et al. Remodelling of a Polypyrimidine Tract-Binding Protein Complex during Apoptosis Activates Cellular IRESs. Cell Death Differ 2014, 21, 161–171. [Google Scholar] [CrossRef]
- Kaneko, S.; Rozenblatt-Rosen, O.; Meyerson, M.; Manley, J.L. The Multifunctional Protein P54nrb/PSF Recruits the Exonuclease XRN2 to Facilitate Pre-mRNA 3’ Processing and Transcription Termination. Genes Dev 2007, 21, 1779–1789. [Google Scholar] [CrossRef]
- Fox, A.H.; Lam, Y.W.; Leung, A.K.L.; Lyon, C.E.; Andersen, J.; Mann, M.; Lamond, A.I. Paraspeckles: A Novel Nuclear Domain. Curr Biol 2002, 12, 13–25. [Google Scholar] [CrossRef]
- Fox, A.H.; Lamond, A.I. Paraspeckles. Cold Spring Harb Perspect Biol 2010, 2, a000687. [Google Scholar] [CrossRef]
- McCluggage, F.; Fox, A.H. Paraspeckle Nuclear Condensates: Global Sensors of Cell Stress? Bioessays 2021, 43, e2000245. [Google Scholar] [CrossRef]
- Bond, C.S.; Fox, A.H. Paraspeckles: Nuclear Bodies Built on Long Noncoding RNA. J Cell Biol 2009, 186, 637–644. [Google Scholar] [CrossRef]
- Takeiwa, T.; Mitobe, Y.; Ikeda, K.; Horie-Inoue, K.; Inoue, S. Roles of Splicing Factors in Hormone-Related Cancer Progression. Int J Mol Sci 2020, 21, 1551. [Google Scholar] [CrossRef]
- Knott, G.J.; Bond, C.S.; Fox, A.H. The DBHS Proteins SFPQ, NONO and PSPC1: A Multipurpose Molecular Scaffold. Nucleic Acids Res 2016, 44, 3989–4004. [Google Scholar] [CrossRef]
- Kiesler, E.; Miralles, F.; Ostlund Farrants, A.-K.; Visa, N. The Hrp65 Self-Interaction Is Mediated by an Evolutionarily Conserved Domain and Is Required for Nuclear Import of Hrp65 Isoforms That Lack a Nuclear Localization Signal. J Cell Sci 2003, 116, 3949–3956. [Google Scholar] [CrossRef] [PubMed]
- Knott, G.J.; Lee, M.; Passon, D.M.; Fox, A.H.; Bond, C.S. Caenorhabditis Elegans NONO-1: Insights into DBHS Protein Structure, Architecture, and Function. Protein Sci 2015, 24, 2033–2043. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.R.; Rubin, G.M. Molecular Analysis of No-on-Transient A, a Gene Required for Normal Vision in Drosophila. Neuron 1990, 4, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Laurenzi, T.; Palazzolo, L.; Taiana, E.; Saporiti, S.; Ben Mariem, O.; Guerrini, U.; Neri, A.; Eberini, I. Molecular Modelling of NONO and SFPQ Dimerization Process and RNA Recognition Mechanism. Int J Mol Sci 2022, 23, 7626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Zhang, L.X.; Busch, R.K.; Farrés, J.; Busch, H. Purification and Characterization of a DNA-Binding Heterodimer of 52 and 100 kDa from HeLa Cells. Biochem J 1993, 290 Pt 1, 267–272. [Google Scholar] [CrossRef]
- Huang, C.-J.; Das, U.; Xie, W.; Ducasse, M.; Tucker, H.O. Altered Stoichiometry and Nuclear Delocalization of NonO and PSF Promote Cellular Senescence. Aging (Albany NY) 2016, 8, 3356–3374. [Google Scholar] [CrossRef]
- Passon, D.M.; Lee, M.; Rackham, O.; Stanley, W.A.; Sadowska, A.; Filipovska, A.; Fox, A.H.; Bond, C.S. Structure of the Heterodimer of Human NONO and Paraspeckle Protein Component 1 and Analysis of Its Role in Subnuclear Body Formation. Proc Natl Acad Sci U S A 2012, 109, 4846–4850. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Casas Garcia, G.P.; Perugini, M.A.; Fox, A.H.; Bond, C.S.; Lee, M. Crystal Structure of a SFPQ/PSPC1 Heterodimer Provides Insights into Preferential Heterodimerization of Human DBHS Family Proteins. J Biol Chem 2018, 293, 6593–6602. [Google Scholar] [CrossRef]
- Schell, B.; Legrand, P.; Fribourg, S. Crystal Structure of SFPQ-NONO Heterodimer. Biochimie 2022, 198, 1–7. [Google Scholar] [CrossRef]
- Feng, P.; Li, L.; Deng, T.; Liu, Y.; Ling, N.; Qiu, S.; Zhang, L.; Peng, B.; Xiong, W.; Cao, L.; et al. NONO and Tumorigenesis: More than Splicing. J Cell Mol Med 2020, 24, 4368–4376. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Dye, B.T.; Pérez, I.; Barnard, D.C.; Thompson, A.B.; Patton, J.G. PSF and P54nrb Bind a Conserved Stem in U5 snRNA. RNA 2002, 8, 1334–1347. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-B.; Xiang, J.-F.; Li, X.; Xu, Y.; Xue, W.; Huang, M.; Wong, C.C.; Sagum, C.A.; Bedford, M.T.; Yang, L.; et al. Protein Arginine Methyltransferase CARM1 Attenuates the Paraspeckle-Mediated Nuclear Retention of mRNAs Containing IRAlus. Genes Dev 2015, 29, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Hupalowska, A.; Jedrusik, A.; Zhu, M.; Bedford, M.T.; Glover, D.M.; Zernicka-Goetz, M. CARM1 and Paraspeckles Regulate Pre-Implantation Mouse Embryo Development. Cell 2018, 175, 1902–1916.e13. [Google Scholar] [CrossRef] [PubMed]
- Fisher, E.; Feng, J. RNA Splicing Regulators Play Critical Roles in Neurogenesis. Wiley Interdiscip Rev RNA 2022, 13, e1728. [Google Scholar] [CrossRef] [PubMed]
- Sury, M.D.; McShane, E.; Hernandez-Miranda, L.R.; Birchmeier, C.; Selbach, M. Quantitative Proteomics Reveals Dynamic Interaction of C-Jun N-Terminal Kinase (JNK) with RNA Transport Granule Proteins Splicing Factor Proline- and Glutamine-Rich (Sfpq) and Non-POU Domain-Containing Octamer-Binding Protein (Nono) during Neuronal Differentiation. Mol Cell Proteomics 2015, 14, 50–65. [Google Scholar] [CrossRef]
- Major, A.T.; Hogarth, C.A.; Young, J.C.; Kurihara, Y.; Jans, D.A.; Loveland, K.L. Dynamic Paraspeckle Component Localisation during Spermatogenesis. Reproduction 2019, 158, 267–280. [Google Scholar] [CrossRef]
- Cheng, R.; Zhu, S.; Guo, S.; Min, L.; Xing, J.; Guo, Q.; Li, P.; Zhang, S. Downregulation of NONO Induces Apoptosis, Suppressing Growth and Invasion in Esophageal Squamous Cell Carcinoma. Oncol Rep 2018, 39, 2575–2583. [Google Scholar] [CrossRef]
- Brown, S.A.; Ripperger, J.; Kadener, S.; Fleury-Olela, F.; Vilbois, F.; Rosbash, M.; Schibler, U. PERIOD1-Associated Proteins Modulate the Negative Limb of the Mammalian Circadian Oscillator. Science 2005, 308, 693–696. [Google Scholar] [CrossRef]
- Luo, W.; Guo, F.; McMahon, A.; Couvertier, S.; Jin, H.; Diaz, M.; Fieldsend, A.; Weerapana, E.; Rosbash, M. NonA and CPX Link the Circadian Clockwork to Locomotor Activity in Drosophila. Neuron 2018, 99, 768–780.e3. [Google Scholar] [CrossRef]
- Hu, Z.; Dong, L.; Li, S.; Li, Z.; Qiao, Y.; Li, Y.; Ding, J.; Chen, Z.; Wu, Y.; Wang, Z.; et al. Splicing Regulator P54nrb /Non-POU Domain-Containing Octamer-Binding Protein Enhances Carcinogenesis Through Oncogenic Isoform Switch of MYC Box-Dependent Interacting Protein 1 in Hepatocellular Carcinoma. Hepatology 2020, 72, 548–568. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Su, X.; Burley, S.K.; Zheng, X.F.S. mTOR Regulates Aerobic Glycolysis through NEAT1 and Nuclear Paraspeckle-Mediated Mechanism in Hepatocellular Carcinoma. Theranostics 2022, 12, 3518–3533. [Google Scholar] [CrossRef] [PubMed]
- Iino, K.; Mitobe, Y.; Ikeda, K.; Takayama, K.-I.; Suzuki, T.; Kawabata, H.; Suzuki, Y.; Horie-Inoue, K.; Inoue, S. RNA-Binding Protein NONO Promotes Breast Cancer Proliferation by Post-Transcriptional Regulation of SKP2 and E2F8. Cancer Sci 2020, 111, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.-I.; Suzuki, T.; Fujimura, T.; Yamada, Y.; Takahashi, S.; Homma, Y.; Suzuki, Y.; Inoue, S. Dysregulation of Spliceosome Gene Expression in Advanced Prostate Cancer by RNA-Binding Protein PSF. Proc Natl Acad Sci U S A 2017, 114, 10461–10466. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, R.; Osawa, T.; Sasaki, Y.; Yamamoto, S.; Anai, M.; Izumi, K.; Matsumura, Y.; Sakai, J.; Aburatani, H.; Mizokami, A.; et al. Overexpression of P54nrb/NONO Induces Differential EPHA6 Splicing and Contributes to Castration-Resistant Prostate Cancer Growth. Oncotarget 2018, 9, 10510–10524. [Google Scholar] [CrossRef]
- Clark, J.; Lu, Y.J.; Sidhar, S.K.; Parker, C.; Gill, S.; Smedley, D.; Hamoudi, R.; Linehan, W.M.; Shipley, J.; Cooper, C.S. Fusion of Splicing Factor Genes PSF and NonO (P54nrb) to the TFE3 Gene in Papillary Renal Cell Carcinoma. Oncogene 1997, 15, 2233–2239. [Google Scholar] [CrossRef]
- Skalsky, Y.M.; Ajuh, P.M.; Parker, C.; Lamond, A.I.; Goodwin, G.; Cooper, C.S. PRCC, the Commonest TFE3 Fusion Partner in Papillary Renal Carcinoma Is Associated with Pre-mRNA Splicing Factors. Oncogene 2001, 20, 178–187. [Google Scholar] [CrossRef]
- Mathur, M.; Das, S.; Samuels, H.H. PSF-TFE3 Oncoprotein in Papillary Renal Cell Carcinoma Inactivates TFE3 and P53 through Cytoplasmic Sequestration. Oncogene 2003, 22, 5031–5044. [Google Scholar] [CrossRef]
- Udayakumar, D.; Bladen, C.L.; Hudson, F.Z.; Dynan, W.S. Distinct Pathways of Nonhomologous End Joining That Are Differentially Regulated by DNA-Dependent Protein Kinase-Mediated Phosphorylation. J Biol Chem 2003, 278, 41631–41635. [Google Scholar] [CrossRef]
- Bladen, C.L.; Udayakumar, D.; Takeda, Y.; Dynan, W.S. Identification of the Polypyrimidine Tract Binding Protein-Associated Splicing Factor.P54(Nrb) Complex as a Candidate DNA Double-Strand Break Rejoining Factor. J Biol Chem 2005, 280, 5205–5210. [Google Scholar] [CrossRef]
- Alfano, L.; Costa, C.; Caporaso, A.; Altieri, A.; Indovina, P.; Macaluso, M.; Giordano, A.; Pentimalli, F. NONO Regulates the Intra-S-Phase Checkpoint in Response to UV Radiation. Oncogene 2016, 35, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Lukong, K.E.; Huot, M.-E.; Richard, S. BRK Phosphorylates PSF Promoting Its Cytoplasmic Localization and Cell Cycle Arrest. Cell Signal 2009, 21, 1415–1422. [Google Scholar] [CrossRef]
- Rajesh, C.; Baker, D.K.; Pierce, A.J.; Pittman, D.L. The Splicing-Factor Related Protein SFPQ/PSF Interacts with RAD51D and Is Necessary for Homology-Directed Repair and Sister Chromatid Cohesion. Nucleic Acids Res 2011, 39, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Lone, B.A.; Siraj, F.; Sharma, I.; Verma, S.; Karna, S.K.L.; Ahmad, F.; Nagar, P.; Sachidanandan, C.; Pokharel, Y.R. Non-POU Domain-Containing Octomer-Binding (NONO) Protein Expression and Stability Promotes the Tumorigenicity and Activation of Akt/MAPK/β-Catenin Pathways in Human Breast Cancer Cells. Cell Commun Signal 2023, 21, 157. [Google Scholar] [CrossRef]
- Roberts, E.C.; Hammond, K.; Traish, A.M.; Resing, K.A.; Ahn, N.G. Identification of G2/M Targets for the MAP Kinase Pathway by Functional Proteomics. Proteomics 2006, 6, 4541–4553. [Google Scholar] [CrossRef] [PubMed]
- Casado, P.; Prado, M.A.; Zuazua-Villar, P.; Del Valle, E.; Artime, N.; Cabal-Hierro, L.; Rupérez, P.; Burlingame, A.L.; Lazo, P.S.; Ramos, S. Microtubule Interfering Agents and KSP Inhibitors Induce the Phosphorylation of the Nuclear Protein P54(Nrb), an Event Linked to G2/M Arrest. J Proteomics 2009, 71, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Shav-Tal, Y.; Cohen, M.; Lapter, S.; Dye, B.; Patton, J.G.; Vandekerckhove, J.; Zipori, D. Nuclear Relocalization of the Pre-mRNA Splicing Factor PSF during Apoptosis Involves Hyperphosphorylation, Masking of Antigenic Epitopes, and Changes in Protein Interactions. Mol Biol Cell 2001, 12, 2328–2340. [Google Scholar] [CrossRef]
- Scully, R.; Panday, A.; Elango, R.; Willis, N.A. DNA Double-Strand Break Repair-Pathway Choice in Somatic Mammalian Cells. Nat Rev Mol Cell Biol 2019, 20, 698–714. [Google Scholar] [CrossRef]
- Lieber, M.R. The Mechanism of Double-Strand DNA Break Repair by the Nonhomologous DNA End-Joining Pathway. Annu Rev Biochem 2010, 79, 181–211. [Google Scholar] [CrossRef]
- Li, S.; Kuhne, W.W.; Kulharya, A.; Hudson, F.Z.; Ha, K.; Cao, Z.; Dynan, W.S. Involvement of P54(Nrb), a PSF Partner Protein, in DNA Double-Strand Break Repair and Radioresistance. Nucleic Acids Res 2009, 37, 6746–6753. [Google Scholar] [CrossRef]
- Salton, M.; Lerenthal, Y.; Wang, S.-Y.; Chen, D.J.; Shiloh, Y. Involvement of Matrin 3 and SFPQ/NONO in the DNA Damage Response. Cell Cycle 2010, 9, 1568–1576. [Google Scholar] [CrossRef] [PubMed]
- Udayakumar, D.; Dynan, W.S. Characterization of DNA Binding and Pairing Activities Associated with the Native SFPQ·NONO DNA Repair Protein Complex. Biochem Biophys Res Commun 2015, 463, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Z.; Shu, F.-J.; Xiong, H.; Phillips, A.C.; Dynan, W.S. Double-Strand Break Repair Deficiency in NONO Knockout Murine Embryonic Fibroblasts and Compensation by Spontaneous Upregulation of the PSPC1 Paralog. Nucleic Acids Res 2014, 42, 9771–9780. [Google Scholar] [CrossRef] [PubMed]
- Krietsch, J.; Caron, M.-C.; Gagné, J.-P.; Ethier, C.; Vignard, J.; Vincent, M.; Rouleau, M.; Hendzel, M.J.; Poirier, G.G.; Masson, J.-Y. PARP Activation Regulates the RNA-Binding Protein NONO in the DNA Damage Response to DNA Double-Strand Breaks. Nucleic Acids Res 2012, 40, 10287–10301. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.; Stark, J.M.; Ommundsen, M.; Jasin, M. Rad51 Overexpression Promotes Alternative Double-Strand Break Repair Pathways and Genome Instability. Oncogene 2004, 23, 546–553. [Google Scholar] [CrossRef]
- Bertrand, P.; Lambert, S.; Joubert, C.; Lopez, B.S. Overexpression of Mammalian Rad51 Does Not Stimulate Tumorigenesis While a Dominant-Negative Rad51 Affects Centrosome Fragmentation, Ploidy and Stimulates Tumorigenesis, in P53-Defective CHO Cells. Oncogene 2003, 22, 7587–7592. [Google Scholar] [CrossRef] [PubMed]
- Morozumi, Y.; Takizawa, Y.; Takaku, M.; Kurumizaka, H. Human PSF Binds to RAD51 and Modulates Its Homologous-Pairing and Strand-Exchange Activities. Nucleic Acids Res 2009, 37, 4296–4307. [Google Scholar] [CrossRef]
- Rajesh, C.; Gruver, A.M.; Basrur, V.; Pittman, D.L. The Interaction Profile of Homologous Recombination Repair Proteins RAD51C, RAD51D and XRCC2 as Determined by Proteomic Analysis. Proteomics 2009, 9, 4071–4086. [Google Scholar] [CrossRef]
- Straub, T.; Grue, P.; Uhse, A.; Lisby, M.; Knudsen, B.R.; Tange, T.O.; Westergaard, O.; Boege, F. The RNA-Splicing Factor PSF/P54 Controls DNA-Topoisomerase I Activity by a Direct Interaction. J Biol Chem 1998, 273, 26261–26264. [Google Scholar] [CrossRef]
- Kawano, S.; Miyaji, M.; Ichiyasu, S.; Tsutsui, K.M.; Tsutsui, K. Regulation of DNA Topoisomerase IIbeta through RNA-Dependent Association with Heterogeneous Nuclear Ribonucleoprotein U (hnRNP U). J Biol Chem 2010, 285, 26451–26460. [Google Scholar] [CrossRef]
- Kuhnert, A.; Schmidt, U.; Monajembashi, S.; Franke, C.; Schlott, B.; Grosse, F.; Greulich, K.O.; Saluz, H.-P.; Hänel, F. Proteomic Identification of PSF and P54(Nrb) as TopBP1-Interacting Proteins. J Cell Biochem 2012, 113, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Klaric, J.A.; Wüst, S.; Panier, S. New Faces of Old Friends: Emerging New Roles of RNA-Binding Proteins in the DNA Double-Strand Break Response. Front Mol Biosci 2021, 8, 668821. [Google Scholar] [CrossRef] [PubMed]
- O’Shaughnessy, A.M.; Grenon, M.; Gilbert, C.; Toh, G.W.-L.; Green, C.M.; Lowndes, N.F. Multiple Approaches to Study S. Cerevisiae Rad9, a Prototypical Checkpoint Protein. Methods Enzymol 2006, 409, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Kai, M.; Furuya, K.; Paderi, F.; Carr, A.M.; Wang, T.S.F. Rad3-Dependent Phosphorylation of the Checkpoint Clamp Regulates Repair-Pathway Choice. Nat Cell Biol 2007, 9, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Luo, W.; Wang, Y. PARP-1 and Its Associated Nucleases in DNA Damage Response. DNA Repair (Amst) 2019, 81, 102651. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, R.; Kraus, W.L. The PARP Side of the Nucleus: Molecular Actions, Physiological Outcomes, and Clinical Targets. Mol Cell 2010, 39, 8–24. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, L.; Li, Z.; Li, S.; Dynan, W.S. SFPQ•NONO and XLF Function Separately and Together to Promote DNA Double-Strand Break Repair via Canonical Nonhomologous End Joining. Nucleic Acids Res 2017, 45, 1848–1859. [Google Scholar] [CrossRef]
- Yang, X.; Qian, K. Protein O-GlcNAcylation: Emerging Mechanisms and Functions. Nat Rev Mol Cell Biol 2017, 18, 452–465. [Google Scholar] [CrossRef]
- Cui, Y.; Xie, R.; Zhang, X.; Liu, Y.; Hu, Y.; Li, Y.; Liu, X.; Yu, X.; Wu, C. OGA Is Associated with Deglycosylation of NONO and the KU Complex during DNA Damage Repair. Cell Death Dis 2021, 12, 622. [Google Scholar] [CrossRef]
- Sakasai, R.; Tibbetts, R. RNF8-Dependent and RNF8-Independent Regulation of 53BP1 in Response to DNA Damage. J Biol Chem 2008, 283, 13549–13555. [Google Scholar] [CrossRef]
- Deshar, R.; Yoo, W.; Cho, E.-B.; Kim, S.; Yoon, J.-B. RNF8 Mediates NONO Degradation Following UV-Induced DNA Damage to Properly Terminate ATR-CHK1 Checkpoint Signaling. Nucleic Acids Res 2019, 47, 762–778. [Google Scholar] [CrossRef] [PubMed]
- Petti, E.; Buemi, V.; Zappone, A.; Schillaci, O.; Broccia, P.V.; Dinami, R.; Matteoni, S.; Benetti, R.; Schoeftner, S. SFPQ and NONO Suppress RNA:DNA-Hybrid-Related Telomere Instability. Nat Commun 2019, 10, 1001. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. RNaseH1 Regulates TERRA-Telomeric DNA Hybrids and Telomere Maintenance in ALT Tumour Cells. Nat Commun 2014, 5, 5220. [Google Scholar] [CrossRef] [PubMed]
- Brieño-Enríquez, M.A.; Moak, S.L.; Abud-Flores, A.; Cohen, P.E. Characterization of Telomeric Repeat-Containing RNA (TERRA) Localization and Protein Interactions in Primordial Germ Cells of the Mouse†. Biol Reprod 2019, 100, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Gigi, V.; Lewis, S.; Shestova, O.; Mijušković, M.; Deriano, L.; Meng, W.; Luning Prak, E.T.; Roth, D.B. RAG2 Mutants Alter DSB Repair Pathway Choice in Vivo and Illuminate the Nature of “Alternative NHEJ. ” Nucleic Acids Res 2014, 42, 6352–6364. [Google Scholar] [CrossRef]
- Chang, H.H.Y.; Pannunzio, N.R.; Adachi, N.; Lieber, M.R. Non-Homologous DNA End Joining and Alternative Pathways to Double-Strand Break Repair. Nat Rev Mol Cell Biol 2017, 18, 495–506. [Google Scholar] [CrossRef]
- de Silva, H.C.; Lin, M.Z.; Phillips, L.; Martin, J.L.; Baxter, R.C. IGFBP-3 Interacts with NONO and SFPQ in PARP-Dependent DNA Damage Repair in Triple-Negative Breast Cancer. Cell Mol Life Sci 2019, 76, 2015–2030. [Google Scholar] [CrossRef]
- Bi, O.; Anene, C.A.; Nsengimana, J.; Shelton, M.; Roberts, W.; Newton-Bishop, J.; Boyne, J.R. SFPQ Promotes an Oncogenic Transcriptomic State in Melanoma. Oncogene 2021, 40, 5192–5203. [Google Scholar] [CrossRef]
- Liu, P.Y.; Erriquez, D.; Marshall, G.M.; Tee, A.E.; Polly, P.; Wong, M.; Liu, B.; Bell, J.L.; Zhang, X.D.; Milazzo, G.; et al. Effects of a Novel Long Noncoding RNA, lncUSMycN, on N-Myc Expression and Neuroblastoma Progression. J Natl Cancer Inst 2014, 106, dju113. [Google Scholar] [CrossRef]
- Zhang, S.; Cooper, J.A.; Chong, Y.S.; Naveed, A.; Mayoh, C.; Jayatilleke, N.; Liu, T.; Amos, S.; Kobelke, S.; Marshall, A.C.; et al. NONO Enhances mRNA Processing of Super-Enhancer-Associated GATA2 and HAND2 Genes in Neuroblastoma. EMBO Rep 2023, 24, e54977. [Google Scholar] [CrossRef]
- Kameoka, S.; Duque, P.; Konarska, M.M. P54(Nrb) Associates with the 5’ Splice Site within Large Transcription/Splicing Complexes. EMBO J 2004, 23, 1782–1791. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
DBHS Family Nomenclature. NonO has also been referred to as 54kDa nuclear RNA binding protein (p54nrb), and as 55kDa nuclear matrix protein (nmt55). SFPQ was originally named PTB-associated splicing factor: PSF. NonO and SFPQ are two members of the DBHS family. The DBHS conserved regions contain two RNA Recognition Motifs (RRMs), a NONA/ParaSpeckle (NOPS) domain, and the Coiled Coil region. SFPQ also contains an N-terminal RGG region and an uncharacterized DNA-binding domain (DBD) at the N-terminus end of the DBHS region. NonO has a highly charged helix-turn-helix (HTH) region C-terminal to its DBHS region which has been suggested to have DNA-binding activity.
Figure 1.
DBHS Family Nomenclature. NonO has also been referred to as 54kDa nuclear RNA binding protein (p54nrb), and as 55kDa nuclear matrix protein (nmt55). SFPQ was originally named PTB-associated splicing factor: PSF. NonO and SFPQ are two members of the DBHS family. The DBHS conserved regions contain two RNA Recognition Motifs (RRMs), a NONA/ParaSpeckle (NOPS) domain, and the Coiled Coil region. SFPQ also contains an N-terminal RGG region and an uncharacterized DNA-binding domain (DBD) at the N-terminus end of the DBHS region. NonO has a highly charged helix-turn-helix (HTH) region C-terminal to its DBHS region which has been suggested to have DNA-binding activity.
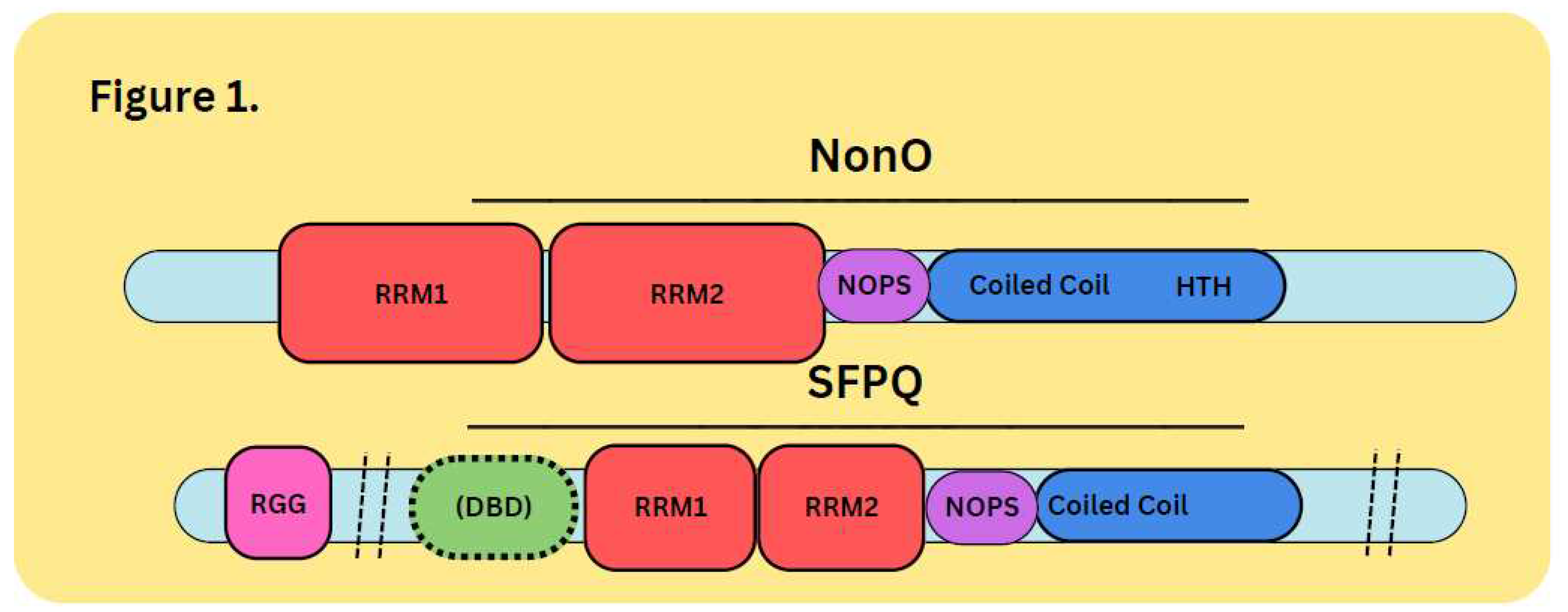
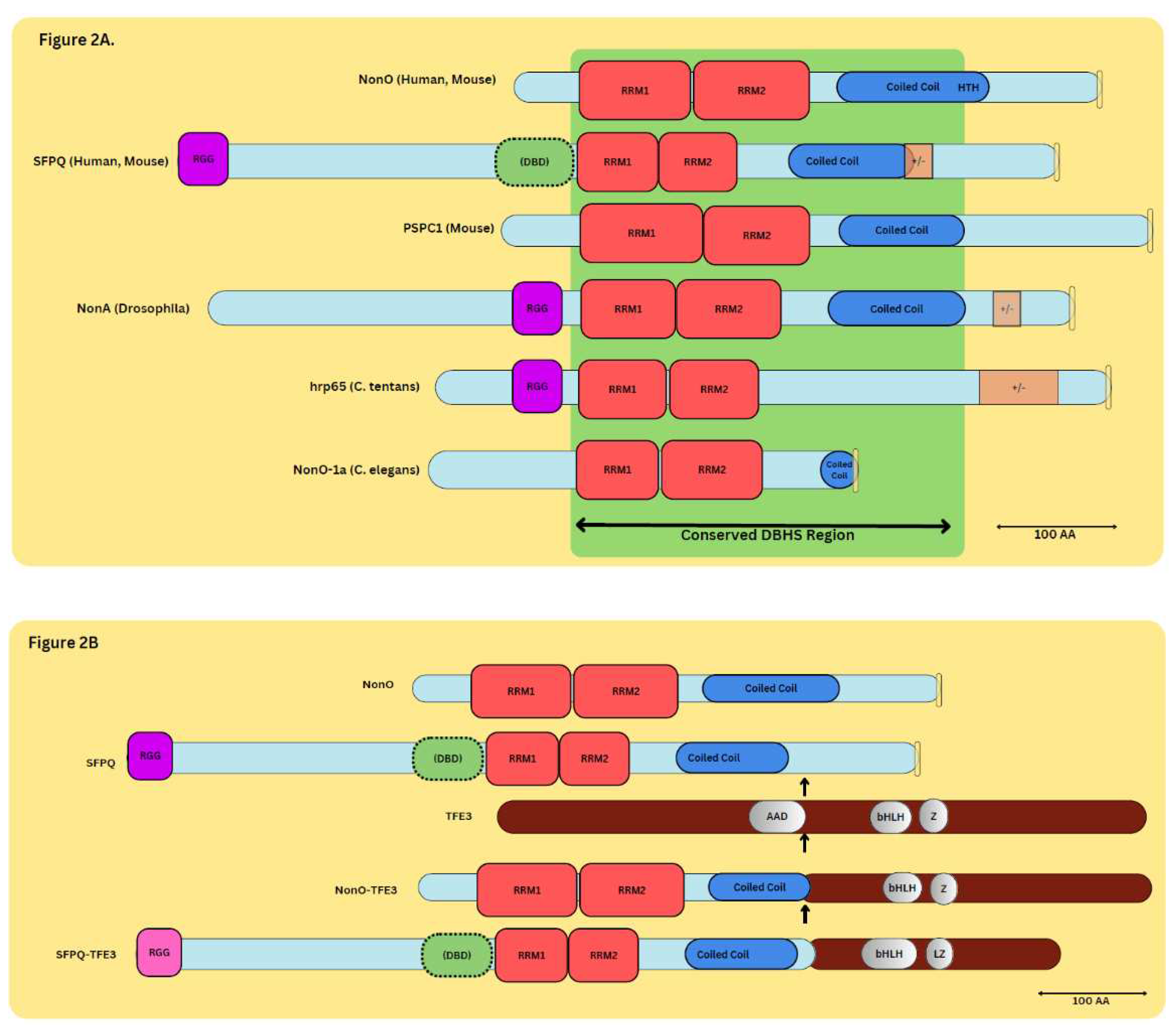
Figure 2.
Alignment of DBHS Family Proteins. (A) Structural elements of DBHS family proteins: NonO, SFPQ, PSPC1, NonA, hrp65, and NonO-1a. The C-terminus contains the conserved DBHS region consisting of (from N-terminus to C-terminus) RNA-Binding Domains (RRM1, 2), NONA/ParaSpeckle domain (NOPS, not pictured), and the Coiled-Coil domain. Also pictured are positively/negatively charged residue regions (+/-), HTH domain (pink), and NLS regions (yellow). (B). NonO, SFPQ, and TFE3 chimera proteins. Structural representations of wild-type NonO, SFPQ, and TFE3 (Transcription Factor Binding to IGHM Enhancer 3) chimera proteins. Scales are indicated with bars representing 100 amino acids (as) in scale bars at lower right.
Figure 2.
Alignment of DBHS Family Proteins. (A) Structural elements of DBHS family proteins: NonO, SFPQ, PSPC1, NonA, hrp65, and NonO-1a. The C-terminus contains the conserved DBHS region consisting of (from N-terminus to C-terminus) RNA-Binding Domains (RRM1, 2), NONA/ParaSpeckle domain (NOPS, not pictured), and the Coiled-Coil domain. Also pictured are positively/negatively charged residue regions (+/-), HTH domain (pink), and NLS regions (yellow). (B). NonO, SFPQ, and TFE3 chimera proteins. Structural representations of wild-type NonO, SFPQ, and TFE3 (Transcription Factor Binding to IGHM Enhancer 3) chimera proteins. Scales are indicated with bars representing 100 amino acids (as) in scale bars at lower right.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



