
Submitted:
14 October 2023
Posted:
18 October 2023
You are already at the latest version
Abstract
(1) Background: Some evidence suggests that cannabidiol (CBD) has potential to help alleviate HIV symptoms due to its antioxidant and anti-inflammatory properties. Here we examined acute CBD effects on various behaviors and the endocannabinoid system in HIV Tat transgenic mice. (2) Methods: Tat transgenic mice (female/male) were injected with CBD (3,10,30 mg/kg) and assessed for antinociception, activity, coordination, anxiety, and recognition memory. Brains were taken to quantify endocannabinoids and related lipids. Additionally, CBD and metabolite 7-hydroxy-CBD were quantified in the plasma and cortex. (3) Results: Tat decreased striatal-related nociception and locomotion, with some sex-dependent effects on coordination. CBD had no effect on nociception and coordination, but increased locomotor activity. For anxiety, differential CBD effects were noted for sex, with decreasing anxiety-like behavior in males only. In the striatum and spinal cord, male sex was associated with lower 2-arachidonoylglycerol and with elevated concentrations of its proinflammatory metabolite arachidonic acid. CBD metabolism was altered by sex and Tat. (4) Conclusion: Findings indicate that acute CBD effects are not altered by HIV Tat, and CBD has minimal effects on behavior and the endocannabinoid system. Interestingly, sex-dependent alterations were noted for endocannabinoids and related lipids, which may be of relevance in view of potential CBD-based treatment options for people living with HIV.
Keywords:
cannabidiol
; neuroHIV
; Tat transgenic mice
; antinociception
; anxiety
; 2-arachidonoylglycerol
; arachidonic acid
1. Introduction
In 2022, 39 million people were living with human immunodeficiency virus (HIV) worldwide, out of which ~75% have access to antiretroviral therapy (ART) [1]. While combination ART (cART) has significantly increased survival rates and quality of life [2,3,4], 30-50% of cART-treated people living with HIV (PLWH) exhibit some degree of cognitive deficits, known as HIV-associated neurocognitive disorders (HAND) [5,6,7,8]. Typical HAND symptoms include deficits in attention, learning and memory, and slowed motor control [5,9,10,11], which can lead to difficulty with activities of daily living [5,12,13,14] as well as unemployment [15,16,17]. Additional central nervous system (CNS)-related symptoms reported by PLWH include neuropathic pain [18] and heightened anxiety [19,20].
HAND and HIV-related CNS symptoms persist in cART-treated PLWH as a result of chronic immune activation, neuroinflammation, and low-level expression of viral proteins [8,21]. One such viral protein is HIV transactivator of transcription (Tat), which is observed rapidly in the CNS after initial HIV infection [22,23], and is particularly elevated in the brains of PLWH with HAND [24,25]. Multiple in vitro and in vivo studies have demonstrated Tat’s role in neurotoxicity in HAND, even in the absence of any other HIV viral proteins [26,27,28,29,30,31,32,33]. HIV Tat is secreted by infected cells [22,34,35,36,37] and can interact with cell surface receptors of uninfected neuronal and glial cells [38,39,40], thus directly damaging neuronal structure and function [35,41,42,43], as well as indirectly by promoting proinflammatory signaling via glial activation [44,45,46]. Further, using the HIV Tat transgenic mouse model, various studies have demonstrated deficits in learning and memory, motivation, and motor activity [28,29,33,47], as well as Tat’s contribution to HIV-related neuropathic pain [31,48,49,50] and anxiety [51,52,53,54,55,56], all of which recapitulate the pathological and behavioral abnormalities associated with HAND.
The endocannabinoid system is a potential source of neuroprotection and anti-inflammatory action [57,58], and has been reported to alleviate HIV-related symptoms, such as pain, anxiety, stress, and loss of appetite [59,60,61,62]. The use of cannabis is highly prevalent in PLWH, with cannabis use being 2-3 times greater than use among the general US population [63,64,65,66]. Cannabis contains exogenous modulators of the endocannabinoid system, including the major psychoactive cannabinoid, Δ9-tetrahydrocannabinol (THC), and the major non-psychoactive component, cannabidiol (CBD). CBD has antioxidant, anti-inflammatory, and neuroprotective properties [67,68,69], and several studies have reported its potential therapeutic role in attenuating memory deficits in various disease conditions [70,71,72,73]. CBD is known to interact with multiple targets within the CNS to exert its effects [74]. CBD has low binding affinity for cannabinoid type 1 and 2 receptors (CB1R and CB2R, respectively) [75], even though CBD can still exert effects mediated by these receptors [76,77,78,79,80,81]. A variety of other non-cannabinoid receptors can be targeted by CBD, including transient receptor potential vanilloid (TRPV) channels [82], serotonin (5-HT1A) receptors [83,84], peroxisome proliferator-activated receptor gamma (PPARγ) [85,86,87], and the cannabinoid-related receptor GPR55 [88,89]. For example, improvement of object recognition memory deficits in antipsychotic mice by CBD was associated with decreases in proinflammatory cytokine levels in the hippocampus, both of which were mediated by PPARγ receptors [87]. On the other hand, anxiolytic and neuroprotective effects of CBD in chronically stressed mice were reported to be related to CB1R and CB2R activation [80,90] due to upregulation of hippocampal endocannabinoid N-arachidonoylethanolamine (AEA) concentrations via fatty acid amide hydrolase (FAAH) inhibition [90,91,92].
In the present study we used HIV-1 Tat transgenic mice to investigate the acute effects of CBD (3, 10, 30 mg/kg) in neuroHIV on multiple behavioral outcomes, including pain sensitivity, motor activity, motor coordination, anxiety, and object recognition memory. Further, acute CBD effects on the endocannabinoid and endocannabinoid-like (“paracannabinoid”) systems in the CNS, including the prefrontal cortex, striatum, cerebellum, and spinal cord, were assessed by quantifying concentrations of AEA, 2-arachidonoylglycerol (2-AG), N-palmitoylethanolamide (PEA), N-oleoylethanolamide (OEA), and their proinflammatory metabolite arachidonic acid (AA) via ultraperformance liquid chromatography/tandem mass spectrometry (UPLC-MS/MS). CBD metabolism was examined via UPLC-MS/MS by quantifying concentrations of CBD and its main oxidative metabolite 7-hydroxy-CBD concentrations (COOH-CBD) in the plasma and cortex of acute CBD-exposed Tat transgenic mice.
2. Results
2.1. Spontaneous heat-evoked nociception
The tail-flick and hot-plate assays were conducted 75 min after drug injections to evaluate effects of acute CBD (3, 10, and 30 mg/kg) exposure on heat-evoked pain-like behaviors in the Tat transgenic mouse model (Figure 1A,B). The tail-flick test was used to assess spinal-related spontaneous nociception (Figure 1A). A three-way ANOVA demonstrated no significant effects or interactions.
The hot-plate test was used to assess supraspinal-related spontaneous nociception (Figure 1B). A three-way ANOVA demonstrated a significant main effect of genotype, F(1, 35) = 4.7, p = 0.037, with Tat(+) mice showing higher latencies to withdrawal or lick their paw (6.7 ± 0.46, n = 19) compared to Tat(−) mice (5.5 ± 0.29, n = 20). No other significant effects or interactions were noted.
Overall, spontaneous heat-evoked nociception was not affected by CBD doses, but the supraspinal-related hot-plate test demonstrated decreased spontaneous nociception for Tat(+) mice compared to Tat(−) mice, independent of acute CBD doses.
2.2. Locomotor activity and rotarod coordination
To understand the effects of acute CBD (3, 10, and 30 mg/kg) exposure on motor function, we assessed locomotor activity and rotarod performance 60 min and 90 min after injections, respectively (Figure 1C,D). The locomotor activity task was conducted to assess effects of acute CBD in Tat transgenic mice on motor activity (Figure 1C). A three-way ANOVA demonstrated a significant main effect of CBD doses, F(3, 105) = 3.3, p = 0.022, with increased locomotion for all three CBD doses compared to vehicle (Vehicle, 75.6 ± 5.53, n = 39; 3 mg/kg CBD, 83.5 ± 5.17, n = 39; 10 mg/kg CBD, 89.2 ± 4.48, n = 39; 30 mg/kg CBD, 88.2 ± 4.78, n = 39), even though Bonferroni’s post hoc comparisons demonstrated no significance between doses. Further, a significant main effect of genotype was noted, F(1, 35) = 5.0, p = 0.031, with Tat(+) mice showing lower locomotor activity compared to Tat(−) mice for both sexes. No other significant effects were noted.
The rotarod task was conducted to investigate effects of acute CBD exposure in Tat transgenic mice on motor coordination and function (Figure 1D). A three-way ANOVA demonstrated a significant sex x genotype interaction, F(1, 35) = 4.9, p = 0.034, with Tat(+) male mice showing lower rotarod performance compared to Tat(−) males, whereas this was not seen in females. Bonferroni’s post hoc comparisons revealed no significant differences between any of the groups.
Overall, an increase in locomotor activity was noted for CBD doses without affecting rotarod performance. Further, Tat expression decreased locomotor activity in female and male mice and motor coordination appeared to be differentially altered by Tat expression depending on sex.
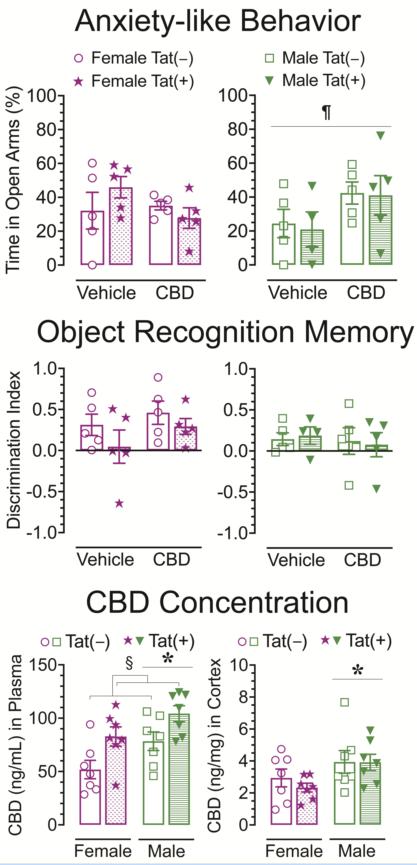
2.3. Anxiety-like behavior
The elevated plus maze task was conducted 60 min after injections to evaluate the effects of acute CBD (0 and 30 mg/kg) exposure in Tat transgenic mice on anxiety-like behavior (Figure 2). The dependent measures included percent time spent in open arms and number of pokes into open arms. For percent time spent in open arms (Figure 2A) a three-way ANOVA revealed a significant sex x drug interaction, F(1, 31) = 5.2, p = 0.029, in which 30 mg/kg CBD decreased anxiety in males but not female mice. Bonferroni’s post hoc comparisons revealed no significant differences between any of the groups.
For number of pokes into open arms (Figure 2A), no effects or interactions were found to be significant.
Overall, CBD appears to have differential effects in male and female mice, with decreasing anxiety-like behavior in male mice but not in female mice.
2.4. Novel object recognition
The NOR task was conducted 60 min after injections to evaluate the effects of acute CBD (0 and 30 mg/kg) exposure in Tat transgenic mice on novel object recognition memory (Figure 3). The dependent measures included total exploration time of the novel and familiar objects and time spent exploring the novel object over the familiar object (discrimination index). For total exploration time (Figure 3A), a three-way ANOVA demonstrated no significant effects.
For object recognition memory, the discrimination index was used, with complete preference for the novel object equal to 1, no preference equal to 0, and complete preference for the familiar object equal to -1 (Figure 3B). A three-way ANOVA demonstrated no significant effects.
Overall, total object exploration time and novel object recognition memory was similar across groups.
2.5. CNS levels of endocannabinoids and related lipids
To assess the impact of acute CBD (0 and 30 mg/kg) exposure on the endogenous cannabinoid system, changes in levels of 2-AG, AEA, PEA, OEA, and AA were assessed 90 min after injections in various CNS regions of Tat transgenic female and male mice (n = 4–5 per group), including the prefrontal cortex, striatum, cerebellum, and spinal cord (Supplemental Table S2, Figure 4). Lipid molecule concentrations (nmol/g) differed significantly between CNS region for AEA, PEA, OEA, and AA, but not for 2-AG. AEA, F(3, 114) = 66.0, p < 0.001, demonstrated differences in expression levels between all CNS regions (p’s < 0.001), except between the striatum and cerebellum, with highest AEA levels found in the prefrontal cortex, followed by the striatum and cerebellum, and the lowest AEA levels being expressed in the spinal cord. The same differences were seen for PEA, F(3, 114) = 162.9, p < 0.001, and OEA, F(3, 114) = 172.2, p < 0.001. AA, F(3, 114) = 122.2, p < 0.001, also showed differences in expression levels between all CNS regions (p’s < 0.001), except between the prefrontal cortex and striatum, with highest AA levels found in the prefrontal cortex and striatum, followed by the cerebellum, and the lowest AA levels being expressed in the spinal cord (Supplemental Table S2, Figure 4). To assess treatment effects, a MANOVA was conducted for each lipid molecule with drug, sex, and genotype as between-subjects factors. No effects or interactions were noted for acute CBD administration on any measure, indicating that acute CBD did not alter the endocannabinoid system and related lipid molecules. The most prominent findings were noted for AA levels with some minor effects for 2-AG (Figure 4), PEA and OEA (Supplemental Table S2), and no effects for AEA (Figure 4).
For 2-AG (Figure 4A), significant sex effects were noted for the striatum, F(1, 31) = 18.5, p < 0.001, and spinal cord, F(1, 31) = 12.7, p = 0.001, with females demonstrating higher 2-AG levels compared to male mice in both CNS regions. No other significant effects or interactions were noted.
For AEA (Figure 4B), no significant effects or interactions were noted.
For AA (Figure 4C), a significant sex effect was noted for the striatum, F(1, 31) = 50.8, p < 0.001, the cerebellum, F(1, 31) = 56.1, p < 0.001, and the spinal cord, F(1, 31) = 31.5, p < 0.001, with males demonstrating higher AA levels compared to female mice for all three CNS regions. Further, for the striatum, a significant sex x genotype interaction was noted, F(1, 31) = 7.8, p = 0.009, with Tat(+) male mice showing higher AA levels compared to Tat(−) males (p = 0.020), which was not noted for females.
Results for PEA and OEA mirrored a few of the findings for AA, specifically the noted sex effects in the striatum and cerebellum, with higher PEA and OEA levels for males compared to females; additionally a genotype effect was demonstrated for PEA and OEA in the spinal cord with lower levels for Tat(+) mice compared to Tat(−) mice (Supplemental Table S2).
Overall, acute CBD administration did not alter endocannabinoids or related lipid molecules in any CNS region. The most prominent effects were noted for AA expression levels with males showing higher AA levels compared to females in all CNS regions except the prefrontal cortex. Whereas no significant effects were noted for AEA, 2-AG levels were higher for females compared to male mice in the striatum and spinal cord.
2.6. Concentration of CBD and its metabolite CBD-7-COOH in plasma and cortex
Concentration of CBD and CBD-7-COOH were assessed 60 min after acute CBD (30 mg/kg) exposure using a different cohort of animals. Tat transgenic female and male mice (n = 7 per group) were subcutaneously injected with 30 mg/kg CBD and sacrificed 60 min later, around the time when behavior was assessed (Figure 5). CBD concentration and its metabolite CBD-7-COOH were detected in all plasma (ng/mL, Figure 5A,B) and cortex (ng/mg, Figure 5C,D) samples. A two-way ANOVA was conducted with sex and genotype as between-subjects factors for plasma and cortex samples separately.
For plasma samples (ng/mL) CBD concentrations (Figure 5A) revealed a significant main effect of sex, F(1, 24) = 8.01, p = 0.009, with higher concentrations in males compared to females. Further, a significant main effect of genotype, F(1, 24) = 11.25, p = 0.003, demonstrated higher CBD concentrations in Tat(+) mice compared to Tat(−) mice. For CBD-7-COOH plasma concentration (Figure 5B), a significant main effect of sex was noted, F(1, 24) = 13.00, p = 0.001, with lower concentrations in males compared to females. Further, a significant main effect of genotype, F(1, 24) = 4.48, p = 0.045, demonstrated higher CBD-7-COOH concentration in Tat(+) mice compared to Tat(−) mice.
For cortex samples (ng/mg), CBD concentration (Figure 5C) revealed a significant main effect of sex, F(1, 24) = 5.62, p = 0.026, with higher concentrations in males compared to females. For CBD-7-COOH concentrations (Figure 5D), a significant main effect of genotype was noted, F(1, 24) = 6.04, p = 0.022, with higher concentration in Tat(+) mice compared to Tat(−) mice.
Overall, CBD and CBD-7-COOH concentrations were higher in plasma compared to cortex concentrations after 60 min acute 30 mg/kg CBD administration. Males demonstrated higher CBD concentrations in the plasma and cortex compared to females, whereas the reverse was true in the plasma for CBD-7-COOH concentrations, with lower CBD-7-COOH concentrations for males compared to females. Interestingly, a genotype effect was noted for CBD and CBD-7-COOH with Tat(+) mice displaying higher concentrations compared to Tat(−) in plasma and/or cortex.
3. Discussion
The present study investigated acute effects of CBD in female and male Tat transgenic mice on various behavioral measures and the endocannabinoid system. HIV Tat induction decreased locomotor activity and lowered pain sensitivity without affecting levels of endocannabinoids and related lipids. Acute CBD overall produced little effects on behavior, only altering locomotor activity and anxiety-like behavior, and had no effects on the endocannabinoid system. Sex effects were specifically noted for the endocannabinoid system and related lipids, including elevated 2-AG levels for females in the striatum and spinal cord, and upregulated AA levels for males in the striatum, cerebellum, and spinal cord. Even though Tat expression did not alter acute CBD effects on any behavioral and endocannabinoid measure, CBD metabolism was significantly affected by sex and Tat induction.
Chronic pain is commonly reported by PLWH [102,103] and significantly interferes with daily function [104]. Preclinical studies using the Tat transgenic mouse model have reported mechanical allodynia, thermal hyperalgesia, and damage to nerve fibers due to Tat expression [31,49,50], but others have found either no effects [105] or decreased pain sensitivity [48]. The current study supports the finding that Tat induction attenuates pain sensitivity but only at the supraspinal level without affecting the spinal-related tail flick task. Based on the variable reports, it is clear that Tat’s effects on nociception are complex and highly dose- and time-dependent [48,106], as well as assay-dependent (i.e., mechanical, thermal, or cold pain sensitivity) [106]. A previous study reported hyposensitivity to mechanical pain in Tat(+) mice following Tat induction for 3 weeks [48], similar to our 1 month Tat induction paradigm. However, hypersensitivity to mechanical pain was reported in transgenic mice when Tat was expressed for more than 1 month [31,106]. It has been suggested that Tat’s neurotoxic effects on nociceptive neurons [31,49] potentially induce nerve dysfunction during an initial period, resulting in diminished pain signaling, however, this later reverses into pain hypersensitivity due to prolonged nerve damage [48]. The finding of reduced pain sensitivity for the supraspinal hot plate test but not the spinal-related tail flick test may be explained by a previous study that found higher Tat mRNA expression in the striatum compared to the spinal cord [105]. For acute CBD exposure, no effects on pain sensitivity in Tat transgenic mice were found at any dose given (3, 10, 30 mg/kg), even though other studies demonstrated attenuation of pain sensitivity by acute CBD treatment of similar doses in various other preclinical rodent models [107,108,109]. To our knowledge, cannabidiol’s effects on pain have not been studied in PLWH and/or neuroHIV mouse models, including the Tat transgenic mouse model. However, it has been reported that cannabis use in PLWH improves nerve pain [59], shrinks the area of painfully sensitive skin [110], and can relief neuropathic pain [111]. Further, CBD has been shown to be necessary for THC-related antinociception in some human trials [112]. Thus, to provide more insights into possible mechanisms underlying CBD’s effects on pain sensitivity, future studies should focus on chronic CBD use as well as investigate the implications of various CBD:THC ratios among PLWH and in preclinical neuroHIV mouse models, including the Tat transgenic mice.
Besides chronic pain as a common comorbidity among PLWH, individuals also exhibit motor deficits, including impaired gait, multitasking, motor strength, and motor coordination, all correlated with HAND status [113,114]. As a result of these motor difficulties, physical activity often suffers, and a majority of PLWH exhibit inadequate levels of daily physical activity, even lower than most other chronic disease populations [115]. These lower levels of activity have been associated with depression, physical pain, and worse cardio-respiratory fitness [115,116]. In accordance with this, preclinical neuroHIV rodent models, including Tat transgenic mice, have been found to exhibit lower levels of locomotor activity [54,117,118]. This finding is supported by the current study that demonstrated lower locomotor activity for Tat(+) mice compared to control Tat(−) mice. Similarly, motor coordination on the rotarod was decreased in Tat(+) male mice compared to control Tat(−) males, but this was not seen in females. The potential heightened vulnerability of males to motor impairments has been previously suggested to be due to substantial synaptic alterations and glia activation detected in the striatum of male mice after chronic HIV-1 Tat exposure [119], even though other studies demonstrate selective vulnerabilities for Tat(+) females in other tasks, such as inhibitory control [120]. For CBD, the present study found that all three CBD doses (3, 10, 30 mg/kg) increased activity independent of Tat expression without affecting rotarod performance. Multiple preclinical studies have reported the lack of CBD effects on locomotor activity in non-diseased mice [121,122] and disease rodent models [123,124,125], including motor performance in the rotarod test [122,126], which is supported by a recent review on effects of cannabidiol on locomotor activity [127]. Even though we did not see differential effects of CBD based on Tat expression, a previous study reported reduction of hypolocomotion by chronic treatment of CBD via 5-HT1A receptors, and thus improvement in motor dysfunction caused by hepatic encephalopathy [128], which demonstrates CBD can increase locomotor activity as shown in the present study. Nevertheless, further research is necessary to better understand CBD’s effects on locomotion in the context of neuroHIV, especially in the light of rodent models that include multiple HIV proteins and not just a single viral protein, such as Tat.
Another noteworthy aspect of CBD is its known anxiolytic effects [70,123,129,130,131], which were partially present in the current study as differential CBD effects were noted for sex, with decreasing anxiety-like behavior in male mice but not in female mice. Sex differences in the effectiveness of CBD as a potential treatment for anxiety in a clinical setting still have to be explored [132], but preclinical studies have shown that anxiolytic effects can be achieved by acute CBD administration in both sexes [131,133]. However, the effectiveness of CBD appears to be sex-dependent as it has been shown that the CBD dose required to produce a decrease in anxiety-like behavior was lower in females compared to male mice [131]. Interestingly, a recent study demonstrated that the responsiveness of female rats to acute CBD depended on the stage of the estrous cycle, with female rats being responsive only in the late diestrus phase at a 10-fold lower dose than males, but females were unresponsive to acute CBD in the proestrus phase [133]. Based on these findings, the majority of females in the current study may have been in the proestrus phase when the elevated plus maze was conducted, as acute CBD treatment (30 mg/kg) did not affect their anxiety-like behavior. Alternatively, a bell-shaped dose-response relationship has been reported for CBD effects on anxiety-like behavior [133,134], and could suggest that females in proestrus may be responsive to CBD at lower doses (i.e., < 30 mg/kg). In the future, it would be interesting to monitor estrous cycle in female mice as well as use a range of CBD doses to get a better understanding of CBD’s effects on anxiety-like behavior in Tat transgenic mice. Additionally, it should be pointed out, that increases in locomotor activity by CBD could have influenced the induced anxiolytic effects in male mice due to increased exploratory behavior and thus, spending more time in the open arms, even though this was not seen in females. Nevertheless, further investigation should use additional measures of anxiety, such as approach behaviors (i.e., stretch attenuated postures—more anxious, and head-dipping for direct exploration—less anxious) [135] that are less confounded by locomotor activity.
An unexpected finding was the lack of Tat and CBD effects on the memory-related object recognition task. Previous research has shown that tests of learning and memory typically show memory deficits in mice exposed to Tat [27,136], and CBD administration has been shown to attenuate disease-associated memory deficits in a variety of contexts [70,71,72,73,137,138,139,140]. The lack of CBD effects may be due to the fact that Tat(+) mice in the current study did not show deficits in the object recognition task. Another caveat was that control mice did not successfully differentiate the novel object from the familiar object, which was noted specifically in Tat(−) male mice. This may be due to similarity between the two objects used. Such similarity can be positive, as it reduces the likelihood of mice having an innate preference for one object, but some mice may have failed to physically differentiate the novel object. Follow-up investigation could use objects that differ significantly in odor, size, color/brightness, or texture to ensure that mice can differentiate the two.
For the endocannabinoids and related lipids, the current study found no acute CBD effects and only a Tat effect for AA in which Tat expression upregulated AA levels in the striatum of male mice. Whereas previous preclinical studies have reported the lack of acute CBD at different doses to affect brain endocannabinoids and related lipids (despite high CBD concentrations in blood and whole brain [141]), multiple studies demonstrated changes in endocannabinoid levels and related lipids following Tat exposure [41,142,143]. Alterations of the endocannabinoid system in postmortem tissue of PLWH has been demonstrated previously with reported changes in CB1R and CB2R expression in the frontal lobe [144,145]. The upregulation of AA levels in the striatum of Tat(+) male mice is not surprising as AA plays an important role in pro- and anti-inflammatory responses [146,147,148], and chronic Tat and HIV-1 infection in the CNS increases inflammatory processes via upregulation of inflammatory mediators and glia activation [8,149,150]. Interestingly, the current study found sex-specific effects in which male mice demonstrated higher AA levels compared to females in all CNS regions except the prefrontal cortex, and females demonstrated higher 2-AG levels in the striatum and spinal cord compared to males. More research is needed to understand how sex alters the endocannabinoid system in PLWH and how it relates to neuroinflammation and HAND, but vulnerabilities to HAND symptoms [151,152,153,154,155] and immune activation [156,157] have been reported to be higher in women compared to men. Whereas a recent study did not find sex differences in various metabolites of the AA cascade of gp120 transgenic mice, transcriptomic analysis revealed sexual dimorphism of AA pathway-related genes, and females appear to be more responsive to the lipoxygenase pathway compared to males [158]. Of note is the finding that females demonstrated higher 2-AG levels in the striatum and spinal cord that in combination with the upregulated AA levels in male mice of the same CNS regions points to potential sex-dependent alterations in endocannabinoid degradation, which has been shown previously [159].
Even though we did not find differential effects of CBD based on Tat expression, CBD metabolite analyses demonstrated that CBD and CBD-7-COOH were differently metabolized by Tat(−) and Tat(+) mice and/or sex. The higher CBD concentration in plasma and cortex for males compared to females, but lower plasma concentration for CBD-7-COOH, suggests that males metabolize acute CBD more slowly. Two recent clinical studies that either administered CBD for 7 days or cannabis acutely did not find concentration differences for CBD and/or CBD-7-COOH in plasma of healthy individuals [160,161]. However, the 7-day oral CBD administration study showed higher accumulation of CBD metabolites in plasma of females over time, which also is supported in a 12-week clinical CBD study [162], suggesting sex differences in CBD metabolism or elimination [161]. In contrast, not much is known about how cannabidiol is metabolized in PLWH or neuroHIV mouse models. The higher drug concentrations in Tat(+) females and males, found in plasma for both CBD and CBD-7-COOH, and for cortex CBD-7-COOH might indicate overall alteration in CBD metabolism. Even though future studies need to investigate this in more detail it suggests that HIV Tat status influences CBD metabolism, which then needs to be considered when thinking about CBD as a potential therapeutic treatment for PLWH.
4. Materials and Methods
4.1. Animals
The present study utilized HIV-1 IIIB Tat1−86 transgenic female and male mice that were developed on a hybrid C57BL/6J background [93,94]. HIV-Tat is brain-specific and expressed in astrocytes using a glial fibrillary acidic protein promoter under the control of a reverse tetracycline transactivator (GFAP-rtTA). Transgenic mice expressing the TRE-tat gene were identified (Transnetyx, Inc., Cordova, TN) as transgenic Tat(+) mice and littermates lacking the tat transgenic gene were used as control Tat(−) mice. To induce Tat expression in Tat(+) mice, animals were fed an ad libitum doxycycline (DOX) chow (6 mg/g; product TD.09282, Envigo, NJ, USA) one month prior to and throughout the study duration. To control for off-target effects, the same DOX-chow was fed to control Tat(−) mice. Animals were kept in group housing, with 3-5 mice per cage, under a reversed 12-h light/dark cycle (with lights off at 6:00 AM) and had free access to water and chow.
Two cohorts of mice were used. First, behavioral and endocannabinoid experiments were conducted on a total of 38 Tat transgenic mice (20 female, 19 male; ~ 9-10 months of age). The second group consisted of 28 Tat transgenic mice (14 female, 14 male; ~ 5 months of age) and was used to confirm CBD and 7-hydroxy-CBD (CBD-7-COOH) concentrations following an acute 30 mg/kg CBD injection.
All research procedures were conducted in strict accordance with the guidelines outlined in the NIH Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23) and approved by the Institutional Animal Care and Use Committee (IACUC) at the University of North Carolina at Chapel Hill.
4.2. Drugs
Cannabidiol (CBD; #90080, Cayman Chemical, Ann Arbor, MI) was dissolved in a vehicle solution composed of a mixture of ethanol, Kolliphor® EL (Sigma-Aldrich, #C5135, St. Louis, MO), and saline (0.9% NaCL, Cytiva, #Z1376, Marlborough, MA), in a 1:1:18 ratio. Administration of vehicle and CBD doses (3, 10, or 30 mg/kg) was carried out through subcutaneous (s.c.) injections at a volume of 10 μL/g of body mass. In all experiments, drug treatments were randomized.
4.3. Experimental design
Mice underwent training in the rotarod task for four days (3 trials per day) and habituated to the testing chambers used for evaluating locomotor activity. Subsequently, CBD doses, including vehicle control (0, 3, 10, 30 mg/kg), were administered over the course of four consecutive days, following a Latin-square design. To ensure proper clearance of CBD, a minimum 48-h interval was maintained between each testing day. The assessment of locomotor activity, spontaneous nociception, and rotarod performance occurred 60 min, 75 min, and 90 min post-injection, respectively. Additionally, the novel object recognition (NOR) and elevated plus maze (EPM) tasks were conducted 4 and 6 days later, respectively, with either vehicle or 30 mg/kg CBD injections administered approximately 60 min prior to these tasks. Drug injections and behavioral measures were typically carried out between 9 am and 1 pm each day.
4.4. Behavioral procedure
4.4.1. Spontaneous heat-evoked nociception
The tail-flick and hot-plate assays were employed to evaluate spontaneous heat-evoked nociception with a focus on spinal and supraspinal pathways, respectively [95].
For the tail-flick test, the distal 1/3 of each mouse’s tail was immersed in a water bath (56 ± 0.1 °C, Thermo Scientific, Precision General-Purpose Water Bath, Model 181, Waltham, MA). The latency to remove the tail from the bath was recorded as an indicator of nociception. To prevent tissue damage, the maximum cut-off time was 10 s.
The hot-plate test was conducted immediately following the tail-flick assay. Mice were placed on the surface of a hot plate (55 ± 0.1 °C; IITC, Inc., MOD 39, Woodland Hills, CA) enclosed within a Plexiglas™ chamber (15 cm height, 10 cm diameter) to prevent escape. The mouse was removed from the hot plate as soon as it withdrew or licked a paw, or exhibited jumping behavior. The time spent on the hot plate was recorded as an indicator of nociception. To prevent tissue damage the maximum cut-off time was 15 s.
4.4.2. Locomotor activity
The locomotor activity task was employed to evaluate spontaneous locomotion in mice. Mice were placed within standard mouse experimental chambers (MED Associates, ENV-307W; 22 cm x 18 cm floor, Fairfax, VT), situated in sound and light attenuating compartments (MED Associates, ENV-022MD). Testing occurred in a darkened room, illuminated by red fluorescent lighting, with white noise from an air conditioning unit inside the room. The ambient temperature in the testing room was maintained at 22 °C with a humidity level of 30%. Locomotor activity was recorded over a duraction of 10 min (600 s) using night vision cameras (Amcrest, FullHD 1080P 2MP Dome, Houston, TX) installed on the ceiling of the soundproof chambers. Videos were recorded to a Security Recorder (Amcrest, AMDV8M16-H5). The chamber floor was divided into four equal sized and shaped quadrants, and the overall locomotor activity of mice was quantified by the number of times they moved their noses across the boundaries between these quadrants. Numerical data were generated from the 10 min video recordings by a team of trained experimenters blinded to treatment conditions. Each video was assessed by two experimenters independently, with an overall interrater reliability (Cronbach’s α) of α = 0.855. The locomotor activity data (# of crossings) represent the average score derived from the assessments of both experimenters.
4.4.3. Rotarod
The accelerating rotarod test was employed to evaluate motor function and coordination [96]. Mice were positioned on a rotarod apparatus (Harvard Apparatus, #76-0770, Holliston, MA) consisting of an elevated, rubber-coated rod (30 mm diameter, elevated 18 cm). The rod was divided into five sections (each 50 mm wide) allowing for testing of five mice simultaneously. Mice were placed on the rod and given 60 s to acclimate prior trial start. Subsequently, rods accelerated from 2 to 60 rpm over a period of 7 min (420 s). The duration (s) each animal stayed on the rotating rod without falling or looping was recorded.
4.4.4. Elevated plus maze (EPM)
The EPM task was employed to evaluate anxiety-like behavior [97]. The elevated, plus-shaped apparatus (San Diego Instruments, #7001-0316, San Diego, CA) consisted of two open arms and two closed arms (15 cm tall walls). Each arm was 30 cm long, 5 cm wide, and 38 cm above the ground. To ensure uniform illumination of all four arms, indirect lighting was employed. Mice were placed in the center of the apparatus and roaming activity was recorded for 10 min (600 s) with a video camera (GoPro, Hero 6 Black, v02.10, San Mateo, CA) mounted overhead. Numerical data were generated from the 10 min video recordings by a team of trained experimenters blinded to treatment conditions. The time spent in the open arms [presented as percent time spent in the open arms from total exploration time (600 s)] and number of nose pokes into the open arms served as indicators of anxiety-like behavior. Each video was assessed by two experimenters independently, with an overall interrater reliability of α = 0.976 for latency and α = 0.971 for number of pokes. For both dependent variables data represent the average score derived from the assessments of both experimenters.
4.4.5. Novel object recognition (NOR)
The NOR task was employed to evaluate object recognition memory which relies on mice’s natural tendency to explore novel stimuli [98,99]. The task was conducted within a hexagonal arena constructed of lightly textured high-density white polyethylene (50 cm wide, 23 cm tall, courtesy of G.F. League Co., Inc., Greenville, SC). The task encompassed three phases: habituation, training, and testing, as previously detailed [98]. Briefly, mice were individually habituated to the testing apparatus for 5 min and returned to their home cage. The training phase started 24-48 h later when mice were placed into the arena containing two identical objects (familiar objects) for 10 min before they were returned to their home cage. Two hours later, mice were injected (s.c.) with vehicle or CBD (30 mg/kg) and kept in their home cage for an additional 60 min before the testing phase started. For the testing phase, mice were placed again for 10 min in the same arena containing one familiar object and one novel object (randomized for each mouse). The duration of time that mice spent exploring the objects (familiar and novel) during the testing session was recorded with a video camera (GoPro Hero 6 Black, v02.10, San Mateo, CA) mounted overhead. Object exploration was defined as being in close proximity to the object and facing it at a distance closer than 1 cm. The glass objects utilized were equal in size (20 cm high, 7 cm wide) but differed in shape, with one being a square prism and the other a curved cylinder. The choice of object to serve as the familiar or novel one was randomized for each mouse. Numerical data were generated from the 10 min video recordings of the testing phase by a team of trained experimenters blinded to treatment conditions. Each video was assessed by two experimenters independently, with an overall interrater reliability of α = 0.949 for the novel object and α = 0.987 for the familiar object. For both dependent variables, data represent the average score derived from the assessments of both experimenters. Total exploration time (s) of both objects (familiar and novel) was calculated to determine object exploration time. A discrimination index (DI) was calculated according to the formula: DI = [t(novel) − t(familiar)]/[t(novel) + t(familiar)] for the testing phase [98,100]. A discrimination index of 0 indicates no preference, 1 indicates a complete preference for the novel object, and -1 indicates a complete preference for the familiar object.
4.5. Analysis of endocannabinoids and related lipids
Upon completion of behavioral experiments (after 48-72 hours), mice received an acute s.c. injection of either vehicle or CBD (30 mg/kg) and were sacrificed 60 min later by rapid decapitation following isoflurane-induced anesthesia. Four CNS regions from the right hemisphere (prefrontal cortex, striatum, cerebellum, and spinal cord) were dissected and snap-frozen in liquid nitrogen within 10 min of decapitation. Samples were stored at −80 °C until use. Using ultraperformance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS), the following lipids were quantified: the two main endocannabinoids N-arachidonoylethanolamine (AEA/anandamide) and 2-arachidonoylglycerol (2-AG), two related eicosanoids N-oleoyl ethanolamide (OEA) and N-palmitoyl ethanolamide (PEA), and arachidonic acid (AA). Details on the extraction and quantification of endocannabinoids and related lipids have been described previously [29,101] and are outlined in Supplemental Methods.
4.6. Analysis of CBD and CBD-7-COOH levels
A new cohort of drug-naive Tat transgenic female and male mice (n = 7 per sex) received an injection of an acute CBD dose (30 mg/kg, s.c.). This was done to confirm the concentrations of CBD and its metabolite 7-hydroxy-CBD (CBD-7-COOH) in both plasma and mouse cortex. 60 min after CBD injections mice were sacrificed under isoflurane-induced anesthesia, and samples were collected within 5 min. Plasma samples were prepared from fresh drawn blood obtained via cardiac puncture (Supplemental Methods). After cardiac puncture, cortex samples were dissected and snap-frozen in liquid nitrogen immediately upon decapitation. Samples were stored at −80 °C until further analysis.
Extraction and quantification of CBD and CBD-7-COOH levels in plasma and cortex samples were carried out as follows. A 100 µL aliquot of plasma was extracted using 600 µL of a mixture of 80:20 MeOH:H2O. Samples were shaken for 15 min and then centrifuged for 10 min at 20,000 rcf. The resulting supernatant was dried down and subsequently reconstituted with 100 µL of MeOH. Cortex tissue was extracted using 600 µL of 80:20 MeOH:H2O, sonicated for 15 min to facilitate tissue breakdown, and then centrifuged for 10 min at 20,000 rcf. Cortex extracts were dried down and reconstituted in 100 µL MeOH.
The analysis of these samples was performed using a Waters ACQUITY Ultra-Performance Liquid Chromatography (UPLC) system tandem to a Thermo Scientific TSQ Vantage triple quadrupole mass spectrometer. Separations were achieved using a Waters BEH C18 150 mm x 2.1 mm column; water with 0.1% formic acid was used for mobile phase A and acetonitrile with 0.1% formic acid was mobile phase B. The flow rate was set at 0.25 mL/min, and the initial starting conditions were 65% A. A linear decrease was performed to 45% A over 2 minutes followed by a hold for 1 minute. Another decrease to 20% A at 7 min was performed followed by another to 5% A at 8 minutes. A sharp decrease was performed from 5 to 0% A over 2 min with a curve of 3 (curve of 6 is linear). There was a hold at 100% B for 3 min followed by a re-equilibration step from 13.5 to 16 min. The injection volume was set at 10 µL. To monitor specific transitions for CBD and CBD-7-COOH, multiple reaction monitoring (MRM) was utilized (Supplemental Table S1).
4.7. Statistical analysis
All data are presented as mean ± the standard error of the mean (SEM). Data sets of body mass, nociception, and motor performance were analyzed by three-way mixed analysis of variances (ANOVAs) with drug dose (4 levels: vehicle, 3, 10, 30 mg/kg CBD) as a within-subjects factor and sex (2 levels: females, males) and genotype [2 levels: Tat(−) mice, Tat(+) mice] as between-subjects factors. Datasets on anxiety-related behavior and novel object recognition were analyzed by three-way ANOVAs with drug (2 levels: vehicle, 30 mg/kg CBD), sex (2 levels: females, males), and/or genotype [2 levels: Tat(−) mice, Tat(+) mice] as between-subjects factors. Significant interactions were followed by follow-up ANOVAs or group comparisons within each sex. Group comparisons were Bonferroni corrected for multiple comparisons with adjusted p values reported. Data sets for endocannabinoids and related lipid molecules were analyzed for each lipid molecule by one-way repeated ANOVAs with CNS regions [4 levels: prefrontal cortex, striatum, cerebellum, spinal cord] as a within-subjects factor or three-way multivariate analysis of variances (MANOVAs) with CNS regions as the multivariate variable and drug (2 levels: vehicle, 10 mg/kg THC), sex (2 levels: females, males), and genotype [2 levels: Tat(−) mice, Tat(+) mice] as between-subjects factors. Datasets from plasma and cortex Δ9-THC levels and its metabolites were analyzed by two-way ANOVAs with sex (2 levels: females, males) and genotype [2 levels: Tat(−) mice, Tat(+) mice] as between-subjects factors. All ANOVAs were followed by Bonferroni post hoc tests when appropriate. An alpha level of p ≤ 0.05 was considered significant for all statistical tests. SPSS Statistics 25 (IBM, Chicago, IL) and Prism GraphPad 8.0 (San Diego, CA) was used for data analysis and data graphing, respectively.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Result: Levels of PEA and OEA in CNS; Materials and Methods: Analysis of endocannabinoids and related lipids, Plasma sample preparation, Table S1: Multiple reaction monitoring transitions for standards. Quantifying transitions were used for quantitation, qualifying transitions were used to ensure proper identity, Table S2: Effects of sex and genotype on PEA and OEA levels (nmol/g) in four CNS regions. Acute CBD administration (0 and 30 mg/kg) did not display any effects or interactions for any of the four CNS regions.
Author Contributions
Conceptualization, B.J.Y.-S., W.J. and S.F.; Methodology, B.J.Y.-S., H.P.R., W.J. and S.F.; Validation, B.J.Y.-S., B.L.G., K.M.B., H.P.R., W.J. and S.F.; Formal Analysis, B.J.Y.-S., B.L.G., H.P.R., W.J. and S.F.; Investigation, B.J.Y.-S., B.L.G., K.M.B., H.P.R., C.J.H., M.R.P., J.L.P. and S.F.; Resources, W.J. and S.F.; Data Curation, B.J.Y.-S., B.L.G., K.M.B., H.P.R., C.J.H., E.D.W., M.R.P., J.L.P., W.J. and S.F.; Writing—Original Draft Preparation, B.J.Y.-S., B.L.G., H.P.R. and S.F.; Writing—Review & Editing, B.J.Y.-S., B.L.G., K.M.B., H.P.R., C.J.H., E.D.W., M.R.P., J.L.P., W.J. and S.F.; Visualization, B.J.Y.-S., B.L.G., H.P.R. and S.F.; Supervision, B.J.Y.-S., W.J. and S.F.; Project Administration, B.J.Y.-S., B.L.G., W.J. and S.F.; Funding Acquisition, J.L.P., W.J. and S.F.
Funding
This research was funded by the National Institute on Drug Abuse (NIDA), R01 DA055523 (SF & WJ), R21 DA041903 (SF), P30 DA033934 (JLP), and the Merit Review Award Number CX002422 (WJ) from the United States (U.S.) Department of Veterans Affairs Office of Research and Development (BLR&D) Service. We also thank the University of North Carolina’s Department of Chemistry Mass Spectrometry Core Laboratory, [especially Dr. Brandie M. Ehrmann], for their assistance with mass spectrometry analysis.
Institutional Review Board Statement
The animal study protocol was conducted in accordance with the ethical guidelines defined by the National Institutes of Health (NIH Publication No. 85- 23) and all procedures were approved by the University of North Carolina at Chapel Hill (UNC) Institutional Animal Care and Use Committee (IACUC) on 28 March 2023 (Protocol ID: 23.056.0, Web ID: 93813).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author (Barkha J. Yadav-Samudrala) upon request.
Acknowledgments
The authors would like to thank Julia Bondareva, Margaret Lostetter, Ava Davis, Caroline Clodfelter, Gabrielle Garner, Neha Palle, Pranathi Dandu, Rich Gray, Susan Lin, and William Lee for helping with scoring and quantifying videos taken for the various behavioral experiments. We thank the University of North Carolina’s Department of Chemistry Mass Spectrometry Core Laboratory, (especially Brandie M Ehrmann, Director), for their assistance with mass spectrometry analysis. The authors would also like to acknowledge the work of animal care technician DeVeda Eubanks for her role in maintaining the welfare of our animals through the studies.
Conflicts of Interest
The authors declare no conflict of interest.
References
- UNAIDS. Global HIV & AIDS statistics - Fact Sheet. Available online: https://home.liebertpub.com/publications/cannabis-and-cannabinoid-research/633/for-authors (accessed on October 13, 2023).
- Harrison, K.M.; Song, R.; Zhang, X. Life expectancy after HIV diagnosis based on national HIV surveillance data from 25 states, United States. J. Acquir. Immune. Defic. Syndr. 2010, 53, 124–130. [Google Scholar] [CrossRef]
- May, M.T.; Sterne, J.A.; Costagliola, D.; Sabin, C.A.; Phillips, A.N.; Justice, A.C.; Dabis, F.; Gill, J.; Lundgren, J.; Hogg, R.S.; et al. HIV treatment response and prognosis in Europe and North America in the first decade of highly active antiretroviral therapy: a collaborative analysis. Lancet 2006, 368, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Marcus, J.L.; Chao, C.R.; Leyden, W.A.; Xu, L.; Quesenberry, C.P., Jr.; Klein, D.B.; Towner, W.J.; Horberg, M.A.; Silverberg, M.J. Narrowing the Gap in Life Expectancy Between HIV-Infected and HIV-Uninfected Individuals With Access to Care. J. Acquir. Immune. Defic. Syndr. 2016, 73, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.; Langford, D.; Masliah, E. HIV and antiretroviral therapy in the brain: neuronal injury and repair. Nat. Rev. Neurosci. 2007, 8, 33–44. [Google Scholar] [CrossRef]
- Heaton, R.K.; Clifford, D.B.; Franklin, D.R., Jr.; Woods, S.P.; Ake, C.; Vaida, F.; Ellis, R.J.; Letendre, S.L.; Marcotte, T.D.; Atkinson, J.H.; et al. HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study. Neurology 2010, 75, 2087–2096. [Google Scholar] [CrossRef]
- Sacktor, N.; Skolasky, R.L.; Seaberg, E.; Munro, C.; Becker, J.T.; Martin, E.; Ragin, A.; Levine, A.; Miller, E. Prevalence of HIV-associated neurocognitive disorders in the Multicenter AIDS Cohort Study. Neurology 2016, 86, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Ajasin, D.; Eugenin, E.A. HIV-1 Tat: Role in Bystander Toxicity. Front. Cell. Infect. Microbiol. 2020, 10, 61. [Google Scholar] [CrossRef]
- Antinori, A.; Arendt, G.; Becker, J.T.; Brew, B.J.; Byrd, D.A.; Cherner, M.; Clifford, D.B.; Cinque, P.; Epstein, L.G.; Goodkin, K.; et al. Updated research nosology for HIV-associated neurocognitive disorders. Neurology 2007, 69, 1789–1799. [Google Scholar] [CrossRef]
- Thames, A.D.; Mahmood, Z.; Burggren, A.C.; Karimian, A.; Kuhn, T.P. Combined effects of HIV and marijuana use on neurocognitive functioning and immune status. AIDS care 2016, 28, 628–632. [Google Scholar] [CrossRef]
- Thames, A.D.; Kuhn, T.P.; Williamson, T.J.; Jones, J.D.; Mahmood, Z.; Hammond, A. Marijuana effects on changes in brain structure and cognitive function among HIV+ and HIV- adults. Drug Alcohol Depend. 2017, 170, 120–127. [Google Scholar] [CrossRef]
- Heaton, R.K.; Grant, I.; Butters, N.; White, D.A.; Kirson, D.; Atkinson, J.H.; McCutchan, J.A.; Taylor, M.J.; Kelly, M.D.; Ellis, R.J.; et al. The HNRC 500--neuropsychology of HIV infection at different disease stages. HIV Neurobehavioral Research Center. J. Int. Neuropsychol. Soc. 1995, 1, 231–251. [Google Scholar] [CrossRef] [PubMed]
- Heaton, R.K.; Marcotte, T.D.; Mindt, M.R.; Sadek, J.; Moore, D.J.; Bentley, H.; McCutchan, J.A.; Reicks, C.; Grant, I.; Group, H. The impact of HIV-associated neuropsychological impairment on everyday functioning. J. Int. Neuropsychol. Soc. 2004, 10, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Marcotte, T.D.; Wolfson, T.; Rosenthal, T.J.; Heaton, R.K.; Gonzalez, R.; Ellis, R.J.; Grant, I.; Group, H.I.V.N.R.C. A multimodal assessment of driving performance in HIV infection. Neurology 2004, 63, 1417–1422. [Google Scholar] [CrossRef] [PubMed]
- Albert, S.M.; Marder, K.; Dooneief, G.; Bell, K.; Sano, M.; Todak, G.; Stern, Y. Neuropsychologic impairment in early HIV infection. A risk factor for work disability. Arch. Neurol. 1995, 52, 525–530. [Google Scholar] [CrossRef] [PubMed]
- van Gorp, W.G.; Baerwald, J.P.; Ferrando, S.J.; McElhiney, M.C.; Rabkin, J.G. The relationship between employment and neuropsychological impairment in HIV infection. J. Int. Neuropsychol. Soc. 1999, 5, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Heaton, R.K.; Velin, R.A.; McCutchan, J.A.; Gulevich, S.J.; Atkinson, J.H.; Wallace, M.R.; Godfrey, H.P.; Kirson, D.A.; Grant, I. Neuropsychological impairment in human immunodeficiency virus-infection: implications for employment. HNRC Group. HIV Neurobehavioral Research Center. Psychosom. Med. 1994, 56, 8–17. [Google Scholar] [CrossRef]
- Martin, C.; Solders, G.; Sonnerborg, A.; Hansson, P. Painful and non-painful neuropathy in HIV-infected patients: an analysis of somatosensory nerve function. Eur. J. Pain 2003, 7, 23–31. [Google Scholar] [CrossRef]
- Mitra, P.; Sharman, T. HIV Neurocognitive Disorders. In StatPearls; Treasure Island (FL), 2022.
- Whetten, K.; Reif, S.; Whetten, R.; Murphy-McMillan, L.K. Trauma, mental health, distrust, and stigma among HIV-positive persons: implications for effective care. Psychosom. Med. 2008, 70, 531–538. [Google Scholar] [CrossRef]
- Mediouni, S.; Darque, A.; Baillat, G.; Ravaux, I.; Dhiver, C.; Tissot-Dupont, H.; Mokhtari, M.; Moreau, H.; Tamalet, C.; Brunet, C.; et al. Antiretroviral therapy does not block the secretion of the human immunodeficiency virus tat protein. Infect. Disord. Drug. Targets 2012, 12, 81–86. [Google Scholar] [CrossRef]
- King, J.E.; Eugenin, E.A.; Buckner, C.M.; Berman, J.W. HIV tat and neurotoxicity. Microbes Infect. 2006, 8, 1347–1357. [Google Scholar] [CrossRef]
- Das, A.T.; Harwig, A.; Berkhout, B. The HIV-1 Tat protein has a versatile role in activating viral transcription. J. Virol. 2011, 85, 9506–9516. [Google Scholar] [CrossRef]
- Hudson, L.; Liu, J.; Nath, A.; Jones, M.; Raghavan, R.; Narayan, O.; Male, D.; Everall, I. Detection of the human immunodeficiency virus regulatory protein tat in CNS tissues. J. Neurovirol. 2000, 6, 145–155. [Google Scholar] [CrossRef]
- Wiley, C.A.; Baldwin, M.; Achim, C.L. Expression of HIV regulatory and structural mRNA in the central nervous system. AIDS 1996, 10, 843–847. [Google Scholar] [CrossRef]
- Bertrand, S.J.; Aksenova, M.V.; Mactutus, C.F.; Booze, R.M. HIV-1 Tat protein variants: critical role for the cysteine region in synaptodendritic injury. Exp. Neurol. 2013, 248, 228–235. [Google Scholar] [CrossRef]
- Carey, A.N.; Sypek, E.I.; Singh, H.D.; Kaufman, M.J.; McLaughlin, J.P. Expression of HIV-Tat protein is associated with learning and memory deficits in the mouse. Behav. Brain Res. 2012, 229, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Fitting, S.; Ignatowska-Jankowska, B.M.; Bull, C.; Skoff, R.P.; Lichtman, A.H.; Wise, L.E.; Fox, M.A.; Su, J.; Medina, A.E.; Krahe, T.E.; et al. Synaptic dysfunction in the hippocampus accompanies learning and memory deficits in human immunodeficiency virus type-1 Tat transgenic mice. Biol. Psychiatry 2013, 73, 443–453. [Google Scholar] [CrossRef] [PubMed]
- League, A.F.; Gorman, B.L.; Hermes, D.J.; Johnson, C.T.; Jacobs, I.R.; Yadav-Samudrala, B.J.; Poklis, J.L.; Niphakis, M.J.; Cravatt, B.F.; Lichtman, A.H.; et al. Monoacylglycerol Lipase Inhibitor MJN110 Reduces Neuronal Hyperexcitability, Restores Dendritic Arborization Complexity, and Regulates Reward-Related Behavior in Presence of HIV-1 Tat. Front. Neurol. 2021, 12, 651272. [Google Scholar] [CrossRef]
- Hahn, Y.K.; Masvekar, R.R.; Xu, R.; Hauser, K.F.; Knapp, P.E. Chronic HIV-1 Tat and HIV reduce Rbfox3/NeuN: evidence for sex-related effects. Curr. HIV Res. 2015, 13, 10–20. [Google Scholar] [CrossRef]
- Wodarski, R.; Bagdas, D.; Paris, J.J.; Pheby, T.; Toma, W.; Xu, R.; Damaj, M.I.; Knapp, P.E.; Rice, A.S.C.; Hauser, K.F. Reduced intraepidermal nerve fibre density, glial activation, and sensory changes in HIV type-1 Tat-expressing female mice: involvement of Tat during early stages of HIV-associated painful sensory neuropathy. Pain Rep. 2018, 3, e654. [Google Scholar] [CrossRef]
- Marks, W.D.; Paris, J.J.; Barbour, A.J.; Moon, J.; Carpenter, V.J.; McLane, V.D.; Lark, A.R.S.; Nass, S.R.; Zhang, J.; Yarotskyy, V.; et al. HIV-1 Tat and Morphine Differentially Disrupt Pyramidal Cell Structure and Function and Spatial Learning in Hippocampal Area CA1: Continuous versus Interrupted Morphine Exposure. eNeuro 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fan, Y.; Vann, P.H.; Wong, J.M.; Sumien, N.; He, J.J. Long-term HIV-1 Tat Expression in the Brain Led to Neurobehavioral, Pathological, and Epigenetic Changes Reminiscent of Accelerated Aging. Aging. Dis. 2020, 11, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Ensoli, B.; Barillari, G.; Salahuddin, S.Z.; Gallo, R.C.; Wong-Staal, F. Tat protein of HIV-1 stimulates growth of cells derived from Kaposi’s sarcoma lesions of AIDS patients. Nature 1990, 345, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, J.M.; Vives, E.; Mabrouk, K.; Benjouad, A.; Rochat, H.; Duval, A.; Hue, B.; Bahraoui, E. Evidence for neurotoxic activity of tat from human immunodeficiency virus type 1. J. Virol. 1991, 65, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Conant, K.; Tornatore, C.; Atwood, W.; Meyers, K.; Traub, R.; Major, E.O. In vivo and in vitro infection of the astrocyte by HIV-1. Adv. Neuroimmunol. 1994, 4, 287–289. [Google Scholar] [CrossRef]
- Kannan, M.; Singh, S.; Chemparathy, D.T.; Oladapo, A.A.; Gawande, D.Y.; Dravid, S.M.; Buch, S.; Sil, S. HIV-1 Tat induced microglial EVs leads to neuronal synaptodendritic injury: microglia-neuron cross-talk in NeuroHIV. Extracell. Vesicles Circ. Nucl. Acids 2022, 3, 133–149. [Google Scholar] [CrossRef]
- Fitting, S.; Knapp, P.E.; Zou, S.; Marks, W.D.; Bowers, M.S.; Akbarali, H.I.; Hauser, K.F. Interactive HIV-1 Tat and morphine-induced synaptodendritic injury is triggered through focal disruptions in Na(+) influx, mitochondrial instability, and Ca(2)(+) overload. J. Neurosci. 2014, 34, 12850–12864. [Google Scholar] [CrossRef]
- Philippon, V.; Vellutini, C.; Gambarelli, D.; Harkiss, G.; Arbuthnott, G.; Metzger, D.; Roubin, R.; Filippi, P. The basic domain of the lentiviral Tat protein is responsible for damages in mouse brain: involvement of cytokines. Virology 1994, 205, 519–529. [Google Scholar] [CrossRef]
- Prendergast, M.A.; Rogers, D.T.; Mulholland, P.J.; Littleton, J.M.; Wilkins, L.H., Jr.; Self, R.L.; Nath, A. Neurotoxic effects of the human immunodeficiency virus type-1 transcription factor Tat require function of a polyamine sensitive-site on the N-methyl-D-aspartate receptor. Brain Res. 2002, 954, 300–307. [Google Scholar] [CrossRef]
- Hermes, D.J.; Yadav-Samudrala, B.J.; Xu, C.; Paniccia, J.E.; Meeker, R.B.; Armstrong, M.L.; Reisdorph, N.; Cravatt, B.F.; Mackie, K.; Lichtman, A.H.; et al. GPR18 drives FAAH inhibition-induced neuroprotection against HIV-1 Tat-induced neurodegeneration. Exp. Neurol. 2021, 341, 113699. [Google Scholar] [CrossRef]
- Bertrand, S.J.; Mactutus, C.F.; Aksenova, M.V.; Espensen-Sturges, T.D.; Booze, R.M. Synaptodendritic recovery following HIV Tat exposure: neurorestoration by phytoestrogens. J. Neurochem. 2014, 128, 140–151. [Google Scholar] [CrossRef]
- Cheng, J.; Nath, A.; Knudsen, B.; Hochman, S.; Geiger, J.D.; Ma, M.; Magnuson, D.S. Neuronal excitatory properties of human immunodeficiency virus type 1 Tat protein. Neuroscience 1998, 82, 97–106. [Google Scholar] [CrossRef]
- Jin, J.; Lam, L.; Sadic, E.; Fernandez, F.; Tan, J.; Giunta, B. HIV-1 Tat-induced microglial activation and neuronal damage is inhibited via CD45 modulation: A potential new treatment target for HAND. Am. J. Transl. Res. 2012, 4, 302–315. [Google Scholar]
- Hahn, Y.K.; Vo, P.; Fitting, S.; Block, M.L.; Hauser, K.F.; Knapp, P.E. β-chemokine production by neural and glial progenitor cells is enhanced by HIV-1 Tat: effects on microglial migration. J. Neurochem. 2010, 114, 97–109. [Google Scholar] [CrossRef]
- Nath, A.; Conant, K.; Chen, P.; Scott, C.; Major, E.O. Transient exposure to HIV-1 Tat protein results in cytokine production in macrophages and astrocytes. A hit and run phenomenon. J. Biol. Chem. 1999, 274, 17098–17102. [Google Scholar] [CrossRef]
- Kesby, J.P.; Markou, A.; Semenova, S. The effects of HIV-1 regulatory TAT protein expression on brain reward function, response to psychostimulants and delay-dependent memory in mice. Neuropharmacology 2016, 109, 205–215. [Google Scholar] [CrossRef]
- Bagdas, D.; Paris, J.J.; Carper, M.; Wodarski, R.; Rice, A.S.C.; Knapp, P.E.; Hauser, K.F.; Damaj, M.I. Conditional expression of HIV-1 Tat in the mouse alters the onset and progression of tonic, inflammatory, and neuropathic hypersensitivity in a sex-dependent manner. Eur. J. Pain 2020. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Amet, T.; Byrd, D.; Chang, K.H.; Shah, K.; Hu, N.; Grantham, A.; Hu, S.; Duan, J.; Tao, F.; et al. Direct effects of HIV-1 Tat on excitability and survival of primary dorsal root ganglion neurons: possible contribution to HIV-1-associated pain. Plos One 2011, 6, e24412. [Google Scholar] [CrossRef] [PubMed]
- Cirino, T.J.; Alleyne, A.R.; Duarte, V.; Figueroa, A.; Simons, C.A.; Anceaume, E.M.; Kendrick, J.; Wallman, O.; Eans, S.O.; Stacy, H.M.; et al. Expression of Human Immunodeficiency Virus Transactivator of Transcription (HIV-Tat1-86) Protein Alters Nociceptive Processing that is Sensitive to Anti-Oxidant and Anti-Inflammatory Interventions. J. Neuroimmune. Pharmacol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Salahuddin, M.F.; Mahdi, F.; Paris, J.J. HIV-1 Tat Dysregulates the Hypothalamic-Pituitary-Adrenal Stress Axis and Potentiates Oxycodone-Mediated Psychomotor and Anxiety-Like Behavior of Male Mice. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Qrareya, A.N.; Mahdi, F.; Kaufman, M.J.; Ashpole, N.M.; Paris, J.J. HIV-1 Tat promotes age-related cognitive, anxiety-like, and antinociceptive impairments in female mice that are moderated by aging and endocrine status. Geroscience 2021, 43, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Hahn, Y.K.; Paris, J.J.; Lichtman, A.H.; Hauser, K.F.; Sim-Selley, L.J.; Selley, D.E.; Knapp, P.E. Central HIV-1 Tat exposure elevates anxiety and fear conditioned responses of male mice concurrent with altered mu-opioid receptor-mediated G-protein activation and beta-arrestin 2 activity in the forebrain. Neurobiol. Dis. 2016, 92, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Joshi, C.R.; Stacy, S.; Sumien, N.; Ghorpade, A.; Borgmann, K. Astrocyte HIV-1 Tat Differentially Modulates Behavior and Brain MMP/TIMP Balance During Short and Prolonged Induction in Transgenic Mice. Front. Neurol. 2020, 11, 593188. [Google Scholar] [CrossRef]
- Paris, J.J.; Fenwick, J.; McLaughlin, J.P. Progesterone protects normative anxiety-like responding among ovariectomized female mice that conditionally express the HIV-1 regulatory protein, Tat, in the CNS. Horm. Behav. 2014, 65, 445–453. [Google Scholar] [CrossRef]
- Kasten, C.R.; Zhang, Y.; Boehm, S.L. , 2nd. Acute and long-term effects of Delta9-tetrahydrocannabinol on object recognition and anxiety-like activity are age- and strain-dependent in mice. Pharmacol. Biochem. Behav. 2017, 163, 9–19. [Google Scholar] [CrossRef]
- Bie, B.; Wu, J.; Foss, J.F.; Naguib, M. An overview of the cannabinoid type 2 receptor system and its therapeutic potential. Curr. Opin. Anaesthesiol. 2018, 31, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Yadav-Samudrala, B.J.; Fitting, S. Mini-review: The therapeutic role of cannabinoids in neuroHIV. Neurosci. Lett. 2021, 750, 135717. [Google Scholar] [CrossRef]
- Woolridge, E.; Barton, S.; Samuel, J.; Osorio, J.; Dougherty, A.; Holdcroft, A. Cannabis use in HIV for pain and other medical symptoms. J. Pain Symptom Manage. 2005, 29, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Wardell, J.D.; Rueda, S.; Fox, N.; Costiniuk, C.T.; Jenabian, M.A.; Margolese, S.; Mandarino, E.; Shuper, P.; Hendershot, C.S.; Cunningham, J.A.; et al. Disentangling Medicinal and Recreational cannabis Use Among People Living with HIV: An Ecological Momentary Assessment Study. AIDS Behav. 2023, 4, 1350–1363. [Google Scholar] [CrossRef]
- Shiau, S.; Arpadi, S.M.; Yin, M.T.; Martins, S.S. Patterns of drug use and HIV infection among adults in a nationally representative sample. Addict. Behav. 2017, 68, 39–44. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Saneei, Z.; Salahuddin, S.; Cox, J.; Routy, J.P.; Rueda, S.; Abdallah, S.J.; Jensen, D.; Lebouche, B.; Brouillette, M.J.; et al. Cannabis Consumption in People Living with HIV: Reasons for Use, Secondary Effects, and Opportunities for Health Education. Cannabis Cannabinoid Res. 2019, 4, 204–213. [Google Scholar] [CrossRef]
- Whitfield, R.M.; Bechtel, L.M.; Starich, G.H. The impact of ethanol and Marinol/marijuana usage on HIV+/AIDS patients undergoing azidothymidine, azidothymidine/dideoxycytidine, or dideoxyinosine therapy. Alcohol Clin. Exp. Res. 1997, 21, 122–127. [Google Scholar] [CrossRef]
- Cristiani, S.A.; Pukay-Martin, N.D.; Bornstein, R.A. Marijuana use and cognitive function in HIV-infected people. J. Neuropsychiatry Clin. Neurosci. 2004, 16, 330–335. [Google Scholar] [CrossRef]
- Pacek, L.R.; Towe, S.L.; Hobkirk, A.L.; Nash, D.; Goodwin, R.D. Frequency of Cannabis Use and Medical Cannabis Use Among Persons Living With HIV in the United States: Findings From a Nationally Representative Sample. AIDS Educ. Prev. 2018, 30, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Wilson, N.; Peterson, S. Cannabis and Inflammation in HIV: A Review of Human and Animal Studies. Viruses 2021, 13, 1521. [Google Scholar] [CrossRef] [PubMed]
- Iuvone, T.; Esposito, G.; Esposito, R.; Santamaria, R.; Di Rosa, M.; Izzo, A.A. Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis sativa, on beta-amyloid-induced toxicity in PC12 cells. J. Neurochem. 2004, 89, 134–141. [Google Scholar] [CrossRef]
- Booz, G.W. Cannabidiol as an emergent therapeutic strategy for lessening the impact of inflammation on oxidative stress. Free Radic. Biol. Med. 2011, 51, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Hughes, B.; Herron, C.E. Cannabidiol Reverses Deficits in Hippocampal LTP in a Model of Alzheimer’s Disease. Neurochem. Res. 2019, 44, 703–713. [Google Scholar] [CrossRef]
- Garcia-Baos, A.; Puig-Reyne, X.; Garcia-Algar, O.; Valverde, O. Cannabidiol attenuates cognitive deficits and neuroinflammation induced by early alcohol exposure in a mice model. Biomed. Pharmacother. 2021, 141, 111813. [Google Scholar] [CrossRef]
- Osborne, A.L.; Solowij, N.; Weston-Green, K. A systematic review of the effect of cannabidiol on cognitive function: Relevance to schizophrenia. Neurosci. Biobehav. Rev. 2017, 72, 310–324. [Google Scholar] [CrossRef]
- Osborne, A.L.; Solowij, N.; Babic, I.; Huang, X.F.; Weston-Green, K. Improved Social Interaction, Recognition and Working Memory with Cannabidiol Treatment in a Prenatal Infection (poly I:C) Rat Model. Neuropsychopharmacology 2017, 42, 1447–1457. [Google Scholar] [CrossRef]
- Cheng, D.; Low, J.K.; Logge, W.; Garner, B.; Karl, T. Chronic cannabidiol treatment improves social and object recognition in double transgenic APPswe/PS1E9 mice. Psychopharmacology (Berl.) 2014, 231, 3009–3017. [Google Scholar] [CrossRef]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Properties of Cannabidiol. Antioxidants (Basel) 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.M.; Duncan, M.; Di Marzo, V.; Pertwee, R.G. Are cannabidiol and Delta(9) -tetrahydrocannabivarin negative modulators of the endocannabinoid system? A systematic review. B.r J. Pharmacol. 2015, 172, 737–753. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: delta9-tetrahydrocannabinol, cannabidiol and delta9-tetrahydrocannabivarin. Br. J. Pharmacol. 2008, 153, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Sagredo, O.; Pazos, M.R.; Satta, V.; Ramos, J.A.; Pertwee, R.G.; Fernandez-Ruiz, J. Neuroprotective effects of phytocannabinoid-based medicines in experimental models of Huntington’s disease. J. Neurosci. Res. 2011, 89, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Galaj, E.; Bi, G.H.; Yang, H.J.; Xi, Z.X. Cannabidiol attenuates the rewarding effects of cocaine in rats by CB2, 5-HT1A and TRPV1 receptor mechanisms. Neuropharmacology 2020, 167, 107740. [Google Scholar] [CrossRef]
- Vilela, L.R.; Lima, I.V.; Kunsch, E.B.; Pinto, H.P.P.; de Miranda, A.S.; Vieira, E.L.M.; de Oliveira, A.C.P.; Moraes, M.F.D.; Teixeira, A.L.; Moreira, F.A. Anticonvulsant effect of cannabidiol in the pentylenetetrazole model: Pharmacological mechanisms, electroencephalographic profile, and brain cytokine levels. Epilepsy & behavior: E&B 2017, 75, 29–35. [Google Scholar] [CrossRef]
- Fogaca, M.V.; Campos, A.C.; Coelho, L.D.; Duman, R.S.; Guimaraes, F.S. The anxiolytic effects of cannabidiol in chronically stressed mice are mediated by the endocannabinoid system: Role of neurogenesis and dendritic remodeling. Neuropharmacology 2018, 135, 22–33. [Google Scholar] [CrossRef]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.; Denovan-Wright, E.M. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef]
- Hassan, S.; Eldeeb, K.; Millns, P.J.; Bennett, A.J.; Alexander, S.P.; Kendall, D.A. Cannabidiol enhances microglial phagocytosis via transient receptor potential (TRP) channel activation. Br. J. Pharmacol. 2014, 171, 2426–2439. [Google Scholar] [CrossRef]
- Zanelati, T.V.; Biojone, C.; Moreira, F.A.; Guimaraes, F.S.; Joca, S.R. Antidepressant-like effects of cannabidiol in mice: possible involvement of 5-HT1A receptors. Br. J. Pharmacol. 2010, 159, 122–128. [Google Scholar] [CrossRef]
- Resstel, L.B.; Tavares, R.F.; Lisboa, S.F.; Joca, S.R.; Correa, F.M.; Guimaraes, F.S. 5-HT1A receptors are involved in the cannabidiol-induced attenuation of behavioural and cardiovascular responses to acute restraint stress in rats. Br. J. Pharmacol. 2009, 156, 181–188. [Google Scholar] [CrossRef]
- Esposito, G.; Scuderi, C.; Valenza, M.; Togna, G.I.; Latina, V.; De Filippis, D.; Cipriano, M.; Carratu, M.R.; Iuvone, T.; Steardo, L. Cannabidiol reduces Abeta-induced neuroinflammation and promotes hippocampal neurogenesis through PPARgamma involvement. PLoS One 2011, 6, e28668. [Google Scholar] [CrossRef]
- Khosropoor, S.; Alavi, M.S.; Etemad, L.; Roohbakhsh, A. Cannabidiol goes nuclear: The role of PPARgamma. Phytomedicine 2023, 114, 154771. [Google Scholar] [CrossRef]
- Sonego, A.B.; Prado, D.D.S.; Guimaraes, F.S. PPARgamma receptors are involved in the effects of cannabidiol on orofacial dyskinesia and cognitive dysfunction induced by typical antipsychotic in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 111, 110367. [Google Scholar] [CrossRef]
- Burstein, S. Cannabidiol (CBD) and its analogs: a review of their effects on inflammation. Bioorg. Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef]
- Ryberg, E.; Larsson, N.; Sjogren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- Campos, A.C.; Ortega, Z.; Palazuelos, J.; Fogaca, M.V.; Aguiar, D.C.; Diaz-Alonso, J.; Ortega-Gutierrez, S.; Vazquez-Villa, H.; Moreira, F.A.; Guzman, M.; et al. The anxiolytic effect of cannabidiol on chronically stressed mice depends on hippocampal neurogenesis: involvement of the endocannabinoid system. Int. J. Neuropsychopharmacol. 2013, 16, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, T.; Hanus, L.; De Petrocellis, L.; Tchilibon, S.; Ponde, D.E.; Brandi, I.; Moriello, A.S.; Davis, J.B.; Mechoulam, R.; Di Marzo, V. Molecular targets for cannabidiol and its synthetic analogues: effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br. J. Pharmacol. 2001, 134, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Boggs, D.L.; Nguyen, J.D.; Morgenson, D.; Taffe, M.A.; Ranganathan, M. Clinical and Preclinical Evidence for Functional Interactions of Cannabidiol and Delta(9)-Tetrahydrocannabinol. Neuropsychopharmacology 2018, 43, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Bruce-Keller, A.J.; Turchan-Cholewo, J.; Smart, E.J.; Geurin, T.; Chauhan, A.; Reid, R.; Xu, R.; Nath, A.; Knapp, P.E.; Hauser, K.F. Morphine causes rapid increases in glial activation and neuronal injury in the striatum of inducible HIV-1 Tat transgenic mice. Glia 2008, 56, 1414–1427. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Turchan, J.; Pocernich, C.B.; Bruce-Keller, A.J.; Roth, S.; Butterfield, D.A.; Major, E.O.; Nath, A. Intracellular human immunodeficiency virus tat expression in astrocytes promotes astrocyte survival but induces potent neurotoxicity at distant sites via axonal transport. J. Biol. Chem. 2003, 278, 13512–13519. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Kongara, K.; Harding, D.; Ward, N.; Dukkipati, V.S.R.; Johnson, C.; Chambers, P. Comparison of electroencephalographic changes in response to acute electrical and thermal stimuli with the tail flick and hot plate test in rats administered with opiorphin. BMC Neurol. 2018, 18, 43. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.J.; Roberts, D.J. The quantiative measurement of motor inco-ordination in naive mice using an acelerating rotarod. J. Pharm. Pharmacol. 1968, 20, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Shoji, H.; Miyakawa, T. Effects of test experience, closed-arm wall color, and illumination level on behavior and plasma corticosterone response in an elevated plus maze in male C57BL/6J mice: a challenge against conventional interpretation of the test. Mol. Brain 2021, 14, 34. [Google Scholar] [CrossRef]
- Lueptow, L.M. Novel Object Recognition Test for the Investigation of Learning and Memory in Mice. J. Vis. Exp. 2017, 126, 55718. [Google Scholar] [CrossRef]
- Ennaceur, A. One-trial object recognition in rats and mice: methodological and theoretical issues. Behav. Brain Res. 2010, 215, 244–254. [Google Scholar] [CrossRef]
- Miedel, C.J.; Patton, J.M.; Miedel, A.N.; Miedel, E.S.; Levenson, J.M. Assessment of Spontaneous Alternation, Novel Object Recognition and Limb Clasping in Transgenic Mouse Models of Amyloid-beta and Tau Neuropathology. J. Vis. Exp. 2017, 123, 55523. [Google Scholar] [CrossRef]
- Dempsey, S.K.; Gesseck, A.M.; Ahmad, A.; Daneva, Z.; Ritter, J.K.; Poklis, J.L. Formation of HETE-EAs and dihydroxy derivatives in mouse kidney tissue and analysis by high-performance liquid chromatography tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2019, 1126-1127, 121748. [Google Scholar] [CrossRef]
- Slawek, D.E. People living with HIV and the emerging field of chronic pain-what is known about epidemiology, etiology, and management. Curr. HIV/AIDS Rep. 2021, 18, 436–442. [Google Scholar] [CrossRef]
- Lu, H.J.; Fu, Y.Y.; Wei, Q.Q.; Zhang, Z.J. Neuroinflammation in HIV-Related Neuropathic Pain. Front. Pharmacol. 2021, 12, 653852. [Google Scholar] [CrossRef]
- Namisango, E.; Harding, R.; Atuhaire, L.; Ddungu, H.; Katabira, E.; Muwanika, F.R.; Powell, R.A. Pain among ambulatory HIV/AIDS patients: multicenter study of prevalence, intensity, associated factors, and effect. J. Pain 2012, 13, 704–713. [Google Scholar] [CrossRef]
- Fitting, S.; Scoggins, K.L.; Xu, R.; Dever, S.M.; Knapp, P.E.; Dewey, W.L.; Hauser, K.F. Morphine efficacy is altered in conditional HIV-1 Tat transgenic mice. Eur. J. Pharmacol. 2012, 689, 96–103. [Google Scholar] [CrossRef]
- Toma, W.; Paris, J.J.; Warncke, U.O.; Nass, S.R.; Caillaud, M.; McKiver, B.; Ondo, O.; Bagdas, D.; Bigbee, J.; Knapp, P.E.; et al. Persistent sensory changes and sex differences in transgenic mice conditionally expressing HIV-1 Tat regulatory protein. Exp. Neurol. 2022, 358, 114226. [Google Scholar] [CrossRef] [PubMed]
- Jesus, C.H.A.; Redivo, D.D.B.; Gasparin, A.T.; Sotomaior, B.B.; de Carvalho, M.C.; Genaro, K.; Zuardi, A.W.; Hallak, J.E.C.; Crippa, J.A.; Zanoveli, J.M.; et al. Cannabidiol attenuates mechanical allodynia in streptozotocin-induced diabetic rats via serotonergic system activation through 5-HT1A receptors. Brain Res. 2019, 1715, 156–164. [Google Scholar] [CrossRef]
- Wanasuntronwong, A.; Kaewsrisung, S.; Rotpenpian, N.; Arayapisit, T.; Pavasant, P.; Supronsinchai, W. Efficacy and mechanism of the antinociceptive effects of cannabidiol on acute orofacial nociception induced by Complete Freund’s Adjuvant in male Mus musculus mice. Arch. Oral Biol. 2022, 144, 105570. [Google Scholar] [CrossRef]
- Genaro, K.; Fabris, D.; Arantes, A.L.F.; Zuardi, A.W.; Crippa, J.A.S.; Prado, W.A. Cannabidiol Is a Potential Therapeutic for the Affective-Motivational Dimension of Incision Pain in Rats. Front. Pharmacol. 2017, 8, 391. [Google Scholar] [CrossRef] [PubMed]
- Abrams, D.I.; Jay, C.A.; Shade, S.B.; Vizoso, H.; Reda, H.; Press, S.; Kelly, M.E.; Rowbotham, M.C.; Petersen, K.L. Cannabis in painful HIV-associated sensory neuropathy: a randomized placebo-controlled trial. Neurology 2007, 68, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Toperoff, W.; Vaida, F.; van den Brande, G.; Gonzales, J.; Gouaux, B.; Bentley, H.; Atkinson, J.H. Smoked medicinal cannabis for neuropathic pain in HIV: a randomized, crossover clinical trial. Neuropsychopharmacology 2009, 34, 672–680. [Google Scholar] [CrossRef]
- Johnson, J.R.; Burnell-Nugent, M.; Lossignol, D.; Ganae-Motan, E.D.; Potts, R.; Fallon, M.T. Multicenter, double-blind, randomized, placebo-controlled, parallel-group study of the efficacy, safety, and tolerability of THC:CBD extract and THC extract in patients with intractable cancer-related pain. J. Pain Symptom. Manage. 2010, 39, 167–179. [Google Scholar] [CrossRef]
- Robinson-Papp, J.; Gensler, G.; Navis, A.; Sherman, S.; Ellis, R.J.; Gelman, B.B.; Kolson, D.L.; Letendre, S.L.; Singer, E.J.; Valdes-Sueiras, M.; et al. Characteristics of Motor Dysfunction in Longstanding Human Immunodeficiency Virus. Clin. Infect. Dis. 2020, 71, 1532–1538. [Google Scholar] [CrossRef]
- Kronemer, S.I.; Mandel, J.A.; Sacktor, N.C.; Marvel, C.L. Impairments of Motor Function While Multitasking in HIV. Front. Hum. Neurosci. 2017, 11, 212. [Google Scholar] [CrossRef] [PubMed]
- Vancampfort, D.; Mugisha, J.; De Hert, M.; Probst, M.; Firth, J.; Gorczynski, P.; Stubbs, B. Global physical activity levels among people living with HIV: a systematic review and meta-analysis. Disabil. Rehabil. 2018, 40, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, G.; Pinacchio, C.; Santinelli, L.; Adami, P.E.; Borrazzo, C.; Cavallari, E.N.; Vullo, A.; Innocenti, G.P.; Mezzaroma, I.; Mastroianni, C.M.; et al. Physical Activity and HIV: Effects on Fitness Status, Metabolism, Inflammation and Immune-Activation. AIDS Behav. 2020, 24, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- June, H.L.; Tzeng Yang, A.R.; Bryant, J.L.; Jones, O.; Royal, W. , 3rd. Vitamin A deficiency and behavioral and motor deficits in the human immunodeficiency virus type 1 transgenic rat. J. Neurovirol. 2009, 15, 380–389. [Google Scholar] [CrossRef]
- Moran, L.M.; Booze, R.M.; Webb, K.M.; Mactutus, C.F. Neurobehavioral alterations in HIV-1 transgenic rats: evidence for dopaminergic dysfunction. Exp. Neurol. 2013, 239, 139–147. [Google Scholar] [CrossRef]
- Hahn, Y.K.; Podhaizer, E.M.; Farris, S.P.; Miles, M.F.; Hauser, K.F.; Knapp, P.E. Effects of chronic HIV-1 Tat exposure in the CNS: heightened vulnerability of males versus females to changes in cell numbers, synaptic integrity, and behavior. Brain Struct. Funct. 2015, 220, 605–623. [Google Scholar] [CrossRef]
- Jacobs, I.R.; Xu, C.; Hermes, D.J.; League, A.F.; Xu, C.; Nath, B.; Jiang, W.; Niphakis, M.J.; Cravatt, B.F.; Mackie, K.; et al. Inhibitory Control Deficits Associated with Upregulation of CB1R in the HIV-1 Tat Transgenic Mouse Model of Hand. J. Neuroimmune Pharmacol. 2019, 14, 661–678. [Google Scholar] [CrossRef]
- Poulia, N.; Delis, F.; Brakatselos, C.; Ntoulas, G.; Asprogerakas, M.Z.; Antoniou, K. CBD Effects on Motor Profile and Neurobiological Indices Related to Glutamatergic Function Induced by Repeated Ketamine Pre-Administration. Front. Pharmacol. 2021, 12, 746935. [Google Scholar] [CrossRef]
- Viudez-Martinez, A.; Garcia-Gutierrez, M.S.; Medrano-Relinque, J.; Navarron, C.M.; Navarrete, F.; Manzanares, J. Cannabidiol does not display drug abuse potential in mice behavior. Acta Pharmacol. Sin. 2019, 40, 358–364. [Google Scholar] [CrossRef]
- Zieba, J.; Sinclair, D.; Sebree, T.; Bonn-Miller, M.; Gutterman, D.; Siegel, S.; Karl, T. Cannabidiol (CBD) reduces anxiety-related behavior in mice via an FMRP-independent mechanism. Pharmacol. Biochem. Behav. 2019, 181, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Coles, M.; Watt, G.; Kreilaus, F.; Karl, T. Medium-Dose Chronic Cannabidiol Treatment Reverses Object Recognition Memory Deficits of APP (Swe)/PS1DeltaE9 Transgenic Female Mice. Front. Pharmacol. 2020, 11, 587604. [Google Scholar] [CrossRef]
- Florensa-Zanuy, E.; Garro-Martinez, E.; Adell, A.; Castro, E.; Diaz, A.; Pazos, A.; Mac-Dowell, K.S.; Martin-Hernandez, D.; Pilar-Cuellar, F. Cannabidiol antidepressant-like effect in the lipopolysaccharide model in mice: Modulation of inflammatory pathways. Biochem. Pharmacol. 2021, 185, 114433. [Google Scholar] [CrossRef]
- Schleicher, E.M.; Ott, F.W.; Muller, M.; Silcher, B.; Sichler, M.E.; Low, M.J.; Wagner, J.M.; Bouter, Y. Prolonged Cannabidiol Treatment Lacks on Detrimental Effects on Memory, Motor Performance and Anxiety in C57BL/6J Mice. Front. Behav. Neurosci. 2019, 13, 94. [Google Scholar] [CrossRef]
- Calapai, F.; Cardia, L.; Calapai, G.; Di Mauro, D.; Trimarchi, F.; Ammendolia, I.; Mannucci, C. Effects of Cannabidiol on Locomotor Activity. Life (Basel) 2022, 5, 652. [Google Scholar] [CrossRef]
- Magen, I.; Avraham, Y.; Ackerman, Z.; Vorobiev, L.; Mechoulam, R.; Berry, E.M. Cannabidiol ameliorates cognitive and motor impairments in bile-duct ligated mice via 5-HT1A receptor activation. Br. J. Pharmacol. 2010, 159, 950–957. [Google Scholar] [CrossRef]
- Blessing, E.M.; Steenkamp, M.M.; Manzanares, J.; Marmar, C.R. Cannabidiol as a Potential Treatment for Anxiety Disorders. Neurotherapeutics 2015, 12, 825–836. [Google Scholar] [CrossRef]
- Bonaccorso, S.; Ricciardi, A.; Zangani, C.; Chiappini, S.; Schifano, F. Cannabidiol (CBD) use in psychiatric disorders: A systematic review. Neurotoxicology 2019, 74, 282–298. [Google Scholar] [CrossRef]
- Huffstetler, C.M.; Cochran, B.; May, C.A.; Maykut, N.; Silver, C.R.; Cedeno, C.; Franck, E.; Cox, A.; Fadool, D.A. Single cannabidiol administration affects anxiety-, obsessive compulsive-, object memory-, and attention-like behaviors in mice in a sex and concentration dependent manner. Pharmacol. Biochem. Behav. 2023, 222, 173498. [Google Scholar] [CrossRef]
- Wright, M.; Di Ciano, P.; Brands, B. Use of Cannabidiol for the Treatment of Anxiety: A Short Synthesis of Pre-Clinical and Clinical Evidence. Cannabis Cannabinoid Res. 2020, 5, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Fabris, D.; Carvalho, M.C.; Brandao, M.L.; Prado, W.A.; Zuardi, A.W.; Crippa, J.A.; de Oliveira, A.R.; Lovick, T.A.; Genaro, K. Sex-dependent differences in the anxiolytic-like effect of cannabidiol in the elevated plus-maze. J. Psychopharmacol. 2022, 36, 1371–1383. [Google Scholar] [CrossRef]
- Salviato, B.Z.; Raymundi, A.M.; Rodrigues da Silva, T.; Salemme, B.W.; Batista Sohn, J.M.; Araujo, F.S.; Guimaraes, F.S.; Bertoglio, L.J.; Stern, C.A. Female but not male rats show biphasic effects of low doses of Delta(9)-tetrahydrocannabinol on anxiety: can cannabidiol interfere with these effects? Neuropharmacology 2021, 196, 108684. [Google Scholar] [CrossRef]
- Cryan, J.F.; Holmes, A. The ascent of mouse: advances in modelling human depression and anxiety. Nat. Rev. Drug Discov. 2005, 4, 775–790. [Google Scholar] [CrossRef]
- Marks, W.D.; Paris, J.J.; Schier, C.J.; Denton, M.D.; Fitting, S.; McQuiston, A.R.; Knapp, P.E.; Hauser, K.F. HIV-1 Tat causes cognitive deficits and selective loss of parvalbumin, somatostatin, and neuronal nitric oxide synthase expressing hippocampal CA1 interneuron subpopulations. J. Neurovirol. 2016. [Google Scholar] [CrossRef]
- Zhornitsky, S.; Potvin, S. Cannabidiol in humans-the quest for therapeutic targets. Pharmaceuticals (Basel) 2012, 5, 529–552. [Google Scholar] [CrossRef]
- Fagherazzi, E.V.; Garcia, V.A.; Maurmann, N.; Bervanger, T.; Halmenschlager, L.H.; Busato, S.B.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Schroder, N. Memory-rescuing effects of cannabidiol in an animal model of cognitive impairment relevant to neurodegenerative disorders. Psychopharmacology (Berl) 2012, 219, 1133–1140. [Google Scholar] [CrossRef]
- Razavi, Y.; Shabani, R.; Mehdizadeh, M.; Haghparast, A. Neuroprotective effect of chronic administration of cannabidiol during the abstinence period on methamphetamine-induced impairment of recognition memory in the rats. Behav. Pharmacol. 2020, 31, 385–396. [Google Scholar] [CrossRef]
- Stark, T.; Ruda-Kucerova, J.; Iannotti, F.A.; D’Addario, C.; Di Marco, R.; Pekarik, V.; Drazanova, E.; Piscitelli, F.; Bari, M.; Babinska, Z.; et al. Peripubertal cannabidiol treatment rescues behavioral and neurochemical abnormalities in the MAM model of schizophrenia. Neuropharmacology 2019, 146, 212–221. [Google Scholar] [CrossRef]
- Mustafa, M.A.; Poklis, J.L.; Karin, K.N.; Elmer, J.A.; Porter, J.H.; Parra, V.; Lu, D.; Schlosburg, J.E.; Lichtman, A.H. Investigation of Cannabidiol in the Mouse Drug Discrimination Paradigm. Cannabis Cannabinoid Res. 2023. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yadav-Samudrala, B.J.; Xu, C.; Nath, B.; Mistry, T.; Jiang, W.; Niphakis, M.J.; Cravatt, B.F.; Mukhopadhyay, S.; Lichtman, A.H.; et al. Inhibitory Neurotransmission Is Sex-Dependently Affected by Tat Expression in Transgenic Mice and Suppressed by the Fatty Acid Amide Hydrolase Enzyme Inhibitor PF3845 via Cannabinoid Type-1 Receptor Mechanisms. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Hermes, D.J.; Xu, C.; Poklis, J.L.; Niphakis, M.J.; Cravatt, B.F.; Mackie, K.; Lichtman, A.H.; Ignatowska-Jankowska, B.M.; Fitting, S. Neuroprotective effects of fatty acid amide hydrolase catabolic enzyme inhibition in a HIV-1 Tat model of neuroAIDS. Neuropharmacology 2018, 141, 55–65. [Google Scholar] [CrossRef]
- Swinton, M.K.; Sundermann, E.E.; Pedersen, L.; J. D., N.; Grelotti, D.J.; Taffe, M.A.; Iudicello, J.E.; Fields, J.A. Alterations in Brain Cannabinoid Receptor Levels Are Associated with HIV-Associated Neurocognitive Disorders in the ART Era: Implications for Therapeutic Strategies Targeting the Endocannabinoid System. Viruses 2021, 13, 1742. [Google Scholar] [CrossRef]
- Cosenza-Nashat, M.A.; Bauman, A.; Zhao, M.L.; Morgello, S.; Suh, H.S.; Lee, S.C. Cannabinoid receptor expression in HIV encephalitis and HIV-associated neuropathologic comorbidities. Neuropathol. Appl. Neurobiol. 2011, 37, 464–483. [Google Scholar] [CrossRef]
- Tallima, H.; El Ridi, R. Arachidonic acid: Physiological roles and potential health benefits - A review. J. Adv. Res. 2018, 11, 33–41. [Google Scholar] [CrossRef]
- Bosetti, F. Arachidonic acid metabolism in brain physiology and pathology: lessons from genetically altered mouse models. J. Neurochem. 2007, 102, 577–586. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: mechanisms and potential therapeutic targets. Signal Transduct. Target. Ther. 2021, 6, 94. [Google Scholar] [CrossRef]
- Hauser, K.F.; El-Hage, N.; Stiene-Martin, A.; Maragos, W.F.; Nath, A.; Persidsky, Y.; Volsky, D.J.; Knapp, P.E. HIV-1 neuropathogenesis: glial mechanisms revealed through substance abuse. J. Neurochem. 2007, 100, 567–586. [Google Scholar] [CrossRef]
- Chen, N.C.; Partridge, A.T.; Sell, C.; Torres, C.; Martin-Garcia, J. Fate of microglia during HIV-1 infection: From activation to senescence? Glia 2017, 65, 431–446. [Google Scholar] [CrossRef]
- Duarte, E.A.C.; Benevides, M.L.; Martins, A.L.P.; Duarte, E.P.; Weller, A.B.S.; de Azevedo, L.O.C.; de Oliveira Thais, M.E.R.; Nunes, J.C. Female sex is strongly associated with cognitive impairment in HIV infection. Neurol. Sci. 2021, 42, 1853–1860. [Google Scholar] [CrossRef]
- Rubin, L.H.; Neigh, G.N.; Sundermann, E.E.; Xu, Y.; Scully, E.P.; Maki, P.M. Sex Differences in Neurocognitive Function in Adults with HIV: Patterns, Predictors, and Mechanisms. Curr. Psychiatry Rep. 2019, 21, 94. [Google Scholar] [CrossRef]
- Sundermann, E.E.; Heaton, R.K.; Pasipanodya, E.; Moore, R.C.; Paolillo, E.W.; Rubin, L.H.; Ellis, R.; Moore, D.J.; Group, H. Sex differences in HIV-associated cognitive impairment. Aids 2018, 32, 2719–2726. [Google Scholar] [CrossRef]
- Maki, P.M.; Martin-Thormeyer, E. HIV, cognition and women. Neuropsychol. Rev. 2009, 19, 204–214. [Google Scholar] [CrossRef]
- Maki, P.M.; Rubin, L.H.; Springer, G.; Seaberg, E.C.; Sacktor, N.; Miller, E.N.; Valcour, V.; Young, M.A.; Becker, J.T.; Martin, E.M.; et al. Differences in Cognitive Function Between Women and Men With HIV. J. Acquir. Immune. Defic. Syndr. 2018, 79, 101–107. [Google Scholar] [CrossRef]
- Santinelli, L.; Ceccarelli, G.; Borrazzo, C.; Innocenti, G.P.; Frasca, F.; Cavallari, E.N.; Celani, L.; Nonne, C.; Mastroianni, C.M.; d’Ettorre, G. Sex-related differences in markers of immune activation in virologically suppressed HIV-infected patients. Biol. Sex. Differ. 2020, 11, 23. [Google Scholar] [CrossRef]
- Ziegler, S.; Altfeld, M. Sex differences in HIV-1-mediated immunopathology. Curr. Opin. HIV AIDS 2016, 11, 209–215. [Google Scholar] [CrossRef]
- Yuan, N.Y.; Maung, R.; Xu, Z.; Han, X.; Kaul, M. Arachidonic Acid Cascade and Eicosanoid Production Are Elevated While LTC4 Synthase Modulates the Lipidomics Profile in the Brain of the HIVgp120-Transgenic Mouse Model of NeuroHIV. Cells 2022, 11. [Google Scholar] [CrossRef]
- Levine, A.; Liktor-Busa, E.; Lipinski, A.A.; Couture, S.; Balasubramanian, S.; Aicher, S.A.; Langlais, P.R.; Vanderah, T.W.; Largent-Milnes, T.M. Sex differences in the expression of the endocannabinoid system within V1M cortex and PAG of Sprague Dawley rats. Biol. Sex Differ. 2021, 12, 60. [Google Scholar] [CrossRef]
- Arkell, T.R.; Kevin, R.C.; Vinckenbosch, F.; Lintzeris, N.; Theunissen, E.; Ramaekers, J.G.; McGregor, I.S. Sex differences in acute cannabis effects revisited: Results from two randomized, controlled trials. Addict. Biol. 2022, 27, e13125. [Google Scholar] [CrossRef]
- MacNair, L.; Kulpa, J.; Hill, M.L.; Eglit, G.M.L.; Mosesova, I.; Bonn-Miller, M.O.; Peters, E.N. Sex Differences in the Pharmacokinetics of Cannabidiol and Metabolites Following Oral Administration of a Cannabidiol-Dominant Cannabis Oil in Healthy Adults. Cannabis Cannabinoid Res. 2023. [Google Scholar] [CrossRef]
- Batinic, A.; Sutlovic, D.; Kuret, S.; Burcul, F.; Kalajzic, N.; Matana, A.; Dujic, G.; Vrdoljak, J.; Kumric, M.; Bozic, J.; et al. Differences in Plasma Cannabidiol Concentrations in Women and Men: A Randomized, Placebo-Controlled, Crossover Study. Int. J. Mol. Sci. 2023, 24, 10273. [Google Scholar] [CrossRef]
Figure 1.
Effects of acute CBD doses (0, 3, 10, and 30 mg/kg) on spontaneous evoked nociception and motor function. (A) For the spinal-related tail-flick assay, no significant effects and/or interactions were noted. (B) For the supraspinal-related hot-plate assay Tat(+) mice showed higher latencies to pain signals compared to Tat(−) mice independent of acute CBD doses. (C) Locomotor activity was decreased by Tat expression and acute CBD doses increased locomotor activity across sex and genotype. (D) Motor coordination was not affected by acute CBD, but Tat expression altered rotarod performance based on sex, with Tat(+) male mice showing lower rotarod performance compared to Tat(−) males, whereas the opposite was seen in females. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; § p < 0.05 main effect of genotype; # p < 0.05 main effect of drug dose; δ p = 0.034 sex x genotype interaction. n = 9-10 mice per group.
Figure 1.
Effects of acute CBD doses (0, 3, 10, and 30 mg/kg) on spontaneous evoked nociception and motor function. (A) For the spinal-related tail-flick assay, no significant effects and/or interactions were noted. (B) For the supraspinal-related hot-plate assay Tat(+) mice showed higher latencies to pain signals compared to Tat(−) mice independent of acute CBD doses. (C) Locomotor activity was decreased by Tat expression and acute CBD doses increased locomotor activity across sex and genotype. (D) Motor coordination was not affected by acute CBD, but Tat expression altered rotarod performance based on sex, with Tat(+) male mice showing lower rotarod performance compared to Tat(−) males, whereas the opposite was seen in females. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; § p < 0.05 main effect of genotype; # p < 0.05 main effect of drug dose; δ p = 0.034 sex x genotype interaction. n = 9-10 mice per group.
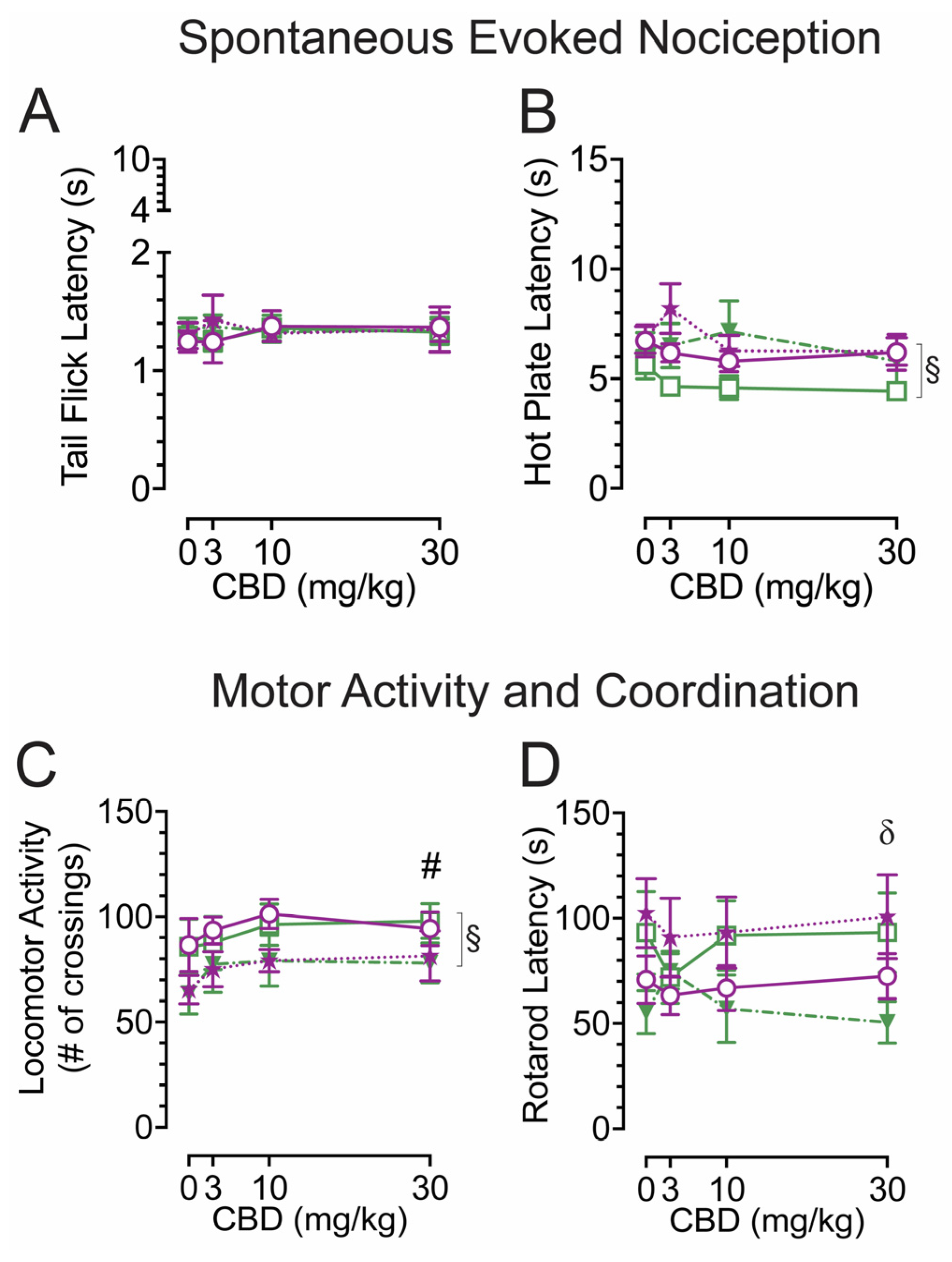
Figure 2.
Anxiety-like behavior assessed by the elevated plus maze was altered by sex and acute CBD (30 mg/kg) exposure. (A) CBD showed differential effects based on sex, with increasing time spent in open arms for males, thus decreasing anxiety in males, but not in females. (B) No significant effects were noted for number of pokes into open arms. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; ¶p = 0.034 sex x drug interaction. CBD dose = 30 mg/kg. n = 4-5 mice per group.
Figure 2.
Anxiety-like behavior assessed by the elevated plus maze was altered by sex and acute CBD (30 mg/kg) exposure. (A) CBD showed differential effects based on sex, with increasing time spent in open arms for males, thus decreasing anxiety in males, but not in females. (B) No significant effects were noted for number of pokes into open arms. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; ¶p = 0.034 sex x drug interaction. CBD dose = 30 mg/kg. n = 4-5 mice per group.

Figure 3.
Novel object recognition memory was not altered by acute CBD (30 mg/kg) exposure but increased object exploration time in male mice only. (A) Total object (novel and familiar objects) exploration time was similar across all groups. (B) Novel object recognition memory, indicated by the discrimination index (0 = no preference, 1 = complete preference for the novel object, -1 = complete preference for the familiar object) demonstrated no significant effects. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; CBD dose = 30 mg/kg. n = 4-5 mice per group.
Figure 3.
Novel object recognition memory was not altered by acute CBD (30 mg/kg) exposure but increased object exploration time in male mice only. (A) Total object (novel and familiar objects) exploration time was similar across all groups. (B) Novel object recognition memory, indicated by the discrimination index (0 = no preference, 1 = complete preference for the novel object, -1 = complete preference for the familiar object) demonstrated no significant effects. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; CBD dose = 30 mg/kg. n = 4-5 mice per group.
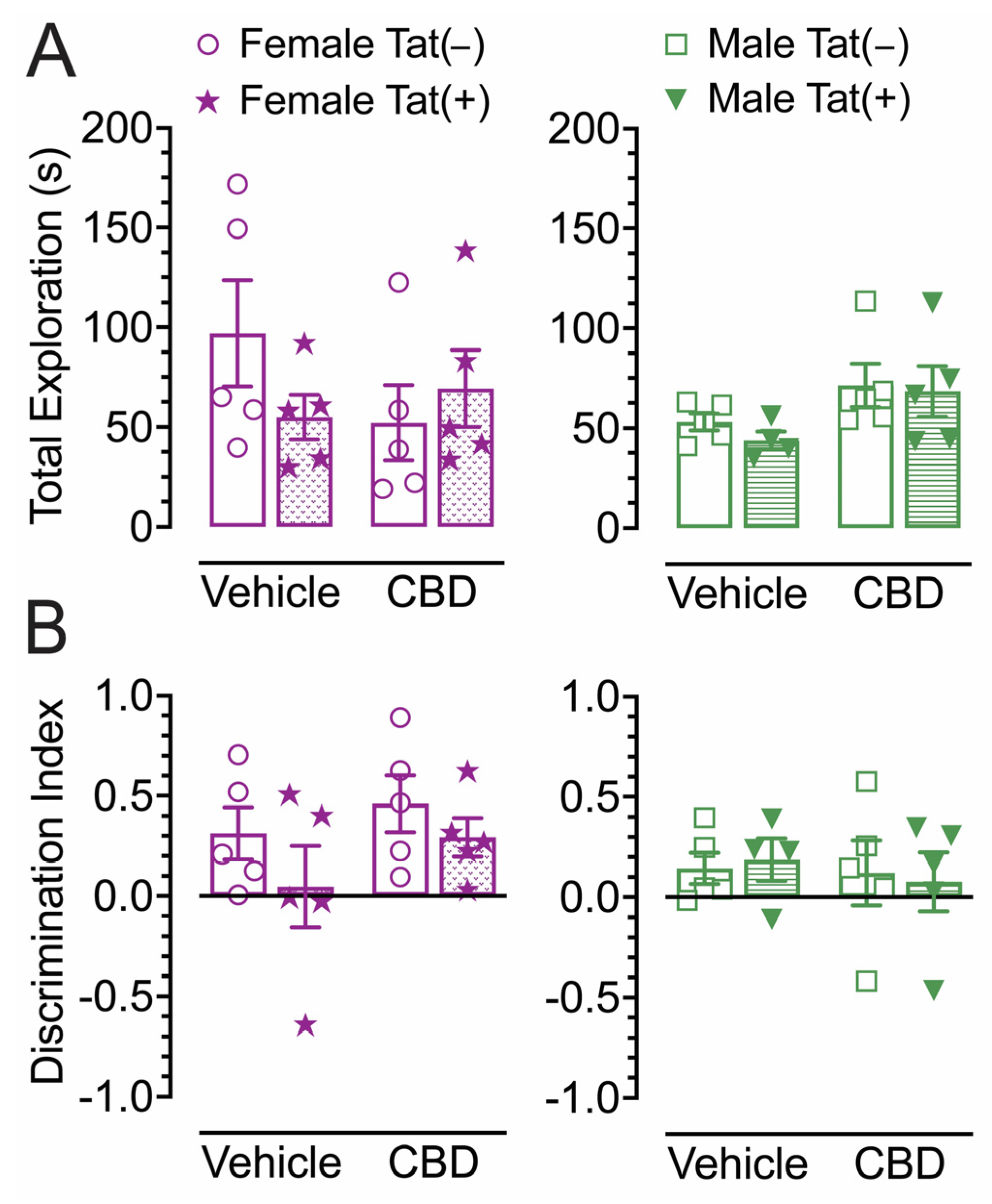
Figure 4.
2-AG, AEA, and AA levels (nmol/g) were not affected by acute CBD (30 mg/kg) exposure but 2-AG and AA demonstrated sex differences in various CNS regions. Concentrations of 2-AG, AEA, and AA were assessed in the prefrontal cortex, striatum, cerebellum, and spinal cord for vehicle and acute CBD-treated Tat transgenic mice via LC/MS/MS. Lipid concentrations were normalized to nmol/g of tissue. Note that no effects or interactions were noted for acute CBD administration on any measure. (A) 2-AG levels in the striatum and cerebellum were significantly lower in male mice compared to females. (B) AEA levels demonstrated no significant effects for any CNS region. (C) AA levels in the striatum, cerebellum, and spinal cord were significantly higher in male mice compared to females, which was noted for most of the groups. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; *p < 0.05 main effect of sex; §p = 0.039 main effect of genotype; δp = 0.009 sex x genotype interaction; ap = 0.020 Tat(+) males vs. Tat(−) males. CBD dose = 30 mg/kg. n = 4-5 mice per group.
Figure 4.
2-AG, AEA, and AA levels (nmol/g) were not affected by acute CBD (30 mg/kg) exposure but 2-AG and AA demonstrated sex differences in various CNS regions. Concentrations of 2-AG, AEA, and AA were assessed in the prefrontal cortex, striatum, cerebellum, and spinal cord for vehicle and acute CBD-treated Tat transgenic mice via LC/MS/MS. Lipid concentrations were normalized to nmol/g of tissue. Note that no effects or interactions were noted for acute CBD administration on any measure. (A) 2-AG levels in the striatum and cerebellum were significantly lower in male mice compared to females. (B) AEA levels demonstrated no significant effects for any CNS region. (C) AA levels in the striatum, cerebellum, and spinal cord were significantly higher in male mice compared to females, which was noted for most of the groups. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; *p < 0.05 main effect of sex; §p = 0.039 main effect of genotype; δp = 0.009 sex x genotype interaction; ap = 0.020 Tat(+) males vs. Tat(−) males. CBD dose = 30 mg/kg. n = 4-5 mice per group.
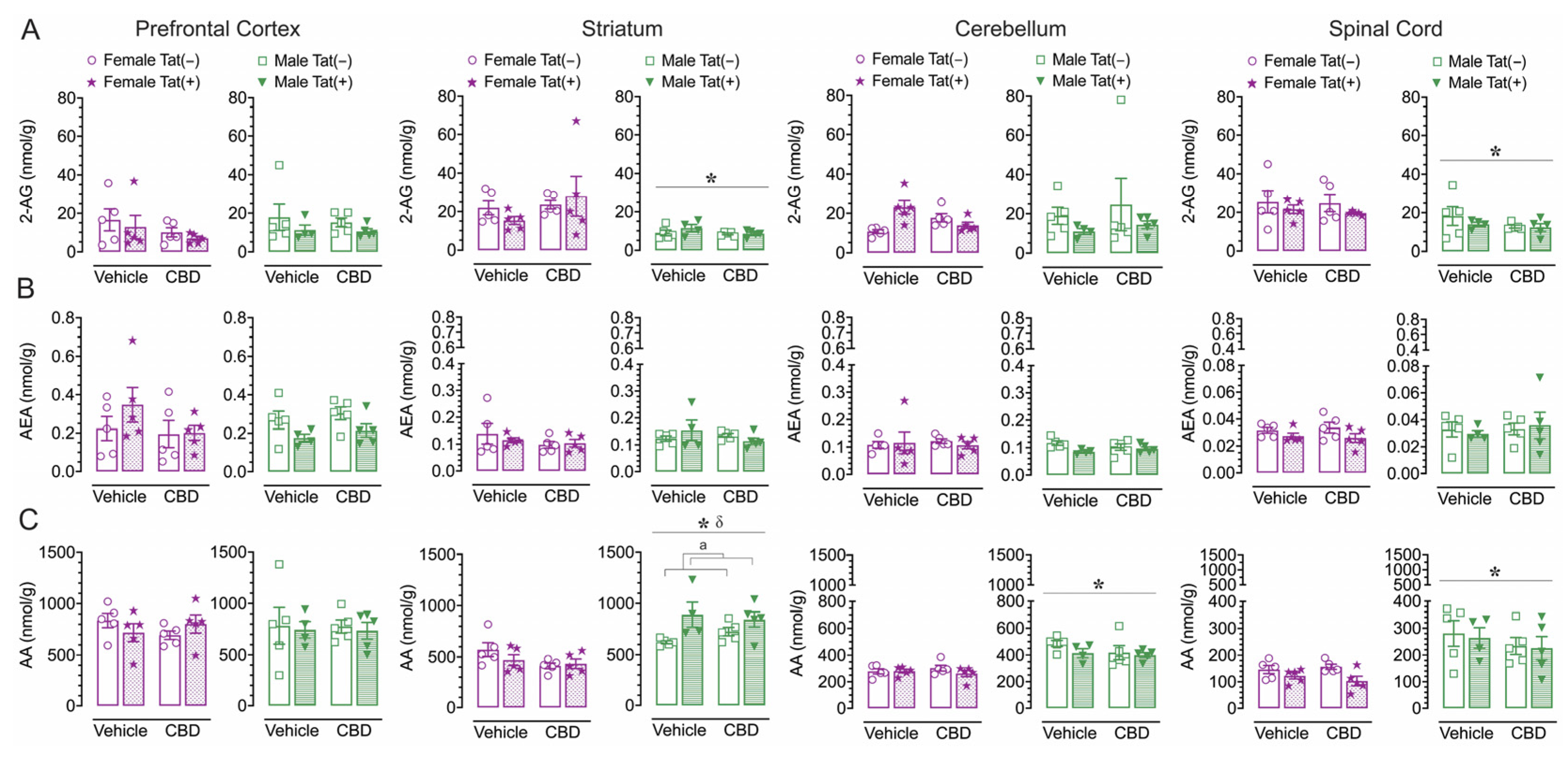
Figure 5.
CBD and the CBD metabolite CBD-7-COOH concentrations after acute CBD (30 mg/kg) exposure are altered by sex and/or genotype. Concentrations of CBD and CBD-7-COOH were assessed 60 min after 30 mg/kg CBD administration. Concentrations are expressed as ng/mL for plasma and ng/mg for cortex samples. Concentrations for all measures were higher in the plasma (A,B) compared to the cortex (C,D). (A) In the plasma, CBD concentrations were higher in males compared to females, and Tat(+) mice displayed higher CBD concentrations compared to their Tat(−) counterparts. (B) CBD-7-COOH plasma concentrations were higher in females compared to males, and Tat(+) mice displayed higher CBD-7-COOH concentrations compared to Tat(−) mice. (C) In the cortex, CBD levels were higher in males compared to female mice. (D) CBD-7-COOH concentrations in the cortex were significantly upregulated in Tat(+) mice compared to Tat(−) mice. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; *p < 0.05 main effect of sex; §p < 0.05 main effect of genotype; CBD dose = 30 mg/kg. n = 7 mice per group.
Figure 5.
CBD and the CBD metabolite CBD-7-COOH concentrations after acute CBD (30 mg/kg) exposure are altered by sex and/or genotype. Concentrations of CBD and CBD-7-COOH were assessed 60 min after 30 mg/kg CBD administration. Concentrations are expressed as ng/mL for plasma and ng/mg for cortex samples. Concentrations for all measures were higher in the plasma (A,B) compared to the cortex (C,D). (A) In the plasma, CBD concentrations were higher in males compared to females, and Tat(+) mice displayed higher CBD concentrations compared to their Tat(−) counterparts. (B) CBD-7-COOH plasma concentrations were higher in females compared to males, and Tat(+) mice displayed higher CBD-7-COOH concentrations compared to Tat(−) mice. (C) In the cortex, CBD levels were higher in males compared to female mice. (D) CBD-7-COOH concentrations in the cortex were significantly upregulated in Tat(+) mice compared to Tat(−) mice. All data are expressed as mean ± the standard error of the mean (SEM). Statistical significance was assessed by ANOVAs followed by Bonferroni’s post hoc tests when appropriate; *p < 0.05 main effect of sex; §p < 0.05 main effect of genotype; CBD dose = 30 mg/kg. n = 7 mice per group.
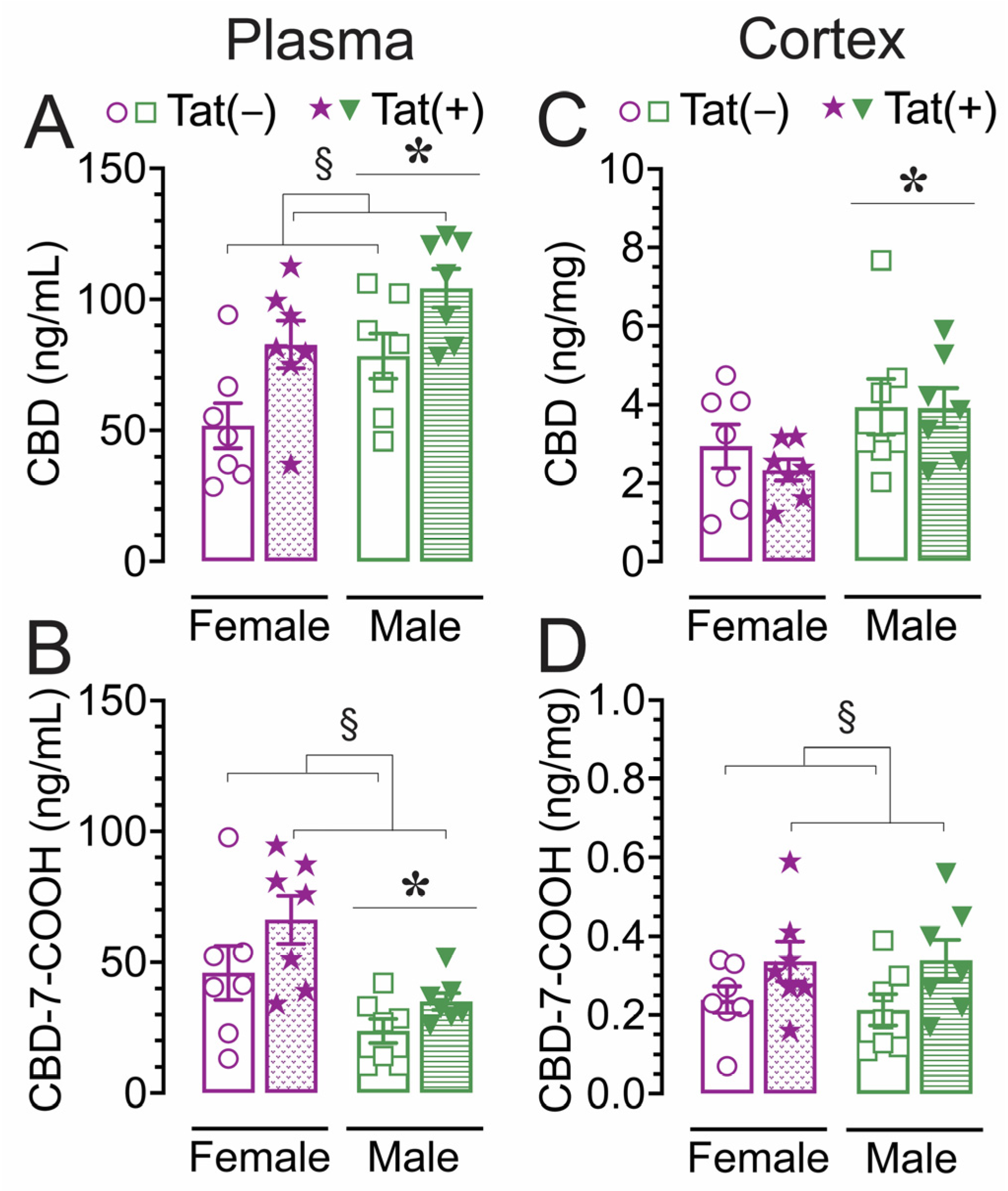
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



