
Submitted:
17 October 2023
Posted:
17 October 2023
You are already at the latest version
Abstract
Viviparity is made possible by the placenta, a structure acquired relatively recently in the evolutionary history of eutherian mammals. Compared to oviparity, it increases the survival rate of the fetus, owing to the eutherian placenta. Questions such as “How was the placenta acquired?” and “Why is there diversity in placental morphology among mammalian species?” remain largely unsolved. Our present understanding of the molecules regulating placental development remains unclear, owing in no small part to the persistent obscurity surrounding the molecular mechanisms underlying placental acquisition. Many genes that characterize eutherian placental morphology likely evolved to be expressed at the fetal-maternal interface concurrently with those during embryogenesis. Therefore, identifying these genes, how they were acquired, and how they came to be expressed specifically at the fetal-maternal interface will shed light on some crucial molecular mechanisms underlying placental evolution. Exhaustive studies support the hypothesis that endogenous retroviruses (ERVs) could be evolutional driving forces for trophoblast cell fusion and placental structure in mammalian placentas including those of the bovine species. This review focuses on bovine ERVs (BERVs) and their expression and function in the placenta.
Keywords:
cattle
; placenta morphology
; ERVs (endogenous retroviruses)
1. Introduction
Animals reproduce their offspring in three different ways: oviparous, in which the eggs hatch from externally laid eggs; ovoviviparous, in which the eggs are retained in the parent’s body and hatch just before birth; and viviparous, in which the eggs are kept in the body of the parent (usually the mother) without a shell and are born alive. Viviparity, which enables the maternal maintenance of the embryonic environment, is a reproductive strategy observed in various groups of vertebrates, including bony fishes, elasmobranchs, amphibians, reptiles, and mammals [1]. This primitive structure and function of the placenta, facilitating viviparity, was acquired relatively early in the course of vertebrate evolution, with a substantial fraction of vertebrate species developing this capability [2,3].
How did mammals evolve from egg-laying to in-utero incubation, and how did the proto-placental function (primitive viviparous) develop into the mature placenta of today? The transition from ovoviviparity to viviparity and the subsequent emergence of placentation must have required considerable changes in the morphology and physiology of the reproductive tract. For example, monotremes, including the platypus and echidna, are oviparous mammals. They lay thin, low-mineral eggs that typically hatch about ten days after being laid. During a six-month period in the uterus, the developing young is nourished by maternal secretions through a simple yolk sac placenta [4]. Therian mammals, a lineage that encompassing marsupials and eutherians (all extant mammals except monotremes and marsupials), evolved the capacity for live birth (viviparity) during this evolutionary process. They abandoned both the mineralized eggshell and yolk and ultimately developed a placenta. It is worth noting that marsupials significantly differ from eutherians in that their gestation period is relatively short (an average of 25 days), with pregnancy essentially completed within a single estrous cycle. In contrast, eutherian pregnancies are prolonged, lasting up to 670 days (with an average of 131 days) without estrous cycles [4]. The oldest eutherian, Juramaia sinensis, acquired a placenta approximately 160 million years ago [5], while the oldest known mammal to date, Adelobasileus cromptoni, has been dated to about 225 million years ago [6]. In the 65 million years between the birth of the first mammals and the acquisition of the placenta, did they acquire new genes to form the placenta? If so, how did mammals acquire those genes? Or did mammals express and exapt existing genes in the placenta? It was thought that placentation occurred only once in the common ancestor of mammals, but it is now thought to have happened independently many times in already-diverged classes and families of mammals [7]. Thus, the acquisition of the placenta in vertebrates represents clear cases of convergent evolution, both at the macro level such as organ acquisition, and at the molecular level such as through gene acquisition and exaptation [8].
Many of the genes that characterize placental morphology in eutherians likely evolved to be expressed at the fetal-maternal boundary, concurrently with embryogenesis. Consequently, identifying these genes and the mechanisms through which they acquired specific expression at the fetal-maternal interface should illuminate essential molecular mechanisms underlying placental evolution. Moreover, conducting a comprehensive comparative study of genes expressed in the placenta across various animal species may unveil common genes necessary for primitive placental formation or placentation, as well as sets of genes that have either been newly acquired or have lost their functions in different species. Mika and colleagues compared the genes used by the human endometrium with those expressed by endometria of 32 other species, including monkeys, marsupials, other mammals, birds, and reptiles [9]. The analysis revealed that humans use nearly 1,000 genes that other animals do not use [9]. These genes are involved in placental invasion, vascular growth, and immune system regulation [9]. Elucidating the genetic and molecular mechanisms underlying these phenotypic differences between species will lead to a better understanding of the diversity of placental morphology.
2. Diversity and Classification of Placenta in Mammals
Although the role of the placenta is to nurture the fetus, no other organ has a structure as different as that of the placenta among mammals. The genes responsible for viviparity could serve as the foundation for the initial development of primitive placenta during mammalian evolution, with subsequent independent acquisitions of new genes in various species leading to species-specific placental morphologies (Figure 1). Eutherian placentas can be classified based on several criteria, including their shape (number and distribution of nutrient exchange regions on the placental surface), invasiveness (number of maternal tissue layers separating maternal blood and fetal tissue), interdigitation (degree of contact between fetal and maternal tissue in nutrient exchange regions), placental weight relative to neonatal weight, and the relative direction of maternal and fetal blood flow [10]. In fact, eutherian placentas can be histologically classified according to the degree of syncytialization (mode of distribution of villi) into epitheliochorial placenta (Diffuse placenta) in pigs and horses, synepitheliochorial placenta (Cotyledonary placenta) in cattle and sheep, endotheliochorial placenta (Zonary placenta) in dogs and cats, and hemochorial placenta (Discoidal placenta) in humans and mice (Table 1). This classification depends on the morphology of the fetal-maternal interface. Syncytialization is the fusion of trophoblast cells to form syncytia (syncytiotrophoblasts) on the surface of the chorioallantoic membrane, as in the human placenta. The most important barriers to elucidating the origin of evolutionarily novel and placental phenotypes are the absence of transient structures among extant lineages and a lack of experimental systems that allow detailed functional studies. Differences in gene expression underlie changes in anatomical structure, suggesting that changes in gene expression at the maternal-fetal interface underlie pregnancy traits specific to each species. On the other hand, genes commonly expressed in connection with placentas of all mammals may be the genes responsible for proto-placentation (Figure 1). In fact, Armstrong et al. have identified a set of 115 core genes expressed in the placenta of 14 mammalian species [11].
Mechanisms underlying placental innovations encompass the recycling of proteins already expressed within the tissue (co-option), the induction of genes expression typically observed elsewhere in the organism (recruitment), and the introduction of novel genes into the organism’s genome through gene duplication or retroviral insertion (horizontal transfer). It was hypothesized that mammals have developed species-specific placental morphology by harnessing the uniquely acquired ERVs within each lineage. Peg10 and Peg11/Rtl1, responsible for fundamental placental function, were present in common ancestral mammals. Subsequently bovine acquired the retrovirus-derived genes individually. We have identified 7,624 ERV-derived genes in the bovine genome ref. [52]. Among these, 284 ERVs, including BERV-K1, BERV-K2, BERV-K3, Syncytin-Rum1, BERV-P, bERVE-A and bERVE-B, are expressed in cotyledonary placenta due to epigenetic regulations, maternal-fetal conflict ref. [27] and gene replacement through a Baton pass ref. [63]. These factors collectively contribute to the development of ruminant-specific placental morphology. Mya: million years ago
3. The Principle of Genomic Rewiring During Waves of Retroviral Infection
There have been several major genome-size expansions in the lineage from the beginning of life to the rise of human beings. Two primary mechanisms responsible for increasing genome size are recognized: DNA duplication and the incorporation of retrotransposons. DNA duplication events have led to major innovations in the history of life, giving rise to eukaryotes (2 billion years ago), multicellular organisms (1 billion years ago), and vertebrates (500 million years ago). Subsequently, there was an explosion of retrotransposons about 50 to 40 million years ago, which is believed to have driven species diversification and increased the complexity of reproductive strategies, including the diversification of placental morphology [12]. Traditionally, retrotransposons have received little attention in discussions of life's evolution. However, it has become increasingly evident that retrotransposons play a significant role in shaping genomes, leading to outcomes such as genome size expansion and various effects on genetic function. These effects encompass the disruption of gene function through insertion and the potential for genes to acquire new functions. Thus, genomes have co-evolved with transposable elements (TEs) and have devised strategies to regulate uncontrollable elements while extracting novel functions from their newly inserted nucleotides.
Although different types of TEs are found in various organisms, all major TEs can be classified into two main categories, elements that can be transposed via a DNA intermediate and a cut-and-paste mechanism (transposons), and those using an RNA and a copy-paste mechanism (retrotransposons) [13]. Given that TEs make up a substantial portion of animal genomes, comprising approximately 27% in the bovine genome [14], they are instrumental in driving evolutionary changes in genome size and composition [15]. Host organisms share numerous repetitive sequences, encompassing genes, cis-regulatory elements, and chromatin domain boundaries. These shared sequences have the potential to alter gene regulatory networks and, moreover, are partially responsible for morphological evolution as in mammals [16]. Endogenous retroviruses (ERVs) belong to the TE and are considered to have originated from long terminal repeat- (LTR-) type retrotransposons [17]. The genetic changes underlying the evolution of gestation appear to be associated with TEs that invaded and became integrated into early mammalian genomes. ERVs, in particular, are thought to have originated from ancient viruses that infected primordial germ cells or their products, including oocytes, sperm, and fertilized eggs in mammals and other vertebrates [18]. ERVs have become integral components of host genome, resulting from waves of retroviral infection in the host's ancestors, insertion of retroviral sequences into germline DNA, and vertical transmission from one generation to the next [18]. Remarkably, retrovirus-derived sequences account for 8.8% of the human genome [19], 10% of the mouse genome [20] and 18% of the bovine genome [21]. A study on the location and expression of bovine ERVs, as well as the characteristics of neighboring genes, has revealed that up to 1,610 bovine genes contain bovine ERVs (BERVs) inserted within their introns [22]. Most of these BERVs are oriented in the antisense direction, and several genes located in proximity to BERVs appear to be associated with viral response and chromatin assembly [22].
ERVs are genomic elements present in a wide range of vertebrates, spanning from basal vertebrates like sharks and rays to mammals [23]. While LTR-type retrotransposons closely resemble ERVs on the sequence structures, ERVs differ because they retain traces of envelope (env) genes in the sequence, which enables particle formation. Many ERVs become inactivated through nucleotide insertions, deletions, substitutions, and epigenetic modifications, including DNA methylation and histone modifications [24,25]. Nevertheless, some ERV open reading frames (ORFs) are expressed as virus-derived proteins within host cells [26]. Over time, hosts have undergone alterations in these sequences, limiting their ability to reintegrate randomly into the genome (transposition) and producing virus-like particles with reduced effects or proteins with functions distinct from those of the host. As Haig [27] points out, the phenotypic effects of a gene in one organism can influence the fate of the same gene in another organism, highlighting the significance of genes' evolutionary roles over the organisms themselves. The appearance of changes in the expression of existing genes is the result of changes in evolutionary interests across species and can be predicted as a functional evolution of the placenta, but this alone cannot explain the full diversity observed in placental morphology. Furthermore, it's possible that maternal-fetal conflicts have led to the removal of once-infected ERVs. Additionally, newly acquired genes, including species-specific ERVs, may further contribute to the generation of morphological diversity in the placenta (as illustrated in Figure 1)."
4. Peg10, an LTR-type Retrotransposon Commonly Expressed in Marsupial and Eutherian Placentas
The placenta is highly diverse among mammalian species, yet little is known about how it has evolved and what consequences the evolution of specific placental structures has for the mother and fetus. The acquisition of placental-like structures is thought to be due to the acquisition of the paternally expressed gene 10 (Peg10) derived from a LTR-type retrotransposon about 160 million years ago [28]. Knockout mice lacking the Peg10 show early embryonic lethality with placental defects, indicating the importance of this gene in placental development [29]. This gene is highly conserved among mammalian species and is evolutionarily new, being present only in marsupials such as kangaroos and eutherian groups such as humans and mice. In addition, an essential paralog of Peg10, Peg11/retrotransposon-like 1 (Rtl1), is conserved only in eutherians [30], and this gene is essential for maintaining fetal capillaries [31]. Peg10 and Peg11/Rtl1 are imprinted genes believed to have originated from a Ty3/Gypsy LTR-type retrotransposon family, which encodes gag- and pol-like domains [28,29,30,31]. Peg10 is found in both marsupials and eutherian lineages and contributes to trophoblast cell growth during early placentation [11]. On the other hand, Peg11/Rtl1 is exclusive to eutherians and plays a critical role in maintaining endothelial cells in fetal capillaries during late embryonic development. It is evident that both eutherian Peg10 and Peg11/Rtl1 are key factors in establishing viviparity. However, it is apparent that these two genes alone place few constraints on the diversity of placental morphology.
5. The env Genes, Syncytin and Syncytin-like, Essential for Placentation in Mammals
Many studies have shown that ERVs play an important role in developing eutherian placentas and trophoblast cells and suggest the possibility that different species may have utilized ERVs of different origins or of the same origin during evolution. It could be hypothesized that utilizing newly acquired ERVs, each species has evolved the placenta independently (Figure 1), although maternal-fetal conflict through the use of TEs other than ERVs and selective pressures on existing and newly acquired genes could not be excluded. The env gene syncytin, essential for placentation, has been inserted into the genomes of humans [32,33,34,35,36], mice [37,38,39], rabbits [40], dogs [41], cat [41], sheep [42,43,44,45,46], cattle [47,48,49,50,51,52] or marsupials [53]. All identified syncytin env genes share functions such as cell fusion and immunosuppression in mammals, although the genomic sequences and loci differ among species. However, both the ERV evolutionary pathway and the extent to which ERVs function in placentation remains unknown.
6. Morphology of the Bovine Placenta
In contrast to primates such as humans and monkeys, as well as mice, in most ruminants, implantation into the endometrial epithelium and subsequent placentation do not occur immediately after blastocyst formation [54]. The fertilized egg spends a long period in the uterine lumen before finally attaching to the endometrial epithelium and prompting the subsequent formation of placental structures. Ruminants such as cows and sheep have a unique placental variation known as synepitheliochorial placentas, that develop pre-determined sites of the uterine wall termed caruncles and are characterized by the presence of about 100 placentomes, a unique structure with both embryonic-maternal tissues [55]. These placentas can be considered to have a semi-invasive type of placentation [55]. The 'syn-' prefix signifies the contribution of BNC-derived syncytial trophoblast, while the term 'epitheliochorial' underscores the presence of extensive regions with simple placental cell attachment in the definitive placenta [56]. Bovidae is divided into seven subfamilies, including the subfamily Bovine, the subfamily Goatinae, and the subfamily Impala, and the placentas of all subfamilies are plexiform polyplacental, consisting of numerous placental segments. Histologically, the fetal side of the placenta has three characteristic cell types in ruminants: uninucleate trophoblast cells (UNC), BNCs, and trinucleate cells (TNCs) [57]. In cattle and other ruminants, BNCs begin to emerge around day 20 of gestation. They originate from UNC through processes such as mitotic polyploidization or endoreduplication [55,58]. This process continues throughout gestation, primarily in cotyledons after day 30, resulting in a consistent 15 to 20% of the trophectoderm comprising BNC at various gestational stages [59]. In the placentome, which consists of maternal curancles and fetal cotyledons, BNC are believed to migrate while maintaining tight junctions with other trophoblast cells. Subsequently they fuse with a uterine luminal epithelium (LE) cell, leading to the formation of TNC in the sub-epithelial-stromal areas [55,58]. This allows secretory granules containing unique placenta-specific hormones, such as placental lactogen and pregnancy-associated glycoproteins, as well as microvesicles, including exosomes, and other factors, to be released via exocytosis into the endometrial stroma throughout gestation [55,60]. Among the bovine family, the form of cell fusion has been considered to differ among animal species, and only the Bovinae subfamily, to which bovines and water buffalo belong, and the Goatinae subfamily, to which goats and sheep belong, exhibit a unique characteristic in which cells on the fetal and maternal sides fuse together. In the bovine subfamily, BNCs and endometrial cells fuse one-to-one to form TNCs, while in the goat subfamily, multiple BNCs fuse with one endometrial cell to form multinucleate cells [58]. This characteristic of cell fusion in the placenta of bovine subfamily animals is thought to be due to bovine endogenous retrovirus K1 (BERVK1, also known as Fematrin-1), described below [51]. It has long been believed that the BNCs fuse with endometrial cells on the surface of the placenta on the maternal side, which invade the endometria and transport hormones to the mother, thereby maintaining pregnancy [58]. It was recently shown by two laboratories that the fusion between trophoblast cells in sheep is not limited to the formation of BNCs, and new BNCs involve the lateral fusion between growing syncytial plaques [61,62].
7. Expression of env-Derived Endogenous Retroviruses in Bovine Placenta
Although the placenta performs the same function in all mammals, the env-derived genes responsible for trophoblast fusion vary among mammalian species and had remained largely unidentified in bovines.
Syncytin-Rum1 [50] is env-derived ERV specifically expressed in binucleated trophoblast cells, of which integration occurred more than 30 million years ago in the common ancestor of ruminants [50]. Syncytin-Rum1 has undergone a purifying selection, remaining conserved in most ruminants, except for Tragulidae (mouse-deer), whose ruminant ancestor diverged from others about 50 million years ago [50] (Table 2). While Syncytin-Rum1 is expressed in both bovine and sheep placentas, its expression level is notably higher in sheep compared to cattle [50]. BERV-K1 env is also expressed in binucleated trophoblasts during cattle gestation and, 25.3 to 18.3 million years ago, BERV-K1 infected only the bovine subfamily ancestors [51]. It is probable that structural similarity seen among ruminants’ placentas could be due to the function of Syncytin-Rum1. The integration of both Syncytin-Rum1 and BERV-K1 into the bovine genome (as shown in Figure 1 and Table 2) suggests the possibility of successive integrations of ERVs with homologous functions, at least in the bovine. Furthermore, while Syncytin-Rum1 exhibits high expression in the colon, cecum, rectum, kidney, mammary gland, and testis, BERV-K1 is expressed in most tissues (Figure 2). These findings imply that newly acquired ERVs could contribute more effectively to placental morphogenesis or cellular function than preexisting genes. Consequently, we refer to these consecutive ERV acquisitions as the 'baton pass' hypothesis. For more detailed information, refer to the following review [63].
While Syncytin-Rum1 exhibits fusogenic activity specifically under acidic conditions [50], BERV-K1 exhibits much higher cell-fusion activity than Syncytin-Rum1 under physiological conditions [51,64]. It is possible that BERV-K1 became a key player in bovine placentation (Table 2). Interestingly, BERV-K1 also played a role in the formation of trophoblast hybrid cells in heterologous trophoblast cells (sheep trophoblast cells) [62]. In addition to Syncytin-Rum1 and BERV-K1, other ERV envs such as BERV-K2 [48], bERVE-A [49], and bERVE-B [49] have been identified. BERV-K1 and BERV-K2 share similar amino acid sequences and belong to the Beta retrovirus genus [65]. BERV-K2 encompasses all coding sequences, including gag, pro-pol, and env, whereas BERV-K1 retains only the env coding sequence [48]. BERV-K1 and BERV-K2 are expressed throughout different stages of pre-implantation development [66]. Their higher expression levels are detected in embryonic blastomeres (from the two-cell to 16-cell stages), with significantly lower ERV expression observed at the more differentiated blastocyst stage [66]. Notably, BERV-K2 env does not induce cell-to-cell fusion with endometrial cells [51,64], attributed to a failure of env glycoprotein maturation [65]. These varying properties of ERV envs may contribute to differences in cell fusion between animal species.
Since the release of the bovine genome information, numerous in silico analyses have been conducted to detect ERVs [22,48,50,52,67,68,69]. A search for intact env genes in the Bos taurus genome (Bos taurus_UMD3.1 version) identified 18 candidates from five endogenous retrovirus families, including Syncytin-Rum1 [50] and another research identified BERV-K1 [48] and BERV-K2 [48]. Three computational tools (BLAST, LTR_STRUC, and Retrotector©) were used for genome-wide detection of endogenous retroviruses (ERVs) in the bovine genome (Bos taurus_Btau_3.1) [67]. A total of 13,622 putative ERVs were initially identified, but only 1,532 were further identified using two or three additional programs [67]. Our group similarly analyzed the bovine genome (Bos taurus_Btau 4.0) and identified 7,624 ERV-derived genes, including 1,542 env-derived genes [52]. Of these BERV envs, 18.4% (284 genes) were expressed in conceptus on day 22 of pregnancy, and approximately 4% (63 genes) of the env-derived genes in the genome were detected on all days studied (days 17, 20, and 22 of pregnancy) [52]. Of these env-derived genes, the sequence of the env-derived gene with the longest ORF (designated BERV-P env) was similar to the Syncytin-Car1 gene found in dogs and cats [52]. It is thought that BERV-P integration occurred about 16.9 to 7 million years ago [52]. However, BERV-P env was not expressed specifically in the peri-implantation conceptus but in almost all tissues examined, like BERV-K1 (Figure 2). These results suggest that placentation depends on various retrovirus-derived genes that may have replaced endogenous predecessor genes during evolution, upon which the question of placental diversity in ruminants could be answered [63].
ERVs comprise three different classes depending on their relationship with their exogenous counterparts [70]: Class I is related to Epsilonretrovirus and Gammaretrovirus; Class II to Alpharetrovirus, Betaretrovirus, Deltaretrovirus and Lentivirus; and Class III to Spumavirus. In cattle, Xiao et al. had identified sequences from different retrovirus families (BERVγ4, γ7, γ9 - class, I.; and BERVβ3 - class II) using a specialized PCR technique targeting the pro/pol complex [68]. These retrovirus families were named based on their resemblance to sheep ERVs [68]. BERVγ4 was the most common family, originating approximately 13.3 million years ago and found to be distantly related to gamma retroviruses [71]. While full-length BERVγ4 provirus sequences were found in specific chromosomes, partial sequences were distributed throughout the bovine genome [71]. The bovine genome also contained a full-length BERVβ1 provirus with a standard organization [72]. The most conserved BERVβ3 provirus contained multiple stop codons and closely resembled the HERV-K human family [73]. Whether these ERVs are expressed in bovine placenta has not been evaluated. Re-examination of the bovine genome (Bos taurus_UMD3.1 version) using LTRharvest and LTRdigest further detected class I BoERV25-BoERV27 and class II BoERV28-BoERV30 [69]. However, whether these ERVs are expressed in the bovine placenta has not been examined. The availability of new bovine genomic data on ERVs will help to confirm or discover new insights into the evolutionary implications of ERVs demonstrated in model animals.
Endogenous Jaagsiekte retroviruses (enJSRV) exist in the bovine genome [47], but whether enJSRV is expressed in the bovine placenta has not been investigated. It has been reported that enJSRV env promotes trophoblast cell fusion in the sheep placenta through the activation of the PKA/MEK/ERK1/2 signaling pathway [74].
bERVE-A envs have been identified as transcripts in bovine placenta and trophectoderm [49]. bERVE-A exhibited approximately 94% homology to a sequence similar to human syncytin and featured an ORF comprising 107 amino acids [49]. bERVE-A env was highly expressed in post-implantation conceptuses (D20 and D22), with no expression was observed in tissues or organs other than the ovary, testis, rectum, or spleen (Figure 2). Expression of bERVE-A was detected at a low level in the conceptuses from day 17 to 19 of gestation, with an increase in expression observed in fetal membranes up to day 30 of gestation [49]. bERVE-A is expressed preferentially in BNCs [57]. In addition, bERVE-A was rarely expressed in the endometrium throughout the estrus cycle [49]. This gene contains a syncytin 1-like SU domain and ASCT2 binding domain, but it does not possess fusogenic activity due to the loss of the fusion peptide [49]. It is suggested that bERVE-A may have arisen as a result of BNC formation rather than serving as an initiator of cell-to-cell fusion, as it lacks an intact envelope sequence [49]. On the other hand, bERVE-B is expressed in most organs, but its expression in conceptuses was very weak (Figure 2). Differences in these retroviral gene integrations and degrees of expression may account for some changes in the placental structures and/or functions among the placentas of different ruminants.
8. Regulatory Mechanisms involved in BERV Gene Expression in the Bovine Placenta
In the human and mouse placentas, Suppressyn (SUPYN) originated from a retrovirus distinct from Syncytin-1, interacts with the syncytin-1 receptor (ASCT2/SLC1A5), leading to the inhibition of cytotrophoblast cell fusion [75]. It should be noted, however, that SUPYN or any SUPYN-like sequences are absent in the bovine genome. Consequently, the relationship between SUPYN and BERVs could not be discussed. In a study by Kitao et al. [76,77], functional syncytin post-transcriptional regulatory elements (SPREs) were identified in several syncytin genes. These elements were suggested to enhance the expression of viral genes previously repressed due to inefficient codon frequencies or repressive elements within the coding sequence. This discovery raises the possibility that SPREs sequences are present in syncytin-like genes, including BERV-K1, Syncytin-rum1, and BERV-P in bovines. Additionally, while a transcription factor binding site for Rxra has been examined in the thyroid gland, its assessment in the uterus or placenta remains unexplored [22]. This underscores the need for further investigation into the regulatory mechanisms governing ERV gene expression in these specific contexts.
The expression of human ERV (HERV) is intricately regulated at the LTR level, where these LTRs act as promoters for HERV expression [78]. They contain essential RNA polymerase II regulatory sequences [79,80] and numerous transcription factor binding sites [81], through which LTRs can interact with nuclear transcription factors [82]. Notably, recombination events have led to the presence of solo LTRs in the human genome, excising the rest of the provirus [83]. Remarkably, as much as 85% of HERVs have been removed through these recombination events [84], and most HERV loci now exist as solo LTRs. These solo LTRs have the ability to serve as promoters in both sense and antisense directions [85], potentially influencing host gene expression [86,87]. As a result, LTRs play a crucial role in epigenetic modifications, ultimately governing the regulation of both HERV and human gene expression. In the human placenta, DNA methylation levels are generally reduced compared to other tissues, reflecting a higher proportion of HERV LTRs functioning as tissue-specific promoters within placental tissue [88]. Particularly, the CpG island of the 5' LTR is hypomethylated in placental cells, contrasting with its hypermethylated state in other tissues [89]. It should be noted that the investigation into ERV silencing and transcriptional regulation has primarily focused on humans. It is essential to acknowledge that while the mechanisms and principles governing ERV transcriptional regulation in one species may provide insights into another, there can be significant differences among species. Therefore, studies on the regulation of BERV expression by LTR must be examined using bovine placenta and bovine cells.
As shown in Figure 2, ERV expression in cattle is not exclusive to the placenta. It is conceivable that ERVs may have played roles in various organs or cell types, with some later adapting to serve in placental functions. To understand their function in the placenta, it is important to identify the cell types expressing these ERVs in different organs and their respective functions. While in silico analysis has identified expression of numerous ERVs in the early bovine placenta, the specific roles of individual ERVs in placentation remain unclear. Additionally, there has been limited research into the regulatory mechanisms governing ERV expression in cattle. One reason for this lack of clarity is the scarcity of studies on the cellular and molecular mechanisms of bovine placentation compared to other animal models. Elucidation of the cellular and molecular mechanisms governing trophoblast differentiation and function in the bovine placenta will undoubtedly contribute to a more de-tailed understanding of the role of ERVs. A recent study [90] demonstrated the mechanism associated with the differentiation of trophoblast MNC into BNC. Additionally, there have been reports of the successful establishment of bovine trophoblast stem cells capable of generating two distinct functional trophoblast cell types [91]. Moreover, our ongoing research focuses on generating trophectoderm cells from bovine induced pluripotent stem (iPS) cells and identifying expressed ERVs. The data we collect will significantly contribute to elucidating the ERVs responsible for placental morphology characteristics in bovine species and potentially beyond.
Total RNA was extracted from tissues of Japanese black cattle (gestation Day 190) at NIAS, Ibaraki, Japan, excluding the uterus, ovary, or testis. mRNA from the uterus, ovary, and testis was sourced from Zyagen, San Diego, USA. Conceptuses (Days 17, 20, and 22, with day 0 denoting the day of estrus) were collected at Zen-noh Embryo Transfer Center, Hokkaido, Japan (For more details, refer to [52]). For reverse transcription, 500 ng of total RNA was used, following the manufacturer's manual of the ReverTra Ace qPCR RT Master Mix with gDNA Remover (Toyobo, Osaka, Japan). The resulting cDNA was diluted 1:10 with DNase/RNase-free water and subsequently employed for qPCR with specific primers. The primer sequences were designed based on previously published information [refs. 49, 50, 52]. qPCR was conducted using the THUNDERBIRD Next SYBR qPCR Mix (Toyobo) and the Thermal Cycler Dice Real Time System (Takara Bio, Shiga, Japan). Relative expression levels of target genes were determined using the Delta-Delta Ct method and normalized to GAPDH mRNA expression. The results were presented relative to Day 17 of the conceptus, which was set as 1.
9. Expression of gag-Derived Endogenous Retroviruses in Cattle Placenta
ERVs consist of three primary genes: gag, encoding the capsid protein; pro-pol, encoding enzymes for maturation, replication, and insertion; and env, encoding the envelope protein [70]. BERV-K1, BERV-K2, bERVE-A, Syncytin-Rum1, and BERV-P have all been reported to originate from env regions homologous to syncytin. However, ERVs from other regions, such as gag and pol, which may also play a role in ruminant placentation, had not been identified or characterized. In a search for ERV-derived nucleotide structures expressed in the bovine peri-implantation conceptuses [52], we identified ten putative ERV gags located between functional genes in the bovine genome [92]. One candidate ERV gene with gag/pol on bovine chromosome 7 exhibited minimal expression on day 20 during uterine epithelium attachment, increased on day 22 at the onset of placentation, and maintained high expression until at least day 150 [92]. Its expression in bovine trophoblast cells was induced by WNT agonists, a common intracellular signal in placental genes [92]. We named this ERV gag/pol as BERV-K3, which, like other BERVs, was expressed not only in the placenta but also in the skin, kidney, liver, and ileum [92]. Therefore, understanding the function of BERV-K3 in these organs may provide insights into divergence and role in placental function. In addition to BERV-K3, we have identified several other ERVs containing the gag/pol region expressed during the peri-implantation period of bovine embryos [92]. Our current research involves gene expression and cDNA cloning of these ERVs with gag/pol in peri-implantation conceptuses and various organs.
10. Conclusions
Throughout the course of evolution, organisms underwent a transition from asexual to sexual reproduction, and subsequently, from oviparity to viviparity. This transition required the emergence of the uterus as a vital organ for nurturing developing fetuses and physiological changes. During this pivotal period, it is conceivable that various retroviruses spread across mammalian species, integrating into their genomes. Prior to the emergence of the placenta, it is plausible that various retroviruses indeed spread across mammalian species and became integrated into their genomes. Over time, these proviruses likely underwent both negative and positive selection, eventually acquiring functional roles in various cell types and organs. It's important to note that most ERVs are not limited to specific tissues; instead, they exhibit varying degrees of expression across a wide range of tissues and organs. In essence, ERVs with pre-existing functions in diverse tissues or organs may have been co-opted to operate at the fetal-maternal boundary within the uterus throughout the course of pregnancy evolution. This adaptation enabled new functions, such as cell fusion activities, to emerge gradually over time, ultimately contributing to the remarkable diversity we observe in placental structures today. Despite the challenges associated with elucidating the functions of ERV genes, particularly due to the absence of homologous genes in model organisms like mice, the shared viral ancestry of ERVs suggests the possibility of shared functional similarities at the molecular level. A comprehensive investigation of this hypothesis could shed light on the evolutionary history of the placenta and unravel the mysteries surrounding its diversity.
Author Contributions
TS: conceptualization, writing, editing, figure making, and funding acquisition; KK: manuscript editing, revision and approval of the final version for submission; KI: supervision and review. All authors have read and agreed to the published version of the manuscript.
Funding
The work is supported by JSPS KAKENHI Grant Number JP 22H02502, awarded to, T.S.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Mr. Robert Moriarty for the English language review.
Conflicts of Interest
The authors declare that the review was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Shine, R. Life-history evolution in reptiles. Annual Review of Ecology Evolution and Systematics 2005, 36, 23–46. [Google Scholar] [CrossRef]
- Van Dyke, J.U.; Brandley, M.C.; Thompson, M.B. The evolution of viviparity: molecular and genomic data from squamate reptiles advance understanding of live birth in amniotes. Reproduction. 2013, 147, R15–R26. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, D.G. Evolution of vertebrate viviparity and specializations for fetal nutrition: A quantitative and qualitative analysis. J Morphol. 2015, 276, 961–990. [Google Scholar] [CrossRef]
- Hughes RL and Hall, L.S. Early development and embryology of the platypus. Philos Trans R Soc Lond B Biol Sci. 1998, 353, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.X.; Yuan, C.X.; Meng, Q.J.; Ji, Q. A Jurassic eutherian mammal and divergence of marsupials and placentals. Nature. 2011, 476, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Zheng, X.; Wang, X.; Cignetti, N.E.; Yang, S.; Wible, J.R. An Early Cretaceous eutherian and the placental-marsupial dichotomy. Nature. 2018, 558, 390–395. [Google Scholar] [CrossRef]
- Roberts, R.M.; Green, J.A.; Schulz, L.C. The evolution of the placenta. Reproduction. 2016, 152, R179–R189. [Google Scholar] [CrossRef]
- Foster, C.S.P.; Van Dyke, J.U.; Thompson, M.B.; Smith, N.M.A.; Simpfendorfer, C.A.; Murphy, C.R.; Whittington, C.M. Different Genes are Recruited During Convergent Evolution of Pregnancy and the Placenta. Mol Biol Evol. 2022, 39, msac077. [Google Scholar] [CrossRef]
- Mika, K.; Marinić, M.; Singh, M.; Muter, J.; Brosens, J.J.; Lynch, V.J. Evolutionary transcriptomics implicates new genes and pathways in human pregnancy and adverse pregnancy outcomes. Elife. 2021, 10, e69584. [Google Scholar] [CrossRef]
- Carter AM and Enders, A.C. Comparative aspects of trophoblast development and placentation. Reprod Biol Endocrinol. 2004, 2, 46. [Google Scholar]
- Armstrong, D.L.; McGowen, M.R.; Weckle, A.; Pantham, P.; Caravas, J.; Agnew, D.; Benirschke, K.; Savage-Rumbaugh, S.; Nevo, E.; Kim, C.J.; et al. The core transcriptome of mammalian placentas and the divergence of expression with placental shape. Placenta. 2017, 57, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Hattori, M.; Yada, T.; Gojobori, T.; Sakaki, Y.; Okada, N. Whole-genome screening indicates a possible burst of formation of processed pseudogenes and Alu repeats by particular L1 subfamilies in ancestral primates. Genome Biol. 2003, 4, R74. [Google Scholar] [CrossRef]
- Kazazian, H.H., Jr. Mobile elements: drivers of genome evolution. Science. 2004, 303, 1626–1632. [Google Scholar] [CrossRef] [PubMed]
- Bovine Genome Sequencing and Analysis Consortium; Elsik, C. G.; Tellam, R.L.; Worley, K.C.; Gibbs, R.A.; Muzny, D.M.; Weinstock, G.M.; Adelson, D.L.; Eichler, E.E.; Elnitski, L.; Guigó, R.; et al. The genome sequence of taurine cattle: a window to ruminant biology and evolution. Science. 2009, 324, 522–528. [Google Scholar]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: from conflicts to benefits. Nat Rev Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Peng, C.; Fang, L.; Wang, Z.; Liu, G.E. Taming transposable elements in livestock and poultry: a review of their roles and applications. Genet Sel Evol. 2023, 55, 50. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Eickbush, T.H. Origin and evolution of retroelements based upon their reverse transcriptase sequences. EMBO J. 1990, 9, 3353–3362. [Google Scholar] [CrossRef]
- Weiss, R.A. The discovery of endogenous retroviruses. Retrovirology. 2006, 3, 67. [Google Scholar] [CrossRef]
- Hoyt, S.J.; Storer, J.M.; Hartley, G.A.; Grady, P.G.S.; Gershman, A.; de Lima, L.G.; Limouse, C.; Halabian, R.; Wojenski, L.; Rodriguez, M.; et al. From telomere to telomere: The transcriptional and epigenetic state of human repeat elements. Science. 2022, 376, eabk3112. [Google Scholar] [CrossRef]
- Jern P and Coffin, J.M. Effects of retroviruses on host genome function. Annu Rev Genet. 2008, 42, 709–732. [Google Scholar] [CrossRef]
- Adelson, D.L.; Raison, J.M.; Edgar, R.C. Characterization and distribution of retrotransposons and simple sequence repeats in the bovine genome. Proc Natl Acad Sci USA. 2009, 106, 12855–12860. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Etxebarria K and Jugo, B.M. Genomic environment and digital expression of bovine endogenous retroviruses. Gene. 2014, 548, 14–21. [Google Scholar] [CrossRef]
- Gifford R and Tristem, M. The evolution, distribution and diversity of endogenous retroviruses. Virus Genes. 2003, 26, 291–315. [Google Scholar] [CrossRef] [PubMed]
- de Parseval, N.; Heidmann, T. Human endogenous retroviruses: from infectious elements to human genes. Cytogenet Genome Res. 2005, 110, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Vargiu, L.; Rodriguez-Tomé, P.; Sperber, G.O.; Cadeddu, M.; Grandi, N.; Blikstad, V.; Tramontano, E.; Blomberg, J. Classification and characterization of human endogenous retroviruses; mosaic forms are common. Retrovirology. 2016, 13, 7. [Google Scholar] [CrossRef]
- Dupressoir, A.; Vernochet, C.; Harper, F.; Guégan, J.; Dessen, P.; Pierron, G.; Heidmann, T. A pair of co-opted retroviral envelope syncytin genes is required for formation of the two-layered murine placental syncytiotrophoblast. Proc Natl Acad Sci USA. 2011, 108, E1164–E1173. [Google Scholar] [CrossRef]
- Haig, D. Genetic conflicts in human pregnancy. Q Rev Biol. 1993, 68, 495–532. [Google Scholar] [CrossRef]
- Kaneko-Ishino T and Ishino, F. The role of genes domesticated from LTR retrotransposons and retroviruses in mammals. Front Microbiol. 2012, 3, 262. [Google Scholar] [CrossRef]
- Ono, R.; Nakamura, K.; Inoue, K.; Naruse, M.; Usami, T.; Wakisaka-Saito, N.; Hino, T.; Suzuki-Migishima, R.; Ogonuki, N.; Miki, H.; et al. Deletion of Peg10, an imprinted gene acquired from a retrotransposon, causes early embryonic lethality. Nat Genet. 2006, 38, 101–106. [Google Scholar] [CrossRef]
- Edwards, C.A.; Mungall, A.J.; Matthews, L.; Ryder, E.; Gray, D.J.; Pask, A.J.; Shaw, G.; Graves, J.A.; Rogers, J.; SAVOIR consortium; Dunham, I. ; Renfree, M.B.; et al. The evolution of the DLK1-DIO3 imprinted domain in mammals. PLoS Biol. 2008, 6, e135. [Google Scholar] [CrossRef]
- Sekita, Y.; Wagatsuma, H.; Nakamura, K.; Ono, R.; Kagami, M.; Wakisaka, N.; Hino, T.; Suzuki-Migishima, R.; Kohda, T.; Ogura, A.; et al. Role of retrotransposon-derived imprinted gene, Rtl1, in the feto-maternal interface of mouse placenta. Nat Genet. 2008, 40, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Blond, J.L.; Lavillette, D.; Cheynet, V.; Bouton, O.; Oriol, G.; Chapel-Fernandes, S.; Mandrand, B.; Mallet, F.; Cosset, F.L. An envelope glycoprotein of the human endogenous retrovirus HERV-W is expressed in the human placenta and fuses cells expressing the type D mammalian retrovirus receptor. J Virol. 2000, 74, 3321–3329. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Lee, X.; Li, X.; Veldman, G.M.; Finnerty, H.; Racie, L.; LaVallie, E.; Tang, X.Y.; Edouard, P.; Howes, S.; et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature. 2000, 403, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Blaise, S.; de Parseval, N.; Bénit, L.; Heidmann, T. Genomewide screening for fusogenic human endogenous retrovirus envelopes identifies syncytin 2, a gene conserved on primate evolution. Proc Natl Acad Sci USA. 2003, 100, 13013–13018. [Google Scholar] [CrossRef] [PubMed]
- Esnault, C.; Priet, S.; Ribet, D.; Vernochet, C.; Bruls, T.; Lavialle, C.; Weissenbach, J.; Heidmann, T. A placenta-specific receptor for the fusogenic, endogenous retrovirus-derived, human syncytin-2. Proc Natl Acad Sci USA. 2008, 105, 17532–17537. [Google Scholar] [CrossRef] [PubMed]
- Lokossou, A.G.; Toudic, C.; Barbeau, B. Implication of human endogenous retrovirus envelope proteins in placental functions. Viruses. 2014, 6, 4609–4627. [Google Scholar] [CrossRef] [PubMed]
- Dupressoir, A.; Marceau, G.; Vernochet, C.; Bénit, L.; Kanellopoulos, C.; Sapin, V.; Heidmann, T. Syncytin-A and syncytin-B, two fusogenic placenta-specific murine envelope genes of retroviral origin conserved in Muridae. Proc Natl Acad Sci USA. 2005, 102, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Dupressoir, A.; Vernochet, C.; Bawa, O.; Harper, F.; Pierron, G.; Opolon, P.; Heidmann, T. Syncytin-A knockout mice demonstrate the critical role in placentation of a fusogenic, endogenous retrovirus-derived, envelope gene. Proc Natl Acad Sci USA. 2009, 106, 12127–12132. [Google Scholar] [CrossRef]
- Dupressoir, A.; Vernochet, C.; Harper, F.; Guégan, J.; Dessen, P.; Pierron, G.; Heidmann, T. A pair of co-opted retroviral envelope syncytin genes is required for formation of the two-layered murine placental syncytiotrophoblast. Proc Natl Acad Sci USA. 2011, 108, E1164–E1173. [Google Scholar] [CrossRef]
- Heidmann, O.; Vernochet, C.; Dupressoir, A.; Heidmann, T. Identification of an endogenous retroviral envelope gene with fusogenic activity and placenta-specific expression in the rabbit: a new “syncytin” in a third order of mammals. Retrovirology. 2009, 6, 107. [Google Scholar] [CrossRef]
- Cornelis, G.; Heidmann, O.; Bernard-Stoecklin, S.; Reynaud, K.; Véron, G.; Mulot, B.; Dupressoir, A.; Heidmann, T. Ancestral capture of syncytin-Car1, a fusogenic endogenous retroviral envelope gene involved in placentation and conserved in Carnivora. Proc Natl Acad Sci USA. 2012, 109, E432–E441. [Google Scholar] [CrossRef]
- Dunlap, K.A.; Palmarini, M.; Adelson, D.L.; Spencer, T.E. Sheep endogenous betaretroviruses (enJSRVs) and the hyaluronidase 2 (HYAL2) receptor in the ovine uterus and conceptus. Biol Reprod. 2005, 73, 271–279. [Google Scholar] [CrossRef]
- Dunlap, K.A.; Palmarini, M.; Varela, M.; Burghardt, R.C.; Hayashi, K.; Farmer, J.L.; Spencer, T.E. Endogenous retroviruses regulate periimplantation placental growth and differentiation. Proc Natl Acad Sci USA. 2006, 103, 14390–14395. [Google Scholar] [CrossRef]
- Dunlap, K.A.; Palmarini, M.; Spencer, T.E. Ovine endogenous betaretroviruses (enJSRVs) and placental morphogenesis. Placenta. 2006, S135–S140. [Google Scholar] [CrossRef]
- Murcia, P.R.; Arnaud, F.; Palmarini, M. The transdominant endogenous retrovirus enJS56A1 associates with and blocks intracellular trafficking of Jaagsiekte sheep retrovirus Gag. J Virol. 2007, 81, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Armezzani, A.; Arnaud, F.; Caporale, M.; di Meo, G.; Iannuzzi, L.; Murgia, C.; Palmarini, M. The signal peptide of a recently integrated endogenous sheep betaretrovirus envelope plays a major role in eluding gag-mediated late restriction. J Virol. 2011, 85, 7118–7128. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Morozov, A.V.; Lagaye, S. Endogenous JSRV-like proviruses in domestic cattle: analysis of sequences and transcripts. Virology. 2007, 367, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Baba, K.; Nakaya, Y.; Shojima, T.; Muroi, Y.; Kizaki, K.; Hashizume, K.; Imakawa, K.; Miyazawa, T. Identification of novel endogenous betaretroviruses which are transcribed in the bovine placenta. J Virol. 2011, 85, 1237–1245. [Google Scholar] [CrossRef]
- Koshi, K.; Suzuki, Y.; Nakaya, Y.; Imai, K.; Hosoe, M.; Takahashi, T.; Kizaki, K.; Miyazawa, T.; Hashizume, K. Bovine trophoblastic cell differentiation and binucleation involves enhanced endogenous retrovirus element expression. Reprod Biol Endocrinol. 2012, 10, 41. [Google Scholar] [CrossRef]
- Cornelis, G.; Heidmann, O.; Degrelle, S.A.; Vernochet, C.; Lavialle, C.; Letzelter, C.; Bernard-Stoecklin, S.; Hassanin, A.; Mulot, B.; Guillomot, M.; et al. Captured retroviral envelope syncytin gene associated with the unique placental structure of higher ruminants. Proc Natl Acad Sci USA. 2013, 110, E828–E837. [Google Scholar] [CrossRef]
- Nakaya, Y.; Koshi, K.; Nakagawa, S.; Hashizume, K.; Miyazawa, T. Fematrin-1 is involved in fetomaternal cell-to-cell fusion in Bovinae placenta and has contributed to diversity of ruminant placentation. J Virol. 2013, 87, 10563–10572. [Google Scholar] [CrossRef]
- Nakagawa, S.; Bai, H.; Sakurai, T.; Nakaya, Y.; Konno, T.; Miyazawa, T.; Gojobori, T.; Imakawa, K. Dynamic evolution of endogenous retrovirus-derived genes expressed in bovine conceptuses during the period of placentation. Genome Biol Evol. 2013, 5, 296–306. [Google Scholar] [CrossRef]
- Cornelis, G.; Vernochet, C.; Carradec, Q.; Souquere, S.; Mulot, B.; Catzeflis, F.; Nilsson, M.A.; Menzies, B.R.; Renfree, M.B.; Pierron, G.; et al. Retroviral envelope gene captures and syncytin exaptation for placentation in marsupials. Proc Natl Acad Sci USA. 2015, 112, E487–E496. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Spencer, T.E.; Johnson, G.A. Interferons and uterine receptivity. Semin Reprod Med. 2009, 27, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Green, J.A.; Geisert, R.D.; Johnson, G.A.; Spencer, T.E. Implantation and Placentation in Ruminants. Adv Anat Embryol Cell Biol. 2021, 234, 129–154. [Google Scholar] [PubMed]
- Wooding, F.B. Current topic: the synepitheliochorial placenta of ruminants: binucleate cell fusions and hormone production. Placenta. 1992, 13, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Klisch, K.; Pfarrer, C.; Schuler, G.; Hoffmann, B.; Leiser, R. Tripolar acytokinetic mitosis and formation of feto-maternal syncytia in the bovine placentome: different modes of the generation of multinuclear cells. Anat Embryol (Berl). 1999, 200, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Wooding, F.B.P. The ruminant placental trophoblast binucleate cell: an evolutionary breakthrough. Biol Reprod. 2022, 107, 705–716. [Google Scholar] [CrossRef]
- Wooding, F.B. Frequency and localization of binucleate cells in the placentomes of ruminants. Placenta. 1983, 4, 527–539. [Google Scholar]
- Wooding, F.B.; Wathes, D.C. Binucleate cell migration in the bovine placentome. J Reprod Fertil. 1980, 59, 425–430. [Google Scholar] [CrossRef]
- Seo, H.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A. Immunohistochemical Examination of Trophoblast Syncytialization during Early Placentation in Sheep. Int J Mol Sci. 2019, 20, 4530. [Google Scholar] [CrossRef]
- Yamada, A.; Ohtsuki, K.; Shiga, N.; Green, J.A.; Matsuno, Y.; Imakawa, K. Epithelial-mesenchymal transition and bi- and multi-nucleated trophoblast cell formation in ovine conceptuses during the peri-implantation period. J Reprod Dev. 2022, 68, 110–117. [Google Scholar] [CrossRef]
- Imakawa, K.; Nakagawa, S.; Miyazawa, T. Baton pass hypothesis: successive incorporation of unconserved endogenous retroviral genes for placentation during mammalian evolution. Genes Cells. 2015, 20, 771–788. [Google Scholar] [CrossRef]
- Nakaya, Y.; Miyazawa, T. Dysfunction of bovine endogenous retrovirus K2 envelope glycoprotein is related to unsuccessful intracellular trafficking. J Virol. 2014, 88, 6896–6905. [Google Scholar] [CrossRef]
- Koshi, K.; Nakaya, Y.; Kizaki, K.; Ishiguro-Oonuma, T.; Miyazawa, T.; Spencer, T.E.; Hashizume, K. Induction of ovine trophoblast cell fusion by fematrin-1 in vitro. Anim Sci. J. 2016, 87, 419–422. [Google Scholar] [CrossRef]
- Khazaee, E.; Farzaneh, N.; Mirshokraei, P.; Tabatabaeizadeh, S.E.; Dehghani, H. Expression of endogenous retroviruses in pre-implantation stages of bovine embryo. Reprod Domest Anim. 2018, 53, 1405–1414. [Google Scholar] [CrossRef]
- Garcia-Etxebarria, K.; Jugo, B.M. Genome-wide detection and characterization of endogenous retroviruses in Bos taurus. J Virol. 2010, 84, 10852–10862. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Park, K.; Lee, H.; Kim, J.; Park, C. Identification and classification of endogenous retroviruses in cattle. J Virol. 2008, 82, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Etxebarria, K.; Jugo, B.M. Evolutionary history of bovine endogenous retroviruses in the Bovidae family. BMC Evol Biol. 2013, 13, 256. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.E. Endogenous Retroviruses in the Genomics Era. Annu. Rev. Virol. 2015, 2, 135–159. [Google Scholar] [CrossRef]
- Xiao, R.; Park, K.; Oh, Y.; Kim, J.; Park, C. Structural characterization of the genome of BERV gamma4, the most abundant endogenous retrovirus family in cattle. Mol Cells. 2008, 26, 404–408. [Google Scholar] [CrossRef]
- Torresi, C.; Casciari, C.; Giammarioli, M.; Feliziani, F.; De Mia, G.M. Characterization of a novel full-length bovine endogenous retrovirus, BERV-β1. Arch Virol. 2015, 160, 3105–3114. [Google Scholar] [CrossRef]
- Xiao, R.; Kim, J.; Choi, H.; Park, K.; Lee, H.; Park, C. Characterization of the bovine endogenous retrovirus beta3 genome. Mol Cells. 2008, 25, 142–147. [Google Scholar] [CrossRef]
- Wang, X.; Liu, S. Endogenous Jaagsiekte sheep retrovirus envelope protein promotes sheep trophoblast cell fusion by activating PKA/MEK/ERK1/2 signaling. Theriogenology. 2022, 193, 58–67. [Google Scholar] [CrossRef]
- Sugimoto, J.; Sugimoto, M.; Bernstein, H.; Jinno, Y.; Schust, D. A novel human endogenous retroviral protein inhibits cell-cell fusion. Sci Rep. 2013, 3, 1462. [Google Scholar] [CrossRef]
- Kitao, K.; Tanikaga, T.; Miyazawa, T. Identification of a post-transcriptional regulatory element in the human endogenous retroviral syncytin-1. J Gen Virol. 2019, 100, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Kitao, K.; Nakagawa, S.; Miyazawa, T. An ancient retroviral RNA element hidden in mammalian genomes and its involvement in co-opted retroviral gene regulation. Retrovirology. 2021, 18, 36. [Google Scholar] [CrossRef]
- Kovalskaya, E.; Buzdin, A.; Gogvadze, E.; Vinogradova, T.; Sverdlov, E. Functional human endogenous retroviral LTR transcription start sites are located between the R and U5 regions. Virology. 2006, 346, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Criscione, S.W.; Zhang, Y.; Thompson, W.; Sedivy, J.M.; Neretti, N. Transcriptional landscape of repetitive elements in normal and cancer human cells. BMC Genomics. 2014, 15, 583. [Google Scholar] [CrossRef]
- Thompson, P.J.; Macfarlan, T.S.; Lorincz, M.C. Long Terminal Repeats: From Parasitic Elements to Building Blocks of the Transcriptional Regulatory Repertoire. Mol Cell. 2016, 62, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Manghera, M.; Douville, R.N. Endogenous retrovirus-K promoter: a landing strip for inflammatory transcription factors? Retrovirology. 2013, 10, 16. [Google Scholar] [CrossRef]
- Akopov, S.B.; Nikolaev, L.G.; Khil, P.P.; Lebedev, Y.B.; Sverdlov, E.D. Long terminal repeats of human endogenous retrovirus K family (HERV-K) specifically bind host cell nuclear proteins. FEBS Lett. 1998, 421, 229–233. [Google Scholar] [CrossRef]
- Hughes, J.F.; Coffin, J.M. Human endogenous retrovirus K solo-LTR formation and insertional polymorphisms: implications for human and viral evolution. Proc Natl Acad Sci U S A. 2004, 101, 1668–1672. [Google Scholar] [CrossRef]
- Belshaw, R.; Watson, J.; Katzourakis, A.; Howe, A.; Woolven-Allen, J.; Burt, A.; Tristem, M. Rate of recombinational deletion among human endogenous retroviruses. J Virol. 2007, 81, 9437–9442. [Google Scholar] [CrossRef]
- Cohen, C.J.; Lock, W.M.; Mager, D.L. Endogenous retroviral LTRs as promoters for human genes: a critical assessment. Gene. 2009, 448, 105–114. [Google Scholar] [CrossRef]
- Dunn, C.A.; Romanish, M.T.; Gutierrez, L.E.; van de Lagemaat, L.N.; Mager, D.L. Transcription of two human genes from a bidirectional endogenous retrovirus promoter. Gene. 2006, 366, 335–342. [Google Scholar] [CrossRef]
- Romanish, M.T.; Lock, W.M.; van de Lagemaat, L.N.; Dunn, C.A.; Mager, D.L. Repeated recruitment of LTR retrotransposons as promoters by the anti-apoptotic locus NAIP during mammalian evolution. PLoS Genet. 2007, 3, e10. [Google Scholar] [CrossRef] [PubMed]
- Reiss, D.; Zhang, Y.; Mager, D.L. Widely variable endogenous retroviral methylation levels in human placenta. Nucleic Acids Res. 2007, 35, 4743–4754. [Google Scholar] [CrossRef]
- Matousková, M.; Blazková, J.; Pajer, P.; Pavlícek, A.; Hejnar, J. CpG methylation suppresses transcriptional activity of human syncytin-1 in non-placental tissues. Exp Cell Res. 2006, 312, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Davenport, K.M.; Ortega, M.S.; Liu, H.; O’Neil, E.V.; Kelleher, A.M.; Warren, W.C.; Spencer, T.E. Single-nuclei RNA sequencing (snRNA-seq) uncovers trophoblast cell types and lineages in the mature bovine placenta. Proc Natl Acad Sci U S A. 2023, 120, e2221526120. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ming, H.; Yu, L.; Li, J.; Zhu, L.; Sun, H.X.; Pinzon-Arteaga, C.A.; Wu, J.; Jiang, Z. Establishment of bovine trophoblast stem cells. Cell Rep. 2023, 42, 112439. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Nakagawa, S.; Bai, H.; Bai, R.; Kusama, K.; Ideta, A.; Aoyagi, Y.; Kaneko, K.; Iga, K.; Yasuda, J.; et al. Novel endogenous retrovirus-derived transcript expressed in the bovine placenta is regulated by WNT signaling. Biochem, J. 2017, 474, 3499–3512. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Hypothesis on the diversity of placental morphology by ERVs.
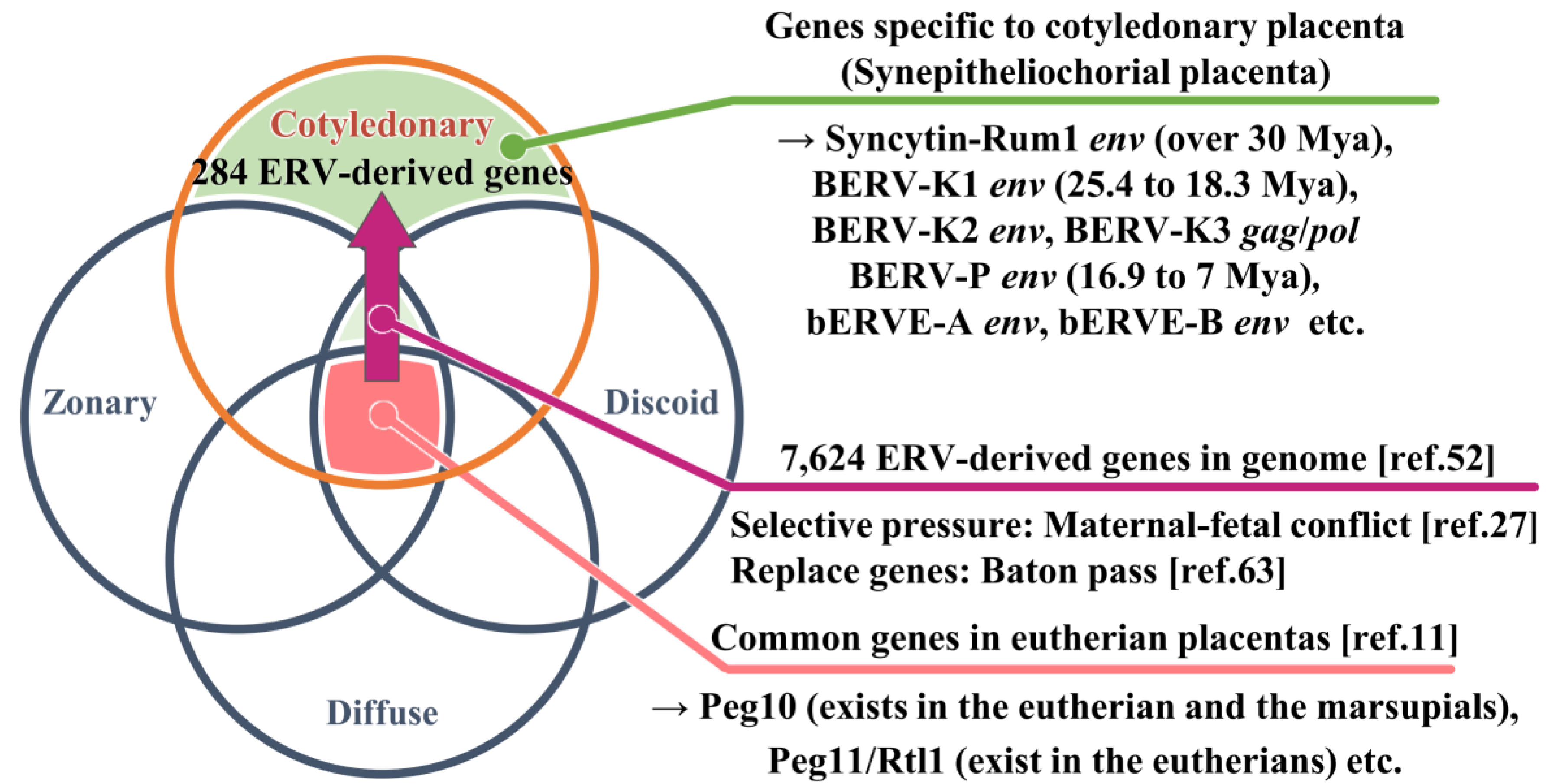
Figure 2.
Relative expression of BERV env genes in bovine tissues and conceptuses.
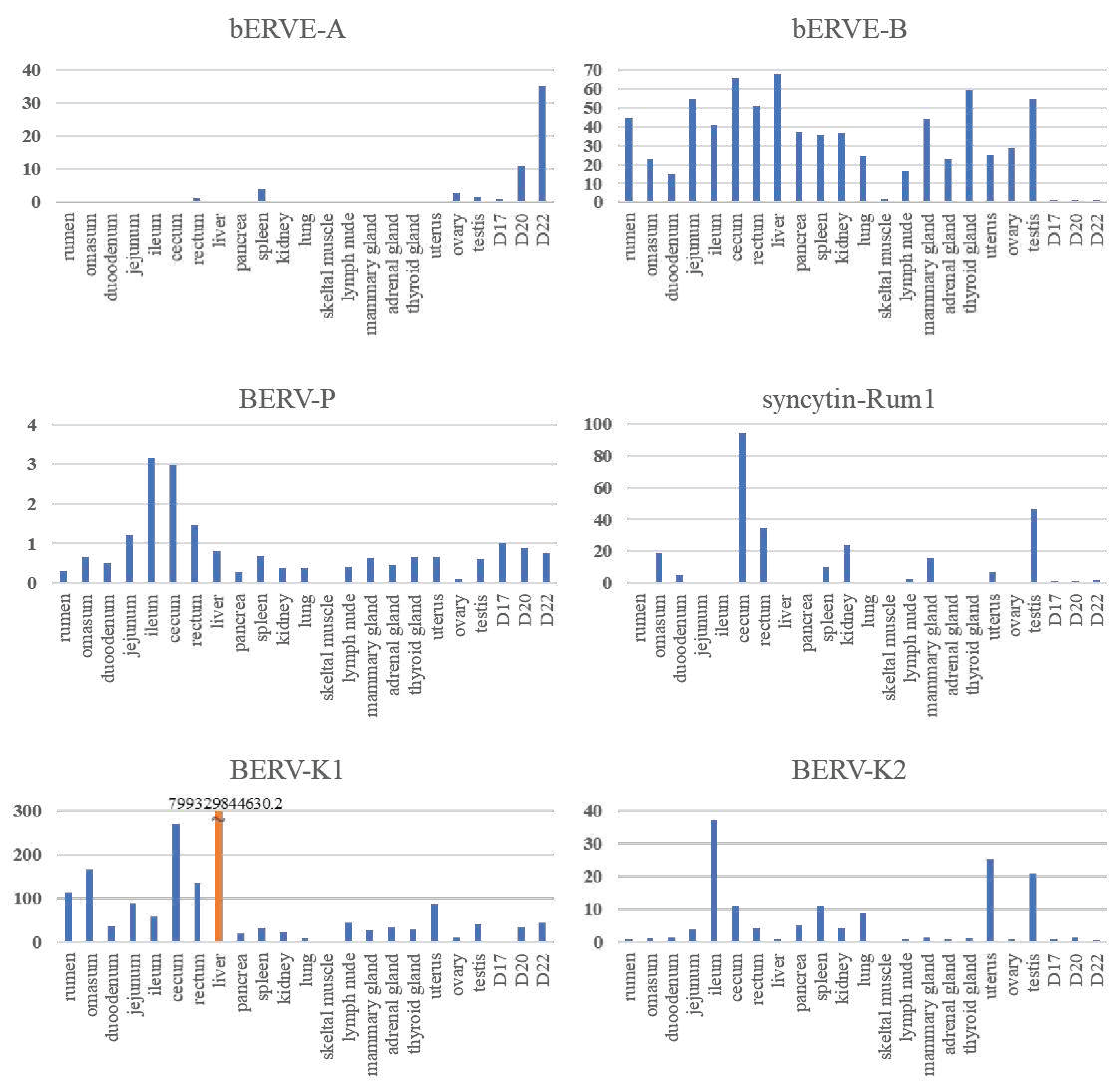

Table 1.
Classification of mammalian placentas.
| Mode of distribution of villi | Degree of syncytialization | Representative species | Type of fused cells |
|---|---|---|---|
| Cotyledonary | Synepitheliochorial | Cow, Sheep | Fetomaternal hybrid |
| Diffuse | Epitheliochorial | Horse, Pig | None |
| Zonary | Endotheliochorial | Dog, Cat | Syncytiotrophoblast |
| Discoid | Hemochorial | Mouse, Human | Syncytiotrophoblast |
Table 2.
ERV genes supposed to play roles in placentation in ruminants.
| Appearance in Host Genome | Species | Expressed Tissues (localization)#1 | Function | References | |
| Syncytin-Rum1 env | over 30 Mya | Most ruminants(except for the family Tragulidae) | Placenta (BNCs) | Fusogenic activity | [50] |
| BERV-K1 env | 25.4 to 18.3 Mya | Bovinae subfamily | Placenta (BNCs), Embryonic blastomeres (two- to 16-cell stage) | Fusogenic activity | [51,64] |
| BERV-K2 env | No data | No data | Emdometrium, Embryonic blastomeres (two- to 16-cell stage) | No data | [48,64] |
| BERV-P env | 16.9 to 7.0 Mya | genus Bos | Conceptus during periattachment (Trophectoderm) | No data | [52] |
| bERVE-A env | No data | Bos taurus | Placenta (BNCs) | No data | [49] |
| bERVE-B env | No data | No data | Endometrium, Placenta | No data | [49] |
| BERV-K3 gag/pol | No data | No data | Conceptus after attachment (Trophectoderm), Placenta | No data | [92] |
#1: Other than showing in Figure 2. Mya: million years ago
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



