
Submitted:
17 October 2023
Posted:
18 October 2023
You are already at the latest version
Abstract
The continuous growth of roots depends on their ability to maintain a balanced ratio between cell production and cell differentiation at the tip. This process was reported to be regulated by the hormonal balance of cytokinin and auxin. However, other important regulators, such as plant folates, have also been reported to play a regulatory role. In this study, we investigated the impact of the folate precursor PABA on root development. Using pharmacological, genetic, and imaging approaches, we show that the growth of Arabidopsis thaliana roots is repressed by either supplementing the growth medium with PABA or overexpressing the PABA synthesis gene GAT-ADCS. This is associated with a smaller root meristem size, which consists of fewer cells. Conversely, reducing the levels of free root endogenous PABA results in longer roots with extended meristems and increased cell numbers. We provide evidence that PABA represses Arabidopsis root growth in a folate-independent manner and likely acts through two mechanisms: (i) the G2/M transition of cell division in the root apical meristem and (ii) promoting premature cell differentiation in the transition zone. These data collectively suggest that PABA plays a role in Arabidopsis root growth at the point where cell division and cell differentiation intersect.
Keywords:
Arabidopsis thaliana
; PABA
; root growth
; folates
; cell cycle
1. Introduction
Plants have evolved sophisticated structures to efficiently access and collect available resources. Thus, the uptake of water and nutrients, as well as the overall performance of a plant, are determined by its root system architecture (RSA) [1]. The latter describes the organization of primary and lateral roots, adventitious roots, and root hairs. The primary root is established from the root apical meristem (RAM) during embryo development, whereas lateral roots are initiated post-embryonically from the lateral root founder cells of the primary root [2].
The entire primary root results from the activity of the stem cell niche in the RAM. The root apex consists of five distinct functional/developmental zones based on their cellular activities: the meristematic zone (MZ) containing fast proliferative cells; the root transition zone (TZ) composed of cells containing small vacuoles and exhibiting slow cell growth in both length and width; the fast elongating zone (EZ) which contains fast elongating cells but no change in width; the growth terminating zone (GTZ) where cells progressively slow down their elongation to reach their mature lengths and exhibit root hair tip growth; and the differentiation zone (DZ) containing fully differentiated cells ([3,4,5,6].
The integrity of the RAM is crucial for continuous root growth, and it relies on a balance between the rate of cell differentiation and the rate of cells undergoing mitosis. This balance is established through the antagonistic effects of auxin, which promotes cell division, and cytokinins, which promote cell differentiation [7,8]. The position of the TZ, at the boundary between cell proliferation and cell differentiation, is tightly regulated. This regulation requires cytokinins to inhibit auxin through ARR1-regulated expression of the auxin inhibitors GH3.17 and SHY2 genes [7,9,10,11,12].
The maintenance of RAM is dependent on the proper functioning of the cell division cycle, which consists of four stages: an increase in cell size (Gap 1 or G1 stage), DNA synthesis (Synthesis or S stage), cell growth and preparation for cell division (Gap 2 or G2 stage), and cell division (Mitosis or M stage) [13]. The G0 stage is a non-proliferative state during which the cell enters a quiescent phase. Heterodimeric Ser/Thr protein kinases, comprising the catalytic subunit cyclin-dependent kinase (CDK) and the activating subunit cyclin (CYC), play a crucial role in driving cells through the different phases (G1/S/G2/M) of the cell cycle [14].
Plants contain a large number of cyclins, the vast majority of which are uncharacterized. During the G2 phase of the cell cycle, B-type cyclins are synthesized and reach their peak levels during prometaphase. This is the stage when the chromosomes condense and the spindle apparatus forms, preparing the cell for mitosis. In Arabidopsis, these cyclins play a regulatory role in root growth. The double mutant cycb1;1 cycb1;2 exhibits a 50% reduction in root growth compared to the wild type [15].
Cyclins are synthesized and degraded in a cyclic manner during the cell cycle, and their concentrations are highest during the specific phase of the cell cycle in which they are necessary. When the level of a specific cyclin decreases, the associated CDKs become less active, which leads to the cell entering the next stage of the cell cycle [13]. CDK-CYC complexes are important targets for both environmental and internal cellular signals. CDKs' cell division-promoting activity is suppressed by various pathways that act in parallel [16].
Several marker lines have been developed in Arabidopsis to visualize the spatiotemporal progression of the cell cycle. The use of a truncated version of the DNA replication origin licensing factor CDT1a, driven by the S-phase specific promoter of a histone 3.1-type gene, has been reported as a reliable marker for the S+G2 phase [17]. Otero et al. [18] demonstrated that the H3.1 protein is maintained during the M and G1 phases in frequently dividing cells of the root meristem. However, it is expelled during the G2 phase in cells undergoing their final cell cycle before differentiation in the root TZ. When used in combination with CyCB1, which is specific to the G2/M transition, it is possible to visualize both the S to G2 and G2 to M cell cycle stages [17,18]. More recently, Desvoyes and colleagues have developed the first Plant Cell Cycle Indicator (PlaCCI) marker line. This marker line expresses CDT1a-CFP, the histone protein H3.1-mCherry, and CYCB1;1-YFP driven by their own specific promoters. This development convincingly enables the identification of cell cycle stages in the Arabidopsis root tip [19]. Besides phytohormones, studies have also revealed that the folate derivative 5-formyl-tetrahydrofolate is an essential regulator of root meristem maintenance [20,21]. Folates are tripartite molecules composed of para-aminobenzoate, pterine, and one or several glutamate moieties [22]. They are essential elements in several important reactions, including biosynthesis of lipids, chlorophyll, and lignin, as well as the regulation of gene and protein expression [23,24]. These components, which are essential parts of the human diet due to their health benefits, are synthesized de novo only in plants, fungi, and bacteria. The first step in the folate biosynthesis pathway takes place in the cytosol, where pterine biosynthesis occurs, leading to the formation of 6-hydroxymethyldihydropterin (HDMDHP). This compound is then phosphorylated in the mitochondria to form 6-hydroxymethyldihydropterin pyrophosphate (HMDHP-P2) (reviewed by [23]).
The second precursor of folate is para-aminobenzoic acid (PABA), which is synthesized in plastids from chorismate and glutamine through a two-step reaction. This reaction is successively mediated by the aminodeoxychorismate lyase (ADCL) and the bifunctional GLUTAMINE AMIDOTRANSFERASE-AMINODEOXYCHORISMATE SYNTHASE (GAT-ADCS) [25,26]. More than 80% of PABA is esterified by uridine diphosphate (UDP)-glucosyltransferase (UGT75B1) to form PABA-glucose, which is then stored in the vacuole [23,27,28]. The remaining portion is utilized in the biosynthesis of folates within the mitochondria, where PABA combines with HMDHP-P2 to produce dihydropteroate (DHP). Afterward, a glutamate residue attaches to the carboxyl group of PABA in a reaction catalyzed by dihydrofolate reductase (DHFR) to produce tetrahydrofolate (THF). THF is then linked to additional glutamate molecules in mitochondria, plastids, or vacuoles [29,30].
It has long been believed that PABA activity is closely associated with the folate biosynthesis pathway. However, in a previous work, we demonstrated that PABA promotes asymmetric root growth during root gravitropism in an independent manner from folate. This suggests that endogenous PABA can act as a signaling molecule to modulate Arabidopsis root growth [31]. In the present study, we further examine the role of PABA in root growth and development. We demonstrate that artificially elevating PABA levels through the external addition of PABA to the growth medium or overexpression of the PABA synthesis gene GAT-ADCS significantly impedes root growth, and this is correlated with a smaller root meristem size due to fewer cells. On the contrary, reducing free PABA levels in the roots by conjugating PABA to glucose results in longer roots with extended meristems and increased cell numbers. In addition, we provide evidence that PABA activity affects root meristem size by deregulating the progression of root cell cycles. The levels of CYCB1;1, a crucial regulator of cell G2/M transition during mitosis, decrease in roots grown in the presence of PABA. This results in cells undergoing premature endoreduplication in the root TZ. This is indicated by the expression of the cyclin CDT1a-CFP marker, whose levels are known to increase in chromatin after mitosis, peak during G1, and rapidly decrease after S-phase initiation [19]. Therefore, PABA inhibits Arabidopsis root growth through two mechanisms: (i) by inhibiting the G2/M transition and (ii) by promoting the premature differentiation of cells in the transition zone. Taken together, this data supports the role of the folate precursor PABA in Arabidopsis root growth at the intersection of cell division and cell differentiation.
2. Results
2.1. PABA reduces the number of cells in the root meristem
We previously reported that the primary root of Arabidopsis is inhibited by PABA treatment in a dose-dependent manner [31]. To investigate the impact of PABA on root elongation, wild-type Arabidopsis (Col-0) seedlings were grown in the presence of PABA concentrations ranging from 0 to 400 µM (Figure 1A). The elongation of the primary root was measured over time, starting when the root radicle broke through the seed coat. Studying the root elongation over time revealed that exogenously applied PABA repressed root growth in a time- and concentration-dependent manner (Figure 1B). Root growth inhibition was delayed in the presence of 50 µM PABA and only occurred on day 6, while concentrations higher than 50 µM significantly repressed root growth starting from day 3 (Figure 1B). These data suggest that root inhibition starts very early, probably already when the radicle emerges.
Since the elongation of the primary root depends on cell production in the Root Apical Meristem (RAM), we quantified the size of the root meristematic zone (MZ) at day 6 by measuring and summing up the length of individual cortex cells in continuity, starting from the cortex cell initial to the first elongated cell (dashed line in Figure 1C). As expected, the inhibition of root elongation by PABA correlated well with the shortening of the MZ. With the highest reduction in meristem size (~33%) observed at 400µM. The application of 200µM, 100µM, and 50µM PABA resulted in a 25%, 8%, and 5% reduction in meristem size, respectively (Figure 1D). In addition, PABA also had a negative impact on the number of root cortex cells. Non-treated roots consisted of a meristem of 43±5 cortex cells (Figure 1E), this number is reduced to 40±5 cells in the presence of both 50µM and 100µM PABA (Figure 1E). The number of cells dramatically dropped by 12 and 16 cells at 200µM and 400µM PABA, respectively (Figure 1E). Because the reduction in both meristem size and meristematic cell number became more pronounced at concentrations of PABA as low as 200µM, this concentration was used for the rest of the study.
2.2. PABA regulation of root growth is independent of folates
To get insights whether the inhibitory effect of PABA on root growth is related to its role as folate precursor [26], Arabidopsis plants were grown for 7 days in the presence of PABA or 5-formyltetrahydrofolic acid (5-FTHF). 5-FTHF is a stable folate derivative that occurs naturally and can be easily converted into metabolically active folates once it is taken up by cells [26,32]. PABA, but not 5-FTHF, repressed root growth and significantly affected both RAM size and the number of meristematic cells (Figure S1). This suggests that PABA and 5-FTHF have different effects on root growth. Interestingly, when 1 mM 5-FTHF was applied, it promoted root growth, suggesting that 5-FTHF acts as a positive regulator of root development.
2.3. PABA negatively affects the G2/M transition of root meristematic cells
To check whether PABA acts on cell proliferation, we used Arabidopsis transgenic lines carrying pCycB1;2::GUS and pCycB1;2::GFP constructs, respectively, where the CYCB1;2 promoter and coding region, including the cyclin destruction box, is fused in frame to GUS [33] or GFP [34]. This enabled the visualization of the G2-M cell cycle transition and the degradation of the resulting CDB-GUS or CDB-GFP protein at the end of mitosis [34]. The number of cells expressing pCycB1;2::GUS was visibly reduced in roots treated with PABA (Figure 2A). The quantification of the GUS signal [35] showed that the intensity of the mean grey values was indeed significantly lower in the presence of PABA (Figure 2B). Similar results were obtained with the pCycB1;2::GFP marker line, where the CYCB1::GFP signal was detected in a significantly lower number of meristematic cells (Figure 2C). Additionally, the intensity of the GFP signal was also notably lower when PABA was present in the medium (Figure 2D). Taken together, these data suggest that PABA negatively impacts the G2/M transition of meristem cells.
To investigate the impact of PABA on other phases of the cell cycle, we utilized the Plant Cell Cycle Indicator (PlaCCI) marker line [19]. PlaCCI and WT were equally sensitive to 200µM PABA, and no significant difference in root size could be measured (Figure S2A,B). Next, in line with Desvoyes and colleagues [19], confocal microscopy images showed that the root tip of non-treated PlaCCI roots can be divided into three main regions (Figure 3): (i) the region 1 containing highly proliferative cells characterized by the expression of all markers, CDT1a-CFP (G1 phase), HTR13-mCherry (G1-S-G2-M phases) and CycB1;1 (G2/M phase); (ii) the region 2 occupied by cells in G1-phase, expressing CDT1a-CFP and H3.1-mCherry, but not CycB1;1-YFP ; (iii) and finally, the region 3 devoid of CycB1 expression, containing cells in G- and S-phases undergoing endocycle (Figure 3A,B and ). In PABA-treated roots, region 1 albeit smaller than in control roots, and region 2 can be easily recognized (Figure 3C,D and ). However, in line with a shorter meristem domain (magenta dashed line), the distal boundary of region 3, as indicated by the cells expressing CDT1a-CFP (white arrow in Figure 3A–D and Figure S2C), is much closer to the root tip. This suggests that meristem cells undergo earlier differentiation. Measuring the distance between QC and the first cell expressing CDT1a-CFP confirmed this assertion (Figure 3E).
Finally, to confirm whether cells had indeed differentiated earlier, we quantified the distance between the tip of the root and the first bulging root hair. We found that the root hairs bulged much closer to the root tip on PABA, as shown in Figure 3F.
2.4. Endogenous PABA is a key component in the regulation of Arabidopsis root growth
PABA is synthesized through a two-step reaction involving the plastidial bifunctional enzyme GLUTAMINE AMIDOTRANSFERASE-AMINODEOXYCHORISMATE SYNTHASE (GAT-ADCS) [25,26]. We previously reported that the GAT-ADCS gene is expressed in the QC, root cap cells, and root stem cells [31]. On the other hand, the DHPS gene, which is involved in PABA incorporation into the folate pathway, is expressed in columella cell initials and root cap cells [31]. This strongly suggests that PABA is indeed produced in the stem cell niche and meristematic zone. To investigate the significance of endogenous PABA on root growth, we created transgenic lines that overexpress the GAT-ADCS genomic sequence under the control of the strong constitutive CaMV35S promoter. Among the 19 lines analyzed (Figure S3A,B), we isolated the 35::ADCS-12 with high levels of ADCS mRNA (Figure S3C) and which accumulate twice as much PABA as the WT Col-0 (Figure S3D). Overexpression of the GAT-ADCS resulted in a reduction of both shoot and root growth (Figure 4A), suggesting that PABA content might be critical for shoot development in addition to its role in the root. All the root length, the size of the root meristem, and the number of cells in the meristem were significantly reduced by 30-40% in this line (Figure 4B–D). Similar results, but not as pronounced, were obtained using a mutant lacking the UDP-GLYCOSYL TRANSFERASE75B1 (UGT75B1; [28]), the enzyme that catalyzes the glycosylation of PABA, resulting in plants over-accumulating free PABA [28]. Compared to the wild type (WT), the Landsberg erecta (Ler) ugt75b1 knock-out mutant developed primary roots that were 15% shorter, with a reduction in meristem size by 5% and a decrease in the number of meristematic cells by 10% (Figure 4E–H).
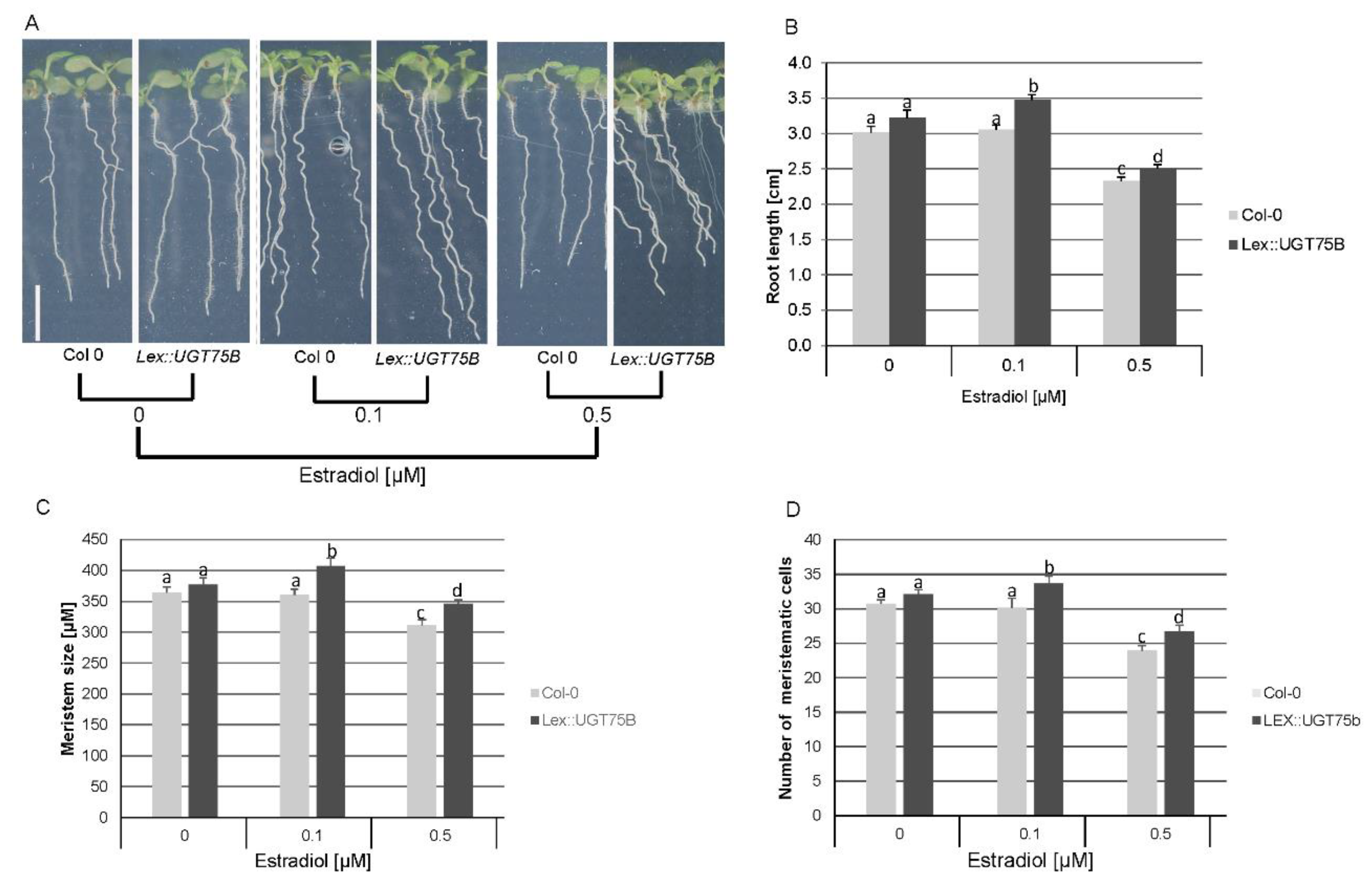
Conversely, to artificially reduce the amount of free PABA, we utilized a stable conditional line that overexpresses UGT75B1 (Lex::UGT75B; [31]). This line, when compared to the wild type (WT), accumulates PABA glucosylates (PABA-Glc) upon β-estradiol induction [31]. In the absence of ß-estradiol, the roots of Lex::UGT75B plants appeared slightly longer, with larger meristems containing more cells than WT plants (Figure 5A–D). This trend can be explained by the reported leakiness of the β-estradiol inducible system [31,36]. At 0.1 µM, ß-estradiol had no impact on the growth of the WT root, but it stimulated a 14% increase in the elongation of the Lex::UGT75B primary root (Figure 5A,B). This stimulation was only 7% greater when compared to non-induced Lex::UGT75B control roots (Figure 5A,B). The promotion of root growth was accompanied by an increase in meristem size and the number of meristematic cells (Figure 5C,D). However, increasing β-estradiol concentration to 0.5 µM significantly impacted the growth of both WT and Lex::UGT75B roots, and the differences reported above for 0.1 µM became less pronounced (Figure 5A–D). The primary root of Lex::UGT75B was only 7% longer than that of ß-estradiol-treated WT plants, and the size of the meristem and the number of meristematic cells were about 10% higher compared to WT (Figure 5B–D). These data suggest that a slight decrease in free PABA levels leads to an increase in meristematic cell number and meristem size, promoting root growth. Conversely, increasing free PABA levels would reduce cell cycle activity, resulting in a decrease in the number of meristematic cells and limiting root growth. Maybe the rate of free PABA conversion to PABA-Glc in the presence of 0.5µM leads to a deficiency of free PABA, which is also necessary for proper cell division, root meristem maintenance, and root development and maintenance.
3. Discussion
The makeup of the plant relies on its ability to convert both biotic and abiotic signals into developmental responses. Therefore, the sustained growth of the root, which results from a balance between the production of new cells in the RAM and their progressive differentiation in the TZ, is a fundamental asset for plant adaptability to various ecological niches. However, the molecular and physiological mechanisms that sustain the continuous growth of the root are not fully understood. In this study, we explored the role of the folate precursor PABA in root growth and development using pharmacological, genetic, and imaging approaches.
3.1. PABA represses G2/M transition and promotes endocycle
Because PABA-treated roots are shorter (this study; [31,37]) and display a smaller meristem, which consists of fewer cells compared to untreated roots, we hypothesized that a defect in cell division alters the development of these roots. We found that the number of cells expressing the B-type cyclin CycB1;1, which is crucial for the G2/M cell cycle transition, was dramatically reduced in PABA-treated roots. As a consequence, the number of cells undergoing the G2/M transition is indeed significantly reduced in the presence of PABA. CycB1;1 is synthesized during the G2 phase, peaks during the prometaphase, and disappears at early anaphase [38]. Hence, by affecting CycB1;1 expression, PABA prevents meristem cells from fully completing their mitotic cycle, thereby causing them to lose their proliferation potential.
In untreated roots, the transition between cell division and cell elongation is progressive as cells enter the elongation zone. They express the DNA replication origin licensing factor CDT1a and the G2 marker H3.1, but not the G2/M marker CycB1;1 (present study; [17,19]), indicating their G1 status and that they are undergoing endocycle [19]. The fact that this transition occurs much closer to the root tip in PABA-treated plants suggests that PABA promotes the shift of meristematic cells from the mitotic cell cycle to the endocycle, an alternate cell cycle during which cells replicate their genetic material but do not split [39]. Thus, by repressing CyCB1;1, PABA inhibits cells from completing mitosis and cytokinesis under conditions that would still allow error free chromosomes replication to occur [40]. Indeed, we never observed apoptotic cells, and the fact that PABA only slows down the growth (even when applied at high concentrations) but never abolishes it also supports this hypothesis. However, it cannot be excluded that when occurring, high concentrations of free PABA might be hindered by increased levels of PABA glucosylation by the UGT75B enzyme. How PABA acts on CycB1;1 is still an open question. However, we showed in a previous study that PABA activity on root gravitropism requires ethylene biosynthesis [31]. This is particularly interesting since ethylene, known to repress the growth of the Arabidopsis root, was reported to act through a mechanism involving the reduction of the CycB1;1 protein level [41]. This highlights the possibility that PABA may modulate root growth in an ethylene-dependent post-translational regulatory mechanism controlling CyCB1;1 stability [41].
3.2. PABA and auxin display distinct activities on root growth
The importance of plant hormones crosstalk in maintaining the continuous growth of the root has been clearly demonstrated [42,43,44]. For instance, the action of cytokinins through the activation of SHY2/IAA3 (SHY2), the negative regulator of PIN auxin transporters genes, by primary cytokinin- response transcription factor ARR1 in the root transition zone, can reduce auxin distribution and thereby promote cell differentiation [10]. The gaseous hormone ethylene also can repress root elongation by two distinct mechanisms. A significant portion of the inhibitory ethylene effect on root growth was suggested to be mediated by the local stimulation of the auxin response in the meristem and EZ epidermis [42,45]. However, the existence of yet to be described auxin independent mechanisms was also evidenced [45].
Interestingly, using theoretical calculations and crystallographic analyses, it was reported that PABA in its anionic form (PABA-) would bind the active site of the auxin receptor TIR1, similarly to the most important naturally occurring auxin, the indole acetic acid (IAA) [37]. This led to the conclusion that PABA might act as a potential auxin-like plant growth regulator [37,46]. IAA indeed inhibits root growth even at very low concentrations (nanomolar range) [47,48,49], whereas PABA inhibition of root growth is visible from 50 μM (this study; [31,37]). This concentration dependent activity, may account from the fact that IAA exhibits a slightly stronger binding to TIR1, than PABA [31]. However, contrary to the previously mentioned inhibitory effect of PABA on cell division, it was observed that elevated concentrations of auxin stimulate the root cell cycle and impede the transition to endocycle, through the activation of CDKs and mitotic cyclin production in unison with cytokinin [10,50,51,52]. Consistent with this, disrupting auxin response in root cell files results in repression of cell cycle genes including CycB1;1 [52,53]. Thus, although both compounds inhibit root growth, one (auxin) positively influences cell division and negatively endoreduplication, whereas the other (PABA) inhibits cell cycle-G2/M-transition and promotes cell differentiation.
Further, we provide evidence that root endogenous PABA levels are important for the balance proliferation/differentiation of cells at the root tip. Artificially increasing or decreasing endogenous PABA levels respectively reduce or promote meristem size. PABA acts in a folate-independent manner since PABA activity on meristem size maintenance is distinct from that of tetrahydrofolate. PABA is synthesized in QC and lateral root cap cells and is probably redistributed at the root tip through diffusion. Indeed, as a weak acid PABA might enter cells in its uncharged form [31].
3.3. Conclusions and future prospects
In this study, we present a mechanism by which the folate precursor PABA modulates root development by regulating the G2/M transition of the cell cycle. The activity of PABA on the cell cycle appears to be similar to that of ethylene, suggesting that PABA may act, at least partially, upstream of the ethylene pathway. It is interesting to note that cells enter the endocycle when they pass through a region with an auxin minimum, which represents the boundary between proliferating and differentiating cells [6]. We do not yet know with certainty whether PABA interacts with auxin in this region. Further studies are needed to determine whether PABA also affects cell elongation, which appears to be the primary target of auxin [42]. However, our previous study demonstrated that PABA's effect on root gravitropic response relies on the auxin biosynthesis TAA1 gene expression in the transition zone, likely through an ethylene-dependent mechanism [31]. Therefore, an increase in PABA-promoted auxin levels could, in theory, result in a PABA-induced auxin-dependent inhibition of cell growth in the root region. Altogether, these observations support the idea that PABA might regulate root growth through two mechanisms involving both auxin and ethylene signaling. Specifically, PABA would act at the point where cell proliferation and cell differentiation intersect by repressing cell growth and inhibiting cell division.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Apart from ugt75b (GT6017) in the Landsberg erecta background, all genetic backgrounds were from the Arabidopsis (Arabidopsis thaliana) ecotype Col-0. pCycB1;2::GUS [33], pCycB1;2::GFP [34], Lex::UGT75b [31], and PlaCCl seeds [19] were kindly provided by Prof. C. Gutierrez (Centro de Biologia Molecular Severo Ochoa, CSIC-UAM, Cantoblanco, Madrid, Spain). Seeds were surface-sterilized using a solution of 5% (w/v) calcium hypochlorite and 0.02% (v/v) Triton X-100 in 80% (v/v) ethanol. Then, they were rinsed twice with 80% (v/v) ethanol and once with 100% (v/v) ethanol. For all experiments, seedlings were grown vertically in Petri dishes tilted at an angle of 60° from the vertical axis. The seedlings were placed on a solid 1.3% (w/v) agar Arabidopsis medium, which was prepared by adding 0.5× basal salt Murashige and Skoog medium supplemented with 1% (w/v) Sucrose and 5 mM MES (pH 5.8). The growth conditions were maintained in a climatic cabinet at 21°C, under long-day conditions (16 hours of light and 8 hours of dark), with a humidity level of 70%.
4.2. Construction of plasmids
Plasmids were constructed according to the protocol described by Sambrook et al. [54]. Gateway® cloning procedures (Invitrogen) were used, and E. coli TOPO10 cells (Invitrogen) were used as the host for the constructed plasmids. GAT-ADCS cDNA was obtained from RIKEN, amplified using cADCS_F (CTGTCGTTATTCGGAATGATGAGT) and cADCS_R (GAAAGCAGCTCTTACATTCCCAC) oligonucleotides and cloned into the pDONR207 vector. Subsequently, it was cloned into the pMDC32 vector to generate the 35S::GAT-ADCS construct.
4.3. Plant Transformation
Arabidopsis plants were transformed using the agrobacterium strain GV3101 (pMP90) according to the standard floral dip method [55]. Transformants were selected on 15 μg/ml Hygromycin B.
4.4. Plant Scanning and Microscopy
Seedlings were imaged using a flatbed CanonScan 9950F scanner while they were growing on a plastic Petri dish. Histological detection of GUS activity and plant preparation for microscopy were performed according to Ditengou et al. [56]. For light microscopy, samples were observed using a Zeiss Axiovert 200M MOT device (Carl Zeiss MicroImaging) to capture high-magnification images. Low-magnification views were obtained using a Zeiss Stemi SV11 Apo stereomicroscope (Carl Zeiss MicroImaging) and observed under differential interference contrast optics. Plants expressing fluorescent proteins were stained with 5 µg/ml FM4-64, a lipophilic probe that binds to cell plasma membranes. The stained plants were then analyzed using Nikon's AZ-C1 Macro Laser Confocal Microscope and the Zeiss LSM 980 laser scanning microscope. To simultaneously monitor DAPI, YFP, and mCherry fluorescence, we employed multitracking in-frame mode, and the emission was separated using the META spectral analyzer online unmixing feature. Images were extracted and analyzed using Zen software (Carl Zeiss MicroImaging) and Imaris 9.8.0 (Oxford Instruments). All figures were created using Inkscape software.
4.5. Quantification of Root Meristem Size and Number of Meristematic Cells
To measure the size of the root meristem and the number of meristematic cells, we collected roots from Arabidopsis seedlings of different genotypes that were grown for 6 days with or without PABA. The collected roots were then stained with 5 µM FM4-64 for 5 minutes. After a quick rinse, the roots were scanned using a Nikon C1 confocal microscope. The size of the meristem and the number of cortex cells were calculated following the method described by Takasuka et al. [57].
4.6. Quantification of GFP Signals
GFP quantifications were performed by selecting a meristem region that included the quiescent center (QC) and stem cells up to the first elongated cortex cell. Fluorescence intensity was measured using ImageJ software.
4.7. Quantification of GUS Signals
GUS quantifications were performed by selecting a meristem region that included the quiescent center (QC) and stem cells up to the first elongated cortex cell. Signal intensity was measured using ImageJ software, as described by Beziat et al. [35].
4.8. Quantification of eCFP/mCherry and YFP Signals
Proliferating cells in the epidermis of the root apical meristem were measured in seven roots growing on medium with or without PABA, according to the method described by Desvoyes et al. [19].
4.9. Quantification of free PABA
Free and total PABA determinations were essentially performed according to the method described by Camara et al. [26].
4.10. Accession Numbers
The sequence data from this article can be found in the GenBank/EMBL database and the Arabidopsis Genome Initiative database under the following accession number: At1g05560 (UGT75B1).
Supplementary Materials
The following supporting information can be downloaded at: at the website of this paper posted on Preprints.org, Figure S1: Comparison of the impact of PABA and 5-FTHF on the root growth; Figure S2: Use of the PlaCCI reporter line to visualize the root cell cycle state. Figure S3: Characterization of ADCS overexpressing lines.
Author Contributions
Conceptualization, F.A.D.; Methodology, H.L., H. N., and F.A.D.; Investigation, H.L., P.K. and F.A.D.; Data curation, H.L., H.N., P.K. and F.A.D.; H.L., H.N., and F.A.D have participated in the redaction of this publication; Supervision, F.A.D.; Funding acquisition, H.L. and F.A.D.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the POLISH NATIONAL SCIENCE CENTRE grant number 2019/03/X/NZ3/01217, and supported by the Medical Faculty, University of Freiburg (Project Number 2021/B3-Fol) and Lighthouse Core Facility (RRID:SCR_023785). Hanna Lasok was supported by the DEUTSCHER AKADEMISCHER AUSTAUSCHDIENST (DAAD) Fellowship.
Data Availability Statement
Data are available upon request from the corresponding author.
Acknowledgments
This work could not have been completed without the support, recommendations, and materials offered by coworkers, collaborators, and friends. The authors specially thank Prof. Klaus Palme for fruitful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Smith, S.; De Smet, I. Root system architecture: insights from Arabidopsis and cereal crops. Philos Trans R Soc Lond B Biol Sci 2012, 367, 1441–1452. [Google Scholar] [CrossRef]
- Tian, H.; De Smet, I.; Ding, Z. Shaping a root system: regulating lateral versus primary root growth. Trends Plant Sci 2014, 19, 426–431. [Google Scholar] [CrossRef]
- Larkin, J.C.; Brown, M.L.; Schiefelbein, J. How do cells know what they want to be when they grow up? Lessons from epidermal patterning in Arabidopsis. Annu Rev Plant Biol 2003, 54, 403–430. [Google Scholar] [CrossRef]
- Verbelen, J.P.; De Cnodder, T.; Le, J.; Vissenberg, K.; Baluska, F. The Root Apex of Arabidopsis thaliana Consists of Four Distinct Zones of Growth Activities: Meristematic Zone, Transition Zone, Fast Elongation Zone and Growth Terminating Zone. Plant Signal Behav 2006, 1, 296–304. [Google Scholar] [CrossRef]
- Salvi, E.; Di Mambro, R.; Sabatini, S. Dissecting mechanisms in root growth from the transition zone perspective. J Exp Bot 2020, 71, 2390–2396. [Google Scholar] [CrossRef]
- Di Mambro, R.; De Ruvo, M.; Pacifici, E.; Salvi, E.; Sozzani, R.; Benfey, P.N.; Busch, W.; Novak, O.; Ljung, K.; Di Paola, L.; et al. Auxin minimum triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Proc Natl Acad Sci U S A 2017, 114, E7641–E7649. [Google Scholar] [CrossRef]
- Dello Ioio, R.; Linhares, F.S.; Scacchi, E.; Casamitjana-Martinez, E.; Heidstra, R.; Costantino, P.; Sabatini, S. Cytokinins determine Arabidopsis root-meristem size by controlling cell differentiation. Curr Biol 2007, 17, 678–682. [Google Scholar] [CrossRef]
- Liu, S.; Strauss, S.; Adibi, M.; Mosca, G.; Yoshida, S.; Dello Ioio, R.; Runions, A.; Andersen, T.G.; Grossmann, G.; Huijser, P.; et al. Cytokinin promotes growth cessation in the Arabidopsis root. Curr Biol 2022, 32, 1974–1985. [Google Scholar] [CrossRef]
- Moubayidin, L.; Perilli, S.; Dello Ioio, R.; Di Mambro, R.; Costantino, P.; Sabatini, S. The rate of cell differentiation controls the Arabidopsis root meristem growth phase. Curr Biol 2010, 20, 1138–1143. [Google Scholar] [CrossRef]
- Dello Ioio, R.; Nakamura, K.; Moubayidin, L.; Perilli, S.; Taniguchi, M.; Morita, M.T.; Aoyama, T.; Costantino, P.; Sabatini, S. A genetic framework for the control of cell division and differentiation in the root meristem. Science 2008, 322, 1380–1384. [Google Scholar] [CrossRef]
- Perilli, S.; Sabatini, S. Analysis of root meristem size development. Methods Mol Biol 2010, 655, 177–187. [Google Scholar] [CrossRef]
- Kong, X.; Liu, G.; Liu, J.; Ding, Z. The Root Transition Zone: A Hot Spot for Signal Crosstalk. Trends Plant Sci 2018, 23, 403–409. [Google Scholar] [CrossRef]
- Pederson, T. Historical review: an energy reservoir for mitosis, and its productive wake. Trends Biochem Sci 2003, 28, 125–129. [Google Scholar] [CrossRef]
- Barnum, K.J.; O'Connell, M.J. Cell cycle regulation by checkpoints. Methods Mol Biol 2014, 1170, 29–40. [Google Scholar] [CrossRef]
- Romeiro Motta, M.; Zhao, X.; Pastuglia, M.; Belcram, K.; Roodbarkelari, F.; Komaki, M.; Harashima, H.; Komaki, S.; Kumar, M.; Bulankova, P.; et al. B1-type cyclins control microtubule organization during cell division in Arabidopsis. EMBO Rep 2022, 23, e53995. [Google Scholar] [CrossRef]
- Bhosale, R.; Maere, S.; De Veylder, L. Endoreplication as a potential driver of cell wall modifications. Current Opinion in Plant Biology 2019, 51, 58–65. [Google Scholar] [CrossRef]
- Yin, K.; Ueda, M.; Takagi, H.; Kajihara, T.; Sugamata Aki, S.; Nobusawa, T.; Umeda-Hara, C.; Umeda, M. A dual-color marker system for in vivo visualization of cell cycle progression in Arabidopsis. Plant J 2014, 80, 541–552. [Google Scholar] [CrossRef]
- Otero, S.; Desvoyes, B.; Peiro, R.; Gutierrez, C. Histone H3 Dynamics Reveal Domains with Distinct Proliferation Potential in the Arabidopsis Root. Plant Cell 2016, 28, 1361–1371. [Google Scholar] [CrossRef]
- Desvoyes, B.; Arana-Echarri, A.; Barea, M.D.; Gutierrez, C. A comprehensive fluorescent sensor for spatiotemporal cell cycle analysis in Arabidopsis. Nat Plants 2020, 6, 1330–1334. [Google Scholar] [CrossRef]
- Reyes-Hernandez, B.J.; Srivastava, A.C.; Ugartechea-Chirino, Y.; Shishkova, S.; Ramos-Parra, P.A.; Lira-Ruan, V.; Diaz de la Garza, R.I.; Dong, G.; Moon, J.C.; Blancaflor, E.B.; et al. The root indeterminacy-to-determinacy developmental switch is operated through a folate-dependent pathway in Arabidopsis thaliana. New Phytol 2014, 202, 1223–1236. [Google Scholar] [CrossRef]
- Srivastava, A.C.; Ramos-Parra, P.A.; Bedair, M.; Robledo-Hernández, A.L.; Tang, Y.; Sumner, L.W.; Díaz de la Garza, R.I.; Blancaflor, E.B. The folylpolyglutamate synthetase plastidial isoform is required for postembryonic root development in Arabidopsis. Plant Physiol 2011, 155, 1237–1251. [Google Scholar] [CrossRef]
- Kołton, A.; Długosz-Grochowska, O.; Wojciechowska, R.; Czaja, M. Biosynthesis Regulation of Folates and Phenols in Plants. Scientia Horticulturae 2022, 291, 110561. [Google Scholar] [CrossRef]
- Gorelova, V.; Ambach, L.; Rebeille, F.; Stove, C.; Van Der Straeten, D. Folates in Plants: Research Advances and Progress in Crop Biofortification. Front Chem 2017, 5, 21. [Google Scholar] [CrossRef]
- Rebeille, F.; Ravanel, S.; Jabrin, S.; Douce, R.; Storozhenko, S.; Van Der Straeten, D. Folates in plants: biosynthesis, distribution, and enhancement. Physiologia Plantarum 2006, 126, 330–342. [Google Scholar] [CrossRef]
- Basset, G.J.C.; Ravanel, S.; Quinlivan, E.P.; White, R.; Giovannoni, J.J.; Rebeille, F.; Nichols, B.P.; Shinozaki, K.; Seki, M.; Gregory, J.F., III; et al. Folate synthesis in plants: the last step of the p-aminobenzoate branch is catalyzed by a plastidial aminodeoxychorismate lyase. Plant Journal 2004, 40, 453–461. [Google Scholar] [CrossRef]
- Camara, D.; Bisanz, C.; Barette, C.; Van Daele, J.; Human, E.; Barnard, B.; Van Der Straeten, D.; Stove, C.P.; Lambert, W.E.; Douce, R.; et al. Inhibition of p-Aminobenzoate and Folate Syntheses in Plants and Apicomplexan Parasites by Natural Product Rubreserine. Journal of Biological Chemistry 2012, 287, 22367–22376. [Google Scholar] [CrossRef]
- Quinlivan, E.P.; Roje, S.; Basset, G.; Shachar-Hill, Y.; Gregory, J.F., III; Hanson, A.D. The folate precursor p-aminobenzoate is reversibly converted to its glucose ester in the plant cytosol. Journal of Biological Chemistry 2003, 278, 20731–20737. [Google Scholar] [CrossRef]
- Eudes, A.; Bozzo, G.G.; Waller, J.C.; Naponelli, V.; Lim, E.-K.; Bowles, D.J.; Gregory, J.F., III; Hanson, A.D. Metabolism of the folate precursor p-aminobenzoate in plants - Glucose ester formation and vacuolar storage. Journal of Biological Chemistry 2008, 283, 15451–15459. [Google Scholar] [CrossRef]
- Ravanel, S.; Cherest, H.; Jabrin, S.; Grunwald, D.; Surdin-Kerjan, Y.; Douce, R.; Rébeillé, F. Tetrahydrofolate biosynthesis in plants: molecular and functional characterization of dihydrofolate synthetase and three isoforms of folylpolyglutamate synthetase in Arabidopsis thaliana. Proc Natl Acad Sci U S A 2001, 98, 15360–15365. [Google Scholar] [CrossRef]
- Orsomando, G.; de la Garza, R.D.; Green, B.J.; Peng, M.; Rea, P.A.; Ryan, T.J.; Gregory, J.F., 3rd; Hanson, A.D. Plant gamma-glutamyl hydrolases and folate polyglutamates: characterization, compartmentation, and co-occurrence in vacuoles. J Biol Chem 2005, 280, 28877–28884. [Google Scholar] [CrossRef]
- Nziengui, H.; Lasok, H.; Kochersperger, P.; Ruperti, B.; Rebeille, F.; Palme, K.; Ditengou, F.A. Root Gravitropism Is Regulated by a Crosstalk between para-Aminobenzoic Acid, Ethylene, and Auxin. Plant Physiol 2018, 178, 1370–1389. [Google Scholar] [CrossRef]
- Li, W.; Liang, Q.; Mishra, R.C.; Sanchez-Mu Oz, R.; Wang, H.; Chen, X.; Van Der Straeten, D.; Zhang, C.; Xiao, Y. The 5-formyl-tetrahydrofolate proteome links folates with C/N metabolism and reveals feedback regulation of folate biosynthesis. Plant Cell 2021, 33, 3367–3385. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, P.M.; Bonetta, D.; Tsukaya, H.; Dengler, R.E.; Dengler, N.G. Cell cycling and cell enlargement in developing leaves of Arabidopsis. Dev Biol 1999, 215, 407–419. [Google Scholar] [CrossRef]
- Ubeda-Tomas, S.; Federici, F.; Casimiro, I.; Beemster, G.T.S.; Bhalerao, R.; Swarup, R.; Doerner, P.; Haseloff, J.; Bennett, M.J. Gibberellin Signaling in the Endodermis Controls Arabidopsis Root Meristem Size. Current Biology 2009, 19, 1194–1199. [Google Scholar] [CrossRef]
- Beziat, C.; Kleine-Vehn, J.; Feraru, E. Histochemical Staining of beta-Glucuronidase and Its Spatial Quantification. Methods Mol Biol 2017, 1497, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Kubo, M.; Imai, A.; Nishiyama, T.; Ishikawa, M.; Sato, Y.; Kurata, T.; Hiwatashi, Y.; Reski, R.; Hasebe, M. System for stable beta-estradiol-inducible gene expression in the moss Physcomitrella patens. PLoS One 2013, 8, e77356. [Google Scholar] [CrossRef]
- Crisan, M.E.; Bourosh, P.; Maffei, M.E.; Forni, A.; Pieraccini, S.; Sironi, M.; Chumakov, Y.M. Synthesis, crystal structure and biological activity of 2-hydroxyethylammonium salt of p-aminobenzoic acid. PLoS One 2014, 9, e101892. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.; Rosenthal, E.T.; Youngblom, J.; Distel, D.; Hunt, T. Cyclin: a protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division. Cell 1983, 33, 389–396. [Google Scholar] [CrossRef]
- Edgar, B.A.; Orr-Weaver, T.L. Endoreplication cell cycles: more for less. Cell 2001, 105, 297–306. [Google Scholar] [CrossRef]
- Desvoyes, B.; Fernandez-Marcos, M.; Sequeira-Mendes, J.; Otero, S.; Vergara, Z.; Gutierrez, C. Looking at plant cell cycle from the chromatin window. Frontiers in Plant Science 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Street, I.H.; Aman, S.; Zubo, Y.; Ramzan, A.; Wang, X.; Shakeel, S.N.; Kieber, J.J.; Schaller, G.E. Ethylene Inhibits Cell Proliferation of the Arabidopsis Root Meristem. Plant Physiol 2015, 169, 338–350. [Google Scholar] [CrossRef]
- Ruzicka, K.; Ljung, K.; Vanneste, S.; Podhorska, R.; Beeckman, T.; Friml, J.; Benkova, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef]
- Takahashi, N.; Kajihara, T.; Okamura, C.; Kim, Y.; Katagiri, Y.; Okushima, Y.; Matsunaga, S.; Hwang, I.; Umeda, M. Cytokinins control endocycle onset by promoting the expression of an APC/C activator in Arabidopsis roots. Curr Biol 2013, 23, 1812–1817. [Google Scholar] [CrossRef]
- Sablowski, R. Coordination of plant cell growth and division: collective control or mutual agreement? Curr Opin Plant Biol 2016, 34, 54–60. [Google Scholar] [CrossRef]
- Stepanova, A.N.; Yun, J.; Likhacheva, A.V.; Alonso, J.M. Multilevel interactions between ethylene and auxin in Arabidopsis roots. Plant Cell 2007, 19, 2169–2185. [Google Scholar] [CrossRef]
- Sumalan, R.L.; Halip, L.; Maffei, M.E.; Croitor, L.; Siminel, A.V.; Radulov, I.; Sumalan, R.M.; Crisan, M.E. Bioprospecting Fluorescent Plant Growth Regulators from Arabidopsis to Vegetable Crops. International Journal of Molecular Sciences 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Serre, N.B.C.; Kralik, D.; Yun, P.; Slouka, Z.; Shabala, S.; Fendrych, M. AFB1 controls rapid auxin signalling through membrane depolarization in Arabidopsis thaliana root. Nat Plants 2021, 7, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, M.; Jowett, J.; Stirnberg, P.; Rouse, D.; Leyser, O. pax1-1 partially suppresses gain-of-function mutations in Arabidopsis AXR3/IAA17. BMC Plant Biol 2007, 7, 20. [Google Scholar] [CrossRef]
- Fendrych, M.; Akhmanova, M.; Merrin, J.; Glanc, M.; Hagihara, S.; Takahashi, K.; Uchida, N.; Torii, K.U.; Friml, J. Rapid and reversible root growth inhibition by TIR1 auxin signalling. Nat Plants 2018, 4, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Himanen, K.; Boucheron, E.; Vanneste, S.; de Almeida Engler, J.; Inze, D.; Beeckman, T. Auxin-mediated cell cycle activation during early lateral root initiation. Plant Cell 2002, 14, 2339–2351. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Lescot, M.; Inze, D.; De Veylder, L. Effect of auxin, cytokinin, and sucrose on cell cycle gene expression in Arabidopsis thaliana cell suspension cultures. Plant Cell Tissue and Organ Culture 2002, 69, 167–176. [Google Scholar] [CrossRef]
- Ishida, T.; Adachi, S.; Yoshimura, M.; Shimizu, K.; Umeda, M.; Sugimoto, K. Auxin modulates the transition from the mitotic cycle to the endocycle in Arabidopsis. Development 2010, 137, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Zhang, F.; Wang, J.; Wang, F.; Liu, J.; Xie, C.; Hu, Y.; Shani, E.; Kong, X.; Ding, Z.; et al. Cell-type action specificity of auxin on Arabidopsis root growth. Plant J 2021, 106, 928–941. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular cloning: a laboratory manual; Cold spring harbor laboratory press: 1989.
- Clough, S.J.; Bent, A.F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 1998, 16. [Google Scholar] [CrossRef]
- Ditengou, F.A.; Teale, W.D.; Kochersperger, P.; Flittner, K.A.; Kneuper, I.; van der Graaff, E.; Nziengui, H.; Pinosa, F.; Li, X.; Nitschke, R.; et al. Mechanical induction of lateral root initiation in Arabidopsis thaliana. Proc Natl Acad Sci U S A 2008, 105, 18818–18823. [Google Scholar] [CrossRef]
- Takatsuka, H.; Higaki, T.; Umeda, M. Actin Reorganization Triggers Rapid Cell Elongation in Roots. Plant Physiol 2018, 178, 1130–1141. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The impact of PABA on Arabidopsis primary root growth. (A) Root size of 6-day-old plants grown in 0 (control), 50, 100, 200, and 400 μM PABA-supplemented media. (B) Primary root elongation over time (n=45). (C) Representative root tips of 6-day-old plants stained with the plasma membrane marker FM4-64 (red), grown in the absence (control) or presence of 200 μM PABA. The white dashed lines depict the limits of the meristematic zone. Bar, 100µm. (D, E) Size of the meristem and number of cortex cells in the meristem. Data are shown as means ± se (n > 20 roots). Meristem size, or cortex cell number, is considered different from the control at P < 0.05. One-way ANOVA with Bonferroni multiple testing corrections was used to attest to the differences between treatment groups. The letters (a, b, c, d) indicate independent groups according to one-way ANOVA.
Figure 1.
The impact of PABA on Arabidopsis primary root growth. (A) Root size of 6-day-old plants grown in 0 (control), 50, 100, 200, and 400 μM PABA-supplemented media. (B) Primary root elongation over time (n=45). (C) Representative root tips of 6-day-old plants stained with the plasma membrane marker FM4-64 (red), grown in the absence (control) or presence of 200 μM PABA. The white dashed lines depict the limits of the meristematic zone. Bar, 100µm. (D, E) Size of the meristem and number of cortex cells in the meristem. Data are shown as means ± se (n > 20 roots). Meristem size, or cortex cell number, is considered different from the control at P < 0.05. One-way ANOVA with Bonferroni multiple testing corrections was used to attest to the differences between treatment groups. The letters (a, b, c, d) indicate independent groups according to one-way ANOVA.
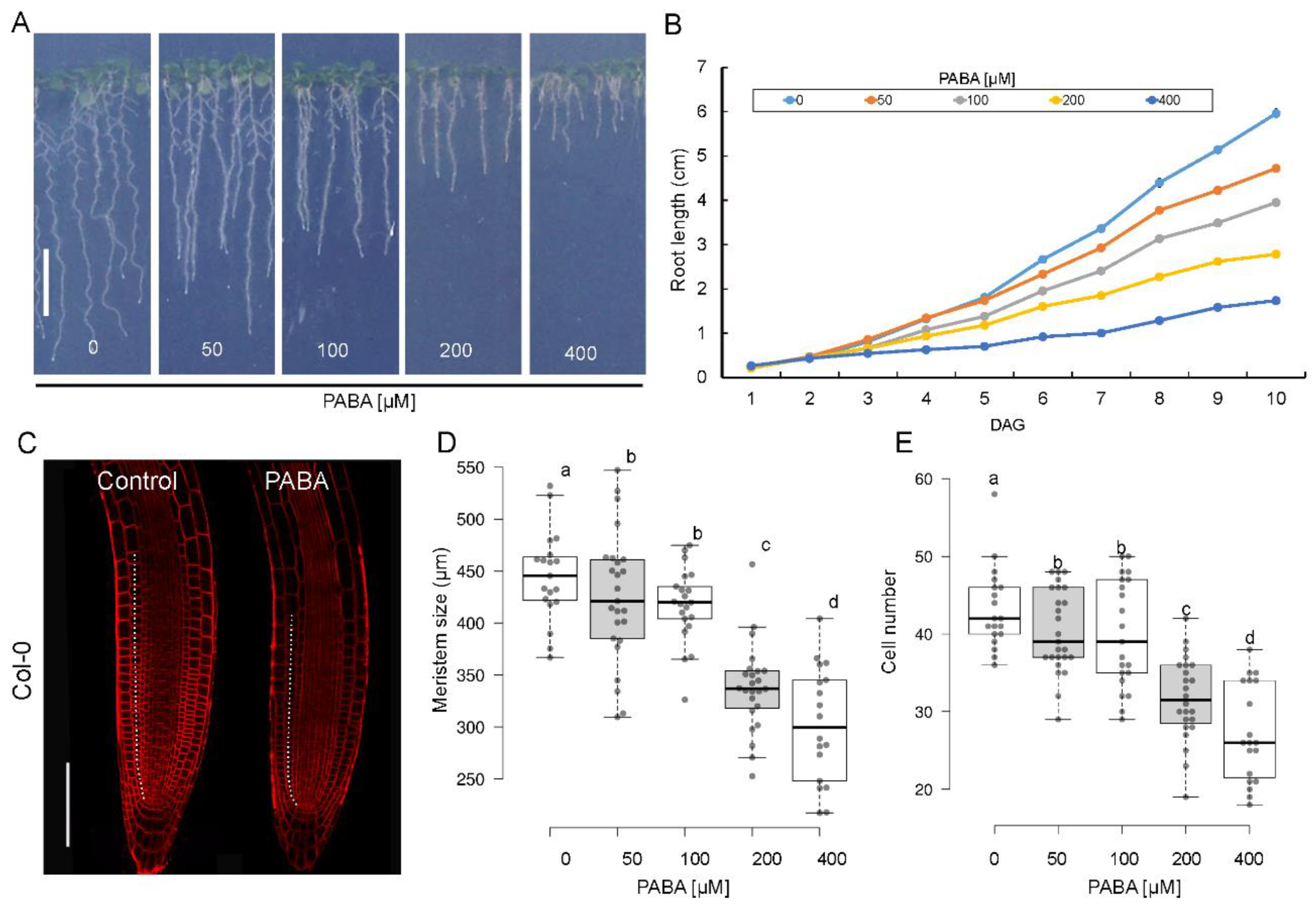
Figure 2.
Expression pattern of CYCB1 at the root tip. (A) Visualization of GUS staining of 6-day-old seedlings expressing the pCycB1;2::GUS-CDB construct grown on control and 200 μM PABA media. (B) Quantification of the GUS signal from plants shown in (A). (C) Visualization of CYCB1;2::CYCB1;2-YFP signal in 6-day-old seedlings grown in the absence (control) and presence of 200 μM PABA. (D) Quantification of the GFP signal from plants shown in (C). Data are shown as means ± se (n = 28 for B and 11 for D). Asterisks indicate statistical significance (P < 0.001 based on Student’s t test). A.u., arbitrary units. Scale, 100µm.
Figure 2.
Expression pattern of CYCB1 at the root tip. (A) Visualization of GUS staining of 6-day-old seedlings expressing the pCycB1;2::GUS-CDB construct grown on control and 200 μM PABA media. (B) Quantification of the GUS signal from plants shown in (A). (C) Visualization of CYCB1;2::CYCB1;2-YFP signal in 6-day-old seedlings grown in the absence (control) and presence of 200 μM PABA. (D) Quantification of the GFP signal from plants shown in (C). Data are shown as means ± se (n = 28 for B and 11 for D). Asterisks indicate statistical significance (P < 0.001 based on Student’s t test). A.u., arbitrary units. Scale, 100µm.
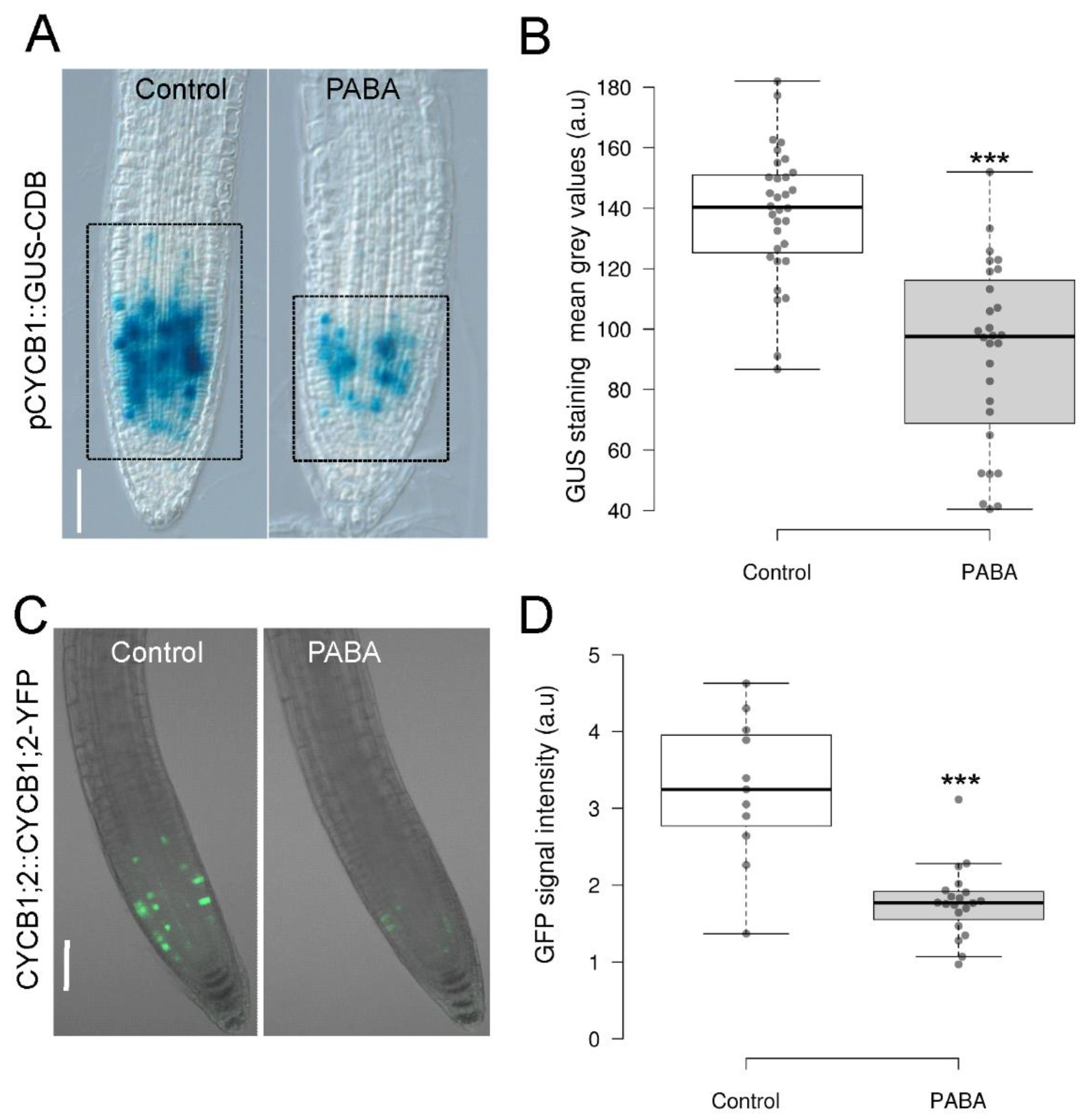
Figure 3.
Use of the PlaCCI reporter line to visualize the root cell cycle state. Confocal images displaying the root tips of 6 day-old PlaCCI plants expressing CDT1a-CFP, HTR13-mCherry, and CycB1;1-YFP constructs and grown in the absence (control) or presence of 200 μM PABA. (A-B), control root. (A) Middle section of a control root. (B) Root epidermis and cortex. (C-D), PABA-treated roots. (C) Middle section of a root. (D) Root epidermis and cortex. Dashed circle line depicts the position of the quiescent centre (QC). Dashed lines mark the limits of the root zones: Magenta (MZ, zone 1), white (TZ, zone 2) and orange (zone 3, differentiation zone). The white arrow indicates the beginning of the differentiation zone. Bar, 20μm.
Figure 3.
Use of the PlaCCI reporter line to visualize the root cell cycle state. Confocal images displaying the root tips of 6 day-old PlaCCI plants expressing CDT1a-CFP, HTR13-mCherry, and CycB1;1-YFP constructs and grown in the absence (control) or presence of 200 μM PABA. (A-B), control root. (A) Middle section of a control root. (B) Root epidermis and cortex. (C-D), PABA-treated roots. (C) Middle section of a root. (D) Root epidermis and cortex. Dashed circle line depicts the position of the quiescent centre (QC). Dashed lines mark the limits of the root zones: Magenta (MZ, zone 1), white (TZ, zone 2) and orange (zone 3, differentiation zone). The white arrow indicates the beginning of the differentiation zone. Bar, 20μm.
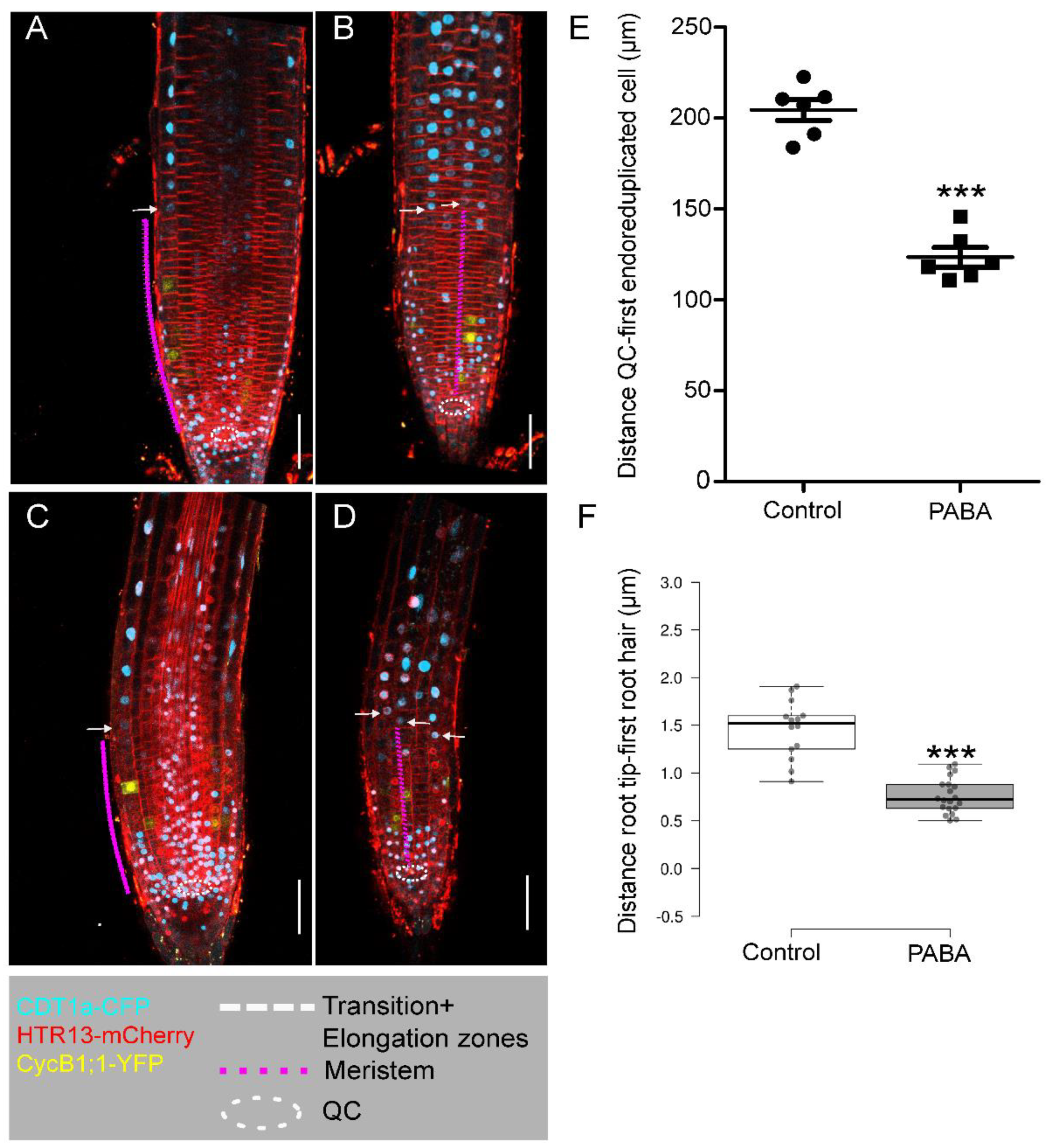
Figure 4.
Endogenous PABA regulates root growth. Analysis of root development of Arabidopsis wild type, 35S::ADCS-12 transgenic and ugt75b1 knock-out lines. (A-D), Phenotype analysis of WT (Col 0) versus 35S::ADCS-12. (A) Col 0 (WT, left panel) and 35::ADCS-12 plants grown in the absence of PABA. (B) Quantifications of the root length of plants shown in (A). (C) Size of the meristem. (D) Number of meristematic cells. (E-H), Phenotype analysis of WT (Ler) versus UGT75B1 loss-of-function mutant. (E) Ler (left panel) and ugt75b1. (B) Quantifications of the root length of plants shown in (A). (C) Size of the meristem. (D) Number of meristematic cells. Data are shown as means ± se (n>25 for B and F; n≥14 for C, D, G, H). The asterisks indicate statistical significance based on Student’s test: *** for P < 0.0001, * for P < 0.05, ns (not significant based on t-test). Bar, 1 cm.
Figure 4.
Endogenous PABA regulates root growth. Analysis of root development of Arabidopsis wild type, 35S::ADCS-12 transgenic and ugt75b1 knock-out lines. (A-D), Phenotype analysis of WT (Col 0) versus 35S::ADCS-12. (A) Col 0 (WT, left panel) and 35::ADCS-12 plants grown in the absence of PABA. (B) Quantifications of the root length of plants shown in (A). (C) Size of the meristem. (D) Number of meristematic cells. (E-H), Phenotype analysis of WT (Ler) versus UGT75B1 loss-of-function mutant. (E) Ler (left panel) and ugt75b1. (B) Quantifications of the root length of plants shown in (A). (C) Size of the meristem. (D) Number of meristematic cells. Data are shown as means ± se (n>25 for B and F; n≥14 for C, D, G, H). The asterisks indicate statistical significance based on Student’s test: *** for P < 0.0001, * for P < 0.05, ns (not significant based on t-test). Bar, 1 cm.
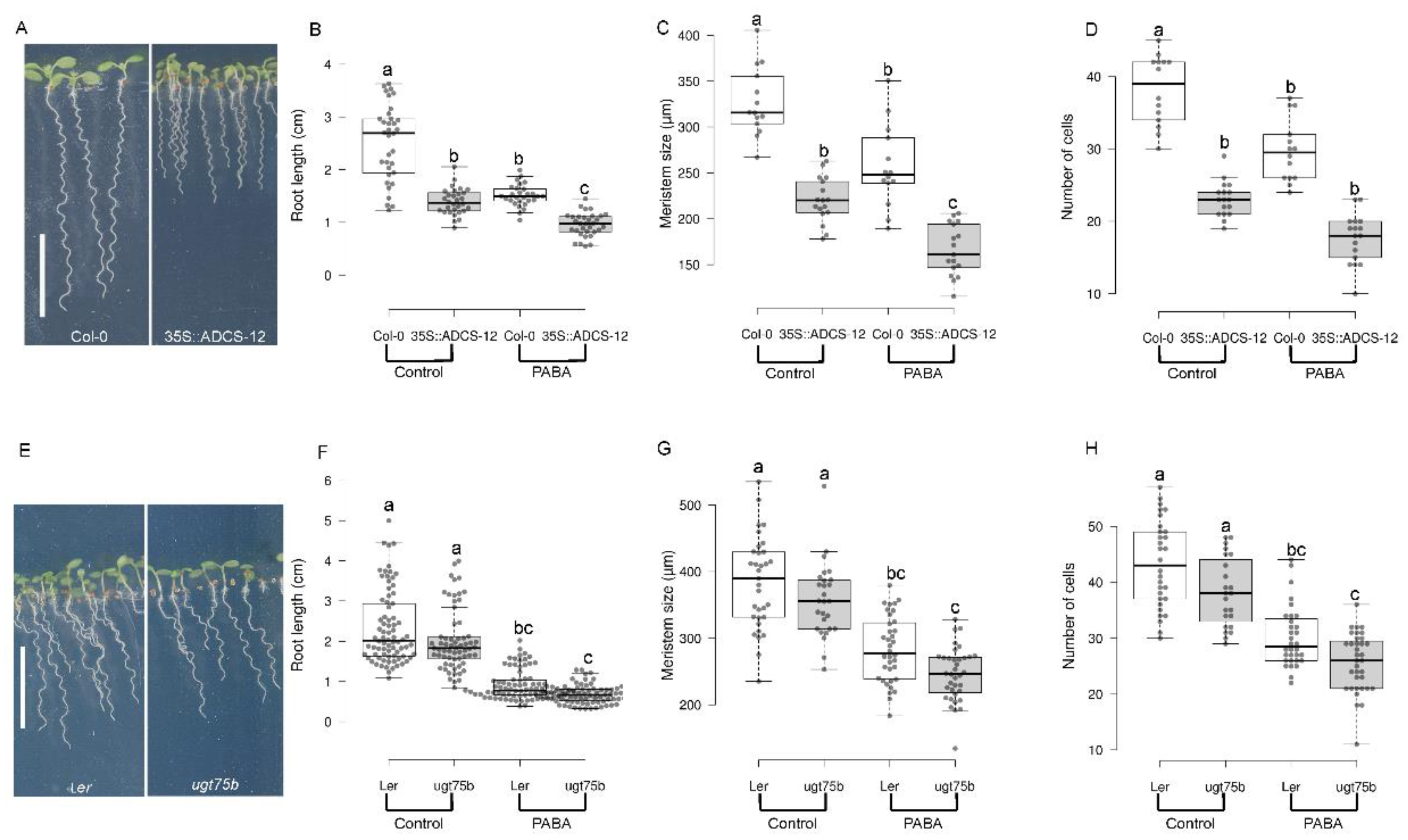
Figure 5.
Effect of reduced free endogenous PABA levels on root development. (A) The phenotype of Arabidopsis WT (Col-0) and Lex::UGT75B plants grown in the absence (0) or in the presence of ß-estradiol (0.1 and 0.5 μM). (B, C, D) Quantification of root length, meristem size, and number of meristematic cells, respectively. Data are shown as means ± se (n > 30 roots). Meristem size or cortex cell number is considered different from the control at P < 0.05. One-way ANOVA with Bonferroni multiple testing corrections was used to attest to the differences between treatment groups. The letters (a, b, c, d) indicate independent groups according to one-way ANOVA. Bar, 1 cm.
Figure 5.
Effect of reduced free endogenous PABA levels on root development. (A) The phenotype of Arabidopsis WT (Col-0) and Lex::UGT75B plants grown in the absence (0) or in the presence of ß-estradiol (0.1 and 0.5 μM). (B, C, D) Quantification of root length, meristem size, and number of meristematic cells, respectively. Data are shown as means ± se (n > 30 roots). Meristem size or cortex cell number is considered different from the control at P < 0.05. One-way ANOVA with Bonferroni multiple testing corrections was used to attest to the differences between treatment groups. The letters (a, b, c, d) indicate independent groups according to one-way ANOVA. Bar, 1 cm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



