
Submitted:
23 October 2023
Posted:
25 October 2023
You are already at the latest version
Abstract
Recently, nanobodies have taken center stage in research on drug discovery and development. Several therapeutic possibilities using nanobodies are presently undergoing clinical trials and waiting for FDA approval. The goal of this study was to emphasize the potential of nanobodies as therapeutic agents by concentrating on the most recent published studies that examined their properties, manufacturing, and possible applications. This article demonstrates that the unique properties of nanobodies in comparison to conventional antibodies, which are based on their small size and offer a number of benefits, make them seem to have a promising future. These advantages include the ability to access complex or hidden target sites that may be elusive to their larger antibody counterparts, and increased resilience against extreme conditions like tempera-ture changes and pH variations. As a result, nanobodies seem to have a bright future as adaptable tools for imaging in cancer and non-malignant diseases, as well as for in vitro and in vivo diagnostic and therapeutic potential for a variety of conditions, including oncology, infectious, metabolic, neurological, and other conditions like ophthalmologic, immune-mediated, and genetic disorders. More research is required to determine their effectiveness and safety in clinical applications. The current analysis offers a thorough overview of the therapeutic uses for nanobody products that are either on the market or undergoing clinical trials.
Keywords:
nanobody
; therapeutic nanobody
; production
; imaging
; diagnosis
; single-domain antibodies
; VHHs
1. Introduction
Glycoproteins termed antibodies, commonly known as immunoglobulins (Ig), are created by B lymphocytes [1]. They are vital components of the immune system that play a crucial role in the body's defense against disease by identifying and inhibiting invading antigens [2]. Conventional antibodies are "Y"-shaped molecules made up of two heavy chains and two light chains (Igκ or Igλ), which are joined together by covalent disulfide bonds to form a tetrameric structure (Figure 1 A) [3]. As seen in Figure 1 A, both light chain classes have two domains: a constant domain and a variable domain [4]. Antibody light chains in humans come in two different varieties: kappa and lambda. They have a similar form and function while having different protein sequences [5]. In contrast, human antibody heavy chains can be one of five isotypes: IgA, IgD, IgE, IgG, or IgM, each with a distinct function within the adaptive immune system [4,6]. IgAs, IgDs, and IgGs contain three constant domains and one variable domain. IgEs and IgMs are composed of one variable domain and four constant domains. IgA and IgM can form dimers and pentamers, respectively, due to the presence of an additional joining chain [4,6]. These moieties are symmetrical with a variable region called the Fab (fragment antigen binding) and a constant region called the Fc (fragment crystallizable) (Figure 1 A) [3,4]. The antigen recognition and the binding specificity of the entire Ig molecule are dependent on the Fab region, especially on the two variable domains on the top, whereas the Fc domain initiates biological processes upon antigen binding [7]. IgA, IgD, IgE, IgG, and IgM are the five main classes of antibodies. They are categorized into alpha, delta, epsilon, gamma, or mu depending on the heavy chain they contain. These differ in terms of hinge structure, the valency of the antibody, and the number and sequence of constant domains as aforementioned [5]. The most abundant immunoglobulin isotype in human serum is IgG [8].
Several stages can be identified in the evolution of antibodies. It is reported that Edward Jenner is responsible for developing the smallpox vaccination [9]. Louis Pasteur then unintentionally discovered the vaccine, opening up new avenues for antibody research [10]. Early research mostly focused on observations of protective effects following direct injection of modest doses of virulent disease vectors; however, tests with crude vaccines intended to treat or prevent the disease were also carried out [11]. However, it took until the 1880s for the germ hypothesis to take the place of the spontaneous generation and miasma theories before inoculation could successfully treat diseases other than smallpox [11]. The concept of the "antibody" and the use of the antiserum to detect pathogens were both inspired by research into natural immunity in the 1890s [11]. Paul Ehrlich's contributions to the field of immunology, which included the development of the "side-chain theory" to explain the specificity of immune reactions are largely responsible for the discovery of antibodies, which further clarified the specific interaction between antibodies and antigens in the blood [12,13]. Emil Fischer added to Ehrlich's initial theory of immunological recognition by providing the framework for mechanisms akin to a lock-and-key system, which allowed for a better understanding of the specific manner in which antibodies bind to antigens [13]. The development of monoclonal antibodies (mAbs) technology by Köhler and Milstein in 1975, which made it possible to produce highly specific antibodies for a variety of uses, gave this idea even more support [14]. Therefore, it is considered that Paul Ehrlich suggested that antibodies might have been employed for therapeutic purposes more than a century ago [15]. In 1948, Astrid Fagraeus reported that plasma B cells have a specialized role in the production of antibodies, and in 1957, Frank Burnet and David Talmage established the clonal selection theory [14]. The molecular structure of immunoglobulin was separately described by Gerald Edelman and Rodney Porter in 1959, for which they eventually shared the Nobel Prize in 1972 [14]. The first high-resolution structure of an antibody fragment was initially reported in 1973 [14,16]. The discovery of mAbs by Georges Köhler and César Milstein in 1975 marked the beginning of the contemporary age of antibody research and discovery [14]. The approval of the first monoclonal antibody (mAb) by the Food and Drug Administration (FDA) in 1986 marked the undeniable evolution of antibody engineering [3]. In recent times, the therapeutic antibodies drug market has experienced an exceptional surge in growth, primarily attributed to the approval of novel drugs for treating a broad range of diseases, including cancer, neurological, infectious, autoimmune, metabolic, genetic, and others [17]. For instance, more than 100 mAbs have received official authorization for commercial purposes [18,19]. In 2022, about 24% of novel medications authorized by the FDA were mAbs [19]. The therapeutic potential of mAbs is because they are developed via a single clone of B cells, a property that makes them monospecific and homogeneous compared with polyclonal antibodies that are produced in vivo [18]. The four types of therapeutic antibodies, depending on the source from which they are produced, are murine, chimeric, humanized, and fully human mAbs [20]. Human anti-mouse antibodies have been created due to the frequent usage of murine antibodies. In order to lessen the immunogenicity of mouse antibodies, chimeric, humanized, and totally human antibodies have been developed [20]. Depending on how they are utilized, there are a variety of therapeutic mAbs types such as unconjugated (naked), conjugated, and bi- and tri-specific [21]. These have been developed as a result of the effective use of IgG mAbs [22]. Unconjugated mAbs are the most common agents in use [21]. Conjugated mAbs are linked with chemotherapy or a radioactive compound. Bispecific mAbs pair two different mAbs and bind to two diverse antigens, simultaneously. While tri-specific mAb can interact with three different antigens [21]. Although conventional antibodies are useful for treating a variety of illnesses[23], there are still a number of issues that need to be resolved. Particularly, drug resistance and poor stability because mAbs are glycoproteins remain to be the main drawbacks [24].
To enhance conventional antibody features, several structural alterations to their structure have been made using recombinant DNA technology and protein engineering [7]. As a result, smaller antibody fragments have been designed, including Fab, single-chain variable fragments (scFvs), mini-, dia-, and tria-bodies, and nanobodies (Nbs) which are also recognized as VHHs (variable domain of heavy-chain-only antibodies (HCAb)) or single-domain antibodies (Figure 1 B, C and D) [7]. Among all of them, scFvs and nanobodies are the most widespread ones (Figure 1 B and D) [7]. The scFvs, which are made up of the Ig heavy and light chain variable regions that bind by a peptide linker, were earlier thought to be the smallest antibody fragment with a molecular weight of around 25–35 kDa and the same antigen-binding selectivity as the entire Ig molecule (Figure 1 B) [7,25]. However, it was shown that a single variable-like domain can maintain the affinity of an entire antibody molecule via the discovery of the camelid VHH [26] and shark variable new antigen receptor [27]. Although scFvs continue to rule the clinic and ten of them have received FDA approval [7], the superior features of Nbs have made them dominant in the field of recombinant antibody engineering in less than three decades since the discovery of HCAb [7].
Nbs were discovered in 1993 when Hamers et al. revealed for the first time that camelids such as llamas, alpacas, and camels naturally have HCAbs that lack light chains (Figure 1 C and D) [26,28]. Although single-domain antibodie were also detected in cartilaginous fish, camelids were used in most single-domain antibodies biotechnological applications studies since they are easier to handle and immunize [29]. In 2018, Caplacizumab received approval from the European Medicines Agency for the management of patients with thrombotic thrombocytopenic purpura and therefore became the first nanobody drug to be marketed [30]. Another nanobody called Ozoralizumab was approved in Japan in September 2022 for the treatment of rheumatoid arthritis that is not adequately controlled by currently existing therapies [31]. Currently, several Nb-based therapeutic candidates are undergoing clinical studies and waiting for FDA approval, such as Sonelokimab and Gontivimab [7].
Since research on therapeutic Nbs has dominated drug discovery and development lately [3], this study conducted a thorough review with a focus on the latest studies that discussed the characteristics, production, and potential indications of Nbs to highlight the future directions of these novel agents.
2. Methods
Literature electronic search was conducted using the most popular search engines: Google Scholar; PubMed; Scopus; and Science Direct, and the following keywords have been used: Nb; imaging; diagnosis; single-domain antibodies; VHHs; and indications of Nbs. The most recent research that met the following selection criteria was included in this review: (1) written in English; and (2) discussed the features, production, or therapeutic potential of Nbs. Studies that have not explored the Nbs were excluded.
3. Results and Discussion
3.1. Nbs Characteristics and Production
The primary characteristic of Nbs lies in their simplified structure, consisting of a sole variable domain encompassing the antigen-binding region. This key attribute results in their notably smaller size with a molecular weight of only 12–15 kDa compared to conventional antibodies that have a molecular weight of around 150 kDa, granting Nbs the unique capacity to access intricate or concealed target sites that may remain elusive to larger antibody counterparts (Figure 1 A and D) [32,33,34]. In addition, their compact size offers several benefits, including heightened resilience against extreme conditions like temperature fluctuations and pH variations [32,33]. Significantly, Nbs exhibit remarkable binding affinity and specificity towards their target antigens, often surpassing the performance of traditional antibody molecules. Their propensity for solubility reduces the risk of aggregation and facilitates simplified manufacturing processes. Moreover, Nbs' exceptional ability to penetrate tissues underscores their potential for diagnostic and therapeutic applications, and their amenability to genetic manipulation allows for tailored enhancements of their properties., Nbs also demonstrate reduced immunogenicity, minimizing the likelihood of undesirable immune responses [28,35]. Despite the aforementioned Nbs advantages, they have some drawbacks including short serum half-life since they are rapidly cleared by the kidneys due to their small size [36]; however, multimeric Nbs, pegylated Nbs, or Nbs that bind to serum albumin can be manufactured to enhance their half-life in blood [29,36]. In addition, it was believed that unmodified Nbs are not able to permeate via the cell membrane [37]. Nevertheless, Singh et al recently have described an unmodified cell penetrating Nb, SBT-100, that enters the cell membrane and gives a therapeutic effect against cancers. Further investigations are being conducted [38].
Nbs are typically acquired from immunological libraries created by animal immunization and involve the administration of the appropriate antigen through animal injection (Figure 2) [39]. Naïve libraries, derived from the blood of non-immunized camelids, are also used in Nbs production. However, in recent years, synthetic libraries of various forms have gained traction as trustworthy Nb sources, delivering significant cost and speed advantages [40]. A synthetic Nb library is defined by two main features: framework selection and the construction of complementarity-determining regions [40]. For an immune library, a healthy young adult camelid is inoculated with 50 to 200 μg of the selected antigen per injection, depending on antigen features. For two months, the animal must be injected with four to eight shots in total [39]. After immunization, B-lymphocytes are isolated from the peripheral blood or lymph nodes of the immunized animal, and they are then used to extract mRNA [39]. After using the mRNA for reverse transcription to create complementary DNA and polymerase chain reaction (PCR) with VHH-specific primers to amplify the VHH gene area, the nanobody sequences are chosen by phage display or any display method such as ribosome, yeast, and bacterial displays, or NestLink-based selection [39]. The oldest and still the most durable of these selection technologies is phage display [39]. The chosen nanobodies are recombinantly expressed at the industrial level in either eukaryotic or prokaryotic systems, including mammalian cells, bacteria such as Escherichia coli, or yeasts such as Pichia pastoris [41,42]. Finally, it is extracted and purified (Figure 2) [41].
3.2. Therapeutic Indications of Nbs
Nbs emerge as a promising and versatile molecular tool applicable across diverse domains such as imaging, diagnostics, therapeutics, and research, with their unique attributes distinguishing them from conventional antibody formats and other antibody fragments [28,35].
3.2.1. Nbs for Imaging
Nbs have lately been considered incredible tools for confocal and high-resolution imaging due to their small size which reduces the distance between the probe and the antigen [43]. Nbs enable acquiring early high-quality images that allow personalized precision therapy by providing a full estimation of diseases such as cancer and many other diseases [44]. A molecular tracer compositing of targeting integrity and a detection label are required to achieve the direction and specific accumulation of the probe and allow the visualization of the target tissue, respectively [45]. Therefore, Nbs must be supplied with appropriate radioisotope or fluorescent dye [45]. Numerous labeling method of Nbs has been applied to enable in vivo imaging including radio-labeling for single photon emission computed tomography (SPECT) and PET imaging [43]. For nuclear imaging uses, positron-emitting isotopes are applied for PET, whereas a gamma-emitting isotope is employed for SPECT [45]. Several approaches have been used for radiolabeling of Nbs including direct and indirect radiohalogenation, and radiometals using either synthetic chelators or proteinogenic chelators [45]. In addition to radiolabeling of Nbs, fluorescent dye labeling of the Nbs has also been used for near-infrared or infrared region imaging [43].
3.2.1.1. Nbs for Imaging in Cancer
Nbs have been used for imaging cancer cell markers [46]. For instance, a modern PET probe, anti-HER2 Nb labeled with 18F for imaging HER2 overexpression in cancer cells, has been shown great potential for clinical translation [46]. The bi-paratopic Nb construct (MSB0010853) is another example of using Nbs for imaging cancer cell markers, but this complex was formulated to target HER3 [47]. It consists of three domains, two of them are directed against the HER3 receptor, and the third is developed to bind to albumin, resulting in extending its serum half-life [47]. This Nb was radiolabeled with 89Zr [47]. It has shown potential to be employed for both therapeutic and imaging applications in cancer [47]. NJB2 is another Nb that was designated specifically to target the Fibronectin-EIIIB splice variant expressed in the extracellular matrix of cancer and fibrosis [48]. They proposed that NJB2 can be used as a powerful tool for the non-invasive detection and targeting of tumors and other metastatic lesions [48].
Besides using Nbs for imaging cancer cell markers, they have also been used for imaging immune checkpoint markers. For example, an Nb-based probe is a potential agent for PET imaging for future clinical measurement of expression of human programmed death-ligand 1, an immune checkpoint, by PET imaging [49]. Envafolimab is a novel anti-programmed death ligand 1 Nb [50,51,52]. It has lately been used for immuno-PET imaging with 89Zr-Df-KN035 and is considered a powerful tool for non-invasive in vivo evaluation of programmed death-ligand 1 expression in non-small-cell lung cancer and other tumors [50,51]. A cytotoxic T-lymphocyte antigen-4-specific Nb-fluorescent carbon quantum dots complex (QDs-Nb36) has currently been designed to detect the cytotoxic T-lymphocyte antigen-4-positives T-cell, cytotoxic T-lymphocyte antigen is a checkpoint inhibitory molecule, via flow cytometry or immunofluorescent staining [53]. Lymphocyte-activation gene 3 is another frequent target in immune checkpoint inhibition [54]. Most recently, Nbs have been developed and radiolabeled to non-invasively measure lymphocyte-activation gene 3 expression by SPECT/CT imaging in tumor-bearing mice [54]. They have demonstrated that 99mTc-labeled Nb is an excellent SPECT probe to assess lymphocyte-activation gene 3 on immune cells and tumor cells [54]. They later assessed the ability of Nb to image lymphocyte-activation gene 3 on tumor-infiltrating lymphocytes by SPECT/CT in various tumor-bearing mice [55]. Their findings suggest that Nbs could be used to non-invasively detect the lymphocyte-activation gene 3 upregulation of tumor-infiltrating lymphocytes [55]. A follow-up trial in tumor-carrying mice treated with anti-PD-1 antibodies concluded that Nb-lymphocyte-activation gene 3 imaging can have prognostic value and could be used to predict therapy outcomes both before and during therapy [55].
Nbs have also been used to target immune markers to image immune responses by imaging particular subgroups of immune cells, such as T cells and macrophages [52]. This can help in predicting the response to treatment. In addition, tracking the infiltration of these immune cells would be beneficial to detect tumors [52]. An anti-CD8 Nb was designed and radiolabeled with 89Zr to image the animals non-invasively, with PET showing clearly the lymphoid organs [56]. A pegylated moiety was attached to this radiolabeled Nb, which enhanced signal-to-noise and reduced kidney absorption [56]. The above-mentioned 89Zr labeled Nb has been later used to investigate the dynamics of the CD8+ T cells and CD11b+ myeloid cells in response to anti-PD-1 therapy in the MC38 colorectal mouse adenocarcinoma model [57]. They demonstrated that non-invasive immuno-PET imaging can track intratumoral CD8+ T cells even when their specificity is unknown and track the development of the anticancer response to anti-PD-1 medication [57]. These findings may help specialists to make better choices regarding the patients’ treatment options [57]. Newly, high radiochemical purity and affinity Nb has been designed to target human CD8 antigen [58]. This novel Nb (68Ga-NOTA-SNA006a) has shown a long retention period, low background, and impressive tumor-to-background ratios compared with other Nbs, allowing precisely gauging the human CD8 positive T cells in mice models, demonstrating excellent ability for immunotherapy observersion and effectiveness measurement [58]. The macrophage mannose receptor (gene name: CD206) is associated with disease development, ranging from oncology to atherosclerosis and rheumatoid arthritis [59]. A [68Ga]Ga-NOTA-anti-macrophage mannose receptor-Nb, a PET tracer targeting CD206, has undergone a phase I clinical study and shown high safety and tolerability and allowed high contrast to noise imaging at 1:30 hours after injection [59]. This Nbs will be moved to a phase II clinical study [59].
3.2.1.2. Nbs for Imaging of Non-Malignant Diseases
Nbs have also been designed to image the biomarkers that are overexpressed in other diseases, such as atherosclerosis, fibrosis, arthritis, and some neurodegenerative disorders [52]. For example, the expression level of vascular cell adhesion molecule-1 has been detected both in mouse models of atherosclerosis and in ex vivo human endarterectomy specimens via anti-vascular cell adhesion molecule-1 Nbs that is linked to microbubbles [60]. This complex is translatable into the clinic and could enhance risk stratification for atherosclerosis [60]. The aggregation of α-synuclein protein underlies the development of several neurodegenerative disorders [61]. Therefore, two anti- α-synuclein Nbs were successfully designed to link to the C-terminal domain of α-synuclein [61,62]. These anti-α-synuclein Nbs also hold promise in the diagnosis and management plans of neurodegenerative diseases [61]. Another study has demonstrated that 64Cu-anti-EIIIB Nb has the promise to image and detect pulmonary fibrosis in a clear, non-invasive, and specific way [48]. Moreover, a 99mTc-labeled anti-DPP6 Nb successfully detects high amounts of EndoC-βH1 cells or human islets grafted in severe combined immunodeficient mice. Moving this Nb to the clinic may enable non-invasive, in vivo monitoring of diabetic management progress [63].
Overall, Nbs have demonstrated great success as imaging tools in cancers and non-malignant diseases.
3.2.2. Nbs for Diagnosis
The adaptability of Nbs in diagnostics has drawn interest [42]. They are promising for a variety of diagnostic systems in both in vivo and in vitro experiments due to their compact structure, stability, and strong binding to target antigens [42,64]. Additionally, Nbs are made specifically for screening chemicals and pesticides in food, to ensure safety [64].
3.2.2.1. Nbs for In Vitro Diagnosis
Nbs have gained prominence in in vitro diagnostic tests, particularly in lateral flow immunoassays (LFIA) and enzyme-linked immunosorbent assays (ELISA) [42]. LFIA employs Nbs to swiftly detect specific antigens, forming a visual "sandwich" at the test line [65]. Nbs' stability, broad binding capabilities, absence of a cross-reactive Fc region, multivalency, and affinity for gold nanoparticles enhance LFIA's sensitivity [66,67,68,69,70]. The Cellular and Molecular Immunology research unit has made significant advancements in LFIA development. Notably, they have successfully detected Trypanosoma congolense pyruvate kinase, a glycolytic enzyme, with the potential for diagnosing active trypanosomosis infections [68]. This involves generating Nbs against Trypanosoma congolense secretome, creating a sandwich ELISA, and adapting it to LFIA, demonstrating high sensitivity (80%) and specificity (92%) [68]. Similar strategies extend to Nbs targeting glycolytic enzymes enolase and aldolase, enhancing Trypanosoma evansi and Trypanosoma congolense diagnosis [71,72,73]. Nb-based sandwich ELISAs also exhibit promise across various antigens, such as cancer biomarkers, toxins, and bacteria, expanding diagnostic possibilities [42], along with a reported competitive Nbs-based ELISA for diagnosing foot and mouth disease antibodies in cattle [74].
Nbs perform well as bioreceptors in biosensors, anchoring onto platforms like nanoparticles or semiconductors. This biosensing approach involves Nbs binding to target molecules, triggering electric potential changes converted into measurable signals [75]. This rapid response enables real-time detection, surpassing traditional methods like ELISAs and LFIAs [42]. For instance, Nbs were applied in a fibrinogen-detecting biosensor, a cardinal cardiovascular disease biomarker [76]. Nbs also show promise as biosensors for identifying spike proteins of SARS-CoV-2 and Middle East respiratory syndrome coronavirus [77].
3.2.2.2. Nbs for In Vivo Diagnostic Imaging
In vitro diagnostics are crucial for disease detection, yet their utility is limited, especially for localized abnormalities in cases like cancer within bodily fluids [42]. In contrast, fluorescent or radioactively labeled antibodies are used in in vivo diagnostics, such as optical and nuclear imaging, to detect abnormalities like tumors as aforementioned. Due to their size, ease of modification, and quick clearing, Nbs are an ideal option for making target-specific tracers [42]. Although optical imaging may not be as accurate as nuclear imaging, it is safer and provides data immediately [78]. A fluorescent Nb with the name of 7D12 demonstrated a quicker and deeper response than the commercial mAb Cetuximab, indicating potential for optical imaging [79]. Nbs against HER2 and carbonic anhydrase IX revealed increased target-to-background ratios for improved tumor diagnosis in a dual tracer method [80]. Nbs are beneficial for cardiovascular molecular imaging due to their lack of an Fc region and quick clearance, as demonstrated by a 99mTc-labeled Nb against vascular cell adhesion molecule-1, a biomarker linked to susceptible atherosclerotic plaques [81,82]. In the field of nuclear imaging (PET/SPECT), Nbs as radioactive tracers against HER2 exhibit high potential [78]. For instance, 68Ga conjugate Nbs against HER2 showed high-contrast PET imaging results [83]. Thus, this promising Nb is undergoing phase 2 trials, which are expected to be completed by 2023 [84]. Moreover, radioactively labeled anti-HER2 Nbs like 18F and 177Lu show promise in the detection of biomarkers not only in cancer but also in other conditions [42,46,85]. Nbs' size, clearance, and ease of functionalization position them as excellent candidates for both in vitro and in vivo imaging, advancing disease diagnosis [42].
3.2.3. Nbs for Oncology
Nbs’ outstanding properties such as their small size and ability to penetrate tumors in vivo enhanced their tumor-targeting ability [86]. Compared with traditional antibodies, Nbs possess the same antigen binding features but with a single Ig variable domain for antigen recognition [87]. Therefore, they can reach epitopes that conventional antibodies cannot access such as cleft on a protein’s surface [87]. Anticancer Nbs can be divided into 3 types: naked monomeric or multimeric Nbs, Nbs as targeting moieties to effector domains, and Nbs binding the surface of liposomes, or other nanoparticles that encapsulate compounds [86].
Regarding oncology therapy, Nbs have been used to target tumor antigens and they have first been examined as antagonists. For instance, Nbs against epidermal growth factor receptor (EGFR) and c-mesenchymal-epithelial transition factor have been developed [88,89]. They have shown in vitro antagonistic effects for either EGF or hepatocyte growth factor [89]. Tumor growth in vivo was suppressed with trivalent biparatopic anti-EGFR Nb 7D12-9G8-Alb [88,89]. In addition to the antagonistic effect of Nbs, they can be produced as allosteric inhibitors to modify the enzymatic action of their interest protein [89]. On the other hand, when only one ligand is responsible for the stimulation of the target receptor, Nbs can be used to bind to the ligands directly [89]. Nbs have also been demonstrated potential effects in enhancing immune checkpoint inhibitors [90,91]. Several trials have developed Nb immune checkpoint inhibitors for programmed death-ligand 1 that can exhibit significant antitumor effects [90,91]. It has also shown promising activity in inhibiting angiogenesis such as recombinant anti-vascular endothelial growth factor bivalent Nb was developed with extended plasma half-life properties that hold promise to inhibit angiogenesis within solid cancers [91,92]. As demonstrated by these findings, Nbs-based treatment in oncology holds tremendous potential.
3.2.4. Nbs for Infectious Diseases
Nbs have become a promising therapeutic approach with the potential to revolutionize the treatment of infectious diseases against viruses, bacteria, and parasites by focusing on different agents [93].
Nbs showed considerable potential in treating viral infections by neutralizing a variety of viruses, such as influenza, respiratory syncytial virus, HIV, and SARS-CoV-2. They disrupted the infection cycle by successfully preventing viral entrance, fusion, and replication [93,94]. With their improved affinity and virus-blocking properties, polyvalent Nbs have the potential to treat a variety of infectious diseases and improve global health outcomes [93,94]. Nbs that target the virus's receptor-binding domain are quite successful in the context of SARS-CoV-2. The viral entry is facilitated by the interaction of SARS-CoV-2 with the human angiotensin-converting enzyme 2 receptor via the receptor binding domain. Multivalent Nbs targeting the SARS-CoV-2 receptor binding domain bind multiple epitopes simultaneously, countering viral mutation evasion [95]. These Nbs can block the interaction, preventing viral invasion and positioning them as prime candidates for immunotherapy [94]. Notable examples include Nb variants like "VHH-72 Fc fusion" and "VHH-114," which demonstrate strong neutralization potency against multiple SARS-CoV-2 variants [96,97]. Beyond SARS-CoV-2, multivalent Nbs show efficacy against infections like HIV and respiratory syncytial virus. Notably, Nb ALX-0171, under clinical trials, targets respiratory syncytial virus and significantly reduces viral load [98]. Nbs also hold potential for influenza, Shigella spp., and other pathogens. Multivalent Nbs offer the ability to simultaneously target pathogens and receptors in vivo. For example, Nbs developed against specific proteins on the H5N1 influenza virus have shown remarkable potential in mice [99]. Moreover, Nbs' potential extends to hepatitis C therapy customization and fighting Rift Valley Fever virus and Schmallenberg virus infections [100,101,102,103]. Overall, these de novo studies highlighted the significant potential of Nbs in combating viral infectious diseases. Their precise targeting, specificity, and potency are leading the way for novel therapeutic and diagnostic strategies, potentially revolutionizing a new approach to viral infections.
In bacterial infections, Nbs have displayed effectiveness by targeting virulence factors essential for the invasion of pathogens like Clostridium difficile, Staphylococcus aureus, and Escherichia coli into host cells. Through their binding, Nbs prevent bacterial attachment, toxin activity, and invasion [104]. An important example is Nbs that target Shigella spp.'s Type III secretion system (T3SS), decreasing their hemolytic activity and possibly enabling novel therapeutic approaches against bacillary dysentery [105]. Furthermore, an Nb has been created to target a bacterial surface antigen as a substitute for antibiotics in the context of treating Streptococcus mutants, the cause of tooth decay [104]. However, the combined Nb and glucose oxidase hybrid (S36-VHH-GOx) exhibited a reduction in bacterial levels in infected rats [104]. Similarly, efforts to create Nb-based treatments for Neisseria and Escherichia coli-related infections aimed to provide alternatives for antibiotic-resistant individuals [104]. The Neisseria-targeting VHH-5G Nb effectively blocked lipopolysaccharide receptor interaction, whereas the treatment targeting Escherichia coli F4 fimbriae did not show success in preventing disease or improving mortality rates in a mouse model [104]. On the other hand, Nbs created specifically for Clostridium difficile infections have shown promise in focusing on bacterial toxins and inhibiting receptor connections [104]. The investigation of Nbs as a potential therapeutic tool is opening up new possibilities for treating bacterial infections in immunocompromised and antibiotic-resistant individuals.
Nbs have been used to treat parasitic infections, including Trypanosoma brucei and Plasmodium species. Researchers designed Rift Valley fever virus and Schmallenberg virus-specific Nbs, which effectively neutralized the respective viruses, reducing morbidity and mortality in mice [100]. In particular, Nbs targeting the Duffy antigen receptor for chemokines inhibited the binding of Plasmodium vivax and Plasmodium knowlesi to human erythrocytes [106]. For Trypanosomes causing sleeping sickness and Chagas disease, Nbs targeting variable surface glycoproteins have been developed, including NbAn33 for parasite detection and cross-reactive Nb392 for diagnosing trypanosomiasis by targeting the paraflagellar rod protein of Trypanosoma brucei [107,108].
In conclusion, multivalent Nbs offer a powerful tool for combating infectious diseases, with diverse strategies to enhance their affinity, neutralization, and therapeutic potential. While challenges remain in designing optimal Nb combinations and forms, ongoing research holds promise for expanding their applications and impact.
3.2.5. Nbs for Metabolic Disorders
Nbs are increasingly offering novel therapeutic and diagnostic possibilities for complex disorders in the field of cardiovascular research such as atherosclerosis, coronary artery disease, and thrombosis [109]. As part of the treatment of heart conditions, Nbs are being explored for targeting amyloidosis-related cardiotoxicity, inhibiting ryanodine receptor 2 phosphorylation to mitigate cardiac dysfunction, and inhibiting G-protein coupled receptors to regulate hypertension. Furthermore, plasminogen activator inhibitor-1 (PAI-1), controls blood clot formation through its crucial role in plasminogen activation [110,111,112,113,114]. A high plasminogen activator inhibitor-1 level increases the risk of developing thrombosis and cardiovascular diseases such as heart attacks and strokes. Plasminogen activator inhibitor-1 activity is inhibited by Nb called VHH-2w-93. Ultimately, this will reduce the excessive clotting and the associated cardiac complications [110,111,112,113,114]. In addition, Nbs may enhance heart function by optimizing calcium ion balance in heart muscle cells, by influencing sarco-endoplasmic reticulum Ca2+-ATPase and phospholamban levels [115]. Anti-von Willebrand factor bivalent Nbs such as Caplacizumab, show potential for disrupting von Willebrand factor activity and platelet aggregation in thrombotic thrombocytopenic purpura [116]. It has received approval from the European Medicines Agency and the US FDA for the management of patients with thrombotic thrombocytopenic purpura [36]. Furthermore, Nbs targeting matrix metalloproteinase-2 and glycoprotein VI, exhibit potential in inhibiting platelet aggregation and thrombus formation, indicating their potential in anti-thrombotic therapy [117,118]. In general, Nbs hold immense promise for revolutionizing diagnostic approaches and unveiling new therapeutic avenues for addressing a range of cardiac challenges [109].
3.2.6. Nbs for Neurological Disorders
Nbs hold promise as therapeutic agents for neurological disorders due to their ability to cross the blood-brain barrier (BBB) and target central nervous system (CNS) antigens [36]. Nbs' small size and specificity address the challenge of BBB delivery, offering the potential for conditions like Alzheimer's and Parkinson's diseases [115]. Methods like attaching BBB-crossing peptides and receptor-mediated transcytosis show potential for targeted drug delivery [115]. Additionally, the "Trojan horse" strategy, utilizing Nbs to target BBB receptors, extends their potential [115].
3.2.6.1. Nbs for Alzheimer’s Disease
Alzheimer’s disease is characterized by amyloid plaques and neurofibrillary tangles as a result of amyloid-β peptide’s accumulation in the brain [119]. Nbs hold significant promise for diagnosing and treating Alzheimer's disease. They hold potential for detecting Alzheimer’s disease biomarkers in body fluids such as cerebrospinal fluid, through simple ELISA tests [120], and identifying early Alzheimer’s disease changes in the retina [121]. Nbs also present a targeted strategy for addressing Alzheimer’s disease pathology, inhibiting amyloid-β and tau aggregation, neutralizing toxic amyloid-β oligomers, and even directly degrading amyloid-β. These Nbs can also be fused with therapeutic molecules for precise brain delivery [122,123,124]. Additionally, Nbs hold therapeutic potential for tau pathology, with Nbs like VHH Z70 effectively inhibiting tau aggregation. Nbs' delivery into the brain via viral vectors like adenoviruses offers a gene therapy potential for Alzheimer’s disease [122,123,124]. Furthermore, a new investigation introduces a novel therapeutic strategy in the form of a multivalent Nb conjugate equipped with a robust scaffold capable of scavenging reactive oxygen species [125]. By integrating an amyloid-β segment into an Nb, it recognizes and inhibits amyloid-β aggregation. Further enhancements involve modifying the Nb with a human interleukin-1β fragment to aid amyloid-β clearance [125]. The resulting conjugate shows significantly improved binding to amyloid-β aggregates and long-term capabilities in reducing oxidative stress and inflammation. Notably, this conjugate mitigates symptoms in Alzheimer’s mouse models, validating its potential as a multi-target therapeutic approach [125]. Although some Nbs showed promise in preclinical studies, further research is crucial to establish their effectiveness and safety in clinical applications. Nonetheless, Nbs' distinct characteristics make them promising for innovative Alzheimer's diagnosis and treatment methods.
3.2.6.2. Nbs for Parkinson's Disease
Nbs have demonstrated significant potential for both diagnosing and treating Parkinson's disease [126]. This progressive neurodegenerative disorder is mainly due to the depleted dopamine in the basal ganglia [127]. Nbs have the potential to target Parkinson's disease-related proteins such as Nbsyn2, Nbsyn87, and Nbalpha-syn01, which have shown promise in inhibiting α-synuclein aggregation and toxicity [128,129,130,131,132]. Additionally, Nbs targeting other Parkinson's disease-related proteins like Leucine-rich repeat kinase 2 have been developed as a potential therapeutic option [133]. In summary, Nbs hold great potential for both diagnostic and therapeutic applications in Parkinson's disease, particularly in targeting α-synuclein and other related proteins. Further research and clinical studies are needed to fully realize their potential in improving Parkinson's disease diagnosis and treatment [134].
3.2.6.3. Nbs for Multiple Sclerosis
Multiple sclerosis is a demyelinating disease that involves inflammatory responses, potentially linked to genetic, environmental, and infectious factors [134]. Nbs have been recently evaluated as a potential therapeutic option for multiple sclerosis. Studies reported that Nbs such as anti-tumor necrosis factor receptor 1 Nb and 3Nb12 Nb that are targeting proteins like tumor necrosis factor receptor 1 and C-X-C motif chemokine ligand 10, respectively, have demonstrated anti-inflammatory effects in experimental models. These Nbs showed promise in reducing neuroinflammation and preserving myelin and neurons [135,136,137].
3.2.6.4. Nbs for Brain Tumor
The therapeutic effects of Nbs in brain tumor therapy is a promising and rapidly developing field, due to their small size, which allows them to cross the BBB and selectively target tumor cells [134]. Conjugating Nbs with therapeutic agents enables precise drug delivery, minimizing damage to healthy brain tissue. Nbs can also be engineered to inhibit tumor growth by targeting specific signaling pathways or antigens. While early preclinical studies show promise, further research and clinical trials are needed to fully explore Nbs' potential in brain tumor treatment [134].
Glioblastomas often exhibit amplification of the EGFR gene, playing a crucial role in their development [138]. Nbs targeting alpha EGFR have been developed to inhibit EGF-induced signaling and cell proliferation in vitro [139]. Combination strategies involving Nbs specific to EGFR and a pro-apoptotic molecule TNF-related apoptosis-inducing ligand, delivered by neural stem cells, effectively reduced glioblastoma growth and invasiveness in laboratory and animal studies [36,138]. Brain metastases, originating from primary tumors in other body parts, present significant clinical challenges [140]. Nbs offer new approaches, such as using them for imaging and therapy in brain metastases originating from HER2-positive breast cancer. Nbs like 2Rs15d, a Nb targeting HER2, labeled with radioactive isotopes, enable improved imaging and treatment outcomes. Nbs-based innovations in liposomal systems have the potential to improve the identification of brain metastases and treatment outcomes by overcoming obstacles including drug resistance and BBB penetration [141,142].
3.2.6.5. Nbs for Infectious Disease Affecting CNS
Nbs are rarely employed in CNS infectious diseases, but the latest studies have shown promising outcomes in this field [143]. Notably, Nbs VHHG9 and VHHF3 were designed to target Neisseria adhesin A, which can disrupt the interaction with its cell receptors. These Nbs reduced Neisseria meningitidis adhesion to human microvascular endothelial cells and hindered its BBB penetration in vitro, offering a potential treatment approach [143]. Additionally, for diseases like rabies, an infectious disease that primarily affects the CNS, Nbs have gained attention. They offer a cost-effective solution with high affinity. Novel bispecific Nbs extend albumin's bloodstream longevity, potentially enhancing post-exposure vaccine prophylaxis [144,145].
3.2.6.6. Nbs for Other Neurological Disorders
Nbs hold potential benefits for other neurodegenerative disorders, including Huntington's disease. This disease is caused by genetic mutations leading to abnormal Huntingtin protein accumulation in nerve cells. In 2015, the first Nb targeting the N-terminal domain of the Huntingtin protein was developed, showing strong affinity for both mutant and wild-type forms of the protein [146,147]. Considering the intracellular aggregation of the Huntingtin protein, Nbs used as intrabodies could be a promising avenue for development, with early evidence of feasibility from other sources of intrabodies [146,147].
Overall, Nbs offer a versatile and promising approach to diagnosing and treating neurological disorders. However, further studies are needed to fully validate their safety and effectiveness in clinical applications.
3.2.7. Nbs for Other Disorders
Nbs are expanding their therapeutic applications in fields like ophthalmic, genetic, immune-mediated, and other diseases [148,149,150]. In ophthalmologic conditions, their unique features reported promise in targeting ocular components like vascular endothelial growth factor and immune-related elements for treating retinal diseases and neovascular disorders [149]. In fungal keratitis, a severe corneal infection, a specialized Nb targeting β-glucan, effectively disrupted fungal growth when combined with natamycin, surpassing traditional treatments in safety and efficacy [150]. Nbs also demonstrated potential in treating autosomal dominant retinitis pigmentosa, stabilizing P23H mutant rhodopsin proteins, and presenting a novel therapeutic approach for this genetic eye condition [151].
In immune-mediated disorders such as psoriasis, inflammatory bowel disease, spondyloarthritis, and knee osteoarthritis Nbs show potential in targeting inflammation and key molecules [148,152]. For instance, in spondyloarthritis, studies indicate their potential to target interleukin-17 (e.g. Sonelokimab) and TNF, offers effective therapeutic options [152]. This trivalent novel Nb, Sonelokimab, exhibited protective effects against cartilage degradation and aggrecan turnover in knee osteoarthritis, as shown in preliminary results from a phase I study [153]. Moreover, Sonelokimab has shown promising safety and effectiveness in other immune-mediated disorders such as psoriasis [148]. On the other hand, Ozoralizumab is a trivalent bispecific albumin-conjugated Nb that targets TNF-α [148]. It has currently undergone a Phase III trial as a potential therapy for rheumatoid arthritis [148] and was authorized in Japan in September 2022 for the treatment of rheumatoid arthritis that is not adequately controlled by currently existing therapies [31].
In the field of genetic diseases, some researchers focused on voltage-gated sodium channels, which play a role in initiating rapid action potentials in skeletal and cardiac muscles [109,154]. Developing Nbs that target voltage-gated sodium channels presents a challenge due to potential cross-reactivity. However, specific Nbs (Nb17 and Nb82) binding with nanomolar affinity to voltage-gated sodium channel 1.4 and voltage-gated sodium channel 1.5 isoforms, crucial for skeletal and cardiac muscles, have been developed, highlighting potential therapeutic methods [109,154].
4. Summary and Conclusions
From naturally occurring HCAb found in camelids in 1993, a particular class of small antibody fragments known as Nbs, VHHs, or single-domain antibodies was created. Their streamlined structure with just one variable domain encompassing the antigen-binding region serves as their primary point of differentiation. Their diminutive size offers a variety of benefits, including the ability to reach intricate or difficult-to-reach target sites that may be inaccessible to larger antibody competitors and enhanced tolerance to harsh environmental conditions including temperature and pH fluctuations.
This in-depth examination emphasizes how important Nbs' therapeutic potential is in the discovery and creation of novel medicines. It emphasizes that Nbs have therapeutic potential in a variety of conditions, including oncology, infectious (viral, bacterial, and parasitic), metabolic, neurological (Alzheimer's, Parkinson's, multiple sclerosis, brain tumor, and other disorders affecting the CNS), ophthalmologic, immune-mediated, and genetic disorders. To fully prove their effectiveness and safety in clinical settings, more study is needed. By providing an up-to-date and comprehensive assessment of the therapeutic indications of Nbs, this study serves as a valuable resource for academics, medical experts, and pharmaceutical industry professionals.
Let's sum up by saying that therapeutic Nbs have recently dominated drug discovery and research. Despite the fact that Nbs and other antibody fragments share the same ability to bind antigens, their unique characteristics—such as their small size—allow them to be more soluble, long-lasting, and capable of penetrating deeper into tissues. Nbs have a promising future ahead of them as versatile molecular tools that may be applied in a range of domains, including imaging, therapies, and diagnosis.
Author Contributions
F.H. designed the review outline; F.H. and G.D. searched the literature; wrote the first and final draft; and reviewed the revised version. S.K. contributed to writing part of the first draft. R.K. wrote the final draft and the revised version. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BBB | Blood-brain barrier |
| CNS | Central nervous system |
| COVID-19 | Coronavirus disease-2019 |
| EGFR | Epidermal growth factor receptor |
| ELISA | Enzyme-linked immunosorbent assays |
| Fab | Fragment antigen binding |
| Fc | Fragment crystallizable |
| HCAb | Heavy-chain-only antibodies |
| HER | Human epidermal growth factor receptor |
| HIV | Human immunodeficiency viruses |
| Ig | Immunoglobulins |
| LFIA | Lateral flow immunoassays |
| Mab | Monoclonal antibody |
| MAbs | Monoclonal antibodies |
| Nb | Nanobody |
| Nbs | Nanobodies |
| PCR | Polymerase chain reaction |
| PET | Positron emission tomography |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| scFvs | Single-chain variable fragments |
| SPECT | Single photon emission computed tomography |
| TNF | Tumor necrosis factor |
| VHHs | Variable domain of heavy chain of HCAb |
References
- Megha, K.; Mohanan, P. Role of immunoglobulin and antibodies in disease management. Int. J. Biol. Macromol. 2020, 169, 28–38. [CrossRef]
- Singh, S.; Kumar, N.K.; Dwiwedi, P.; Charan, J.; Kaur, R.; Sidhu, P.; Chugh, V.K. Monoclonal Antibodies: A Review. Curr. Clin. Pharmacol. 2018, 13, 85–99. [CrossRef]
- Lu, R.-M.; Hwang, Y.-C.; Liu, I.-J.; Lee, C.-C.; Tsai, H.-Z.; Li, H.-J.; Wu, H.-C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1–30. [CrossRef]
- Wang, Q.; Chen, Y.; Park, J.; Liu, X.; Hu, Y.; Wang, T.; McFarland, K.; Betenbaugh, M.J. Design and Production of Bispecific Antibodies. Antibodies 2019, 8, 43. [CrossRef]
- Rajpal, A.; Strop, P.; Yeung, Y.A.; Chaparro-Riggers, J.; Pons, J. Introduction: Antibody Structure and Function. 2013.
- Chiu, M.L.; Goulet, D.R.; Teplyakov, A.; Gilliland, G.L. Antibody Structure and Function: The Basis for Engineering Therapeutics. Antibodies 2019, 8, 55. [CrossRef]
- Asaadi, Y.; Jouneghani, F.F.; Janani, S.; Rahbarizadeh, F. A comprehensive comparison between camelid nanobodies and single chain variable fragments. Biomark. Res. 2021, 9, 1–20. [CrossRef]
- Liu, S.; Liu, X. IgG N-glycans. Advances in Clinical Chemistry 2021, 105, 1-47.
- Jenner, E. On the Origin of the Vaccine Inoculation. The Medical and physical journal 1801, 5, 505–508.
- Rees, A. Antibodies: A History of Their Discovery and Properties. In Introduction to Antibody Engineering, Rüker, F., Wozniak-Knopp, G., Eds. Springer International Publishing: Cham, 2021; 10.1007/978-3-030-54630-4_2pp. 5-39.
- Packer, D. The history of the antibody as a tool. Acta Histochem. 2021, 123, 151710. [CrossRef]
- Davies, D.R.; Chacko, S. Antibody structure. Accounts Chem. Res. 1993, 26, 421–427. [CrossRef]
- Lemieux, R.U.; Spohr, U. How Emil Fischer was led to the lock and key concept for enzyme specificity. Adv Carbohydr Chem Biochem 1994, 50, 1-20.
- Hosseini, S.; Vázquez-Villegas, P.; Rito-Palomares, M.; Martinez-Chapa, S.O.; Hosseini, S.; Vázquez-Villegas, P.; Rito-Palomares, M.; Martinez-Chapa, S.O. Fundamentals and history of ELISA: the evolution of the immunoassays until invention of ELISA. Enzyme-linked Immunosorbent Assay (ELISA) From A to Z 2018, 1-18.
- Grillo-López, A.J. The first antibody therapy for cancer: a personal experience. Expert Rev. Anticancer. Ther. 2013, 13, 399–406. [CrossRef]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [CrossRef]
- Mahmuda, A.; Bande, F.; Al-Zihiry, K.J.K.; Abdulhaleem, N.; Majid, R.A.; Hamat, R.A.; Abdullah, W.O.; Unyah, Z. Monoclonal antibodies: A review of therapeutic applications and future prospects. Trop. J. Pharm. Res. 2017, 16, 713–722. [CrossRef]
- Castelli, M.S.; McGonigle, P.; Hornby, P.J. The pharmacology and therapeutic applications of monoclonal antibodies. Pharmacol. Res. Perspect. 2019, 7, e00535. [CrossRef]
- Erstad, B.L.; Davis, L.E. Fixed Versus Body-Sized-Based Dosing of Monoclonal Antibodies. Ann. Pharmacother. 2023, 58, 91–95. [CrossRef]
- Gupta, S.K.; Chaudhary, P. Therapeutic human monoclonal antibodies. In Biomedical Translational Research: Drug Design and Discovery, Springer: 2022; pp. 401-418.
- Behl, A.; Wani, Z.A.; Das, N.N.; Parmar, V.S.; Len, C.; Malhotra, S.; Chhillar, A.K. Monoclonal antibodies in breast cancer: A critical appraisal. Crit. Rev. Oncol. 2023, 183, 103915. [CrossRef]
- Jin, S.; Sun, Y.; Liang, X.; Gu, X.; Ning, J.; Xu, Y.; Chen, S.; Pan, L. Emerging new therapeutic antibody derivatives for cancer treatment. Signal Transduct. Target. Ther. 2022, 7, 1–28. [CrossRef]
- Suzuki, M.; Kato, C.; Kato, A. Therapeutic antibodies: their mechanisms of action and the pathological findings they induce in toxicity studies. J. Toxicol. Pathol. 2015, 28, 133–139. [CrossRef]
- Ioele, G.; Chieffallo, M.; Occhiuzzi, M.A.; De Luca, M.; Garofalo, A.; Ragno, G.; Grande, F. Anticancer Drugs: Recent Strategies to Improve Stability Profile, Pharmacokinetic and Pharmacodynamic Properties. Molecules 2022, 27, 5436. [CrossRef]
- Navabi, P.; Ganjalikhany, M.R.; Jafari, S.; Dehbashi, M.; Ganjalikhani-Hakemi, M. Designing and generating a single-chain fragment variable (scFv) antibody against IL2Rα (CD25): An in silico and in vitro study. Iranian Journal of Basic Medical Sciences 2021, 24, 360–368. [CrossRef]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hammers, C.; Songa, E.B.; Bendahman, N.; Hammers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448.
- Streltsov, V.A.; Carmichael, J.A.; Nuttall, S.D. Structure of a shark IgNAR antibody variable domain and modeling of an early-developmental isotype. Protein Sci. 2005, 14, 2901–2909. [CrossRef]
- Pillay, T.S.; Muyldermans, S. Application of single-domain antibodies (“nanobodies”) to laboratory diagnosis. Ann Lab Med 2021, 41, 549-558.
- Siontorou, C.G. Nanobodies as novel agents for disease diagnosis and therapy. Int. J. Nanomed. 2013, 8, 4215–27. [CrossRef]
- Duggan, S. Caplacizumab: First Global Approval. Drugs 2018, 78, 1639–1642. [CrossRef]
- Keam, S.J. Ozoralizumab: First Approval. Drugs 2022, 83, 87–92. [CrossRef]
- Muyldermans, S. Nanobodies: Natural Single-Domain Antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [CrossRef]
- Abbady, A.; Al-Daoude, A.; Al-Mariri, A.; Zarkawi, M.; Muyldermans, S. Chaperonin GroEL a Brucella immunodominant antigen identified using Nanobody and MALDI-TOF-MS technologies. Veter- Immunol. Immunopathol. 2012, 146, 254–263. [CrossRef]
- Wang, W.; Yuan, J.; Jiang, C. Applications of nanobodies in plant science and biotechnology. Plant Mol. Biol. 2020, 105, 43–53. [CrossRef]
- Tang, H.; Gao, Y.; Han, J. Application Progress of the Single Domain Antibody in Medicine. Int. J. Mol. Sci. 2023, 24, 4176. [CrossRef]
- Jovcevska, I.; Muyldermans, S. The Therapeutic Potential of Nanobodies. BioDrugs 2020, 34, 11–26. [CrossRef]
- Beghein, E.; Gettemans, J. Nanobody Technology: A Versatile Toolkit for Microscopic Imaging, Protein–Protein Interaction Analysis, and Protein Function Exploration. Front. Immunol. 2017, 8, 771. [CrossRef]
- Singh, S.; Murillo, G.; Richner, J.; Singh, S.P.; Berleth, E.; Kumar, V.; Mehta, R.; Ramiya, V.; Parihar, A.S. A Broad-Based Characterization of a Cell-Penetrating, Single Domain Camelid Bi-Specific Antibody Monomer That Targets STAT3 and KRAS Dependent Cancers. Int. J. Mol. Sci. 2022, 23, 7565. [CrossRef]
- Muyldermans, S. A guide to: Generation and design of nanobodies. FEBS J. 2021, 288, 2084–2102. [CrossRef]
- Contreras, M.A.; Serrano-Rivero, Y.; González-Pose, A.; Salazar-Uribe, J.; Rubio-Carrasquilla, M.; Soares-Alves, M.; Parra, N.C.; Camacho-Casanova, F.; Sánchez-Ramos, O.; Moreno, E. Design and Construction of a Synthetic Nanobody Library: Testing Its Potential with a Single Selection Round Strategy. Molecules 2023, 28, 3708. [CrossRef]
- de Marco, A. Recombinant expression of nanobodies and nanobody-derived immunoreagents. Protein expression and purification 2020, 172, 105645.
- Jin, B.-k.; Odongo, S.; Radwanska, M.; Magez, S. Nanobodies: a review of generation, diagnostics and therapeutics. International Journal of Molecular Sciences 2023, 24, 5994.
- Harmand, T.J.; Islam, A.; Pishesha, N.; Ploegh, H.L. Nanobodies as in vivo, non-invasive, imaging agents. RSC Chemical Biology 2021, 2, 685-701.
- Bao, G.; Tang, M.; Zhao, J.; Zhu, X. Nanobody: a promising toolkit for molecular imaging and disease therapy. EJNMMI Res. 2021, 11, 1–13. [CrossRef]
- Küppers, J.; Kürpig, S.; Bundschuh, R.A.; Essler, M.; Lütje, S. Radiolabeling Strategies of Nanobodies for Imaging Applications. Diagnostics 2021, 11, 1530. [CrossRef]
- Xavier, C.; Blykers, A.; Vaneycken, I.; D'Huyvetter, M.; Heemskerk, J.; Lahoutte, T.; Devoogdt, N.; Caveliers, V. 18F-nanobody for PET imaging of HER2 overexpressing tumors. Nucl. Med. Biol. 2016, 43, 247–252. [CrossRef]
- Warnders, F.J.; van Scheltinga, A.G.T.; Knuehl, C.; van Roy, M.; de Vries, E.F.; Kosterink, J.G.; de Vries, E.G.; Lub-de Hooge, M.N. Human epidermal growth factor receptor 3–specific tumor uptake and biodistribution of 89Zr-MSB0010853 visualized by real-time and noninvasive PET imaging. Journal of Nuclear Medicine 2017, 58, 1210-1215.
- Jailkhani, N.; Ingram, J.R.; Rashidian, M.; Rickelt, S.; Tian, C.; Mak, H.; Jiang, Z.; Ploegh, H.L.; Hynes, R.O. Noninvasive imaging of tumor progression, metastasis, and fibrosis using a nanobody targeting the extracellular matrix. Proc. Natl. Acad. Sci. USA 2019, 116, 14181–14190. [CrossRef]
- Bridoux, J.; Broos, K.; Lecocq, Q.; Debie, P.; Martin, C.; Ballet, S.; Raes, G.; Neyt, S.; Vanhove, C.; Breckpot, K.; et al. Anti-Human PD-L1 Nanobody for Immuno-PET Imaging: Validation of a Conjugation Strategy for Clinical Translation. Biomolecules 2020, 10, 1388. [CrossRef]
- Li, D.; Cheng, S.; Zou, S.; Zhu, D.; Zhu, T.; Wang, P.; Zhu, X. Immuno-PET Imaging of 89Zr Labeled Anti-PD-L1 Domain Antibody. Mol. Pharm. 2018, 15, 1674–1681. [CrossRef]
- Li, D.; Zou, S.; Cheng, S.; Song, S.; Wang, P.; Zhu, X. Monitoring the Response of PD-L1 Expression to Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors in Nonsmall-Cell Lung Cancer Xenografts by Immuno-PET Imaging. Mol. Pharm. 2019, 16, 3469–3476. [CrossRef]
- Berland, L.; Kim, L.; Abousaway, O.; Mines, A.; Mishra, S.; Clark, L.; Hofman, P.; Rashidian, M. Nanobodies for medical imaging: about ready for prime time? Biomolecules 2021, 11, 637.
- Wang, W.; Hou, X.; Yang, X.; Liu, A.; Tang, Z.; Mo, F.; Yin, S.; Lu, X. Highly sensitive detection of CTLA-4-positive T-cell subgroups based on nanobody and fluorescent carbon quantum dots. Oncology Letters 2019, 18, 109-116.
- Lecocq, Q.; Zeven, K.; De Vlaeminck, Y.; Martens, S.; Massa, S.; Goyvaerts, C.; Raes, G.; Keyaerts, M.; Breckpot, K.; Devoogdt, N. Noninvasive Imaging of the Immune Checkpoint LAG-3 Using Nanobodies, from Development to Pre-Clinical Use. Biomolecules 2019, 9, 548. [CrossRef]
- Lecocq, Q.; Awad, R.M.; De Vlaeminck, Y.; de Mey, W.; Ertveldt, T.; Goyvaerts, C.; Raes, G.; Thielemans, K.; Keyaerts, M.; Devoogdt, N.; et al. Single-Domain Antibody Nuclear Imaging Allows Noninvasive Quantification of LAG-3 Expression by Tumor-Infiltrating Leukocytes and Predicts Response of Immune Checkpoint Blockade. J. Nucl. Med. 2021, 62, 1638–1644. [CrossRef]
- Rashidian, M.; Ingram, J.R.; Dougan, M.; Dongre, A.; Whang, K.A.; LeGall, C.; Cragnolini, J.J.; Bierie, B.; Gostissa, M.; Gorman, J.; et al. Predicting the response to CTLA-4 blockade by longitudinal noninvasive monitoring of CD8 T cells. J. Exp. Med. 2017, 214, 2243–2255. [CrossRef]
- Rashidian, M.; LaFleur, M.W.; Verschoor, V.L.; Dongre, A.; Zhang, Y.; Nguyen, T.H.; Kolifrath, S.; Aref, A.R.; Lau, C.J.; Paweletz, C.P.; et al. Immuno-PET identifies the myeloid compartment as a key contributor to the outcome of the antitumor response under PD-1 blockade. Proc. Natl. Acad. Sci. 2019, 116, 16971–16980. [CrossRef]
- Zhao, H.; Wang, C.; Yang, Y.; Sun, Y.; Wei, W.; Wang, C.; Wan, L.; Zhu, C.; Li, L.; Huang, G.; et al. ImmunoPET imaging of human CD8+ T cells with novel 68Ga-labeled nanobody companion diagnostic agents. J. Nanobiotechnology 2021, 19, 1–11. [CrossRef]
- Gondry, O.; Xavier, C.; Raes, L.; Heemskerk, J.; Devoogdt, N.; Everaert, H.; Breckpot, K.; Lecocq, Q.; Decoster, L.; Fontaine, C.; et al. Phase I Study of [68Ga]Ga-Anti-CD206-sdAb for PET/CT Assessment of Protumorigenic Macrophage Presence in Solid Tumors (MMR Phase I). J. Nucl. Med. 2023, 64, 1378–1384. [CrossRef]
- Punjabi, M.; Xu, L.; Ochoa-Espinosa, A.; Kosareva, A.; Wolff, T.; Murtaja, A.; Broisat, A.; Devoogdt, N.; Kaufmann, B.A. Ultrasound molecular imaging of atherosclerosis with nanobodies: translatable microbubble targeting murine and human VCAM (vascular cell adhesion molecule) 1. Arteriosclerosis, Thrombosis, and Vascular Biology 2019, 39, 2520-2530.
- Iljina, M.; Hong, L.; Horrocks, M.H.; Ludtmann, M.H.; Choi, M.L.; Hughes, C.D.; Ruggeri, F.S.; Guilliams, T.; Buell, A.K.; Lee, J.-E.; et al. Nanobodies raised against monomeric ɑ-synuclein inhibit fibril formation and destabilize toxic oligomeric species. BMC Biol. 2017, 15, 57–57. [CrossRef]
- Gerdes, C.; Waal, N.; Offner, T.; Fornasiero, E.F.; Wender, N.; Verbarg, H.; Manzini, I.; Trenkwalder, C.; Mollenhauer, B.; Strohäker, T.; et al. A nanobody-based fluorescent reporter reveals human α-synuclein in the cell cytosol. Nat. Commun. 2020, 11, 1–13. [CrossRef]
- Demine, S.; Ribeiro, R.G.; Thevenet, J.; Marselli, L.; Marchetti, P.; Pattou, F.; Kerr-Conte, J.; Devoogdt, N.; Eizirik, D.L. A nanobody-based nuclear imaging tracer targeting dipeptidyl peptidase 6 to determine the mass of human beta cell grafts in mice. Diabetologia 2019, 63, 825–836. [CrossRef]
- Muyldermans, S. Applications of nanobodies. Annual review of animal biosciences 2021, 9, 401-421.
- Koczula, K.M.; Gallotta, A. Lateral flow assays. Essays in biochemistry 2016, 60, 111-120.
- De Genst, E.; Silence, K.; Decanniere, K.; Conrath, K.; Loris, R.; Kinne, J.; Muyldermans, S.; Wyns, L. Molecular basis for the preferential cleft recognition by dromedary heavy-chain antibodies. Proc. Natl. Acad. Sci. 2006, 103, 4586–4591. [CrossRef]
- Mohseni, A.; Molakarimi, M.; Taghdir, M.; Sajedi, R.H.; Hasannia, S. Exploring single-domain antibody thermostability by molecular dynamics simulation. J. Biomol. Struct. Dyn. 2018, 37, 3686–3696. [CrossRef]
- Torres, J.E.P.; Goossens, J.; Ding, J.; Li, Z.; Lu, S.; Vertommen, D.; Naniima, P.; Chen, R.; Muyldermans, S.; Sterckx, Y.G.-J.; et al. Development of a Nanobody-based lateral flow assay to detect active Trypanosoma congolense infections. Sci. Rep. 2018, 8, 1–15. [CrossRef]
- Vincke, C.; Gutiérrez, C.; Wernery, U.; Devoogdt, N.; Hassanzadeh-ghassabeh, G.; Muyldermans, S. Generation of Single Domain Antibody Fragments Derived from Camelids and Generation of Manifold Constructs. In Antibody Engineering: Methods and Protocols; Second Edition 2012; Humana Press: Totowa, NJ, USA, 2012; pp. 145–176.
- Goossens, J.; Sein, H.; Lu, S.; Radwanska, M.; Muyldermans, S.; Sterckx, Y.G.-J.; Magez, S. Functionalization of gold nanoparticles with nanobodies through physical adsorption. Anal. Methods 2017, 9, 3430–3440. [CrossRef]
- Li, Z.; Torres, J.E.P.; Goossens, J.; Vertommen, D.; Caljon, G.; Sterckx, Y.G.-J.; Magez, S. An Unbiased Immunization Strategy Results in the Identification of Enolase as a Potential Marker for Nanobody-Based Detection of Trypanosoma evansi. Vaccines 2020, 8, 415. [CrossRef]
- Odongo, S.; Sterckx, Y.G.J.; Stijlemans, B.; Pillay, D.; Baltz, T.; Muyldermans, S.; Magez, S. An Anti-proteome Nanobody Library Approach Yields a Specific Immunoassay for Trypanosoma congolense Diagnosis Targeting Glycosomal Aldolase. PLOS Neglected Trop. Dis. 2016, 10, e0004420. [CrossRef]
- Pinto, J.; Odongo, S.; Lee, F.; Gaspariunaite, V.; Muyldermans, S.; Magez, S.; Sterckx, Y.G.-J. Structural basis for the high specificity of a Trypanosoma congolense immunoassay targeting glycosomal aldolase. PLOS Neglected Trop. Dis. 2017, 11, e0005932. [CrossRef]
- Gelkop, S.; Sobarzo, A.; Brangel, P.; Vincke, C.; Romão, E.; Fedida-Metula, S.; Strom, N.; Ataliba, I.; Mwiine, F.N.; Ochwo, S.; et al. The Development and Validation of a Novel Nanobody-Based Competitive ELISA for the Detection of Foot and Mouth Disease 3ABC Antibodies in Cattle. Front. Veter- Sci. 2018, 5, 250. [CrossRef]
- Katz, H.E. Antigen sensing via nanobody-coated transistors. Nat. Biomed. Eng. 2021, 5, 639–640. [CrossRef]
- Campuzano, S.; Salema, V.; Moreno-Guzmán, M.; Gamella, M.; Yáñez-Sedeño, P.; Fernández, L.; Pingarrón, J. Disposable amperometric magnetoimmunosensors using nanobodies as biorecognition element. Determination of fibrinogen in plasma. Biosens. Bioelectron. 2014, 52, 255–260. [CrossRef]
- Guo, K.; Wustoni, S.; Koklu, A.; Díaz-Galicia, E.; Moser, M.; Hama, A.; Alqahtani, A.A.; Ahmad, A.N.; Alhamlan, F.S.; Shuaib, M.; et al. Rapid single-molecule detection of COVID-19 and MERS antigens via nanobody-functionalized organic electrochemical transistors. Nat. Biomed. Eng. 2021, 5, 666–677. [CrossRef]
- Noltes, M.E.; van Dam, G.M.; Nagengast, W.B.; van der Zaag, P.J.; Slart, R.H.J.A.; Szymanski, W.; Kruijff, S.; Dierckx, R.A.J.O. Let’s embrace optical imaging: a growing branch on the clinical molecular imaging tree. Eur. J. Nucl. Med. 2021, 48, 4120–4128. [CrossRef]
- Oliveira, S.; Van Dongen, G.a.M.S.; Stigter-Van Walsum, M.; Roovers, R.C.; Stam, J.C.; Mali, W.; Van Diest, P.J.; Van Bergen En Henegouwen, P.M.P. Rapid Visualization of Human Tumor Xenografts through Optical Imaging with a Near-Infrared Fluorescent Anti–Epidermal Growth Factor Receptor Nanobody. Mol. Imaging 2012, 11, 33–46. [CrossRef]
- Kijanka, M.M.; van Brussel, A.S.; van der Wall, E.; Mali, W.P.; van Diest, P.J.; van Bergen en Henegouwen, P.M.; Oliveira, S. Optical imaging of pre-invasive breast cancer with a combination of VHHs targeting CAIX and HER2 increases contrast and facilitates tumour characterization. Ejnmmi Research 2016, 6, 1-13.
- Broisat, A.; Hernot, S.; Toczek, J.; De Vos, J.; Riou, L.M.; Martin, S.; Ahmadi, M.; Thielens, N.; Wernery, U.; Caveliers, V.; et al. Nanobodies Targeting Mouse/Human VCAM1 for the Nuclear Imaging of Atherosclerotic Lesions. Circ. Res. 2012, 110, 927–937. [CrossRef]
- Broisat, A.; Toczek, J.; Dumas, L.S.; Ahmadi, M.; Bacot, S.; Perret, P.; Slimani, L.; Barone-Rochette, G.; Soubies, A.; Devoogdt, N. 99mTc-cAbVCAM1-5 imaging is a sensitive and reproducible tool for the detection of inflamed atherosclerotic lesions in mice. Journal of Nuclear Medicine 2014, 55, 1678-1684.
- Xavier, C.; Vaneycken, I.; D’huyvetter, M.; Heemskerk, J.; Keyaerts, M.; Vincke, C.; Devoogdt, N.; Muyldermans, S.; Lahoutte, T.; Caveliers, V. Synthesis, preclinical validation, dosimetry, and toxicity of 68Ga-NOTA-anti-HER2 Nanobodies for iPET imaging of HER2 receptor expression in cancer. Journal of Nuclear Medicine 2013, 54, 776-784.
- Keyaerts, M.; Xavier, C.; Everaert, H.; Vaneycken, I.; Fontaine, C.; Decoster, L.; Vanhoeij, M.; Caveliers, V.; Lahoutte, T. Phase II trial of HER2-PET/CT using 68Ga-anti-HER2 VHH1 for characterization of HER2 presence in brain metastases of breast cancer patients. Ann. Oncol. 2019, 30, iii25–iii26. [CrossRef]
- D'Huyvetter, M.; Vincke, C.; Xavier, C.; Aerts, A.; Impens, N.; Baatout, S.; De Raeve, H.; Muyldermans, S.; Caveliers, V.; Devoogdt, N.; et al. Targeted Radionuclide Therapy with A 177Lu-labeled Anti-HER2 Nanobody. Theranostics 2014, 4, 708–720. [CrossRef]
- Bannas, P.; Hambach, J.; Koch-Nolte, F. Nanobodies and Nanobody-Based Human Heavy Chain Antibodies As Antitumor Therapeutics. Front. Immunol. 2017, 8, 1603. [CrossRef]
- Verhaar, E.R.; Woodham, A.W.; Ploegh, H.L. Nanobodies in cancer. In Proceedings of Seminars in immunology; p. 101425.
- nbsp;Roovers, R.C.; Vosjan, M.J.W.D.; Laeremans, T.; El Khoulati, R.; De Bruin, R.C.G.; Ferguson, K.M.; Verkleij, A.J.; Van Dongen, G.A.M.S.; Van Bergen en Henegouwen, P.M.P.A biparatopic anti-EGFR nanobody efficiently inhibits solid tumour growth. Int. J. Cancer 2011, 129, 2013–2024. [CrossRef]
- Kijanka, M.; Dorresteijn, B.; Oliveira, S.; van Bergen en Henegouwen, P.M.P. Nanobody-based cancer therapy of solid tumors. Nanomedicine 2015, 10, 161–174. [CrossRef]
- Broos, K.; Lecocq, Q.; Xavier, C.; Bridoux, J.; Nguyen, T.T.; Corthals, J.; Schoonooghe, S.; Lion, E.; Raes, G.; Keyaerts, M.; et al. Evaluating a Single Domain Antibody Targeting Human PD-L1 as a Nuclear Imaging and Therapeutic Agent. Cancers 2019, 11, 872. [CrossRef]
- Yang, E.Y.; Shah, K. Nanobodies: Next Generation of Cancer Diagnostics and Therapeutics. Front. Oncol. 2020, 10, 1182. [CrossRef]
- Sadeghi, A.; Behdani, M.; Muyldermans, S.; Habibi-Anbouhi, M.; Kazemi-Lomedasht, F. Development of a mono-specific anti-VEGF bivalent nanobody with extended plasma half-life for treatment of pathologic neovascularization. Drug testing and analysis 2020, 12, 92-100.
- Sanaei, M.; Setayesh, N.; Sepehrizadeh, Z.; Mahdavi, M.; Yazdi, M.H. Nanobodies in Human Infections: Prevention, Detection, and Treatment. Immunol. Investig. 2019, 49, 875–896. [CrossRef]
- Mei, Y.; Chen, Y.; Sivaccumar, J.P.; An, Z.; Xia, N.; Luo, W. Research progress and applications of nanobody in human infectious diseases. Front. Pharmacol. 2022, 13, 963978. [CrossRef]
- Xiang, Y.; Nambulli, S.; Xiao, Z.; Liu, H.; Sang, Z.; Duprex, W.P.; Schneidman-Duhovny, D.; Zhang, C.; Shi, Y. Versatile and multivalent nanobodies efficiently neutralize SARS-CoV-2. Science 2020, 370, 1479-1484.
- Zupancic, J.M.; Schardt, J.S.; Desai, A.A.; Makowski, E.K.; Smith, M.D.; Pornnoppadol, G.; Garcia de Mattos Barbosa, M.; Cascalho, M.; Lanigan, T.M.; Tessier, P.M. Engineered multivalent nanobodies potently and broadly neutralize SARS-CoV-2 variants. Advanced therapeutics 2021, 4, 2100099.
- Saied, A.A.; Metwally, A.A.; Alobo, M.; Shah, J.; Sharun, K.; Dhama, K. Bovine-derived antibodies and camelid-derived nanobodies as biotherapeutic weapons against SARS-CoV-2 and its variants: a review article. International Journal of Surgery 2022, 98, 106233.
- Detalle, L.; Stohr, T.; Palomo, C.; Piedra, P.A.; Gilbert, B.E.; Mas, V.; Millar, A.; Power, U.F.; Stortelers, C.; Allosery, K.; et al. Generation and Characterization of ALX-0171, a Potent Novel Therapeutic Nanobody for the Treatment of Respiratory Syncytial Virus Infection. Antimicrob. Agents Chemother. 2016, 60, 6–13. [CrossRef]
- Ibañez, L.I.; De Filette, M.; Hultberg, A.; Verrips, T.; Temperton, N.; Weiss, R.A.; Vandevelde, W.; Schepens, B.; Vanlandschoot, P.; Saelens, X. Nanobodies With In Vitro Neutralizing Activity Protect Mice Against H5N1 Influenza Virus Infection. J. Infect. Dis. 2011, 203, 1063–1072. [CrossRef]
- Wichgers Schreur, P.J.; van de Water, S.; Harmsen, M.; Bermudez-Mendez, E.; Drabek, D.; Grosveld, F.; Wernike, K.; Beer, M.; Aebischer, A.; Daramola, O. Multimeric single-domain antibody complexes protect against bunyavirus infections. Elife 2020, 9, e52716.
- Kennedy, P.J.; Oliveira, C.; Granja, P.L.; Sarmento, B. Monoclonal antibodies: technologies for early discovery and engineering. Crit. Rev. Biotechnol. 2017, 38, 394–408. [CrossRef]
- Li, H.-C.; Lo, S.-Y. Hepatitis C virus: Virology, diagnosis and treatment. World journal of hepatology 2015, 7, 1377.
- Liver, E.A.f.T.S.o.T. EASL recommendations on treatment of hepatitis C 2016. Journal of hepatology 2017, 66, 153-194.
- Wilken, L.; McPherson, A. Application of camelid heavy-chain variable domains (VHHs) in prevention and treatment of bacterial and viral infections. Int. Rev. Immunol. 2017, 37, 69–76. [CrossRef]
- Barta, M.L.; Shearer, J.P.; Arizmendi, O.; Tremblay, J.M.; Mehzabeen, N.; Zheng, Q.; Battaile, K.P.; Lovell, S.; Tzipori, S.; Picking, W.D.; et al. Single-domain antibodies pinpoint potential targets within Shigella invasion plasmid antigen D of the needle tip complex for inhibition of type III secretion. J. Biol. Chem. 2017, 292, 16677–16687. [CrossRef]
- Smolarek, D.; Hattab, C.; Hassanzadeh-Ghassabeh, G.; Cochet, S.; Gutiérrez, C.; de Brevern, A.G.; Udomsangpetch, R.; Picot, J.; Grodecka, M.; Wasniowska, K.; et al. A recombinant dromedary antibody fragment (VHH or nanobody) directed against human Duffy antigen receptor for chemokines. Cell. Mol. Life Sci. 2010, 67, 3371–3387. [CrossRef]
- Stijlemans, B.; Caljon, G.; Natesan, S.K.A.; Saerens, D.; Conrath, K.; Pérez-Morga, D.; Skepper, J.N.; Nikolaou, A.; Brys, L.; Pays, E.; et al. High Affinity Nanobodies against the Trypanosome brucei VSG Are Potent Trypanolytic Agents that Block Endocytosis. PLOS Pathog. 2011, 7, e1002072. [CrossRef]
- Obishakin, E.; Stijlemans, B.; Santi-Rocca, J.; Vandenberghe, I.; Devreese, B.; Muldermans, S.; Bastin, P.; Magez, S. Generation of a Nanobody Targeting the Paraflagellar Rod Protein of Trypanosomes. PLOS ONE 2014, 9, e115893. [CrossRef]
- Bocancia-Mateescu, L.-A.; Stan, D.; Mirica, A.-C.; Ghita, M.G.; Stan, D.; Ruta, L.L. Nanobodies as Diagnostic and Therapeutic Tools for Cardiovascular Diseases (CVDs). Pharmaceuticals 2023, 16, 863. [CrossRef]
- Broggini, L.; Giono, M.; Speranzini, V.; Barzago, M.; Palladini, G.; Diomede, L.; Pappone, C.; Ricagno, S. Nanobodies as novel potential drugs to target cardiac light chain amyloidosis. Cardiovasc. Res. 2022, 118. [CrossRef]
- Wingler, L.M.; Feld, A.P. Nanobodies as Probes and Modulators of Cardiovascular G Protein–Coupled Receptors. J. Cardiovasc. Pharmacol. 2021, 80, 342–353. [CrossRef]
- McMahon, C.; Staus, D.P.; Wingler, L.M.; Wang, J.; Skiba, M.A.; Elgeti, M.; Hubbell, W.L.; Rockman, H.A.; Kruse, A.C.; Lefkowitz, R.J. Synthetic nanobodies as angiotensin receptor blockers. Proc. Natl. Acad. Sci. 2020, 117, 20284–20291. [CrossRef]
- Li, T.; Shen, Y.; Lin, F.; Fu, W.; Liu, S.; Wang, C.; Liang, J.; Fan, X.; Ye, X.; Tang, Y.; et al. Targeting RyR2 with a phosphorylation site–specific nanobody reverses dysfunction of failing cardiomyocytes in rats. FASEB J. 2019, 33, 7467–7478. [CrossRef]
- Sillen, M.; Declerck, P.J. Targeting PAI-1 in Cardiovascular Disease: Structural Insights Into PAI-1 Functionality and Inhibition. Front. Cardiovasc. Med. 2020, 7. [CrossRef]
- De Genst, E.; Foo, K.S.; Xiao, Y.; Rohner, E.; de Vries, E.; Sohlmér, J.; Witman, N.; Hidalgo, A.; Kolstad, T.R.S.; Louch, W.E.; et al. Blocking phospholamban with VHH intrabodies enhances contractility and relaxation in heart failure. Nat. Commun. 2022, 13, 1–18. [CrossRef]
- Callewaert, F.; Roodt, J.; Ulrichts, H.; Stohr, T.; van Rensburg, W.J.; Lamprecht, S.; Rossenu, S.; Priem, S.; Willems, W.; Holz, J.-B. Evaluation of efficacy and safety of the anti-VWF Nanobody ALX-0681 in a preclinical baboon model of acquired thrombotic thrombocytopenic purpura. Blood The Journal of the American Society of Hematology 2012, 120, 3603–3610. [CrossRef]
- Marturano, A.; Hendrickx, M.L.; Falcinelli, E.; Sebastiano, M.; Guglielmini, G.; Hassanzadeh-Ghassabeh, G.; Muyldermans, S.; Declerck, P.J.; Gresele, P. Development of anti-matrix metalloproteinase-2 (MMP-2) nanobodies as potential therapeutic and diagnostic tools. Nanomedicine: Nanotechnology, Biol. Med. 2019, 24, 102103. [CrossRef]
- Jooss, N.J.; Smith, C.W.; Slater, A.; Montague, S.J.; Di, Y.; O'shea, C.; Thomas, M.R.; Henskens, Y.M.; Heemskerk, J.W.; Watson, S.P. Anti-GPVI nanobody blocks collagen-and atherosclerotic plaque–induced GPVI clustering, signaling, and thrombus formation. Journal of Thrombosis and Haemostasis 2022, 20, 2617-2631.
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789. [CrossRef]
- Knopman, D.S.; Amieva, H.; Petersen, R.C.; Chételat, G.; Holtzman, D.M.; Hyman, B.T.; Nixon, R.A.; Jones, D.T. Alzheimer disease. Nature reviews Disease primers 2021, 7, 33.
- Habiba, U.; Descallar, J.; Kreilaus, F.; Adhikari, U.K.; Kumar, S.; Morley, J.W.; Bui, B.V.; Koronyo-Hamaoui, M.; Tayebi, M. Detection of retinal and blood Aβ oligomers with nanobodies. Alzheimer's Dementia: Diagn. Assess. Dis. Monit. 2021, 13, e12193. [CrossRef]
- Danis, C.; Dupré, E.; Zejneli, O.; Caillierez, R.; Arrial, A.; Bégard, S.; Mortelecque, J.; Eddarkaoui, S.; Loyens, A.; Cantrelle, F.-X.; et al. Inhibition of Tau seeding by targeting Tau nucleation core within neurons with a single domain antibody fragment. Mol. Ther. 2022, 30, 1484–1499. [CrossRef]
- Marino, M.; Zhou, L.; Rincon, M.Y.; Callaerts-Vegh, Z.; Verhaert, J.; Wahis, J.; Creemers, E.; Yshii, L.; Wierda, K.; Saito, T. AAV-mediated delivery of an anti-BACE1 VHH alleviates pathology in an Alzheimer's disease model. EMBO molecular medicine 2022, 14, e09824.
- Marino, M.; Holt, M.G. AAV Vector-Mediated Antibody Delivery (A-MAD) in the Central Nervous System. Front. Neurol. 2022, 13, 870799. [CrossRef]
- Zhao, L.; Meng, F.; Li, Y.; Liu, S.; Xu, M.; Chu, F.; Li, C.; Yang, X.; Luo, L. Multivalent Nanobody Conjugate with Rigid, Reactive Oxygen Species Scavenging Scaffold for Multi-Target Therapy of Alzheimer's Disease. Advanced Materials 2023, 2210879.
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson's disease. The Lancet 2021, 397, 2284-2303.
- Haddad, F.; Sawalha, M.; Khawaja, Y.; Najjar, A.; Karaman, R. Dopamine and Levodopa Prodrugs for the Treatment of Parkinson’s Disease. Molecules 2017, 23, 40. [CrossRef]
- Vuchelen, A.; O’day, E.; De Genst, E.; Pardon, E.; Wyns, L.; Dumoulin, M.; Dobson, C.M.; Christodoulou, J.; Hsu, S.-T.D. 1H, 13C and 15N assignments of a camelid nanobody directed against human α-synuclein. Biomol. NMR Assignments 2009, 3, 231–233. [CrossRef]
- De Genst, E.J.; Guilliams, T.; Wellens, J.; O'Day, E.M.; Waudby, C.A.; Meehan, S.; Dumoulin, M.; Hsu, S.-T.D.; Cremades, N.; Verschueren, K.H.; et al. Structure and Properties of a Complex of α-Synuclein and a Single-Domain Camelid Antibody. J. Mol. Biol. 2010, 402, 326–343. [CrossRef]
- Guilliams, T.; El-Turk, F.; Buell, A.K.; O'Day, E.M.; Aprile, F.A.; Esbjörner, E.K.; Vendruscolo, M.; Cremades, N.; Pardon, E.; Wyns, L.; et al. Nanobodies Raised against Monomeric α-Synuclein Distinguish between Fibrils at Different Maturation Stages. J. Mol. Biol. 2013, 425, 2397–2411. [CrossRef]
- El-Turk, F.; Newby, F.N.; De Genst, E.; Guilliams, T.; Sprules, T.; Mittermaier, A.; Dobson, C.M.; Vendruscolo, M. Structural Effects of Two Camelid Nanobodies Directed to Distinct C-Terminal Epitopes on α-Synuclein. Biochemistry 2016, 55, 3116–3122. [CrossRef]
- Hmila, I.; Vaikath, N.N.; Majbour, N.K.; Erskine, D.; Sudhakaran, I.P.; Gupta, V.; Ghanem, S.S.; Islam, Z.; Emara, M.M.; Abdesselem, H.B. Novel engineered nanobodies specific for N-terminal region of alpha-synuclein recognize Lewy-body pathology and inhibit in-vitro seeded aggregation and toxicity. The FEBS Journal 2022, 289, 4657-4673.
- Singh, R.K.; Soliman, A.; Guaitoli, G.; Störmer, E.; von Zweydorf, F.; Dal Maso, T.; Oun, A.; Van Rillaer, L.; Schmidt, S.H.; Chatterjee, D. Nanobodies as allosteric modulators of Parkinson’s disease–associated LRRK2. Proceedings of the National Academy of Sciences 2022, 119, e2112712119.
- Zheng, F.; Pang, Y.; Li, L.; Pang, Y.; Zhang, J.; Wang, X.; Raes, G. Applications of nanobodies in brain diseases. Front. Immunol. 2022, 13, 978513. [CrossRef]
- Steeland, S.; Puimège, L.; Vandenbroucke, R.E.; Van Hauwermeiren, F.; Haustraete, J.; Devoogdt, N.; Hulpiau, P.; Leroux-Roels, G.; Laukens, D.; Meuleman, P.; et al. Generation and Characterization of Small Single Domain Antibodies Inhibiting Human Tumor Necrosis Factor Receptor 1. J. Biol. Chem. 2015, 290, 4022–4037. [CrossRef]
- Steeland, S.; Van Ryckeghem, S.; Van Imschoot, G.; De Rycke, R.; Toussaint, W.; Vanhoutte, L.; Vanhove, C.; De Vos, F.; Vandenbroucke, R.E.; Libert, C. TNFR1 inhibition with a Nanobody protects against EAE development in mice. Sci. Rep. 2017, 7, 13646. [CrossRef]
- Sadeghian-Rizi, T.; Behdani, M.; Khanahmad, H.; Sadeghi, H.M.; Jahanian-Najafabadi, A. Generation and characterization of a functional nanobody against inflammatory chemokine CXCL10, as a novel strategy for the treatment of multiple sclerosis. CNS & Neurological Disorders-Drug Targets (Formerly Current Drug Targets-CNS & Neurological Disorders) 2019, 18, 141-148.
- van de Water, J.A.J.M.; Bagci-Onder, T.; Agarwal, A.S.; Wakimoto, H.; Roovers, R.C.; Zhu, Y.; Kasmieh, R.; Bhere, D.; Henegouwen, P.M.P.V.B.E.; Shah, K. Therapeutic stem cells expressing variants of EGFR-specific nanobodies have antitumor effects. Proc. Natl. Acad. Sci. 2012, 109, 16642–16647. [CrossRef]
- Figarella-Branger, D.; Appay, R.; Metais, A.; Tauziède-Espariat, A.; Colin, C.; Rousseau, A.; Varlet, P. La classification de l’OMS 2021 des tumeurs du système nerveux central. In Proceedings of Annales de Pathologie; pp. 367-382.
- Ellingson, B.M.; Boxerman, J.; Barboriak, D.; Erickson, B.J.; Smits, M.; Nelson, S.J.; Gerstner, E.; Alexander, B.; Goldmacher, G.; Wick, W.; et al. Consensus recommendations for a standardized Brain Tumor Imaging Protocol in clinical trials. Neuro-Oncology 2015, 17, 1188–1198. [CrossRef]
- Zhou, Z.; Vaidyanathan, G.; McDougald, D.; Kang, C.M.; Balyasnikova, I.; Devoogdt, N.; Ta, A.N.; McNaughton, B.R.; Zalutsky, M.R. Fluorine-18 Labeling of the HER2-Targeting Single-Domain Antibody 2Rs15d Using a Residualizing Label and Preclinical Evaluation. Mol. Imaging Biol. 2017, 19, 867–877. [CrossRef]
- Vaneycken, I.; Devoogdt, N.; Van Gassen, N.; Vincke, C.; Xavier, C.; Wernery, U.; Muyldermans, S.; Lahoutte, T.; Caveliers, V. Preclinical screening of anti-HER2 nanobodies for molecular imaging of breast cancer. FASEB J. 2011, 25, 2433–2446. [CrossRef]
- Kulkarni, A.; Mochnáčová, E.; Majerova, P.; Čurlík, J.; Bhide, K.; Mertinková, P.; Bhide, M. Single Domain Antibodies Targeting Receptor Binding Pockets of NadA Restrain Adhesion of Neisseria meningitidis to Human Brain Microvascular Endothelial Cells. Front. Mol. Biosci. 2020, 7. [CrossRef]
- Terryn, S.; Francart, A.; Lamoral, S.; Hultberg, A.; Rommelaere, H.; Wittelsberger, A.; Callewaert, F.; Stohr, T.; Meerschaert, K.; Ottevaere, I.; et al. Protective Effect of Different Anti-Rabies Virus VHH Constructs against Rabies Disease in Mice. PLOS ONE 2014, 9, e109367. [CrossRef]
- Terryn, S.; Francart, A.; Rommelaere, H.; Stortelers, C.; Van Gucht, S. Post-exposure Treatment with Anti-rabies VHH and Vaccine Significantly Improves Protection of Mice from Lethal Rabies Infection. PLOS Neglected Trop. Dis. 2016, 10, e0004902. [CrossRef]
- Schut, M.H.; Pepers, B.A.; Klooster, R.; van der Maarel, S.M.; el Khatabi, M.; Verrips, T.; Dunnen, J.T.D.; van Ommen, G.-J.B.; van Roon-Mom, W.M.C. Selection and characterization of llama single domain antibodies against N-terminal huntingtin.. Neurol. Sci. 2014, 36, 429–34. [CrossRef]
- Colby, D.W.; Chu, Y.; Cassady, J.P.; Duennwald, M.; Zazulak, H.; Webster, J.M.; Messer, A.; Lindquist, S.; Ingram, V.M.; Wittrup, K.D. Potent inhibition of huntingtin aggregation and cytotoxicity by a disulfide bond-free single-domain intracellular antibody. Proc. Natl. Acad. Sci. 2004, 101, 17616–17621. [CrossRef]
- Wang, J.; Kang, G.; Yuan, H.; Cao, X.; Huang, H.; de Marco, A. Research Progress and Applications of Multivalent, Multispecific and Modified Nanobodies for Disease Treatment. Front. Immunol. 2022, 12, 838082. [CrossRef]
- Beltran, W.A.; Ripolles-Garcia, A.; Sato, Y.; Gray, A.; Wolfe, J.H.; Luz-Madrigal, A.; Philipps, M.J.; Gamm, D.M.; Aguirre, G.K.; Aguirre, G.D. Structural and functional assessment after subretinal transplantation of human iPSC-derived photoreceptor precursors in a canine model of end stage retinal degeneration. Investigative Ophthalmology & Visual Science 2023, 64, 766-766.
- Liu, X.; Sui, J.; Li, C.; Wang, Q.; Peng, X.; Meng, F.; Xu, Q.; Jiang, N.; Zhao, G.; Lin, J. The preparation and therapeutic effects of β-glucan-specific nanobodies and nanobody-natamycin conjugates in fungal keratitis. Acta Biomater. 2023, 169, 398–409. [CrossRef]
- Wu, A.; Salom, D.; Sander, C.L.; Pardon, E.; Steyaert, J.; Kiser, P.D.; Palczewski, K. NANOBODY rescues adRP phenotype in a P23H adRP cell model, suggesting therapeutic potential. Investigative Ophthalmology & Visual Science 2022, 63, 88–A0061-0088–A0061.
- Navarro-Compán, V.; Ermann, J.; Poddubnyy, D. A glance into the future of diagnosis and treatment of spondyloarthritis. Ther. Adv. Musculoskelet. Dis. 2022, 14. [CrossRef]
- Li, S.; Cao, P.; Chen, T.; Ding, C. Latest insights in disease-modifying osteoarthritis drugs development. Ther. Adv. Musculoskelet. Dis. 2023, 15. [CrossRef]
- Srinivasan, L.; Alzogaray, V.; Selvakumar, D.; Nathan, S.; Yoder, J.B.; Wright, K.M.; Klinke, S.; Nwafor, J.N.; Labanda, M.S.; Goldbaum, F.A.; et al. Development of high-affinity nanobodies specific for NaV1.4 and NaV1.5 voltage-gated sodium channel isoforms. J. Biol. Chem. 2022, 298, 101763. [CrossRef]
Figure 1.
Structure of conventional antibody (A), single-chain variable fragment (scFv) (B), camelid heavy chain only antibody (HCAb) (C), and nanobody (D). CH1 (constant domain 1 of the heavy chain), CH2 (constant domain 2 of the heavy chain), CH3 (constant domain 3 of the heavy chain), CL (constant domain of the light chain), Fab (fragment antigen binding), Fc ( fragment crystallizable), VH (variable domain of heavy chain), VL (variable domain of light chain), and VHHs (variable domain of HCAb).
Figure 1.
Structure of conventional antibody (A), single-chain variable fragment (scFv) (B), camelid heavy chain only antibody (HCAb) (C), and nanobody (D). CH1 (constant domain 1 of the heavy chain), CH2 (constant domain 2 of the heavy chain), CH3 (constant domain 3 of the heavy chain), CL (constant domain of the light chain), Fab (fragment antigen binding), Fc ( fragment crystallizable), VH (variable domain of heavy chain), VL (variable domain of light chain), and VHHs (variable domain of HCAb).
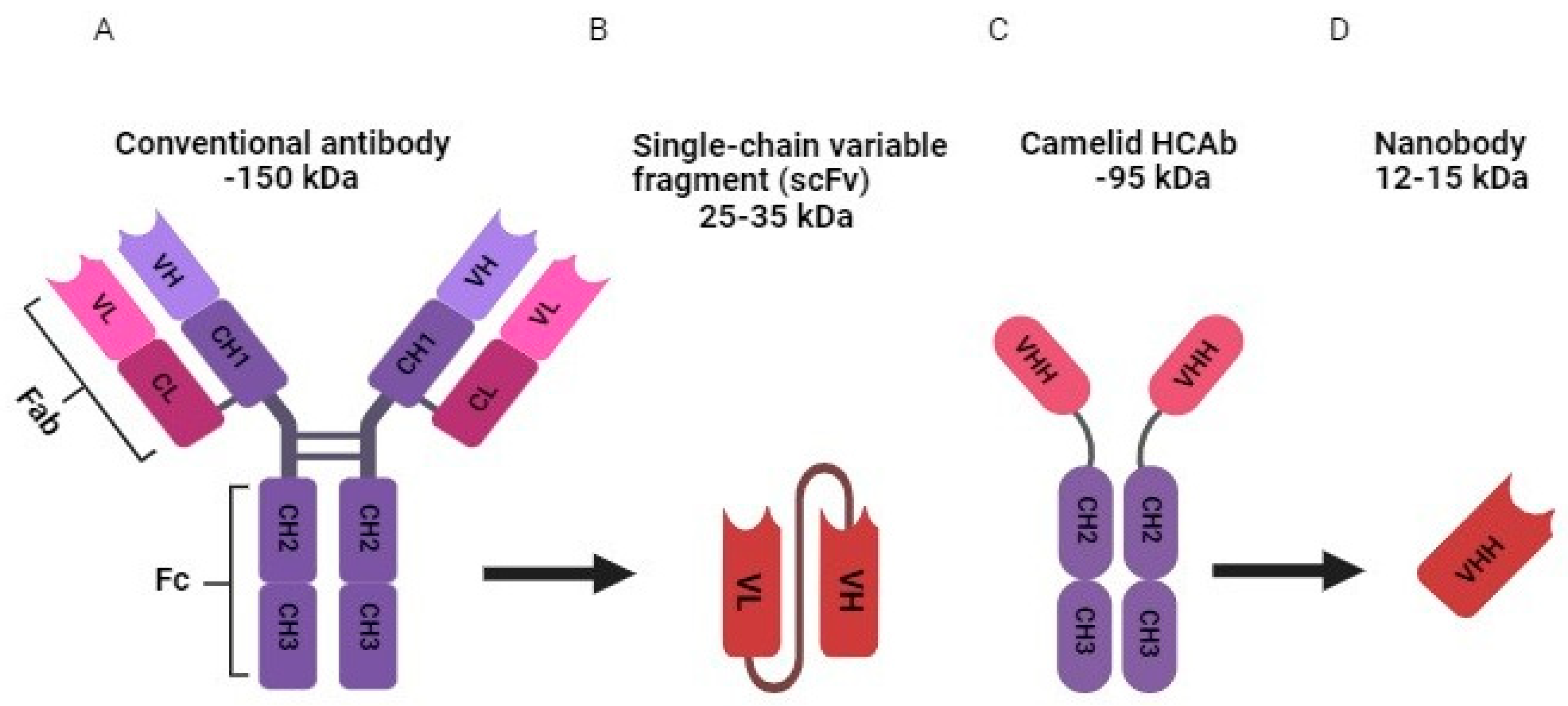
Figure 2.
Representation of nanobody production procedure.
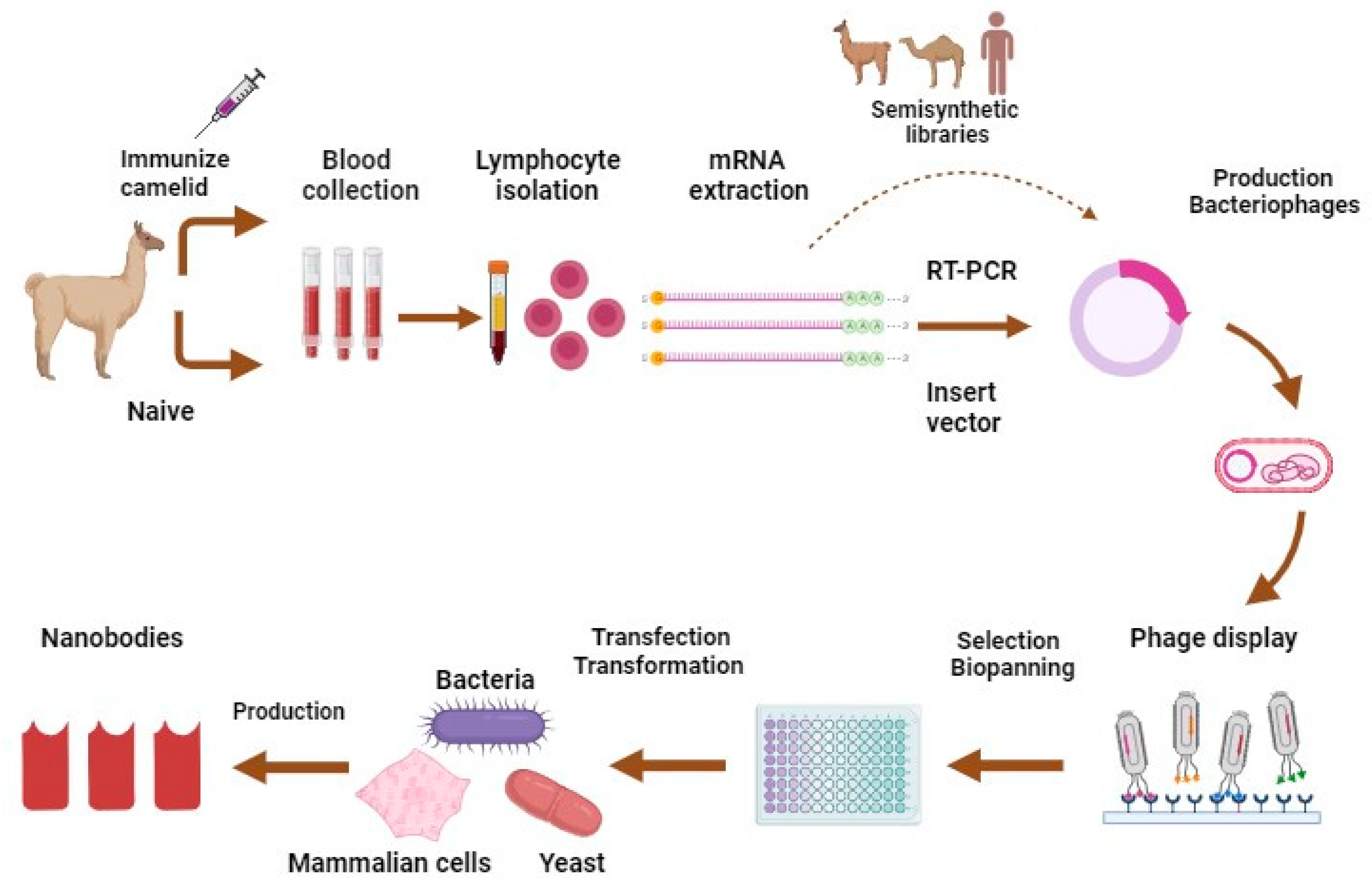
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



