Submitted:
22 October 2023
Posted:
23 October 2023
You are already at the latest version
Abstract
Ferroptosis is a recent form of non-apoptotic cell death, which occurs due to imbalance of iron homeostasis (iron overload). Oxidative stress due to the impairment of the antioxidant system is a major pathophysiology during ferroptosis, which eventually results in lipid peroxidation. The cellular and physiological biomarker of ferroptotic cell death includes major alteration in the glutathione peroxidase 4 (GPX4) antioxidant system and lipid peroxidation induced loss of plasma membrane integrity. This review elaborates the mechanism of oxidative stress which triggers ferroptosis.
Keywords:
Oxidative stress
; Ferroptosis
; Iron
; Cell death
; Lipid peroxidation
Introduction
Programmed Cell Death (PCD) can be described as sequential events that lead to the controlled and organized destruction of the cell (Lockshin and Zakeri, 2004). PCD is required for maintaining the homoeostasis and normal development which broadly includes autophagy, apoptosis, necrosis, necroptosis (Liu et al., 2013; Dixon et al., 2012; Fuchs and Steller, 2011; Degterevet al., 2005; Brennan et al., 2000). Ferroptosis is established as a non-apoptotic pathway, which is triggered by imbalance in iron homeostasis and eventually by iron overload in the cells (Dixon et al., 2012). The oxidative stress is pathophysiologically controlled by molecules, which are the products of oxidative metabolism and provoke oxidative injury, are collectively known as Reactive Oxygen Species (ROS). Ferroptotic cell death is induced by oxidative stress and triggered by the accumulation of toxic Lipid-Reactive Oxygen Species (L-ROS) by inhibiting or inactivating glutathione biosynthesis or the glutathione-dependent antioxidant enzyme, glutathione peroxidase 4 (GPX4) (Dixon et al. 2012; Skouta et al., 2014; Cao and Dixon, 2016).
Cysteine uptake, along with the export of glutamate, is required for the biosynthesis of GPX4 which occurs by the antiport named System Xc-. Biochemically, it shows increase in iron- dependent ROS accumulation and oxidation of Poly Unsaturated Fatty Acids (PUFAs) in the plasma membrane and hence it alters iron homoeostasis and lipid peroxidation metabolism in cells (Chen et al., 2020). Accumulation of L-ROS causing oxidative stress eventually depletes the plasma membrane integrity. Regulatory genes of lipid and amino acid metabolism play crucial role in ferroptosis (Cao and Dixon, 2016). Ferroptosis causes neurological issues, ischemic-reperfusion, acute renal failure among others (Munshi and Bhattacharya 2022; Chen et al. 2020; Gao et al., 2015; Linkermann et al., 2014; Angeli et al., 2014; Skouta et al., 2014; Dixon et al., 2012).
Approximately 65% of the iron is bound to hemoglobin and 10% is the constituent of myoglobin, cytochrome and iron containing enzymes and at about 25% of iron is bound to ferritin and haemosiderin. The storage capacity of ferritin is 4500 atoms of iron / molecule. Transferrin is the other molecule that helps the iron to remain in bound state in plasma (2 atoms of iron/ molecule). Only little amount iron is remained in free state. This inner pool of iron should be maintained to prevent the potential health disturbances produced due to both iron deficiency and iron overload. Normal iron homeostasis is maintained by the balance between absorbed dietary iron and required iron of body (Valko, 2005). Iron changes its ionic states between +3 (ferric ion, oxidized state) and +2 (ferrous state, reduced state) (Chatterjee et al., 2014b). Iron plays an important role in maintaining physiological function. It is present in heme which is essential for haemoglobin formation and serves as coenzyme for several catalytic reactions. There are two states of iron in human body: a. Bound Fe2+; remains in form of ferritin such as haemoglobin and iron sulfur nanoclusters and b. Free Fe2+; remains in heme or non-iron sulfur nanoclusters. Free irons remain in both ferrous and ferric form. Excessive free ferrous iron causes harm to cell by generating hydroxyl radicals (OH°) via Fenton reaction (Chen et al., 2020) and then induces oxidative stress which eventually cause lipid peroxidation.
Fe2+ mediated toxicity may be aggravated by increased oxidative stress and DNA damage. Interestingly, Fe2+ mediated cell death does not always occur via apoptotic pathway (Chew et al., 2011). It also has the capacity to induce hemolytic cleavage of hydrogen peroxide (H2O2), iron overload may cause the generation of aggressive hydroxyl radical (OH°) or other iron centered radicals and in such case Fe2+ becomes hazardous metal (Gutteridge, 1982; Eaton et al., 2002). As most of the enzymes that cause site specific oxidation of the lipid molecules are either iron containing or iron dependent (lipoxygenase), iron pool catalyze the formation of alcoxy radicals (L-O°) that cause PUFA fragmentation (Cheng and Li, 2007). These enzymes also inhibit lipophilic iron chelators that can chelate intracellular free iron molecules and induce ferroptosis (Kuhn et al., 2015; Abeysinghe et al., 1996; Barradas et al., 1989).
Stress mechanism of ferroptosis
Imbalance in the antioxidant system results in excessive ROS production and can trigger lipid peroxidation to damage plasma membrane and induce ferroptosis (Hassannia et al., 2019). Certain concentration of ROS can increase the repair of damages DNA, strand breaks and promote cell survival, but excessive ROS severely damages biofilm, protein and nucleic acid and finally leads to cell death (Chen et al., 2020). The generative mechanism of ferroptosis involves iron overloading, lipid peroxidation and downstream execution whereas the regulatory mechanism includes Glu/GPX4 pathway as well as mevalonate pathway and trans-sulfuration pathway (Chen et al., 2020).
Role of system Xc-
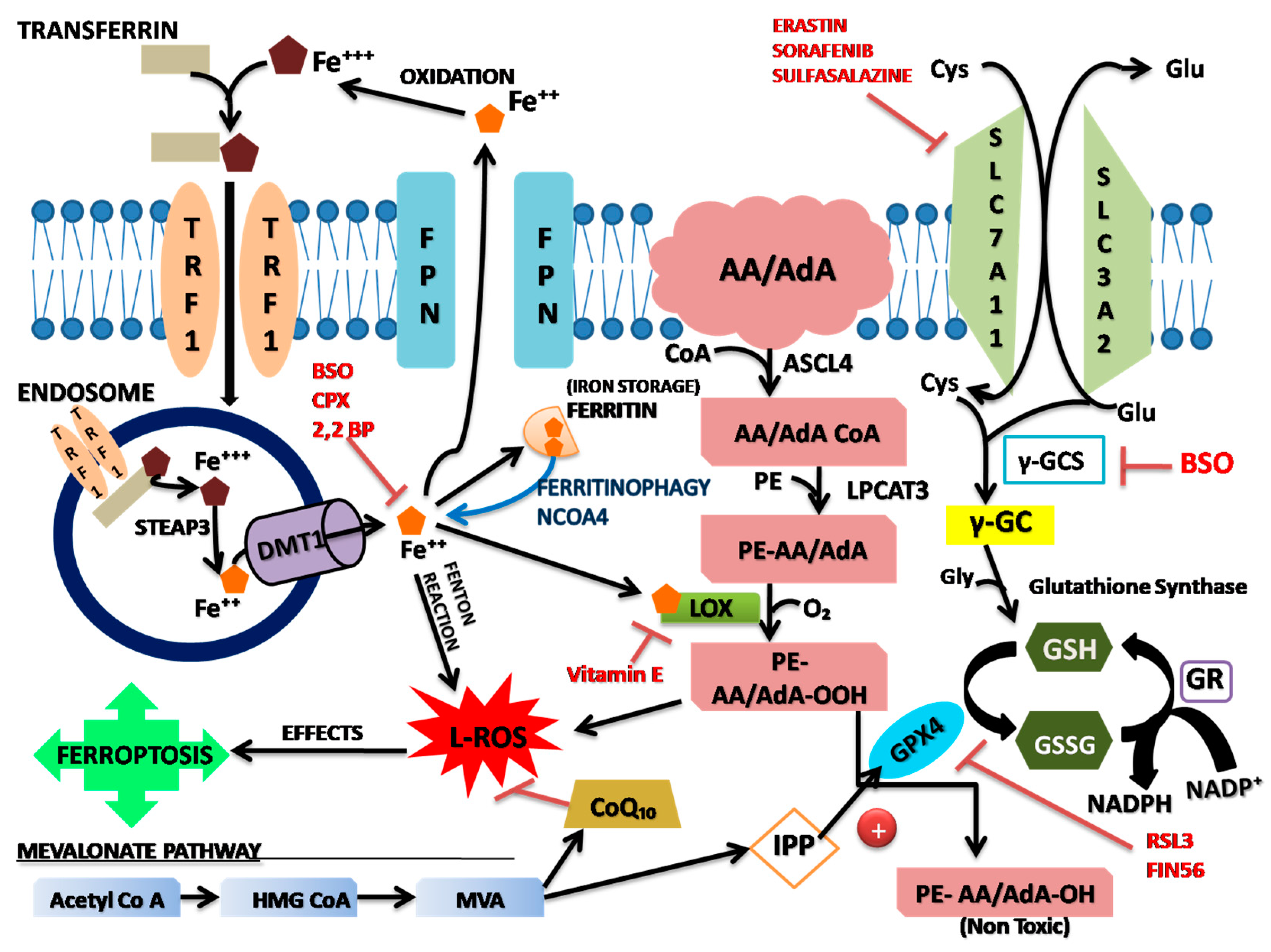
Amino acids, vitamins that are required for the growth and proliferation of mammalian cells in culture and found that cystine (Cys2) which is the oxidized thiol-containing form of cysteine (Cys) one of the most essential amino acids. In absence of Cys2, cells fail to grow as there will be rapid depletion of Cys-containing antioxidant tripeptide GSH (c-L-glutamyl-L-cysteinglycine) and subsequently the cells die due to the accumulation of L-ROS (Banni et al., 1977; Eagle et al., 1961; Eagle, 1959; Eagle, 1955). System X-C is a Cystine/Glutamate (Cys/Glu) antiporter which play an important role in ferroptosis. It causes uptake of Cys along with the export of Glu (Dixon et al., 2014; Dixon et al. 2012). It is a heterodimeric, amino acid antiporter which is present at the cell surface and composed of twelve pass transmembrane transporter protein named SLC7A11 (Sato et al., 1999). This system plays important role in Glutathione synthesis. Glutathione is a tripeptide made of glutamine, cysteine and glysine. Glu is linked to Cys by GCL to form glutamylcysteine. This dipeptide, Glutamylcysteine is linked to Gly by the GSS to form the final GSH molecule (Griffith, 1999). Free GSH has antioxidant property (Griffith, 1999; Staal, 1998). GPX4 is the GSH dependent enzyme that helps in reduction of hydroperoxidase (LOOH) to lipid alcohols (L-OH) (Seiler et al., 2008; Ursini et al., 1985). Inactivation of GPX4 is required for induction of ferroptosis as it involves lipid peroxidation of PUFA chains of Membrane lipids and causes iron dependent accumulation of L-ROS (Yang et al., 2014; Skouta et al., 2014; Angeliet al., 2014, Dixon et al., 2012; Cheng and Li, 2007). Oxygen and Fe2+ also help in lipid peroxidation and cause L-ROS accumulation (Cao and Dixon, 2016). This accumulation can be prevented by continuous uptake of Cys2 and GSH synthesis. It was found that ferroptosis can only happen when there is direct or indirect inactivation of GPX4 and when highly oxidizable PUFA arachchidonic acid is present in the membrane (Cao and Dixon, 2016). Subsequent studies demonstrated that both lipophilic antioxidants and iron chelators can block this cell death (Yonezawa et al., 1996; Murphy et al., 1990; Murphy et al., 1989; De Brabanderet al., 1979).
Iron metabolism pathway
The iron present in ferric state (Fe3+) forms a complex with transferrin and then the complex bind to Transferrin Receptor 1 (TFR1) on the cell membrane and then it is transplanted into the cell via endocytosis (Hao et al., 2018). By the action of Six-Transmembrane Protein of prostate 3 (STEAP3) Fe3+ is degraded into highly reactive Fe2+ and this ferrous form then translocate from endosome to the cytoplasm via Divalent Metal Transporter 1 (DMT1) (Chen et al., 2020). Thus, an unstable pool of iron is generated inside the cytoplasm. Under normal condition, part of this Fe2+ pool is stirred in ferritin to protect cells and tissues from iron mediated damage, while another part of the Fe2+ are pumped out through ferroportin on the cell membrane. Thus, the intracellular iron concentration remains stable (Chen et al., 2020). When the iron overload occurs, this balance is disrupted. As a result, Fe3+ and hydroxyl radicals can be directly catalyzed by Fenton chemical reaction. As hydroxyl radicals are most unstable oxygen free radicals and are active lethal ROS, they can easily get electrons to form other molecules (like O2-) which can cause lipid peroxidation and ferroptosis. Fe3+ can be reduced to Fe2+ by the superoxide radical (O2-) reaction (Hassannia et al., 2019). Under stress condition Ferritin is self-degraded to Fe2+ by the process called iron autophagy and then induces ferroptosis (Chen et al., 2020). Increase of iron uptake by TFR1 or reduction of stable iron by self-degradation of ferritin, leads to overload of iron inside the cell which causes oxidative damage and cause ferroptosis (Yang et al., 2008). This iron overload also induces noncanonical pathway of ferroptosis (Hassannia et al., 2018; Li et al., 2017). Iron export protein CDGSH iron sulfur domain 1 (CISD1) in the mitochondrial membrane reduces the accumulation of iron and production of L-ROS in the mitochondria thus prevents ferroptosis.
Lipid metabolism pathway and accumulation of lipid peroxides
Newly synthesized fatty acids must transform the long chain fatty acids into coenzyme A (CoA) for getting into the phospholipids (PLs) and this is mediated by Acetyl-CoA synthetase (Hassannia et al., 2019). Acetyl-CoA Synthetase Long Chain Family Member 4 (ACSL4) catalyzes the acylation reaction of Arachidonic Acid (AA) and Adrenic Acid (AdA) and this enzyme is important to execute ferroptosis (Doll et al., 2017). Now this acylation products combine with Phosphatidylethanolamines (PE) into membrane phospholipid by the action of Lypophosphatidylcholine Acetyltransferase 3 (LPCTA3) and produce PE-AA and PE-AdA (Kagan et al., 2017). ACSL4 and LPCTA3 make the cell membrane rich in sensitive PUFAs and lipoxygenase (LOX) especially 15-LOX (Yang et al., 2016) and then oxidize PUFAs into lipid hydroperoxides (PE-AA-O-OH or PE-AdA-O-OH) (Wenzel et al., 2017). When there is accumulation of Fe2+ in the cytoplasm the lipid hydroperoxide forms toxic L-ROS and cause cell damage. These radicals capture electrons from adjacent PUFA and launch a new round of lipid oxidation reaction and cause more serious damage (Kagan et al., 2017). ACSL (Kagan et al., 2017) and vitamin E (Wenzel et al., 2017) can block ferroptosis. Phosphatidylethanolamine-Binding Protein (PEBP1) increases the binding of LOX15 and increases the PUFAs in the cell membrane and promotes ferroptosis (Wenzel et al., 2017). The integration of PUFAs into the PLs after acylation and the production of lipid free radicals determine the advancement of ferroptosis (Dixon et al., 2019). PUFA can be incorporated into the PLs in different ways (phosphatidylcholine, phosphatidylinositol or phosphatidylethanolamine, PE) and it is dependent on the length of the carbon chain and the degree of unsaturation (Magtanong et al., 2016). Acylated chains of AA and AdA should be integrated with PE to induce ferroptosis (Kagan et al., 2017). L-ROS attack the mammalian phospholipid bilayer membrane and functions through 2 different pathways (Hassannia et al., 2019).
Non-enzymatic lipid peroxidation pathway
In such case free radicals (OH°) captures the hydrogen ions of PUFAs which are present in the plasma membrane and form a phospholipid free radical which acts with the oxygen molecule (O2) to produce the Phospholipid hydrogen peroxide radical (PLOO°) which can also capture hydrogen atoms from PUFAs and produces Phospholipid hydroperoxide (PLOOH) and a new PL°. This newly synthesized phospholipid radical can again start a new oxidation reaction (Hassannia et al., 2019).
Lipid peroxidation pathway
Lipoxigenase (LOX) plays major role in this pathway, and it causes dehydrogenation of PUFAs to form PLOOH. PLOOH is decomposed into alcoxy radical (PLO°) and in the presence of Fe2+ it can attack other PUFAs to trigger a chain reaction of lipid peroxidation. PLOOH through decomposition can produce 4-Hydroxynonenal (4-HNE) and Malondialdehyde (MDA). They along with PUFA-PLs reduce the stability of the cell membrane and increase permeability and thus resulting into cell death via ferroptosis (Hassannia et al., 2019).
GPX4 pathway
GPX4 is a selenoprotein, synthesized in the presence of selenium (Se) and it can eliminate PLOOH of PUFAs by transforming it into inactivated phosphatidylcholine (PLOH) because of this the lipid peroxidation process be prevented. Thus, ferroptosis will be inhibited (Hassannia et al., 2019). On the other hand, Cys uptake inside the cell is required to synthesize GSH and this GSH is an essential cofactor of GPX4 and maintains its function. GPX4 and GSH are antioxidants and eliminate ROS. GPX4 eliminates PLOOH and converts them into alcohol (PL-OH) under the assistance of GSH. GPX4 has antagonistic function of active Fe2+ and converts H2O2 into H2O (Imai et al., 2003). When, there is high concentration of extracellular glutamate, the synthesis of GSH gets affected (Chen et al., 2020). Latest research indicates that, Interferon γ (IFNγ), which is released by CD8+ T-cells, can downregulate the expression of Glutathione antiporter and reduce the uptake of cystine by tumour cell which affects the production and activity of GPX4 and enhance the occurrence of Ferroptosis to have an anti-tumour effect (Wang et al., 2019).
Mevalonate (MVA) pathway
It regulates the cholesterol synthesis and isoprene modification of the small G-protein after translation. In this pathway at first, Acetyl-CoA forms 3-Hydroxyl-3-Methylglutaryl Coenzyme A (HMGCoA) by the action of 3-Hydroxyl-3-Methulreductase (HMGCR). HMGCoA reduces to MVA then produces Isoprenyl Pyrophosphate (IPP) under the action of a series of enzymes. This IPP produces Famesyl Pyrophosphate (FPP) under the action of famesyl phosphate synthetase. This FPP generates squalene by the action of Squalene Synthease (SQS). Then cholesterol is formed under the action of squalene cyclase (Kim et al., 2014). This plays important role in GPX4 maturation. During the synthesis of GPX4, selenocysteine must be inserted into its catalytic center to function as antioxidant. IPP plays important role in this step (Warner, 2000). It promotes the formation of isoprene transferase and thus promotes the integration of selenocysteine on the GPX4 catalytic subunit (Kim et al., 2014).
Other pathways
p53 reduces the uptake of cystine by decreasing the expression of SLC7A11 which inhibits the transporter activity, and this decreases GSH synthesis, inhibit GPX4 activity, increase L-ROS and cause ferroptosis (Jiang et al., 2015). Activation of MAPK pathway can induce ferroptosis in cancer cells when RAS/RAF/MEK/ERK will be activated (Xie et al., 2016).
Figure 1.
Schematic diagram of oxidative stress during ferroptotic cell death.
References
- Abeysinghe, R.D.; Roberts, P.J.; Cooper, C.E.; MacLean, K.H.; Hider, R.C.; Porter, J.B. The environment of the lipoxygenase iron binding site explored with novel hydroxypyridinone iron chelators. Journal of Biological Chemistry 1996, 271, 7965–7972. [Google Scholar] [CrossRef]
- Andrews, N.C.; Schmidt, P.J. Iron homeostasis. Annu. Rev. Physiol. 2007, 69, 69–85. [Google Scholar] [CrossRef]
- Angeli, J.P.F.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; Basavarajappa, D. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nature cell biology 2014, 16, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Barradas, M.A.; Jeremy, J.Y.; Kontoghiorghes, G.J.; Mikhailidis, D.P.; Hoflbrand, A.V.; Dandona, P. Iron chelators inhibit human platelet aggregation, thromboxane A2 synthesis and lipoxygenase activity. FEBS letters 1989, 245, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.A.; Cookson, B.T. Salmonella induces macrophage death by caspase-1-dependent necrosis. Molecular microbiology 2000, 38, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.Y.; Dixon, S.J. Mechanisms of ferroptosis. Cellular and Molecular Life Sciences 2016, 73, 2195–2209. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Sarkar, S.; Bhattacharya, S. Toxic metals and autophagy. Chemical research in toxicology 2014, 27, 1887–1900. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, S.; Li, J.; Li, Z.; Quan, J.; Liu, X.; Tang, Y.; Liu, B. The latest view on the mechanism of ferroptosis and its research progress in spinal cord injury. Oxidative Medicine and Cellular Longevity 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Li, Y. What is responsible for the initiating chemistry of iron-mediated lipid peroxidation: an update. Chemical Reviews 2007, 107, 748–766. [Google Scholar] [CrossRef]
- Chew, K.C.; Ang, E.T.; Tai, Y.K.; Tsang, F.; Lo, S.Q.; Ong, E.; Ong, W.Y.; Shen, H.M.; Lim, K.L.; Dawson, V.L.; Dawson, T.M. Enhanced autophagy from chronic toxicity of iron and mutant A53T α-synuclein: implications for neuronal cell death in Parkinson disease. Journal of Biological Chemistry 2011, 286, 33380–33389. [Google Scholar] [CrossRef]
- Crespo-López, M.E.; Macêdo, G.L.; Pereira, S.I.; Arrifano, G.P.; Picanço-Diniz, D.L.; do Nascimento, J.L.M.; Herculano, A.M. Mercury and human genotoxicity: critical considerations and possible molecular mechanisms. Pharmacological research 2009, 60, 212–220. [Google Scholar] [CrossRef]
- De Brabander, M.; Van Belle, H.; Aerts, F.; Van De Veire, R.; Geuens, G. Protective effect of levamisole and its sulfhydryl metabolite OMPI against cell death induced by glutathione depletion. International journal of immunopharmacology 1979, 1, 93–100. [Google Scholar] [CrossRef]
- Degterev, A.; Huang, Z.; Boyce, M.; Li, Y.; Jagtap, P.; Mizushima, N.; Cuny, G.D.; Mitchison, T.J.; Moskowitz, M.A.; Yuan, J. Chemical inhibitor of nonapoptotic cell death with therapeutic potential for ischemic brain injury. Nature chemical biology 2005, 1, 112–119. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; Morrison III, B. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; Prokisch, H. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nature chemical biology 2017, 13, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H.; Piez, K.A.; Oyama, V.I. The biosynthesis of cystine in human cell cultures. Journal of Biological Chemistry 1961, 236, 1425–1428. [Google Scholar] [CrossRef] [PubMed]
- Eagle, H. Nutrition needs of mammalian cells in tissue culture. Science 1955, 122, 501–504. [Google Scholar] [CrossRef]
- Eagle, H. Amino acid metabolism in mammalian cell cultures. Science 1959, 130, 432–437. [Google Scholar] [CrossRef]
- Eaton, J.W.; Qian, M. Molecular bases of cellular iron toxicity. Free Radical Biology and Medicine 2002, 32, 833–840. [Google Scholar] [CrossRef]
- Fuchs, Y.; Steller, H. Programmed cell death in animal development and disease. Cell 2011, 147, 742–758. [Google Scholar] [CrossRef]
- Gao, M.; Monian, P.; Quadri, N.; Ramasamy, R.; Jiang, X. Glutaminolysis and transferrin regulate ferroptosis. Molecular cell 2015, 59, 298–308. [Google Scholar] [CrossRef]
- Griffith, O.W. Biologic and pharmacologic regulation of mammalian glutathione synthesis. Free Radical Biology and Medicine 1999, 27, 922–935. [Google Scholar] [CrossRef]
- Gutteridge, J.M. The role of superoxide and hydroxyl radicals in phospholipid peroxidation catalysed by iron salts. FEBS letters 1982, 150, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Liang, B.; Huang, Q.; Dong, S.; Wu, Z.; He, W.; Shi, M. Metabolic networks in ferroptosis. Oncology letters 2018, 15, 5405–5411. [Google Scholar] [CrossRef]
- Hassannia, B.; Vandenabeele, P.; Berghe, T.V. Targeting ferroptosis to iron out cancer. Cancer cell 2019, 35, 830–849. [Google Scholar] [CrossRef]
- Hassannia, B.; Wiernicki, B.; Ingold, I.; Qu, F.; Van Herck, S.; Tyurina, Y.Y.; Bayır, H.; Abhari, B.A.; Angeli, J.P.F.; Choi, S.M.; Meul, E. Nano-targeted induction of dual ferroptotic mechanisms eradicates high-risk neuroblastoma. The Journal of clinical investigation 2018, 128, 3341–3355. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Nakagawa, Y. Biological significance of phospholipid hydroperoxide glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radical Biology and Medicine 2003, 34, 145–169. [Google Scholar] [CrossRef]
- Jiang, L.; Kon, N.; Li, T.; Wang, S.J.; Su, T.; Hibshoosh, H.; Baer, R.; Gu, W. Ferroptosis as a p53-mediated activity during tumour suppression. Nature 2015, 520, 57–62. [Google Scholar] [CrossRef]
- Kagan, V.E.; Mao, G.; Qu, F.; Angeli, J.P.F.; Doll, S.; St Croix, C.; Dar, H.H.; Liu, B.; Tyurin, V.A.; Ritov, V.B.; Kapralov, A.A. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nature chemical biology 2017, 13, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Kim, Y.B.; Kim, J.K.; Kim, S.U.; Park, S.U. Molecular cloning and characterization of mevalonic acid (MVA) pathway genes and triterpene accumulation in Panax ginseng. Journal of the Korean Society for Applied Biological Chemistry 2014, 57, 289–295. [Google Scholar] [CrossRef]
- Kuhn, H.; Banthiya, S.; Van Leyen, K. Mammalian lipoxygenases and their biological relevance. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids 2015, 1851, 308–330. [Google Scholar] [CrossRef]
- Li, F.; Qiu, Z.; Zhang, J.; Liu, W.; Liu, C.; Zeng, G. Investigation, pollution mapping and simulative leakage health risk assessment for heavy metals and metalloids in groundwater from a typical brownfield, middle China. International journal of environmental research and public health 2017, 14, 768. [Google Scholar] [CrossRef]
- Lieu, P.T.; Heiskala, M.; Peterson, P.A.; Yang, Y. The roles of iron in health and disease. Molecular aspects of medicine 2001, 22, 1–87. [Google Scholar] [CrossRef]
- Linkermann, A.; Skouta, R.; Himmerkus, N.; Mulay, S.R.; Dewitz, C.; De Zen, F.; Prokai, A.; Zuchtriegel, G.; Krombach, F.; Welz, P.S.; Weinlich, R. Synchronized renal tubular cell death involves ferroptosis. Proceedings of the National Academy of Sciences 2014, 111, 16836–16841. [Google Scholar] [CrossRef]
- Liu, Y.; Shoji-Kawata, S.; Sumpter, R.M.; Wei, Y.; Ginet, V.; Zhang, L.; Posner, B.; Tran, K.A.; Green, D.R.; Xavier, R.J.; Shaw, S.Y. Autosis is a Na+, K+-ATPase–regulated form of cell death triggered by autophagy-inducing peptides, starvation, and hypoxia–ischemia. Proceedings of the National Academy of Sciences 2013, 110, 20364–20371. [Google Scholar] [CrossRef]
- Lockshin, R.A.; Zakeri, Z. Apoptosis, autophagy, and more. The international journal of biochemistry & cell biology 2004, 36, 2405–2419. [Google Scholar]
- Munshi, C.; Bhattacharya, S. The “Irony” of Ferroptosis: A Review on Neurological Challenges. 2022. [Google Scholar]
- Murphy 2022, T.H.; Schnaar, R.L.; Coyle, J.T. Immature cortical neurons are uniquely sensitive to glutamate toxicity by inhibition of cystine uptake. The FASEB Journal 1990, 4, 1624–1633. [Google Scholar] [CrossRef]
- Murphy, T.H.; Miyamoto, M.; Sastre, A.; Schnaar, R.L.; Coyle, J.T. Glutamate toxicity in a neuronal cell line involves inhibition of cystine transport leading to oxidative stress. Neuron 1989, 2, 1547–1558. [Google Scholar] [CrossRef]
- Seiler, A.; Schneider, M.; Förster, H.; Roth, S.; Wirth, E.K.; Culmsee, C.; Plesnila, N.; Kremmer, E.; Rådmark, O.; Wurst, W.; Bornkamm, G.W. Glutathione peroxidase 4 senses and translates oxidative stress into 12/15-lipoxygenase dependent-and AIF-mediated cell death. Cell metabolism 2008, 8, 237–248. [Google Scholar] [CrossRef]
- Skouta, R.; Dixon, S.J.; Wang, J.; Dunn, D.E.; Orman, M.; Shimada, K.; Rosenberg, P.A.; Lo, D.C.; Weinberg, J.M.; Linkermann, A.; Stockwell, B.R. Ferrostatins inhibit oxidative lipid damage and cell death in diverse disease models. Journal of the American Chemical Society 2014, 136, 4551–4556. [Google Scholar] [CrossRef]
- Staal, F.J. Glutathione and HIV infection: reduced reduced, or increased oxidized? European journal of clinical investigation 1998, 28, 194–196. [Google Scholar] [CrossRef]
- Valko, M.M.H.C.M.; Morris, H.; Cronin, M.T.D. Metals, toxicity and oxidative stress. Current medicinal chemistry 2005, 12, 1161–1208. [Google Scholar] [CrossRef]
- Warner, G.J.; Berry, M.J.; Moustafa, M.E.; Carlson, B.A.; Hatfield, D.L.; Faust, J.R. Inhibition of selenoprotein synthesis by selenocysteine tRNA [Ser] Sec lacking isopentenyladenosine. Journal of Biological Chemistry 2000, 275, 28110–28119. [Google Scholar] [CrossRef]
- Wang, W.; Green, M.; Choi, J.E.; Gijón, M.; Kennedy, P.D.; Johnson, J.K.; Liao, P.; Lang, X.; Kryczek, I.; Sell, A.; Xia, H. CD8+ T cells regulate tumour ferroptosis during cancer immunotherapy. Nature 2019, 569, 270–274. [Google Scholar] [CrossRef]
- Woo, J.H.; Shimoni, Y.; Yang, W.S.; Subramaniam, P.; Iyer, A.; Nicoletti, P.; Martínez, M.R.; López, G.; Mattioli, M.; Realubit, R.; Karan, C. Elucidating compound mechanism of action by network perturbation analysis. Cell 2015, 162, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: process and function. Cell Death & Differentiation 2016, 23, 369–379. [Google Scholar]
- Yagoda, N.; von Rechenberg, M.; Zaganjor, E.; Bauer, A.J.; Yang, W.S.; Fridman, D.J.; Wolpaw, A.J.; Smukste, I.; Peltier, J.M.; Boniface, J.J.; Smith, R. RAS–RAF–MEK-dependent oxidative cell death involving voltage-dependent anion channels. Nature 2007, 447, 865–869. [Google Scholar] [CrossRef]
- Yang, W.S.; Stockwell, B.R. Ferroptosis: death by lipid peroxidation. Trends in cell biology 2016, 26, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; Brown, L.M. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef]
- Yang, W.S.; Stockwell, B.R. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chemistry & biology 2008, 15, 234–245. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



