
Submitted:
20 October 2023
Posted:
23 October 2023
You are already at the latest version
Abstract
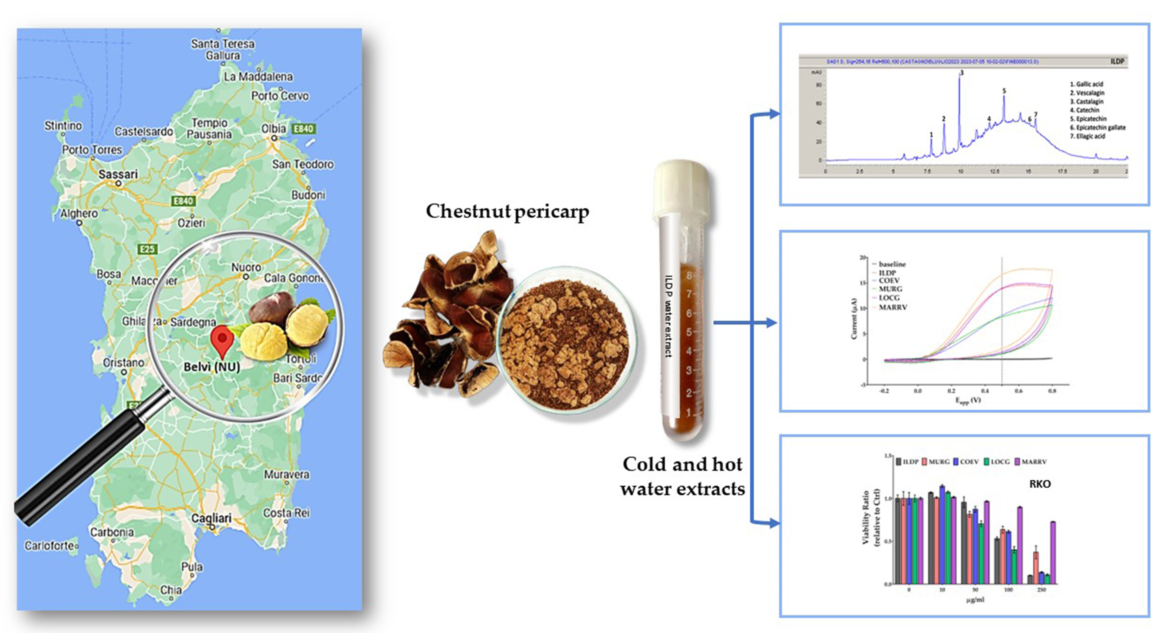
The residue of chestnut processing generates a large amount of waste material, a resource not adequately exploited. The phenolic composition of water extracts from discarded pericarp of four (MURG, LOCG, ILDP and COEV) chestnut accessions and one marron variety belonging to the Sardinian biodiversity was studied. The antioxidant capacity of cold and hot water extracts was determined by DPPH, ABTS and cyclic voltammetry tests. The antiproliferative effect on normal cells (fibroblasts), and colon (RKO and SW48), breast (MCF7) and melanoma (B16F10) cancer cells, was evaluated by biological assays. MTT test demonstrated that temperature and different extraction times significantly influenced the growth of cells, both normal and tumor. The fibroblast viability was significantly reduced by moderate doses of cold extracts, but only by doses greater than 250 µg/ml of hot extracts, regardless of the accession or cultivar. An even more marked effect was observed when RKO and SW48 were treated with cold extracts, while treatments on B16F10 melanoma cells resulted less effective. Differently, the cold extracts of all accessions induced a significant increase in MCF7 cell viability starting from moderate doses (50 µg/ml), whereas hot extracts had a similar, but less evident, effect compared to reference fibroblasts.
Keywords:
Chestnut biodiversity
; pericarp
; water extraction
; antioxidant activity
; cancer cells
1. Introduction
The sweet chestnut tree (Castanea sativa Mill.), which belongs to the Fagaceae family, is widely distributed in European countries, Italy, Spain, France, Greece, Portugal and Turkey for wood, fruit, honey as tannins source [1,2]. Over the last decade, the chestnut sector has constantly grown in Europe, since fruits, mainly consumed boiled or roasted, are an excellent source of energy, due to their high starch content, minerals (K, Mg, Mn, and Cu), unsaturated fatty acids (omega-3), fiber and vitamins E and C [3,4]. A variety of preparations have been added to the traditional use of the fruit: marron glacé, purées, frozen nuts, and jams, and, more recently, different gluten-free products based on chestnut flour for celiac patients [5,6].
Burs, leaves and the residue of chestnut processing generates a large amount of waste material, about the 15% of the total production. Inner and outer shell resulting from chestnut peeling, represent about 20% of the total fruit weight [5]. This residue, habitually burned as fuel in factories, is the target of new strategies of value assessment and reuse: heavy metal absorbents or wood adhesives, raw materials for lignin and bioethanol production and pigment source for dyeing [7,8]. At present, there are new attractive applications for utilizing chestnut waste as a source of bioactive compounds with healthy effects [9]; the chestnut pericarp (outer shell; husk) and integument (inner shell; pellicle) are a rich source of nutraceuticals, ascorbic acid, carotenoids, phenolics like gallic and ellagic acid, flavonoids and condensed tannins, with antioxidant, antimicrobial, and anticancer properties [4,10,11,12]. The green recovery of these bioactive compounds from chestnut shell becomes decisive, both to preserve their healthy properties for food, cosmetics and pharmaceutical industries and, not less important, to reduce the environmental impact of organic solvents [6]. Hence the decision to use, in this work, only deionized water as an extractant of phenolic compounds. Cold and hot water, alone or in combinations with organic solvents, has been employed by our group, with different purposes and on different matrices, since 2010 [13,14,15,16,17,18], but this work is the first on chestnut. Other researchers used cold water with Na2SO3 and NaOH to extract phenolic compounds, tannins and ellagitannins from Portuguese chestnut peels [19]; bur and shell extracts of Castanea sativa were obtained using water and hydroalcoholic solutions at 25, 50 and 75 °C to make the extraction faster, selective, and more effective [20]; Jung et al. [21] used water and a variety of organic solvents, at 60 °C, to extract polyphenols from inner shells of Japanese chestnut (Castanea crenata); boiling water aimed at maximizing the recovery of bioactive compounds from inner and outer shells of Italian chestnut [22,23]. The use of water alone as a solvent is expected to provide extracts with lower yield of phenolic compounds than organic solvents or hydroalcoholic mixtures [20]. The chemical composition of these extracts, their antioxidant and antitumoral properties, that are the subject of this research, are also expected to change [20,21,24]. In this work, the antioxidant capacity of chestnut extracts has been quantified with analytical systems such as DPPH and ABTS, spectrophotometric tests capable of indirectly measuring, with a single assay, the sum of the antioxidant activities of a pool of molecules [25]. A direct electrochemical determination of antioxidants was also provided according to recent studies on outer shell extracts [26] and other plant and fruit extracts [27,28]; the redox properties of single molecules, such as gallic, ellagic and tannic acid, were also investigated [29,30]. Since the phenolic compounds can be easily oxidized on carbon electrodes [31,32,33], and their ionization potential determines their efficiency as antioxidants [34,35], the cyclic voltammetry (CV) was used to show the potential at which the oxidation starts and to provide a measure of antioxidant capacity [17].
Various researches, over the last 20 years, have provided a characterization of the phenolic compounds present in chestnut shell extracts, attributing them antioxidant and anti-tumor properties [36,37,38]. Although there's a general concern in indicating the polyphenolic fraction of various chestnut organs responsible for anti-tumor action [12,21,39], even in this case it is yet to be established which molecules are actually responsible and which are not. Moreover, there is some evidence that the same molecules that perform antioxidant activity at certain concentrations act as pro-oxidants at others [17]. Chestnut extracts, derived from flesh and inner- and outer-shell, induced apoptosis in human gastric cancer cells [36]; apoptosis was also induced by chestnut honey on prostate cancer cells [40], by bark extracts on neuroblastoma cells [41], and by shell extracts on prostate, breast and hepatocellular cancer cells [12]. In this work, we tried to understand how the extraction method could affect the extraction yield of some compounds with presumed anti-tumor activity in different chestnut accessions of Sardinian biodiversity, and then we focused on the ability of the different extracts to inhibit the growth of cells belonging to different tumor lines.
2. Results
The ability to recover bioactive molecules of two extraction techniques, one with cold water (20°C), the other with hot water (95°C), from the pericarp of different accessions and varieties of chestnut was investigated. It has been demonstrated that, at the same extraction time, hot water is a more effective extractant [20]. So, in this work, the contact time between matrix and solvent was diversified: 4 hours with cold water and 1 hour with hot water. The results of this experimental plan were reported in table 1 and table 2, where it has been shown that the two extraction techniques sometimes lead to the same result, other times to significant and even important quantitative and qualitative differences.
2.1. Phytochemical content of cold and hot water pericarp extracts
The phytochemical content of water extracts at 20 and 95 °C, of pericarp of four ancient Sardinian chestnut accessions and of one commercial marron cultivar was determined, together with their antioxidant capacity (Table 1). The content of total phenolics (TP), the total flavonoids (TF) and the condensed tannins (CT), varied significantly among chestnut accessions and the marron: this was expected since they were cultivated in different pedoclimatic conditions, at different altitude and harvested at different times, as specified in the Material and Methods section. So, we mainly focused the attention on the differences between treatments (20 °C vs 95 °C) within the same accession or variety.
MURG, ILDP and LOCG accessions were characterized by a high level of TP, TF, and CT, both when they were extracted at 20 °C and 95 °C. COEV had the lowest TP, FT and CT content of all the accessions. The commercial MARRV cultivar was characterized by a low content of phenolic compounds, at least compared to the four accessions. Unlike the latter, however, its total content of phenolic compounds and the content of tannins are strongly influenced by the extraction temperature, with TP and CT values much higher at 95 °C.
2.2. Antioxidant capacity determination by DPPH and ABTS tests
The results of the DPPH and ABTS tests on antioxidant capacity reflect the phenol content detected in the different accessions and in the marron. The treatments did not determine statistically significant differences in MURG, LOCG and ILDP accessions, while the antioxidant activity of the extracts at 95 °C was higher than at 20 °C, both for COEV and MARRV. The results of the electrochemical tests, however, did not follow the same trend as the other two assays: this was expected since the radicals (DPPH* and ABTS*+) react with almost all the compounds present in the extracts able of giving up electrons. The AUC values, which refer only to compounds that release electrons at an applied potential (Eapp) of +0.5 V, were higher at 20°C in MURG, at 95°C in LOCG and MARRV, and not statistically different in ILDP and COEV. The difference among results obtained with different methods will be discussed, with appropriate bibliographical references, in the "Discussion" chapter.
2.4. Electrochemical characterization of chestnut pericarp extracts
The cyclic voltammetric patterns recorded for chestnut pericarp samples were displayed in Figure 1.
The voltammograms show that the oxidation in the extracts started from + 0.06 to + 0.2 V, both at 20 °C and 95 °C. The shapes of the voltammograms were similar but the currents, recorded at +0.5 V, by different accessions, varied from 8 to 14.5 µA at 20 °C, and from 7.5 to 17 µA at 95 °C. These differences should be ascribed to the activity of compounds with different redox potential (see also AUC values in table 1). If we consider cold extraction first, and we refer the voltammetries to a gallic acid calibration curve (Figure S1a and S1b, in Supplementary Materials), COEV has the lowest quantity of compounds that oxidize at +0.5 V. LOCG and MURG have similar voltammograms but while MURG has a higher concentration of compounds that oxidize at a potential lower than +0.5 V, the molecules that oxidize at an Eapp greater than this threshold appear more represented in LOCG. ILDP has the highest redox peak of all the other cold extracts, and also has the highest polyphenol component that oxidizes at less than +0.5 V. MARRV has a voltammogram comparable to MURG. Differently, when we consider the hot extracts, the COEV and MURG voltammograms are superimposable. They indicate that the two samples have the lowest concentration of compounds, no matter whether they oxidize at a potential lower or higher than + 0.5 V. The voltammograms of ILDP and LOCG are also very similar to each other, but the extracts have a concentration of antioxidant compounds higher than the other two Sardinian accessions. Finally, the marron MARRV has a higher antioxidant capacity than all four accessions.
2.4. Chemical characterization of chestnut pericarp extracts
The concentration of the main phenolic compounds in the water extracts of chestnut pericarp was determined by High Performance Liquid Chromatography (HPLC). The research was focused on those phenolic compounds, present in the pericarp of chestnut, which were reported to have cytoprotective effects towards normal cells and/or antiproliferative effects on cancer cells. Molecules like gallic acid and ellagic acid, two phenolics of low molecular weight, the ellagitannin isomers castalagin and vescalagin, and flavanols represented by catechin, epicatechin and epigallocatechin [4,39,42] were identified and quantified, and the results were reported in Table 2; the chromatogram of ILDP can be found, as an example, in Figure S2 in Supplementary materials.
Gallic acid, ellagic acid, vescalagin, castalagin, catechin, epicatechin and epigallocatechin were the seven main phenolic compounds identified in the four accessions and in the marron. Water at 20 °C was more efficient than hot water to extract gallic and ellagic acid from the epicarp of all the accessions, but less efficient in the marron. The ellagic acid was the most represented compound in all the samples and its content was 5-fold higher when the extraction was done with cold water. Ellagic acid aside, each accession is distinguished by the prevalence of certain compounds:
- -
- vescalagin, castalagin and epicatechin characterize the MURG extracts, regardless of the temperature used for extraction;
- -
- LOCG cold extracts are characterized by gallic acid, vescalagin and castalagin, while the hot ones have a high content only of vescalagin, castalagin and, particularly, of epicatechin;
- -
- ILDP cold extracts can be distinguished by their content of gallic acid, catechin and epicatechin, whereas the hot extracts by vescalagin, castalagin and epicatechin;
- -
- COEV is distinguished by a low content of all compounds (thus confirming the results of the spectrophotometric assay) if the extraction takes place in cold water, and by a prevalence of castalagin and vescalagin when the extractant is hot water.
The marron MARRV showed a different phenolic composition from the Sardinian accessions: in cold water extracts, low quantities of all seven molecules were detected, except for ellagic acid, whose value is however the lowest among the samples tested. Differently, the hot extracts are characterized by moderate quantities of vescalagin and epicatechin and by a high quantity of catechin, higher than that found in the four accessions.
2.5. Contribution of main polyphenols to the total antioxidant activity of chestnut pericarp extracts
The electrochemical and redox properties of the identified compounds were investigated based on a comparison with previous studies of our group [43,44] and specific literature on chestnut [26]. The redox potential of all the molecules listed in Table 2 was reported in Table 3.
The oxidation potential of a molecule may have small variations as a function of several variables: the working electrode [43,45], the reference electrode (or pseudo-reference) [46], the supporting electrolyte [47] and various other differences in experimental protocols; furthermore, sometimes the main oxidation peak is taken into consideration [26], sometimes the point at which the molecule's voltammogram separates from the baseline is considered (i.e. the moment in which a molecule begins to give up its electrons). Due to these differences, different authors may record different values. The oxidation potential of the seven compounds in table 3 are all in the range within + 0.1 and + 0.4 V, which is the range of interest for the most active molecules according to the parameters reported for antioxidants phenolic compounds [31,32,33].
2.6. Antiproliferative activity of chestnut pericarp extracts on normal and cancer cells
The antiproliferative effects of the five chestnut pericarp extracts on the four cancer cell lines were reported in the figures from 2 to 6, where they were compared to the effect on normal cells.
The MURG cold water extract (Figure 2A) significantly reduced the viability of fibroblasts starting from 100 µg/ml up to a -46% at 250 µg/ml. Treatments became effective reducing the viability of SW48 at 100 µg/ml and, at the highest concentration, on RKO and B216F10; differently they increased the viability of MCF7 starting from 50 µg/ml. The hot water extract (Figure 2B) appeared less effective both on fibroblast and cancer cells but increased the viability of B16F10 at 100 and 250 µg/ml.
The LOCG cold water extract (Figure 3A) reduced the viability of fibroblasts starting from 50 µg/ml. The viability of SW48 and RKO was progressively reduced from 50 to 250 µg/ml, that of B16F10 from 100 to 250 µg/ml. The viability of MCF7 was significantly higher than fibroblasts at 50, 100 and 250 µg/ml. The hot water extracts were less effective than the cold ones (Figure 3B) on fibroblasts, since a 26% significant reduction of the viability was observed only at the highest concentration. A significant effect on cancer cells was observed on RKO (-21% vs fibroblasts) and on MCF7 (+36% vs fibroblasts).
Treatments with cold and hot ILDP extracts (Figure 4) did not appear to have particular efficacy on fibroblasts, except for the cold ones at 250 µg/ml (-41% of viability). Differently, the cold-water extracts (Figure 4A) seriously reduced the viability of SW48 and RKO starting from 100 µg/ml, while significantly increased that of MCF7 starting from 10 µg/ml. The effect of hot water extracts was limited to SW48 cells (Figure 4B).
Treatments with cold and hot water COEV extracts affected the viability of fibroblasts at 100 and 250 µg/ml, respectively (Figure 5). The same concentrations of cold-water extract significantly reduced the viability of colon cancer cells (Figure 5A) and slightly increased that of MCF7; the hot water extract was effective also on B16F10 (Figure 5B).
The marron MARRV extract had different effects, both on fibroblasts and cancer cells, compared to the four Sardinian chestnut accessions (Figure 6). The viability of fibroblasts was significantly and progressively reduced, starting from 10 µg/ml, by treatments with cold water extracts; no significant reduction, except for melanoma cells at 250 mg/ml, was observed on cancer cells compared to fibroblasts (Figure 6A). Hot extract treatments reduced the viability of normal cells only at 250 µg/ml. Instead, unlike all Sardinian accessions, hot extracts of MARRV are the only ones to have shown significant antiproliferative activity on breast cancer MCF7 cells (Figure 6B).
3. Discussion
This work aimed to investigate some healthy properties of polyphenolic waste extracts obtained with sustainable and eco-friendly methods for industrial applications. The research focused on water extracts of outer shell of four chestnut accessions and one marron variety, with emphasis on their phenolic composition, antioxidant activity and antiproliferative effect on normal cells (fibroblasts), and on two colon (RKO and SW48), one breast (MCF7) and one melanoma (B16F10) cancer cell lines.
The extraction of phenolic compounds from plant materials has been carried out with organic solvents (methanol, ethanol, acetone, etc.) for years, and it was the same for the chestnut [48,49]. Vasquez et al. [50] compared the yields resulting from different extraction methods, with organic solvents (96% n-hexane, acetone, ethyl acetate, ethanol and methanol), with water or with a 2.5% Na2SO3 aqueous solution heated to 90 °C for one hour: the yield of extractable compounds increased with the polarity of the solvent and the highest yield corresponded to water. The same authors, studying the influence of extraction conditions (water and different alkaline solutions at 70 and 90 °C) observed an increase of total phenols and antioxidant activity when the temperature increased. The use of water alone or combined in hydroalcoholic solutions is more recent [19,20,22,23], and there are green technologies, like subcritical water extraction, used to maximize the extraction yield and polyphenol content [51], which offer a more economically sustainable and environmentally friendly alternative for large-scale industrial applications. In this work, we took into account previous results which demonstrated that the use of hot water, for the same extraction time, increased the extraction capacity of the solvent [20]. The use of hot water has two important critical issues: first, heating the water has a cost for industries; second, the thermolabile phenolic component is affected [51]. We have shown that this is not always true, at least in terms of yield of total polyphenols, flavonoids and tannins: the extracts from the four Sardinian accessions contained statistically non-different quantities of TP, TF and CT after 4 hours of extraction in cold water or one hour of extraction in hot water at 95 °C. For marron the results were different, and hot water proved to be able to extract a greater quantity of total polyphenols and condensed tannins than cold water, but not of flavonoids.
The DPPH and ABTS tests for antioxidant capacity determination seemed to mirror the analytical tests for TP, TF and CT. No significant differences were determined by the extraction at different temperatures for MURG, LOCG and ILDP accessions, while a higher antioxidant activity was measured for COEV and MARRV extracts at 95 °C. The electrochemical tests, however, showed a different trend: the antioxidant capacity of MURG extracts was higher at 20 °C, the one of LOCG and MARRV was higher at 95 °C, and ILDP and COEV did not statistically differ. A comparison between DPPH assay and CV cyclic voltammetry can be found in a previous study [26]: what the two works have in common, and which gives us the possibility of having a valid term of comparison, is the threshold of +0.5 V established a priori in order to discriminate the real antioxidant capacity of the extracts. Ricci's group attributes two important limitations to the DPPH assay, long incubation times and difficulties with colored samples: we completely agree and, furthermore, we also believe that the DPPH and ABTS assays tend to overestimate the antioxidant capacity because they take into account species in solution, which oxidize at a potential greater than +0.5 V [34]. Fadda and Sanna [52], in a dissertation on advantages and pitfalls of methods for antioxidant activity evaluation, affirmed that the capacity of antioxidants to reduce the radical cation ABTS•+ to ABTS is directly related to the reduction potential of the antioxidants, and only those with E0 values lower than that of the couple ABTS•+/ABTS (E0 = 0.68 V) are able to perform this reduction. Obtaining a good correlation between spectrophotometric and electrochemical methods is possible if similar trends are observed, but this does not mean that comparable antioxidant capacity values are obtained. The antioxidant capacity of the outer shell extracts is linked to the concentration of phenolic compounds able of oxidizing at Eapp lower than +0.5 V but, in the absence of a mass spectrometric analysis, we do not have the complete profile of all molecules in the investigated accessions, nor do we know their specific reducing power. According to our HPLC analysis, we can state that all seven compounds identified and quantified have a redox potential lower than +0.4 V and that all of them contributed to the antioxidant capacity according to their concentration in the extracts. Even though the obtained qualitative phenolic profile was similar to that reported by other authors, differences in quantitative content was found [4,39,42]: TP values similar to our MARVV and lower than those of Sardinian accessions, but with shorter extraction times were recorded by De Vasconcelos et al. [4], while CT values were comparable to our results. A content of TP approximately ten-fold higher than ours was found in chestnut waste consisting of inner and outer shell water extract [24]. This high variability assessed in the literature could be attributed to varietal differences, although a wide number of factors influence the phytochemical levels in vegetal matrices, such as geographical origin, pedoclimatic conditions, irrigation, seasonality etc. Beyond all this, the role of temperature must be considered. The HPLC analysis showed that the temperature distinguishes between the different molecules: the ellagic acid showed the highest concentration in the cold extracts, ellagitannins in the hot ones and, for the other molecules, specific cultivar-dependent trends were recorded. Vella et al. [23] found a TP and FC content much lower than ours in water extract at boiling temperature (for 40 min) of outer shell chestnut Campania cultivars. An increase in the castalagin and gallic acid concentration, consequence of an increase in the extraction temperature, was reported by De Vasconcelos et al. [4] as for our cultivar MARRV. The thermal stability at 60, 80 and 100 °C of water solutions of gallic acid, catechin, and vanillic acid was studied: it was demonstrated that the degradation of the phenolic compounds in grape seeds and spruce bark vegetal extracts was lower, for all the temperatures, than individual standard solutions, confirming that the complex chemical composition of vegetal extracts has to take into account to evaluated thermal stability of phenolic compounds [53]. It was reported that high temperature and long extraction times can promote the phenols oxidations and reduce antioxidant properties [7], that temperatures up to 90 °C increase the extraction of phenolic compounds [4,23,54], and that this increase is not always associated with the maximum antioxidant capacity [20] due to additive and synergic effects of individual phytochemicals.
All the considerations made so far are based on in vitro studies, chemical and electrochemical tests, but to better understand the role of phenolic compounds their biological activity on normal and cancer cells has been investigated. It is known that polyphenols decrease DNA damage induced by various carcinogens acting as ROS scavengers, chelating transition metals, or modulating the expression and the activity of the enzymes related to oxidative stress [55]. The antioxidant activity of polyphenols could be essential but not sufficient for chemoprevention [56] and there is evidence that naturally occurring antioxidants elicit different redox responses according to a dose-response mechanism and the intracellular redox state [57,58,59].
As many other species, the chestnut phenolic compounds have been correlated with anticancer activity. Unfortunately, studies on the anticancer properties of chestnut are still limited to a few articles: one tumor (DU 145) and one normal prostate epithelial (PNT2) cell line were exposed to increasing concentration of shell polyphenols extracts, thus showing an increase of apoptotic cells after 48 and 72 h treatments [12]; anticancer activity vs. prostate cancer cells (DU 145 and PC3) was attributed to a quinoline-pyrrolidine alkaloid present in chestnut honey, and the inhibition of clonogenic activity was congruent with apoptotic events [40]. Apoptotic-induced effects were also observed on neuroblastoma cells (SH-SY5Y) when exposed to increasing concentration extract obtained by bark of Castanea sativa containing high amounts of vescalagin, castalagin gallic and ellagic acid [41].
In our work, normal cells and four cancer cell lines, SW48, RKO, MCF7 and B16F10, were exposed to cold and hot water pericarp extracts of four Sardinian accessions and of one marron cultivar. The temperature and different extraction times significantly influenced the growth of cells, both normal and tumor. The fibroblast viability was significantly reduced, at least by 20%, by treatments with cold extracts starting from 100 µg/ml of MURG and COEV, 50 µg/ml of LOCG and ILDP, and even 10 µg/ml of MARRV. According to Floris et al. [60], a 20% reduction in viability is the threshold to consider a treatment to be effective. Such a reduction, with hot extracts, was observed only with doses greater than 250 µg/ml, regardless of the accession or cultivar to which they belong. This clearly indicates that the pool of molecules extracted at 95 °C exerts a cytotoxic effect significantly lower than that of cold extracts. Even on cancer cells, the most evident cytotoxic effects were observed when the cells were treated with the cold extracts.
The viability of metastatic colon cancer cells, SW48 and RKO, was strongly reduced by 100 µg/ml (if not even 50 µg/ml of LOCG) of all the Sardinian accessions extracts, while MARRV had no effect; also in this case, the cytotoxic action of the hot extracts was decidedly lower, and only at the highest doses. The activity against colon cancer cells of chestnut extracts was also investigated by other research groups. The capacity of inner shell extracts of Japanese chestnut (Castanea crenata Siebold & Zucc., 1846), to inhibit the viability of LoVo and HT29, two human colon adenocarcinoma cell lines, was explored [21]: the water extracts (24 h at 60 °C), containing 2.78 mg g-1 of gallic acid (higher than ours) and 1.07 mg g-1 of ellagic acid (lower than ours), induced a viability reduction, varying between 20 and 60%, starting from 125 µg/ml, a result comparable to those obtained by our group on RKO and SW48. On the other hand, the bioactive compounds of shells of Chinese chestnut (Castanea mollissima Blume) seemed to have a moderate anti-cancer activity on human COLO 320 DM colon cancer cells, but results cannot be compared with ours since a 90:10 (v/v) ethanol-water solution was used as extractant [61].
Treatments on B16F10 melanoma cells resulted less effective: a decrease in viability by cold MURG extracts was induced only by the maximum dose, while the same dose of hot extract increased cell growth. The cold extracts of LOCG, ILDP and COEV also had mild effects, while the hot extracts appeared almost ineffective. The same goes for MARRV, with reduction in cell viability only at the highest dose of hot and cold extract. Unfortunately, we were not able to compare our data with the literature since we did not find any other pertinent study concerning melanoma cancer cells and chestnut derivatives.
A separate discussion must be made for breast cancer MCF7 cells: cold extracts of MURG, LOCG, ILDP and COEV induced a significant increase in cell viability starting from moderate doses (50 µg/ml); hot extracts had a similar, but less evident effect compared to reference fibroblasts. What was surprising, compared with the Sardinian accessions, was the effect induced by the hot extracts of MARRV which induced a reduction in cell viability of approximately 50% at the highest treatment dose. Cacciola et al. [12], worked with chestnut shell water extract, rich in gallic acid but with a much lower ellagic acid content than that found in our extracts; they observed that treatments with 100 µg/ml of extract induced a significant reduction in the viability of MCF7 cells, but less than 20%, and no effect on MDA-MB-231, another breast cancer cell line. This is in agreement with our results, since MARRV hot extracts were not effective at 100 µg/ml, and became effective only at 250. A similar result, 18% inhibition of cell viability, on the MDA-MB-231 line was obtained by treating the cells for 24 h with doses of 200 µg/ml of chestnut powder [36]. Finally, a study is worth reporting that suggests that chestnut leaf extracts of Castanea crenata could increase the susceptibility of MCF7-derived cancer stem cells to paclitaxel, an anticancer drug [62].
4. Materials and Methods
This study was carried out in "Barbagia" (Nu), an area historically devoted to chestnut cultivation, in the vast mountainous region of central Sardinia which extends along the sides of the "Gennargentu" massif. Samples were collected from four chestnut (Castanea sativa Miller) accessions of local biodiversity and one Marron variety (Table 3). MURG, LOCG, ILDP, COEV and MARRV were all harvested in situ in the municipality of Belvì (39°57′38.19″N 9°11′02.91″E) [63].

Table 3.
Origin, elevation and harvest time of the chestnut accession and of the marron.
| Accession/variety | Origin | Elevation (m a.s.l) |
Harvest time | Acronym |
|---|---|---|---|---|
| MIGHELI URRU G | Belvì | 822 | October, second decade |
MURG |
| LOCCHEDDU G | Belvì | 812 | October, second decade |
LOCG |
| ILDUBBA P | Belvì | 849 | October, second decade |
ILDP |
| COESERRA V | Belvì | 686 | October, third decade |
COEV |
| MARRONE di Marradi V | Belvì | 780 | October, second decade |
MARRV |
4.1. Plant Material and Fruit Sampling
Chestnuts were collected at commercial maturity in October 2021. Fruits, free of defects and mechanical damage, were divided in three replicates of 30 chestnuts for each accession. The chestnut pericarp (CP) was manually separated and oven-dried at 40 °C until reaching constant weight, to reduce and standardize the moisture content. The dried samples were ground in a mill to a fine powder (particle sizes 1mm), and stored under vacuum in total darkness, until further analyses.
4.2. Extraction of phenolic compounds
Two different extractions, using water as a solvent, were carried out for the recovery of the bioactive molecules, according to [12] with some modification:
i) a conventional liquid extraction (CLE) in cold water at 20 °C for 4h. 2.5 grams of CP powder were put in contact with 25 ml of milli-Q water in a thermostatic bath. The liquid and solid phases were separated by centrifugation at 3220× g for 15 minutes. The supernatant was recovered by filtration through a filter paper (Whatman n. 1) under vacuum, while the solid residue was added of another 25 ml. The extraction procedure was repeated, the resulting supernatant was added to the first (solvent/sample final ratio 20:1 (v/w)) and all the liquid was stored at -20 °C until analysis.
ii) a CLE in hot water at 95 °C for 1h. 2.5 g of dried sample were extracted with 50 ml milli-Q water (solvent/sample ratio 20:1 (v/w)), for 1h, under continuous stirring in a thermostatic bath; then the suspension was cooled on ice and centrifuged (15 min at 3220× g). The supernatant was recovered by filtration through a filter paper (Whatman n. 1) under vacuum and stored at -20 °C until analysis.
4.3. Analytical tests
Water extracts were analyzed for, total phenolics (TP), total flavonoids (TF), condensed tannins (CT), individual phenolic compounds and antioxidant capacity.
4.3.1. Determination of total phenolic content
The total phenolic content (TPC) was assessed by Folin-Ciocalteu assay according to previous procedure [18]. Aliquots of the diluted samples were mixed in a 25 mL volumetric flask with Folin–Ciocalteu reagent (1:1) and a 10 ml sodium carbonate solution 7.5%, and incubated for 2 h at room temperature. TPC was determined by spectrophotometric analysis (8453 UVVisible Spectrophotometer, Agilent Technologies, USA) at an absorbance of 750 nm; it was expressed as milligrams of gallic acid equivalents (GAE) per g of dried matter (DW) referring to a gallic acid calibration curve (10-100 mg l-1, R2= 0.990). Samples were analyzed in triplicate.
4.3.2. Determination of total flavonoid content
The total flavonoid content (TFC) was quantified by colorimetric assay according to the AlCl3 assay and previously reported procedures [64]. Quantification was carried using a catechin (C) calibration curve (2.5 – 20 µg ml-1, r2 = 0.995). Results were expressed as mg of catechin equivalent (CE) x g-1 of dried matter (mg CE g-1 DW).
4.3.3. Determination of condensed tannin content
The vanillin assay was used to quantify the extractable condensed tannin content of the samples, according to [18]. The absorbance of vanillin-tannin adducts was spectrophotometrically detected at 500 nm, and concentrations were calculated referring to a catechin calibration curve (1 – 6 μg ml-1, R2 = 0.998). Results were expressed as mg CE g-1 DW.
4.3.4. Determination of antioxidant capacity by DPPH and ABTS assays
The antioxidant capacity of CP extracts was evaluated by DPPH (1,1-diphenyl-2-picrylhydrazyl) and ABTS (2,2-azinobis (3-ethylbenzothiazoline-6-sulphonic acid) diammonium salt) assays, according to [65]. Briefly, for each assay, 0.1 mL of appropriately diluted PPE was mixed with 3.9 mL of 60 μM DPPH or 7 mM ABTS, and then stored in the dark for 120 or 6 min, respectively. The Trolox equivalent antioxidant capacity (TEAC), was expressed as mmol Trolox equivalents·100 x g−1 of dry matter (mmol TEAC·100 g−1 DM).
4.3.5. Determination of antioxidant capacity by electrochemical method
The antioxidant capacity (AAox) determination of the chestnut pericarp extracts was also performed by cyclic voltammetry (CV) as previously reported [17,44] with some modifications. Measures were carried out by screen-printed sensors purchased by GSI Technologies (Burr Ridge, IL, USA), consisting of a 4 mm carbon working electrode (WE), an Ag/AgCl pseudo reference electrode (RE), and a carbon auxiliary electrode (AE). Currents were recorded by Quadstat, a commercial four-channel potentiostat (eDaQ Quadstat, e-Corder 410 and Echem software, eDAQ Europe Poland, Warsaw Poland). Cyclic voltammograms (CVs) were performed from −0.2 V to +0.8 V (vs. Ag/AgCl pseudo-RE) at a scan rate of 0.1 V/s. A first aliquot of 70 μL, containing only PBS (used as a supporting electrolyte), was deposited on the screen-printed WE with a graduated micropipette in order to obtain a baseline. Once the baseline current was recorded, the PBS drop was dried with absorbent paper without touching the surface of the sensor, and 70 μL aliquot of 2 mg/ml chestnut outer shell extract solution was deposited on the sensor surface, thus obtaining the corresponding CV pattern (the experiment was performed in triplicate). In order to provide a quantitative comparison among the CV patterns of extracts of different origin, the voltammograms were integrated and the area under curve (AUC) was calculated at +0.5 V and expressed in microcoulombs (μC). The redox potential of +0.5 V is used as a threshold to detect the antioxidant activity of chestnut extracts in accordance with previous studies [17]. As already reported [43,45,66], oxidation potentials higher than +0.5 V refer to polyphenols with low reducing power which, in this work, were not accounted as antioxidants.
4.4. HPLC analysis of phenolic compounds
Chromatographic separation of phenolic compounds was carried out with reverse-phase HPLC method using an Agilent 1100 Liquid Chromatography (LC) system (Agilent Technologies, Palo Alto, CA, USA) equipped with a quaternary pump (G1311A), degasser, column thermostat, auto-sampler (G1313A), and a diode array detector (G1315 B, DAD). The column was a Luna C18 column (250 x 4.6 mm, 5 µm) from Phenomenex (Torrance, CA, USA) with a security guard cartridge (4 × 2 mm). The flow rate was set at 0.8 ml min-1, and the column temperature was set to 30 °C. Elution was carried out with a binary mobile phase of solvent A (water and 0.1% trifluoracetic acid) and solvent B (acetonitrile) and the gradient elution program was according to [18]: 0 min, 99% A; 5 min, 95% A; 6 min, 93% A; 10 min, 85% A; 15 min, 75% A; 20 min, 10% A; 25 min, 99% A, with a post-time of 3 min. Detection was performed at 254, 280, 350, and 360 nm. Phenolic molecules were identified according to the retention time of a mixture of standards, selected from the literature concerning CP phenolic, their UV absorption spectra, as well as by adding standard solutions to the sample composition and quantified using the respective calibration curves. Samples were appropriately diluted before injection. The results are presented as milligrams per gram of dry weight (mg g-1 DW).
4.5. Cell culture and biological assays
Five different cell lines, obtained from ATCC, had been selected to be tested with cold and hot water chestnut extracts: 1) Human fibroblast (normal cells); 2) B16F10 (murine melanoma cells; N-ras, Braf and p53 wt); 3) MCF7 (human metastatic breast cancer cells, estrogen receptor (ER) +, progesterone receptor (PR)+, epithelial receptor 2 (HER2) -, BRCA1 wt); 4) RKO (human metastatic colon cancer cells; P53, K-ras and PTEN wt, Braf p.V600E, PIK3CA p.H1047R); 5) SW48 (human metastatic colon cancer cells; P53, K-ras and PTEN wt, Braf p.R347Xc, PIK3CA p.G914Rc).
Human fibroblasts, B16F10 and MCF7 were grown in DMEM, RKO cells in EMEM and SW48 in RPMI1640 at 37 °C under humidified 5% CO2/air. All media were completed with 10% FBS, 1% penicillin (100 U mL -1)/streptomycin (100 µg mL-1), and 1% L-glutamine. For experimental studies, cells were plated in 96 well plates at a density of about 2 x 104 mL-1 and, 24 h post seeding, exposed to increasing concentrations of hot and cold extract (10–50-100-250 µg mL-1) for 24 h.
4.5.1. MTT assay
The MTT assay was used to assess the cell viability as previously reported [17]: cells were incubated with 100 µL (0.05 mg mL-1) of MTT, and the cultures were allowed to incubate at 37 °C for 3 h. The MTT was removed and the formazan crystals were dissolved in 100 µL of isopropanol. The color was read at 570 nm using a microplate reader (EMax® Plus, Molecular Devices). The percentage of cell growth and metabolic activity was calculated by normalizing the absorbance of the treated cells to corresponding control. All the experiments were done in quadruplicate and repeated at least three times.
4.6. Data Analysis
The statistical analysis was performed by GraphPad Prism 5 for Windows software (Graph-Pad Software, Inc., La Jolla, CA 92037, USA). A two-way ANOVA was used to estimate how the chemical parameters (TP, TF, CT, HPLC quantification of phenols and antioxidant capacity) of the different accessions, changed according to the temperature of extraction (20 °C and 95 °C). Data were reported as mean ± standard deviation (SD). In order to compare means obtained at 20 °C and 95 °C, a Student’s t-test was performed for every parameter within each chestnut accession and for the marron. The level of significance was fixed at p ≤ 0.05.
Biological tests were performed to compare the effects of different extract concentration (0, 10, 50, 100 and 250 µg/ml) on cancer cells, with the effect of the same concentration on fibroblasts. For each accession, a Student's t-test was used to compare the effects of different extract concentration on fibroblasts, setting the significance level at p ≤ 0.01 vs Ctrl. Then, a one-way ANOVA was performed to compare the effects of different extract concentration on cancer cells, setting the significance level at p ≤ 0.01 vs fibroblasts. The mean value ± SE was reported in the figures.
5. Conclusions
This paper deals with the antioxidant and anticancer properties of cold and hot water pericarp extracts from chestnut accessions belonging to the Sardinian biodiversity. The study showed that the extracts contained bioactive compounds whose properties are attributable, at least in part, to molecules that are extracted in different quantities, depending on temperature and contact times with the solvent. Cold extraction gives rise to mixtures of molecules that are apparently more effective than hot extracts in reducing the cellular viability of tumor cells but which, at the same time, have a greater impact on the viability of fibroblasts; for this reason, doses and duration of treatments must be carefully evaluated, regardless of the type of extract. As far as we know, there are only few studies reporting the use of chestnut extracts as potential antitumor agents in the treatment of colon and breast cancer, and this is the first time that chestnut extracts were tested on melanoma cells. The reported results are preliminary, but preparatory for further studies (i) to distinguish the different role of each of the main phenolic classes and, (ii) to combine the extracts with clinically standardized anti-tumor therapies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: cyclic voltammograms of growing concentration of gallic acid (a) and gallic acid calibration curve (b); Figure S2: HPLC characterization of cold (a) and hot (b) pericarp water extracts.
Author Contributions
Conceptualization, G.D. and A.M.; methodology, M.G.M. and Y.S.; formal analysis, G.R.S., G.S., M.G.M. and Y.S.; investigation, A.B.; data curation, M.G.M. and Y.S. writing—original draft preparation, A.B.; writing—review and editing, A.B., G.D., M.G.M. and Y.S.; project administration, G.D.; funding acquisition, G.D., A.B., M.G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This publication was funded by two different projects: (1) It is partially carried out within the Agritech National Research Center and received funding from the European Union Next-Generation EU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR)—MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4—D.D. 1032 17/06/2022, CN00000022). (2) It is partially carried out within the "ONFOODS project funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.3—Call for proposals No. 341 of 15 March 2022 of Italian Ministry of University and Research funded by the European Union—_Next-Generation EU. This manuscript reflects only the authors’ views and opinions, neither the European Union nor the European Commission can be considered responsible for them.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
The authors acknowledge Mr. Pietro Pisano and Mr. Antonello Petretto for their technical support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Sample Availability
Samples of the compounds are available from the authors.
References
- Poljak, I.; Idžojtić, M.; Šatović, Z.; Ježić, M.; Ćurković-Perica, M.; Simovski, B.; Acevski, J.; Liber, Z. Genetic Diversity of the Sweet Chestnut (Castanea Sativa Mill.) in Central Europe and the Western Part of the Balkan Peninsula and Evidence of Marron Genotype Introgression into Wild Populations. Tree Genetics & Genomes 2017, 13, 18–18. [CrossRef]
- Massantini, R.; Moscetti, R.; Frangipane, M.T. Evaluating Progress of Chestnut Quality: A Review of Recent Developments. Trends in Food Science & Technology 2021, 113, 245–254. [CrossRef]
- Piccolo, E.L.; Landi, M.; Ceccanti, C.; Mininni, A.N.; Marchetti, L.; Massai, R.; Guidi, L.; Remorini, D. Nutritional and Nutraceutical Properties of Raw and Traditionally Obtained Flour from Chestnut Fruit Grown in Tuscany. Eur Food Res Technol 2020, 246, 1867–1876. [CrossRef]
- De Vasconcelos, M.D.C.B.M.; Bennett, R.N.; Quideau, S.; Jacquet, R.; Rosa, E.A.S.; Ferreira-Cardoso, J.V. Evaluating the Potential of Chestnut (Castanea Sativa Mill.) Fruit Pericarp and Integument as a Source of Tocopherols, Pigments and Polyphenols. Industrial Crops and Products 2010, 31, 301–311. [CrossRef]
- Cerulli, A.; Napolitano, A.; Masullo, M.; Hošek, J.; Pizza, C.; Piacente, S. Chestnut Shells (Italian Cultivar “Marrone Di Roccadaspide” PGI): Antioxidant Activity and Chemical Investigation with in Depth LC-HRMS/MSn Rationalization of Tannins. Food Research International 2020, 129, 108787. [CrossRef]
- Braga, N.; Rodrigues, F.; P.P. Oliveira, M.B. Castanea Sativa by-Products: A Review on Added Value and Sustainable Application. Natural Product Research 2015, 29, 1–18. [CrossRef]
- Hu, M.; Yang, X.; Chang, X. Bioactive Phenolic Components and Potential Health Effects of Chestnut Shell: A Review. J. Food Biochem. 2021, 45. [CrossRef]
- Zhu, T.; Shen, Q.; Xu, Y.; Li, C. Ionic Liquid and Ultrasound-Assisted Extraction of Chestnut Shell Pigment with Good Hair Dyeing Capability. Journal of Cleaner Production 2022, 335, 130195. [CrossRef]
- Santos, M.J.; Pinto, T.; Vilela, A. Sweet Chestnut (Castanea Sativa Mill.) Nutritional and Phenolic Composition Interactions with Chestnut Flavor Physiology. Foods 2022, 11, 4052. [CrossRef]
- Pinto, D.; Moreira, M.M.; Vieira, E.F.; Švarc-Gajić, J.; Vallverdú-Queralt, A.; Brezo-Borjan, T.; Delerue-Matos, C.; Rodrigues, F. Development and Characterization of Functional Cookies Enriched with Chestnut Shells Extract as Source of Bioactive Phenolic Compounds. Foods 2023, 12, 640. [CrossRef]
- Silva, V.; Falco, V.; Dias, M.I.; Barros, L.; Silva, A.; Capita, R.; Alonso-Calleja, C.; Amaral, J.S.; Igrejas, G.; C. F. R. Ferreira, I.; et al. Evaluation of the Phenolic Profile of Castanea Sativa Mill. By-Products and Their Antioxidant and Antimicrobial Activity against Multiresistant Bacteria. Antioxidants 2020, 9, 87. [CrossRef]
- Cacciola, N.A.; Squillaci, G.; D’Apolito, M.; Petillo, O.; Veraldi, F.; La Cara, F.; Peluso, G.; Margarucci, S.; Morana, A. Castanea Sativa Mill. Shells Aqueous Extract Exhibits Anticancer Properties Inducing Cytotoxic and Pro-Apoptotic Effects. Molecules 2019, 24, 3401. [CrossRef]
- Schirra, M.; Palma, A.; Barberis, A.; Angioni, A.; Garau, V.L.; Cabras, P.; D’Aquino, S. Postinfection Activity, Residue Levels, and Persistence of Azoxystrobin, Fludioxonil, and Pyrimethanil Applied Alone or in Combination with Heat and Imazalil for Green Mold Control on Inoculated Oranges. Journal of Agricultural and Food Chemistry 2010, 58, 3661–3666. [CrossRef]
- Fadda, A.; Barberis, A.; D’Aquino, S.; Palma, A.; Angioni, A.; Lai, F.; Schirra, M. Residue Levels and Performance of Potassium Sorbate and Thiabendazole and Their Co-Application against Blue Mold of Apples When Applied as Water Dip Treatments at 20 or 53 Degrees C. Postharvest Biology and Technology 2015, 106, 33–43. [CrossRef]
- Fadda, A.; Virdis, A.; Barberis, A.; Melito, S. Variation in Secondary Metabolites Contents of Spinoso Sardo Artichoke (Cynara Cardunculus l.) under Different Day Lengths. Turkish Journal of Agriculture and Forestry 2018, 42, 372–381. [CrossRef]
- Ramli, I.; Posadino, A.M.; Zerizer, S.; Spissu, Y.; Barberis, A.; Djeghim, H.; Azara, E.; Bensouici, C.; Kabouche, Z.; Rebbas, K.; et al. Low Concentrations of Ambrosia Maritima L. Phenolic Extract Protect Endothelial Cells from Oxidative Cell Death Induced by H2O2 and Sera from Crohn’s Disease Patients. Journal of Ethnopharmacology 2023, 300, 115722. [CrossRef]
- Spissu, Y.; Gil, K.A.; Dore, A.; Sanna, G.; Palmieri, G.; Sanna, A.; Cossu, M.; Belhadj, F.; Gharbi, B.; Pinna, M.B.; et al. Anti- and Pro-Oxidant Activity of Polyphenols Extracts of Syrah and Chardonnay Grapevine Pomaces on Melanoma Cancer Cells. Antioxidants 2023, 12. [CrossRef]
- Salim, A.; Deiana, P.; Fancello, F.; Molinu, M.G.; Santona, M.; Zara, S. Antimicrobial and Antibiofilm Activities of Pomegranate Peel Phenolic Compounds: Varietal Screening Through a Multivariate Approach. Journal of Bioresources and Bioproducts 2023, S2369969823000154. [CrossRef]
- Aires, A.; Carvalho, R.; Saavedra, M.J. Valorization of Solid Wastes from Chestnut Industry Processing: Extraction and Optimization of Polyphenols, Tannins and Ellagitannins and Its Potential for Adhesives, Cosmetic and Pharmaceutical Industry. Waste Management 2016, 48, 457–464. [CrossRef]
- Fernández-Agulló, A.; Freire, M.S.; Antorrena, G.; Pereira, J.A.; González-Álvarez, J. Effect of the Extraction Technique and Operational Conditions on the Recovery of Bioactive Compounds from Chestnut ( Castanea Sativa ) Bur and Shell. Separation Science and Technology 2014, 49, 267–277. [CrossRef]
- Jung, B.S.; Lee, N.-K.; Na, D.S.; Yu, H.H.; Paik, H.-D. Comparative Analysis of the Antioxidant and Anticancer Activities of Chestnut Inner Shell Extracts Prepared with Various Solvents. Journal of the Science of Food and Agriculture 2016, 96, 2097–2102. [CrossRef]
- Squillaci, G.; Apone, F.; Sena, L.M.; Carola, A.; Tito, A.; Bimonte, M.; Lucia, A.D.; Colucci, G.; Cara, F.L.; Morana, A. Chestnut ( Castanea Sativa Mill.) Industrial Wastes as a Valued Bioresource for the Production of Active Ingredients. Process Biochemistry 2018, 64, 228–236. [CrossRef]
- Vella, F.M.; De Masi, L.; Calandrelli, R.; Morana, A.; Laratta, B. Valorization of the Agro-Forestry Wastes from Italian Chestnut Cultivars for the Recovery of Bioactive Compounds. Eur Food Res Technol 2019, 245, 2679–2686. [CrossRef]
- Galiñanes, C.; Freire, M.S.; González-Álvarez, J. Antioxidant Activity of Phenolic Extracts from Chestnut Fruit and Forest Industries Residues. Eur. J. Wood Prod. 2015, 73, 651–659. [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Evaluation of The Antioxidant Capacity of Food Products: Methods, Applications and Limitations. Processes 2022, 10. [CrossRef]
- Ricci, A.; Parpinello, G.P.; Teslić, N.; Kilmartin, P.A.; Versari, A. Suitability of the Cyclic Voltammetry Measurements and DPPH• Spectrophotometric Assay to Determine the Antioxidant Capacity of Food-Grade Oenological Tannins. Molecules 2019, 24. [CrossRef]
- Alam, M.W.; Najeeb, J.; Naeem, S.; Usman, S.M.; Nahvi, I.; Alismail, F.; Abuzir, A.; Farhan, M.; Nawaz, A. Electrochemical Methodologies for Investigating the Antioxidant Potential of Plant and Fruit Extracts: A Review. Antioxidants 2022, 11. [CrossRef]
- Zheng, Y.; Karimi-Maleh, H.; Fu, L. Evaluation of Antioxidants Using Electrochemical Sensors: A Bibliometric Analysis. Sensors 2022, 22. [CrossRef]
- Raja, A.N.; Annu; Singh, K.; Jain, R. Ultrasensitive Quantification of Ellagic Acid Using Gr/Bi2O3/GCE as Voltammetric Sensor. International Journal of Electrochemical Science 2020, 15, 10040–10057. [CrossRef]
- Ghoreishi, S.M.; Behpour, M.; Khayatkashani, M.; Motaghedifard, M.H. NEW APPLIED METHOD FOR SIMULTANEOUS DETERMINATION OF ELLAGIC AND TANNIC ACID BY MULTI-WALL CARBON NANOTUBE PASTE ELECTRODE: APPLICATION IN QUANTIFICATION PUNICA GRANATUM AND QUERCUS INFECTORIA. Digest Journal of Nanomaterials & Biostructures (DJNB) 2011, 6.
- Buratti, S.; Scampicchio, M.; Giovanelli, G.; Mannino, S. A Low-Cost and Low-Tech Electrochemical Flow System for the Evaluation of Total Phenolic Content and Antioxidant Power of Tea Infusions. Talanta 2008, 75, 312–316. [CrossRef]
- Spissu, Y.; Barberis, A.; D’hallewin, G.; Orrù, G.; Scano, A.; Serra, G.R.; Pinna, M.; Pinna, C.; Marceddu, S.; Serra, P.A. An Ascorbate Bluetooth© Analyzer for Quality Control of Fresh-Cut Parsley Supply Chain. Antioxidants 2021, 10. [CrossRef]
- Spissu, Y.; Barberis, A.; Bazzu, G.; D’hallewin, G.; Rocchitta, G.; Serra, P.A.; Marceddu, S.; Vineis, C.; Garroni, S.; Culeddu, N. Functionalization of Screen-Printed Sensors with a High Reactivity Carbonaceous Material for Ascorbic Acid Detection in Fresh-Cut Fruit with Low Vitamin C Content. Chemosensors 2021, 9. [CrossRef]
- Percevault, L.; Limanton, E.; Nicolas, P.; Paquin, L.; Lagrost, C. Electrochemical Determination and Antioxidant Capacity Modulation of Polyphenols in Deep Eutectic Solvents. ACS Sustainable Chemistry & Engineering 2021, 9, 776–784. [CrossRef]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of Methods to Determine Antioxidant Capacities. Food Analytical Methods 2009, 2, 41–60. [CrossRef]
- Lee, H.S.; Kim, E.J.; Kim, S.H. Chestnut Extract Induces Apoptosis in AGS Human Gastric Cancer Cells. nrp 2011, 5, 185–191. [CrossRef]
- Lee, J.H.; Khor, T.O.; Shu, L.; Su, Z.-Y.; Fuentes, F.; Kong, A.-N.T. Dietary Phytochemicals and Cancer Prevention: Nrf2 Signaling, Epigenetics, and Cell Death Mechanisms in Blocking Cancer Initiation and Progression. Pharmacology & therapeutics 2013, 137, 153–171. [CrossRef]
- Dashwood, R.H. Frontiers in Polyphenols and Cancer Prevention. The Journal of nutrition 2007, 137, 267S-269S. [CrossRef]
- Sorice, A.; Siano, F.; Capone, F.; Guerriero, E.; Picariello, G.; Budillon, A.; Ciliberto, G.; Paolucci, M.; Costantini, S.; Volpe, M. Potential Anticancer Effects of Polyphenols from Chestnut Shell Extracts: Modulation of Cell Growth, and Cytokinomic and Metabolomic Profiles. Molecules 2016, 21, 1411. [CrossRef]
- Beretta, G.; Moretti, R.M.; Nasti, R.; Cincinelli, R.; Dallavalle, S.; Montagnani Marelli, M. Apoptosis-Mediated Anticancer Activity in Prostate Cancer Cells of a Chestnut Honey (Castanea Sativa L.) Quinoline–Pyrrolidine Gamma-Lactam Alkaloid. Amino Acids 2021, 53, 869–880. [CrossRef]
- Santulli, C.; Brizi, C.; Durante, M.; Micucci, M.; Budriesi, R.; Chiarini, R.; Frosini, M. Apoptotic-Induced Effects of Castanea Sativa Bark Extract in Human SH-SY5Y Neuroblastoma Cells. NPC - Natural Product Communications 2018, 13, 887–890. [CrossRef]
- Sangiovanni, E.; Piazza, S.; Vrhovsek, U.; Fumagalli, M.; Khalilpour, S.; Masuero, D.; Di Lorenzo, C.; Colombo, L.; Mattivi, F.; De Fabiani, E.; et al. A Bio-Guided Approach for the Development of a Chestnut-Based Proanthocyanidin-Enriched Nutraceutical with Potential Anti-Gastritis Properties. Pharmacological Research 2018, 134, 145–155. [CrossRef]
- Barberis, A.; Spissu, Y.; Fadda, A.; Azara, E.; Bazzu, G.; Marceddu, S.; Angioni, A.; Sanna, D.; Schirra, M.; Serra, P.A. Simultaneous Amperometric Detection of Ascorbic Acid and Antioxidant Capacity in Orange, Blueberry and Kiwi Juice, by a Telemetric System Coupled with a Fullerene- or Nanotubes-Modified Ascorbate Subtractive Biosensor. Biosensors and Bioelectronics 2015, 67, 214–223. [CrossRef]
- Barberis, A.; Deiana, M.; Spissu, Y.; Azara, E.; Fadda, A.; Serra, P.A.; D’hallewin, G.; Pisano, M.; Serreli, G.; Orrù, G.; et al. Antioxidant, Antimicrobial, and Other Biological Properties of Pompia Juice. Molecules (Basel, Switzerland) 2020, 25, 3186. [CrossRef]
- Barberis, A.; Spissu, Y.; Bazzu, G.; Fadda, A.; Azara, E.; Sanna, D.; Schirra, M.; Serra, P.A. Development and Characterization of an Ascorbate Oxidase-Based Sensor–Biosensor System for Telemetric Detection of AA and Antioxidant Capacity in Fresh Orange Juice. Analytical Chemistry 2014, 86, 8727–8734. [CrossRef]
- Barberis, A.; Bazzu, G.; Calia, G.; Puggioni, G.M.G.; Rocchitta, G.G.; Migheli, R.; Schirra, M.; Desole, M.S.; Serra, P.A. New Ultralow-Cost Telemetric System for a Rapid Electrochemical Detection of Vitamin C in Fresh Orange Juice. Analytical Chemistry 2010, 82, 5134–5140. [CrossRef]
- Pedotti, S.; Patti, A.; Dedola, S.; Barberis, A.; Fabbri, D.; Dettori, M.A.; Serra, P.A.; Delogu, G. Synthesis of New Ferrocenyl Dehydrozingerone Derivatives and Their Effects on Viability of PC12 Cells. Polyhedron 2016, 117, 80–89. [CrossRef]
- Ham, J.-S.; Kim, H.-Y.; Lim, S.-T. Antioxidant and Deodorizing Activities of Phenolic Components in Chestnut Inner Shell Extracts. Industrial Crops and Products 2015, 73, 99–105. [CrossRef]
- Barreira, J.C.M.; Ferreira, I.C.F.R.; Oliveira, M.B.P.P. Bioactive Compounds of Chestnut (Castanea Sativa Mill.) BT - Bioactive Compounds in Underutilized Fruits and Nuts. In; Murthy, H.N., Bapat, V.A., Eds.; Springer International Publishing: Cham, 2020; pp. 303–313 ISBN 978-3-030-30182-8.
- Vázquez, G.; Fontenla, E.; Santos, J.; Freire, M.S.; González-Álvarez, J.; Antorrena, G. Antioxidant Activity and Phenolic Content of Chestnut (Castanea Sativa) Shell and Eucalyptus (Eucalyptus Globulus) Bark Extracts. Industrial Crops and Products 2008, 28, 279–285. [CrossRef]
- Cravotto, C.; Grillo, G.; Binello, A.; Gallina, L.; Olivares-Vicente, M.; Herranz-López, M.; Micol, V.; Barrajón-Catalán, E.; Cravotto, G. Bioactive Antioxidant Compounds from Chestnut Peels through Semi-Industrial Subcritical Water Extraction. Antioxidants 2022, 11. [CrossRef]
- Fadda, A.; Sanna, D. Advantages and Pitfalls of the Methods for the Antioxidant Activity Evaluation. In Advances in Food Analysis Research; A. Haynes (Ed.), Ed.; Nova Science Publishers, 2015; pp. 65–88.
- Volf, I.; Ignat, I.; Neamtu, M.; Popa, V. Thermal Stability, Antioxidant Activity, and Photo-Oxidation of Natural Polyphenols. Chemical Papers 2014, 68. [CrossRef]
- Vázquez, G.; González-Alvarez, J.; Santos, J.; Freire, M.S.; Antorrena, G. Evaluation of Potential Applications for Chestnut (Castanea Sativa) Shell and Eucalyptus (Eucalyptus Globulus) Bark Extracts. Industrial Crops and Products 2009, 29, 364–370. [CrossRef]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-Oxidant Activity of Polyphenols and Its Implication on Cancer Chemoprevention and Chemotherapy. Biochemical Pharmacology 2015, 98, 371–380. [CrossRef]
- Khan, H.Y.; Zubair, H.; Ullah, M.F.; Ahmad, A.; Hadi, S.M. A Prooxidant Mechanism for the Anticancer and Chemopreventive Properties of Plant Polyphenols. Current Drug Targets 2012, 13, 1738–1749. [CrossRef]
- Pasciu, V.; Posadino, A.M.; Cossu, A.; Sanna, B.; Tadolini, B.; Gaspa, L.; Marchisio, A.; Dessole, S.; Capobianco, G.; Pintus, G. Akt Downregulation by Flavin Oxidase–Induced ROS Generation Mediates Dose-Dependent Endothelial Cell Damage Elicited by Natural Antioxidants. Toxicological Sciences 2010, 114, 101–112. [CrossRef]
- Giordo, R.; Cossu, A.; Pasciu, V.; Hoa, P.T.; Posadino, A.M.; Pintus, G. Different Redox Response Elicited by Naturally Occurring Antioxidants in Human Endothelial Cells. The Open Biochemistry Journal 2013, 7, 44–53. [CrossRef]
- Posadino, A.M.; Giordo, R.; Cossu, A.; Nasrallah, G.K.; Shaito, A.; Abou-Saleh, H.; Eid, A.H.; Pintus, G. Flavin Oxidase-Induced ROS Generation Modulates PKC Biphasic Effect of Resveratrol on Endothelial Cell Survival. Biomolecules 2019, 9. [CrossRef]
- Floris, A.; Mazarei, M.; Yang, X.; Robinson, A.E.; Zhou, J.; Barberis, A.; D’hallewin, G.; Azara, E.; Spissu, Y.; Iglesias-Ara, A.; et al. SUMOylation Protects FASN Against Proteasomal Degradation in Breast Cancer Cells Treated with Grape Leaf Extract. Biomolecules 2020, 10. [CrossRef]
- Zhang, L.; Gao, H.; Baba, M.; Okada, Y.; Okuyama, T.; Wu, L.; Zhan, L. Extracts and Compounds with Anti-Diabetic Complications and Anti-Cancer Activity from Castanea Mollissina Blume (Chinese Chestnut). BMC Complementary and Alternative Medicine 2014, 14, 422–422. [CrossRef]
- Woo, Y.; Oh, J.; Kim, J.-S. Suppression of Nrf2 Activity by Chestnut Leaf Extract Increases Chemosensitivity of Breast Cancer Stem Cells to Paclitaxel. Nutrients 2017, 9. [CrossRef]
- Muroni, A.; D’hallewin, G.; Satta, D.; De Pau, L. La Biodiversità Castanicola Della Sardegna Centrale. In Proceedings of the VIII Convegno Nazionale del Castagno; Di Vaio, C., Cirillo, C., Eds.; Italian Society for Horticultural Sciences (SOI) ISBN: 978-88-32054-04-0: Portici (Napoli), 2023; pp. 46–46.
- Molinu, M.G.; Sulas, L.; Campesi, G.; Re, G.A.; Sanna, F.; Piluzza, G. Subterranean Clover and Sulla as Valuable and Complementary Sources of Bioactive Compounds for Rainfed Mediterranean Farming Systems. Plants 2023, 12, 417. [CrossRef]
- Piluzza, G.; Campesi, G.; D’hallewin, G.; Molinu, M.G.; Re, G.A.; Sanna, F.; Sulas, L. Antioxidants in Fruit Fractions of Mediterranean Ancient Pear Cultivars. Molecules 2023, 28, 3559. [CrossRef]
- José Jara-Palacios, M.; Hernanz, D.; Luisa Escudero-Gilete, M.; Heredia, F.J. Antioxidant Potential of White Grape Pomaces: Phenolic Composition and Antioxidant Capacity Measured by Spectrophotometric and Cyclic Voltammetry Methods. Food Research International 2014, 66, 150–157. [CrossRef]
Figure 1.
Cyclic voltammetry, with a scanned potential range (Eapp) comprised between –0.2 V and +0.8 V vs. carbon pseudoreference, in the absence (PBS black line) and in the presence of 2 mg ml−1 of MURG (green line), LOCG (purple line), ILDP (red line), COEV (blue line) and MARRV (yellow line) chestnut pericarp cold water extracts (A) and hot water extracts (B).
Figure 1.
Cyclic voltammetry, with a scanned potential range (Eapp) comprised between –0.2 V and +0.8 V vs. carbon pseudoreference, in the absence (PBS black line) and in the presence of 2 mg ml−1 of MURG (green line), LOCG (purple line), ILDP (red line), COEV (blue line) and MARRV (yellow line) chestnut pericarp cold water extracts (A) and hot water extracts (B).
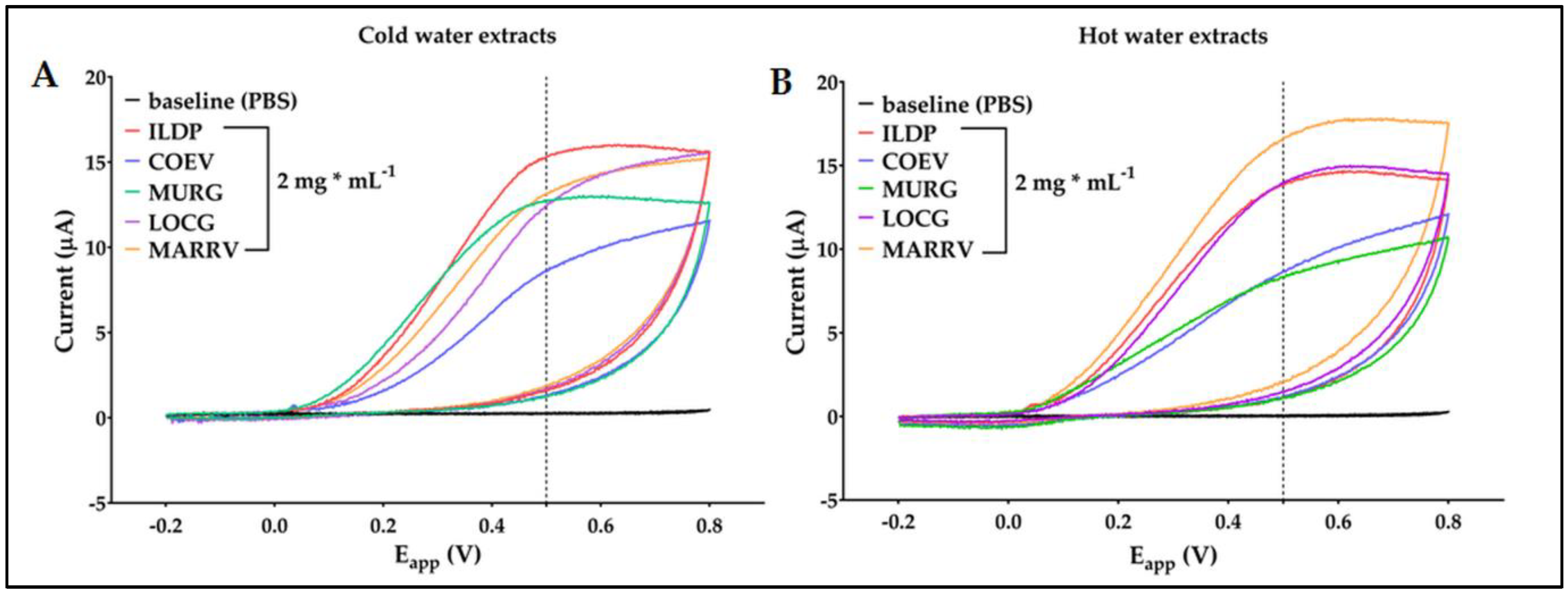
Figure 2.
Effect of growing concentrations (from 10 to 250 µg/ml) of MURG chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
Figure 2.
Effect of growing concentrations (from 10 to 250 µg/ml) of MURG chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
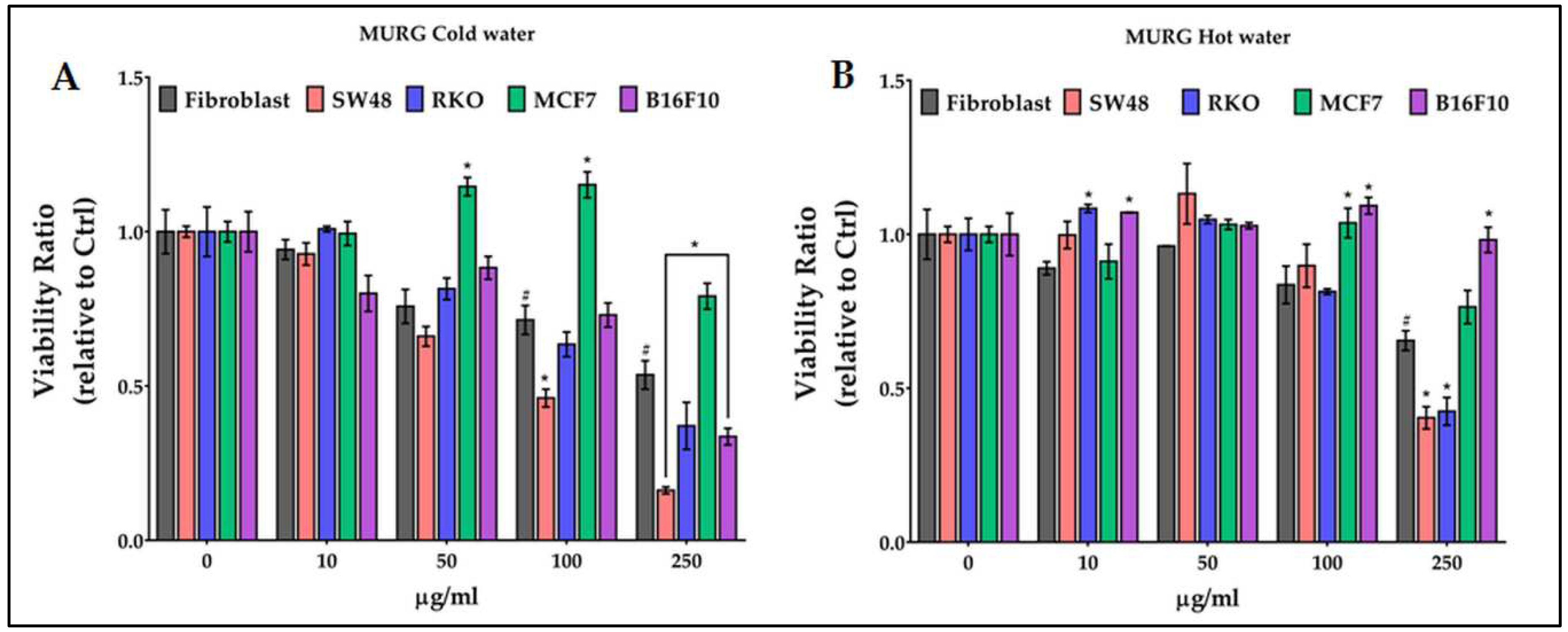
Figure 3.
Effect of growing concentrations (from 10 to 250 µg/ml) of LOCG chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
Figure 3.
Effect of growing concentrations (from 10 to 250 µg/ml) of LOCG chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
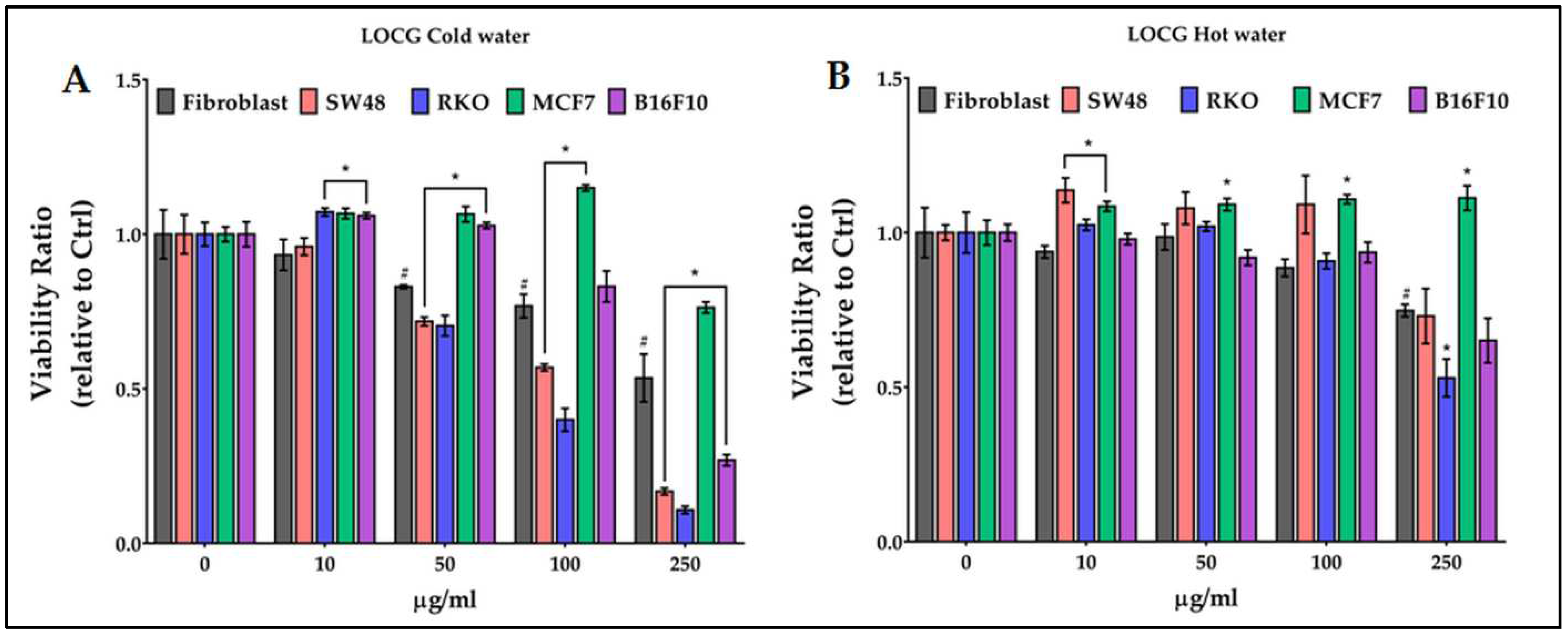
Figure 4.
Effect of growing concentrations (from 10 to 250 µg/ml) of ILDP chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
Figure 4.
Effect of growing concentrations (from 10 to 250 µg/ml) of ILDP chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
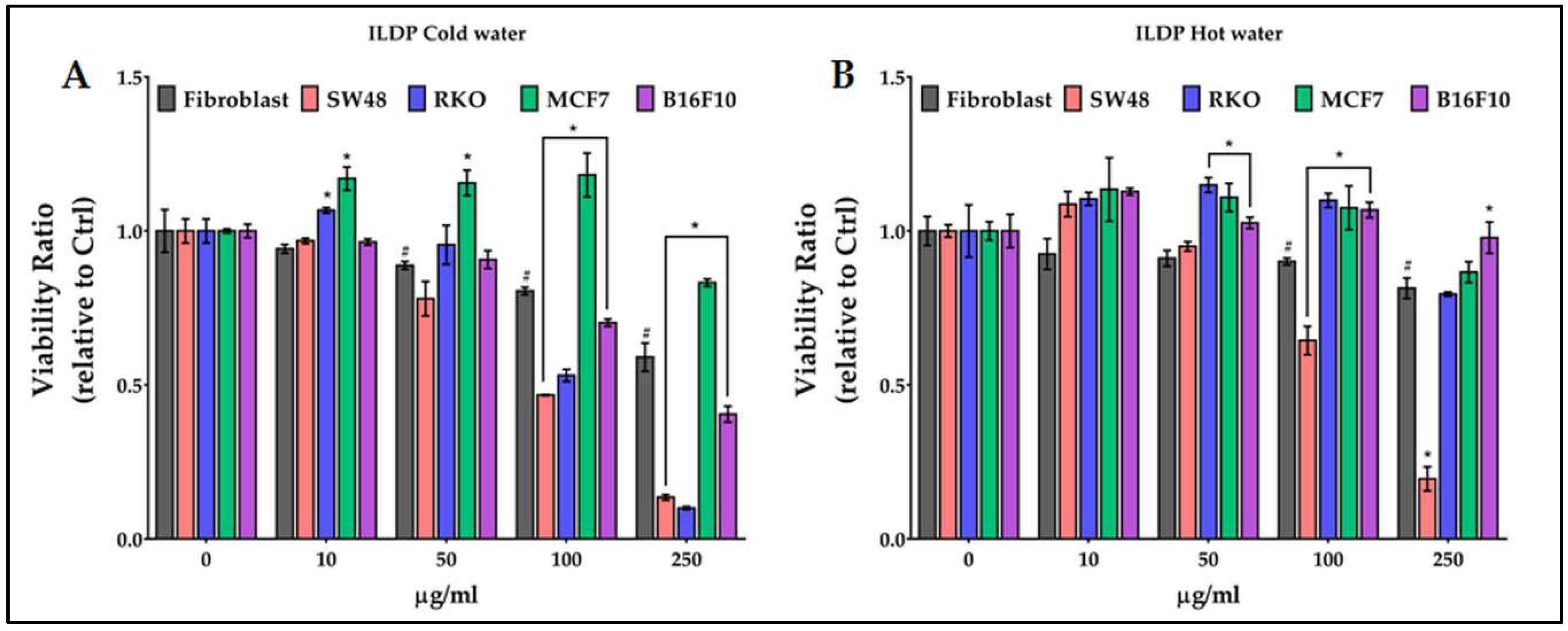
Figure 5.
Effect of growing concentrations (from 10 to 250 µg/ml) of COEV chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
Figure 5.
Effect of growing concentrations (from 10 to 250 µg/ml) of COEV chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
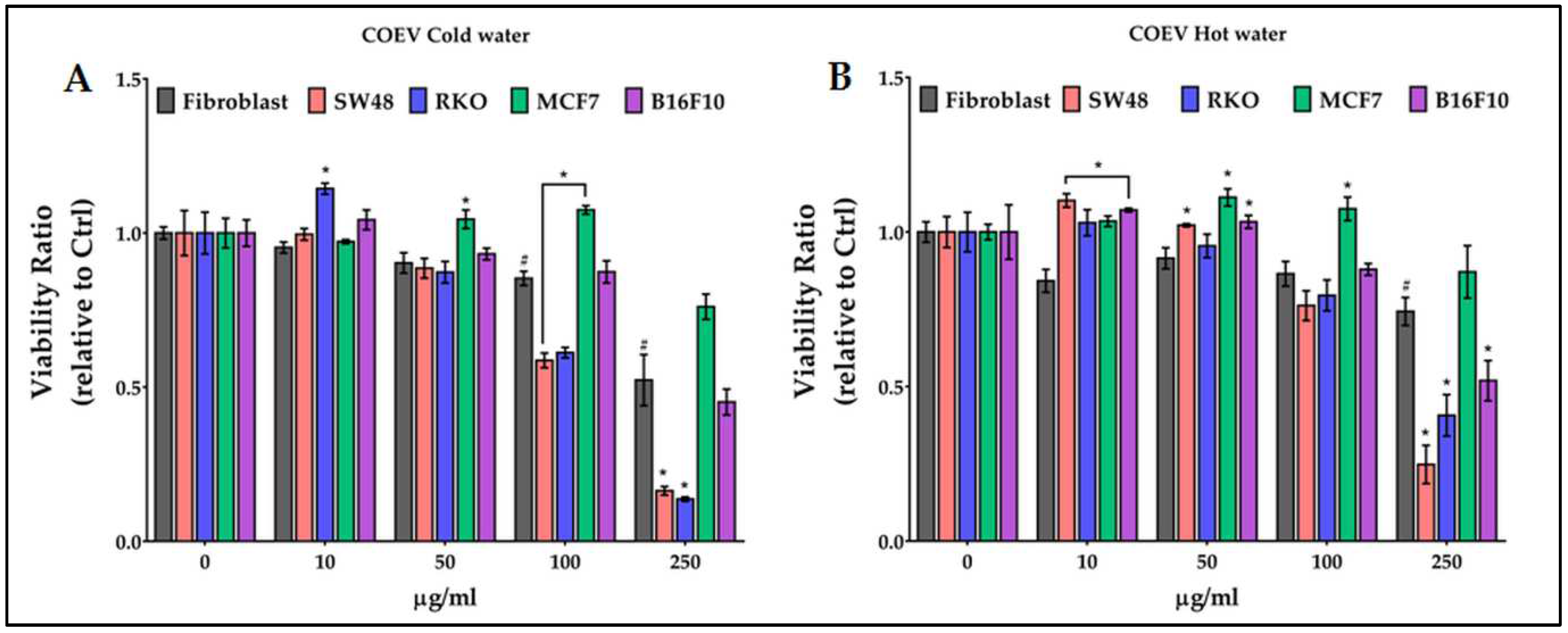
Figure 6.
Effect of growing concentrations (from 10 to 250 µg/ml) of MARRV chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
Figure 6.
Effect of growing concentrations (from 10 to 250 µg/ml) of MARRV chestnut pericarp extract on viability of normal cells (fibroblasts), and of colon (SW48 and RKO), breast (MCF7) and melanoma (B16F10) cancer cells. # = p ≤ 0.01 vs Ctrl; * = p ≤ 0.01 vs fibroblasts.
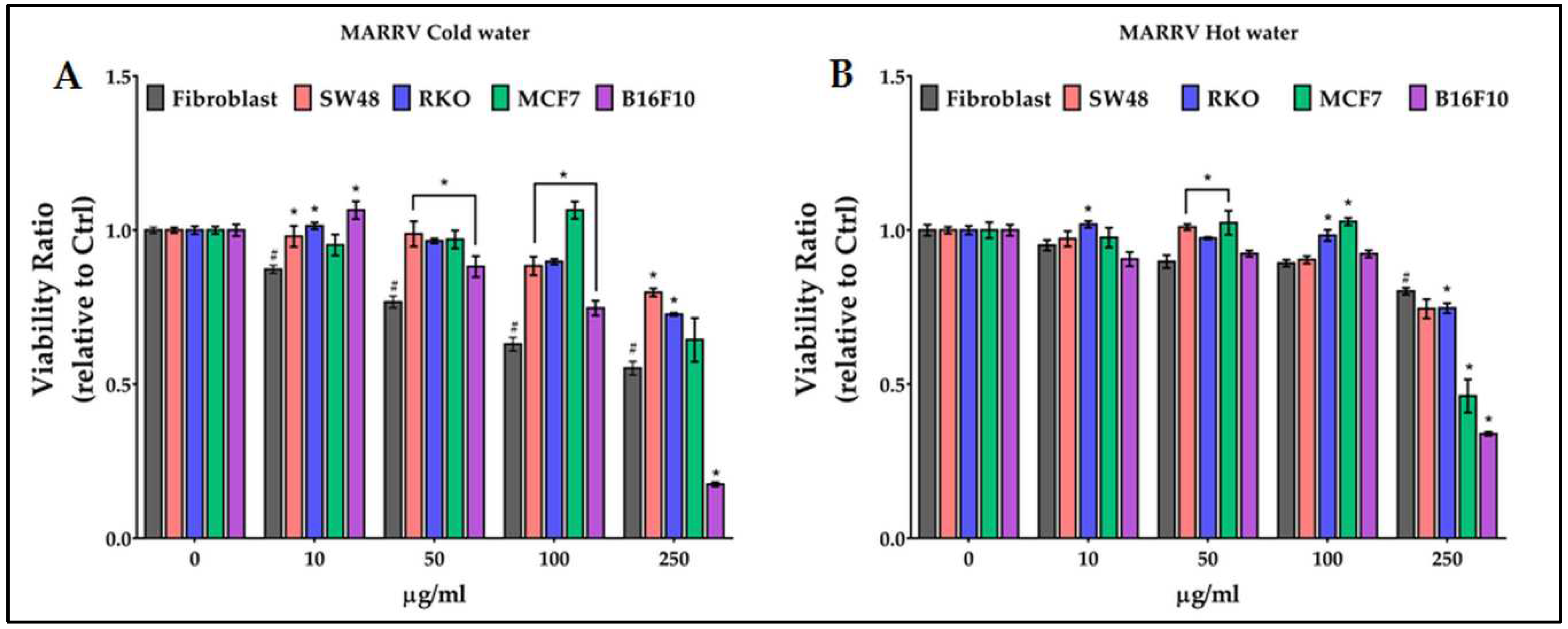
Table 1.
Total phenolics (TP), total flavonoids (TF), condensed tannins (CT), and antioxidant capacity (TEAC) determined by DPPH, ABTS and by electrochemical method (Cyclic Voltammetry), in the extracts of the pericarp chestnut samples.
Table 1.
Total phenolics (TP), total flavonoids (TF), condensed tannins (CT), and antioxidant capacity (TEAC) determined by DPPH, ABTS and by electrochemical method (Cyclic Voltammetry), in the extracts of the pericarp chestnut samples.
| Accessions or variety |
T °C | Total Phenols | Total Flavonoids | Condensed Tannins | DPPH | ABTS | CV |
|---|---|---|---|---|---|---|---|
| mg GAE g-1 DW | mg CE g-1 DW | mg CE g-1 DW | mmol TEAC 100 g-1 DM | AUC (µC) | |||
| MURG | 20 95 |
39.58 ± 1.69 37.35 ± 0.64 |
18.69 ± 0.42* 16.52 ± 0.14* |
9.10 ± 0.72 9.20 ± 0.20 |
24.29 ± 1.16* 31.77 ± 2.40* |
33.33 ± 1.24 34.02 ± 0.67 |
3.02 ± 0.12* 2.04 ± 0.07* |
| LOCG | 20 95 |
44.33 ± 2.89 40.10 ± 2.16 |
12.92 ± 1.70* 16.44 ± 2.3* |
6.37 ± 1.17 7.55 ± 1.38 |
23.91 ± 1.34 27.52 ± 2.84 |
31.17 ± 0.31 29.57 ± 0.92 |
2.16 ± 0.13* 2.97 ± 0.19* |
| ILDP | 20 95 |
42.96 ± 1.25 38.92 ± 2.89 |
11.37 ± 1.14* 16.26 ± 1.62* |
7.05 ± 1.25 7.99 ± 0.96 |
27.20 ± 2.89 28.17 ± 2.39 |
35.71 ± 1.50 34.51 ± 0.91 |
3.15 ± 0.15 3.17 ± 0.31 |
| COEV | 20 95 |
25.06 ± 0.46 22.70 ± 0.43 |
6.38 ± 0.21* 9.29 ± 1.66* |
4.73 ± 0.59 4.02 ± 0.51 |
15.31 ± 0.36* 20.07 ± 2.94* |
21.03 ± 0.37* 25.11 ± 2.49* |
1.52 ± 0.37 1.87 ± 0.28 |
| MARRV | 20 95 |
15.04 ± 3.38* 31.47 ± 1.58* |
4.33 ± 1.73 3.47 ± 0.68 |
1.28 ± 0.11* 3.53 ± 0.09* |
9.12 ± 1.74* 14.22 ± 0.24* |
11.60 ± 0.20* 18.59 ± 0.05* |
2.61 ± 0.20* 3.78 ± 0.25* |
| A x T | # | # | n.s. | # | # | # | |
According to the two-way Anova, the # indicates the presence of interaction between accessions and temperature of extraction. n.s. = not significant A Student’s t-test was used to compare data obtained at 20 °C and 95 °C within each chestnut accession and for the marron. When present, the * indicates that means significantly differ at p ≤ 0.05.
Table 2.
Main phenolic compounds (mg g−1 DM) quantified by HPLC analysis in the pericarp extracts of the four chestnut accessions and of the marron variety.
Table 2.
Main phenolic compounds (mg g−1 DM) quantified by HPLC analysis in the pericarp extracts of the four chestnut accessions and of the marron variety.
| Phenolic compound | T °C | MURG | LOCG | ILDP | COEV | MARRV |
|---|---|---|---|---|---|---|
| Gallic acid | 20 95 |
0.402 ± 0.003* 0.300 ± 0.007* |
0.553 ± 0.001* 0.390 ± 0.027* |
0.929 ± 0.125* 0.349 ± 0.025* |
0.299 ± 0.022 0.280 ± 0.029 |
0.131 ± 0.040* 0.255 ± 0.009* |
| Ellagic acid | 20 95 |
2.864 ± 0.032* 0.573 ± 0.019* |
4.732 ± 0.190* 0.727 ± 0.090* |
4.469 ± 0.492* 0.684 ± 0.075* |
3.395 ± 0.434* 0.363 ± 0.050* |
1.659 ± 0.068* 0.869 ± 0.033* |
| Vescalagin | 20 95 |
0.525 ± 0.058* 0.768 ± 0.007* |
0.406 ± 0.013* 0.998 ± 0.069* |
0.364 ± 0.037* 0.629 ± 0.012* |
0.237 ±0.001* 0.409 ±0.025* |
0.168 ± 0.029* 0.463 ± 0.007* |
| Castalagin | 20 95 |
0.631 ± 0.041 0.714 ± 0.015 |
0.449 ± 0.021* 0.980 ± 0.167* |
0.364 ± 0.037* 0.629 ± 0.012* |
0.246 ± 0.012* 0.419 ±0.064* |
0.238 ±0.060 0.374 ± 0.050 |
| Catechin | 20 90 |
0.219 ± 0.008 0.231 ± 0.009 |
0.261 ± 0.022 0.268 ± 0.016 |
0.564 ± 0.028* 0.248 ± 0.002* |
0.161 ± 0.005 0.180 ± 0.017 |
0.199 ± 0.027* 0.670 ± 0.013* |
| Epicatechin | 20 95 |
0.595 ± 0.077 0.665 ± 0.016 |
0.271 ± 0.033* 2.232 ± 0.279* |
0.732 ± 0.067 0.900 ± 0.009 |
0.287 ± 0.044 0.285 ± 0.065 |
0.204 ±0.011* 0.521 ± 0.008* |
| Epigallocatechin | 20 95 |
0.067 ± 0.012 0.052 ± 0.015 |
0.081 ± 0.011 0.063 ± 0.001 |
0.091 ± 0.020 0.075 ± 0.003 |
0.055 ± 0.001 0.041 ± 0.000 |
0.046 ± 0.004 0.056 ± 0.006 |
A Student’s t-test was used to compare data obtained at 20 °C and 95 °C within each chestnut accession and for the marron. When present, the * indicates that means significantly differ at p ≤ 0.05.
Table 3.
Redox potential values of standards of the seven main phenolic compounds found in the chestnut epicarp extracts of MURG, ILDP, LOCG, COEV and MARRV.
Table 3.
Redox potential values of standards of the seven main phenolic compounds found in the chestnut epicarp extracts of MURG, ILDP, LOCG, COEV and MARRV.
| Phenolic compound | Redox potential (V) | Reference |
|---|---|---|
| Gallic acid | + 0.391 + 0.180 |
[26,43] |
| Ellagic acid | + 0.367 | [26] |
| Vescalagin | + 0.384 | [26] |
| Castalagin | + 0.384 | [26] |
| Catechin | + 0.391 + 0.300 + 0.120 |
[26,43,44] |
| Epicatechin | + 0.120 | [44] |
| Epigallocatechin | + 0.080 | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



