
Submitted:
17 October 2023
Posted:
18 October 2023
You are already at the latest version
Abstract
Microbial cell factories offer an eco-friendly alternative for transforming raw materials into commercially valuable products, primarily because of their reduced carbon impact compared to conventional industrial procedures. These systems often depend on lignocellulosic materials, mainly pentose and hexose sugars, richly available carbon reservoirs. Nevertheless, these resources might not always be efficient due to the limited supply, significant cost, and other considerable obstacles. One major hurdle when working with sugars derived from lignocellulosic biomass, especially glucose, is balancing carbon allocation to satisfy energy, cofactor, and other essential component needs for cellular proliferation while maintaining a robust yield. Furthermore, nearly half or more of this carbon is inevitably lost as CO2 during the biosynthesis processes, which serves to generate the energy necessary for maintaining cell growth and other regular metabolic activities. This carbon loss lowers the theoretical production yield and compromises the benefit of reducing greenhouse gas emissions – a fundamental advantage of biomanufacturing. This review paper posits the perspectives of using CO2 from atmosphere, industrial wastes, or the exhausted gases generated in microbial fermentation as a feedstock for biomanufacturing. Both one-step direct CO2 fixation or two-step indirect CO2 fixation and conversion strategies are discussed, which can minimize the carbon loss, significantly increase the carbon yield, and eventually achieve the carbon-neutral or -negative goals. The one-step strategy uses novel metabolic pathway design and engineering approaches to directly fix the CO2 via the pathway toward the synthesis of the desired fermentation products. Due to the limitation of the yield and efficiency in one-step CO2 fixation, the two-step strategy aims to completely avoid carbon loss in biomanufacturing by integrating a first electrochemical fixation of the exhausted CO2 into C1/C2 products such as formate, methanol, acetate, and ethanol and a second fermentation unit to utilize the CO2-derived C1/C2 chemicals or co-utilize both C5/C6 sugars and C1/C2 chemicals for product formation. The great potentials and challenges of using CO2 as a feedstock for further biomanufacturing are also discussed.
Keywords:
Metabolic engineering
; CO2 fixation
; feedstock
; biomanufacturing
; electrochemical catalysis
; microbial electrosynthesis
1. Introduction
Carbon emission to our ecosystem and its accumulation in its highly oxidized state, carbon dioxide (CO2), is a primary contributing factor to global climate change [1]. Since the 1960s, the total CO2 emissions have rapidly increased with a net annual escalation rate of 2.11% in recent years [2]. The push for carbon neutrality necessitates reimagining our feedstock sources. With over 90% of our chemical and fuel needs rooted in fossil feedstocks, there's an impetus to transition towards a more circular industry model. G20 economies have implemented carbon emission taxes ranging from $3 to $60 per ton, which can incentivize the capture of CO2 from industrial processes [3]. The cost of carbon capture can vary widely depending on the CO2 source, with concentrated CO2 streams from natural gas processing and ethanol production having lower capture costs than dilute streams from cement production and power generation [4]. This suggests that, in some countries, it may be possible to obtain CO2 at zero cost. Therefore, it is essential to explore the potential of capturing and utilizing CO2 to mitigate the impact of global warming.
Green plants and algae are capable of using sunlight via the photosynthesis process to capture CO2 from atmosphere and fix it into sugars or carbohydrates, which can then be used as the feedstocks for microbial cells to produce fuels and chemicals. Therefore, biomanufacturing is considered more sustainable than chemical manufacturing with petroleum-based feedstocks. However, using agricultural resources to provide feedstocks for biomanufacturing still poses a sustainability challenge as it hinders food production and threatens biodiversity when natural areas are used for agricultural purposes. To circumvent these limitations, using non-consumable biomass such as lignocellulose and algae could avoid competing with food sources although production of biomass through the photosynthesis process still suffers the challenge in high-cost processing and low energy efficiency (less than 1% of the sunlight energy in chemicals ) [5].
As the feedstock and raw materials significantly contribute to the overall cost of biomanufacturing [6], reducing the cost is critical and can be achieved by using more economical raw materials and designing new microbial cell factories that can efficiently utilize alternative feedstocks. Glucose is the most widely used substrate for biomanufacturing in laboratory and industrial settings for historical and practical reasons as it can be obtained from many economical agricultural crops or wild plants, such as corn starch, sugar canes, and switch grass, or their waste residues. However, employing glucose may repress gene expression and specific biosynthetic pathways for certain biomanufacturing products. In most cases, glucose may also cause several limitations in cell metabolism, resulting in carbon loss as CO2 [7]. This is particularly noticeable when the product of interest requires long synthetic routes from the starting carbon source when it has chemical properties distinct from the substrate or when unfavorable substrates are used, ultimately leading to low product yield [8]. To overcome these limitations, researchers are exploring alternative carbon sources for biomanufacturing, such as agricultural residues and waste plastics [9].
Despite the predominant dependence of current industrial biomanufacturing processes on carbon-intensive carbohydrate substrates including the C5/C6 sugars such as xylose and glucose derived from agricultural biomass, it is noteworthy to acknowledge the presence of microorganisms that exhibit the inherent capability or possess the potential to metabolize C1 and C2 substrates [10]. These C1 substrates, comprising CO2, carbon monoxide (CO), methane (CH4), methanol (CH3OH), and formate (CHOO-) [11], and C2 substrates, comprising mainly ethanol and acetate [12], possess the gains of being inexpensive, naturally abundant, and straightforward manufacturing along with their abundant availability as by-products and industrial wastes [10]. Owing to the worldwide attention to continuous conversion of greenhouse gases, specifically CO2 [13] to recover its diminished economic worth, scientists have a special interest in designing innovative CO2 fixation ways in microbial entities, thereby assisting them in the synthesis of crucial substrate precursors (C1 and C2 chemicals) having the inherent capability to serve as a direct carbon and energy substrate in numerous biomanufacturing processes [14,15]. Particularly, CO2 can be electrochemically converted into the C1/C2 chemicals that can be reused as substrates for microbial fermentation. Further metabolic engineering can enhance the microbial utilization efficiency of these C1~C2 chemicals to the desired products.
However, the utilization of CO2-derived C1/C2 chemicals for biomanufacturing is challenged by the inherent inefficiency of converting C1/C2 into desired bioproducts by native microorganisms, resulting in relatively lower productivity and deprived carbon yield, as compared with the utilization of C5/C6 sugars [15]. Autotrophs, for instance, rely on light as in photoautotrophs or reduced compounds like hydrogen sulfide in chemoautotrophs. Moreover, microbes like acetogenic, carboxydotrophic, and methanotrophic bacteria demonstrate the ability to convert gaseous C1 substrates into multi-carbon compounds, but their potential remains partially unrealized due to inherent growth and conversion efficiency limitations. Carbon fixation pathways' low efficiency and limited energy availability also represent significant bottlenecks. While photosynthesis is a marvel of nature, its energy efficiency seldom surpasses 3%, constraining its industrial applicability. To address the associated challenges, several efforts have been made in the field of synthetic biology and metabolic engineering to evolve both natural microbes (e.g. methanotrophs, a subset of methylotrophs with an ability to utilize methane as a sole carbon and energy source) [16] and/or heterologous microorganisms by engineering and integrating the pathways or enzymes to improve their C1 and C2 substrate-utilizing capabilities [15,17,18,19,20]. Pathway engineering has emerged as a game-changer in this domain. By altering, augmenting, or introducing new metabolic pathways, scientists can manipulate microbial processes to improve carbon fixation efficiency. Such interventions may range from enhancing native pathways to integrating entirely novel ones crafted from a deep understanding of metabolic networks and enzymology [20].
Furthermore, as we delve into microbial fermentation for carbon fixation, we stumble upon its nuanced challenges. One of the pivotal concerns is the significant carbon loss, especially in the format of CO2 during microbial fermentation [21,22], which comprises the advantageous of using biomanufacturing as one of the major efforts in reducing greenhouse gas emission [23]. Therefore, recycling the exhausted CO2 back to the microbial fermentation process is also critical to the success of biomanufacturing.
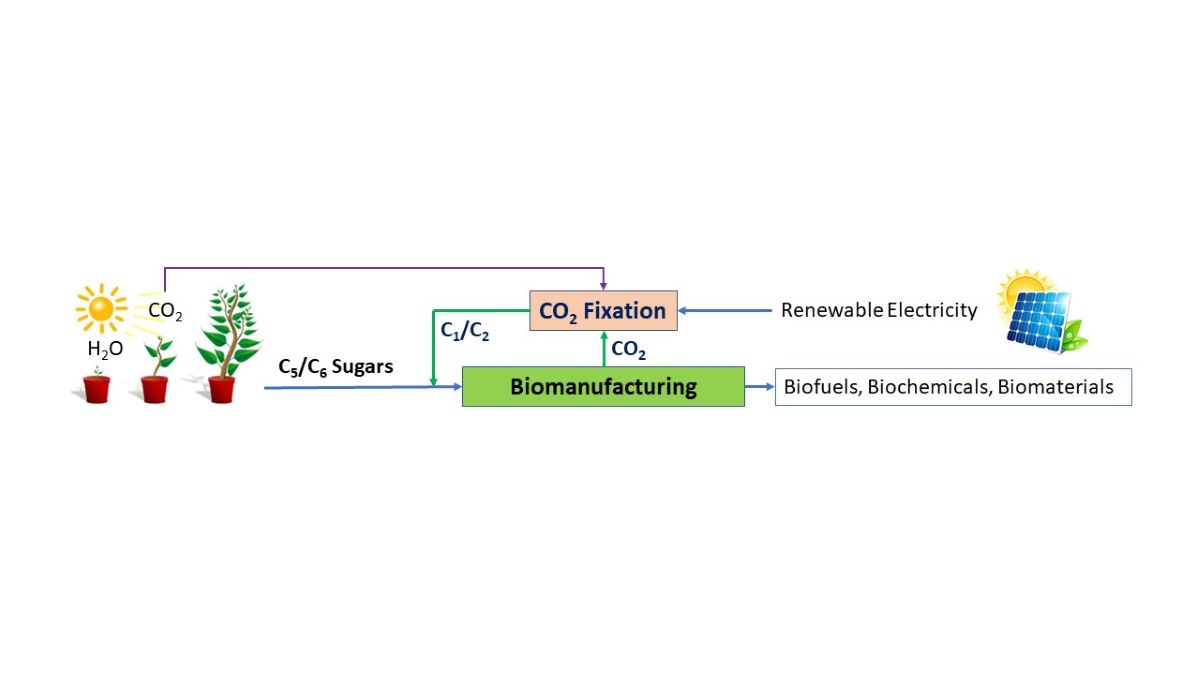
This review delves into the perspectives for using CO2 as a feedstock for biomanufacturing in future. First, the one-step strategy is discussed, which uses novel metabolic pathway design in microbes and engineering approaches to directly fix CO2 and convert it into desired fermentation products. Due to the limitation of the efficiency of one-step CO2 fixation, we further discuss the two-step strategy, which aims to integrate a first electrochemical fixation of CO2 into C1/C2 products such as formate, methanol, acetate, and ethanol and a second fermentation unit co-fed with the original C5/C6 sugars and the CO2-derived C1/C2 chemicals. The great potentials and challenges of using CO2 as a feedstock for future biomanufacturing of various fermentation products are discussed. An overview of the CO2 conversion approaches and using CO2-derived C1/C2 chemicals for biomanufacturing of common products is shown in Figure 1.
2. Current Technologies and State-Of-Art
The conversion of CO2 into value-added chemicals using microbes as biocatalysts is an exciting field of research with the potential to revolutionize biomanufacturing processes [24]. For using CO2 as the feedstock for biomanufacturing, both one-step and two-step strategies can be applied. Table 1 summarizes the general strategies for fixation of CO2 for biomanufacturing. The one-step strategy uses the native or engineered pathways to directly fix CO2 and convert it into desired fermentation products, typically with multiple carbons. Since CO2 has the lowest energy format, producing high-value chemicals with a higher energy format require extra energy, this can be achieved by either plants, algae, or cyanobacteria via photosynthesis process by using light as the energy source or by other microorganisms with cofeeding higher energy-intensive chemicals such as hydrogen gas. The two-step strategy uses a hybrid electrochemical and biochemical conversion approach to fix CO2 and convert it to the desired fermentation products at higher yield and efficiency, where the first step uses an electrochemical catalysis process to covert CO2 into C1/C2 chemicals, followed by a second fermentation step to further convert C1/C2 chemicals into desired products by native or engineered microorganisms.
2.1. One-Step Strategy – Direct Conversion
Internal carbon sequestration has taken many different forms throughout history. Even before the evolution of eukaryotic plants utilizing photosynthesis and light to convert CO2 and energy from light to compose simple sugars, single-celled organisms had already developed mechanisms to capture atmospheric CO2 and transform it into essential compounds for the cell’s development. These primitive mechanisms, especially those in microorganisms like acetogens and methanogens, have shown to be highly efficient, utilizing unique proteins and metabolic pathways for carbon sequestration [1]. Furthermore, microorganisms, especially microalgae and cyanobacteria, exhibit significant advantages over higher plants in their capacity for CO2 fixation as they can yield higher solar energy retention and the potential for year-round growth compared to their more complex plant counterparts [25]. While microalgae are well-recognized for their CO2 fixation capabilities, bacteria present advantages that cannot be overlooked [26]. Microalgae cultivation can be subject to biocontamination over prolonged use from fungal and bacterial species and often run into issues pertaining to even distribution of sun exposure over larger microalgae ponds due to their preferred growth environments, vastly limiting their ability to be utilized on an industrial scale without major alternations to the water infrastructure the microalgae is grown on. Bacteria and some yeasts, on the other hand, have been widely used in biotechnology industry due to inherent compatibility to produce chemicals and their rapid growth rates and life cycles. Further, they are more inclined to accept DNA during genetic modification in the form of plasmids and genomic alternations. This ability allows bacteria and yeast to have DNA introduced into their cells of enzymes to complete metabolic pathways previously incompletely represented in the cells and allow production of specialized products, including bio-alcohols and essential fatty acids. Through this biotechnological approach, CO2 can be directly converted into value-added products, offering an advantage over traditional methods like catalytic conversion, which demand energy-intensive conditions [24].
In this section, we will provide an overview of the one-step strategy for directly using CO2 as the feedstock for biomanufacturing, which includes (1) natural CO2 fixation pathways, (2) synthetic CO2 fixation pathways, (3) host selection and reducing power required for biomanufacturing with CO2, and (4) Using microbial electrosynthesis to utilize CO2 for biomanufacturing.
2.1.1. Natural CO2 fixation pathways
Several pathways facilitate the assimilation of atmospheric CO2 into organic materials, as shown in Figure 2. Among all natural CO2 fixation pathways, the Calvin-Benson-Bassham (CBB) cycle dominates, and is responsible for 90% of global CO2 uptake, primarily driven by the enzyme Ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) [27]. This enzyme catalyzes the transformation of ribulose 1,5-bisphosphate (RuBP) into 3-phosphoglycerate (3-PGA), but its efficiency is occasionally halved due to its tendency to favor O2 during photorespiration [28]. Additionally, pathways such as the Wood-Ljungdahl (WLP), reductive glycine pathway (rGlyP), reductive tricarboxylic acid (rTCA) cycle, 3-hydroxypropionate bi-cycle (HP), 3-hydroxypropionate/4-hydroxybutyrate (HP/HB) cycle, and dicarboxylate/4-hydroxybutyrate (DC/HB) cycle play significant roles in CO2 utilization [29]. These processes, predominantly in autotrophic microorganisms, often lead to vital metabolites like pyruvate or acetyl-CoA, each with unique energy efficiency concerning ATP consumption [30].
Calvin-Benson-Bassham (CBB) Cycle: The CBB cycle stands as the premier identified CO2 biofixation route and remains the primary carbon fixation method in nature. Since it shares numerous metabolites and enzymes with the pentose phosphate pathway (PP pathway), leading to its alternate naming as the reductive pentose phosphate pathway. Found in a variety of organisms such as plants, algae, cyanobacteria, and specific chemoautotrophic microorganisms, this cycle fundamentally operates through the enzymatic action of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO). This enzyme facilitates the electrophilic addition of CO2 to ribulose 1,5-bisphosphate (RuBP), producing two molecules of 3-phosphoglycerate (3-PG). Interestingly, RuBisCO can also introduce oxygen (O2) instead of CO2, forming 3-PG and glyoxylate. This alternative incorporation initiates photorespiration, a series of reactions that release rather than assimilate CO2. As the cycle progresses, 3-PG is subsequently converted to glyceraldehyde 3-phosphate (G3P) with the assistance of enzymes from the gluconeogenic pathway. From there, some G3P molecules fuel central carbon metabolism, while others contribute to the regeneration of RuBP, which is crucial for ongoing CO2 fixation [31]. While RuBisCO's central role in the CBB cycle is undeniable, its efficiency is often questioned. Known for its limited catalytic activity, RuBisCO prefers O2 over CO2, complicating efforts aimed at engineering it for enhanced kinetics largely due to the intricate nature of its substrate-binding pocket [32]. However, efforts to enhance the cycle's efficiency have centered on engineering RuBisCO. For instance, a heterologous cyanobacterial RuBisCO, thanks to its carbon fixation efficiency, was successfully overexpressed in Ralstonia eutropha (Cupriavidus necator), bolstering autotrophic growth and CO2 fixation capabilities [33]. Furthermore, a comprehensive in vitro examination of 143 RuBisCO enzyme activities unveiled a promising type-II RuBisCO variant from Gallionella sp., which is iron oxidizing chemolithotrophic bacteria [34]. Such advancements underscore the potential to amplify CO2 assimilation rates by harnessing superior RuBisCO variants.
Wood-Ljungdahl Pathway (WLP): The WLP, referred to as the reductive acetyl-CoA (rAc-CoA) pathway, is an exemplar of efficient non-photosynthetic carbon fixation. Requiring only one ATP molecule to produce pyruvate is notably more energy-conserving than the CBB cycle, which expends seven ATPs for the same result [5]. The WLP, primarily recognized in acetogens, operates exclusively under anaerobic conditions [35]. This is attributed to the oxygen sensitivity of its key enzymes: carbon monoxide dehydrogenase (CODH). In the rAC-CoA pathway, two CO2 molecules are reduced and converted into acetyl-CoA. A bifunctional enzyme, CO dehydrogenase/acetyl-CoA synthase (CODH/ACS), operates this by catalyzing both the reduction of CO2 to CO and the subsequent condensation of coenzyme A, methyl group, and CO to produce acetyl-CoA. The same rTCA cycle pathways then assimilate this acetyl-CoA further. Microbes utilizing the rAC-CoA pathway often produce acetate or methane as end products [36].
Reductive Glycine Pathway (rGlyP): The initial CO2 assimilation steps in WLP parallel the reductive glycine pathway (rGlyP), wherein formate dehydrogenase reduces CO2 by employing formate dehydrogenase (FDH) to formate, which is subsequently incorporated into the THF cycle to yield 5,10-methylene-THF. This is then reduced further to generate 5-methyl-THF in WLP. Eventually, a combination of this methyl group with CoA and CO produces acetyl-CoA. rGlyP, instead, employs glycine cleavage/synthase system (GCS) to incorporate CO2 and ammonium into 5,10-methylene-THF to produce l-glycine and recycle THF back [30]. Highlighting their potential in microbial CO2 utilization, the WLP and the rGlyP stand out for their ATP efficiency in carbon fixation [37]. The most important advantage of the rGly pathway over WLP is that rGlyP can be operate both in aerobic and anaerobic microorganisms. [38]. Strategies such as overexpressing the essential enzymes can further augment CO2 assimilation efficiency. For instance, enzymes related with THF cycle overexpression in Acetobacterium woodii to create a more productive WLP resulted in a 14% rise in acetate production [39]. Similarly, Eubacterium limosum, when introduced with the GCS, exhibited an improved growth rate and acetate production [40]. Taking it further, even heterologous microbes like E. coli have been successfully engineered with WLP and rGlyP, broadening their carbon conversion capabilities [41,42].
Reductive Tricarboxylic Acid Cycle (rTCA): Initially discovered in the green sulfur bacterium Chlorobium limicola, the rTCA functions as the reverse counterpart to the traditional TCA (or Krebs cycle), primarily in strictly anaerobic or microaerobic autotrophic eubacteria [43]. Key to the rTCA cycle's operation are various enzymes, such as fumarate reductase, 2-oxoglutarate: ferredoxin oxidoreductase (OGOR), ATP-citrate lyase, pyruvate: ferredoxin oxidoreductase (PFOR), and either PEP or pyruvate carboxylase [44]. The rTCA cycle begins with the reductive carboxylation of acetyl-CoA to pyruvate by PFOR, which uses reduced ferredoxin as an electron donor in an oxygen-sensitive process. From pyruvate, several transformations lead to acetyl-CoA and oxaloacetate, completing the cycle. Interestingly, in thermophilic species such as Hydrogenobacter thermophilus, there's a unique conversion of 2-oxoglutarate to isocitrate by enzymes that avoid the accumulation of the unstable intermediate succinyl-CoA [45]. Another variation, the reverse oxidative tricarboxylic acid (roTCA) cycle, is akin to the rTCA cycle but stands out in its enzyme utilization for citrate cleavage, which results in an energy-efficient pathway, despite its thermodynamic challenges [46]. Both cycles highlight the intricate ways organisms assimilate carbon dioxide, ultimately contributing to pyruvate biosynthesis. Although studies on the rTCA cycle's application in metabolic engineering remain limited, emerging research, such as one involving E. coli, has shown promising results in recycling CO2 and optimizing the production of acetate and ethanol [47].
3-Hydroxypropionate (3HP) Bi-Cycle: The 3HP bi-cycle, or Fuchs-Holo bicycle, was first discovered in the thermophilic phototrophic bacterium Chloroflexus aurantiacus [48]. This cycle is considered unique due to its two cyclic CO2 assimilation pathways that collaboratively share initial reactions for CO2 assimilation, forming a complex bicyclic system. Distinctively, acetyl-CoA is first carboxylated to malonyl-CoA and, through a sequence of reactions involving characteristic intermediates like 3-hydroxypropionate and (S)-malyl-CoA, leads to the formation of propionyl-CoA and glyoxylate. The latter subsequently enters a second cycle, culminating in the formation of pyruvate. The 3HP bicycle consumes approximately 2.3 mol ATP to reduce 1 mole of CO2 to pyruvate, similar to the CBB cycle [49].
3-Hydroxypropionate/4-Hydroxybutyrate (HP/HB) Cycle and Dicarboxylate/4-Hydroxybutyrate (DC/HB) Cycle: Beyond the 3HP bi-cycle, other carbon fixation pathways like the 3-hydroxypropionate/4-hydroxybutyrate (HP/HB) cycle and the dicarboxylate/4-hydroxybutyrate (DC/HB) cycle have also been identified. Notably, the HP/HB and DC/HB cycles, prevalent in certain archaea, demonstrate higher energy efficiency in anaerobic environments, with the DC/HB cycle being particularly efficient, requiring only 1.6 mol ATP to reduce one mol CO2 to pyruvate [49]. From an evolutionary perspective, the capability of the 3HP bicycle and the HP/HB cycle to assimilate bicarbonate rather than CO2 is notable. This adaptability likely stems from the higher intracellular concentration of bicarbonate compared to CO2. This feature and oxygen tolerance potentially contribute to their evolutionary survival [45]. From an application standpoint, there have been attempts to harness these pathways for biotechnological purposes. The 3HP bi-cycle’s key enzymes, such as propionyl-CoA synthase and malonyl-CoA reductase, have been leveraged to construct efficient cell factories for 3-hydroxypropionic acid [50]. Similarly, parts of the HP/HB cycle have been expressed in Pyrococcus furiosus, highlighting advancements in CO2 reduction to acetyl-CoA [51]. However, attempts to fully recreate and utilize these pathways in common microbial hosts like E. coli have faced challenges [5].
2.1.2. Synthetic CO2 fixation pathways
Synthetic CO2 fixation pathways have garnered significant attention as potential alternatives to enhance carbon assimilation efficiency, transcending the inherent constraints observed in natural pathways. The focus lies in developing pathways with optimized thermodynamic and kinetic properties while overcoming difficulties associated with key enzymes like RuBisCO [32,34]. One noteworthy example is the crotonyl-CoA/ethylmalonyl-CoA/hydroxybutyryl-CoA (CETCH) cycle. Assembled using 17 enzymes derived from nine distinct organisms, the CETCH cycle has displayed a greater rate of CO2 fixation and a reduced ATP requirement compared to the CBB cycle [27]. Its efficiency is partly attributed to the use of the enoyl-CoA carboxylase/reductase enzyme, which showcases high carboxylation activity. However, translating the in vitro success of the CETCH cycle into in vivo applications remains a challenge [45].
Another synthetic CO2 assimilation route is the Gnd-Entner-Doudoroff (GED) pathway. By inducing specific gene deletions in E. coli, researchers demonstrated the energy-efficient reductive carboxylation of ribulose-5-phosphate via this pathway. Despite its potential, the complete cyclic GED pathway has only been partially shown in vivo [52]. Another advancement was made when researchers synthesized starch from CO2 and hydrogen in a cell-free system. This process coined the artificial starch anabolic pathway (ASAP), comprised 11 core reactions, and showcased an impressive CO2-to-starch conversion rate. This rate was approximately 8.5 times faster than starch synthesis observed in corn [53]. Since pathway length also generates problems for energy efficiency, novel pathways like the POAP cycle and the ICE-CAP pathway have been proposed [54]. The POAP cycle, comprising merely four steps, potentially offers a more streamlined and efficient approach to carbon sequestration. The ICE-CAP pathway, on the other hand, utilizes CO2 alongside high-energy C1 compounds, such as methanol or formaldehyde, obviating the need for ATP and cofactors like NAD(P)H [55].
One computational study, utilizing a repository of around 5,000 known enzymes, unveiled the Malonyl-CoA-Oxaloacetate-Glyoxylate (MOG) pathways. These proposed pathways, which display ATP efficiency over the conventional CBB, might be revolutionary. They use rapid carboxylases and are oxygen-tolerant. However, some enzymes in MOG pathways are thermally sensitive, and their end-product, glyoxylate, when integrated into central metabolism, could revert to CO2, causing this study performed only in in silico [56]. Nevertheless, designing and implementing synthetic pathways isn't without its challenges. When introduced into diverse microbes, these synthetic pathways can disrupt the metabolic balance, necessitating further optimization to realign central metabolic fluxes. Despite this, the capabilities of these synthetic pathways, especially when combined with other technological advancements like biocompatible semiconductor materials or cell-free systems, offer promising avenues for the future of carbon sequestration and utilization [57].
2.1.3. Host selection and reducing power
2.1.3.1. CO2-fixing autotrophs and synthetic hosts
Microorganisms that can synthesize organic substances by fixing inorganic carbon, leveraging energy from either light or inorganic chemicals, are classified as autotrophs. Depending on their energy source, these autotrophs bifurcate into two groups: photoautotrophs, which harness energy via photosynthesis, and chemoautotrophs, which extract energy from chemical reactions [58].
Photoautotrophs, such as cyanobacteria and microalgae, derive energy from photosynthesis. These organisms house photosynthetic pigments, allowing them to harness energy from light and water [59]. Notably, they assimilate CO2 primarily via the Calvin-Benson-Bassham (CBB) cycle. Due to their superior solar energy utilization and rapid growth rates compared to terrestrial plants, they have gained considerable attention as potential bio-production platforms [60]. Cyanobacterial strains like Synechocystis spp. and Synechococcus spp., for instance, have made significant strides in metabolic engineering, that these advancements enable them to produce valuable chemicals [61,62]. Furthermore, certain eukaryotic microalgae have been explored for lipid and alkane production, though their genetic manipulation is somewhat restricted due to limited transformation efficiencies and genetic tool availability [63].
On the other hand, chemoautotrophs, including certain bacteria, obtain energy through chemical reactions. A prominent example is the hydrogen oxidizing bacteria Cupriavidus necator, which can oxidize substances like H2 [64] or formate [65]. This bacterium is known for its ability to naturally accumulate polyhydroxybutyrate (PHB), a precursor for bioplastics, comprising up to 70% of its biomass [66]. Furthermore, genetic engineering has expanded its repertoire to produce chemicals such as branched-chain alcohols and alkanes [67,68]. Another chemoautotroph of interest is Acidithiobacillus ferrooxidans, which can absorb electrons from Fe2+ or directly from a cathode in bioelectrochemical systems [69].
Acetogens represent another subset of chemoautotrophs, which are strictly anaerobic bacteria and use specifically WLP. Certain acetogens, like Clostridium ljungdahlii, Clostridium autoethanogenum, and Acetobacterium woodii, are naturally equipped to produce chemicals such as acetate, ethanol, and 2,3-butanediol [70]. Genetic tools have been applied to acetogens to expand their production portfolio, with some species even being utilized for large-scale industrial applications [71]. Yet, their ATP regeneration capacity poses challenges in producing ATP-intensive products.
In heterotrophic hosts, organisms like E. coli and S. cerevisiae do not initially possess functional CO2 fixation pathways or photosystems. However, scientific endeavors have partially succeeded in transplanting such systems into these hosts, thus ushering in a mixotrophic mode of nutrition [72]. Shifting the spotlight to synthetic autotrophic microorganisms, model organisms like E. coli, Saccharomyces cerevisiae, and Corynebacterium glutamicum have been engineered to metabolize CO2. For instance, E. coli has been engineered to fix CO2 by co-expressing RuBisCO, phosphoribulokinase, and FDH, using formate as a reducing agent [73]. On the other hand, S. cerevisiae, despite the successful expression of RuBisCO from Cupriavidus necator, it has failed to grow on sole CO2 [74]. Recent advances have also demonstrated that autotrophic production platforms can effectively integrate autotrophic and heterotrophic hosts, melding their beneficial traits. A notable instance involves the non-engineered autotrophic acetogen Sporomusa ovata paired with engineered E. coli strains. S. ovata, harnessing semiconductor nanowires, fixes CO2 and excretes acetate – a substrate-engineered E. coli strains that can produce valuable compounds like n-butanol or PHB under aerobic conditions, up to 52% of acetate-to-product yield was reported for PHB production by E. coli [75]. Similarly, another two-reactor system combines the thermophilic acetogen Moorella thermoacetica and yeast Yarrowia lipolytica, where the former's acetate output serves as a feedstock for the latter, engineered for increased lipid synthesis [76]. Such systems still need improvement converting CO2 into valuable end products, achieving sustainable energy conversion efficiencies.
Successfully applying microbial hosts with CO2 fixation capabilities depends on deeply understanding their physiology, biochemistry, and genetics. Both photoautotrophic and chemoautotrophic microbes offer unique opportunities for bio-production, with advances in genetic tools and metabolic engineering paving the way for more efficient autotrophic cell factories. These microbial systems, in combination with advances in metabolic engineering, hold immense potential to revolutionize the sustainable production of value-added compounds.
2.1.3.2. Energy supplies for microbial CO2 fixation
Reducing powers such as NAD(P)H, FADH, ferredoxin red (FdRED), and menaquinol serve as driving forces in microbial CO2 fixation which is pivotal for metabolism. Regeneration of these reducing powers entails the extraction of high-energy electrons from either organic and/or inorganic compounds, or light. Light remains the most prevalent energy source utilized by photoautotrophs like plants, algae, and photosynthetic microorganisms [77]. Photosystems I and II (PS I and PS II) are the primary photo-reaction complexes in photolithotrophic organisms like plants, algae, and cyanobacteria [31]. They absorb light wavelengths ranging from 400 to 700 nm, facilitating the photocatalytic splitting of water to produce ATP and NADPH, thereby providing the requisite energy for CO2 fixation [78]. PS I absorbs light and uses it to excite a low-energy electron from chlorophyll, which then produces FdRED and eventually NADPH. PS II compensates for the electron extracted from PS I by a subsequent electron transfer, originally sourced from a water-splitting reaction [31]. Recently, Chroococcidiopsis thermalis has demonstrated growth in far-red light through specialized photosystems, highlighting the potential for engineering increased efficiency in light utilization [79]. However, there's an inherent energy loss of around 60% in the electron transfer between PS I and II, limiting the efficiency of this system [80]. Efforts to address this inefficiency include the integration of artificial photosensitizers, such as the incorporation of cadmium sulfide nanoparticles with Moorella thermoaceticato facilitate the photosynthesis of acetic acid from CO2 [81].
On the other hand, chemolithotrophs utilize inorganic compounds to extract high-energy electrons for regenerating their reducing powers. The hydrogen-oxidizing bacteria, for instance, employ hydrogenases to consume H2 and regenerate reducing powers. These hydrogenases come in two known varieties: membrane-bound, which uptake hydrogen to produce ATP, and soluble NAD-reducing hydrogenases, which produce NADH [82,83]. For example, E. coli possesses membrane-bound hydrogenases, with Hyd-1 or Hyd-2 catalyzing hydrogen uptake to generate ATP [84]. Ralstonia eutropha, a natural hydrogen-utilizing autotroph, has been studied for its hydrogenase-driven ATP and NADH generation, which, expressed as in the soluble hydrogenase form in E. coli, have shown promise in enhancing intracellular NADH levels [85]. As another example for inorganic compounds to exploit high-energy electrons, iron-oxidizing bacteria oxidize Fe2+ ions to generate NADH [86]. Meanwhile, nitrifying bacteria like ammonia-oxidizing bacteria and nitrite-oxidizing bacteria obtain high-energy electrons by oxidizing nitrogen compounds [87,88]. Notably, sulfur-oxidizing bacteria, derive their electrons from the oxidation of various sulfur compounds through intricate pathways to regenerate reducing powers such as menaquinol, NADH, and FdRED [89]. A smaller group of bacteria focuses on the oxidation of PO33- to PO43-, using phosphite dehydrogenase to transfer electrons and regenerate NADH [90].
In summary, microbial CO2 fixation relies heavily on various pathways to regenerate essential reducing powers, utilizing light and chemicals as energy sources. Whether through photosystems in photoautotrophs or hydrogenases in chemolithotrophs, these microorganisms have developed diverse mechanisms to ensure efficient CO2 fixation, underpinning their importance in the planet's carbon cycle. To regenerate more reducing power, using renewable electricity can also be one of the options for both keeping the carbon neutral environment and regenerate more reducing power, as mentioned in detail in the next subsection.
2.1.4. Microbial electrosynthesis
As shown in Figure 3, microbial electrosynthesis (MES) is an innovative bioelectrochemical approach that leverages electroactive microorganisms to convert renewable electrical energy into value-added products [91,92]. Rooted in bioelectrochemical systems (BES) principles, MES offers a sustainable route to harness CO2 for the synthesis of biofuels and commodity chemicals, some of which include methane, acetate, formic acid, and ethanol, among others, potentially mitigating the detrimental impacts of CO2 emissions [93]. At its core, MES operates by utilizing a biofilm on an electrode as a catalyst, which contrasts with traditional methods that employ chemical catalysts [24].
The MES architecture is intricate [94]. The anodic chamber operates abiotically, where water undergoes splitting to generate protons, electrons, and oxygen. Electrons generated in this chamber are channeled towards the biocathode via an external circuit when an external voltage is applied to the electrochemical cell. Conversely, electrophilic bacteria, primarily acetogens, inhabit the cathodic chamber, which maintains anaerobic, biotic conditions. CO2 acts as an electron acceptor in the MES system, undergoing fixation and conversion at the cathode [95]. Certain electroactive microbes have demonstrated the ability to shuttle electrons intra- and extra-cellularly in this environment [96]. Herein, specialized microbes like Sporomusa species and engineered strains of Clostridium have exhibited the potential to generate biofuels directly from CO2 [97,98]. A classic example demonstrates an acetate production rate of 142.2 mg/L/d and a carbon conversion efficiency of 84% when utilizing enriched mixed homoacetogenic bacteria [99]. Notably, other microbes such as Clostridium scatologenes ATCC 25,775 employ the WLP pathway for CO2 fixation, generating acetic acid, butyric acid, and ethanol by using H2 as reducing power [100].
The true potential of MES lies in its scalability and flexibility. The efficiency and spectrum of products from MES can be influenced by adaptive measures like improved electrode materials, specialized bioreactor designs, and genetically engineered biocatalysts [101]. Indeed, bioreactor optimization, which included strategies like increasing biomass retention and media dilution rate, showcased an acetate production with a titer of 13.5 g/L [102]. Beyond acetate, MES also promises the generation of other valuable bioproducts like butyrate, caproate, and polyhydroxybutyrate (PHB) [103,104,105].
However, MES also faces challenges for more wide applications. Current systems grapple with issues like low CO2 conversion rates, high energy input, and the nuances of maintaining effective microbial communities [106]. Fortunately, recent innovations have exhibited promise to enhance system efficiency. For instance, thermal conditions have been found to influence these processes; Moorella thermoautotrophica exhibited an enhanced rate of acetate and formate production at 55°C as opposed to 25°C [107]. The microbes' biodiversity in MES also plays a pivotal role in its efficiency. Notably, autotrophic sulfate-reducing bacteria (SRM) have displayed potential as excellent biocatalysts, elevating the performance of BES in CO2 fixation [108]. These bacteria hold the potential to improve hydrogen production and water sulfate removal. In a recent study, a co-culture of Desulfopila corrodens and Methanococcus maripaludisco magnified methane production twenty-fold compared to M. maripaludisco alone [109]. Electro-catalyst-assisted MES systems have been developed with electrical-biological hybrid cathodes to improve product rates and variety. Here, Zn-based electrodes have outperformed others; one system achieved an acetic acid production rate of 1.23 g/L [110].
Overall, the CO2 bioelectrorefinery concept, as heralded by MES, is an embodiment of a circular bioeconomy, envisioning an integration of CO2 capture, renewable energy, and sustainable production of chemicals and fuels [111]. While strides have been made, the commercial realization of MES awaits advancements in electrode materials, microbial communities, and process optimization to rival traditional biomass-based processes. Nevertheless, the trajectory of MES research promises a sustainable and innovative path to a cleaner, greener future [112].
2.2. Two-Step Strategy – Fixing CO2 into C1/C2 Chemicals via Electrochemical Catalysis and Converting C1/C2 Chemicals into Bioproducts via Biomanufacturing
The two-step/indirect CO2 fixation and conversion strategy takes the advantages of the current advances from both electrochemical CO2 fixation into C1/C2 chemicals and the synthetic biology to further convert the derived C1/C2 chemicals into the fuels, chemicals, and pharmaceuticals via biomanufacturing process. Even though industrial bioprocesses are predominantly dependent on carbohydrate-based substrates such as the C5/C6 sugars, glucose and xylose, C1 (including carbon monoxide, methane, methanol, formate, and formic acid) and C2 chemicals (mainly ethanol and acetate) can be new candidates for utilization by microbes. A primary advantage of these substrates is their non-competitive nature with alimentary resources, which contributes to an economically sustainable framework while diminishing carbon efflux into the biosphere [113]. Nevertheless, it has been widely studied that the C1/C2 substrates can be produced from CO2 via an electrochemical catalysis process [114,115], which uses renewable electricity from solar, wind, or hydraulic power to capture and fix CO2 into specific C1/C2 products at high yield and selectivity. This two-step CO2 fixation and conversion approach can potentially reduce the dependence on fossil oil-based fuels and chemicals and mitigate the impact of greenhouse gas emissions on the environment.
2.2.1. Using CO2-derived C1 chemicals for biomanufacturing
One-carbon (C1) substrates like CO and CH4 are gaseous C1 substrates from industrial wastes like steel mills and biomass gasification, while liquid C1 substrates, formate, and methanol, are derived from CO2 or waste gas conversions [116]. As the direct CO2 splitting into CO and oxygen is a thermodynamically unfavorable reaction due to the stability of CO2 at ambient temperatures, the response demands a large amount of energy for initiation [117]. Although this reaction was attempted to be feasible by membrane reactor systems by lowering the energy input, the conversion rates are too low to be efficient at an industrial scale. Moreover, conversion efficiencies might cause futile separation of the resultant products, CO and O2, to handle at higher temperatures [118]. Initiatives have been undertaken to capture CO2 and transform it catalytically into a range of high-value products by employing hydrogenation and oxidation processes. However, these chemical conversions of C1 compounds pose significant challenges, including costly catalysts, extreme conditions such as high temperatures (around 450°C) and pressures (approximately 30 MPa), and the emission of hazardous by-products such as carbon monoxide. These factors contribute to the overall expense and unsustainability of the technology [119].
The liquid C1 substrates are advantageous as they're storable and fully soluble, supporting higher production. Microbes can transform C1 substrates into products like alcohols, acids, and plastic components. Specific bacteria can process CO, or CH4 and create multi-carbon compounds [120]. Some also use formate and methanol, which are essential in the C1 pathway [116]. In the following section, natural autotrophs and industrial strains that have been engineered to fix CO2 and recent advances in molecular biology and metabolic engineering for creating more effective CO2 fixation pathways will be discussed. Typical C1 chemical fixation pathways are shown in Figure 4.
2.2.1.1. Carbon monoxide
Carbon monoxide (CO) is a relatively rare gas in the atmosphere, but novel electrochemical CO2 conversion approaches can effectively produce CO from CO2 [121]. Waste gases from industrial processes partially oxidizing carbon-containing compounds or gasifying waste streams can also yield CO [5]. The co-electrolysis of CO2 and H2O can also produce CO. One of the primary concerns of using CO is its high toxicity and difficulty in detection because it is colorless, odorless, and tasteless [122]. Although CO has the potential to impair oxygen transport and mitochondrial function in many organisms, it can be an advantageous carbon and energy source for a phylogenetically diverse array of bacteria and archaea known as carboxydotrophs [123]. Carboxydotrophs have evolved to assimilate CO using carbon monoxide dehydrogenase (CODH), which catalyzes CO oxidation to CO2, providing reducing power to the cell and employing either molybdenum (for aerobes) or nickel (for anaerobes) as essential metal cofactors to facilitate electron transport [124,125].
In aerobic carboxydotrophs, the generated CO2 is typically assimilated via the Calvin–Benson–Bassham (CBB) cycle to produce biomass. Aerobic CO oxidation, which is more exothermic and possesses higher free energy (ΔG0 = -514 kJ) than anaerobic CO oxidation (ΔG0 = -174 kJ), is advantageous for synthesizing ATP-intensive complex products, thereby facilitating higher ATP availability and resulting in increased biomass concentrations [126]. Recent studies have shown the potential of aerobic production of complex molecules, such as the production of C15 sesquiterpene (E)-α-bisabolene from synthesis gas (syngas), a composite of CO, H2, CO2, and trace amounts of impurities such as H2S and NH3—in Hydrogenophaga pseudoflava, although there are challenges due to the potentially explosive mixture of O2 and CO [127].
Anaerobic carboxydotrophs predominantly employ the WLP pathway, also known as the reductive acetyl-CoA pathway, for CO and CO2 assimilation [125]. The WLP bifurcates into two branches: the carbonyl branch, which reduces CO2 to CO, and the methyl branch, which transforms CO2 into formate and its subsequent products. This pathway has garnered significant attention in biotechnological research and genetic and metabolic engineering, particularly in relation to acetogens, microorganisms that use the WLP as their signature pathway [128]. Despite some progress, it remains challenging to demonstrate growth in CO and nonacetogenic hosts. Initial attempts failed to demonstrate CODH/acetyl CoA synthase (ACS) activity in E. coli by expressing genes from Morella thermoacetica [129]. Success was later achieved following the incubation of ACS in NiCl2 solution, although growth using CO as a substrate remained elusive. One major obstacle is the inadequate intracellular conditions and genetic framework of traditional hosts, such as E. coli or yeast, which limits the production and assembly of essential cofactors and sensitive metal centers [121]. As a different strategy, hosts and gene sources with closer phylogenetic relationships have been employed. In recent study, a group of genes from Clostridium ljungdahlii, responsible for encoding CODH/ACS, in conjunction with a methylenetetrahydrofolate reductase gene from E. coli, were integrated into C. acetobutylicum [130]. This reconstruction enabled functional WLP, thereby underscoring the crucial role played by metal clusters. Another study demonstrated increased CO oxidation rates ( 3.1-fold) through overexpression of the endogenous CODH/ACS complex in Eubacterium limosum [131]. In addition, specific adaptive laboratory evolution (ALE) attempts in CODH or ACS have been proven to enhance the activity of the CODH/ACS complex for CO oxidation, showing higher growth and CO gas uptake rates [132]. Nonetheless, despite these advancements, the complete transformation of non-acetogenic microorganisms into carboxydotrophs requires further research.
2.2.1.2. Methane
Methane (CH4) is a potent greenhouse gas, ubiquitous in natural and shale gas reserves. Anthropogenic methane sourced from human activities, including landfills, agricultural practices such as animal livestock emissions, paddy rice cultivation, coal mining, and wastewater treatment, contributes significantly to global warming [133]. According to estimates from the Environmental Defense Fund, at least 25% of present-day global warming is attributable to anthropogenic CH4 emissions. This is a significant concern because CH4, over the initial two decades following its release into the atmosphere, exhibits a warming effect over 80 times greater than CO2 [134]. Consequently, cultivating CH4 for biotechnological applications has dual implications: it not only enhances its value beyond traditional uses, such as generating heat or electricity (termed revalorization), but also plays a pivotal role in curbing greenhouse gas emissions.
CH4 assimilation is initiated by converting methane monooxygenase (MMO) to methanol. Methanotrophs, organisms capable of metabolizing methane exclusively as their carbon source in oxygen-rich and oxygen-deprived environments, have two separate versions of MMO. One is a soluble intracellular variant (sMMO), and the other is a particulate form attached to the membrane (pMMO) [135]. Once methanol is produced, it undergoes further oxidation to form formaldehyde. This compound can then be broken down into CO2, which involves specific enzymes, notably formaldehyde dehydrogenase and formate dehydrogenase [136]. Some intermediate formate or formaldehyde is integrated via serine or ribulose monophosphate (RuMP) cycles, serving as a carbon source in the biomass. Formaldehyde is utilized in the RuMP cycle, transforming it into hexulose-6-phosphate and later into ribulose-5-phosphate to complete the cycle. Through the (tetrahydromethanopterin) H4MPT pathway, formaldehyde undergoes a conversion process to become formate. Meanwhile, the serine cycle incorporates formate through the (tetrahydrofolate) H4F pathway and finally converts serine into glycine to close the cycle [137].
sMMO is recognized for its extensive substrate specificity; however, high copper concentrations may adversely affect its performance. Conversely, pMMO, owing to its proximity to the membrane, has superior accessibility to methane compared to sMMO. The linkage of pMMO with the membrane indicates its ability to accelerate catalysis in CH4 oxidation mechanisms [138]. The phenomenon of anaerobic CH4 oxidation first came to light within microbial consortia. In these communities, the transition of methane to CO2 was paired with the reduction of specific elements, such as sulfate, nitrate, nitrite, iron, or manganese [139,140,141,142]. However, owing to difficulties in obtaining pure cultures, all methanotrophs identified to date are aerobic bacteria [143]. Methanotrophs have been metabolically engineered to yield value-added chemicals from CH4, such as lactate, succinate, and astaxanthin [144]. Despite the slower development and growth rates of methanotrophs, non-native hosts, such as Escherichia coli, offer promising potential for CH4 utilization owing to a deeper understanding of their physiology and established metabolic engineering systems [145].
Utilizing industrially relevant strains, such as E. coli, for methane bioconversion is a promising strategy because of its superior growth rate, in-depth understanding of its physiology, wide range of system/synthetic tools available, and well-established metabolic engineering system for value-added products. However, achieving the full activity expression of methane monooxygenase (MMO) in non-native hosts has proven to be a substantial challenge, thus far, largely unsuccessful [146]. Protein engineering endeavors have used P450 monooxygenase as an alternative to MMO for converting methane to methanol; however, these attempts have garnered very limited success [147,148]. The only progress made includes the expression of the β-subunit of pMMO in E. coli, albeit with merely detectable activity [149]. This underscores that the principal challenge in synthesizing methanotrophs in non-native hosts depends on the functional expression of the enzyme responsible for methane oxidation. Despite these obstacles, recent breakthroughs have led to promising outcomes. For example, the β-subunit of pMMO and the catalytic domains of pMMO from Methylococcus capsulatus have been effectively expressed as soluble enzymes in E. coli. By reassembling these enzymes in a framework built from apoferritin particles, a pMMO-mimetic enzyme particle was generated. This assembly exhibits in vitro methanol production kinetics that rival those of native pMMO [150]. Additionally, heterologous expression of sMMO from M. capsulatus and the GroESL chaperone CH4 was converted to acetone in an E. coli strain previously engineered for methanol-based acetone production [151]. These advances indicate the proof-of-concept and feasibility of synthetic microbes for CH4 bioconversion, suggesting that further strain engineering could significantly enhance the conversion rates and yields, potentially fulfilling the industrial potential of microbial CH4 bioconversion.
2.2.1.3. Methanol
As of 2018, the worldwide methanol (CH3OH) production capacity stood at around 100 million metric tons annually, demonstrating a steady increase in the capacity to convert CH4 into methanol and a concurrent decrease in methanol prices [152]. Today, methanol's cost is already comparable to glucose, an outcome largely influenced by its production predominantly from natural gas, crude oil, and coal via methods such as steam reforming of natural gas, biomass-derived synthesis gas, or through hydrogenation of CO2; thus making its price ($150-300/ton) generally lower than that of sugar ($300-400/ton) [10,11,153]. Methanol, significantly more reduced than most sugars, is an attractive substrate or co-substrate alongside sugars for producing various metabolites, including alcohols, carboxylic acids, fatty acids, and hydrocarbons, given its high reductivity. It boasts a reduction degree of six per carbon, compared to glucose's four, denoting that methanol possesses 50% more electrons per carbon atom, thus housing a high energy content. This abundance of electrons can be harnessed to boost product yields in fermentations, further accentuating methanol's appeal as a substrate [154].
Among all identified native methylotrophy groups, aerobic methylotrophy is the largest, encompassing both prokaryotic and eukaryotic forms, represented by well-studied bacteria such as Bacillus methanolicus and the Methylobacterium extorquens, as well as certain yeast species like Pichia pastoris [155]. These aerobic methylotrophs employ two key methanol utilization pathways. The initial pathway involves the oxidation of methanol to formaldehyde, facilitated by three classes of oxidoreductases, each distinguished by their electron acceptors: PQQ-dependent methanol dehydrogenases (MDHs), NAD+-dependent MDHs, O2-dependent alcohol oxidases (AODs) [156]. The first two are primarily found in methylotrophic bacteria, while the latter is characteristic of methylotrophic yeasts [157]. NAD+-dependent MDHs stand out for their ability to use a universal cofactor, NAD+, to transfer electrons for metabolite production, creating reducing equivalents of NADH. O2-dependent AODs, identified mainly in yeasts, convert methanol into hydrogen peroxide and formaldehyde [158]. The second pathway entails the incorporation of formaldehyde into central carbon metabolism via one of three identified assimilation pathways in aerobic methylotrophs: the xylulose monophosphate (XuMP) cycle (as known as dihydroxyacetone (DHA) cycle), the RuMP cycle, and the serine cycle. The XuMP pathway predominantly occurs in yeasts, while the RuMP and serine pathways are observed in B. methanolicus and M. extorquens, respectively [159,160]. The serine pathway stands out for its carbon efficiency, fixing 3 mol CO2 and merging 3 mol formaldehyde to produce 3 mol acetyl-CoA, although it is also the most ATP-costly. In contrast, the RuMP pathway exhibits the highest energy efficiency, generating 2 mol of NADH and 1 mol of ATP per mole of acetyl-CoA. The XuMP pathway, meanwhile, yields 2 mol of NADH but consumes 1 mol of ATP per mole of acetyl-CoA produced [161].
Anaerobic methylotrophy is mainly limited to methanogenic archaea and acetogenic bacteria, with the latter gaining interest due to their metabolic capacity for high acetate or butyrate production [162]. In methylotrophic acetogens, the methyl-THF produced by the methyltransferase system enters the WLP pathway to generate cell mass and conserve energy [163]. The WLP consists of two separate branches, the methyl, and the carbonyl, each handling one CO2 molecule. In the methyl branch, CO2 converts to formate, which merges with auxiliary tetrahydrofolate and reduces to the methyl group of tetrahydrofolates, a precursor for the methyl group of acetyl-CoA. Conversely, in the carbonyl branch, CO2 transforms to CO, merging with methyl-THF from methanol to produce acetyl-CoA via the CO dehydrogenase/acetyl-CoA synthase (CODH/ACS) [164]. This resultant acetyl-CoA can be used for pyruvate synthesis, biomass production, or acetate generation, enabling ATP production [58]. With higher energy efficiency in converting methanol to biomass or products than aerobic methylotrophs, anaerobic acetogens can also assimilate other C1-compounds such as CO2 and CO due to the presence of the WLP pathway. This methanol assimilation also involves CO2 fixation, making acetogens attractive platform microbes for methanol bioconversion [120].
Native methylotrophs hold the potential for generating high-value chemicals from methanol, but methanol assimilation rates curb the efficiency [165]. Expanding these rates to produce target compounds is an insistent need. While the limited availability of genetic tools poses a challenge, the strides made in synthetic biology now enable the development of these tools to engineer native methylotrophs [166]. For example, B. methanolicus was modified to generate L-lysine by implementing the CRISPRi system [167]. Similarly, M. extorquens has been enabled to produce itaconate by heterologously introducing the cis-aconitic acid decarboxylase gene from Aspergillus terreus [168]. For the aerobic methylotrophs, intermediate metabolite formaldehyde accumulation may cause cellular toxicity due to the macromolecule interactions [169]. Anaerobic acetogens are favorable to avoid formaldehyde toxicity since methanol is directly assimilated through WLP [156]. Similarly, methanol assimilation is conducted within the peroxisome in methylotrophic yeasts, and this might have an advantage over other microbes in keeping the formaldehyde away from other cell components [10]. For example, P. pastoris could produce free fatty acids with superior efficiency from methanol by boosting the availability of precursors and cofactors and minimizing the build-up of formaldehyde through optimized methanol metabolism engineering [170]. Another known methylotrophic yeast, Ogataea polymorpha growth, was also restrained by formaldehyde formation. Engineering pentose phosphate (PP) and gluconeogenesis pathways and further ALE efforts overcome those problems and implemented efficient free fatty acid production with a titer of 15.9 g/L [171]. Nonetheless, genetically modifying and engineering native methylotrophs requires more effort due to constraints like the insufficient understanding of cellular metabolic pathways and a confined set of genetic tools for such engineering [163].
Initiatives have been directed toward creating synthetic methylotrophs to navigate the above-mentioned challenges. For instance, by integrating the heterologous methanol assimilation pathway from B. methanolicus MGA3 into Bacillus subtilis, a methanol-dependent engineered strain that can process 4.09 g/L methanol was produced [172]. In addition to integrating natural methanol-utilization pathways into non-native hosts, unique enzymatic conversions have been employed in synthetic pathway development, boosting the potential for methanol conversion into valuable compounds [173]. Nevertheless, the performance of synthetic methylotrophs falls short of those observed in native methylotrophs. For example, when comparing growth and acetate production from methanol between the most efficient synthetic methylotrophic Escherichia coli and Eubacterium limosum, it was evident that both growth and product yield was markedly lower in E. coli than in E. limosum [174]. As a different strategy, Nguyen et al. employed a comprehensive, genome-scale approach that incorporated mutagenesis, 13C tracer analysis, flux balance examination, and comparative transcriptomic and metabolomic studies to present the metabolism of Methylotuvimicrobium alcaliphilum by implementing robust in silico and in vivo methodologies illuminated the mechanism behind efficient methanol consumption and formaldehyde resilience [175].
Significant advances have been made recently in synthetic methylotrophy in model organisms like E. coli, with the groundwork laid by pinpointing the most likely genes for methanol metabolism from methylotrophs: mdh, hps, and phi. Isotopic incorporation tests with 13C-methanol resulted in a 40% label integration into central carbon metabolites, notably hexose 6-phosphate (H6P), in E. coli expressing these three genes, confirming the functionality of the RuMP pathway established by Hps and Phi [176]. By physically co-localizing crucial enzymes like Mdh, Hps, and Phi into a unified complex, methanol oxidation, and formaldehyde assimilation were enhanced, resulting in a 50-fold rise in methanol to F6P conversion [177]. Once these methanol assimilation pathways were set up, research efforts shifted to tackle the complexities of utilizing methanol as the sole carbon source for E. coli growth and energy. One significant issue is the cofactor imbalance, as methanol oxidation through Mdh is impeded when the cellular NADH to NAD+ ratio rises [178]. A 3.6-fold enhancement in methanol to formaldehyde conversion was achieved by linking this step to an NADH consumption cycle [177]. Alternatively, the concentration of cellular NADH was decreased by removing maldh that encodes NAD+-dependent malate dehydrogenase, which mimicked the strategy used by natural methylotrophs to reduce TCA cycle activity [179]. Another common strain, S. cerevisiae, has been explored by performing ALE on laboratory strain CEN.PK, which has an uncharacterized transcriptional regulator Ygr067cp. It was found that deletion of alcohol oxidation (ADH2) and acetyl-CoA synthetase (ACS1) had severely hindered the growth on methanol [180]. On the other hand, the exact methanol assimilation mechanism in S. cerevisiae is still unknown. Beside conventional host, a nonconventional yeast Yarrowia lipolytica has also been engineered for methanol utilization by introducing RuMP and XuMP pathway genes and ALE efforts [181]
2.2.1.4. Formate
Formate (CHOO-) is a valuable biotechnology substrate because of its high solubility in water and polar solvents, a higher degree of reduction than CO2 and CO, and non-flammability [182]. Despite being less abundant than methanol, rapid advancements in synthesis technology, particularly in electrochemical, photochemical, and catalytic methods, promise to increase its availability. Economic efficiency is also improving, with cost predictions suggesting that formate can compete with glucose as feedstock [183].
Microbial formate assimilation employs two primary strategies naturally. The first oxidizes formate to CO2, extracting and reducing the power that supports carbon fixation and provides the cell with energy [184]. This process is ideally supported by formate due to its low reduction potential [185]. The known carbon-fixation pathways facilitating formatotrophic growth through complete formate oxidation include the ATP-costly CBB cycle (i.e., reductive pentose phosphate pathway) [186] and the highly ATP-efficient, albeit oxygen-sensitive, WLP (i.e., reductive acetyl-CoA pathway) [187]. Despite the latter path is energetically most efficiently utilize formate, product variability and anaerobic growth conditions may limit for use in biotechnological applications [160,188].
The second strategy adopted by microbes to utilize formate as the only carbon source entails the fusion of formate with another intermediary metabolic product, though a portion may still undergo oxidation to furnish the cell with reduction potential and energy [189]. Formate is combined with tetrahydrofolate (THF) to promote such growth, using energy from ATP hydrolysis, resulting in formyl-THF. This compound is then transformed into methylene-THF. Methylene-THF contributes its formaldehyde component to glycine, generating serine, which changes into glycerate. Subsequently, conversions result in the regeneration of acetyl-CoA, which can be either a biomass or valuable product precursor. While the serine pathway's capability to directly incorporate formate, and oxygen insensitivity, it still consumes three ATP to produce one acetyl-CoA from one formate molecule, which causes a kinetic inefficiency [18,184].
In formate bioconversion, it has been suggested that exchanging these inefficient formate assimilation pathways, with ATP-efficient alternatives could improve yield and energy efficiency. The rGly pathway was suggested as a most convenient alternative to the other ATP-infeasible and low-biomass-yielded carbon fixation pathways [190]. One such experiment involved replacing the CBB cycle Cupriavidus necator with the reductive glycine pathway (rGly), which, despite requiring further improvements, could convert formate into valuable chemicals offers a streamlined process that bypasses costly formate separation and prevents harmful formate accumulation [191]. Recently, Sánchez-Andrea et al. [192] showed the sulfate-reducing bacterium Desulfovibrio desulfuricans (strain G11), which can utilize sulfate and hydrogen as energy sources, harness an autotrophic (and formatotrophic) carbon fixation mechanism through the reductive glycine (rGly) pathway and using formate. Its pathway coincides with the WLP route, starting from CO2/CO and producing 5,10-methylene-THF. Then, under the action of the glycine cleavage/synthase system (GCS), a process that includes CO2, NH3, and 5,10-methylene-THF, glycine is synthesized and undergoes further assimilation into pyruvate and biomass [10]. GCS was also introduced to Clostridium pasteurianum to create non-model industrial host by heterologous expression and anaerobic formate utiliziation was successfully demonstrated [193].
As a common industrial strain, E. coli was employed a lot to achieve the most optimal formate utilizer strain. rGly pathway, one of the most promising pathway, was introduced into E. coli together with the serine-threonine cycle to develop a double-direction strategy, and formate was used both as an intermediate (endogenous) and as a carbon source (exogenous) [194]. Then the same group further developed their strategy and applied all homologous and heterologous expressions possible to produce the whole glycine and serine the cell needs from formate and CO2 [195]. Another approach was proposed to improve obstacles in the previous work ([194]), by introducing the THF cycle and reverse glycine cleavage (gcv) pathway together and to obtain a final strain that could utilize both formic acid (FA) and CO2 [42]. As a next step, they engineered E. coli by solving the NADPH generation problem by optimizing cytochrome bo3 and bd-I ubiquinol oxidase levels to acquire full growth on sole FA and CO2 and as high OD600 as 7.38 in 450 h [41].
Developing autotrophic organisms in formate utilization is also an important goal. For instance, Tashiro et al. used an electrochemical-biological system to reduce CO2 into formate in the first place and synthesized L-serine from formate using GCS-introduced E. coli [196]. Gleizer et al. obtained an E. coli strain which has CBB established to utilize formate to cover metabolic activities and uses CO2 as sole carbon source [73]. They employed ALE to convert modified strain from heterotroph to full autotroph in chemostat. In another study, E. coli has been successfully engineered to grow on formate via the rGly pathway. Further ALE boosted the biomass yield of the engineered E. coli strain to 2.3 g CDW/mol formate and halved the doubling time [197]. rGly pathway was also employed in S. cerevisiae to increase tolerance (up to 500 mM) against formate by overexpression of only native enzymes [198].
It is claimed that formate assimilation pathways of natural formatotrophs remain suboptimal for biotechnological applications and present limitations compared to industrially optimized strains, such as S. cerevisiae and E. coli, due to the costlier cultivation requirements, slower growth rates, higher sensitivity to environmental conditions, challenges in genetic manipulation due to less understood metabolic networks, and lacking optimization techniques. [15]. Despite these limitations, certain species, like Acetobacterium woodii, show the potential to bridge this gap. Recently, studies highlighted that A. woodii, when cultivated solely with formate as the carbon and energy source, demonstrated greater efficiency and speed in transforming formate to acetate than when using gaseous substrates [199,200]. This research also undertook a comparative analysis of the energy efficiencies of various acetogens and other microbes, such as formatotrophs or engineered strains capable of utilizing formate or methanol, during the growth and product formation on C1 or sugar substrates. The results revealed that acetogens displayed superior energy efficiency across all substrates tested, specifically C1 substrates, with formate demonstrating even more significant energy potential than gaseous substrates [70]. Among the acetogens, Eubacterium limosum emerged as promising biocatalysts for transforming formate into acetate, primarily producing acetate, during formate-fueled growth [201].
While industrially utilized microbes demonstrate enhanced yield potential and genetic manipulability than above-mentioned nonconventional organisms, cytotoxicity associated with elevated formate concentrations poses a formidable challenge [202]. The tolerance threshold for formate varies widely among organisms and is mainly dictated by formate dehydrogenase activity [116]. For instance, Escherichia coli encounters significant growth disruption at formate concentrations exceeding 100 mM, indicating minimal formate dehydrogenase activity [203]. Conversely, organisms such as Saccharomyces cerevisiae, which exhibit heightened formate dehydrogenase activity, endure and capitalize on elevated formate concentrations, underscoring the differential formate tolerance across diverse organisms [204]. Besides, formate consumption can lead to a slight increase in medium pH, and the resulting alcohols can be toxic to certain microbes at high concentrations, potentially damaging the cell membrane and inhibiting glycolytic enzymes [205]. Therefore, strategies such as metabolic, evolutionary, and rational engineering, proven effective for other inhibitory feedstock compounds or fermentation products, could enhance microbial resilience to formate toxicity [197,206,207].
2.2.2. Using CO2-derived C2 chemicals for biomanufacturing
C2 chemicals, mainly ethanol and acetate, have garnered interest as alternative substrates for biomanufacturing, especially in the production of biofuels, pharmaceuticals, and biopolymers [208]. One of the key challenges in utilizing C2 chemicals as substrates in biomanufacturing is the need to expand and engineer the native metabolic pathways of microorganisms to efficiently convert these substrates into value-added products. This is achieved through various metabolic engineering strategies, such as overexpressing native or heterologous enzymes, redirecting carbon flux, and eliminating competing pathways [5]. The common C2 chemical assimilation pathways are shown in Figure 5.
2.2.2.1. Acetate
Acetate (CH3COO-) typically denotes the disassociated form of acetic acid (CH3COOH), holds significant economic value for biomanufacturing, and the production volume worldwide is expected to be approximately 24.5 million metric tons annually by the year 2025 [12]. Its cost ranges between $350-450 per ton, making it slightly more economical than traditional sugars like glucose, which cost about $500 per ton [209]. The acetate production, with around 75% of it accomplished through chemical catalysis, encompasses methods such as methanol carbonylation, ethylene oxidation, and alkane oxidation [210]. Additional methods for acetate synthesis include the hydrolysis of lignocellulosic biomass, anaerobic digestion, syngas fermentation, and microbial electrosynthesis. One of the green sides of these routes is using waste streams. For instance, lignocellulosic biomass and anaerobic digestion could leverage waste biomass and industrial or agricultural wastes as substrates. Furthermore, processes like syngas fermentation, microbial electrosynthesis and chemical catalysis utilize CO2 as their primary raw material in C1 gas conversion [211]. This highlights the considerable potential of using acetate as potential feedstock regarding environmental friendliness and sustainability, particularly pertinent to achieving carbon neutrality.
The process of utilizing and metabolizing acetate for biochemical production starts with the transportation of acetate from the external environment into the cell, continues with the assimilation of acetate to acetyl-CoA, and at the end, the chemicals formatted from acetyl-CoA. The acidity level within the moderately basic cellular fluid, typically with a pH value between 7.5 and 7.6, significantly exceeds the pKa value of HAc. Thus, acetic acid increases intracellular acidity to some extent by dividing into an acetate anion (Ac-) and a hydrogen ion (H+) [12]. Acetic acid can be toxic to cells, even at concentrations less than 5 g/L [210]. Other than the toxicity and proton imbalance, there are more theories explaining the inhibitory effect of acetate on cell growth. These include (i) alterations in membrane permeability and integrity; (ii) changes in amino acid metabolism, where weak acids hinder the use of specific amino acids and the production of methionine, leading to the buildup of toxic cysteine; and (iii) induced programmed cell death, where high-concentration acetate causes accumulation of reactive oxygen species and impairs energy metabolism in mitochondria, leading to chromatin and nuclear DNA denaturation and subsequent programmed cell death [212].
When microorganisms utilize acetate as the sole carbon source, it is first converted to acetyl-CoA, primarily achieved through two enzymatic routes. The first route involves the formation of an intermediate, acetyl-adenosine monophosphate (acetyl-AMP), which subsequently converts to acetyl-CoA [210]. This pathway requires two moles of ATP due to forming AMP and ADP, marking it as a more energetically expensive route. On the other hand, the acetate kinase-phosphotransacetylase (AckA-Pta) catalyzes acetate to acetyl-phosphate first and then converts it to acetyl-CoA. It represents a reversible mechanism allowing bidirectional conversion between acetate and acetyl-CoA. This pathway consumes only one mole of ATP, making it less energy-demanding than the ACS pathway. Nevertheless, the ACS pathway possesses a high affinity for acetate, around 35 times higher than the AckA-Pta pathway, thus playing a critical role in efficient acetate assimilation, particularly in low-acetate conditions. Besides, despite its role in acetate production and consumption, the AckA-Pta pathway exhibits a lower affinity for acetate, primarily contributing to acetate production overconsumption [12].
An alternative route exists in certain bacteria, such as Pseudomonas sp. and acetic acid bacteria, involving the enzyme succinyl-CoA: acetate CoA-transferase (SCACT). This mechanism eliminates ATP consumption, using succinyl-CoA to convert acetate into acetyl-CoA. Therefore, the SCACT pathway is a significant supplement or alternative to the ACS and AckA-Pta pathways, especially under conditions where these two are non-functional or absent. These acetate assimilation pathways, including aerobic and anaerobic mechanisms, are widespread across several microbial species and constitute the first step of acetate metabolism [210]. Understanding these metabolic routes and their energetic requirements enriches our knowledge of microbial physiology and aids in industrial biotechnology applications by optimizing acetate metabolism in host organisms like E. coli [213].
Acetyl-CoA, derived from acetate, plays a pivotal role as a precursor for extending carbon length, and it is primarily incorporated into two metabolic pathways: the tricarboxylic acid (TCA) cycle or the glyoxylate shunt, leading to an increase from C2 to C4 carbon compounds. Notably, the glyoxylate shunt significantly impacts cell growth when acetate is utilized as the primary carbon source. This pathway divides isocitrate into glyoxylate and succinate via isocitrate lyase (ICL). Following this, glyoxylate is transformed into malate using another acetyl-CoA molecule [214]. Both succinate and malate are crucial to the TCA cycle, being further oxidized to form fumarate and oxaloacetate (OAA), which aid energy generation and higher carbon compound synthesis. Within the TCA cycle, the transition from isocitrate to α-ketoglutarate, facilitated by isocitrate dehydrogenase (IDH), represents an essential step that vies with ICL, thereby affecting carbon flux distribution. Both the glyoxylate shunt and the TCA cycle play a crucial role in acetate absorption due to their role in energy generation and carbon movement [215].
Nevertheless, acetate is less preferable to glucose as a carbon source when generating ATP and NADPH for most acetate-utilizing bacteria. It is worth noting that acetate yields a significantly lower energy content, with 10 ATPs per mol, compared to 38 ATPs per mol for glucose [216]. Consequently, acetate's low energy content might be a limiting factor for its absorption and cell growth. Given that ATP or NADPH is required for most chemical synthesis from acetate, ensuring efficient energy supplementation through metabolic engineering or other techniques is vital for acetate assimilation and biochemical production. To manufacture biochemicals effectively from acetate, certain traits are indispensable in these strains: a high level of acetate tolerance, improved activation of acetate to acetyl-CoA, enhanced acetate assimilation, and efficient chemical production [217].
In recent years, various microbes have been metabolically engineered to create biochemicals, with acetate serving as the carbon source, producing various substances, including acids, alcohols, esters, and other chemicals. In the context of C2-biomanufacturing, the use of acetate as a feedstock has been extensively researched, including native acetogens and genetically modified organisms, can utilize acetate. These include strains such as E. coli [218], Corynebacterium glutamicum [219], Pseudomonas putida [220], S. cerevisiae [221], Cryptococcus curvatus [222], Rhodotorula glutinis [223], Yarrowia lipolytica [224], and Aspergillus oryzae [225], among others. Besides, over 20 value-added chemicals have been produced, with acetate as the main carbon source. Notable examples include poly(3-hydroxybutyrate) (PHB) [226], aromatic amino acids [227], lipids [228], acetate esters [229], and natural products such as isoprenoids that are derived from acetyl-CoA [230]. However, a significant challenge in using acetate is its low concentration (typically 20-30 g/L) when produced from numerous upstream waste utilization processes. Such dilute feedstock solutions further dilute when added into the microbial culture, potentially leading to a low product titer, particularly in batch operations. In response to this challenge, Xu et al. proposed an innovative approach using a continuous bioreactor with a cell recycling unit to produce triacylglycerides (TAGs), which are intracellular products that accumulate in the bioreactor with host cells such as Yarrowia lipolytica [7]. Another known strategy is ALE to increase the acetate tolerance in microorganisms. This leans with the fact that microorganisms produce acetic acid during glucose fermentation and consume this acetic acid when carbon is limited. This consumption may be increased by adding suitable acetic acid salts to balance pH, which makes candidate organisms tolerate and consume acetate more [215].
2.2.2.2. Ethanol
Ethanol (C2H5OH), a simple alcoholic compound has a broad range of applications spanning various industries, including chemical, food, medical, and health. It represents an economically viable raw material. Nonetheless, its primary sources, such as corn, are starch-based, raising concerns due to their competition with food production and considerable CO2 emissions during processing. The compound can be generated from renewable sources such as biomass fermentation, using sugars, starch, or cellulose as raw materials [231]. It has been established in product manufacturing of beverages, flavors, fuels, dyes, disinfectants, antifreeze, and paint [232]. Despite its versatile utilization, the exploration of ethanol as a metabolic engineering feedstock is not yet thoroughly investigated [233].
The conversion of ethanol into productive biochemical pathways typically ensues through two main mechanisms. The initial route involves the enzymatic action of alcohol dehydrogenase and acetaldehyde dehydrogenase, transforming ethanol into acetaldehyde and subsequently into acetyl-CoA. Alternatively, a route more prevalent in eukaryotes, such as S. cerevisiae, initiates the transformation of ethanol to acetate using acetaldehyde as an intermediary, which is then integrated into acetyl-CoA. Microbial species like Clostridium acetobutylicum and E. coli predominantly utilize the former pathway, whereas in S. cerevisiae, the oxidation of ethanol to acetaldehyde is facilitated by alcohol dehydrogenase enzymes, specifically Adh2 or Adh4. This acetaldehyde is then converted to acetate via aldehyde dehydrogenase enzymes, namely Ald4 and Ald5. These processes generate NADH, which is crucial for ATP regeneration, thus providing a higher theoretical yield from ethanol than acetate for reducing product production. However, a significant caveat is that ethanol assimilation can be heat-intensive and oxygen-dependent, potentially amplifying the overall production expenditure [10,234].
In some synthetic hosts like E. coli, metabolic engineering has been deployed to optimize ethanol assimilation. This includes the manipulation of the acetaldehyde dehydrogenase and alcohol dehydrogenase enzymes for efficient ethanol growth [235]. These engineered strains can be further refined to produce valuable products like polyhydroxy butyrate (PHB) or prenol from ethanol [226]. Ethanol can also be utilized as the sole or co-substrate with glucose to produce valuable compounds like the artemisinin precursor in S. cerevisiae [236]. Further strategies have explored the genetic modification of E. coli strains for ethanol utilization. These modifications often introduce ethanol catabolism pathways into E. coli, such as those found in Aspergillus nidulans [237]. By expressing different alcohol dehydrogenases and aldehyde dehydrogenases in E. coli, there's potential for efficient ethanol utilization and production of value-added products from ethanol [235]. One example is introducing a two-step ethanol utilization pathway (EUP) into E. coli has shown promising results in generating polyhydroxy butyrate (PHB), an acetyl-CoA-derived product [233]. The engineered strain demonstrated robust growth on ethanol as the sole carbon source. It produced 1.1 g/L of PHB from 10 g/L of ethanol in 96 h with supplementation of a small amount of amino acids. To expand the range of potential acetyl-CoA-derived compounds from ethanol, this EUP was coupled with a prenol biosynthetic pathway. The resulting strain produced 24 mg/L of prenol from a medium containing ten g/L of ethanol in 48 h. As an exciting new approach, C2-biomanufacturing using ethanol as the sole carbon source has opened the possibility of producing acetyl-CoA-derived Significantly, this strategy has led to a higher theoretical yield for producing acetyl-CoA-derived chemicals from ethanol than other sources. For example, the PHB yield from ethanol was 2-fold higher than that from acetate [226]. Further technological developments and metabolic engineering strategies will likely enhance these processes, making CO2-derived ethanol an abundant, renewable, and affordable substrate to fuel ethanol-based fermentation processes [238].
Taking together, both C1 and C2 chemicals derived from the electrochemical fixation of CO2 can serve as the carbon and energy sources for further biomanufacturing with various microorganisms. The major biochemical reactions to generate ATP from the most common C1/C2 substrates are summarized in Table 2, which may potentially provide guidance for further pathway design and bioconversion yield predictions in future.
2.2.3. Biomanufacturing with syngas via gas fermentation
In addition to the CO2-derived liquid C1/C2 chemicals that can be used as the alternative feedstock for biomanufacturing of fuels and chemicals, synthesis gas, or syngas, which consists of carbon monoxide (CO), hydrogen (H2), carbon dioxide (CO2), nitrogen (N2), and some higher hydrocarbons can also be used as an economical feedstock option. The percentage of CO in syngas can range between 5 to 60%, and the gas can be steam reformed to enrich the H2 content [239]. CO can be obtained from CO2 via electrochemical conversion and H2 can be produced as a product of electrolysis process with water. Syngas can also be produced from biomass gasification, an endothermic process that occurs at temperatures of 750–800 °C and utilizes materials like lignocellulosic biomass and municipal solid waste as feedstocks [240,241]. Despite its promise, the process has some drawbacks. It requires a considerable input of heat energy, and the feedstock must maintain a degree of homogeneity for efficient operation [242]. Heterogeneous feedstocks can lead to wide variations in product composition, necessitating pre-treatment and post-treatment steps that can escalate operational costs [243]. Conversely, the thermochemical process involves gasifying carbonaceous materials into syngas and converting it into biofuels [244]. Syngas can be converted to diesel, methanol, or ethanol using the Fischer-Tropsch (FT) process, which uses chemical catalysts. Still, this method requires high temperatures and pressures, making it less feasible [243]. Another option is using microbial catalysts to convert syngas into a variety of products, like alcohols and carboxylic acids, at milder conditions [245,246].
However, each of these platforms presents unique advantages and disadvantages. Biochemical conversion, for example, struggles with high production costs and energy demands. On the other hand, the thermochemical conversion process, while capable of utilizing all biomass components (including lignin), faces challenges like gas-liquid mass transfer limitation, low productivity, and elevated production costs [247]. Combining the two conversion processes, such as electrochemical conversion CO2 into CO, biomass gasification, and syngas fermentation, could be a solution to these problems. Syngas fermentation, compared to Fischer-Tropsch Synthesis (FTS), is seen as a superior option due to its operational flexibility, end product variety, and cost-effectiveness. This technology could serve as a sustainable way of supplying feedstock for fermentation. Integrating gasification with syngas fermentation could bring together the benefits of thermochemical (full conversion of lignocellulosic biomass) and biochemical (flexibility in CO/H2 ratio of the substrate and end products) technologies, mitigating the complexity of pre-treatment steps and the high enzyme and operational costs of biomass valorization [248]. This approach has potential to be directly implemented in industries that release high levels of exhaust gases, like steel manufacturing, oil refining, and petrochemistry.
However, syngas fermentation processes still have challenges to overcome, such as bacterial biomass washout, low gas solubility, and limited mass transfer rates at the gas-liquid interface. These challenges demand further research and innovation to boost microbial activity or limit the exposure of microorganisms to excessive shear stress, ultimately reducing operational costs [249]. Microbial conversion of CO, H2, and CO2 to acids and alcohols via acetogenic bacteria operates via the reductive acetyl-CoA or WLP, as mentioned in earlier sections. These biological methods offer several advantages such as high tolerance to trace contaminants, high product specificity, and being sustainable, environmentally friendly, and cost-effective [250]. Despite these obstacles, gas fermentation offers a promising route for sustainable fuel production and waste recycling. It provides feedstock flexibility, non-food biomass utilization, and total carbon utilization, including lignin from woody biomass, offering significant advantages over sugar fermentation. Moreover, if the process limitations can be overcome, gas fermentation could provide a more selective, robust, flexible, and cost-effective option than the thermocatalytic Fischer-Tropsch synthesis, suggesting it's a promising technology for mitigating global warming and fulfilling increased liquid fuel demand, especially in transportation [251].
3. Challenges and Future Perspectives
3.1. Challenges for Biomanufacturing with Direct Fixation of CO2
The conversion of inorganic carbon (CO2) into organic compounds offers a promising strategy to mitigate the greenhouse effect and furnish sustainable resources. This method has potential implications for addressing climate change and utilizing CO2 as an economical substrate for producing fuels, chemicals, food ingredients, pharmaceuticals, and industrial materials. The rapid advances in chemical, electrochemical, and biotechnological research methods and tools indicate the imminent identification of novel carbon-fixing enzymes and pathways, which makes it feasible for directly fixing and converting CO2 into desired fuels or chemical products. However, despite these discoveries, the current natural or engineered carbon fixation systems are plagued by inefficiencies and a lack of adaptability for genetic modifications, making them inadequate for industrial applications. There are several major challenges to be addressed before the one-step or direct fixation of CO2 strategy can be applied in large scale applications for high-yield production of fuels and chemicals from CO2:
- 1)
- Only low energy utilization efficiency can be achieved when light is used as the energy source to fix CO2. Green plants, algae, and certain bacteria are capable of using sunlight via the photosynthesis process to capture and fix CO2 into carbohydrates, but at low energy efficiency, with less than 1% of the sunlight energy stored in the biosynthesized chemicals [5].
- 2)
- Energy-intensive chemicals such as H2 gas can be used to fix CO2 and providing the reducing power to convert CO2 into desired carbohydrate products, but there are concerns of extra material cost, technical challenges of using gas for fermentation, increased process complexity, and operating safety due to the use of H2 gas or similar energy-intensive materials.
- 3)
- A very limited number of microbial hosts, genetic manipulation methods and tools, and pathway engineering strategies are available for more generalized applications of direct CO2 fixation and conversion. Many synthetic pathways for direct CO2 fixation face major challenges, such as enzymes with toxicity to host cells or with non-compatible optimum temperatures. Innovations such as the allyl-CoA carboxylase/reductase, which boasts an activity rate 37 times that of the CBB cycle, show promise in addressing this [27]. Introducing mechanisms to concentrate carbon also seems to be a viable strategy to enhance the carbon flux in these pathways. With synthetic biology's progress, exploring and designing novel pathways might be the key. Predictions even suggest that certain pathways, like those using phosphoenolpyruvate carboxylase, could potentially offer 2 to 3 times the carbon fixation rate of the Calvin cycle [56].
- 4)
- Microbial electrosynthesis (MES) can be used to produce certain fuels or valuable organic acids [91,92,93] by utilizing a biofilm on an electrode as a catalyst to directly reduce CO2 to the products [24], but the species of the microorganisms and the categories of the fuels and chemicals that can be produced are very limited. Acetate is the current major product and its production titer and yield are still too low, which significantly increases the downstream recovery cost [252]. In addition, there is strict requirement for the materials that can be used for cathode. More challenges for further process design and scale-up are expected for large-scale applications in future [252].
3.2. Challenges for Biomanufacturing with CO2-Derived C1/C2 Chemicals
Due to the overall low energy efficiency and/or product yield from the biomanufacturing process with one-step/direct CO2 fixation, the two-step CO2 fixation and conversion strategy is considered more promising for future biomanufacturing of various fuels and chemicals, which uses C1/C2 substrates derived from CO2 via electrochemical catalysis. However, there are also several major challenges need to be addressed:
- 1)
- Mass transfer challenges limits the microbial fermentation productivity when the CO2-derived C1 gases, such as CO or CH4, are used as the substrate. Metabolic engineering strategies for using appropriate microorganisms to metabolize the C1 gases are also to be established and further optimized. In addition, safety concerns are also another challenge that may limit the use of CO for biomanufacturing.
- 2)
- Though formic acid and acetic acid can be used as the substrate for biomanufacturing, most current electrochemical catalysis processes can only fix CO2 into the form of formate or acetate salts in aqueous solution, which need to be further treated with acid and base and go through a complicated purification process to obtain the acid products so that they can be fed into the bioreactor for microbial fermentation. There has been progress in electrochemically fix CO2 into nearly pure formic acid [253], but the productivity needs to be further improved for large scale application. Comparing to the electrochemical reduction of CO2 into formic acid, converting CO2 into acetic acid at high yield is still a challenge [254].
- 3)
- Direct feeding too much formic acid or acetic acid into a bioreactor may cause sudden acidic pH spikes in fermentation and kill the microbial cells. Therefore, new formic/acetic acid feeding strategies should be developed to avoid/minimize pH spikes in a bioreactor while providing enough substrate(s) for cell growth and product formation [255,256].
- 4)
- Methanol and ethanol can be used as the fermentation substrate with high energy densities, but high concentrations of the alcohol substrates may cause toxicity to the microbial cells. In addition, further metabolic engineering strategies for efficient assimilation of methanol and/or ethanol should be explored for significantly higher product yield.
3.3. Future Perspectives for Biomanufacturing with CO2
The overuse of fossil oil-based or -derived fuels, chemicals, and materials has led to increased carbon emissions, which becomes one of the major contributors to global climate change. Biomanufacturing with renewable or waste feedstocks is considered as a promising and sustainable route to replace the current petrochemical methods for producing all fuels, chemicals, and materials that are needed in our daily life. Feedstock or raw materials, typically obtained from land-based biomass in the format of starch, sugars, and fats, contribute to a significant portion of the biomanufacturing product cost. Using CO2 or CO2-derived chemicals as biomanufacturing feedstock not only reduces the material cost, but also contributes to the global effort in reducing carbon emissions and achieving the carbon-neutral or -negative goal. While significant progresses have been achieved to demonstrate the feasibility of using one-step or two-step strategies for biomanufacturing with CO2, major challenges and technical barriers still exist, as described earlier. Figure 6 shows a brief summary of using various methods that have been developed or will be developed for using CO2 as feedstock for biomanufacturing. The following research efforts and perspectives will be expected in future:
- 1)
- Using advanced synthetic biology to creating new microbial cell factories to utilize CO2 and CO2-derived chemicals for high-yield biomanufacturing: Researchers are now at the forefront of devising more efficient synthetic systems. This involves engineering pivotal enzymes and transferring whole or partial carbon fixation pathways into heterotrophic cells, enabling them to perform carbon fixation. A testament to these efforts includes the creation of pathways like the MCG pathway and the CETCH cycle using different carboxylases [27]. Although the enhancement in carbon fixation rate remains modest, these innovations may lead to designing more adept systems. Host selection also serves a challenge to keep CO2 fixation sustainable. For example, most CO2-fixing microbes cannot tolerate high CO2 concentrations, necessitating research into strains that can endure and efficiently process higher levels of CO2 or CO2-derived substrates. Adaptive laboratory evolution (ALE) methods may be applied to help develop more robust production strains that are suitable for large scale applications.
- 2)
- Using artificial intelligence (AI) to guide the discoveries of new strains, metabolic pathways, enzymes, and fermentation process controls that may lead to complete bioconversion of CO2 or CO2-derived substrates [257,258,259]: This may also help discover new valuable products that may be produced from the pathways using CO2 or having CO2 as the major intermediates. More advanced process, such as continuous biomanufacturing with extremely high yield and productivity, can also be developed [6].
- 3)
- Exploring a cofeeding strategy that uses a mixed C1 and C2 substrates for biomanufacturing: Current electrochemical reduction of CO2 focuses on maximizing the production of a single C1/C2 product at high yield and selectivity. However, the microbial cells may be capable of using a mixed C1 and C2 feed for producing a desired fermentation product. This may help relieve the burden in the electrochemical catalysis system and significantly reduce its cost. More strain engineering and fermentation process development work should be conducted to use a medium or feed with mixed C1/C2 substrates, including methanol, formic acid, ethanol, and acetic acid, for various biomanufacturing purposes. A joint research effort between the electrochemists, biologist, and chemical engineers are expected to achieve the goal.
- 4)
- Developing an advanced process control strategy based on online monitoring/measurements of dissolved CO2 in aqueous medium, exhausted CO2 in off-gas flow, and the cellular redox levels. Technologies for measuring dissolved CO2 in liquid and gas-phase CO2 have been well established and become commercially available. Monitoring redox cofactor (NAD/NADH, NADP/NADPH, FAD/FADH2) balance has also been investigated and demonstrated capability for advanced fermentation control to further improve the biomanufacturing yield [260,261,262]. In particular, nutrient-induced metabolic shift for high productivity and low-waste generation has been demonstrated in cultures of various cell lines and products. However, as the cells rapidly respond to culture conditions, it is crucial to closely monitor their metabolism for a controlled balance between the target metabolic pathway and unfavorable consequences. In particular, during biosynthesis of bioproducts from CO2-derived C1/C2 substrates, additional reduction power (NADH, NADPH, FADH2) has to be supplied to produce compounds whose degree of reduction is higher than that of the substrate [263,264]. Therefore, adjusting the metabolic status and pathways for improved NADH/NADPH in microbial cells is an effective method to enhance the biosynthesis of many bioproducts [263,265,266].
- 5)
- Developing a novel biomanufacturing platform that can produce fuels and chemicals from sugars at zero or near zero life cycle carbon emissions via in-situ CO2 recycling: Most microbial fermentation processes that use C5/C6 sugars as substrates have nearly 50% or more carbon loss due to the need for metabolizing a portion of the sugar substrate into CO2 to generate energy (ATP) and cofactors for cell growth and biosynthesis. To date, there has been very rare research aiming for biomanufacturing with direct recycling of the exhausted CO2. There is an urgent need for developing a transformative technology that can capture the exhausted CO2 from biomanufacturing and recycle it as high-energy chemicals such as the liquid C1/C2 chemicals that can be reused as feedstocks for microbial fermentation. The capturing and fixation of CO2 into C1/C2 chemicals can be achieved via similar electrochemical catalysis processes [115,267]. The developed new biomanufacturing platform should employ newly engineered strains that can co-utilize C5/C6 sugars and CO2-derived C1/C2 chemicals for producing the desired fermentation products. As shown in Figure 7. Recycling the exhausted CO2 back to fermentation not only avoid/minimize the CO2 release from the biomanufacturing processes, but also maximize the use of the renewable feedstocks for significantly higher product yield.
4. Conclusions
This review summarized the most recent advancements and strategies in CO2 fixation and conversion into industrially valuable chemicals. The path to efficient CO2 fixation is fraught with challenges, ranging from biological to technical. Nonetheless, the rapid advancements in synthetic biology and multi-disciplinary collaborations offer a promising future for the field. Addressing these challenges will provide avenues for sustainable resource generation and significantly contribute to climate change mitigation. Continued research and innovation are vital to bring these promising laboratory-level techniques to commercial reality and industrially available candidates in addressing GHG emissions.
Author Contributions
Conceptualization, E.K. and D.X.; writing—original draft preparation, E.K., J.Q., A.W., and D.X.; writing—review and editing E.K., J.Q., Y.Z., and D.X.; supervision, D.X.; project administration, D.X.; funding acquisition, D.X. and Y.Z. All authors have read and agreed to the published version of the manuscript.
Acknowledgement
The authors would like to thank the financial support from the US. National Science Foundation (Award No. 2133660) and NIH/NCCIH (Award No. 2R44AT010840).
Conflicts of Interest
No conflicts of interest are declared by the authors.
References
- Lemaire, O.N.; Jespersen, M.; Wagner, T. CO2-Fixation Strategies in Energy Extremophiles: What Can We Learn From Acetogens? Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef]
- Hossain, M.F. Extreme Level of CO2 Accumulation into the Atmosphere Due to the Unequal Global Carbon Emission and Sequestration. Water, Air, & Soil Pollution 2022, 233, 105. [Google Scholar] [CrossRef]
- Bhat, P. Carbon needs to cost at least $100/tonne now to reach net zero by 2050: Reuters poll. 2021.
- Baylin-Stern, A.; Berghout, N. Is carbon capture too expensive? IEA, Paris 2021.
- Claassens, N.J.; Sousa, D.Z.; Dos Santos, V.A.; de Vos, W.M.; van der Oost, J. Harnessing the power of microbial autotrophy. Nat Rev Microbiol 2016, 14, 692–706. [Google Scholar] [CrossRef]
- Xie, D. Continuous biomanufacturing with microbes — upstream progresses and challenges. Current Opinion in Biotechnology 2022, 78, 102793. [Google Scholar] [CrossRef]
- Park, J.O.; Liu, N.; Holinski, K.M.; Emerson, D.F.; Qiao, K.; Woolston, B.M.; Xu, J.; Lazar, Z.; Islam, M.A.; Vidoudez, C. Synergistic substrate cofeeding stimulates reductive metabolism. Nature Metabolism 2019, 1, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.C.; Mi, L.; Pontrelli, S.; Luo, S. Fuelling the future: microbial engineering for the production of sustainable biofuels. Nature Reviews Microbiology 2016, 14, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.-H.; Mou, J.-H.; Chao, C.Y.H.; Chopra, S.S.; Daoud, W.; Leu, S.-y.; Ning, Z.; Tso, C.Y.; Chan, C.K.; Tang, S.; et al. Biotechnology of Plastic Waste Degradation, Recycling, and Valorization: Current Advances and Future Perspectives. ChemSusChem 2021, 14, 4103–4114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ottenheim, C.; Weingarten, M.; Ji, L. Microbial Utilization of Next-Generation Feedstocks for the Biomanufacturing of Value-Added Chemicals and Food Ingredients. Front Bioeng Biotechnol 2022, 10, 874612. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Schilling, M.; Holtmann, D.; Sell, D.; Filho, M.V.; Marx, A.; Vorholt, J.A. Methanol-based industrial biotechnology: current status and future perspectives of methylotrophic bacteria. Trends in Biotechnology 2009, 27, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, D.; Merkel, M.; Lilge, L.; Henkel, M.; Hausmann, R. From acetate to bio-based products: underexploited potential for industrial biotechnology. Trends in Biotechnology 2021, 39, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Peter, S.C. Reduction of CO2 to Chemicals and Fuels: A Solution to Global Warming and Energy Crisis. ACS Energy Letters 2018, 3, 1557–1561. [Google Scholar] [CrossRef]
- Gomez Vidales, A.; Bruant, G.; Omanovic, S.; Tartakovsky, B. Carbon dioxide conversion to C1 - C2 compounds in a microbial electrosynthesis cell with in situ electrodeposition of nickel and iron. Electrochimica Acta 2021, 383, 138349. [Google Scholar] [CrossRef]
- Bae, J.; Jin, S.; Kang, S.; Cho, B.-K.; Oh, M.-K. Recent progress in the engineering of C1-utilizing microbes. Current Opinion in Biotechnology 2022, 78, 102836. [Google Scholar] [CrossRef]
- Iguchi, H.; Yurimoto, H.; Sakai, Y. Interactions of Methylotrophs with Plants and Other Heterotrophic Bacteria. Microorganisms 2015, 3, 137–151. [Google Scholar] [CrossRef]
- Chou, A.; Lee, S.H.; Zhu, F.; Clomburg, J.M.; Gonzalez, R. An orthogonal metabolic framework for one-carbon utilization. Nature Metabolism 2021, 3, 1385–1399. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liao, J.C. A modified serine cycle in Escherichia coli coverts methanol and CO2 to two-carbon compounds. Nature Communications 2018, 9, 3992. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.Y.H.; Jung, H.-W.; Tsuei, C.-Y.; Liao, J.C. Converting Escherichia coli to a Synthetic Methylotroph Growing Solely on Methanol. Cell 2020, 182, 933–946. [Google Scholar] [CrossRef]
- Antonovsky, N.; Gleizer, S.; Noor, E.; Zohar, Y.; Herz, E.; Barenholz, U.; Zelcbuch, L.; Amram, S.; Wides, A.; Tepper, N.; et al. Sugar Synthesis from CO2 in Escherichia coli. Cell 2016, 166, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.-F.; Zhang, G.-C.; Walker, B.; Seo, S.-O.; Kwak, S.; Liu, J.-J.; Kim, H.; Ort, D.R.; Wang, S.-G.; Jin, Y.-S. Recycling Carbon Dioxide during Xylose Fermentation by Engineered Saccharomyces cerevisiae. ACS Synthetic Biology 2017, 6, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.-l.; Chen, K.-q.; Li, J.; Fang, X.-j.; Zheng, X.-y.; Sui, S.-s.; Jiang, M.; Wei, P. Optimization of culture conditions in CO2 fixation for succinic acid production using Actinobacillus succinogenes. Journal of Industrial Microbiology and Biotechnology 2011, 38, 1605–1605. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Nurhayati; Cheng, C.-L.; Nagarajan, D.; Chang, J.-S.; Hu, J.; Lee, D.-J. Carbon capture and utilization of fermentation CO2: Integrated ethanol fermentation and succinic acid production as an efficient platform. Applied Energy 2017, 206, 364–371. [CrossRef]
- Salehizadeh, H.; Yan, N.; Farnood, R. Recent advances in microbial CO2 fixation and conversion to value-added products. Chemical Engineering Journal 2020, 390, 124584. [Google Scholar] [CrossRef]
- Morales, M.; Sánchez, L.; Revah, S. The impact of environmental factors on carbon dioxide fixation by microalgae. FEMS Microbiology Letters 2018, 365, fnx262. [Google Scholar] [CrossRef]
- Ray, S.; Abraham, J.; Jordan, N.; Lindsay, M.; Chauhan, N. Synthetic, Photosynthetic, and Chemical Strategies to Enhance Carbon Dioxide Fixation. C 2022, 8. [Google Scholar] [CrossRef]
- Schwander, T.; Schada von Borzyskowski, L.; Burgener, S.; Cortina, N.S.; Erb, T.J. A synthetic pathway for the fixation of carbon dioxide in vitro. Science 2016, 354, 900–904. [Google Scholar] [CrossRef]
- Erb, T.J.; Zarzycki, J. A short history of RubisCO: the rise and fall (?) of Nature's predominant CO2 fixing enzyme. Current Opinion in Biotechnology 2018, 49, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Cai, Z.; Li, Y. Synthetic biology for CO2 fixation. Science China Life Sciences 2016, 59, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.R.; Ahn, Y.-J.; Lee, S.Y. Bacterial conversion of CO2 to organic compounds. Journal of CO2 Utilization 2022, 58, 101929. [Google Scholar] [CrossRef]
- Johnson, M.P. Photosynthesis. Essays Biochem 2016, 60, 255–273. [Google Scholar] [CrossRef]
- Flamholz, A.I.; Prywes, N.; Moran, U.; Davidi, D.; Bar-On, Y.M.; Oltrogge, L.M.; Alves, R.; Savage, D.; Milo, R. Revisiting Trade-offs between Rubisco Kinetic Parameters. Biochemistry 2019, 58, 3365–3376. [Google Scholar] [CrossRef]
- Li, Z.; Xin, X.; Xiong, B.; Zhao, D.; Zhang, X.; Bi, C. Engineering the Calvin–Benson–Bassham cycle and hydrogen utilization pathway of Ralstonia eutropha for improved autotrophic growth and polyhydroxybutyrate production. Microbial cell factories 2020, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Davidi, D.; Shamshoum, M.; Guo, Z.; Bar-On, Y.M.; Prywes, N.; Oz, A.; Jablonska, J.; Flamholz, A.; Wernick, D.G.; Antonovsky, N.; et al. Highly active rubiscos discovered by systematic interrogation of natural sequence diversity. The EMBO Journal 2020, 39, e104081. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F. Hydrogen, metals, bifurcating electrons, and proton gradients: The early evolution of biological energy conservation. FEBS Letters 2012, 586, 485–493. [Google Scholar] [CrossRef]
- Pu, X.; Han, Y. Promotion of Carbon Dioxide Biofixation through Metabolic and Enzyme Engineering. Catalysts 2022, 12. [Google Scholar] [CrossRef]
- Claassens, N.J.; Satanowski, A.; Bysani, V.R.; Dronsella, B.; Orsi, E.; Rainaldi, V.; Yilmaz, S.; Wenk, S.; Lindner, S.N. Engineering the reductive glycine pathway: a promising synthetic metabolism approach for c1-assimilation. In One-Carbon Feedstocks for Sustainable Bioproduction; Springer: 2022; pp. 299-350.
- Gonzales, J.N.; Matson, M.M.; Atsumi, S. Nonphotosynthetic Biological CO2 Reduction. Biochemistry 2019, 58, 1470–1477. [Google Scholar] [CrossRef]
- Straub, M.; Demler, M.; Weuster-Botz, D.; Dürre, P. Selective enhancement of autotrophic acetate production with genetically modified Acetobacterium woodii. Journal of Biotechnology 2014, 178, 67–72. [Google Scholar] [CrossRef]
- Song, Y.; Lee, J.S.; Shin, J.; Lee, G.M.; Jin, S.; Kang, S.; Lee, J.-K.; Kim, D.R.; Lee, E.Y.; Kim, S.C. Functional cooperation of the glycine synthase-reductase and Wood–Ljungdahl pathways for autotrophic growth of Clostridium drakei. Proceedings of the National Academy of Sciences 2020, 117, 7516–7523. [Google Scholar] [CrossRef]
- Bang, J.; Hwang, C.H.; Ahn, J.H.; Lee, J.A.; Lee, S.Y. Escherichia coli is engineered to grow on CO2 and formic acid. Nature Microbiology 2020, 5, 1459–1463. [Google Scholar] [CrossRef]
- Bang, J.; Lee, S.Y. Assimilation of formic acid and CO2 by engineered Escherichia coli equipped with reconstructed one-carbon assimilation pathways. Proceedings of the National Academy of Sciences 2018, 115, E9271–E9279. [Google Scholar] [CrossRef]
- Berg Ivan, A. Ecological Aspects of the Distribution of Different Autotrophic CO2 Fixation Pathways. Applied and Environmental Microbiology 2011, 77, 1925–1936. [Google Scholar] [CrossRef]
- Chen, P.Y.; Li, B.; Drennan, C.L.; Elliott, S.J. A reverse TCA cycle 2-oxoacid:ferredoxin oxidoreductase that makes C-C bonds from CO(2). Joule 2019, 3, 595–611. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, K.; Chen, Y.; Tan, T.; Nielsen, J. Third-generation biorefineries as the means to produce fuels and chemicals from CO2. Nature Catalysis 2020, 3, 274–288. [Google Scholar] [CrossRef]
- Steffens, L.; Pettinato, E.; Steiner, T.M.; Eisenreich, W.; Berg, I.A. Tracking the Reversed Oxidative Tricarboxylic Acid Cycle in Bacteria. Bio Protoc 2022, 12, e4364. [Google Scholar] [CrossRef]
- Chen, C.-H.; Tseng, I.-T.; Lo, S.-C.; Yu, Z.-R.; Pang, J.-J.; Chen, Y.-H.; Huang, C.-C.; Li, S.-Y. Manipulating ATP Supply Improve in situ CO2 recycling by reductive TCA cycle in engineered Escherichia coli. 3 Biotech 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, G. Alternative pathways of carbon dioxide fixation: insights into the early evolution of life? Annu Rev Microbiol 2011, 65, 631–658. [Google Scholar] [CrossRef]
- Erb Tobias, J. Carboxylases in Natural and Synthetic Microbial Pathways. Applied and Environmental Microbiology 2011, 77, 8466–8477. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, M.; Chen, Y.; Wang, Z.; Nielsen, J.; Liu, Z. Engineering yeast mitochondrial metabolism for 3-hydroxypropionate production. Biotechnology for Biofuels and Bioproducts 2023, 16, 64. [Google Scholar] [CrossRef]
- Keller, M.W.; Schut, G.J.; Lipscomb, G.L.; Menon, A.L.; Iwuchukwu, I.J.; Leuko, T.T.; Thorgersen, M.P.; Nixon, W.J.; Hawkins, A.S.; Kelly, R.M. Exploiting microbial hyperthermophilicity to produce an industrial chemical, using hydrogen and carbon dioxide. Proceedings of the National Academy of Sciences 2013, 110, 5840–5845. [Google Scholar] [CrossRef]
- Satanowski, A.; Dronsella, B.; Noor, E.; Vögeli, B.; He, H.; Wichmann, P.; Erb, T.J.; Lindner, S.N.; Bar-Even, A. Awakening a latent carbon fixation cycle in Escherichia coli. Nature Communications 2020, 11, 5812. [Google Scholar] [CrossRef]
- Cai, T.; Sun, H.; Qiao, J.; Zhu, L.; Zhang, F.; Zhang, J.; Tang, Z.; Wei, X.; Yang, J.; Yuan, Q. Cell-free chemoenzymatic starch synthesis from carbon dioxide. Science 2021, 373, 1523–1527. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.; Gong, F.; Zhu, H.; Zhang, Y.; Cai, Z.; Li, Y. A minimized synthetic carbon fixation cycle. ACS Catalysis 2021, 12, 799–808. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, H.; Xu, Y.; Meng, H.; Zeng, A.-P. Turn air-captured CO2 with methanol into amino acid and pyruvate in an ATP/NAD (P) H-free chemoenzymatic system. Nature Communications 2023, 14, 2772. [Google Scholar] [CrossRef]
- Bar-Even, A.; Noor, E.; Lewis, N.E.; Milo, R. Design and analysis of synthetic carbon fixation pathways. Proceedings of the National Academy of Sciences 2010, 107, 8889–8894. [Google Scholar] [CrossRef]
- Liu, Z.; Shi, S.; Ji, Y.; Wang, K.; Tan, T.; Nielsen, J. Opportunities of CO2-based biorefineries for production of fuels and chemicals. Green Carbon 2023, 1, 75–84. [Google Scholar] [CrossRef]
- Humphreys, C.M.; Minton, N.P. Advances in metabolic engineering in the microbial production of fuels and chemicals from C1 gas. Current Opinion in Biotechnology 2018, 50, 174–181. [Google Scholar] [CrossRef]
- Kato, Y.; Inabe, K.; Hidese, R.; Kondo, A.; Hasunuma, T. Metabolomics-based engineering for biofuel and bio-based chemical production in microalgae and cyanobacteria: A review. Bioresource Technology 2022, 344, 126196. [Google Scholar] [CrossRef] [PubMed]
- Farrokh, P.; Sheikhpour, M.; Kasaeian, A.; Asadi, H.; Bavandi, R. Cyanobacteria as an eco-friendly resource for biofuel production: a critical review. Biotechnology progress 2019, 35, e2835. [Google Scholar] [CrossRef] [PubMed]
- Ramey, C.J.; Barón-Sola, A.n.; Aucoin, H.R.; Boyle, N.R. Genome engineering in cyanobacteria: where we are and where we need to go. ACS synthetic biology 2015, 4, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microbial cell factories 2018, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Gimpel, J.A.; Henríquez, V.; Mayfield, S.P. In metabolic engineering of eukaryotic microalgae: potential and challenges come with great diversity. Frontiers in microbiology 2015, 6, 1376. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Colón, B.C.; Ziesack, M.; Silver, P.A.; Nocera, D.G. Water splitting–biosynthetic system with CO2 reduction efficiencies exceeding photosynthesis. Science 2016, 352, 1210–1213. [Google Scholar] [CrossRef]
- Li, H.; Opgenorth, P.H.; Wernick, D.G.; Rogers, S.; Wu, T.-Y.; Higashide, W.; Malati, P.; Huo, Y.-X.; Cho, K.M.; Liao, J.C. Integrated electromicrobial conversion of CO2 to higher alcohols. Science 2012, 335, 1596–1596. [Google Scholar] [CrossRef]
- Nybo, S.E.; Khan, N.E.; Woolston, B.M.; Curtis, W.R. Metabolic engineering in chemolithoautotrophic hosts for the production of fuels and chemicals. Metabolic Engineering 2015, 30, 105–120. [Google Scholar] [CrossRef]
- Lu, J.; Brigham, C.J.; Gai, C.S.; Sinskey, A.J. Studies on the production of branched-chain alcohols in engineered Ralstonia eutropha. Applied microbiology and biotechnology 2012, 96, 283–297. [Google Scholar] [CrossRef]
- Bi, C.; Su, P.; Müller, J.; Yeh, Y.-C.; Chhabra, S.R.; Beller, H.R.; Singer, S.W.; Hillson, N.J. Development of a broad-host synthetic biology toolbox for Ralstonia eutropha and its application to engineering hydrocarbon biofuel production. Microbial cell factories 2013, 12, 1–10. [Google Scholar] [CrossRef]
- Kernan, T.; Majumdar, S.; Li, X.; Guan, J.; West, A.C.; Banta, S. Engineering the iron-oxidizing chemolithoautotroph Acidithiobacillus ferrooxidans for biochemical production. Biotechnology and bioengineering 2016, 113, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Bae, J.; Jin, S.; Kang, S.; Cho, B.-K. Engineering Acetogenic Bacteria for Efficient One-Carbon Utilization. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef]
- Liew, F.E.; Nogle, R.; Abdalla, T.; Rasor, B.J.; Canter, C.; Jensen, R.O.; Wang, L.; Strutz, J.; Chirania, P.; De Tissera, S. Carbon-negative production of acetone and isopropanol by gas fermentation at industrial pilot scale. Nature biotechnology 2022, 40, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Oh, M.-K. Strategies for Biosynthesis of C1 Gas-derived Polyhydroxyalkanoates: A review. Bioresource technology 2022, 344, 126307. [Google Scholar] [CrossRef]
- Gleizer, S.; Ben-Nissan, R.; Bar-On, Y.M.; Antonovsky, N.; Noor, E.; Zohar, Y.; Jona, G.; Krieger, E.; Shamshoum, M.; Bar-Even, A. Conversion of Escherichia coli to generate all biomass carbon from CO2. Cell 2019, 179, 1255–1263. e1212. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Kim, S.-J.; Kim, S.-K.; Seo, S.-O.; Park, S.; Shin, J.; Kim, J.-S.; Park, B.-R.; Jin, Y.-S.; Chang, P.-S.; et al. Yeast metabolic engineering for carbon dioxide fixation and its application. Bioresource Technology 2021, 346. [Google Scholar] [CrossRef]
- Liu, C.; Gallagher, J.J.; Sakimoto, K.K.; Nichols, E.M.; Chang, C.J.; Chang, M.C.; Yang, P. Nanowire–bacteria hybrids for unassisted solar carbon dioxide fixation to value-added chemicals. Nano letters 2015, 15, 3634–3639. [Google Scholar] [CrossRef]
- Hu, P.; Chakraborty, S.; Kumar, A.; Woolston, B.; Liu, H.; Emerson, D.; Stephanopoulos, G. Integrated bioprocess for conversion of gaseous substrates to liquids. Proceedings of the National Academy of Sciences 2016, 113, 3773–3778. [Google Scholar] [CrossRef]
- Meng, X.; Liu, L.; Chen, X. Bacterial photosynthesis: state-of-the-art in light-driven carbon fixation in engineered bacteria. Current Opinion in Microbiology 2022, 69, 102174. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, F.; Meng, H.; Zhang, Y.; Li, Y. Introducing extra NADPH consumption ability significantly increases the photosynthetic efficiency and biomass production of cyanobacteria. Metabolic engineering 2016, 38, 217–227. [Google Scholar] [CrossRef]
- Nürnberg, D.J.; Morton, J.; Santabarbara, S.; Telfer, A.; Joliot, P.; Antonaru, L.A.; Ruban, A.V.; Cardona, T.; Krausz, E.; Boussac, A. Photochemistry beyond the red limit in chlorophyll f–containing photosystems. Science 2018, 360, 1210–1213. [Google Scholar] [CrossRef] [PubMed]
- Work, V.H.; D’Adamo, S.; Radakovits, R.; Jinkerson, R.E.; Posewitz, M.C. Improving photosynthesis and metabolic networks for the competitive production of phototroph-derived biofuels. Current Opinion in Biotechnology 2012, 23, 290–297. [Google Scholar] [CrossRef]
- Sakimoto, K.K.; Wong, A.B.; Yang, P. Self-photosensitization of nonphotosynthetic bacteria for solar-to-chemical production. Science 2016, 351, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Xu, Y.; Wang, X.; Christie, P. Function of biohydrogen metabolism and related microbial communities in environmental bioremediation. Frontiers in microbiology 2019, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, W.; Ogata, H.; Rudiger, O.; Reijerse, E. Hydrogenases. Chemical reviews 2014, 114, 4081–4148. [Google Scholar] [CrossRef] [PubMed]
- Sargent, F. The model [NiFe]-hydrogenases of Escherichia coli. Advances in microbial physiology 2016, 68, 433–507. [Google Scholar]
- Lamont, C.M.; Sargent, F. Design and characterisation of synthetic operons for biohydrogen technology. Archives of Microbiology 2017, 199, 495–503. [Google Scholar] [CrossRef]
- White, G.F.; Edwards, M.J.; Gomez-Perez, L.; Richardson, D.J.; Butt, J.N.; Clarke, T.A. Mechanisms of bacterial extracellular electron exchange. Advances in microbial physiology 2016, 68, 87–138. [Google Scholar]
- Van Kessel, M.A.; Speth, D.R.; Albertsen, M.; Nielsen, P.H.; Op den Camp, H.J.; Kartal, B.; Jetten, M.S.; Lücker, S. Complete nitrification by a single microorganism. Nature 2015, 528, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nature Reviews Microbiology 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lin, J.-Q.; Liu, X.-M.; Pang, X.; Zhang, C.-J.; Yang, C.-L.; Gao, X.-Y.; Lin, C.-M.; Li, Y.-Q.; Li, Y. Sulfur oxidation in the acidophilic autotrophic Acidithiobacillus spp. Frontiers in microbiology 2019, 9, 3290. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, I.; Coates, J. Microbial phosphite oxidation and its potential role in the global phosphorus and carbon cycles. Advances in Applied Microbiology 2017, 98, 93–117. [Google Scholar] [PubMed]
- Butler, C.S.; Lovley, D.R. How to sustainably feed a microbe: strategies for biological production of carbon-based commodities with renewable electricity. Frontiers in Microbiology 2016, 7, 1879. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Simoska, O.; Lim, K.; Grattieri, M.; Yuan, M.; Dong, F.; Lee, Y.S.; Beaver, K.; Weliwatte, S.; Gaffney, E.M. Fundamentals, applications, and future directions of bioelectrocatalysis. Chemical Reviews 2020, 120, 12903–12993. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, S.; Srikanth, S.; Mohanakrishna, G.; Zacharia, R.; Strik, D.P.; Pant, D. Biotransformation of carbon dioxide in bioelectrochemical systems: State of the art and future prospects. Journal of Power Sources 2017, 356, 256–273. [Google Scholar] [CrossRef]
- Bhagchandanii, D.D.; Babu, R.P.; Sonawane, J.M.; Khanna, N.; Pandit, S.; Jadhav, D.A.; Khilari, S.; Prasad, R. A comprehensive understanding of electro-fermentation. Fermentation 2020, 6, 92. [Google Scholar] [CrossRef]
- Zhen, G.; Lu, X.; Kumar, G.; Bakonyi, P.; Xu, K.; Zhao, Y. Microbial electrolysis cell platform for simultaneous waste biorefinery and clean electrofuels generation: current situation, challenges and future perspectives. Progress in Energy and Combustion Science 2017, 63, 119–145. [Google Scholar] [CrossRef]
- Das, S.; Ghangrekar, M. Value added product recovery and carbon dioxide sequestration from biogas using microbial electrosynthesis. 2018. [Google Scholar]
- Gong, Y.; Ebrahim, A.; Feist, A.M.; Embree, M.; Zhang, T.; Lovley, D.; Zengler, K. Sulfide-driven microbial electrosynthesis. Environmental science & technology 2013, 47, 568–573. [Google Scholar]
- Leang, C.; Ueki, T.; Nevin, K.P.; Lovley, D.R. A genetic system for Clostridium ljungdahlii: a chassis for autotrophic production of biocommodities and a model homoacetogen. Applied and environmental microbiology 2013, 79, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Mohanakrishna, G.; Vanbroekhoven, K.; Pant, D. Impact of dissolved carbon dioxide concentration on the process parameters during its conversion to acetate through microbial electrosynthesis. Reaction Chemistry & Engineering 2018, 3, 371–378. [Google Scholar]
- Liu, H.; Song, T.; Fei, K.; Wang, H.; Xie, J. Microbial electrosynthesis of organic chemicals from CO 2 by Clostridium scatologenes ATCC 25775 T. Bioresources and Bioprocessing 2018, 5, 1–10. [Google Scholar] [CrossRef]
- Bajracharya, S.; Yuliasni, R.; Vanbroekhoven, K.; Buisman, C.J.; Strik, D.P.; Pant, D. Long-term operation of microbial electrosynthesis cell reducing CO2 to multi-carbon chemicals with a mixed culture avoiding methanogenesis. Bioelectrochemistry 2017, 113, 26–34. [Google Scholar] [CrossRef]
- Gildemyn, S.; Verbeeck, K.; Slabbinck, R.; Andersen, S.J.; Prévoteau, A.; Rabaey, K. Integrated Production, Extraction, and Concentration of Acetic Acid from CO2 through Microbial Electrosynthesis. Environmental Science & Technology Letters 2015, 2, 325–328. [Google Scholar] [CrossRef]
- Sciarria, T.P.; Batlle-Vilanova, P.; Colombo, B.; Scaglia, B.; Balaguer, M.D.; Colprim, J.; Puig, S.; Adani, F. Bio-electrorecycling of carbon dioxide into bioplastics. Green Chemistry 2018, 20, 4058–4066. [Google Scholar] [CrossRef]
- Jourdin, L.; Raes, S.M.; Buisman, C.J.; Strik, D.P. Critical biofilm growth throughout unmodified carbon felts allows continuous bioelectrochemical chain elongation from CO2 up to caproate at high current density. Frontiers in Energy Research 2018, 6, 7. [Google Scholar] [CrossRef]
- Batlle-Vilanova, P.; Ganigue, R.; Ramió-Pujol, S.; Baneras, L.; Jiménez, G.; Hidalgo, M.; Balaguer, M.D.; Colprim, J.; Puig, S. Microbial electrosynthesis of butyrate from carbon dioxide: Production and extraction. Bioelectrochemistry 2017, 117, 57–64. [Google Scholar] [CrossRef]
- Chu, N.; Hao, W.; Wu, Q.; Liang, Q.; Jiang, Y.; Liang, P.; Ren, Z.J.; Zeng, R.J. Microbial Electrosynthesis for Producing Medium Chain Fatty Acids. Engineering 2022, 16, 141–153. [Google Scholar] [CrossRef]
- Yu, L.; Yuan, Y.; Tang, J.; Zhou, S. Thermophilic Moorella thermoautotrophica-immobilized cathode enhanced microbial electrosynthesis of acetate and formate from CO2. Bioelectrochemistry 2017, 117, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Agostino, V.; Rosenbaum, M.A. Sulfate-reducing electroautotrophs and their applications in bioelectrochemical systems. Frontiers in Energy Research 2018, 6, 55. [Google Scholar] [CrossRef]
- Deutzmann, J.S.; Spormann, A.M. Enhanced microbial electrosynthesis by using defined co-cultures. The ISME journal 2017, 11, 704–714. [Google Scholar] [CrossRef]
- Jiang, Y.; Chu, N.; Zhang, W.; Ma, J.; Zhang, F.; Liang, P.; Zeng, R.J. Zinc: A promising material for electrocatalyst-assisted microbial electrosynthesis of carboxylic acids from carbon dioxide. Water research 2019, 159, 87–94. [Google Scholar] [CrossRef]
- Lee, S.Y.; Oh, Y.-K.; Lee, S.; Fitriana, H.N.; Moon, M.; Kim, M.-S.; Lee, J.; Min, K.; Park, G.W.; Lee, J.-P.; et al. Recent developments and key barriers to microbial CO2 electrobiorefinery. Bioresource Technology 2021, 320, 124350. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Singh, R.; Bose, A. Microbial electron uptake in microbial electrosynthesis: a mini-review. Journal of Industrial Microbiology and Biotechnology 2019, 46, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Santala, S.; Stephanopoulos, G. Mixed carbon substrates: a necessary nuisance or a missed opportunity. Current Opinion in Biotechnology 2020, 62, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Nitopi, S.; Bertheussen, E.; Scott, S.B.; Liu, X.; Engstfeld, A.K.; Horch, S.; Seger, B.; Stephens, I.E.L.; Chan, K.; Hahn, C.; et al. Progress and Perspectives of Electrochemical CO2 Reduction on Copper in Aqueous Electrolyte. Chemical Reviews 2019, 119, 7610–7672. [Google Scholar] [CrossRef] [PubMed]
- Costentin, C.; Robert, M.; Savéant, J.-M. Catalysis of the electrochemical reduction of carbon dioxide. Chemical Society Reviews 2013, 42, 2423–2436. [Google Scholar] [CrossRef] [PubMed]
- Cotton, C.A.R.; Claassens, N.J.; Benito-Vaquerizo, S.; Bar-Even, A. Renewable methanol and formate as microbial feedstocks. Current Opinion in Biotechnology 2020, 62, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.; Lougou, B.G.; Shuai, Y.; Wang, Z.; Tan, H. Current technology development for CO2 utilization into solar fuels and chemicals: A review. Journal of Energy Chemistry 2020, 49, 96–123. [Google Scholar] [CrossRef]
- Rayne, S. Thermal Carbon Dioxide Splitting: A Summary of the Peer-Reviewed Scientific Literature. Nature Precedings 2008. [Google Scholar] [CrossRef]
- Kamkeng, A.D.N.; Wang, M.; Hu, J.; Du, W.; Qian, F. Transformation technologies for CO2 utilisation: Current status, challenges and future prospects. Chemical Engineering Journal 2021, 409, 128138. [Google Scholar] [CrossRef]
- Bae, J.; Song, Y.; Lee, H.; Shin, J.; Jin, S.; Kang, S.; Cho, B.-K. Valorization of C1 gases to value-added chemicals using acetogenic biocatalysts. Chemical Engineering Journal 2022, 428, 131325. [Google Scholar] [CrossRef]
- Liew, F.; Martin, M.E.; Tappel, R.C.; Heijstra, B.D.; Mihalcea, C.; Köpke, M. Gas Fermentation—A Flexible Platform for Commercial Scale Production of Low-Carbon-Fuels and Chemicals from Waste and Renewable Feedstocks. Frontiers in Microbiology 2016, 7. [Google Scholar] [CrossRef]
- Herranz, J.; Pătru, A.; Fabbri, E.; Schmidt, T.J. Co-electrolysis of CO2 and H2O: From electrode reactions to cell-level development. Current Opinion in Electrochemistry 2020, 23, 89–95. [Google Scholar] [CrossRef]
- King, G.M.; Weber, C.F. Distribution, diversity and ecology of aerobic CO-oxidizing bacteria. Nature Reviews Microbiology 2007, 5, 107–118. [Google Scholar] [CrossRef]
- Can, M.; Armstrong, F.A.; Ragsdale, S.W. Structure, Function, and Mechanism of the Nickel Metalloenzymes, CO Dehydrogenase, and Acetyl-CoA Synthase. Chemical Reviews 2014, 114, 4149–4174. [Google Scholar] [CrossRef] [PubMed]
- Oelgeschlager, E.; Rother, M. Carbon monoxide-dependent energy metabolism in anaerobic bacteria and archaea. Arch Microbiol 2008, 190, 257–269. [Google Scholar] [CrossRef]
- Takors, R.; Kopf, M.; Mampel, J.; Bluemke, W.; Blombach, B.; Eikmanns, B.; Bengelsdorf, F.R.; Weuster-Botz, D.; Dürre, P. Using gas mixtures of CO, CO2 and H2 as microbial substrates: the do's and don'ts of successful technology transfer from laboratory to production scale. Microbial Biotechnology 2018, 11, 606–625. [Google Scholar] [CrossRef] [PubMed]
- Grenz, S.; Baumann, P.T.; Rückert, C.; Nebel, B.A.; Siebert, D.; Schwentner, A.; Eikmanns, B.J.; Hauer, B.; Kalinowski, J.; Takors, R.; et al. Exploiting Hydrogenophaga pseudoflava for aerobic syngas-based production of chemicals. Metabolic Engineering 2019, 55, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Diender, M.; Stams, A.J.; Sousa, D.Z. Pathways and Bioenergetics of Anaerobic Carbon Monoxide Fermentation. Front Microbiol 2015, 6, 1275. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.L.; James-Hagstrom, J.E.; Garvin, D.K.; Gorst, C.M.; Runquist, J.A.; Baur, J.R.; Haase, F.C.; Ragsdale, S.W. Cloning and expression of the gene cluster encoding key proteins involved in acetyl-CoA synthesis in Clostridium thermoaceticum: CO dehydrogenase, the corrinoid/Fe-S protein, and methyltransferase. Proc Natl Acad Sci U S A 1989, 86, 32–36. [Google Scholar] [CrossRef]
- Fast Alan, G.; Papoutsakis Eleftherios, T. Functional Expression of the Clostridium ljungdahlii Acetyl-Coenzyme A Synthase in Clostridium acetobutylicum as Demonstrated by a Novel In Vivo CO Exchange Activity En Route to Heterologous Installation of a Functional Wood-Ljungdahl Pathway. Applied and Environmental Microbiology 2018, 84, e02307–02317. [Google Scholar] [CrossRef]
- Kang, H.; Park, B.; Oh, S.; Pathiraja, D.; Kim, J.-Y.; Jung, S.; Jeong, J.; Cha, M.; Park, Z.-Y.; Choi, I.-G.; et al. Metabolism perturbation Causedby the overexpression of carbon monoxide dehydrogenase/Acetyl-CoA synthase gene complex accelerated gas to acetate conversion rate ofEubacterium limosumKIST612. Bioresource Technology 2021, 341, 125879. [Google Scholar] [CrossRef]
- Jin, S.; Kang, S.; Bae, J.; Lee, H.; Cho, B.-K. Development of CO gas conversion system using high CO tolerance biocatalyst. Chemical Engineering Journal 2022, 449, 137678. [Google Scholar] [CrossRef]
- Karakurt, I.; Aydin, G.; Aydiner, K. Sources and mitigation of methane emissions by sectors: A critical review. Renewable Energy 2012, 39, 40–48. [Google Scholar] [CrossRef]
- (EDF), E.D.F. Reducing methane will help hit the brakes on runaway global warming. EDF Voices blog 6/2/2023.
- Semrau, J.D.; DiSpirito, A.A.; Yoon, S. Methanotrophs and copper. FEMS Microbiology Reviews 2010, 34, 496–531. [Google Scholar] [CrossRef]
- Sahoo, K.K.; Goswami, G.; Das, D. Biotransformation of Methane and Carbon Dioxide Into High-Value Products by Methanotrophs: Current State of Art and Future Prospects. Front Microbiol 2021, 12, 636486. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-y.; Kim, C.-g. Application and development of methanotrophs in environmental engineering. Journal of Material Cycles and Waste Management 2019, 21, 415–422. [Google Scholar] [CrossRef]
- Ge, X.; Yang, L.; Sheets, J.P.; Yu, Z.; Li, Y. Biological conversion of methane to liquid fuels: Status and opportunities. Biotechnology Advances 2014, 32, 1460–1475. [Google Scholar] [CrossRef] [PubMed]
- Boetius, A.; Ravenschlag, K.; Schubert, C.J.; Rickert, D.; Widdel, F.; Gieseke, A.; Amann, R.; Jørgensen, B.B.; Witte, U.; Pfannkuche, O. A marine microbial consortium apparently mediating anaerobic oxidation of methane. Nature 2000, 407, 623–626. [Google Scholar] [CrossRef]
- Beal, E.J.; House, C.H.; Orphan, V.J. Manganese-and iron-dependent marine methane oxidation. Science 2009, 325, 184–187. [Google Scholar] [CrossRef]
- Haroon, M.F.; Hu, S.; Shi, Y.; Imelfort, M.; Keller, J.; Hugenholtz, P.; Yuan, Z.; Tyson, G.W. Anaerobic oxidation of methane coupled to nitrate reduction in a novel archaeal lineage. Nature 2013, 500, 567–570. [Google Scholar] [CrossRef]
- Ettwig, K.F.; Butler, M.K.; Le Paslier, D.; Pelletier, E.; Mangenot, S.; Kuypers, M.M.; Schreiber, F.; Dutilh, B.E.; Zedelius, J.; de Beer, D. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature 2010, 464, 543–548. [Google Scholar] [CrossRef]
- Lee, O.K.; Hur, D.H.; Nguyen, D.T.N.; Lee, E.Y. Metabolic engineering of methanotrophs and its application to production of chemicals and biofuels from methane. Biofuels, Bioproducts and Biorefining 2016, 10, 848–863. [Google Scholar] [CrossRef]
- Hwang, I.Y.; Nguyen, A.D.; Nguyen, T.T.; Nguyen, L.T.; Lee, O.K.; Lee, E.Y. Biological conversion of methane to chemicals and fuels: technical challenges and issues. Applied Microbiology and Biotechnology 2018, 102, 3071–3080. [Google Scholar] [CrossRef]
- Dürre, P.; Eikmanns, B.J. C1-carbon sources for chemical and fuel production by microbial gas fermentation. Current Opinion in Biotechnology 2015, 35, 63–72. [Google Scholar] [CrossRef]
- Nguyen, A.D.; Lee, E.Y. Engineered Methanotrophy: A Sustainable Solution for Methane-Based Industrial Biomanufacturing. Trends in Biotechnology 2021, 39, 381–396. [Google Scholar] [CrossRef]
- Zilly, F.E.; Acevedo, J.P.; Augustyniak, W.; Deege, A.; Häusig, U.W.; Reetz, M.T. Tuning a P450 Enzyme for Methane Oxidation. Angewandte Chemie International Edition 2011, 50, 2720–2724. [Google Scholar] [CrossRef] [PubMed]
- Meinhold, P.; Peters, M.W.; Chen, M.M.Y.; Takahashi, K.; Arnold, F.H. Direct Conversion of Ethane to Ethanol by Engineered Cytochrome P450 BM3. ChemBioChem 2005, 6, 1765–1768. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, R.; Smith, S.M.; Rawat, S.; Yatsunyk, L.A.; Stemmler, T.L.; Rosenzweig, A.C. Oxidation of methane by a biological dicopper centre. Nature 2010, 465, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Huh, J.; Kwon, Y.W.; Park, D.; Yu, Y.; Jang, Y.E.; Lee, B.-R.; Jo, E.; Lee, E.J.; Heo, Y. Biological conversion of methane to methanol through genetic reassembly of native catalytic domains. Nature Catalysis 2019, 2, 342–353. [Google Scholar] [CrossRef]
- Bennett, R.K.; Nyaradzo, D.; Michael, D.; Stephanie, J.; Kelley, H.; Baolong, Z.; Noah, H.; Derek, G.; Elizabeth, C.; Eleftherios, T.P. Expression of soluble methane monooxygenase in Escherichia coli enables methane conversion. bioRxiv 2021, 2021.2008.2005.455234. [Google Scholar] [CrossRef]
- Raili, K.; Markku, H.; Pekka, O. Methanol-Managing greenhouse gas emissions in the production chain by optimizing the resource base. AIMS Energy 2018, 6, 1074–1102. [Google Scholar] [CrossRef]
- Du, X.-L.; Jiang, Z.; Su, D.S.; Wang, J.-Q. Research Progress on the Indirect Hydrogenation of Carbon Dioxide to Methanol. ChemSusChem 2016, 9, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, W.B.; Sandoval, N.R.; Bennett, R.K.; Fast, A.G.; Papoutsakis, E.T. Synthetic methylotrophy: engineering the production of biofuels and chemicals based on the biology of aerobic methanol utilization. Current Opinion in Biotechnology 2015, 33, 165–175. [Google Scholar] [CrossRef]
- Chen, A.Y.; Lan, E.I. Chemical Production from Methanol Using Natural and Synthetic Methylotrophs. Biotechnology Journal 2020, 15, 1900356. [Google Scholar] [CrossRef]
- Pfeifenschneider, J.; Brautaset, T.; Wendisch, V.F. Methanol as carbon substrate in the bio-economy: Metabolic engineering of aerobic methylotrophic bacteria for production of value-added chemicals. Biofuels, Bioproducts and Biorefining 2017, 11, 719–731. [Google Scholar] [CrossRef]
- Antoniewicz, M.R. Synthetic methylotrophy: strategies to assimilate methanol for growth and chemicals production. Current opinion in biotechnology 2019, 59, 165–174. [Google Scholar] [CrossRef]
- Yurimoto, H.; Sakai, Y. Methylotrophic yeasts: current understanding of their C1-metabolism and its regulation by sensing methanol for survival on plant leaves. Current Issues in Molecular Biology 2019, 33, 197–210. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Nicaud, J.M. Metabolic Engineering for Expanding the Substrate Range of Yarrowia lipolytica. Trends Biotechnol 2016, 34, 798–809. [Google Scholar] [CrossRef]
- Lv, X.; Yu, W.; Zhang, C.; Ning, P.; Li, J.; Liu, Y.; Du, G.; Liu, L. C1-based biomanufacturing: Advances, challenges and perspectives. Bioresource Technology 2023, 367, 128259. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Li, X.; Yang, Y.; Nielsen, J.; Bai, Z.; Chen, Y. Strategies and challenges with the microbial conversion of methanol to high-value chemicals. Biotechnology and Bioengineering 2021, 118, 3655–3668. [Google Scholar] [CrossRef] [PubMed]
- Kremp, F.; Müller, V. Methanol and methyl group conversion in acetogenic bacteria: biochemistry, physiology and application. FEMS Microbiology Reviews 2021, 45, fuaa040. [Google Scholar] [CrossRef] [PubMed]
- Gregory, G.J.; Bennett, R.K.; Papoutsakis, E.T. Recent advances toward the bioconversion of methane and methanol in synthetic methylotrophs. Metabolic Engineering 2022, 71, 99–116. [Google Scholar] [CrossRef]
- Song, Y.; Bae, J.; Shin, J.; Jin, S.; Kang, S.; Lee, H.; Cho, S.; Cho, B.-K. Systems Biology on Acetogenic Bacteria for Utilizing C1 Feedstocks. In One-Carbon Feedstocks for Sustainable Bioproduction; Springer: 2022; pp. 57-90.
- Bennett, R.K.; Steinberg, L.M.; Chen, W.; Papoutsakis, E.T. Engineering the bioconversion of methane and methanol to fuels and chemicals in native and synthetic methylotrophs. Current Opinion in Biotechnology 2018, 50, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qin, R.; Guo, Y.; Ma, C.; Wang, X.; Chen, K.; Ouyang, P. Engineering the native methylotrophs for the bioconversion of methanol to value-added chemicals: current status and future perspectives. Green Chemical Engineering 2023, 4, 199–211. [Google Scholar] [CrossRef]
- Schultenkämper, K.; Gütle, D.D.; López, M.G.; Keller, L.B.; Zhang, L.; Einsle, O.; Jacquot, J.-P.; Wendisch, V.F. Interrogating the Role of the Two Distinct Fructose-Bisphosphate Aldolases of Bacillus methanolicus by Site-Directed Mutagenesis of Key Amino Acids and Gene Repression by CRISPR Interference. Frontiers in Microbiology 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.K.; Villada, J.C.; Chalifour, A.; Duran, M.F.; Lu, H.; Lee, P.K. Designing and engineering Methylorubrum extorquens AM1 for itaconic acid production. Frontiers in Microbiology 2019, 10, 1027. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.Y.-H.; Jung, H.-W.; Tsuei, C.-Y.; Liao, J.C. Converting Escherichia coli to a synthetic methylotroph growing solely on methanol. Cell 2020, 182, 933–946. e914. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Wu, X.; Deng, J.; Gao, L.; Shen, Y.; Yao, L.; Zhou, Y.J. Methanol biotransformation toward high-level production of fatty acid derivatives by engineering the industrial yeast Pichia pastoris. Proceedings of the National Academy of Sciences 2022, 119, e2201711119. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, Y.; Yu, W.; Zhou, Y.J. Rescuing yeast from cell death enables overproduction of fatty acids from sole methanol. Nature Metabolism 2022, 4, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhao, N.; Deng, J.; Gu, Y.; Jia, S.; Hou, Y.; Lv, X.; Liu, L. Constructing a methanol-dependent Bacillus subtilis by engineering the methanol metabolism. Journal of Biotechnology 2022, 343, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Zhao, T.; Bankefa, O.E.; Li, Y. Engineering unnatural methylotrophic cell factories for methanol-based biomanufacturing: Challenges and opportunities. Biotechnology advances 2020, 39, 107467. [Google Scholar] [CrossRef]
- Sanford, P.A.; Woolston, B.M. Synthetic or natural? Metabolic engineering for assimilation and valorization of methanol. Current Opinion in Biotechnology 2022, 74, 171–179. [Google Scholar] [CrossRef]
- Nguyen, A.D.; Park, J.Y.; Hwang, I.Y.; Hamilton, R.; Kalyuzhnaya, M.G.; Kim, D.; Lee, E.Y. Genome-scale evaluation of core one-carbon metabolism in gammaproteobacterial methanotrophs grown on methane and methanol. Metabolic Engineering 2020, 57, 1–12. [Google Scholar] [CrossRef]
- Müller, J.E.; Meyer, F.; Litsanov, B.; Kiefer, P.; Potthoff, E.; Heux, S.; Quax, W.J.; Wendisch, V.F.; Brautaset, T.; Portais, J.-C. Engineering Escherichia coli for methanol conversion. Metabolic engineering 2015, 28, 190–201. [Google Scholar] [CrossRef]
- Price, J.V.; Chen, L.; Whitaker, W.B.; Papoutsakis, E.; Chen, W. Scaffoldless engineered enzyme assembly for enhanced methanol utilization. Proceedings of the National Academy of Sciences 2016, 113, 12691–12696. [Google Scholar] [CrossRef] [PubMed]
- Woolston, B.M.; King, J.R.; Reiter, M.; Van Hove, B.; Stephanopoulos, G. Improving formaldehyde consumption drives methanol assimilation in engineered E. coli. Nature communications 2018, 9, 2387. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Keller, P.; Hartl, J.; Gröninger, O.G.; Kiefer, P.; Vorholt, J.A. Methanol-essential growth of Escherichia coli. Nature communications 2018, 9, 1508. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, M.I.; Gonzalez-Garcia, R.A.; Valgepea, K.; Plan, M.R.; Scott, C.; Pretorius, I.S.; Marcellin, E.; Paulsen, I.T.; Williams, T.C. Adaptive laboratory evolution of native methanol assimilation in Saccharomyces cerevisiae. Nature Communications 2020, 11, 5564. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Olofsson-Dolk, M.; Hansson, F.G.; Donati, S.; Li, X.; Chang, H.; Cheng, J.; Dahlin, J.; Borodina, I. Engineering Yeast Yarrowia lipolytica for Methanol Assimilation. ACS Synthetic Biology 2021, 10, 3537–3550. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Hernandez Villamor, D.; Peng, H.; Chen, J.; Liu, L.; Haritos, V.; Ledesma-Amaro, R. Metabolic engineering strategies to enable microbial utilization of C1 feedstocks. Nat Chem Biol 2021, 17, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Yishai, O.; Lindner, S.N.; Gonzalez de la Cruz, J.; Tenenboim, H.; Bar-Even, A. The formate bio-economy. Current Opinion in Chemical Biology 2016, 35, 1–9. [Google Scholar] [CrossRef]
- Bar-Even, A.; Noor, E.; Flamholz, A.; Milo, R. Design and analysis of metabolic pathways supporting formatotrophic growth for electricity-dependent cultivation of microbes. Biochimica et Biophysica Acta (BBA)-Bioenergetics 2013, 1827, 1039–1047. [Google Scholar] [CrossRef]
- Bar-Even, A.; Flamholz, A.; Noor, E.; Milo, R. Thermodynamic constraints shape the structure of carbon fixation pathways. Biochimica et Biophysica Acta (BBA)-Bioenergetics 2012, 1817, 1646–1659. [Google Scholar] [CrossRef]
- Grunwald, S.; Mottet, A.; Grousseau, E.; Plassmeier, J.K.; Popović, M.K.; Uribelarrea, J.-L.; Gorret, N.; Guillouet, S.E.; Sinskey, A. Kinetic and stoichiometric characterization of organoautotrophic growth of Ralstonia eutropha on formic acid in fed-batch and continuous cultures. Microbial Biotechnology 2015, 8, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Xu, S.; Liu, Z.; Fu, X.; Zhao, H.; Shi, S. Challenges and opportunities in C1-based biomanufacturing. Bioresource Technology 2022, 364, 128095. [Google Scholar] [CrossRef] [PubMed]
- Claassens, N.J.; He, H.; Bar-Even, A. Synthetic Methanol and Formate Assimilation Via Modular Engineering and Selection Strategies. Current Issues in Molecular Biology 2019, 33, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Bar-Even, A. Formate Assimilation: The Metabolic Architecture of Natural and Synthetic Pathways. Biochemistry 2016, 55, 3851–3863. [Google Scholar] [CrossRef] [PubMed]
- Claassens, N.J. Reductive Glycine Pathway: A Versatile Route for One-Carbon Biotech. Trends in Biotechnology 2021, 39, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Claassens, N.J.; Bordanaba-Florit, G.; Cotton, C.A.; De Maria, A.; Finger-Bou, M.; Friedeheim, L.; Giner-Laguarda, N.; Munar-Palmer, M.; Newell, W.; Scarinci, G. Replacing the Calvin cycle with the reductive glycine pathway in Cupriavidus necator. Metabolic Engineering 2020, 62, 30–41. [Google Scholar] [CrossRef]
- Sánchez-Andrea, I.; Guedes, I.A.; Hornung, B.; Boeren, S.; Lawson, C.E.; Sousa, D.Z.; Bar-Even, A.; Claassens, N.J.; Stams, A.J.M. The reductive glycine pathway allows autotrophic growth of Desulfovibrio desulfuricans. Nature Communications 2020, 11, 5090. [Google Scholar] [CrossRef]
- Hong, Y.; Arbter, P.; Wang, W.; Rojas, L.N.; Zeng, A.-P. Introduction of glycine synthase enables uptake of exogenous formate and strongly impacts the metabolism in Clostridium pasteurianum. Biotechnology and Bioengineering 2021, 118, 1366–1380. [Google Scholar] [CrossRef]
- Yishai, O.; Goldbach, L.; Tenenboim, H.; Lindner, S.N.; Bar-Even, A. Engineered assimilation of exogenous and endogenous formate in Escherichia coli. ACS Synthetic Biology 2017, 6, 1722–1731. [Google Scholar] [CrossRef]
- Yishai, O.; Bouzon, M.; Döring, V.; Bar-Even, A. In vivo assimilation of one-carbon via a synthetic reductive glycine pathway in Escherichia coli. ACS synthetic biology 2018, 7, 2023–2028. [Google Scholar] [CrossRef]
- Tashiro, Y.; Hirano, S.; Matson, M.M.; Atsumi, S.; Kondo, A. Electrical-biological hybrid system for CO2 reduction. Metabolic engineering 2018, 47, 211–218. [Google Scholar] [CrossRef]
- Kim, S.; Lindner, S.N.; Aslan, S.; Yishai, O.; Wenk, S.; Schann, K.; Bar-Even, A. Growth of E. coli on formate and methanol via the reductive glycine pathway. Nature chemical biology 2020, 16, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Machens, F.; Messerschmidt, K.; Bar-Even, A. Core Catalysis of the Reductive Glycine Pathway Demonstrated in Yeast. ACS Synthetic Biology 2019, 8, 911–917. [Google Scholar]
- Neuendorf, C.S.; Vignolle, G.A.; Derntl, C.; Tomin, T.; Novak, K.; Mach, R.L.; Birner-Grünberger, R.; Pflügl, S. A quantitative metabolic analysis reveals Acetobacterium woodii as a flexible and robust host for formate-based bioproduction. Metabolic Engineering 2021, 68, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.; Dönig, J.; Kramer, S.; Poehlein, A.; Daniel, R.; Müller, V. Formate metabolism in the acetogenic bacterium Acetobacterium woodii. Environmental Microbiology 2021, 23, 4214–4227. [Google Scholar] [CrossRef]
- Wood, J.C.; Marcellin, E.; Plan, M.R.; Virdis, B. High methanol-to-formate ratios induce butanol production in Eubacterium limosum. Microbial Biotechnology 2022, 15, 1542–1549. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, P. Formate as an inhibitor of cytochrome c oxidase. Biochemical and biophysical research communications 1975, 67, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, T.; Gill, R.T. Organic acid toxicity, tolerance, and production in Escherichia coli biorefining applications. Microbial Cell Factories 2005, 4, 25. [Google Scholar] [CrossRef]
- Overkamp, K.M.; Kötter, P.; van der Hoek, R.; Schoondermark-Stolk, S.; Luttik, M.A.; van Dijken, J.P.; Pronk, J.T. Functional analysis of structural genes for NAD+-dependent formate dehydrogenase in Saccharomyces cerevisiae. Yeast 2002, 19, 509–520. [Google Scholar] [CrossRef]
- Litty, D.; Müller, V. Butyrate production in the acetogen Eubacterium limosum is dependent on the carbon and energy source. Microbial Biotechnology 2021, 14, 2686–2692. [Google Scholar] [CrossRef]
- Du, C.; Li, Y.; Xiang, R.; Yuan, W. Formate dehydrogenase improves the resistance to formic acid and acetic acid simultaneously in Saccharomyces cerevisiae. International Journal of Molecular Sciences 2022, 23, 3406. [Google Scholar] [CrossRef]
- Babel, W. The Auxiliary Substrate Concept: From simple considerations to heuristically valuable knowledge. Engineering in Life Sciences 2009, 9, 285–290. [Google Scholar] [CrossRef]
- Nielsen, J.; Keasling, Jay D. Engineering Cellular Metabolism. Cell 2016, 164, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.G.; Lee, J.H.; Noh, M.H.; Jung, G.Y. Rediscovering Acetate Metabolism: Its Potential Sources and Utilization for Biobased Transformation into Value-Added Chemicals. Journal of Agricultural and Food Chemistry 2018, 66, 3998–4006. [Google Scholar] [CrossRef]
- Kim, Y.; Lama, S.; Agrawal, D.; Kumar, V.; Park, S. Acetate as a potential feedstock for the production of value-added chemicals: Metabolism and applications. Biotechnology Advances 2021, 49, 107736. [Google Scholar] [CrossRef]
- Mutyala, S.; Kim, J.R. Recent advances and challenges in the bioconversion of acetate to value-added chemicals. Bioresource Technology 2022, 364, 128064. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sun, X.; Yuan, Q. Strategies for enhancing microbial tolerance to inhibitors for biofuel production: a review. Bioresource Technology 2018, 258, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Kutscha, R.; Pflugl, S. Microbial Upgrading of Acetate into Value-Added Products-Examining Microbial Diversity, Bioenergetic Constraints and Metabolic Engineering Approaches. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Zhou, S.; Lama, S.; Jiang, J.; Sankaranarayanan, M.; Park, S. Use of acetate for the production of 3-hydroxypropionic acid by metabolically-engineered Pseudomonas denitrificans. Bioresource Technology 2020, 307, 123194. [Google Scholar] [CrossRef]
- Seong, W.; Han, G.H.; Lim, H.S.; Baek, J.I.; Kim, S.-J.; Kim, D.; Kim, S.K.; Lee, H.; Kim, H.; Lee, S.-G.; et al. Adaptive laboratory evolution of Escherichia coli lacking cellular byproduct formation for enhanced acetate utilization through compensatory ATP consumption. Metabolic Engineering 2020, 62, 249–259. [Google Scholar] [CrossRef]
- Gong, G.; Wu, B.; Liu, L.; Li, J.; Zhu, Q.; He, M.; Hu, G. Metabolic engineering using acetate as a promising building block for the production of bio-based chemicals. Engineering Microbiology 2022, 2, 100036. [Google Scholar] [CrossRef]
- Deng, Y.; Ma, N.; Zhu, K.; Mao, Y.; Wei, X.; Zhao, Y. Balancing the carbon flux distributions between the TCA cycle and glyoxylate shunt to produce glycolate at high yield and titer in Escherichia coli. Metabolic engineering 2018, 46, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Lama, S.; Kim, Y.; Nguyen, D.T.; Im, C.H.; Sankaranarayanan, M.; Park, S. Production of 3-hydroxypropionic acid from acetate using metabolically-engineered and glucose-grown Escherichia coli. Bioresource Technology 2021, 320, 124362. [Google Scholar] [CrossRef]
- Chang, Z.; Dai, W.; Mao, Y.; Cui, Z.; Zhang, Z.; Wang, Z.; Ma, H.; Chen, T. Enhanced 3-hydroxypropionic acid production from acetate via the malonyl-CoA pathway in Corynebacterium glutamicum. Frontiers in Bioengineering and Biotechnology 2022, 9, 808258. [Google Scholar] [CrossRef]
- Yang, S.; Li, S.; Jia, X. Production of medium chain length polyhydroxyalkanoate from acetate by engineered Pseudomonas putida KT2440. Journal of Industrial Microbiology and Biotechnology 2019, 46, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Quarterman, J.; Kim, S.R.; Cate, J.H.; Jin, Y.-S. Enhanced biofuel production through coupled acetic acid and xylose consumption by engineered yeast. Nature communications 2013, 4, 2580. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-f.; Shen, Y.; Luo, H.-j.; Liu, J.-n.; Liu, J. Enhancement of extracellular lipid production by oleaginous yeast through preculture and sequencing batch culture strategy with acetic acid. Bioresource Technology 2018, 247, 395–401. [Google Scholar] [CrossRef]
- Gong, G.; Zhang, X.; Tan, T. Simultaneously enhanced intracellular lipogenesis and β-carotene biosynthesis of Rhodotorula glutinis by light exposure with sodium acetate as the substrate. Bioresource technology 2020, 295, 122274. [Google Scholar] [CrossRef]
- Chen, L.; Yan, W.; Qian, X.; Chen, M.; Zhang, X.; Xin, F.; Zhang, W.; Jiang, M.; Ochsenreither, K. Increased lipid production in Yarrowia lipolytica from acetate through metabolic engineering and Cosubstrate fermentation. ACS synthetic biology 2021, 10, 3129–3138. [Google Scholar] [CrossRef]
- Kövilein, A.; Umpfenbach, J.; Ochsenreither, K. Acetate as substrate for l-malic acid production with Aspergillus oryzae DSM 1863. Biotechnology for Biofuels 2021, 14, 1–15. [Google Scholar] [CrossRef]
- Sun, S.; Ding, Y.; Liu, M.; Xian, M.; Zhao, G. Comparison of glucose, acetate and ethanol as carbon resource for production of poly (3-hydroxybutyrate) and other acetyl-CoA derivatives. Frontiers in bioengineering and biotechnology 2020, 8, 833. [Google Scholar] [CrossRef]
- Jo, M.; Noh, M.H.; Lim, H.G.; Kang, C.W.; Im, D.-K.; Oh, M.-K.; Jung, G.Y. Precise tuning of the glyoxylate cycle in Escherichia coli for efficient tyrosine production from acetate. Microbial Cell Factories 2019, 18, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, N.; Qiao, K.; Vogg, S.; Stephanopoulos, G. Application of metabolic controls for the maximization of lipid production in semicontinuous fermentation. Proc Natl Acad Sci U S A 2017, 114, E5308–E5316. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, Y.; Desai, S.H.; Atsumi, S. Two-dimensional isobutyl acetate production pathways to improve carbon yield. Nature communications 2015, 6, 7488. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Nie, Q. Engineering Escherichia coli to convert acetic acid to β-caryophyllene. Microbial cell factories 2016, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.H.; Turanlı-Yıldız, B.; Liu, D.; Resch, M.G.; Fink, G.R.; Stephanopoulos, G. Engineered yeast tolerance enables efficient production from toxified lignocellulosic feedstocks. Science Advances 2021, 7, eabf7613. [Google Scholar] [CrossRef] [PubMed]
- Hahn-Hägerdal, B.; Galbe, M.; Gorwa-Grauslund, M.-F.; Lidén, G.; Zacchi, G. Bio-ethanol–the fuel of tomorrow from the residues of today. Trends in biotechnology 2006, 24, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Ma, X.; Ning, W.; Liu, Y.; Sinskey, A.J.; Stephanopoulos, G.; Zhou, K. Constructing an ethanol utilization pathway in Escherichia coli to produce acetyl-CoA derived compounds. Metabolic Engineering 2021, 65, 223–231. [Google Scholar] [CrossRef]
- Liu, Y.; Bai, C.; Liu, Q.; Xu, Q.; Qian, Z.; Peng, Q.; Yu, J.; Xu, M.; Zhou, X.; Zhang, Y.; et al. Engineered ethanol-driven biosynthetic system for improving production of acetyl-CoA derived drugs in Crabtree-negative yeast. Metabolic Engineering 2019, 54, 275–284. [Google Scholar] [CrossRef]
- Cao, Y.; Mu, H.; Guo, J.; Liu, H.; Zhang, R.; Liu, W.; Xian, M.; Liu, H. Metabolic engineering of Escherichia coli for the utilization of ethanol. J Biol Res (Thessalon) 2020, 27, 1. [Google Scholar] [CrossRef]
- Westfall, P.J.; Pitera, D.J.; Lenihan, J.R.; Eng, D.; Woolard, F.X.; Regentin, R.; Horning, T.; Tsuruta, H.; Melis, D.J.; Owens, A. Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin. Proceedings of the National Academy of Sciences 2012, 109, E111–E118. [Google Scholar] [CrossRef] [PubMed]
- Felenbok, B.; Flipphi, M.; Nikolaev, I. Ethanol catabolism in Aspergillus nidulans: a model system for studying gene regulation. 2001.
- Ricci, L.; Seifert, A.; Bernacchi, S.; Fino, D.; Pirri, C.F.; Re, A. Leveraging substrate flexibility and product selectivity of acetogens in two-stage systems for chemical production. Microbial Biotechnology 2023, 16, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Rouboa, A.; Silva, V.; Monteiro, E.; Bouziane, K. Influence of the biomass gasification processes on the final composition of syngas. Energy procedia 2013, 36, 596–606. [Google Scholar] [CrossRef]
- Molino, A.; Chianese, S.; Musmarra, D. Biomass gasification technology: The state of the art overview. Journal of Energy Chemistry 2016, 25, 10–25. [Google Scholar] [CrossRef]
- Ahmad, A.A.; Zawawi, N.A.; Kasim, F.H.; Inayat, A.; Khasri, A. Assessing the gasification performance of biomass: A review on biomass gasification process conditions, optimization and economic evaluation. Renewable and Sustainable Energy Reviews 2016, 53, 1333–1347. [Google Scholar] [CrossRef]
- Griffin, D.W.; Schultz, M.A. Fuel and chemical products from biomass syngas: a comparison of gas fermentation to thermochemical conversion routes. Environmental progress & sustainable energy 2012, 31, 219–224. [Google Scholar]
- Daniell, J.; Köpke, M.; Simpson, S.D. Commercial biomass syngas fermentation. Energies 2012, 5, 5372–5417. [Google Scholar] [CrossRef]
- Akhtar, A.; Krepl, V.; Ivanova, T. A combined overview of combustion, pyrolysis, and gasification of biomass. Energy & Fuels 2018, 32, 7294–7318. [Google Scholar]
- Phillips, J.R.; Huhnke, R.L.; Atiyeh, H.K. Syngas fermentation: a microbial conversion process of gaseous substrates to various products. Fermentation 2017, 3, 28. [Google Scholar] [CrossRef]
- Lemgruber, R.d.S.P.; Valgepea, K.; Tappel, R.; Behrendorff, J.B.; Palfreyman, R.W.; Plan, M.; Hodson, M.P.; Simpson, S.D.; Nielsen, L.K.; Köpke, M. Systems-level engineering and characterisation of Clostridium autoethanogenum through heterologous production of poly-3-hydroxybutyrate (PHB). Metabolic engineering 2019, 53, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Pavan, M.; Reinmets, K.; Garg, S.; Mueller, A.P.; Marcellin, E.; Köpke, M.; Valgepea, K. Advances in systems metabolic engineering of autotrophic carbon oxide-fixing biocatalysts towards a circular economy. Metabolic Engineering 2022, 71, 117–141. [Google Scholar] [CrossRef]
- Pala, L.P.R.; Wang, Q.; Kolb, G.; Hessel, V. Steam gasification of biomass with subsequent syngas adjustment using shift reaction for syngas production: An Aspen Plus model. Renewable Energy 2017, 101, 484–492. [Google Scholar] [CrossRef]
- Gunes, B. A critical review on biofilm-based reactor systems for enhanced syngas fermentation processes. Renewable and Sustainable Energy Reviews 2021, 143, 110950. [Google Scholar] [CrossRef]
- Stoll, I.K.; Boukis, N.; Sauer, J. Syngas Fermentation to Alcohols: Reactor Technology and Application Perspective. Chemie Ingenieur Technik 2020, 92, 125–136. [Google Scholar] [CrossRef]
- Köpke, M.; Simpson, S.D. Pollution to products: recycling of ‘above ground’carbon by gas fermentation. Current Opinion in Biotechnology 2020, 65, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Diels, L.; Pant, D.; Patil, S.A.; Ghangrekar, M.M. Review—Microbial Electrosynthesis: A Way Towards The Production of Electro-Commodities Through Carbon Sequestration with Microbes as Biocatalysts. Journal of The Electrochemical Society 2020, 167, 155510. [Google Scholar] [CrossRef]
- Fan, L.; Xia, C.; Zhu, P.; Lu, Y.; Wang, H. Electrochemical CO2 reduction to high-concentration pure formic acid solutions in an all-solid-state reactor. Nature Communications 2020, 11, 3633. [Google Scholar] [CrossRef]
- Ahmad, W.; Koley, P.; Dwivedi, S.; Lakshman, R.; Shin, Y.K.; van Duin, A.C.T.; Shrotri, A.; Tanksale, A. Aqueous phase conversion of CO2 into acetic acid over thermally transformed MIL-88B catalyst. Nature Communications 2023, 14, 2821. [Google Scholar] [CrossRef]
- Xu, J.; Liu, N.; Qiao, K.; Vogg, S.; Stephanopoulos, G. Application of metabolic controls for the maximization of lipid production in semicontinuous fermentation. Proceedings of the National Academy of Sciences 2017, 114, E5308–E5316. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Tran, V.G.; Qin, J.; Olson, A.; Mishra, S.; Schultz John, C.; Huang, C.; Xie, D.; Zhao, H. Metabolic engineering of oleaginous yeast Rhodotorula toruloides for overproduction of triacetic acid lactone. Biotechnology and Bioengineering 2022, 119, 2529–2540. [Google Scholar] [CrossRef]
- Lawson, C.E.; Martí, J.M.; Radivojevic, T.; Jonnalagadda, S.V.R.; Gentz, R.; Hillson, N.J.; Peisert, S.; Kim, J.; Simmons, B.A.; Petzold, C.J.; et al. Machine learning for metabolic engineering: A review. Metabolic Engineering 2021, 63, 34–60. [Google Scholar] [CrossRef] [PubMed]
- Helmy, M.; Smith, D.; Selvarajoo, K. Systems biology approaches integrated with artificial intelligence for optimized metabolic engineering. Metab Eng Commun 2020, 11, e00149. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.D.; Setati, M.E.; Divol, B. Redox cofactor metabolism in Saccharomyces cerevisiae and its impact on the production of alcoholic fermentation end-products. Food Research International 2023, 163, 112276. [Google Scholar] [CrossRef] [PubMed]
- Bloem, A.; Sanchez, I.; Dequin, S.; Camarasa, C. Metabolic Impact of Redox Cofactor Perturbations on the Formation of Aroma Compounds in Saccharomyces cerevisiae. Applied and Environmental Microbiology 2016, 82, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-G.; Xue, C.; Lin, Y.-H.; Bai, F.-W. Redox potential control and applications in microaerobic and anaerobic fermentations. Biotechnology Advances 2013, 31, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Scalcinati, G.; Oldiges, M.; Vemuri, G.N. Metabolic impact of increased NADH availability in Saccharomyces cerevisiae. Appl Environ Microbiol 2010, 76, 851–859. [Google Scholar] [CrossRef]
- Qiao, K.; Wasylenko, T.M.; Zhou, K.; Xu, P.; Stephanopoulos, G. Lipid production in Yarrowia lipolytica is maximized by engineering cytosolic redox metabolism. Nature Biotechnology 2017, 35, 173–177. [Google Scholar] [CrossRef]
- Kim, J.-E.; Jang, I.-S.; Sung, B.H.; Kim, S.C.; Lee, J.Y. Rerouting of NADPH synthetic pathways for increased protopanaxadiol production in Saccharomyces cerevisiae. Scientific Reports 2018, 8, 15820. [Google Scholar] [CrossRef]
- Qiao, K.; Imam Abidi, S.H.; Liu, H.; Zhang, H.; Chakraborty, S.; Watson, N.; Kumaran Ajikumar, P.; Stephanopoulos, G. Engineering lipid overproduction in the oleaginous yeast Yarrowia lipolytica. Metabolic Engineering 2015, 29, 56–65. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, S.-X.; Gandionco, K.A.; Bond, A.M.; Zhang, J. Electrocatalytic carbon dioxide reduction: from fundamental principles to catalyst design. Materials Today Advances 2020, 7, 100074. [Google Scholar] [CrossRef]
Figure 1.
An overview of the CO2 conversion approaches and using CO2-derived C1/C2 chemicals for biomanufacturing of common products. Energy conversion and sources used in the conversion are summarized on the left. After CO2 is converted from inorganic to organic carbon substrates, various valuable chemicals can be biomanufactured through natural and synthetic microorganisms. The figure was generated using Biorender.
Figure 1.
An overview of the CO2 conversion approaches and using CO2-derived C1/C2 chemicals for biomanufacturing of common products. Energy conversion and sources used in the conversion are summarized on the left. After CO2 is converted from inorganic to organic carbon substrates, various valuable chemicals can be biomanufactured through natural and synthetic microorganisms. The figure was generated using Biorender.
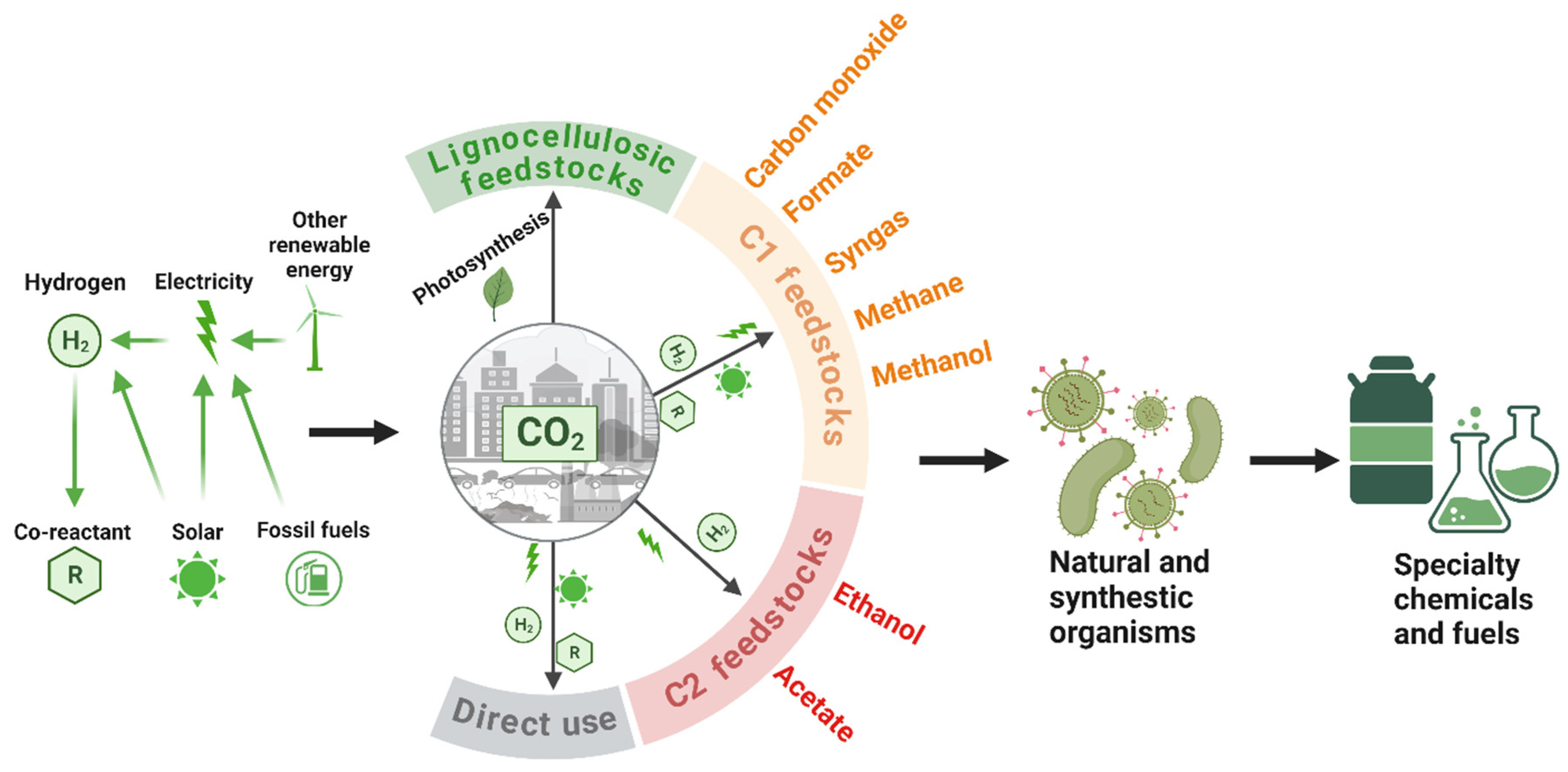
Figure 2.
An overview of natural direct CO2 fixation pathways. Metabolites: ribulose 5-phosphate, Ru5P; ribulose 1,5-bisphosphate, RuBP; 3-phosphoglycerate, 3PG; 1,3-bisphosphoglycerate, 1,3BPG; glyceraldehyde 3-phosphate, GAP; fructose 1,6-bisphosphate, FBP; fructose 6-phosphate, F6P; xylulose 5-phosphate, Xu5P; dihydroxyacetone phosphate, DHAP; erythrose 4-phosphate, E4P; sedoheptulose 1,7-bisphosphate, SBP; sedoheptulose 7-phosphate, S7P; ribose 5-phosphate, R5P; tetrahydrofolate, THF; (3S)-citramalyl-CoA, CM-CoA; mesaconyl-C4-CoA, MC-C4-CoA; mesaconyl-C1-CoA, MC-C1-CoA; beta-methylmalyl-CoA, βMM-CoA; propionyl-CoA, P-CoA; 3-hydroxypropionyl-CoA, 3-HP-CoA; 3-hydroxypropionate, 3-HP; malonyl-CoA, Mal-CoA; (S)-malyl-CoA, M-CoA; (S)-methylmalonyl-CoA, S-MM-CoA; (R)-methylmalonyl-CoA, R-MM-CoA; acetoacetyl-CoA, AcAc-CoA; acryloyl-CoA, Acr-CoA; crotonyl-CoA, C-CoA; 4-hydroxybutyrate, 4-HB; 4-hydroxybutyryl-CoA, 4-HB-CoA; succinate semialdehyde, SSA; (S)-3-hydroxybutyryl-CoA, (S) 3-HB-CoA; malonate semialdehyde, MSA; phosphoenolpyruvate, PEP. Enzymes: Ribulose-1,5-bisphosphate carboxylase, RuBisCo; phosphoribulokinase, prkA; carbon monoxide dehydrogenase, CODH; acetyl CoA synthase, ACS; formate dehydrogenase, FDH; serine hydroxymethyltransferase, GlyA; pyruvate synthase, PFOR; ATP-citrate lyase, ACL; acetyl-CoA carboxyltransferase, ACC; propionyl-CoA carboxylase, PCC; phosphoenolpyruvate carboxylase, PPC. Multi-step reactions are presented by continuous arrows. Special parts of WLP are shown dashed arrows. The figure was created with BioRender.
Figure 2.
An overview of natural direct CO2 fixation pathways. Metabolites: ribulose 5-phosphate, Ru5P; ribulose 1,5-bisphosphate, RuBP; 3-phosphoglycerate, 3PG; 1,3-bisphosphoglycerate, 1,3BPG; glyceraldehyde 3-phosphate, GAP; fructose 1,6-bisphosphate, FBP; fructose 6-phosphate, F6P; xylulose 5-phosphate, Xu5P; dihydroxyacetone phosphate, DHAP; erythrose 4-phosphate, E4P; sedoheptulose 1,7-bisphosphate, SBP; sedoheptulose 7-phosphate, S7P; ribose 5-phosphate, R5P; tetrahydrofolate, THF; (3S)-citramalyl-CoA, CM-CoA; mesaconyl-C4-CoA, MC-C4-CoA; mesaconyl-C1-CoA, MC-C1-CoA; beta-methylmalyl-CoA, βMM-CoA; propionyl-CoA, P-CoA; 3-hydroxypropionyl-CoA, 3-HP-CoA; 3-hydroxypropionate, 3-HP; malonyl-CoA, Mal-CoA; (S)-malyl-CoA, M-CoA; (S)-methylmalonyl-CoA, S-MM-CoA; (R)-methylmalonyl-CoA, R-MM-CoA; acetoacetyl-CoA, AcAc-CoA; acryloyl-CoA, Acr-CoA; crotonyl-CoA, C-CoA; 4-hydroxybutyrate, 4-HB; 4-hydroxybutyryl-CoA, 4-HB-CoA; succinate semialdehyde, SSA; (S)-3-hydroxybutyryl-CoA, (S) 3-HB-CoA; malonate semialdehyde, MSA; phosphoenolpyruvate, PEP. Enzymes: Ribulose-1,5-bisphosphate carboxylase, RuBisCo; phosphoribulokinase, prkA; carbon monoxide dehydrogenase, CODH; acetyl CoA synthase, ACS; formate dehydrogenase, FDH; serine hydroxymethyltransferase, GlyA; pyruvate synthase, PFOR; ATP-citrate lyase, ACL; acetyl-CoA carboxyltransferase, ACC; propionyl-CoA carboxylase, PCC; phosphoenolpyruvate carboxylase, PPC. Multi-step reactions are presented by continuous arrows. Special parts of WLP are shown dashed arrows. The figure was created with BioRender.
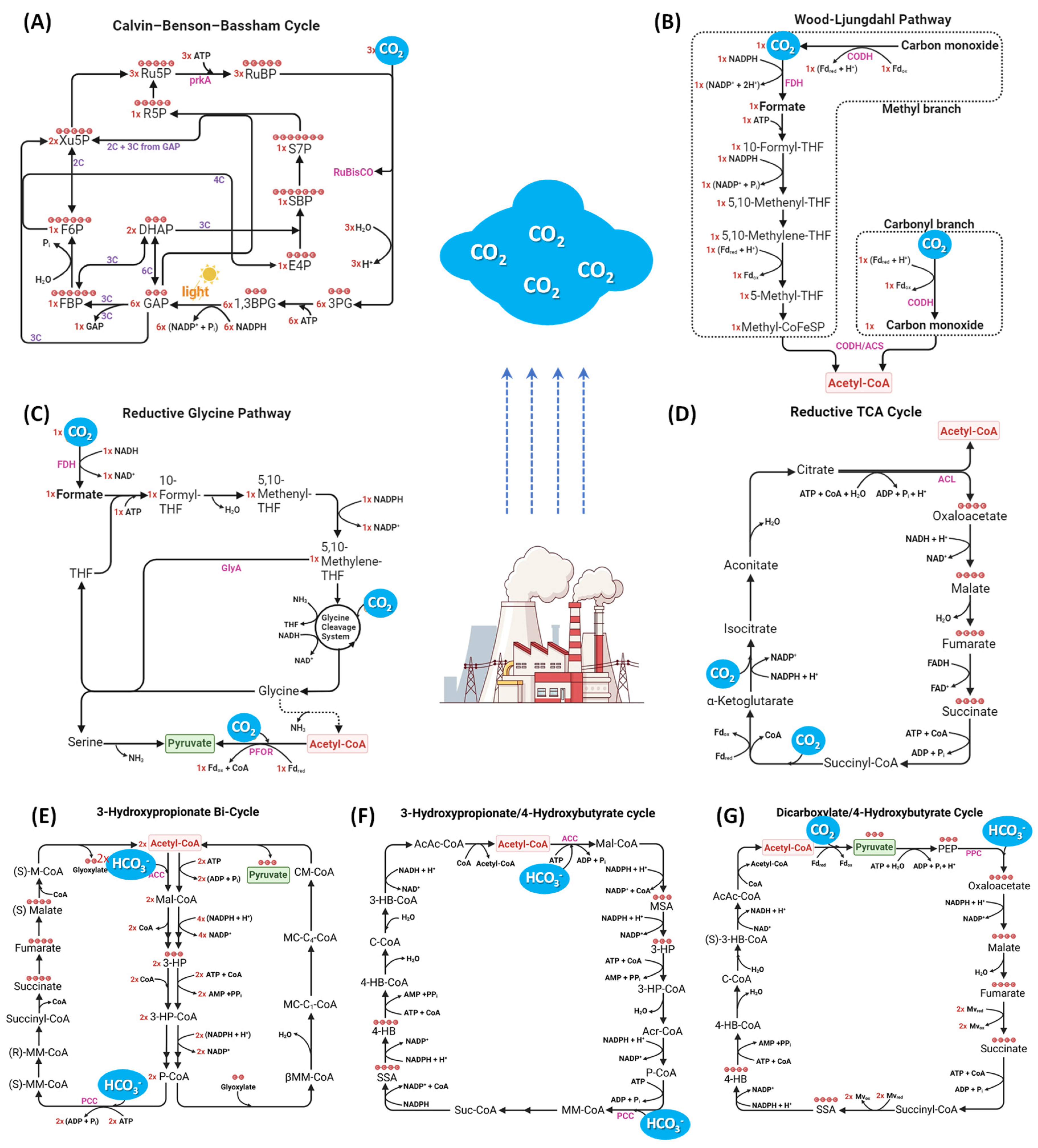
Figure 3.
A brief summary of the mechanism of microbial electrosynthesis that can be used for one-step CO2 fixation and conversion.
Figure 3.
A brief summary of the mechanism of microbial electrosynthesis that can be used for one-step CO2 fixation and conversion.
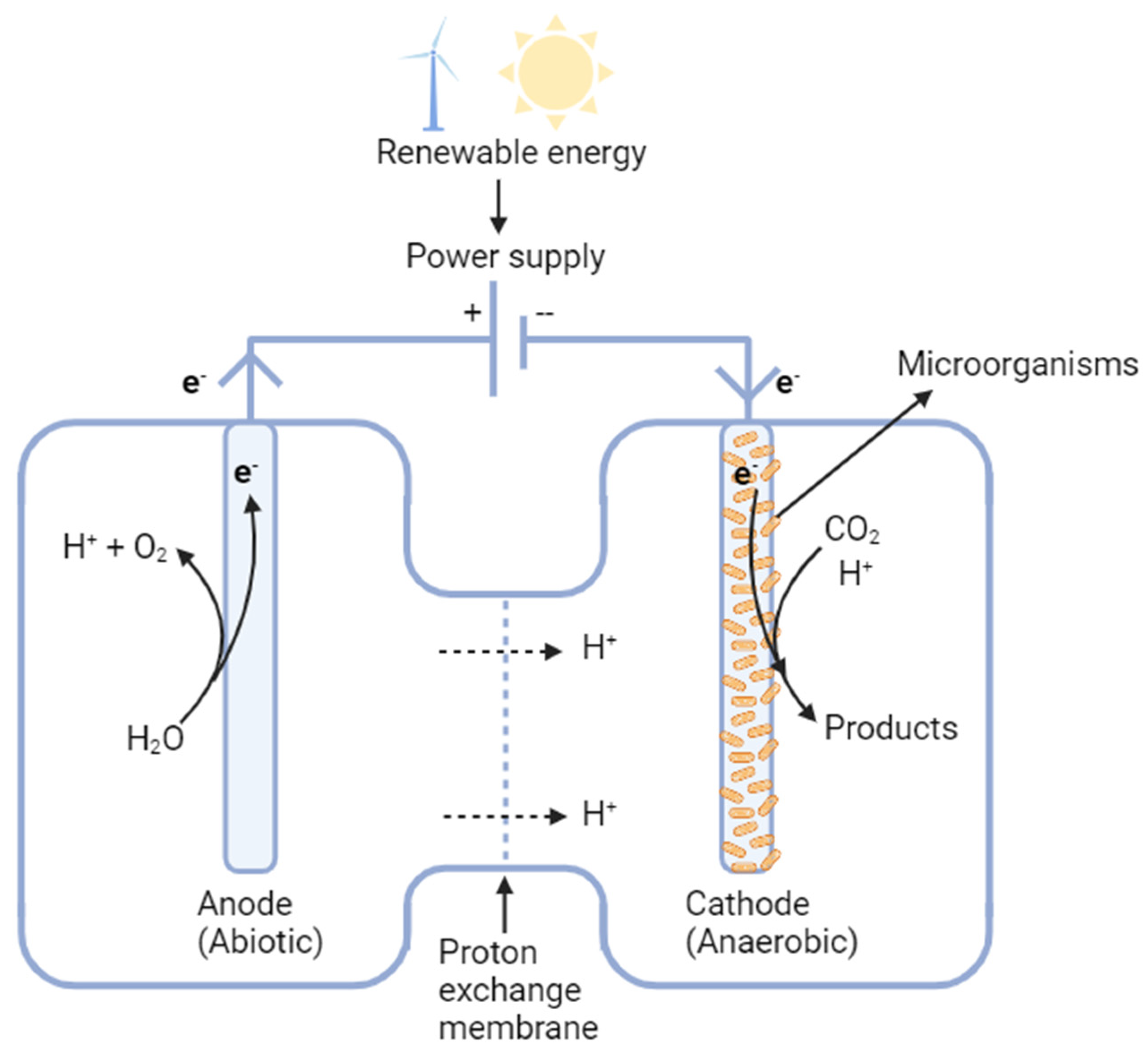
Figure 4.
Typical C1 utilization pathways. Metabolites: ribulose 5-phosphate, Ru5P; hexulose 6-phosphate, H6P; glyceraldehyde 3-phosphate, GAP; fructose 6-phosphate, F6P; fructose 1,6-bisphosphate, FBP; xylulose 5-phosphate, Xu5P; dihydroxyacetone, DHA; ribulose-1,5-bisphosphate, RuBP; tetrahydrofolate, THF; 3-phosphoglycerate, 3PG; 1,3-diphosphoglycerate, 1,3DPG; phosphoenolpyruvate, PEP; pyruvate, Pyr. Enzymes: carbon monoxide dehydrogenase, CODH; acetyl CoA synthase, ACS; membrane-bound methane monooxygenase, pMMO; cytoplasmic methane monooxygenase, sMMO; alcohol oxidase, Aox; methanol dehydrogenase, Mdh; formaldehyde dehydrogenase, FADH; formate dehydrogenase, FDH; serine hydroxymethyltransferase, GlyA; Ribulose-1,5-bisphosphate carboxylase, RuBisCo. Multi-step reactions are presented by dashed arrows. Special parts of WLP are shown in faded dashed arrows in the related color. The Figure was created with BioRender.
Figure 4.
Typical C1 utilization pathways. Metabolites: ribulose 5-phosphate, Ru5P; hexulose 6-phosphate, H6P; glyceraldehyde 3-phosphate, GAP; fructose 6-phosphate, F6P; fructose 1,6-bisphosphate, FBP; xylulose 5-phosphate, Xu5P; dihydroxyacetone, DHA; ribulose-1,5-bisphosphate, RuBP; tetrahydrofolate, THF; 3-phosphoglycerate, 3PG; 1,3-diphosphoglycerate, 1,3DPG; phosphoenolpyruvate, PEP; pyruvate, Pyr. Enzymes: carbon monoxide dehydrogenase, CODH; acetyl CoA synthase, ACS; membrane-bound methane monooxygenase, pMMO; cytoplasmic methane monooxygenase, sMMO; alcohol oxidase, Aox; methanol dehydrogenase, Mdh; formaldehyde dehydrogenase, FADH; formate dehydrogenase, FDH; serine hydroxymethyltransferase, GlyA; Ribulose-1,5-bisphosphate carboxylase, RuBisCo. Multi-step reactions are presented by dashed arrows. Special parts of WLP are shown in faded dashed arrows in the related color. The Figure was created with BioRender.
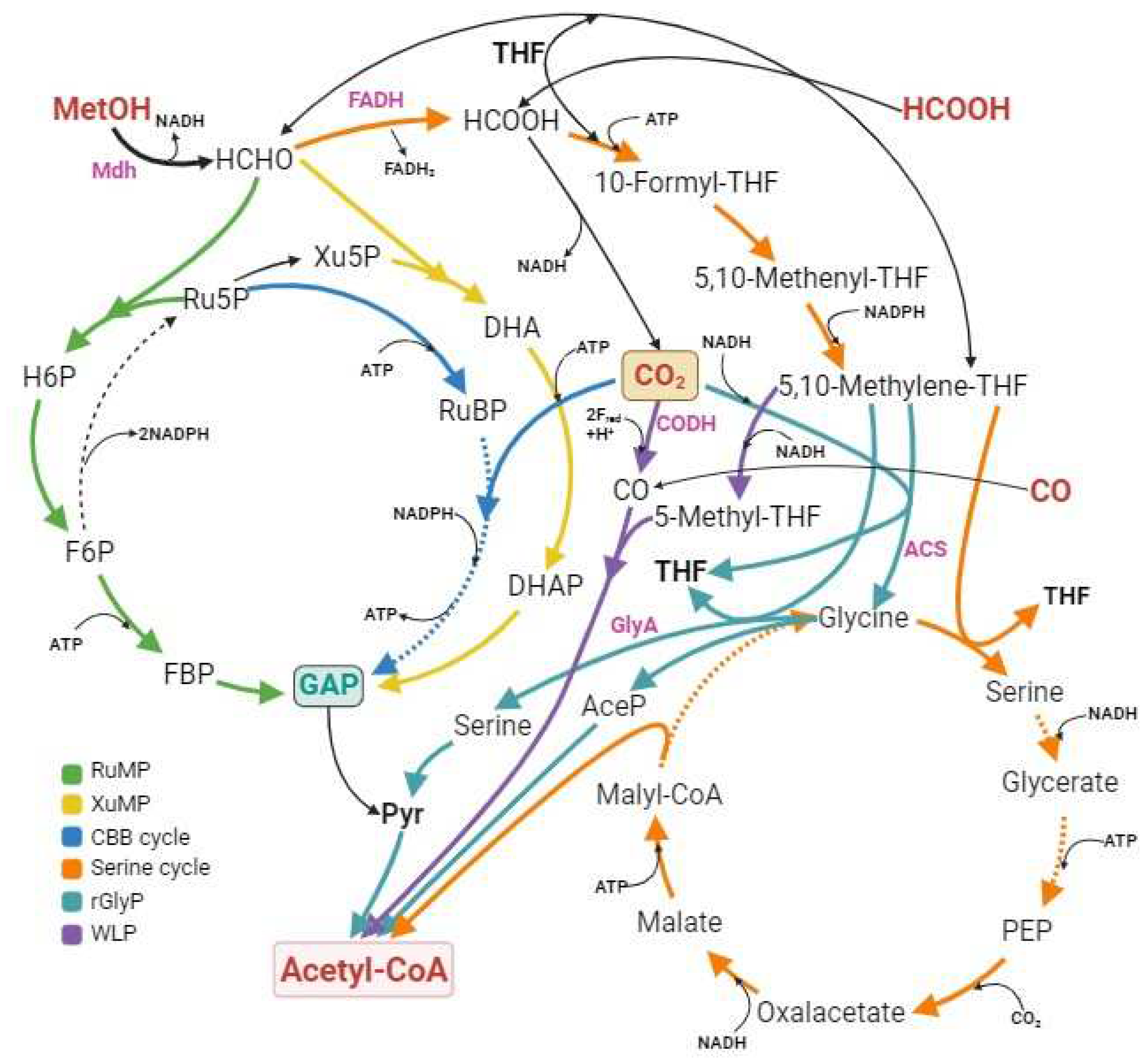
Figure 5.
Common C2 chemical assimilation pathways. Metabolites: Glyceraldehyde 3-phosphate, GAP; phosphoenolpyruvate, PEP. Enzymes: acetyl-CoA carboxylase, ACC; alcohol dehydrogenase, ADH; aldehyde dehydrogenase, ALD; acetaldehyde dehydrogenase, ALDH; acetyl-CoA synthetase, ACS; cytochrome P4502E1, CYP2E1; ATP-citrate lyase, ACL; glycerol-3-phosphate dehydrogenase, GPD; malic enzyme, ME; pyruvate dehydrogenase complex, PDC; phosphoenolpyruvate carboxykinase, PCK; pyruvate carboxylase, PYC; pyruvate kinase, PYK. Multi-step reactions are presented by dashed arrows in related color. Black dashed arrows represent metabolite transfer. The Figure was created with BioRender.
Figure 5.
Common C2 chemical assimilation pathways. Metabolites: Glyceraldehyde 3-phosphate, GAP; phosphoenolpyruvate, PEP. Enzymes: acetyl-CoA carboxylase, ACC; alcohol dehydrogenase, ADH; aldehyde dehydrogenase, ALD; acetaldehyde dehydrogenase, ALDH; acetyl-CoA synthetase, ACS; cytochrome P4502E1, CYP2E1; ATP-citrate lyase, ACL; glycerol-3-phosphate dehydrogenase, GPD; malic enzyme, ME; pyruvate dehydrogenase complex, PDC; phosphoenolpyruvate carboxykinase, PCK; pyruvate carboxylase, PYC; pyruvate kinase, PYK. Multi-step reactions are presented by dashed arrows in related color. Black dashed arrows represent metabolite transfer. The Figure was created with BioRender.
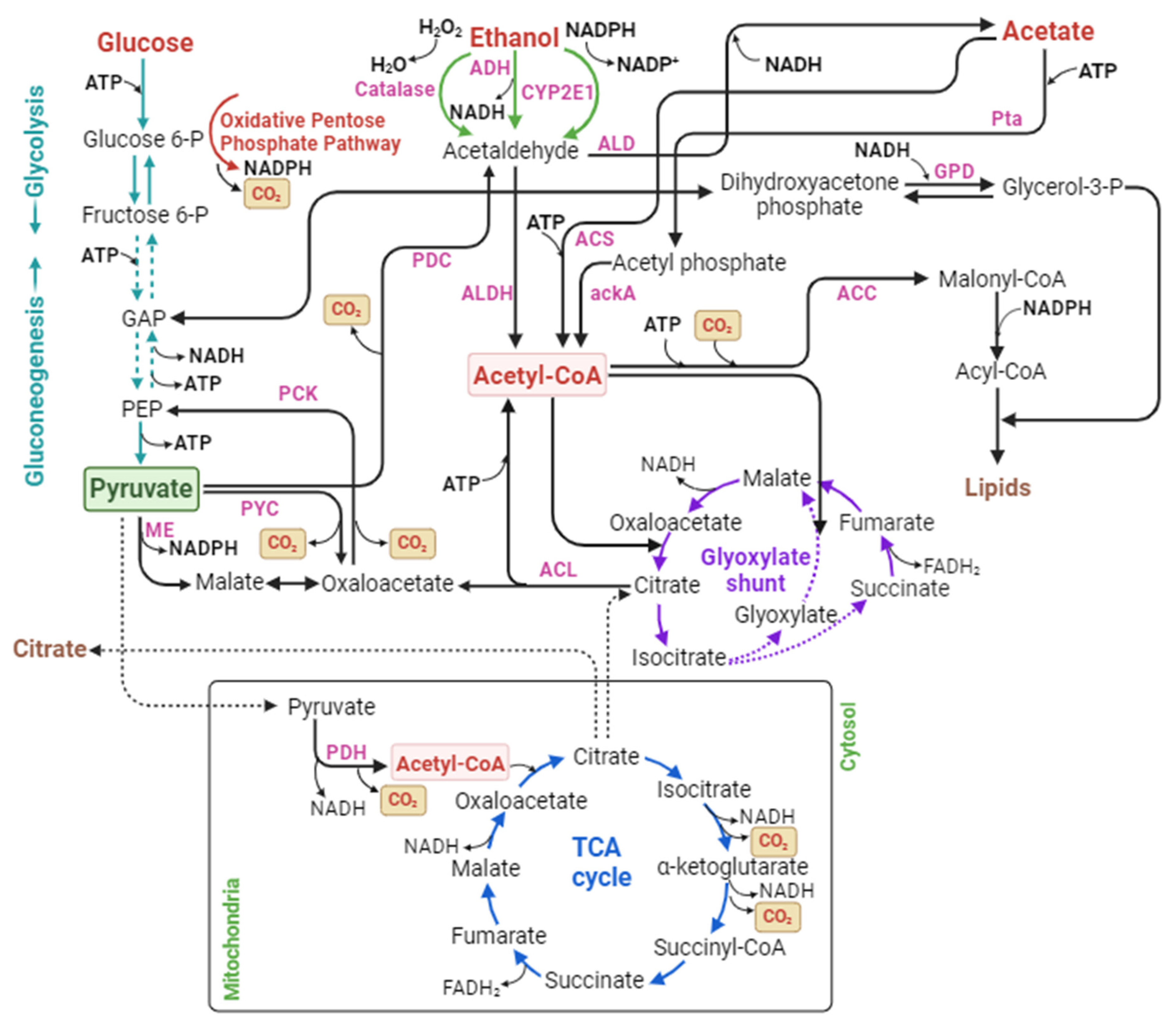
Figure 6.
A brief summary of various methods for using CO2 as feedstock for biomanufacturing. All these methods serve promising approach to fix CO2 more efficiently and obtain desired product by carbon-neutral or -negative biomanufacturing.
Figure 6.
A brief summary of various methods for using CO2 as feedstock for biomanufacturing. All these methods serve promising approach to fix CO2 more efficiently and obtain desired product by carbon-neutral or -negative biomanufacturing.
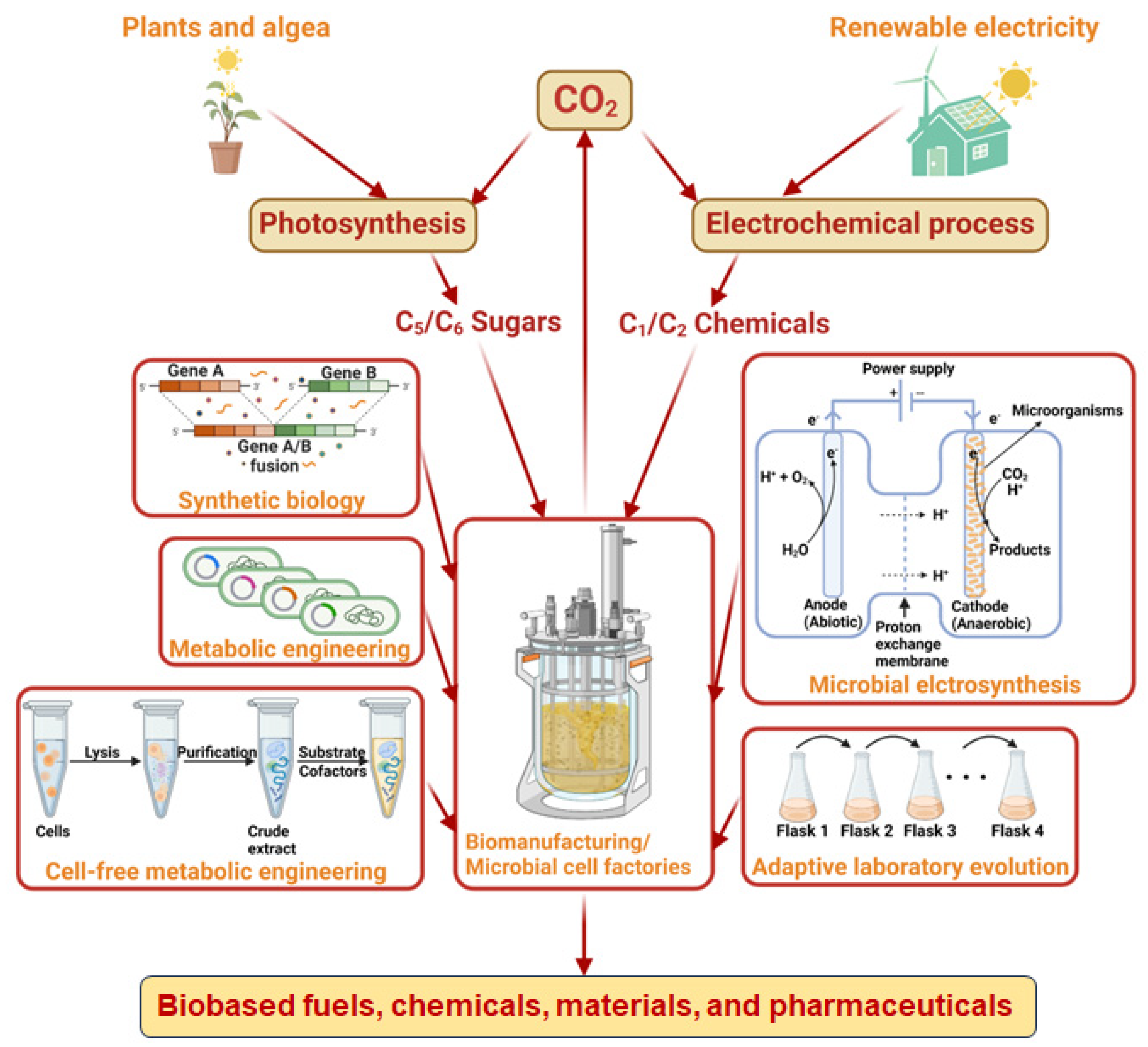
Figure 7.
A conceptual diagram for a novel biomanufacturing platform that can produce fuels and chemicals from C5/C6 sugars at zero or near zero life cycle carbon emissions via in-situ CO2 recycling.
Figure 7.
A conceptual diagram for a novel biomanufacturing platform that can produce fuels and chemicals from C5/C6 sugars at zero or near zero life cycle carbon emissions via in-situ CO2 recycling.

Table 1.
General strategies for biotechnological fixation of CO2.
| Methods | Major steps and overall reaction of CO2 fixation | |
|---|---|---|
| One-step/Direct CO2 fixation & conversion |
|
|
| Two-step CO2 fixation & conversion | Step 1 (electrochemical catalysis): CO2 + H2O + electricity → C1/C2 chemicals |
Step 2 (biomanufacturing): C1/C2 → biofuels and chemicals |
|
|
|
Table 2.
ATP balance for the most common C1 and C2 chemicals, calculated regarding Figure 4 and Figure 5.
| Substrate | Key enzyme | Major biochemical reactions | Eq. ATP/ substrate | ||
|---|---|---|---|---|---|
| Reaction 1 | Reaction 2 | Reaction 3 | |||
| CO2 | N/A | CO2 + RuBP + 2NADPH + 2ADP + 2Pi → 2GAP + 2NADP + 2ATP | N/A | N/A | 3.3 |
| CO | N/A | CO + 5-Methyl-THF → AcCoA | N/A | N/A | 6.0 |
| Methane (CH4) | N/A | CH4 + O2 + NADH → HCHO + NAD | HCHO + Xu5P + ATP → 2GAP + ADP + Pi | N/A | 8.7 |
| Methanol (CH3OH or MeOH) | RuMp | MeOH + NADH → HCHO + NAD | HCHO + Ru5P + ATP → 2GAP + ADP + Pi | 2GAP + 8ADP + Pi + 8NAD → 2AcCoA + 8ATP + 8NADH + 2CO2 | 8.7 |
| XuMp | HCHO + Xu5P + ATP → 2GAP + ADP + Pi | N/A | 9.2 | ||
| Serine | HCHO + FAD + 3ATP + 2NADPH + 2NADH + Glycine + CO2 → AcCoA + FADH2 + 3ADP + 2NADP + 2NAD + Glyoxylate | N/A | -6.0 | ||
| Formate (HCOOH) | CBB | HCOOH + NAD → CO2 + NADH | RuBP + CO2 + 2NADPH + 2ADP + Pi → 2GAP + 2NADP + 2ATP | 2GAP + 8ADP + Pi + 8NAD → 2AcCoA + 8ATP + 8NADH + 2CO2 | 9.2 |
| HCOOH + ATP → 10-Formyl-THF + ADP + Pi | 10-Formyl-THF + NADPH + NADH + CO2 + FADH2 → AcCoA + NADP + NAD + FAD | N/A | 5.0 | ||
| Acetate (CH3COOH or OAc) | Pta/ackA | OAc + ATP → ADP + AcP | AcP + CoA → AcCoA + pi | N/A | 11 |
| ACS | OAc + ATP + CoA → AcCoA + AMP + PPi | N/A | N/A | 11 | |
| Ethanol (CH3CH2OHor EtOH) | CYP2E1 | EtOH + NADPH + H + O2 → MeCHO + NADP + H2O | MeCHO + NADH → NAD + OAc | OAc + ATP → ADP + AcP; AcP + CoA → AcCoA + pi | 5 |
| OAc + ATP + CoA → AcCoA + AMP + PPi | 5 | ||||
| ADH | EtOH + NAD → MeCHO + NADH | OAc + ATP → ADP + AcP; AcP + CoA→ AcCoA + pi | 11 | ||
| OAc + ATP + CoA → AcCoA + AMP + Ppi | 11 | ||||
| Catalase | EtOH + H2O2 → MeCHO + H2O | OAc + ATP → ADP + AcP; AcP + CoA → AcCoA + pi | 8 | ||
| OAc + ATP + CoA → AcCoA + AMP + PPi | 8 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



