Submitted:
23 October 2023
Posted:
23 October 2023
You are already at the latest version
Abstract
To study the fermentation process of biocatalyzing decanoic acid to synthesize trans-2-decenoic acid, this study was based on the engineering E.coli constructed in the laboratory in the early stage and studied the effect of seed liquid culture time, culture temperature, inoculum amount, induction temperature, dissolution effects of substrate solvent, metal ions, and substrate loading on the yield of trans-2-decenoic acid. Based on single-factor experimental optimization, the Box–Behnken design was used to conduct response surface testing using the substrate feeding concentration, inducer concentration, and MnCl2 concentration as response variables and trans-2-decenoic acid production as the response value. The optimal fermentation process was determined through experiments: seed culture time 20 h, culture temperature 37°C, inoculation amount 1%, induction temperature 30°C, substrate flow amount 0.15 g/L, inducer concentration 5.60 g/L, MnCl2 concentration was 0.10 mM. Under these conditions, the average production of trans-2-decenoic acid was 1.982 ± 0.110 g/L. Compared with the basic LB medium, the production of trans-2-decenoic acid increased by 1.042 g/L; compared with the previous period, the yield of trans-2-decenoic acid studied increased by 1.501 ± 0.110 g/L, which provides a reliable basis for further research on the fermentation process of biocatalytic decanoic acid synthesis of trans-2-decenoic acid.
Keywords:
decanoic acid
; trans-2-decenoic acid
; engineered E.coli
; optimization
; response surface
1. Introduction
Trans-2-decenoic acid is a medium-chain fatty acid with ten carbon atoms and an unsaturated double bond at the α and β carbon positions. Its molecular formula is C10H18O2, and its relative molecular mass is 170.25. The specific structural formulas are shown in Figure 1. As an essential and uniquely structured medium-chain α, β-unsaturated fatty acid, trans-2-decenoic acid has a wide range of applications, so it is essential to explore its synthesis method. The most common way to produce α, β-unsaturated fatty acid is through chemical synthesis. Chemical methods include dehydrogenating established carbonyl compounds [1], aldol condensation [2], and Wittig-type [3] reactions. However, when using various raw materials to synthesize target substances through chemical synthesis, there are common shortcomings, such as serious environmental pollution, low reaction controllability, and a low safety guarantee factor. Therefore, compared with traditional methods, the biocatalytic synthesis of target substances has attracted widespread attention because of its advantages, such as high safety, strong specificity, and low environmental pollution [4].
α, β-unsaturated fatty acids generally have an aromatic smell and are primarily found in some animal fats [5] and some plant fruits. In the biosynthesis of royal jelly acid (10-hydroxy-2-decenoic, 10-HDA), trans-2-decenoic acid can be used as a precursor for the biosynthesis of 10-HDA [6]. As a key factor in royal jelly, 10-HDA plays a role in antibacterial activity [7,8], cancer prevention and treatment, and immunity regulation [9]. The biosynthesis of trans-2-decenoic acid provides a basis for future biosynthetic pathways of 10-HDA. Trans-2-decenoic acid can also be used as a ligand to synthesize anisotropic nanocrystalline materials. Lu et al [9] and others confirmed that using organic carboxylic acid ligands plays an essential role in the size and morphology of the synthesized one-dimensional structured nanocrystals, and only when trans-2-decenoic acid is used as the ligand does crystal growth become quicker, producing elongated nanocrystals.
Whether a substance is antibacterial is usually closely related to the special functional groups, chain length, and substituent group position [10,11]. Usually, the special functional groups, chain length, and substituent group position determine whether it is bacteriostatic; Marques et al. [12] reported that 2-decenoic acid is responsible for induction in a series of Gram-negative and Gram-positive bacteria and yeast. The dispersion of natural biofilms increases microbial metabolic activity, significantly enhancing the bactericidal effects of conventional antibacterial drugs. Cai et al. [13] reported that trans-2-decenoic acid plays a vital role in inducing the disintegration of aerobic particles. After adding trans-2-decenoic acid, the surfaces of aerobic particles, bacteria, and extracellular polymers are stripped away. Therefore, trans-2-decenoic acid is widely used in medicine.
The biosynthesis of fatty acids has gradually become a research hotspot [14]. Liu et al. synthesized crotonic acid through the fatty acid biosynthesis pathway in E. coli strains using a specific thioesterase (bTE) derived from Bacteroides thetaiotaomicron to catalyze the hydrolysis of crotonyl-ACP to synthesize crotonyl-ACP butyric acid [15]. However, there are few reports on the biocatalytic synthesis of trans-2-decenoic acid. The first report on the biocatalytic synthesis of decenoic acid was on the metabolic pathway of biosynthesis of crotonic acid by modifying the β-oxidation pathway; 2-decenoic acid is a by-product of its metabolic pathway, and only trace amounts of 2-decenoic acid can be detected in the by-products, with a yield of only 9.45 mg/L. However, the successful synthesis of 2-decenoic acid has laid an important foundation for microbial production of α, β-unsaturated fatty acids [16,17].
The biocatalytic synthesis of trans-2-decenoic acid relies on the β-oxidative decarboxylation process of fatty acids [18,19]. As shown in Figure 2, the β-oxidation pathway was modified in E. coli in the early stage of the laboratory, relying on fatty acyl-CoA synthase FadD, fatty acyl-CoA dehydrogenase FadE, and fatty acyl-CoA thioesterase YdiI to achieve the addition of double bonds and knocking. In addition to the genes encoding enoyl-CoA synthase FadB and the protein operon FadR [20], they inhibit the consumption of the intermediate product branch, thereby catalyzing the synthesis of trans-2-decenoic acid from decanoic acid [6,21]. Based on this metabolic process, FadD (MaMACS) from Mycobacterium avium was screened to activate the substrate decanoic acid because it has the highest activity on decanoic acid compared to caprylic acid or dodecanoic acid and longer chain fatty acids [22], so it was selected to connect CoA to the saturated fatty acid decanoic acid; the fatty acyl-CoA dehydrogenase (PpFadE) from Pseudomonas putida KT2440 was screened. PpFadE has broad substrate specificity; fatty acyl CoA with a chain length of 14 carbon atoms is more preferred [23], catalyzing the dehydrogenation of decanoyl-CoA to generate trans-2-decenoyl-CoA [24]; lipids from Cronobacter turicensis were screened for acyl-CoA thioesterase (CtYdiI) due to its higher catalytic ability for mid-chain trans-2-decenoyl, achieving higher efficiency in hydrolysing CoA in trans-2-decenoyl-CoA, obtaining trans-2-decenoic acid [25,26].
Owing to the extensive role of trans-2-decenoic acid in medicine, food, health care, etc., the industrial production of trans-2-decenoic acid has become particularly important. Although microbial fermentation has been widely researched and applied in recent years, owing to its low cost and ease of operation, there have been few studies on medium-chain unsaturated fatty acids [27]. Therefore, we selected the engineering strain (E. coli BL21(DE3)-△fadB-△fadJ-△fadR/pET28a-sumo-CtydiI/pCDFDuet-1-MaMACS-PpfadE) that was constructed and saved in the laboratory in the early stage to design a reasonable fermentation method. The culture medium and conditions were optimized to increase the yield and achieve the goal of high industrial production of trans-2-decenoic acid.
The response surface method (RSM) was used to obtain the optimal parameters of complex systems by establishing mathematical models, regression analysis, and variance analysis [28,29]. It can optimize submerged fermentation media and culture conditions [30,31]. The statistical model based on the Box–Behnken design (BBD) is a response surface design method [32]. To optimize the fermentation medium and conditions, we used the BBD to explore the effects of different nutrients, such as the substrate dissolution method, substrate flow amount, carbon source, and metal ions, on the synthesis of trans-2-decenoic acid. Through preliminary screening of single-factor experiments, the key factors affecting fermentation were selected, and a fed-batch fermentation experiment was conducted in a 1 L fermentation tank, efficiently increasing the fermentation yield of trans-2-decenoic acid.
2. Materials and Methods
2.1. Strains
The engineering strain (E. coli BL21(DE3)-△fadB-△fadJ-△fadR/pET28a-sumo-CtydiI/pCDFDuet-1-MaMACS-PpfadE) was constructed in the early stage of the experiment.
2.2. Activation and culture of engineered Escherichia coli.
2.2.1. Strain activation
Solid LB seed culture medium was prepared and sterilized at 121°C for 20 min. The preserved engineering strain (E. coli BL21(DE3)-△fadB-△fadJ-△fadR/pET28a-sumo-CtydiI/pCDFDuet-1-MaMACS-PpfadE) was streaked on a solid LB plate and cultured at a biological constant temperature 37°C for 16 h.
2.2.2. Strain culture and preservation.
A single colony strain grown on the plate was added to and grown in a 50 mL triangular flask and 40 μL streptomycin and 50 μL kanamycin were added. A 50 mL triangular flask was placed and incubated in a constant-temperature shaking incubator at 37°C (200 rpm) for 12 h. After 12 h, the bacterial solution was removed, aliquoted, and stored at −80°C.
2.3. Determination of substrates and target products based on Gas Chromatography.
2.3.1. Sample pretreatment
The removed sample was dispensed into a 2 mL EP tube, and 1 mL of 4 M hydrochloric acid solution was added. The sample was placed in a shaking incubator (30°C, 200 rpm) for 1 h, boiled in boiling water for 10 min, and then frozen at −80°C for 15 min. The sample was then divided into. Ethyl acetate (750 μL) was added to two 2 mL EP tubes and vortexed vigorously for 20 s for extraction. The sample was centrifuged at 8000 rpm for 10 min to separate the aqueous and organic layers. The above procedure was repeated twice, and the supernatant from the two extractions was used. The liquid was collected in a 2 mL centrifuge tube. The collected samples were dried using a nitrogen blower (37°C), and 500 μL ethyl acetate, 500 μL n-hexane, and 100 μL DTFA-TMCS (99:1) were added and mixed. After being left at room temperature for 5 min, the sample was placed in a 70°C oven for 50 min to react, and silanization was performed. The obtained liquid was filtered using a 0.22 μm organic filter membrane, then stored and transferred to a gas chromatography vial for later use.
2.3.2. Gas Chromatography Detection
Six standard samples with different concentration gradients were prepared as the substrates and products. The concentrations were 0.0625 g/L, 0.125 g/L, 0.25 g/L, 0.5 g/L, 1 g/L, and 2 g/L. An island Jin GC-2030 gas chromatograph was used for analysis, with the chromatographic capillary column as HP-5 and the instrument carrier gas. The inlet conditions were set to constant flow mode, the injection volume was 1 μL, the split injection mode was selected, the injection split ratio was maintained at 1:50, the injection temperature was 250°C, and the constant flow rate was set to 1 mL/min. Based on the experimental exploration and product characteristics, the temperature rise program was maintained at 50°C for 1 min, raised to 250°C at 15°C/min, and held for 10 min.
2.4. Single-factor analysis
Based on the preliminary laboratory work, the seed culture temperature, seed liquid culture time, inoculation amount, induction temperature, substrate dissolution method, inducer concentration, substrate flow concentration, and reaction of different metal compounds were determined through single-factor experiments on the effects on the trans-2-decenoic acid yield and conversion rate. The seed culture temperatures were set to 25°C, 28°C, 30°C, 33°C, and 37°C; the seed liquid culture times were set to 8 h, 10 h, 12 h, 16 h, and 20 h; the inoculum amounts were set to 1%, 3%, 5%, 8%, and 10%; the induction temperatures were set to 20°C, 25°C, 28°C, 30°C, and 37°C; absolute ethanol, Tween-80, decane, N, N-Dimethylformamide (DMF) and Dimethyl sulfoxide (DMSO) were used as organic solvents to dissolve capric acid [33,34,35]; the substrate flow concentrations were 0, 0.1 g/L, 0.2g/ L, 0.3 g/L, and 0.4g/L; the metal compounds [36,37] were MnCl2, CaCl2, MgCl2·6H2O, ZnCl2, and CuCl2.
2.5. Box–Behnken design
This experiment used a BBD to design and optimize fermentation conditions and adopted a three-factor and three-level experimental method. Based on the results of previous single-factor experiments, three factors that had a significant impact on the production of trans-2-decenoic acid were selected, as shown in Table 1: substrate flow concentration, inducer concentration, and MnCl2. The RSM was used to evaluate the fermentation conditions. Optimization was performed to determine the optimal fermentation conditions.
2.6. Comparison of expression of E. coli transporter proteins before and after optimization.
Owing to the complex metabolic pathways in E. coli, the substrate decanoic acid not only participates in the biosynthetic pathway but also partially in the metabolic cycle. We removed fatty acids of specific chain lengths from CoA by enhancing YdiI, FadD, and FadE expression levels to promote the β-oxidation reaction, promoting the large-scale synthesis of product intermediates, and increasing the production of trans-2-decenoic acid. A series of catalytic fermentation reactions were carried out on successfully constructed recombinant engineered bacteria, and the protein expression levels before and after optimization were compared. By comparing the expression levels of FadD, FadE, and YdiI, we determined whether the protein expression levels significantly improved after optimizing the fermentation process. This improved, verifying the feasibility of the fermentation optimization results.
3. Results
3.1. Gas chromatography determination of substrate and product results
According to the gas chromatography results, the peak time of the six different concentration gradients of the substrate decanoic acid was at approximately 11.221 min; the peak time of the six different concentration gradients of the product trans-2-decenoic acid was at approximately 11.731 min. Determining the peak time of the substrate and product provided a reliable basis for detecting substrate consumption and product production in the next step.
3.1.1. Effect of seed culture temperature on trans-2-decenoic acid production
The role of temperature in E. coli fermentation is critical because temperatures outside the optimal range can affect cell growth, biomass, and yield. Temperature plays a key role in the synthesis of metabolites, regulation of enzymes, and biosynthesis of target products during the middle and late stages of fermentation [38]. As shown in Figure 3A, in terms of seed culture temperature, the yield of trans-2-decenoic acid is highest when the seeds are cultured at 37°C, with a yield of 1.01 ± 0.03 g/L, consistent with the optimal temperature for E. coli growth; therefore, we took 37°C as the optimal seed cultivation temperature.
3.1.2. Effect of seed culture time on trans-2-decenoic acid production
Seed activity played a decisive role in the reaction system. Usually, strains in the exponential phase are used for the reaction; therefore, a reasonable seed growth time is the key to reaction efficiency. As shown in Figure 3B, when the seed culture time was 16 h, the yield of trans-2-decenoic acid was highest, 0.711 ± 0.011 g/L, so we took 16 h as the optimal seed culture time.
3.1.3. Effect of inoculation amount on trans-2-decenoic acid production
If the inoculation amount is too small, the growth of the bacteria will be slow, and the metabolic efficiency will be low. If the inoculation amount is too large, the ability of the bacteria to produce enzymes is inhibited, and the metabolic reaction efficiency will be affected. As shown in Figure 3-C, when the inoculation amount was 1%, the yield of trans-2-decenoic acid was highest, 1.212 ± 0.017 g/L; therefore, we selected 1% inoculation amount as optimal.
3.1.4. Effect of induction temperature on trans-2-decenoic acid production.
The key to this reaction is using inducers for bacterial enzyme production. Appropriate induction timing and temperature can increase the accuracy and efficiency of enzyme production. Early induction may result in a metabolic burden related to protein overexpression in the host strains. Cell product formation occurs at higher cell densities after the exponential phase. However, after the exponential phase, the metabolic state of cells may not be conducive to protein expression, as they may be under stressful conditions, triggering reactions that increase protease levels, which may reduce heterologous protein production [39]. Li et al. [40] used whole cell catalysis production experiments to select the OD600 = 0.8–1.2 range for induction; as shown in Figure 3-D. When the induction temperature was 30°C, the trans-2-decenoic acid yield was the highest, 1.226 ± 0.022 g/L, so we chose 30°C as optimal.
3.1.5. Effects of different solvents dissolving substrates on production of trans-2-decenoic acid
Decanoic acid crystals are white with a special odor. They are insoluble in water and are soluble in most organic solvents, meaning it is particularly important to choose a reasonable organic solvent to bring decanoic acid into the reaction system and allow it to fully react. As shown in Figure 4–E, among the reactions using five different solvents, the yield of trans-2-decenoic acid was highest when absolute ethanol was used, yielding 1.28 ± 0.03 g/L. This may be attributed to the unique characteristics of absolute ethanol; the bactericidal properties reduce the risk of bacterial contamination and the good compatibility of the decanoic acid substrate with absolute ethanol ensures that the substrate can carry out the transformation and fermentation reactions normally; therefore, we chose absolute ethanol as the solvent to dissolve the substrate.
3.1.6. Effect of feeding different concentrations of substrate on production of trans-2-decenoic acid
To ensure the continuous growth of trans-2-decenoic acid production, the product production and substrate consumption must maintain a high dynamic equilibrium; therefore, the substrate consumption rate requires special attention during the fermentation process. Continuity of the fermentation process was ensured via the fed substrate. As shown in Figure 4-F, the yield of trans-2-decenoic acid was the highest when the fed concentration was 0.2 g/L, yielding 1.902 ± 0.032 g/L, so we chose a fed concentration of 0.2 g/L as the bottom stream plus the concentration.
3.1.7. Effect of inducer concentration on trans-2-decenoic acid production.
The concentration of the inducer is an important factor that affects the expression of recombinant proteins from the inducible promoters. If the concentration is too low, the amount of inducer may be insufficient to bind to the repressor protein, reducing the expression level. If the concentration of the inducer is too high, the bacteria may be damaged. Growth is limited, and inclusion bodies are produced if the recombinant protein is expressed too quickly; therefore, it is particularly important to choose the appropriate concentration of the inducer. As shown in Figure 4-G, when the inducer concentration was 5 g/L, the yield of trans-2-decenoic acid was highest, 1.296 ± 0.019 g/L, so we chose the inducer concentration of 5 g/L.
3.1.8. Effects of adding different metal ions to culture medium on production of trans-2-decenoic acid.
Mn2+ is crucial in enhancing the activities of various enzymes involved in sugar metabolism and nucleotide processing. It also facilitates the adenylation of amino acids in antibiotics, a prerequisite for the production of many peptide antibiotics. Consequently, the content of Mn2+ significantly influences the population size of organisms and the yield of products. However, adding Mg2+ boosts the activity of essential enzymes such as hexose phosphorylase, citrate dehydrogenase, and carboxylase. This, in turn, affects processes such as matrix oxidation and protein synthesis. Additionally, magnesium ions are necessary for the proper functioning of most enzymes that act on nucleotides. Furthermore, Mn2+ enhances the tolerance of certain antibiotic-producing strains to the generated antibiotics. Based on the findings in Figure 4-H, the highest production rate of trans-2-decenoic acid was observed when MnCl2 was added to the reaction system, reaching 1.307 ± 0.014 g/L. Therefore, we decided to incorporate MnCl2 into the culture medium for further experiments.
3.2. Response Surface Testing
A BBD was used based on a single-factor experiment, with the flow rate, inducer concentration, and MnCl2 concentration as independent variables. The response value was the yield of trans-2-decenoic acid, and the results are presented in Table 2. Multiple regression analysis was performed on Table 2 using Design Expert 8.0 software to obtain the regression equation.
Y = +1.77 + 0.19A + 0.032B − 0.059C − 0.017AB + 0.028AC − 0.026BC − 0.083A2 − 0.090B2 + 0.066C2
Seventeen experiments were performed using three independent variables at three levels (1, 0, and +1). Table 2 lists the results of the 17 experiments. The results were analyzed using Design Expert software. An analysis of variance (ANOVA) was used to test the statistical significance of the quadratic regression equations (Table 3). The P-value of the regression model was <0.0001, indicating that the regression equation obtained using this model was extremely significant. The p-value for the missing FT was 0.2998, and the difference was not significant. The regression coefficient (R2) was used to determine the model fitting efficiency. R2 and the corrected quadratic model were 0.9963 and 0.9916, respectively, which were very close to 1. This shows that the model fits the data well, indicating that 99% of the variation in response can be described by this polynomial model.
This equation explained more than 99.63% of the experimental values. Each factor had a different effect on trans-2-decenoic acid production. The first one affects A (<0.0001) > C (<0.0001) > B (0.0005). The interaction effects among AB, AC, and BC were not statistically significant. The second-factor-affected items A2, B2, and C2 have an extremely significant impact (P < 0.001).
3.3. Response Surface Interaction
To further study the effects of the substrate feeding concentration, inducer concentration, and MnCl2 concentration on the production of trans-2-decenoic acid, the response surface and contours were drawn according to the regression equation, as shown in Figure 5. The steeper the curve trend, the stronger the interaction and the greater the impact on trans-2-decenoic acid production. The contour plot intuitively reflects the significance of the interactions between various factors. As any two variables increased, the production of trans-2-decenoic acid also increased. After the interaction between the two reached a peak, the surface area decreased. The interaction between the substrate-feeding concentration and the inducer concentration was the most significant, with the steepest curve and an elliptical contour.
3.4. Response Surface Results Optimization
After the response surface analysis, the optimal fermentation conditions were screened out as follows: substrate flow addition amount of 0.2 g/L, inducer concentration of 4.72 g/L, and MnCl2 concentration 0.15 mM. Fermentation verification was carried out in a 1 L fermentation tank under optimal conditions, and three sets of parallel experiments were conducted.
Through analyzing the regression model equation, we found the optimal process conditions for trans-2-decenoic acid were substrate flow addition amount 0.14 g/L, inducer concentration 5.63 g/L, and MnCl2 concentration 0.11 mM. Under these conditions, the yield of trans-2-decenoic acid was 1.9 g/L. The actual operation feasibility was improved, and the fermentation process was determined as follows: inoculum amount 1%, fermentation temperature 30°C, seed liquid culture time 20 h, substrate flow addition amount 0.15 g/L, inducer concentration 5.60 g/L, and MnCl2 concentration was 0.10 mM. Three repeated experiments were performed under these conditions, and the results are shown in Figure 6-B. The average yield of trans-2-decenoic acid after optimization was 1.982 ± 0.110 g/L, which is 0.082 g/L different from the theoretical value, indicating that the model is reliable and that the regression equation can be used for the fermentative production via biocatalytic synthesis of trans-2-decenoic acid from decanoic acid.
We also compared the production of trans-2-decenoic acid before and after fermentation optimization. Production using LB medium before optimization was 0.940 ± 0.104 g/L, and after optimization, the production was 1.982 ± 0.110 g/L. By optimizing trans, the production of -2-decenoic acid was increased by 1.04 g/L. Simultaneously, we monitored the OD600 during fermentation. As shown in Figure 6-A, the highest OD600 before optimization was 6.46, as shown in Figure 6-B. After optimization, the highest OD600 reached 8.46. Optimization promoted the growth of bacteria in the reaction system. However, as shown in Figure 6, the OD600 declined after 56 h. This may be because as the reaction proceeded, the bacteria continued to grow and metabolize, and the metabolic waste accumulated negatively impacted the bacteria. It also inhibited bacterial growth. After the bacteria enter a stable period, the external environment becomes increasingly unfavorable to their growth. The intracellular catabolic rate greatly exceeds the anabolic rate, the reproduction rate slows down, the morphology changes significantly, and even the bacteria spontaneously lyse, reducing the number of bacteria.
3.5. Comparison before and after optimization of SDS-PAGE protein expression of engineered E. coli.
A comparative analysis of the samples before and after the optimization of trans-2-decenoic acid fermentation was performed. The samples were analyzed using sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Lane M represents the protein marker, and lanes 1, 2, and 3 are three sets of parallel bands in Figure 7-A. Figure 7-B shows a protein gel image of the fermentation broth sample under optimal fermentation conditions; the protein size of the transporter Fad was 59 kDa, FadE was 45 kDa, and YdiI was 30 kDa. Subsequently, the two images were compared. The protein amounts of the three bands in Figure 7-A were significantly lower than those in Figure 7-B, which shows that the protein expression effect in the reaction system after fermentation optimization was better, making the reaction system more efficient.
4. Discussion
Based on the E. coli BL21(DE3)-△fadB-△fadJ-△fadR/pET28a-sumo-CtydiI/pCDFDuet-1-MaMACS-PpfadE) designed by the previous research group, we used the biological fermentation method for the first time to catalyze the synthesis of trans-2-decenoic acid from decanoic acid. The reaction system was put into a fermentation tank for amplification. Based on the aerobic characteristics of E. coli, better ventilation and stirring of the fermentation tank were used to improve the reaction system. Efficient reactions were performed. In the experiment, the purpose of controlling the substrate concentration at 0.5 g/L and 1% inoculum volume was to enable the bacteria to grow better in the system and maintain a relatively strong metabolic activity for better biocatalytic reactions.
For reaction optimization, we conducted eight single-factor experiments: seed culture temperature, seed liquid culture time, inoculum amount, induction temperature, solvent type, feed concentration, inducer concentration, and metal compound type. Based on the influence of different factors on the production of trans-2-decenoic acid, the three factors with the greatest influence on trans-2-decenoic acid were determined as the response variables: feed, inducer, and MnCl2 concentrations. The BBD was used to optimize the fermentation conditions. These three factors have different effects on trans-2-decenoic acid. According to the order of significance, they can be arranged as fed concentration > inducer concentration > MnCl2; after optimizing each factor, the predicted value of the optimal fermentation conditions was 1.9 g/L, the fed concentration was 0.14 g/L, the inducer concentration was 5.63 g/L, and the MnCl2 concentration was 0.11 mM. We conducted three repeated experiments based on the optimized optimal conditions and found that the yield of trans-2-decenoic acid was 1.982 ± 0.110 g/L, which was 1.042 g/L higher than the production of LB medium before optimization. With a substrate concentration of 0.5 g/L, the yield was 0.479 g/L higher than the average yield achieved by Li et al. [40]It increased by 1.503 ± 0.110 g/L. Evidently, the fermentation condition optimization plan we carried out had a significant effect.
During the catalytic reaction of engineered E. coli, the fadD gene encodes acyl-CoA synthase, which has broad substrate specificity and catalyzes the conversion of free fatty acids into highly metabolically active coenzyme A thioesters. Through the action of CoA synthase, free fatty acids form fatty acyl-CoAs with high metabolic activity [41,42,43]. Subsequently, FadE catalyzes the oxidation of acyl-CoA to trans-2-enoyl CoA [44] and finally removes CoA to form α, β-unsaturated fatty acids through the catalysis of acyl-CoA thioesterase YdiI. Therefore, the three enzymes FadD, FadE, and YdiI play vital roles in this reaction process. While optimizing the fermentation conditions, we explored the changes in the expression levels of the three enzymes before and after fermentation. The bands in the optimized samples were more apparent than those in the unoptimized samples, further confirming the necessity of optimizing the fermentation conditions.
One problem that arose during fermentation was that the amount of trans-2-decenoic acid did not increase regularly or stably. In the early stages of the reaction, the level of trans-2-decenoic acid gradually grew. As the reaction continued, the amounts of the products and substrates gradually decreased or even disappeared, as confirmed through many repeated experiments. We speculate that this is due to the continuous growth and metabolism of the bacteria during the reaction process, which required a large amount of nutrients, including carbon sources. Various substances, such as nitrogen sources and biotin, are gradually consumed or even exhausted as the bacterial cells grow, resulting in the culture medium being unable to provide sufficient nutrients to the bacterial cells. Therefore, we suspected that bacterial cells were preferentially consumed at this stage. The substrates and products were used as carbon sources to ensure growth and metabolic activity. For this phenomenon, we need to formulate a more accurate and efficient plan for the next step by adding nutrients and energy during the reaction process to maintain a stable biological reaction while also considering that long-term fermentation leads to bacterial aging. The aging of bacterial cells decreases enzyme production owing to a decline in metabolic capacity. Therefore, we believe that after the reaction has been carried out for a certain period, new culture media must be added to dilute the bacterial cells so that more new bacterial cells can grow to ensure reaction success.
Due to the various drawbacks of the chemical synthesis of α, β-unsaturated fatty acids currently on the market, attention to their biosynthesis has gradually increased and has developed into an alternative to traditional chemical methods. There have been few previous studies on trans-2-decenoic acid, strongly limiting the current research on this substance. This limitation is reflected in the instability during the fermentation process and the low conversion rate and yield; how to produce it efficiently and even industrially mass produce it remains difficult. At present, the synthesis of trans-2-decenoic acid relies mainly on chemical synthesis; however, this method involves a long process, has low reaction controllability, and can easily cause environmental pollution. Therefore, exploring biosynthetic methods with the advantages of high safety, strong specificity, and low environmental pollution to catalyze the synthesis of trans-2-decenoic acid has become a research hotspot. This study improved the production of trans-2-decenoic acid by improving the metabolic pathway of trans-2-decenoic acid modified by a previous research group and by improving the limiting factors in the whole-cell catalysis process. Compared with whole-cell catalysis, the production indeed improved, which provides a basis for the future synthesis of trans-2-decenoic acid catalyzed by growing cells.
5. Conclusions
This study is based on engineered E. coli constructed in the laboratory in the early stage and uses decanoic acid as the substrate for biological fermentation to catalyze the synthesis of trans-2-decenoic acid from decanoic acid. First, the peak times of decanoic acid and trans-2-decenoic acid production were determined using gas chromatography. Eight sets of single-factor experiments were conducted, and the three groups that had the greatest impact on the yield of trans-2-decenoic acid were screened. A BBD and RSA were performed on these factors: feed concentration, inducer concentration, and MnCl2 concentration, for further optimization. The construction of this model can better predict and explain the effects of the three different factors on the fermentation of trans-2-decenoic acid. The final optimization results show that the optimal process conditions for trans-2-decenoic acid are substrate flow addition amount 0.14 g/L, inducer concentration 5.63 g/L, and MnCl2 concentration 0.11 mM (1 L under these conditions). The fermentation tank conditions were verified by three parallel experiments, and the average of the final results of the three parallel experiments was 1.98 g/L. Compared to the LB medium before optimization, the output increased by 1.04 g/L; compared with trans-, the output of -2-decenoic acid increased by 1.501 g/L. Finally, we compared the protein expression levels of the samples before and after optimization; the enzyme expression in the optimized samples was significantly higher than that in the non-optimized samples. This study improved the production of trans-2-decenoic acid and provided an important reference for future generations to study biosynthetic fatty acid technology.
Author Contributions
Conceptualization, P.L. and J.S.; methodology, S.N.; software, S.N., K.L. and B.L; validation, P.L. and J.S.; formal analysis, S.N and K.L.; investigation, S.N.,P.L. and J.S.; writing—original draft preparation, S.N.; writing—review and editing, S.N,P.L. and J.S.; supervision, P.L. and J.S.; project administration, P.L. and J.S.; funding acquisition, P.L. and J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Science Foundation of Shandong Province (grant nos. ZR2023MC089) and the Focuson Research and Development Plan in Shandong Province (grant no.2023CXGC010714).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hirao, T. Synthetic strategy: palladium-catalyzed alpha,beta-dehydrogenation of carbonyl compounds. J. Org. Chem. 2019, 84(4), 1687-1692. [CrossRef]
- Perrin, C.L.; Chang, K.L. The complete mechanism of an aldol condensation. J. Org. Chem. 2016, 81(13), 5631–5635. [CrossRef]
- Farfan, P.; Gomez, S.; Restrepo, A. Dissection of the mechanism of the Wittig reaction. J. Org. Chem. 2019, 84(22), 14644–14658. [CrossRef]
- Bonnard, I.; Rolland, M.; Salmon, J. M.; Debiton, E; Barthomeuf, C.; Banaigs, B. Total structure and inhibition of tumor cell proliferation of laxaphycins. J. Med. Chem. 2017, 50(6), 1266-1279. [CrossRef]
- Wang, L.H.; He, Y.; Gao, Y.; Wu, J.E.; Dong, Y.H,; He, C.; Wang, S.X.; Weng, L.X.; Xu, J.L.; Tay, L.; Fang, R.X.; Zhang, L.H. A bacterial cell-cell communication signal with cross-kingdom structural analogues. Mol. Microbiol. 2004, 51(3): 903-912. [CrossRef]
- Li, Y.; Wang, J.; Wang, F.; Wang, L.; Wang, L.; Xu, Z.; Yuan, H.; Yang, X.; Li, P.; Su, J.; & Wang, R. Production of 10-Hydroxy-2-Decenoic Acid from Decanoic Acid via Whole-Cell Catalysis in Engineered Escherichia coli. ChemSusChem. 2022, 15(9): e202102152. [CrossRef]
- Nascimento, A.P.; Moraes, L.R.; Ferreira, N.U.; Moreno, G.; Uahib, F.M.; Barizon, E.A.; Berretta, A.A. The lyophilization process maintains the chemical and biological characteristics of royal jelly. Evid Based Complement Alternat Med. 2015: 825068. [CrossRef]
- Townsend, G.F.; Morgan, J.F.; Hazlett, B. Activity of 10-hydroxydecenoic acid from royal jelly against experimental leukemia and ascitic tumours. Nature. 1959, 183(4670), 1270–1271. [CrossRef]
- Pavel, Al, C.I.; L.; Mărghitaş, O. Bobiş, & MN Mădaş. Lucrari Stiint. Zootehnie Biotecnol. 2011. Biological activities of royal jelly -review, 44 (2).
- Koh, W.K.; Bartnik, A.C.; Wise, F.W.; & Murray, C.B. Synthesis of monodisperse pbse nanorods: a case for oriented attachment. J Am Chem Soc. 2010, 132(11):3909-13. [CrossRef]
- Sarkar, A.; Middya, T.R.;Jana, A.D. A qsar study of radical scavenging antioxidant activity of a series of flavonoids using dft based quantum chemical descriptors - the importance of group frontier electron density. J Mol Model. 2012, (6):2621-31. [CrossRef]
- Wei, D.X.; Dao, J.W.; Chen, G.Q. A micro-ark for cells: highly open porous polyhydroxyalkanoate microspheres as injectable scaffolds for tissue regeneration. Adv Mater. 2018, (31):e1802273. [CrossRef]
- Marques, C.N., Davies, D.G., & Sauer, K. Control of biofilms with the fatty acid signaling molecule cis-2-decenoic acid. Pharmaceuticals. 2015, 8(4), 816-835. [CrossRef]
- Cai, P.J.; Xiao, X.; He, Y.R.; Li, W.W.; Yu, L.; Yu, H.Q. Disintegration of aerobic granules induced by trans-2-decenoic acid. Bioresour. Technol. 2013, 128(Complete), 823-826. [CrossRef]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Qin, J.; Siewers, V.; Nielsen, J. Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat. Commun. 2016, 7(1), 11709. [CrossRef]
- Liu, X.; Yu, H.; Jiang, X.; Ai, G.; Yu, B.; & Zhu, K. Biosynthesis of butenoic acid through fatty acid biosynthesis pathway in Escherichia coli. Appl Microbiol Biotechnol. 2015, 99(4), 1795-1804. [CrossRef]
- Kim, S.; Cheong, S.; & Gonzalez, R. Engineering Escherichia coli for the synthesis of short- and medium-chain α,β-unsaturated carboxylic acids. Metab Eng. 2016, 36, 90-98. [CrossRef]
- Cronan, J. E., Jr, & Rock, C. O. Biosynthesis of Membrane Lipids. EcoSal Plus. 2008, 3(1). [CrossRef]
- Jawed, K.; Mattam, A.J.; Fatma, Z.; Wajid, S.; Abdin, M.Z.; Yazdani, S.S. Engineered production of short chain fatty acid in Escherichia coli using fatty acid synthesis pathway. Plos One. 2016, 11(7), e0160035. [CrossRef]
- Iram, S.H.; Cronan, J.E. The beta-oxidation systems of Escherichia coli and Salmonella enterica are not functionally equivalent. J Bacteriol. 2006 Jan;188(2):599-608. [CrossRef]
- Lee, T.S.; Krupa, R.A.; Zhang, F.; Hajimorad, M.; Holtz, W. J.; Prasad, N.; Lee, S. K.; Keasling, J.D. Bglbrick vectors and datasheets: a synthetic biology platform for gene expression. J Biol Eng. 2011, 5(1), 12. [CrossRef]
- Morsczeck, C.; Berger, S.; Plum, G. The macrophage-induced gene (mig) of mycobacterium avium encodes a medium-chain acyl-coenzyme a synthetase - sciencedirect. Biochim Biophys Acta. 2001, 1521(1), 59-65. [CrossRef]
- Guzik, M.W.; Narancic, T.; Ilic-Tomic, T.; Vojnovic, S.; Kenny, S.T.; Casey, W.T.; Duane, G.F.; Casey, E.; Woods, T.; Babu, R.P.; Nikodinovic-Runic, J.; O'Connor, K.E. Identification and characterization of an acyl-CoA dehydrogenase from Pseudomonas putida KT2440 that shows preference towards medium to long chain length fatty acids. Microbiology. 2014, 160(8):1760-1771. [CrossRef]
- Campbell, J.W.; Cronan, J.E. The enigmatic Escherichia coli fade gene is yafh. J Bacteriol. 2002. 184(13):3759-64. [CrossRef]
- Poirier, Y.; Antonenkov, V.D.; Glumoff, T.; Hiltunen, J.K. Peroxisomal beta-oxidation--a metabolic pathway with multiple functions. Biochim Biophys Acta. 2006. 1763(12), 1413-26. [CrossRef]
- Yan, Q.; Simmons, T.R.; Cordell, W.T.; Hernández Lozada, N.J.; Breckner, C.J.; Chen, X., Jindra, M.A.; Pfleger, B.F. Metabolic engineering of β-oxidation to leverage thioesterases for production of 2-heptanone, 2-nonanone, and 2-undecanone. Metab Eng. 2020. 61:335-343. [CrossRef]
- Tao, A.; Feng, X., Sheng, Y., et al. Optimization of the Artemisia polysaccharide fermentation process by Aspergillus niger. Front Nutr. 2022, 9, 842766. [CrossRef]
- Kuo, C.H.; Hsiao, F.W.; Chen, J.H.; Hsieh, C.W.; Liu, Y.C.; Shieh, C. J. Kinetic aspects of ultrasound-accelerated lipase catalyzed acetylation and optimal synthesis of 4′-acetoxyresveratrol. Ultrason Sonochem. 2013, 20(1):546-52. [CrossRef]
- Gu, F.; Xu, F.; Tan, L.; Wu, H.; Chu, Z.; Wang, Q. Optimization of enzymatic process for vanillin extraction using response surface methodology. Molecules. 2012, 25;17(8):8753-61. [CrossRef]
- Wang, Y.H.; Xuan, Z.H.; Tian, S.; Du, G.H. Echinacoside Protects against 6-Hydroxydopamine-Induced Mitochondrial Dysfunction and Inflammatory Responses in PC12 Cells via Reducing ROS Production. Evid Based Complement Alternat Med. 2015:1-9. [CrossRef]
- Chen, W.; Lin, H.R.; Wei, C.M.; Luo, X.H.; Sun, M.L.; Yang, Z.Z.; Chen, X.Y.; Wang, H.B. Echinacoside, a phenylethanoid glycoside from Cistanche deserticola, extends lifespan of Caenorhabditis elegans and protects from Aβ-induced toxicity." Biogerontology. 2018, 19(1): 47-65. [CrossRef]
- Peiran, L.; Ying, L.; Mingzhuo, Z.; Ye, Y.; Xiuming, C. The development of a Panax notoginseng medicinal liquor processing technology using the response surface method and a study of its antioxidant activity and its effects on mouse melanoma B16 cells." Food Funct. 2017, 8. [CrossRef]
- Oh, H.J., Shin, K.C., Oh, D.K. Production of 10-hydroxy-12,15(z,z)-octadecadienoic acid from α-linolenic acid by permeabilized cells of recombinant escherichia coli expressing the oleate hydratase gene of stenotrophomonas maltophilia. Biotechnol Lett. 2013, Sep;35(9):1487-93. [CrossRef]
- Jung, D.H.; Jung, J.H.; Seo, D.H.; Ha, S.J.; Kweon, D.K.; Park, C.S. One-pot bioconversion of sucrose to trehalose using enzymatic sequential reactions in combined cross-linked enzyme aggregates. Bioresour Technol. 2013, 130:801-4. [CrossRef]
- Rajendran V, Simab K, Aran I. Non-ionic surfactant integrated extraction of exopolysaccharides from engineered Synechocystis sp. PCC 6803 under fed-batch mode facilitates the sugar-rich syrup production for ethanol fermentation. Algal Research, 2022,66. [CrossRef]
- Mackenzie, S.; Zachary, W.; Lauren, W. Manganese Homeostasis in Bacteria: Interaction of the Small Protein MntS and Manganese Exporter MntP in E. coli. The FASEB Journal, 2022,36. [CrossRef]
- Letuta, U.G.; Binder, A. S; Tikhonova, T. A. (2020). Effect of magnesium isotopes on antibiotic sensitivity of e. coli. Microbiology, 89(3). [CrossRef]
- Liszkowska, W.; Berlowska, J. Yeast fermentation at low temperatures: adaptation to changing environmental conditions and formation of volatile compounds. Molecules. 2021 Feb 16;26(4):1035. [CrossRef]
- Sónia Carneiro, Eugénio C. Ferreira, Rocha I. Metabolic responses to recombinant bioprocesses in Escherichia coli[J].Journal of Biotechnology, 2013, 164(3):396-408. [CrossRef]
- Wang, L.; Wang, L.; Wang, R.; Wang, Z.; Wang, J.; Yuan, H.; et al. Efficient biosynthesis of 10-hydroxy-2-decenoic acid using a nad(p)h regeneration p450 system and whole-cell catalytic biosynthesis. ACS Omega. 2022, 19;7(21):17774-17783. [CrossRef]
- Ford, T.J.; Way, J.C. Enhancement of e. coli acyl-coa synthetase fadd activity on medium chain fatty acids. PeerJ, 3, 2015, e1040. [CrossRef]
- Black, P.N.; Dirusso, C.C.; Metzger, A.K.; Heimert, T.L. Cloning, sequencing, and expression of the fadd gene of escherichia coli encoding acyl coenzyme a synthetase. J Biol Chem. 1992 Dec 15;267(35):25513-20.
- Kameda, K.; Nunn, W.D. Purification and characterization of acyl coenzyme a synthetase from escherichia coli. J Biol Chem. 1981 Jun 10;256(11):5702-7.
- Campbell, J.W.; Cronan, J. Jr. The enigmatic Escherichia coli fadE gene is yafH. J Bacteriol. 2002 Jul;184(13):3759-64. [CrossRef]
Figure 1.
Source and function of trans-2-decenoic acid.
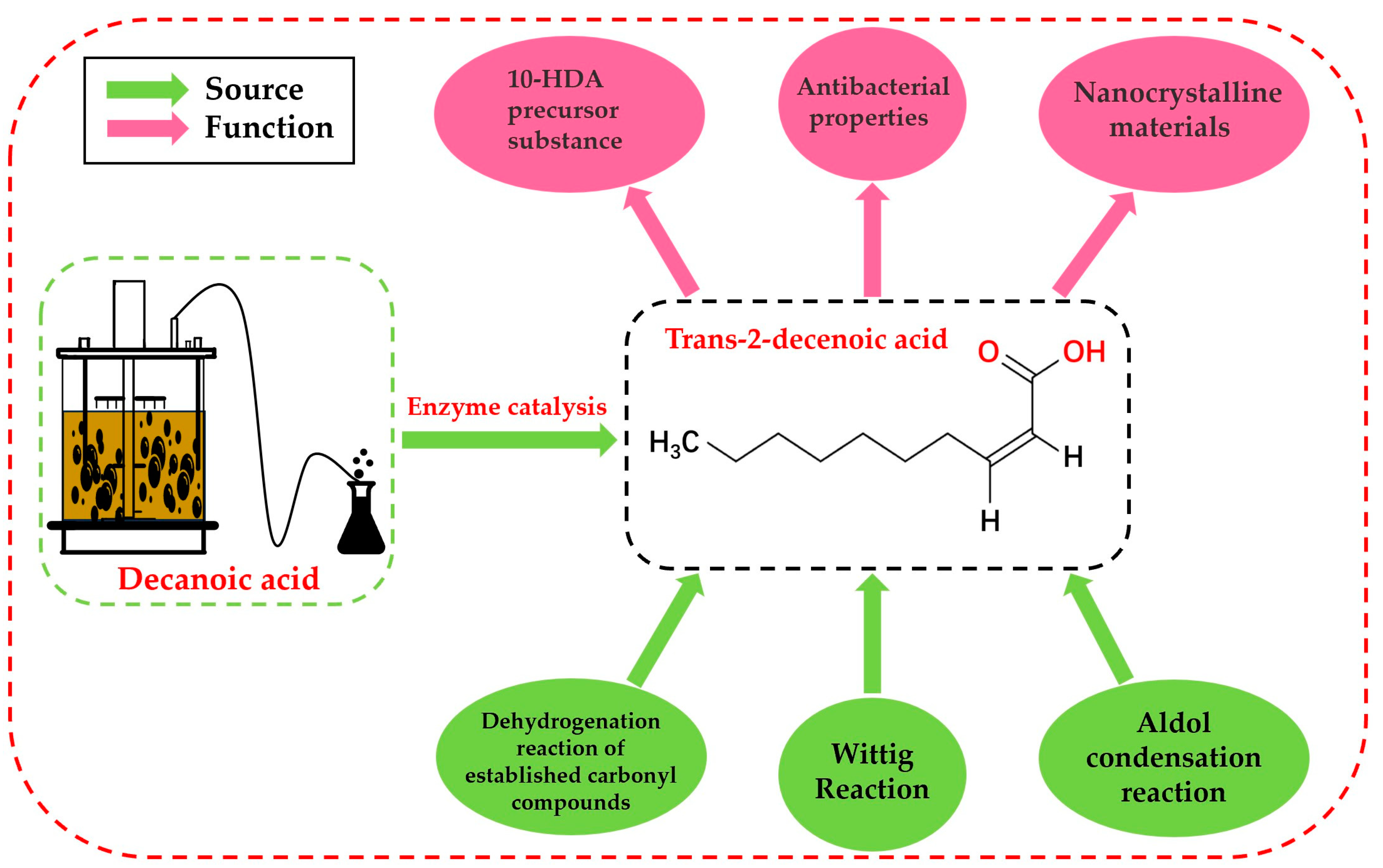
Figure 2.
Synthetic pathway of trans-2-decenoic acid. Note: FadD: acyl-CoA synthetase; FadE: acyl-CoA dehydrogenase; YdiI: acyl-CoA thioesterase; FadR: enoyl-CoA hydratase homologous protein gene; FadJ: enoyl-CoA hydratase homologous protein gene; FadB: enyl-CoA hydratase homologous protein gene.
Figure 2.
Synthetic pathway of trans-2-decenoic acid. Note: FadD: acyl-CoA synthetase; FadE: acyl-CoA dehydrogenase; YdiI: acyl-CoA thioesterase; FadR: enoyl-CoA hydratase homologous protein gene; FadJ: enoyl-CoA hydratase homologous protein gene; FadB: enyl-CoA hydratase homologous protein gene.
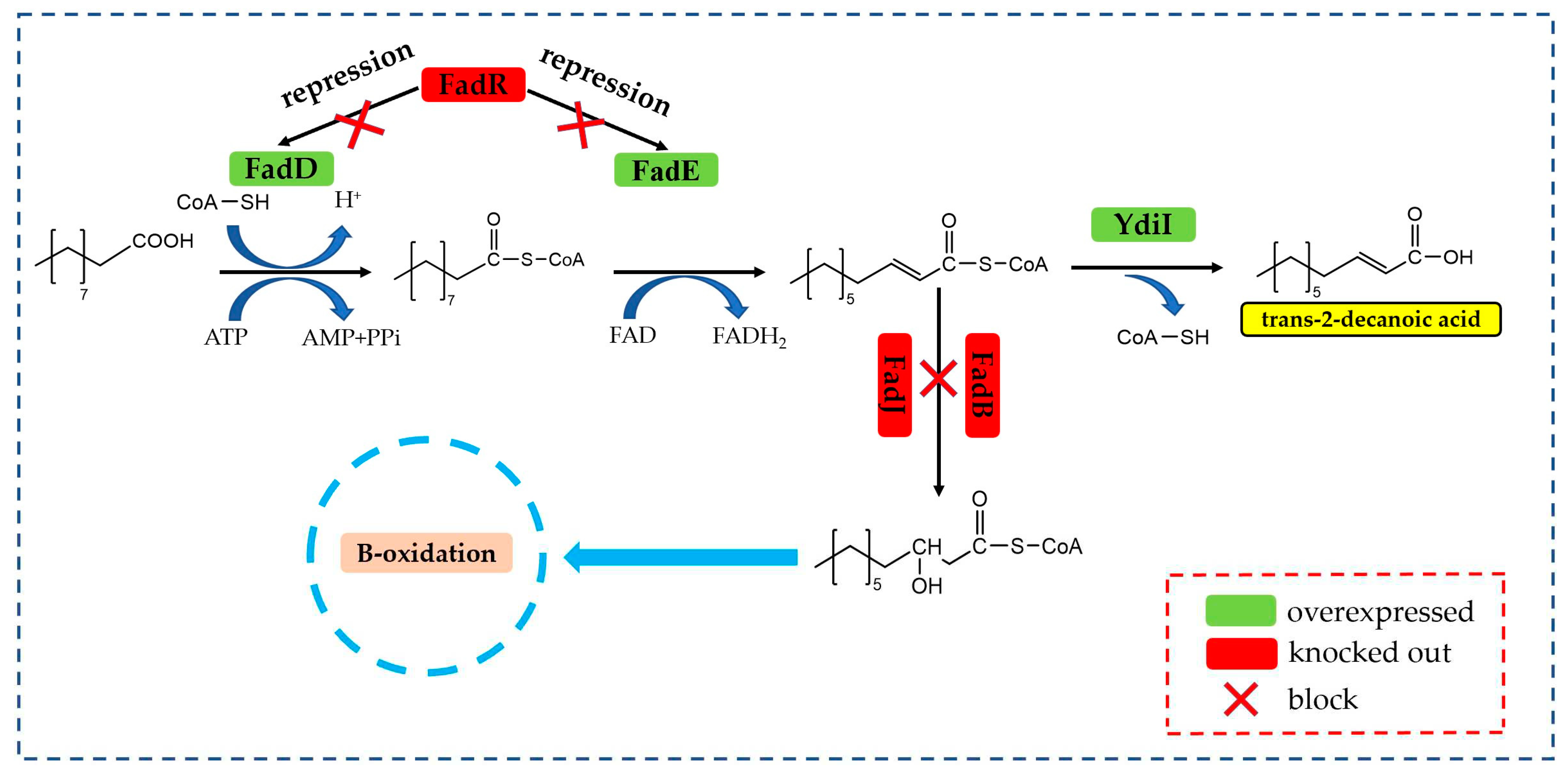
Figure 3.
Effect of different factors on trans-2-decenoic acid production. Fermentation for 72 h. (A) Temperature; (B) seed culture time; (C) inoculum amount; (D) induction temperature.
Figure 3.
Effect of different factors on trans-2-decenoic acid production. Fermentation for 72 h. (A) Temperature; (B) seed culture time; (C) inoculum amount; (D) induction temperature.
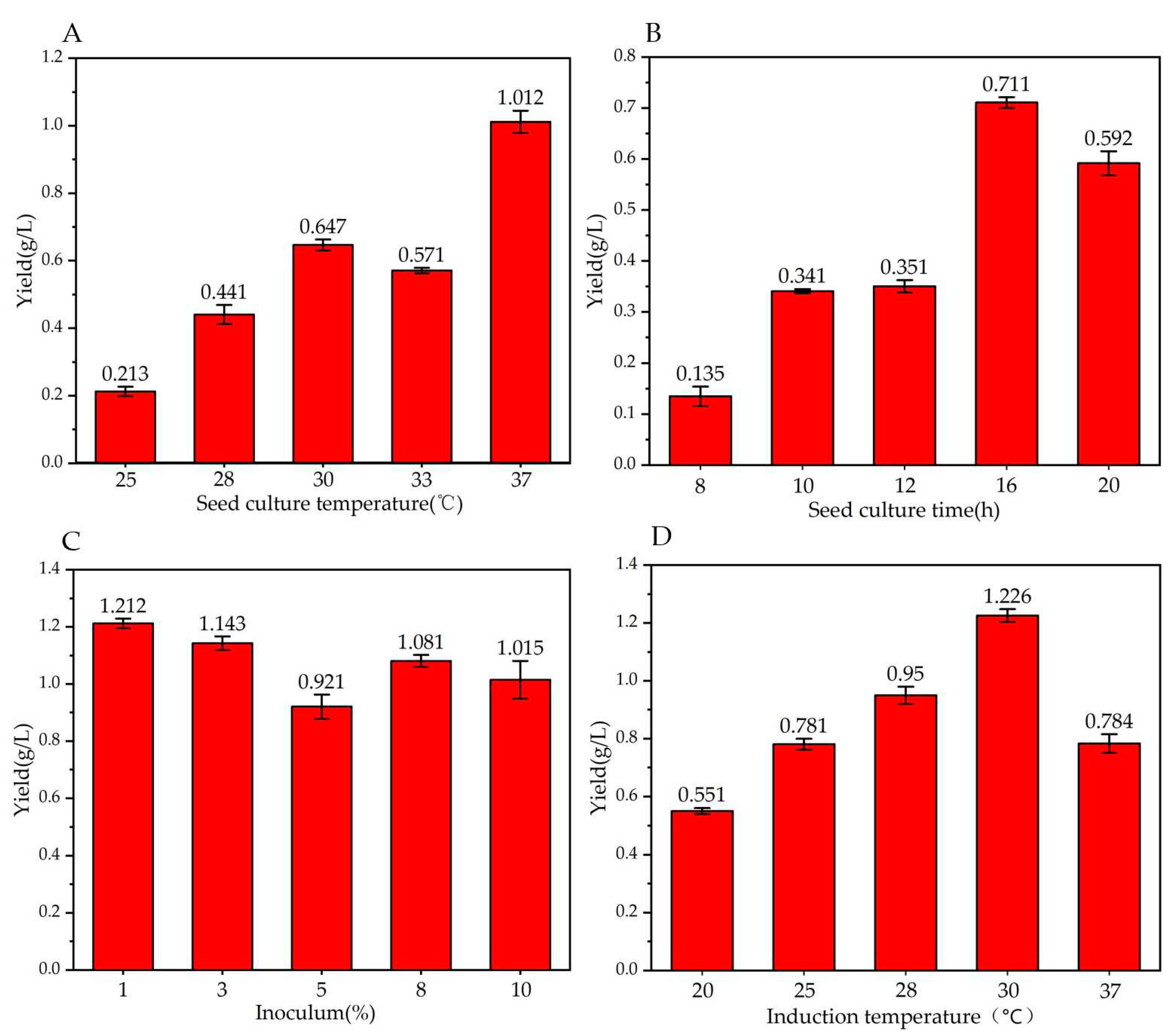
Figure 4.
Effect of different factors on trans-2-decenoic acid production. Fermentation for 72 h. (E) solvent; (F) feed volume; (G) inducer concentration; (H) metal compound.
Figure 4.
Effect of different factors on trans-2-decenoic acid production. Fermentation for 72 h. (E) solvent; (F) feed volume; (G) inducer concentration; (H) metal compound.
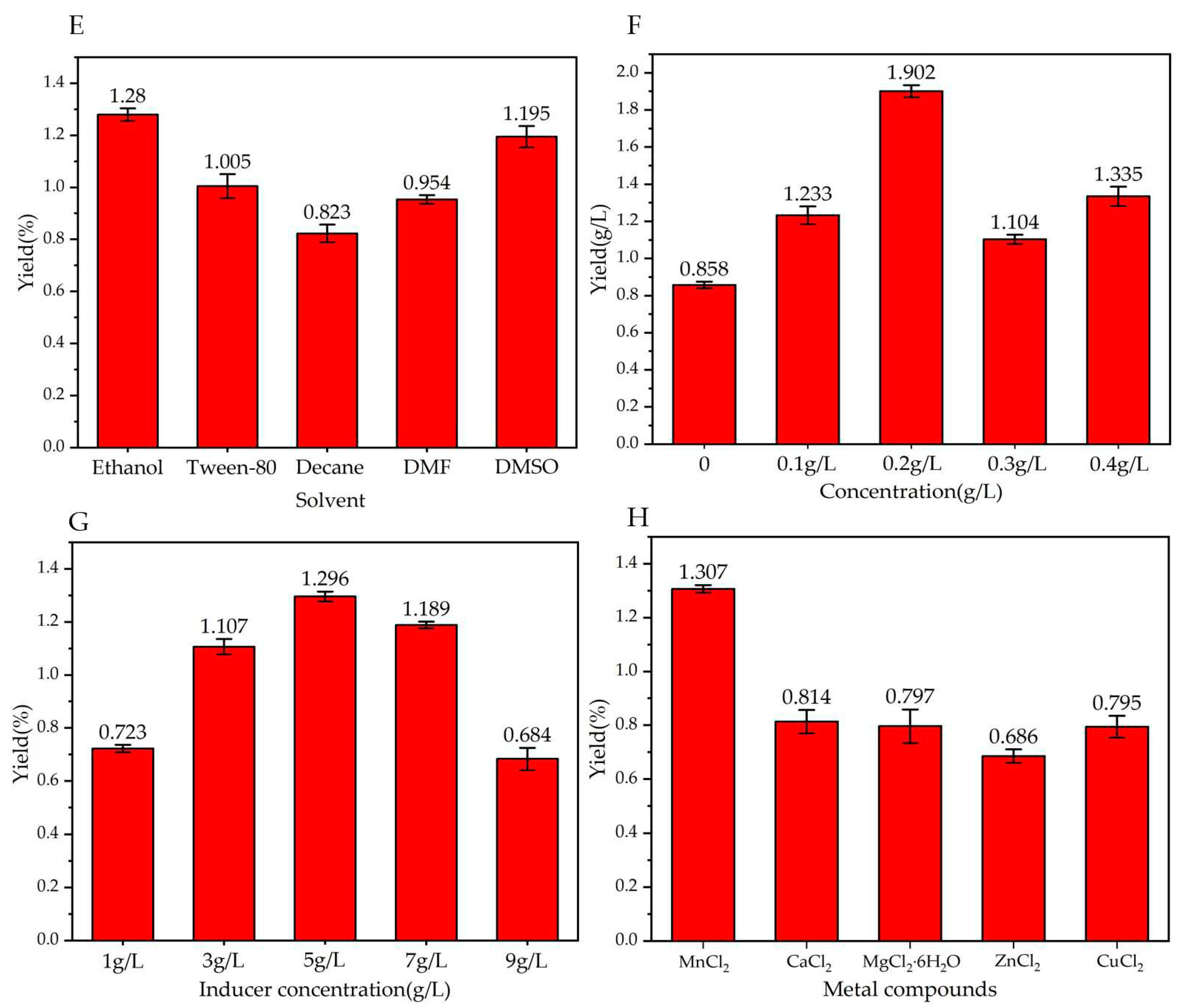
Figure 5.
Response surface and contour plots for trans-2-decenoic acid production. (A,a) Effect of flow rate and inducer concentration. (B,b) Effect of flow rate and MnCl2 concentration. (C,c) Effect of inducer concentration and MnCl2.
Figure 5.
Response surface and contour plots for trans-2-decenoic acid production. (A,a) Effect of flow rate and inducer concentration. (B,b) Effect of flow rate and MnCl2 concentration. (C,c) Effect of inducer concentration and MnCl2.
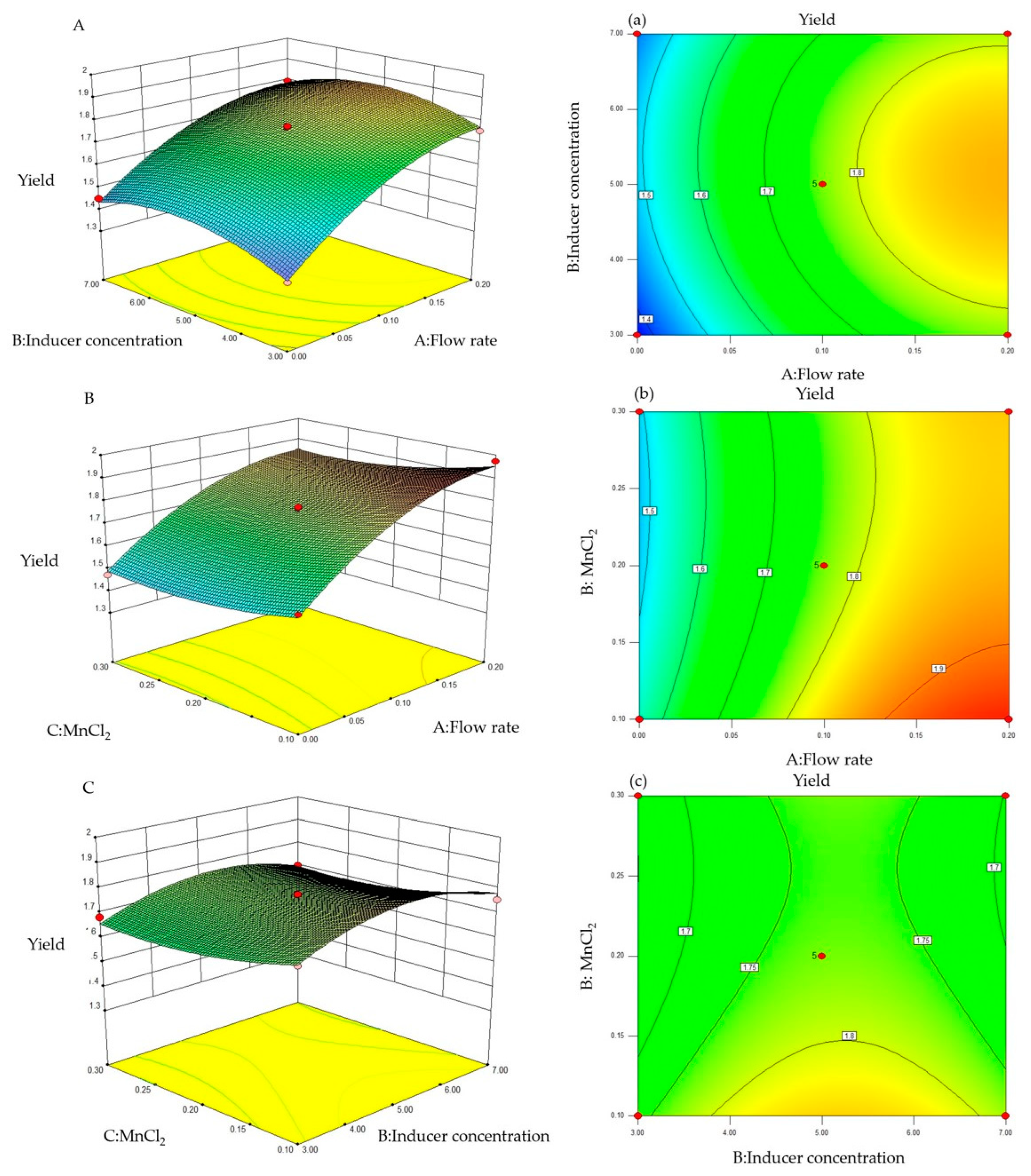
Figure 6.
Comparison of trans-2-decenoic acid production and OD600 before and after optimization. (A) Yield and OD600 before optimization (B) Yield and OD600 after optimization.
Figure 6.
Comparison of trans-2-decenoic acid production and OD600 before and after optimization. (A) Yield and OD600 before optimization (B) Yield and OD600 after optimization.
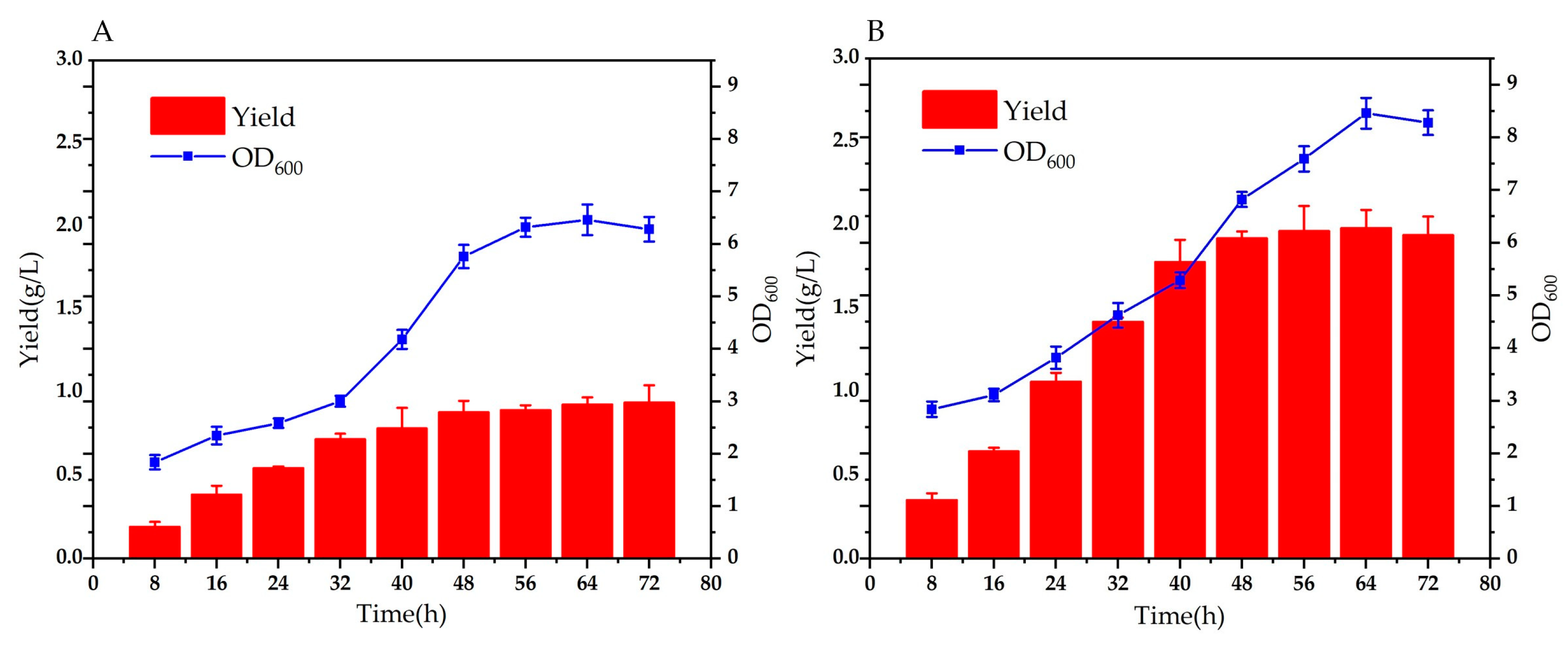
Figure 7.
Comparison of protein expression in samples before and after optimization. (A) Protein expression before optimization (B) Protein expression after optimization.
Figure 7.
Comparison of protein expression in samples before and after optimization. (A) Protein expression before optimization (B) Protein expression after optimization.
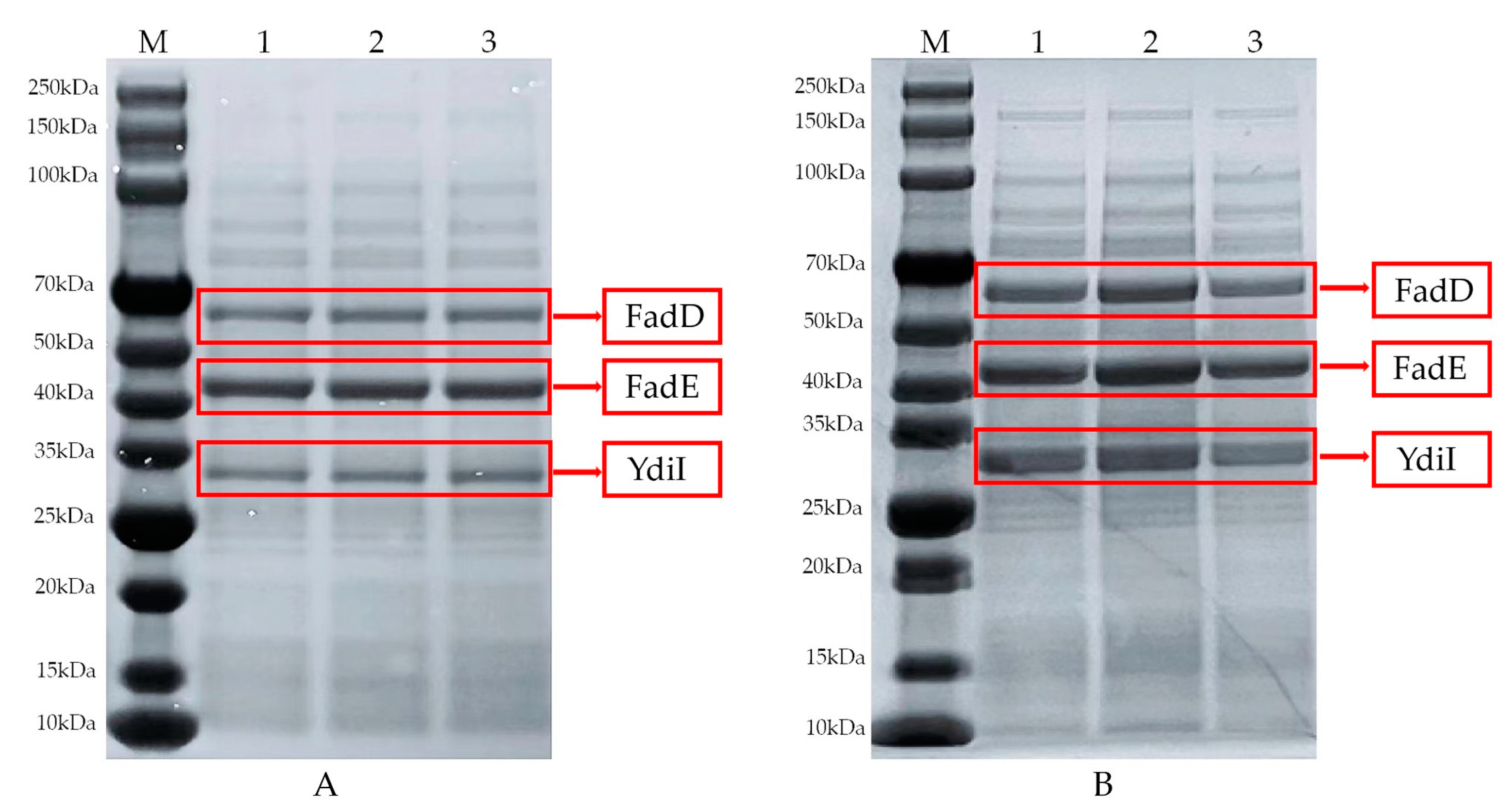

Table 1.
Coded and actual values of factors in BBD.
| Factor | Level | ||
|---|---|---|---|
| -1 | 0 | 1 | |
| A flow rate | 0 | 0.1 | 0.2 |
| B. Inducer concentration | 3 | 5 | 7 |
| C MnCl2 | 0.1 | 0.2 | 0.3 |
Table 2.
Response surface test design and results.
| Test number | Flow rate (g/L) | Inducer concentration (g/L) | MnCl2 (mM) | Yield (g/L) |
|---|---|---|---|---|
| 1 | 0.20 | 5.00 | 0.30 | 1.841 |
| 2 | 0.10 | 3.00 | 0.10 | 1.733 |
| 3 | 0.10 | 5.00 | 0.20 | 1.774 |
| 4 | 0.20 | 7.00 | 0.20 | 1.794 |
| 5 | 0.00 | 3.00 | 0.20 | 1.360 |
| 6 | 0.20 | 3.00 | 0.20 | 1.771 |
| 7 | 0.10 | 5.00 | 0.20 | 1.769 |
| 8 | 0.10 | 5.00 | 0.20 | 1.776 |
| 9 | 0.10 | 7.00 | 0.30 | 1.702 |
| 10 | 0.00 | 5.00 | 0.10 | 1.664 |
| 11 | 0.10 | 5.00 | 0.20 | 1.745 |
| 12 | 0.00 | 5.00 | 0.30 | 1.472 |
| 13 | 0.10 | 3.00 | 0.30 | 1.684 |
| 14 | 0.00 | 7.00 | 0.20 | 1.450 |
| 15 | 0.20 | 5.00 | 0.10 | 1.972 |
| 16 | 0.10 | 7.00 | 0.10 | 1.854 |
| 17 | 0.10 | 5.00 | 0.20 | 1.769 |
Table 3.
Response surface regression model analysis of variance.
| Source | Sum of Squares | DF | Mean Square | F-value | P-value | Significance |
|---|---|---|---|---|---|---|
| Model | 0.40 | 9 | 0.044 | 210.02 | <0.0001 | *** |
| A | 0.27 | 1 | 0.27 | 1304.41 | <0.0001 | *** |
| B | 7.938E-003 | 1 | 7.938E-003 | 37.72 | 0.0005 | *** |
| C | 0.028 | 1 | 0.028 | 133.44 | <0.0001 | *** |
| AB | 1.122E-003 | 1 | 1.122E-003 | 5.33 | 0.0543 | |
| AC | 3.080E-003 | 1 | 3.080E-003 | 14.64 | 0.0065 | |
| BC | 2.652E-003 | 1 | 2.652E-003 | 12.60 | 0.0093 | |
| A2 | 0.029 | 1 | 0.029 | 139.40 | <0.0001 | *** |
| B2 | 0.034 | 1 | 0.034 | 161.95 | <0.0001 | *** |
| C2 | 0.018 | 1 | 0.018 | 87.21 | <0.0001 | *** |
| Residual | 1.473E-003 | 7 | 2.105E-004 | |||
| Lack of Fit | 8.305E-004 | 3 | 2.768E-004 | 1.72 | 0.2998 | |
| Pure Error | 6.428E-004 | 4 | 1.607E-004 | |||
| Cor Total | 0.40 | 16 | ||||
| R2 = 0.9963 | R2adj = 0.9916 | R2pre = 0.9642 | ||||
| Adeq Precision | 54.485 |
Values of “Prob. > F” values less than 0.001, 0.01, and 0.05 indicate that model terms were extremely significant (***), highly significant (**), and significant (*), respectively.
Table 4.
Optimal fermentation conditions.
| Flow rate (g/L) | Inducer concentration (g/L) | MnCl2 (mM) | Yield (g/L) |
|---|---|---|---|
| 0.14 | 5.63 | 0.11 | 1.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



