
Submitted:
10 October 2023
Posted:
11 October 2023
You are already at the latest version
Abstract
Bacillus species produce different classes of antimicrobial and antioxidant substances: peptides or proteins with different structural composition and molecular mass and a broad range of volatile organic compounds (VOCs), some of which may serve as biomarkers for microorganism identification. The aim of this study is the identification of biologically active compounds synthesized by five Bacillus species using gas chromatography coupled to mass spectrometry (GC–MS). The current study profoundly enhances the knowledge of antibacterial and antioxidant metabolites ensuring the unambiguous identification of VOCs produced by some Bacillus species, which were isolated from vegetable samples of potato, carrot, and tomato. Phylogenetic and biochemical studies were used to identify the bacterial isolates after culturing. Phylogenetic analysis proved that five bacterial isolates BSS12, BSS13, BSS16, BSS21, and BSS25 showed 99% nucleotide sequence similarities with Bacillus safensis AS-08, Bacillus cereus WAB2133, and Bacillus acidiproducens NiuFun, Bacillus toyonesis FORT 102 and Bacillus thuringiensis F3, respectively. The crude extract was prepared from bacterial isolates to assess the antibiotic resistance potency and the antimicrobial potential against various targeted multidrug-resistant strains, including Candida albicans, Candida krusei, Enterococcus hirae, Escherichia coli, Klebsiella aerogenes, Klebsiella pneumoniae, Staphylococcus aureus, Staphylococcus epidermidis, Streptococcus group B, Streptococcus mutans, Shigella sonnei, Salmonella enteritidis, Serratia marcescens, Pseudomonas aeruginosa, and Proteus vulgaris. GC–MS analysis of bacterial strains found that VOCs from Bacillus species come in a variety of chemical forms, such as ketones, alcohols, terpenoids, alkenes, etc. Overall, 69 volatile organic compounds were identified from five Bacillus species, and all five were found to share different chemical classes of volatile organic components, which have a variety of pharmacological applications. However, 8 antibacterial compounds with different concentrations were commonly found in all five species: acetoin, acetic acid, butanoic acid, 2-methyl-, oxime-, methoxy-phenyl, phenol, 1,2-benzenedicarboxylic acid, bis(2-methylpropyl) ester, nonanoic acid, and hexadecanoic acid, methyl. The biology and physiology of Bacillus can be better understood using these results, which can also be used to create novel biotechnological procedures and applications. Moreover, because of its exceptional ability to synthesize and produce a variety of different antibacterial compounds, Bacillus species can serve as natural and universal carriers for antibiotic compounds in the form of probiotic cultures and strains to fight different pathogens, including mycobacteria.
Keywords:
antimicrobial activity
; Bacillus subtilis
; Bacillus thuringiensis
; Bacillus toyonensis
; Bacillus acidiproducens
; Bacillus cereus
; Bacillus safensis
; GC–MC analysis
; volatile organic compounds.
1. Introduction
The emergence of bacterial strains that previously were susceptible to existing antibiotics but now cause serious infectious diseases makes it necessary to find and create novel treatments for these illnesses [1]. The most well-known and clinically significant example of this issue is the rise in multidrug-resistant strains of Staphylococcus aureus (MRSA), which is most pathogenic and leads to the formation of an abscess. Moreover, it can cause pneumonia, endocarditis, and osteomyelitis. According to some investigations, MRSA is resistant not only to some antibiotics such as methicillin, macrolides, tetracycline, aminoglycosides, and chloramphenicol but also to some disinfectants [2].
In order to create new antibiotic treatments or disinfectants, it is also necessary to look for and analyze compounds that have bactericidal or bacteriostatic capabilities against human and animal infections. Currently, analysis of the potential of natural compounds from various sources as antimicrobials has received significant attention in addition to the synthesis of new chemical substances. The observation of antagonism, or the interaction between microorganisms, is frequently the starting point for the development of antibiotics with activity against human infections. The creation and release of chemicals that impede or entirely stop the growth of other species serve as the physical manifestation of this hostility. Under natural circumstances, an agent released by a microbe that prevents the growth of another organism has an edge in the competition for environmental resources. The majority of antibiotics used in medicine are secreted by or derived from bacteria. Hence, it is a fact that the bacterial world has a vast repository of potentially antimicrobial chemicals that have not yet been identified or exploited. In this respect, members of the genus Bacillus are recognized as manufacturers of a wide variety of enzymes and antibacterial substances. For example, 23 peptide antibiotics are produced by Bacillus brevis, while Bacillus subtilis produces roughly 70 antimicrobials which are ribosomal peptides, non-ribosomal peptides, polyketides, hybrids, and volatile compounds [3,4]. Consequently, there is increased interest in taking these compounds into consideration as disjunctive antimicrobials for the healing of human infections [5,6,7,8,9].
Nowadays, a novel strategy for the management and prevention of numerous infectious illnesses is the use of bacterial probiotic strains and their metabolic products. Probiotics from the Bacillus genus have been shown to exhibit antibacterial effects in experiments on animals. As an affordable and infrequently resistant alternative to antibiotics, the application of bacteriocins and antimicrobial peptides synthesized by probiotic strains is recommended [13,14]. They hold promise for clinical usage since many of these molecules are efficient and affordable [15]. Due to their desirable medicinal qualities, such as their antibacterial, antiviral, anticancer, and contraceptive effects, a few natural peptides from bacterial isolates have demonstrated potential. Furthermore, when combined with traditional antibiotics, they have been demonstrated to offer protection against systemic and topical infections. Therefore, the justification for using probiotics in medicine is founded on the notion that giving oral or topically applied probiotics could restore the depleted state of the human microbiome [16].
Development, registration, and commercialization of biocontrol drugs based on microbial antagonists have advanced significantly during the past few decades. Although their use has side effects on both human and animal health. Bacillus species compete directly with fungal pathogens for resources and habitats and through a variety of processes such as the generation of siderophores. Hence, they also indirectly create systemic resistance or stimulate the growth of plants [17,18]. Moreover, they produce a vast array of volatile organic compounds with strong inhibitory potential against plant pathogens: alcohols, alkenes, benzenoids, terpenoids, ketones, sulfur-containing compounds, and others [19,20]. Non-volatile components of these metabolites have gotten a lot of scientific interest, whereas volatile components are examined less frequently. Numerous applications in biology, environmental sciences, health, the food industry, and national security include the study and detection of volatile organic compounds (VOCs) that come from or interact with creatures ranging from bacteria to people. Low-molecular-weight organic molecules with a lipophilic nature and a low boiling point are known as volatile organic compounds (VOCs) [21]. According to several studies, VOCs released by bacteria may help plants by fostering development, triggering defence mechanisms, and inhibiting or removing dangerous infections [22,23,24,25,26]. Furthermore, VOCs released by microorganisms are biodegradable as they are naturally occurring compounds. As a result, using VOCs produced by microorganisms is a sustainable method of crop protection and promotion. According to some recent studies, because they can stop certain pathogenic fungi's mycelial growth and spore germination, VOCs produced by B. subtilis have been suggested as an alternate control approach for postharvest fruit illnesses [27]. For instance, different VOCs produced by B. subtilis TB09 and TB72, such as nonan-2-one, β-benzeneethanamine, and 2-methyl-1,4-diazine effectively controlled the anthracnose pathogen on postharvest mangoes [28]. Likewise, some VOCs synthesized by B. subtilis PPCB001 helped to evaluate its antagonistic activity and it was found that B. subtilis PPCB001 reduces growth one of the imperfect fungi which is called Penicillium crustosum [29]. Additionally, the GC–MS analysis of three bacterial isolates of Bacillus subtilis Md1-42, Bacillus subtilis O-3, and Bacillus subtilis Khozestan2 samples proved the presence of phenol, benzoic acid, 1,2-benzenedicarboxylic acid, bis(2-methylpropyl), methoxyphenyl-oxime, and benzaldehyde, which are known for their antimicrobial and other properties [30].
Overall, representatives of the genus Bacillus are found as producers of a wide range of antimicrobial compounds. The synergetic mechanism of antimicrobial compounds such as VOCs, polyketides, ribosomal peptides, and others explains why they have an increased industrial interest as therapeutic agents, food preservatives and biopesticides. Since Bacillus species have the unique ability to produce a variety of diverse antibacterial chemicals, they can serve as a natural carrier for antibiotics in the form of probiotic cultures and strains to fight different pathogens, including mycobacteria. Nevertheless, the dangerous effects of several antibacterials on humans and animals have prevented some of them from being used medically despite their promising in vitro antimycobacterial activity [31,32].
Another instance of a Bacillus species' employment is in the manufacture of food-grade amylase, glucoamylase, protease, pectinase, and cellulase in a variety of foods [33,34,35,36]. Additionally, many species of Bacillus have been employed to synthesize a number of dietary supplements for human use, including vitamins (such as riboflavin, cobalamin, and inositol) and carotenoids [36,37,38,39]. However, despite the foregoing advantages, these strains have not attracted much interest in the contemporary functional food market because of their relationships with a limited number of human diseases.
This study focused on isolating potential Bacillus species from vegetable samples (potato, carrot, and tomato) and preparing a crude extract from isolated bacterial strains to assess antimicrobial activities against the most common human pathogens. In addition, bacterial isolates were tested for antibiotic resistance using an inhibition zone diameter when determined via the disk diffusion method. GC–MC analysis was performed to determine bioactive compounds from the bacterial isolates. This study will facilitate the development of novel antibiotics against MDR bacterial strains and help to explore possible probiotics typical for the representatives of the genus Bacillus.
2. Results
2.1. Isolation and Identification
A total of n = 25 bacteria strains were isolated and identified using colony morphology, microscopy, biochemical properties, and sugar fermentation. Among all, Gram-strain-positive, rod-shaped, mycelial, and spore-forming bacterial strains were chosen for further verifying tests. The molecular analysis further validated the bacterial strains (BSS25, BSS21, BSS16, BSS13, and BSS12) as Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08.
2.2. Microbial Morphology and Colony Characteristics
The morphology of each colony by different bacterial isolates showed regular, irregular, slightly raised, flat, white, and cream-coloured colonies. By motility test, bacterial isolates were motile and possessed terminal and subterminal spores (Table 1).
2.3. Antimicrobial Potency Evaluation
Five bacterial cultures of BSS25, BSS21, BSS16, BSS13 and BSS12 were tested for their antagonistic activity against 15 pathogens such as Salmonella enterica ATCC 35664, Klebsiella aerogenes ATCC 13048, Serratia marcescens ATCC 13880, Klebsiella pneumoniae ATCC 13883, Streptococcus group B, Escherichia coli ATCC 25922, Candida krusei ATCC 14243, Shigella sonnei ATCC 25931, Streptococcus mutans ATCC 25175, Enterococcus hirae ATCC 10541, Proteus Vulgaris ATCC 6380, Staphylococcus epidermidis ATCC 12228, Pseudomonas aeruginosa ATCC 9027, Staphylococcus aureus ATCC 29213 and Candida albicans ATCC 2091 (Figure 1).
The five extracts showed antibacterial activity against all the bacterial pathogens except Staphylococcus epidermidis ATCC 12228 and Enterococcus hirae ATCC 10541 (Table 2). The strain of Bacillus thuringiensis F3 (BSS25), showed a better zone of inhibition for Staphylococcus aureus ATCC 29213 (35±1,27 mm),Staphylococcus epidermidis ATCC 12228 (37±1,47 mm),Candida albicans ATCC 2091 (36±1,43 mm),Candida krusei ATCC 14243 (37±1,41 mm), Klebsiella aerogenes ATCC 13048 (37±1,27 mm), and Enterococcus hirae ATCC 10541 (37±1,25 mm). Additionally, the strain of Bacillus toyonensis FORT 102 (BSS21) was effective against the pathogens Staphylococcus epidermidis ATCC 12228 (36±1,27), Candida albicans ATCC 2091 (38±1,21 mm), Candida krusei ATCC 14243 (36±1,28 mm), Klebsiella aerogenes ATCC 13048 (36±1,37 mm), and Enterococcus hirae ATCC 10541 (38±1,27 mm).
2.4. Antibiotic Susceptibility Profile of the Isolates
With the exception of bacitromycin (B, 10), polymyxin (PB, 300), and cloxacillin (CX, 5), none of the five Bacillus species examined were resistant to any antibiotics, according to the analysis of the antibiogram (Table 3). All five strains, which are BSS25, BSS21, BSS16, BSS13 and BSS13 strains showed the highest vulnerability to gentamicin (CN, 120) with 39 ± 0,37 mm, 38 ± 0,43 mm, 40± 0,12 mm, 41 ± 0,23 and 39 ± 0,31 sensitivity diameters, respectively
2.5. GC–MS Analysis
According to the results of the GC–MS analysis, crude extracts from different Bacillus bacterium species contained a variety of compounds. Table 4, Table 5, Table 6, Table 7 and Table 8 explain the most important and plentiful components identified in the crude extracts that were subjected to the GC–MS analysis, as well as information about where the chemicals found in this study had previously been identified. These substances exhibited similarities to natural products of a variety of organisms, such as bacterial, plant and fungi origin. In a study, the majority of the compounds detected were derived from volatile substances. Different Bacillus species have been found to produce volatile compounds belonging to various classes, such as alcohols, ketones, fatty acids, and aromatic compounds, in addition to esters and ethers. In strain BSS25, ethyl acetate extraction showed the presence of 33 compounds (Table 4) compared to 37 compounds arising out of the same extraction strain BSS21 (Table 5). In the ethyl acetate extract of the BSS16 bacterial strain were identified 23 compounds (Table 6). Acetoin, benzaldehyde, 3(2H)-thiophenone, dihydro-2-methyl-, propanoic acid, 2-methyl-, and oleic acid were identified in the BSS16 extract with important concentrations of 8.440992%, 4.751143%, 6.072668%, 13.97511%, and 9.677555%, respectively. In the BSS25 bacterial isolate, the major compounds were butanoic acid, 2-methyl- at 29.39467% and 9,12-octadecadienoic acid (Z, Z)- at 11.09688% and other three compounds such as 3(2H)-thiophenone, dihydro-2-methyl-, benzoic acid, tridecyl ester and pentadecanoic acid were identified only in this bacterial extract. In bacterial strain BSS13, some compounds such as acetone, acetic acid, benzaldehyde, hexadecanoic acid, octadecanoic acid, 2-hydroxy-1,3-propanediyl ester, 9-Octadecenoic acid, (E)-, and 9,12-Octadecadienoic acid (Z,Z)- were found with high concentrations 3.660169%, 6.313543%, 6.242524%, 4.45127%, 3.793011%, 9.952046%, and 5.86713%, respectively (Table 7). The solvent with metabolites for isolate BSS12 was ethyl acetate, which also contained 38 chemicals (Table 8), whereas the same solvent with metabolites for isolate BSS13 was found to contain 33 compounds (Table 7). GC–MS analysis for five bacterial (BSS25, BSS21, BSS16, BSS13 and BSS12) analyses also confirmed the presence of the same volatile organic compounds, while some components were found only in some bacterial isolates.
The GC–MS based metabolite profiling of the ethyl acetate extracts of BSS25, BSS21, BSS16, BSS13, and BSS12 bacterial isolates revealed a total of 69 volatile organic substances. Based on the analysis of bacterial isolates, all five isolates were found to share a similar composition of volatile organic components, such as acetoin, acetic acid, butanoic acid, 2-methyl-, oxime-, methoxy-phenyl, phenol, 1,2-benzenedicarboxylic acid, bis(2-methylpropyl) ester, nonanoic acid, and hexadecanoic acid, methyl ester. Their chemical structure details are available, as illustrated in Figure 2.

Table 9.
List of various classes of compounds identified from five Bacillus spp. and their pharmacological activities.
Table 9.
List of various classes of compounds identified from five Bacillus spp. and their pharmacological activities.
| No. | Name | Chemical classes | Known pharmacological activities |
|---|---|---|---|
| 1 | 3-Pentanol, 2-methyl- | alcohols | – |
| 2 | 2,3-Butanediol, [S-(R*,R*)]- | alcohols | – |
| 3 | 1,6-Octadien-3-ol, 3,7-dimethyl- | monoterpene alcohols | anti-inflammatory, anticancer, anti-hyperlipidemic, antimicrobial, antinociceptive, analgesic, anxiolytic, anti-depressive and neuroprotective [40] |
| 4 | 1-Hepten-4-ol | alcohols | – |
| 5 | 1-Nonanol | alcohols | antifungal [41] and antibacterial[42] |
| 6 | (S)-(+)-6-Methyl-1-octanol | alcohols | – |
| 7 | 1-Decanol | alcohols | antibacterial[42], antioxidant and neuroprotective [43] |
| 8 | E-3-Pentadecen-2-ol | alcohols | – |
| 9 | 2-Octanol | alcohols | – |
| 10 | 3-Penten-1-ol | alcohols | – |
| 11 | 2-Nonen-1-ol | alcohols | – |
| 12 | 2,3-Butanediol | alcohols | CNS depressant [44], antimicrobial and antagonistic [45] |
| 13 | 1-Hexanol, 2-ethyl- | alcohols | – |
| 14 | 1-Dodecanol | alcohols | antibacterial [42] |
| 15 | Hexanal | aldehydes | antimicrobial [46] |
| 16 | Nonanal | aldehydes | anti-fungal [47] |
| 17 | Decanal | aldehydes | anti-fungal [48] |
| 18 | Dodecanal | aldehydes | – |
| 19 | 2,4-Decadienal, (E,E)- | aldehydes | flavoring agent, fragrance agent, toxic [49] |
| 20 | Acetic acid | aromatic aldehydes | antibacterial and antifungal, anticancer [50] |
| 21 | Benzaldehyde | carboxylic acids (simple acids) | denaturant and a flavoring agent [51] |
| 22 | Butanoic acid, 2-methyl- | carboxylic acids (simple acids) | laxative [52] |
| 23 | Hexanoic acid | carboxylic acids (fatty acids) | – |
| 24 | (R)-(-)-4-Methylhexanoic acid | carboxylic acids (fatty acids) | – |
| 25 | Hexanoic acid, 2-ethyl- | carboxylic acids (fatty acids) | – |
| 26 | Neodecanoic acid | carboxylic acids (fatty acids) | – |
| 27 | Octanoic acid | carboxylic acids (fatty acids) | anticancer [53], antibacterial [54], antimicrobial [55] |
| 28 | Nonanoic acid | carboxylic acids (fatty acids) | skin-conditioning agent [56], anti-fungal [57] |
| 29 | Propanoic acid, 2-methyl- | carboxylic acids (fatty acids) | – |
| 30 | Butanoic acid | carboxylic acids (fatty acids) | the main energetic substrate of the colonocyte [58] |
| 31 | Pentadecanoic acid | carboxylic acids (fatty acids) | a JAK2/STAT3 signaling inhibitor in breast cancer cells [59], anti-biofilm agent [60] |
| 32 | Oleic Acid | carboxylic acids (fatty acids) | anticancer, anti-inflammatory, wound healing [61] |
| 33 | Hexadecanoic acid | carboxylic acids (fatty acids) | anti-inflammatory [62], antibacterial [63], |
| 34 | Octadecanoic acid | carboxylic acids (fatty acids) | anticancer [64] |
| 35 | 9,12-Octadecadienoic acid (Z,Z)- | carboxylic acids (fatty acids) | used for the treatment or prevention of cardiac arrhythmias [65] |
| 36 | Tiglic acid | carboxylic acids (fatty acids) | – |
| 37 | Decanoic acid | carboxylic acids (fatty acids) | enhances antibacterial effect [66], anti-inflammatory [67] |
| 38 | 9-Octadecenoic acid, (E)- | carboxylic acids (fatty acids) | – |
| 39 | Pentanoic acid | carboxylic acids (fatty acids) | neuroprotective agent and suppresses oxidative stress [68] |
| 40 | Acetone | ketones | – |
| 41 | 2,3-Butanedione | ketones | – |
| 42 | Acetoin | ketones | CNS depressant [44] |
| 43 | 5,9-Undecadien-2-one, 6,10-dimethyl-, (E)- | ketones(sesquiterpenoid) | – |
| 44 | 2,3-Pentanedione | ketones | – |
| 45 | 3-Buten-2-one, 4-(1-cyclopenten-1-yl)-, (E)- | ketones(cyclic) | – |
| 46 | 2-Hydroxy-3-pentanone | ketones (acyloins) | – |
| 47 | Oxime-, methoxy-phenyl | Esters | – |
| 48 | 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | Esters | – |
| 49 | Propanoic acid, 2-methyl-, 3-hydroxy-2,4,4-trimethylpentyl ester | fatty acid esters | – |
| 50 | Hexadecanoic acid, methyl ester | fatty acid esters | shows cardioprotective effect against the ischemia/reperfusion (I/R) injury [70], antibacterial [71], counteracts cyclophosphamide cardiotoxicity [72] |
| 51 | Octadecanoic acid, 2-hydroxy-1,3-propanediyl ester | fatty acid esters | – |
| 52 | Ethane-1,1-diol dibutanoate | fatty acid esters | – |
| 53 | Benzoic acid, 2-ethylhexyl ester | benzoic acid esters | – |
| 54 | 2-Octyl benzoate | benzoic acid esters | – |
| 55 | Benzoic acid 2-methylpentyl ester | benzoic acid esters | – |
| 56 | Benzoic acid, heptyl ester | benzoic acid esters | – |
| 57 | Benzoic acid, tridecyl ester | benzoic acid esters | – |
| 58 | Benzoic acid, undecyl ester | benzoic acid esters | – |
| 59 | 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | phthalate esters | – |
| 60 | Diibutyl phthalate | phthalate esters | – |
| 61 | Formic acid, octyl ester | fatty alcohol esters | – |
| 62 | 1-Propen-2-ol, acetate | fatty alcohol esters | – |
| 63 | Oxirane, (methoxymethyl)- | heterocyclic ethers | – |
| 64 | (2-Aziridinylethyl)amine | amines | – |
| 65 | Cetene | alkenes | – |
| 66 | Benzoic acid | benzenoids | antibacterial and antifungal [73] |
| 67 | Phenol | phenols | desinfectant [74] |
| 68 | 3(2H)-Thiophenone, dihydro-2-methyl- | tetrahydrothiophenes | – |
| 69 | 1,4-Benzenediol, 2,6-bis(1,1-dimethylethyl)- | quinones | – |
2.5. GC–MS Analysis
Five (n=5) bacterial isolates with increased antibacterial activity were isolated from distinct samples. Phylogenetic analysis of the 16S rRNA gene sequences indicated that all the five candidate bacterial isolates, BSS25, BSS21, BSS16, BSS13 and BSS12, belong to five different Bacillus spp., respectively (Figure 3) as they are related to the aforementioned bacterial species in the phylogenetic tree.
Bacillus species, i.e., Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08 were identified as having the highest hit sequence similarity for these bacterial isolates (Table 10). High bootstrap values were obtained following a phylogenetic analysis and tree topology both served to confirm the presumably described taxonomy.
3. Discussion
Microorganisms, including bacteria, archaea, fungi, and even viruses, inhabit diverse environments and contribute to the cycling of nutrients and the production of a wide array of metabolites. Especially extreme microbial diversity, abundance, and structure can have significant implications for the production of various metabolites with diverse functions, including anti-parasitic, antimicrobial, anti-pesticidal anti-cancerous functions. Consequently, these metabolites can have important applications in various fields. The goal of the present study was to investigate the possibility of particular vegetable microbial communities displaying antibacterial properties and to establish the possible relationship between isolated compounds through GC–MS and the antagonistic activity of studied bacterial strains. As a result of various phases of isolation, the identification of diverse general objectives with the selection of bacterial growth conditions, and biochemical tests, nineteen (n=19) different bacterial isolates were detected. In recent years, the ongoing exploration of microbial diversity, along with advancements in culturing techniques, genomics, and metagenomics, has rejuvenated the search for new antibiotics. This is a promising development in the fight against infectious diseases and antibiotic resistance, as it offers the potential for a new generation of antimicrobial agents to tackle previously untreatable infections [75,76]. However, both, mobile genetic elements and inherent characteristics (natural phenotypic traits) causes contribute to the development and spread of antibiotic resistance, making it a complex and evolving problem in healthcare and public health. The inappropriate use of antibiotics, both in clinical settings and in agriculture, accelerates the selection and dissemination of antibiotic-resistant bacteria by providing a selective advantage to those carrying resistance genes. Addressing antibiotic resistance requires a multifaceted approach that includes responsible antibiotic use, surveillance, development of new antibiotics, and strategies to prevent the spread of resistant bacteria [77]. Nearly all antibiotics, with the exception of bacitracin, polymyxin, and cloxacillin, were found to be effective against Bacillus species, i.e., Bacillus thuringiensis F3 (BSS25), Bacillus toyonensis FORT 102 (BSS21), Bacillus acidiproducens NiuFun (BSS16), Bacillus cereus WAB2133 (BSS13), and Bacillus safensis AS-08 (BSS12) (Table 3). Similar findings on the susceptibility of various antibiotic-susceptible Bacillus species were observed in some recent studies [30,78,79,80,81]. Here, resistance in specific Bacillus strains to particular antibiotics can result from both inherent (natural) mechanisms and acquired resistance due to the presence of resistance genes associated with the production of resistance enzymes [82]. The probability of passing on resistance genes to other bacteria, particularly dangerous pathogens, may be lower when resistance is due to inherent (natural) resistance mechanisms rather than acquired resistance through the acquisition of resistance genes. This distinction is important in the context of antibiotic resistance transmission and the potential for the development of multidrug-resistant or extensively drug-resistant bacteria. Since antibiotic resistance has indeed become a serious global concern, and the spread of resistant bacteria through the food chain is one of the pathways contributing to this problem [83]. Although, Isolated Bacillus strains may not necessarily harbor antibiotic resistance genes that can be horizontally transferred to dangerous pathogens, but they can still exhibit natural resistance or insensitivity to a wide variety of antibiotics due to their inherent characteristics. Indeed, further research into Bacillus strains, especially those with unique characteristics or inherent resistance to antibiotics, can be valuable for various applications, including the development of probiotic starter cultures and the production of high-quality, medicinal, and health-promoting substances.
Gas Chromatography–Mass Spectrometry (GC–MS) is a powerful analytical technique commonly used to detect and identify various compounds in biological samples, including microbial cells and their metabolites helped in this study to detect markers in biological material, such as components of microbial cells and metabolites like fatty acids, aldehydes, and phenolic compounds. Additionally, using GC–MS without the need for the preliminary isolation of pure cultures of microorganisms offers several advantages in the case of both endogenous and exogenous microflora, which is especially important when considering the difficulties in cultivating anaerobes. The method's unique benefits were quick analytical times and the capacity to quantify marker content. According to the GC–MS analysis, the Bacillus species produce a variety of chemicals (Table 9), which possess different pharmacological activities such as antiviral, antibacterial, antifungal, antioxidant, anticancer, anti-inflammatory, hyperlipidemic, antimicrobial, antinociceptive, analgesic, anxiolytic, anti-depressive, neuroprotective and so on. Overall, 69 compounds were determined by the GC–MS analysis of their crude metabolites from five Bacillus species. 8 biologically active compounds such as acetoin, acetic acid, butanoic acid, 2-methyl-, oxime-, methoxy-phenyl, phenol, 1,2-benzenedicarboxylic acid, bis(2-methylpropyl) ester, nonanoic acid, and hexadecanoic acid, methyl ester were found as common for all five strains (Table 4, Table 5, Table 6, Table 7, and Table 8 and Figure 2).
Previously, we found that acetic acid is present in bacterial isolates such as Bacillus subtilis O-3, Bacillus subtilis Md1-42, and Bacillus subtilis Khozestan2 [30]. According to the present study acetic acid seems a common organic compound almost for all Bacillus species as its presence is confirmed in other five Bacillus species i.e. Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08. BSS13 has the highest concentration of acetic acid at 6.313543%, while the other four strains were found to share similar low concentrations. Acetic acid is a common organic acid and a component of the volatile organic compounds (VOCs) produced by some bacterial species, including certain strains of Bacillus. Acetic acid is a byproduct of microbial metabolism, particularly in bacteria that undergo fermentation processes or produce acetic acid as part of their metabolic pathways. It has been known for antibacterial and antifungal, anticancer activities [50]. The second common compound for all five strains was acetoin, and its concentration for BSS25, BSS21, BSS16, BSS13, and BSS12 were different as follows 44.063%, 38.25336%, 8.440992%, 0.745862%, and 0.263788%, respectively. Acetoin is a common compound produced by various bacteria, and its concentration can vary among different strains. It is not typically used as a central nervous system (CNS) depressant in medical practice or for recreational purposes. However, in one recent study, it was proven that acetoin has a potent CNS depressant effect [44]. It was found that the third component that all bacteria share is butanoic acid, 2-methyl-, which has an application as a laxative. BSS16 and BSS13 have the highest concentrations of butanoic acid, 2-methyl- at 29.3946% and 31.6913%, while BSS16, BSS13, and BSS12 share very low concentrations at 2.237883%, 3.572379, and 1.894643, respectively. Other 4 compounds common for all five strains, which are oxime-, meth-oxy-phenyl, 1,2-benzenedicarboxylic acid, bis(2-methylpropyl) ester, nonanoic acid, and hexadecanoic acid, methyl ester, were found at low concentrations similarly to each other.
The GC-MS analysis of Bacillus thuringiensis F3 has revealed the presence of several chemical compounds of 2,3-butanedione, 2,3-butanediol, [S-(R*,R*)]-, 1-hepten-4-ol, hexanoic acid, hexanoic acid, 2-ethyl-, octanoic acid, benzaldehyde, (S)-(+)-6-methyl-1-octanol in relatively high concentrations at 15.84782%, 4.077882%, 3.418443%, 1.862169%, 2.90418%, 2.465807%, 2.222227%, and 1.248967%, respectively. The compositional study of Bacillus toyonensis FORT 102 established that it differs from other bacterial strains by composing of 2,3-butanedione (16,97044%), propanoic acid, 2-methyl- (1.406486%), hexadecanoic acid (4.002248%), oleic acid (5.853397%), 9,12-octadecadienoic acid (Z,Z)- (3.796889%), 2,3-pentanedione (2,035411%), 3-pentanol, 2-methyl- (2.916132%), oxirane, (methoxymethyl)- (2.409396%), 2,3-butanediol, [S-(R*,R*)]- (7.796162%), hexadecanoic acid (4.002248%), 9,12-octadecadienoic acid (Z,Z)- (3.796889%). The presence of some valuable compounds in Bacillus acidiproducens NiuFun also was confirmed, which are benzaldehyde (4.751143%); 3(2H)-thiophenone, dihydro-2-methyl- (6.072668%), propanoic acid, 2-methyl- (13.97511%), hexadecanoic acid (3.60544%), octadecanoic acid (1.638403%), oleic acid (9.677555%), and 9,12-octadecadienoic acid (Z,Z)- (11.09688%). The presence of some valuable compounds in Bacillus acidiproducens NiuFun also was confirmed, which are benzaldehyde (4.751143%); 3(2H)-thiophenone, dihydro-2-methyl- (6.072668%), propanoic acid, 2-methyl- (13.97511%), hexadecanoic acid (3.60544%), octadecanoic acid (1.638403%), oleic acid (9.677555%), and 9,12-octadecadienoic acid (Z,Z)- (11.09688%) and the presence of these chemical constituents indicates a diverse metabolic capacity of this bacterium. Finally, the specific VOCs with high concentrations produced by Bacillus safensis AS-08 were as 2,3-butanedione (21.42702%), 3-pentanol, 2-methyl- (36.53327%), benzaldehyde (2.170694%), 2,3-butanediol (4.672893%), propanoic acid, 2-methyl- (3.156132%), oleic acid (4.585822%), and 9,12-octadecadienoic acid (Z,Z)- (3.576243).
Additionally, the GC–MS analysis showed that all five bacterial isolates contained various fatty acids along with other volatile organic compounds. It is well-known that fatty acids and their derivatives can exhibit powerful antibacterial and antifungal activities [52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68]. Indeed, fatty acids are known for their biodegradability, low toxicity, and resistance to extremes in pH, salinity, and temperature, which make them environmentally friendly compounds. These properties have led to their acceptance and use as food additives in various applications. Antifungal fatty acids, particularly those found in natural sources, may have a lower likelihood of inducing resistance in pathogenic fungi compared to some synthetic antifungal drugs [84]. The identification of volatile compounds, including esters, alkaloids, ethers, and phenolics, in five different Bacillus species, is noteworthy and suggests a diverse array of secondary metabolites produced by these bacteria. The presence of volatile organic compounds (VOCs) as major constituents of a bacterial strain with properties against phytopathogens is significant and highlights the potential of these bacteria for various agricultural and environmental applications. The presence of common volatile compounds among different Bacillus species suggests the existence of conserved metabolic pathways or biochemical processes within the genus Bacillus. These shared compounds may be indicative of fundamental metabolic activities that are essential for the survival and growth of these bacteria. Our findings support prior research on the chemical composition of bacterial strains using GC–MS and show that Bacillus spp. share similar volatile chemicals [30,85,86,87,88].
The antibacterial characteristic of certain microorganisms, including Bacillus strains, plays a crucial role in various therapeutic activities. The perpendicular streak method is a common laboratory technique used to assess the antibacterial properties of bacterial isolates against selected human bacterial pathogens. In the present study, this method was used to analyze the antibacterial potency of the bacterial isolates labeled as BSS25, BSS21, BSS16, BSS13 and BSS12 against the selected human bacterial pathogens. It can be explained with the help of bioactive compounds produced by bacteria, which can have various effects on other organisms, including antimicrobial. The perpendicular streak method is recognized as a first-pass qualitative screening method for the detection of microbial activity, particularly when assessing the antibacterial properties of bacterial isolates. The research demonstrated strong antagonistic action against human pathogens such as Enterococcus hirae and Staphylococcus epidermidis by all bacterial isolates. It is obvious that the synergistic contribution of antibacterial potency refers to the combined effect of two or more antimicrobial agents (such as antibiotics or other antimicrobial com-pounds) that work together to produce a stronger inhibitory or bactericidal effect against bacteria than the individual agents would achieve on their own. According GC–MS analysis, many compounds out 69 were identified with antibacterial potency (Table 9) and their synergetic contribution explains how all bacterial strains have shown strong antagonistic action against Enterococcus hirae and Staphylococcus epidermidis. Moreover, the current research provided indicates that bacterial isolates BSS25, BSS21, and BSS16 exhibit strong antagonistic activity against Candida albicans while BSS25, BSS21, and BSS13 suppress the growth or activity of Candida krusei. Additionally, only BSS25 and BSS21 were found to show being studied exhibited strong inhibitory effects against Klebsiella aerogenes. This may be due to the high concentrations of acetoin in BSS25 and BSS21 and butanoic acid, 2-methyl- in BSS16 and BSS13. For sure acetoin and butanoic acid, 2-methyl- enhances the antibacterial potency of bacterial strains along with other organic volatile compounds, peptides, etc.
These are positive results, suggesting that these bacterial isolates may have potential applications in controlling or preventing infections caused by these human pathogens. Our results showed that Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08 are effective at inhibiting the growth of some multidrug-resistant bacterial strains and this is similar to findings of our previous study [30]. Previuos researches found that the inhibitory impact of Bacillus species on other microorganisms, including pathogens, can indeed be attributed to various factors, including the pH of the growing medium and the generation of volatile chemicals. Bacillus species are known to produce a variety of polypeptide antibiotic substances, including bacitracin, polymyxin, gramicidin S, and tyrothricin. These antibiotic substances have shown effectiveness against a wide range of bacteria, including both Gram-positive and Gram-negative bacteria [88].
Bacterial extract investigation is a versatile and interdisciplinary field with far-reaching implications for science, medicine, agriculture, and industry. It involves the discovery and characterization of compounds and biological activities that can address various challenges and opportunities in these domains. Bacterial extracts are screened to discover novel antibiotic compounds, and antimicrobial peptides (AMPs) with potential therapeutic applications, as bacteria produce a wide variety of substances. Moreover, bacterial extracts can be a source of potential drug candidates for the treatment of various diseases, including infectious diseases and cancer. According to recent studies, bacterial extracts are also used in bioassays to evaluate the biological activity of compounds, including screening for enzyme inhibitors or activators while more of the studies proved the value of bacterial extracts' use in agriculture to manage plant diseases and pests. For example, Bacillus species are known for their significant roles in agriculture and biotechnology, primarily due to their ability to produce various bioactive compounds that can benefit plant health and promote agricultural sustainability [89,90]. Moreover, bacterial extracts are analyzed to identify probiotic strains with potential benefits for gut health.
Molecular investigations can provide valuable insights into the taxonomy and genetic relatedness of different bacterial isolates and our results revealed the taxonomy of five different isolated species of Bacillus, which are Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08. It was determined that the five most viable candidates of bacterial isolates BSS25, BSS21, BSS16, BSS13 and BSS12 belong to Bacillus thuringiensis F3 (99%), Bacillus toyonensis FORT 102 (99%), Bacillus acidiproducens NiuFun (99%), Bacillus cereus WAB2133 (99%), and Bacillus safensis AS-08 (99%), respectively, based on top hit sequence similarity results and phylogenetic analysis.
The identification of the five separate bacterial strains (Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08) and their antibacterial activity can significantly facilitate microbial screening and the isolation of active metabolites, especially against multidrug-resistant strains. Consequently, knowing the specific strains that exhibit antibacterial activity allows for targeted screening of these strains against multidrug-resistant bacterial strains. This targeted approach saves time and resources compared to screening a wide range of microorganisms. As the antibacterial activity is confirmed, the isolation and purification of bioactive metabolites from these strains can be prioritized. This is crucial for identifying the specific compounds responsible for the antibacterial effects. Moreover, the isolated bioactive compounds may have potential as antibiotic adjuvants, especially against multidrug-resistant strains. This can be a valuable contribution to the fight against antibiotic resistance. Additionally, these compounds may serve as lead compounds for drug discovery efforts, where further modifications or structural optimization can be performed to enhance their efficacy and reduce potential side effects. Overall, the identification of metabolites from bacterial strains and evaluation of their antibacterial and antibiotic activity provides a strong foundation for focused research and applications in various fields.
4. Materials and Methods
4.1. Isolation of Potential Strains of the Genus Bacillus spp.
Five Bacillus strains (BSS25, BSS21, BSS16, BSS13 and BSS12) have been isolated from different vegetable samples such as tomato, potato, and carrot. A vegetable sample with a mass equal to 15 g was homogenized in a solvent with a volume of 100 ml of NaCl by shaking at 150 rpm for 20 min. After, the sample was steadily diluted and incubated for 10 min at 90 ◦C. After the incubation the sample has been cooled to room temperature. The sample with a volume of 0.1 mL was loaded onto nutrient agar/meat peptone agar (NA/MPA) plates, which serves as a fertile medium for the growth of undemanding microorganisms. NA/MPA plates were consisted of bacteriological agar (BA, 15 g/L), gelatin peptone (GP, 5 g/L), and meat extract (ME, 3 g/L). The plates were incubated at 37°C for 48 hours. The isolated pure strains were refrigerated at −20 ◦C in nutrient broth (NB) media supplemented with 20% (v/v) glycerin. Pure strains were stored at −20 ◦C in nutrient broth (NB), which contains 20% (v/v) glycerin as a supplement. Then, morphological identification was performed on the newly created culture. Slightly raised, flat, white, and cream-colored colonies were chosen for further study. Strain isolates were helpful in further studies, particularly in the preparation of ethyl acetate extract for subjecting GC–MS analysis.
4.2. Antagonistic Activity Study
On Mueller-Hinton agar (MHA) plates, a preliminary antibacterial investigation of the isolates was carried out using the perpendicular streak method against potent human pathogens. Overall, n=15 bacterial pathogens were used in antagonistic activity study: Salmonella enterica ATCC 35664, Klebsiella aerogenes ATCC 13048, Serratia marcescens ATCC 13880, Klebsiella pneumoniae ATCC 13883, Streptococcus group B, Escherichia coli ATCC 25922, Candida krusei ATCC 14243, Shigella sonnei ATCC 25931, Streptococcus mutans ATCC 25175, Enterococcus hirae ATCC 10541, Proteus Vulgaris ATCC 6380, Staphylococcus epidermidis ATCC 12228, Pseudomonas aeruginosa ATCC 9027, Staphylococcus aureus ATCC 29213 and Candida albicans ATCC 2091. On the basis of the perpendicular streak method, an exponential culture of the studied pathogens was streaked on the surface of an agar medium and incubated at 30 ± 4 ◦C for 24 h [91]. Then, an exponential culture of the test strain was inoculated perpendicularly from the edge of the cup to the stroke of the grown culture of the antagonist with a stroke by slightly touching the stroke of the antagonist strain. The plate was once more incubated in a setting that encouraged test culture growth. The samples were then processed using the first technique.
The second method an agar well-diffusion method with a few minor modifications was used to measure antimicrobial activity [92]. On the plate, bacterial suspensions were applied with turbidity that was calibrated to the McFarland 0.5 standard (about 108 colony forming units, or CFU per milliliter). Using the back end of a sterile 1-mL pipette tip, a 7-mm diameter well was punched aseptically onto Mueller-Hinton agar (Oxoid, Basingstoke, UK). The positive control was streptomycin (1 g/mL). Each well received 100 L of test agent in total. The diameter of the clear zone was measured during an incubation period of 16 to 24 hours at 37 °C.
4.3. Antibiotic Susceptibility of the Bacillus Isolates
Using the disk diffusion method, the antibiotic susceptibility test of all five Bacillus strains (BSS25, BSS21, BSS16, BSS13 and BSS12) was done in accordance with the guidelines of the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2019). Using an aliquot of 1 mL for each strain and a concentration of 106 CFU/mL (0.5 McFarland, Hi-media, India), Bacillus strains were spread-plated on Mueller-Hinton (MH) agar using sterile beads. The plates were subjected to drying for an hour. Consequently, antibiotic disks were inserted into the agar plates containing an inoculated strain. After incubation period (24 h) at 37 °C, the widths of the inhibition zones surrounding the antibiotic disks were measured using an electronic digital vernier caliper micrometer measuring instrument caliber digital ruler (ZHHRHC LCD, Hardened, China). This helped to identify following parameters such as the strain’s antibiotic susceptibility (S), intermediate resistance (I), or resistance (R) according to the CLSI guidelines (2012) [93,94]. Antibiotic disks (n=18) contained a sample each of ampicillin (AMP, 10), amoxycillin (AMOX, 30), amoxycillin-clavulanic acid (AMC, 30), azithromycin (AZM, 15), bacitromycin (B, 10), carbenicillin (CAR, 100), cefepime (FEP, 30), cefepime/clavulanic acid FEC-40, cephalatin (KF, 30), cefotaxime (CTX, 30), cloxacillin (CX, 5), erythromycin (ERO, 15), gentamicin (CN, 120), polymyxin (PB, 300), penicillin G (PEN, 10), streptomycin (STR, 10), tobramycin (TOB, 10), tetracycline (TET, 30).
4.4. Gas Chromatography–Mass Spectrum Analysis
The volatile substances extraction for each bacterial isolate (Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, Bacillus safensis AS-08) was carried out separately two times from the culture broth with 25 mL ethyl acetate (Sigma-Aldrich, Germany) for 20 min and two extracts were combined. After that, 1.5 mL of the extract was transferred into plastic vials with a capacity of 2 mL, which were then set on the autosampler tray for GC-MS analysis. Bacterial secondary metabolites were examined through a GC-MS analysis on a Thermo Scientific GC Focus Series DSQ. As the carrier gas, helium gas was used at a constant flow rate of 1 mL per minute and the injection volume of sample was equal to 1 mL. The injector and hot oven were kept at 250 ◦C and 110 ◦C, respectively, with the temperature increasing by 10 ◦C per minute up to 200 ◦C, 5 ◦C per minute up to 280 ◦C, and shutting down after 9 min at a temperature of 280 ◦C [30]. The retention durations of several chemical peaks that were eluted from the GC column were recorded. After matching the data with the mass spectra of the compounds, the database was searched for compounds with comparable molecular masses and retention times. The current investigation discovered a parallel pattern in the bioactivities of previously studied natural compounds.
4.5. Molecular Characterization of the Bacterial Isolates
Isolated bacterial strains were molecularly characterized using universal bacterial primers and 16S rRNA conserved gene sequences. Using the conventional PCR method, the targeted gene sequence was amplified. The final result was then processed via 1% gel electrophoresis to determine the size of the amplified fragments. The amplified materials were delivered for sequencing along with the pertinent sequencing fragments. The nucleotide sequences were phylogenetically analyzed using MEGA software (MEGA-11). The bacterial isolates were subsequently validated and categorized at the species level using GenBank NCBI's BLAST search (National Center for Biotechnology Information). The accession numbers (MF135173, MG561363, MF446886, MH169322, and JX849661) correspond to the 16S rRNA gene sequences of the probiotic strains and were used to retrieve and reference the sequences in the GenBank database. (www.ncbi.nlm.nih.gov/projects/genome/clone/, accessed on 9 July 2023).
4.6. Statistical Analysis
The XLSAT software version 2016.02.27444 was used to perform the one-factor analysis of variance at the significance level (α = 0.05). The Newman-Keuls test was used to rank the means when there were substantial differences between the parameters under study.
5. Conclusions
In a current study, vegetable bacterial isolates obtained from five different species of Bacillus demonstrated the ability to inhibit the growth of multidrug-resistant bacterial strains. As a result of a screening process, three potent isolates named BSS25, BSS21, BSS16, BSS13 and BSS12 were identified. GC–MS was used to identify and quantify the chemical compounds present in bacterial species, despite the fact that they are all members of the same Bacillus subspecies. The observation was that volatile organic compounds (VOCs) differ among members of the same Bacillus subspecies despite their taxonomic similarity highlighting the chemical diversity that can exist within closely related bacterial strains. This study confirmed a variety of volatile inhibitory substances, including esters, phenolics, and ethers, which are believed to play a role in antimicrobial activity. The volatile compounds differ in chemical composition among the tested samples suggesting that these variations may have an impact on antimicrobial activity and antibiotic potency. Strains of the Bacillus subtilis group are known to have the capability of producing a wide range of secondary metabolites that contribute to their antimicrobial characteristics and this was also confirmed by our findings. In addition to volatile organic compounds, strains within the Bacillus subtilis group are known to produce a variety of other bioactive compounds, including bacteriocins, polyketides, peptides, and more. Hence, it can be concluded that the discovered organic volatile substances i.e. acetoin and butanoic acid, 2-methyl- enhance the antimicrobial properties of Bacillus spp. together with the above substances. The remarkable metabolic capacity and adaptive biochemistry of Bacillus species, i.e., Bacillus thuringiensis F3 (BSS25), Bacillus toyonensis FORT 102 (BSS21), Bacillus acidiproducens NiuFun (BSS16), Bacillus cereus WAB2133 (BSS13), and Bacillus safensis AS-08 (BSS12) make these strains valuable for various commercial and biotechnological applications, as they have the potential to generate a wide range of bioactive chemical substances. Bacterial extracts, which contain bioactive chemicals produced by these Bacillus strains, have the potential to be used as antimicrobial agents. The anticipation of conducting a thorough investigation, similar to the one described, holds great promise in uncovering new microbiological possibilities and discovering previously unknown substances or metabolites with strong antibacterial potential. Such research endeavors are essential for addressing the burden and danger posed by bacterial strains that have developed resistance to multiple drugs.
Author Contributions
Conceptualization: M.K., A.A., K.W. and Z.S.; methodology: M.K., Z.S., A.A., K.M., G.U., K.W. and A.T.; software: M.K., Z.S. and A.A.; validation: T.A., Z.S. and M.A.; formal analysis: R.B., M.A. and G.K.; investigation: M.K. and Z.S.; resources: M.K. and A.A.; data curation: M.K. and A.A.; writing—original draft: M.K. and Z.S.; writing—review and editing: M.K. and Z.S.; visualization: A.K., Z.A., Y.K. and D.B.; supervision: G.U.; project administration: G.U.; funding acquisition: G.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds are not available from the authors.
References
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal Transfer of Antibiotic Resistance Genes in Clinical Environments. Canadian Journal of Microbiology 2019, 65, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Multidrug resistance in bacteria. Annual review of biochemistry 2009, 78, 119–146. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Chitinase production by Bacillus thuringiensis and Bacillus licheniformis: their potential in antifungal biocontrol. Journal of microbiology 2012, 50, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Caulier S, Nannan C, Gillis A, Licciardi F, Bragard C, Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Frontiers in microbiology 2019, 10, 302. [Google Scholar] [CrossRef]
- Lawton, E.M.; Ross, R.P.; Hill, C.; Cotter, P.D. Two-Peptide Lantibiotics: A Medical Perspective. Mini-Rev. Med. Chem. 2007, 7, 1236–1247. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Nishie, M.; Nagao, J.I.; Sonomoto, K. Antibacterial Peptides “Bacteriocins”: An Overview of Their Diverse Characteristics and Applications. Biocontrol Sci. 2012, 17, 1–16. [Google Scholar] [CrossRef]
- Yang, S.C.; Lin, C.H.; Sung, C.T.; Fang, J.Y. Antibacterial Activities of Bacteriocins: Application in Foods and Pharmaceuticals. Front. Microbiol. 2014, 5, 241. [Google Scholar] [CrossRef]
- Magashi, A.M.; Bukar, A.; Omola, E.M.; Halima, B.A.; Hadiza, M.S. Bacteriocin and its application—A review. Int. J. Adv. Acad. Res. Sci. Technol. Eng. 2019, 5, 242–256, https://www.ijaar.org/articles/Volume5-Number11/Sciences-Technology-Engineering/ijaar-ste-v5n11-nov19-p29.pdf. [Google Scholar]
- Wiegers, C.; van de Burgwal, L.H.M.; Larsen, O.F.A. Probiotics for the Management of Infectious Diseases: Reviewing the State of the Art. Front Microbiol. 2022, 13, 877142. [Google Scholar] [CrossRef]
- Ramirez-Olea, H.; Reyes-Ballesteros, B.; Chavez-Santoscoy, R.A. Potential Application of the Probiotic Bacillus Licheniformis as an Adjuvant in the Treatment of Diseases in Humans and Animals: A Systematic Review. Front. Microbiol. 2022, 13, 993451. [Google Scholar] [CrossRef] [PubMed]
- Hallaj-Nezhadi, S.; Hamdipour, R.; Shahrvirani, M.; Zare tin, R.; Chapeland-leclerc, F.; Ruprich-Robert, G.; Esnaashari, S.; Elyasi Far, B.; Dilmaghani, A. Antimicrobial Activity of Bacillus sp. Isolated Strains of Wild Honey. BMC Complement. Med. Ther. 2022, 22, 78. [Google Scholar] [CrossRef] [PubMed]
- Joerger, R.D. Alternatives to Antibiotics: Bacteriocins, Antimicrobial Peptides and Bacteriophages. Poult. Sci. 2003, 82, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Jach, M.E.; Serefko, A.; Szopa, A.; Sajnaga, E.; Golczyk, H.; Santos, L.S.; Borowicz-Reutt, K.; Sieniawska, E. The Role of Probiotics and Their Metabolites in the Treatment of Depression. Molecules 2023, 28, 3213. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Shi, X.; Wang, B.; Li, M.; Wang, Q.; Zhang, S. Antifungal Effect of Volatile Organic Compounds from Bacillus velezensis CT32 against Verticillium dahlia and Fusarium oxysporum. Processes 2020, 8, 1674. [Google Scholar] [CrossRef]
- Reddy, K.V.R.; Yedery, R.D.; Aranha, C. Antimicrobial Peptides: Premises and Promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef]
- Emmert, E.A.B.; Handelsman, J. Biocontrol of plant disease: A (Gram-) positive perspective. FEMS Microbiol. Lett. 1999, 171, 1–9. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Fira, D.; DimkiC, I.; Beric, T.; Lozo, J.; Stankovic, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Effmert, U.; Kalderás, J.; Warnke, R.; Piechulla, B. Volatile Mediated Interactions Between Bacteria and Fungi in the Soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef]
- Farag, M.A.; Zhang, H.; Ryu, C.-M. Dynamic Chemical Communication between Plants and Bacteria through Airborne Signals: Induced Resistance by Bacterial Volatiles. J. Chem. Ecol. 2013, 39, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Fincheira, P.; Quiroz, A. Microbial volatiles as plant growth inducers. Microbiol. Res. 2018, 208, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Kanchiswamy, C.N.; Emalnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Farag, M.A.; Park, H.B.; Kloepper, J.W.; Lee, S.H.; Ryu, C. Induced Resistance by a Long-Chain Bacterial Volatile: Elicitation of Plant Systemic Defense by a C13 Volatile Produced by Paenibacillus polymyxa. PLoS ONE 2012, 7, e48744. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Kloepper, J.W.; Pare, P.W. Bacterial Volatiles Induce Systemic Resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiao, X.; Wang, J.; Wu, L.; Zheng, Z.; Yu, Z. Antagonistic effects of volatiles generated by Bacillus subtilis on spore germination and hyphal growth of the plant pathogen Botrytis cinerea. Biotechnol. Lett. 2008, 30, 919–923. [Google Scholar] [CrossRef]
- Zheng, M.; Shi, J.Y.; Shi, J.; Wang, Q.G.; Li, Y.H. Antimicrobial effects of volatiles produced by two antagonistic Bacillus strains on the anthracnose pathogen in postharvest mangos. Biol. Control 2013, 65, 200–206. [Google Scholar] [CrossRef]
- Arrebola, E.; Sivakumar, D.; Korsten, L. Effect of volatile compounds produced by Bacillus strains on postharvest decay in citrus. Biol. Control 2010, 53, 122–128. [Google Scholar] [CrossRef]
- Koilybayeva, M.; Shynykul, Z.; Ustenova, G.; Abzaliyeva, S.; Alimzhanova, M.; Amirkhanova, A.; Turgumbayeva, A.; Mustafina, K.; Yeleken, G.; Raganina, K.; et al. Molecular Characterization of Some Bacillus Species from Vegetables and Evaluation of Their Antimicrobial and Antibiotic Potency. Molecules 2023, 28, 3210. [Google Scholar] [CrossRef]
- Haque, M.A.; Wang, F.; Chen, Y.; Hossen, F.; Islam, M.A.; Hossain, M.A.; Siddique, N.; He, C.; Ahmed, F. Bacillus spp. Contamination: A Novel Risk Originated From Animal Feed to Human Food Chains in South-Eastern Bangladesh. Front Microbiol. 2022, 12, 783103. [Google Scholar] [CrossRef] [PubMed]
- Aruna, K.; Shah, J.; Birmole, R. Production and partial characterization of alkaline protease from Bacillus tequilensis strains CSGAB 0139 isolated from spoilt cottage cheese. Int J Appl Biol Pharm. 2014, 5, 201–21. [Google Scholar]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of Microbial Enzymes in Food Industry. Food technology and biotechnology 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- van der Maarel, M.J.; van der Veen, B.; Uitdehaag, J.C.; Leemhuis, H.; Dijkhuizen, L. Properties and applications of starch-converting enzymes of the alpha-amylase family. Journal of biotechnology 2002, 94, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Soccol, C.R.; Rojan, P.J.; Patel, A.K.; Woiciechowski, A.L.; Vandenberghe, L.P.S.; Pandey, A. Glucoamylase. In Enzyme Technology; Pandey, A., Webb, C., Soccol, C.R., Larroche, C., Eds.; Asiatech Publishers Inc.: New Delhi, India, 2005; pp. 221–38. [Google Scholar]
- Gurung, N.; Ray, S.; Bose, S.; Rai, V. A broader view: Microbial enzymes and their relevance in industries, medicine, and beyond. BioMed Res Int. 2013, 2013, 329121. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Nigam, P.; Soccol, C.R.; Soccol, V.T.; Singh, D.; Mohan, R. Advances in microbial amylases. Biotechnol Appl Biochem. 2000, 31 Pt 2, 135–52. [Google Scholar] [CrossRef]
- de Souza, P.M.; de Oliveira Magalhães, P. Application of microbial α-amylase in industry - A review. Brazilian journal of microbiology: [publication of the Brazilian Society for Microbiology] 2010, 41, 850–861. [Google Scholar] [CrossRef]
- James, J.; Simpson, B.K.; Marshall, M.R. Application of enzymes in food processing. Crit Rev Food Sci Nutr. 1996, 36, 437–63. [Google Scholar] [CrossRef]
- Pereira, I.; Severino, P.; Santos, A.C.; Silva, A.M.; Souto, E.B. Linalool bioactive properties and potential applicability in drug delivery systems. Colloids Surf B Biointerfaces 2018, 171, 566–578. [Google Scholar] [CrossRef]
- Zhang, S.B.; Qin, Y.L.; Li, S.F.; Lv, Y.Y.; Zhai, H.C.; Hu, Y.S.; Cai, J.P. Antifungal mechanism of 1-nonanol against Aspergillus flavus growth revealed by metabolomic analyses. Applied microbiology and biotechnology 2021, 105, 7871–7888. [Google Scholar] [CrossRef]
- Togashi, N.; Shiraishi, A.; Nishizaka, M.; Matsuoka, K.; Endo, K.; Hamashima, H.; Inoue, Y. Antibacterial activity of long-chain fatty alcohols against Staphylococcus aureus. Molecules (Basel, Switzerland) 2007, 12, 139–148. [Google Scholar] [CrossRef]
- Eom, M.R.; Weon, J.B.; Jung, Y.S.; Ryu, G.H.; Yang, W.S.; Ma, C.J. Neuroprotective compounds from Reynoutria sachalinensis. Archives of pharmacal research 2017, 40, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Izumo, T.; Ichiki, C.; Saitou, K.; Otsuka, M.; Ohmori, S.; Kamei, C. Effects of ethanol, acetoin and 2,3-butanediol on EEG power spectra in conscious rats. Biological & pharmaceutical bulletin 1998, 21, 29–33. [Google Scholar] [CrossRef]
- Kim, H.S.; Arellano, K.; Park, H.; Todorov, S.D.; Kim, B.; Kang, H.; Park, Y.J.; Suh, D.H.; Jung, E.S.; Ji, Y.; Holzapfel, W.H. Assessment of the safety and anti-inflammatory effects of three Bacillus strains in the respiratory tract. Environmental microbiology 2021, 23, 3077–3098. [Google Scholar] [CrossRef] [PubMed]
- Dhakshinamoorthy, D.; Sundaresan, S.; Iyadurai, A.; Subramanian, K.S.; Janavi, G.J.; Paliyath, G.; Subramanian, J. Hexanal Vapor Induced Resistance against Major Postharvest Pathogens of Banana (Musa acuminata L.). The plant pathology journal 2020, 36, 133–147. [Google Scholar] [CrossRef]
- Zhang, J.H.; Sun, H.L.; Chen, S.Y.; Zeng, L.; Wang, T.T. Anti-fungal activity, mechanism studies on α-Phellandrene and Nonanal against Penicillium cyclopium. Botanical studies 2017, 58, 13. [Google Scholar] [CrossRef]
- Tao, N.; Jia, L.; Zhou, H. Anti-fungal activity of Citrus reticulata Blanco essential oil against Penicillium italicum and Penicillium digitatum. Food chemistry 2014, 153, 265–271 [. [Google Scholar] [CrossRef]
- Chan, P.C. NTP toxicity studies of toxicity studies of 2,4-decadienal (CAS No. 25152-84-5) administered by gavage to F344/N Rats and B6C3F1 mice. Toxicity report series 2011, 76, 1–94 [https://pubmedncbinlmnihgov/21445102/]. [Google Scholar]
- Armand, J. , De Forni, M., Recondo, G., Cals, L., Cvitkovic, E., & Munck, J. Flavonoids: a new class of anticancer agents? Preclinical and clinical data of flavone acetic acid. Progress in clinical and biological research 1988, 280, 235–241 [https://pubmedncbinlmnihgov/3051027/]75. [Google Scholar]
- Andersen, A. Final report on the safety assessment of benzaldehyde. International journal of toxicology 2006, 25 Suppl 1, 11–27. [Google Scholar] [CrossRef]
- Tangavelou, A.C.; Viswanathan, M.B.; Balakrishna, K.; Patra, A. Phytochemical Analysis in the Leaves of Chamaecrista nigricans (Leguminosae). Pharmaceutica Analytica Acta 2018, 09, 3 [. [Google Scholar] [CrossRef]
- Altinoz, M.A.; Ozpinar, A.; Seyfried, T.N. Caprylic (Octanoic) Acid as a Potential Fatty Acid Chemotherapeutic for Glioblastoma. Prostaglandins, leukotrienes, and essential fatty acids 2020, 159, 102142 [. [Google Scholar] [CrossRef]
- Kim, S.A.; Rhee, M.S. Antibacterial activity of caprylic acid for potential application as an active antiseptic ingredient in consumer antiseptics. International journal of antimicrobial agents 2016, 48, 765–767 [. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dolan, H.L.; Ding, Q.; Wang, S.; Tikekar, R.V. Antimicrobial action of octanoic acid against Escherichia coli O157:H7 during washing of baby spinach and grape tomatoes. Food research international (Ottawa, Ont.) 2019, 125, 108523 [. [Google Scholar] [CrossRef]
- Johnson, W. Jr.; Heldreth, B.; Bergfeld, W.F.; Belsito, D.V.; Klaassen, C.D.; Hill, R.; Liebler, D.; Marks, J.G. Jr.; Shank, R.C.; Slaga, T.J.; Snyder, P.W.; Andersen, F.A. Final report of the Cosmetic Ingredient Review Expert Panel on the safety assessment of pelargonic acid (nonanoic acid) and nonanoate esters. International Journal of Toxicology 2011, 30 (6 Suppl), 228S–269S. [Google Scholar] [CrossRef]
- Jang, Y.W.; Jung, J.Y.; Lee, I.K.; Kang, S.Y.; Yun, B.S. Nonanoic Acid, an Antifungal Compound from Hibiscus syriacus Ggoma. Mycobiology 2012, 40, 145–146. [Google Scholar] [CrossRef] [PubMed]
- Manrique Vergara, D.; González Sánchez, M.E. Ácidos grasos de cadena corta (ácido butírico) y patologías intestinales [Short chain fatty acids (butyric acid) and intestinal diseases]. Nutricion hospitalaria 2017, 34 (Suppl. 4), 58–61. [Google Scholar] [CrossRef] [PubMed]
- To, N.B.; Nguyen, Y.T.; Moon, J.Y.; Ediriweera, M.K.; Cho, S.K. Pentadecanoic Acid, an Odd-Chain Fatty Acid, Suppresses the Stemness of MCF-7/SC Human Breast Cancer Stem-Like Cells through JAK2/STAT3 Signaling. Nutrients 2020, 12, 1663. [Google Scholar] [CrossRef]
- Galdiero, E.; Ricciardelli, A.; D'Angelo, C.; de Alteriis, E.; Maione, A.; Albarano, L.; Casillo, A.; Corsaro, M.M.; Tutino, M.L.; Parrilli, E. Pentadecanoic acid against Candida albicans-Klebsiella pneumoniae biofilm: towards the development of an anti-biofilm coating to prevent polymicrobial infections. Research in microbiology 2021, 172, 103880. [Google Scholar] [CrossRef]
- Sales-Campos, H.; Souza, P.R.; Peghini, B.C.; da Silva, J.S.; Cardoso, C.R. An overview of the modulatory effects of oleic acid in health and disease. Mini reviews in medicinal chemistry 2013, 13, 201–210 https://pubmedncbinlmnihgov/23278117/. [Google Scholar]
- Aparna, V.; Dileep, K.V.; Mandal, P.K.; Karthe, P.; Sadasivan, C.; Haridas, M. Anti-inflammatory property of n-hexadecanoic acid: structural evidence and kinetic assessment. Chemical biology & drug design 2012, 80, 434–439. [Google Scholar] [CrossRef]
- Shaaban, M.T.; Ghaly, M.F.; Fahmi, S.M. Antibacterial activities of hexadecanoic acid methyl ester and green-synthesized silver nanoparticles against multidrug-resistant bacteria. Journal of basic microbiology 2021, 61, 557–568 [. [Google Scholar] [CrossRef] [PubMed]
- Habib, N.A.; Wood, C.B.; Apostolov, K.; Barker, W.; Hershman, M.J.; Aslam, M.; Heinemann, D.; Fermor, B.; Williamson, R.C.; Jenkins, W.E. Stearic acid and carcinogenesis. British journal of cancer 1987, 56, 455–458. [Google Scholar] [CrossRef] [PubMed]
- den Hartigh, L.J. Conjugated Linoleic Acid Effects on Cancer, Obesity, and Atherosclerosis: A Review of Pre-Clinical and Human Trials with Current Perspectives. Nutrients 2019, 11, 370 [. [Google Scholar] [CrossRef]
- Shen, T.; Chen, L.; Liu, Y.; Shi, S.; Liu, Z.; Cai, K.; Liao, C.; Wang, C. Decanoic acid modification enhances the antibacterial activity of PMAP-23RI-Dec. European journal of pharmaceutical sciences: official journal of the European Federation for Pharmaceutical Sciences 2021, 157, 105609 [. [Google Scholar] [CrossRef]
- Huang, W.C.; Tsai, T.H.; Chuang, L.T.; Li, Y.Y.; Zouboulis, C.C.; Tsai, P.J. Anti-bacterial and anti-inflammatory properties of capric acid against Propionibacterium acnes: a comparative study with lauric acid. Journal of dermatological science 2014, 73, 232–240 [. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Beiram, R.; Azimullah, S.; Mf, N.M.; Ojha, S.K.; Adem, A.; Jalal, F.Y. Valeric Acid Protects Dopaminergic Neurons by Suppressing Oxidative Stress, Neuroinflammation and Modulating Autophagy Pathways. International journal of molecular sciences 2020, 21, 7670 [. [Google Scholar] [CrossRef]
- Felczykowska, A.; Pastuszak-Skrzypczak, A.; Pawlik, A.; Bogucka, K.; Herman-Antosiewicz, A.; Guzow-Krzemińska, B. Antibacterial and anticancer activities of acetone extracts from in vitro cultured lichen-forming fungi. BMC complementary and alternative medicine 2017, 17, 300 [. [Google Scholar] [CrossRef]
- Shen, Y.J.; Shen, Y.C.; Lee, W.S.; Yang, K.T. Methyl palmitate protects heart against ischemia/reperfusion-induced injury through G-protein coupled receptor 40-mediated activation of the PI3K/AKT pathway. European journal of pharmacology 2021, 905, 174183 [. [Google Scholar] [CrossRef]
- Shaaban, M.T.; Ghaly, M.F.; Fahmi, S.M. Antibacterial activities of hexadecanoic acid methyl ester and green-synthesized silver nanoparticles against multidrug-resistant bacteria. Journal of basic microbiology 2021, 61, 557–568 [. [Google Scholar] [CrossRef]
- El-Agamy, D.S.; Elkablawy, M.A.; Abo-Haded, H.M. Modulation of cyclophosphamide-induced cardiotoxicity by methyl palmitate. Cancer chemotherapy and pharmacology 2017, 79, 399–409 [. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Yang, Q.; Chen, D.; Yu, B.; He, J. Benzoic Acid Used as Food and Feed Additives Can Regulate Gut Functions. BioMed research international 2019, 2019, 5721585 [. [Google Scholar] [CrossRef] [PubMed]
- Assadian, O. From antiseptics to antibiotics - and back? GMS Krankenhaushygiene interdisziplinar 2007, 2, Doc26 [https://wwwncbinlmnihgov/pmc/articles/PMC2831499/]. [Google Scholar]
- Ueda, K.; Beppu, T. Antibiotics in microbial coculture. J. Antibiot. 2017, 70, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 386. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8839143/ (accessed on 7 January 2023). [CrossRef]
- Jagannath, S.; Matthew, W.; Alan, M.; Batter, H.; Spears, I.R. Gait Retraining and Incidence of Medial Tibial Stress Syndrome in Army Recruits. Off. J. Am. Coll. Sport. Med. 2014, 46, 1684–1692. [Google Scholar]
- Sharma, P.; Kumar, S.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic resistance among commercially available probiotics. Food Res. Int. J. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Citron, D.M.; Appleman, M.D. In vitro activities of daptomycin, ciprofloxacin, and other antimicrobial agents against the cells and spores of clinical isolates of Bacillus species. J. Clin. Microbiol. 2006, 44, 3814–3818. Available online: https://pubmed.ncbi.nlm.nih.gov/17021118/ (accessed on 15 September 2022). [CrossRef]
- Jensen, L.B.; Baloda, S.; Boye, M.; Aarestrup, F.M. Antimicrobial resistance among Pseudomonas spp. and the Bacillus cereus group isolated from Danish agricultural soil. Environ. Int. 2001, 26, 581–587. Available online: https://pubmed.ncbi.nlm.nih.gov/11485227/ (accessed on 27 June 2022). [CrossRef]
- Fiedler, G.; Schneider, C.; Igbinosa, E.O.; Kabisch, J.; Brinks, E.; Becker, B.; Stoll, D.A.; Cho, G.-S.; Huch, M.; Franz, C.M.A.P. Antibiotics resistance and toxin profiles of Bacillus cereus-group isolates from fresh vegetables from German retail markets. BMC Microbiol. 2019, 19, 250. [Google Scholar] [CrossRef]
- Adimpong, D.B.; Sørensen, K.I.; Thorsen, L.; Stuer-Lauridsen, B.; Abdelgadir, W.S.; Nielsen, D.S.; Derkx, P.M.; Jespersen, L. Antimicrobial susceptibility of Bacillus strains isolated from primary starters for African traditional bread production and characterization of the bacitracin operon and bacitracin biosynthesis. Appl. Environ. Microbiol. 2012, 78, 7903–7914. Available online: https://pubmed.ncbi.nlm.nih.gov/22941078/ (accessed on 28 December 2022). [CrossRef] [PubMed]
- Compaore, C.S.; Jensen, L.B.; Diawara, B.; Ouedraogo, G.A.; Jakobsen, M.; Ouoba, L.I. Resistance to antimicrobials and acid and bile tolerance of Bacillus spp isolated from Bikalga, fermented seeds of Hibiscus sabdariffa. Afr. J. Food Sci. 2013, 7, 408–414. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642 Available online: https://pubmedncbinlmnihgov/19956944/. [Google Scholar] [CrossRef]
- Dongkun, W.; Yichi, L.; Yuan, Y.; Depeng, C.h.; Jianmin, C.; Guangjun, S.; Yongfeng, A.; Zhiyan, C.; Yongfeng, Z.h.; Fenglong, W.; et al. Identification of non-volatile and volatile organic compounds produced by Bacillus siamensis LZ88 and their antifungal activity against Alternaria alternata. Biol. Control 2022, 169, 104901. [Google Scholar]
- Raj, A.; Krishna Reddy, M.M.; Chandra, R. Identification of low molecular weight aromatic compounds by gas chromatography–mass spectrometry (GC–MS) from kraft lignin degradation by three Bacillus sp. Int. Biodeterior. Biodegrad. 2007, 59, 292–296. [Google Scholar] [CrossRef]
- Surya, M.; Thiruvudainambi, S.; Ebenezar, E.G.; Vanniarajan, C.; Kumutha, K.; Vellaikumar, S. GC-MS Analysis of antimicrobial compounds produced by Bacillus spp. against rice sheath rot pathogen Sarocladium oryzae. J. Entomol. Zool. Stud. 2020, 8, 1417–1423. [Google Scholar]
- Balcázar, J.L.; Rojas-Luna, T. Inhibitory activity of probiotic Bacillus subtilis UTM 126 against Vibrio species confers protection against vibriosis in juvenile shrimp (Litopenaeus vannamei). Curr. Microbiol. 2007, 55, 409–412. Available online: https://pubmed.ncbi.nlm.nih.gov/17680306/ (accessed on 18 December 2022). [CrossRef]
- Radhakrishnan, R.; Lee, I.J. Gibberellins producing Bacillus methylotrophicus KE2 supports plant growth and enhances nutritional metabolites and food values of lettuce. Plant Physiol. Biochem. 2016, 109, 181–189. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef]
- Sabu, R.; Radhakrishnan, E.K. Bioprospecting of endophytic bacteria from zingiber officinale with antibacterial activities. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 462–467. [Google Scholar] [CrossRef]
- Yilmaz, M.; Soran, H.; Beyatli, Y. Antimicrobial Activities of Some Bacillus spp. Strains Isolated from the Soil. Microbiol. Res. 2006, 161, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Beiranvand, M.; Amin, M.; Hashemi-Shahraki, A.; Romani, B.; Yaghoubi, S.; Sadeghi, P. Antimicrobial activity of endophytic bacterial populations isolated from medical plants of Iran. Iran J. Microbiol. 2017, 9, 11–18. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5533999/ (accessed on 22 June 2022). [PubMed]
- Khan, Z.A.; Siddiqui, M.F.; Park, S. Current and Emerging Methods of Antibiotic Susceptibility Testing. Diagnostics 2019, 9, 49. Available online: https://pubmed.ncbi.nlm.nih.gov/31058811/ (accessed on 15 May 2021). [CrossRef]
Figure 1.
Antagonistic activity of the bacteria of the genus Bacillus against pathogens. The antagonistic efficacy of all five isolates was examined against pathogenic bacteria, such as Salmonella enterica ATCC 35664, Serratia marcescens ATCC 13880, Klebsiella aerogenes ATCC 13048, Candida krusei ATCC 14243, Shigella sonnei ATCC 25931, Streptococcus mutans ATCC 25175, Klebseiella pneumoniae ATCC 13883, Group B Streptococcus, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 9027, Staphylococcus aureus ATCC 29213, Candida albicans ATCC 2091, Enterococcus hirae ATCC 10541, Proteus vulgaris ATCC 6380, and Staphylococcus epidermidis ATCC 12228. (A–A2)—BSS25, (B–B2)—BSS21, (C–C2)—BSS16, (D–D2)—BSS13, and (E–E2)—BSS12.
Figure 1.
Antagonistic activity of the bacteria of the genus Bacillus against pathogens. The antagonistic efficacy of all five isolates was examined against pathogenic bacteria, such as Salmonella enterica ATCC 35664, Serratia marcescens ATCC 13880, Klebsiella aerogenes ATCC 13048, Candida krusei ATCC 14243, Shigella sonnei ATCC 25931, Streptococcus mutans ATCC 25175, Klebseiella pneumoniae ATCC 13883, Group B Streptococcus, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 9027, Staphylococcus aureus ATCC 29213, Candida albicans ATCC 2091, Enterococcus hirae ATCC 10541, Proteus vulgaris ATCC 6380, and Staphylococcus epidermidis ATCC 12228. (A–A2)—BSS25, (B–B2)—BSS21, (C–C2)—BSS16, (D–D2)—BSS13, and (E–E2)—BSS12.
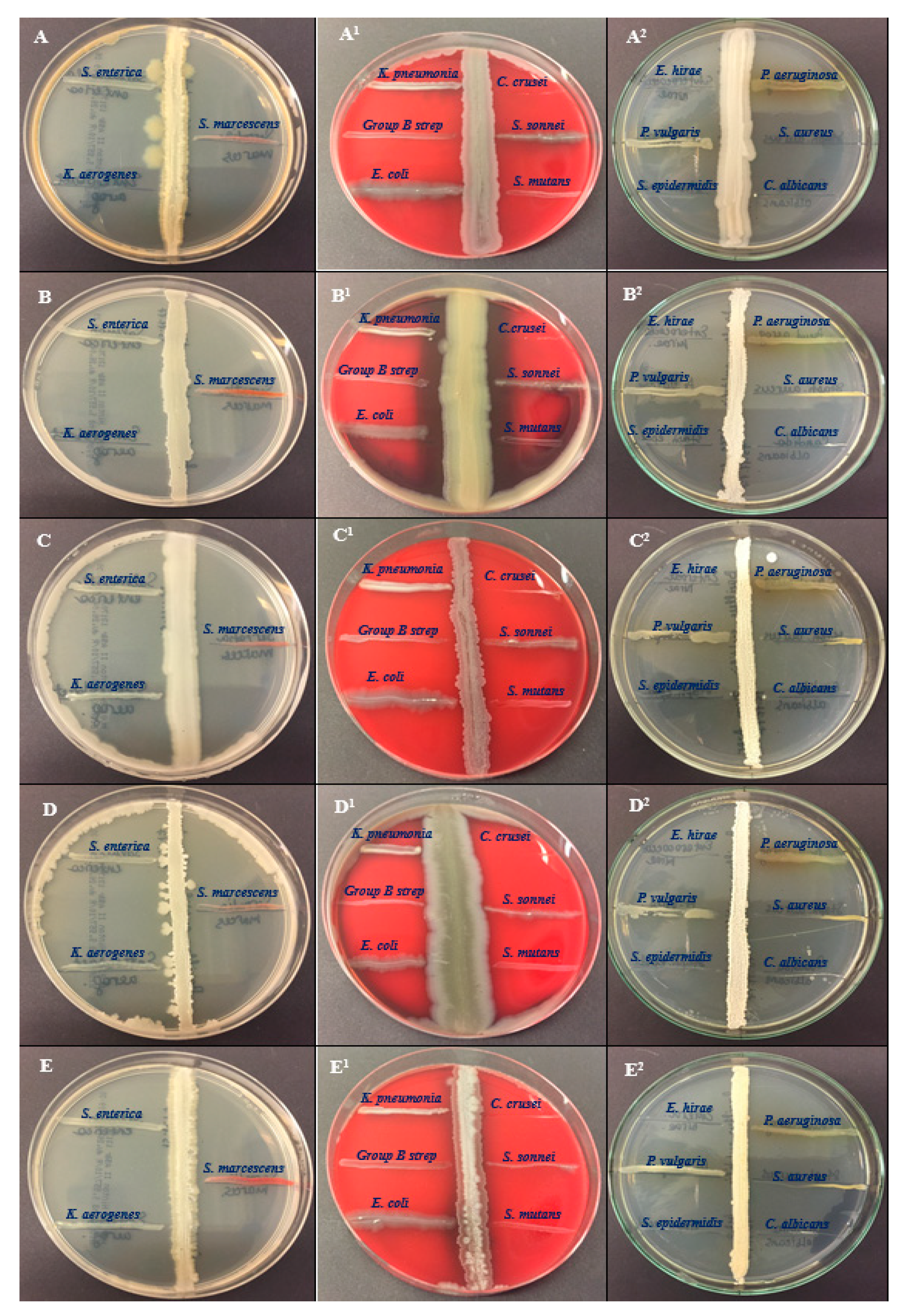
Figure 2.
Structure of common components identified from Bacillus spp. isolates.

Figure 3.
The phylogenetic tree using the Neighbour-joining model was constructed according to 16S rRNA gene sequences representing different Bacillus species, i.e., Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08, respectively. E. coli JCM 1649 (AB242910) was used as an outgroup in a phylogenetic tree.
Figure 3.
The phylogenetic tree using the Neighbour-joining model was constructed according to 16S rRNA gene sequences representing different Bacillus species, i.e., Bacillus thuringiensis F3, Bacillus toyonensis FORT 102, Bacillus acidiproducens NiuFun, Bacillus cereus WAB2133, and Bacillus safensis AS-08, respectively. E. coli JCM 1649 (AB242910) was used as an outgroup in a phylogenetic tree.
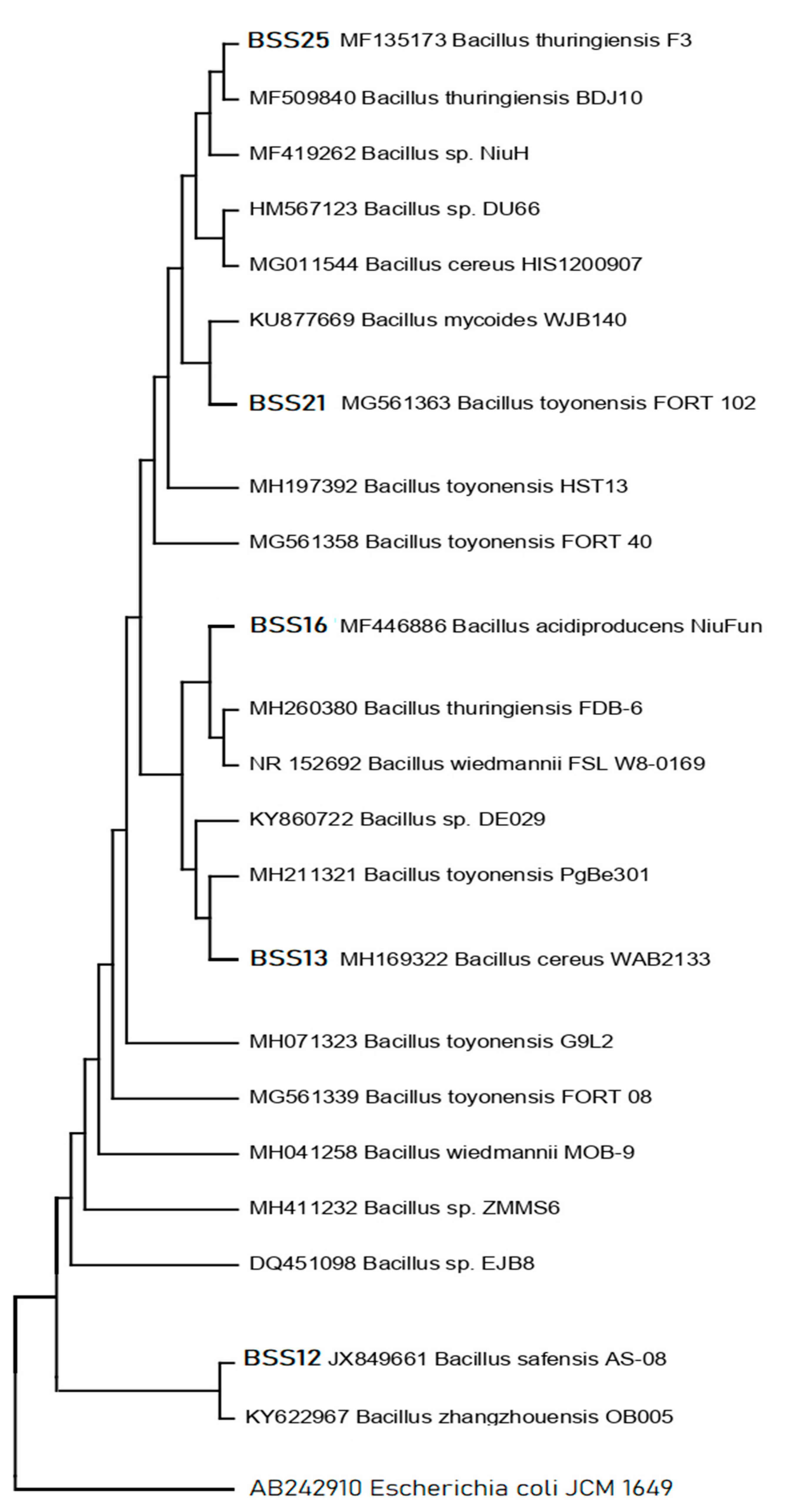
Table 1.
Colony morphology and microscopic presentation of isolated bacterial species.
| Bacterial Species | Media | Colony Color and Texture | Microscopic Presentation |
|---|---|---|---|
| Bacillus thuringiensis F3 (BSS25) | Bacillus Medium. | White, irregular, flat. | Gram-strain-positive, spore-forming, rod. |
| Bacillus toyonensis FORT 102 (BSS21) | Bacillus Medium. | White, irregular, flat. | Gram-strain-positive, spore-forming, rod. |
| Bacillus acidiproducens NiuFun (BSS16) | Bacillus Medium. | White, irregular, flat. | Gram-strain-positive, spore-forming, rod. |
| Bacillus cereus WAB2133 (BSS13) | Bacillus Medium. | White, irregular, flat. | Gram-strain-positive, spore-forming, rod. |
| Bacillus safensis AS-08 (BSS12) | Bacillus Medium. | White, irregular, flat. | Gram-strain-positive, spore-forming, rod. |
1 Tables may have a footer.
Table 2.
Antibacterial activity of the bacterial culture extracts against pathogenic strains.
| Species of Microorganism | BSS25 | BSS21 | BSS16 | BSS13 | BSS12 | Control (Streptomycin) |
|---|---|---|---|---|---|---|
| Staphylococcus aureus ATCC 29213 | 35±1,27 | 9±1,53* | 23±0,50* | 20±2,50* | 20±1,54 | 24±0,33*** |
| Staphylococcus epidermidis ATCC 12228 | 37±1,47 | 36±1,27 | 38±1,27 | 37±1,07 | 35±1,47 | 22±0.33*** |
| Streptococcus group B | 18±1,56* | 19±1,31* | 19±1,23* | 18±1,56* | 17±1,39* | 17±0.33*** |
| Streptococcus mutans ATCC 25175 | 20±1.47* | 19±1,27* | 23±1,33 | 20±1,33 | 20±1,37* | 19±0.33*** |
| Candida albicans ATCC 2091 | 36±1,43 | 38±1,21 | 38±1,27 | 35±1,26 | 34±1,22 | 31±0.33*** |
| Candida krusei ATCC 14243 | 37±1,41 | 36±1,28 | 8±1,38* | 36±1,27 | 13±1,27* | 30±0.33*** |
| Pseudomonas aeruginosa ATCC 9027 | 18±0,53* | 17±1,27* | 17±0,33* | 17±1,10* | 16±1,33* | 15±0.33*** |
| Shigella sonnei ATCC 25931 | 20±1,27* | 33±1,37* | 21±1,57* | 21±1,37* | 21±1,06* | 19±0.33*** |
| Klebsiella pneumonia ATCC 13883 | 9±1,53* | 9±1,27* | 8±1,27* | 8±1,37* | 8±1,37* | 12±0.33*** |
| Salmonella enterica ATCC 35664 | 17±1,36* | 15±1,25* | 18±1,27* | 15±1,27* | 17±1,27* | 19±0.33*** |
| Klebsiella aerogenes ATCC 13048 | 37±1,27 | 36±1,37 | 32±1,33* | 35±0,63* | 33±1,33* | 23±0.33*** |
| Enterococcus hirae ATCC 10541 | 37±1,25 | 38±1,27 | 38±1,27 | 39±1,27 | 38±1,27 | 22±0.33*** |
| Escherichia coli ATCC 25922 | 17±1,37* | 16±1,07* | 19±1,44* | 18±1,33* | 20±1,08* | 16±0.33*** |
| Serratia marcescens ATCC 13880 | 27±0,56* | 29±1,36* | 25±1,32* | 28±0,53* | 27±1,53* | 22±0.33*** |
| Proteus Vulgaris ATCC 6380 | 20±0,31* | 19±1,31* | 21±1,33* | 20±1,23* | 21±0,33* | 22±0.33*** |
* Data are represented as the means ± SE (n = 3). Values with the same superscript symbols are not statistically different. Significance level * < ***.
Table 3.
Colony morphology and microscopic presentation of isolated bacterial species.
| Antibiotic (AB, charge in μg) used | Bacillus strains | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BSS25 | BSS21 | BSS16 | BSS13 | BSS12 | ||||||
| Diameter (mm) | S/R | Diameter (mm) | S/R | Diameter (mm) | S/R | Diameter (mm) | S/R | Diameter (mm) | S/R | |
| Penicillins: | ||||||||||
| Penicillin G (PEN, 10) | 37± 0,19a | S | 20 ± 0,35ab | S | 34 ± 0,48abc | S | 22 ± 1,43a | S | 22 ± 1,43a | S |
| Ampicillin (AMP, 10) | 30 ± 0,21ab | S | 20 ± 0,35ab | S | 36 ± 0,36ab | S | 32 ± 0,98ab | S | 32 ± 0,98ab | S |
| Amoxycillin (AMOX, 30) | 40 ± 0,21a | S | 20 ± 0,35ab | S | 38 ± 0,41abc | S | 35 ± 1,81abc | S | 35 ± 1,81abc | S |
| Amoxycillin-clavulanic acid (AMC, 30) | 35 ± 0,98c | S | 20 ± 0,35abc | S | 36 ± 0,98abc | S | 30 ± 1,45ab | S | 30 ± 1,45b | S |
| Carbenicillin (CAR, 100) | 35 ± 0,32abs | S | 20 ± 0,35ab | S | 34 ± 0,56abc | S | 35 ± 1,43abc | S | 35 ± 1,43abc | S |
| Cloxacillin (CX, 5) | 14 ± 0,23a | R | 14 ± 0,23a | R | 14 ± 0,23ab | R | 14 ± 0,23abc | R | 14 ± 0,23a | R |
| Macrolides: | ||||||||||
| Erythromycin (ERO, 15) | 30 ± 0,21a | S | 20 ± 0,11abc | S | 40 ± 0,39ab | S | 40 ± 0,28a | S | 30 ± 0,21a | S |
| Azithromycin (AZM, 15) | 30 ± 0,22a | S | 20 ± 0,35a | S | 35 ± 0,59ab | S | 37 ± 1,52a | S | 30 ± 0,22a | S |
| Cephalosporins: | ||||||||||
| Cefepime (FEP, 30) | 30 ± 0,35bcd | S | 30 ± 0,35abc | S | 30 ± 0,36ab | S | 20 ± 1,43ab | S | 20 ± 1,43ab | S |
| Cefepime/clavulanic acid FEC-40 | 30 ± 0,35a | S | 30 ± 0,35ab | S | 33 ± 0,28a | S | 26 ± 0,23a | S | 26 ± 0,23a | S |
| Cephalatin (KF, 30) | 34 ± 0,21ab | S | 30 ± 0,35a | S | 30 ± 0,54a | S | 25 ± 0,98ab | S | 25 ± 0,98abs | S |
| Cefotaxime (CTX, 30) | 27 ± 0,35a | S | 28 ± 0,11a | S | 25 ± 0,28a | S | 35 ± 1,29ab | S | 35 ± 1,29a | S |
| Aminoglycosides: | ||||||||||
| Gentamicin (CN, 120) | 39 ± 0,37ab | S | 38 ± 0,43ab | S | 40± 0,12ab | S | 41 ± 0,23ab | S | 39 ± 0,31ab | S |
| Streptomycin (STR, 10) | 23 ± 0,36ab | S | 25 ± 0,31ab | S | 28 ± 1,41ab | S | 23 ± 1,41ab | S | 25 ± 0,31ab | S |
| Tobramycin (TOB, 10) | 32 ± 0,32a | S | 25 ± 0,31a | S | 34± 1,18a | S | 35 ± 0,98ab | S | 35 ± 1,29a | S |
| Tetracyclines: | ||||||||||
| Tetracycline (TET, 30) | 30 ± 0,52a | S | 26 ± 0,15a | S | 36 ± 1,43a | S | 30 ± 0,23a | S | 30 ± 0,52ab | S |
| Polypeptides: | ||||||||||
| Polymyxin (PB, 300) | 7 ± 0,28b | R | 0 ± 0,00b | R | 8 ± 1,49b | R | 10± 0,23b | R | 0 ± 0,00b | R |
| Bacitromycin (B, 10) | 0 ± 0,00b | R | 0 ± 0,00b | R | 0 ± 0,00b | R | 0 ± 0,00b | R | 0 ± 0,00b | R |
* The Newman–Keuls test was used to compare means in a data set, and the results suggest that there are statistically significant differences among some of the groups. Here, “±” is a standard error; legend: D and S/R are dimension and sensible/resistant, respectively.
Table 4.
The main constituents of bacterial extract BSS25 were identified through a GC–MS analysis.
| Bacillus thuringiensis F3 (BSS25) | |||||||
|---|---|---|---|---|---|---|---|
| No. | Name | Molecular formula | Molecular Mass, g/mol | Retention Time (min) | PubСhemCompound CID | Similarities | Area, % |
| 1 | Acetone | C3H6O | 58.08 | 1.642 | 180 | 87 | 0.170794 |
| 2 | 2,3-Butanedione | C4H6O | 86.09 | 2.64 | 650 | 93 | 15.84782 |
| 3 | Hexanal | C6H12O | 100.16 | 3.617 | 6184 | 65 | 0.343629 |
| 4 | Acetoin | C4H8O2 | 88.11l | 6.239 | 179 | 78 | 44.063 |
| 5 | 3-Pentanol, 2-methyl- | C6H14O | 102.17 | 7.012 | 11264 | 80 | 1.373128 |
| 6 | Oxirane, (methoxymethyl)- | C4H8O2 | 88.11 | 7.219 | 13589 | 79 | 1.360574 |
| 7 | Acetic acid | C2H4O2 | 60.05 | 8.368 | 176 | 86 | 0.887442 |
| 8 | Decanal | C10H20O | 156.26 | 8.504 | 8175 | 73 | 0.3926 |
| 9 | 1-Hexanol, 2-ethyl- | C8H18O | 130,229 | 8.896 | 7720 | 87 | 0.244481 |
| 10 | Benzaldehyde | C7H6O | 106.12 | 9.411 | 240 | 94 | 2.222227 |
| 11 | 2,3-Butanediol, [S-(R*,R*)]- | C4H10O | 90.12 | 9.491 | 439888 | 82 | 4.077882 |
| 12 | 1,6-Octadien-3-ol, 3,7-dimethyl- | C10H18O | 154.25 | 9.64 | 6549 | 85 | 0.781074 |
| 13 | 1-Hepten-4-ol | C7H14O | 114.19 | 9.847 | 19040 | 70 | 3.418443 |
| 14 | 1-Nonanol | C9H20O | 144.25 | 10.47 | 8914 | 78 | 0.731353 |
| 15 | (S)-(+)-6-Methyl-1-octanol | C9H20O | 144.25 | 10.639 | 13548104 | 85 | 1.248967 |
| 16 | Butanoic acid, 2-methyl- | C5H10O2 | 102.13 | 11.091 | 8314 | 81 | 2.237883 |
| 17 | Oxime-, methoxy-phenyl | C8H9NO2 | 151.16 | 12.063 | 9602988 | 73 | 1.136385 |
| 18 | 1-Decanol | C10H22O | 158.28 | 12.21 | 8174 | 60 | 0.516912 |
| 19 | Hexanoic acid | C6H12O2 | 116.16 | 13.085 | 8892 | 91 | 1.862169 |
| 20 | 5,9-Undecadien-2-one, 6,10-dimethyl-, (E)- | C13H22O | 194.31 | 13.329 | 1549778 | 60 | 0.661766 |
| 21 | Propanoic acid, 2-methyl-, 3-hydroxy-2,4,4-trimethylpentyl ester | C12H24O | 216.32 | 13.462 | 551387 | 65 | 0.99794 |
| 22 | 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | C16H30O4 | 286.41 | 13.63 | 23284 | 81 | 0.906841 |
| 23 | (R)-(-)-4-Methylhexanoic acid | C7H14O2 | 130.18 | 13.965 | 12600623 | 70 | 0.615997 |
| 24 | Hexanoic acid, 2-ethyl- | C8H16O2 | 144.21 | 14.226 | 8697 | 89 | 2.90418 |
| 25 | Cetene | C16H32 | 224.42 | 14.477 | 12395 | 81 | 0.750762 |
| 26 | Phenol | C6H6O | 94.11 | 14.825 | 996 | 89 | 0.81691 |
| 27 | Neodecanoic acid | C10H20O2 | 172.26 | 15.305 | 62838 | 61 | 0.428921 |
| 28 | Octanoic acid | C8H16O2 | 144.21 | 15.386 | 379 | 91 | 2.465807 |
| 29 | 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | C16H22O4 | 278.34 | 16.151 | 6782 | 77 | 0.437683 |
| 30 | Nonanoic acid | C9H18O2 | 158.24 | 16.478 | 8158 | 90 | 2.797785 |
| 31 | Benzoic acid, 2-ethylhexyl ester | C15H22O2 | 234.33 | 17.083 | 94310 | 61 | 0.172171 |
| 32 | Hexadecanoic acid, methyl ester | C17H34O2 | 270.5 | 17.161 | 8181 | 91 | 1.221765 |
| 33 | 2-Octyl benzoate | C15H22O2 | 234.33 | 17.531 | 243800 | 69 | 0.908645 |
Table 5.
The main constituents of bacterial extract BSS21 were identified through a GC–MS analysis.
| Bacillus toyonensis FORT 102 (BSS21) | |||||||
|---|---|---|---|---|---|---|---|
| No. | Name | Molecular formula | Molecular Mass, g/mol | Retention Time (min) | PubСhemCompound CID | Similarities | Area, % |
| 1 | Acetone | C3H6O | 58.08 | 1.661 | 180 | 79 | 0,551252 |
| 2 | 2,3-Butanedione | C4H6O2 | 86.09 | 2.652 | 650 | 92 | 16,97044 |
| 3 | 2,3-Pentanedione | C5H8O2 | 100.12 | 3.404 | 11747 | 68 | 2,035411 |
| 4 | Acetoin | C4H8O2 | 88.11 | 6.246 | 179 | 84 | 38.25336 |
| 5 | 3-Pentanol, 2-methyl- | C6H14O | 102.17 | 7.008 | 11264 | 81 | 2.916132 |
| 6 | Oxirane, (methoxymethyl)- | C4H8O2 | 88.11 | 7.216 | 13589 | 80 | 2.409396 |
| 7 | Nonanal | C9H18O | 142.24 | 7.73 | 31289 | 85 | 0.746341 |
| 8 | Acetic acid | C2H4O2 | 60.05 | 8.343 | 176 | 97 | 2.456759 |
| 9 | 1-Hexanol, 2-ethyl- | C8H18O | 130.229 | 8.889 | 7720 | 88 | 0.154366 |
| 10 | E-3-Pentadecen-2-ol | C15H30O | 226.4 | 9.048 | 5363322 | 65 | 0.135906 |
| 11 | 2,3-Butanediol, [S-(R*,R*)]- | C4H10O2 | 90.12 | 9.486 | 439888 | 88 | 7.796162 |
| 12 | Formic acid, octyl ester | C9H18O2 | 158.24 | 9.751 | 8176 | 69 | 0.371511 |
| 13 | Propanoic acid, 2-methyl- | C4H8O2 | 88.11 | 9.832 | 6590 | 76 | 1.406486 |
| 14 | 2-Octanol | C8H18O | 130.229 | 9.928 | 20083 | 74 | 0.263263 |
| 15 | (S)-(+)-6-Methyl-1-octanol | C9H20O | 144.25 | 10.631 | 13548104 | 82 | 0.282974 |
| 16 | 1-Nonanol | C9H20O | 144.25 | 11.003 | 8914 | 78 | 0.129921 |
| 17 | Butanoic acid, 2-methyl- | C5H10O2 | 102.13 | 11.073 | 8314 | 87 | 3.572379 |
| 18 | Dodecanal | C12H24O | 184.32 | 11.714 | 8194 | 92 | 0.683743 |
| 19 | Oxime-, methoxy-phenyl-_ | C8H9NO2 | 151.16 | 12.052 | 9602988 | 73 | 0.348646 |
| 20 | 2,4-Decadienal, (E,E)- | C10H16O | 152.23 | 12.851 | 5283349 | 77 | 0.282056 |
| 21 | Pentanoic acid | C5H10O2 | 102.13 | 13.071 | 7991 | 79 | 0.233567 |
| 22 | 3-Buten-2-one, 4-(1-cyclopenten-1-yl)-, (E)- | C9H12O | 136.19 | 13.461 | 5370075 | 76 | 0.401939 |
| 23 | Hexanoic acid, 2-ethyl- | C8H16O2 | 144.21 | 14.199 | 8697 | 80 | 0.315248 |
| 24 | 1-Dodecanol | C12H26O | 186.33 | 14.446 | 8193 | 81 | 0.150351 |
| 25 | Phenol | C6H6O | 94.11 | 14.774 | 996 | 90 | 0.191971 |
| 26 | Octanoic acid | C8H16O2 | 144.21 | 15.314 | 379 | 79 | 0.398877 |
| 27 | Nonanoic acid | C9H18O2 | 158.24 | 16.359 | 8158 | 88 | 0.730917 |
| 28 | Hexadecanoic acid, methyl ester | C17H34O2 | 270.5 | 17.01 | 8181 | 88 | 0.315637 |
| 29 | 1,4-Benzenediol, 2,6-bis(1,1-dimethylethyl)- | C14H22O2 | 222.32 | 17.298 | 75550 | 63 | 0.17709 |
| 30 | Decanoic acid | C10H20O2 | 172.26 | 17.356 | 2969 | 64 | 0.307782 |
| 31 | Benzoic acid, heptyl ester | C14H20O2 | 220.31 | 18.369 | 81591 | 73 | 0.161383 |
| 32 | Benzoic acid | C7H6O2 | 122.12 | 18.739 | 243 | 85 | 0.221304 |
| 33 | 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | C16H22O4 | 278.34 | 19.85 | 6782 | 81 | 0.333842 |
| 34 | Dibutyl phthalate | C16H22O4 | 278.34 | 21.099 | 3026 | 67 | 0.325750 |
| 35 | Hexadecanoic acid | C16H32O2 | 256.42 | 22.611 | 985 | 76 | 4.002248 |
| 36 | Oleic Acid | C18H34O2 | 282.5 | 24.54 | 445639 | 86 | 5.853397 |
| 37 | 9,12-Octadecadienoic acid (Z,Z)- | C18H32O2 | 280.4 | 25.063 | 5280450 | 87 | 3.796889 |
Table 6.
The main constituents of bacterial extract BSS16 were identified through a GC–MS analysis.
| Bacillus acidiproducens NiuFun (BSS16) | |||||||
|---|---|---|---|---|---|---|---|
| No. | Name | Molecular formula | Molecular Mass, g/mol | Retention Time (min) | PubChemCompound CID | Similarities | Area, % |
| 1 | Acetone | C3H6O | 58.08 | 1.67 | 180 | 93 | 0.745862 |
| 2 | Acetoin | C4H8O2 | 88.11 | 6.49 | 179 | 81 | 8.440992 |
| 3 | Acetic acid | C2H4O2 | 60.05 | 8.363 | 176 | 95 | 2.398269 |
| 4 | Benzaldehyde | C7H6O | 106.12 | 9.411 | 240 | 94 | 4.751143 |
| 5 | 3(2H)-Thiophenone, dihydro-2-methyl- | C5H8OS | 116.18 | 9.463 | 61664 | 84 | 6.072668 |
| 6 | Propanoic acid, 2-methyl- | C4H8O2 | 88.11 | 9.839 | 6590 | 92 | 13.97511 |
| 7 | Butanoic acid | C4H8O2 | 88.11 | 10.581 | 264 | 83 | 0.612913 |
| 8 | Butanoic acid, 2-methyl- | C5H10O2 | 102.13 | 11.084 | 8314 | 83 | 29.39467 |
| 9 | Oxime-, methoxy-phenyl-_ | C8H9NO2 | 151.16 | 12.063 | 9602988 | 76 | 1.137091 |
| 10 | Phenol | C6H6O | 94.11 | 14.788 | 996 | 90 | 0.318667 |
| 11 | Nonanoic acid | C9H18O2 | 158.24 | 16.372 | 8158 | 85 | 0.544397 |
| 12 | Hexadecanoic acid, methyl ester | C17H34O2 | 270.5 | 17.016 | 8181 | 91 | 0.720873 |
| 13 | 2-Octyl benzoate | C15H22O2 | 234.33 | 17.363 | 243800 | 68 | 0.683898 |
| 14 | Benzoic acid 2-methylpentyl ester | C13H18O2 | 206.28 | 17.813 | 570433 | 66 | 0.529643 |
| 15 | Benzoic acid, heptyl ester | C14H20O2 | 220.31 | 18.084 | 81591 | 80 | 0.501309 |
| 16 | Benzoic acid, tridecyl ester | C20H32O2 | 304.5 | 18.375 | 9814973 | 75 | 0.560423 |
| 17 | Benzoic acid | C7H6O2 | 122.12 | 18.752 | 243 | 79 | 0.652983 |
| 18 | 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | C16H22O4 | 278.34 | 19.859 | 6782 | 91 | 0.940027 |
| 19 | Pentadecanoic acid | C15H30O2 | 242.4 | 21.427 | 13849 | 63 | 0.410235 |
| 20 | Hexadecanoic acid | C16H32O2 | 256.42 | 22.604 | 985 | 88 | 3.605440 |
| 21 | Octadecanoic acid | C18H36O2 | 284.5 | 24.256 | 5281 | 72 | 1.638403 |
| 22 | Oleic acid | C18H34O2 | 282.5 | 24.542 | 445639 | 90 | 9.677555 |
| 23 | 9,12-Octadecadienoic acid (Z,Z)- | C18H32O2 | 280.4 | 25.067 | 5280450 | 91 | 11.09688 |
Table 7.
The main constituents of bacterial extract BSS13 were identified through a GC–MS analysis.
| Bacillus cereus WAB2133 (BSS13) | |||||||
|---|---|---|---|---|---|---|---|
| No. | Name | Molecular formula | Molecular Mass, g/mol | Retention Time (min) | PubСhemCompound CID | Similarities | Area, % |
| 1 | Acetone | C3H6O | 58.08 | 1.644 | 180 | 94 | 3.660169 |
| 2 | Acetoin | C4H8O2 | 88.11 | 6.49 | 179 | 81 | 0.745862 |
| 3 | Acetic acid | C2H4O2 | 60.05 | 8.335 | 176 | 97 | 6.313543 |
| 4 | Decanal | C10H20O | 156.26 | 9.114 | 8175 | 70 | 0.631996 |
| 5 | Benzaldehyde | C7H6O | 106.12 | 9.402 | 240 | 95 | 6.242524 |
| 6 | Propanoic acid, 2-methyl- | C4H8O2 | 88.11 | 9.828 | 6590 | 92 | 11,51283 |
| 7 | Butanoic acid | C4H8O2 | 88.11 | 10.569 | 264 | 86 | 0.911113 |
| 8 | Butanoic acid, 2-methyl- | C5H10O2 | 102.13 | 11.081 | 8314 | 83 | 31.6913 |
| 9 | Oxime-, methoxy-phenyl-_ | C8H9NO2 | 151.16 | 12.054 | 9602988 | 67 | 1.284529 |
| 10 | Tiglic acid | C5H8O2 | 100.12 | 13.078 | 125468 | 81 | 1.676206 |
| 11 | (R)-(-)-4-Methylhexanoic acid | C7H14O2 | 130.18 | 13.945 | 12600623 | 84 | 0.532951 |
| 12 | Hexanoic acid, 2-ethyl- | C8H16O2 | 144.21 | 14.197 | 8697 | 88 | 1.032308 |
| 13 | Phenol | C6H6O | 94.11 | 14.777 | 996 | 87 | 0.439273 |
| 14 | Octanoic acid | C8H16O2 | 144.21 | 15.317 | 379 | 90 | 1.111877 |
| 15 | Nonanoic acid | C9H18O2 | 158.24 | 16.362 | 8158 | 90 | 2.401952 |
| 16 | Hexadecanoic acid, methyl ester | C17H34O2 | 270.5 | 17.016 | 8181 | 80 | 1.091995 |
| 17 | Decanoic acid | C10H20O2 | 172.26 | 17.359 | 2969 | 66 | 0.942792 |
| 18 | Benzoic acid 2-methylpentyl ester | C13H18O2 | 206.28 | 17.809 | 570433 | 64 | 0.556621 |
| 19 | Benzoic acid, heptyl ester | C14H20O2 | 220.31 | 18.079 | 81591 | 78 | 0.427025 |
| 20 | Benzoic acid | C7H6O2 | 122.12 | 18.742 | 243 | 87 | 0.568841 |
| 21 | 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | C16H22O4 | 278.34 | 19.852 | 6782 | 93 | 1.113468 |
| 22 | Dibutyl phthalate | C16H22O4 | 278.34 | 21.116 | 3026 | 69 | 0.936291 |
| 23 | Hexadecanoic acid | C16H32O2 | 256.42 | 22.596 | 985 | 84 | 4.45127 |
| 24 | Octadecanoic acid, 2-hydroxy-1,3-propanediyl ester | C39H76O5 | 625 | 24.244 | 101269 | 67 | 3.793011 |
| 25 | 9-Octadecenoic acid, (E)- | C18H34O2 | 282.5 | 24.53 | 637517 | 86 | 9.952046 |
| 26 | 9,12-Octadecadienoic acid (Z,Z)- | C18H32O2 | 280.4 | 25.052 | 5280450 | 83 | 5.86713 |
Table 8.
The main constituents of bacterial extract BSS12 were identified through a GC–MS analysis.
| Bacillus safensis AS-08 (BSS12) | |||||||
|---|---|---|---|---|---|---|---|
| No. | Name | Molecular formula | Molecular Mass, g/mol | Retention Time (min) | PubСhemCompound CID | Similarities | Area, % |
| 1 | (2-Aziridinylethyl)amine | C4H10N2 | 86.14 | 1.162 | 97697 | 96 | 0.389922 |
| 2 | 1-Propen-2-ol, acetate | C5H8O2 | 100.12 | 1.667 | 7916 | 65 | 0.784972 |
| 3 | 2,3-Butanedione | C4H6O2 | 86.09 | 2.647 | 650 | 93 | 21.42702 |
| 4 | 3-Penten-1-ol | C5H10O | 86.13 | 3.411 | 510370 | 69 | 2.637362 |
| 5 | Acetoin | C4H8O2 | 88.11 | 5.703 | 179 | 72 | 0.263788 |
| 6 | 3-Pentanol, 2-methyl- | C6H14O | 102.17 | 6.259 | 11264 | 72 | 36.53327 |
| 7 | 2-Nonen-1-ol | C9H18O | 142.24 | 7.011 | 61896 | 82 | 1.230591 |
| 8 | 2-Hydroxy-3-pentanone | C5H10O2 | 102.13 | 7.109 | 521790 | 73 | 0.422042 |
| 9 | Ethane-1,1-diol dibutanoate | C10H18O4 | 202.25 | 7.215 | 551339 | 83 | 1.183593 |
| 10 | Acetic acid | C2H4O2 | 60.05 | 8.354 | 176 | 90 | 0.783519 |
| 11 | 1-Hexanol, 2-ethyl- | C8H18O | 130.229 | 8.888 | 7720 | 93 | 0.440399 |
| 12 | Benzaldehyde | C7H6O | 106.12 | 9.397 | 240 | 96 | 2.170694 |
| 13 | 2,3-Butanediol | C4H10O2 | 90.12 | 9.484 | 262 | 89 | 4.672893 |
| 14 | 1,6-Octadien-3-ol, 3,7-dimethyl- | C10H18O | 154.25 | 9.632 | 6549 | 87 | 0.782704 |
| 15 | Propanoic acid, 2-methyl- | C4H8O2 | 88.11 | 9.832 | 6590 | 67 | 3.156132 |
| 16 | 2,3-Butanediol, [R-(R*,R*)]- | C4H10O2 | 90.12 | 9.925 | 225936 | 74 | 0.530592 |
| 17 | 1-Nonanol | C9H20O | 144.25 | 10.366 | 8914 | 82 | 0.418021 |
| 18 | (S)-(+)-6-Methyl-1-octanol | C9H20O | 144.25 | 10.635 | 13548104 | 89 | 0.984823 |
| 19 | Butanoic acid, 2-methyl- | C5H10O2 | 102.13 | 11.082 | 8314 | 82 | 1.894643 |
| 20 | Oxime-, methoxy-phenyl-_ | C8H9NO2 | 151.16 | 12.053 | 9602988 | 67 | 0.518031 |
| 21 | 2,4-Decadienal | C10H16O | 152.23 | 12.853 | 5283349 | 79 | 0.322369 |
| 22 | 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | C16H30O4 | 286.41 | 13.131 | 23284 | 82 | 0.168580 |
| 23 | (R)-(-)-4-Methylhexanoic acid | C7H14O2 | 130.18 | 13.315 | 12600623 | 62 | 0.133451 |
| 24 | Phenol | C6H6O | 94.11 | 13.711 | 996 | 75 | 0.205061 |
| 25 | Octanoic acid | C8H16O2 | 144.21 | 14.194 | 379 | 76 | 0.185012 |
| 26 | Nonanoic acid | C9H18O2 | 158.24 | 14.587 | 8158 | 76 | 0.138322 |
| 27 | Hexadecanoic acid, methyl ester | C17H34O2 | 270.5 | 14.766 | 8181 | 96 | 2.885995 |
| 28 | 2-Octyl benzoate | C15H22O2 | 234.33 | 15.31 | 243800 | 70 | 0.17637 |
| 29 | Benzoic acid, heptyl ester | C14H20O2 | 220.31 | 15.742 | 81591 | 75 | 0.10189 |
| 30 | Benzoic acid, undecyl ester | C18H28O2 | 276.4 | 16.355 | 229159 | 88 | 0.402725 |
| 31 | Benzoic acid | C7H6O2 | 122.12 | 17.008 | 243 | 89 | 0.281133 |
| 32 | 1,2-Benzenedicarboxylic acid, bis(2-methylpropyl) ester | C16H22O4 | 278.34 | 18.074 | 6782 | 69 | 0.097665 |
| 33 | Oleic Acid | C18H34O2 | 282.5 | 18.364 | 445639 | 76 | 0.140625 |
| 34 | Dibutyl phthalate | C16H22O4 | 278.34 | 19.845 | 3026 | 89 | 0.325177 |
| 35 | Hexadecanoic acid | C16H32O2 | 256.42 | 22.582 | 985 | 82 | 2.377302 |
| 36 | Octadecanoic acid | C18H36O2 | 284.5 | 24.235 | 5281 | 69 | 2.671248 |
| 37 | Oleic Acid | C18H34O2 | 282.5 | 24.517 | 445639 | 87 | 4.585822 |
| 38 | 9,12-Octadecadienoic acid (Z,Z)- | C18H32O2 | 280.4 | 25.039 | 5280450 | 83 | 3.576243 |
Table 10.
Colony morphology and microscopic presentation of isolated bacterial species.
| No. | Isolates | 16S rRNA Amplified Region Length | Bacterial Species | NCBI Accession No |
|---|---|---|---|---|
| 1 | BSS25 | 1420 bp | 99% with Bacillus thuringiensis F3 | MF135173 |
| 2 | BSS21 | 1492 bp | 99% with Bacillus toyonensis FORT 102 | MG561363 |
| 3 | BSS16 | 1452 bp | 99% with Bacillus acidiproducens NiuFun | MF446886 |
| 4 | BSS13 | 1474 bp | 98% with Bacillus cereus WAB2133 | MH169322 |
| 5 | BSS12 | 1449 bp | 99% with Bacillus safensis AS-08 | JX849661 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



