
Submitted:
24 October 2023
Posted:
25 October 2023
You are already at the latest version
Abstract
AML is a highly aggressive and heterogeneous form of hematological cancer. Proteomics-based stratification of patients into more refined subgroups may contribute to a more precise characterization of the patient-derived AML cells. Here, we reanalyzed liquid chromatography-tandem mass spectrometry (LC-MS/MS) generated proteomic and phosphoproteomic data from 26 chemoresistant/relapse (RELAPSE) and 15 relapse-free (REL_FREE) AML patients. We considered not only the RELAPSE and REL_FREE characteristics but also integrated the French-American-British (FAB) classification, along with considering the presence of nucleophosmin 1 (NPM1) mutation and cytogenetically normal AML. We found a significant number of differentially enriched proteins (911) and phosphoproteins (257) between the various FAB subtypes in RELAPSE patients. Patients with the myeloblastic M1/M2 subtype showed higher levels of RNA processing-related routes and lower levels of signaling related to terms like translation and degranulation, when compared to the M4/M5 subtype. Moreover, we found that high abundance of proteins associated to mitochondrial translation and oxidative phosphorylation, particularly observed in the RELAPSE M4/M5 NPM1 mutated subgroup, distinguishes relapsing from non-relapsing AML patient cells with the FAB subtype M4/M5. Thus, the discovery of subtype-specific biomarkers through proteomic profiling may complement the existing classification system for AML and potentially aid in selecting personalized treatment strategies for individual patients.
Keywords:
acute myeloid leukemia
; mass spectrometry
; proteomic
; phosphoproteomic
; relapse
; mitochondria
; FAB subtypes
1. Introduction
Acute myeloid leukemia (AML) is a highly aggressive type of blood cancer that arises from hematopoietic stem or progenitor cells. Its heterogeneity is attributed to several factors, including different mutations, potential cytogenetic abnormalities, changes in gene and protein expression, and disrupted signaling transduction [1,2]. Prior to the identification of genetic and cytogenetic abnormalities, the classification of AML into subtypes relied primarily on the morphological characteristics of the leukemic cells. In the 1970s, the French-American-British (FAB) Cooperative Group proposed a classification system for AML, which divided AML patients into eight FAB subgroups (M0-M7) based on morphological, cytochemical, and maturation characteristics of the leukemic cells [3]. Later, the importance of cytogenetics and molecular genetic features in the stratification of patients into risk groups, such as those with favorable prognosis and high complete remission (CR) rates, as well as intermediate and poor/adverse outcome, was recognized [4,5]. Based on new knowledge of clinical and genetical abnormalities, the World Health Organization (WHO) and the European Leukemia Network (ELN) have recently updated their risk classification and treatment recommendations [1,2]. At initial AML diagnosis, patients with nucleophosmin 1 (NPM1) mutation without Fms related receptor tyrosine kinase 3-internal tandem duplication (FLT3-ITD) are categorized as favorable, whereas mutated NPM1 along with FLT3-ITD are now classified as intermediate risk in the revised ELN risk classification [1]. Additionally, mutated NPM1 with adverse-risk cytogenetics are now classified as adverse.
In adults, the NPM1 mutation and morphological signs of differentiation, along with the expression of the CD33 differentiation marker and absence of the CD34 stem cell marker, are more commonly observed in the monocytic FAB-M4/M5 subgroups and less frequently seen in the myeloblastic FAB-M0/M1/M2 subgroups [6,7,8]. Monocytic differentiation is also associated with generally high constitutive cytokine release [9,10], i.e., these cells differ with regard to the communication with neighboring stromal cells in their common bone marrow microenvironment. Gene expression profiling of AML has provided valuable insight into distinct gene expression signatures observed in different patient subgroups characterized by specific genetic and cytogenetic abnormalities [7,11]. These profiles include unique gene expression patterns associated with NPM1 mutations and also the expression levels of three genes (annexin A3, ANXA3; protein S100-A9, S100A9 and Wilms tumor 1, WT1) that can differentiate between AML FAB subtypes M1 from M2 [12,13]. Similarly, proteomic profiling was conducted to compare differences in protein expression between two subtypes of myeloblastic AML: M1 without maturation and M2 with maturation. The study identified five proteins (ANXA A1; ANXA A3; plastin-2, PLSL; 6-phosphogluconate dehydrogenase, 6PGD; actin cytoplasmatic 2, ACTG) that exhibited differential expression, allowing for the distinction between the two subtypes [14].
Recent advancements in quantitative proteomics, especially those based on liquid chromatography-tandem mass spectrometry (LC-MS/MS), have made it possible to accurately quantify AML-disease related proteins and phosphorylation sites in a substantial number of patients with different disease characteristics and treatment responses [15,16,17,18,19,20,21,22,23,24,25]. In a previous study, we employed quantitative LC-MS/MS analysis to compare the proteome and phosphoproteome of pretreatment AML cells obtained at the time of diagnosis. Specifically, we focused on two distinct patient subgroups: 15 patients who achieved leukemia-free survival for more than five years and 26 patients who experienced relapse despite undergoing intensive chemotherapy (henceforth REL_FREE and RELAPSE patients, respectively) [26]. The heterogeneity of AML emphasizes the importance of stratification of patients into disease subgroups. Here, we further categorized the original cohort of 41 AML patients into more refined subgroups, considering not only relapse categories but also incorporating the FAB-M1/M2 and FAB-M4/M5 classification subgroups (i.e., morphological signs of AML cell differentiation), as well as NPM1 mutation and cytogenetically normal AML. The aim of our present study was thus to identify at the first time of diagnosis the heterogeneity of proteomic and phosphoproteomic AML cell profiles for patients that later develop leukemia relapse after intensive and potentially curative chemotherapy.
2. Materials and Methods
2.1. Patient and Sample Collection
We here reanalyzed our previously published LC-MS/MS-based proteomic and phosphoproteomic cohort of primary cells from 41 AML patients at time of diagnosis [26]. These patients represent a consecutive group of relatively young patients that received intensive and potentially curative antileukemic treatment. The study should be regarded as population-based because our department was responsible for diagnosis and treatment of AML in a defined geographical area during the defined time period. The patients received only intensive induction and consolidation cytotoxic therapy without stem cell transplantation as their initial therapy, and REL_FREE patients after this treatment were classified after observation for at least five years.
Primary AML cells were density gradient-separated from peripheral blood (PB) of untreated patients with blast counts (leukemic cells) exceeding 80% of the circulating leukocytes. The cells were cryopreserved and stored in liquid nitrogen until analyzed. Quantification was performed by combining protein lysates with a heavy-labeled AML-super SILAC (stable isotope labeling by amino acids in cell culture) mixture [27]. Detailed methods and patient information on FAB type, cytogenetic and mutational analysis from the time of first diagnosis is comprehensively given in our previously described cohort of 41 AML patients [26,28]. The criteria for FAB classification of patients have been described in detail previously [29]; this system is regarded as a standardized and well-described system to characterize and classify AML patients with regard to the differentiation status of their leukemic cells [30,31]. All raw data and MaxQuant output files can be found in the ProteomeXchange consortium with the dataset identifier PXD014997.
2.2. Data Analysis
Patients were grouped after clinical progression as RELAPSE and REL_FREE if they had not relapsed after a five-years observation time from the initial induction chemotherapy/consolidation therapy. The Perseus 2.0.7.0 bioinformatics platform was used for functional and statistical analysis of the proteomics and phosphoproteomics data [32]. SILAC ratios were inverted and log2 transformed. Categorical annotation rows were used for stratification of patients into disease subgroups based on FAB classification, mutated NPM1 and cytogenetic status (Table 1). The generated AML subgroups included eight RELAPSE M1/M2 (REL_M1/2_all), 12 RELAPSE M4/M5 (REL_M4/5_all) and 14 REL_FREE M4/M5 (REL_F_M4/5_all). M0 patients were left out as there were no M0 REL_FREE patients and four out of the five M0 RELAPSE patients did not showed NPM1 mutations.
Within the REL_M1/2_all subgroup, only two patients showed a 4-base pair insertion (Ins) mutation in NPM1 and five patients had a normal cytogenetic status. This subgroup was not further stratified for analysis. Within the M4/M5 patients, five RELAPSE M4/M5 and eight REL_FREE M4/M5 showed the Ins NPM1 mutation (REL_M4/5_mut and REL_F_M4/5_mut, respectively); and seven RELAPSE M4/M5 and nine REL_FREE M4/M5 displayed a normal cytogenetic status 46, XY or XX (REL_M4/5_CN and REL_F_M4/5_CN, respectively). These subgroups were normalized by using width adjustment. Proteins and phosphosites (localization probability > 0.75) with minimum three individual SILAC ratios for each patient subgroup were selected for statistical analysis. ANOVA multiple sample test was performed with a threshold P-value < 0.05 to test for significant difference between means for the proteins and phosphosites between the subgroups. Hierarchical clustering of significantly differential proteins with ANOVA was done using the Euclidean function and complete linkage. A post hoc Turkey’s honest significance difference (HSD) test with FDR < 0.05 was performed on the ANOVA significant different pairs of protein and phosphosites. Reactome pathway, Gene Ontology (GO) and KEGG pathway enrichment analyses were obtained with the Enrichr gene set search engine [33,34,35]. Protein-protein interaction (PPI) network analysis was performed with STRING database version 11.5 [36]. Networks were visualized using the Cytoscape platform v3.10.0 [37]. The ClusterONE plugin was used to identify protein groups of high cohesiveness [38]. Phosphosite motif analysis was done with the web-based WebLogo application [39]. Venn diagrams were generated by BioVenn [40].
3. Results
Given the high number of possible individual comparisons among the patient subgroups (Table 1), proteomic data results will be presented as a Venn diagram of the regulated proteins from different subgroup comparisons. Enrichment of Reactome pathways and PPI of overlapped and subgroup-specific regulated proteins will be shown as well. Phosphoproteomic data results will be additionally presented as sequence logos of the surrounding amino acids of the differentially regulated phosphorylation sites obtained from overlapped and subgroup-specific phosphosites.
3.1. Distinct Protein Expression and Site Phosphorylation Patterns in RELAPSE Patients for AML FAB Subtypes M1/M2 and M4/M5
The heterogeneity of AML stresses the importance of categorizing patients into disease subgroups based on various considerations such as cytogenetic, genetic mutations, protein expression and aberrant PTM patterns. Based on information about clinical progression and pathological processes, we recategorized our original cohort with 41 AML patients [26] into more defined subgroups based on FAB, cytogenetic and NPM1 mutation parameters (Table 1). We quantified a total of 6,781 proteins and 12,309 class I protein phosphorylation sites. Among these, 4,601 proteins and 3,148 phosphosites had at least three valid SILAC ratios in each patient subgroup (Supplementary file 1 and 2). By comparing the different AML disease subgroups for patients that relapse after chemotherapy, we found a substantial number of proteins (911) and phosphosites (257) that exhibited statistically significant (ANOVA, post hoc Turkey’s HSD with FDR < 0.05) differences between the FAB classes (RELAPSE M1/M2 vs RELAPSE M4/M5 subgroups, Table 1): 162 proteins and 32 phosphosites were upregulated, and 268 proteins and 94 phosphosites were downregulated for the comparison REL_M1/2_all vs REL_M4/5_all; 34 proteins and 24 phosphosites were upregulated, and 283 proteins and 86 phosphosites were downregulated for REL_M1/2_all vs REL_M4/5_mut; 131 proteins and 44 phosphosites were upregulated, and 382 proteins and 89 phosphosites were downregulated for REL_M1/2_all vs REL_M4/5_CN (Supplementary File S3).
Seventy-eight regulated proteins were identified in the REL_M1/2_all comparisons against the different REL_M4/5 subgroups (Figure 1a). Through the utilization of Reactome pathways and PPI network analyses we observed distinct patterns of protein expression linked to the AML FAB subtypes M1/M2 and M4/M5. The basal transcription machinery such as RNA processing and RNA polymerase transcription were significantly enriched in RELAPSE patients with the myeloblastic subtype M1/M2 (Figure 1b,c), whereas terms like translation, neutrophil degranulation and intracellular protein/vesicle transport were more abundant terms for RELAPSE patients with the monocytic subtypes M4/M5 (Figure 2b,c). Additionally, we found that hematopoietic stem cell differentiation was enriched for the M4/M5 subtype (Figure 2c). We observed a high number of regulated proteins (166) that were detected in the different REL_M4/5 comparisons against REL_M1/2_all (Figure 2a).
Thirteen differentially regulated phosphorylation sites were identified in the REL_M1/2_all comparisons against the different REL_M4/5 subgroups (Figure 3a). By conducting GO and KEGG pathway analyses on the differently regulated phosphorylation sites, we confirmed the enrichment of basal transcription machinery, as well as RNA and DNA binding processes, in relapse patients with M1/M2 subtype compared to the M4/M5 subtypes (Figure 3b). Forty-two differentially regulated phosphorylation sites were identified in the different REL_M4/5 comparisons against REL_M1/2_all (Figure 4a). The relapse M4/M5 patient subgroups exhibited higher site-specific phosphorylation on proteins linked to RNA binding and high translational activity when compared to the M1/M2 subgroup (Figure 4b). PPI network analysis confirmed the higher phosphorylation of translational proteins in the REL_M4/5_all, REL_ M4/5_mut and REL_M4/5_CN patient groups (Figure 4c). Additionally, the REL_M4/5_all and REL_M4/5_CN subgroups showed increased phosphorylation of proteins involved in DNA damage response and protein synthesis.
To identify the potential kinases responsible for phosphorylating the differently regulated phosphosites, we conducted WebLogo substrate motif analysis. As illustrated in Figure 3c, we observed a prevalent pSP followed by a pSXXE motif in REL_M1/2_all compared to all RELAPSE M4/M5 subtypes, suggesting higher activity of extracellular signal-regulated kinases (ERK1/2) and casein kinase 2 (CSNK2) in the M1/M2 subtype. The pSP motif for ERK1/2 and RXXpS motif for protein kinase A (PRKA) and C (PRKC) were prominent in REL_M1/2_all compared to REL_M4/5_all and REL_M4/5_CN. Multiple kinase substrate motifs, i.e., pSP, pSXXE/D and RXXpS, were found in the REL_M1/2_all vs REL_M4/5_CN comparison.
The pSP and RXXpS substrate motifs were found in all the REL_M4/M5 subgroups compared to REL_M1/2 indicating an increased activity of ERK1/2 and PRKA/PRKC in AML patients with the subtype M4/M5 (Figure 4d). However, the acidic amino acids in close proximity to the differentially regulated phosphorylation sites of the CSNK2 substrate motif was found in the overlapping sequences flanking the regulated phosphosites from the REL_M4/5_all vs REL_M1/2_all and REL_M4/5_CN vs REL_M1/2_all and in the separated REL_M4/5_CN vs REL_M1/2_all and REL_M4/5_mut vs REL_M1/2_all comparisons.
3.2. High Mitochondrial Protein Expression Splits Relapsing from Non-Relapsing AML Patients with the FAB Subtypes M4/M5
We conducted a comparison between the proteome and phosphoproteome profiles of patients with the monocytic FAB-M4/M5 subtypes who later experienced relapse to those who did not. Among the 4,601 proteins and 3,148 phosphosites which had at least three valid SILAC ratios in each patient subgroup, we found a substantial number of proteins (850) and phosphosites (294) that exhibited statistically significant (ANOVA, post hoc Turkey’s HSD with FDR < 0.05) differences between the relapse status (RELAPSE M4/M5 vs REL_FREE M4/M5 subgroups; Table 1, Supplementary file 4 and 5).
For the FAB-M4/M5 group with NPM1 mutation we quantified 193 differently expressed proteins and 34 differentially regulated phosphorylation sites for RELAPSE and REL_FREE patient comparisons (ANOVA, post hoc Turkey’s HSD with FDR < 0.05). Among these, 145 proteins and 17 phosphosites were more abundant, while 48 proteins and 17 phosphosites were less abundant for the REL_M4/5_mut vs REL_F_M4/5_mut comparison (Supplementary file 3). The number of regulated proteins and phosphosites for the FAB-M4/M5 subgroup with normal cytogenetics or without any other stratification for RELAPSE compared to REL_FREE patients was low (Figure 5a). Through Reactome pathways enrichment analysis, we discovered that terms like mitochondrial translation were significantly enriched in the REL_M4/5_mut subgroup (Figure 5b). Importantly, proteins associated to mitochondrial translational activity were also more abundant in REL_M4/5_all and REL_M4/5_CN when compared to the corresponding REL_FREE subgroups. Moreover, significant enrichment of PPI networks required for mitochondrial translation, electron transport and ATP synthesis were found in RELAPSE patients with FAB subtypes M4/M5 (Figure 5c, Supplementary Table S1). The largest network cluster consisted of 37 mitochondrial translational elongation proteins observed in the REL_ M4/5_mut subgroup. The mitochondrial protein interaction networks reflect differences in the mitochondrial energy metabolism and included 11 proteins that are important for the mitochondrial ATP synthase (the final step in the ATP generating electron chain) as well as proteins important for Complex 1 (NDUFB3, NDUFC2, NDUFST, NDUFV2), Complex 3 (UQCRC2, UQCRQ) and cytochrome c oxidase (COX15, COX5B) of the mitochondrial electron chain [41,42,43]. Additionally, this NPM1 mutated REL_M4/5 subgroup exhibited functional PPI clusters for transcription elongation from RNA polymerase II promotor. Hierarchical clustering analyses showed that mitochondrial ribosomal proteins of the translation elongation network and ATP synthases appeared to be better discriminators than electron transport proteins (NADH dehydrogenases) between REL_M4/5 and REL_F_M4/5 patients (Figures S1–S3).
On the other hand, we did not find any regulated protein from the REL_F_M4/5_all vs REL_M4/5_all comparison (Figure 6a, Supplementary File S3) whereas REL_F_M4/5_mut and REL_F_M4/5_CN subgroups were generally more prevalent in terms related to metabolism of nucleotides when compared to the RELAPSE counterparts. Moreover, endosomal sorting, Golgi-to-ER retrograde traffic, and regulation of actin cytoskeleton were also enriched in the REL_F_M4/5_mut subgroup (Figure 6b,c).
Regarding the phosphoproteome, few differentially regulated phosphorylation sites were identified from the different subgroup comparisons (Figure 7a and Figure 8a, Supplementary File S3). However, we observed higher site-specific phosphorylation on proteins associated to transcription and translation such as 90S preribosome, mRNA splicing and poly (A)+ mRNA export in the REL_M4/5_mut subgroup compared to the REL_F_M4/5_mut (Figure 7b). Additionally, GO terms like chromatin organization and DNA binding were significantly enriched in the REL_M4/5_mut subgroup. Through kinase substrate motif analyses, we identified a prevailing pSP motif for ERK1/2 seconded by the pSXXD/E motif for CSK2 in the REL_M4/5_mut subgroup (Figure 7c) while the RXXpS motif for PRKA/PRKC was most prominent in the REL_F_M4/5_mut subgroup (Figure 8c). Interestingly, this subgroup exhibited significantly enriched site-specific phosphorylation on proteins associated to glycolysis and gluconeogenesis (Figure 8b). Moreover, positive regulation of autophagy of mitochondrion was also overrepresented.
4. Discussion
The FAB classification system provides a standardized and well-described system to characterize and classify AML patients with regard to the differentiation status of their leukemic cells [30,31]. The FAB classification has been replaced by the WHO and ELN classification and is no longer considered to have a prognostic role when the mutation status of NPM1 and CCAAT/enhancer-binding protein alpha (CEPBA) is known [44].
Two previous studies have investigated the possible impact of FAB/differentiation on survival for AML patients receiving allogeneic stem cell transplantation. A small early study included 39 patients (median age 14 years) transplanted in the period November 1976 to July 1983. These authors described an adverse prognostic impact of high peripheral blood leukocyte counts at the time of diagnosis (i.e., ≥ 20 x 109/L; P= 0.001) and monocyte morphology (i.e., FAB M4/M5, P=0.05) [45]. It should also be mentioned that most patients in the FAB-M4/M5 groups died from relapse whereas most of the other patients died in remission, but the numbers of patients are low and a reliable statistical comparison is therefore not possible. Another study was based on 1,690 patients transplanted in first complete remission [46]. The patients were classified as having AML not otherwise specified according to the 2016 WHO classification, and the authors described an association between FAB M6/M7 and adverse prognosis, i.e., increased nonrelapse mortality. Finally, the possible association between differentiation and survival after allogeneic stem cell transplantation may not only reflect an association between differentiation and susceptibility to antileukemic treatment; posttransplant survival is possibly also influenced by the immunomodulatory effects (i.e., inhibition of antileukemic immune reactivity) through the expression of immune checkpoint ligands by the AML cells [47].
Even though experimental studies suggest that FAB subtypes differ with regard to the antileukemic effects of daunorubicin and cytarabine (i.e., two drugs commonly combined in the initial induction treatment of AML) [48], the overall results from the clinical studies discussed above suggest that FAB classification/AML cell differentiation has a limited prognostic impact for patients receiving conventional antileukemic treatment. However, the studies of AML in general do not exclude a possible impact of differentiation for patient subsets and/or other types of antileukemic treatment.
Recently, Wojtuszkiewicz et al., found that there is maturation state-specific differential splicing of genes associated with cell cycle control and DNA damage in FLT3-ITD and NPM1 mutated AML blasts. Intriguingly, the number of genes that displayed differential splicing was significantly higher in the FAB M4 subtype, with a total of 1,438 splicing events, compared to the FAB M1 and M2 subtypes, each with about 200 splicing events [49].
The FAB-M4/M5 subset of AML patients is a heterogeneous group with regard to genetic abnormalities and includes mutations associated with both adverse and favorable prognosis for patients receiving intensive treatment based on conventional cytotoxic drugs [45,50,51,52,53]. The present study demonstrates that distinct patterns of protein expression and phosphorylation, as well as signaling pathways, are associated with each of the different FAB subtypes, specifically M1/M2 and M4/M5. In total, we found 911 proteins and 257 phosphosites that exhibited differential regulation when comparing the RELAPSE M1/M2 subtype to all the different subtypes of RELAPSE M4/M5. In RELAPSE patients with the myeloblastic subtype M1/M2, RNA-related processes like transcription and splicing were significantly increased. On the other hand, in RELAPSE patients with the more differentiated monocytic subtype M4/M5, there was a higher prevalence of signaling pathways involved in translation and degranulation. Additionally, the kinase-substrate analysis demonstrated enrichment of ERK1/2 and CSK2 kinases in the RELAPSE M1/M2 subgroup, whereas PKRA and PKRC kinases exhibited higher activity in the M4/M5 subgroups. In a study by Kornblau et al., reverse-phase protein array (RPPA) was used to distinguish between different AML FAB subtypes in 256 patients. They identified 24 proteins that showed differential expression among the 51 assayed proteins, effectively separating the myeloblastic subtype M1/M2 from the monocytic subtype M4/M5 [54]. Thus, the unique expression patterns of proteins and phosphoproteins among the different FAB subtypes, as identified in both current and previous studies [54,55], points out the importance of incorporating FAB classification into proteomic and phosphoproteomic studies. A recent study suggested that malignant hematological cells share biological characteristics with their normal counterparts [56]; this seems to be true also for monocytic AML cells (i.e., FAB-M4/M5 subclassification that show high levels of constitutive release of several cytokines/chemokines as well as other soluble mediators [10,56,57,58]. Normal macrophages seem to be reprogrammed by AML cells and thereby support leukemogenesis/chemosensitivity of the leukemic cells through their release of chemokines/cytokines [59]; when these mediators are released by the AML cells they may become a part of the intrinsic mechanisms for chemoresistance/disease progression caused by autocrine mechanisms and associated with differentiation in primary human AML cells.
We have previously reported that phosphoproteins and proteins linked to ribosome biogenesis and rRNA processing exhibit higher abundance in AML cells obtained from patients who experience relapse after intensive chemotherapy, compared to patient who achieve leukemia-free survival (> 5 years) [26]. In this study, we performed further stratification of this initial cohort by incorporating FAB classification and subsets with NPM1 mutation and normal cytogenetics as additional selection criteria. We found 850 proteins and 294 phosphosites that exhibited statistically significant differences between the relapse status, RELAPSE M4/M5 vs REL_FREE M4/M5 subgroups. In addition to conforming previous findings of term enrichment related to transcription and high CSK2 kinase activity, our current study also revealed a significant enrichment of proteins associated with mitochondrial translation and oxidative phosphorylation, particularly in the REL_M4/5_mut subgroup, as well as in all M4/M5 subgroups of patients who experienced relapse. The proteins found to be enriched include various mammalian mitochondrial ribosomal proteins (MRPL, MRPS), the mitochondrial intermediate peptidase (MIPEP) involved in the processing of oxidative phosphorylation-related proteins within the mitochondria [60], and NADH:ubiquinone oxidoreductase complex assembly factor 2 (NDUFAF2), a constituent of the NADH:ubiquinone oxidoreductase (complex I). Complex I is responsible for catalyzing the transfer of electrons from NADH to ubiquinone, which is the initial step in the mitochondrial respiratory chain [61]. Moreover, individual proteomic profiling showed that mitochondrial ribosomal proteins and ATP synthases could be used as relapse predictors in FAB M4/M5-classified patients with the NPM1 Ins mutations. However, these findings require further validation with external cohorts. Another intriguing observation in the present study is the significant enrichment of site-specific phosphorylation sites associated with glycolysis and autophagy of the mitochondrion and the involvement of PRKA/PRKC kinases in the REL_F_M4/5_mut subgroup, as compared to the REL_ M4/5_mut patients.
Patients with AML FAB-M4/M5 are heterogeneous with regard to their genetic abnormalities and include abnormalities with both favorable and adverse prognostic impact [51]. In our present study we observed that FAB-M4/M5 patients with later relapse had a proteomic profile that differed both from FAB-M4/M5 patients without relapse and from other relapse patients (i.e., FAB-M1/M2 patients). Despite their genetic heterogeneity, relapsed FAB-M4/M5 patients had in common proteomic differences with regard to mitochondrial function when compared with other FAB-M4/M5 patients. Even though previous studies have failed to demonstrate a prognostic impact of FAB-M4/M5 in AML patients receiving intensive and potentially curative cytotoxic therapy [44], our present study suggests that the molecular mechanisms behind relapse differ between patients and for certain subsets (at least partly) depend on the AML cell differentiation.
Mitochondria are important regulators of both cellular metabolism and survival; these two regulatory systems are characterized by a similar compartmentalization but also by molecular crosstalk/interactions, and the apoptotic machinery (including apoptosis regulator BCL2) is involved in the regulation of mitochondrial metabolism [62]. Our observation that the mitochondrial function/metabolism at the first time of diagnosis differs between AML-FAB-M4/M5 patients with and without later relapse is also consistent with other observations suggesting that mitochondrial function/energy metabolism is important for susceptibility to antileukemic therapy. First, monocytic differentiation reflected by the FAB classification is associated with resistance to venetoclax-based (i.e., a BCL2 inhibitor); and this resistance seems to be due to decreased functional importance of BCL2 and thereby altered regulation of apoptosis and mitochondrial energy metabolism in monocytic AML cells, including leukemic stem cells [30,31,63,64]. Second, differential expression of mitochondria-related genes is important for chemoresistance and seems to have an independent prognostic impact in AML [65]. Third, a subset of AML patients show mutations in genes that encode proteins in the electron transport complexes (Complex I/III/IV, ATP synthase), and mutations in the mitochondrial NADH dehydrogenase subunit 4 (a component of Complex I) seems to have a prognostic impact in adult AML [66,67]. Finally, monocytic differentiation is associated with response to BET (bromodomain and extraterminal domain protein family) inhibitors [68]. Taken together these studies show that monocytic AML cell differentiation together with mitochondrial functions are important for the responsiveness to various forms of antileukemic strategies, and this is also the reason why oxidative phosphorylation is regarded as a possible therapeutic target in cancer therapy [42]. Our present study suggests that the importance of altered mitochondrial function/metabolism for the development of AML relapse after intensive cytotoxic treatment differs between patients and is of particular importance in AML cells showing monocytic differentiation.
We were the first to demonstrate that chemoresistant relapsed AML cells have transitioned to a state characterized by higher expression of mitochondrial proteins, in adults [69]. In line with our findings, Stratman et al., recently published a comprehensive proteogenomic study reporting that the proteome at relapse is enriched with mitochondrial ribosomal proteins and subunits of the mitochondrial respiratory chain complex, not only in adults but also in children [24]. Interestingly, recent evidence suggests the existence of a proteomic subtype called Mito-AML, characterized by elevated expression of mitochondrial proteins and associated with a poor outcome. Moreover, Mito-AML cells exhibit a strong reliance on complex I-dependent mitochondrial respiration, which can be targeted by drugs like venetoclax [21]. Thus, both current and previous proteomics studies indicate that increased mitochondrial translational activity and oxidative phosphorylation are associated with poor prognosis, including higher relapse rates, and reduced overall survival.
5. Conclusions
This and recently published proteomics studies demonstrate that high mitochondrial protein abundance and respiration are associated with higher relapse rates and reduced overall survival [21,24,69]. The identification of distinct protein expression and phosphorylation profiles for each AML subtype, as observed in the present study for the FAB subtypes M1/M2 and M4/M5, holds promise for the discovery of subtype-specific biomarkers. These biomarkers could serve as predictors of prognosis and potential targets for personalized therapies. In particular, the high expression of mitochondrial ribosomal proteins and associated respiratory chain complexes emerges as a reliable predictor of a high relapse risk in AML patients with M4 and M5 subtypes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary File S1: Proteomics data REL_M1_M2_ and REL_M4_M5 subgroups; Supplementary File S2: Phosphoproteomics data REL_M1_M2_ and REL_M4_M5 subgroups; Supplementary File S3: Number of regulated phosphosites and proteins in the different comparison studies; Supplementary File S4: Proteomics data REL_F_M4_M5_ and REL_M4_M5 subgroups; Supplementary File S5: Phosphoproteomics data REL_F_M4_M5_ and REL_M4_M5 subgroups.
Author Contributions
F.S., E.A., Ø.B. and M.H.-V. contributed to experimental design and scientific discussion. F.S. and M.H.-V. performed the data analysis. F.S., Ø.B. and M.H.-V. wrote the article. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Kreftforeningen, the Norwegian Cancer Society (grant no. 100933) and by the Research Council of Norway INFRASTRUKTUR-program (project no. 295910).
Informed Consent Statement
Patient consent was awarded in our previous Cancers publication (DOI: 10.3390/cancers12030709).
Data Availability Statement
All raw data and MaxQuant output files from the original cohort can be found in the ProteomeXchange consortium with the dataset identifier PXD014997.
Acknowledgments
Mass spectrometry-based proteomic acquisition were performed at the Proteomics Unit at the University of Bergen (PROBE). PROBE is a member of the National Network of Advanced Proteomics Infrastructure (NAPI), which is funded by the Research Council of Norway.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dohner, H.; Wei, A.H.; Appelbaum, F.R.; Craddock, C.; DiNardo, C.D.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Godley, L.A.; Hasserjian, R.P.; et al. Diagnosis and management of AML in adults: 2022 recommendations from an international expert panel on behalf of the ELN. Blood 2022, 140, 1345–1377. [Google Scholar] [CrossRef] [PubMed]
- Khoury, J.D.; Solary, E.; Abla, O.; Akkari, Y.; Alaggio, R.; Apperley, J.F.; Bejar, R.; Berti, E.; Busque, L.; Chan, J.K.C.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Myeloid and Histiocytic/Dendritic Neoplasms. Leukemia 2022, 36, 1703–1719. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.M.; Catovsky, D.; Daniel, M.T.; Flandrin, G.; Galton, D.A.; Gralnick, H.R.; Sultan, C. Proposals for the classification of the acute leukaemias. French-American-British (FAB) co-operative group. Br. J. Haematol. 1976, 33, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Dohner, H.; Estey, E.H.; Amadori, S.; Appelbaum, F.R.; Buchner, T.; Burnett, A.K.; Dombret, H.; Fenaux, P.; Grimwade, D.; Larson, R.A.; et al. Diagnosis and management of acute myeloid leukemia in adults: Recommendations from an international expert panel, on behalf of the European LeukemiaNet. Blood 2010, 115, 453–474. [Google Scholar] [CrossRef] [PubMed]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef] [PubMed]
- Boissel, N.; Renneville, A.; Biggio, V.; Philippe, N.; Thomas, X.; Cayuela, J.M.; Terre, C.; Tigaud, I.; Castaigne, S.; Raffoux, E.; et al. Prevalence, clinical profile, and prognosis of NPM mutations in AML with normal karyotype. Blood 2005, 106, 3618–3620. [Google Scholar] [CrossRef] [PubMed]
- Rau, R.; Brown, P. Nucleophosmin (NPM1) mutations in adult and childhood acute myeloid leukaemia: Towards definition of a new leukaemia entity. Hematol. Oncol. 2009, 27, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Tsykunova, G.; Reikvam, H.; Hovland, R.; Bruserud, O. The surface molecule signature of primary human acute myeloid leukemia (AML) cells is highly associated with NPM1 mutation status. Leukemia 2012, 26, 557–559. [Google Scholar] [CrossRef]
- Reikvam, H.; Aasebo, E.; Brenner, A.K.; Bartaula-Brevik, S.; Gronningsaeter, I.S.; Forthun, R.B.; Hovland, R.; Bruserud, O. High Constitutive Cytokine Release by Primary Human Acute Myeloid Leukemia Cells Is Associated with a Specific Intercellular Communication Phenotype. J. Clin. Med. 2019, 8. [Google Scholar] [CrossRef]
- Brenner, A.K.; Tvedt, T.H.; Nepstad, I.; Rye, K.P.; Hagen, K.M.; Reikvam, H.; Bruserud, O. Patients with acute myeloid leukemia can be subclassified based on the constitutive cytokine release of the leukemic cells; the possible clinical relevance and the importance of cellular iron metabolism. Expert Opin. Ther. Targets 2017, 21, 357–369. [Google Scholar] [CrossRef]
- Bullinger, L.; Dohner, K.; Bair, E.; Frohling, S.; Schlenk, R.F.; Tibshirani, R.; Dohner, H.; Pollack, J.R. Use of gene-expression profiling to identify prognostic subclasses in adult acute myeloid leukemia. N. Engl. J. Med. 2004, 350, 1605–1616. [Google Scholar] [CrossRef]
- Alcalay, M.; Tiacci, E.; Bergomas, R.; Bigerna, B.; Venturini, E.; Minardi, S.P.; Meani, N.; Diverio, D.; Bernard, L.; Tizzoni, L.; et al. Acute myeloid leukemia bearing cytoplasmic nucleophosmin (NPMc+ AML) shows a distinct gene expression profile characterized by up-regulation of genes involved in stem-cell maintenance. Blood 2005, 106, 899–902. [Google Scholar] [CrossRef]
- Handschuh, L.; Kazmierczak, M.; Milewski, M.C.; Goralski, M.; Luczak, M.; Wojtaszewska, M.; Uszczynska-Ratajczak, B.; Lewandowski, K.; Komarnicki, M.; Figlerowicz, M. Gene expression profiling of acute myeloid leukemia samples from adult patients with AML-M1 and -M2 through boutique microarrays, real-time PCR and droplet digital PCR. Int. J. Oncol. 2018, 52, 656–678. [Google Scholar] [CrossRef]
- Luczak, M.; Kazmierczak, M.; Handschuh, L.; Lewandowski, K.; Komarnicki, M.; Figlerowicz, M. Comparative proteome analysis of acute myeloid leukemia with and without maturation. J. Proteomics 2012, 75, 5734–5748. [Google Scholar] [CrossRef]
- Casado, P.; Cutillas, P.R. Proteomic Characterization of Acute Myeloid Leukemia for Precision Medicine. Mol. Cell. Proteom. 2023, 22, 100517. [Google Scholar] [CrossRef] [PubMed]
- Casado, P.; Wilkes, E.H.; Miraki-Moud, F.; Hadi, M.M.; Rio-Machin, A.; Rajeeve, V.; Pike, R.; Iqbal, S.; Marfa, S.; Lea, N.; et al. Proteomic and genomic integration identifies kinase and differentiation determinants of kinase inhibitor sensitivity in leukemia cells. Leukemia 2018, 32, 1818–1822. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, M.; Smith, R.; Rajeeve, V.; Bessant, C.; Cutillas, P.R. Reconstructing kinase network topologies from phosphoproteomics data reveals cancer-associated rewiring. Nat. Biotechnol. 2020, 38, 493–502. [Google Scholar] [CrossRef]
- Hernandez-Valladares, M.; Wangen, R.; Aasebo, E.; Reikvam, H.; Berven, F.S.; Selheim, F.; Bruserud, O. Proteomic Studies of Primary Acute Myeloid Leukemia Cells Derived from Patients before and during Disease-Stabilizing Treatment Based on All-Trans Retinoic Acid and Valproic Acid. Cancers 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Aasebo, E.; Brenner, A.K.; Hernandez-Valladares, M.; Birkeland, E.; Berven, F.S.; Selheim, F.; Bruserud, O. Proteomic Comparison of Bone Marrow Derived Osteoblasts and Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Aasebo, E.; Brenner, A.K.; Birkeland, E.; Tvedt, T.H.A.; Selheim, F.; Berven, F.S.; Bruserud, O. The Constitutive Extracellular Protein Release by Acute Myeloid Leukemia Cells-A Proteomic Study of Patient Heterogeneity and Its Modulation by Mesenchymal Stromal Cells. Cancers 2021, 13. [Google Scholar] [CrossRef]
- Jayavelu, A.K.; Wolf, S.; Buettner, F.; Alexe, G.; Haupl, B.; Comoglio, F.; Schneider, C.; Doebele, C.; Fuhrmann, D.C.; Wagner, S.; et al. The proteogenomic subtypes of acute myeloid leukemia. Cancer Cell 2022, 40, 301–317.e312. [Google Scholar] [CrossRef]
- Caplan, M.; Wittorf, K.J.; Weber, K.K.; Swenson, S.A.; Gilbreath, T.J.; Willow Hynes-Smith, R.; Amador, C.; Hyde, R.K.; Buckley, S.M. Multi-omics reveals mitochondrial metabolism proteins susceptible for drug discovery in AML. Leukemia 2022, 36, 1296–1305. [Google Scholar] [CrossRef]
- Kramer, M.H.; Zhang, Q.; Sprung, R.; Day, R.B.; Erdmann-Gilmore, P.; Li, Y.; Xu, Z.; Helton, N.M.; George, D.R.; Mi, Y.; et al. Proteomic and phosphoproteomic landscapes of acute myeloid leukemia. Blood 2022, 140, 1533–1548. [Google Scholar] [CrossRef]
- Stratmann, S.; Vesterlund, M.; Umer, H.M.; Eshtad, S.; Skaftason, A.; Herlin, M.K.; Sundstrom, C.; Eriksson, A.; Hoglund, M.; Palle, J.; et al. Proteogenomic analysis of acute myeloid leukemia associates relapsed disease with reprogrammed energy metabolism both in adults and children. Leukemia 2023, 37, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Emdal, K.B.; Palacio-Escat, N.; Wigerup, C.; Eguchi, A.; Nilsson, H.; Bekker-Jensen, D.B.; Ronnstrand, L.; Kazi, J.U.; Puissant, A.; Itzykson, R.; et al. Phosphoproteomics of primary AML patient samples reveals rationale for AKT combination therapy and p53 context to overcome selinexor resistance. Cell Rep. 2022, 40, 111177. [Google Scholar] [CrossRef]
- Aasebo, E.; Berven, F.S.; Bartaula-Brevik, S.; Stokowy, T.; Hovland, R.; Vaudel, M.; Doskeland, S.O.; McCormack, E.; Batth, T.S.; Olsen, J.V.; et al. Proteome and Phosphoproteome Changes Associated with Prognosis in Acute Myeloid Leukemia. Cancers 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Aasebo, E.; Vaudel, M.; Mjaavatten, O.; Gausdal, G.; Van der Burgh, A.; Gjertsen, B.T.; Doskeland, S.O.; Bruserud, O.; Berven, F.S.; Selheim, F. Performance of super-SILAC based quantitative proteomics for comparison of different acute myeloid leukemia (AML) cell lines. Proteomics 2014, 14, 1971–1976. [Google Scholar] [CrossRef]
- Hernandez-Valladares, M.; Aasebo, E.; Selheim, F.; Berven, F.S.; Bruserud, O. Selecting Sample Preparation Workflows for Mass Spectrometry-Based Proteomic and Phosphoproteomic Analysis of Patient Samples with Acute Myeloid Leukemia. Proteomes 2016, 4. [Google Scholar] [CrossRef]
- Vardiman, J.W.; Thiele, J.; Arber, D.A.; Brunning, R.D.; Borowitz, M.J.; Porwit, A.; Harris, N.L.; Le Beau, M.M.; Hellstrom-Lindberg, E.; Tefferi, A.; et al. The 2008 revision of the World Health Organization (WHO) classification of myeloid neoplasms and acute leukemia: Rationale and important changes. Blood 2009, 114, 937–951. [Google Scholar] [CrossRef]
- Kuusanmaki, H.; Leppa, A.M.; Polonen, P.; Kontro, M.; Dufva, O.; Deb, D.; Yadav, B.; Bruck, O.; Kumar, A.; Everaus, H.; et al. Phenotype-based drug screening reveals association between venetoclax response and differentiation stage in acute myeloid leukemia. Haematologica 2020, 105, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Pei, S.; Pollyea, D.A.; Gustafson, A.; Stevens, B.M.; Minhajuddin, M.; Fu, R.; Riemondy, K.A.; Gillen, A.E.; Sheridan, R.M.; Kim, J.; et al. Monocytic Subclones Confer Resistance to Venetoclax-Based Therapy in Patients with Acute Myeloid Leukemia. Cancer Discov. 2020, 10, 536–551. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Nepusz, T.; Yu, H.; Paccanaro, A. Detecting overlapping protein complexes in protein-protein interaction networks. Nat. Methods 2012, 9, 471–472. [Google Scholar] [CrossRef]
- Schneider, T.D.; Stephens, R.M. Sequence logos: A new way to display consensus sequences. Nucleic Acids Res. 1990, 18, 6097–6100. [Google Scholar] [CrossRef]
- Hulsen, T.; de Vlieg, J.; Alkema, W. BioVenn—A web application for the comparison and visualization of biological lists using area-proportional Venn diagrams. BMC Genom. 2008, 9, 488. [Google Scholar] [CrossRef] [PubMed]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Ashton, T.M.; McKenna, W.G.; Kunz-Schughart, L.A.; Higgins, G.S. Oxidative Phosphorylation as an Emerging Target in Cancer Therapy. Clin. Cancer Res. 2018, 24, 2482–2490. [Google Scholar] [CrossRef]
- Papa, S.; Martino, P.L.; Capitanio, G.; Gaballo, A.; De Rasmo, D.; Signorile, A.; Petruzzella, V. The oxidative phosphorylation system in mammalian mitochondria. Adv. Exp. Med. Biol. 2012, 942, 3–37. [Google Scholar] [CrossRef]
- Walter, R.B.; Othus, M.; Burnett, A.K.; Lowenberg, B.; Kantarjian, H.M.; Ossenkoppele, G.J.; Hills, R.K.; van Montfort, K.G.; Ravandi, F.; Evans, A.; et al. Significance of FAB subclassification of “acute myeloid leukemia, NOS” in the 2008 WHO classification: Analysis of 5848 newly diagnosed patients. Blood 2013, 121, 2424–2431. [Google Scholar] [CrossRef]
- Canaani, J.; Beohou, E.; Labopin, M.; Socie, G.; Huynh, A.; Volin, L.; Cornelissen, J.; Milpied, N.; Gedde-Dahl, T.; Deconinck, E.; et al. Impact of FAB classification on predicting outcome in acute myeloid leukemia, not otherwise specified, patients undergoing allogeneic stem cell transplantation in CR1: An analysis of 1690 patients from the acute leukemia working party of EBMT. Am. J. Hematol. 2017, 92, 344–350. [Google Scholar] [CrossRef]
- Bostrom, B.; Brunning, R.D.; McGlave, P.; Ramsay, N.; Nesbit, M., Jr.; Woods, W.G.; Hurd, D.; Krivit, W.; Kim, T.; Goldman, A.; et al. Bone marrow transplantation for acute nonlymphocytic leukemia in first remission: Analysis of prognostic factors. Blood 1985, 65, 1191–1196. [Google Scholar] [CrossRef]
- Saultz, J.N.; Tyner, J.W. Chasing leukemia differentiation through induction therapy, relapse and transplantation. Blood Rev. 2023, 57, 101000. [Google Scholar] [CrossRef]
- Norgaard, J.M.; Olesen, L.H.; Olesen, G.; Meyer, K.; Kristensen, J.S.; Bendix, K.; Pedersen, B.; Kjeldsen, E.; Hokland, P. FAB M4 and high CD14 surface expression is associated with high cellular resistance to Ara-C and daunorubicin: Implications for clinical outcome in acute myeloid leukaemia. Eur. J. Haematol. 2001, 67, 221–229. [Google Scholar] [CrossRef]
- Wojtuszkiewicz, A.; van der Werf, I.; Hutter, S.; Walter, W.; Baer, C.; Kern, W.; Janssen, J.; Ossenkoppele, G.J.; Haferlach, C.; Cloos, J.; et al. Maturation State-Specific Alternative Splicing in FLT3-ITD and NPM1 Mutated AML. Cancers 2021, 13. [Google Scholar] [CrossRef]
- Alfayez, M.; Issa, G.C.; Patel, K.P.; Wang, F.; Wang, X.; Short, N.J.; Cortes, J.E.; Kadia, T.; Ravandi, F.; Pierce, S.; et al. The Clinical impact of PTPN11 mutations in adults with acute myeloid leukemia. Leukemia 2021, 35, 691–700. [Google Scholar] [CrossRef]
- Cheng, Z.; Hu, K.; Tian, L.; Dai, Y.; Pang, Y.; Cui, W.; Zhao, H.; Qin, T.; Han, Y.; Hu, N.; et al. Clinical and biological implications of mutational spectrum in acute myeloid leukemia of FAB subtypes M4 and M5. Cancer Gene Ther. 2018, 25, 77–83. [Google Scholar] [CrossRef]
- Miyajima, T.; Onozawa, M.; Yoshida, S.; Miyashita, N.; Kimura, H.; Takahashi, S.; Yokoyama, S.; Matsukawa, T.; Goto, H.; Sugita, J.; et al. Clinical implications of NUP98::NSD1 fusion at diagnosis in adult FLT3-ITD positive AML. Eur. J. Haematol. 2023, 111, 620–627. [Google Scholar] [CrossRef]
- Sano, H.; Shimada, A.; Taki, T.; Murata, C.; Park, M.J.; Sotomatsu, M.; Tabuchi, K.; Tawa, A.; Kobayashi, R.; Horibe, K.; et al. RAS mutations are frequent in FAB type M4 and M5 of acute myeloid leukemia, and related to late relapse: A study of the Japanese Childhood AML Cooperative Study Group. Int. J. Hematol. 2012, 95, 509–515. [Google Scholar] [CrossRef]
- Kornblau, S.M.; Tibes, R.; Qiu, Y.H.; Chen, W.; Kantarjian, H.M.; Andreeff, M.; Coombes, K.R.; Mills, G.B. Functional proteomic profiling of AML predicts response and survival. Blood 2009, 113, 154–164. [Google Scholar] [CrossRef]
- Aasebo, E.; Forthun, R.B.; Berven, F.; Selheim, F.; Hernandez-Valladares, M. Global Cell Proteome Profiling, Phospho-signaling and Quantitative Proteomics for Identification of New Biomarkers in Acute Myeloid Leukemia Patients. Curr. Pharm. Biotechnol. 2016, 17, 52–70. [Google Scholar] [CrossRef]
- Majumder, M.M.; Leppa, A.M.; Hellesoy, M.; Dowling, P.; Malyutina, A.; Kopperud, R.; Bazou, D.; Andersson, E.; Parsons, A.; Tang, J.; et al. Multi-parametric single cell evaluation defines distinct drug responses in healthy hematologic cells that are retained in corresponding malignant cell types. Haematologica 2020, 105, 1527–1538. [Google Scholar] [CrossRef]
- Attenhofer, C.; Vuilliomenet, A.; Richter, M.; Kaufmann, U.; Metzger, U.; Bertel, O. [Heart contusions: Pathological findings and clinical course]. Schweiz. Med. Wochenschr. 1992, 122, 1593–1599. [Google Scholar]
- Rundgren, I.M.; Ryningen, A.; Anderson Tvedt, T.H.; Bruserud, O.; Ersvaer, E. Immunomodulatory Drugs Alter the Metabolism and the Extracellular Release of Soluble Mediators by Normal Monocytes. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Miari, K.E.; Guzman, M.L.; Wheadon, H.; Williams, M.T.S. Macrophages in Acute Myeloid Leukaemia: Significant Players in Therapy Resistance and Patient Outcomes. Front. Cell. Dev. Biol. 2021, 9, 692800. [Google Scholar] [CrossRef]
- Chew, A.; Buck, E.A.; Peretz, S.; Sirugo, G.; Rinaldo, P.; Isaya, G. Cloning, expression, and chromosomal assignment of the human mitochondrial intermediate peptidase gene (MIPEP). Genomics 1997, 40, 493–496. [Google Scholar] [CrossRef]
- Ogilvie, I.; Kennaway, N.G.; Shoubridge, E.A. A molecular chaperone for mitochondrial complex I assembly is mutated in a progressive encephalopathy. J. Clin. Investig. 2005, 115, 2784–2792. [Google Scholar] [CrossRef]
- Basak, N.P.; Banerjee, S. Mitochondrial dependency in progression of acute myeloid leukemia. Mitochondrion 2015, 21, 41–48. [Google Scholar] [CrossRef]
- Cherry, E.M.; Abbott, D.; Amaya, M.; McMahon, C.; Schwartz, M.; Rosser, J.; Sato, A.; Schowinsky, J.; Inguva, A.; Minhajuddin, M.; et al. Venetoclax and azacitidine compared with induction chemotherapy for newly diagnosed patients with acute myeloid leukemia. Blood Adv. 2021, 5, 5565–5573. [Google Scholar] [CrossRef]
- Pollyea, D.A.; Stevens, B.M.; Jones, C.L.; Winters, A.; Pei, S.; Minhajuddin, M.; D’Alessandro, A.; Culp-Hill, R.; Riemondy, K.A.; Gillen, A.E.; et al. Venetoclax with azacitidine disrupts energy metabolism and targets leukemia stem cells in patients with acute myeloid leukemia. Nat. Med. 2018, 24, 1859–1866. [Google Scholar] [CrossRef]
- Tong, X.; Zhou, F. Integrated bioinformatic analysis of mitochondrial metabolism-related genes in acute myeloid leukemia. Front. Immunol. 2023, 14, 1120670. [Google Scholar] [CrossRef]
- Wu, S.; Akhtari, M.; Alachkar, H. Characterization of Mutations in the Mitochondrial Encoded Electron Transport Chain Complexes in Acute Myeloid Leukemia. Sci. Rep. 2018, 8, 13301. [Google Scholar] [CrossRef]
- Damm, F.; Bunke, T.; Thol, F.; Markus, B.; Wagner, K.; Gohring, G.; Schlegelberger, B.; Heil, G.; Reuter, C.W.; Pullmann, K.; et al. Prognostic implications and molecular associations of NADH dehydrogenase subunit 4 (ND4) mutations in acute myeloid leukemia. Leukemia 2012, 26, 289–295. [Google Scholar] [CrossRef]
- Romine, K.A.; Nechiporuk, T.; Bottomly, D.; Jeng, S.; McWeeney, S.K.; Kaempf, A.; Corces, M.R.; Majeti, R.; Tyner, J.W. Monocytic differentiation and AHR signaling as Primary Nodes of BET Inhibitor Response in Acute Myeloid Leukemia. Blood Cancer Discov. 2021, 2, 518–531. [Google Scholar] [CrossRef]
- Aasebo, E.; Berven, F.S.; Hovland, R.; Doskeland, S.O.; Bruserud, O.; Selheim, F.; Hernandez-Valladares, M. The Progression of Acute Myeloid Leukemia from First Diagnosis to Chemoresistant Relapse: A Comparison of Proteomic and Phosphoproteomic Profiles. Cancers 2020, 12. [Google Scholar] [CrossRef]
Figure 1.
The AML cell proteome shows increased abundance of rRNA metabolism and modification proteins for RELAPSE patients with FAB classification M1 and M2. (a) Venn diagram of regulated proteins (ANOVA, post hoc Turkey’s HSD with FDR < 0.05) obtained from REL_M1/2_all vs REL_M4/5_all ( ), REL_M1/2_all vs REL_M4/5_mut (
), REL_M1/2_all vs REL_M4/5_mut ( ) and REL_M1/2_all vs REL_M4/5_CN (
) and REL_M1/2_all vs REL_M4/5_CN ( ) comparisons. (b) Reactome pathways and (c) protein-protein interaction (PPI) network analyses of comparison-specific (61 and 25) and comparison-overlapping (78 and 22) regulated proteins.
) comparisons. (b) Reactome pathways and (c) protein-protein interaction (PPI) network analyses of comparison-specific (61 and 25) and comparison-overlapping (78 and 22) regulated proteins.
), REL_M1/2_all vs REL_M4/5_mut () and REL_M1/2_all vs REL_M4/5_CN () comparisons. (b) Reactome pathways and (c) protein-protein interaction (PPI) network analyses of comparison-specific (61 and 25) and comparison-overlapping (78 and 22) regulated proteins.
Figure 1.
The AML cell proteome shows increased abundance of rRNA metabolism and modification proteins for RELAPSE patients with FAB classification M1 and M2. (a) Venn diagram of regulated proteins (ANOVA, post hoc Turkey’s HSD with FDR < 0.05) obtained from REL_M1/2_all vs REL_M4/5_all (), REL_M1/2_all vs REL_M4/5_mut () and REL_M1/2_all vs REL_M4/5_CN () comparisons. (b) Reactome pathways and (c) protein-protein interaction (PPI) network analyses of comparison-specific (61 and 25) and comparison-overlapping (78 and 22) regulated proteins.
), REL_M1/2_all vs REL_M4/5_mut () and REL_M1/2_all vs REL_M4/5_CN () comparisons. (b) Reactome pathways and (c) protein-protein interaction (PPI) network analyses of comparison-specific (61 and 25) and comparison-overlapping (78 and 22) regulated proteins.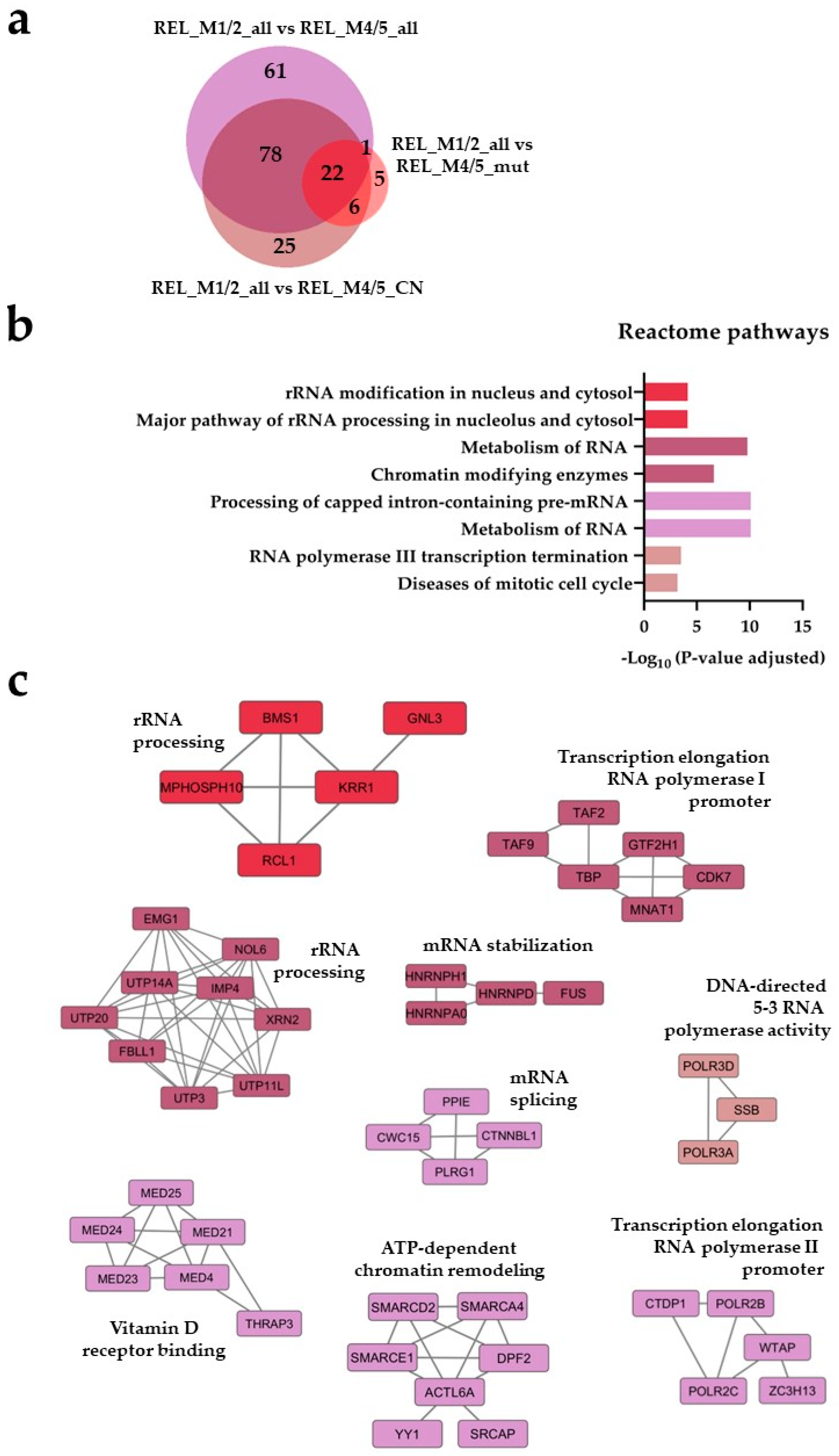
Figure 2.
The AML cell proteome shows increased translation, neutrophil degranulation and intracellular protein/vesicle transport proteins for RELAPSE patients with FAB classification M4 and M5. (a) Venn diagram of regulated proteins obtained from REL_M4/5_all vs REL_M1/2_all ( ), REL_M4/5_mut vs REL_M1/2_all (
), REL_M4/5_mut vs REL_M1/2_all ( ) and REL_M4/5_CN vs REL_M1/2_all (
) and REL_M4/5_CN vs REL_M1/2_all ( ) comparisons. (b) Reactome pathways and (c) PPI network analyses of comparison-specific (30, 37 and 68) and comparison-overlapping (70, 166 and 78) regulated proteins. * It stands for Reactome pathways with unadjusted P-value < 0.05.
) comparisons. (b) Reactome pathways and (c) PPI network analyses of comparison-specific (30, 37 and 68) and comparison-overlapping (70, 166 and 78) regulated proteins. * It stands for Reactome pathways with unadjusted P-value < 0.05.
), REL_M4/5_mut vs REL_M1/2_all () and REL_M4/5_CN vs REL_M1/2_all () comparisons. (b) Reactome pathways and (c) PPI network analyses of comparison-specific (30, 37 and 68) and comparison-overlapping (70, 166 and 78) regulated proteins. * It stands for Reactome pathways with unadjusted P-value < 0.05.
Figure 2.
The AML cell proteome shows increased translation, neutrophil degranulation and intracellular protein/vesicle transport proteins for RELAPSE patients with FAB classification M4 and M5. (a) Venn diagram of regulated proteins obtained from REL_M4/5_all vs REL_M1/2_all (), REL_M4/5_mut vs REL_M1/2_all () and REL_M4/5_CN vs REL_M1/2_all () comparisons. (b) Reactome pathways and (c) PPI network analyses of comparison-specific (30, 37 and 68) and comparison-overlapping (70, 166 and 78) regulated proteins. * It stands for Reactome pathways with unadjusted P-value < 0.05.
), REL_M4/5_mut vs REL_M1/2_all () and REL_M4/5_CN vs REL_M1/2_all () comparisons. (b) Reactome pathways and (c) PPI network analyses of comparison-specific (30, 37 and 68) and comparison-overlapping (70, 166 and 78) regulated proteins. * It stands for Reactome pathways with unadjusted P-value < 0.05.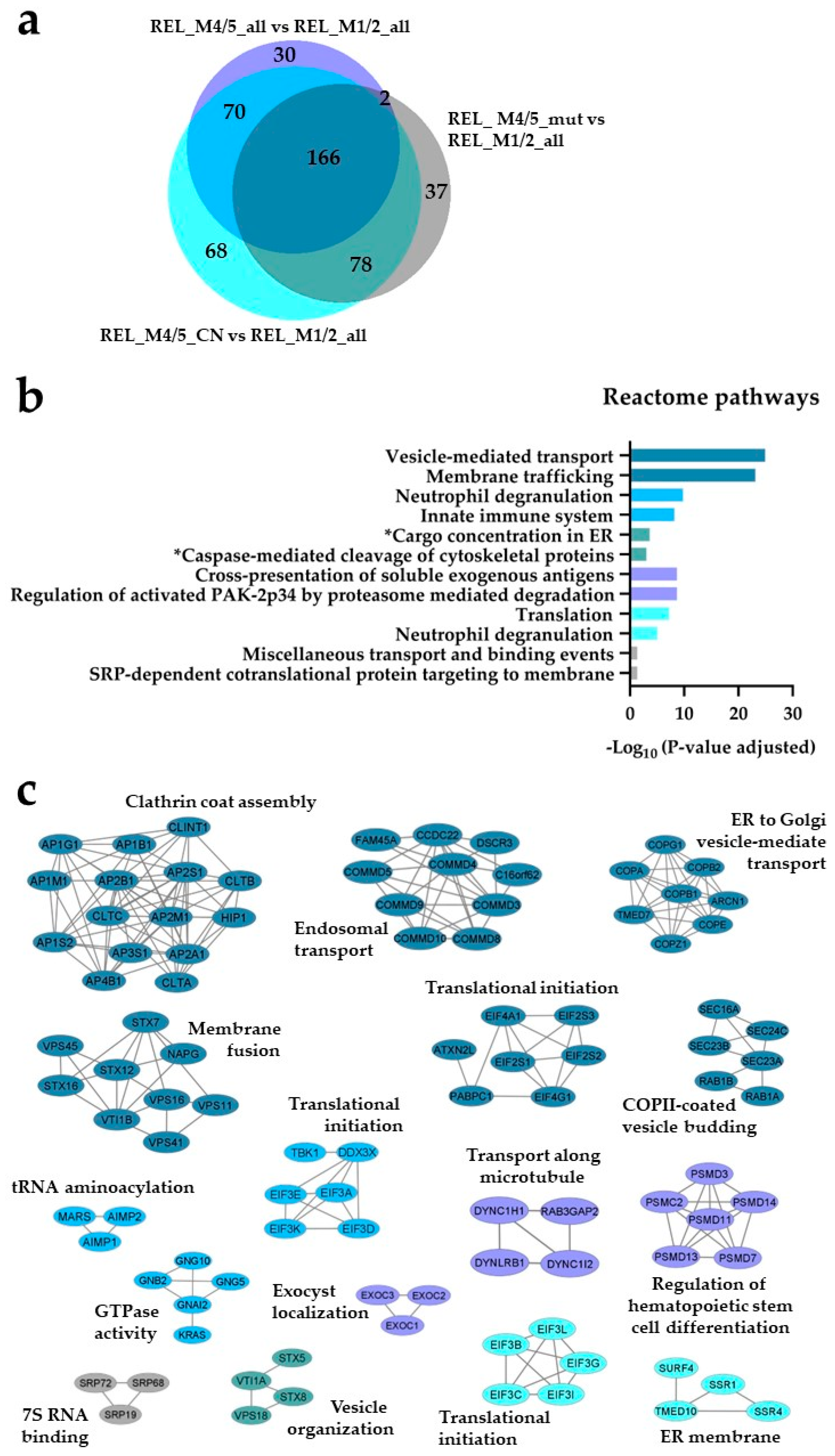
Figure 3.
The AML cell phosphoproteome shows increased phosphorylation of RNA and DNA binding proteins for RELAPSE patients with FAB classification M1 and M2. (a) Venn diagram of differentially regulated phosphorylation sites (ANOVA, post hoc Turkey’s HSD with FDR < 0.05) obtained from REL_M1/2_all vs REL_M4/5_all ( ), REL_M1/2_all vs REL_M4/5_mut (
), REL_M1/2_all vs REL_M4/5_mut ( ) and REL_M1/2_all vs REL_M4/5_CN (
) and REL_M1/2_all vs REL_M4/5_CN ( ) comparisons. (b) Gene ontology (GO) with molecular function terms and KEGG pathway analyses of comparison-specific (16) and comparison-overlapping (11 and 13) differentially phosphorylated proteins. (c) Sequence motif analysis of the ±5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (16) and comparison-overlapping (11 and 13) datasets.
) comparisons. (b) Gene ontology (GO) with molecular function terms and KEGG pathway analyses of comparison-specific (16) and comparison-overlapping (11 and 13) differentially phosphorylated proteins. (c) Sequence motif analysis of the ±5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (16) and comparison-overlapping (11 and 13) datasets.
), REL_M1/2_all vs REL_M4/5_mut () and REL_M1/2_all vs REL_M4/5_CN () comparisons. (b) Gene ontology (GO) with molecular function terms and KEGG pathway analyses of comparison-specific (16) and comparison-overlapping (11 and 13) differentially phosphorylated proteins. (c) Sequence motif analysis of the ±5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (16) and comparison-overlapping (11 and 13) datasets.
Figure 3.
The AML cell phosphoproteome shows increased phosphorylation of RNA and DNA binding proteins for RELAPSE patients with FAB classification M1 and M2. (a) Venn diagram of differentially regulated phosphorylation sites (ANOVA, post hoc Turkey’s HSD with FDR < 0.05) obtained from REL_M1/2_all vs REL_M4/5_all (), REL_M1/2_all vs REL_M4/5_mut () and REL_M1/2_all vs REL_M4/5_CN () comparisons. (b) Gene ontology (GO) with molecular function terms and KEGG pathway analyses of comparison-specific (16) and comparison-overlapping (11 and 13) differentially phosphorylated proteins. (c) Sequence motif analysis of the ±5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (16) and comparison-overlapping (11 and 13) datasets.
), REL_M1/2_all vs REL_M4/5_mut () and REL_M1/2_all vs REL_M4/5_CN () comparisons. (b) Gene ontology (GO) with molecular function terms and KEGG pathway analyses of comparison-specific (16) and comparison-overlapping (11 and 13) differentially phosphorylated proteins. (c) Sequence motif analysis of the ±5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (16) and comparison-overlapping (11 and 13) datasets.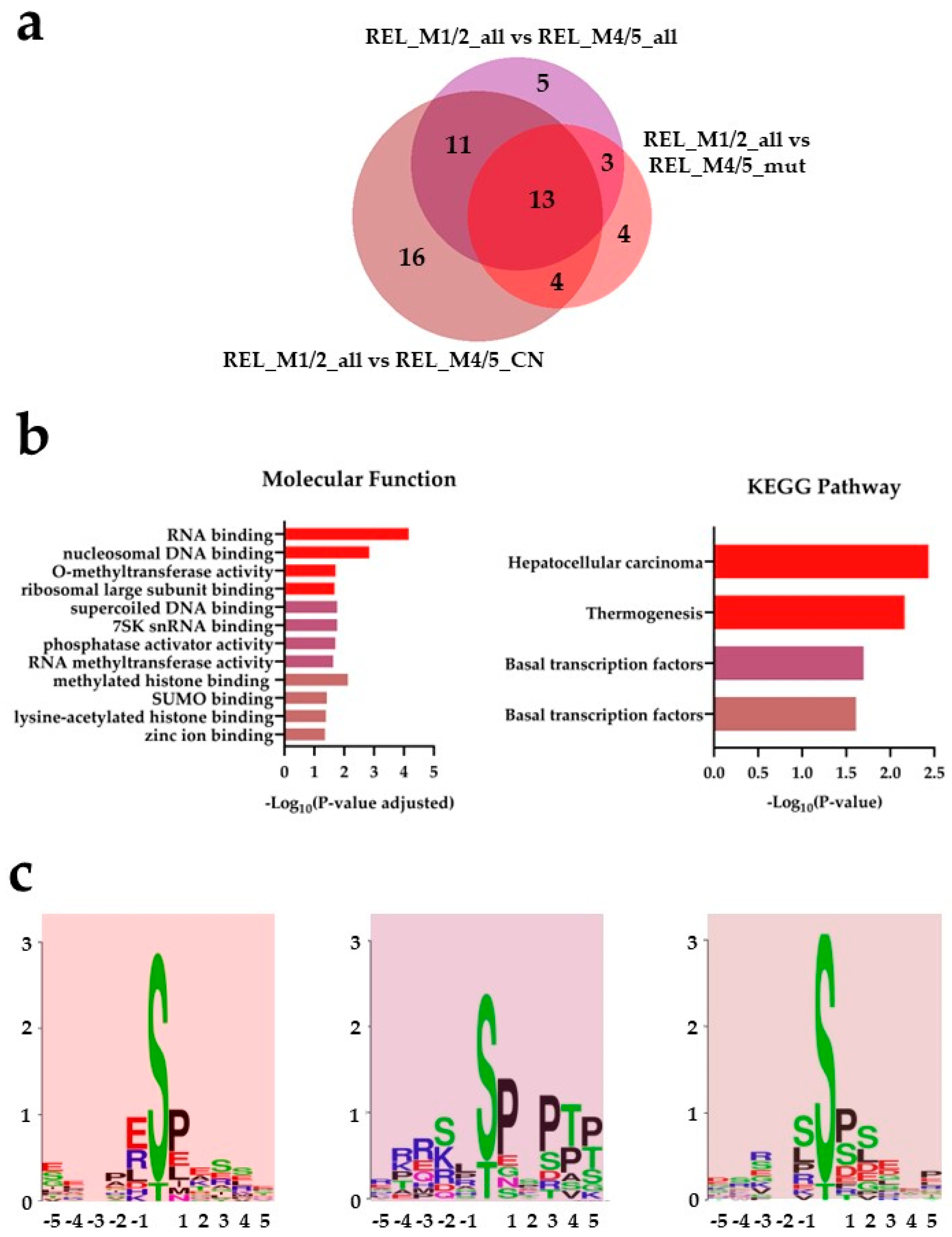
Figure 4.
The AML cell phosphoproteome shows increased phosphorylation of RNA binding and transcriptional proteins for RELAPSE patients with FAB classification M4 and M5. (a) Venn diagram of differentially regulated phosphorylation sites obtained from REL_M4/5_all vs REL_M1/2_all ( ), REL_M4/5_mut vs REL_M1/2_all (
), REL_M4/5_mut vs REL_M1/2_all ( ) and REL_M4/5_CN vs REL_M1/2_all (
) and REL_M4/5_CN vs REL_M1/2_all ( ) comparisons. (b) GO with molecular function terms and KEGG pathway analyses of comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) differentially phosphorylated proteins. (c) PPI network analyses of overlapping (42 and 24) differentially phosphorylated proteins. (d) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) datasets. * A shorter name for “Regulation of lipolysis in adipocytes” KEGG pathway is added for space purposes.
) comparisons. (b) GO with molecular function terms and KEGG pathway analyses of comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) differentially phosphorylated proteins. (c) PPI network analyses of overlapping (42 and 24) differentially phosphorylated proteins. (d) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) datasets. * A shorter name for “Regulation of lipolysis in adipocytes” KEGG pathway is added for space purposes.
), REL_M4/5_mut vs REL_M1/2_all () and REL_M4/5_CN vs REL_M1/2_all () comparisons. (b) GO with molecular function terms and KEGG pathway analyses of comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) differentially phosphorylated proteins. (c) PPI network analyses of overlapping (42 and 24) differentially phosphorylated proteins. (d) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) datasets. * A shorter name for “Regulation of lipolysis in adipocytes” KEGG pathway is added for space purposes.
Figure 4.
The AML cell phosphoproteome shows increased phosphorylation of RNA binding and transcriptional proteins for RELAPSE patients with FAB classification M4 and M5. (a) Venn diagram of differentially regulated phosphorylation sites obtained from REL_M4/5_all vs REL_M1/2_all (), REL_M4/5_mut vs REL_M1/2_all () and REL_M4/5_CN vs REL_M1/2_all () comparisons. (b) GO with molecular function terms and KEGG pathway analyses of comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) differentially phosphorylated proteins. (c) PPI network analyses of overlapping (42 and 24) differentially phosphorylated proteins. (d) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) datasets. * A shorter name for “Regulation of lipolysis in adipocytes” KEGG pathway is added for space purposes.
), REL_M4/5_mut vs REL_M1/2_all () and REL_M4/5_CN vs REL_M1/2_all () comparisons. (b) GO with molecular function terms and KEGG pathway analyses of comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) differentially phosphorylated proteins. (c) PPI network analyses of overlapping (42 and 24) differentially phosphorylated proteins. (d) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites from the comparison-specific (22, 12 and 27) and comparison-overlapping (42 and 24) datasets. * A shorter name for “Regulation of lipolysis in adipocytes” KEGG pathway is added for space purposes.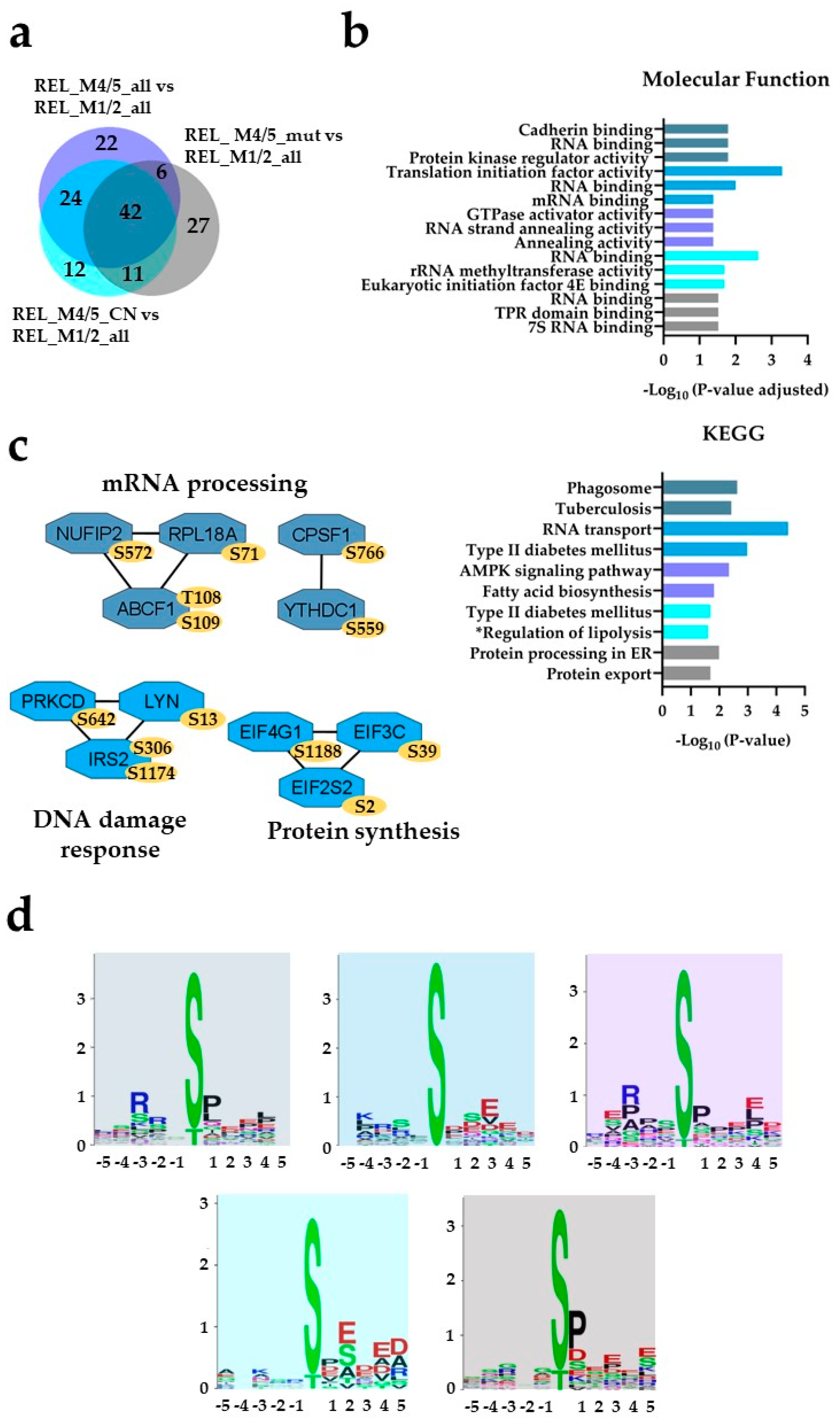
Figure 5.
The AML cell proteome shows increased abundance of mitochondrial translational proteins for RELAPSE patients when compared to REL_FREE with FAB classification M4 and M5. (a) Venn diagram of regulated proteins obtained from REL_M4/5_all vs REL_F_M4/5_all ( ), REL_M4/5_mut vs REL_F_M4/5_mut (
), REL_M4/5_mut vs REL_F_M4/5_mut ( ) and REL_M4/5_CN vs REL_F_M4/5_CN (
) and REL_M4/5_CN vs REL_F_M4/5_CN ( ) comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (119) and comparison-overlapping (15, 7 and 4) regulated proteins.
) comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (119) and comparison-overlapping (15, 7 and 4) regulated proteins.
), REL_M4/5_mut vs REL_F_M4/5_mut () and REL_M4/5_CN vs REL_F_M4/5_CN () comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (119) and comparison-overlapping (15, 7 and 4) regulated proteins.
Figure 5.
The AML cell proteome shows increased abundance of mitochondrial translational proteins for RELAPSE patients when compared to REL_FREE with FAB classification M4 and M5. (a) Venn diagram of regulated proteins obtained from REL_M4/5_all vs REL_F_M4/5_all (), REL_M4/5_mut vs REL_F_M4/5_mut () and REL_M4/5_CN vs REL_F_M4/5_CN () comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (119) and comparison-overlapping (15, 7 and 4) regulated proteins.
), REL_M4/5_mut vs REL_F_M4/5_mut () and REL_M4/5_CN vs REL_F_M4/5_CN () comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (119) and comparison-overlapping (15, 7 and 4) regulated proteins.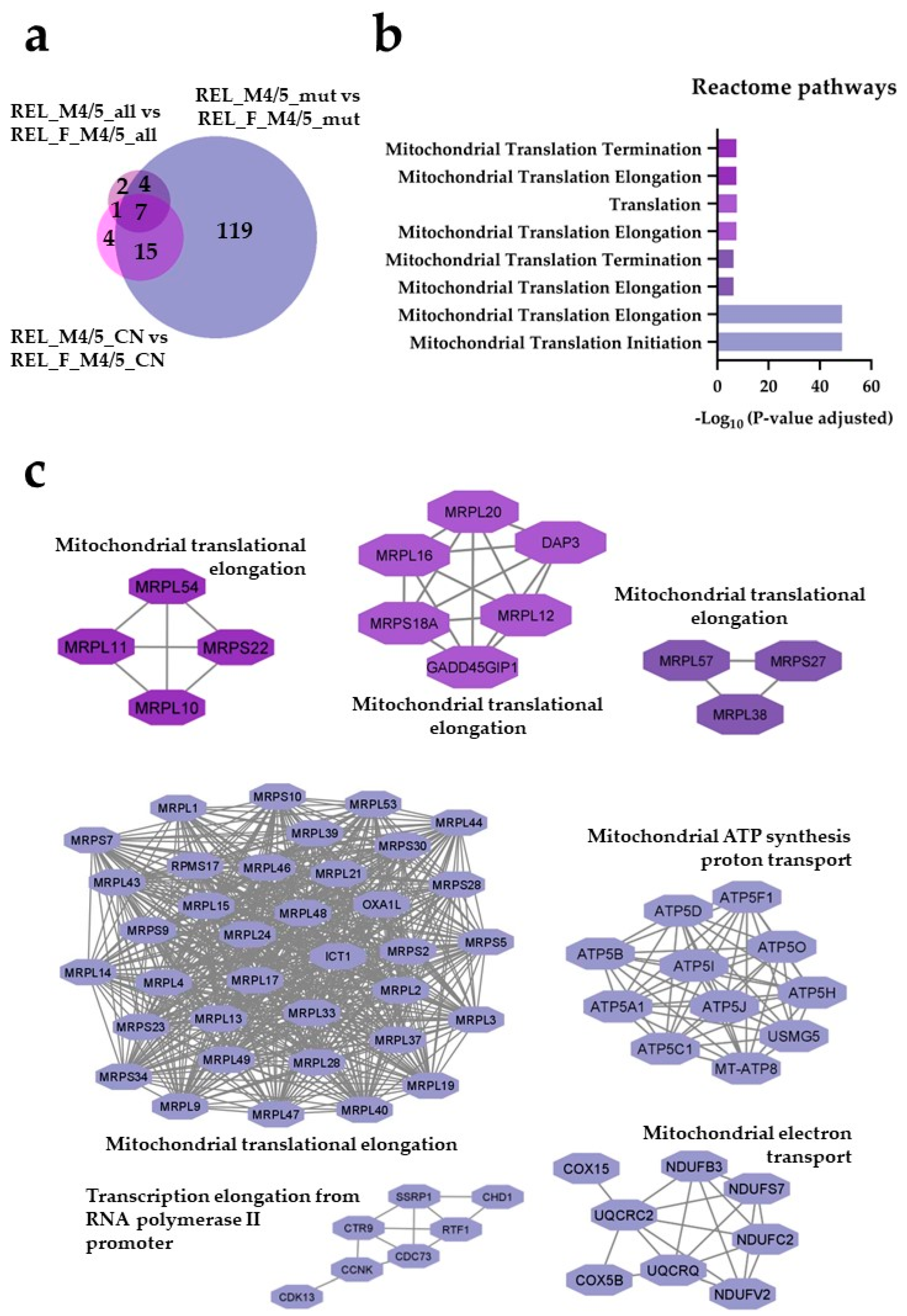
Figure 6.
The AML cell proteome shows increased of proteins involved in the metabolism of nucleotides for REL_FREE patients when compared to RELAPSE with FAB classification M4 and M5. (a) Venn diagram of regulated proteins obtained from REL_F_M4/5_mut vs REL_M4/5_mut ( ) and REL_F_M4/5_CN vs REL_M4/5_CN (
) and REL_F_M4/5_CN vs REL_M4/5_CN ( ) comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (38 and 11) and comparison-overlapping (10) regulated proteins. * It stands for Reactome pathways with unadjusted P-value <0.05.
) comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (38 and 11) and comparison-overlapping (10) regulated proteins. * It stands for Reactome pathways with unadjusted P-value <0.05.
) and REL_F_M4/5_CN vs REL_M4/5_CN () comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (38 and 11) and comparison-overlapping (10) regulated proteins. * It stands for Reactome pathways with unadjusted P-value <0.05.
Figure 6.
The AML cell proteome shows increased of proteins involved in the metabolism of nucleotides for REL_FREE patients when compared to RELAPSE with FAB classification M4 and M5. (a) Venn diagram of regulated proteins obtained from REL_F_M4/5_mut vs REL_M4/5_mut () and REL_F_M4/5_CN vs REL_M4/5_CN () comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (38 and 11) and comparison-overlapping (10) regulated proteins. * It stands for Reactome pathways with unadjusted P-value <0.05.
) and REL_F_M4/5_CN vs REL_M4/5_CN () comparisons. (b) Reactome pathway and (c) PPI network analyses of comparison-specific (38 and 11) and comparison-overlapping (10) regulated proteins. * It stands for Reactome pathways with unadjusted P-value <0.05.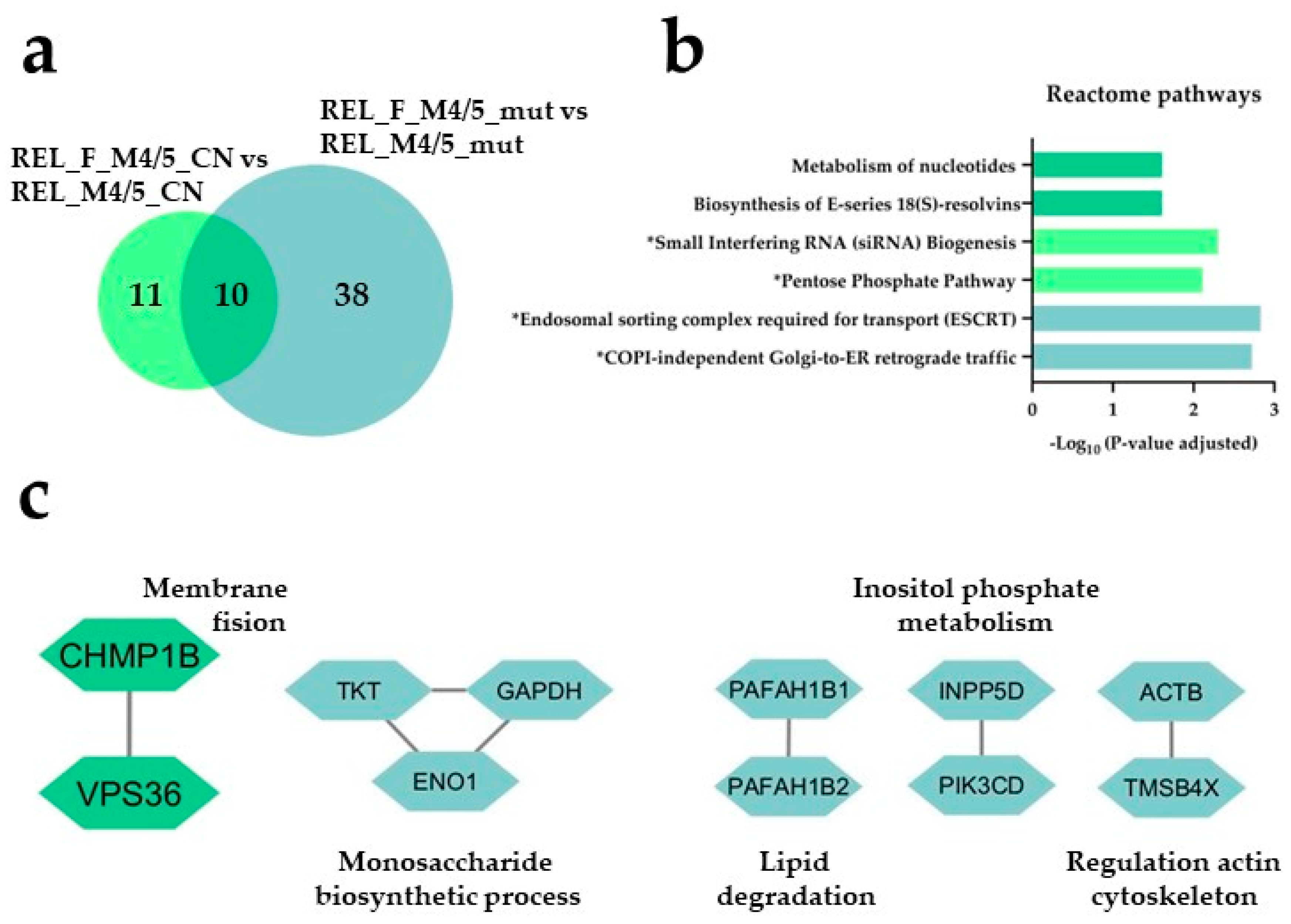
Figure 7.
The AML cell phosphoproteome shows increased phosphorylation of RNA and DNA binding proteins for RELAPSE patients with FAB classification M4 and M5 and with the NPM1 Ins mutation. (a) Venn diagram of differentially regulated phosphorylation sites obtained from REL_M4/5_all vs REL_F_M4/5_all ( ), REL_M4/5_mut vs REL_F_M4/5_mut (
), REL_M4/5_mut vs REL_F_M4/5_mut ( ) and REL_M4/5_CN vs REL_F_M4/5_CN (
) and REL_M4/5_CN vs REL_F_M4/5_CN ( ) comparisons. (b) GO with biological process (BP), molecular function (MF) and cellular compartment (CC) terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. * It stands for KEGG and Reactome pathways with unadjusted P-value < 0.05.
) comparisons. (b) GO with biological process (BP), molecular function (MF) and cellular compartment (CC) terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. * It stands for KEGG and Reactome pathways with unadjusted P-value < 0.05.
), REL_M4/5_mut vs REL_F_M4/5_mut () and REL_M4/5_CN vs REL_F_M4/5_CN () comparisons. (b) GO with biological process (BP), molecular function (MF) and cellular compartment (CC) terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. * It stands for KEGG and Reactome pathways with unadjusted P-value < 0.05.
Figure 7.
The AML cell phosphoproteome shows increased phosphorylation of RNA and DNA binding proteins for RELAPSE patients with FAB classification M4 and M5 and with the NPM1 Ins mutation. (a) Venn diagram of differentially regulated phosphorylation sites obtained from REL_M4/5_all vs REL_F_M4/5_all (), REL_M4/5_mut vs REL_F_M4/5_mut () and REL_M4/5_CN vs REL_F_M4/5_CN () comparisons. (b) GO with biological process (BP), molecular function (MF) and cellular compartment (CC) terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. * It stands for KEGG and Reactome pathways with unadjusted P-value < 0.05.
), REL_M4/5_mut vs REL_F_M4/5_mut () and REL_M4/5_CN vs REL_F_M4/5_CN () comparisons. (b) GO with biological process (BP), molecular function (MF) and cellular compartment (CC) terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_M4/5_mut vs REL_F_M4/5_mut comparison. * It stands for KEGG and Reactome pathways with unadjusted P-value < 0.05.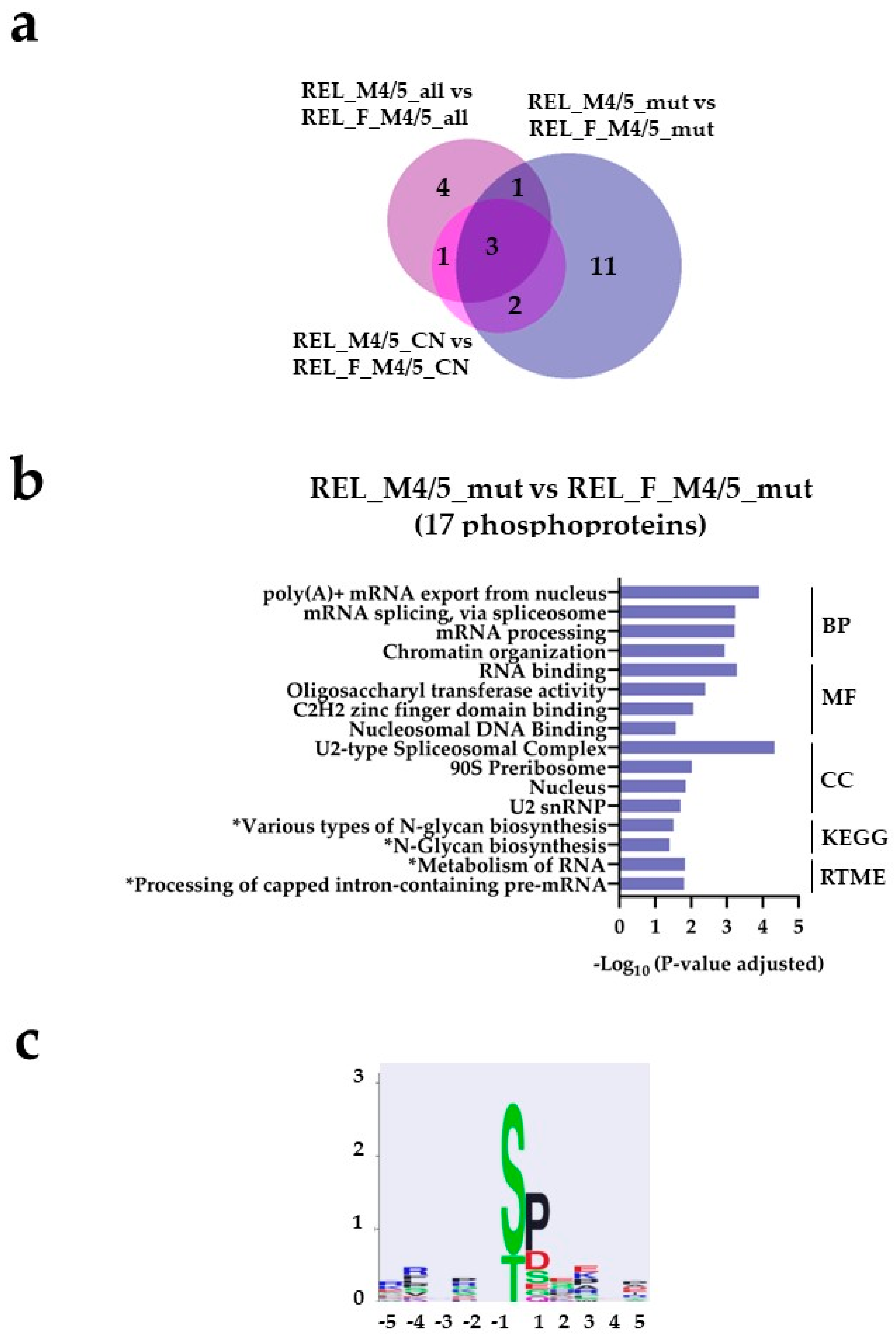
Figure 8.
The AML cell phosphoproteome shows increased phosphorylation of glycolysis and gluconeogenesis proteins for RELAPSE_FREE patients with FAB classification M4 and M5 and with the NPM1 Ins mutation. (a) Venn diagram of differentially regulated phosphorylation sites obtained from REL_F_M4/5_all vs REL_M4/5_all ( ), REL_F_M4/5_mut vs REL_M4/5_mut (
), REL_F_M4/5_mut vs REL_M4/5_mut ( ) and REL_F_M4/5_CN vs REL_M4/5_CN (
) and REL_F_M4/5_CN vs REL_M4/5_CN ( ) comparisons. (b) GO with BP, MF and CC terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_F_M4/5_mut vs REL_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_F_M4/5_mut subgroup when compared to REL_M4/5_mut patients.
) comparisons. (b) GO with BP, MF and CC terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_F_M4/5_mut vs REL_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_F_M4/5_mut subgroup when compared to REL_M4/5_mut patients.
), REL_F_M4/5_mut vs REL_M4/5_mut () and REL_F_M4/5_CN vs REL_M4/5_CN () comparisons. (b) GO with BP, MF and CC terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_F_M4/5_mut vs REL_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_F_M4/5_mut subgroup when compared to REL_M4/5_mut patients.
Figure 8.
The AML cell phosphoproteome shows increased phosphorylation of glycolysis and gluconeogenesis proteins for RELAPSE_FREE patients with FAB classification M4 and M5 and with the NPM1 Ins mutation. (a) Venn diagram of differentially regulated phosphorylation sites obtained from REL_F_M4/5_all vs REL_M4/5_all (), REL_F_M4/5_mut vs REL_M4/5_mut () and REL_F_M4/5_CN vs REL_M4/5_CN () comparisons. (b) GO with BP, MF and CC terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_F_M4/5_mut vs REL_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_F_M4/5_mut subgroup when compared to REL_M4/5_mut patients.
), REL_F_M4/5_mut vs REL_M4/5_mut () and REL_F_M4/5_CN vs REL_M4/5_CN () comparisons. (b) GO with BP, MF and CC terms, KEGG and Reactome pathway analyses of 17 differentially higher phosphorylated proteins in the REL_F_M4/5_mut vs REL_M4/5_mut comparison. (c) Sequence motif analysis of the ± 5 amino acids flanking the differentially regulated phosphorylation sites in the REL_F_M4/5_mut subgroup when compared to REL_M4/5_mut patients.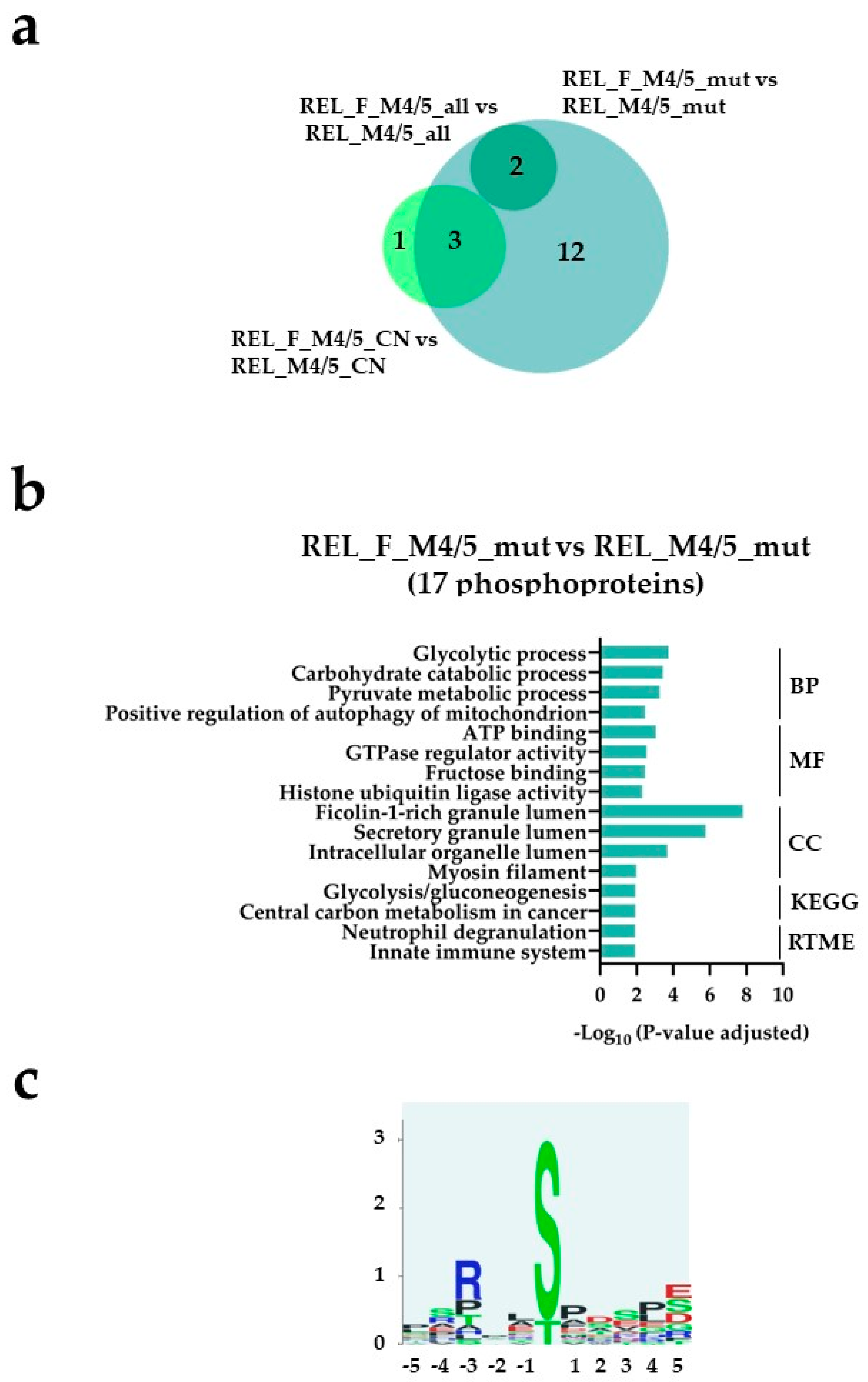

Table 1.
Characteristics of acute myeloid leukemia (AML) disease subgroups.
| Characteristic | RELAPSE | REL_FREE | |
|---|---|---|---|
| FAB classification | M1/M2 | 8 | 1 |
| M4/M5 | 12 | 14 | |
| NPM1 | WT | 6 | 6 |
| Ins | 5 | 8 | |
| CN | 46, XY or XX | 7 | 9 |
FAB: French-American-British; NPM1: nucleophosmin 1; CN: cytogenetically normal; WT: wild type; Ins a 4 bp-insertion.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



