
Submitted:
17 October 2023
Posted:
18 October 2023
You are already at the latest version
Abstract
Nuclear bodies (NBs) are dynamic structures present in eukaryotic cell nuclei. They are not bounded by membranes and are often considered biomolecular condensates, defined structurally and functionally by the localisation of core components. Nuclear architecture can be reorganised during normal cellular processes such as the cell cycle as well as in response to cellular stress. Many plant and animal viruses target their proteins to NBs, in some cases triggering their structural disruption and redistribution. Although not all such interactions have been well characterised, subversion of NBs and their functions may form a key part of the life cycle of eukaryotic viruses that require the nucleus for their replication. This review will focus on Cajal bodies (CBs) and the viruses that target them. Since CBs are dynamic structures, other NBs (principally nucleoli and promyelocytic leukaemia, PML, bodies), whose components interact with CBs, will also be considered. As well as providing important insights into key virus: host cell interactions, studies on Cajal and associated NBs may identify novel cellular targets for development of anti-viral compounds.
Keywords:
nuclear bodies
; cajal bodies
; PML bodies
; nucleolus
; snRNPs
; animal viruses
; plant viruses
; nuclear remodelling
1. Introduction
The eukaryotic cell nucleus is a highly organised yet dynamic intracellular environment in which most cellular DNA is condensed into chromatin and assembled into chromosomes. In addition, there are many structures that reside within the nucleus, known as nuclear bodies (NBs; Figure 1). NBs perform multiple functions, several of which are associated with gene expression and DNA replication. NBs are not bounded by membranes but are defined, both functionally and structurally, by the localisation and enrichment of nuclear factors. They have been referred to as biomolecular condensates [1]. For example, the Cajal body (CB) is defined by the presence of several key components, including p80-coilin [2,3,4] , Survival of Motor Neuron 1 (SMN [5]), TOE1 (Target of EGR1) [6]) and WRAP53 (WD40 encoding RNA Antisense to p53) [7]). Functional links between CBs and other NBs exist. For example, fibrillarin and Nopp140 are present in both NBs and nucleoli and Nopp140 mediates shuttling of proteins between the two bodies [8,9,10,11].
CBs are targeted during infection by a number of plant and animal viruses, often in conjunction with the targeting of nucleoli and other NBs such as PML (Promyelocytic Leukaemia) bodies and nuclear speckles. Viruses frequently sequester, re-organise or degrade host cell components to facilitate their replication and establish systemic infections [12]. Furthermore, viruses may target NBs to inactivate anti-virus functions that they may possess. This would be beneficial for the virus, for example in the regulation of apoptosis by PML bodies [13]. Therefore, by studying the structure and function of NBs during virus infections, it may be possible gain insights into the functions of NBs in uninfected cells. A deeper understanding of the interaction between viruses and NBs may open new therapeutic approaches to infections by targeting key cellular components or structures that facilitate virus replication.
In this review, current research on NBs will be considered, with a particular focus on CBs and the viruses that target them. There is an often-recurring motif of nuclear architecture disruption in virus-infected cells, particularly of CBs, which provides evidence for functional associations between NBs and infection in virus-infected eukaryotic cells.
1.1. Cajal Bodies
Cajal Bodies (CBs), also termed coiled bodies, were first discovered and characterised as a nucleolar accessory bodies (named due to their localisation with the nucleolar periphery) in vertebrate neuronal cells, first described by Ramon y Cajal in 1903 [14]. CBs are localised in the nucleoplasm and are approx. 1µm in diameter. In the light or electron microscope, CBs appear as a structured tangle of fibrillar threads [15]. There are generally between 1-6 CBs per cell, dependent on the cell type [16]. CBs are ubiquitous in foetal tissues. In these tissues they display greater distinction from the closely-associated NB, Gemini of coiled bodies (gems), whereas in later development or in adult tissues the two bodies are more closely associated and, in some cases, indistinguishable [17]. However, both CBs and gems are less common in adult tissues. Their abundance seems to decrease with differentiation, and during later development they are more apparent in metabolically-active cells [18]. CBs have been detected in motor neuron cells, where they have a larger volume. However, they are present in all cells of the central nervous system except blood vessels. They have been detected in epithelial cells of the liver, pancreas, duodenum and oesophagus, skeletal muscle, Islets of Langerhans, as well as liver parenchymal cells [18].
CBs are mobile bodies and traverse the nucleoplasm of both plant and animal cell nuclei, including to and from the nucleolus. They are capable of both fusing together and dividing into smaller bodies [19,20]. This discovery infers the existence of a regulatory or feedback system that functions to determine the volume and number of CBs in each nucleus. The volume and number of CBs per cell is also dependent on the stage of the cell cycle. For example, CBs are most abundant in the late G1 phase but appear smaller in volume. In the S and G2 phases, the numerous smaller CBs that exist in G1 appear to fuse into fewer, larger, CBs. During mitosis (M phase) no CBs are observed, although levels of cellular p80-coilin remain constant [21]. This suggests that CBs disassemble during M phase and reassemble at the beginning of interphase.
The assembly of CBs is dependent on VRK1, an abundant nuclear serine-threonine protein kinase. VRK1 phosphorylates p80-coilin at two serine residues, Ser184 and Ser489. VRK1-mediated phosphorylation protects p80-coilin from ubiquitination and subsequent proteasomal degradation following CB disassembly during mitosis [22].
CBs respond to cellular stress and can be disrupted by UV-C irradiation, causing some of their components e.g., p80-coilin and snRNPs to be displaced and re-distributed in the nucleus. However, some factors, including the fibrillarin and Nopp140 proteins remain in CBs under these conditions [23]. CBs are also linked to ageing by their co-localisation with telomerase RNA, which is a key component of the telomerase complex (TERC), particularly during S phase [16]. TERC is a ribonucleoprotein enzyme responsible for stabilising chromosomes by adding DNA repeat sequences (telomere repeats) to their ends. Shortening of telomeres over cell generations due to reduction in TERC activity is critical for the process of cell senescence in normal cells, a process that is subverted in cancer cells [24].
Plant CBs contain several homologues of animal CB components [25], such as p80-coilin [2], small nuclear RNP (snRNP) maturation and splicing factors such as SMN [26] and small nucleolar RNP (snoRNP) maturation factors such as fibrillarin [27]. Furthermore, plant CBs facilitate different aspects of RNA processing, including assembly and modification of the spliceosome [28,29], as well as containing small Cajal body-specific RNAs (scaRNAs) and small nucleolar RNAs (snoRNAs) that function in RNA metabolism which affects plant growth and development [30]. In Arabidopsis thaliana, components of the plant gene silencing machinery, such as Argonaute 4, co-localise with CBs, suggesting a role for CBs in the siRNA transcriptional silencing pathway [31]. The dynamics and mobility of CBs also exist in plants. In A. thaliana and tobacco BY-2 cells, CBs fuse, fragment and traverse the nucleoplasm in a similar fashion to animal cells [19].
1.2. The Nucleolus
The nucleolus comprises three compartments, the fibrillar centre (FC), the dense fibrillar component (DFC) and the granular component (GC) [32]. Genes that encode ribosomal RNAs (rDNA) and their rRNA transcripts, as well as many nucleolar proteins are localised within nucleoli [33]. The three compartments and associated proteins correspond to the major functions of the nucleolus. For example, pre-rRNA is transcribed from rDNA in the FC or at the border between the FC and DFC [34]. Thus, the RNA polymerase I machinery, including DNA topoisomerase and RNA polymerase I is localised in the FC [35,36]. Pre-rRNA processing factors associate with the DFC, including fibrillarin [37] and the proteins of snoRNPs. The GC envelopes the FC and DFC and it is here where the predominant function of the nucleolus occurs, as the site of pre-ribosomal subunit biogenesis [34]. Chromatin localises with the nucleolus or perinucleolar regions, however this co-localisation of chromatin may reflect a functional role for the nucleolus in silencing gene transcription at particular loci [38].
The nucleolus has also been linked with ageing [39], cell cycle control [40,41], preventing initiation of DNA replication following the cellular stress response and the generation of RNPs [42,43,44]. A regulatory role for the nucleolus in DNA damage repair has been proposed, as well as in other cellular and nuclear processes [45]. During cellular stress caused by DNA damage, the nucleolus undergoes segregation of its nucleolar components. This was first observed in cells in which Topoisomerase II was chemically inhibited, resulting in the reduction of rRNA synthesis [46]. Nucleolar segregation (the process of movement of nucleolar proteins into the nucleoplasm following cellular stress) was subsequently observed in cells in which DNA had been damaged by UV-irradiation [47]. The nucleolus was further linked to apoptosis in UV-irradiated cells [48]. Rearrangement of nucleolar architecture has also been associated with cell cycle checkpoints. For example, prior to mitosis, the nucleolus is disassembled and then reassembled at the end of mitosis by the re-localisation of its core components [49].
1.3. Promyelocytic leukaemia (PML) bodies
Promyelocytic leukaemia (PML) bodies are composed of an outer insoluble scaffold surrounding an inner core of over 50 constitutively and transiently associated proteins, including the protein product of the p53 tumour suppressor gene. The PML protein accumulates in PML bodies and is often used as a marker protein for this NB [50]. PML bodies cannot assemble in the absence of SUMO-1-modified PML protein, which is essential for the co-localisation of several proteins with PML bodies [51]. PML belongs to a family of proteins that are defined by the presence of an RBCC (RING-B-Box-Coiled-coil) motif [52]. This motif consists of a RING finger (C3HC4 zinc finger), which forms tetramer torus structures, B-boxes composed of 1 or 2 cysteine rich regions and a coiled-coil formed from a leucine chain [13,51].
PML bodies respond to cellular stress by undergoing changes in their native architecture and in so doing regulate stress-induced sumoylation, the process whereby a small ubiquitin-like modifier (SUMO) protein is covalently attached to target proteins, altering their biological functions [53]. Ablation of PML in mice has shown that PML is a regulator of haemopoietic differentiation, growth and tumorigenesis. This discovery led to the PML protein being recognised as a tumour suppressor, specifically in acute promyelocytic leukaemia, due to the close association of the PML gene with the retinoic acid receptor-alpha (RAR-α) gene in this disease type [13]. PML is also critical for multiple apoptotic pathways including Fas and caspase-dependent DNA-damage induced apoptosis [54]. PML bodies also have a functional role in antimicrobial defence, since biosynthesis of the PML protein is induced by all the major Interferon (IFN) types (α, β and γ), and over-expression of PML inhibits the replication of several viruses, including influenza and vesicular stomatitis viruses [55].
1.4. Components of Cajal Bodies
1.4.1. p80-coilin
CBs are characterised by the enrichment of a marker protein p80-coilin (a nuclear autoantigen of molecular mass 80kDa in humans) [56,57]. Although p80-coilin is present in the nucleoplasm, antibody staining for p80-coilin and subsequent immuno-electron microscopy showed that the protein localises to distinct bodies, corresponding to CBs [56]. Germline knockout of p80-coilin in mice [4], zebrafish [3] and A. thaliana [2] revealed that p80-coilin has an important role in assembly and integrity of CBs, as well as for growth and development of the organism. For example, during gestation in p80-coilin knockout mice, there was prenatal semi-lethality in homozygous animals, which suggested that, while p80-coilin is important for development, it is not an essential protein the mouse [58]. In addition, as previously mentioned, p80-coilin is phosphorylated at two amino acids during mitosis when CBs are disassembled, which further indicates that p80-coilin is associated with the structural integrity of CBs [59].
p80-coilin is a self-interacting protein, with serine phosphorylation sites and two nuclear localisation signals (NLS) located in the central domain. However, these consensus targeting motifs are not sufficient to target p80-coilin to CBs but are responsible for targeting p80-coilin to the nucleus [60]. The self-interacting domain of p80-coilin has been mapped to within the N-terminal 92 amino acids. p80-coilin self-interaction activity is reduced by hyperphosphorylation of the p80-coilin protein [61]. Furthermore, p80-coilin has a cryptic Nucleolar Localisation Signal (NoLS) (Figure 2), which may be indicative of the protein’s involvement in a specific, as yet undefined, cellular response, which is also regulated by phosphorylation [61]. The C-terminus of p80-coilin is conserved and folded into a Tudor domain. Tudor domains have been shown to have preferential binding to DNA, RNA and modified amino acids. p80-coilin also contains an RG-box, a region enriched in arginine and glycine (Figure 2). The RG-box has been shown to form direct interactions with the survival of motor neuron (SMN) protein, thus localising SMN to CBs [62].
1.4.2. SMN
The SMN protein is encoded by the Survival of Motor Neuron 1 gene (SMN1) and, despite its name, is found in all cells [63]. Gems (gemini of coiled bodies). which often co-localise with CBs, particularly in non-foetal tissues, are characterised by the enrichment of SMN protein [5]. SMN is also localised in CBs in many cell types [18]. The SMN protein is critical for the formation of the SMN complex which, along with seven Gemin proteins, is required for spliceosomal snRNP maturation [64]. This might explain the functional and spatial overlapping of snRNP-enriched Cajal bodies with gems. SMN and the SMN complex function in the pre-mRNA splicing cycle and splicing complex recycling [65]. Over-expression of SMN leads to the disruption of CBs, suggesting a potential regulatory function for the protein in defining the number and volume of CBs within the nucleus [66].
Reduced abundance of SMN protein results in the neuromuscular disorder Spinal Muscular Atrophy (SMA) [67]. The disorder is caused by a mutation in, or homozygous deletion of the SMN1 gene. Symptomatic phenotypes in individuals suffering with SMA can be severe. This includes loss of motor neurons, muscle weakness, immobility, respiratory failure and death. It is thought that SMA results from a disturbance in snRNP assembly, caused by loss of SMN chaperoning capabilities [67]. Furthermore, it has been demonstrated that SMN directly interacts with p53, both of which colocalise with CBs, suggesting the potential for an additional role for CBs in the regulation of cellular stress [68].
1.4.3. TOE1
Target of Egr1 (TOE1) is a highly conserved protein with roles in cell proliferation (maintenance of cellular p21, an inhibitor of cell proliferation) [6] and is a target of Early Growth Response 1 (EGR1) protein [69]. It co-localises with p80-coilin and SMN in CBs. TOE1 is important for the integrity of CBs, acting in concert with p80-coilin and TOE1 knockdown resulted in reduced SMN recruitment to CBs, with the protein instead accumulating in cytoplasmic foci. This suggests that TOE1 may be involved in the localisation of SMN to CBs. TOE1-deficient cells also showed reduced cell proliferation and splicing capabilities [6].
1.4.4. WRAP53
The WD40 encoding RNA Antisense to p53 (WRAP53) gene encodes a natural antisense transcript of p53, (WRAP53a), that regulates endogenous p53 mRNA [70], as well as a scaffold WD40 protein, WRAP53P (also known as WRAP53). This latter protein functions to guide factors to CBs as well as to telomeres and DNA double-strand breaks [70,71,72]. WRAP53 is critical for CB maintenance as well as directing SMN to CBs. Knockdown of WRAP53 was disruptive to existing CBs in the nucleus, preventing assembly of new CBs, and resulted in the relocation of SMN and p80-coilin to the nucleolus. This highlights the importance of WRAP53 to the integrity of CBs [7]. Overexpression of WRAP53 is disruptive to CBs, resulting in their disassembly and the displacement of p80-coilin and SMN to the nucleoplasm [7].
1.4.5. Nopp140 and fibrillarin
Nopp140 (Nucleolar phosphoprotein 140) and fibrillarin are protein components of CBs. p80-coilin directly interacts with Nopp140, which shuttles between the nucleolus and CBs, as well as between the nucleolus and the cytoplasm, inferring a dynamic relationship between the nucleolus and CBs [8,10]. Nopp140, along with fibrillarin, is also found in the nucleolus, which suggests a functional relationship between the two nuclear bodies. Fibrillarin, a highly conserved nucleolar protein, interacts with the SMN protein [9,11]. Furthermore, both Nopp140 and fibrillarin interact with snoRNPs, with fibrillarin binding C/D box snoRNAs and Nopp140 binding C/D and H/ACA snoRNAs [16].
1.4.6. snRNPs, snoRNPs and scaRNPs
Almost all small non-coding RNAs (snRNAs) present in the cell, except tRNAs and miRNAs, directly interact with p80-coilin [73]. It is widely accepted that CBs are critical in the biogenesis of snRNPs, however, beyond this function, the association with snRNAs implies that CBs may also act as a central hub or coordinator for several processes in RNA metabolism.
Spliceosomal snRNPs are closely associated with CBs in both plant and animal cells. They are composed of a small nuclear RNA, (of approx. 150 nucleotides), 1-12 specific proteins per snRNP and an Sm protein, or Sm-like protein, in a heptameric ring. Currently, five major snRNPs (U1, U2, U4, U5 and U6) and four minor snRNPs (U11, U12, U4atac and U6atac) have been identified based on the presence of the snRNA [74]. The intracellular distribution of snRNPs, detected using fluorescently-labelled antisense RNA probes, revealed co-localisation of snRNPs and snRNAs with specific nuclear foci that were later defined as CBs. Certain snRNPs also co-localise with the nucleolus. Furthermore, U1 snRNA is distributed throughout the nucleoplasm and, although it co-localises with CBs, it is not concentrated there [75,76]. The two major modifications to snRNAs are pseudouridylation and 2'-O-methylation. These modifications are guided by two classes of snoRNPs, which are defined by their snoRNA component. Box H/ACA snoRNPs contain dyskerin that completes pseudouridylation and C/D snoRNPs contain fibrillarin, which effects ribose methylation [77].
Small CB-specific RNAs (scaRNAs) form a family of eight snRNAs that co-localise specifically with CBs. They direct the pseudouridylation and ribose methylation modifications of U1, U2, U4 and U5 snRNAs, which are transcribed by RNA polymerase II [78,79,80]. In contrast, U6 RNA, transcribed by RNA polymerase III, undergoes the same modifications in the nucleolus by alternative snRNAs [81,82]. The H/ACA scaRNAs (and C/D-H/ACA mixed domain scaRNAs) are targeted to CBs by the cis-acting sequence, the CAB box, of which H/ACA has two copies [83]. Furthermore, a conserved WD40 protein, originally isolated in Drosophila melanogaster, termed WDR79 in humans (also known as WRAP53: see section 1.4.4), is required for the localisation of scaRNAs in CBs. It functions by binding to cellular scaRNAs, acting in a CAB box-dependent interaction [84].
snRNP biogenesis is a multi-step process that takes place in the cytoplasm and nucleus of animal and plant cells. The complete pathway has been reviewed [85]. snRNPs colocalise in CBs during their generation cycle, prior to translation, processing and assembly of the SMN complex in the cytoplasm. Furthermore, the integrator complex, which is involved in the initial cleavage to form pre-snRNA and terminal 3’-end processing of snRNAs [86], is critical for CB integrity and homeostasis [87]. Following assembly, the import complex is transported into the nucleus where it again passes through CBs [85,88]. This localisation to CBs following import coincides with modifications of snRNPs, including 2'-O-methylation and pseudouridylation, implying a functional role for CBs in the biogenesis pathway, a process guided by scaRNAs [89]. By binding its associated proteins, CBs facilitate snRNP assembly by bringing the factors in close enough proximity to interact. CBs are involved in the generation of U4/U6 di-snRNPs and U4/U6.U5 tri-snRNPs [85]. The integrator complex mediates processing of snRNAs, An activity-deficient integrator complex had a negative impact on the co-localisation of CB core components. This included a depletion of snRNPs in CBs, the re-localisation of some p80-coilin to the nucleoplasm (subsequently forming CB-like foci) or nucleoli and SMN to the cytoplasm [87].
snRNP biogenesis is linked to growth and development as well as stress responses in plants. An example of this is the diverse developmental-defective phenotypes observed in snRNP-mutant A. thaliana, including effects on female gametogenesis and flowering, as well as enhanced stress sensitivity [90].
2. Viruses that Interact with Nuclear Bodies
There is much yet to be discovered concerning the function and mechanisms of NBs and how they contribute to the complex system of nuclear and cellular functions. In both animals and plants, certain viruses target these bodies, pointing towards potentially conserved cellular targets of diverse viruses. Investigation into the mechanism(s) of action and functional consequences of virus targeting of NBs may provide further insights into our understanding of the cell biology of NBs, virus: host cell interactions and the pathogenesis of infections.
2.1. Animal Viruses interacting with Nuclear Bodies
This section will discuss how viruses that infect members of the animal kingdom interact with NBs, in particular with Cajal bodies. Viruses that interact with Cajal bodies and other NBs are summarised in Table 1.
2.2. Animal Viruses that target Cajal Bodies
2.2.1. Minute virus of Mice
Minute Virus of Mice (MVM) is an autonomous parvovirus that possesses a linear ssDNA genome. The genome encodes three major classes of mRNA, R1-3. The virus capsid proteins VP-1 and VP-2/3 are encoded by R3 while the two non-structural polypeptides, NS1 and NS2, are encoded by R1 [91,92]. MVM, in a similar manner to the related parvovirus H-1 [93], has the capability to form novel subnuclear structures, termed autonomous parvovirus- associated replication (APAR) bodies during infection, which are distinct from host nuclear bodies. They appear to be sites of virus replication due to the co-localisation and accumulation of replication factors [94]. NS1 specifically interacts with host SMN protein and co-localises with CBs [95]. This interaction was detected late in infection, coinciding with major virus-induced nuclear reorganisation, including components from CBs and APAR bodies, which reconstitute into multiple large nuclear bodies termed SMN-associated APAR bodies (SAABs). NS1 is critical for virus replication and transcription regulation and its accumulation in SAABs suggests that these bodies may be important in the virus life cycle and/or may indicate the initiation of cell death mediated by the virus via NS1 cytotoxicity [95].
2.2.2. African Swine Fever Virus
African Swine Fever Virus (ASFV) is the only member of the Asfaviridae genus within the Nucleocytoplasmic Large DNA Viruses superfamily and causes severe disease in pigs [96]. It possesses a dsDNA genome containing between 150 and 167 ORFs [97]. As well as other pathogenic effects within the host nucleus, ASFV infection causes the structural rearrangement of host NBs. In terms of nuclear speckles, ASFV infection results in their enlargement and reduction in number per nucleus [96]. Depletion of PML resulted in mal-formed virus factories and a reduction in progeny virus, indicating that PML facilitates ASFV infection. In addition, PML bodies were disrupted during virus infection, increasing in size and decreasing in number [96]. CBs are remodelled during infection by ASFV, being fragmented into a greater number of ‘comma-shaped’ bodies [96]. This virus-induced structural reorganisation of host nuclear bodies may contribute to ASFV control of host transcriptional machinery [96,98].
2.3. Human Viruses interacting with Cajal Body components to induce anti-virus defence
Human Immunodeficiency Virus
Human Immunodeficiency Virus (HIV) isolates are classified into two types, HIV-1 and HIV-2, which differ in the organisation of their genome, with HIV-1 being the most prevalent worldwide. It is a member of the Lentivirus genus within the Retroviridae family. The HIV genome comprises two identical positive-sense ssRNA molecules [99]. TOE1, a conserved multifunctional nuclear protein that is localised to CBs, inhibits replication of HIV-1 at the transcriptional level. This occurs by direct interaction between TOE1 and HIV-1 TAR, thus interfering with Tat transactivation, which is critical for the virus RNA elongation step of transcription. This interaction occurs via a Lysine/Arginine-rich NLS domain in TOE1. It has been proposed that TOE1 acts to competitively bind TAR in place of Tat to inhibit RNA transcription of the virus genome [100]. TOE1 has also been linked to the host CD8+ T cell mediated immune response [100].
2.4. Human viruses interacting with Cajal Body components
2.4.1. Influenza a Virus
Influenza A virus (a member of the Orthomyxoviridae family) is one type of influenza virus (the other less common types being B and C) that cause recurrent epidemics and occasional pandemics in humans and other mammalian and avian species. Their persistence in these populations is largely due to continuous evolution of the viruses, which is driven in Influenza A by the natural virus gene reservoir in aquatic birds [101]. As in other influenza viruses, influenza A has a negative-sense ssRNA genome, which is segmented into eight RNAs that exist as virus ribonucleoproteins (vRNPs) in the virus capsid [101,102,103] . Replication and transcription of the virus genome takes places in the host nucleus, by sequestering components of the nuclear machinery [104]. Influenza A targets NBs in several ways, including the accumulation of non-structural protein 1 (NS1) and matrix protein (M1) in PML bodies in order to interfere with the host interferon-induced anti-virus pathway, thus suppressing apoptosis [105]. Over-expression of the PML protein resulted in cellular resistance to influenza virus infection [106]. The host nucleolus is remodelled during influenza A infection, in association with virus replication [107,108,109]. This restructuring of the nucleolus was found to be mediated by the NS1 and M1 virus proteins, and may subvert host nucleolar functions [109]. Influenza A nucleoprotein (NP) and NS1, the latter of which interacts with nucleolin, localise to the nucleolus at different times during infection, via nucleolar localisation signals (NoLs) present within the virus proteins [110,111,112]. As well as co-localisation with nucleoli, NP has been shown to tightly associate with regions of the nucleus adjacent to PML bodies and CBs [102]. Influenza A virus infection causes an increase in number of CBs per nucleus and a concurrent reduction in their size [113]. Influenza A NP co-localises with CBs, tightly associating with them, and in doing so follows the same rearrangement as CBs. It was proposed that this association between NP and nucleoli and CBs, as well as the relative fragmentation of both, may be related to trafficking between the two NBs or there may be a further functional link [102].
2.4.2. Zika Virus
Zika virus (ZIKV) is an arthropod-borne virus (arbovirus) within the Flaviviridae family (Flavivirus genus) of viruses and has a positive-sense ssRNA genome [114]. The virus RNA genome encodes a single polypeptide, which is subsequently proteolytically cleaved to produce three structural proteins (capsid (C), envelope (E) and membrane precursor (prM)) and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5) [115]. ZIKV proteins target cytoplasmic bodies, including the endoplasmic reticulum and lysosomes, as well as nuclear bodies [114]. The ZIKV NS5 protein associates with host CBs via interaction with p80-coilin, causing disruption of CBs [114]. Furthermore, NS5 expression coincided with an increase in the number of CBs per nucleus and a corresponding reduction in their volume [114].
2.4.3. Adenoviruses
There are at least 100 types of human adenoviruses (Ads), classified into seven species, A-G [116]. The adenovirus genome is a linear double stranded DNA molecule. It encodes five early transcriptional units (E1A, E1B, E2, E3 and E4), four intermediate transcription units (IX, IVa2, L4 intermediate and E2 late) and one major late transcriptional unit (MLTU). Five families of late mRNAs (L1-5) are generated from a single pre-mRNA by differential pre-mRNA splicing. A common tripartite leader sequence is spliced to the coding regions for mRNAs encoding structural proteins of the virus and non-structural proteins that have functions in virus assembly and transcriptional regulation [116].
Ad5 infection results in the redistribution of CBs into smaller foci termed rosettes or microfoci, as defined by immunofluorescence with anti-p80 coilin [117,118,119]. The microfoci appear to be unique structures. They occur adjacent to, but do not co-localise with virus DNA replication centres (containing the virus E2-72K protein) or with areas of virus pre-mRNA splicing and mRNA export (containing the ASF/SF2 protein) [118] or with with Aly, a component of the major mRNA export TREX complex [119]. The formation of these CB microfoci takes place at the onset of Ad5 DNA replication and at the intermediate/late phase of infection; blocking of Ad DNA replication prevents CB reorganisation [118]. CBs are associated with a population of nuclear actin in uninfected cells and in the early phase of Ad5 infection. However, late in Ad5 infection, actin disassociates from CBs as they are rearranged, suggesting that actin is involved in the formation of CB microfoci [120]. p80-coilin facilitates Ad5 infection as, following p80-coilin siRNA-mediated reduction, virus yield is reduced by up to 80% and the levels of most virus proteins are reduced in p80-coilin siRNA-treated cells compared to controls [119]. The underlying mechanism in reduction of virus protein and virus production in p80-coilin-depleted cells appears to be a decrease in export of Ad5 mRNA transcripts from the nucleus to the cytoplasm [119].
Immunofluorescent antibody staining of Ad5-infected cells revealed co-localisation of p80-coilin in microfoci with areas of immunoreactivity defined by an antibody that recognised the virus L4-22K and L-33K proteins. The L4-22K protein shares an N-terminal domain with L4-33K but has a unique carboxy-terminal region due to differential pre-mRNA splicing. The L4-22K protein has multiple functions in Ad infection, including transcriptional and post-transcriptional regulation of virus gene expression [121,122]. Co-immunoprecipitation analysis showed that only the L4-22K protein formed a stable complex with p80-coilin in Ad5-infected cells and in cells co-transfected with plasmids encoding epitope-tagged p80-coilin and either the L4-22K or -33K protein [119].
Overall, the data described above suggest that, during the late phase of Ad5 infection, CBs are reorganised into microfoci by a currently unknown mechanism involving nuclear actin. One consequence of production of microfoci may be to increase the surface area of the CB and/or to release CB proteins such as p80-coilin into the nucleoplasm. The virus L4-22K protein forms a complex with p80-coilin, potentially facilitating and directing virus transcripts to complexes such as Aly/Ref/TREX or CRM1 for mRNA export [119].
PML bodies are reorganised in Ad-infected cells [123]. The two major E1A proteins co-localise with PML bodies in Ad-infected cells [123]. Furthermore, the E4-ORF3-encoded 11kDa protein appears to be the sole protein responsible for the reorganisation of PML bodies during infection [123]. E4-ORF3-dependent rearrangement of PML bodies has been shown to activate PML-dependent, interferon-induced anti-virus innate immune defence [124]. Infection of cells by Ad5 also induces the redistribution of the nucleolus, involving the displacement of chromatin and several nucleolar components, including fibrillarin [117].
2.4.4. Herpesviruses
The Herpesviruses form a large group of DNA viruses causing a range of infections in humans and animals. They are divided into three sub-families, Alpha (e.g. herpes simplex virus 1, HSV-1), Beta (e.g. Cytomegalovirus, CMV) and Gamma (Epstein-Barr virus, EBV), all of which have a large coding capacity of between 80 and 200 proteins. They can establish latent and lytic infections and have different cell tropisms and lifecycles. In a large-scale, genome-wide screen of proteins encoded by these viruses, several expressed ORFs were found to associate with or disrupt CBs, PML bodies or nucleoli [125]. Ninety-three virus proteins out of 269 recovered as molecular clones encoded by the three viruses were located in the nucleus. No virus protein associated with CBs, however cells expressing the CMV UL3 or UL30 ORFs contained significantly fewer CBs compared to control cells, suggesting that these virus gene products disrupt CBs. The avian alphaherpesvirus, MDV (Marek’s disease virus) encodes oncogenic proteins Meq and a fusion of Meq with a virus homologue of interleukin 8, Meq/vIL8. In cells transfected with either construct (expressed as a fluorescent fusion protein), around 20% of cells showed co-localisation of p80-coilin with either Meq or Meq/vIL8, with generalised nuclear or nucleolar localisation found in the remainder of the cells [126,127]. The CMV UL3, UL30 and the Meq gene products associated with or reorganised PML bodies, providing further evidence of the inter-relationships between these two NBs [125,126,127].
2.5. Plant Viruses interacting with Nuclear Bodies
This section will discuss how viruses that infect members of the plant kingdom interact with NBs, in particular with Cajal bodies. Viruses that interact with Cajal bodies and other NBs are summarised in Table 2.
2.5.1. Virus interactions with the Cajal Body component, Fibrillarin
Potato Virus A
Potato Virus A (PVA) belongs to the Potyvirus genus (family Potyviridae). It is part of a large group of positive- sense ssRNA viruses like picornaviruses in animals. In order to generate proteins, of which they can produce up to ten, a large polyprotein is translated and cleaved by three virus-encoded proteinases [128]. The multifunctional virus protein Nuclear inclusion protein-a (NIa) is targeted to CBs [128]. NIa is conserved among several other plant viruses including Tobacco Etch virus (TEV), also belonging to the Potyviridae family [129]. NIa is a polyprotein with a C-terminal proteinase domain and an N-terminal virus genome-linked protein (VPg) domain, the latter of which contains the NLS required for co-localisation of the protein with the host nucleus [129]. In PVA, the VPg domain of NIa was found to contain both nuclear and nucleolar localisation signals [128]. Furthermore, this same domain, more specifically NLS I (amino acids 4-9), was shown to target and direct the accumulation of NIa to CBs, where the VPg domain interacted with fibrillarin. The functional consequences of this interaction was demonstrated by the depletion of fibrillarin, which resulted in reduced infectivity of PVA in Nicotiana benthamiana (tobacco) plants [128].
Citrus Tristeza Virus
Citrus Tristeza Virus (CTV) is a member of the Closteroviridae family and contains a large, approx. 20kb ssRNA positive-stranded genome that encodes approx. 17 proteins. The CTVp23 protein is a major pathogenicity determinant of CTV, acting by RNA silencing suppression. CTVp23 accumulates in Cajal bodies and in nucleoli, where it co-localises with fibrillarin [130]. Expression of CTVp24 alone recapitulates many of the pathogenic properties of the whole virus and formed the basis for mutational analysis of the protein. This identified a bipartite nucleolar localisation sequence and revealed that most of the CTVp23 sequence is required for CB and nucleolar localisation [130].
Groundnut Rosette Virus
Groundnut Rosette Virus (GRV) is a member of the Umbravirus genus (Tombusviridae family), and has a genome consisting of a linear segment of positive-sense ssRNA. The genome is divided into four open reading frames (ORF1-ORF4). ORF1 and ORF2 are located at the 5’ end, slightly overlapping with each other and are translated by a frameshift mechanism as a single fusion protein. Following ORF2 is a short untranslated region, which separates it from ORF3. Finally, ORF3 and ORF4 are at located at the 3’ end and overlap almost entirely [131]. ORF4 encodes a 28kDa protein involved in cell-to-cell movement and localises to, or in close proximity to, the plasmodesmata (channels connecting neighbouring plant cells), thus fulfilling a role similar to movement proteins (MPs) in other plant viruses [132]. ORF3 encodes a 27kDa protein which is involved in long distance movement and stabilisation of virus RNA [131,133]. ORF3 may utilise existing trafficking pathways, through the nucleolus and involving the rearrangement of CBs. Once the ORF3 protein enters the nucleus it targets CBs and reorganizes them into multiple CB-like structures (CBLs), potentially by interacting with an SMN homologue [134]. This reorganisation probably occurs by displacing CB components, such as p80-coilin and fibrillarin and redistributing them with ORF3 into the new CBL structures. ORF3 then localises to the nucleolus by fusing CBLs with the nucleolus in a novel pathway [134]. This fusion of CBLs with the nucleolus relies on the interaction of ORF3 with fibrillarin, shown by silencing the fibrillarin gene, which prevented long-distance, but not cell-to-cell movement [135]. Furthermore, reduction in expression of fibrillarin caused ORF3 to accumulate in CBLs which were unable to fuse with the nucleolus [135]. However, the ORF3:fibrillarin interaction does not appear to be critical in either targeting of ORF3 to CBs or in their reorganisation [135]. There is a further function of ORF3, in which ORF3 redistributes fibrillarin to the cytoplasm in order to be used for RNP particle generation [135]. ORF3 interacts with virus RNA and subsequently forms filamentous RNP particles, protecting the virus RNA during long distance transport through the host phloem, thus aiding virus spreading in the plant [135,136].
2.5.2. Virus interaction with the Cajal Body protein p80-coilin, resulting in increased pathogenicity
Poa Semilatent Virus
Poa Semilatent Virus (PSLV) is a member of the Hordeivirus genus and has a positive sense RNA genome [137]. The genome comprises three RNAs (α, β,and γ). RNAβ encodes the first ORF encoding the coat protein, followed by several overlapping ORFs, which together form the triple gene block (TGB). The two further ORFs encode small hydrophobic proteins [138]. The first gene of the triple gene block (TGBp1) encodes the Movement Protein (MP) of PSLV, which interacts with CBs during virus infection. TGBp1, analogously to GRV ORF3, is involved in interacting with virus RNAs to form an RNP complexes for cell-to-cell movement which is also critical for long distance transport and establishing a systemic infection in the plant [139,140]. A direct interaction between p80-coilin and PSLV MP was shown both in vitro using purified recombinant proteins and in vivo, following introduction of a TGBp1 expression construct into N. benthamania plants. Deletion analysis revealed a major p80-coilin binding site in the positively-charged N-terminal domain of TGBp1 [140].
2.5.3. Virus interaction with Cajal Body marker protein p80-coilin, resulting in decreased pathogenicity
Tobacco Rattle Virus
The involvement of p80-coilin in Tobacco Rattle Virus (TRV) infection produces a different outcome than in most plant viruses in that it has a paradoxical role in host N. benthamiana anti-virus defence. TRV is a bipartite (RNA1 and RNA2) virus, belonging to the Tobravirus genus (Virgaviridae family), with a positive-sense ssRNA genome [141]. RNA1 encodes a 29-30kDa MP in the 1a ORF and a 12-16kDa cysteine-rich silencing suppressor protein (termed 16K) in the 1b ORF. Both 1a and 1b are translated from subgenomic RNA (sgRNA) [141]. RNA1 also encodes the helicase and RNA-dependent RNA polymerase (RdRp) proteins. RNA2, the smaller of the two RNAs, encodes the virus coat protein as well as several proteins involved in virus transmission to nematodes [141]. TRV-infected wild-type (WT) plants are able to recover from infection [142], suggesting the existence of a host anti-virus pathway. The host defence system comprises both RNA silencing and salicylic acid (SA) pathways, which require the interaction of the virus 16K protein and host p80-coilin. This was demonstrated by the knockdown of host p80-coilin or deletion of 16K in the virus, which prevents plant recovery and results in an exacerbation of symptoms [143]. RNA silencing is a major plant anti-virus response. In this case it was demonstrated in p80-coilin knockdown plants where, contrary to expectation, depletion of p80-coilin increased TRV accumulation and TRV-specific small interfering RNAs (siRNAs), suggesting the RNA silencing pathway is negatively affected by p80-coilin knockdown [142]. However, the discovery that RNA silencing mechanisms can persist in p80-coilin deficient plants suggested that another host anti-virus defence pathway was also involved [142]. In terms of the SA-dependent, anti-virus response against TRV, p80-coilin knockdown and/or deletion of the virus 16K resulted in similar effects, namely the accumulation of SA and expression of SA-responsive genes. This suggests that the two mechanisms are working together to produce an anti-virus response leading to plant recovery [142,143]. A stable interaction of 16K and p80-coilin has been demonstrated in vitro, as well as the nucleolar localisation of p80-coilin via interaction with 16K in WT plants [143]. A molecule, poly(ADP-ribose) polymerase 1 (PARP-1) that links the processes of SA-activated gene expression and plant resistance to TRV infection has recently been identified and can also participate in the p80-coilin:16K complex [144].
Barley Stripe Mosaic Virus
Barley Stripe Mosaic Virus (BSMV) induces plant anti-virus responses in a similar way to TRV. BSMV is a member of the Hordeivirus genus (Virgaviridae family) and has a tripartite (RNAα, β and γ) positive-sense ssRNA genome [145] encoding seven proteins [138]. RNAβ contains four ORFs (βa-d) with βa encoding for the virus coat protein [145], as well as three MPs (TGB1, TGB2 and TGB3) [146]. TGB MP expression is mediated by two sgRNAs, sgRNAβ1 and sgRNAβ2. TGB1, the virus protein which interacts with p80-coilin, is expressed from sgRNAβ1 [138]. TGB1 is a multifunctional protein, possessing RNA helicase, ATPase and RNA binding activities. It is thought to facilitate several virus processes such as cell-to-cell movement and the establishment of a systemic infection [146]. When studied in N. benthamiana, WT plants infected with BSMV showed no recovery, with mild systemic symptoms persisting [142]. However, like TRV, plants with p80-coilin knockdown presented with an increase in severity of symptoms and an increase in accumulation of BSMV, implying that p80-coilin was acting in host anti-virus defence [142]. However, in contrast to TRV, p80-coilin-dependent anti-virus defence did not appear to lead to plant recovery, but instead resulted in reduced plant susceptibility to BSMV in virus-infected leaves [142].
Rice Stripe Virus
Rice stripe virus (RSV) is member of the Tenuivirus genus and has a ssRNA genome consisting of four segments that encode at least seven proteins [147]. One of these proteins, p2, is responsible for long-range movement in the plant. Interestingly p2 is targeted to both nucleoli and CBs and interacts both physically and functionally with p80-coilin and fibrillarin without disrupting these NBs [148,149].
Grapevine red blotch-associated virus
Grapevine red blotch-associated virus (GrBaV) is a single-stranded DNA virus of the Geminivirus family and contains a 3206 nucleotide genome encoding at least six proteins [150]. The V2 ORF, when expressed as a fluorescent fusion protein in A. thaliana localises to the nucleoplasm and structures that may be CBs. On co-expression with a fluorescent fibrillarin construct, the V2 fusion protein becomes directed into nucleoli [150]. Thus, V2 may have the ability, like RSV p2, to target both CBs and nucleoli or may shuttle between these two NBs in complex with fibrillarin.
2.5.4. Targeting Argonaute 4
De novo DNA methylation at cytosine residues is a conserved, protective epigenetic mark in eukaryotic cells, mediated in plants by RNA-directed DNA methylation (RdDM) of which Argonaut 4 (AGO4) is a component [151].
Tomato Yellow Leaf Curl Virus
Tomato Yellow Leaf Curl Virus (TYLCV) is a member of the Begomovirus genus (Geminivirus family). The TYLCV genome encodes six ORFs, two on the virus sense strand (V1 and V2) and four on the complementary strand (C1-4). This genome consists of circular ssDNA [152]. Mini-chromosomes are formed during geminivirus infection by the association of virus dsDNA with host histones [153] which are methylated by host enzymes in order to mount an anti-virus defence [154]. Hypermethylation of virus DNA led to plant recovery. Plant recovery in this context is indicative of transcriptional gene silencing (TGS) and posttranscriptional TGS (PTGS) of host pathways. [155,156]. However, TYLCV protein V2 suppresses both TGS and PTGS pathways by the host, potentially via different domains, thus fulfilling an analogous role to RNA silencing suppressor proteins discovered in several other viruses [157,158]. In TYLCV-infected cells, there is an interaction between V2 and host histone deacetylase 6 (HDAC6), which resulted in interference in binding of methyltransferase 1 (MET1) to and subsequent reduction in methylation of virus DNA via TGS, thus increasing host susceptibility to TYLCV infection [159]. An additional TGS suppression capability of TYLCV V2 is present in N. benthamania cells, where V2 interacts with AGO4 and interrupts its binding to virus DNA.
TYLCV anti-virus methylation takes place in CBs. The link between CBs and TYLCV infection was further demonstrated by the co-localisation of the V2-AGO4 complex with host CBs, which was shown to be critical for the interaction between V2 and AGO4 [151]. A similar mechanism of anti-virus DNA methylation is employed by V2 of Cotton leaf curl Multan virus (CLCMV), via its interaction with AGO4, although the intra-nuclear localisation of CLCMV V2 has not been reported [160].
Cucumber mosaic virus
Cucumber mosaic virus (CMV) is a member of the Bromoviridae family and is a tripartite, ssRNA virus that encodes five proteins, of which protein 2b has host-silencing suppression functions. The 2b protein is located in Cajal bodies and nucleoli and interacts with AGO1 and 4, reducing their methylation activity and PTGS [161,162].
3. Concluding remarks: common threads and future perspectives
3.1. Nuclear Architecture can be re-modelled in similar ways by diverse viruses
Nuclear bodies (NBs) can rearrange their architecture and redistribute their core components in response to various physiological conditions. For example, in response to cellular stress or DNA damage the nucleolus undergoes nucleolar segregation [46,47]. A similar phenomenon has been demonstrated in CBs during cellular stress [23]. Likewise, PML bodies are linked to the regulation of stress-induced sumoylation by undergoing changes in their structure [53]. A number of diverse viruses disrupt Cajal bodies in analogous ways. Following infection with ASFV, influenza A virus or ZIKV, CBs increase in number while simultaneously decreasing in volume [96,113,114]. These viruses have either RNA or DNA as their genomes, they are members of different virus families and genera and have different virus lifecycles, but appear to adopt a conserved strategy of disruption of CBs.
In some virus infections, CB rearrangement does not occur in isolation, for example ASFV infection also causes the structural rearrangement of PML bodies as well as the production of enlarged nuclear speckles. ASFV infection is PML-dependent, since PML knockdown resulted in mal-formed virus factories and reduced yields of virus progeny [96], which implies that host PML bodies play an important role in virus replication. Similarly, Adenoviruses reorganise CBs, PMLs and nucleoli during infection. Fragmentation of the nucleolus takes place in Influenza A virus infection, providing a link between the nucleolus and CBs [102]. However, although influenza A virus NP is tightly associated with both the nucleolus and CBs in infected cells, it does not appear to induce their fragmentation [102]. Another possibility is that re-arrangement of NBs such as CBs and nucleoli occur in response to the cellular stress that is stimulated by virus infection, as seen in host cells [107,108,109].
GRV in plants, and MVM and adenoviruses in mammals, all cause disruption of NBs during infection, although their mechanisms for doing so follow unique pathways. For example, MVM and GRV redistribute components of CBs to form novel bodies that also contain virus proteins. In GRV infection, CBs are reorganised, displacing p80-coilin and fibrillarin into CB-like (CBL) structures that contain the virus ORF3 protein, which then fuse with the nucleolus. Fusion of CBLs with the nucleolus may facilitate the export of fibrillarin and ORF3 to the cytoplasm where they are used in the generation of virus RNPs. This process has been proposed to aid the establishment of a systemic infection [134,135].
In MVM, disruption of CBs arises following the formation of APAR (autonomous parvovirus-associated replication) bodies which recombine with nuclear components, including from CBs, into SMN-associated APAR bodies or SAABs – both of these bodies are sites of virus replication [95]. This infers a link between the rearrangement of CBs and the nucleolus in order to support replication of the virus.
Adenoviruses (Ad5) disrupt CBs in a novel manner, by redistributing their components into CB microfoci [118,119] potentially triggered by the dissociation of nuclear actin from CBs [120]. This rearrangement was observed to occur at a specific time in the virus lifecycle and is associated with late phase virus DNA replication [118]. CB microfoci localised to the periphery of virus replication centres, characterised by the localisation of virus protein E2A-72K [118]. A virus protein, L4-22K, forms a stable complex with p80-coilin and is located in some CB microfoci. This co-localisation strongly indicates that the virus is rearranging the host nuclear architecture in order to serve its own replication, likely in order to sequester critical factors present in CBs. PML bodies and the nucleolus are similarly disrupted upon infection by Ad5, displacing their core components [117,123]. It is also worth noting that the human Ad L4-22K protein, the TRV 16K, the PSLV MP and the RSV p2 proteins of plant viruses all form physical complexes with p80-coilin, suggesting a common target of diverse viruses in their infectious cycles.
Taken together, this information further strengthens a proposed link between virus appropriation of genome replication machinery and rearrangement of NBs. This link may be redistributing components of these nuclear bodies to be in close proximity to virus replication sites, thus supplying them with required factors, such as snRNPs or host proteins etc. This may be a common function of these viruses. The existence of virus proteins that are conserved between virus species, e.g., V2 in TYLCV and V2 in Cotton leaf curl Multan virus that both target AGO4 [151,160] or NIa in PVA and TEV that target CBs [129] suggests that these viruses may, at least in some cases, share a common mechanism.
3.2. Involvement of Nuclear Bodies in Anti-virus Defence
Several viruses interact with CBs and other NBs in both plant and mammalian cells, inducing a host anti-virus response. Although PML body involvement in these antimicrobial defence pathways is well documented, less is known concerning CBs. In TRV and BSMV, anti-virus defence is mediated by virus interaction with p80-coilin, resulting in recovery of the former virus and reduced severity of symptoms in the latter [142]. The interaction of TRV 16K protein with p80-coilin was shown to facilitate both RNA silencing and SA pathways in order to coordinate an effective anti-virus response [142,143]. In both TRV and BSMV, knockdown of p80-coilin resulted in increased accumulation of the virus and an exacerbation of symptoms [142]. In addition, in plants, the TYLCV-targeted anti-virus defence mechanism has been shown to encompass transcriptional and post-transcriptional gene silencing pathways, leading to plant recovery [154,155,156]. The anti-virus methylation modifications required for these pathways were shown to occur in CBs [151]. TYLCV is protected from these pathways by interaction of TYLCV V2 with host histone deacetylase 6, resulting in reduced methylation of virus DNA [159].
TYLCV V2 also interacts with AGO4, a component of the RdDM pathway which localises with CBs. This interaction, and the subsequent formation of the V2-AGO4 complex, was further shown to rely on co-localisation with CBs [159]. Taken together, this implies an association between CBs and protection against several plant viruses, suggesting a functional role for CBs in anti-virus defence.
In humans, HIV-1 replication is inhibited at the level of transcription by the CB component protein TOE1 [100]. During Adenovirus and Influenza A virus infection, NBs have been linked to contributing to an anti-virus immune response. Adenoviruses target apoptosis pathways via interactions with PML body components and disrupt both PML bodies and the nucleolus, displacing their core components [123,163,164,165]. This suggests that the disruption of NBs is a core process in the virus life cycle, that has evolved to protect the virus from host defence mechanisms. In influenza A virus infection, the host anti-virus defence is mediated by PML bodies, which are involved in the interferon-induced anti-virus pathway [105]. This is subverted by influenza A virus by accumulating NS1 and M1 in PML bodies. This proposed function for PML bodies in influenza A anti-virus defence is strengthened by the discovery that host-resistance to influenza A virus is facilitated by overexpression of PML [106].
It is possible that greater understanding of these mechanisms may inform the discovery of novel targets for anti-virus therapies, targeting NBs such as CBs, PML bodies and nucleoli and thus disrupting the replication cycle of several viruses of plant and animal origin. It will be of great interest to have greater knowledge of other virus types and the extent to which they disrupt NBs. For example, viruses that replicate only in the cytoplasm without an obvious nuclear involvement would be interesting to study, since they may induce cellular stress that might result in re-organisation or disruption of NBs.
References
- Sharp, P.A., Chakraborty, A. K., Henninger, J. E., & Young, R. A, RNA in formation and regulation of transcriptional condensates. Rna 2022, 28, 52–57. [CrossRef]
- Collier, S., et al., A distant coilin homologue is required for the formation of cajal bodies in Arabidopsis. Molecular biology of the cell 2006, 17, 2942–2951. [CrossRef]
- Strzelecka, M., et al., Coilin-dependent snRNP assembly is essential for zebrafish embryogenesis. Nature structural molecular biology of the cell 2010, 17, 403–409. [CrossRef]
- Tucker, K.E., et al., Residual Cajal bodies in coilin knockout mice fail to recruit Sm snRNPs and SMN, the spinal muscular atrophy gene product. The Journal of cell biology 2001, 154, 293–308. [CrossRef]
- Liu, Q.a.D., G, A novel nuclear structure containing the survival of motor neurons protein. The EMBO journal 1996, 15, 3555–3565. [CrossRef]
- Fong, K.-w., et al., Whole-genome screening identifies proteins localized to distinct nuclear bodies. Journal of Cell Biology 2013, 203, 149–164. [CrossRef]
- Mahmoudi, S., et al., WRAP53 is essential for Cajal body formation and for targeting the survival of motor neuron complex to Cajal bodies. PLoS biology 2010, 8, e1000521.
- Isaac, C., Yang, Y., Thomas M. U, Nopp140 functions as a molecular link between the nucleolus and the coiled bodies. The Journal of cell biology 1998, 142, 319–329. [CrossRef]
- Jones, K., Gorzynski, K., Hales, C., Fischer, U., Badbanchi, F., Terns, R. and Terns, M, Direct Interaction of the Spinal Muscular Atrophy Disease Protein SMN with the Small Nucleolar RNA-associated Protein Fibrillarin. Journal of Biological Chemistry 2001, 276, 38645–38651. [CrossRef]
- Meier, U.T. and G. Blobel, Nopp 140 shuttles on tracks between nucleolus and cytoplasm. Cell 1992, 70, 127–138. [CrossRef]
- Pellizzoni, L., et al., The survival of motor neurons (SMN) protein interacts with the snoRNP proteins fibrillarin and GAR1. Current Biology 2001, 11, 1079–1088. [CrossRef]
- Rampersad, S. and P. Tennant, Replication and expression strategies of viruses. Viruses, 2018: p. 55.
- Wang, P., et al., RING tetramerization is required for nuclear body biogenesis and PML sumoylation. Nature communications 2018, 9, 1277.
- S, R.y.C., A simple method of selective staining of the protoplasmic reticulum and its effects on the various nervous organs of vertebrates and invertebrates. . Trab. Lab. Invest. Biol. (Madrid), 1903(2): 129–221.
- Lafarga, M., Herv's, J., Santa-Cruz, M., Villegas, J. and Crespo, D, The "Accessory Body" of Cajal in the Neuronal Nucleus. Anatomy and Embryology 1983, 166, 19–30. [CrossRef]
- Cioce, M. and A.I. Lamond, Cajal bodies: a long history of discovery. Annual Review of Cell and Developmental Biology 2005, 21, 105–131.
- Young, P., Le, T., Dunckley, M., thi Man, N., Burghes, A. and Morris, G, Nuclear Gems and Cajal (Coiled) Bodies in Fetal Tissues: Nucleolar Distribution of the Spinal Muscular Atrophy Protein, SMN. Experimental Cell Research 2001, 265, 252–261. [CrossRef]
- Young, P., Le, T., thi Man, N., Burghes, A. and Morris, G, The Relationship between SMN, the Spinal Muscular Atrophy Protein, and Nuclear Coiled Bodies in Differentiated Tissues and Cultured Cells. Experimental Cell Research 2000, 256, 365–374. [CrossRef]
- Boudonck, K., Dolan, L. and Shaw, P, The Movement of Coiled Bodies Visualized in Living Plant Cells by the Green Fluorescent Protein. Molecular biology of the cell 1999, 10, 2297–2307. [CrossRef]
- Platani, M., Goldberg, I., Swedlow, J. and Lamond, A, In Vivo Analysis of Cajal Body Movement, Separation, and Joining in Live Human Cells. Journal of Cell Biology 2000, 151, 1561–1574. [CrossRef]
- Andrade, L.T., E.; Chan, E., Immunocytochemical Analysis of the Coiled Body in the Cell Cycle and During Cell Proliferation. Proceedings of the National Academy of Sciences 1993, 90, 1947–1951.
- Cantarero, L., Sanz-García, M., Vinograd-Byk, H., Renbaum, P., Levy-Lahad, E. and Lazo, P, VRK1 regulates Cajal body dynamics and protects coilin from proteasomal degradation in cell cycle. Scientific Reports 2015. 5(1). [CrossRef]
- Cioce, M., et al., UV-induced fragmentation of Cajal bodies. The Journal of cell biology 2006, 175, 401–413. [CrossRef]
- Satyanarayana, A., Manns, M. and Rudolph, K, Telomeres, Telomerase and Cancer: An Endless Search to Target the Ends. Cell Cycle 2004, 3, 1136–1148. [CrossRef]
- Love, A., Yu, C., Petukhova, N., Kalinina, N., Chen, J. and Taliansky, M, Cajal bodies and their role in plant stress and disease responses. RNA Biology 2016, 14, 779–790.
- Kroiss, M., Schultz, J., Wiesner, J., Chari, A., Sickmann, A. and Fischer, U, Evolution of an RNP assembly system: A minimal SMN complex facilitates formation of UsnRNPs in Drosophila melanogaster. Proceedings of the National Academy of Sciences 2008, 105, 10045–10050. [CrossRef]
- Barneche, F.S., F;. Echeverrı́a, M., Fibrillarin Genes Encode Both a Conserved Nucleolar Protein and a Novel Small Nucleolar RNA Involved in Ribosomal RNA Methylation in Arabidopsis Thaliana. Journal of Biological Chemistry 2000, 275, 27212–27220.
- Beven, A.F.S., G. G.; Brown, J. W. S.; Shaw, P. J., The Organization of Spliceosomal Components in the Nuclei of Higher Plants. Journal of Cell Science 1995, 108, 509–518. [CrossRef]
- Bassett, C., Cajal Bodies and Plant RNA Metabolism. Critical Reviews in Plant Sciences 2012, 31, 258–270.
- Kim, S., Spensley, M., Choi, S., Calixto, C., Pendle, A., Koroleva, O., Shaw, P. and Brown, J, Plant U13 orthologues and orphan snoRNAs identified by RNomics of RNA from Arabidopsis nucleoli. Nucleic Acids Research 2010, 38, 3054–3067. [CrossRef]
- Li, C.F., et al., An ARGONAUTE4-containing nuclear processing center colocalized with Cajal bodies in Arabidopsis thaliana. Cell 2006, 126, 93–106. [CrossRef]
- Scheer, U. and R. Hock, Structure and function of the nucleolus. Current opinion in cell biology 1999, 11, 385–390. [CrossRef]
- Iarovaia, O.V., et al., Nucleolus: a central hub for nuclear functions. Trends in cell biology 2019, 29, 647–659. [CrossRef]
- Boulon, S., et al., The nucleolus under stress. Molecular cell 2010, 40, 216–227. [CrossRef]
- Guldner, H.-H., et al., Scl 70 autoantibodies from scleroderma patients recognize a 95 kDa protein identified as DNA topoisomerase I. Chromosoma 1986, 94, 132–138. [CrossRef]
- Reimer, G., et al., Autoantibody to RNA polymerase I in scleroderma sera. The Journal of clinical investigation 1987, 79, 65–72. [CrossRef]
- Masson, C., et al., A 116000 M r nucleolar antigen specific for the dense fibrillar component of the nucleoli. Journal of Cell Science 1990, 95, 371–381. [CrossRef]
- Padeken, J. and P. Heun, Nucleolus and nuclear periphery: velcro for heterochromatin. Current opinion in cell biology 2014, 28, 54–60. [CrossRef]
- Guarente, L., Link between aging and the nucleolus. Genes development 1997, 11, 2449–2455. [CrossRef]
- Visintin, R., E.S. Hwang, and A. Amon, Cfi1 prevents premature exit from mitosis by anchoring Cdc14 phosphatase in the nucleolus. Nature 1999, 398, 818–823. [CrossRef]
- Weber, J.D., et al., Nucleolar Arf sequesters Mdm2 and activates p53. Nature cell biology 1999, 1, 20–26. [CrossRef]
- Daniely, Y. and J.A. Borowiec, Formation of a complex between nucleolin and replication protein A after cell stress prevents initiation of DNA replication. The Journal of cell biology 2000, 149, 799–810. [CrossRef]
- Higashiura, M., et al., Immunolocalization of Ku-proteins (p80/p70): localization of p70 to nucleoli and periphery of both interphase nuclei and metaphase chromosomes. Experimental cell research 1992, 201, 444–451.
- Boisvert, F.-M., et al., The multifunctional nucleolus. Nature reviews Molecular cell biology 2007, 8, 574–585.
- Ogawa, L.a.B., S., Crosstalk between the nucleolus and the DNA damage response. Molecular BioSystems 2017, 13, 443–455. [CrossRef]
- Govoni, M., Farabegoli, F., Pession, A. and Novello, F., Inhibition of Topoisomerase II Activity and Its Effect on Nucleolar Structure and Function. Experimental Cell Research 1994, 211, 36–41. [CrossRef]
- Al-Baker, E.B., J.; Harry, R.; Kill, I., A p53-independent pathway regulates nucleolar segregation and antigen translocation in response to DNA damage induced by UV irradiation. Exp Cell Res 2004, 292, 179–186.
- Miller, M., Andringa, A., Dixon, K. and Carty, M., Insights into UV-induced apoptosis: ultrastructure, trichrome stain and spectral imaging. Micron 2002, 33, 157–166. [CrossRef]
- Hernandez-Verdun, D., Nucleolus: from structure to dynamics. Histochemistry and Cell Biology 2005, 125, 127–137. [CrossRef]
- Lallemand-Breitenbach, V.a.d.T., H, PML nuclear bodies: from architecture to function. Current Opinion in Cell Biology 2018, 52, 154–161.
- Zhong, S., Müller, S., Ronchetti, S., Freemont, P., Dejean, A. and Pandolfi, P, Role of SUMO-1–modified PML in nuclear body formation. Blood 2000, 95, 2748–2752. [CrossRef]
- Borden, K., Lally, J., Martin, S., O'Reilly, N., Solomon, E. and Freemont, P and pp.1601-1606., In vivo and in vitro characterization of the B1 and B2 zinc-binding domains from the acute promyelocytic leukemia protooncoprotein PML. Proceedings of the National Academy of Sciences 1996, 93(4). [CrossRef]
- Sahin, U., Ferhi, O., Jeanne, M., Benhenda, S., Berthier, C., Jollivet, F., Niwa-Kawakita, M., Faklaris, O., Setterblad, N., de Thé, H. and Lallemand-Breitenbach, V, Oxidative stress–induced assembly of PML nuclear bodies controls sumoylation of partner proteins. Journal of cell biology 2014, 204, 931–945. [CrossRef]
- Wang, Z., Ruggero, D., Ronchetti, S., Zhong, S., Gaboli, M., Rivi, R. and Pandolfi, P, Pml is essential for multiple apoptotic pathways. Nature Genetics 1998, 20, 266–272. [CrossRef]
- Regad, T.a.C.-A., M, Role and fate of PML nuclear bodies in response to interferon and viral infections. Oncogene 2001, 20, 7274–7286.
- Raška, I., Andrade, L., Ochs, R., Chan, E., Chang, C., Roos, G. and Tan, E, Immunological and ultrastructural studies of the nuclear coiled body with autoimmune antibodies. Experimental Cell Research 1991, 195, 27–37. [CrossRef]
- Andrade, L.C., E.; Raska, I.; Peebles, C.; Roos, G.; Tan, E., Human autoantibody to a novel protein of the nuclear coiled body: immunological characterization and cDNA cloning of p80-coilin. Journal of Experimental Medicine 1991, 173, 1407–1419.
- Walker, M., Tian, L. and Matera, A, Reduced Viability, Fertility and Fecundity in Mice Lacking the Cajal Body Marker Protein, Coilin. PLoS ONE 2009, 4, 6171. [CrossRef]
- Carmo-Fonseca, M., Ferreira, J. and Lamond, A, Assembly of snRNP-containing coiled bodies is regulated in interphase and mitosis--evidence that the coiled body is a kinetic nuclear structure. Journal of Cell Biology 1993, 120, 841–852.
- Bohmann, K., Ferreira, J. and Lamond, A and pp.. Mutational analysis of p80 coilin indicates a functional interaction between coiled bodies and the nucleolus. Journal of Cell Biology 1995, 131, 817–831. [CrossRef]
- Hebert, M.a.M., A, Self-association of Coilin Reveals a Common Theme in Nuclear Body Localization. Molecular Biology of the Cell 2000, 11, 4159–4171.
- Hebert, M., Szymczyk, P., Shpargel, K. and Matera, A, Coilin forms the bridge between Cajal bodies and SMN, the Spinal Muscular Atrophy protein. Genes & Development 2001, 15, 2720–2729. [CrossRef]
- Meister, G., Characterization of a nuclear 20S complex containing the survival of motor neurons (SMN) protein and a specific subset of spliceosomal Sm proteins. Human Molecular Genetics 2000, 9, 1977–1986. [CrossRef]
- Gubitz, A., Feng, W. and Dreyfuss, G, The SMN complex. Experimental Cell Research 2004, 296, 51–56.
- Pellizzoni, L., Kataoka, N., Charroux, B. and Dreyfuss, G, A Novel Function for SMN, the Spinal Muscular Atrophy Disease Gene Product, in Pre-mRNA Splicing. Cell 1998, 95, 615–624. [CrossRef]
- Sleeman, J., Cajal body proteins SMN and Coilin show differential dynamic behaviour in vivo. Journal of Cell Science 2003, 116, 2039–2050. [CrossRef]
- Lanfranco, M., Vassallo, N. and Cauchi, R, Spinal Muscular Atrophy: From Defective Chaperoning of snRNP Assembly to Neuromuscular Dysfunction. Frontiers in MolecularBiosciences 2017. 4. [CrossRef]
- Young, P.J., et al., A direct interaction between the survival motor neuron protein and p53 and its relationship to spinal muscular atrophy. Journal of Biological Chemistry 2002, 277, 2852–2859. [CrossRef]
- de Belle, I., Wu, J., Sperandio, S., Mercola, D. and Adamson, E, In Vivo Cloning and Characterization of a New Growth Suppressor Protein TOE1 as a Direct Target Gene of Egr1. Journal of Biological Chemistry 2003, 278, 14306–14312. [CrossRef]
- Mahmoudi, S., Henriksson, S., Corcoran, M., Méndez-Vidal, C., Wiman, K. and Farnebo, M, Wrap53, a Natural p53 Antisense Transcript Required for p53 Induction upon DNA Damage. Molecular cell 2009, 33, 462–471. [CrossRef]
- Henriksson, S.a.F., M, On the road with WRAP53β: guardian of Cajal bodies and genome integrity. Frontiers in Genetics 2015, 6.
- Venteicher, A., Abreu, E., Meng, Z., McCann, K., Terns, R., Veenstra, T., Terns, M. and Artandi, S, A Human Telomerase Holoenzyme Protein Required for Cajal Body Localization and Telomere Synthesis. Science 2009, 323, 644–648. [CrossRef]
- Machyna, M., Kehr, S., Straube, K., Kappei, D., Buchholz, F., Butter, F., Ule, J., Hertel, J., Stadler, P. and Neugebauer, K, The Coilin Interactome Identifies Hundreds of Small Noncoding RNAs that Traffic through Cajal Bodies. Molecular Cell 2014, 56, 389–399. [CrossRef]
- Staněk, D., Cajal bodies and snRNPs - friends with benefits. RNA Biology 2017, 14, 671–679. [CrossRef]
- Carmo-Fonseca, M., Pepperkok, R., Sproat, B., Ansorge, W., Swanson, M. and Lamond, A, In vivo detection of snRNP-rich organelles in the nuclei of mammalian cells. The EMBO Journal 1991, 10, 1863–1873.
- Matera, A.a.W., D, Nucleoplasmic organization of small nuclear ribonucleoproteins in cultured human cells. Journal of Cell Biology 1993, 121, 715–727. [CrossRef]
- Burke, M., Logan, M. and Hebert, M, Identification of additional regulatory RNPs that impact rRNA and U6 snRNA methylation. Biology Open 2018, 7, bio036095. [CrossRef]
- Darzacq, X., et al., Cajal body-specific small nuclear RNAs: a novel class of 2′-O-methylation and pseudouridylation guide RNAs. The EMBO journal 2002, 21, 2746–2756. [CrossRef]
- Jády, B.E. and T. Kiss, A small nucleolar guide RNA functions both in 2′-O-ribose methylation and pseudouridylation of the U5 spliceosomal RNA. The EMBO journal 2001, 20, 541–551. [CrossRef]
- Kiss, A.M., et al., A Cajal body-specific pseudouridylation guide RNA is composed of two box H/ACA snoRNA-like domains. Nucleic acids research 2002, 30, 4643–4649.
- Tycowski, K., You, Z., Graham, P. and Steitz, J, Modification of U6 Spliceosomal RNA Is Guided by Other Small RNAs. Molecular Cell. 1998, 2, 629–638. [CrossRef]
- Ganot, P., Jády, B., Bortolin, M., Darzacq, X. and Kiss, T, Nucleolar Factors Direct the 2′-O-Ribose Methylation and Pseudouridylation of U6 Spliceosomal RNA. Molecular and Cellular Biology 1999, 19, 6906–6917. [CrossRef]
- Richard, P., Darzacq, X., Bertrand, E., Jady, B., Verheggen, C. and Kiss, T, A common sequence motif determines the Cajal body-specific localization of box H/ACA scaRNAs. The EMBO Journal 2003, 22, 4283–4293. [CrossRef]
- Tycowski, K., Shu, M., Kukoyi, A. and Steitz, J, A Conserved WD40 Protein Binds the Cajal Body Localization Signal of scaRNP Particles. Molecular Cell 2009, 34, 47–57.
- Fischer, U., Englbrecht, C. and Chari, A, Biogenesis of spliceosomal small nuclear ribonucleoproteins. Wiley Interdisciplinary Reviews: RNA 2011, 2, 718–731. [CrossRef]
- Skaar, J., Ferris, A., Wu, X., Saraf, A., Khanna, K., Florens, L., Washburn, M., Hughes, S. and Pagano, M, The Integrator complex controls the termination of transcription at diverse classes of gene targets. Cell Research 2015, 25, 288–305. [CrossRef]
- Takata, H., Nishijima, H., Maeshima, K. and Shibahara, K, The integrator complex is required for integrity of Cajal bodies. Journal of Cell Science 2012, 125, 166–175. [CrossRef]
- Matera, A.a.W., Z, A day in the life of the spliceosome. Nature Reviews Molecular Cell Biology 2014, 15, 108–121.
- Jady, B., Darzacq, X., Tucker, K., Matera, A., Bertrand, E. and Kiss, T, Modification of Sm small nuclear RNAs occurs in the nucleoplasmic Cajal body following import from the cytoplasm. The EMBO Journal 2003, 22, 1878–1888. [CrossRef]
- Tsukaya, H., et al., How do ‘housekeeping’genes control organogenesis?—Unexpected new findings on the role of housekeeping genes in cell and organ differentiation. Journal of plant research 2013, 126, 3–15.
- Cotmore, S., Sturzenbecker, L. and Tattersall, P, The autonomous parvovirus MVM encodes two nonstructural proteins in addition to its capsid polypeptides. Virology 1983, 129, 333–343. [CrossRef]
- Cotmore, S.a.T., P, Organization of nonstructural genes of the autonomous parvovirus minute virus of mice. Journal of Virology 1986, 58, 724–732. [CrossRef]
- Cziepluch, C., Lampel, S., Grewenig, A., Grund, C., Lichter, P. and Rommelaere, J, H-1 Parvovirus-Associated Replication Bodies: a Distinct Virus-Induced Nuclear Structure. Journal of Virology 2000, 74, 4807–4815. [CrossRef]
- Bashir, T.R., J.; Cziepluch, C., In Vivo Accumulation of Cyclin A and Cellular Replication Factors in Autonomous Parvovirus Minute Virus of Mice-Associated Replication Bodies. Journal of Virology 2001, 75, 4394–4398.
- Young, P., Jensen, K., Burger, L., Pintel, D. and Lorson, C, Minute Virus of Mice NS1 Interacts with the SMN Protein, and They Colocalize in Novel Nuclear Bodies Induced by Parvovirus Infection. Journal of Virology 2002, 76, 3892–3904. [CrossRef]
- Simões, M., Rino, J., Pinheiro, I., Martins, C. and Ferreira, F, Alterations of Nuclear Architecture and Epigenetic Signatures during African Swine Fever Virus Infection. Viruses 2015, 7, 4978–4996. [CrossRef]
- Dixon, L., Chapman, D., Netherton, C. and Upton, C, African swine fever virus replication and genomics. Virus Research 2013, 173, 3–14. [CrossRef]
- Sánchez, E., Quintas, A., Nogal, M., Castelló, A. and Revilla, Y, African swine fever virus controls the host transcription and cellular machinery of protein synthesis. Virus Research 2013, 173, 58–75.
- Fanales-Belasio, E., Raimondo, M., Suligoi, B. and Buttò, S, HIV virology and pathogenetic mechanisms of infection: a brief overview. Annali dell’Istituto Superiore di Sanità 2010, 46(1). [CrossRef]
- Sperandio, S., Barat, C., Cabrita, M., Gargaun, A., Berezovski, M., Tremblay, M. and de Belle, I, TOE1 is an inhibitor of HIV-1 replication with cell-penetrating capability. Proceedings of the National Academy of Sciences 2015, 112, E3392–E3401. [CrossRef]
- Webster, R.G., Bean, W. J., Gorman, O. T., Chambers, T. M., & Kawaoka, Y Evolution and ecology of Influenza A viruses. Microbiological Reviews 1992, 56, 152– 179.
- Höfer, C., Jolmes, F., Haralampiev, I., Veit, M. and Herrmann, A Influenza A virus nucleoprotein targets subnuclear structures. Cellular Microbiology 2016, 19, p. p.e12679. [CrossRef]
- Josset, L., Frobert, E. and Rosa-Calatrava, M, Influenza A replication and host nuclear compartments: Many changes and many questions. Journal of Clinical Virology 2008, 43, 381–390. [CrossRef]
- Engelhardt, O.a.F., E, Functional association between viral and cellular transcription during influenza virus infection. Reviews in Medical Virology 2006, 16, 329–345.
- Sato, Y., Yoshioka, K., Suzuki, C., Awashima, S., Hosaka, Y., Yewdell, J. and Kuroda, K, Localization of influenza virus proteins to nuclear dot 10 structures in influenza virusinfected cells. Virology 2003, 310, 29–40. [CrossRef]
- Chelbi-Alix, M., Quignon, F., Pelicano, L., Koken, M. and de Thé, H, Resistance to Virus Infection Conferred by the Interferon-Induced Promyelocytic Leukemia Protein. Journal of Virology 1998, 72, 1043–1051. [CrossRef]
- Compans, R.a.D., N, An electron microscopic study of single-cycle infection of chick embryo fibroblasts by influenza virus. Virology 1969, 39, 499–515.
- Shaw, M.a.C., R, Isolation and characterization of cytoplasmic inclusions from influenza A virus-infected cells. Journal of Virology 1978, 25, 608–615. [CrossRef]
- Terrier, O., Moules, V., Carron, C., Cartet, G., Frobert, E., Yver, M., Traversier, A., Wolff, T., Riteau, B., Naffakh, N., Lina, B., Diaz, J. and Rosa-Calatrava, M, The influenza fingerprints: NS1 and M1 proteins contribute to specific host cell ultrastructure signatures upon infection by different influenza A viruses. Virology 2012, 432, 204–218. [CrossRef]
- Davey, J., Colman, A. and Dimmock, N, Location of Influenza Virus M, NP and NS1 Proteins in Microinjected Cells. Journal of General Virology 1985, 66, 2319–2334. [CrossRef]
- Murayama, R., Harada, Y., Shibata, T., Kuroda, K., Hayakawa, S., Shimizu, K. and Tanaka, T, Influenza A virus non-structural protein 1 (NS1) interacts with cellular multifunctional protein nucleolin during infection. Biochemical and Biophysical Research Communications 2007, 362, 880–885. [CrossRef]
- Ozawa, M., Fujii, K., Muramoto, Y., Yamada, S., Yamayoshi, S., Takada, A., Goto, H., Horimoto, T. and Kawaoka, Y, Contributions of Two Nuclear Localization Signals of Influenza A Virus Nucleoprotein to Viral Replication. Journal of Virology 2007, 81, 30–41. [CrossRef]
- Fortes, P., Lamond, A. and Ortin, J, Influenza virus NS1 protein alters the subnuclear localization of cellular splicing components. Journal of General Virology 1995, 76, 1001–1007. [CrossRef]
- Coyaud, E., Ranadheera, C., Cheng, D., Gonçalves, J., Dyakov, B., Laurent, E., St-Germain, J., Pelletier, L., Gingras, A., Brumell, J., Kim, P., Safronetz, D. and Raught, B, Global Interactomics Uncovers Extensive Organellar Targeting by Zika Virus. Molecular & Cellular Proteomics 2018, 17, 2242–2255. [CrossRef]
- Shankar, A., Patil, A. and Skariyachan, S, Recent Perspectives on Genome, Transmission, Clinical Manifestation, Diagnosis, Therapeutic Strategies, Vaccine Developments, and Challenges of Zika Virus Research. Frontiers in Microbiology 2017, 8. [CrossRef]
- Hearing, P., Adenoviridae: The Viruses and Their Replication. 7th ed. In Fields Virology: DNA Viruses., ed. P.M. Howley, Knipe, D. M. Vol. 2. 2021, Philadelphia: Wolters Kluwer Health.
- Rodrigues S. H., S.N.P., Delício L. R., Granato C., Andrade L. E, The behaviour of the coiled body in cells infected with adenovirus in vitro. Molecular Biolology Rep 1996. 23(3-4): 183–189.
- James, N., Howell, G., Walker, J. and Blair, G, The role of Cajal bodies in the expression of late phase adenovirus proteins. Virology 2010, 399, 299–311. [CrossRef]
- White, L.E., B.; Blair, G.E., The Cajal Body Protein p80-coilin Forms a Complex with the Adenovirus L4-22K Protein and Facilitates the Nuclear Export of Adenovirus mRNA. mBio 2023, 0, 01459–23.
- Gedge, L., Morrison, E., Blair, G. and Walker, J, Nuclear actin is partially associated with Cajal bodies in human cells in culture and relocates to the nuclear periphery after infection of cells by adenovirus 5. Experimental Cell Research 2005, 303, 229–239. [CrossRef]
- Guimet, D., & Hearing, P., The adenovirus L4-22K protein has distinct functions in the posttranscriptional regulation of gene expression and encapsidation of the viral genome. Journal of virology 2013, 87, 7688–7699. [CrossRef]
- Biasiotto, R., & Akusjärvi, G., Regulation of human adenovirus alternative RNA splicing by the adenoviral L4-33K and L4-22K proteins. International journal of molecular sciences 2015, 16, 2893–2912.
- Carvalho, T., Seeler, J., Ohman, K., Jordan, P., Pettersson, U., Akusjärvi, G., Carmo-Fonseca, M. and Dejean, A, Targeting of adenovirus E1A and E4-ORF3 proteins to nuclear matrix-associated PML bodies. Journal of Cell Biology 1995, 131, 45–56. [CrossRef]
- Ullman, A.a.H., P, Cellular Proteins PML and Daxx Mediate an Innate Antiviral Defense Antagonized by the Adenovirus E4 ORF3 Protein. Journal of Virology 2008, 82, 7325–7335.
- Salsman, J., Zimmerman, N., Chen, T., Domagala, M. and Frappier, L, Genome-Wide Screen of Three Herpesviruses for Protein Subcellular Localization and Alteration of PML Nuclear Bodies. PLoS Pathogens 2008, 4, p. p.e1000100. [CrossRef]
- Anobile, J.A., V.; Downs, D.; Czymmek, K.; Parcells, M.; Schmidt, C., Nuclear Localization and Dynamic Properties of the Marek's Disease Virus Oncogene Products Meq and Meq/vIL8. Journal of Virology 2006, 80, 1160–1166.
- Liu, J., Lee, L., Ye, Y., Qian, Z. and Kung, H, Nucleolar and nuclear localization properties of a herpesvirus bZIP oncoprotein MEQ. Journal of virology 1997, 71, 3188–3196. [CrossRef]
- Rajamäki, M.a.V., J, Control of Nuclear and Nucleolar Localization of Nuclear Inclusion Protein a of Picorna-Like Potato virus A in Nicotiana Species. The Plant Cell 2009, 21, 2485–2502.
- Schaad, M., Haldeman-Cahill, R., Cronin, S. and Carrington, J, Analysis of the VPgproteinase (NIa) encoded by tobacco etch potyvirus: effects of mutations on subcellular transport, proteolytic processing, and genome amplification. Journal of virology 1996, 70, 7039–7048.
- Ruiz-Ruiz, S., Soler, N., Sánchez-Navarro, J., Fagoaga, C., Lopez, C., Navarro, L., ... & Flores, R, Citrus tristeza virus p23: determinants for nucleolar localization and their influence on suppression of RNA silencing and pathogenesis. Molecular Plant-Microbe Interactions 2013, 26, 306–318. [CrossRef]
- Taliansky, M.a.R., D, Molecular biology of umbraviruses: phantom warriors. Journal of General Virology 2003, 84, 1951–1960.
- Ryabov, E., Oparka, K., Santa Cruz, S., Robinson, D. and Taliansky, M, Intracellular Location of Two Groundnut Rosette Umbravirus Proteins Delivered by PVX and TMV Vectors. Virology 1998, 242, 303–313. [CrossRef]
- Ryabov, E., Robinson, D. and Taliansky, M, A plant virus-encoded protein facilitates long-distance movement of heterologous viral RNA. Proceedings of the National Academy of Sciences 1999, 96, 1212–1217. [CrossRef]
- Kim, S., Ryabov, E., Kalinina, N., Rakitina, D., Gillespie, T., MacFarlane, S., Haupt, S., Brown, J. and Taliansky, M, Cajal bodies and the nucleolus are required for a plant virus systemic infection. The EMBO Journal 2007, 26, 2169–2179. [CrossRef]
- Kim, S., MacFarlane, S., Kalinina, N., Rakitina, D., Ryabov, E., Gillespie, T., Haupt, S., Brown, J. and Taliansky, M, Interaction of a plant virus-encoded protein with the major nucleolar protein fibrillarin is required for systemic virus infection. Proceedings of the National Academy of Sciences 2007, 104, 11115–11120. [CrossRef]
- Taliansky, M., Roberts, I., Kalinina, N., Ryabov, E., Raj, S., Robinson, D. and Oparka, K, An Umbraviral Protein, Involved in Long-Distance RNA Movement, Binds Viral RNA and Forms Unique, Protective Ribonucleoprotein Complexes. Journal of Virology 2003, 77, 3031–3040. [CrossRef]
- Solovyev, A., Savenkov, E., Agranovsky, A. and Morozov, S, Comparisons of the Genomiccis-Elements and Coding Regions in RNAβ Components of the Hordeiviruses Barley Stripe Mosaic Virus, Lychnis Ringspot Virus, and Poa Semilatent Virus. Virology 1996, 219, 9–18.
- Jackson, A., Lim, H., Bragg, J., Ganesan, U. and Lee, M, Hordeivirus Replication, Movement, and Pathogenesis. Annual Review of Phytopathology 2009, 47, 385–422.
- Lim, H., Bragg, J., Ganesan, U., Lawrence, D., Yu, J., Isogai, M., Hammond, J. and Jackson, A, Triple Gene Block Protein Interactions Involved in Movement of Barley Stripe Mosaic Virus. Journal of Virology 2008, 82, 4991–5006. [CrossRef]
- Semashko, M., Rakitina, D., González, I., Canto, T., Kalinina, N. and Taliansky, M, Movement protein of hordeivirus interacts in vitro and in vivo with coilin, a major structural protein of Cajal bodies. Doklady Biochemistry and Biophysics 2012, 442, 57–60. [CrossRef]
- MacFarlane, S., Tobraviruses-plant pathogens and tools for biotechnology. Molecular Plant Pathology 2010, 11, 577–583. [CrossRef]
- Shaw, J., Love, A., Makarova, S., Kalinina, N., Harrison, B. and Taliansky, M, Coilin, the signature protein of Cajal bodies, differentially modulates the interactions of plants with viruses in widely different taxa. Nucleus 2014, 5, 85–94. [CrossRef]
- Shaw, J., Yu, C., Makhotenko, A., Makarova, S., Love, A., Kalinina, N., MacFarlane, S., Chen, J. and Taliansky, M, Interaction of a plant virus protein with the signature Cajal body protein coilin facilitates salicylic acid-mediated plant defence responses. New Phytologist 2019, 224, 439–453. [CrossRef]
- Spechenkova, N., Samarskaya, V. O., Kalinina, N. O., Zavriev, S. K., MacFarlane, S., Love, A. J., & Taliansky, M, Plant Poly (ADP-ribose) Polymerase 1 Is a Potential Mediator of Cross-Talk between the Cajal Body Protein Coilin and Salicylic Acid-Mediated Antiviral Defence. Viruses 2023, 15, 1282. [CrossRef]
- Zhou, H.a.J., A, Expression of the Barley Stripe Mosaic Virus RNAβ “Triple Gene Block”. Virology 1996, 216, 367–379.
- Li, Z., Zhang, Y., Jiang, Z., Jin, X., Zhang, K., Wang, X., Han, C., Yu, J. and Li, D, Hijacking of the nucleolar protein fibrillarin by TGB1 is required for cell-to-cell movement of Barley stripe mosaic virus. Molecular Plant Pathology 2017, 19, 1222–1237. [CrossRef]
- Zheng, L., Du, Z., Lin, C., Mao, Q., Wu, K., Wu, J., Wei, T., Wu, Z. and Xie, L, Rice stripe tenuivirus p2 may recruit or manipulate nucleolar functions through an interaction with fibrillarin to promote virus systemic movement. Molecular Plant Pathology 2015, 16, 921–930. [CrossRef]
- Zheng, L., He, J., Ding, Z., Zhang, C. and Meng, R, Identification of Functional Domain(s) of Fibrillarin Interacted with p2 of Rice stripe virus. Canadian Journal of Infectious Diseases and Medical Microbiology 2018, 2018, 1–6. [CrossRef]
- Zheng, L., Hong, P., Guo, X., Li, Y. and Xie, L, Rice stripe virus p2 Colocalizes and Interacts with Arabidopsis Cajal Bodies and Its Domains in Plant Cells. BioMed Research International 2020, 2020, 1–7. [CrossRef]
- Guo, T., Vimalesvaran, D., Thompson, J., Perry, K. and Krenz, B, Subcellular localization of grapevine red blotch-associated virus ORFs V2 and V3. Virus Genes 2015, 51, 156–158. [CrossRef]
- Wang, L., Ding, Y., He, L., Zhang, G., Zhu, J. and Lozano-Duran, R, A virus-encoded protein suppresses methylation of the viral genome through its interaction with AGO4 in the Cajal body. eLife 2020. 9.
- Czosnek, H., Eybishtz, A., Sade, D., Gorovits, R., Sobol, I., Bejarano, E., ... & Lozano-Durán, R, Discovering host genes involved in the infection by the Tomato Yellow Leaf Curl Virus complex and in the establishment of resistance to the virus using Tobacco Rattle Virus-based post transcriptional gene silencing. Viruses 2013, 5, 998–1022. [CrossRef]
- Ceniceros-Ojeda, E., Rodríguez-Negrete, E. and Rivera-Bustamante, R, Two Populations of Viral Minichromosomes Are Present in a Geminivirus-Infected Plant Showing Symptom Remission (Recovery). Journal of Virology 2016, 90, 3828–3838. [CrossRef]
- Deuschle, K., Kepp, G. and Jeske, H, Differential methylation of the circular DNA in geminiviral minichromosomes. Virology 2016, 499, 243–258. [CrossRef]
- Raja, P., Sanville, B., Buchmann, R. and Bisaro, D, Viral Genome Methylation as an Epigenetic Defense against Geminiviruses. Journal of Virology 2008, 82, 8997–9007. [CrossRef]
- Rodríguez-Negrete, E., Carrillo-Tripp, J. and Rivera-Bustamante, R, RNA Silencing against Geminivirus: Complementary Action of Posttranscriptional Gene Silencing and Transcriptional Gene Silencing in Host Recovery. Journal of Virology 2008, 83, 1332–1340. [CrossRef]
- Wang, B., Li, F., Huang, C., Yang, X., Qian, Y., Xie, Y. and Zhou, X, V2 of tomato yellow leaf curl virus can suppress methylation-mediated transcriptional gene silencing in plants. Journal of General Virology 2014, 95, 225–230. [CrossRef]
- Zrachya, A., Glick, E., Levy, Y., Arazi, T., Citovsky, V. and Gafni, Y, Suppressor of RNA silencing encoded by Tomato yellow leaf curl virus-Israel. Virology 2007, 358, 159–165. [CrossRef]
- Wang, B., Yang, X., Wang, Y., Xie, Y. and Zhou, X, Tomato Yellow Leaf Curl Virus V2 Interacts with Host Histone Deacetylase 6 To Suppress Methylation-Mediated Transcriptional Gene Silencing in Plants. Journal of Virology 2018. 92(18). [CrossRef]
- Wang, Y., Wu, Y., Gong, Q., Ismayil, A., Yuan, Y., Lian, B., Jia, Q., Han, M., Deng, H., Hong, Y., Hanley-Bowdoin, L., Qi, Y. and Liu, Y, Geminiviral V2 Protein Suppresses Transcriptional Gene Silencing through Interaction with AGO4. Journal of Virology 2019. 93(6). [CrossRef]
- Hamera, S., Song, X., Su, L., Chen, X., & Fang, R, Cucumber mosaic virus suppressor 2b binds to AGO4-related small RNAs and impairs AGO4 activities. The Plant Journal 2012, 69, 104–115. [CrossRef]
- Duan, C.G., Fang, Y. Y., Zhou, B. J., Zhao, J. H., Hou, W. N., Zhu, H., ... & Guo, H. S, Suppression of Arabidopsis ARGONAUTE1-mediated slicing, transgene-induced RNA silencing, and DNA methylation by distinct domains of the Cucumber mosaic virus 2b protein. The Plant Cell 2012, 24, 259–274.
- Lin, J., Chen, J., Elenbaas, B. and Levine, A, Several hydrophobic amino acids in the p53 amino-terminal domain are required for transcriptional activation, binding to mdm-2 and the adenovirus 5 E1B 55-kD protein. Genes & Development 1994, 8, 1235–1246. [CrossRef]
- Schreiner, S., Wimmer, P., Sirma, H., Everett, R., Blanchette, P., Groitl, P. and Dobner, T, Proteasome-Dependent Degradation of Daxx by the Viral E1B-55K Protein in Human Adenovirus-Infected Cells. Journal of Virology 2010, 84, 7029–7038. [CrossRef]
- Zhao, L., Colosimo, A., Liu, Y., Wan, Y. and Liao, D, Adenovirus E1B 55-Kilodalton Oncoprotein Binds to Daxxand Eliminates Enhancement of p53-Dependent Transcription byDaxx. Journal of Virology 2003, 77, 11809–11821. [CrossRef]
Figure 1.
A schematic representation of the eukaryotic nucleus. Major domains and Nuclear Bodies (NBs) are labelled. The NBs upon which this review is focussed are nucleoli, Cajal bodies and PML bodies (marked in red ovals). See text for details.
Figure 1.
A schematic representation of the eukaryotic nucleus. Major domains and Nuclear Bodies (NBs) are labelled. The NBs upon which this review is focussed are nucleoli, Cajal bodies and PML bodies (marked in red ovals). See text for details.
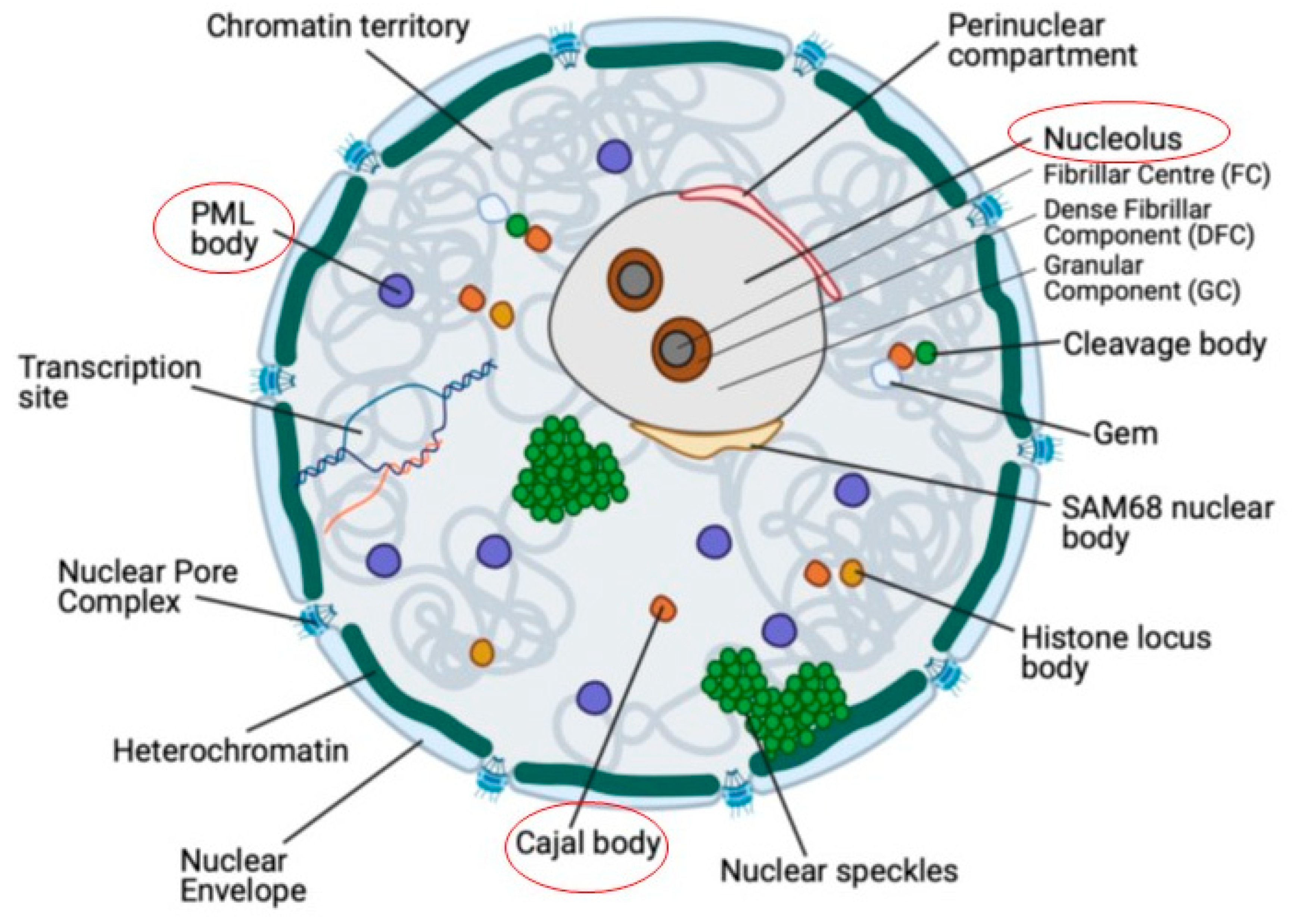
Figure 2.
A schematic representation of the domain structure of p80-coilin. The self-interacting domain is located at the N terminus. NLSa and NLSb refer to the two nuclear localisation signals present in coilin and NoLS refers to the nucleolar localisation signal, all of which have positive charge. The stippled boxes represent acidic serine-rich patches. The RG box domain is located at the C-terminus. The C terminus of coilin is folded into a Tudor domain. Modified from [62].
Figure 2.
A schematic representation of the domain structure of p80-coilin. The self-interacting domain is located at the N terminus. NLSa and NLSb refer to the two nuclear localisation signals present in coilin and NoLS refers to the nucleolar localisation signal, all of which have positive charge. The stippled boxes represent acidic serine-rich patches. The RG box domain is located at the C-terminus. The C terminus of coilin is folded into a Tudor domain. Modified from [62].
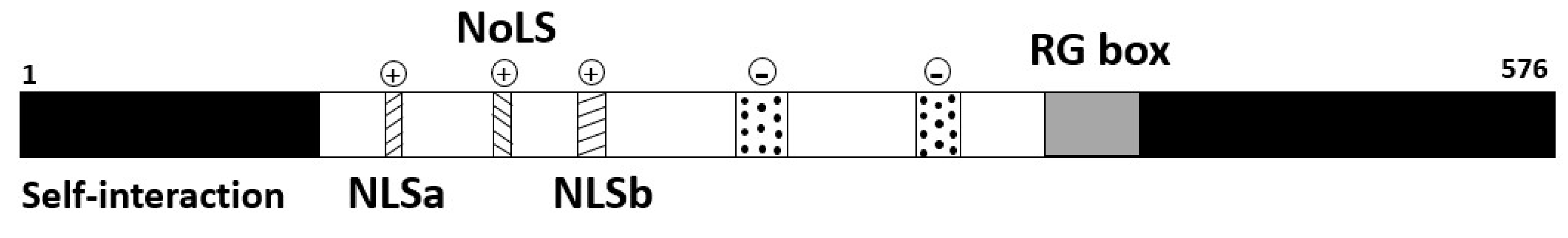

Table 1.
Viruses that infect members of the animal kingdom and interact with Cajal bodies as well as other relevant nuclear bodies.
Table 1.
Viruses that infect members of the animal kingdom and interact with Cajal bodies as well as other relevant nuclear bodies.
| Species infected by virus | Name of Virus | Virus protein that interacts with Cajal bodies | Cajal body protein that Interacts with virus | Function of interaction | Other Nuclear Bodies that interact with virus (along with CBs) | References |
|---|---|---|---|---|---|---|
| Aves (Birds),principally Gallus gallus domesticus (Chicken) | Marek's Disease Virus (MDV) (Avian alphaherpesvirus) | Meq/ Meq/vIL8 | p80-Coilin | Unknown | Nucleoplasm, nucleolus | [91,92] |
| Several mammalian species, including Homo sapiens (Human) | Adenovirus Type 5 (Ad5) | L4-22K | p80-coilin | Ad5 infection disrupts CBs, redistributed into CB microfoci; L4-22K forms complex with p80-coilin. Depletion of p80-coilin reduces export of virus mRNAs from nucleus in infected cells. | PML bodies (E4-ORF3 11kDa), PML bodies (E1A) | [93,94,95,96] |
| Several animal species, including Homo sapiens (Human) | Zika Virus (ZIKV) | NS5 | p80-coilin | Unknown, disrupt CBs, resulting in an increased number of smaller CBs | Unknown | [98] |
| Murinae (Murine), principally Mus musculus (Mouse) | Minute Virus of Mice (MVM) | NS1 | SMN | Disrupt components of CBs, recombined into SAABs (sites of virus replication) | Unknown | [99] |
| Homo sapiens (Human) | Human Cytomegalovirus (HCMV) | UL3 and UL30 | Unknown | Unknown, expression of UL3 and UL30 decreased number of CBs in cell | PML bodies | [100,101,102,103] |
| Several animal species, including Homo sapiens (Human) | Influenza A virus | NP | Unknown | Unknown, expression of NP results in an increased number of smaller CBs | PML bodies, nucleolus (virus NS1) | [97,104,105,106,107,108,109] |
| Sus scrofa domesticus (Swine) | African Swine Fever Virus (AFSV) | Unknown | Unknown | Unknown, disrupt CBs, results in an increased number of smaller CBs | Nuclear speckles, PML bodies | [110] |
| Homo sapiens (Human) | Human Immunodeficiency Virus (HIV) | TAR | TOE1 | Host anti-virus defence | Unknown | [111] |
Table 2.
Viruses that infect members of the plant kingdom and interact with Cajal bodies as well as other relevant nuclear bodies.
Table 2.
Viruses that infect members of the plant kingdom and interact with Cajal bodies as well as other relevant nuclear bodies.
| Species infected by virus |
Name of Virus | Virus protein that interacts with Cajal bodies |
Cajal body protein that Interacts with virus |
Function of interaction | Other Nuclear Bodies that interact with virus (along with CBs) | References |
|---|---|---|---|---|---|---|
| Nicotiana benthamiana | Citrus tristeza virus (CTV) | P23 | Fibrillarin | Suppression of RNA silencing, enhancing systemic infection and virus accumulation | Nucleolus | [131] |
| Solanum tuberosum (Potato) | Potato Virus A (PVA) | NIa | Fibrillarin | Unknown, reduced fibrillarin resulted in reduced accumulation of PVA | Nucleolus | [132] |
|
Arachis hypogaea (Groundnut) |
Groundnut Rosette Virus (GRV) |
ORF3 | Fibrillarin | Long-distance movement, establishing systemic infection, stabilisation of virus RNA | Nucleolus | [133,134] |
| Vitis (Grapevine) | Grapevine Red Blotch-associated Virus (GRBaV) |
V2 | Fibrillarin | Unknown | Nucleolus, inclusions in the nucleoplasm | [135] |
| Oryza sativa (Rice) | Rice Stripe Virus (RSV) |
P2 | Coilin | Unknown | Nucleolus (via fibrillarin to establish systemic infection) |
[136,137,138] |
| Poaceae (Grass) | Poa Semilatent Virus (PSLV) |
TGBp1 | Coilin | Cell-to-cell movement, long distance transport, establishing a systemic infection | Nucleolus | [139,140] |
| A wide variety of species |
Tobacco Rattle Virus (TRV) |
16K | Coilin | Host anti-virus defence (RNA and SA silencing pathways) |
Nucleolus | [141,142] |
|
Hordeum vulgare (Barley) |
Barley Stripe Mosaic Virus (BSMV) | Unknown | Coilin | Host anti-virus defence | Unknown | [141] |
| Solanum ycopersicum (Tomato) | Tomato Yellow Leaf Curl Virus (TYLCV) |
V2 | AGO4 | Suppression of host anti-virus defence (Inhibit TGS and PTGS pathways) |
Nucleoplasm | [143] |
| A wide variety of species | Cucumber mosaic virus (CMV) | 2b | AGO4 | RNA-directed DNA methylation | Nucleolus | [144,145] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



