
Submitted:
05 December 2023
Posted:
06 December 2023
You are already at the latest version
Abstract
African swine fever virus (ASFV) is a large double stranded DNA arbovirus that is highly contagious and seriously endangers the lives of domestic and wild pigs. In the past decade, African swine fever (ASF) has spread in many countries in the Caucasus, Russian Federation, Eastern Europe and Asia, causing significant losses to the pig industry. At present, there is a lack of effective vaccine and treatment for ASF. Therefore, the rapid and accurate detection is crucial for ASF prevention and control. In this study, we have developed a portable lateral flow strip (LFS) detection method mediated by recombinase polymerase amplification (RPA) and DNA enzyme (LwCas13a), which is performed at 37 ℃ and visualized by eyes without the need for complex instruments. This RPA-LwCas13a-LFS is based on the ASFV structural protein p17 gene (D117L), with a detection sensitivity up to 2 gene copies. This method is highly specific and has no cross reactivity to 7 other pig viruses. In the detection of 33 clinical samples, the p17 (D117L) RPA-LwCas13a-LFS had 100% coincidence with conventional quantitative PCR (qPCR). These findings demonstrate the potential of this simple, rapid, sensitive, and specific ASFV detection method for on-site ASFV detection.
Keywords:
African swine fever virus (ASFV)
; LwCas13a
; recombinase polymerase amplification (RPA)
; lateral flow strip (LFS)
; visual detection
1. Introduction
African swine fever (ASF) is an acute, febrile and highly contagious infectious diseases caused by African swine fever virus (ASFV) infecting domestic pigs and wild boars [1,2]. The acute type ASF is characterized by high fever, depression, anorexia, skin cyanosis, splenomegaly and bleeding, with very high morbidity and mortality [3]. In recent years, ASFV has been widely spread, causing a huge impact on the global pig industry [4]. Currently, there is no effective vaccine for this disease, and neither is effective therapeutic to treat ASF [5]. Therefore, early monitoring, diagnosis, and biosafety measures are crucial for ASF prevention and control [6]. Many molecular and serological detection methods of ASFV have been developed, such as fluorescence quantitative PCR, conventional PCR, fluorescence antibody test, enzyme-linked immunosorbent assay (ELISA) [7,8]. However, these methods require expensive equipment, standardized laboratory and/or professional technical personnel to operate, resulting in long reaction time and difficult to meet the front-line requirements of pig farms. Further, early monitoring cannot be achieved for cases with extremely low virus content [7,8]. How to detect ASFV more quickly, simply, accurately and cheaply is the future direction for ASFV detection development [7].
ASFV encodes more than 150 proteins, and p17 is a conserved protein necessary for the formation of icosahedron from the viral precursor membrane, which is crucial to the viral vitality [9]. When the expression of p17 is inhibited, the hydrolytic processing of polyprotein pp220 and pp62 is blocked, the mature core shell containing viral genome cannot be assembled and the formed icosahedron particles with defective core shell have impaired virus infectivity [9]. In the ASFV particles, p17, as an advanced transmembrane protein, is located in the viral capsid and inner lipid envelope, and is one of the main structural proteins of ASFV [10]. Confirmed by indirect ELISA experiment, p17 has good immunogenicity and can induce the production of specific antibodies during ASFV infection [11]. The viral protein p17 is encoded by the conservative D117L gene, which can be used as target site for nucleic acid detection [9,12].
The CRISPR/LwCas13a system, via guide RNA (gRNA), recognizes the RNA of the protospacer flanking site (PFS) sequence (composed of A, U, and C at the 3' end), demonstrating efficient degradation ability towards the target RNA [13,14]. LwCas13a protein consists of a CRISPR RNA (crRNA) recognition leaf (REC) and nuclease leaf (NUC) [15]. This double leaf structure enables LwCas13a to have two kinds of RNA enzyme activities [15,16]. The Helical-1 domain of LwCas13a protein is responsible for cleaving pre-crRNA to form mature crRNA [16]. On the other hand, the HEPN domain of LwCas13a protein is responsible for specific cis cleavage of targeted RNA, followed by non-specific trans cleavage of other nearby RNAs [16]. LwCas13a recognizes the pre spacer sequence, PFS, which is equivalent to the protospacer adjacent motif (PAM) sequence of DNA recognized by Cas9 [17,18]. When the LwCas13a crRNA complex recognizes PFS sequence, the crRNA guiding sequence undergoes base complementary pairing with the target RNA, while inducing a synergistic conformational change of LwCas13a, resulting in HEPN-RNase activity of the LwCas13a crRNA complex [13,14,16]. The RNA guided RNA endonuclease LwCas13a was additionally used to exert non-specific cleavage activity and degrade single stranded RNA probes for detection [19]. In addition, CRISPR/LwCas13a was combined with recombinant polymerase chain reaction (RPA) and/or lateral flow test strip (LFS) to create a fast, accurate, sensitive, and specific visual detection platform [20,21,22].
At present, only three literatures have reported the detection of ASFV based on the CRISPR/LwCas13a system, with all the detection target genes being the major capsid protein p72 gene B646L [20,22,23]. In this study, we developed CRISPR/Cas13a method and utilized p17 gene (D117L) as the detection target to detect ASFV. Further, our study designed and optimized a specific combination of RPA primers, crRNA, and probe for p17 gene D117L detection by RPA-LwCas13a-LFS. Our results demonstrated that this p17/D117L-RPA-LwCas13a-LFS method can be effectively applied for clinical ASFV detection, thus providing an important tool for timely monitoring and prevention of ASF.
2. Materials and Methods
2.1. Reagents and specimens
The virus DNA extraction kit Hipure Tissue DNA Mini Kit (D3121-02) and Hipure Blood DNA Mini Kit (D3111-02) were purchased from Magen Biotechnology Co., Ltd (Guangzhou, China). CRISPR/LwCas13a protein (C005M) was from Meige Biotechnology Co., Ltd (Guangzhou, China). Rosseta-2 (DE3/BL21) competent E.coli was from Tiangen BioTech Co., Ltd (Beijing China). The FastPure Gel DNA Extraction Mini Kit (DC301-01) and T7 High Yield RNA Transcription Kit (TR101-01) were both from Nanjing Vazyme Biotech Co., Ltd (Nanjing, China). RNA Clean & Concentrator-5 (R1013) was from Zymo Corporation (Irvine, CA, USA). The recombinant polymerase amplification (RPA) kit (TwistAmp Basic, Cat #TABAS03KIT) was from TwistDx Limited (Maidenhead, UK). LwCas13a test strip (M20801-F007) was from Bio-Lifesci Company (Guangzhou, China). The other conventional reagents and p17 expression plasmid pCold-I-D117L were all stored in our laboratory. The classical swine fever virus (CSFV) attenuated strain (CVCC AV1412), porcine pseudorabies virus attenuated strain (Bartha-K61) were from Jiangsu Nannong Hi Tech Co., Ltd (Nanjing China). The inactivated porcine parvovirus (S-1 strain) was purchased from Shangdong Huahong Biological Engineering Co., Ltd (Binzhou, China). In this study, the nucleic acids were extracted from 3 hearts, 3 livers, 3 spleens, 3 lungs, 3 kidneys, 3 small intestines, 3 lymph nodes, 2 oral swabs, 5 blood samples, and 5 serum samples of pigs (Guangzhou Aisu Testing Technology Research Institute Henan Branch) by using High Tissue DNA Mini Kit (D3121-02) and High Blood DNA Mini Kit (D3111-02) according to the instructions.
2.2. Design and preparation of D117L gene crRNA
Three crRNAs targeting the p17 gene D117L were designed based on the special PFS sequence identified by LwCas13a (consisting of A, U, and C at the 3 'end). Each crRNA sequences (5’-3’) consist of a T7 promoter (GAAATTAATACGATACTATATATAGG) + LwCas13a anchoring sequence (GATTTAGACTACCCAAAAAACGAAGGGACTAAAAAC) + target sequence (18-28 bp after the PFS sequence, with the 3 'end as A, U, and C). The synthesized complementary encoding DNAs (Table S1, Qingke Biotechnology Co., Ltd.) were annealed, gel purified, and the corresponding specific crRNAs were obtained through in vitro transcription (IVT) followed by RNA purification using ZYMO RNA Clean&Concentrator-5.
2.3. Preparation of D117L gene derived target RNA (T-RNA)
Based on LwCas13a specific PFS sequences (consisting of A, U, and C at the 3 'end) in D117L gene sequence, the PCR primers were designed (Table S1) to amplify the target double stranded DNA from the ASFV-p17 plasmid pCold-I-D117L constructed in our laboratory. Subsequently, the CRISRP-Cas13a target RNA (T-RNA) were obtained from the PCR products by in vitro transcription (IVT) via the T7 promoter (TAATACGACCACTACTAGAGG) added to the 5 'end of the forward PCR primer.
2.4. Design of single stranded RNA (ssRNA) probe for CRISPR/LwCas13a reaction
The LwCas13a reporter ssRNA probe was designed as 5'-6-FAM-rUrUrUrUrArUrU-BHQ-3' (synthesized by Shanghai Shenggong Biotechnology Co., Ltd.). The 5' and 3' ends of LwCas13a probe are conjugated with the fluorescent group 6-FAM and the quenching group BHQ-1, respectively. The synthetic probe was centrifuged at 5000 rpm 4 ℃ for 5 minutes, diluted to 100 µM with sterile enzyme free water, and stored at -20 ℃ for experimental use.
2.5. CRISPR/LwCas13a reaction and its detection
LwCas13a and crRNA mediate CRISPR response, recognize the transcribed T-RNA, cleaving T-RNA as well as probe RNA to generate signaling (Figure 1). Specifically, the LwCas13a protein was mixed with crRNA, target T-RNA, and RNA probe in a sterile enzyme free PCR tube in a detection buffer (20mM Tris HCl, 100 mM KCl, 6 mM MgCl2, 1 mM DTT, 5% glycerin, 50g /mL heparin medium, pH 8.0), and subjected to reaction at 37 ℃ for 30 minutes. The amounts of reaction components of CRISPR/LwCas13a are as follows: 1 µL LwCas13a (40 nM), 1 µL crRNA (250 ng/µL), 1 µL T-RNA (100 ng/µL), 1 µL ssRNA probe (40 nM), and 46 µL buffer, totaling 50 µL. Blue light (wavelength 450 nm), ultraviolet light (wavelength 320 nm) and fluorescence signal (excitation wavelength 492 nm, emission wavelength 521 nm) were used to detect the activity of LwCas13a RNA endonuclease (Figure 1). Additionally, the cleavage of T-RNA by CRISPR/LwCas13a without RNA probes can be further verified by SDS-PAGE together with silver staining.
2.6. Recombinase polymerase amplification (RPA)
According to the design principle of RPA primers: the primer length is between 28 - 35 bp, and the optimal amplification fragment size is 150-200 bp. Two pairs of RPA primers for D117L gene were designed, with the 5 'end of each forward primer appended with the T7 promoter sequence (GAAATTAATACGACTACATAGG) (Table S1). According to the TwistAmp Basic Kit (TwistDx), each RPA reaction contains 29.5 μL buffer, reaction microspheres, 0.48 μM forward and reverse primers, appropriate amount of purified DNA templates, 2.5 μL magnesium acetate (MgAc) and up to 12.2 μL sterile water, approximately 50 μL in total. RPA reaction was run in a conventional water bath at 39 ℃ for 15-20 min, and the amplified D117L gene DNA was examined by gel electrophoresis. Two pairs of RPA primers were evaluated and selected based on better performance. The final p17 RPA product was subjected for T7 in vitro transcription to produce target RNA for subsequent detection of LwCas13a mediated lateral flow strip (LFS).
2.7. RPA-CRISPR/LwCas13a Lateral Flow Strip (LFS) detection method
Synthetic biotin labeled ssRNA probe 5'-6-FAM-UUUUUUUUAUUUUUU-Biotin-3' was synthesized by Shanghai Shenggong Biotechnology Co., Ltd. One microliter (1 μL) RPA derived target RNA was added into the CRISPR/LwCas13a reaction system (1 μL LwCas13a, 1 μ L D117L-crRNA1, 1 μL 0.2 nM biotin ssRNA probe and 46 μL detection buffer, a total of 50 μL), for a moment at room temperature to allow the CRISPR/LwCas13a reaction. Subsequently, the binding pad of the test strip with arrow end was inserted into the CRISPR reaction solution for 1-2 minutes, the binding pad area will be completely soaked in the reaction solution. The FAM antibody cross-linked gold particles on the test strip bind to the ssRNA probe in the reaction solution, forming a 5'-gold particle-anti-FAM-6-FAM-ssRNA-biotin-3' detection probe. As the liquid diffuses, streptavidin on the quality control (C) line captures biotin labeled gold particle probes, presenting a gold line. Finally, the test strip was taken out and test results were read directly based on whether there is gold wire on the test strip (T) line (Figure 1).
2.8. Sensitivity and specificity of the D117L gene RPA-LwCas13a-LFS detection method
The plasmid pCold-I-D117L was 10 fold serially diluted where the plasmid concentration was from 2×1011 copies/µL to 2×100 copies/µL. Plasmids with different dilutions were subjected to RPA amplification, and the amplified products were subjected to in vitro transcription (IVT), followed by CRISPR/LwCas13a-LFS detection. The LFS assay sensitivity was determined by the minimum copy number of plasmid DNA in the positive results.
Virus RNA or DNA was extracted from 7 pig virus samples. Specifically, RNA was from porcine reproductive and respiratory syndrome virus (PRRSV), porcine epidemic diarrhea virus (PEDV), classical swine fever virus (CSFV) and swine influenza virus (SIV), respectively. DNA was from porcine parvovirus (PPV), porcine pseudorabies virus (PRV), porcine circovirus type 3 (PCV3), respectively. Both viral RNA and DNA were used as viral target RNA for the CRISPR/LwCas13a reaction mediated by D117L-crRNA1, followed by LFS detection to determine the reaction specificity of D117L-LwCas13a-LFS detection.
3. Results
3.1. CRISPR/LwCas13a reaction mediated by ASFV structural gene D117L crRNA and its optimization
LwCas13a is an RNA guided RNase and T7 transcription mediated Cas13a assay has been proven to be successful for DNA detection [24]. We used this CRISPR/Cas13a system with RNA cleavage characteristic to develop a molecular testing method successfully. As shown in Figure 2A-B, all three ASFV D117L crRNAs (crRNA1, crRNA2, and crRNA3) can mediate LwCas13a cleavage of the target RNA of the D117L gene. The RNA probe cleaved by CRISPR/LwCas13a generates consistent light signals under blue light, ultraviolet light, or fluorescence activation (Figure 2C-D).
3.2. Probe concentration optimization
We optimized the probe concentration by measuring the blue light, ultraviolet light, and fluorescence signals generated by LwCas13a cleaved ssRNA probe in the CRISPR reaction. Starting from 10 nM, as the probe concentration increases, the intensities of blue light, ultraviolet light, and fluorescence signals continue to increase (Figure S1). Considering the signal-to-noise balance, a probe concentration of 40-80 nM is appropriate (Figure S1).
3.3. Optimization of concentration ratios between LwCas13a protein and crRNA1
The initial concentrations of LwCas13a protein and crRNA1 were set to 40 nM and 250 ng/μL, respectively. The CRISPR/LwCas13a reaction was performed at different concentrations of LwCas13a : crRNA1 (8:1, 4:1, 2:1, 1:1, 1:2, 1:4, and 1:8). Different LwCas13a/crRNA1 ratios led to variation of LwCas13a activity. At the LwCas13a/crRNA1 ratio of 2:1, LwCas13a exhibited higher blue, ultraviolet, and fluorescence signals than other ratios (Figure S2).
3.4. Detection sensitivity of CRISPR/LwCas13a reaction
The plasmid pCold-I-D117L was 10 fold serially diluted where the plasmid concentration was from 2×1011 copies/µL to 2×100 copies/µL. The D117L gene was directly subjected for in vitro transcription, and the transcribed RNA was used as the detection target to determine the sensitivity of CRISPR/LwCas13a reaction system. The results indicated that the detection limit is 2 × 1010 copies/µL (Figure 3A-B).
3.5. Establishment of CRISPR/LwCas13a mediated lateral flow test strip (LFS) method
The FAM antibody cross-linked gold particles on the test strip react with the RNA probe to form a gold particle RNA probe (5'-gold particles-anti-FAM-6-FAM-ssRNA-Biotin-3'). If the sample to be tested contains the ASFV target gene, the CRISPR/LwCas13a system will cut off the ssRNA probe and separate the FAM gold particles from biotin. Thus, the FAM gold particles are able to cross the streptavidin coated C-line, continue to migrate forward, and will be captured by secondary antibody on the test line (T), presenting a positive gold line (Figure 1). On the contrary, the ssRNA report probe in the negative sample reaction is not cleaved and will be completely captured by C-line streptavidin (Figure 1).
Our experimental results demonstrated that the gold particle signals appeared on the T-line of the lateral flow strip only when LwCas13a, crRNA, target RNA, and gold particle probes are all present (Figure 4A). The detection results of LwCas13a-LFS were completely consistent with those of blue light, ultraviolet light, and fluorescence signals. (Figure 4B-C). Further, we optimized the concentrations of biotin probes (0.1-1.2 nM) and the results showed that the 0.1 nM probe could effectively display positive gold particle bands on the T line of the colloidal gold test strip (Figure 4D).
3.6. Sensitivity of RPA-CRISPR/LwCas13a-LFS method
In order to improve the sensitivity of the CRISPR/LwCas13a-LFS detection method, we designed two pairs of RPA primers (Table S1) to amplify the ASFV structural gene D117L before in vitro transcription. Based on the specificity of RPA amplified products, the optimal RPA primers would be selected for subsequent experiments, and our results suggested that both RPA primers exhibited comparable performance in the D117L gene amplification (not shown).
Next, the gel recovered PCR products of D117L gene were 10 fold serially diluted, with the calculated molecular copy numbers ranging from 2×1011 copies/µL to 2×100 copies/µL. The above D117L gene products were subjected for RPA amplification (Figure 5A) and subsequent T7 in vitro transcription (Figure 5A). The transcribed RNAs were then used for CRISPR/LwCas13a-LFS detection. The results showed that, with the pre-amplification by RPA, the detection limits of CRISPR/LwCas13a reaction and CRISPR/LwCas13a-LFS method could both reached to 2×100 copies/µL (Figure 5B-D).
3.7. Specificity of CRISPR/LwCas13a-LFS detection method
In order to verify the specificity of CRISPR/LwCas13a-LFS detection method, DNA or RNA samples of seven pig viruses other than ASFV were obtained, including the RNAs from porcine reproductive and respiratory syndrome virus (PRRSV), porcine epidemic diarrhea virus (PEDV), classical swine fever virus (CSFV), and swine influenza virus (SIV), and the DNAs from porcine parvovirus (PPV), porcine pseudorabies virus (PRV), and porcine circovirus type 3 (PCV3). The nucleic acid samples of seven pig viruses and ASFV were all confirmed by PCR or RT-PCR (Figure 6A) using specific PCR primers (Table S1). The D117L-LwCas13a-LFS detection results showed that the T lines of all 7 pig viruses were absent of gold particles whereas the T line of ASFV samples showed gold particle signals (Figure 6B). Similar results were obtained from CRISPR/LwCas13a reaction detection using blue light, ultraviolet, and fluorescence detection (Figure 6C-D). In summary, the CRISPR/LwCas13a-LFS method has very high specificity for detection of ASFV.
3.8. RPA-CRISPR/LwCas13a-LFS detection of clinical samples
Using the established RPA-LwCas13a-LFS method, we conducted preliminary detection of ASFV in 33 clinical samples. The results showed that out of 33 clinical samples, 12 were strongly positive (No.1-12), 6 were weakly positive (No.13-18), and 15 were negative (No.19-33) (Figure 7). In order to validate the reliability of the test strip for detecting ASFV, the gold standard quantitative PCR (qPCR) recommended by the World Organization for Animal Health (WOAH) was used for verification detection. The results showed that the RPA-LwCas13a-LFS detection results were completely consistent with the qPCR detection results (Table 1). This result further confirmed the reliability of the RPA-LwCas13a-LFS method for detecting clinical ASFV.
4. Discussion
This study provides a rapid, low-cost and visual CRISPR/LwCas13a-LFS nucleic acid test for ASFV detection by selecting the conserved structural gene D117L of ASFV as the detection target [9,12]. Currently, most of the CRISPR/Cas12 system reported in the literatures for detecting ASFV utilized target structural protein p72 gene B646L, except one report using ASFV pp220 gene or DNA Pol gene as detection targets [25]. Further, only three articles have reported the Cas13a system for detecting ASFV, which all targeted the p72/B646L gene [20,22,23]. We recently established CRISPR/LbCas12a detection and RPA-Cas12a-LFS detection of ASFV by targeting the structural gene D117L gene [26]. As a conserved ASFV structural gene, D117L not only has multiple PAM sequences (TTTN) recognized by LbCas12a, but also has several PFS sequences recognized by LwCas13a. Therefore, the designed crRNAs based on PFS in this study were able to mediated Cas13a reaction specifically and effectively (Figure 2).
Even though a single crRNA and a single targeted gene is sufficient for detection of ASFV, ASFV is subjected to high mutation in the field [1,2]. In order to avoid the missed detection due to the target ASFV gene mutation, multiple crRNAs targeting different ASFV genes (such as both p17 and p72 structural genes and others) could be considered for multiplex detection [23].
By combining with RPA pre-amplification, the D117L-CRISPR/LwCas13a reaction and the corresponding LFS detection were able to detect target gene as low as 2 copies/ μL (Figure 5). Importantly, the RPA-Cas13a-LFS was able to differentiate strongly positive and weakly positive ASF clinical samples (Figure 7). The positive detection results vary depending on the efficiency of crRNA mediated Cas13a cleavage. In strongly positive samples, the cutting efficiency of Cas13a is high, and the ssDNA probe is completely cut, leading to a high intense gold band at the T line, but no band at the C line. In weakly positive samples, the ssDNA probe is not completely cleaved by Cas13a, with only a faint gold band at T line and most gold band at C line. The D117L-CRISPR/LwCas13a-LFS detection results from 33 clinical samples including 12 strongly positive (Nos 1-12) and 6 weakly positive (Nos 13-18) samples were completely consistent with those by WOAH recommended gold standard TaqMan qPCR [27]., further confirming reliability of the LwCas13a-LFS detection (Figure 7).
In order to effectively monitor and manage virus transmission of ASFV, in addition to sensitivity and specificity, an ideal molecular detection method that meets the following standards is urgently needed [28,29]: (1) fast, providing diagnostic information before ASF occurs; (2) simple, allowing unskilled personnel to use; (3) field applicable, which is very important for on-site ASFV testing. Our D117L-CRISPR/LwCas13a-LFS detection method is sensitive, specific, fast and simple, but the field applicability has still room to be improved, where the nucleic acids extraction and RPA need to be performed separately. In the follow-up research, it is necessary to solve the problem by using nucleic acid extraction free reagents and converging RPA and CRISPR/Cas13 reaction to achieve "sample results" detection of ASFV nucleic acid [30].
In summary, the CRISPR/Cas13a bio-sensing system as an innovative technology has a significant impact on detection and diagnostic capabilities in many fields [31,32,33]. The RPA-CRISPR/Cas13a-LFS method for ASFV detection we developed is fast, simple, sensitive, specific and has potential for field application.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
J.Zhu and WZ conceived and designed the experiments; D.Z, S.J, N.X, J.Zhang, A.L, D.D, C.Z, performed the experiments; W.Z, N.C, X.K, Z.P, J.Zhu analyzed the data; D.Z, W.Z and J.Zhu wrote the paper. All authors contributed to the article and approved the submitted version.
Acknowledgments
The work was partly supported by partly supported by the National Key Research and Development Program of China (2021YFD1801201-2), Jiangsu agricultural science and technology independent innovation fund project [CX(21)2035], Jiangsu provincial key R & D plan (BE2020398), the 111 Project D18007, and A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD). W.Z was supported by the Open Project Program of Jiangsu Key Laboratory of Zoonosis (No. R2304).
Conflicts of Interest
The authors declare no any potential conflict of interest
References
- Li, Z.; Chen, W.; Qiu, Z.; Li, Y.; Fan, J.; Wu, K.; Li, X.; Zhao, M.; Ding, H.; Fan, S.; Chen, J., African Swine Fever Virus: A Review. Life (Basel) 2022, 12, (8). [CrossRef]
- Galindo, I.; Alonso, C., African Swine Fever Virus: A Review. Viruses 2017, 9, (5). [CrossRef]
- Wang, Y.; Kang, W.; Yang, W.; Zhang, J.; Li, D.; Zheng, H., Structure of African Swine Fever Virus and Associated Molecular Mechanisms Underlying Infection and Immunosuppression: A Review. Front Immunol 2021, 12, 715582. [CrossRef]
- Ata, E. B.; Li, Z. J.; Shi, C. W.; Yang, G. L.; Yang, W. T.; Wang, C. F., African swine fever virus: A raised global upsurge and a continuous threaten to pig husbandry. Microb Pathog 2022, 167, 105561. [CrossRef]
- Urbano, A. C.; Ferreira, F., African swine fever control and prevention: an update on vaccine development. Emerg Microbes Infect 2022, 11, (1), 2021-2033. [CrossRef]
- Denstedt, E.; Porco, A.; Hwang, J.; Nga, N. T. T.; Ngoc, P. T. B.; Chea, S.; Khammavong, K.; Milavong, P.; Sours, S.; Osbjer, K.; Tum, S.; Douangngeun, B.; Theppanya, W.; Van Long, N.; Thanh Phuong, N.; Tin Vinh Quang, L.; Van Hung, V.; Hoa, N. T.; Le Anh, D.; Fine, A.; Pruvot, M., Detection of African swine fever virus in free-ranging wild boar in Southeast Asia. Transbound Emerg Dis 2021, 68, (5), 2669-2675. [CrossRef]
- Oura, C. A.; Edwards, L.; Batten, C. A., Virological diagnosis of African swine fever--comparative study of available tests. Virus Res 2013, 173, (1), 150-8. [CrossRef]
- Qiu, Z.; Li, Z.; Yan, Q.; Li, Y.; Xiong, W.; Wu, K.; Li, X.; Fan, S.; Zhao, M.; Ding, H.; Chen, J., Development of Diagnostic Tests Provides Technical Support for the Control of African Swine Fever. Vaccines (Basel) 2021, 9, (4). [CrossRef]
- Suarez, C.; Gutierrez-Berzal, J.; Andres, G.; Salas, M. L.; Rodriguez, J. M., African swine fever virus protein p17 is essential for the progression of viral membrane precursors toward icosahedral intermediates. J Virol 2010, 84, (15), 7484-99. [CrossRef]
- Wang, N.; Zhao, D.; Wang, J.; Zhang, Y.; Wang, M.; Gao, Y.; Li, F.; Wang, J.; Bu, Z.; Rao, Z.; Wang, X., Architecture of African swine fever virus and implications for viral assembly. Science 2019, 366, (6465), 640-644. [CrossRef]
- Li, L.; Qiao, S.; Li, G.; Tong, W.; Dong, S.; Liu, J.; Guo, Z.; Zheng, H.; Zhao, R.; Tong, G.; Zhou, Y.; Gao, F., The Indirect ELISA and Monoclonal Antibody against African Swine Fever Virus p17 Revealed Efficient Detection and Application Prospects. Viruses 2022, 15, (1). [CrossRef]
- Xia, N.; Wang, H.; Liu, X.; Shao, Q.; Ao, D.; Xu, Y.; Jiang, S.; Luo, J.; Zhang, J.; Chen, N.; Meurens, F.; Zheng, W.; Zhu, J., African Swine Fever Virus Structural Protein p17 Inhibits Cell Proliferation through ER Stress-ROS Mediated Cell Cycle Arrest. Viruses 2020, 13, (1). [CrossRef]
- Cox, D. B. T.; Gootenberg, J. S.; Abudayyeh, O. O.; Franklin, B.; Kellner, M. J.; Joung, J.; Zhang, F., RNA editing with CRISPR-Cas13. Science 2017, 358, (6366), 1019-1027. [CrossRef]
- Liu, L.; Pei, D. S., Insights Gained from RNA Editing Targeted by the CRISPR-Cas13 Family. Int J Mol Sci 2022, 23, (19). [CrossRef]
- Liu, L.; Li, X.; Wang, J.; Wang, M.; Chen, P.; Yin, M.; Li, J.; Sheng, G.; Wang, Y., Two Distant Catalytic Sites Are Responsible for C2c2 RNase Activities. Cell 2017, 168, (1-2), 121-134 e12. [CrossRef]
- Knott, G. J.; East-Seletsky, A.; Cofsky, J. C.; Holton, J. M.; Charles, E.; O'Connell, M. R.; Doudna, J. A., Guide-bound structures of an RNA-targeting A-cleaving CRISPR-Cas13a enzyme. Nat Struct Mol Biol 2017, 24, (10), 825-833. [CrossRef]
- Kleinstiver, B. P.; Prew, M. S.; Tsai, S. Q.; Topkar, V. V.; Nguyen, N. T.; Zheng, Z.; Gonzales, A. P.; Li, Z.; Peterson, R. T.; Yeh, J. R.; Aryee, M. J.; Joung, J. K., Engineered CRISPR-Cas9 nucleases with altered PAM specificities. Nature 2015, 523, (7561), 481-5. [CrossRef]
- Jiang, F.; Doudna, J. A., CRISPR-Cas9 Structures and Mechanisms. Annu Rev Biophys 2017, 46, 505-529. [CrossRef]
- Liu, L.; Li, X.; Ma, J.; Li, Z.; You, L.; Wang, J.; Wang, M.; Zhang, X.; Wang, Y., The Molecular Architecture for RNA-Guided RNA Cleavage by Cas13a. Cell 2017, 170, (4), 714-726 e10. [CrossRef]
- Hu, F.; Liu, Y.; Zhao, S.; Zhang, Z.; Li, X.; Peng, N.; Jiang, Z., A one-pot CRISPR/Cas13a-based contamination-free biosensor for low-cost and rapid nucleic acid diagnostics. Biosens Bioelectron 2022, 202, 113994. [CrossRef]
- Liu, H.; Chang, S.; Chen, S.; Du, Y.; Wang, H.; Wang, C.; Xiang, Y.; Wang, Q.; Li, Z.; Wang, S.; Qiu, S.; Song, H., Highly sensitive and rapid detection of SARS-CoV-2 via a portable CRISPR-Cas13a-based lateral flow assay. J Med Virol 2022, 94, (12), 5858-5866. [CrossRef]
- Wei, N.; Zheng, B.; Niu, J.; Chen, T.; Ye, J.; Si, Y.; Cao, S., Rapid Detection of Genotype II African Swine Fever Virus Using CRISPR Cas13a-Based Lateral Flow Strip. Viruses 2022, 14, (2). [CrossRef]
- Tian, T.; Qiu, Z.; Jiang, Y.; Zhu, D.; Zhou, X., Exploiting the orthogonal CRISPR-Cas12a/Cas13a trans-cleavage for dual-gene virus detection using a handheld device. Biosens Bioelectron 2022, 196, 113701. [CrossRef]
- Khan, H.; Khan, A.; Liu, Y.; Wang, S.; Bibi, S.; Xu, H.; Liu, Y.; Durrani, S.; Jin, L.; He, N.; Xiong, T., CRISPR-Cas13a mediated nanosystem for attomolar detection of canine parvovirus type 2. Chin Chem Lett 2019, 30, (12), 2201-2204. [CrossRef]
- Lu, S.; Li, F.; Chen, Q.; Wu, J.; Duan, J.; Lei, X.; Zhang, Y.; Zhao, D.; Bu, Z.; Yin, H., Rapid detection of African swine fever virus using Cas12a-based portable paper diagnostics. Cell Discov 2020, 6, 18. [CrossRef]
- Zhang, D.; Jiang, S.; Xia, N.; Zhang, Y.; Zhang, J.; Liu, A.; Zhang, C.; Chen, N.; Meurens, F.; Zheng, W.; Zhu, J., Rapid Visual Detection of African Swine Fever Virus with a CRISPR/Cas12a Lateral Flow Strip Based on Structural Protein Gene D117L. Animals 2023, 13, (23), 3712. [CrossRef]
- King, D. P.; Reid, S. M.; Hutchings, G. H.; Grierson, S. S.; Wilkinson, P. J.; Dixon, L. K.; Bastos, A. D.; Drew, T. W., Development of a TaqMan PCR assay with internal amplification control for the detection of African swine fever virus. J Virol Methods 2003, 107, (1), 53-61. [CrossRef]
- Lu, S.; Tong, X.; Han, Y.; Zhang, K.; Zhang, Y.; Chen, Q.; Duan, J.; Lei, X.; Huang, M.; Qiu, Y.; Zhang, D. Y.; Zhou, X.; Zhang, Y.; Yin, H., Fast and sensitive detection of SARS-CoV-2 RNA using suboptimal protospacer adjacent motifs for Cas12a. Nat Biomed Eng 2022, 6, (3), 286-297. [CrossRef]
- Liu, S.; Tao, D.; Liao, Y.; Yang, Y.; Sun, S.; Zhao, Y.; Yang, P.; Tang, Y.; Chen, B.; Liu, Y.; Xie, S.; Tang, Z., Highly Sensitive CRISPR/Cas12a-Based Fluorescence Detection of Porcine Reproductive and Respiratory Syndrome Virus. ACS Synth Biol 2021, 10, (10), 2499-2507. [CrossRef]
- Samanta, D.; Ebrahimi, S. B.; Ramani, N.; Mirkin, C. A., Enhancing CRISPR-Cas-Mediated Detection of Nucleic Acid and Non-nucleic Acid Targets Using Enzyme-Labeled Reporters. J Am Chem Soc 2022, 144, (36), 16310-16315. [CrossRef]
- Li, Y.; Li, S.; Wang, J.; Liu, G., CRISPR/Cas Systems towards Next-Generation Biosensing. Trends Biotechnol 2019, 37, (7), 730-743. [CrossRef]
- Phan, Q. A.; Truong, L. B.; Medina-Cruz, D.; Dincer, C.; Mostafavi, E., CRISPR/Cas-powered nanobiosensors for diagnostics. Biosens Bioelectron 2022, 197, 113732. [CrossRef]
- van Dongen, J. E.; Berendsen, J. T. W.; Steenbergen, R. D. M.; Wolthuis, R. M. F.; Eijkel, J. C. T.; Segerink, L. I., Point-of-care CRISPR/Cas nucleic acid detection: Recent advances, challenges and opportunities. Biosens Bioelectron 2020, 166, 112445. [CrossRef]
Figure 1.
Schematic diagram of RPA-CRISPR/LwCas13a reaction and its different detection methods.
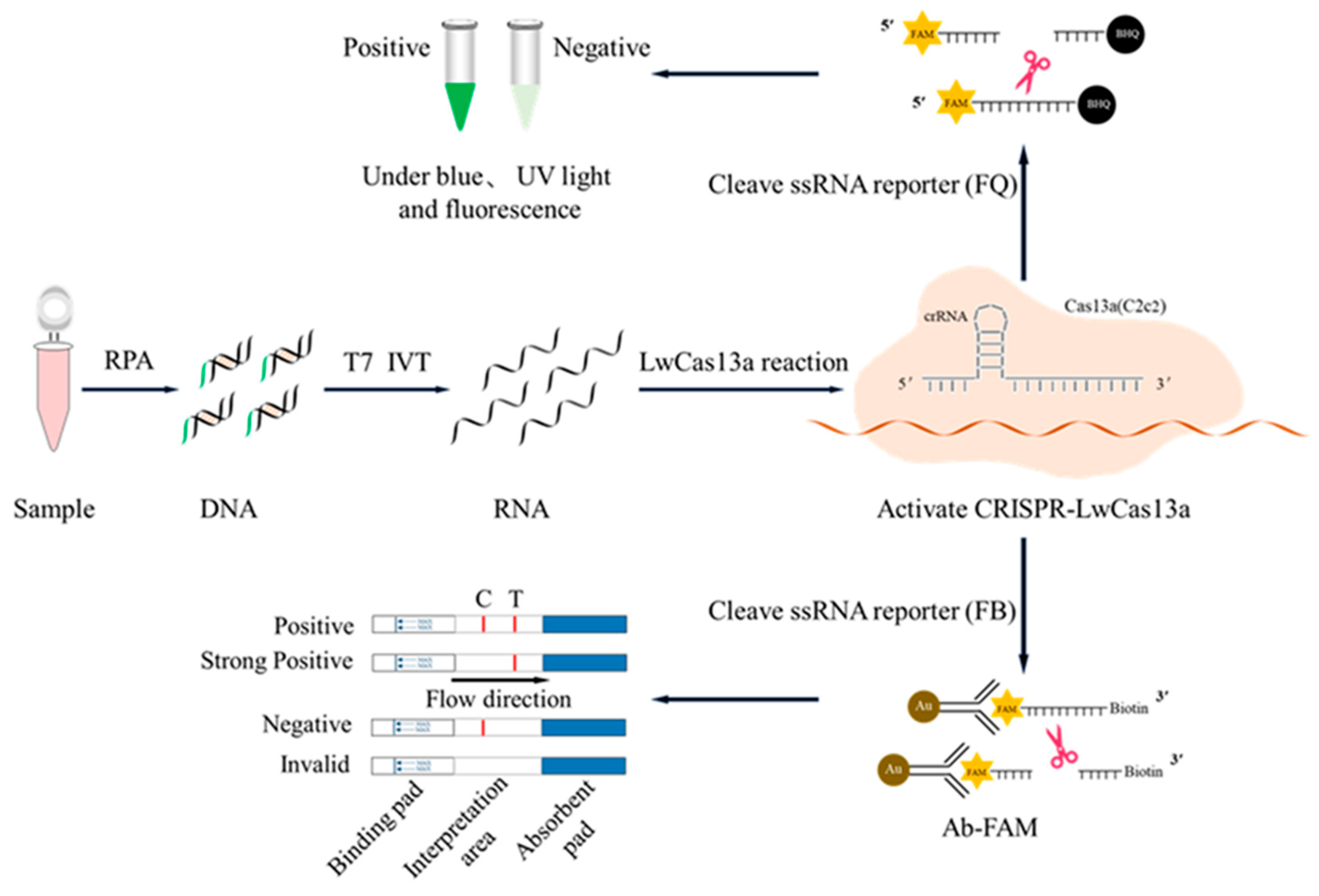
Figure 2.
Establishment of CRISPR/LwCas13a reaction system. A, Three D117L gene crRNAs and the target T-RNA of D117L gene from vitro transcription (IVT) were verified by agarose gel electrophoresis. B, Three crRNA mediated cleavage of D117L T-RNA by guide LwCas13a was validated through SDS-PAGE plus silver staining. C, The blue and ultraviolet light signals from the three crRNA mediated CRISPR/LwCas13a reaction. D, The fluorescence signal intensity was measured using microplate reader, and the strongest fluorescence signal of LwCas13a cleavage mediated by crRNA1. ** P<0.01, target RNA sample (PC) versus no target RNA sample (NC) (n=3). The results in the figure are representative of three similarly repeated experiments.
Figure 2.
Establishment of CRISPR/LwCas13a reaction system. A, Three D117L gene crRNAs and the target T-RNA of D117L gene from vitro transcription (IVT) were verified by agarose gel electrophoresis. B, Three crRNA mediated cleavage of D117L T-RNA by guide LwCas13a was validated through SDS-PAGE plus silver staining. C, The blue and ultraviolet light signals from the three crRNA mediated CRISPR/LwCas13a reaction. D, The fluorescence signal intensity was measured using microplate reader, and the strongest fluorescence signal of LwCas13a cleavage mediated by crRNA1. ** P<0.01, target RNA sample (PC) versus no target RNA sample (NC) (n=3). The results in the figure are representative of three similarly repeated experiments.
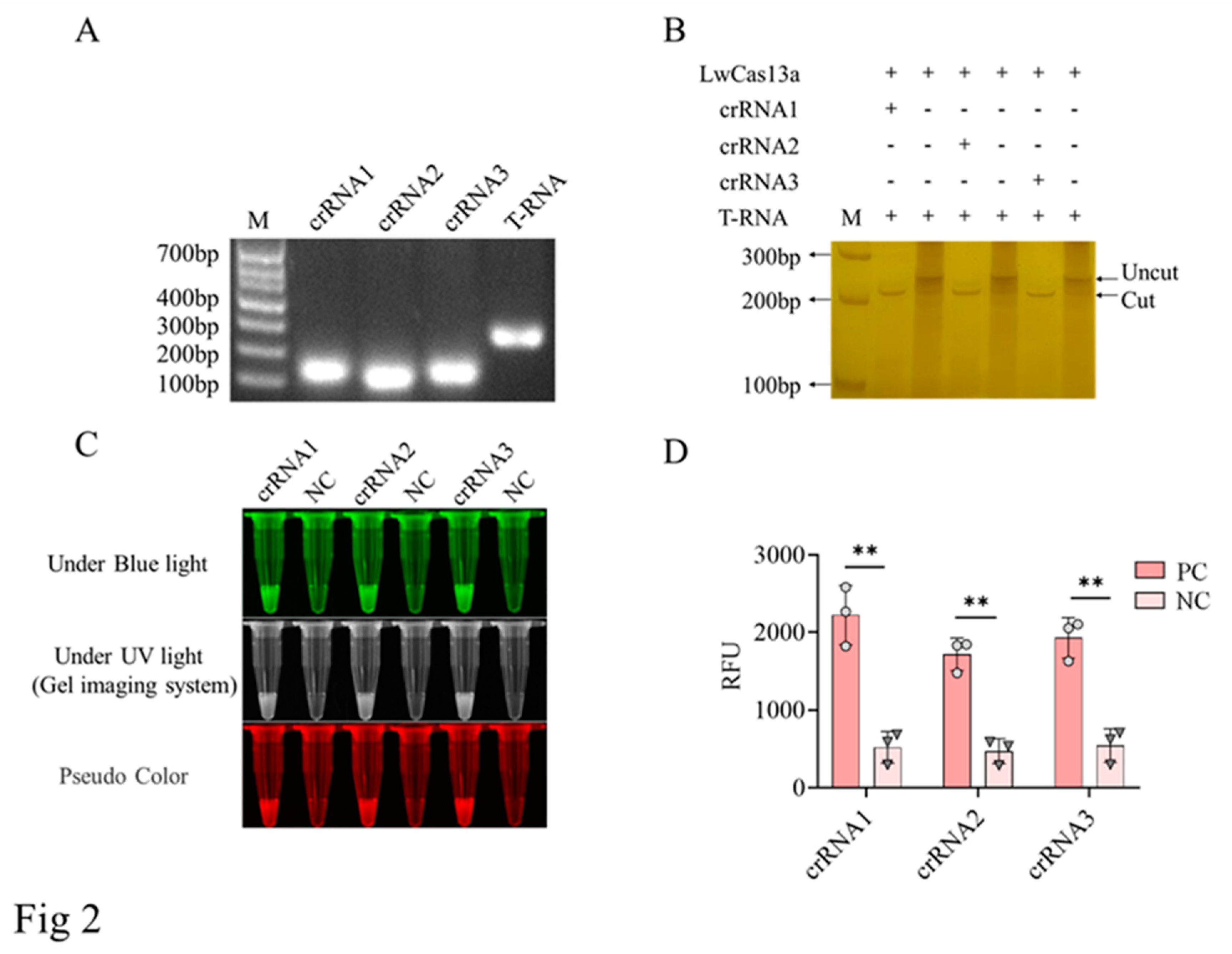
Figure 3.
The sensitivity of CRISPR/LwCas13a reaction to detect the D117L gene. A, Blue and ultraviolet light signals detected based on CRISPR/LwCas13a reaction. B, The fluorescence signal intensity by using Microplate reader. * P<0.05 versus no target RNA control (NC) (n=3).
Figure 3.
The sensitivity of CRISPR/LwCas13a reaction to detect the D117L gene. A, Blue and ultraviolet light signals detected based on CRISPR/LwCas13a reaction. B, The fluorescence signal intensity by using Microplate reader. * P<0.05 versus no target RNA control (NC) (n=3).
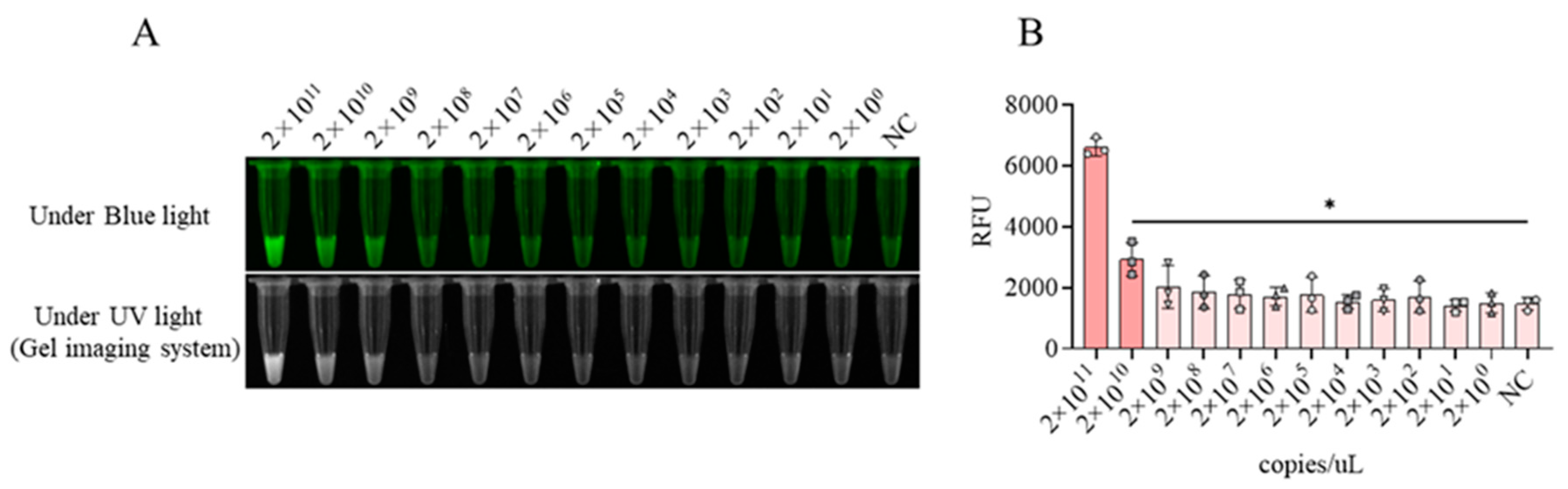
Figure 4.
Establishment of CRISPR/LwCas13a-LFS method. A, The assembly of CRISPR/LwCas13a-LFS reaction system. 1, with LwCas13a, crRNA1, T-RNA, and gold particle RNA probes all present; 2, lack of crRNA1; 3, lack of LwCas13a; 4, lack of LwCas13a and crRNA1; 5, lack of LwCas13a, crRNA1 and target RNA; NC, lack of target RNA. B, The blue and ultraviolet light signals mediated by crRNA1 from the CRISPR/LwCas13a reaction. C, The fluorescence signal intensity mediated by crRNA1 from the CRISPR/LwCas13a reaction. **P<0.01 versus no target RNA control (NC) (n=3). D, The CRISPR/LwCas13a-LFS reaction with different concentrations of biotin RNA probe. NC is a negative control without target RNA, and ns is not statistically significant.
Figure 4.
Establishment of CRISPR/LwCas13a-LFS method. A, The assembly of CRISPR/LwCas13a-LFS reaction system. 1, with LwCas13a, crRNA1, T-RNA, and gold particle RNA probes all present; 2, lack of crRNA1; 3, lack of LwCas13a; 4, lack of LwCas13a and crRNA1; 5, lack of LwCas13a, crRNA1 and target RNA; NC, lack of target RNA. B, The blue and ultraviolet light signals mediated by crRNA1 from the CRISPR/LwCas13a reaction. C, The fluorescence signal intensity mediated by crRNA1 from the CRISPR/LwCas13a reaction. **P<0.01 versus no target RNA control (NC) (n=3). D, The CRISPR/LwCas13a-LFS reaction with different concentrations of biotin RNA probe. NC is a negative control without target RNA, and ns is not statistically significant.
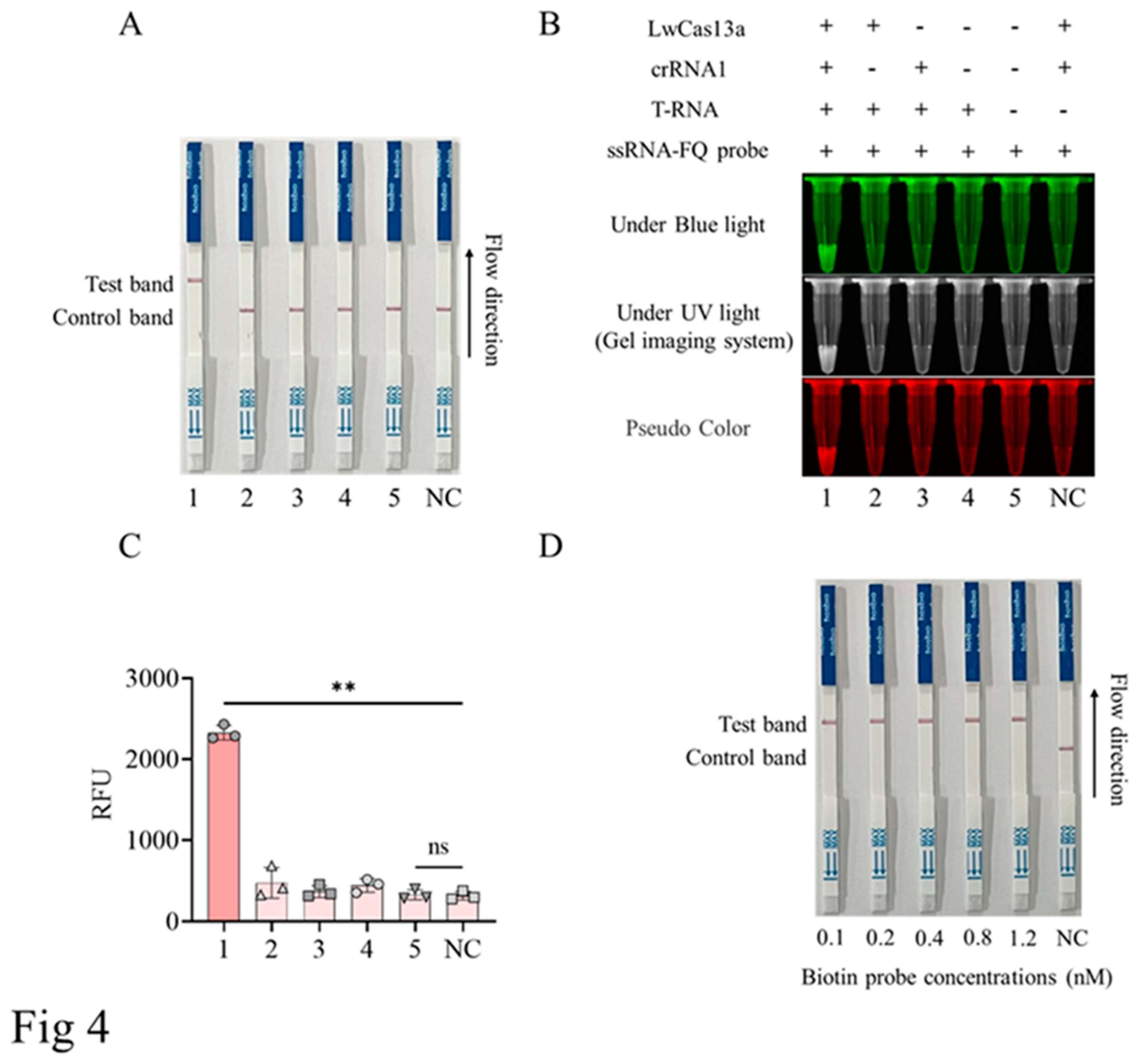
Figure 5.
Sensitivity of RPA-CRISPR/LwCas13a-LFS for detecting D117L gene. A, The RPA products of D117L target gene of different concentrations (2×1011 copies/µL to 2×100 copies/µL). B, The in vitro transcribed RNA from RPA product was applied to CRSIRP/LwCas13a-LFS detection, with the sensitivity of 2 copies/µL. C, The blue and ultraviolet light signals detected from CRISPR/LwCas13a reaction by using same target RNA as in B. D, The fluorescence signal intensity detected from CRISPR/LwCas13a reaction by using same target RNA as in B. ** P<0.01 versus no target RNA control (NC) (n=3).
Figure 5.
Sensitivity of RPA-CRISPR/LwCas13a-LFS for detecting D117L gene. A, The RPA products of D117L target gene of different concentrations (2×1011 copies/µL to 2×100 copies/µL). B, The in vitro transcribed RNA from RPA product was applied to CRSIRP/LwCas13a-LFS detection, with the sensitivity of 2 copies/µL. C, The blue and ultraviolet light signals detected from CRISPR/LwCas13a reaction by using same target RNA as in B. D, The fluorescence signal intensity detected from CRISPR/LwCas13a reaction by using same target RNA as in B. ** P<0.01 versus no target RNA control (NC) (n=3).
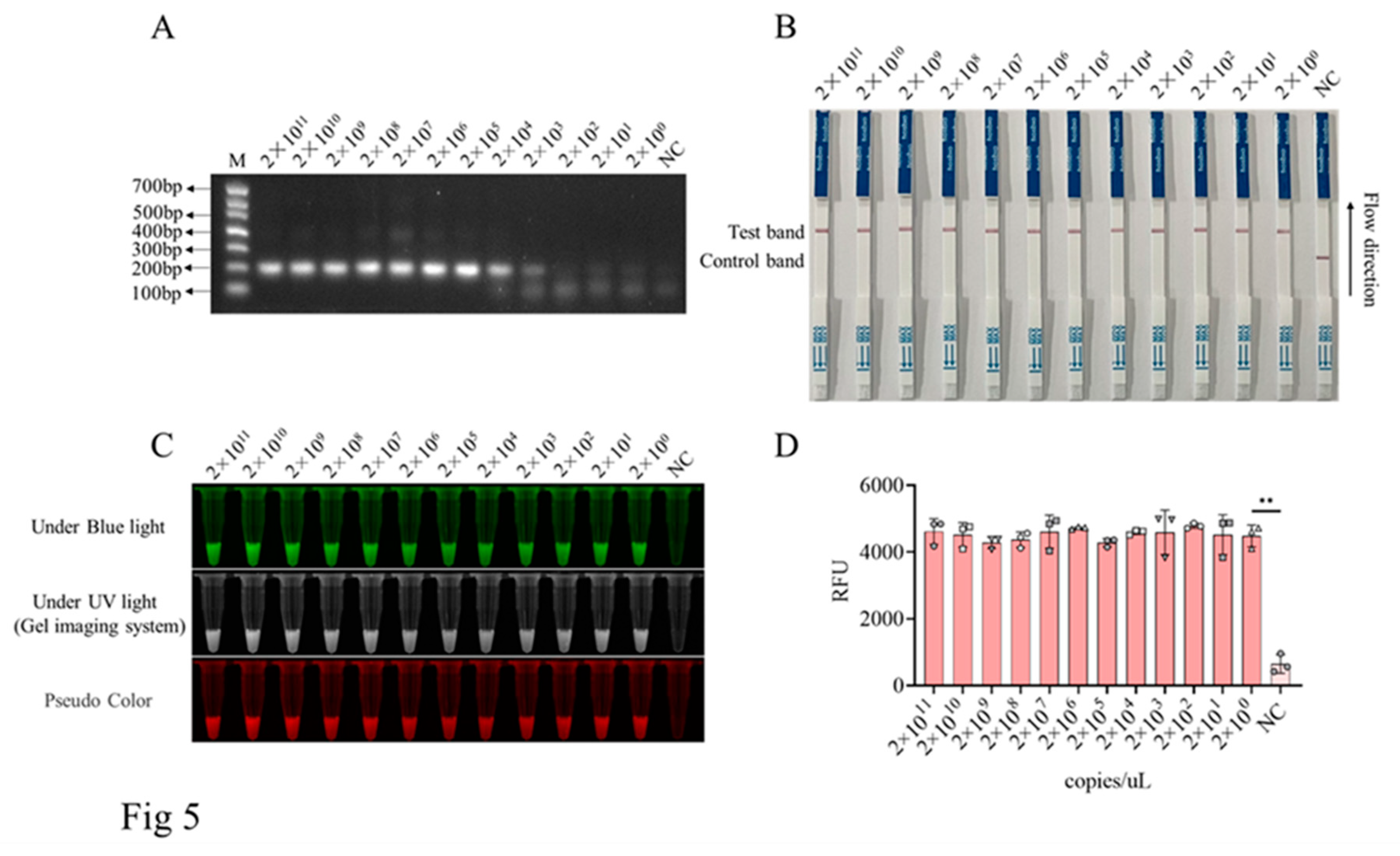
Figure 6.
The specificity of CRISPR/LwCas13a-LFS method. A, PCR or RT-PCR were performed to confirm the gene fragments of ASFV and 7 other pig viruses. B, Detection of nucleic acids of different viruses through CRISPR/LwCas13a-LFS method. C, The blue and ultraviolet light signals from CRISPR/LwCas13a reaction detecting nucleic acids of different viruses. D, The fluorescence signal intensity from CRISPR/LwCas13a reaction detecting nucleic acids of different viruses. **P<0.01 versus no target RNA control (n=3).
Figure 6.
The specificity of CRISPR/LwCas13a-LFS method. A, PCR or RT-PCR were performed to confirm the gene fragments of ASFV and 7 other pig viruses. B, Detection of nucleic acids of different viruses through CRISPR/LwCas13a-LFS method. C, The blue and ultraviolet light signals from CRISPR/LwCas13a reaction detecting nucleic acids of different viruses. D, The fluorescence signal intensity from CRISPR/LwCas13a reaction detecting nucleic acids of different viruses. **P<0.01 versus no target RNA control (n=3).
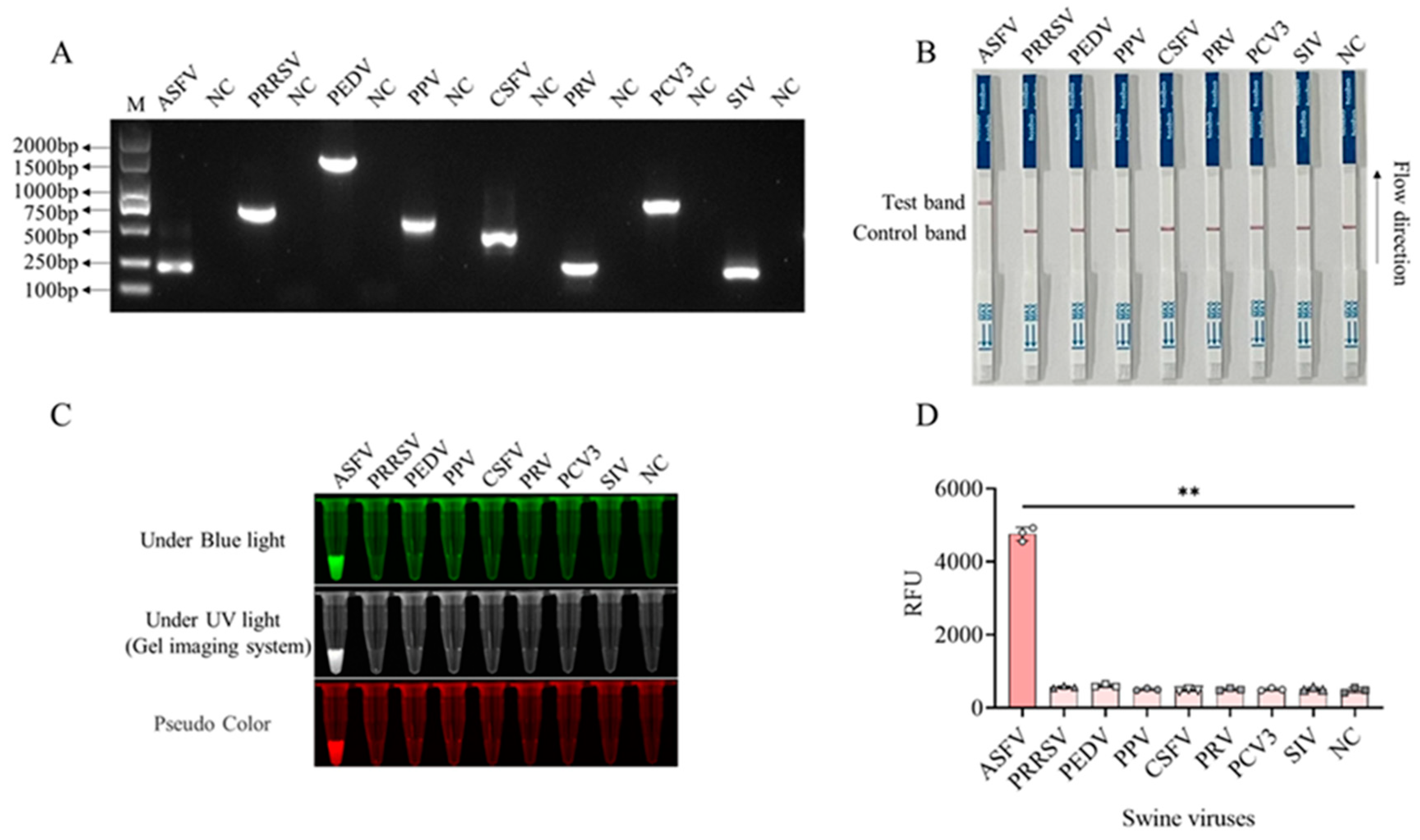
Figure 7.
RPA-CRISPR/LwCas13a-LFS detection of 33 clinical samples. 33 clinical samples were detected by RPA-LwCas13a-LFS for ASFV, with strong positive (Nos. 1-12), weak positive (Nos. 13-18), and negative (Nos. 19-33).
Figure 7.
RPA-CRISPR/LwCas13a-LFS detection of 33 clinical samples. 33 clinical samples were detected by RPA-LwCas13a-LFS for ASFV, with strong positive (Nos. 1-12), weak positive (Nos. 13-18), and negative (Nos. 19-33).


Table 1.
Comparison between RPA-LwCas13a-LFS and qPCR detections of clinical samples.
| Sample type | Samples numbers | Detection results (positive / negative) | |
| RPA-LwCas13a-LFS | qPCR | ||
| Heart | 3 | 2/3 | 2/3 |
| Liver | 3 | 2/3 | 2/3 |
| Spleen | 3 | 2/3 | 2/3 |
| Lung | 3 | 3/3 | 3/3 |
| Kidney | 3 | 2/3 | 2/3 |
| Intestinum tenue | 3 | 2/3 | 2/3 |
| Lymph node | 3 | 1/3 | 1/3 |
| Oral swab | 2 | 0/2 | 0/2 |
| Blood | 5 | 2/5 | 2/5 |
| Serum | 5 | 2/5 | 2/5 |
| Total | 33 | 18/33 | 18/33 |
| Positive ratios | 54.5 % | 54.5 % | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



