
Submitted:
17 October 2023
Posted:
17 October 2023
You are already at the latest version
Abstract
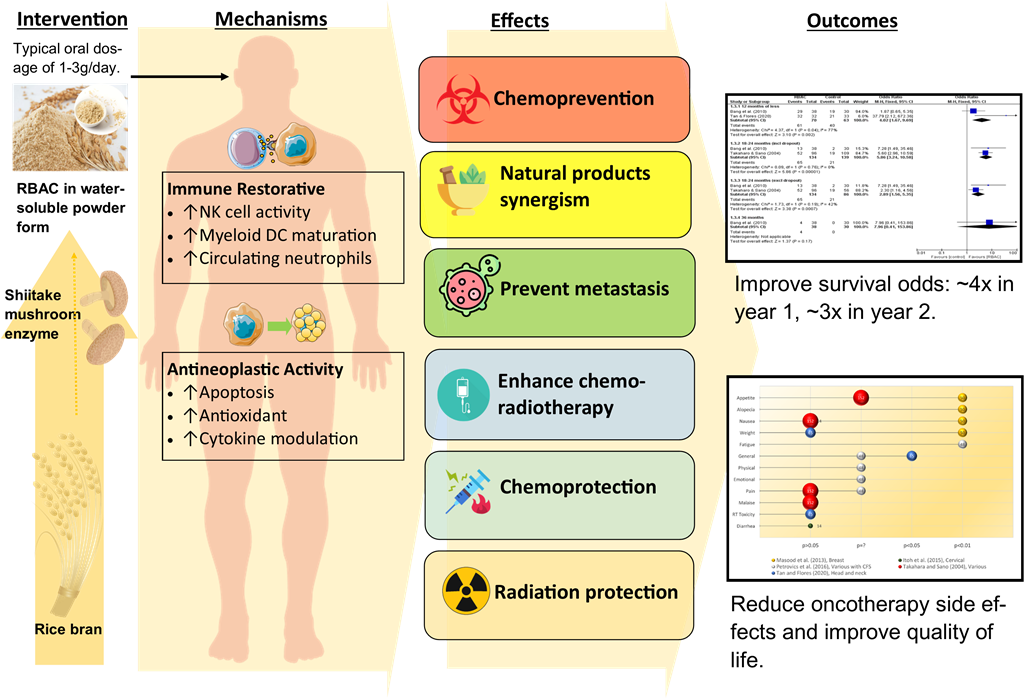
Natural products, especially biologically active compounds derived from plants, have been and continue to be invaluable in anticancer research and therapeutic discoveries. Rice bran arabinoxylan compound (RBAC) is a heteropolysaccharide extract of defatted rice bran obtained through enzymatic treatment with Lentinus Edodes mycelium. RBAC exhibited antitumour properties by arresting in vivo tumour growth in several murine models. In vitro experiments showed RBAC to promote apoptosis in cancer cells by increasing the susceptibility of CD95 (Fas/APO-1) ligands and downregulating Bcl-2 proteins to trigger the permeabilisation of the mitochondrial membrane. RBAC may restore immune dysfunction in cancer patients, most notably by upregulating natural killer cell activity and modulating cytokine production to enhance inflammatory and cytotoxic responses. As such, RBAC acts on the immune system to enhance chemopreventative activity and block tumourigenesis. In treatment, RBAC works synergistically with other natural products and chemotherapeutic agents, such as daunorubicin and paclitaxel, by enhancing drug transport to increase DNA damage in tumours and reduce proliferation. In a clinical trial, combining RBAC with chemoembolization in treating liver cancer showed improved response, reduced recurrence, and prolonged survival time. RBAC has been shown to augment the endogenous antioxidant system to prevent oxidative stress and thus protect against the harmful side effects of radiation therapy. RBAC also has chemoprotective effects, with animal and human studies showing reduced toxicity and side effects from chemotherapy, leading to improved health-related quality of life. Meta-analysing the best available evidence indicates that RBAC treatment increases the odds of survival among cancer patients by 4.02 times (95% CI: 1.67, 9.69) in the first year and 2.89 (95% CI: 1.56, 5.35) times in the second year after commencing treatment, compared to those not treated with RBAC. Notwithstanding, additional research is needed to characterise, quantify, and standardise the active ingredients in RBAC responsible for the anticancer effects. More well-designed, large-scale clinical trials are required to further substantiate the treatment efficacies.
Keywords:
Biobran
; Biologics response modifier
; Polysaccharide
; Immunomodulator
; MGN-3
; Natural killer cells
; Proapoptotic agent.
Introduction
Cancer is a disease that often evokes an image of ‘dread and death’ in the mind of most people (Kathryn et al., 2014). Indeed, according to the global mortality data estimates in 2019, cancer caused 3 out of 10 premature deaths of non-communicable diseases (Bray et al., 2021). With an estimated 19.3 million new cancer cases and almost 10.0 million cancer deaths in 2020 worldwide, cancer is a global concern (Sung et al., 2021). Also called malignancy, cancer refers to any pathophysiological conditions resulting from abnormal and uncontrolled cell growth that can become invasive to other organs or parts of the body through the circulatory and lymphatic systems (National Cancer Institute, n.d.). From a philosophical perspective, such disordered growth signifies the breakdown of the natural selection within the host tissue that defines the order of life itself (Lemoine, 2022). Cancer is thus not a disease introduced by some entity foreign to the body, but rather the host cells turning rogue to become agents of destruction (Hausman, 2019).
Generally, cancer is named based on the primary site, and the most commonly diagnosed are female breast, lung, colorectal, prostate, and stomach cancers (Sung et al., 2021). The aetiology of cancer can be many, ranging from infectious agents (such as viruses, parasites, fungi, and bacteria) to environmental exposures (such as pollutants, radiation, ultraviolet rays from the sunlight, chemical exposure, etc.) and lifestyle factors (such as cigarette smoking, an unhealthy diet with excessive fried foods and red meat, alcohol drinking, stress, obesity, and physical inactivity) (Blackadar, 2016). Essentially, any endogenous or exogenous substances capable of inducing deoxyribonucleic acid (DNA) damage can lead to cancer, and these substances are termed carcinogens (Barnes et al., 2018). Moreover, hereditary genetic predispositions can also increase the relative risks of one or more types of cancer in some individuals (Knudson, 2002).
At the cellular level, cancer is developed from a single cell following genetic damage, possibly through exposure to a carcinogen, starting to grow and divide abnormally. This proliferation then leads to the selective clonal expansion of the initiated cells and gives rise to a small benign neoplasm. However, further selective and rapid cell mass growth increases the risk of genetic mutations in clonal cells to express the malignant phenotypes and become a cancerous tumour. Malignant cells acquire more aggressive characteristics through additional genetic and epigenetic changes, including the activation of protooncogenes and the functional loss of tumour suppressor genes (Wang et al., 2018). These changes lead to tumour progression and metastasis to other body parts (Cooper, 2000; Weston & Harris, 2003).
In terms of treatment, the conventional oncological options are surgical intervention to provide definitive locoregional control of the primary tumour (Dare et al., 2015), chemotherapy for inhibiting cell proliferation and tumour growth, thus avoiding invasion and metastasis (Amjad et al., 2023), and radiation therapy to deprive cancer cells of the multiplication potential (Baskar et al., 2012). Although modalities such as immunotherapy, targeted therapy, hormonal therapy, and gene therapy are existing systematic therapeutic alternatives (Arruebo et al., 2011), chemotherapy and radiotherapy remain the mainstays for cancer treatment in the foreseeable future. The global demand for first-course chemotherapy was projected to increase from 9.8 million patients annually in 2018 to 15.0 million in 2040 (Wilson et al., 2019). Also, the optimal radiotherapy utilisation rate was estimated to be almost half (48.3%) of all patients with cancer indicated for irradiation treatment (Delaney & Barton, 2015).
Conventional chemo and irradiation treatments are known for their undesirable side effects. Nausea, vomiting, fatigue, anorexia, dysgeusia, hair loss, dry mouth, and constipation are among chemotherapy’s most common concomitant complaints (Altun & Sonkaya, 2018). Incidents of severe toxicity requiring medical intervention are not uncommon, and some can even be life-threatening. One study reported that 76.1% of participants with lung cancer from two clinical trials experienced severe toxicity during chemotherapy (Kristina et al., 2020). Moreover, the cytotoxicity of chemotherapy not only destroys malignant cells but also causes immunogenic cell death, making the host susceptible to opportunistic pathogenic infection that further weakens the immune system (Nesher & Rolston, 2014). Cancer can also develop resistance to chemotherapy, reducing the administered drugs’ efficacy and causing treatment complications (Bukowski et al., 2020). Patients receiving radiotherapy also commonly experience fatigue and localised radiation-induced adverse events such as inflammation or ulceration (head and nose), dyspnoea and chronic lung fibrosis (thoracic), and gastrointestinal (GI) symptoms (pelvic). Furthermore, depression and anxiety are common among cancer patients during treatment and may linger for years in cancer survivors (Götze et al., 2020).
To improve the therapeutic efficacy of cancer treatment while reducing the potential toxicity, researchers often look to nature for ingredients and inspiration. Substances produced naturally from living organisms, such as plants, animals, and microbes, often possess pharmacological or biological properties worth harnessing for disease treatment. Unsurprisingly, natural products, especially biologically active compounds derived from plants, have been and continue to be invaluable in anticancer research and therapeutic discoveries (Ali Abdalla et al., 2022; Ma et al., 2021; Muhammad et al., 2022; Naeem et al., 2022). Among the better-known plant-based natural products with chemopreventive and anticancer properties include curcumin in turmeric (Curcuma longa), indole-3-carbinol from cruciferous vegetables, resveratrol in grapes and wine, epigallocatechin gallate from green tea, and genistein in soybeans (Muhammad et al., 2022). Another source of natural products that has gained much interest is rice bran, the hard outer layer of rice grain when removed during milling. Rice bran extracts, fermented rice bran products, and γ-oryzanol in rice bran have all been researched for their anticancer potentials (Yu et al., 2019).
Rice bran arabinoxylan compound (RBAC) is a heteropolysaccharide extract of defatted rice bran obtained through enzymatic treatment with Lentinus Edodes mycelium (Ooi et al., 2021). The most studied RBAC is Biobran MGN-3 developed by Daiwa Pharmaceutical Co., Ltd. (Tokyo, Japan), which has been marketed as a dietary supplement for the immune system and used by cancer patients during and after treatment (Clark, 1999; Ghoneum, 2000, 2016; Igari, 2020). A previous review by the authors (SLO and SCP) has found RBAC to be an effective immunomodulator for complementing conventional cancer treatment with favourable effects, including enhancing the immune profile, reducing side effects, improving treatment outcomes, and increasing survival rates (Ooi et al., 2018). However, the physiological process of RBAC wielding such synergistic anticancer effects has not been critically assessed.
Furthermore, according to the guidelines of the American Society of Clinical Oncology (1996), the primary outcomes of cancer treatment are survival, especially disease-free survival, and health-related quality of life (QoL), including overall QoL, as well as its physical, psychologic, and social dimensions. Other outcome measures, such as toxicity, tumour response, and biomarkers, are means to assess or predict the survival or QoL of cancer patients. Hence, when considering a potential adjuvant therapeutic option for cancer, it is essential to consider the best available evidence based on the outcomes of survival and QoL.
Thus, to inform evidence-based clinical practice, the present study aims to critically evaluate the available evidence to elucidate the following two-part research questions: (1) What are the mechanisms and biological pathways that RBAC exerts synergistic effects to prevent cancer development and support cancer treatment? and in cancer patients, (2) what are the changes in survival and QoL outcomes associated with RBAC as a complementary therapy compared to no RBAC?
Methods
Sources of evidence
The current study is a secondary analysis of the evidence gathered from a previous scoping review which systematically identified all RBAC preclinical and clinical studies published until the end of 2022. The characteristics of all included studies (n=98) with bibliographic and network analyses were reported in an earlier manuscript (Ooi et al., 2023). An additional two recent RBAC studies published after the scoping study completion were also considered in the present review (Ghoneum et al., 2023; Hajto et al., 2022).
Selection criteria
To answer the first research question, the reviewers screened and shortlisted the sources of evidence (n=100) using the following concept-population-context criteria: (a) any studies of RBAC evaluating the mechanisms and biological pathways (concept); (b) on healthy or malignant cells, tissues, animal models or human participants including cancer patients (population); (c) concerning any synergistic effects to prevent cancer development or support cancer treatment (context). The reviewers excluded all case reports or series as they are not rigorous research to investigate an intervention’s effects and mechanisms of action.
From the included studies, the reviewers further shortlisted the best available evidence for RBAC as an intervention for cancer to address the second research question based on the following patient-intervention-comparator-outcome criteria: (a) a randomised controlled trial (RCT); (b) includes patients of any malignancies; (c) uses RBAC as an intervention; (d) with any comparators; and (e) outcome measures include survival and/or QoL assessment. We included trials with outcome measures based on cancer treatment-related side effects since treatment-related side effects may predict QoL (Mazzotti et al., 2012).
Evidence synthesis, analysis, and presentation
Data and results from selected articles were extracted with specific details about the citation, study design, concept, context, methodology, outcome measures and key findings relevant to the topic. The evidence synthesis is illustrated graphically, diagrammatically, or in tabular form, accompanying narrative summaries to demonstrate how the results relate to the first part of the research question.
For analysis of the survival rate, the sample sizes and survival events of RBAC and placebo groups of selected studies under similar time points were combined for analysis (published data only). The data from each study were weighed, such that studies with a smaller 95% confidence interval (CI) or a larger sample size contributed more heavily to the odds ratio (OR) estimate (Mantel-Haenszel) with a fixed effect model (Deeks et al., 2021). Review Manager 5.3 (The Nordic Cochrane Center, The Cochrane Collaboration, Copenhagen, Denmark) was used to calculate and display the meta-analysis results in a forest plot.
Due to the dissimilarity in the QoL assessment across studies, performing meta-analyses to estimate the effect sizes is not feasible. Instead, the visualisation of the evidence is achieved on a bubble chart, with QoL outcome measures as the Y-axis and statistical significance (p-value) of the outcome as the X-axis. Should the p-value of a continuous variable not be available, the reviewers will use the standard deviation or 95% CI to estimate. Fisher’s exact test was used to calculate the p-value if not reported for dichotomous outcome variables, such as alopecia events. All charting and calculations were performed with Microsoft Excel 365 (Microsoft Corp, WA, USA).
Quality assessment
Assessment of the methodological quality of the evidence was based on the Quality Assessment Tool for Controlled Interventional Studies published by the National Heart Lung and Blood Institute (2013). The assessment tool consists of 14 items covering all the essential quality criteria of an RCT, including randomisation, allocation concealment, blinding, baseline similarity, dropout, adherence, concomitant avoidance, outcome validity, power, and intention-to-treat analysis. A copy of the assessment tool is available as Supplementary S2. Two authors (PSM and SCP) and an independent assessor evaluated the study quality independently, with consensus achieved through the Delphi method (Nasa et al., 2021). The third author (SLO) was the facilitator, aggregating and sharing the responses to the checklist anonymously with the group after each assessment round. The assessors could adjust their answers at each iteration based on how they interpret the group response until the agreement is reached. The 14 quality assessment items can be further grouped for the detection of six types of bias, namely selection (items 1-3), performance (item 4), detection (item 5), attrition (items 7-8), reporting (items 11,13-14) and other biases (items 6, 9-10, 12), summarised in a percentage-stacked bar chart. The clinical effects of the best available evidence and the assessed quality formed the basis for final recommendations.
Results
The flow of article selection is depicted in Figure 1. Out of the 100 preidentified RBAC sources of evidence, 50 non-cancer-related articles did not fulfil the inclusion criteria, 11 case reports/series on cancer patients and one study protocol were excluded. Hence, 38 articles were included for evidence synthesis. Of these, 24 were preclinical studies (16 animal and 8 in vitro), and 14 were human clinical trials (RCT=7, non-RCT=1, before and after=5, cross-sectional=1). The characteristics of all included articles are summarised in Table S2.1 (Supplementary S2). Note that 89.5% (34/38) of all included studies are based on Biobran MGN-3, and the rest (10.5%, 4/38) are based on other RBAC products produced by Erom Co., Ltd. (Chuncheon, South Korea).
The following sections synthesise the evidence on how RBAC exerts synergistic effects to prevent cancer development or support cancer treatment and the potential mechanisms.
Immune restorative effects
The immune system plays an essential role in suppressing cancer growth through immunosurveillance by cytotoxic lymphocytes, including the natural killer cells (NKC) and cluster differentiation (CD) 8+ T cells. As such, developing a malignant tumour from initiation to proliferation requires cancer cells to evade the immune system attacks by avoiding recognition and instigating an immunosuppressive microenvironment conducive to tumour growth (Gonzalez et al., 2018). Restoring and harnessing the body’s antitumour immune response to control and eliminate tumours thus becomes a viable therapeutic option in cancer known as immunotherapy (Wu et al., 2021). RBAC was known as an immunomodulator, with the available evidence on the immune restorative effects in cancer listed in Table 1.
Low numbers of dysfunctional NK cells are often observed in the microenvironment of advanced solid tumours due to the production of soluble modulators, low nutrient levels, and hypoxic conditions that negatively affect the maturation, proliferation, activation, and cytolytic function of NK cells (Melaiu et al., 2020). This phenomenon has prompted the call for NKC-based immunotherapy for cancer treatment (Riggan et al., 2021). Available evidence has demonstrated RBAC to be a potent activator of NKC cytolytic activity against malignant cells in cancer patients. Ghoneum and Brown (1999) first reported in a single-arm study of 32 patients with various malignancies (prostate, breast, multiple myeloma [MM], and leukaemia) who had completed one or more conventional therapies (surgery, chemotherapy, and radiation). Low NKC activity levels were prevalent among these patients. After taking RBAC (3g/day) orally for one to two weeks, significant increases (p<0.001) in NKC activity of up to 10-fold compared to baseline were detected (Ghoneum & Brown, 1999). In a separate but possibly related article, Ghoneum (1999) reported that 86 out of 90 cancer patients (95.5%) with various malignancies who received 3g/day of RBAC after completion of conventional therapies demonstrated 2 to 10-fold increases in NKC cytolytic activity at 1-2 weeks posttreatment. However, since Ghoneum (1999) is a conference abstract, insufficient detail is presented, and the data was not peer-reviewed.
Further examination of NKC granularity by Ghoneum and Brown (1999) with cytocentrifuge preparation of a patient’s peripheral blood lymphocytes (PBL) at baseline found low or absent granularity, indicating dysfunctional NK cell populations. Increased NKC granularity was subsequently observed in the same patient after RBAC treatment for one week. These NKC demonstrated increased capacity in binding and killing tumour cells (K562) in vitro compared to the low granular NKC isolated before treatment. Testing of T and B lymphocyte proliferation after one month of RBAC treatment in five selected patients also showed statistically significant (p<0.001) increases in responses with phytohaemagglutinin (B cell mitogen), concanavalin A (T cell mitogen) and pokeweed (T and B cell mitogen) tests compared to baseline, all of which demonstrated signs of restoration of the adaptive immunity (Ghoneum & Brown, 1999).
An in vivo experiment by Badr El-Din, Abdel Fattah, et al. (2016) also showed that oral administration (p.o.) of RBAC at 40 mg/kg body weight (BW) every other day was able to prevent lymphocyte depletion in male Wistar rats exposed to the carcinogen methylnitronitrosoguanidine (MNNG). After eight months, rats administered with MNNG alone had a significantly lower percentage of lymphocytes (↓23.3%, p<0.01) compared to healthy controls. However, the group treated with RBAC after MNNG administration showed lymphocyte recovery, with the levels returning to normal, a significant difference from the untreated MNNG group (p<0.05).
Takahara and Sano (2004) analysed the relationship between NKC cytolytic activity and survival rate in an RCT with two groups of cancer patients. All participants had progressive cancer of late stages (III-IV) with recurrence, unresectable lesions, or metastasis after surgery. The intervention group (n=96) received 3g/day of RBAC oral supplement plus complementary therapies, whereas the control group (n=109) received only the complementary therapies. Fifty patients in the control group could not complete the study due to cancer progression or pessimism in the treatment. After 18 months, a higher survival rate (p<0.019) was observed in the RBAC group (54.2%, 52/96) compared to the control group (33.9%, 19/56). The difference in survival rate is more significant (p<0.001) based on intention-to-treat analysis, which includes all dropouts (control=53, RBAC=0). The study found no patients who dropped out had survived at 18 months. Hence, the survival rate for the control group was only 17.4% (19/109).
When categorising the participants based on initial NKC activity of low (<20%), medium (20% to 40%), and high (>40%), the study found significantly higher rates of participants with low or medium NKC activity in the RBAC group survived, compared to the control group (Low: 42.5% vs 12.5%, p<0.01; Medium: 51.4% vs 28.0%, p<0.05). Hence, RBAC appeared to upregulate the dysfunctional NKC in late-stage cancer patients to prolong survival.
Conversely, an exploratory RCT by Itoh et al. (2015) did not detect any significant difference in NKC activities between the RBAC (n=7) and the control (n=7) groups in cervical cancer patients receiving chemoradiotherapy. The trial was conducted over three weeks of one treatment cycle, with the participants starting either oral RBAC (3g/day) or placebo powder up to one week before treatment commencement. Both groups experienced NKC activity decline after chemoradiotherapy compared to the baseline values. Hence, RBAC could not prevent the depletion of NKC activity during chemoradiotherapy in this trial. Nonetheless, with the small sample size and short duration, the study may not have sufficient statistical power to detect the treatment effects.
Cholujova et al. (2013) studied the immunomodulatory effects of RBAC on the innate immunity of MM patients in a double-blind placebo-RCT. Admitted to this study were MM patients (n=48) under observation and those receiving or completed chemotherapy. Participants were randomly assigned to take 2g/day of RBAC (n=32) or a matching placebo (n=16) orally for three months, and their blood samples collected at baseline and monthly intervals. The study found significant increases in NKC cytolytic activity of the RBAC group compared to the baseline (30.8±7.4 lytic unit [LU]) in the first (47.0±8.5 LU, p=0.045) and second (56.6±12.2, p=0.029) months but not the third month. No significant differences in NKC cytolytic activity were observed in the placebo group throughout the trial. Additionally, Cholujova et al. (2013) also detected a substantial increase in the percentage of circulating myeloid dendritic cells (DC) after three months of RBAC treatment compared to baseline (25.8±3.6 % vs 17.6±2.6 %, p=0.036). The myeloid-to-plasmacytoid DC ratio in the RBAC group also significantly increased (p=0.030). In contrast, no significant changes in both DC markers were detected in the placebo group over time.
The myeloid DC capture and present antigens on their surface to T lymphocytes, thus bridging the innate immunity to adaptive immune responses (Chistiakov et al., 2015). The plasmacytoid DC, on the other hand, are crucial to antiviral immunity by specialising in the secretion of high levels of interferons (IFN) type I (Ye et al., 2020). These DC also play a role in immunosuppression by recruiting regulatory CD4+CD25+ T lymphocytes (Treg) into the tumour microenvironment (Zhou et al., 2021). Treg lymphocytes are characterised by the forkhead box protein p3 expression, a master transcription factor that suppresses anticancer immunity and thus promotes proliferation (Li et al., 2020). In MM patients, myeloid and plasmacytoid DC populations were inversely correlated with disease progression (Pasiarski et al., 2013). The increase in myeloid DC levels after the three-month RBAC supplementation coincided with the tapering of NKC cytolytic activity. Such observations could signify a switch from innate immunity to more lasting adaptive immunity as part of the immune restorative process in MM patients.
The Treg lymphocytes are immune regulatory cells that tightly regulate immune activation to prevent response to self-antigens, permit tolerance for weak antigens, and limit collateral damage in inflammation. Treg are essential to prevent autoimmune diseases, but they also suppress myeloid DC maturation and prevent T and B cell differentiation and proliferation, allowing cancer to escape detection (Ohue & Nishikawa, 2019; Sojka et al., 2008; Togashi et al., 2019). Lissoni et al. (2008) studied the changes in total NKC, total T lymphocytes and the T cell subpopulations (CD3+, CD4+CD25+, CD4+, and CD8+) in 24 consecutive cancer patients who had received RBAC for two months (2g/day for the first month and 1g/day after). Among the participants, 18 did not respond to conventional treatment for solid metastatic tumours and had no other effective standard treatment. The remaining 6 had surgery only, for locally limited neoplasms. Two participants died due to disease progression before the end of the study, leaving the results of 22 participants for evaluation.
The study by Lissoni et al. (2008) found no substantial changes in the mean number of lymphocytes, T lymphocytes (CD3+), T cytotoxic (CD8+) lymphocytes and NKC before and after RBAC intervention. The mean cell count of T helper (Th, CD4+) and Treg increased and decreased, respectively, but without reaching statistical significance. Notwithstanding, a statistically significant change in the ratio of Th/Treg was detected (p=0.025), and the increase in Th/Treg was more pronounced in participants with low Th/Treg at baseline (Lissoni et al., 2008). Hence, RBAC treatment appeared to inhibit the immunosuppressive Treg while restoring the adaptive immune responses facilitated by CD4+ Th in the fight against cancer.
Neutropenia is a common complication among cancer patients, especially those treated with chemotherapy, with almost one-third of patients developing low neutrophil count during treatment (Salako et al., 2021). The reduction of circulating neutrophils in the bloodstream increases the risk of infections. Neutropenia is even more common in patients with haematological malignancies, and the risk of bloodstream infection is more pronounced (Carvalho et al., 2020). The combination of fever and neutropenia (febrile neutropenia) is one of the most common causes of oncological emergencies, which can be fatal (Yi et al., 2020). The risks of further infections and mortality among patients with febrile neutropenia remained high for six months after the initial episode (Nordvig et al., 2018).
Golombick et al. (2016) reported the potential restorative effects of RBAC on the depleted neutrophil count of patients with early B-cell lymphoid malignancies in a preliminary single-arm study. Recruiting patients with monoclonal gammopathy of undetermined significance (MGUS)/smouldering multiple myeloma (SMM) who had been on oral curcumin therapy (6g/day) for six months or more, this study added RBAC (2g/day). Inflammatory and immunologic markers were monitored every two months for six months. Half of the MGUS/SMM patients (n=10) were neutropenia at baseline. The study found an increased neutrophil count between 10% and 90% among 8 participants after consuming RBAC. Such observations are encouraging but require validation through a larger controlled clinical trial.
As summarised in Figure 2, RBAC appears to be a biological modifier that could prevent or restore immune dysfunction in cancer patients by upregulating NKC cytolytic activity, improving the maturation of myeloid DC, inhibiting the immunosuppressive Treg, and reversing neutropenia. All these effects help to neutralise or eliminate immunity suppression triggered by tumour-associated inflammation, thus restoring the effectiveness of antitumour immune responses (Shalapour & Karin, 2015).
Anticancer effects and pathways
Anticancer effects in vivo
RBAC has been shown to arrest tumour growth and demonstrate anticancer activity directly. Table 2 shows a list of murine models investigating RBAC’s anticancer effects in halting and reversing in vivo tumour growth and extending survival rates of treated animals.
Bae et al. (2004) compared RBAC to polysaccharide peptide (PSP) extracted from the mycelium of basidiomycetes, a known natural anticancer product, in an experiment with ICR mice injected with sarcoma 180 (S-180) squamous cells. The mice were orally fed with either RBAC or PSP (1.5mg/day) as treatment or saline as a control for 23 days. RBAC was effective in inhibiting tumour growth by 66.2% based on tumour weight (TW) at the end of the study compared to untreated control mice (0.51 ±0.34g vs 3.40+1.46g, p<0.01). In contrast, TW reduction by PSP was lesser (↓49.0%, p<0.05), albeit was statistically significant compared to untreated control. The mean BW of the RBAC and PSP groups were also significantly lower (p<0.01) than that of the control mice from day eight onwards.
Similarly, Badr El-Din et al. (2008) also found RBAC to have in vivo anticancer effects in female Swiss albino mice inoculated with Ehrlich ascites carcinoma cells intramuscularly. After eight days, mice bearing a solid Ehrlich carcinoma (SEC) mass of ∼100 mm3 were randomly divided into receiving RBAC (40mg/kg BW) either intraperitoneally (i.p., 3x/weeks from day 10) for three weeks or intratumorally (i.t., 2x/week from day 11) for five weeks. SEC-bearing mice receiving saline injections were used as controls. The delay in tumour development was apparent in RBAC-treated mice. In the i.p. group, the mean tumour volume (TV) became significantly lower than the control group starting from day 14 (p<0.05), with between-group differences increasing throughout the study period. By day 35, the percentage difference in mean TV was 63.27% (p<0.001) in favour of the i.p. group. The mean TW of the i.p. group at day 35 was also significantly lower (3.63±0.45 vs 6.62±0.38g, p<0.01) than the control. In parallel, the i.t. group also demonstrated a significant TV reduction trend starting from day 28, reaching a -44.83% (p<0.01) difference on day 45. Through flow cytometry analysis of SEC, the study also found a 1.8-fold increase in the percentage of apoptotic cells in RBAC-treated mice (74.68±4.22%) compared to the control mice (42.61±5.56%, p<0.0001) with the increased apoptosis, further confirmed through histopathological examinations of the tumours.
The results of the anticancer effects of RBAC in SEC-bearing mice were also validated by Badr El-Din et al. (2019) in a similar study with female Swiss albino mice. Treatment with RBAC at 40mg/kg BW i.p. (3x/weeks from day 11) for three weeks was shown to significantly prevent BW loss in SEC-bearing mice (↓4.1% vs 18%, p<0.01) and reduce TW (↓46.3%, p<0.01) compared to control at day 30. Continuous suppression of TV throughout the study was detected: On day 14, the TV of RBAC-treated mice was 33.7% (p<0.01) less than untreated mice, and the reduction reached 49.9% (p<0.01) at the end of the study.
Noaman et al. (2008) performed another study with SEC-bearing mice to compare low-dosage RBAC treatment (25mg/kg BW i.p.) of two schedules on tumour growth. The early treatment schedule started from day four and continued to day 25 (19 injections, 6x/week), whereas the late treatment began from day 11 up to day 25 (13 injections). Early treatment significantly retarded TV by 54% relative to the control, compared to only 24% in the late treatment group (p<0.01). Both treatment schedules also showed markedly reduced mean TW compared to the control, with -34% (p<0.01) for the early group versus -12% (p<0.05) for the late group.
Another RBAC product, Erom’s rice bran bio-exopolymer (RBEP), also shows anticancer effects in vivo. Kim et al. (2007) conducted experiments on RBEP with two different models: (1) Survival time of ICR mice inoculated with S-180 sarcoma to induce malignant ascites, and (2) Solid tumour growth in C57/Bl6 mice transplanted with B16/Bl6 melanoma. In the first experiment, mice were treated with RBEP of different dosages (50mg/kg BW i.p. or p.o., 250mg/kg BW p.o.). RBEP was found to prolong the mean survival time of mice with malignant ascites, compared to the untreated mice (27.4 days), by 14.6% (31.4 days) and 30.3% (35.7 days) with 50mg/kg and 250mg/kg p.o. treatment, respectively. Further prolonging lifespan by 38.0% (37.8 days) was observed in mice treated with 50mg/kg i.p., demonstrating that i.p. could be the preferred therapeutic route for RBEP (Kim et al., 2007). In the mice transplanted with B16/Bl6 solid tumour, RBEP significantly (p<0.05) inhibited TW by 35.6% (2.38g vs 3.70g of control mice) with the 50mg/kg p.o. treatment, 41.7% (2.155g) with 250mg/kg p.o., and 55.1% (1.66g) with 50mg/kg i.p. For comparison, another group of mice was treated with fluorouracil, a pyrimidine antagonist (antimetabolite), and the TW was 0.851g at the end of the study. Thus, fluorouracil inhibited tumour growth by 77% compared to no treatment. Comparatively, RBEP was not as effective as fluorouracil in tumour growth inhibition.
An (2011) also confirmed that treatment with 250mg/kg BW of RBEP p.o. and i.p. daily for two weeks effectively extended survival and reduced cancer growth of S-180 inoculated ICR mice. The study found higher survival rates in RBEP-treated mice than the controls, with 5.3% higher in the p.o. group (19.9 vs 18.9 days) and 23.2% higher in the i.p. group (23 vs 18.7 days). Notably, on day 23, all i.p. mice treated with RBEP remained alive, but none in the control group survived. Evaluating tumour growth by BW, the study found significantly (p<0.05) lower BW than the control in the p.o. group starting from day 13. For the p.o. group, significantly lower BW was detected as early as day 10, and the difference continued to widen until the end of the study (p<0.001).
RBAC derived from a specific black rice cultivar called fermented SuperC3GHi bran (C3G-F) was also tested for its anticancer properties by Kim et al. (2011) on mice models with malignant ascites (ICR mice + S-180 cells) and solid tumour (C57BL/6 mice + B16/BI6 melanoma). The study found 250mg/kg BW C3G-F administered orally reduced the BW gain of the ascites-bearing mice compared to the control mice. The between-group mean BW difference reached statistical significance (p<0.05) from day eight onward. At day 15, the BW of the C3G-F group was about 60% lower than that of the control group (6.5 g vs 11.8g). In the second experiment, mice fed with 250mg/kg BW C3G-F also had solid tumours with 19.4% lower mass than untreated control mice three weeks after transplantation (0.514±0.129g vs 0.635±0.241g, p<0.05). Haematologic investigations found that C3G-F-treated mice had significantly higher white blood cell count than the control mice (4.24±0.71 vs 2.63±1.26, p<0.05). Accordingly, it was inferred that the in vivo antitumour effects of RBAC products could be via strengthening the immune system.
To demonstrate that NKC activated by RBAC could have a direct role in tumour suppression, Pérez-Martínez et al. (2015) conducted an in vivo experiment with NOD-scid IL-2Rgnull mice inoculated with NB-1691luc neuroblastoma cells. Intravenous NK cellular therapy, with either fresh NKC or NKC activated with RBAC (100mg/ml) overnight, began after seven days of tumour cell transplant for four weeks (2x/week). Another group of cancer cells-inoculated mice received only saline injections as controls. Through bioluminescence imaging, the study found tumours in mice receiving RBAC-activated NKC treatment had significantly lower TV (p<0.05) than that of the two control groups at day 42. Furthermore, through Kaplan-Meier analysis, mice in the RBAC group were shown to survive significantly longer (p<0.05) than the other two cohorts. RBAC, therefore, could activate NKC to reduce TV and increase the chance of survival in cancer-bearing mice.
RBAC acts not only on the host immune system but also has been shown to directly affect cancer cells to arrest tumour growth. The potential mechanisms investigated in the literature include impacts on the proapoptotic pathway, oxidative stress, and cytokine signalling are shown in Table 1.
Promote cancer cell apoptosis
Ghoneum et al. (2000) reported that incubation of squamous cell carcinoma (SCC13) cell line with RBAC showed a 30% decrease in cell numbers after 48 hours and 50% at 72 hours. In contrast, untreated SCC13 cells continued to grow over time. Coculturing of RBAC with human breast cancer cells (MCF-7) showed significant decreases in cell survival rates of 75%, 70% and 63% after three days, at concentrations of 100, 500 and 1000mg/ml, respectively (Gollapudi & Ghoneum, 2008). The half maximal inhibitory concentration (IC50), a measure of the potency of RBAC against MCF-7 cells, was estimated to be approximately 800 μg/ml at 24 hours and about 1000 μg/ml at 48 hours. The effect of RBAC against murine breast cancer cells (4T1) was even more remarkable, with IC50 being 700 μg/ml at 24 hours and 580 μg/ml at 48 hours (Ghoneum et al., 2014). Likewise, Brush et al. (2010) found RBAC to significantly decrease (p<0.05) the proliferation of human prostate cancer cell lines (PC3 and LNCaP) in a dose-dependent manner after culturing the cells for 24, 48, and 72 hours with different doses of RBAC (0-1000 µg/ml).
Given that RBAC is known to be non-cytotoxic to healthy cells with no direct effect on healthy mouse fibroblast (L929) cell line (An, 2011) and did not affect microbial cell viability in vitro (Ghoneum et al., 2008), the mechanism of how RBAC inhibits malignant cell growth is worth exploring. To this end, Ghoneum et al. (2000) examined cytokine secretion by the SCC13 cells cultured with RBAC. There was an 8-fold increase in interleukin (IL)-10 and a 3-fold increase in IL-12 after 16 hours, but no apparent change in interferon-gamma (INF-γ) content was detected. Thus, the reduction in SCC13 cell count could be due to increased secretion of IL-10 and IL-12 triggered by RBAC, as these are cytokines known to induce programmed cell death via the CD95 (APO-1/Fas) receptor/ligand pathway (Fan et al., 2002; Schmidt et al., 2000).
To validate the proapoptotic mechanism, Ghoneum and Gollapudi (2003) studied the effect of RBAC on CD95 death receptor-induced apoptosis in the human HUT 78 T lymphocyte cell line (leukaemia). The study found that HUT 78 cells treated with RBAC (100–1000 μg/ml) alone induced about 2.5–4.5% of specific apoptosis (over and above spontaneous programmed cell death) after 24 hours. Anti-CD95 antibodies, on the other hand, induced about 20% specific apoptosis. Most importantly, pretreatment of HUT 78 cells with RBAC (for 3 hours) before incubating with anti-CD95 antibodies increased the specific apoptosis significantly (p<0.01) by 35-42%, about double that of anti-CD95 antibodies alone. Such an increase was not associated with the upregulation of death receptors on the HUT 78 cells, as the percentage of cells expressing CD95 and the density of CD95 on the cell surface were not different between treated and untreated cells. Additional experiments by Ghoneum and Gollapudi (2003) also found that, compared to untreated control, activation of intracellular caspases 3, 8, and 9 increased significantly (p<0.001) in cells treated with RBAC and anti-CD95 antibodies. Moreover, a marked decrease in membrane potential and significant downregulation of the Bcl-2 antiapoptotic molecule in RBAC-treated HUT 78 cells compared to untreated cells were also detected. The results confirm that RBAC could increase the susceptibility of cancer cells to undergo apoptosis mediated by the CD95 (APO-1/Fas) death ligands.
Badr El-Din, Abdel Fattah, et al. (2016) performed cell-cycle analyses of the stomach tumour cells of male Wistar rats induced with carcinogen MNNG to understand the proapoptotic actions of RBAC further. Significant differences were detected in cells in G0/G1, SubG1, and S phases between rats fed with RBAC (40 mg/kg BW every other day) for eight months and those that were not. In particular, RBAC mitigated the carcinogenic effects of MNNG by causing cell-cycle arrest in the subG1 phase with a 115.8% increase in the hypodiploid cell population (p<0.01) compared to the MNNG group. Furthermore, comparing the ratio of the apoptotic index over the proliferation index (AI/PrI), the MNNG+RBAC group showed a 1.67-fold increase in AI/PrI over the MNNG group. AI/PrI is a prognostic marker for cancer proliferation, with a higher value indicating a much higher apoptotic rate of tumour cells, slowing the growth (Liu et al., 2001). Quantification of apoptosis confirmed that the addition of RBAC increased the apoptotic cancer cells in tumour tissues by 63.7% (p<0.01) compared to treatment of MNNG alone, most prominently during the early apoptosis with a 230.1% (p<0.01) increase to eliminate unwanted cells damaged by MNNG. In terms of the expression of apoptotic regulators in gastric tumour cells, RBAC was shown to induce apoptosis via the mitochondria-dependent pathways through downregulation of Bcl-2 (↓15.1%, p<0.05) and upregulation of p53 (↑37.3%, p<0.05), Bax (↑49.3%, p<0.01), Bax/Bcl-2 ratio (↑75.7%, p<0.01) and caspase-3 (↑34.8%, p<0.01). The upregulation of the p53 gene indicates that RBAC could enhance the tumour suppressor protein production to stop the division of mutated cells.
RBAC’s effects on NDEA+CCI4-induced hepatocarcinogenesis based on cell cycle analysis of liver tissues were also reported by Badr El-Din et al. (2020). Cell-cycle arrest in the sub-G1 phase was markedly increased by 126% and 99% (p<0.01) through pretreatment and posttreatment of RBAC, respectively, compared to no treatment. Flow cytometric analysis of apoptosis also showed that RBAC treatment (pre, post) significantly reduced (p<0.01) viable cells (↓74.51%, ↓72.54%) and necrosis (↑89%, ↑75.47%) while increasing early (↑316%, ↑309%) and late (↑255%, ↑237%) apoptosis, compared to carcinogen-untreated rats. The analysis of apoptotic gene regulators also showed treatment of RBAC significantly (p<0.01) upregulated p53, Bax, and caspase-3 while downregulated Bcl-2 gene expression levels relative to untreated rats. The study also found marked downregulation of nuclear factor kappa B (NFκB)/p65 inflammatory pathways in the liver of the RBAC-treated rats by reversing the decrease of IκB-α (nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha) gene expression caused by NDEA+CCI4. Detection of DNA damage of the liver tissues by gel electrophoresis also showed increased fragmentation by 364.83% and 477.35% in RBAC treatment groups (pre, post), respectively, compared to the untreated rats. Hence, RBAC inhibited hepatocarcinogenesis through induced apoptosis, suppressed inflammation and reduced tumour cell proliferation (Badr El-Din et al., 2020).
Similarly, in female Swiss albino mice inoculated with SEC, Badr El-Din et al. (2019) showed that RBAC treatment (40 mg/kg BW i.p. 3x/weeks) for three weeks increased apoptosis of the tumour cells. A marked increase of cell-cycle arrest in the sub-G1 phase was detected by 102% (p<0.01) in the RBAC group compared to the control after 30 days of treatment. RBAC also increased the AI/PrI ratio by 2-fold (p<0.01). Through quantitative histochemical analysis, the study also showed reduced viable cells (28.2±1.25% vs 74.5±2.25%) and increased apoptotic cells (53.1 ± 1.21% vs 18.2±1.68%) in the tumour tissues of RBAC-treated mice compared to control. Additionally, RBAC also significantly (p<0.01) upregulated p53 (↑113.78%), Bax (↑114.1%), and caspase-3 (↑123.22%), while downregulated Bcl-2 (↓53.32%) gene expression. The Bax/Bcl-2 ratio increased by 358.9% in the RBAC-treated mice compared to those without treatment.
Prevent oxidative stress
Reactive oxygen species (ROS) such as superoxide radicals (O2•−), hydrogen peroxide (H2O2), and hydroxyl radicals (•OH) are genotoxins that can cause DNA damage leading to malignancy (Phillips & Arlt, 2009). High levels of ROS accompany the hyperproliferation of cancer cells and deplete endogenous antioxidants, causing oxidative stress, which can harm the surrounding healthy tissues. Cancer cells, however, adapt and thrive under oxidative stress. As such, ROS greatly assist in initiating, promoting and progressing tumour growth (Hayes et al., 2020). Antioxidants, such as endogenous glutathione (GSH), could modulate DNA-repair activity to suppress tumour progression (Chatterjee, 2013). Several plant-based antioxidants, including resveratrol, baicalein, and genistein, have been shown to be genotoxic but not mutagenic and could selectively kill multidrug-resistant cancer cells (Fox et al., 2012). Hence, the ability to enhance the endogenous antioxidant system could be another mechanism by which RBAC impairs tumour growth.
Noaman et al. (2008) evaluated the antioxidant status of the SEC-bearing mice and the corresponding effects of RBAC treatment. The study found significant elevations in malondialdehyde (MDA), a measurement for lipid peroxidation, in the plasma (↑58.96%, p<0.05) and liver (↑44.54%, p<0.01) of SEC-bearing mice compared to that of healthy mice at day 25 after Ehrlich carcinoma cells inoculation. Not surprisingly, SEC-bearing mice also had significantly lower levels of GSH in the blood (↓25.96%, p<0.05) and liver (↓59.31%, p<0.01) than the control values. Marked depletion (p<0.05) of endogenous antioxidant enzymes, including glutathione peroxidase (GPx), glutathione-S-transferase (GST), superoxide dismutase (SOD) and catalase (CAT), were also detected with corresponding downregulation of gene expressions. Such results confirmed the increased ROS attack and the presence of oxidative stress in mice with cancer.
For comparison, two groups of SEC-bearing mice were treated with RBAC (25 mg/kg BW starting from day 4 [early, E] or day 8 [late, L] after injection of Ehrlich ascites). These mice did not show signs of oxidative stress, with the MDA values in the blood (E↑1.73% and L↑7.52%) and liver (E↓21.57% and L↓9.03%) not significantly different from the control. Furthermore, when comparing the MDA levels within the tumour tissue, the early and late treatment group had significantly lower values than the untreated SEC-bearing mice, showing -39.34% (p<0.01) and -36.43% (p<0.05) reductions, respectively. The GSH levels of RBAC-treated SEC-bearing mice in the blood (E↑39.0%, L↑3.67%), liver (E↑40.97%, L↑14.04%) and tumour (E↑74.41%, L↑59.12%) were at normal or above normal values, and significantly higher (p<0.01) than those of the untreated SEC-bearing mice. Similarly, the levels of GPx, GST, SOD, and CAT and the related gene expressions in both RBAC groups were shown to be significantly higher (p<0.01) than the untreated mice and not deviating much from the control values. Hence, RBAC could attenuate oxidative stress to minimise tumour growth by instigating higher endogenous antioxidant productions, thus averting collateral damage to healthy cells.
Modulate cytokine production
The anticancer effects of RBAC could also be linked to the ability to influence the cytokine production of immune cells. Badr El-Din et al. (2008) reported that SEC-bearing mice treated with RBAC (40 mg/kg BW 3x/day) for three weeks had significantly higher levels (p<0.01) of tumour necrosis factor (TNF)-α (↑15.63% over control) and IFN-γ (↑154.54% over control) compared to both untreated tumour-bearing mice (TNF-α ↑4.17%, IFN-γ ↓10.46%) and healthy control mice after 35 days. Additionally, untreated tumour-bearing mice were observed to have elevated IL-10 compared to tumour-free mice by 111.71%, whereas only a minor change was detected in RBAC-treated mice (↑14.75%). The difference between the treated and untreated groups was statistically significant (p<0.01). Both TNF-α and IFN-γ are secreted by Th1 cells and exert proinflammatory and anticancer activity, whereas IL-10 is a type of antiinflammatory cytokines of Th2 cell response that mediates humoral immunity. High levels of Th2 response relative to low Th1 could favour tumour growth (Lin et al., 2019; Zhao et al., 2019).
Cholujova et al. (2013) confirmed that a group of MM patients (n=45) had a predominant Th2 response over Th1 by analysing the ratios of plasma concentration of Th1 cytokines (IL-1β, IL-2, IL-12, IL-15, and IFN-γ) to Th2 (IL-4, IL-5, IL-6, IL-9, IL-10, and IL-13) compared to healthy donors (n=30). Healthy donors were observed to have 20 Th1/Th2 ratios that were greater than 1.0 (Score: 20:10), whereas, in MM patients, there were only 14 such ratios (Score: 14:16). RBAC was shown to elevate (p<0.05) significantly the plasma concentration of several Th1 cytokines in MM patients over placebo, especially IL-12, IL-17, and TNF-α, consistently when measured at one month and after three months. IL-1β was also elevated at one month (p=0.047) but not after three months, whereas IFN-γ was significantly higher (p<0.018) after three months. However, after three months of RBAC treatment, the levels of Th2 cytokines, including IL-4, IL-6, IL-9, IL-10, and IL-13, were significantly increased (p<0.05) compared to the placebo (Cholujova et al., 2013). Thus, RBAC supplementation appeared to affect both Th1 and Th2 cytokines, demonstrating immunomodulating effects, but how it could influence the disease progression of MM remained unclear.
In a non-randomised clinical trial, Kim et al. (2020) found that cancer patients (n=10, with various malignancies) consuming an oral nutritional supplement containing 0.4g of RBEP for eight weeks showed significantly lower IL-1β, IL-6 and IL-8 (p<0.05) compared to the control group (n=24) receiving standard care in nutritional counselling only. The RBEP group, however, had a significantly higher IL-12p70 (p<0.05) level than the control while no difference in TNF-α levels was detected. The authors also reported a marginally significant rise (p=0.056) in the IL-10 level in the RBEP group at week eight compared to baseline, but the between-group difference was not significant. Notably, the cytokine levels in this study were measured from the PBL of patients after being stimulated by lipopolysaccharides (LPS) to determine the levels of inflammatory responses. Again, supplementation of RBAC appeared to affect both Th1 (IL-12p70, IL-1β and IL-8) and Th2 (IL-6 and IL-10) bi-directionally with no clear indication of whether how such cytokine modulation could influence the inflammatory responses of the body and its antitumour mechanisms and impact. More research in this area is needed.
Overall, RBAC appears to exert anticancer effects through multiple pathways, including selectively promoting apoptosis in cancer cells via both intrinsic and extrinsic pathways, acting as an antioxidant, and modulating antitumor cytokines secretion, as summarised in Figure 3.
Chemoprevention
Cancer chemoprevention is the use of natural, synthetic, or biological chemical agents to reverse, suppress, or prevent carcinogenic progression (Tsao et al., 2004). There has been a strong interest in leveraging natural products as a risk-modifying strategy to prevent, delay, or suppress tumour development or recurrence, especially in high-risk patients (G et al., 2022; Haque et al., 2021). RBAC was shown to be a prophylactic agent against carcinogenesis in vivo, with results summarised in Table 4.
Badr El-Din, Abdel Fattah, et al. (2016) studied the chemoprevention activity of RBAC against chemical-induced glandular stomach carcinogenesis in rats. Male Wistar rats were given carcinogen MNNG (200 mg/kg BW p.o. daily) for two weeks to instigate cancer growth. Along with chemical induction, the rats (n=12) were given RBAC at 40 mg/kg BW every other day for eight months. Another group of rats (n=10) were treated with MNNG alone. After eight months, histopathological examination of the gastric mucosa of the rats showed that 80% of the rats treated with MNNG only developed mild- and high-grade gastric glandular dysplasia (6/10, 60%) and invasive well-differentiated keratinising cell carcinoma (2/10, 20%). The MNNG+RBAC group, however, showed significantly lower incidence (p<0.01) of mild dysplasia, of which were patchy and small (3.5/12, 29.2%) and carcinoma in situ only (1/12, 8.3%). In addition, the MNNG+RBAC group also had significantly lower (p<0.01) Ki-67 tumour proliferation marker expression at 39.8% compared to 50.8% in the MNNG-only group. Hence, RBAC considerably lowered the risk of developing gastric dysplasia and adenocarcinoma while exposed to MNNG.
Another study by Badr El-Din, Ali and Othman (2016) explored the in vivo chemopreventive effects of RBAC (25 mg/kg BW i.p. 5x/week) on liver cancer under two treatment regimes. Male albino rats were administered with N-nitrosodiethyamine (NDEA, 200 mg/kg BW, i.p.) at week 2 to induce hepatocarcinogenesis, followed by weekly subcutaneous injections of carbon tetrachloride (CCI4, 3ml/kg BW for 6 weeks) as a promoter. Pretreatment of RBAC for a group of mice (n=20) commenced two weeks prior to the injections of NDEA+CCI4 and lasted for another 20 weeks. Conversely, the posttreatment group only received RBAC from week 10 to week 22. The study found NDEA+CCI4 induced significant BW loss (↓39.54%, p<0.01) and increased liver mass (↑24.73%, p<0.01) in untreated mice compared to healthy control at the end of 22 weeks. Both RBAC treatment regimes kept the liver weight at the normal range and significantly reduced (p<0.01) the percentage of BW loss caused by the carcinogens, with pretreatment (↓17%) faring better than posttreatment (↓23.44%). Histopathological studies of the liver tissues of the NDEA+CCI4 mice showed signs of inflammation and hepatocarcinogenesis with fatty infiltration of hepatocytes, loss of architecture, necrosis and fibrosis. As for rats pretreated with RBAC, the liver tissues showed minimal changes in hepatocyte morphology and histology with no inflammation. Moderate liver damage was observed in the posttreatment group but with only a few degenerated hepatocytes. Testing of liver enzymes also showed similar findings between two treatment regimes. Thus, RBAC treatment prevented carcinogenesis in the liver even with the
Enhance chemotherapy
Combining two or more therapeutic agents in oncological treatments is a common practice as it can reduce the risk of acquired resistance and enhance efficacy through the synergistic or additive effects of the agents (Palmer & Sorger, 2017). For instance, evidence of combining immunotherapy and chemotherapy showed improvements in overall progression-free survival, response rates and duration, as well as clinical benefit for MM, breast cancer, and lung cancer (Morse et al., 2023). With its immunomodulation and proapoptotic effects, RBAC could be a safe and effective addition to combination treatment, with evidence listed in Table 5.
Gollapudi and Ghoneum (2008) explored the sensitising activity of RBAC with daunorubicin, an anthracycline class of antibiotics, against human breast cancer cells (MCF-7 and HCC70) in vitro. Coculturing RBAC with daunorubicin for three days lowered the IC50 values against MCF-7 cells by 3-, 5- and 5.5-fold, at 100, 500 and 1000 µg/ml, respectively. The IC50 of daunorubicin for HCC70 cells was also consistently decreased by 2.5-fold with RBAC of all concentrations. RBAC appeared to enhance the drug transport with evidence of increased accumulation of daunorubicin in both MCF-7 and HCC70 cells observed under flow cytometry. The administration of RBAC (500 mg/ml) enhanced drug accumulation in MCF-7 cells over time, with differences compared to daunorubicin-only uptake starting 45 minutes after culturing and reaching 26.21% higher at the hour.
RBAC was also tested for its synergistic effects with paclitaxel, a mitotic inhibiting taxane, on breast cancer cell (non-metastatic MCF-7 and metastatic 4T1) growth in vitro. Ghoneum et al. (2014) showed that the IC50 values of paclitaxel against MCF-7 at 24 hours were lowered by a factor of over 100 with the addition of 600, 750, and 1000 μg/ml of RBAC, compared to paclitaxel alone. Compared to paclitaxel alone against 4T1 cells, the IC50 value for paclitaxel at 24 hours decreased by a factor of ~3 at 600 μg/ml of RBAC and up to a factor of ~100 at 1000 μg/ml. Additional in vitro experiments also showed that paclitaxel plus RBAC (500 and 600 μg/ml) significantly increased DNA damage, reduced proliferation, and induced apoptosis of 4T1 cells, compared to either agent alone.
Badr El-Din, Ali, Alaa El-Dein, et al. (2016) followed up with an in vivo study to examine the treatment effects of combining RBAC (40 mg/kg BW) and low-dose paclitaxel (2 mg/kg BW) in a murine model. They utilised female Swiss albino mice (n=36) that were inoculated with Ehrlich ascites carcinoma. The mice received no treatment, RBAC only, paclitaxel only, or RBAC plus paclitaxel, every other day. At day 30 post-inoculation, the study found the combination therapy significantly reduced (p<0.01) TV by 88.3% compared to no treatment. The reduction in TV was more pronounced than the effects of either paclitaxel (↓58.9%) or RBAC (↓77.1%) alone. RBAC plus paclitaxel also inhibited tumour cell proliferation at a higher propensity (↓35.4%, p<0.01 vs untreated mice) compared to only 11.6% and 27.0% of paclitaxel or RBAC alone, respectively. RBAC plus paclitaxel also maximised the downregulation of Ki-67 expression by 85.7% (p<0.01) compared to no treatment, with paclitaxel or RBAC alone reduced Ki-67 expression by 51.7% and 80.6%, respectively. Significant increases (p<0.01) in the percentage of cancer cell apoptosis were also detected in all treatment groups, showing 20.9% for paclitaxel only, 76.1% for RBAC only, and 93.2% for paclitaxel+RBAC. Analyses of DNA damage and cell cycle phases also showed a similar trend, with paclitaxel+RBAC superior in causing much more DNA damage and maximising the AI/PrI ratio compared to either agent alone.
The effectiveness of RBAC in improving the treatment outcomes of conventional antineoplastic drugs has been studied in an RCT by Bang et al. (2010). Patients (n=68) with hepatocellular carcinoma (stages I and II) participated in this study, with the intervention group (n=38) receiving RBAC (1g/day) as a dietary supplement for 12 months while receiving oncological treatment simultaneously. The control group (n=30) received only the standard therapies. The oncological therapies were mainly transarterial oily chemoembolisation (TOCE, n=24) or TOCE in combination with percutaneous ethanol injection treatment (TOCE+PEIT, n=34). A few participants received PEIT only (n=6) or PEIT plus radiofrequency ablation (n=4). Hence, all participants received antineoplastic drugs directly delivered to their tumour sites.
RBAC was reported to significantly improve (p<0.01) the treatment response rate of standard therapies for liver cancer, with 89% of patients in the RBAC group responding to oncological treatment compared to only 80% in the control group. The mean posttreatment alpha-fetoprotein (AFP) tumour marker in the RBAC group significantly decreased by 38% compared to baseline (p<0.001), a favourable contrast over the non-significant 7% increase in AFP in the control group. Furthermore, combining RBAC with standard therapies significantly decreased (p<0.01) the average TV in patients by 36% compared to almost no change in the control group (↑0.2%). After the treatment, the patients were followed up every six months for up to 3 years, and the tumour recurrence rate in the RBAC was lower at 32% compared to 47% in the control group. In terms of survival, 63% of patients receiving only standard treatment survived the first year, only 6.7% lasted at least two years, and none survived after 30 months. In contrast, the RBAC group maintained a much higher survival rate at 76%, 35%, and 11% at the end of 1, 2, and 3 years, respectively. In particular, patients receiving RBAC in addition to TOCE+PEIT survived, on average, ten months longer than those treated with TOCE+PEIT only. Hence, evidence from this RCT supported the synergistic anticancer effects of RBAC in enhancing the effectiveness of TOCE and/or PEIT in increasing treatment response, reducing TV, lowering the AFP marker, and prolonging the survival of liver cancer patients (Bang et al., 2010).
Chemoprotection
Chemoprotection refers to protecting healthy cells and tissues from toxicity and side effects of chemotherapy. Several studies have demonstrated that RBAC could be a promising source to achieve such protection (see Table 6).
Jacoby et al. (2001) explored the in vivo effects of RBAC in reducing the toxicity of cisplatin (an alkylating agent) and doxorubicin (an anthracycline antibiotic like daunorubicin) with a murine model. Sprague-Dawley-derived albino rats (n=80) were orally fed with 0, 5, or 50 mg/kg BW of RBAC daily for 11 days. On day 3, rats were administered cisplatin (9 mg/kg BW), doxorubicin (10 mg/kg BW), or vehicle control by a single i.p. injection. The study found that RBAC could prevent weight loss induced by the chemotherapeutic agents. Rats administered cisplatin alone showed weight loss at day 11 (98.5±0.06% of initial BW). In contrast, weight gains were observed in both low and high-dose RBAC plus cisplatin groups (L: 111.5±0.13%, H: 144.0±0.15%) with significant differences compared to the cisplatin-only group (p<0.05). The doxorubicin-only group also showed BW gain (132±0.13%±0.05) but was significantly lower (p<0.05) than the gains in RBAC plus doxorubicin groups (L: 146.6±0.08%, H: 143.5±0.06%).
The toxicity of cisplatin was severe, with 50% deaths, 70% had gross GI mucosal pathology, and 100% showed signs of diarrhea (Jacoby et al., 2001). The corresponding proportion in the low-dose RBAC plus cisplatin group was 10% death (p<0.05), 40% GI pathology, and 50% diarrhea (p<0.05). The high-dose group reported 40% death, 50% GI pathology, and 40% diarrhea (p<0.05). Compared to cisplatin, doxorubicin had less toxicity, no death, and mostly non-significant differences in diarrhea across all doxorubicin-treated groups. Notwithstanding, 50% of the doxorubicin-only group experienced GI pathology compared to only 10% in the low-dose RBAC plus doxorubicin group (p<0.05) and 30% in the low-dose RBAC plus doxorubicin group (p>0.05). Hence, RBAC at 5 mg/kg BW appeared more effective than at the higher dose of 50 mg/kg in preventing the toxicity and side effects of cisplatin and doxorubicin (Jacoby et al., 2001).
Endo and Kanbayashi (2003) investigated the chemoprotective effects of RBAC (1 mg/day p.o. and i.p.) against BW loss due to cisplatin in BALB/c female mice over a longer duration. One shot of cisplatin (15 mg/kg i.p.) was administered after the mice had received RBAC for one week. The mice were weighed daily for 28 days. Control substances were either drinking water (p.o.) or phosphate saline (i.p.). Analysis of variance (ANOVA) was conducted at weekly intervals corresponding to the (I) initial phase, (II) weight loss phase, (III) weight gain phase, and (IV) weight stabilising phase. Statistically significant differences (p<0.05) in BW were detected in phases II, III and IV of both groups of RBAC (i.p. and p.o.) compared to their respective control groups, with the RBAC groups showing trends of reduced BW loss and faster BW recovery over time. When comparing the two groups of RBAC, there was no significant difference in the protective effect of the administration route on weight loss induced by cisplatin.
In humans, the chemoprotective effects of RBAC were validated by Masood et al. (2013) in an RCT among breast cancer patients (n=50) receiving chemotherapy. One group of patients (n=25) were assigned to take RBAC (3g/day) as a dietary supplement one week before and one week after chemotherapy. Another control group (n=25) received only chemotherapy. The trial lasted for six cycles of chemotherapy, with the patients completing questionnaires before each treatment cycle to assess any chemotherapy-induced side effects. The study found significant differences (p<0.001) in the proportions of patients experiencing anorexia/tiredness (RBAC vs control: 20% vs 88%), nausea/vomiting (40% vs 100%), hair loss (28% vs 100%) between the two groups. Furthermore, the distribution of patients having weight gain or loss significantly differed with weight gain among 64% in the RBAC but none in the control group. Instead, 84% of the control group experienced weight loss but none in the RBAC group. Hence, RBAC reduced the chemotherapy-induced side effects of anorexia/tiredness, nausea/vomiting, hair loss, and weight loss among breast cancer patients.
Radioprotection and radiotherapy enhancement
With antioxidant capacity, RBAC has also been shown to protect against the harmful effects of radiation treatment, as shown in Table 7.
Ghoneum et al. (2013) explored how RBAC could protect mice against whole-body γ-irradiation. Female Swiss albino mice were irradiated with an acute single dose level of 5 Gy at a rate of 0.45 Gy/min. One group of mice (n=6) received RBAC (40 mg/kg BW i.p.) every other day for two weeks before irradiation and continued receiving RBAC until four weeks after. Compared to irradiated mice that did not receive RBAC, the RBAC group showed less weight loss relative to control nonirradiated mice when measured at week 1 (↓1.41% vs. ↓20.03%, p<0.01) and week 4 (↓0.54% vs. ↓7.79%, p<0.05) after irradiation. RBAC prevented the radiation-induced weight loss and helped maintain regular BW throughout the trial. Significant differences in the liver (RBAC vs irradiation: ↓8.58% vs ↓25.51%, p<0.05) and kidney (↑5.04% vs ↓23.19%, p<0.05) weight were also observed between the two groups at week 1, although the organ weights for all groups return to normal at week 4.
Exposure to γ-radiation also caused anaemia in the mice showing significantly lower (p<0.05) than normal red blood cell (RBC) count and haemoglobin (Hb) levels measured after 1 and 4 weeks (Ghoneum et al., 2013). Moreover, irradiation also caused significant (p<0.01) leukopenia, lymphopenia, neutrophilia, and thrombocytopenia in mice compared to healthy control at week one before normalising at week four except for platelet count, which remained significantly lower than normal (p<0.5). Histopathological examination of the bone marrow revealed haematopoietic tissue damage with the absence of cellularity in irradiated mice and a significant decrease (p<0.01) in spleen size (↓60%) and megakaryocyte density (↓75%) compared to control mice at week 1, which only partially recovered at week 4. In contrast, RBAC prevented anaemia from radiation exposure and maintained normal white blood cells, lymphocytes, neutrophils, and platelets in the treated mice. The preservation of haematopoietic tissues by RBAC was evident in histopathological examination showing normal bone marrow cellularity, spleen size and megakaryocyte density despite exposure to harmful irradiation.
The beneficial effects of RBAC against γ-irradiation could be due to its ability to protect against ROS by enhancing the endogenous antioxidant system discussed earlier. Oxidative stress was observed in irradiated mice, with the MDA level spiking at 106.34% (p<0.01) above normal at week 1, accompanied by a significant decline in the GSH level (↓40%, p<0.01). MDA remained high (43.44%) at week 4, while endogenous GSH content was restored over time. The RBAC, however, showed only a slightly elevated MDA level at week 1, which was not significantly different from the healthy control. The GSH content of RBAC-treated mice remained high throughout the trial.
The potential mechanisms for RBAC’s radioprotective effect were investigated by Zhao et al. (2020) in an animal study with radiation-induced intestinal injury. One group of C57BL/6 mice was pretreated with RBAC (40 mg/kg BW i.p.) every other day for two weeks before undergoing a local high-dose abdominal precision irradiation at 2 Gy/min for five minutes (10 Gy single dose). RBAC treatment continued every other day for another four weeks. A separate group of mice received only irradiation. At the end of the study, the jejunal and colonic segments of the mice were collected for analysis. Irradiation was found to disrupt cellular respiration with significant reductions (p<0.05) in mitochondrial respiratory chain complexes, resulting in the depletion of intercellular adenosine triphosphate (ATP) content in the jejunal and colonic mucosa compared to healthy control. However, in mice treated with RBAC, the mitochondrial respiratory chain complex activity and intercellular ATP content remained normal. Moreover, the abundance of mitochondria-encoded genes and mitochondrial copy numbers in the jejunal and colonic mucosa of irradiated mice treated with RBAC increased significantly (p<0.05) compared to the reduction observed in the irradiation-only mice. Thus, RBAC was able to preserve mitochondrial function from the harmful effects of radiation.
Zhao et al. (2020) also evaluated the oxidative status of the intestinal epithelium after radiation by assessing the levels of ROS, reactive nitrogen species (RNS), MDA, and H2O2. As expected, all oxidative status markers were significantly elevated (p<0.05) in the irradiation-only mice compared to the healthy control. Analysis of the antioxidative amplitude of SOD, GPx, CAT, and total antioxidant capacity in serum and intestinal mucosa also showed significant depletion (p<0.05) after irradiation in mice. Conversely, RBAC protected the intestinal epithelium from oxidative stress by strengthening the endogenous antioxidative activities and total antioxidant capacity to neutralise radiation-induced free radicals, thus maintaining oxidative status at normal levels. The study also found evidence of a significant increase (p<0.05) in intestinal permeability and disruption of the barrier function of mucosa after irradiation. However, RBAC restored these components to the levels of the control mice. As such, RBAC could protect against irradiation-induced intestinal damage through its antioxidant capacity.
RBAC could not only protect against the adverse effects of radiation therapy but also enhance the efficacy of the treatment. Badr El-Din et al. (2019) demonstrated the benefits of combining RBAC (40 mg/kg BW i.p. 5x/week for three weeks) and X-ray irradiation (3x 2Gy dose with a dose rate of 0.85 Gy/min) in female Swiss albino mice inoculated with SEC. At the experiment endpoint, the study found the combined treatment reduced the TV by 77.3% and TW by 56.9% compared to no treatment. The reduction was significantly more than the effects of RBAC (TV↓66.4%, TW↓46.3%, p<0.05) or radiation treatment (TV↓49.9%, TW↓30.7%, p<0.01) alone, evidence of the synergistic effects of the two therapies. The increased efficacy was also accompanied by reduced adverse effects of irradiation as the addition of RBAC managed to significantly arrest the BW loss in RBAC+radiation-treated mice compared to radiation-only mice (↓17.9% vs 31.2%, p<0.01). Badr El-Din et al. (2019) also conducted quantitative histochemical analysis and reported that tumour tissues from RBAC+radiation treated mice contained only 4.6±0.93% viable cells, 64.0±1.47% apoptotic cells, and 21.4±1.7% necrotic cells. In comparison, the tumour tissues of RBAC-only (viable: 28.2±1.25%, apoptotic: 53.1±1.21%, necrotic: 18.8±0.96%) and radiation-only (viable: 30.3±1.23%, apoptotic: 41.3±1.22%, necrotic: 28.4±0.89%) groups contained more viable cells and less apoptotic cells. The increase in apoptosis of the tumour cells by RBAC+radiation treatment was also confirmed with the highest cell-cycle arrest at the sub-G1 phase, peak AI/PrI ratio, while maximising the levels of apoptotic regulators (p53, Bax, caspase-3) and the corresponding apoptotic gene expressions (Badr El-Din et al., 2019).
Tan and Flores (2020) confirmed the radioprotective effects of RBAC in a double-blind placebo-RCT with head and neck cancer patients (n=65) undergoing radiotherapy and/or concurrent chemotherapy. The patients were mainly prescribed a total radiation dose of 70 Gy and randomly assigned to either the RBAC (n=32) or placebo (n=33) groups. The oral supplementation dosage was 3g/day, commenced two weeks before the start of oncological treatment, during chemoradiotherapy, and for two months after completion. The study found reductions in the haematological parameters in both groups during chemoradiotherapy. Two months after treatment, significant between-group differences (p<0.05) were detected in Hb, haematocrit, RBC, platelets, neutrophils, and lymphocytes, with the RBAC group showing favourable recovery compared to the placebo group. However, the study did not detect any statistical differences in radiation toxicities assessment between the two groups based on the Radiation Therapy Oncology Group (RTOG) severity grading. Notwithstanding, participants in the RBAC group reported significantly better mean scores in health-related QoL than that of the placebo group (1.53±0.24 vs. 1.72±0.33, p=0.019). Clinical outcomes of the RBAC group were also significantly better than the placebo group in mortality (0% vs 33.3%, p<0.001), blood transfusion (51.5% vs 3.1%, p<0.001), hospitalisation (63.6% vs 6.2%), and metastasis (15.2% vs 0%, p<0.05). The placebo group also reported marginally higher infection cases than the RBAC group (12.1% vs 0%, p=0.06). The results showed the superiority of RBAC over placebo in radiation protection, subjective QoL, and objective treatment outcomes.
Synergistic effects with other natural products and complementary therapies
RBAC was reported to work synergistically with other natural products or complementary therapies, including yeast, curcumin, mistletoe lectin, and oncothermia, as shown in Table 8.
Malignant cells may develop phagocytic behaviour against host cells or other microorganisms, especially in aggressive and invasive tumours (Lugini et al., 2003). Heat-killed Saccharomyces cerevisiae, commonly known as baker’s or brewer’s yeast, could cause apoptosis in breast cancer cells after being engulfed by phagocytic tumour cells (Ghoneum & Gollapudi, 2004). S. cerevisiae has also been explored as a probiotic and natural product for antitumour actions (Badr El-Din et al., 2018; Shamekhi et al., 2020).
RBAC was shown by Ghoneum and Gollapudi (2005a) to enhance yeast-induced apoptosis synergistically against breast cancer cells in vitro. Tumour cells (MCF-7) were cocultured with yeast (1:10 ratio) in the presence (100, 500, 1000 µg/ml) or absence of RBAC. Treatment with RBAC showed a 2-fold increase (54% vs 27%, p<0.001) in the percentage of yeast attachment to the MCF-7 cells at 0.5-hour post incubation, which was accompanied by a significant increase (p<0.01) in the rate of phagocytosis of MCF-7 cells by yeast (72%), as compared to cells cultured with yeast without RBAC (23%). Increased apoptosis in RBAC-treated MCF-7 cells was also detected, with 32% undergoing apoptosis compared to only 18.7% in untreated cells with yeast alone at 0.5 hours. The apoptosis rates continued to rise for both cultures, reaching 31% for yeast alone and 39.3% in the presence of RBAC. However, with more cell death, the yeast attachment percentage declined for RBAC, whereas untreated MCF-7 cells continued to attract yeasts. By 2 hours, the attachment rates were reversed with a significant between-group difference (p<0.001). Moreover, compared to MCF-7 cells and yeast only, RBAC caused dose-dependent increases in phagocytosis-induced cell death of 35.4%, 40.1%, and 33.04% at 100, 500, and 1000 µg/ml, respectively. Further experiments with other cell lines (ZR-75 and HCC70) also showed consistent results of RBAC-enhancing yeast-induced apoptosis. The effect of RBAC was also shown to associate with increased activation of caspases 8 and 9 in MCF-7 cells and caspases 8, 9 and 3 in HCC70 cells (Ghoneum & Gollapudi, 2005a).
Similar experiments were repeated by Ghoneum and Gollapudi (2005b) using MCF-7 cells in monolayer culture instead of suspension culture. Non-cancerous breast epithelial cells (MCF-10A) were used as controls. Cells were cultured with heat-killed S. cerevisiae (1:10 ratio) in the presence or absence of RBAC (100 µg/ml). Monolayer MCF-7 cells also exhibited properties of phagocytosis with increased attachment to yeast observed over time (↑13.4% at 1 hour and ↑25% at 4 hours) compared to the control MCF-10A cells which showed no phagocytosis behaviours. RBAC increased the magnitude of phagocytising of yeast by MCF-7 cells by 2- to 3-fold after one to four hours. The percentage of dead MCF-7 cells after treatment with RBAC, yeast, or yeast+RBAC for four hours showed 58%, 85%, and 92%, respectively, compared to only 9.5% in untreated MCF-7 cells. Hence, RBAC could improve the effectiveness of yeast-included apoptosis in MCF-7 cells regardless of how the cell culture is maintained.
Ghoneum and Gollapudi (2011) also reported that RBAC had a synergistic apoptotic effect with curcumin. The in vitro study found that treatment of MM cell line U266 with curcumin alone (2.5-10 μM) and RBAC alone (50 and 100 μg/ml) resulted in a decreased cell survival rate that followed a dose-dependent manner. However, RBAC+curcumin caused a significant decrease (p<0.0005) in cell survival compared to either agent alone and achieved an 87% decrease in cell count at 100 μg/ml RBAC and 10 μM curcumin. Apoptosis determined by hypodiploid DNA showed that RBAC alone at 50 μg/ml caused about 10% of apoptosis in U266 cells, which was no different from background apoptosis. However, combining RBAC (50 μg/ml) with curcumin significantly increased (p<0.05) to 20.0, 22.0, and 24.7% at 2.5, 5, 10 μM, respectively. Cell cycle analysis showed that RBAC+curcumin caused a significant decrease in the G0 phase from 36 % to 17% (p<0.0005), a slight decline in the S phase (from 15.5% to 13%) and the G2-M phase (from 15% to 12%). Additionally, RBAC+curcumin treatment altered the Bax/Bcl-2 ratio to 2.5 compared to 0.9, 1.5, and 1.6 in control, RBAC alone, and curcumin alone. Hence, combining RBAC and curcumin could better promote cell apoptosis in malignant U266 cells.
RBAC has also been applied in cancer treatment with mistletoe lectin (Viscum album L.) extract, another natural product known to possess cytotoxic and immunostimulating effects (Majeed et al., 2021). Hajtó, Horváth and Papp (2016) surveyed the QoL of 35 patients (20 females, 15 males) with advanced (stage II-IV) cancer of various malignancies after being treated with RBAC (12 to 45 mg/kg BW 2x/week) as an oral supplement and a standardised mistletoe extract (Iscador M 5 mg 2x/week) as subcutaneous injections for six months or more. The patients were asked to complete a questionnaire about the combined therapy’s perceived impacts on anxiety, physical activity, appetite, sleep, digestion, side effects of cancer treatment, and disease progression. Improvement in physical activity (71%) and appetite (66%) were two of the most important benefits reported by the patients. Among the 35 patients, 24 were concurrently treated with conventional cancer treatment, with 70.8% (17/24) also citing the reduction of side effects as a benefit of RBAC and mistletoe extract. Thus, RBAC works synergistically with mistletoe extract to improve cancer QoL according to the cross-sectional survey by Hajtó, Horváth and Papp (2016). However, mistletoe extract is known to improve the QoL of cancer patients, with a meta-analysis reporting a significant, medium-sized effect (Loef & Walach, 2020). Hence, without a controlled study, the impact of RBAC on enhancing the effectiveness of mistletoe extract is unknown.
Oncothermia, also called modulated electro-hyperthermia, is a complementary therapy that applies low-radiofrequency heat to the tumour site to direct energy absorption in the extracellular liquid and destroy the cell membrane of cancer (Andocs et al., 2009; Gabriella et al., 2013). By overheating the malignant tissues locally in a targeted and controlled manner, oncothermia is used to improve the efficacy of conventional cancer treatment while improving the QoL and survival rate of cancer patients (Alshaibi et al., 2020). The combined therapy of RBAC and oncothermia has also been shown to work synergistically to reduce cancer patients’ symptoms with chronic fatigue syndrome (CFS) in an RCT.
Petrovics et al. (2016) recruited cancer patients with various malignancies diagnosed with CFS (n=50). One group of participants (n=25) were randomised to consume RBAC (1 g/day for 24 weeks) and undergo oncothermia (60 min with 140 W energy weekly for 15 times) concurrently while being treated by chemo- or radiotherapy. Another group served only as controls receiving only conventional oncological treatment. The fatigue symptoms were measured with Chalder Fatigue Scale (CFQ), and the subjective clinical outcome of the combined treatment was also assessed by the Patient Global Impression of Change Scale (PGIC). At 24 weeks, the RBAC+oncothermia group reported a significantly lower mean CFQ score compared to baseline (14.6±2.3 vs 23.9±2.3, p<0.01), while the control group recorded no significant change in mean CFQ. The reduction in fatigue is also clinically significant as the RBAC+oncothermia group reported a mean PGIC score of 2.1±0.5 at 24 weeks, indicating the patients’ perceiving ‘much improved’ CFS symptoms after treatment. In contrast, the control group’s mean PGIC score was 4.3±0.9, which means ‘no change’ (Petrovics et al., 2016). Notwithstanding, the study by Petrovics et al. (2016) did not include oncothermia-only or RBAC-only treatment as a third and fourth arm. Therefore, there was insufficient data to determine the effects of each therapy and any multiplicative impact on the combination.
Metastasis prevention
Metastasis refers to the development of a secondary tumour at a distance from the primary cancer site. Cancer metastasis is a major cause of disease morbidity and accounts for about 90% of cancer deaths (Guan, 2015). Metastatic cascade is a complex process. Tumour cells must detach from the primary tumour and migrate through the microenvironment before entering the blood or lymph vessels (Schegoleva et al., 2022). Once in the bloodstream, these circulating tumour cells (CTCs) must survive the immunosurveillance while being transported at a distance at which attachment to the endothelium in a target organ happens. The CTCs then invade the surrounding parenchyma and form new tumours (Lin et al., 2021). As such, CTCs play a major role in cancer metastasis and are suggested as a biomarker for cancer diagnosis, prognosis and a therapeutic target for eradication (Deng et al., 2022; Schegoleva et al., 2022). Specifically, a high CTC count is considered clinically relevant in non-metastasised breast cancer for early detection of recurrence (Fridrichova et al., 2022). Whereas in metastasised breast cancer, CTC and CTC cluster count are indicators of disease progression and therapy response (Fridrichova et al., 2022).
In a single-arm human study by Pescatore et al. (2022), RBAC has been shown to lower CTC count in cancer patients. The study evaluated the CTC counts of 12 participants (males to females = 1:1, aged 56-79) before and after taking RBAC (1g/day for 10 to 19 weeks). The initial mean CTC count was 8.33±8.89 for the group and was significantly reduced (p<0.047) to 2.33±3.50 at the end of the study. Of the 12 participants, two had a CTC count of zero throughout the study. The rest (n=10) all recorded lower CTC counts. In parallel, decreased tumour markers, including prostate-specific antigen (PSA, for prostate cancer, n=6) and cancer antigens 125, 27.29, and 15-3 (CA125, CA27.29, and CA15-3 for breast [n=2], ovarian [n=1], and uterine [n=1]), were detected in nine participants with one of them experiencing remission (Pescatore et al., 2022). Therefore, the evidence suggests that RBAC could prevent disease progression and potential metastasis of the primary tumours by reducing CTC. However, such observations require validation with a larger controlled study in future research.
To summarise, research evidence has shown that the potential beneficial effects of RBAC chemoprotection against carcinogenesis, enhancement of chemotherapy and radiation treatment efficacies, protection against toxicity and side effects of oncological treatment, synergism with other natural and complementary cancer treatment, and prevention of metastasis. These effects are illustrated in Figure 4.
Best available evidence of RBAC treatment in cancer patients from clinical trials
RCT is the best available evidence in clinical research to establish causal associations between interventions and outcomes (Zabor et al., 2020). As such, in evidence-based practice, RCT is considered the gold standard for evaluating the efficacy of any new treatment (Hariton & Locascio, 2018). This review identified a total of seven RCTs of RBAC. However, only six fulfilled the inclusion criterion of having survival and/or QoL as outcome measures for evaluation (Bang et al., 2010; Itoh et al., 2015; Masood et al., 2013; Petrovics et al., 2016; Takahara & Sano, 2004; Tan & Flores, 2020). The excluded RCT by Cholujova et al. (2013) investigated only the impact of RBAC based on changes in the immune profile of the participants without any treatment efficacy outcomes.
The characteristics of all the included RCTs are summarised in Table 9. Four RCTs recruited patients with a specific cancer type, including liver (Bang et al., 2010), breast (Masood et al., 2013), cervical (Itoh et al., 2015), plus head and neck (Tan & Flores, 2020). In contrast, Takahara and Sano (2004) and Petrovics et al. (2016) recruited patients with various cancer types. Additionally, Petrovics et al. (2016) only included participants diagnosed with CFS. All these RCTs used Biobran MGN-3 as the RBAC intervention with a dose of either 1g or 3g per day. RBAC was used as an adjunct therapy for conventional cancer treatment except for Takahara and Sano (2004), where the participants were treated with complementary therapies plus anticancer drugs with fewer side effects. All studies assessed RBAC as a sole intervention except Petrovics et al. (2016), which assessed the combination therapy of RBAC with oncorthemia. Comparator-wise, only two studies were placebo-controlled (Itoh et al., 2015; Tan & Flores, 2020) while others used no-RBAC invention as controls.
Survival rate analysis
Three RCTs reported the survival outcomes of participants (Bang et al., 2010; Takahara & Sano, 2004; Tan & Flores, 2020). Figure 5 shows a forest plot of the survival event analysis from these studies. Based on survival data (n=133) from patients with lung cancer (n=68) of Bang et al. (2010) and head and neck cancer (n=65) from Tan and Flores (2020), the OR for RBAC treatment compared to control for one year or less is 4.02 (95% CI: 1.67, 9.69) in favour of RBAC. Thus, cancer patients under RBAC treatment have 4.02 times better odds of surviving one year or less than control.
Meta-analysis of data with durations between 18 to 24 months shows that the OR of survival in the RBAC group is 5.86 (95% CI: 3.24, 10.58) over control based on intention-to-treat analysis that includes all dropouts (n=273, RBAC=134, control=139). Excluding dropouts (n=53 from the control group), the OR per protocol analysis is lower at 2.89 (95% CI: 1.56, 5.35) based on data from 223 participants (RBAC=134, control=86). The study by Takahara and Sano (2004) is larger (n=205) and included adult (mean age=54.7) progress-cancer patients in the late III-IV stages of different malignancies (lung, liver, uterus, breast, prostate, rectum, stomach, lymph node, and others) who had completed conventional treatment. Hence, the meta-analysis shows that adult cancer patients, particularly those with lung cancer or in later stages (II-IV), are at least 2.89 times more likely to survive longer than 18 to 24 months after starting RBAC treatment than those not treated with RBAC.
Only Bang et al. (2010) reported survival data up to 36 months, with the OR of RBAC over control being 7.96 (95% CI: 0.41, 153.86). However, these values are not statistically significant since the lower values of 95% CI are less than 1.0. With only a small number of lung cancer participants surviving up to 36 months, a large sample size is needed to confirm the life-prolonging effect of RBAC for the longer term.
Quality of life assessment
Participant QoL was assessed in five of the included RCTs (Itoh et al., 2015; Masood et al., 2013; Petrovics et al., 2016; Takahara & Sano, 2004; Tan & Flores, 2020), albeit with different outcome measures. A bubble chart for visualisation of the available evidence is shown in Figure 6. Each bubble in the plot represents an outcome assessed in one study, with the study sample size defining the bubble’s diameter. Also, each study is assigned a different colour.
The most significant results were from Masood et al. (2013), with RBAC markedly (p<0.01) improved anorexia, alopecia, nausea, and weight loss in breast cancer patients during chemotherapy compared to control. Takahara and Sano (2004) also stated improvement in appetite after RBAC treatment but reported no p-value and data for statistical significance estimation, which weakened the claim. For nausea, unlike Masood et al. (2013), both Takahara and Sano (2004) and Itoh et al. (2015) found no significant difference between the RBAC and control groups post-treatment. Moreover, Tan and Flores (2020) also found no between-group difference in weight loss among head and neck cancer patients two months after chemoradiotherapy.
Among cancer patients with CFS (n=48), Petrovics et al. (2016) confirmed that the combination therapy of RBAC and oncothemia significantly lowered (p<0.01) fatigue scores compared to control based on two validated instruments (CFQ and PGIC). However, other beneficial QoL outcomes stated by Petrovics et al. (2016) were improvement in pain (based on a visual analogue scale), general QoL, physical and emotional functioning (based on the European Organisation for Research and Treatment of Cancer’s [EORTC] Quality of Life Questionnaire version 3.0 [QLQ-C3]). Although the outcomes were assessed with validated instruments, the lack of statistical analysis for these items in the published report rendered the assertion or benefits unconvincing.
Tan and Flores (2020) also reported general QoL outcome improvement by RBAC treatment based on the EORTC QoL instrument specific for head and neck cancer (QLQ-H&N35). The mean QoL scores of patients treated with RBAC were significantly better than the control group two months after chemoradiotherapy (p<0.05). Nevertheless, the study did not detect any between-group difference in radiation toxicities. Likewise, no between-group differences were reported for pain and malaise by Takahara and Sano (2004) and diarrhea by Itoh et al. (2015).
Overall, QoL enhancement by RBAC was shown only in selected types of cancer and for specific measures, most notably in reduction of anorexia, alopecia, nausea, and weight loss in breast cancer patients during chemotherapy (Masood et al., 2013), improvement of general QoL for head and neck cancer patients after radiation (Tan & Flores, 2020) and ameliorating the fatigue syndrome in cancer patients with CFS during oncological treatment when used together with oncothermia (Petrovics et al., 2016). However, it remains unclear whether these QoL benefits apply to other cancer patients.
Quality assessment of the included studies
The quality assessment results for each study are summarised in Figure 7. None of the studies were considered good quality in the assessors’ opinion. Three studies, namely Bang et al. (2010), Itoh et al. (2015), and Tan and Flores (2020), are rated as fair quality as these studies adequately adhered to some of the required standards in reporting the RCT results to allow for a level of confidence on the study’s internal validity. The remaining three studies are considered poor quality. Takahara and Sano (2004), in particular, suffer from high dropout rates and unclear baseline patient characteristics and allocation methods. For Masood et al. (2013), many required items were not reported, which placed the study’s internal validity in doubt. As for Petrovics et al. (2016), many of the findings, such as the QoL responses from the participants, were not presented correctly, making it difficult to compare outcomes between groups.
Based on the quality assessment outcomes, the risk of bias in all included studies is summarised in Figure 8. Overall, these RCTs are assessed to have a low risk of reporting bias. The assessors find all studies use valid and reliable outcome measures, and the intention-to-treat analysis has been adequately carried out and documented. However, the risk of selection, performance, and detection biases are moderate. Although all studies were randomised trials, most did not report how random sequence generation was performed and whether there was any allocation concealment. There was some potential risk of attrition bias as there were high dropout rates in two studies (Itoh et al., 2015; Takahara & Sano, 2004). The risk of other biases also exists with the lack of specific power calculations to detect a between-group difference in all these studies. Also, all studies did not report any data on adherence to treatment.
Discussion
This review identifies RBAC as an immunomodulating natural substance capable of restoring immune dysfunction due to tumour development. Specifically, by increasing the NKC granularity and cytolytic activity, RBAC could improve the immunosurveillance ability of the host to destroy malignant cells (Cholujova et al., 2013; Ghoneum & Brown, 1999). These findings are consistent with the numerous in vivo and in vitro evidence that confirms the capacity of RBAC to enhance the NKC activity in healthy subjects (Badr El-Din et al., 2008; Ghoneum, 1998; Ghoneum & Abedi, 2004; Ghoneum & Jewett, 2000; Giese et al., 2008; Kim et al., 2007; Pérez-Martínez et al., 2015). Hence, RBAC could be a potential natural product candidate for NKC immunotherapy, which seeks to alleviate the inhibited state of NKC in the tumour microenvironment (Du et al., 2021). The existing NKC therapy includes allogeneic NKC infusion, cytokine treatment (IL-2 and IL-5), and immune checkpoint inhibitors (Du et al., 2021). Ghoneum (2023), who has extensively investigated the NKC augmentation capacity of RBAC, proclaims the superiority of RBAC, compared to other NKC therapies, in terms of the duration and extent of NKC activation, as well as the lack of hyperresponsiveness overtime. However, such an assertion requires further investigation as this study found no published human trials comparing the effects of RBAC to any existing NKC therapy.
Other immune restorative effects of RBAC include promoting the increase of myeloid DC and CD4+ Th cell counts while lowering the level of the immunosuppressive Treg (Lissoni et al., 2008). As Treg is known to promote tumour cell proliferation through inhibiting antigen-presenting cells such as the myeloid DC (Shohei & Hiroyoshi, 2021), reducing the abundance of Treg could increase activation of CD4+ Th cells by matured myeloid DC, which in turn could prime the CD8+ T cells for antitumour activity in cancer patients. Earlier research has shown that RBAC could stimulate the maturation of DC in healthy hosts by downregulating the immature surface markers (CD14, CD1a, and CD11c), reducing endocytic activities, while upregulating the maturation markers (CD12 and CD80) (Cholujova et al., 2009; Ghoneum & Agrawal, 2011, 2014). Healthy CD8+ T cells cultured with RBAC-activated DC also had increased cytotoxicity against tumour cell targets compared to those treated with unstimulated DC (Ghoneum & Agrawal, 2014). Hence, an alternative pathway that RBAC could exert the anticancer immune response is via indirect activation of cytotoxic CD8+ T cells—the most potent effectors among all the immune cells to be harnessed in immunotherapy (Raskov et al., 2021). CD8+ T cells and NKC are parallel and complementary cytotoxic effectors crucial for the immunosurveillance of tumour cells (Rosenberg & Huang, 2018). The depletion of CD8+ T cells and NKC can lead to the escape of CTCs to form metastasis as CTC count is negatively correlated with the lymphocyte ratios of CD4+/CD8+ and NKC in late-stage cancer patients (Ye et al., 2017). Therefore, RBAC could augment immunosurveillance to prevent cancer metastasis by lowering CTCs (Pescatore et al., 2022).
Beyond immunomodulation, RBAC also possesses direct anticancer effects achieved through increased susceptibility of CD95 (Fas/APO-1) ligands on cancerous cells to extracellular apoptotic signals (the extrinsic pathway) and downregulating anti-apoptotic Bcl-2 proteins (the intrinsic pathway) to lower mitochondrial membrane potentials while upregulating the tumour suppressing P53 gene, and increasing Bax and caspase-3 proteins to initiate programmed cell death (Badr El-Din, Abdel Fattah, et al., 2016; Badr El-Din et al., 2020; Ghoneum & Gollapudi, 2003). Hence, RBAC is not cytotoxic but rather proapoptotic to malignant cells. Conventional antineoplastic agents can be classified into many families. The first group are structural analogues of nucleobases, nucleosides, or folic analogues called antimetabolites. Antimetabolites act as decoys to inhibit the synthesis of DNA components (Guichard et al., 2017). The second family includes DNA-interactive molecules, such as alkylating agents, which directly alter DNA replication and transcription processes. The third group consists of those acting on mitosis, such as antitubulin agents that cause cell death by blocking the division of the nucleus (Guichard et al., 2017). Other drug families include antitumour antibiotics, such as anthracyclines, that block DNA to RNA transcription (Martins-Teixeira & Carvalho, 2020); topoisomerase inhibitors that stop DNA unwinding (Kim & Khan, 2020); and tyrosine kinase inhibitors which suppress growth factor receptors (Huang et al., 2020). However, RBAC does not belong to any of these classes of conventional antineoplastic agents but could be a member of a promising new class of plant-derived anticancer therapies that activate the apoptotic pathway (Pfeffer & Singh, 2018).
Unlike most antineoplastic agents, which affect malignant and fast-growing normal cells, RBAC’s proapoptotic effect does not cause normal cell death unnecessarily, as it has been shown to be non-cytotoxic (An, 2011). As most antineoplastic drugs exploit the intact mitochondrial apoptotic signalling pathways to trigger cancer cell death, RBAC could improve the efficacy of these antineoplastic agents and thus prevent tumour resistance to therapies (Pistritto et al., 2016). It is not surprising that RBAC was shown to work synergistically with several commonly used antineoplastic agents, including daunorubicin and paclitaxel, to increase the efficacy of chemotherapy (Badr El-Din, Ali, Alaa El-Dein, et al., 2016; Ghoneum et al., 2014; Gollapudi & Ghoneum, 2008). RBAC may thus allow lower drug concentrations in chemotherapy, reducing cytotoxicity and unwanted side effects while achieving similar therapeutic objectives.
RBAC can also benefit cancer treatment through the antioxidant pathways to protect healthy tissues against increased oxidative stress from carcinogens, tumourigenesis, antineoplastic agents, and radiation treatment (Badr El-Din, Abdel Fattah, et al., 2016; Badr El-Din, Ali, & Othman, 2016; Badr El-Din et al., 2019; Endo & Kanbayashi, 2003; Ghoneum et al., 2013; Jacoby et al., 2001; Zhao et al., 2020). This review found evidence that RBAC could upregulate the endogenous GSH and antioxidant enzymes (GPx, GST, SOD, and CAT) in cancer and healthy tissues in vivo (Noaman et al., 2008). Other studies also showed RBAC to have potent scavenging capacities towards ROS with an Oxygen Radical Absorption Capacity level higher than broccoli, a known high-antioxidant food (An, 2011; Tazawa et al., 2000). The antioxidant effects of RBAC have also been shown to protect against neurodegeneration due to sporadic Alzheimer’s disease in a mice model (Ghoneum & El Sayed, 2021). Interestingly, RBAC’s protective effect against brain tissue damage was via the suppression of amyloid-beta-induced apoptosis through upregulation of the antiapoptotic protein Bcl-2 while downregulating the proapoptotic protein Bax and caspase-3 cleavage (Ghoneum & El Sayed, 2021). Therefore, the effect of RBAC on the apoptosis pathway is selective and works against malignant cells but protects healthy cells, most likely through the antioxidant activity which delays and inhibits cell damage (Redza-Dutordoir & Averill-Bates, 2016).
With immune restorative, proapoptotic, and antioxidant effects, RBAC was shown to work synergistically with other complementary therapies, such as Baker’s yeast, curcumin, mistletoe lectin, and oncothermia, to improve treatment effects in cancer (Ghoneum & Gollapudi, 2005a, 2005b, 2011; Hajtó, Horváth, & Papp, 2016; Petrovics et al., 2016). Hence, RBAC could be a highly versatile plant-based therapeutic option in cancer treatment. An approach that combined RBAC with mistletoe lectin and wheat germ extract as plant-based immunomodulators to increase the sensitivity of tumour cells to low-dose anticancer drugs (such as gemcitabine or growth factor inhibitors) is being advocated by Hajtó et al. (2013) as a novel oncological strategy. Favourable clinical responses of the combined treatment in different cancer metastases, including breast (Hajtó, 2018; Hajtó & Kirsch, 2013), colon (Hajtó & Kirsch, 2013), ovarian (Hajtó, 2018; Hajtó et al., 2015), lung (Hajtó, Horváth, Baranyai, et al., 2016), and bile duct (Hajtó, 2017), have been reported. However, good quality research evidence remains lacking. Hajtó (2023) claimed difficulty obtaining permission for clinical trials even though RBAC is considered “the most supported evidence-based and standardised plant immunomodulator without any side effects” as the compound remains registered as a food supplement and not an oncological therapy, which hindered clinical research (p. 71).
The lack of RCTs severely restricts RBAC’s application in evidence-based practice. This review found only six RCTs that evaluated survival and/or QoL cancer patients with RBAC as an intervention. There is evidence that RBAC could improve the survival rate of cancer patients (OR=2.89, 95% CI: 1.56, 5.35) for the first two years, compared to no RBAC. However, with only limited data (3 studies, n=338), the confidence in this OR is low. Further research will very likely change the estimate. Regarding QoL, there is evidence from small RCTs showing that RBAC could reduce fatigue and side effects of chemotherapy (anorexia, alopecia, nausea, and weight loss) and improve general QoL after radiation treatment. However, again, with results obtained from only a small number of trials conducted on patients with specific cancer types and conditions, to draw any conclusion on RBAC’s effects on cancer patients’ QoL, in general, is premature. Moreover, there could be risks of bias in many of these trials, with half of the included RCTs regarded as poor quality. Note that this review’s risk of bias assessment is consistent with a previous evaluation that all RBAC RCTs have either unclear or high risk of selection, performance, detection, attrition, and other biases (Ooi et al., 2018). Hence, the current level of evidence for RBAC as a credible oncological therapeutic option remains low. Notwithstanding, RBAC is considered safe to consume at the typical dosage of 1-3g/day, with no RBAC-related adverse events reported in any of the human studies.
Moving forward, RBAC research in cancer should focus on two areas. The first is to identify, quantify and standardise the active ingredients of RBAC. Miura et al. (2004/2013) had previously identified the immune active compounds in RBAC, based on macrophage activation, to be complex heteropolysaccharides with arabinoxylan as its primary structure while also containing galactan and glucan. However, no follow-up research was conducted to ascertain the detailed molecular structure of the compound nor the components that are most relevant in activating NKC, promoting apoptosis in cancer cells, and scavenging free radicals. For example, could RBAC’s antioxidant capacity be due to the contents of ferulic acids or γ-oryzanol commonly found in rice bran?—an intriguing hypothesis that needs investigation. Most importantly, clarifying the active ingredient in RBAC can also lead to an in-depth understanding of the molecular pathways in which RBAC induces the cellular mechanisms such as proapoptotic cascade or myeloid DC maturation in the tumour microenvironment.
The second focus area in RBAC research should be clinical trials. There is a need for more well-designed RCTs to substantiate the therapeutic use of RBAC in cancer treatment for various cancer types with different therapeutic concomitants. These RCTs should have a sufficient sample size to detect the effects of outcomes in terms of treatment efficacies and side effects. Studies with longer-term follow-up periods of three years or more are also needed to confirm RBAC’s long-term impact on recurrence, QoL, and survival. Only with sufficient high-quality, favourable evidence from RCTs could RBAC become mainstream in oncological treatment.
Conclusion
Current preclinical and clinical research evidence suggests RBAC is a natural product with immense potential in cancer treatment. RBAC could be an immunomodulator priming the cytotoxic immune response against tumorigenesis, promoting apoptosis in malignant cells. Working in synergy with chemoradiation therapies, RBAC could potentially increase treatment effectiveness while reducing side effects, improving patients’ QoL, and prolonging survival. Notwithstanding, more research evidence is needed before RBAC can be considered a viable therapeutical option in cancer.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org
Author Contributions
SLO: Conceptualisation, Methodology, Software, Formal analysis, Investigation, Data Curation, Writing—Original Draft, Visualisation. PSM: Conceptualisation, Methodology, Validation, Writing—Review & Editing. JK: Resources, Writing—Review & Editing. SCP: Conceptualisation, Methodology, Validation, Resources, Writing—Review & Editing, Supervision, Project administration.
Funding
This review received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors. SLO is a recipient of the Australian Government Research Training Program scholarship for his PhD study.
Declarations
This study contributes toward the PhD degree for SLO.
Conflicts of Interest
JK is an employee of is an employee of STR Biotech Co., Ltd., which develops and markets variants of RBAC products commercially. The other authors declare no conflicts of interest.
References
- Abdalla, Y.O.A.; Subramaniam, B.; Nyamathulla, S.; Shamsuddin, N.; Arshad, N.M.; Mun, K.S.; Awang, K.; Nagoor, N.H. Natural Products for Cancer Therapy: A Review of Their Mechanism of Actions and Toxicity in the Past Decade. J. Trop. Med. 2022, 2022, 1–20. [Google Scholar] [CrossRef]
- Alshaibi, H.F.; Al-Shehri, B.; Hassan, B.; Al-Zahrani, R.; Assiss, T. Modulated Electrohyperthermia: A New Hope for Cancer Patients. BioMed Res. Int. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Altun, I.; Sonkaya, A. The Most Common Side Effects Experienced by Patients Were Receiving First Cycle of Chemotherapy. 2018, 47, 1218–1219.
- Outcomes of cancer treatment for technology assessment and cancer treatment guidelines. American Society of Clinical Oncology. J. Clin. Oncol. 1996, 14, 671–679. [Google Scholar] [CrossRef]
- Amjad, M. T. , Chidharla, A., & Kasi, A. (2023, 27 Feb, 2023). Cancer chemotherapy. StatPearls Publishing. https://www.ncbi.nlm.nih. 5643. [Google Scholar]
- An, S. Y. (2011). Immune-enhance and anti-tumor effect of exo-biopolymer extract from submerged culture of Lentinus edodes with rice bran [Doctoral Thesis, Korea University]. Seoul, Republic of Korea.
- Andocs, G.; Szasz, O.; Szasz, A. Oncothermia Treatment of Cancer: From the Laboratory to Clinic. Electromagn. Biol. Med. 2009, 28, 148–165. [Google Scholar] [CrossRef]
- Arruebo, M.; Vilaboa, N.; Sáez-Gutierrez, B.; Lambea, J.; Tres, A.; Valladares, M.; González-Fernández, Á. Assessment of the Evolution of Cancer Treatment Therapies. Cancers 2011, 3, 3279–3330. [Google Scholar] [CrossRef]
- El-Din, N.K.B.; Fattah, S.M.A.; Pan, D.; Tolentino, L.; Ghoneum, M. Chemopreventive Activity of MGN-3/Biobran Against Chemical Induction of Glandular Stomach Carcinogenesis in Rats and Its Apoptotic Effect in Gastric Cancer Cells. Integr. Cancer Ther. 2016, 15, NP26–NP34. [Google Scholar] [CrossRef]
- El-Din, N.K.B.; Ali, D.A.; El-Dein, M.A.; Ghoneum, M. Enhancing the Apoptotic Effect of a Low Dose of Paclitaxel on Tumor Cells in Mice by Arabinoxylan Rice Bran (MGN-3/Biobran)*. Nutr. Cancer 2016, 68, 1010–1020. [Google Scholar] [CrossRef]
- El-Din, N.K.B.; Ali, D.A.; Othman, R.M. INHIBITION OF EXPERIMENTAL CARCINOGENESIS BY THE BIOACTIVE NATURAL PRODUCT BIOBRAN. J. Plant Prot. Pathol. 2016, 7, 85–91. [Google Scholar] [CrossRef]
- El-Din, N.K.B.; Ali, D.A.; Othman, R.; French, S.W.; Ghoneum, M. Chemopreventive role of arabinoxylan rice bran, MGN-3/Biobran, on liver carcinogenesis in rats. Biomed. Pharmacother. 2020, 126, 110064. [Google Scholar] [CrossRef]
- El-Din, N.K.B.; Areida, S.K.; O Ahmed, K.; Ghoneum, M. Arabinoxylan rice bran (MGN-3/Biobran) enhances radiotherapy in animals bearing Ehrlich ascites carcinoma†. J. Radiat. Res. 2019, 60, 747–758. [Google Scholar] [CrossRef] [PubMed]
- El-Din, N.K.B.; Mahmoud, A.Z.; Hassan, T.A.; Ghoneum, M. Baker’s Yeast Sensitizes Metastatic Breast Cancer Cells to Paclitaxel In Vitro. Integr. Cancer Ther. 2017, 17, 542–550. [Google Scholar] [CrossRef] [PubMed]
- El-Din, N.K.B.; Noaman, E.; Ghoneum, M. In Vivo Tumor Inhibitory Effects of Nutritional Rice Bran Supplement MGN-3/Biobran on Ehrlich Carcinoma-Bearing Mice. Nutr. Cancer 2008, 60, 235–244. [Google Scholar] [CrossRef]
- The Effects of the Arabinoxylane and the Polysaccharide Peptide (PSP) on the Antiallergy, Anticancer. J. Korean Soc. Food Sci. Nutr. 2004, 33, 469–474. [CrossRef]
- Bang, M.H.; Van Riep, T.; Thinh, N.T.; Song, L.H.; Dung, T.T.; Van Truong, L.; Van Don, L.; Ky, T.D.; Pan, D.; Shaheen, M.; et al. Arabinoxylan rice bran (MGN-3) enhances the effects of interventional therapies for the treatment of hepatocellular carcinoma: a three-year randomized clinical trial. . 2010, 30, 5145–51. [Google Scholar]
- Barnes, J.L.; Zubair, M.; John, K.; Poirier, M.C.; Martin, F.L. Carcinogens and DNA damage. Biochem. Soc. Trans. 2018, 46, 1213–1224. [Google Scholar] [CrossRef]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.-W. Cancer and Radiation Therapy: Current Advances and Future Directions. Int. J. Med Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef]
- Blackadar, C.B. Historical review of the causes of cancer. World J. Clin. Oncol. 2016, 7, 54–86. [Google Scholar] [CrossRef]
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef]
- Brush, T.P.; Trinh, S.; Brawner, C.M.; Ali, K.H.; Hauke, R.J.; Elkahwaji, J.E. Abstract 5662: RBAC, a modified form of Arabinoxylan from rice bran, impairs prostate cancer cell line proliferation, adhesion, and invasion in vitro. Cancer Res 2010, 70, 5662–5662. [Google Scholar] [CrossRef]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Carvalho, A.S.; Lagana, D.; Catford, J.; Shaw, D.; Bak, N. Bloodstream infections in neutropenic patients with haematological malignancies. Infect. Dis. Heal. 2019, 25, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A. Reduced Glutathione: A Radioprotector or a Modulator of DNA-Repair Activity? Nutrients 2013, 5, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Sobenin, I.A.; Orekhov, A.N.; Bobryshev, Y.V. Myeloid dendritic cells: Development, functions, and role in atherosclerotic inflammation. Immunobiology 2015, 220, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Cholujova, D.; Jakubikova, J.; Czako, B.; Martisova, M.; Hunakova, L.; Duraj, J.; Mistrik, M.; Sedlak, J. MGN-3 arabinoxylan rice bran modulates innate immunity in multiple myeloma patients. Cancer Immunol. Immunother. 2012, 62, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Cholujova, D.; Jakubikova, J.; Sedlak, J. BioBran-augmented maturation of human monocyte-derived dendritic cells. Neoplasma 2009, 56, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Clark, H. R. (1999, May-1999). The natural pharmacy. Winning the war against cancer: MGN-3: mushroom ammunition. Alternative Medicine Magazine, (29), 26-28.
- Cooper, G. M. (2000). The development and causes of cancer. In The Cell: A Molacular Approach. (2nd ed.). Sinauer Associates.
- Dare, A. J. , Anderson, B. O., Sullivan, R., Pramesh, C. S., Andre, I., Adewole, I. F., Badwe, R. A., & Gauvreau, C. L. (2015). Surgical services for cancer care. In H. Gelband, P. Jha, R. Sankaranarayanan, & S. Horton (Eds.), Cancer: Disease Control Priorities (3rd ed., Vol. 3). The International Bank for Reconstruction and Development / The World Bank. [CrossRef]
- Deeks, J. J. , Higgins, J. P., & Altman, D. G. (2021). Chapter 10: Analysing data and undertaking meta-analyses. In J. Higgins, J. Thomas, J. Chandler, M. Cumpston, T. Li, M. Page, & V. Welch (Eds.), Cochrane Handbook for Systematic Reviews of Interventions version 6.2 (updated 21). Cochrane. 20 February.
- Delaney, G.; Barton, M. Evidence-based Estimates of the Demand for Radiotherapy. Clin. Oncol. 2014, 27, 70–76. [Google Scholar] [CrossRef]
- Deng, Z.; Wu, S.; Wang, Y.; Shi, D. Circulating tumor cell isolation for cancer diagnosis and prognosis. EBioMedicine 2022, 83, 104237. [Google Scholar] [CrossRef]
- Du, N.; Guo, F.; Wang, Y.; Cui, J. NK Cell Therapy: A Rising Star in Cancer Treatment. Cancers 2021, 13, 4129. [Google Scholar] [CrossRef]
- Endo, Y.; Kanbayashi, H. Modified Rice Bran Beneficial for Weight Loss of Mice as a Major and Acute Adverse Effect of Cisplatin. Basic Clin. Pharmacol. Toxicol. 2003, 92, 300–303. [Google Scholar] [CrossRef]
- Fan, H.; Walters, C.S.; Dunston, G.M.; Tackey, R. IL-12 PLAYS A SIGNIFICANT ROLE IN THE APOPTOSIS OF HUMAN T CELLS IN THE ABSENCE OF ANTIGENIC STIMULATION. Cytokine 2002, 19, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.T.; Sakamuru, S.; Huang, R.; Teneva, N.; Simmons, S.O.; Xia, M.; Tice, R.R.; Austin, C.P.; Myung, K. High-throughput genotoxicity assay identifies antioxidants as inducers of DNA damage response and cell death. Proc. Natl. Acad. Sci. 2012, 109, 5423–5428. [Google Scholar] [CrossRef]
- Fridrichova, I.; Kalinkova, L.; Ciernikova, S. Clinical Relevancy of Circulating Tumor Cells in Breast Cancer: Epithelial or Mesenchymal Characteristics, Single Cells or Clusters? Int. J. Mol. Sci. 2022, 23, 12141. [Google Scholar] [CrossRef]
- G. , M.S.; Swetha, M.; Keerthana, C.K.; Rayginia, T.P.; Anto, R.J. Cancer Chemoprevention: A Strategic Approach Using Phytochemicals. Front. Pharmacol. 2022, 12, 809308. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, G.; Szasz, O.; Szasz, A. A New Paradigm and Promising Method in Cancer Therapies Oncothermia: a New Paradigm and Promising Method in Cancer Therapies. Acupunct. Electro-Therapeutics Res. 2013, 38, 161–197. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M. (1998). Enhancement of human natural killer cell activity by modified arabinoxylane from rice bran (MGN-3). International Journal of Immunotherapy, 14(2), 89-99.
- Ghoneum, M. (1999, Dec 11-13). Immunostimulation and cancer prevention [Abstract]. 7th International Congress on Anti-Aging & Biomedical Technologies, Las Vegas, NV, USA.
- Ghoneum, M. (2000, Jan). One sizeable step for immunology, one giant leap for cancer patients. Townsend Letter for Doctors and Patients, (198), 58-62.
- Ghoneum, M. (2016). From bench to bedside : The growing use of arabinoxylan rice bran (MGN-3/Biobran) in cancer immunotherapy. Austin Immunology, 1(2), 1006.
- Ghoneum, M.; Abdulmalek, S.; Fadel, H.H. Biobran/MGN-3, an Arabinoxylan Rice Bran, Protects against Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): An In Vitro and In Silico Study. Nutrients 2023, 15, 453. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; Abedi, S. Enhancement of natural killer cell activity of aged mice by modified arabinoxylan rice bran (MGN-3/Biobran). J. Pharm. Pharmacol. 2004, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; Agrawal, S. Activation of Human Monocyte-Derived Dendritic Cells in Vitro by the Biological Response Modifier Arabinoxylan Rice Bran (MGN-3/BIOBRAN). Int. J. Immunopathol. Pharmacol. 2011, 24, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; Agrawal, S. MGN-3/Biobran Enhances Generation of Cytotoxic CD8+ T Cells VIA Upregulation of DEC-205 Expression on Dendritic Cells. Int. J. Immunopathol. Pharmacol. 2014, 27, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; El-Din, N.K.B.; Fattah, S.M.A.; Tolentino, L. Arabinoxylan rice bran (MGN-3/Biobran) provides protection against whole-body -irradiation in mice via restoration of hematopoietic tissues. J. Radiat. Res. 2013, 54, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Ghoneum, M.; El-Din, N.K.B.; Ali, D.A.; El-Dein, M.A. Modified arabinoxylan from rice bran, MGN-3/biobran, sensitizes metastatic breast cancer cells to paclitaxel in vitro. . 2014, 34, 81–7. [Google Scholar]
- Ghoneum, M. , & Brown, J. (1999). NK Immunorestoration and cancer patients by MGN-3, a modified arabinoxylan rice bran (Study of 32 patients followed for up to 4 years). In R. M. Klatz & R. Goldman (Eds.), Anti-aging Medical Therapeutics (Vol. III, pp. 217-226). Health Quest Publications.
- Ghoneum, M.H.; El Sayed, N.S. Protective Effect of Biobran/MGN-3 against Sporadic Alzheimer’s Disease Mouse Model: Possible Role of Oxidative Stress and Apoptotic Pathways. Oxidative Med. Cell. Longev. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Ghoneum, M.; Gollapudi, S. Modified arabinoxylan rice bran (MGN-3/Biobran) sensitizes human T cell leukemia cells to death receptor (CD95)-induced apoptosis. Cancer Lett. 2003, 201, 41–49. [Google Scholar] [CrossRef]
- Ghoneum, M. , & Gollapudi, S. (2004). Induction of apoptosis in breast cancer cells by saccharomyces cerevisiae the baker’s yeast, in vitro. Anticancer Research, 24(3A), 1455.
- Ghoneum, M. , & Gollapudi, S. (2005a). Modified arabinoxylan rice bran (MGN-3/Biobran) enhances yeast-induced apoptosis in human breast cancer cells in vitro. Anticancer Research, 25(2a), 859-870.
- Ghoneum, M.; Gollapudi, S. Synergistic role of arabinoxylan rice bran (MGN-3/Biobran) in S. cerevisiae-induced apoptosis of monolayer breast cancer MCF-7 cells.. 2005, 25, 4187–96. [Google Scholar]
- Ghoneum, M.; Gollapudi, S. Synergistic apoptotic effect of arabinoxylan rice bran (MGN-3/Biobran) and curcumin (turmeric) on human multiple myeloma cell line U266 in vitro. Neoplasma 2011, 58, 118–123. [Google Scholar] [CrossRef]
- Ghoneum, M.; Jewett, A. Production of tumor necrosis factor-alpha and interferon-gamma from human peripheral blood lymphocytes by MGN-3, a modified arabinoxylan from rice bran, and its synergy with interleukin-2 in vitro. . 2000, 24, 314–24. [Google Scholar]
- Ghoneum, M.; Matsuura, M.; Gollapudp, S. Modified Arabinoxylan Rice Bran (Mgn-3/Biobran) Enhances Intracellular Killing of Microbes by Human Phagocytic Cellsin Vitro. Int. J. Immunopathol. Pharmacol. 2008, 21, 87–95. [Google Scholar] [CrossRef]
- Ghoneum, M. , Tachiki, K. H., Ueyama, K., Makinodan, T., Makhijani, N., & Yamaguchi, D. (2000, Dec 14-17). Natural biological response modifier (MGN-3) shown to be effective against tumor cell growth [Abstract]. 8th International Congress on Anti-Aging & Biomedical Technologies, Las Vegas, NV, USA.
- Ghoneum, M. H. (2023). Enhancing natural killer cell activity. In S. C. Pak, S. L. Ooi, P. S. Micalos, & M. H. Ghoneum (Eds.), Modified Rice Bran Arabinoxylan : Therapeutic Applications in Cancer and Other Diseases (pp. 15-25). Springer Nature. [CrossRef]
- Giese, S.; Sabell, G.R.; Coussons-Read, M. Impact of Ingestion of Rice Bran and Shitake Mushroom Extract on Lymphocyte Function and Cytokine Production in Healthy Rats. J. Diet. Suppl. 2008, 5, 47–61. [Google Scholar] [CrossRef]
- Gollapudi, S.; Ghoneum, M. MGN-3/Biobran, modified arabinoxylan from rice bran, sensitizes human breast cancer cells to chemotherapeutic agent, daunorubicin. Cancer Detect. Prev. 2008, 32, 1–6. [Google Scholar] [CrossRef]
- Golombick, T.; Diamond, T.H.; Manoharan, A.; Ramakrishna, R. Addition of Rice Bran Arabinoxylan to Curcumin Therapy May Be of Benefit to Patients With Early-Stage B-Cell Lymphoid Malignancies (Monoclonal Gammopathy of Undetermined Significance, Smoldering Multiple Myeloma, or Stage 0/1 Chronic Lymphocytic Leukemia). Integr. Cancer Ther. 2016, 15, 183–189. [Google Scholar] [CrossRef]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: from tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef]
- Götze, H.; Friedrich, M.; Taubenheim, S.; Dietz, A.; Lordick, F.; Mehnert, A. Depression and anxiety in long-term survivors 5 and 10 years after cancer diagnosis. Support. Care Cancer 2019, 28, 211–220. [Google Scholar] [CrossRef]
- Guan, X. Cancer metastases: challenges and opportunities. Acta Pharm. Sin. B 2015, 5, 402–418. [Google Scholar] [CrossRef]
- Guichard, N.; Guillarme, D.; Bonnabry, P.; Fleury-Souverain, S. Antineoplastic drugs and their analysis: a state of the art review. Anal. 2017, 142, 2273–2321. [Google Scholar] [CrossRef]
- Hajto, T. Can a standardized plant immunomodulator (rice bran arabinoxylan concentrate/MGN-3) increase the effects of MEK and BRAF inhibitors with clinical benefit? Case report of a patient with carcinoma in biliary duct. Res. Rev. Insights 2017, 1. [Google Scholar] [CrossRef]
- Hajtó, T. (2018). New perspectives to improve the MHC-I unrestricted immune mechanisms against malignant tumors. Advances in Clinical and Translational Research, 2(3), 100014.
- Hajtó, T. (2023). The therapeutic application of RBAC in cancer. In S. C. Pak, S. L. Ooi, P. S. Micalos, & M. H. Ghoneum (Eds.), Modified Rice Bran Arabinoxylan : Therapeutic Applications in Cancer and Other Diseases (pp. 67-78). Springer Nature. [CrossRef]
- Hajtó, T. , Adámy, A., Langmár, Z., Kirsch, A., Ábrahám, L., Perjési, P., & Németh, P. (2013). Enhanced effectiveness of conventional oncotherapy with plant immunomodulators: Overview of recent advances. Advancement in Medicinal Plant Research, 1(3), 56-65.
- Hajto, T.; Baranyai, L.; Kirsch, A.; Kuzma, M.; Perjési, P. Can a synergistic activation of pattern recognition receptors by plant immunomodulators enhance the effect of oncologic therapy? Case Report of a patient with uterus and ovary sarcoma. Clin. Case Rep. Rev. 2015, 1. [Google Scholar] [CrossRef]
- Hajto, T.; Horváth, A.; Baranyai, L.; Kuzma, M.; Perjési, P. Can the EGFR inhibitors increase the immunomodulatory effects of standardized plant extracts (mistletoe lectin and arabonoxylan) with clinical benefit? Case report of a patient with lung adenocarcinoma. Clin. Case Rep. Rev. 2016, 2. [Google Scholar] [CrossRef]
- Hajto, T.; Horvath, A.; Papp, S. Improvement of Quality of Life in Tumor Patients after an Immunomodulatory Treatment with Standardized Mistletoe Lectin and Arabinoxylan Plant Extracts. Int. J. Neurorehabilit. 2016, 03. [Google Scholar] [CrossRef]
- Hajto, T.; Kirsch, A. Case Reports of Cancer Patients with Hepatic Metastases Treated by Standardized Plant Immunomodulatory Preparations. J. Cancer Res. Updat. 2013, 2. [Google Scholar] [CrossRef]
- Tibor, H.; László, P.; Monika, K.; Edina, H.; Tamás, O.; Pál, J.; Lilla, B.; Zoltán, K. Enhancing effect of streptozotocin-induced insulin deficit on antitumor innate immune defense in rats. Res. Rev. Insights 2021, 6. [Google Scholar] [CrossRef]
- Haque, A.; Brazeau, D.; Amin, A.R. Perspectives on natural compounds in chemoprevention and treatment of cancer: an update with new promising compounds. Eur. J. Cancer 2021, 149, 165–183. [Google Scholar] [CrossRef]
- Hariton, E.; Locascio, J.J. Randomised controlled trials—the gold standard for effectiveness research: Study design: Randomised controlled trials. BJOG 2018, 125, 1716. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative Stress in Cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Huang, L.; Jiang, S.; Shi, Y. Tyrosine kinase inhibitors for solid tumors in the past 20 years (2001–2020). J. Hematol. Oncol. 2020, 13, 1–23. [Google Scholar] [CrossRef]
- Igari, N. (2020). Biobran, immuno modulating rice bran arabinoxylan compound: functions and evidence. Food processing and ingredients [食品と開発], 55(10), 24-26.
- Itoh, Y.; Mizuno, M.; Ikeda, M.; Nakahara, R.; Kubota, S.; Ito, J.; Okada, T.; Kawamura, M.; Kikkawa, F.; Naganawa, S. A Randomized, Double-Blind Pilot Trial of Hydrolyzed Rice Bran versus Placebo for Radioprotective Effect on Acute Gastroenteritis Secondary to Chemoradiotherapy in Patients with Cervical Cancer. Evidence-Based Complement. Altern. Med. 2015, 2015, 1–6. [Google Scholar] [CrossRef]
- Jacoby, H.I.; Wnorowski, G.; Sakata, K.; Maeda, H.; D, P.; Mba, G.W.B.; Mba, B. The Effect of MGN-3 on Cisplatin and Doxorubicin Induced Toxicity in the Rat. J. Nutraceuticals, Funct. Med Foods 2001, 3, 3–11. [Google Scholar] [CrossRef]
- Robb, K.A.; Simon, A.E.; Miles, A.; Wardle, J. Public perceptions of cancer: a qualitative study of the balance of positive and negative beliefs. BMJ Open 2014, 4, e005434–e005434. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-J.; Ryu, S.-N.; Han, S.-J.; Kim, H.-Y.; Kim, J.-H.; Hong, S.-G. In Vivo Immunological Activity in Fermentation with Black Rice Bran. Korean J. Food Nutr. 2011, 24, 273–281. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Kim, J.-H.; Yang, S.-B.; Hong, S.-G.; Lee, S.-A.; Hwang, S.-J.; Shin, K.-S.; Suh, H.-J.; Park, M.-H. A Polysaccharide Extracted from Rice Bran Fermented withLentinus edodesEnhances Natural Killer Cell Activity and Exhibits Anticancer Effects. J. Med. Food 2007, 10, 25–31. [Google Scholar] [CrossRef]
- Kim, J.-M.; Hong, S.-G.; Song, B.-S.; Sohn, H.-J.; Baik, H.; Sung, M.-K. Efficacy of Cereal-based Oral Nutrition Supplement on Nutritional Status, Inflammatory Cytokine Secretion and Quality of Life in Cancer Patients Under Cancer Therapy. J. Cancer Prev. 2020, 25, 55–63. [Google Scholar] [CrossRef]
- Kim, K.; Khang, D. Past, Present, and Future of Anticancer Nanomedicine. Int. J. Nanomed. 2020, ume 15, 5719–5743. [Google Scholar] [CrossRef]
- Knudson, A.G. Cancer genetics. Am. J. Med Genet. 2002, 111, 96–102. [Google Scholar] [CrossRef]
- Sjøgren, K.; Jacobsen, K.A.; Grønberg, B.H.; Halvorsen, T.O. Timing of Severe Toxicity from Chemotherapy in Patients With Lung Cancer. Anticancer. Res. 2020, 40, 6399–6406. [Google Scholar] [CrossRef]
- Lemoine, M. Anya Plutynski’s Explaining Cancer: Finding Order in Disorder. Philos. Med. 2022, 3. [Google Scholar] [CrossRef]
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory T cells in tumor microenvironment: new mechanisms, potential therapeutic strategies and future prospects. Mol. Cancer 2020, 19, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Shen, L.; Luo, M.; Zhang, K.; Li, J.; Yang, Q.; Zhu, F.; Zhou, D.; Zheng, S.; Chen, Y.; et al. Circulating tumor cells: biology and clinical significance. Signal Transduct. Target. Ther. 2021, 6, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Lin, W. , Niu, Z., Zhang, H., Kong, Y., Wang, Z., Yang, X., & Yuan, F. (2019). Imbalance of Th1/Th2 and Th17/Treg during the development of uterine cervical cancer. International Journal of Clinical and Experimental Pathology, 12(9), 3604-3612.
- Lissoni, P. , Messina, G., Brivio, F., Fumagalli, L., Rovelli, F., Maruelli, L., Miceli, M., Marchiori, P., Porro, G., Held, M., Fede, G., & Uchiyamada, T. (2008). Modulation of the anticancer immunity by natural agents: inhibition of T regulatory lymphocyte generation by arabinoxylan in patients with locally limited or metastatic solid tumors. Cancer Therapy, 6(2), 1011-1016.
- Liu, S.; Edgerton, S.M.; Moore, D.H.; Thor, A.D. Measures of cell turnover (proliferation and apoptosis) and their association with survival in breast cancer. . 2001, 7, 1716–23. [Google Scholar] [PubMed]
- Loef, M.; Walach, H. Quality of life in cancer patients treated with mistletoe: a systematic review and meta-analysis. BMC Complement. Med. Ther. 2020, 20, 227. [Google Scholar] [CrossRef] [PubMed]
- Lugini, L.; Lozupone, F.; Matarrese, P.; Funaro, C.; Luciani, F.; Malorni, W.; Rivoltini, L.; Castelli, C.; Tinari, A.; Piris, A.; et al. Potent Phagocytic Activity Discriminates Metastatic and Primary Human Malignant Melanomas: A Key Role of Ezrin. Mod. Pathol. 2003, 83, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, M.; Zhao, R.; Wang, D.; Ma, Y.; Ai, L. Plant Natural Products: Promising Resources for Cancer Chemoprevention. Molecules 2021, 26, 933. [Google Scholar] [CrossRef] [PubMed]
- Majeed, M.; Hakeem, K.R.; Rehman, R.U. Mistletoe lectins: From interconnecting proteins to potential tumour inhibiting agents. Phytomedicine Plus 2021, 1, 100039. [Google Scholar] [CrossRef]
- Martins-Teixeira, M.B.; Carvalho, I. Antitumour Anthracyclines: Progress and Perspectives. ChemMedChem 2020, 15, 933–948. [Google Scholar] [CrossRef]
- Masood, A. I. , Sheikh, R., & Anwer, R. A. (2013). “BIOBRAN MGN-3”; Effect of reducing side effects of chemotherapy in breast cancer patients. The Professional Medical Journal, 20(1), 13-16.
- Mazzotti, E.; Cappellini, G.C.A.; Buconovo, S.; Morese, R.; Scoppola, A.; Sebastiani, C.; Marchetti, P. Treatment-related side effects and quality of life in cancer patients. Support. Care Cancer 2012, 20, 2553–2557. [Google Scholar] [CrossRef]
- Melaiu, O.; Lucarini, V.; Cifaldi, L.; Fruci, D. Influence of the Tumor Microenvironment on NK Cell Function in Solid Tumors. Front. Immunol. 2020, 10, 3038. [Google Scholar] [CrossRef]
- Miura, T. , Chiba, M., Miyazaki, Y., Kato, Y., & Maeda, H. (2004/2013). Chemical structure of the component involved in immunoregulation. In BioBran/MGN-3 (Rice Bran Arabinoxylan Coumpound): Basic and clinical application to integrative medicine (2nd ed., pp. 14-22). BioBran Research Foundation. (Reprinted from a report of 2004 Annual Meeting of the Japanese society of Applied Glycoscience.
- Morse, W.; Nawaz, H.; Choudhry, A.A. Combination of chemotherapeutic agents and biological response modifiers (immunotherapy) in triple-negative/Her2( +) breast cancer, multiple myeloma, and non-small-cell lung cancer. J. Egypt. Natl. Cancer Inst. 2022, 34, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, N.; Usmani, D.; Tarique, M.; Naz, H.; Ashraf, M.; Raliya, R.; Tabrez, S.; Zughaibi, T.A.; Alsaieedi, A.; Hakeem, I.J.; et al. The Role of Natural Products and Their Multitargeted Approach to Treat Solid Cancer. Cells 2022, 11, 2209. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Hu, P.; Yang, M.; Zhang, J.; Liu, Y.; Zhu, W.; Zheng, Q. Natural Products as Anticancer Agents: Current Status and Future Perspectives. Molecules 2022, 27, 8367. [Google Scholar] [CrossRef] [PubMed]
- Nasa, P.; Jain, R.; Juneja, D. Delphi methodology in healthcare research: How to decide its appropriateness. World J. Methodol. 2021, 11, 116–129. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute. (n.d.). Cancer. In Dictionary of Cancer Terms. https://www.cancer.
- National Heart, Lung, and Blood Institute (NHLBI) Quality Assessment Tool for Observational Cohort and Cross-Sectional Studies. Available online: https://www.nhlbi.nih.gov/health-topics/study-quality-assessment-tools (accessed on 23 February 2022).
- Nesher, L.; Rolston, K.V.I. The current spectrum of infection in cancer patients with chemotherapy related neutropenia. Infection 2013, 42, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Noaman, E.; El-Din, N.K.B.; Bibars, M.A.; Mossallam, A.A.A.; Ghoneum, M. Antioxidant potential by arabinoxylan rice bran, MGN-3/biobran, represents a mechanism for its oncostatic effect against murine solid Ehrlich carcinoma. Cancer Lett. 2008, 268, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Nordvig, J.; Aagaard, T.; Daugaard, G.; Brown, P.; Sengeløv, H.; Lundgren, J.; Helleberg, M. Febrile Neutropenia and Long-term Risk of Infection Among Patients Treated With Chemotherapy for Malignant Diseases. Open Forum Infect. Dis. 2018, 5, ofy255. [Google Scholar] [CrossRef] [PubMed]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) cells in cancer: Can Treg cells be a new therapeutic target? Cancer Sci. 2019, 110, 2080–2089. [Google Scholar] [CrossRef]
- Ooi, S.L.; McMullen, D.; Golombick, T.; Nut, D.; Pak, S.C. Evidence-Based Review of BioBran/MGN-3 Arabinoxylan Compound as a Complementary Therapy for Conventional Cancer Treatment. Integr. Cancer Ther. 2017, 17, 165–178. [Google Scholar] [CrossRef]
- Record #1671 is using a reference type undefined in this output style.
- Ooi, S.L.; Pak, S.C.; Micalos, P.S.; Schupfer, E.; Lockley, C.; Park, M.H.; Hwang, S.-J. The Health-Promoting Properties and Clinical Applications of Rice Bran Arabinoxylan Modified with Shiitake Mushroom Enzyme—A Narrative Review. Molecules 2021, 26, 2539. [Google Scholar] [CrossRef]
- Palmer, A.C.; Sorger, P.K. Combination Cancer Therapy Can Confer Benefit via Patient-to-Patient Variability without Drug Additivity or Synergy. Cell 2017, 171, 1678–1691. [Google Scholar] [CrossRef]
- Pasiarski, M.; Grywalska, E.; Kosmaczewska, A.; Góźdź, S.; Roliński, J. The frequency of myeloid and lymphoid dendritic cells in multiple myeloma patients is inversely correlated with disease progression. 67. [CrossRef]
- Pérez-Martínez, A.; Valentín, J.; Fernández, L.; Hernández-Jiménez, E.; López-Collazo, E.; Zerbes, P.; Schwörer, E.; Nuñéz, F.; Martín, I.G.; Sallis, H.; et al. Arabinoxylan rice bran (MGN-3/Biobran) enhances natural killer cell–mediated cytotoxicity against neuroblastoma in vitro and in vivo. Cytotherapy 2014, 17, 601–612. [Google Scholar] [CrossRef]
- Pescatore, F. M. , Přestáno, C. ( 28(1), 8–10. [PubMed]
- Petrovics, G.; Szigeti, G.; Hamvas, S.; Máté. ; Betlehem, J.; Hegyi, G. Controlled pilot study for cancer patients suffering from chronic fatigue syndrome due to chemotherapy treated with BioBran (MGN-3-Arabinoxylane) and targeted radiofrequency heat therapy. Eur. J. Integr. Med. 2016, 8, 29–35. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D. H. , & Arlt, V. M. (2009). Genotoxicity: damage to DNA and its consequences. In A. Luch (Ed.), Molecular, Clinical and Environmental Toxicology: Volume 1: Molecular Toxicology (pp. 87-110). Birkhäuser Basel. [CrossRef]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. Br. J. Cancer 2020, 124, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Riggan, L.; Shah, S.; E O’sullivan, T. Arrested development: suppression of NK cell function in the tumor microenvironment. Clin. Transl. Immunol. 2021, 10. [Google Scholar] [CrossRef]
- Rosenberg, J.; Huang, J. CD8+ T cells and NK cells: parallel and complementary soldiers of immunotherapy. Curr. Opin. Chem. Eng. 2018, 19, 9–20. [Google Scholar] [CrossRef]
- Salako, O.; Okunade, K.S.; Adeniji, A.A.; Fagbenro, G.T.; Afolaranmi, O.J. Chemotherapy induced neutropenia and febrile neutropenia among breast cancer patients in a tertiary hospital in Nigeria. ecancermedicalscience 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Schegoleva, A.A.; Khozyainova, A.A.; Gerashchenko, T.S.; Zhuikova, L.D.; Denisov, E.V. Metastasis prevention: targeting causes and roots. Clin. Exp. Metastasis 2022, 39, 505–519. [Google Scholar] [CrossRef]
- Schmidt, M.; Lügering, N.; Pauels, H.-G.; Schulze-Osthoff, K.; Domschke, W.; Kucharzik, T. IL-10 induces apoptosis in human monocytes involving the CD95 receptor/ligand pathway. Eur. J. Immunol. 2000, 30, 1769–1777. [Google Scholar] [CrossRef]
- Shalapour, S.; Karin, M. Immunity, inflammation, and cancer: an eternal fight between good and evil. J. Clin. Investig. 2015, 125, 3347–3355. [Google Scholar] [CrossRef] [PubMed]
- Shamekhi, S.; Abdolalizadeh, J.; Ostadrahimi, A.; Mohammadi, S.A.; Barzegari, A.; Lotfi, H.; Bonabi, E.; Zarghami, N. Apoptotic Effect of Saccharomyces cerevisiae on Human Colon Cancer SW480 Cells by Regulation of Akt/NF-ĸB Signaling Pathway. Probiotics Antimicrob. Proteins 2019, 12, 311–319. [Google Scholar] [CrossRef]
- Koyama, S.; Nishikawa, H. Mechanisms of regulatory T cell infiltration in tumors: implications for innovative immune precision therapies. J. Immunother. Cancer 2021, 9, e002591. [Google Scholar] [CrossRef]
- Sojka, D.K.; Huang, Y.; Fowell, D.J. Mechanisms of regulatory T-cell suppression—a diverse arsenal for a moving target. Immunology 2008, 124, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Takahara, K. , & Sano, K. (2004). The life prolongation and QOL improvement effect of rice bran arabinoxylan derivative (MGN-3, Biobran) for progressive cancer. Clinical Pharmacology and Therapy, 14(3), 267-271.
- Tan, D. F. S. , & Flores, J. A. S. (2020). The immunomodulating effects of arabinoxylan rice bran ( Lentin ) on hematologic profile, nutritional status and quality of life among head and neck carcinoma patients undergoing radiation therapy: A double blind randomized control trial. Radiology Journal, The Official Publication of the Philippine College of Radiology, 12(February), 11-16.
- Tazawa, K. , Namikawa, H., Oida, N., Itoh, K., Yatsuzuka, M., Koike, J., Masada, M., & Maeda, H. (2000). Scavenging activity of MGN-3 (arabinoxylane from rice bran) with natural killer cell activity on free radicals. Biotherapy: Official journal of Japanese Society of Biological Response Modifiers, 14(5), 493-495.
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T cells in cancer immunosuppression — implications for anticancer therapy. Nat. Rev. Clin. Oncol. 2019, 16, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Tsao, A.S.; Kim, E.S.; Hong, W.K. Chemoprevention of Cancer. CA: A Cancer J. Clin. 2004, 54, 150–180. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-H.; Wu, C.-F.; Rajasekaran, N.; Shin, Y.K. Loss of Tumor Suppressor Gene Function in Human Cancer: An Overview. Cell. Physiol. Biochem. 2018, 51, 2647–2693. [Google Scholar] [CrossRef]
- Weston, A. , & Harris, C. C. (2003). Multistage carcinogenesis. In D. W. Kufe, R. E. Pollock, R. R. Weichselbaum, R. C. J. Bast, T. S. Gansler, J. F. Holland, & E. Frei (Eds.), Holland-Frei Cancer Medicine (6th ed.). BC Decker.
- E Wilson, B.; Jacob, S.; Yap, M.L.; Ferlay, J.; Bray, F.; Barton, M.B. Estimates of global chemotherapy demands and corresponding physician workforce requirements for 2018 and 2040: a population-based study. Lancet Oncol. 2019, 20, 769–780. [Google Scholar] [CrossRef]
- Wu, Z.; Li, S.; Zhu, X. The Mechanism of Stimulating and Mobilizing the Immune System Enhancing the Anti-Tumor Immunity. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, F.; Li, H.; Yang, L.; Lv, T.; Gu, W.; Song, Y. Circulating Tumor Cells Were Associated with the Number of T Lymphocyte Subsets and NK Cells in Peripheral Blood in Advanced Non-Small-Cell Lung Cancer. Dis. Markers 2017, 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Gaugler, B.; Mohty, M.; Malard, F. Plasmacytoid dendritic cell biology and its role in immune-mediated diseases. Clin. Transl. Immunol. 2020, 9. [Google Scholar] [CrossRef]
- Ba, Y.; Shi, Y.; Jiang, W.; Feng, J.; Cheng, Y.; Xiao, L.; Zhang, Q.; Qiu, W.; Xu, B.; Xu, R.; et al. Current management of chemotherapy-induced neutropenia in adults: key points and new challenges. Cancer Biol. Med. 2020, 17, 896–909. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, J.; Wang, J.; Sun, B. The anti-cancer activity and potential clinical application of rice bran extracts and fermentation products. RSC Adv. 2019, 9, 18060–18069. [Google Scholar] [CrossRef]
- Zabor, E.C.; Kaizer, A.M.; Hobbs, B.P. Randomized Controlled Trials. Chest 2020, 158, S79–S87. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, J.; Ge, S.; Chen, C.; Li, S.; Wu, X.; Feng, X.; Wang, Y.; Cai, D. Saikosaponin A Inhibits Breast Cancer by Regulating Th1/Th2 Balance. Front. Pharmacol. 2019, 10, 624. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Cheng, W.; Qu, W.; Wang, K. Arabinoxylan rice bran (MGN-3/Biobran) alleviates radiation-induced intestinal barrier dysfunction of mice in a mitochondrion-dependent manner. Biomed. Pharmacother. 2020, 124, 109855. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Lawrence, T.; Liang, Y. The Role of Plasmacytoid Dendritic Cells in Cancers. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A flow diagram of evidence selection.
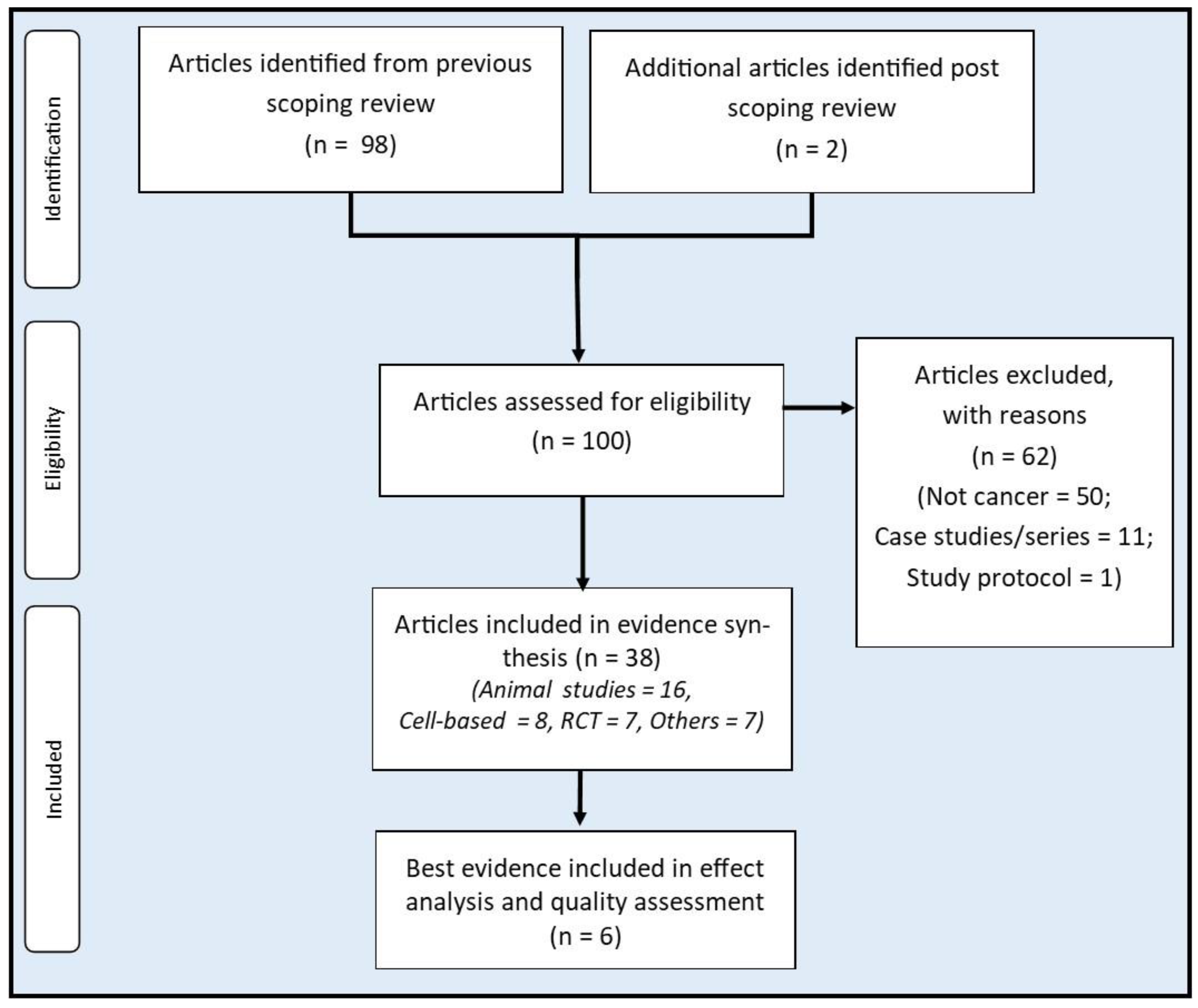
Figure 2.
The immune restorative effects of RBAC. (A) shows some immune functions that are affected by tumour growth resulting in the suppression of antitumour activity. (B) shows the biological modifier effects of RBAC, which restore the immune dysfunction in cancer patients.
Figure 2.
The immune restorative effects of RBAC. (A) shows some immune functions that are affected by tumour growth resulting in the suppression of antitumour activity. (B) shows the biological modifier effects of RBAC, which restore the immune dysfunction in cancer patients.
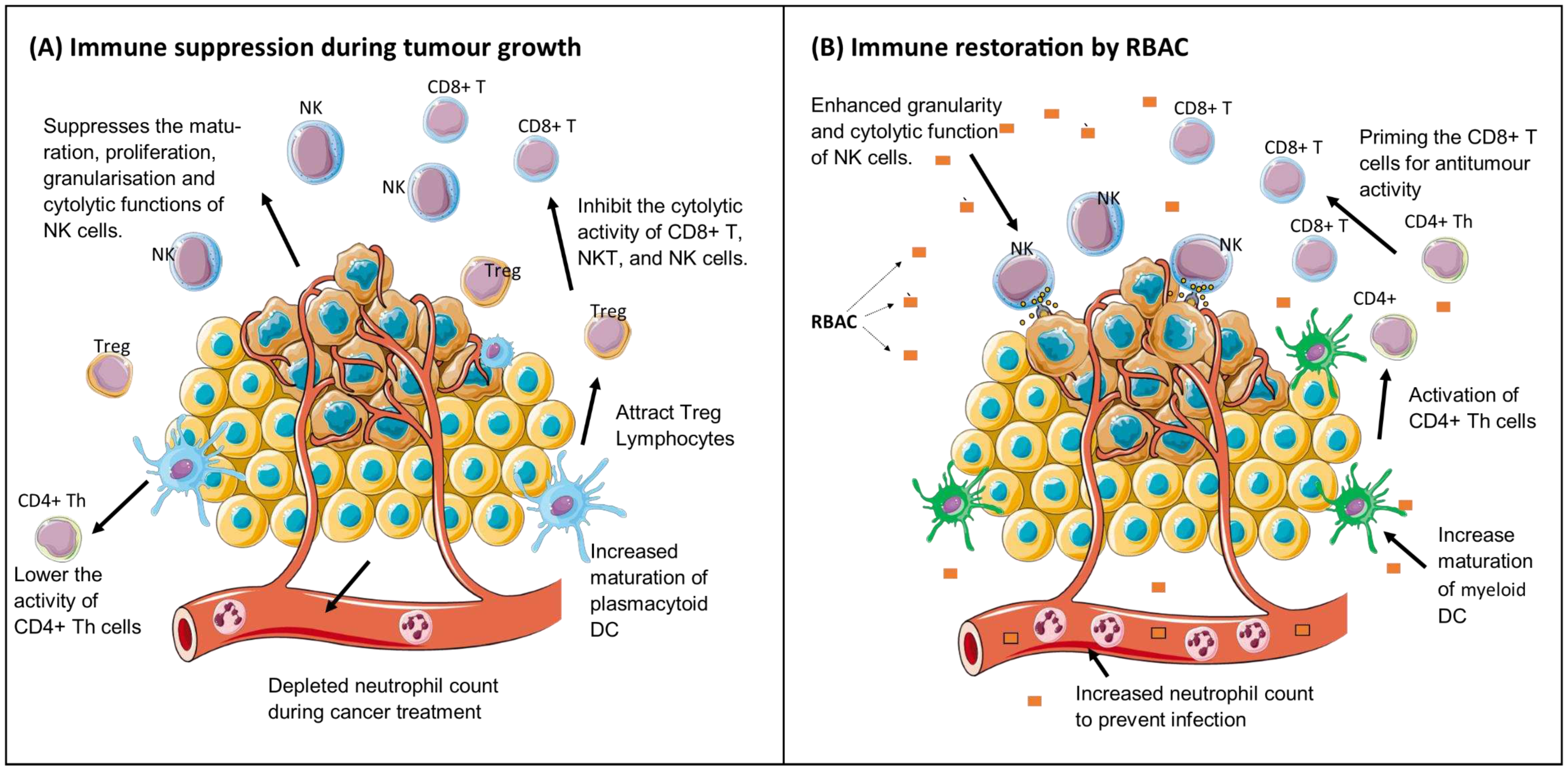
Figure 3.
The anticancer effect of RBAC is achieved through both the intrinsic pathway via increased susceptibility of CD95 (Fas/APO-1) ligands on the cancerous cells to promote apoptosis and the extrinsic pathway through downregulating the antiapoptotic Bcl-2 proteins to lower membrane potentials leading to upregulation of tumour suppressing P53 gene, and increased production of the apoptotic Bax and caspase-3 signalling proteins. Malignant cell proliferation is arrested with evidence of increased hypodiploid cells of the SubG1 phase in cell cycle analysis. RBAC’s antioxidant and cytokine-modulating capacities also augment proapoptotic activity.
Figure 3.
The anticancer effect of RBAC is achieved through both the intrinsic pathway via increased susceptibility of CD95 (Fas/APO-1) ligands on the cancerous cells to promote apoptosis and the extrinsic pathway through downregulating the antiapoptotic Bcl-2 proteins to lower membrane potentials leading to upregulation of tumour suppressing P53 gene, and increased production of the apoptotic Bax and caspase-3 signalling proteins. Malignant cell proliferation is arrested with evidence of increased hypodiploid cells of the SubG1 phase in cell cycle analysis. RBAC’s antioxidant and cytokine-modulating capacities also augment proapoptotic activity.
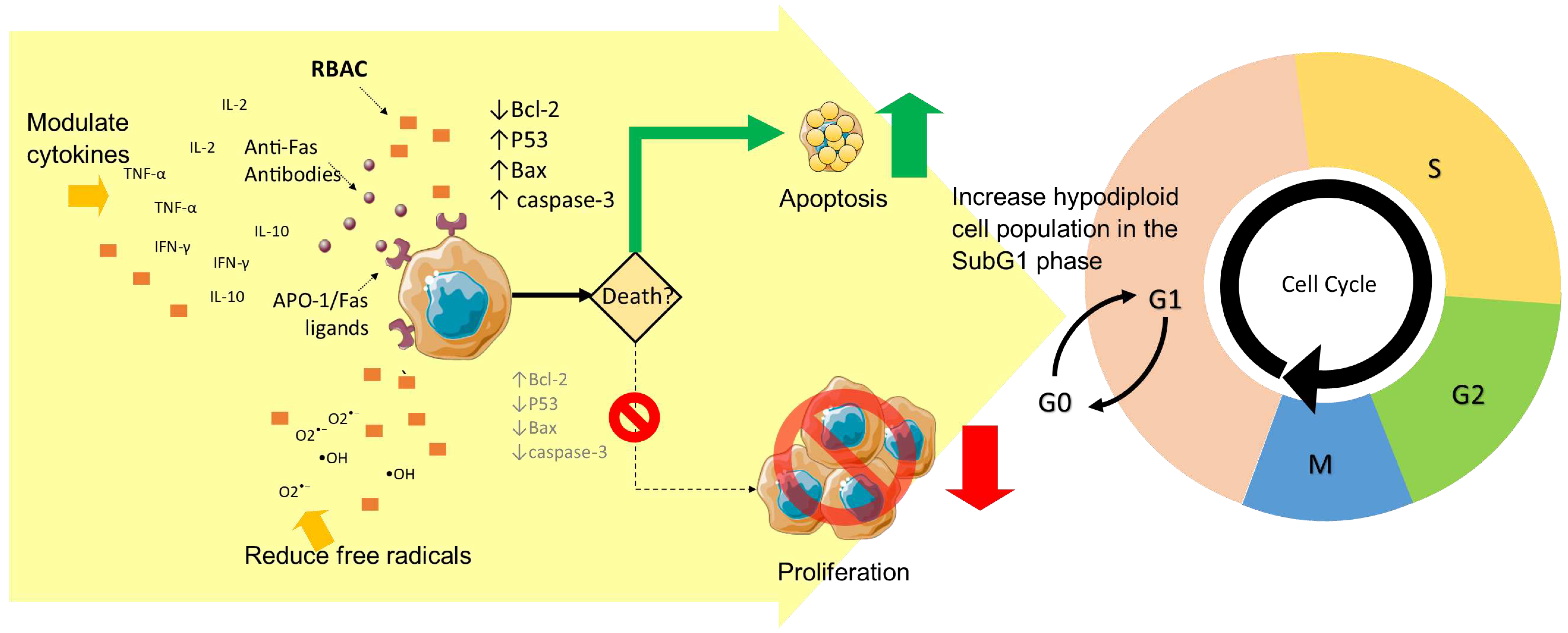
Figure 4.
A summary of the potential benefits of RBAC in cancer treatment.
Figure 5.
Forest plot of survival events of RBAC treatment compared to control.
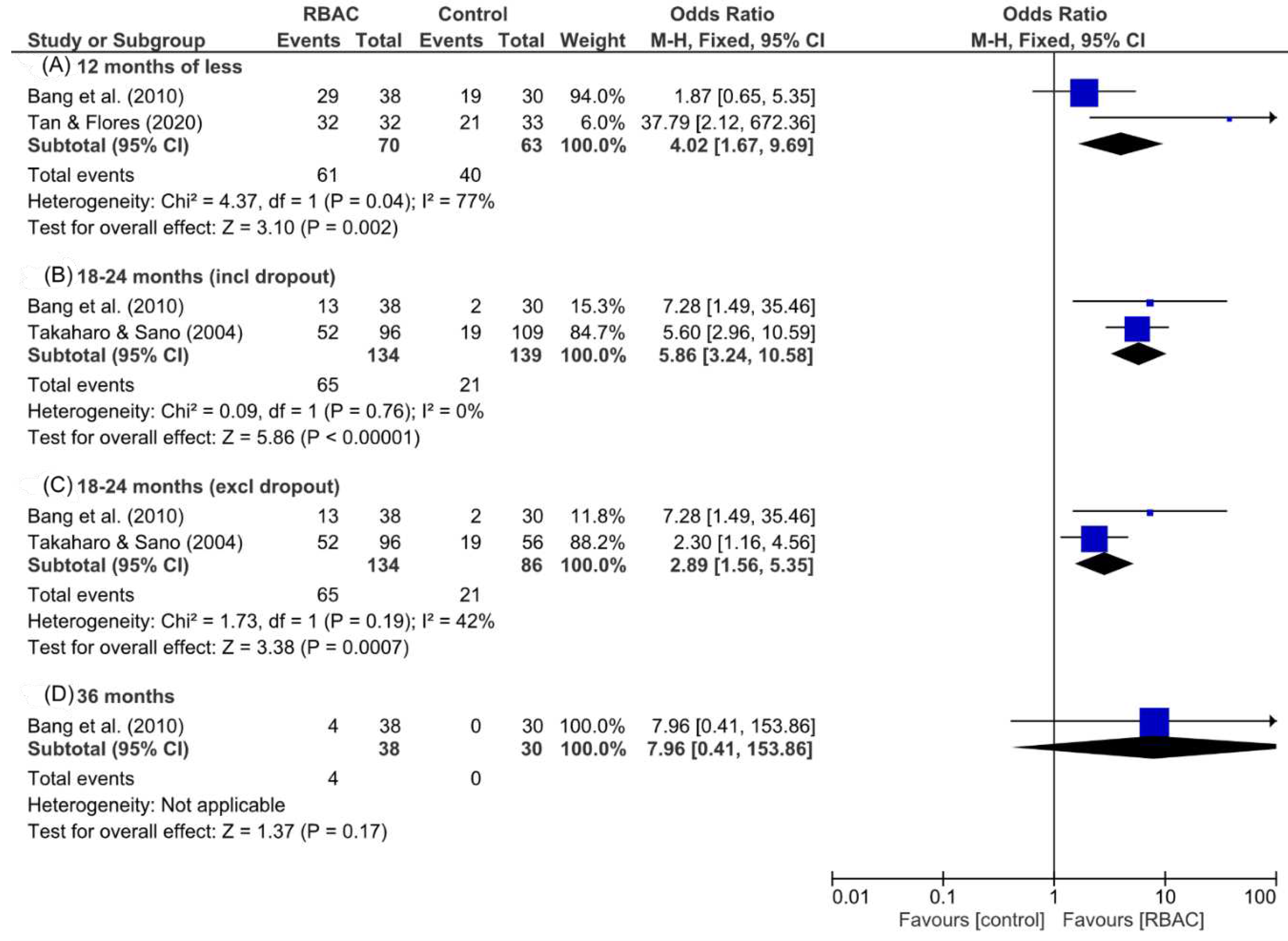
Figure 6.
A bubble chart visualising the available evidence on RBAC’s effect on cancer patients’ QoL. Each bubble represents an outcome assessed in one study, with the sample size defining the bubble’s diameter. Each study is assigned a different colour. X-axis: Statistical significance (p-value) of the outcome; Y-axis: QoL outcome measures.
Figure 6.
A bubble chart visualising the available evidence on RBAC’s effect on cancer patients’ QoL. Each bubble represents an outcome assessed in one study, with the sample size defining the bubble’s diameter. Each study is assigned a different colour. X-axis: Statistical significance (p-value) of the outcome; Y-axis: QoL outcome measures.
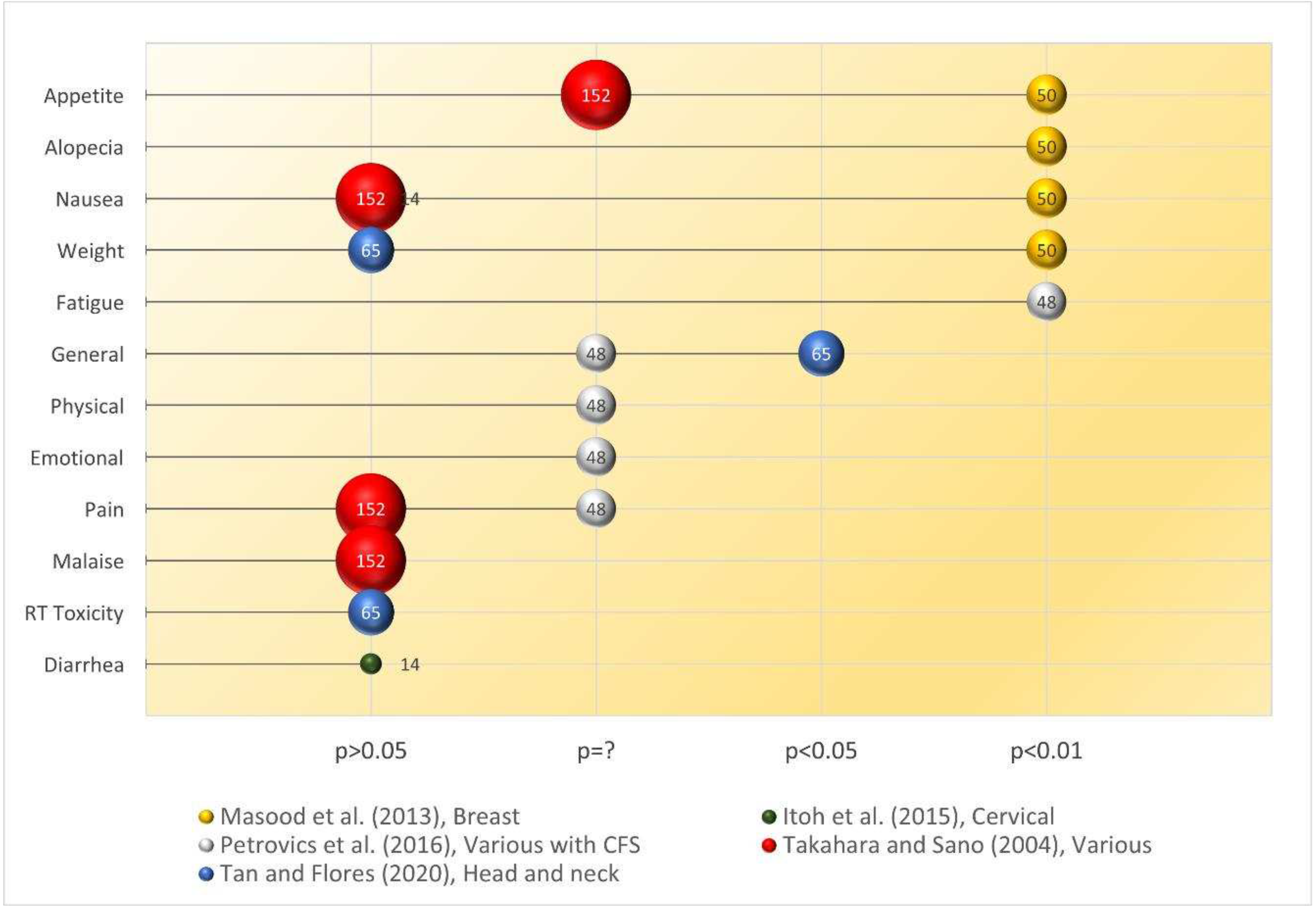
Figure 7.
A summary of the quality assessment results by study.
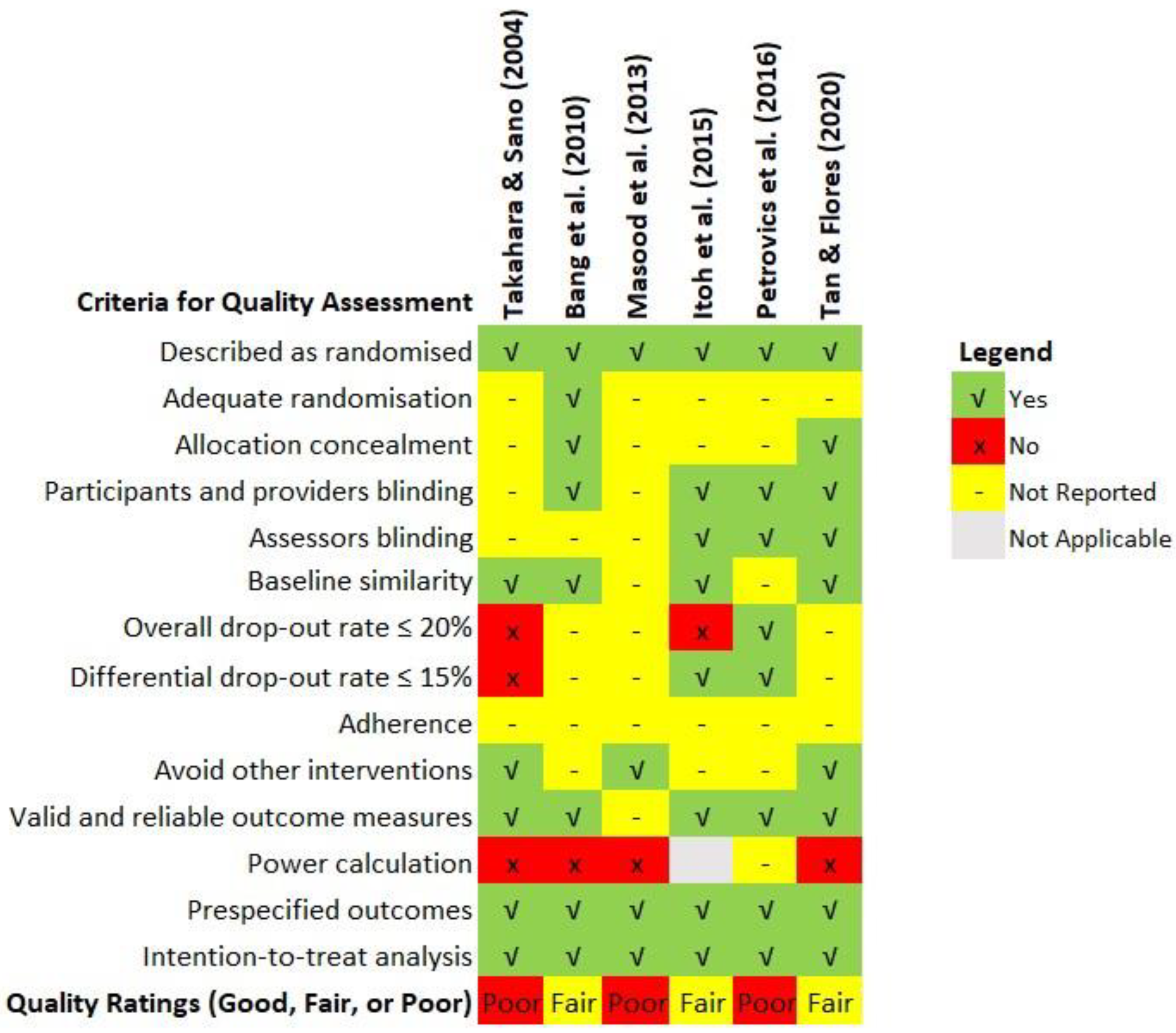
Figure 8.
A chart summarising the quality assessment outcomes by types of bias.
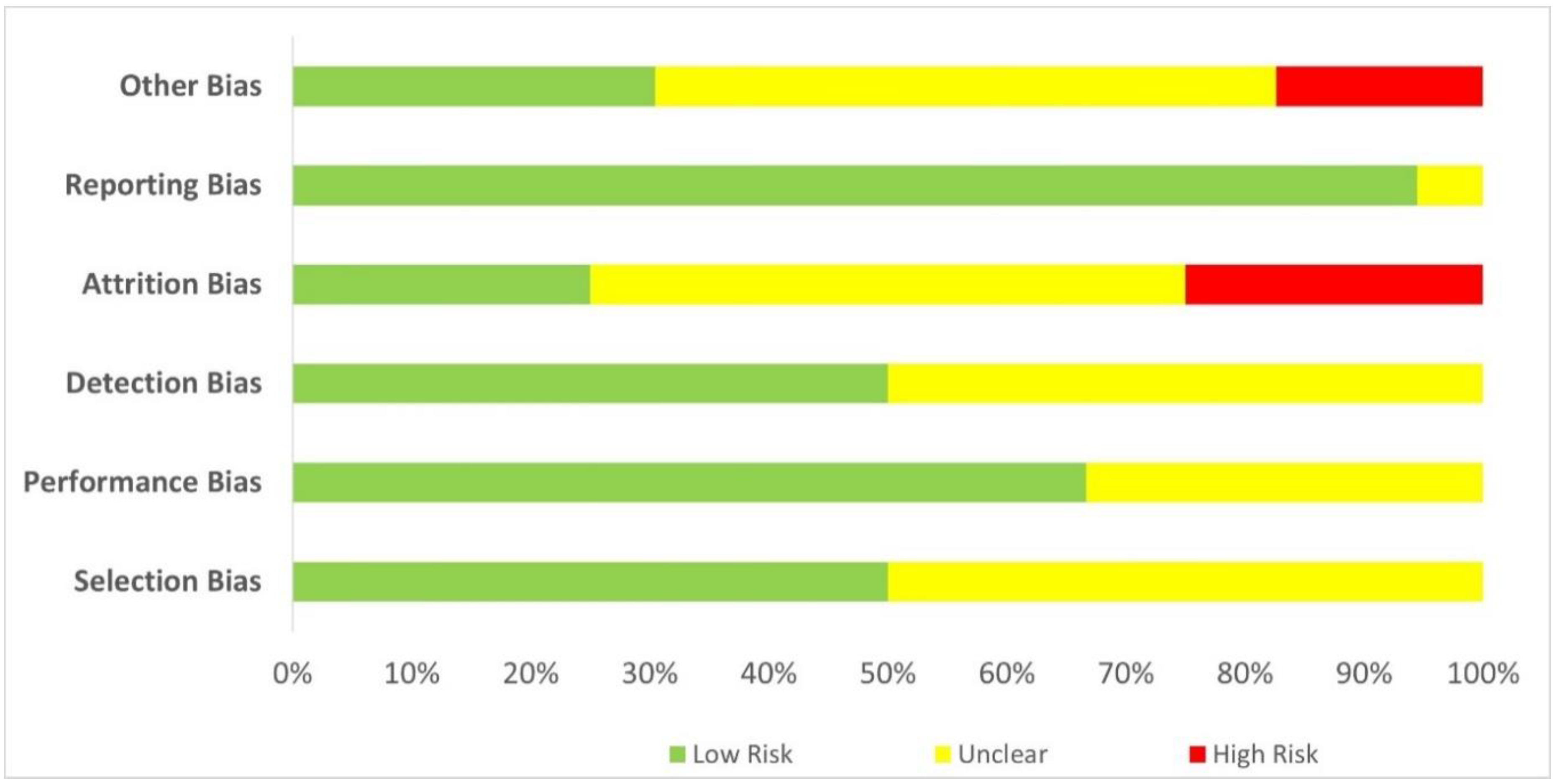

Table 1.
Results from human and animal studies on RBAC’s immune restorative capacity in cancer.
| # | RBAC (dose) | Study Design | Key Findings | Reference |
|---|---|---|---|---|
| 1 | Biobran MGN-3 (3g/day for 1 to 2 weeks) |
Before & after study. Various malignancies (n=32) | Biobran MGN-3 significantly increased (p<0.001) NKC cytolytic activity up to 10-fold compared to baseline. Increased NKC granularity with enhanced tumour-killing capacity was observed posttreatment. | (Ghoneum & Brown, 1999) |
| 2 | Biobran MGN-3 (3g/day for 1 month) | Before & after study. Various malignancies (n=5) | Statistically significant (p<0.001) increases in responses with T & B cell mitogen tests compared to baseline demonstrated signs of restoration of the adaptive immunity. | (Ghoneum & Brown, 1999) |
| 3 | Biobran MGN-3 (3g/day for 1 to 2 weeks) | Before & after study. Various malignancies (n=90) | 95.5% of patients demonstrated 2 to 10-fold increases in NKC cytolytic activity at 1-2 weeks posttreatment. | (Ghoneum, 1999) |
| 4 | Biobran MGN-3 (3g/day for 18 months) | RCT. Patients with progressive cancer of late stages (n=152, RBAC=96, Control=56). | A significantly higher portion of participants with low or medium NKC activity in the RBAC group survived than the control group (Low: 42.5% vs 12.5%, p<0.01; Medium: 51.4% vs 28.0%, p<0.05). | (Takahara & Sano, 2004) |
| 5 | Biobran MGN-3 (2g/day for 1st month, 1g/day for 2nd month) | Before & after study. Various malignancies (n=22) | A statistically significant change in the ratio of Th/Treg was detected (p=0.025), and the increase in Th/Treg was more pronounced in participants with low Th/Treg at baseline. | (Lissoni et al., 2008) |
| 6 | Biobran MGN-3 (2g/day for 3 months) | RCT. MM patients (n=48, RBAC=32, placebo=12) | Significant increases in NKC activity of the RBAC group compared to the baseline in the first (p=0.045) and second (p=0.029) months. The circulating mDC percentage (p=0.036) and mDC/pDC ratio (p=0.030) increased substantially after 3 months. | (Cholujova et al., 2013) |
| 7 | Biobran MGN-3 (3g/day for 4 weeks) | RCT. Cervical cancer patients (n=14, RBAC=7, placebo=7) | Both groups experienced declines in NKC activity after chemoradiotherapy compared to the baseline values, with no significant difference between the two groups. | (Itoh et al., 2015) |
| 8 | Biobran MGN-3 (40mg/kg BW p.o. every other day for 8 months) | Wistar rats + carcinogen (MNNG) | MNNG caused significant lymphocyte depletion (↓23.3%, p<0.01) after 8 months compared to healthy controls. RBAC+MNNG promoted lymphocyte recovery to normal levels (p<0.05). | (Badr El-Din, Abdel Fattah, et al., 2016) |
| 9 | Biobran MGN-3 (2g/day for 6 months) | Before & after study. MGUS/SMM patients (n=10) | Half of the patients had neutropenia at baseline. After consuming RBAC, eight participants showed an increased neutrophil count between 10% and 90%. | (Golombick et al., 2016) |
| Abbreviations: BW, body weight; CD, cluster of differentiation; mDC, myeloid dendritic cells; MM, multiple myeloma; MNNG, methylnitronitrosoguanidine; NKC, natural killer cells; p.o., per oral; pDC, plasmacytoid dendritic cells; RBAC, rice bran arabinoxylan compound; RCT, randomised controlled trial; Th, T helper cells; Treg, Regulatory T cells; MGUS/SMM, monoclonal gammopathy of undetermined significance/smoldering multiple myeloma. | ||||
Table 2.
The in vivo anticancer effects of RBAC from murine models with tumour growth inhibition and life prolongation as outcome measures.
Table 2.
The in vivo anticancer effects of RBAC from murine models with tumour growth inhibition and life prolongation as outcome measures.
| # | RBAC (dose) | Model | Key Findings | Reference |
|---|---|---|---|---|
| 1 | Biobran MGN-3 (1.5mg/day p.o. for 23 days) | ICR mice + S-180 cells | TW ↓66.2% (0.51 ±0.34g vs 3.40+1.46g, p<0.01) compared to the control. Better than PSP with TW ↓49.0% (p<0.05). | (Bae et al., 2004) |
| 2 | RBEP (50mg/kg BW p.o. & i.p., 250mg/kg BW p.o. for 14 days) | (1) ICR mice + S-180; (2) C57/Bl6 mice + B16/Bl6 melanoma | (1) RBEP prolonged survival by 14.6% (31.4 days), 30.3% (35.7 days), and 38.0% (37.8 days) with 50 mg/kg, 250 mg/kg p.o., and 50 mg/kg i.p., respectively. (2) TW ↓35.6% with 50 mg/kg p.o., ↓41.7% (2.155g) 250 mg/kg p.o., and ↓55.1% (1.66g) with 50 mg/kg i.p. |
(Kim et al., 2007) |
| 3 | Biobran MGN-3 (40mg/kg BW i.p. over 3 weeks & i.t. over 5 weeks) | Swiss albino mice + SEC | TV (i.p.) was significantly lower than control starting from day 14 (p<0.05) and ↓63.27% (p<0.01) at day 35; TW (i.p.) was also significantly lower (3.63±0.45 vs 6.62±0.38, p<0.01) compared to the control at day 35. Significant TV (i.t.) reduction from observed from day 28, reaching a ↓44.83% (p<0.01) difference with control on day 45. | (Badr El-Din et al., 2008) |
| 4 | Biobran MGN-3 (25mg/kg BW i.p. over 25 days) | Swiss albino mice + SEC | Early treatment (from day 4) TV ↓54% & TW ↓34% (p<0.01) relative to the control. Late treatment (from day 11) TV ↓24% & TW ↓12% (p<0.05). | (Noaman et al., 2008) |
| 5 | RBEP (250mg/kg p.o. or i.p. for > 2 weeks | ICR mice + S-180 cells | Survival rate: p.o. 5.3% higher (19.9 vs 18.9 days), and i.p. 23.2% higher (23 vs 18.7 days), compared to controls. RBEP-treated mice have significantly lower BW than controls starting from day 13 (p.o.) and day 10 (i.p.) | (An, 2011) |
| 6 | C3G-F (250mg/kg p.o. for 2 or 3 weeks | (1) ICR mice + S-180; (2) C57/Bl6 mice + B16/Bl6 melanoma | (1) BW: significant difference (p<0.05) since day 8. C3G-F prevented BW gain (6.5 g vs 11.8 g, ↓60%) on day 15. (2) TW: ↓19.4% in C3G-F group vs control (0.514±0.129 g vs 0.635±0.241 g, p<0.05) at 3 weeks. |
(Kim et al., 2011) |
| 7 | NKC activated with 100mg/mL Biobran MGN-3 i.v. 2x/week for 4 weeks | NOD-scid IL-2Rgnull mice + NB-1691luc | RBAC-activated NKC treatment significantly lowered TV (p<0.05) under bioluminescence imaging and extended the survival time of the mice than fresh NKC therapy or no treatment. | (Pérez-Martínez et al., 2015) |
| 8 | Biobran MGN-3 (40mg/kg BW i.p. 3x/weeks for 3 weeks) | Swiss albino mice + SEC | RBAC significantly reduced BW loss in SEC-bearing mice (↓4.1% vs 18%, p<0.01) and TW (↓46.3%, p<0.01) compared to control by day 30. Continuous suppression of TV was detected (day 14: ↓33.7%, day 30: ↓49.9%, p<0.01). | (Badr El-Din et al., 2019) |
| Abbreviations: BW, body weight; C3G-F, fermented SuperC3GHi bran; i.p., intraperitoneal injection; i.t., intratumoral injection; i.v., intravenous; NKC, natural killer cells; p.o., per oral; RBAC, rice bran arabinoxylan compound; RBEP, rice bran exo-biopolymer; SEC, solid Ehrlich carcinoma; TV, tumour volume in mm3; TW, tumour weight in g. | ||||
Table 3.
The potential mechanisms of RBAC’s direct anticancer effects.
| # | RBAC (dose) | Model | Key Findings | Reference |
|---|---|---|---|---|
| A. Promote cancer cell apoptosis | ||||
| 1 | Biobran MGN-3 (concentration not reported) | In vitro. SCC13 cell line. | A 30% decrease in cell numbers after 48 hours and 50% at 72 hours after incubation with RBAC was detected. Untreated SCC13 cells continued to grow over time. Also found was increased secretion of IL-10 and IL-12 of SCC13 cells by RBAC. | (Ghoneum et al., 2000) |
| 2 | Biobran MGN-3 (100, 500 and 1000 mg/ml) | In vitro. MCF-7 cell line. | Cell survival rates were dose-dependent at 75%, 70% and 63% after 3 days of incubation with 100, 500 and 1000 mg/ml of RBAC, respectively. | (Gollapudi & Ghoneum, 2008) |
| 3 | Biobran MGN-3 (100–1000 μg/ml) | In vitro. MCF-7 & 4T1 cell lines. | IC50 (MCF-7) was 800 μg/ml at 24 hours and about 1000 μg/ml at 48 hours. IC50 (4T1) being 700 μg/ml at 24 hours and 580 μg/ml at 48 hours. | (Ghoneum et al., 2014) |
| 4 | Biobran MGN-3 (0–1000 μg/ml) | In vitro. PC3 & LNCaP cell lines. | Significantly decreases (p<0.05) in the cancer cell proliferation in a dose- and time-dependent manner (24, 48, and 72h). | (Brush et al., 2010) |
| 5 | Biobran MGN-3 (100–1000 μg/ml) | In vitro. HUT 78 cell line. Anti-CD95 antibodies. | Treatment of HUT 78 cells with RBAC (for 3 hours) before incubating with anti-CD95 antibodies increased the specific apoptosis significantly (p<0.01) by 35-42%, about double that of Anti-CD95 antibodies alone. The escalation in apoptosis was not associated with the upregulation of death receptor expression but through sensitising the receptor. | (Ghoneum & Gollapudi, 2003) |
| 6 | Biobran MGN-3 (40 mg/kg BW p.o. every other day for 8 months) | Wistar rats + carcinogen (MNNG) | RBAC mitigated the carcinogenesis effects of MNNG by causing a significant increase in cell-cycle arrest in the subG1 phase (p<0.01) compared to the control, with the AI/PrI ratio increased by 1.67-fold. RBAC increased the apoptotic cancer cells in tumour tissues by 63.7% (p<0.01), most prominently in early apoptosis (230.1%, p<0.01). Downregulation of Bcl-2 and upregulation of P53, Bax, Bax/Bcl-2 ratio, and caspase-3 were detected. | (Badr El-Din, Abdel Fattah, et al., 2016) |
| 7 | Biobran MGN-3 (25 mg/kg BW i.p. 5x/week, either for 12 or 22 weeks) | Wistar albino rats + carcinogen (NDEA+CCl4) | Cell-cycle arrest in the sub-G1 phase was markedly increased by 126% and 99% (p<0.01) through pretreatment and posttreatment of RBAC. RBAC treatment (pre, post) significantly reduced (p<0.01) viable cells (↓74.51%, ↓72.54%) and necrosis (↑89%, ↑75.47%) while increased early (↑316%, ↑309%) and late (↑255%, ↑237%) apoptosis, compared to carcinogen-untreated rats. RBAC significantly (p<0.01) upregulated p53, Bax, and caspase-3 while downregulated Bcl-2 gene expression. | (Badr El-Din et al., 2020) |
| 8 | Biobran MGN-3 (40 mg/kg BW i.p. 3x/weeks for 3 weeks) | In vivo. Swiss albino mice + SEC | RBAC markedly increased cell-cycle arrest in the sub-G1 phase was detected by 102% (p<0.01) in the RBAC group compared to the control. RBAC treatment also increased the AI/PrI ratio by 2-fold (p<0.01). The quantitative histochemical analysis also showed reduced viable cells (28.2±1.25% vs 74.5±2.25%) and increased apoptotic cells (53.1 ± 1.21% vs 18.2±1.68%) in the tumour tissues of RBAC-treated mice than control. RBAC significantly (p<0.01) upregulated p53, Bax, and caspase-3 while downregulated Bcl-2 gene expression. | (Badr El-Din et al., 2019) |
| B. Prevent oxidative stress | ||||
| 9 | Biobran MGN-3 (25 mg/kg BW i.p. 6x per weeks for 25 days) | In vivo. Swiss albino mice + SEC | Mice treated with RBAC did not show elevated MDA like untreated mice and had significantly higher GSH levels (p<0.01) in the blood, liver, and tumour. GPx, GST, SOD, and CAT and the related gene expressions in RBAC-treated mice were also significantly higher (p<0.01) than in untreated mice. | (Noaman et al., 2008) |
| C. Modulate cytokine production | ||||
| 10 | RBAC (40 mg/kg BW i.p. over 3 wks & i.t. over 5 wks) | In vivo. Swiss albino mice + SEC | RBAC treatment showed a significantly increased TNF-α (↑15.63%) and IFN-γ (↑154.54%) compared to control. Untreated SEC mice showed elevated IL-10 (↑111.71%), but the increase was dampened in RBAC-treated mice (↑14.75%, p<0.01). | (Badr El-Din et al., 2008) |
| 11 | Biobran MGN-3 (2g/day p.o. for 3 months) | RCT. MM patients (n=48, RBAC=32, placebo=12) | RBAC significantly elevated (p<0.05) both Th1 cytokines (IFN-γ, IL-12, IL-17, TNF-α) and Th2 cytokines (IL-4, IL-6, IL-9, IL-10, and IL-13) over placebo after 3 months. | (Cholujova et al., 2013) |
| 12 | ONS with 0.4g of RBEP p.o. for 8 weeks | NRCT. Various malignancies (n=34, RBAC=10, control=24) | RBAC significantly lowered (p<0.05) IL-1β, IL-6 and IL-8 and increased IL-12p70 (p<0.05) compared to the control group. A marginally significant rise (p=0.056) in the IL-10 level in the RBAC group compared to baseline was also detected. | (Kim et al., 2020) |
| Abbreviations: AI/PrI, the ratio of the apoptotic index over the proliferation index; BW, body weight; CAT, catalase; CCI4, carbon tetrachloride; CD, cluster of differentiation; GPx, glutathione peroxidase; GST, glutathione S-transferases; GSH, glutathione; IC50, half maximal inhibitory concentration; IFN, interferon; IL, interleukin; i.p., intraperitoneal; i.t., intratumoral; MDA, malondialdehyde; MM, multiple myeloma; MNNG, methylnitronitrosoguanidine; NDEA, N-nitrosodiethyamine; NRCT, nonrandomised controlled trial. ONS, oral nutritional supplement; p.o., per oral; RBAC, rice bran arabinoxylan compound; RBEP, rice bran exo-biopolymer; RCT, randomised controlled trial; SEC, solid Ehrlich carcinoma; SOD, superoxide dismutase; TNF, tumour necrosis factor. | ||||
Table 4.
In vivo studies investigating the chemoprevention activities of RBAC.
| # | RBAC (dose) | Model | Key Findings | Reference |
|---|---|---|---|---|
| 1 | Biobran MGN-3 (40 mg/kg BW p.o. every other day for 8 months) | Wistar rats + carcinogen (MNNG) | Untreated rats developed mild- and high-grade gastric glandular dysplasia (6/10, 60%) and invasive carcinoma (2/10, 20%). RBAC-treated rats had significantly lower incidence (p<0.01) of mild dysplasia, of which were patchy and small (3.5/12, 29.2%) and carcinoma in situ only (1/12, 8.3%). RBAC also significantly lower Ki-67 tumour proliferation marker expression (39.8% vs 50.8%, p<0.001). | (Badr El-Din, Abdel Fattah, et al., 2016) |
| 2 | Biobran MGN-3 (25 mg/kg BW i.p. 5x/week, either for 12 or 22 weeks) | Wistar rats + carcinogen (NDEA+CCl4) | Both RBAC treatment regimes kept the liver weight at the normal range and significantly reduced (p<0.01) weight loss caused by the carcinogens. Among RBAC-pretreated rats, the liver tissues showed minimal changes in hepatocyte morphology and histology with no inflammation. In contrast, moderate liver damage was observed in the posttreatment group but with only a few degenerated hepatocytes. | (Badr El-Din, Ali, & Othman, 2016) |
| Abbreviations: BW, body weight; CCI4, carbon tetrachloride; i.p., intraperitoneal; MNNG, methylnitronitrosoguanidine; NDEA, N-nitrosodiethyamine; p.o., per oral; RBAC, rice bran arabinoxylan compound. | ||||
Table 5.
Results from in vitro, in vivo and human studies on RBAC’s synergistic effects with chemotherapeutic agents.
Table 5.
Results from in vitro, in vivo and human studies on RBAC’s synergistic effects with chemotherapeutic agents.
| # | RBAC (dose) | Study Design | Key Findings | Reference |
|---|---|---|---|---|
| 1 | Biobran MGN-3 (100, 500 and 1000 µg/ml) | In vitro. MCF-7 and HCC70 + daunorubicin | RBAC+daunorubicin lowered the IC50 values against MCF-7 cells by 3-, 5- and 5.5-fold at 100, 500 and 1000 µg/ml, respectively. The IC50 of daunorubicin for HCC70 cells was also consistently decreased by 2.5-fold with RBAC. RBAC enhanced drug transport with increased accumulation of daunorubicin in cells. | (Gollapudi & Ghoneum, 2008) |
| 2 | Biobran MGN-3 (1g/day p.o. for 1 year) | RCT. Liver cancer (n=68, RBAC=38, control=30) + TOCE+PEIT | RBAC significantly improved (p<0.01) the treatment response rate (89% vs 80%), lowered the AFP marker (↓38% vs ↑7%), and decreased TV (↓36% vs ↑0.2%). During the 3 years follow-up, the RBAC group showed lower recurrence and higher survival rates. | (Bang et al., 2010) |
| 3 | Biobran MGN-3 (600, 750 and 1000 µg/ml) | In vitro. MCF-7 and 4T1 + paclitaxel | RBAC+paclitaxel lowered the IC50 values against MCF-7 cells by a factor of 100. For 4T1 cells, the IC50 value for paclitaxel at 24 hours decreased by a factor of ~3 at 600 μg/ml of RBAC and up to a factor of ~100 at 1000 μg/ml. | (Ghoneum et al., 2014) |
| 4 | Biobran MGN-3 (40 mg/kg BW for 30 days) | Swiss albino mice + SEC + paclitaxel (2 mg/kg BW) | The combination therapy significantly reduced (p<0.01) TV by 88.3% compared to no treatment. The TV reduction was more pronounced than the effects of either paclitaxel (↓58.9%) or RBAC (↓77.1%) alone. RBAC+paclitaxel also increased inhibition of tumour proliferation, cancer cell apoptosis, and downregulation of Ki-67 expression. | (Badr El-Din, Ali, Alaa El-Dein, et al., 2016) |
| Abbreviations: AFP, alpha-fetoprotein; IC50, half maximal inhibitory concentration; PEIT, percutaneous ethanol injection treatment; p.o., per oral; RBAC, rice bran arabinoxylan compound; RCT, randomised controlled trial; TOCE, transarterial oily chemoembolization; TV, tumour volume. | ||||
Table 6.
Results from animal and human studies on RBAC’s protective effects against the toxicity of chemotherapeutic agents.
Table 6.
Results from animal and human studies on RBAC’s protective effects against the toxicity of chemotherapeutic agents.
| # | RBAC (dose) | Study Design | Key Findings | Reference |
| 1 | Biobran MGN-3 (0, 5, or 50 mg/kg BW p.o. daily for 11 days) | Sprague-Dawley-derived albino rats + cisplatin (9 mg/kg BW) or doxorubicin (10 mg/kg BW) | RBAC prevented weight loss induced by the chemotherapeutic agents (p<0.05). RBAC at 5 mg/kg BW appeared more effective than at the higher dose of 50 mg/kg in preventing the toxicity and side effects of cisplatin and doxorubicin. | (Jacoby et al., 2001) |
| 2 | Biobran MGN-3 (1 mg/day p.o. and i.p. for 28 days) | BALB/c mice + cisplatin (15 mg/kg BW i.p.) | Statistically significant differences (p<0.05) in BW were detected in phases II (weight loss), III (weight gain) and IV (weight stabilising) of both groups of RBAC (i.p. and p.o.) compared to their respective control groups with the RBAC groups showing trends of reduced weight loss and faster weight recovery over time. | (Endo & Kanbayashi, 2003) |
| 3 | Biobran MGN-3 (3g/day p.o. 1 week before and 1 week after chemo cycle for 6 cycles) | RCT. Breast cancer patients (n=50) receiving chemotherapy | The study found significant differences (p<0.001) in the proportions of patients experiencing anorexia/tiredness (RBAC vs control: 20% vs 88%), nausea/vomiting (40% vs 100%), hair loss (28% vs 100%) between the two groups. 84% of the control group experienced weight loss but none in the RBAC group. | (Masood et al., 2013) |
| Abbreviations: BW, body weight; i.p., intraperitoneal injection; p.o., per oral; RBAC, rice bran arabinoxylan compound; RCT, randomised controlled trial. | ||||
Table 7.
Results from animal and human studies on RBAC’s synergy with and protection against the adverse effects of radiation treatment.
Table 7.
Results from animal and human studies on RBAC’s synergy with and protection against the adverse effects of radiation treatment.
| # | RBAC (dose) | Study Design | Key Findings | Reference |
|---|---|---|---|---|
| 1 | Biobran MGN-3 (40 mg/kg BW i.p. q.o.d. for 6 weeks) | Swiss albino mice (n=6) with single dose whole-body γ-Rad | Significantly lesser (p<0.05) BW loss at weeks 1 and 4 after Rad than the control group. RBAC reduced (p<0.05) the kidney and liver organ weight loss induced by Rad at week 1 and prevented anaemia, lymphopenia, neutrophilia, and thrombocytopenia caused by Rad damage. | (Ghoneum et al., 2013) |
| 2 | Biobran MGN-3 (40 mg/kg BW i.p. 5x/week for 3 weeks) | Swiss albino mice + SEC with whole-body X-ray Rad (3 doses) | RBAC+Rad significantly prevented Rad-induced BW loss (p<0.01). RBAC+Rad achieved the highest reduction in TV and TW compared to Rad alone (p<0.01) and RBAC alone (p<0.05). RBAC+Rad increased apoptosis in tumour tissues with the highest cell-cycle arrest while maximising the AI/PrI ratio at 2.2-fold (p<0.01) compared to untreated SEC-bearing mice. Increased apoptotic regulators and their corresponding gene expression were also detected. | (Badr El-Din et al., 2019) |
| 3 | Biobran MGN-3 (40 mg/kg BW i.p. q.o.d. for 6 weeks) | C57BL/6 mice (n=6) with single dose abdominal precision irradiation | After irradiation, RBAC prevented the depletion of mitochondrial respiratory chain complexes and intercellular ATP content in mice’s jejunal and colonic tissues by strengthening the endogenous antioxidative activities and total antioxidant capacity. | (Zhao et al., 2020) |
| 4 | Biobran MGN-3 (3g/day 2 weeks before and 2 months after) | RCT. Head & neck cancer patients undergoing chemoradiotherapy (n=65, RBAC=32, placebo=33) | Significant between-group differences (p<0.05) were in Hb, haematocrit, RBC, platelets, neutrophils, and lymphocytes after 2 months, favouring RBAC. The RBAC group reported significantly (p=0.05) better QoL and lower mortality, blood transfusion, hospitalisation, and metastasis. | (Tan & Flores, 2020) |
| Abbreviations: ATP, adenosine triphosphate; BW, body weight; Hb, haemoglobin; i.p., intraperitoneal injection; q.o.d., every other day; QoL, quality of life; Rad, radiation therapy; RBAC, rice bran arabinoxylan compound; RBC, red blood cells; RCT, randomised controlled trial; TV, tumour volume; TW, tumour weight. | ||||
Table 8.
Results from animal and human studies on RBAC’s synergistic effects with other natural products.
Table 8.
Results from animal and human studies on RBAC’s synergistic effects with other natural products.
| # | RBAC (dose) | Study Design | Key Findings | Reference |
|---|---|---|---|---|
| 1 | Biobran MGN-3 (100, 500 and 1000 mg/ml) | In vitro. MCF-7 cell line + yeast (1:10 ratio) | RBAC significantly increased yeast attachment (54% vs 27%, p<0.001) and phagocytosis rate (72% vs 23%, p<0.01) than control at 0.5 hour. RBAC caused dose-dependent increases in phagocytosis-induced cell death of 35.4%, 40.1%, and 33.04% at 100, 500, and 1000 µg/ml, respectively. | (Ghoneum & Gollapudi, 2005a) |
| 2 | Biobran MGN-3 (100 mg/ml) | In vitro. Monolayer MCF-7 cells + yeast (1:10 ratio) | RBAC increased the magnitude of phagocytising of yeast by MCF-7 cells by 2- to 3-fold after 1 to 4 hours. Culturing with RBAC, yeast, or yeast+RBAC caused 58%, 85%, and 92% cell death, respectively, compared to only 9.5% in untreated MCF-7 cells. | (Ghoneum & Gollapudi, 2005b) |
| 3 | Biobran MGN-3 (50 and 100 μg/ml) | In vitro. MM U266 cell line + curcumin (2.5-10 μM) | RBAC+curcumin caused a significant decrease (p<0.0005) in cell survival compared to either agent alone and achieved an 87% decrease in cell count at 100 μg/ml RBAC and 10 μM curcumin. Combining RBAC (50 μg/ml) with curcumin significantly increased apoptosis (p<0.05) to 20.0, 22.0, and 24.7% at 2.5, 5, 10 μM, respectively, compared to control. | (Ghoneum & Gollapudi, 2011) |
| 4 | Biobran MGN-3 (12 to 45 mg/kg BW 2x/week) for ≥6 months | Cross-section survey. Patients with advanced cancer (n=35) + mistletoe extract (5 mg 2x/week) | Improvement in physical activity (71%) and appetite (66%) were two of the most important effects reported by the patients. For those concurrently treated with conventional treatment (n=24), 70.8% (17/24) also cited reducing side effects as a benefit. | (Hajtó, Horváth, & Papp, 2016) |
| 5 | Biobran MGN-3 (1 g/day for 24 weeks) | RCT. Cancer patients with CFS (n=48, RBAC=24, control=24) + oncothermia | RBAC+oncothermia significantly lowered the posttreatment mean CFQ score (14.6±2.3 vs 23.9±2.3, p<0.01) from baseline. The control group with no treatment reported no significant change in mean CFQ. The mean PGIC score of the RBAC group was 2.1±0.5 (much improved after treatment) compared to 4.3±0.9 (no change) of the control group. | (Petrovics et al., 2016) |
| Abbreviations: BW, body weight; CFQ, Chalder Fatigue Scale; CFS, chronic fatigue syndrome; MM, multiple myeloma; PGIC, Patient Global Impression of Change Scale; QoL, quality of life; RBAC, rice bran arabinoxylan compound. | ||||
Table 9.
A summary of all RBAC randomised controlled trials with survival and/or quality of life as outcome measures.
Table 9.
A summary of all RBAC randomised controlled trials with survival and/or quality of life as outcome measures.
| Authors (Year) | Conditions | N (male/female)age (year) | Interventions | Concomitants | Time points | Outcome measures | Results | ||
|---|---|---|---|---|---|---|---|---|---|
| RBAC | Control | RBAC | Control | ||||||
| Takahara and Sano (2004) | Progressive cancer (multiple types) stage III-IV | 96 (55/41) µ=56.0 |
109 (59/50) µ=53.5 |
Biobran MGN-3 (3g/day p.o., 18 months) | NC | CAT + anticancer drugs with fewer side-effects | 18 months | Survival. QoL: pain, malaise, nausea, appetite. |
SR: T (54.2%, 52/96) > C (33.9%, 19/56, PP), p<0.05 > C (17.4%, 19/109, ITT), p<0.001. QoL (T vs C): pain (-15.9% ≈ -14.0%), malaise (-17.3% ≈ -17.1%), nausea (-13.3% ≈ -14.6%), appetite (+24.2% > +15.2%). |
| Bang et al. (2010) | Liver cancer | 38 (30/8) µ=49±19 |
30 (24/6) µ=51±17 |
Biobran MGN-3 (1g/day, 1 year) | NC | TOCE+PEIT | 12,24,36 months | Survival. | SR (12, 24, 36 months): T (76%, 35%, 11%) > C (63%, 6.7%, 0%), p<0.01 (12 & 24 months) |
| (Masood et al., 2013) | Locally advanced breast cancer | 25 (0/25) |
25 (0/25) |
Biobran MGN-3 (3g/day, before & after each cycle) | NC | CT x 6 cycles | ~18 weeks | QoL: anorexia, nausea, alopecia, weight. | QoL (T vs C): anorexia (20% < 88%), nausea (40% < 100%), alopecia (28% < 100%), weight gain (64% > 0%), weight loss (0% < 84%), p<0.001. |
| Itoh et al. (2015) | Cervical cancer | 7 (0/7) µ=49.9 |
7 (0/7) µ=57 |
Biobran MGN-3 (3g/day, 4 weeks) | Placebo | CT+RT (50.4 Gy in 28 fractions) | 4 weeks | QoL: nausea, diarrhea, diarrhea agent. | QoL: T < C in nausea and diarrhea, diarrhea agent but p>0.05. |
| Petrovics et al. (2016) | Cancer (multiple types) with CFS | 24 | 24 | Biobran MGN-3 (1 g/day, 24 weeks) + Oncothermia | NC | CT and/or RT | 24 weeks | QoL: pain, QLQ-C3 (physical, emotional, general), fatigue (CFQ, PGIC). | QoL: T < C in pain, physical, emotional & general QoL, but no data reported. Fatigue (T vs C): CFQ (14.6±2.3 < 23.2±7.2), PGIC (2.1±0.5 < 4.3±0.9), p<0.001. |
| (20/28) M=66 | |||||||||
| Tan and Flores (2020) | H&N cancer (stage II-IV) | 32 (24/8) M=49 |
33 (29/4) M=54.5 |
Biobran MGN-3 (3g/day 2 weeks before and 2 months after RT) | Placebo | RT or CT+RT (60-70 Gy in 30-35 fractions) | ~18 weeks | Survival. QoL: weight, QLQ-C3 H&N35 (general), radiation toxicity. |
SR: T (0%, 0/32) > C (33.3%, 11/33), p<0.001. QoL: p>0.05 for weight loss and radiation toxicity. General QoL, T (1.53±0.24) < C (1.72±0.33), p<0.019. |
| Abbreviations: C, comparator; CAT, complementary and alternative therapies; CFQ, Chalder Fatigue Scale; CFS, chronic fatigue syndrome; H&N, head and neck; NC, no comparator; CT, chemotherapy; ITT, intention to treat; M, median; µ, mean; PEIT, percutaneous ethanol injection treatment; PGIC, Patient Global Impression of Change Scale; PP, per protocol; QLQ-C3, European Organisation for Research and Treatment of Cancer’s Quality of Life Questionnaire version 3.0; QoL, quality of life; RT, radiation therapy; SR, survival rate; T, treatment; TOCE, transarterial oily chemoembolization. | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



