Submitted:
12 October 2023
Posted:
13 October 2023
You are already at the latest version
Abstract
Eimeria tenella rhoptry protein has the properties of a protective antigen. EtROP27 is a pathogenic related gene detected by transcriptome, but its expression pattern, immunogenicity and potency have been unknown. Therefore, a gene segment of EtROP27 was amplified and transplanted to pET28a prokaryotic vector for recombinant protein expression and purified to generate polyclonal antibody. Then, RT-PCR, Western blotting were performed to know the expression pattern of EtROP27. Subsequently, animal experiments were conducted to evaluate the immunoprotective effect of the recombinant protein using different immunizing doses (50, 100, 150 μg). The results showed that the expression of EtROP27 gradually increased with the prolongation of infection time, reaching the highest level after 96 hours and then decreased. And EtROP27 was a natural antigen of coccidia which can stimulate the body to produce high levels of IgY. As recombinant protein vaccines, the immune protection evaluation tests of EtROP27 showed that the average weight gain rate of the immune challenge groups were significantly higher than that of the infection control group. The average lesion scores were significantly lower than that of the control group. The oocyst excretion decreased by 81.25%, 86.21%, 80.01% and the anticoccidial index were 159.45, 171.47, 166.75. EtROP27 is a promising candidate antigen gene for development of coccidiosis vaccine.
Keywords:
Eimeria tenella
; recombinant EtROP27 protein
; expression changes
; immunoprotective
1. Introduction
Chicken coccidiosis is a parasitic protozoonosis caused by one or more of the seven species of Eimeria that are parasitic within the epithelial cells of the chicken intestinal tract [1,2]. Coccidiosis is characterized by intestinal injury, diarrhea, or bloody stools [3]. Eimeria tenella is the most virulent, widely distributed, and most harmful of the seven chicken coccidia species, mainly attacking the cecum[4]. The highest incidence rate was found in 3~6-week-old chicks. The symptoms of excreting blood, stool, or even blood appear within 4~5 days of infection. Large numbers of deaths begin one to two days after the appearance of bloody stools, with a mortality rate of up to 80% in severe cases [5,6,7].
Drug control has been the main method for chicken coccidiosis for a long time. However, many problems such as the emergence of drug resistant strains, drug residues, and increased treatment costs during the prevention and treatment process have greatly limited drug control [8]. Therefore, the key strategy and measure for effective prevention and control of this disease is to develop a safe, effective, low toxicity and environmental protection vaccine against coccidiosis in chickens. Currently, live coccidian oocysts vaccines with virulent and precocious virulence are widely used. However, traditional coccidiosis vaccines have shortcomings which limit their wide application such as high production costs, cumbersome production processes, environmental pollution caused by the spread of pathogens to the outside world, and the improper use of wild virus live worm vaccines can cause outbreaks of coccidiosis [9]. Recent studies have demonstrated that recombinant protein vaccines can be used as effective control measures against coccidiosis [10].
Rhoptry proteins (ROPs) are conservative and immune-protective, and some members are among the main virulence factors of Toxoplasma gondii [11]. Therefore, this protein family is considered a potential candidate molecule in the development of anti-toxoplasmosis vaccines. As one of the key virulence factors of the T. gondii type I strain, ROP18 participates in multiple mechanisms to escape host immune response. ROP5 controls the activity of ROP18 and affects IFN- γ and Irgm3 dependent other effectors to regulate the acute toxicity of T. gondii [12]. Similarly, ROPs play an important role in the process of E. tenella invading host cells. It has the property of a protective antigen, and the corresponding antibody can inhibit the infection of coccidia to the host [13]. Research has proven that EtROP17 can inhibit host cell apoptosis [14]. In ROPs such as ROP14, ROP30, ROP52, and ROP53, E. tenella was detected in the proteome at different developmental stages; they have some degree of homology with the ROPs of T. gondii [14], but their functions are unclear. Zhang Li used RNA-Seq to detect and compare the transcriptome of the precocious strain and the virulent strain at the mitotic stage. q PCR was used to verify and screen the pseudopathogenic-related gene ROP27 of E. tenella, which is presumed to have good immunogenicity [15,16]. However, it is not yet known whether the recombinant protein can effectively prevent and treat E. tenella disease.
Therefore, this study aims to investigate whether chicken EtROP27 can serve as a potential candidate factor for anti E.tenella disease by expressing the recombinant protein ROP27 and detecting its immune protective effect. The research results provide new ideas for the prevention of E. tenella disease and the development of vaccines, which have important scientific significance.
2. Materilas and Methods
2.1. Ethics statement
All experiments involving animals were carried out in accordance with national regulations on the protection of animal welfare and strictly follow the guidelines. Additionally, it was approved by the Animal Protection and Utilization Committee of Guangxi University, China.
2.2. Experimental animals and parasite
Firstly, 15-day-old SPF chicken embryos were obtained from Beijing Meri Avigon Laboratory Animal Technology Co., Ltd. (Beijing, China). The E. tenella Shanxi virulent and precocious strains were used in this experiment, which were donated by the Veterinary Pathology Laboratory of the College of Veterinary Medicine, Shanxi Agricultural University.
2.3. Reagents
PrimeSTAR® Max DNA Polymerase, DL2,000 DNA Marker, DL5,000 DNA Marker, pMD™ 18-T Vector Cloning Kit, E.coli BL21(DE3) Competent Cells, E.coli DH5α Competent Cells, QuickCut™ EcoR I, QuickCut™ Xho I, T4 DNA Ligase, TritonX-100, Tris, Glycine, SDS, Acrylamide, SYBR Green qPCR Kit were purchased from TaKaRa (Ōsaka shi, Japan); pET-28a (+) DNA was purchased from Sangon Biotech (Shanghai, China); isopropyl β-D-thiogalactoside (IPTG), LB broth powder, LB agar powder, Ampicillin, Kanamycin were purchased from Solarbio (Beijing, China); 6*His, His-Tag Monoclonal antibody, HRP-conjugated Affinipure Goat Anti-Rabbit IgG(H+L), HRP-conjugated Affinipure Goat Anti-Mouse IgG(H+L), Fluorescein (FITC)–conjugated Affinipure Goat Anti-Rabbit IgG(H+L), ECL detection kit were purchased from Proteintech (Chicago, IL).
2.4. Plasmid construction
Design specific primers to measure the ROP27 sequence (Cluster-10347.5943) based on transcriptome and send it to the company for synthesis. Primer sequences were as follows: ROP27 sense primer, 5′-GAATTCAGGTAACGAGTCTCTGC-3; and anti-sense primer, 5′-TTGCCAGAATTGGCTCTACTACG-3’. The PCR product was purified and cloned into the pMD18-T vector according to the operation steps of the gel extraction kit. A single bacterial colony was selected for amplification, culture, and sequencing, and the bacterial solution with the most correct sequencing was selected for plasmid extraction. The plasmid and pET-28a (+) DNA were digested using EcoR I and Xho I endonucleases, and the digested products were purified. The target fragment was cloned into the eukaryotic expression vector, and then a single colony was selected for amplification, culture, and sequencing. The plasmid with the correct sequencing was named pET-EtROP27.
2.5. Protein expression and purification
The recombinant plasmid pET-EtROP27 was transformed into BL21 (DE3) Competitive Cells. Subsequently, positive colonies were selected for enrichment. IPTG with a final concentration of 1 mmol/L was added to induce expression for 8 hours when the OD600 value of the bacterial solution was about 0.5. The heavy suspension liquid was placed on ice for ultrasonic crushing treatment, then the sediment and supernatant were collected. We followed the method of protein purification as described by Barkhordari F. Protein purity and concentration were determined through 12% (w/v) sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and the BCA protein determination kit. The purified protein was stored at - 80℃ and used for subsequent experiments.
2.6. Preparation of anti-E. tenella and anti-rEtROP27 positive serum
1×104 sporulated E. tenella oocysts were intragastrically administered to 2-week-old SPF chickens. After 3 days, each chicken was infected with 5000 sporulated oocysts every 3 days for a total of four times. Blood was collected by cardiac puncture and centrifuged to prepare anti-E. tenella positive serum; this was then stored at -80℃ for standby.
Three 2-month-old New Zealand white rabbits were immunized with rEtROP27 which emulsified with Freund's adjuvant at a dose of 200 µg per rabbit three times, once every two weeks. Before each immunization, 30 µL of rabbit blood was taken from the ear vein, and the serum was isolated to detect the antibody titer. The heart took blood and centrifuged to collect serum after the third immunization. The collected serum was deactivated in a water bath at 56℃ for 30 min and stored at - 80℃ for standby.
2.7. Indirect ELISA determination of antibody titer
Rabbit anti-EtROP27 serum is the sample to be tested, and the non-immunized serum is negative. The purified EtROP27 recombinant protein was used as the detection antibody. We used PBST to test the serum according to 1:500, 1:1000, 1:2000, 1:4000, 1: 8000, 1:16000, and 1:32000 gradient dilution. Serum antibody titer testing was performed according to the method reported by Dong-chao Zhang [17].
2.8. Western blotting analysis
The protein samples were separated using a 12% polyacrylamide gels and were transferred to 0.22 µm polyvinylidene fluoride membranes using the Bio-Rad wet transfer system. After blocking at 37°C with 5% BSA in PBS, the membranes were probed with anti-E. tenella (1:400), anti-rEtROP27(1:500), anti-HIS (1:2000), respectively, with an overnight incubation at 4°C. Next, the membranes were washed with PBS containing tween-20 (PBST) four times and were incubated with horseradish peroxidase-linked secondary anti-chicken, anti-rabbit or anti-mouse IgG antibodies for 1 h at 37°C. After washing with PBST, the signal was visualized using an ECL detection kit.
2.9. Primary culture of chicken embryo caecal epithelial cells
The cecum was removed from 15-day-old SPF embryos and striped the mesentery carefully; the cecum was fully washed by the PBS buffer, then cut into 1 mm3 size tissue and washed; the tissue was resuspended and mixed by thermolysin (50 mg/L), and digested at 41℃ for 2 h; PBS solution was used to stop the digestion, centrifugation at 1200 r/min for 5 min, and PBS and enzyme solution were discarded; the cell pellet was resuspended in 10% FBS low-glycemic DMEM cell culture medium, seeded in a cell culture flask and adhered to the culture for 70 min; the liquid in the flask was collected, centrifuged at 1200 r/min for 5 min, and then resuspended and counted in DMEM/F12 culture medium containing 2.5% fetal bovine serum (FBS); the cells were seeded in a cell culture plate and could be used for subsequent experiments when the cell attachment rate was reached about 85%.
2.10. E. tenella sporozoite preparation
E. tenella sporulated oocysts were centrifuged at 2000 r/min for 5 min, the supernatant was discarded, and PBS was added to wash thoroughly. The oocysts were suspended in 2 mL PBS and ground with a homogenizer until the decapsulation rate reached 80%. The above solution was centrifuged at 1800 r/min for 5min. Precipitates were then resuspended with sporocyst digestion solution which contains 0.75% trypsin and 10% chicken bile and digest with shaking at 41℃ (150 r/min) until 80% of the sporozoites are released. After filtration, the digestion solution was removed by centrifugation at 3000 r/min for 10 min, and the precipitate was suspended in DMEM solution. The amount of sporozoites used in this experiment was 1×104.
2.11. RT-PCR analysis
Total RNA from chicken cecum or primary cecal epithelial cells of chicken embryo cells infected with E. tenella virulent and precocious strains was extracted using TRIzol Reagent according to the manufacturer’s instructions. The cDNA was generated by reverse transcription using PrimeScript reverse transcriptase according to the manufacturer’s instructions. ROP27 mRNA was assessed by using quantitative qPCR with a TaKaRa SYBR Green. qPCR Kit on a CFX96 Touch real-time PCR detection system. Primer sequences were as follows: ROP27 sense primer, 5′-AGCTACGACACTCCTGTTGC-3′; and anti-sense primer, 5′-ACTCAAGACGGAGTTGCTGG-3′. PCR products were purified using a Gel Extraction Kit and cloned into the pMD18-T vector. Plasmids were diluted serially and used as standards for quantitative analysis. The initial copy number of ROP27 gene in each group was calculated using the following formula: X = -K log Ct + b, where X is the initial copy number and K, Ct, and b refer to the slope rate, cycle threshold, and constant, respectively.
2.12. Animal experiment
Firstly, 14-day-old chickens were randomly divided into 5 groups, with 10 chickens in each group. Three experimental groups of chicken legs were subcutaneously injected with recombinant proteins emulsified using Freund's adjuvant (50 μg, 100 μg, 150 μg, respectively). The infection control group and the uninfection control group were injected with PBS emulsified with complete Freund's adjuvant. After 7 days, emulsify with incomplete Freund's adjuvant and undergo a second immunization at the same dose. After 7 days, each chicken in the experimental group and the infection control group received oral administration of 5×104 sporulation oocysts of E. tenella were obtained, and PBS was given to the uninfection control group.
2.13. Concentration of serum antibody
Concentration of IgY antibody level in serum were detected by utilizing an indirect enzyme-linked immunosorbent assay (ELISA) commercial kits named "chick cytokine ELISA Quantitation Kits" (catalog numbers: CSB-E11635Ch for IgY CUSABIO, Wuhan, China), according to manufacturer’s instructions.
2.14. Immunoprotective parameters in vivo
In vivo immunoprotective parameters of the rEtROP27 protein include clinical (weight, survival rate and mortality), pathological (cecum lesion score), and parasitological (ACI and oocyst output) factors [18]. Weight gain and survival rate were calculated directly. Cecum lesion score is a continuous number from 0 (none) to 4 (severe) representing different levels of cecum lesion, according to Johnson and Reid [19], which is evaluated by three independent observers. Furthermore, cecal content was collected in order to calculate oocysts per gram (OPG) using McMaster,s counting technique [20]. ACI as a synthetic criterion, which responded to holistic health conditions of chickens, and represents various degrees of vaccine protection: ACI > 180 is good protection, 160 < ACI < 179 is moderate protection, 120 < ACI < 159 is limited protection, and ACI < 120 was no protection [21].
2.15. Image and statistical analyses
Values are presented as the arithmetic mean ± standard error. Each experiment was repeated at least three times. The SPSS 17.0 statistical software package (Chicago, USA) was used to perform ANOVA analysis of all data. Histograms were prepared via GraphPad Prism 5.0 software (San Diego, CA, USA). Fluorescence intensity was analyzed using ImageJ software (National Institutes of Health, USA). All results were considered statistically significant at a p-value < 0.05.
3. Results
3.1. Bioinformatics analysis of EtROP27
EtROP27 showed a specific band at 1068 bp after PCR amplification. ExPASy ProtParam tool analysis showed that the sequence encodes 355 amino acids, predicting a protein size of 38.07 kD, an isoelectric point of 6.82, and a hydrophobicity prediction GRAVY of 0.248. SignalP-5.0 Server sequence analysis shows that there is no secretion signal sequence present (Figure 1A). TMHMM analysis shows that the protein has no transmembrane region (Figure 1B)
3.2. Gene clone and plasmid construction
The PCR product was then cloned into the pMD-18T vector, and positive colonies were selected for bacterial liquid PCR and sequencing analysis (Figure 2A-B). We cut out the target fragment from the T vector and cloned it into the pET-28a vector. We then selected the positive colonies for bacterial liquid PCR, double digestion identification, and sequencing analysis (Figure 2C-D). The prokaryotic expression vector pET-EtROP27 was constructed successfully.
3.3. Expression and purification of rEtROP27 protein
Coomassie brilliant blue staining showed that the highest expression was observed after 6 h of IPTG induction (Figure 3A). After purification, the target band appears at about 48 kDa, and Elution buffer with a concentration of 150 mmol/L imidazole has the best elution effect (Figure 3B). Western blotting results show that His tag anti body and chicken serum infected with coccidia were primary antibodies that could produce clear and sized target bands, while healthy chicken serum as primary antibody did not show any bands (Figure 3C). Chicken naturally infected with coccidia produced antibodies against EtROP27. The results indicate that EtROP27 is a natural antigen of E. tenella and has the basis of a candidate vaccine protein.
3.4. Determination of rEtROP27 polyclonal antibody titer
The rabbit anti-EtROP27 serum was the sample chosen to be tested; the non immunized serum was negative. The purified EtROP27 recombinant protein was used as the detection antibody. After three immunizations with EtROP27 fusion protein, blood and serum were collected to determine the antibody titer. From Table 1, it can be seen that the rabbit serum titer reaches 1:2.56×104 after three immunizations. This indicates that the immunogen injection has achieved a good immune effect, and the positive serum can be used for subsequent experiments. Western blotting results show that rEtROP27 protein can be detected by EtROP27 polyclonal antibody (Figure 4).
3.5. Changes in EtROP27 expression
The expression of EtROP27 mRNA and protein was detected at different time points in primary chicken embryo cecal epithelial cells or 14-day-old chickens infected with E. tenella. The result shows that the expression of EtROP27 mRNA in the primary cecal epithelial cells of chicken embryos infected with E. tenella gradually increased with the prolongation of infection time, reaching a peak at 96 h, and then decreased. The protein expression pattern was also consistent with the above (Figure 5A-D). The results showed that EtROP27 had the highest expression level during the first generation of schizogenesis.
3.6. Immunoprotective of rEtROP27 protein in vivo
Four chickens died in the challenged control group, while no deaths were observed in the other groups. Average and relative body weight gains of chickens immunised rEtROP27 protein and was significantly higher than in the infection control groups (Figure 6A-B). Likewise, chickens vaccinated with either rEtROP27 protein showed reduced oocyst output, lower lesion scores and bloody stool compared with infection control groups (Figure 6C-F). The ACI value of low-dose groups (50 μg) of rEtROP27 protein was more than 155, medium-dose groups (100 μg) and high-dose groups (150 μg) were more than 160 (Table 2). The results indicate that the optimal immune dose for protein is 100 μg.
The IgY antibody levels in the serum of different doses of EtROP27 immunized groups were significantly higher than those of the control group 7 days after the second immunization (0 d post infection), with the 100 μg group having the highest antibody level (Figure 6G). The serum IgY levels in the EtROP27 immune group with different doses and the challenged group at 7 d post infection were significantly higher than those at 0 d post infection (Figure 6G). These results indicate that EtROP27 is an antigen of E. tenella and can induce a higher level of humoral immune response in order to resist E. tenella infection.
4. Discussion
Chicken coccidiosis is one of the most important diseases threatening the health and welfare of poultry caused by Eimeria. In poultry, there are mainly seven different types of Eimeria, of which E. tenella is the most pathogenic and widely distributed species [22]. The life cycle of Eimeria tenella is relatively complex, and can be generally divided into two developmental stages: the exogenous stage (spore-stage reproduction) and the endogenous stage (schizogeny and gametogenesis). The predation of host cell nutrients by worms, and the formation and release of merozoites and gametophytes destroy cell structures and functions, causing hemorrhagic inflammation, emaciation, and death in the cecum during E. tenella schizogeny and gametogenesis [22]. At present, the control of coccidiosis mainly relies on anti coccidiosis drugs, but there are many shortcomings. Therefore, there is an urgent need to develop a safe and effective vaccine against chicken coccidia to effectively prevent and control this disease.
E. tenella is a member of the apicomplexan protozoa [23]. The protozoa of the apicomplexan have conservative and unique secretory organelles, including rod-shaped bodies, dense particles, and microlines, which have relatively conservative invasion mechanisms against host cells [24]. Rod-shaped bodies are rod-shaped organelles and large vesicles containing dense substances, and are composed of osmiophilic substances. Secretion of rhoptry neck protein (RON) related to parasite invasion in the early stage is crucial for the formation and function of mobile connections between parasites and host cell membranes, such as RON1, RON2, and RON3 [25]. The release of ROPs from the base of the rod-shaped body into the PV changes the environment of the PV and enters the host cell [26,27]. They interact with the host cell through signal transduction, causing a series of downstream effects, becoming a key determinant of protozoan virulence. After entering the cell, ROPs can disrupt host cell signaling and defense mechanisms, and assist in recruiting host organelles [28].
EtROP plays an important role in parasitic invasion of host cells, modification of host vacuoles, hijacking, and regulation of host cells, and has been proven to be a key virulence factor. It also has the properties of a protective antigen, and corresponding antibodies can inhibit the infection of coccidia to the host [28]. Studies have confirmed the expression of EtROP17 in E. tenella merozoites. Western blotting analysis showed that EtROP17 can be recognized by the chicken immune system and induce antibody responses. Vaccination of animals with rEtROP17 can significantly reduce egg sac output and reduce cecal lesions [29]. These indicate that EtROP17 can be used as an effective candidate vaccine for E. tenella. Similarly, studies have shown that EtROP30 and EtROP35 are natural antigens of coccidia, which have good immunogenicity and can be used as the basis for candidate vaccine proteins [30,31].
EtROP27 is a differential gene detected by the transcriptome which is presumed to be a pathogenic-related gene [15,16]. However, its expression and function are still unclear. The different expression levels of secreted proteins at different stages of the parasite's life cycle are often closely related to their function. High expression proteins in the mitotic stage of invading host cells are usually associated with invading host cells, or with parasite development and host–parasite interactions [32]. The extracellular stage (sporozoite and merozoite) of Eimeria is immunologically fragile, but it is crucial for the invasion and development of the parasite [33]. The high expression of proteins in the sporozoite and/or merozoite stage makes it an ideal candidate vaccine. The highly expressed ROP Et ROPK Eten5-A of E. tenella during the sporozoite stage has good immunogenicity [34]. In this study, we first conducted bioinformatics analysis on EtROP27. The analysis results indicate that the protein is a secreted protein with potential as an antigen. Subsequently, a prokaryotic expression recombinant plasmid for EtROP27 was successfully constructed and purified, and polyclonal antibodies against EtROP27 were successfully obtained. RT-PCR and Western blotting results indicate that EtROP27 protein is naturally present and expressed at all stages of coccidian development, with the highest expression level in the first generation of schizogenesis. Therefore, it may be a good candidate protein for vaccines.
Subunit vaccines are expected to become an alternative control strategy for avoiding defects in anti coccidian drugs and live egg sac vaccines, and the screening of effective immune protective proteins is a key focus of subunit vaccine research. Animal experiments showed that the rEtROP27 protein could cause increased average body weight gain, increased IgY titer in serum, decreased oocyst output and bloody stool, and lower mean lesion scores compared with the infection control. The ACI value of rEtROP27 protein 100 μg or 150 μg reached more than 160. These results indicate that EtROP27 can effectively induce immune protection against E. tenella in chickens.
This study provides a basis for determining the key genes that dominate or regulate the pathogenicity of coccidia, for searching for drug action targets that control the pathogenicity of coccidia, for developing gene deletion vaccines with strong fecundity and immunogenicity but weak adverse reactions, and for further reducing the adverse effects of precocious strains. They will also provide new ideas and scientific bases for further revealing the pathogenic mechanism of coccidia against the host.
5. Conclusions
This study successfully constructed the eukaryotic expression vector of E. tenella ROP27 and purified the recombinant protein. Our findings suggest that ROP27 is a natural protein of E. tenella and is highly expressed during the first generation of schizogenesis. The results of animal experiments have shown that medium to high doses of rEtROP27 can have a moderate effect against coccidia, indicating that this gene can serve as a potential candidate factor for resistance to chicken E. tenella disease, providing new ideas for the prevention of E. tenella disease and the development of vaccines.
Author Contributions
Meng-gang Li carried out most of the experiments, wrote the manuscript, and should be considered as primary author. Ying-yi Wei critically revised the manuscript and the experiment design. Xiao-ling Lv, Ming-xue Zheng helped with the experiment. All the authors read and approved the final version of the manuscript.
Funding
This study was funded by the Science and Technology Innovation Fund Project of Shanxi Agricultural University (Grant No. 2020BQ08), the Scientific research project of Shanxi Province outstanding doctoral work award fund (Grant No. SXYBKY2019023), Innovation Projects of College of Veterinary Medicine, Shanxi Agricultural University (Grant No. J202111305).
Acknowledgments
Special thanks to Li Zhang, Rui Bai, Yong-bin Li and Kui-hao Liu for their help during the study.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Lv, X.L.; Wang, Y.Y.; Zheng, M.X.; Bai, R.; Zhang, L.; Duan, B.T.; Lei, X.; Zhang, X.S.; Zhao, Y.J.; Cui, K.L.; et al. The role of Ca2+ in the injury of host cells during the schizogenic stage of E. tenella. Poultry Sci 2022, 101. [CrossRef]
- Lv, X.L.; Chen, Z.Y.; Zheng, M.X.; Bai, R.; Zhang, L.; Zhang, X.S.; Duan, B.T.; Zhao, Y.J.; Yin, L.Y.; Fan, B.L.; et al. The interaction between free Ca2+ in host cells and invasion of E. tenella. Parasitol Res 2022, 121, 965-972. [CrossRef]
- Zhang, X.; Li, S.; Zheng, M.; Zhang, L.; Bai, R.; Li, R.; Hao, S.; Bai, B.; Kang, H. Effects of the PI3K/Akt signaling pathway on the apoptosis of early host cells infected with Eimeria tenella. Parasitol Res 2020, 119, 2549-2561. [CrossRef]
- Zhang, Y.; Duan, B.T.; Zhao, Y.J.; Cui, K.L.; Xu, T.; Zhang, X.S.; Lv, X.L.; Guo, L.L.; Zheng, M.X.; Bai, R. Pathogenic mechanism of Eimeria tenella autophagy activation of chicken embryo cecal epithelial cells induced by Eimeria tenella. Poult Sci 2023, 102, 102535. [CrossRef]
- Xu, Z.; Zheng, M.; Zhang, L.; Zhang, X.; Zhang, Y.; Cui, X.; Gong, X.; Xi, R.; Bai, R. Effect of ATP and Bax on the apoptosis of Eimeria tenella host cells. BMC Vet Res 2017, 13, 399. [CrossRef]
- Xu, Z.Y.; Zheng, M.X.; Zhang, L.; Gong, X.; Xi, R.; Cui, X.Z.; Bai, R. Dynamic expression of death receptor adapter proteins tradd and fadd in Eimeria tenella-induced host cell apoptosis. Poult Sci 2017, 96, 1438-1444. [CrossRef]
- Li, S.; Zheng, M.X.; Xu, H.C.; Cui, X.Z.; Zhang, Y.; Zhang, L.; Yang, S.S.; Xu, Z.Y.; Bai, R.; Sun, X.G. Mitochondrial pathways are involved in Eimeria tenella-induced apoptosis of chick embryo cecal epithelial cells. Parasitol Res 2017, 116, 225-235. [CrossRef]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet Res 2020, 51, 115. [CrossRef]
- Jenkins, M.C.; Parker, C.; Ritter, D. Eimeria Oocyst Concentrations and Species Composition in Litter from Commercial Broiler Farms During Anticoccidial Drug or Live Eimeria Oocyst Vaccine Control Programs. Avian Dis 2017, 61, 214-220. [CrossRef]
- Lee, Y.; Lu, M.; Lillehoj, H.S. Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies. Vaccines (Basel) 2022, 10. [CrossRef]
- Possenti, A.; Di Cristina, M.; Nicastro, C.; Lunghi, M.; Messina, V.; Piro, F.; Tramontana, L.; Cherchi, S.; Falchi, M.; Bertuccini, L.; et al. Functional Characterization of the Thrombospondin-Related Paralogous Proteins Rhoptry Discharge Factors 1 and 2 Unveils Phenotypic Plasticity in Toxoplasma gondii Rhoptry Exocytosis. Front Microbiol 2022, 13, 899243. [CrossRef]
- Sparvoli, D.; Delabre, J.; Penarete-Vargas, D.M.; Kumar Mageswaran, S.; Tsypin, L.M.; Heckendorn, J.; Theveny, L.; Maynadier, M.; Mendonca Cova, M.; Berry-Sterkers, L.; et al. An apical membrane complex for triggering rhoptry exocytosis and invasion in Toxoplasma. EMBO J 2022, 41, e111158. [CrossRef]
- Bermudez, M.; Arevalo-Pinzon, G.; Rubio, L.; Chaloin, O.; Muller, S.; Curtidor, H.; Patarroyo, M.A. Receptor-ligand and parasite protein-protein interactions in Plasmodium vivax: Analysing rhoptry neck proteins 2 and 4. Cell Microbiol 2018, 20, e12835. [CrossRef]
- Oakes, R.D.; Kurian, D.; Bromley, E.; Ward, C.; Lal, K.; Blake, D.P.; Reid, A.J.; Pain, A.; Sinden, R.E.; Wastling, J.M.; et al. The rhoptry proteome of Eimeria tenella sporozoites. Int J Parasitol 2013, 43, 181-188. [CrossRef]
- Zhang, L.; Zheng, M.X.; Xi, R.; Xu, Z.Y.; Zhang, X.S.; Zheng, L.L.; Bai, R.; Mi, C.L.; Hao, F.F.; Feng, Y.P. Comparison of the host cells apoptosis induced by precocious strains and virulent strains of Eimeria tenella. Poult Sci 2019, 98, 4384-4390. [CrossRef]
- Zhang, L. Study on pathogenicity differences and pathogenic genes of E. tenella precocious strains and virulent strains. 2020.
- Dongchao Zhang, N.J., Qijun Chen. Vaccination with recombinant adenoviruses expressing Toxoplasma gondii MIC3, ROP9, and SAG2 provide protective immunity against acute toxoplasmosis in mice. Vaccine 2019, 37. [CrossRef]
- Morehouse NF, B.R. Coccidiosis: Evaluation of coccidiostats by mortality, weight gains, and fecal scores. Exp Parasitol 1970, 28. [CrossRef]
- Johnson J, R.W. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp Parasitol 1970, 28. [CrossRef]
- P L Long, B.J.M., L P Joyner, C C Norton. A guide to laboratory techniques used in the study and diagnosis of avian coccidia. Folia Vet Lat 1976, 6, doi:1010500.
- HD, C. Evaluation of the efficacy of anticoccidial drugs against Eimeria species in the fowl. Int J Parasitol 1998, 28. [CrossRef]
- Fatoba AJ, A.M. ransgenic Eimeria parasite: A potential control strategy for chicken coccidiosis. Acta Trop 2020, 205. [CrossRef]
- Huang J, C.S., Zhou Z, Sun X, Haseeb M, Lakho SA, Zhang Y, Liu J, Shah MAA, Song X, Xu L, Yan R, Li X. Poly (D, L-lactide-co-glycolide) delivery system improve the protective efficacy of recombinant antigen TA4 against Eimeria tenella infection. Poult Sci 2021, 100. [CrossRef]
- Patarroyo MA, M.-F.J., Gómez M, Arévalo-Pinzón G, Patarroyo ME. Hotspots in Plasmodium and RBC Receptor-Ligand Interactions: Key Pieces for Inhibiting Malarial Parasite Invasion. Int J Mol Sci 2020, 21. [CrossRef]
- Lim DC, C.B., Doerig C, Saeij JP. Toxoplasma and Plasmodium protein kinases: Roles in invasion and host cell remodelling. Int J Parasitol 2012, 42. [CrossRef]
- Pan M, Z.Y., Wang Y, Li L, Song Y, Hou L, Zhao J. Screening and Identification of the Host Proteins Interacting with Toxoplasma gondii Rhoptry Protein ROP16. Front Microbiol 2017, 8. [CrossRef]
- Ihara F, N.Y. Toxoplasma gondii manipulates host cell signaling pathways via its secreted effector molecules. Parasitol Int 2021, 83. [CrossRef]
- Meng YJ, M.B., Liu XX, Yu LM, Zheng WB, Xie SC, Gao WW, Zhu XQ, Liu Q. Transcriptional changes in LMH cells induced by Eimeria tenella rhoptry kinase family protein 17. Front Vet Sci 2022, 9. [CrossRef]
- Liu X, M.B., Zheng W, Meng Y, Yu L, Gao W, Zhu X, Liu Q. Identification and Protective Efficacy of Eimeria tenella Rhoptry Kinase Family Protein 17. Animals (Basel) 2022, 12. [CrossRef]
- Wang B, Z.N., Sun J, Sun L, Li H, Wu Z, Li H, Zhang X, Zhao X. Identification and Characterization of Eimeria tenella Rhoptry Protein 35 (EtROP35). Vet Sci 2022, 29. [CrossRef]
- Wang B, Z.N., Sun J, Sun L, Li H, Wu Z, Li H, Zhang X, Zhao X. Characterization of the Eimeria tenella rhoptry protein with a nuclear localization sequence (EtROP30). Parasitol Res 2022, 121. [CrossRef]
- Fernandez-Becerra C., B.M., Castellanos A., Correa B. R., Obadia T., Ramirez M., Rui E., Hentzschel F., Lopez-Montanes M., Ayllon-Hermida A., Martin-Jaular L., ElizaldeTorrent A., Siba P., Vencio R. Z., Arevalo-Herrera M., Herrera S., Alonso P. L., Mueller I. and Del Portillo H. A. Plasmodium vivax spleen-dependent genes encode antigens associated with cytoadhesion and clinical protection. Proc Natl Acad Sci U S A 2020, 117. [CrossRef]
- Rezaei F., S.S., Sharif M., Hejazi S. H., Pagheh A. S., Aghayan S. A. and Daryani A. A systematic review of Toxoplasma gondii antigens to find the best vaccine candidates for immunization. Microb Pathog 2019, 126. [CrossRef]
- Song X., Y.X., Zhang T., Liu J. and Liu Q. A Novel Rhoptry Protein as Candidate Vaccine against Eimeria tenella Infection. Vaccines (Basel) 2020, 8. [CrossRef]
Figure 1.
Bioinformatics analysis of EtROP27. (A) Signal peptide prediction of EtROP27 by SignalP-5.0 Server. (B) Transmembrane region prediction of EtROP27 by TMHMM Server v. 2.0.
Figure 1.
Bioinformatics analysis of EtROP27. (A) Signal peptide prediction of EtROP27 by SignalP-5.0 Server. (B) Transmembrane region prediction of EtROP27 by TMHMM Server v. 2.0.
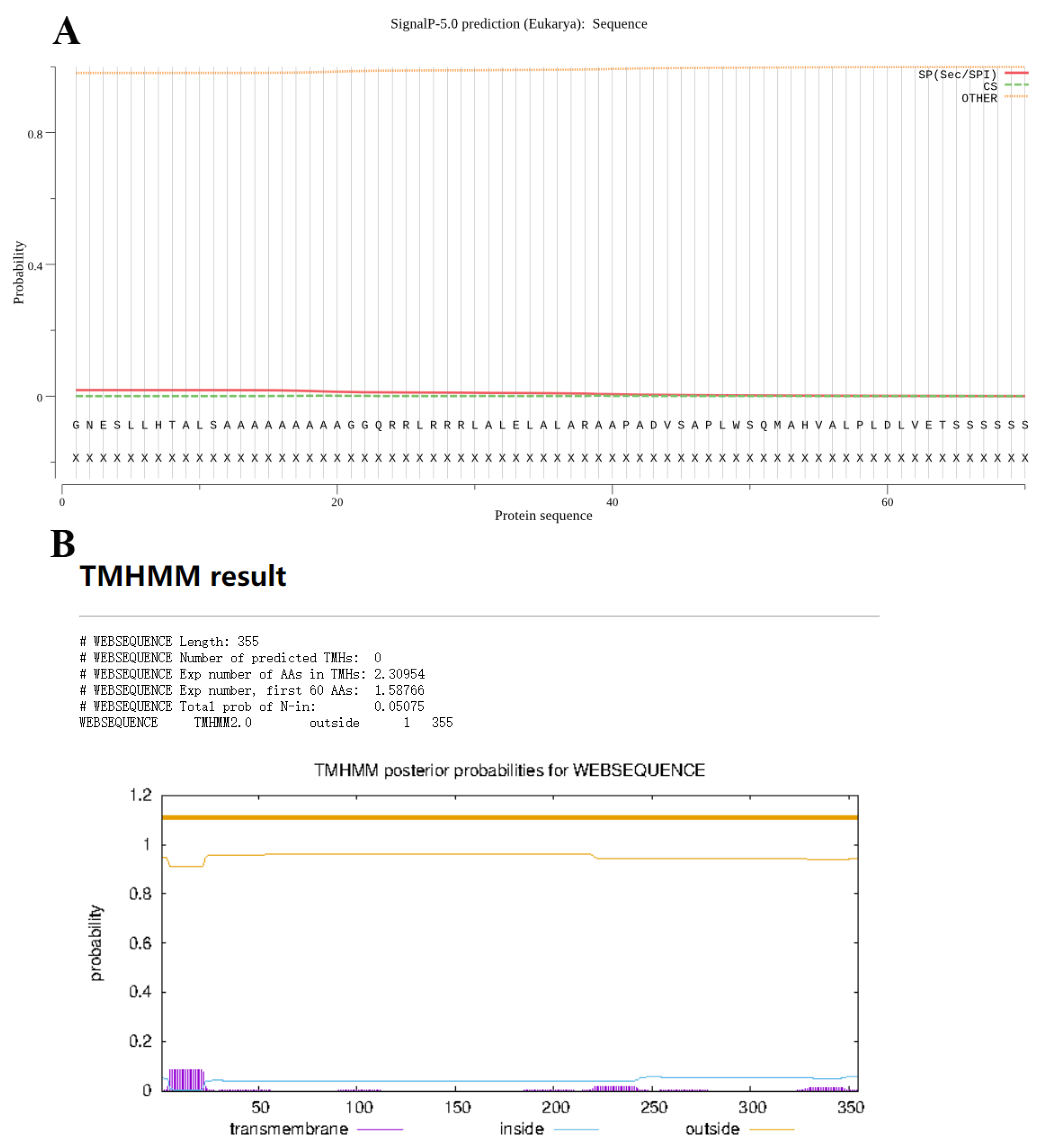
Figure 2.
The construction of pET-EtROP27 Expression Vector. (A) EtROP27 gene amplification.1-4:annealing temperatures are 52 ℃, 54 ℃, 56 ℃, and 58℃, respectively. 5:negative control. (B) The bacterial solution PCR results of pMD-EtROP27. 1-3:Monoclonal colony amplified bacterial liquid. (C) The bacterial solution PCR results of pET-EtROP27. (D) The results of double enzyme digestion of pET-EtROP27. M: DNA marker.
Figure 2.
The construction of pET-EtROP27 Expression Vector. (A) EtROP27 gene amplification.1-4:annealing temperatures are 52 ℃, 54 ℃, 56 ℃, and 58℃, respectively. 5:negative control. (B) The bacterial solution PCR results of pMD-EtROP27. 1-3:Monoclonal colony amplified bacterial liquid. (C) The bacterial solution PCR results of pET-EtROP27. (D) The results of double enzyme digestion of pET-EtROP27. M: DNA marker.
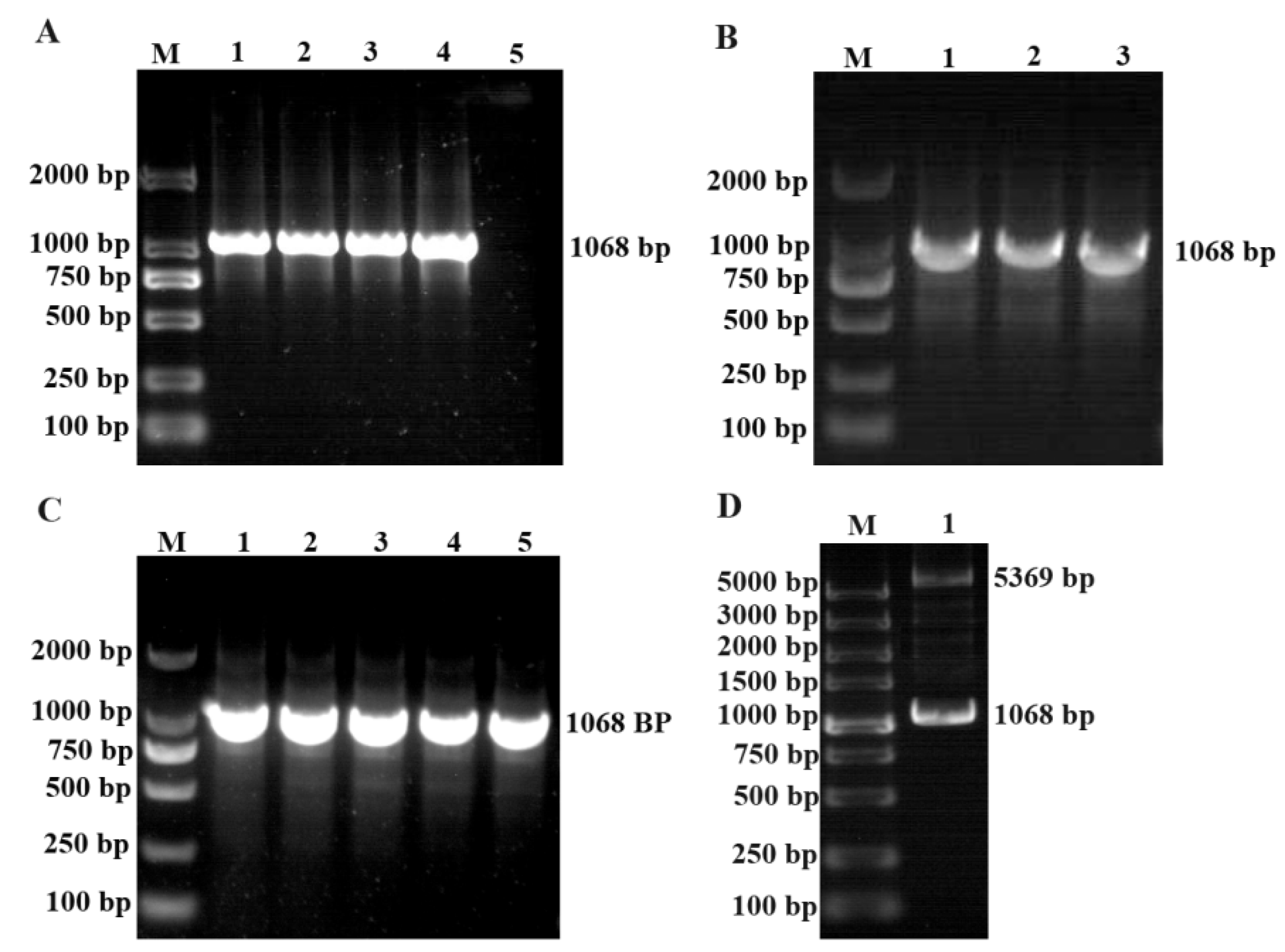
Figure 3.
The expression, purification and identification of rEtROP27. (A) Coomassie bright blue staining of rEtROP27 protein. (B) Coomassie brilliant blue staining of purified rEtROP27 protein. 1:100 mmol/L imidazole. 2:150 mmol/L imidazole. (C) Western blotting identification results.
Figure 3.
The expression, purification and identification of rEtROP27. (A) Coomassie bright blue staining of rEtROP27 protein. (B) Coomassie brilliant blue staining of purified rEtROP27 protein. 1:100 mmol/L imidazole. 2:150 mmol/L imidazole. (C) Western blotting identification results.
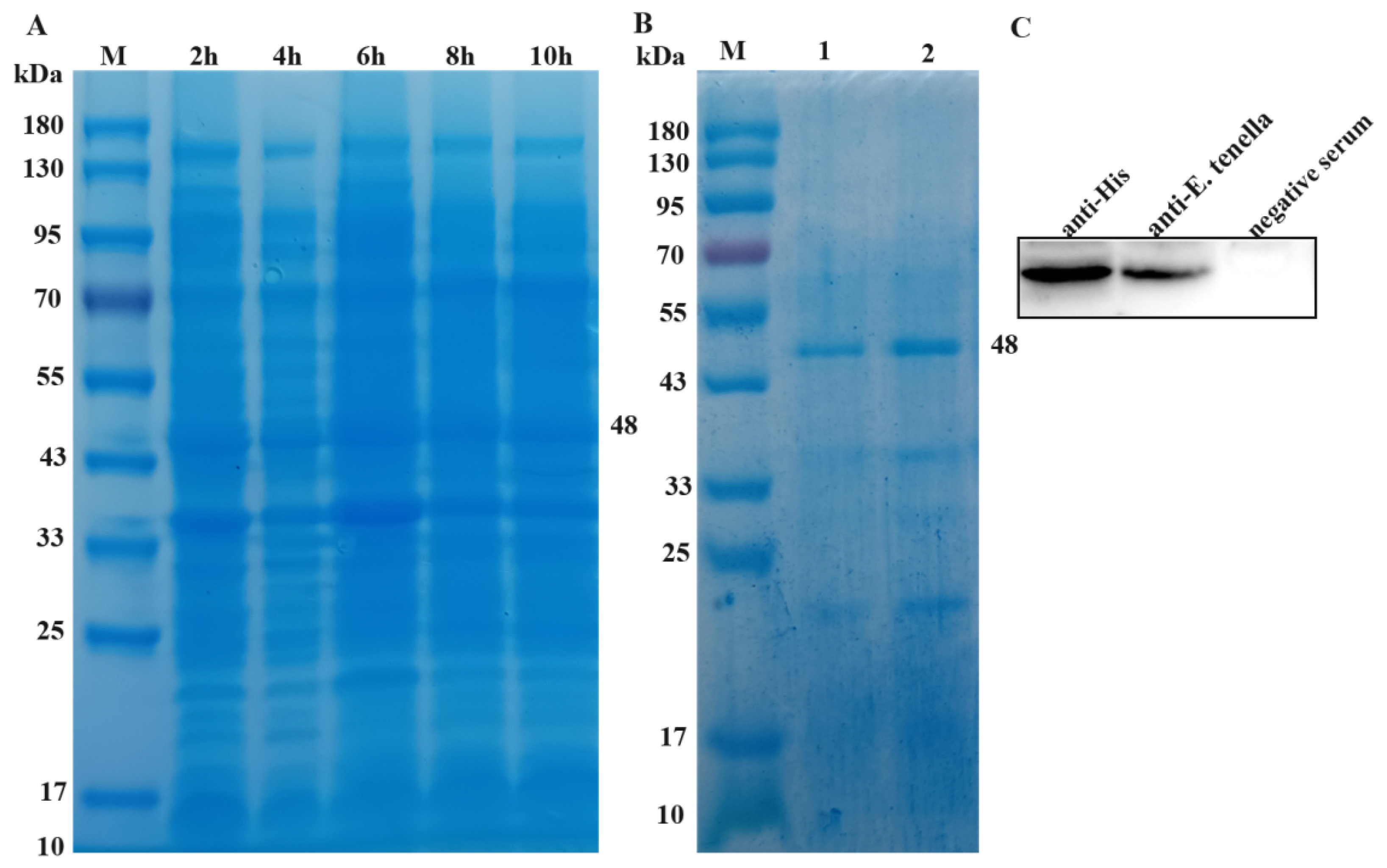
Figure 4.
EtROP27 polyclonal antibody specific detection.

Figure 5.
The expression of EtROP27 inin cells and tissues. (A-B) The expression of EtROP27 mRNA was detected by qRT-PCR. A: chicken embryo cecal epithelial cells. B:cecal tissue. (C-D) The expression of EtROP27 protein was detected by western blotting. C: chicken embryo cecal epithelial cells. D:cecal tissue. All of the data are representative of at least three independent experiments.
Figure 5.
The expression of EtROP27 inin cells and tissues. (A-B) The expression of EtROP27 mRNA was detected by qRT-PCR. A: chicken embryo cecal epithelial cells. B:cecal tissue. (C-D) The expression of EtROP27 protein was detected by western blotting. C: chicken embryo cecal epithelial cells. D:cecal tissue. All of the data are representative of at least three independent experiments.
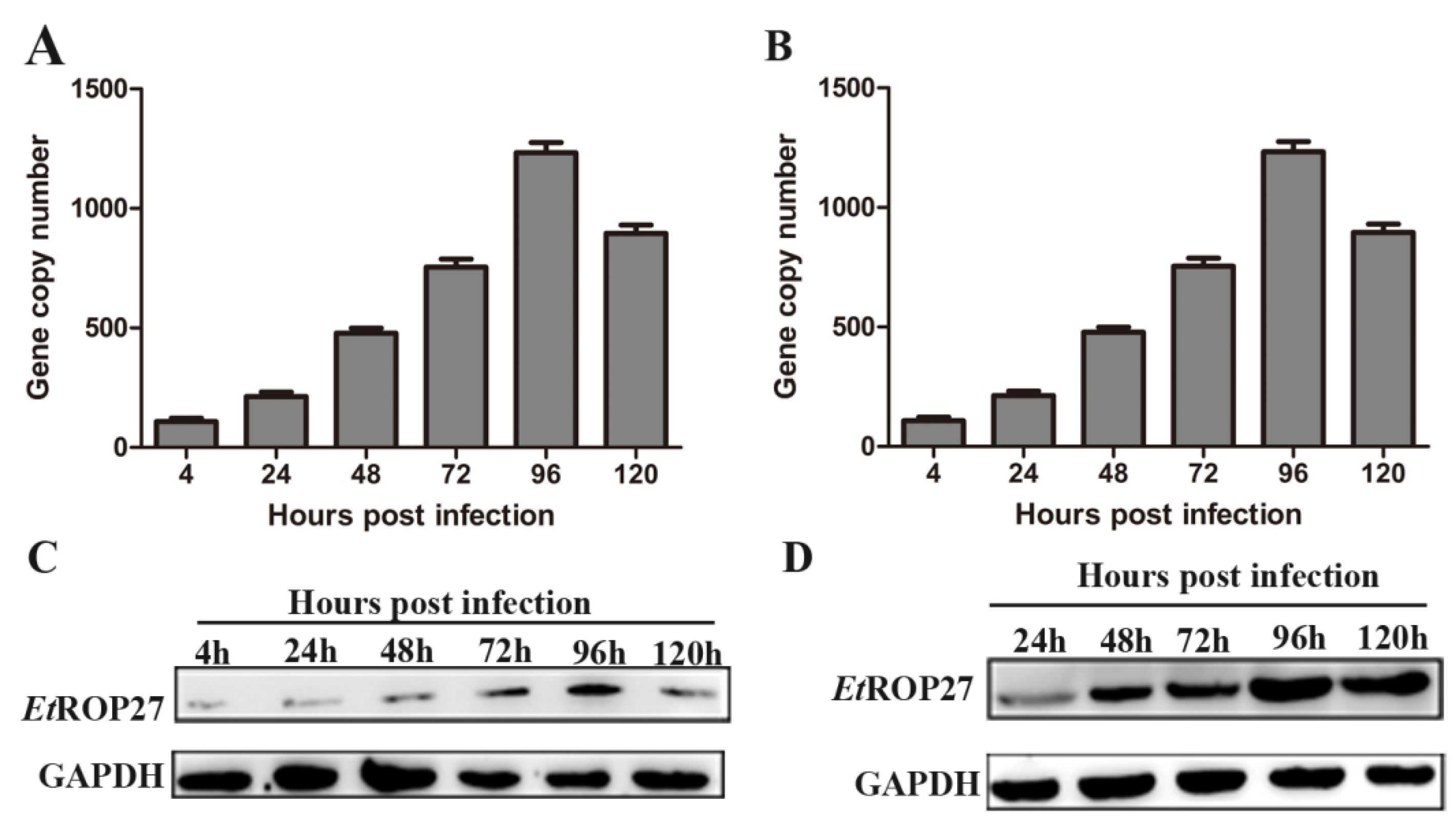
Figure 6.
Efects of recombinant EtROP27 protein against E. tenella challenge. (A)Body weight gain. (B) Relative body weight gain. (C) OPG. (D) Oocyst reduction ratio. (E) bloody stool. The red box indicates bloody stool. (F) Lesion scores. Red arrow : red cecal core; black arrow : bleeding point. (G) IgY concentration. All of the data are representative of at least three independent experiments. The same column of shoulder markers with the same lowercase letters indicate no significant difference (P>0.05), while shoulder markers with different lowercase letters indicate significant differences (P<0.05).
Figure 6.
Efects of recombinant EtROP27 protein against E. tenella challenge. (A)Body weight gain. (B) Relative body weight gain. (C) OPG. (D) Oocyst reduction ratio. (E) bloody stool. The red box indicates bloody stool. (F) Lesion scores. Red arrow : red cecal core; black arrow : bleeding point. (G) IgY concentration. All of the data are representative of at least three independent experiments. The same column of shoulder markers with the same lowercase letters indicate no significant difference (P>0.05), while shoulder markers with different lowercase letters indicate significant differences (P<0.05).


Table 1.
Titer determination of EtROP27 polyclonal antibody (OD450).
| Antibody dilution ratio | Negative serum | ||||||||
| 1:100 | 1:200 | 1:400 | 1:800 | 1:1600 | 1:3200 | 1:6400 | 1:12800 | 1:25600 | |
| * | * | * | 3.442 | 2.532 | 2.015 | 1.503 | 1.022 | 0.516 | 0.36 |
Note: * indicates that the OD450 value exceeds 3.5.
Table 2.
Experimental chicken ACI value.
| Groups | Relative body weight gain (%) | Oocysts index | Cecum lesion scores | Anti-coccidial index |
| Unchallenged control | 100 | 0 | 0 | 200.00 |
| Challenged control | 60.25 | 5 | 29 | 87.25 |
| Et ROP27(50μg) | 80.45 | 1 | 20 | 159.45 |
| Et ROP27(100μg) | 89.47 | 1 | 17 | 171.47 |
| Et ROP27(150μg) | 86.75 | 1 | 19 | 166.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



