
Submitted:
24 October 2023
Posted:
25 October 2023
You are already at the latest version
Abstract
Despite the extensive research conducted on ruminal methanogens and anti-methanogenic intervention strategies over the last 50 years, most of the currently researched enteric CH4 abatement approaches have shown limited efficacy. This is largely because of the complex nature of animal production and the ruminal environment, host genetic variability of CH4 production, and an incomplete understanding of the role of the ruminal microbiome in enteric CH4 emissions. Recent sequencing-based studies suggest the presence of methanogenic archaea in extra-gastrointestinal tract tissues, including respiratory and reproductive tracts of cattle. While these sequencing data require further verification by culture-dependent methods, the consistent identification of methanogens with relatively greater frequency in the airway and urogenital tract of cattle plus increasing appreciation of the microbiome-gut-organ axis highlight the potential interactions between ruminal and extra-gastrointestinal methanogenic communities. Thus, a traditional singular focus on ruminal methanogens may not be sufficient, and a holistic approach which takes into consideration of the transfer of methanogens between ruminal, extra-gastrointestinal, and environmental microbial communities is of necessity to develop more efficient and long-term ruminal CH4 mitigation strategies. In the present review, we provide a holistic survey of the methanogenic archaea present in different anatomical sites of cattle and discuss potential seeding sources of the ruminal methanogens.
Keywords:
Methanogens
; enteric methane emission
; cattle
; microbiome
; holistic
; extra-intestinal microbial communities
1. Introduction
Global warming is projected to have major consequences on food security worldwide, exacerbating the expected increase in food demand by 70% to 100% by 2050 due to the population growth [1,2,3]. Anthropogenic greenhouse gas (GHG) emissions, particularly CH4, are major contributors to global warming. Methane has a global warming potential approximately 28 times greater than CO2 [2]. The agricultural sector is considered to be one of the major sources of CH4 emissions, with ruminant animals, particularly domestic ruminants being significant contributors [4,5]. Ruminal methanogens are responsible for over 87% of total CH4 emissions from ruminants and about 26% in comparison to other sources of CH4 production in the environment [3,6]. Methane is produced in the rumen during the normal fermentation process by methanogenic archaea which use either CO2 and hydrogen (H2), methylamines or methanol, or acetate and H2 to produce CH4 [7]. Other microorganisms residing within the rumen such as bacteria, protozoa, and fungi can provide methanogens with excessive H2, either directly or indirectly, and thereby promote methanogenic activity [8,9]. Although utilizing excess H2 benefits ruminal fermentation by preventing H2 build-up and feedback inhibition, ruminal CH4 production represents 2%- 12% of gross energy loss [10]. Therefore, there is an impetus need for developing approaches to mitigate CH4 emissions from ruminant livestock to reduce energy loss from animals while improving environmental health.
Over the last five decades, ruminal methanogens and anti-methanogenic intervention strategies have been extensively explored as a means of mitigating CH4 emissions [11]. These strategies involve alterations in animal management, dietary composition, and ruminal fermentation, as well as direct inhibition of the growth and metabolic activity of methanogens using anti-methanogenic compounds and substances [12,13]. However, most of these approaches have shown limited efficacy due to the complex nature of animal management and ruminal physiology [14,15]. Additionally, recent research has focused on reducing CH4 emissions from ruminants by genetic selection and manipulation of the ruminal microbiota, and the latter has become an active area of research due to advances in next-generation sequencing technologies [16]. While genomic selection could provide a long-term solution to CH4 emissions [17,18,19], the impact of the selected anti-methanogenic traits on ruminal nutrient metabolism, fermentation, and the microbial community is yet to be defined. Recent identification of a distinctive ruminal microbiota between cattle and sheep with high and low CH4 emitting phenotypes [20] suggest that the manipulation of the ruminal microbiota to mitigate livestock CH4 production may provide long-lasting solutions.
While most of the current research has focused on characterizing the taxonomic diversity and abundance of methanogenic archaea in the rumen and the potential role of the ruminal microbiome in methanogenesis, emerging evidence suggests that microbial communities associated with extra-ruminal sites of animals including the reproductive and respiratory tracts and mammary glands may harbor methanogenic archaeal species [7,21,22,23,24] and these extra-gastrointestinal methanogens may interact with ruminal methanogens. Some of the methanogenic taxa (e.g., Methanobrevibacter spp.) have been reported to be shared as core taxa across ruminal, respiratory, and reproductive tract-associated microbial communities in cattle [21,25]. This, coupled with increased appreciation of the microbiome-gut-organ (respiratory/reproductive) axis [25,26] highlight the possible existence of interactions between methanogenic archaea in the rumen with methanogens and microbiomes present in extra-gastrointestinal organs. Such interactions between the rumen and other organs may be responsible for seeding the rumen or other organs with methanogenic species and/or influencing metabolic activities of the ruminal methanogens. The potential seeding of the rumen with methanogens by methanogens present in the reproductive tract is further supported by the recent identification of methanogenic archaeal species in fetal fluid and intestines of bovine fetuses [21,27]. Thus, these new developments point out that focusing solely on the methanogens present in the rumen and developing CH4 mitigation strategies targeted at only ruminal methanogens could be too narrow of an approach. The ruminal and extra-gastrointestinal methanogens and their interactions on ruminal CH4 emissions should be considered. In this review, we first provide a holistic survey of the methanogenic archaea present in different anatomical sites of cattle. We then discuss potential seeding sources of the ruminal methanogenic archaea in cattle. In addition, we highlight some challenges and future research directions associated with studying ruminal and extra-gastrointestinal methanogens in cattle.
2. General Overview of Methanogens
Methanogens are a diverse group of microorganisms that produce CH4 as a metabolic byproduct of their energy conservation processes [28,29]. They are commonly present in a variety of environments including the digestive tracts of animals, predominantly ruminants, wetlands, and other anaerobic environments [11,30]. Methanogens are classified in the domain archaea and are phylogenetically diverse [31]. Methane production is an essential part of the global carbon cycle, accounting for approximately 18% of anthropogenic GHG emissions [32].
Methanogens are classified within the archaeal phyla Euryarchaeota, Crenarchaeota, and Korarchaeota. They are further subdivided into several orders and families based on their phylogenetic relationships and metabolic pathways [33,34]. Recent research has revealed the diversity of methanogens in different environments, with new genera and species being identified through metagenomic analysis and other non-culture based techniques [35,36]. High-throughput sequencing techniques have enabled the discovery of novel methanogen species and lineages from diverse environments, such as the animal gut and reproductive tracts, rice paddies, and deep-sea sediments [11,37,38]. For example, a recent study using metagenomic analysis identified a new genus of methanogen from the digestive tract of a giant panda, which was found to be closely related to Methanobrevibacter smithii, a common human gut methanogen [39,40,41].
Methanogens are believed to be one of the earliest forms of life on Earth, with evidence suggesting that they were present over 3.5 billion years ago [42]. Their unique metabolic capabilities are thought to have evolved in response to the anaerobic conditions of the early Earth, where they played a significant role in the carbon cycle [43]. Based on the substrates used to produce CH4, methanogens can be classified as hydrogenotrophic, acetoclastic and methylotrophic methanogens [44,45]. The hydrogenotrophic class uses H2 for the reduction of CO2 into CH4. Examples of such methanogens are Methanobrevibacter, Methanobacterium, and Methanomicrobiales. The rumen is mainly inhabited by hydrogenotrophic methanogens [46]. The second class, acetoclastic methanogens which are most commonly present in freshwater sediments and anaerobic digestors, but use alcohols such as ethanol or 2-propanol as electron donors to produce CH4 [41,42,47,48,49]. The methylotrophic class which is predominant in freshwater and wetland soils [50,51,52] relies on methyl groups such as methanol and methylamines to produce CH4, and encompasses the order Methanococcales and Methanosarcinales. Of note, hydrogenotrophic methanogens are the focus of the present review as they are the main class of methanogens involved in enteric CH4 emissions from cattle. The other two classes of methanogens will be briefly discussed as potential environmental sources that could seed ruminal methanogens in cattle.
3. Main Methanogenic Species in the Rumen
The methanogenic community in the rumen of ruminant animals is dominated by two main phyla: Euryarchaeota and Crenarchaeota [53]. The Euryarchaeota phylum is the most abundant and taxonomically diverse group of methanogens in the rumen. This phylum includes the orders Methanobacteriales, Methanomicrobiales, Methanosarcinales, and Methanocellales. The Crenarchaeota phylum contains the order Thermoproteales and is present in some ruminants but is less abundant as compared to the Euryarchaeota phylum [54] (Table 1).
Methanobrevibacter is the most abundant genus of ruminal methanogens in cattle and it belongs to Euryarchaeota phylum [54]. Multiple species of Methanobrevibacter including M. ruminantium, M. smithii, and M. gottschalkii have been reported in the rumen of cattle other ruminant species (Table 1). The Methanobrevibacter spp. produce CH4 from H2 and CO2, and they can also utilize formate and acetate as alternative substrates [55]. Another common archaeal genus found in the rumen is Methanosphaera [56], and species within this genus consume H2 and CO2, as well as methanol and methylamines to produce CH4 [56]. Multiple species within Methanosphaera (e.g., M. stadtmanae and M. cuniculi) have also been detected in the rumen, and believed to contribute to the ruminal CH4 production [57,58]. Guzman and colleagues identified Methanomicrobiales mobile, Methanoccocales votae, and Methanobrevibacter spp. from the gastrointestinal tract of neonatal dairy calves sampled within five minutes of birth [59]. Methanomassiliicoccus, within the phylum Euryarchaeota, is a relatively recent discovered genus from the rumen and feces of ruminants [57]. This genus is unique in a way that it produces CH4 through the reduction of methanol and methylamines, rather than H2 and CO2 [57]. Methanomassiliicoccushas been suggested as a potential target for inhibiting ruminal CH4 emissions due to its ability to outcompete hydrogenotrophic methanogens [60]. Also, Methanospirillum, classified within the order Methanomicrobiales and the phylum Euryarchaeota, has been found in the rumen and feces of cattle and sheep [61]. Additionally, Methanospirillum hungatei and Methanospirillum lacunae are also present in the rumen [62]. Overall, the rumen is home to taxonomically and metabolically diverse methanogenic archaeal species.
4. Pro-and Anti-Methanogenic Ruminal Microorganisms
Methanogens undergo methanogenesis; an energy-intensive process that requires specific substrates and environmental conditions [63]. Syntrophic bacteria are among the most extensively studied microorganisms that enhance the activity of methanogens in the rumen. Syntrophic bacteria, such as Syntrophomonas spp. and Smithella spp., form symbiotic associations with methanogens, by degrading complex organic matter to simpler compounds that methanogens can utilize [57,64]. Examples of syntrophic bacteria are Syntrophomonas wolfei, Syntrophobacter fumaroxidans, and Pelotomaculum thermopropionicum. In this relationship, syntrophic bacteria oxidized short chain fatty acids (SCFAs) to produce H2 and CO2, both of which are used by methanogens to produce CH4 [65].
In addition to syntrophic bacteria, some other bacterial species may enhance the activity of methanogens through various mechanisms in the rumen. For example, exopolysaccharide-producing bacterial species can promote the aggregation of methanogens, creating microenvironments favoring methanogenesis [55]. Other bacteria species that produce secondary metabolites including formic acid and ethanol can facilitate methanogenesis [66]. Furthermore, Pelobacter spp. and Bacteroides spp. can promote methanogenesis by enhancing the growth of specific methanogenic species [67].
While some bacteria promote methanogens, there are some bacteria in the rumen that can inhibit methanogenesis. Sulfate-reducing bacteria (SRB) such as Desulfovibrio spp. consume organic matter and produce hydrogen sulfide (H2S). Both SRB and methanogens compete for H2, as SRB require it to reduce sulfate to sulfide; while methanogens need H2 to reduce CO2 for CH4 production. Because of this competition, the presence of SRB indirectly decreases methane production in the rumen [20]. Ruminal SRB utilize various forms of sulfur (S), including sulfate, sulfite, thiosulfate, and elemental S, as optional H2 sinks [68]. As the end-product of the sulfate reduction pathway, H2S can inhibit methanogenic activity, consequently reducing ruminal CH4 production [69]. In addition, abundance of bacterial genera such as Prevotella, Fibrobacter, and Ruminococcuswere found to be negatively correlated with methanogenic activity in dairy cows [14]. These bacteria that are involved in the degradation of fiber and the production of propionate can compete with methanogenesis for H2. Fumarate-reducing and lactate-producing bacteria may also inhibit methanogens, which was suggested by a study where nitrate supplementation reduced CH4 emission in grazing steers by promoting fumarate-reducing and lactate-producing bacteria [70]. Jenayathan and colleagues [12] also reported that direct-fed microbials comprising of Propionibacterium and Lactobacillus spp. were able to mitigate CH4 emissions in sheep. The genus Lactobacillus has a long history of being used as a probiotic and has recently been proposed to mitigate ruminal CH4 emissions [71]. Other genera such as Prevotella and Succinivibrio have been negatively associated with ruminal methanogenesis [56,72,73]. The family Succinivibrionaceae is a key gut microbial member in the Tammar wallaby that produce only 20% of the CH4 emissions of cattle [74]. This is because Succinivibrionaceae produce succinate through the fumarate-succinate pathway, which is an intermediate product of propionate, that are easily absorbed by the animal for energy [75].
Certain viral species can infect methanogens, called methanogenic viruses or methanophages. Methanophages against Methanosarcina, Methanococcus, and Methanobacterium [76,77,78] have been reported in methanogen abundant anoxic environments. Viruses that infect bacterial species can compete with methanogens for substrates [76] and have also been reported in methanogenic environments. Accordingly, it is plausible that viruses infecting methanogenic archaea or anti-methanogenic bacteria are present in the bovine rumen.
Methane produced in the rumen can be utilized by other ruminal community members. Methanotrophic archaea, also known as methanotrophs, are a group of archaea that can utilize CH4 as the sole source of carbon and energy [79]. Under aerobic conditions, methanotrophs combine O2 and CH4 to form formaldehyde, which is then incorporated into organic compounds via the ribulose monophosphate pathway by type I methanotrophs (γ -proteobacteria) or the serine pathway by type II methanotrophs (α proteobacteria) [79]. The methanotrophic bacteria including Methylobacterium, Methylomonas and Methylomicrobium genera have been detected in the bovine rumen and have been shown to influence methanogens [80].
Fungi species have been shown to promote methanogenic activity by producing various enzymes such as cellulases and hemicellulases, and these enzymes can break down complex polysaccharides (cellulose and hemicellulose) into simpler compounds that can be utilized by methanogens [81]. Anaerobic fungi, such as Neocallimastix spp., and Piromyces spp., have been extensively studied for their ability to enhance the activity of methanogens in the rumen [81]. These fungi species are commonly found in the rumen and play a crucial role in the degradation of plant biomass [82]. As discussed above, it is apparent that the activities of methanogens in the rumen are interdependent on other microbes and their respective activities in the ruminal environment. Thus, understanding the interactions between the methanogens and other microorganisms within the rumen is important as such interactions could be harnessed for enteric methane mitigations from cattle.
5. Methanogens in the Reproductive Tract: Vagina, Uterus and Semen
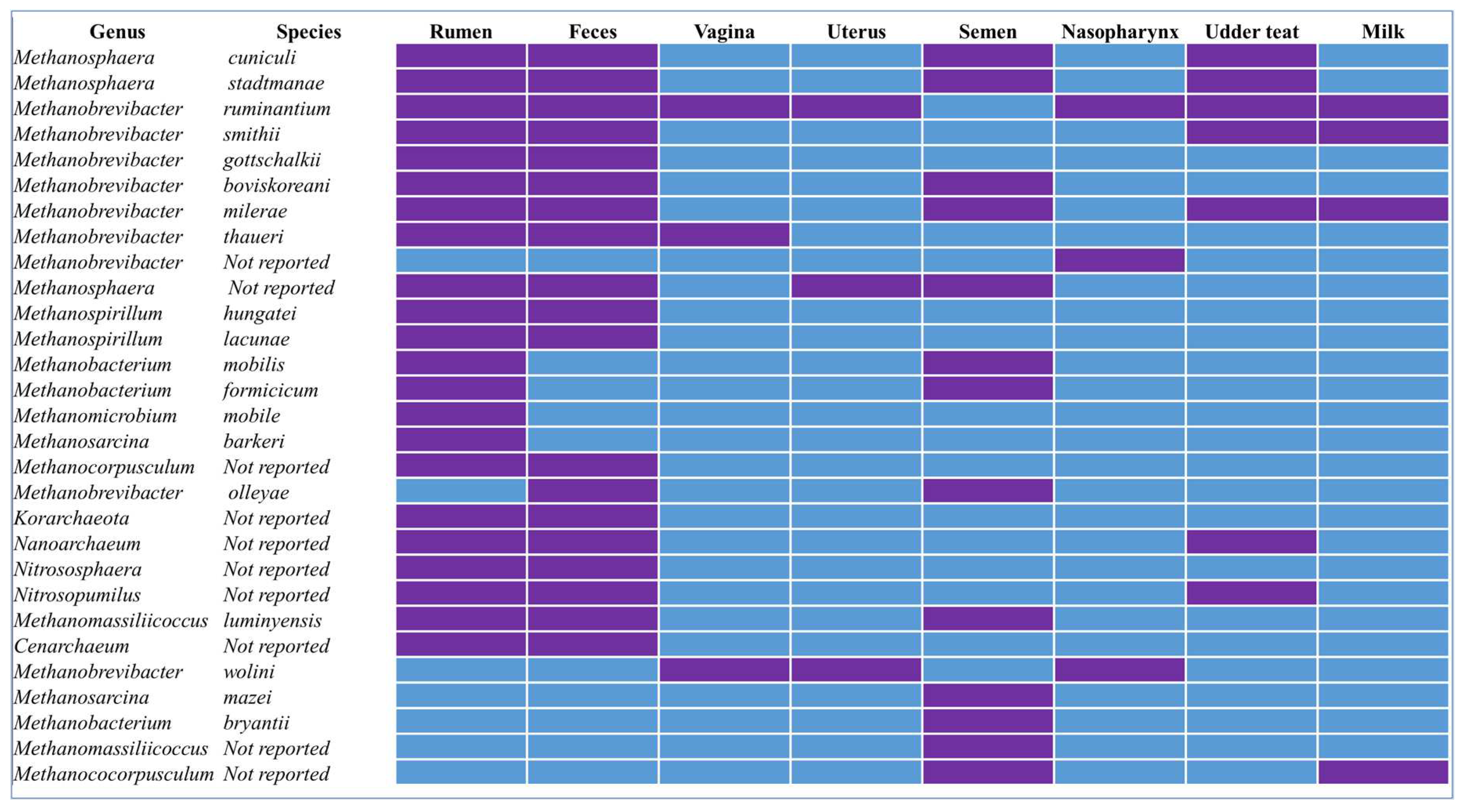
Methanogen presence has been reported in both the lower and upper reproductive tracts of cattle (Figure 1, Table 2). The microbial community associated with the bovine vaginal tract has been relatively well characterized as compared to the cervical and uterine microbiota [83]. Overall, there is less species richness and community diversity in the vaginal microbiota as compared to the bovine gut, and this community has been shown to influence reproductive health and fertility of female cattle [83,84]. Although the archaeal members of the vaginal microbiome have not yet been as extensively characterized as the bacterial members, the presence of some methanogens in the cattle vagina have been reported from 16S rRNA gene amplicon-based studies. The Methanobrevibacter genus has been reported to be the predominant methanogen genus in the vaginal samples of Nellore cows [85]. We recently reported 6 amplicon sequence variants (ASVs) that were classified as Methanobrevibacter spp. (5 of these taxa were unclassified at species level and the remaining one was M. ruminantium) present from vaginal swabs of both virgin yearling heifers and pregnant beef heifers [21]. The Methanobrevibacter ruminantium taxa was most frequently identified from the vaginal swab samples with greater abundance than the other Methanobrevibacter taxa [21].
While there are no culture-based studies reporting the isolation and identification of methanogens in the vaginal tract of cattle, methanogenic archaeal species have been cultured and isolated from the vaginal swabs [86] and urine specimens [87] of women with urinary tract infections. Belay et al. [86] cultured two different Methanobrevibacter smithii strains from the vaginal swabs of women diagnosed with bacterial vaginosis using traditional anaerobic culturing technique. Briefly, vaginal swabs were enriched in a medium containing various salts and additives, and yeast extract and tryptone in anaerobic serum tubes pressurized with H2-CO2 and supplemented with cysteine hydrochloride and Na2S. 9H20. Methane production in the serum tube was monitored using gas chromatography. Following enrichment, the cultures that produced CH4 gas were plated on medium with 1% Gelrite to isolate methanogens. Two methanogen isolates were isolated and taxonomically identified as Methanobrevibacter smithii PS and ALI based on the morphological, cultural, and immunological features. Likewise, Grine and colleagues [87] isolated Methanobrevibacter smithii strains from the urine samples of women suffering from urinary tract infections using a Hungate culture tube containing SAB medium, a versatile medium that supports the growth of most of methanogen species [88]. Based on their methanogenic archaeal culturing results from 383 urine specimens (61% of them from women) prospectively collected for diagnosing urinary tract infection, these authors suggest that M. smithii is part of the urinary microbiota of some individuals. Thus, both sequencing and culturing-based results obtained from bovine and human studies discussed above highlight that the microbial community in the bovine lower reproductive tract could harbor methanogenic archaea. The physiological function of methanogens in the lower reproductive tract of cattle remains to be explored even though some studies have suggested potential association of methanogens with reproductive health [21,85,89,90]. In the human vaginal tract, methanogens are known to play a significant role in prevention of acid accumulation (increase in vaginal pH) which disrupts the vaginal microbiota in bacterial vaginosis patients [86,87,91].
The in-utero environment has long been viewed as sterile as the cervix was thought to prevent ascending bacteria from the lower reproductive tract into the uterus (Wang et al., 2018). However, culture-independent high-throughput sequencing technologies have enabled the identification of commensal microbiota presence in the bovine uterus during both pregnancy and after parturition [93]. The potential role of the uterine microbiome in reproductive health, conception, and embryo development is increasingly appreciated [83,94,95]. Within the bovine uterine microbial community, methanogenic archaeal species have been reported. A sequencing-based study identified several methanogens in both pregnant and non-pregnant cows, including Methanosphaera stadtmanae, Methanobrevibacter ruminantium, and Methanobacterium congolense [85] (Table 2; Figure 1). Our lab has also identified Methanobrevibacter ruminantium and Methanobrevibacter wolini, which accounted for 0.05% of the total microbiota present in the uterus of virgin beef heifers (21-months old) [25]. We also observed greater abundance of Methanobrevibacter ruminantium (ASV330) in the uterine microbiota of beef cows that became pregnant than those remained open following artificial insemination, suggesting positive association of Methanobrevibacter ruminantium with fertility [96]. In contrary, methanogens have been shown to be present in uterus of cows with uterine infections, suggesting a potential role in pathogenesis [97]. The functional and taxonomic characteristics of methanogens in the bovine uterus, like those found in the vagina, are yet to be fully explored.
Recent sequencing-based studies revealed that there are diverse and dynamic microbial communities residing within the bull reproductive tract [98]. Methanogenic archaeal species are identified as commensal microbiota associated with bull semen and the abundance of methanogens in the semen may have an association with bull fertility. For example, Koziol and colleagues identified Methanobrevibacter, Methanosphaera, and Methanomassiliicoccus as the predominant methanogens present in the semen of breeding beef bulls [99] using 16S rRNA gene amplicon sequencing. Likewise, Methanocorpusculum spp. was identified in bovine bull semen, and the relative abundance of this methanogenic genus was inversely correlated with seminal commensal genera Ruminoccoceae and Rikenellaceae RC9 Gut group [100]. Our research group detected more than a dozen methanogenic archaeal taxa (Listed in Figure 1; Table 2) from semen samples collected at three different stages of yearling beef bull development [101]. These studies together demonstrated that the male reproductive tract is colonized by methanogens.
Although the functional features of the seminal microbiota including methanogenic archaeal members are yet to be characterized, a negative correlation of methanogenic archaeal abundance with bull fertility has been reported [99,100]. The bulls with low fertility had greater abundance of seminal Methanocorpusculum as compared to bulls with high fertility [100]. Semen samples with greater abundance of methanogens had significantly lower sperm motility compared to semen samples with lower levels of methanogens, suggesting a potential negative correlations between methanogens and sperm motility and development [99].
6. Methanogens in the Respiratory Tract
Bovine respiratory microbial communities, particularly bacterial microbiota in the upper and lower respiratory tracts have been well studied using both culturing and sequencing-based methods due to their role in protecting or predisposing animal to bovine respiratory disease (BRD), which is one of the costliest diseases affecting commercial feedlot cattle [102,103,104,105]. While most of these sequencing methods used to characterize bovine respiratory microbiota are mainly limited to the 16S rRNA amplicon (V4) and (V3-V4) sequencing (which is more specific to bacterial populations), several studies have reported the presence of methanogenic archaea in the upper respiratory tract (Table 2; Figure 1). Amat and colleagues reported the presence of the Methanobrevibacter genus in the nasopharynx of feedlot steers [21]. Several taxa within the Methanobrevibacter, including Methanobrevibacter ruminantium, have also been detected in the nasopharynx of virgin and pregnant beef cattle [21]. Two Methanobrevibacter species (wolinii and ruminantium) have also been reported in the nasopharyngeal microbiota of finishing feedlot heifers [25]. The lower airway is also colonized by a microbial community and bacterial microbiota associated with the trachea [103,106,107] and lung tissue have been relatively well documented in cattle. However, presence of archaeal species in the lower airway has not been reported in any of these studies. Since the 16S rRNA gene (V4 region) was used by most of these studies and neither archaeal targeted amplicon sequencing nor shotgun metagenomic sequencing approaches have been employed to characterize the lung tissue samples, it is challenging to make a conclusive statement on the presence or absence of methanogenic archaea in the bovine lower respiratory tract.
Figure 1.
Presence (purple color) or absence (blue) of methanogenic archaeal species reported in different anatomical sites of cattle. This figure was generated based on the data presented in the references listed in Table 1 and Table 2.
However, evidence derived from human studies suggests that the lower respiratory tract may harbor methanogenic archaea. Methanobrevibacter spp. (oralis and smithii) have been cultured and isolated from sputum, trachea-bronchial, and broncho-alveolar samples collected from humans [108].
Methanogenic archaeal cell presence in the upper respiratory tract of cattle raises an important question about the survival mechanisms of methanogens in such an O2 rich environment given that archaeal species involved in methanogenesis are believed to be strict anaerobes. It is not uncommon to identify anaerobic bacterial species some of which are associated with the ruminal commensal microbiota such as Ruminococcus and Thermodesulfovibrio (sulfate reducing bacteria) in the upper respiratory tract of cattle [109]. Anaerobic bacteria are predominant components of the upper respiratory tract, and mixed anaerobic-aerobic agents are often responsible for respiratory infections in humans[110]. Thus, despite that some of the methanogenic species in the nasopharyngeal and lung tissue samples of cattle can be transient and from eructation and inhalation of ruminal gas cap, the aerosols generated from the dust and soil particles [106,111,112], the airway mucosal surface of the cattle can harbor methanogens as part of their commensal microbiota. These airway specific methanogens could have evolved to adapt and survive in the microenvironments along the respiratory tract, especially those parts with reduced O2 levels. One of the potential methanogen-promoting factors in the airway can be associated with biofilm forming bacterial cells, as biofilms can provide localized anaerobic conditions. Biofilms can form in different areas of the respiratory tract [113]. In addition, the presence of oxygen-utilizing bacteria in the respiratory tract may support the growth of methanogens by consuming O2 and thereby creating anaerobic pockets where methanogens can persist [114,115]. Future culture-dependent studies are needed to isolate methanogens from the respiratory tract and explore metabolic features of these methanogens, and their interactions with the methanogens present in the rumen.
7. Methanogens in the Udder
The mammary gland is another site that has been reported to harbor relatively diverse and rich microbial communities [116]. Distinctive and site-specific microbial communities are present in various niches of the udder including the teat apex, teat canal, milk, and colostrum [116]. Some of these communities encompass archaeal members (Table 2; Figure 1). Guo and colleagues characterized the archaeal community shared between the maternal rumen and milk in grazing yak calves. They identified two archaeal phyla and 11 different common archaeal genera [60]. The primary archaeal phyla found on the skin of the teat were Euryarchaeota (76.8%) and Thaumarchaeota (23.1%) with dominant genera being Methanobrevibacter (61.7%) and Candidatus nitrocosmicus (12.5%). Similar findings have been reported for the skin of dairy cows, where Methanobrevibacter and Methanosphaera were identified as the primary genera [117]. Species within Methanobrevibacter, including M. ruminantium, M. smithii, M. millerae, and some Methanocorpusculum spp. have also been detected in the bovine milk [23,60,117]. No data is available regarding the presence of methanogenic archaea in bovine colostrum. However, Methanobrevibacter oralis and M. smithii species have been found in human colostrum [118], suggesting that methanogenic archaea may also be present in the cattle colostrum.
Similar to the respiratory tract, the presence of methanogens on teat skin and udder raises an important question regarding the strict anaerobic nature of these methanogens. Methanobrevibacter and Candidatus nitrocosmicus are known to be strict anaerobes and are typically unable to survive with exposure to ambient air for more than 10 minutes [119]. Therefore, their survival on the teat skin would be unlikely unless there is a wound present that creates an anaerobic environment suitable for methanogens. Further, research is needed to investigate the mechanisms and conditions that allow for the presence of methanogens on the skin and explore their potential role in the overall microbial ecology of the teat as related to milk production.
8. Potential Seeding Sources of the Ruminal Methanogens
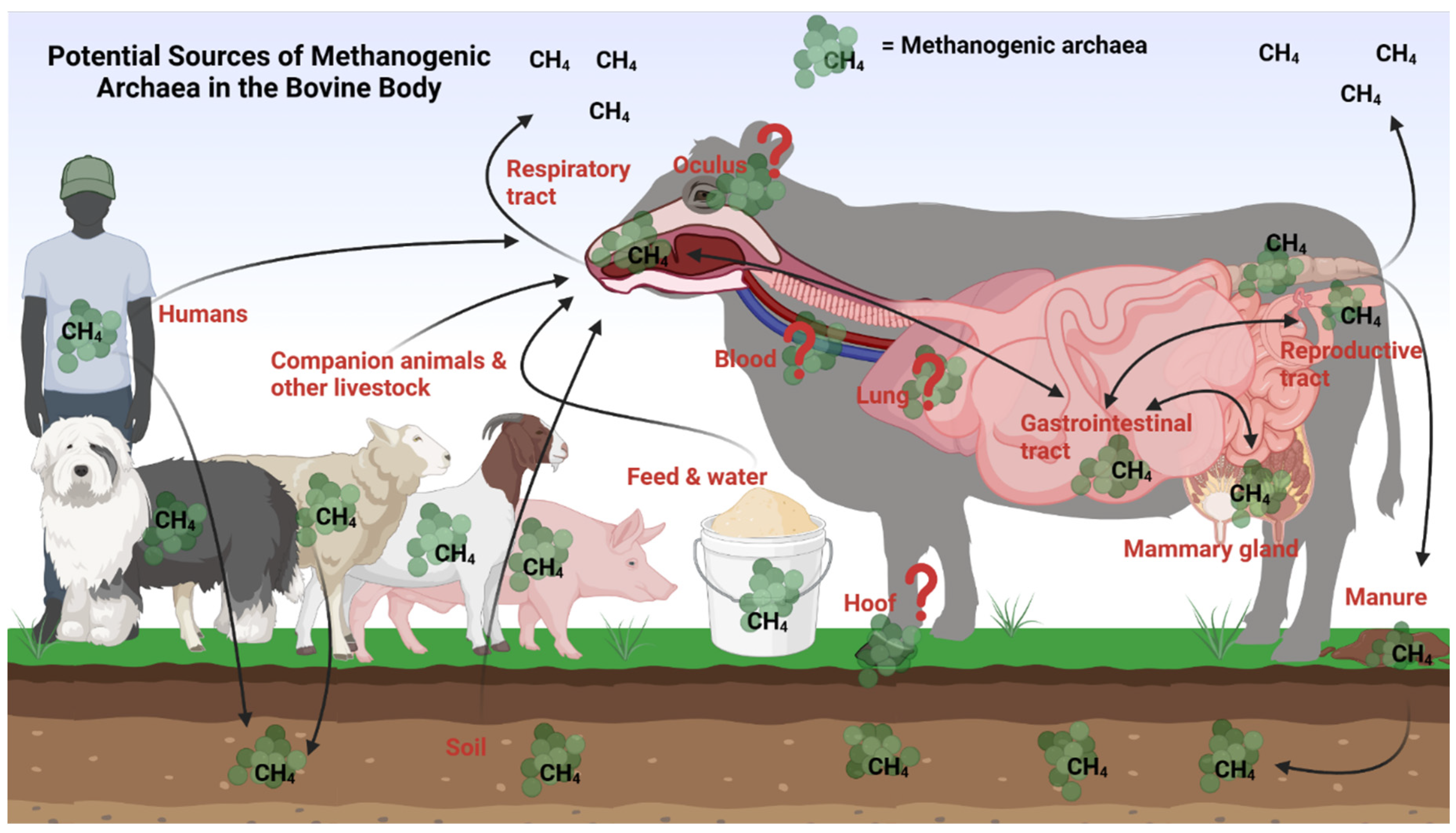
As illustrated in the Figure 2, there are multiple potential seeding sources for the ruminal methanogenic archaea in cattle, and these sources could be the microbial communities present in the different anatomical sites of bovine body, and other external sources which will be discussed in detail below.
8.1. Within the Bovine Body
Ruminal methanogens have the potential to be seeded from the reproductive system of the cow, particularly from the vagina, during calving. Our research team observed that certain methanogenic taxa are present in both the rumen and reproductive tract (vagina and uterus) of beef cattle [21,25]. This may suggest that there could be methanogenic archaea transfer between the gastrointestinal and reproductive tracts within the same animal. Likewise, the methanogenic archaeal species associated with the bovine upper respiratory tract may also be involved in introducing the rumen with methanogenic species. Our recent research revealed the presence of methanogenic taxa shared by the rumen and nasopharynx as core taxa [21,25]. Another important seeding source for the ruminal methanogens could be the udder and mammary gland of female cattle. As discussed above, methanogens can be present in bovine teat and milk. When neonatal calves are nursing, it becomes a direct route for transferring methanogens into the calf gut [86]. Additionally, bull semen harbors methanogens, and thus it is highly likely that the sperm containing methanogens traveling through the female reproductive tract could potentially introduce methanogens into the uterus and reproductive system of the female [99]. Therefore, it would not be surprising to find the same genera of methanogens in both the sperm and milk and in the vagina, uterus, and gut of animals. In addition, diverse and dynamic bacterial microbiota has been reported to be present on the ocular surface of healthy newborn calves [98] and weaned beef calves [120], and cattle hooves are also home to commensal microbiota [98,121]. These studies have not reported the presence of methanogenic archaea in bovine eye and feet. However, until the absence of the methanogens from these sites is confirmed with studies using archaeal or methanogens specific amplicon sequencing or shotgun metagenomic sequencing, the possibility of the methanogenic archaeal exchange between ocular and hoof microbial communities with the bovine gut could not be ruled out.
8.2. Other Sources
There are several external sources that could introduce cattle rumen with methanogens (Figure 2). Among which, the pasture, soil and drinking water that cattle consume may transfer methanogens to cattle rumen [59,72]. Methanogens are commonly found in muddy soils [122,123]. The common and dominant genera of the soil associated methanogens include Methanoregula, Methanobacterium, Methanosarcina, Methanolinea spp. present in wetlands and water [124,125,126]. Presence of Methanosarcina and Methanocella spp. in cattle, sheep and swine grazing land [127], wet soils and desert soils [128] have been documented. Methanosarcina spp. have been identified as the major methanogen in pasture soils compacted by cattle [22,129] and arable soils [130]. Cattle are known to consume approximately 350 kg of soil per cow per year through geophagia which refers to the act of eating soil and dirt [131,132]. Thus, it is most likely that methanogens associated with soil can be transferred into the cattle rumen. Methanogens could be transmitted to cattle via farm dust and dust particles. A study was conducted by Bønløkke and colleagues to investigate the exposure of livestock farmers to archaea[133]. For this, they analyzed the number of 16S rRNA gene copies from archaea and bacteria present in the personal filter samples obtained from 327 farmers working on 89 Danish farms including cattle and chicken farms. Both archaea and bacteria were detected in all types of farm environments. Methanobrevibacter and Methanosarcina species were found to be dominant in aerosols from both pig and cattle farms [133]. The aerosols likely consisted of a mixture of nasal fluid from farmers and other farm workers, as well as dust from the soil. When these aerosols are inhaled by cattle, they could potentially transfer methanogens into the airway systems and ultimately to the cattle rumen. Farmers and farm workers could also be a source for introducing the cattle with methanogens. In the human gut and other parts of the body harbor methanogens. Methaninobrevibacter smithii and Methanobrevibacter oralis are found in the intestine and sputum, while Methanobrevibacter smithii has been reported in bronchoalveolar [108], milk [134], and urine [87]. Methanospaera stadmagnae [135] and Methanobrevibacter oralis are associated with periodontal disease [108,136]. The methanogens present in the human body can be first transferred into the environment through spit and urine, and then ultimately to cattle.
Figure 2.
Holistic view of the bovine methanogens and potential sources of ruminal methanogens in cattle. Potential seeding sources of methanogenic archaea in the bovine body can include the soil, feed, water, farm laborers, other livestock species, or pets. Methanogens from the respiratory tract, reproductive tract, and mammary gland may seed the gastrointestinal tract, and vice versa. It is unclear if methanogens inhabit the oculus, blood, liver, lung, or hoof tissue, but the presence of other microorganisms at these sites could indicate the potential presence of methanogens here as well. Figure created using Biorender.com.
Figure 2.
Holistic view of the bovine methanogens and potential sources of ruminal methanogens in cattle. Potential seeding sources of methanogenic archaea in the bovine body can include the soil, feed, water, farm laborers, other livestock species, or pets. Methanogens from the respiratory tract, reproductive tract, and mammary gland may seed the gastrointestinal tract, and vice versa. It is unclear if methanogens inhabit the oculus, blood, liver, lung, or hoof tissue, but the presence of other microorganisms at these sites could indicate the potential presence of methanogens here as well. Figure created using Biorender.com.
Apart from bovine, other animal species (e.g., sheep, chicken, pig and dogs) raised on the same farm with cattle could inoculating cattle rumen with methanogens. Guindo et al. [137] conducted a study where fecal samples were analyzed from pigs, dogs, cats, sheep, and horses using PCR-sequencing targeted methanogens. Seven different species of methanogens including, Methanobrevibacter smithii, Methanobrevibacter millerae and Methanomassiliicoccus luminyensis, some of which are known to be present in the human digestive tract. Methanobrevibacter smithii were present in all the animal species studied [137]. Specifically, Methanobrevibacter smithii was present 50% of cases in pigs, 25% of cases in dogs, 16.7% of cases in cats, and 4.2% in both sheep and horses [137]. These findings indicate that Methanobrevibacter smithii is core archaeon shared by multiple animal species and humans, and it could be transferred between human and different animal species.
9. Challenges Associated with the Ruminal Methanogens and Future Directions
In vitro culturing techniques allow researchers to isolate and study specific microorganisms in a controlled environment and can provide insights into the physiology and ecology of these organisms. Isolation and culturing of methanogens from the rumen is a particularly challenging task due to their strict anaerobic nature and growth requirements for special culturing media, culturing apparatus, and H2 gas supplementation, all of which have hindered the progress of isolation, and metabolic and genomic characterization of the methanogenic archaea associated with ruminal CH4 production [138,139]. While recent advances in culture-independent high-throughput sequencing techniques have enabled characterization of ruminal microbiota and the potential involvement in methanogenic activity, the information is mostly limited to the bacterial population of the ruminal microbial ecosystem as most of the sequencing is based on 16S rRNA gene amplicon sequencing, which is more specific to bacteria and captures only a small fraction of the archaeal population. Thus, to understand the complete taxonomic characterization of the methanogens and other archaeal populations in the rumen, and other extra-gastrointestinal sites of the bovine body, archaea targeting amplicon sequencing such as 16S rRNA V2-V3 primers and mcrA gene-based primers should be applied. To gain insights beyond the taxonomic properties of bovine methanogens, the metabolic features, and their interactions with other microorganisms such as fungi, bacteria and viruses, shotgun metagenomic sequencing should be performed. Also, comparative genomics on methanogenic isolates from different cattle body sites (e.g., respiratory, reproductive, ruminal and udder) should be performed to understand the genomic adaptative features of the methanogens from different anatomical sites and identify the seeding sources of the ruminal methanogens. Given the several external sources including the feed, soil and water consumed by cattle, and other farm animals and farmers who could exchange methanogens with cattle rumen and other body sites, comprehensive survey of the methanogens present in these external sources and their interactions with ruminal methanogens in cattle warrants further research.
While these culture-independent sequencing methods can provide taxonomic composition and functional features of the methanogenic archaeal members and their interactions with other microbial partners in a particular niche, one of the limitations of these sequencing techniques is that they could not provide information on the viability of the methanogenic archaeal cells. Considering the anaerobic nature of methanogens and their presence in non-anaerobic body sites including respiratory and lower reproductive tracts of cattle, it is critical to use culture- dependent techniques to isolate methanogens from these sites and verify findings from metagenomic sequencing. It is plausible that some of the genomic DNA extracted from the samples associated with nasopharynx, lung tissue and vagina of cattle might be originated from transient and dead methanogens. Nevertheless, identification of methanogenic archaeal genera and species with relatively high abundance, and consistently over multiple sampling timepoints in bovine respiratory and reproductive tracts presented above highlights that the methanogenic archaea could colonize and survive aerobic mucosal surfaces along the airway and urogenital tract of cattle. Thus, further research is warranted to isolate methanogens from extra-gastrointestinal tracts of cattle and explore the survival mechanisms of the methanogens in the environment where oxygen is present. Some anaerobic bacterial species can have evolved mechanisms that can either minimize the extent to which oxygen disrupts their metabolism [140,141] and/or rely on potential anaerobic and aerobic species co-existence mechanisms [142].
Emerging evidence from humans [141], bovine fetal fluids [21,27], the fetal intestine [21,27,143], as well as the human fetal lung [144], suggests that microbial colonization of calves may begin in utero. This, coupled with rodent studies which demonstrate that fetal metabolic and nervous system development is impacted by the maternal microbiota during pregnancy [145,146], highlight the potential and extended role, of the maternal microbiome in calf microbiome development. Recent studies reported the presence of methanogenic archaeal species in fetal fluids and fetal intestine at 4-8 months of gestation. Thus, these recent developments warrant a re-consideration of the timing and mechanisms involved in the first colonization of calf ruminal methanogens.
10. Conclusions
Methanogens are responsible for enteric CH4 emissions in cattle, which contributes to a significant amount of dietary energy loss to the host and GHG emissions. The species associated with ruminal CH4 production are not only present in the cattle rumen, but they could also present in extra-gastrointestinal organs of cattle such as the reproductive and respiratory tracts, udder, eye, and hoof. Recent sequencing-based studies revealed that certain methanogenic taxa are shared by the ruminal, respiratory, and reproductive tracts of cattle as core taxa, suggesting potential interactions between the ruminal and extra-intestinal methanogens. The seeding of the rumen with methanogenic archaea could be from many possible sources within the bovine body (e.g., reproductive and respiratory tracts, and other microbial communities) and external sources such as pasture, soil, water, and farm animals. Therefore, a singular focus on the methanogens in the rumen may not be sufficient and a holistic approach which takes into consideration the transfer of methanogens between ruminal, extra-gastrointestinal, and environmental microbial communities. Taxonomic, genomic, and metabolic characteristics of the methanogenic archaeal species in the rumen and other bovine body sites should be investigated to gain more holistic insights into the methanogens in cattle. It is anticipated that the holistic understanding of the methanogens in the rumen and their interactions with the extra-gastrointestinal methanogens, and identification of the seeding sources (both within and external), and the colonization timing of the ruminal methanogens are important for the development of more effective CH4 mitigation strategies in cattle.
Author Contributions
G.A. and S.A.: conceptualizing and outlining the review. G.A. conducted the literature search. G.A. and S.A. drafted the initial manuscript. G.A., S.M.L., C.R.D., K.C.S. and S.A.: manuscript writing, revision, editing, and finalizing. All authors have read and agreed to the published version of the manuscript.
Funding
The work presented in this study was financially supported in part by the North Dakota Agricultural Experiment Station as part of a start-up package for SA, and the USDA National Institute of Food and Agriculture grant (2022-67016-37092).
Data Availability Statement
No new data were used for the research described in the article.
Acknowledgments
The authors would like to thank Dr. Robin Anderson, Research Microbiologist, at USDA-ARS College Station, Texas, United States for proofreading the manuscript and providing constructive feedback.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Bank Climate Explainer: Food Security and Climate Change. Available online: https://www.worldbank.org/en/news/feature/2022/10/17/what-you-need-to-know-about-food-security-and-climate-change (accessed on 17 May 2023).
- IPCC, I.P. on C.C. IPCC — Intergovernmental Panel on Climate Change 2022.
- Ripple, W.J.; Smith, P.; Haberl, H.; Montzka, S.A.; McAlpine, C.; Boucher, D.H. Ruminants, Climate Change and Climate Policy. Nat. Clim. Change 2014, 4, 2–5. [Google Scholar] [CrossRef]
- FAO Sustainable Agriculture | Sustainable Development Goals | Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/sustainable-development-goals/overview/fao-and-the-2030-agenda-for-sustainable-development/sustainable-agriculture/en/ (accessed on 21 February 2023).
- Clark, M.; Tilman, D. Comparative Analysis of Environmental Impacts of Agricultural Production Systems, Agricultural Input Efficiency, and Food Choice. Environ. Res. Lett. 2017, 12, 064016. [Google Scholar] [CrossRef]
- US EPA, O. Overview of Greenhouse Gases. Available online: https://www.epa.gov/ghgemissions/overview-greenhouse-gases (accessed on 20 June 2023).
- Buddle, B.M.; Denis, M.; Attwood, G.T.; Altermann, E.; Janssen, P.H.; Ronimus, R.S.; Pinares-Patiño, C.S.; Muetzel, S.; Neil Wedlock, D. Strategies to Reduce Methane Emissions from Farmed Ruminants Grazing on Pasture. Vet. J. 2011, 188, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Choudhury, P.K.; Carro, M.D.; Griffith, G.W.; Dagar, S.S.; Puniya, M.; Calabro, S.; Ravella, S.R.; Dhewa, T.; Upadhyay, R.C.; et al. New Aspects and Strategies for Methane Mitigation from Ruminants. Appl. Microbiol. Biotechnol. 2014, 98, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.; Yang, C. Ruminal Methane Production: Associated Microorganisms and the Potential of Applying Hydrogen-Utilizing Bacteria for Mitigation. Sci. Total Environ. 2019, 654, 1270–1283. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Johnson, D.E. Methane Emissions from Cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty Years of Research on Rumen Methanogenesis: Lessons Learned and Future Challenges for Mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [PubMed]
- Jeyanathan, J.; Martin, C.; Morgavi, D.P. Screening of Bacterial Direct-Fed Microbials for Their Antimethanogenic Potential in Vitro and Assessment of Their Effect on Ruminal Fermentation and Microbial Profiles in Sheep. J. Anim. Sci. 2016, 94, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.O.; Fox, D.G.; Tylutki, T.P. Potential Environmental Benefits of Ionophores in Ruminant Diets. J. Environ. Qual. 2003, 32, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Muñoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M.; et al. Addressing Global Ruminant Agricultural Challenges Through Understanding the Rumen Microbiome: Past, Present, and Future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Króliczewska, B.; Pecka-Kiełb, E.; Bujok, J. Strategies Used to Reduce Methane Emissions from Ruminants: Controversies and Issues. Agriculture 2023, 13, 602. [Google Scholar] [CrossRef]
- Malik, P.K.; Bhatta, R.; Gagen, E.J.; Sejian, V.; Soren, N.M.; Prasad, C.S. Alternate H2 Sinks for Reducing Rumen Methanogenesis. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C., Eds.; Springer India: New Delhi, 2015; pp. 303–320. ISBN 978-81-322-2265-1. [Google Scholar]
- de Haas, Y.; Veerkamp, R.F.; de Jong, G.; Aldridge, M.N. Selective Breeding as a Mitigation Tool for Methane Emissions from Dairy Cattle. Animal 2021, 15, 100294. [Google Scholar] [CrossRef] [PubMed]
- Jalil Sarghale, A.; Moradi Shahrebabak, M.; Moradi Shahrebabak, H.; Nejati Javaremi, A.; Saatchi, M.; Khansefid, M.; Miar, Y. Genome-Wide Association Studies for Methane Emission and Ruminal Volatile Fatty Acids Using Holstein Cattle Sequence Data. BMC Genet. 2020, 21, 129. [Google Scholar] [CrossRef]
- Zetouni, L.; Kargo, M.; Norberg, E.; Lassen, J. Genetic Correlations between Methane Production and Fertility, Health, and Body Type Traits in Danish Holstein Cows. J. Dairy Sci. 2018, 101, 2273–2280. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, G. Decreasing Ruminal Methane Production through Enhancing the Sulfate Reduction Pathway. Anim. Nutr. 2022, 9, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Amat, S.; Holman, D.B.; Schmidt, K.; Menezes, A.C.B.; Baumgaertner, F.; Winders, T.; Kirsch, J.D.; Liu, T.; Schwinghamer, T.D.; Sedivec, K.K.; et al. The Nasopharyngeal, Ruminal, and Vaginal Microbiota and the Core Taxa Shared across These Microbiomes in Virgin Yearling Heifers Exposed to Divergent In Utero Nutrition during Their First Trimester of Gestation and in Pregnant Beef Heifers in Response to Mineral Supplementation. Microorganisms 2021, 9, 2011. [Google Scholar] [CrossRef]
- Kroeger, M.E.; Meredith, L.K.; Meyer, K.M.; Webster, K.D.; de Camargo, P.B.; de Souza, L.F.; Tsai, S.M.; van Haren, J.; Saleska, S.; Bohannan, B.J.M.; et al. Rainforest-to-Pasture Conversion Stimulates Soil Methanogenesis across the Brazilian Amazon. ISME J. 2021, 15, 658–672. [Google Scholar] [CrossRef]
- Guindo, C.O.; Drancourt, M.; Grine, G. Dairy Products as Sources of Methanogens for Humans 2021, 2021. 11.16.46 8822.
- de Souza, J.; Leskinen, H.; Lock, A.L.; Shingfield, K.J.; Huhtanen, P. Between-Cow Variation in Milk Fatty Acids Associated with Methane Production. PLoS ONE 2020, 15, e0235357. [Google Scholar] [CrossRef]
- Winders, T.M.; Holman, D.B.; Schmidt, K.N.; Luecke, S.M.; Smith, D.J.; Neville, B.W.; Dahlen, C.R.; Swanson, K.C.; Amat, S. Feeding Hempseed Cake Alters the Bovine Gut, Respiratory and Reproductive Microbiota. Sci. Rep. 2023, 13, 8121. [Google Scholar] [CrossRef]
- Welch, C.B.; Ryman, V.E.; Pringle, T.D.; Lourenco, J.M. Utilizing the Gastrointestinal Microbiota to Modulate Cattle Health through the Microbiome-Gut-Organ Axes. Microorganisms 2022, 10, 1391. [Google Scholar] [CrossRef] [PubMed]
- Guzman, C.E.; Wood, J.L.; Egidi, E.; White-Monsant, A.C.; Semenec, L.; Grommen, S.V.H.; Hill-Yardin, E.L.; De Groef, B.; Franks, A.E. A Pioneer Calf Foetus Microbiome. Sci. Rep. 2020, 10, 17712. [Google Scholar] [CrossRef] [PubMed]
- Buan, N.R. Methanogens: Pushing the Boundaries of Biology. Emerg. Top. Life Sci. 2018, 2, 629–646. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Rotaru, A.-E.; Pimentel, M.; Zhang, C.-J.; Rittmann, S.K.-M.R. Editorial: The Methane Moment - Cross-Boundary Significance of Methanogens: Preface. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.M.; de Lurdes Nunes Enes Dapkevicius, M.; Borba, A.E.S. Alternative Pathways for Hydrogen Sink Originated from the Ruminal Fermentation of Carbohydrates: Which Microorganisms Are Involved in Lowering Methane Emission? Anim. Microbiome 2022, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Taxonomy of Methanogens. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin, Heidelberg, 2010; pp. 547–558. ISBN 978-3-540-77587-4. [Google Scholar]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.; Castel, V.; Rosales, M.; de Haan, C. Livestock’s Long Shadow. 2006. [Google Scholar]
- Gribaldo, S.; Brochier-Armanet, C. The Origin and Evolution of Archaea: A State of the Art. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1007–1022. [Google Scholar] [CrossRef]
- Lyu, Z.; Liu, Y. Diversity and Taxonomy of Methanogens. In Biogenesis of Hydrocarbons; Stams, A.J.M., Sousa, D.Z., Eds.; Handbook of Hydrocarbon and Lipid Microbiology; Springer International Publishing: Cham, 2019; pp. 19–77. ISBN 978-3-319-78108-2. [Google Scholar]
- DasSarma, S.; Coker, J.A.; DasSarma, P. Archaea (Overview). In Encyclopedia of Microbiology (Third Edition); Schaechter, M., Ed.; Academic Press: Oxford, 2009; pp. 1–23. ISBN 978-0-12-373944-5. [Google Scholar]
- Angelidaki, I.; Karakashev, D.; Batstone, D.J.; Plugge, C.M.; Stams, A.J.M. Chapter Sixteen - Biomethanation and Its Potential. In Methods in Enzymology; Rosenzweig, A.C., Ragsdale, S.W., Eds.; Methods in Methane Metabolism, Part A; Academic Press, 2011; Vol. 494, pp. 327–351. [Google Scholar]
- Misiukiewicz, A.; Gao, M.; Filipiak, W.; Cieslak, A.; Patra, A.K.; Szumacher-Strabel, M. Review: Methanogens and Methane Production in the Digestive Systems of Nonruminant Farm Animals. Anim. Int. J. Anim. Biosci. 2021, 15, 100060. [Google Scholar] [CrossRef]
- Prasitwuttisak, W.; Hoshiko, Y.; Maeda, T.; Haraguchi, A.; Yanagawa, K. Microbial Community Structures and Methanogenic Functions in Wetland Peat Soils. Microbes Environ. 2022, 37, ME22004. [Google Scholar] [CrossRef]
- Lewis, W.H.; Tahon, G.; Geesink, P.; Sousa, D.Z.; Ettema, T.J.G. Innovations to Culturing the Uncultured Microbial Majority. Nat. Rev. Microbiol. 2021, 19, 225–240. [Google Scholar] [CrossRef]
- Tahon, G.; Geesink, P.; Ettema, T.J.G. Expanding Archaeal Diversity and Phylogeny: Past, Present, and Future. Annu. Rev. Microbiol. 2021, 75, 359–381. [Google Scholar] [CrossRef]
- Cheng, L.; Wu, K.; Zhou, L.; Tahon, G.; Liu, L.; Li, J.; Zhang, J.; Zheng, F.; Deng, C.; Han, W.; et al. Isolation of a Methyl-Reducing Methanogen Outside the Euryarchaeota; In Review; 2023. [Google Scholar]
- Mei, R.; Kaneko, M.; Imachi, H.; Nobu, M.K. The Origin and Evolution of Methanogenesis and Archaea Are Intertwined. PNAS Nexus 2023, 2, pgad023. [Google Scholar] [CrossRef]
- Martínez-Espinosa, R.M. Microorganisms and Their Metabolic Capabilities in the Context of the Biogeochemical Nitrogen Cycle at Extreme Environments. Int. J. Mol. Sci. 2020, 21, 4228. [Google Scholar] [CrossRef]
- Berghuis, B.A.; Yu, F.B.; Schulz, F.; Blainey, P.C.; Woyke, T.; Quake, S.R. Hydrogenotrophic Methanogenesis in Archaeal Phylum Verstraetearchaeota Reveals the Shared Ancestry of All Methanogens. Proc. Natl. Acad. Sci. 2019, 116, 5037–5044. [Google Scholar] [CrossRef]
- Vanwonterghem, I.; Evans, P.N.; Parks, D.H.; Jensen, P.D.; Woodcroft, B.J.; Hugenholtz, P.; Tyson, G.W. Methylotrophic Methanogenesis Discovered in the Archaeal Phylum Verstraetearchaeota. Nat. Microbiol. 2016, 1, 1–9. [Google Scholar] [CrossRef]
- Gaci, N.; Borrel, G.; Tottey, W.; O’Toole, P.W.; Brugère, J.-F. Archaea and the Human Gut: New Beginning of an Old Story. World J. Gastroenterol. WJG 2014, 20, 16062–16078. [Google Scholar] [CrossRef]
- Ferry, J.G.; Kastead, K.A. Methanogenesis. In Archaea; John Wiley & Sons, Ltd., 2007; pp. 288–314. ISBN 978-1-68367-168-8. [Google Scholar]
- Kurade, M.B.; Saha, S.; Salama, E.-S.; Patil, S.M.; Govindwar, S.P.; Jeon, B.-H. Acetoclastic Methanogenesis Led by Methanosarcina in Anaerobic Co-Digestion of Fats, Oil and Grease for Enhanced Production of Methane. Bioresour. Technol. 2019, 272, 351–359. [Google Scholar] [CrossRef]
- Noel, S.J.; Højberg, O.; Urich, T.; Poulsen, M. Draft Genome Sequence of “Candidatus Methanomethylophilus” Sp. 1R26, Enriched from Bovine Rumen, a Methanogenic Archaeon Belonging to the Methanomassiliicoccales Order. Genome Announc. 2016, 4, e01734-15. [Google Scholar] [CrossRef]
- Knief, C. Diversity of Methane Cycling Microorganisms in Soils and Their Relation to Oxygen. Curr. Issues Mol. Biol. 2019, 33, 23–56. [Google Scholar] [CrossRef]
- Narrowe, A.B.; Borton, M.A.; Hoyt, D.W.; Smith, G.J.; Daly, R.A.; Angle, J.C.; Eder, E.K.; Wong, A.R.; Wolfe, R.A.; Pappas, A.; et al. Uncovering the Diversity and Activity of Methylotrophic Methanogens in Freshwater Wetland Soils. mSystems 2019, 4, e00320-19. [Google Scholar] [CrossRef]
- Smith, G.J.; Angle, J.C.; Solden, L.M.; Borton, M.A.; Morin, T.H.; Daly, R.A.; Johnston, M.D.; Stefanik, K.C.; Wolfe, R.; Gil, B.; et al. Members of the Genus Methylobacter Are Inferred To Account for the Majority of Aerobic Methane Oxidation in Oxic Soils from a Freshwater Wetland. mBio 2018, 9, e00815-18. [Google Scholar] [CrossRef]
- Greening, C.; Geier, R.; Wang, C.; Woods, L.C.; Morales, S.E.; McDonald, M.J.; Rushton-Green, R.; Morgan, X.C.; Koike, S.; Leahy, S.C.; et al. Diverse Hydrogen Production and Consumption Pathways Influence Methane Production in Ruminants. ISME J. 2019, 13, 2617–2632. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.N.; Boyd, J.A.; Leu, A.O.; Woodcroft, B.J.; Parks, D.H.; Hugenholtz, P.; Tyson, G.W. An Evolving View of Methane Metabolism in the Archaea. Nat. Rev. Microbiol. 2019, 17, 219–232. [Google Scholar] [CrossRef]
- Enzmann, F.; Mayer, F.; Rother, M.; Holtmann, D. Methanogens: Biochemical Background and Biotechnological Applications. AMB Express 2018, 8, 1. [Google Scholar] [CrossRef]
- Tapio, I.; Snelling, T.J.; Strozzi, F.; Wallace, R.J. The Ruminal Microbiome Associated with Methane Emissions from Ruminant Livestock. J. Anim. Sci. Biotechnol. 2017, 8, 7. [Google Scholar] [CrossRef]
- Hoedt, E.C.; Parks, D.H.; Volmer, J.G.; Rosewarne, C.P.; Denman, S.E.; McSweeney, C.S.; Muir, J.G.; Gibson, P.R.; Cuív, P.Ó.; Hugenholtz, P.; et al. Culture- and Metagenomics-Enabled Analyses of the Methanosphaera Genus Reveals Their Monophyletic Origin and Differentiation According to Genome Size. ISME J. 2018, 12, 2942–2953. [Google Scholar] [CrossRef] [PubMed]
- Poehlein, A.; Schneider, D.; Soh, M.; Daniel, R.; Seedorf, H. Comparative Genomic Analysis of Members of the Genera Methanosphaera and Methanobrevibacter Reveals Distinct Clades with Specific Potential Metabolic Functions. Archaea 2018, 2018, 7609847. [Google Scholar] [CrossRef]
- Guzman, C.E.; Bereza-Malcolm, L.T.; Groef, B.D.; Franks, A.E. Presence of Selected Methanogens, Fibrolytic Bacteria, and Proteobacteria in the Gastrointestinal Tract of Neonatal Dairy Calves from Birth to 72 Hours. PLOS ONE 2015, 10, e0133048. [Google Scholar] [CrossRef]
- Guo, W.; Bi, S.S.; Wang, W.W.; Zhou, M.; Neves, A.L.A.; Degen, A.A.; Guan, L.L.; Long, R.J. Maternal Rumen and Milk Microbiota Shape the Establishment of Early-Life Rumen Microbiota in Grazing Yak Calves. J. Dairy Sci. 2023, 106, 2054–2070. [Google Scholar] [CrossRef]
- Janssen, P.H. Influence of Hydrogen on Rumen Methane Formation and Fermentation Balances through Microbial Growth Kinetics and Fermentation Thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Gunsalus, R.P.; Cook, L.E.; Crable, B.; Rohlin, L.; McDonald, E.; Mouttaki, H.; Sieber, J.R.; Poweleit, N.; Zhou, H.; Lapidus, A.L.; et al. Complete Genome Sequence of Methanospirillum Hungatei Type Strain JF1. Stand. Genomic Sci. 2016, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, K.; Chang, C.; Pimentel, M. Methanogens, Methane and Gastrointestinal Motility. J. Neurogastroenterol. Motil. 2014, 20, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Schink, B. Energetics of Syntrophic Cooperation in Methanogenic Degradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280. [Google Scholar] [PubMed]
- Hao, L.; Michaelsen, T.Y.; Singleton, C.M.; Dottorini, G.; Kirkegaard, R.H.; Albertsen, M.; Nielsen, P.H.; Dueholm, M.S. Novel Syntrophic Bacteria in Full-Scale Anaerobic Digesters Revealed by Genome-Centric Metatranscriptomics. ISME J. 2020, 14, 906–918. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, B.; Chávez, A.; Forero, A.; García-Huante, Y.; Romero, A.; Sánchez, M.; Rocha, D.; Sánchez, B.; Rodríguez-Sanoja, R.; Sánchez, S.; et al. Production of Microbial Secondary Metabolites: Regulation by the Carbon Source. Crit. Rev. Microbiol. 2010, 36, 146–167. [Google Scholar] [CrossRef] [PubMed]
- Rotaru, A.-E.; Calabrese, F.; Stryhanyuk, H.; Musat, F.; Shrestha, P.M.; Weber, H.S.; Snoeyenbos-West, O.L.O.; Hall, P.O.J.; Richnow, H.H.; Musat, N.; et al. Conductive Particles Enable Syntrophic Acetate Oxidation between Geobacter and Methanosarcina from Coastal Sediments. mBio 2018, 9, e00226-18. [Google Scholar] [CrossRef]
- Hinsley, A.P.; Berks, B.C. Specificity of Respiratory Pathways Involved in the Reduction of Sulfur Compounds by Salmonella Enterica. Microbiol. Read. Engl. 2002, 148, 3631–3638. [Google Scholar] [CrossRef]
- van Zijderveld, S.M.; Gerrits, W.J.J.; Apajalahti, J.A.; Newbold, J.R.; Dijkstra, J.; Leng, R.A.; Perdok, H.B. Nitrate and Sulfate: Effective Alternative Hydrogen Sinks for Mitigation of Ruminal Methane Production in Sheep. J. Dairy Sci. 2010, 93, 5856–5866. [Google Scholar] [CrossRef]
- Granja-Salcedo, Y.T.; Fernandes, R.M.; de Araujo, R.C.; Kishi, L.T.; Berchielli, T.T.; de Resende, F.D.; Berndt, A.; Siqueira, G.R. Long-Term Encapsulated Nitrate Supplementation Modulates Rumen Microbial Diversity and Rumen Fermentation to Reduce Methane Emission in Grazing Steers. Front. Microbiol. 2019, 10, 614. [Google Scholar] [CrossRef]
- Doyle, N.; Mbandlwa, P.; Kelly, W.J.; Attwood, G.; Li, Y.; Ross, R.P.; Stanton, C.; Leahy, S. Use of Lactic Acid Bacteria to Reduce Methane Production in Ruminants, a Critical Review. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Danielsson, R.; Dicksved, J.; Sun, L.; Gonda, H.; Müller, B.; Schnürer, A.; Bertilsson, J. Methane Production in Dairy Cows Correlates with Rumen Methanogenic and Bacterial Community Structure. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Ramayo-Caldas, Y.; Zingaretti, L.; Popova, M.; Estellé, J.; Bernard, A.; Pons, N.; Bellot, P.; Mach, N.; Rau, A.; Roume, H.; et al. Identification of Rumen Microbial Biomarkers Linked to Methane Emission in Holstein Dairy Cows. J. Anim. Breed. Genet. 2019. [Google Scholar] [CrossRef]
- Pope, P.B.; Smith, W.; Denman, S.E.; Tringe, S.G.; Barry, K.; Hugenholtz, P.; McSweeney, C.S.; McHardy, A.C.; Morrison, M. Isolation of Succinivibrionaceae Implicated in Low Methane Emissions from Tammar Wallabies. Science 2011, 333, 646–648. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, P.K.; Jena, R.; Tomar, S.K.; Puniya, A.K. Reducing Enteric Methanogenesis through Alternate Hydrogen Sinks in the Rumen. Methane 2022, 1, 320–341. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Huang, X.; Ma, R.; Li, J.; Wang, F.; Jiao, N.; Zhang, R. Potential Metabolic and Genetic Interaction among Viruses, Methanogen and Methanotrophic Archaea, and Their Syntrophic Partners. ISME Commun. 2022, 2, 1–10. [Google Scholar] [CrossRef]
- Namonyo, S.; Wagacha, M.; Maina, S.; Wambua, L.; Agaba, M. A Metagenomic Study of the Rumen Virome in Domestic Caprids. Arch. Virol. 2018, 163, 3415–3419. [Google Scholar] [CrossRef] [PubMed]
- Pfister, P.; Wasserfallen, A.; Stettler, R.; Leisinger, T. Molecular Analysis of Methanobacterium Phage ΨM2. Mol. Microbiol. 1998, 30, 233–244. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic Bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef]
- Auffret, M.; Stewart, R.; Dewhurst, R.; Duthie, C.-A.; Rooke, J.; Wallace, R.; Freeman, T.; Snelling, T.; Watson, M.; Roehe, R. Identification, Comparison and Validation of Robust Rumen Microbial Biomarkers for Methane Emissions Using Diverse Bos Taurus Breeds and Basal Diets. Front. Microbiol. 2018, 8. [Google Scholar] [CrossRef]
- Li, Y.; Meng, Z.; Xu, Y.; Shi, Q.; Ma, Y.; Aung, M.; Cheng, Y.; Zhu, W. Interactions between Anaerobic Fungi and Methanogens in the Rumen and Their Biotechnological Potential in Biogas Production from Lignocellulosic Materials. Microorganisms 2021, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The Rumen Microbiome: A Crucial Consideration When Optimising Milk and Meat Production and Nitrogen Utilisation Efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef]
- Luecke, S.M.; Webb, E.M.; Dahlen, C.R.; Reynolds, L.P.; Amat, S. Seminal and Vagino-Uterine Microbiome and Their Individual and Interactive Effects on Cattle Fertility. Front. Microbiol. 2022, 13, 1029128. [Google Scholar] [CrossRef]
- Adnane, M.; Chapwanya, A. A Review of the Diversity of the Genital Tract Microbiome and Implications for Fertility of Cattle. Anim. Open Access J. MDPI 2022, 12, 460. [Google Scholar] [CrossRef]
- Laguardia-Nascimento, M.; Branco, K.M.G.R.; Gasparini, M.R.; Giannattasio-Ferraz, S.; Leite, L.R.; Araujo, F.M.G.; Salim, A.C. de M.; Nicoli, J.R.; Oliveira, G.C. de; Barbosa-Stancioli, E.F. Vaginal Microbiome Characterization of Nellore Cattle Using Metagenomic Analysis. PLOS ONE 2015, 10, e0143294. [Google Scholar] [CrossRef]
- Belay, N.; Mukhopadhyay, B.; Conway de Macario, E.; Galask, R.; Daniels, L. Methanogenic Bacteria in Human Vaginal Samples. J. Clin. Microbiol. 1990, 28, 1666–1668. [Google Scholar] [CrossRef] [PubMed]
- Grine, G.; Drouet, H.; Fenollar, F.; Bretelle, F.; Raoult, D.; Drancourt, M. Detection of Methanobrevibacter Smithii in Vaginal Samples Collected from Women Diagnosed with Bacterial Vaginosis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1643–1649. [Google Scholar] [CrossRef] [PubMed]
- Khelaifia, S.; Raoult, D.; Drancourt, M. A Versatile Medium for Cultivating Methanogenic Archaea. PLoS ONE 2013, 8, e61563. [Google Scholar] [CrossRef] [PubMed]
- Swartz, J.D.; Lachman, M.; Westveer, K.; O’Neill, T.; Geary, T.; Kott, R.W.; Berardinelli, J.G.; Hatfield, P.G.; Thomson, J.M.; Roberts, A.; et al. Characterization of the Vaginal Microbiota of Ewes and Cows Reveals a Unique Microbiota with Low Levels of Lactobacilli and Near-Neutral pH. Front. Vet. Sci. 2014, 1. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Wang, W.; Gu, S.; Gu, T.; Yang, H.; Zheng, E.; Xu, Z.; Huang, S.; Li, Z.; Cai, G.; et al. Interaction between Microbes and Host in Sow Vaginas in Early Pregnancy. mSystems 2023, 0, e01192-22. [Google Scholar] [CrossRef]
- Abou Chacra, L.; Fenollar, F. Exploring the Global Vaginal Microbiome and Its Impact on Human Health. Microb. Pathog. 2021, 160, 105172. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Li, H.; Fu, K.; Pang, B.; Yang, Y.; Liu, Y.; Tian, W.; Cao, R. Characterization of the Cervical Bacterial Community in Dairy Cows with Metritis and during Different Physiological Phases. Theriogenology 2018, 108, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Galvão, K.N.; Bicalho, R.C.; Jeon, S.J. Symposium Review: The Uterine Microbiome Associated with the Development of Uterine Disease in Dairy Cows. J. Dairy Sci. 2019, 102, 11786–11797. [Google Scholar] [CrossRef]
- Ault-Seay, T.B.; Moorey, S.E.; Mathew, D.J.; Schrick, F.N.; Pohler, K.G.; McLean, K.J.; Myer, P.R. Importance of the Female Reproductive Tract Microbiome and Its Relationship with the Uterine Environment for Health and Productivity in Cattle: A Review. Front. Anim. Sci. 2023, 4. [Google Scholar] [CrossRef]
- Pascottini, O.B.; Van Schyndel, S.J.; Spricigo, J.F.W.; Rousseau, J.; Weese, J.S.; LeBlanc, S.J. Dynamics of Uterine Microbiota in Postpartum Dairy Cows with Clinical or Subclinical Endometritis. Sci. Rep. 2020, 10, 12353. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.M.; Holman, D.B.; Schmidt, K.N.; Pun, B.; Sedivec, K.K.; Hurlbert, J.L.; Bochantin, K.A.; Ward, A.K.; Dahlen, C.R.; Amat, S. Sequencing and Culture-Based Characterization of the Vaginal and Uterine Microbiota in Beef Cattle That Became Pregnant or Non-Pregnant via Artificial Insemination 2023, 2023. 07.31.55 1361.
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen Methanogens and Mitigation of Methane Emission by Anti-Methanogenic Compounds and Substances. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef]
- Luecke, S.M.; Holman, D.B.; Schmidt, K.N.; Gzyl, K.E.; Hurlbert, J.L.; Menezes, A.C.B.; Bochantin, K.A.; Kirsch, J.D.; Baumgaertner, F.; Sedivec, K.K.; et al. Whole-Body Microbiota of Newborn Calves and Their Response to Prenatal Vitamin and Mineral Supplementation. 2023. [Google Scholar]
- Koziol, J.H.; Sheets, T.; Wickware, C.L.; Johnson, T.A. Composition and Diversity of the Seminal Microbiota in Bulls and Its Association with Semen Parameters. Theriogenology 2022, 182, 17–25. [Google Scholar] [CrossRef]
- Cojkic, A.; Niazi, A.; Guo, Y.; Hallap, T.; Padrik, P.; Morrell, J.M. Identification of Bull Semen Microbiome by 16S Sequencing and Possible Relationships with Fertility. Microorganisms 2021, 9, 2431. [Google Scholar] [CrossRef]
- Webb, E.M.; Holman, D.B.; Schmidt, K.N.; Crouse, M.S.; Dahlen, C.R.; Cushman, R.A.; Snider, A.P.; McCarthy, K.L.; Amat, S. A Longitudinal Characterization of the Seminal Microbiota and Antibiotic Resistance in Yearling Beef Bulls Subjected to Different Rates of Gain. Microbiol. Spectr. 2023, 11, e0518022. [Google Scholar] [CrossRef] [PubMed]
- Alexander, T.W.; Timsit, E.; Amat, S. The Role of the Bovine Respiratory Bacterial Microbiota in Health and Disease. Anim. Health Res. Rev. 2020, 21, 168–171. [Google Scholar] [CrossRef] [PubMed]
- McMullen, C.; Alexander, T.W.; Orsel, K.; Timsit, E. Progression of Nasopharyngeal and Tracheal Bacterial Microbiotas of Feedlot Cattle during Development of Bovine Respiratory Disease. Vet. Microbiol. 2020, 248, 108826. [Google Scholar] [CrossRef]
- Timsit, E.; McMullen, C.; Amat, S.; Alexander, T.W. Respiratory Bacterial Microbiota in Cattle: From Development to Modulation to Enhance Respiratory Health. Vet. Clin. North Am. Food Anim. Pract. 2020, 36, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Zeineldin, M.; Lowe, J.; Aldridge, B. Contribution of the Mucosal Microbiota to Bovine Respiratory Health. Trends Microbiol. 2019, 27, 753–770. [Google Scholar] [CrossRef]
- Amat, S.; Holman, D.B.; Timsit, E.; Schwinghamer, T.; Alexander, T.W. Evaluation of the Nasopharyngeal Microbiota in Beef Cattle Transported to a Feedlot, With a Focus on Lactic Acid-Producing Bacteria. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Timsit, E.; Hallewell, J.; Booker, C.; Tison, N.; Amat, S.; Alexander, T.W. Prevalence and Antimicrobial Susceptibility of Mannheimia Haemolytica, Pasteurella Multocida, and Histophilus Somni Isolated from the Lower Respiratory Tract of Healthy Feedlot Cattle and Those Diagnosed with Bovine Respiratory Disease. Vet. Microbiol. 2017, 208, 118–125. [Google Scholar] [CrossRef]
- Hassani, Y.; Brégeon, F.; Aboudharam, G.; Drancourt, M.; Grine, G. Detection of Methanobrevobacter Smithii and Methanobrevibacter Oralis in Lower Respiratory Tract Microbiota. Microorganisms 2020, 8, 1866. [Google Scholar] [CrossRef]
- McMullen, C.; Orsel, K.; Alexander, T.W.; van der Meer, F.; Plastow, G.; Timsit, E. Evolution of the Nasopharyngeal Bacterial Microbiota of Beef Calves from Spring Processing to 40 Days after Feedlot Arrival. Vet. Microbiol. 2018, 225, 139–148. [Google Scholar] [CrossRef]
- Kedzia, A.; Kwapisz, E.; Wierzbowska, M. [Incidence of anaerobic bacteria in respiratory tract infections]. Pneumonol. Alergol. Pol. 2003, 71, 68–73. [Google Scholar]
- Alsayed, A.; Al-Doori, A.; Al-Dulaimi, A.; Alnaseri, A.; Abuhashish, J.; Aliasin, K.; Alfayoumi, I. Influences of Bovine Colostrum on Nasal Swab Microbiome and Viral Upper Respiratory Tract Infections - A Case Report. Respir. Med. Case Rep. 2020, 31, 101189. [Google Scholar] [CrossRef]
- Centeno-Martinez, R.E.; Glidden, N.; Mohan, S.; Davidson, J.L.; Fernández-Juricic, E.; Boerman, J.P.; Schoonmaker, J.; Pillai, D.; Koziol, J.; Ault, A.; et al. Identification of Bovine Respiratory Disease through the Nasal Microbiome. Anim. Microbiome 2022, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’toole, G.A. Microbial Biofilms: From Ecology to Molecular Genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond Risk: Bacterial Biofilms and Their Regulating Approaches. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Spoering, A.L.; Lewis, K. Biofilms and Planktonic Cells of Pseudomonas Aeruginosa Have Similar Resistance to Killing by Antimicrobials. J. Bacteriol. 2001, 183, 6746–6751. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, H.; Fehr, K.B.; Sepehri, S.; Francoz, D.; De Buck, J.; Barkema, H.W.; Plaizier, J.C.; Khafipour, E. Invited Review: Microbiota of the Bovine Udder: Contributing Factors and Potential Implications for Udder Health and Mastitis Susceptibility. J. Dairy Sci. 2018, 101, 10605–10625. [Google Scholar] [CrossRef] [PubMed]
- Ekman, L.; Bagge, E.; Nyman, A.; Waller, K.P.; Pringle, M.; Segerman, B. A Shotgun Metagenomic Investigation of the Microbiota of Udder Cleft Dermatitis in Comparison to Healthy Skin in Dairy Cows. PLOS ONE 2020, 15, e0242880. [Google Scholar] [CrossRef] [PubMed]
- Togo, A.H.; Grine, G.; Khelaifia, S.; des Robert, C.; Brevaut, V.; Caputo, A.; Baptiste, E.; Bonnet, M.; Levasseur, A.; Drancourt, M.; et al. Culture of Methanogenic Archaea from Human Colostrum and Milk. Sci. Rep. 2019, 9, 18653. [Google Scholar] [CrossRef]
- Drancourt, M.; Djemai, K.; Gouriet, F.; Grine, G.; Loukil, A.; Bedotto, M.; Levasseur, A.; Lepidi, H.; Bou-Khalil, J.; Khelaifia, S.; et al. Methanobrevibacter Smithii Archaemia in Febrile Patients With Bacteremia, Including Those With Endocarditis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 73, e2571–e2579. [Google Scholar] [CrossRef]
- Bartenslager, A.C.; Althuge, N.D.; Loy, J.D.; Hille, M.M.; Spangler, M.L.; Fernando, S.C. Longitudinal Assessment of the Bovine Ocular Bacterial Community Dynamics in Calves. Anim. Microbiome 2021, 3, 16. [Google Scholar] [CrossRef]
- Caddey, B.; Orsel, K.; Naushad, S.; Derakhshani, H.; De Buck, J. Identification and Quantification of Bovine Digital Dermatitis-Associated Microbiota across Lesion Stages in Feedlot Beef Cattle. mSystems 2021, 6, e00708-21. [Google Scholar] [CrossRef]
- Guerrero-Cruz, S.; Vaksmaa, A.; Horn, M.A.; Niemann, H.; Pijuan, M.; Ho, A. Methanotrophs: Discoveries, Environmental Relevance, and a Perspective on Current and Future Applications. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Wu, D.; Zhao, C.; Bai, H.; Feng, F.; Sui, X.; Sun, G. Characteristics and Metabolic Patterns of Soil Methanogenic Archaea Communities in the High-latitude Natural Forested Wetlands of China. Ecol. Evol. 2021, 11, 10396–10408. [Google Scholar] [CrossRef] [PubMed]
- Ayotte, J.B.; Parker, K.L.; Gillingham, M.P. Use of Natural Licks by Four Species of Ungulates in Northern British Columbia. J. Mammal. 2008, 89, 1041–1050. [Google Scholar] [CrossRef]
- Mahaney, W.C.; Krishnamani, R. Understanding Geophagy in Animals: Standard Procedures for Sampling Soils. J. Chem. Ecol. 2003, 29, 1503–1523. [Google Scholar] [CrossRef]
- Tong, C.; Cadillo-Quiroz, H.; Zeng, Z.H.; She, C.X.; Yang, P.; Huang, J.F. Changes of Community Structure and Abundance of Methanogens in Soils along a Freshwater–Brackish Water Gradient in Subtropical Estuarine Marshes. Geoderma 2017, 299, 101–110. [Google Scholar] [CrossRef]
- Mhuireach, G.Á.; Dietz, L.; Gillett, T. One or Many? Multi-Species Livestock Grazing Influences Soil Microbiome Community Structure and Antibiotic Resistance Potential. Front. Sustain. Food Syst. 2022, 6. [Google Scholar] [CrossRef]
- Angel, R.; Claus, P.; Conrad, R. Methanogenic Archaea Are Globally Ubiquitous in Aerated Soils and Become Active under Wet Anoxic Conditions. ISME J. 2012, 6, 847–862. [Google Scholar] [CrossRef]
- Radl, V.; Gattinger, A.; Chroňáková, A.; Němcová, A.; Čuhel, J.; Šimek, M.; Munch, J.C.; Schloter, M.; Elhottová, D. Effects of Cattle Husbandry on Abundance and Activity of Methanogenic Archaea in Upland Soils. ISME J. 2007, 1, 443–452. [Google Scholar] [CrossRef]
- Gattinger, A.; Höfle, M.G.; Schloter, M.; Embacher, A.; Böhme, F.; Munch, J.C.; Labrenz, M. Traditional Cattle Manure Application Determines Abundance, Diversity and Activity of Methanogenic Archaea in Arable European Soil. Environ. Microbiol. 2007, 9, 612–624. [Google Scholar] [CrossRef]
- Healy, W.B. Ingestion of Soil by Dairy Cows. N. Z. J. Agric. Res. 1968, 11, 487–499. [Google Scholar] [CrossRef]
- Jurjanz, S.; Feidt, C.; Pérez-Prieto, L.A.; Ribeiro Filho, H.M.N.; Rychen, G.; Delagarde, R. Soil Intake of Lactating Dairy Cows in Intensive Strip Grazing Systems. Anim. Int. J. Anim. Biosci. 2012, 6, 1350–1359. [Google Scholar] [CrossRef]
- Bønløkke, J.H.; Duchaine, C.; Schlünssen, V.; Sigsgaard, T.; Veillette, M.; Basinas, I. Archaea and Bacteria Exposure in Danish Livestock Farmers. Ann. Work Expo. Health 2019, 63, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Lagier, J.-C.; Khelaifia, S.; Alou, M.T.; Ndongo, S.; Dione, N.; Hugon, P.; Caputo, A.; Cadoret, F.; Traore, S.I.; Seck, E.H.; et al. Culture of Previously Uncultured Members of the Human Gut Microbiota by Culturomics. Nat. Microbiol. 2016, 1, 16203. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the Human Intestinal Microbial Flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.L.; Wolin, M.J. Stability of Methanobrevibacter Smithii Populations in the Microbial Flora Excreted from the Human Large Bowel. Appl. Environ. Microbiol. 1983, 45, 317–318. [Google Scholar] [CrossRef] [PubMed]
- Guindo, C.O.; Davoust, B.; Drancourt, M.; Grine, G. Diversity of Methanogens in Animals’ Gut. Microorganisms 2020, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Battumur, U.; Lee, M.; Bae, G.S.; Kim, C.-H. Isolation and Characterization of a New Methanoculleus Bourgensis Strain KOR-2 from the Rumen of Holstein Steers. Asian-Australas. J. Anim. Sci. 2018, 32, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Dagar, S.S.; Puniya, A.K. Isolation and Characterization of Methanogens from Rumen of Murrah Buffalo. Ann. Microbiol. 2012, 62, 345–350. [Google Scholar] [CrossRef]
- André, A.C.; Debande, L.; Marteyn, B.S. The Selective Advantage of Facultative Anaerobes Relies on Their Unique Ability to Cope with Changing Oxygen Levels during Infection. Cell. Microbiol. 2021, 23, e13338. [Google Scholar] [CrossRef]
- Lu, Z.; Imlay, J.A. When Anaerobes Encounter Oxygen: Mechanisms of Oxygen Toxicity, Tolerance and Defence. Nat. Rev. Microbiol. 2021, 19, 774–785. [Google Scholar] [CrossRef]
- Zakem, E.J.; Mahadevan, A.; Lauderdale, J.M.; Follows, M.J. Stable Aerobic and Anaerobic Coexistence in Anoxic Marine Zones. ISME J. 2020, 14, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Rackaityte, E.; Halkias, J.; Fukui, E.M.; Mendoza, V.F.; Hayzelden, C.; Crawford, E.D.; Fujimura, K.E.; Burt, T.D.; Lynch, S.V. Viable Bacterial Colonization Is Highly Limited in the Human Intestine in Utero. Nat. Med. 2020, 26, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Al Alam, D.; Danopoulos, S.; Grubbs, B.; Ali, N.A.B.M.; MacAogain, M.; Chotirmall, S.H.; Warburton, D.; Gaggar, A.; Ambalavanan, N.; Lal, C.V. Human Fetal Lungs Harbor a Microbiome Signature. Am. J. Respir. Crit. Care Med. 2020, 201, 1002–1006. [Google Scholar] [CrossRef]
- Kimura, I.; Miyamoto, J.; Ohue-Kitano, R.; Watanabe, K.; Yamada, T.; Onuki, M.; Aoki, R.; Isobe, Y.; Kashihara, D.; Inoue, D.; et al. Maternal Gut Microbiota in Pregnancy Influences Offspring Metabolic Phenotype in Mice. Science 2020, 367, eaaw8429. [Google Scholar] [CrossRef]
- Vuong, H.E.; Pronovost, G.N.; Williams, D.W.; Coley, E.J.L.; Siegler, E.L.; Qiu, A.; Kazantsev, M.; Wilson, C.J.; Rendon, T.; Hsiao, E.Y. The Maternal Microbiome Modulates Fetal Neurodevelopment in Mice. Nature 2020, 586, 281–286. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Methanogenic archaeal species present in the rumen and feces of cattle.
| Phylum | Family | Genus | Species | Sequencing method | Host | Reference |
|---|---|---|---|---|---|---|
| Rumen | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium, smithii,gottschalkii, boviskoreani, milerae | 16S rRNA Sequencing V4 | Korean native cattle | Lee et al. 2013 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium, smithii,gottschalkii, boviskoreani, milerae | 16S rRNA Sequencing V6-V8 | Sheep | Snelling et al.2014 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium, smithii,gottschalkii, boviskoreani, milerae,thaueri | 16S rRNA Sequencing V4 | Dairy cow | Danielsson et al. 2017 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium | 16S rRNA Sequencing V4 | Beef heifers | Samat et al. 2021 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | Not reported | 16S rRNA | Ruminants | Hodet et al. 2018 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | stadtmanae, cuniculi | 16S rRNA Sequencing V4 | Breeding bulls | Koziol et al. 2022 |
| Thermoplasmatota | Methanomassiliicoccaceae | Methanomassiliicoccus | luminyensis | qPCR | Sheep, Cow | Guindo et al. 2020 |
| Euryarchaeota | Methanospirillaceae | Methanospirillum | hungatei and lacunae | 16S rRNA Sequencing V4 | Dairy cow | Tamsin Lyons et al. 2018 |
| Euryarchaeota | Methanobacteriales | Methanobacterium | mobilis,formicicum,barkaeri | 16S &18S | Grazing cattle | Jarvis et al. 2000 |
| Euryarchaeota | Methanobacteriaceae | Methanomicrobium | mobile | 16S rRNA Sequencing V1-V2, V2-V3, culturing | Sheep | Belanche et al.2019 |
| Euryarchaeota | Methanosarcinaceae | Methanosarcina | barkeri | 16S rRNA Sequencing, culturing | Dairy cow | Lambie et al. 2015 |
| Euryarchaeota | Methanocorpusculaceae | Methanocorpusculum | Not reported | 16S rRNA Sequencing V3-V4 | Dairy cow | Dingxing Jin et al. 2017 |
| Nitrososphaerota | Nitrososphaeraceae | Nitrososphaera | Not reported | 16S rRNA Sequencing V3-V4 | Dairy cow | Dingxing Jin et al. 2017 |
| Thermoproteota | Cenarchaeaceae | Cenarchaeum | Not reported | Shotgun metagenomics | Dairy cattle | Ngetich et al. 2022 |
| Nitrososphaerota | Nitrosopumilaceae | Nitrosopumilus | Not reported | Shotgun metagenomics | Dairy cattle | Ngetich et al. 2022 |
| Korarchaeota | Korarchaeales | Korarchaeota | Not reported | Shotgun metagenomics | Dairy cattle | Ngetich et al. 2022 |
| Nanoarchaeota | Nanoarchaeaceae | Nanoarchaeum | Not reported | Shotgun metagenomics | Dairy cattle | Ngetich et al. 2022 |
| Feces | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | stadtmanae, cuniculi | 16S rRNA Sequencing V4 | Breeding bulls | Koziol et al. 2022 |
| Euryarchaeota | Methanospirillaceae | Methanospirillum | hungatei and lacunae | 16S rRNA Sequencing V4 | Dairy cow | Guindo et al. 2020 |
| Euryarchaeota | Methanocorpusculaceae | Methanocorpusculum | Not reported | 16S rRNA Sequencing V3-V4 | Sika deer, Dairy cow | Tamsin Lyons et al. 2018 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | smithii, millaerae, labreanum,aggregans | PCR | Sheep | Dingxing Jin et al.2017 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | smithii, millaerae, labreanum,aggregans, thaueri | PCR | cow | Dingxing Jin et al. 2017 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | boviskoreoni, millerae, olleyae,ruminantium,wolini | 16S rRNA Sequencing V3-V4 | Beef | Dingxing Jin et al. 2017 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | Not reported | 16S rRNA Sequencing V3-V4 | Beef | Ngetich et al. 2022 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | cuniculi | 16S rRNA Sequencing V3-V4 | Beef | Ngetich et al. 2022 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | Not reported | 16S rRNA Sequencing V3-V4 | Beef | Ngetich et al. 2022 |
| Euryarchaeota | Methanobacteriaceae | Not reported | Not reported | 16S rRNA Sequencing V3-V4 | Beef | Ngetich et al. 2022 |
| Euryarchaeota | Methanosarcinaceae | Methanosarcina | mazei | 16S rRNA Sequencing V3-V4 | Beef | Guindo et al. 2020 |
| Halobacterota | Methanomicrobia | Methanococorpusculum | Not reported | 16S rRNA Sequencing V3-V4 | Beef | Guindo et al. 2020 |
| Korarchaeota | Korarchaeales | Korarchaeota | Not reported | Shotgun metagenomics | Dairy cattle | Webb et al. 2023 |
| Nanoarchaeota | Nanoarchaeaceae | Nanoarchaeum | Not reported | Shotgun metagenomics | Dairy cattle | Webb et al. 2023 |
| Nitrososphaerota | Nitrososphaeraceae | Nitrososphaera | Not reported | 16S rRNA Sequencing V4 | Dairy cow | Webb et al. 2023 |
| Nitrososphaerota | Nitrososphaeraceae | Nitrososphaera | Not reported | 16S rRNA Sequencing V3-V4 | Dairy cow | Webb et al. 2023 |
| Nitrososphaerota | Nitrosopumilaceae | Nitrosopumilus | Not reported | Shotgun metagenomics | Dairy cattle | Webb et al. 2023 |
| Thermoplasmatota | Methanomassiliicoccaceae | Methanomassiliicoccus | luminyensis | qPCR | Sheep,Cow | Guindo et al. 2020 |
| Thermoplasmatota | Methanomethylophilaceae | Methanomassiliicoccus | Not reported | 16S rRNA Sequencing V3-V4 | Beef | Webb et al. 2023 |
| Thermoproteota | Cenarchaeaceae | Cenarchaeum | Not reported | Shotgun metagenomics | Dairy cattle | Webb et al. 2023 |
Table 2.
Methanogenic archaeal species present in the reproductive and respiratory tracts of cattle.
Table 2.
Methanogenic archaeal species present in the reproductive and respiratory tracts of cattle.
| Phylum | Family | Genus | Species | Sequencing platform | Host | Reference |
|---|---|---|---|---|---|---|
| Vagina | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium | 16S rRNA Sequencing V3-V4 | Beef heifers | Amat et al. 2021 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium | 16S rRNA Sequencing V3-V4 | Beef heifers | Winders et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | wolini | 16S rRNA Sequencing V3-V4 | Beef heifers | Winders et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | not reported | 16S rRNA Sequencing V3 | Beef cattle | Laguardia-Nascimento et al., 2015 |
| Uterus | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium | 16S rRNA Sequencing V3-V4 | Beef heifers | Winders et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | wolini | 16S rRNA Sequencing V3-V4 | Beef heifers | Winders et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | not reported | 16S rRNA Sequencing V3-V4 | Beef heifers | Winders et al. 2023 |
| Semen | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | stadtmanae, cuniculi | 16S rRNA Sequencing V4 | Beef bulls | Koziol et al. 2022 |
| Thermoplasmatota | Methanomassiliicoccaceae | Methanomassiliicoccus | luminyensis | 16S rRNA Sequencing V4 | Beef bulls | Koziol et al. 2022 |
| Crenarchaeota | Nitrososphaeraceae | Not reported | Not reported | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanobacterium | mobilis,formicicum,bryantii, | 16S rRNA Sequencing V4 | Beef bulls | Koziol et al. 2022 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | boviskoreoni, millerae,olleyae, ruminatium, wolini | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | Not reportd | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | cuniculi | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | Not reported | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Not reported | Not reported | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Euryarchaeota | Methanosarcinaceae | Methanosarcina | mazei | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Thermoplasmatota | Methanomethylophilaceae | Methanomassiliicoccus | Not reported | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Halobacterota | Methanomicrobia | Methanococorpusculum | Not reported | 16S rRNA Sequencing V3-V4 | Beef bulls | Webb et al. 2023 |
| Halobacterota | Methanomicrobia | Methanococorpusculum | Not reported | 16S rRNA Sequencing V3-V4 | Beef bulls | Cojkic et al.2021 |
| Milk | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium, smithii, milerae | Shotgun metagenomics | Dairy cow | Ekman et al. 2020; Guindo et al. 2020 |
| Euryarchaeota | Methanocorpusculaceae | Methanocorpusculum | Not reported | Shotgun metagenomics | Dairy cow | Ekma et al.2020 |
| Nasopharynx | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium | 16S rRNA Sequencing V3-V4 | Beef heifers | Winders et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | wolinii | 16S rRNA Sequencing V3-V5 | Beef heifers | Winders et al. 2023 |
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | Not reported | 16S rRNA Sequencing V4 | Beef cattle | Amat et al. 2019 |
| Udder teat | ||||||
| Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | ruminantium, smithii, milerae | Shotgun metagenomics | Dairy cow | Ekman et al. 2020; Guindo et al. 2020 |
| Euryarchaeota | Methanobacteriaceae | Methanosphaera | stadtmanae, cuniculi | Shotgun metagenomics | Dairy cow | Ekma et al.2020 |
| Nitrososphaerota | Nitrosopumilaceae | Nitrosopumilus | Not reported | Shotgun metagenomics | Yak calves | Guo et al. 2023 |
| Nanoarchaeota | Nanoarchaeaceae | Nanoarchaeum | Not reported | Shotgun metagenomics | Yak calves | Guo et al. 2023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



