
Submitted:
24 October 2023
Posted:
26 October 2023
You are already at the latest version
Abstract
Next-generation cancer and oncology research needs to take full advantage of the multi-modal structured, or graph, information, with the graph datatypes ranging from molecular structures to spatially resolved imaging and digital pathology to biological networks to knowledge graphs. Graph Neural Networks (GNNs) efficiently combine the graph structure representations with the high predictive performance of deep learning, especially on the large multi-modal datasets. In this review article, we survey the landscape of recent (2020-present) GNN applications in the context of cancer and oncology research, and delineate six currently predominant research areas. Subsequently, we identify the most promising directions for future research. We compare GNNs with graphical models and "non-structured" deep learning, and devise the guidelines for cancer and oncology researchers or physician-scientists asking the question of whether they should adopt the GNN methodology in their research pipelines.
Keywords:
Graph Neural Network
; GNN
; Deep Learning
; Cancer
; Oncology
; Graphical Model
; Bayesian Network
1. SRC (Sexual Reproductive Cycle)
Sexual reproduction is a major way of eukaryotic organism proliferation in the current world. SRC is a SRC indispensable for sexually reproducing organisms (eukaryotes). During sexual reproduction, two gametes meet and fuse as a zygote, which is the first cell and starting point of diploid sporophyte generation (having 2n chromosomes in a cell). When mature, a cell of diploid sporophyte undergoes meiosis, giving rise to four haploid spores (the first cells and starting points of haploid gametophyte generation), each of which has n chromosomes in a cell and produces gametes when mature. When two different gametes (egg and sperm) from gametophytes meet (fertilization), they fuse as a zygote. This completes a sexual reproductive cycle, SRC[1].
In the initial stage of sexual reproduction, both gametes are equal in morphology, and both haploid and diploid individuals are unicellular. A unicellular diploid undergoes meiosis, giving rise to four haploid cells (gametes). When two unicellular gametes meet, they fuse as a single cell (zygote), and thus restart an entire SRC. This is the most primitive mode of SRC, as shown in Figure 1.
2. Serial Modifications Added to SRC
The above described SRC mode is the simplest one. Although simple, it places the foundation for later evolution of plants (and animals). During later evolution, plants add various modifications on this foundation and thus give rise to various plant groups bearing different characteristic organs.
In a favorable condition, an infinite number of the above simple SRC can be connected head-and-tail and thus forms the so-called phylogeny. The negative effects inflicting such a simple SRC mode include that unicellularity restricts the organisms from complications and diversifications. The actual biological history is that organisms did add various modifications to this simple SRC foundation and thus enabled the diversification of organisms.
The first important modification added to SRC is differentiation of gametes, namely, the former isomorphic gametes in Figure 1 differentiate into two categories, as shown in Figure 2. The differences between these two categories of gametes may be in various aspects, e.g. size, form, chemical features, or gene composition. Such differences make two types of gametes distinguishable, which correspond closely to male and female gametes, namely, smaller and more mobile gametes are male, while bigger and more immobile gametes are female. Needless to say, fertilization is restricted between two different gametes. Relative to isomorphic gametes, this differentiation allocates more resources to female gametes, which play a more involving role in the development of offspring. This modification is significant in the history of organisms as it initiates the history of sex and gender in common sense.
The second important modification added to SRC is multicellularization, which may have occurred multiple times in the history. Compared to the frequently seen complicated morphs of life, the unique feature of the organisms in Figure 1 is their unicellularity and simplicity. To derive more complicated morphs of life, mutlicellularity is a must. To carry out this process, a serial unicellular-colonial-multicellular transformations in cell-to-cell adhesion, communication, coordination, and specialization[2], and probably a multinucleate unicellular progenitor are required[3]. Multicellularization requires formerly free, independent, simple individual cells to be aggregated into a single organism that has all of its cells integrated, behave and function collectively and coordinately (Figure 3) [2]. Apparently, the overhead of such aggregating is that the behaviors of the cells must be coordinated for optimal function. Extracellular matrix (ECM, = cell wall) is a medium that units these cells. ECM may be taken as a gel mixture of water, polysaccharides, and fibrous proteins, which coordinates cell growth to maintain the morphology of organisms and buffers the effects of local stressors. The macromolecules secreted by adjacent cells are scaffolded into polymers that lend the matrix a viscous consistency. The matrix prevents cellular structures from collapsing and keeps the cells in a tissue separate and physically distinct, although these cells actually remain more or less connected with its adjacent peers through plasmodesmata.
It is inconceivable that organisms always live in favorable environments. In case of unfavorable environment or stress, organisms suspend their physiological activity and generate cysts, in which spores are saved, to preserve their germs for later life. These cysts are cases enclosing spores, homologous to sporangia in spore plants. The occurrence of cysts signifies the third important modification added to SRC, differentiation between somatic and reproductive parts in organisms, namely, parts of organisms are assigned different functions, becoming somatic parts and reproductive parts, respectively (Figure 4). The differentiation between these parts can be traced at least back to the Ediacara (600 Ma ago)[4,5].
Differentiation of vegetative parts A noticeable feature of earliest vascular plants is that almost all bifurcated branches are fertile, namely, almost every branch bears a sporangium on its terminal. Ensuing evolution introduces differentiation among these initially equal branches, and some of these branches became sterilized and lost their reproductive functions. These sterilized branches underwent various metamorphosis and gave rise to megaleaves. Although this process appears in a direction opposite to what observed in the individual development, it is a historical fact. The functions of these vegetative parts are not restricted to photosynthesis, instead they are recruited to play various auxiliary roles in forming reproductive organs (as seen below).
The differentiation of reproductive parts makes the occurrence of sporangia in the earliest land plants rather expected. In the Rhynie Chert Flora, Cooksonia plants comprise only branches and sporangia. Cooksonia and its allies are representatives of land plant pioneers[6,7]. Sporangia in these plants are the precursors of sporangia, sori, pollen sacs, anthers, megasporangia, metamegasporangia (= ovules), and the core part in primary, secondary, and various tertiary metamegasporangium complexes. The following is a brief characterization of these novelties in plants.
Sporangia are almost universally isomorphic in sporophytes of early land plants. Just as cysts mentioned above, sporangia protect and ensure the spores within to survive harsh times and revive later, through nutrition supplied through tapetum. The isomorphy is not restricted to general morphology, spores in the sporangia are universally trilete and hard to distinguish each other. When mature, spores escape through a sporangium slit. Those spores that perch in favorable niches germinate and give rise to gametophytes. A gametophyte of early spore plants usually is simple, bearing antheridia/archegonia on its terminals (Figure 5). When a gametophyte matures, sperms from the antheridia swim through a film of water, fuse with and fertilize the eggs in archegonia. The fusion of a sperm and an egg is termed as fertilization, a process that produces a zygote. A zygote is the starting point of a new sporophyte, which bears terminal sporangia when mature. This completes the SRC of early spore plants (Figure 5).
Figure 5.
A simple spore plant and its SRC. Note that both sporophyte and gametophyte are similar in morphology. Compared to the SRC primitive mode in Figure 1, a spore is connected to a gamete by a curve (instead of a straight line as in Figure 1), as multicellularity is inserted in between.
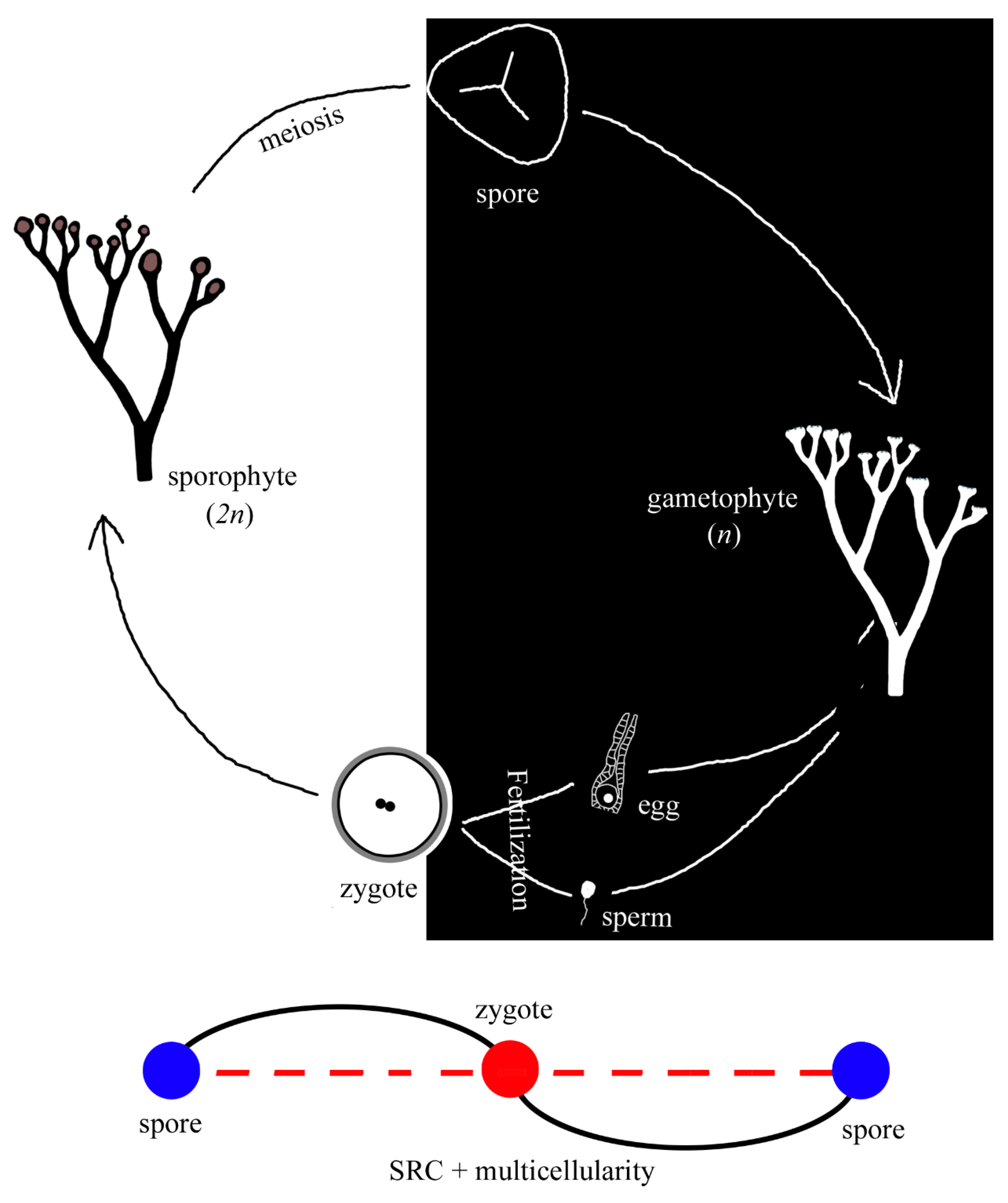
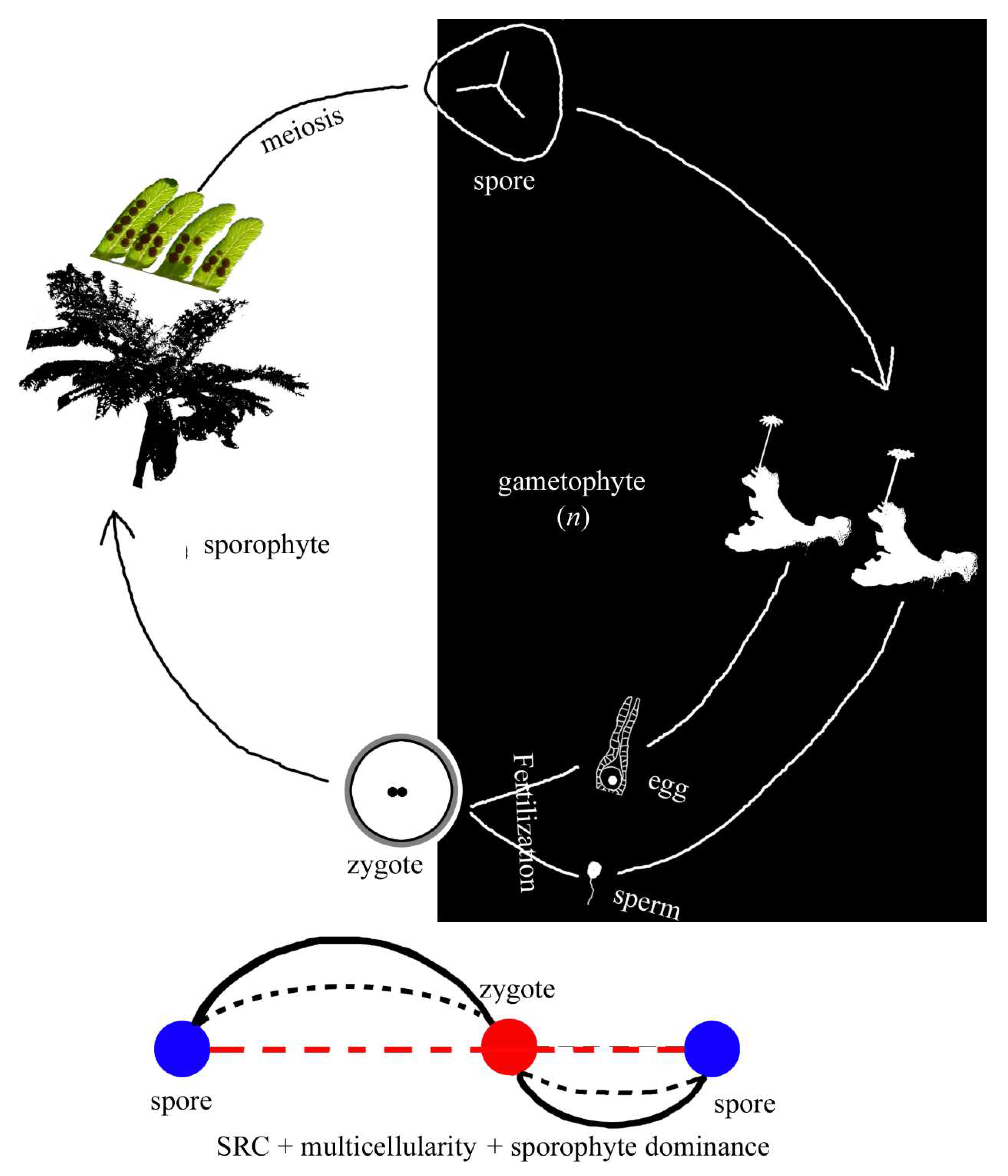
Figure 6.
A fern and its SRC. Note that the sporophyte (generation) is dominant, and the gametophyte (generation) is reduced. Compared to the SRC mode in Figure 2, the dominance of sporophyte (generation) is added to the SRC.
Figure 6.
A fern and its SRC. Note that the sporophyte (generation) is dominant, and the gametophyte (generation) is reduced. Compared to the SRC mode in Figure 2, the dominance of sporophyte (generation) is added to the SRC.
The spores in these pioneering land plants are termed homospores, against those later derived spores of two distinct morphologies, which are termed heterospores. The occurrence of heterospores in plants signifies the occurrence of heterospory. Thenceafter, the differentiation of former identical spores into spores of two distinct morphologies (especially sizes) is completed, correspondingly, former sporangia differentiate into microsporangia and megasporangia (Figure 7). The megaspores in megasporangia are fewer in number and each allocated more resources as they are destined to play a more involving role and take more responsibility in the development of offspring. In contrast, microspores in microsporangia are greater in number and each allocated less nutrition as the responsibility for them in reproduction is restricted to passing genes in 1n chromosomes to ovule to form a diploid zygote.
Although each megaspore in megasporangia is allocated more resources, the amount of resources allocated to each megaspore is limited by the dimensions of megaspore itself, as megaspores leave their parent plants, live independently, and are cut from further nutrition supply afterward. Such a limit apparently sets up an upper limit for the vitality of the megaspore and its offspring, endangering the continuity of a phylogeny. Plants lift such a resource limit by evolving a character termed endospory, which means that mature megaspores are retained in sporangia on their mother plants and mature megaspores remain nutritionally bonded with their mother plant. Endospory extends the time length of the nutritional bond between mother plant and megaspores, providing extra physical protection for megaspores and making the developmental environment of female gametophytes more congenial and controllable until seed maturation. This novel nutritional relationship turns a former megaspore into a metamegaspore (= an ovule), which is the developmental precursor of a seed (Figure 8), and thus gives rise to a new feature of plants, seed, and initiates a new phylogeny of plants, seed plants. The occurrence of ovule/seed may be the single most significant event in the evolution of plants, as it led to the great diversity of seed plants (including more than 300,000 species in angiosperms) in the current earth ecosystem. Besides the above mentioned extended nutritional bond, another significant concomitant change is that sporophyte generation becomes more dominant in SRC and of longer time in a SRC, and correspondingly gametophyte generation is more reduced, parasitic on the sporophytes, and of shorter time in a SRC.
An ovule is not strictly homologous or equivalent to a megaspore. Instead, an ovule is homologous or equivalent to a megasporangium and its adjacent accessories (integument or integuments), which are sterilized sporangia and play a protective role for the nucellus (a megasporangium now centrally positioned) [8,9]. Previously independently living gametophytes (in spore plants) are now highly reduced and develop within the nucellus, which continuously derives its nutrition through the nutritional bond from the parent plant (sporophyte) and thus supports the parasitic gametophyte. In this way, a gametophyte, including egg-bearing archegonia, develops and matures within the nucellus. Various pollination mechanisms transfer pollen (= microspores) to the termini of nucellus, where pollen germinate and complete male gametophyte development and finally produce sperms that swim to egg cell in the archegonia within nucellus to fulfill fertilization process. The fusion between a sperm and an egg results in a zygote. This process (fertilization) gives a start to a new sporophyte (zygote) of seed plants. This completes a SRC of a primitive seed plant, such as Ginkgo and other early seed plants.
Mature ovules (seeds) are propagules of seed plants. Ovules/seeds in Ginkgo and early seed plants are all borne on termini of branches and exposed (naked) to the exterior environment, which may be harsh and harmful for the development of ovules/seeds sometimes. Buffering ovules/seeds against potential harms is apparently conducive to their success in survival struggle. It is not surprising that some seed plants, for example, cycads, develop some strategy to shelter their ovules/seeds (especially in the early developmental period). For example, seeds are aggregated on certain branches (frequently termed as sporophylls) of various forms that in turn are aggregated into strobili, which shelter seeds from harms. In the meantime, seeds in some other seed plants are covered and protected by structures including cupules (Gnetopsis, Caytoniales, Petriellales, Peltaspermales), interseminal scales (Bennettitales), seed-scales (Coniferales) etc[10]. Compared to those in Ginkgo, seeds in these groups are better and more protected (Figure 9). We term these structures primary metamegasporangium complexes.
Angiosperms are the single most diversified group of plants in the current world. The key innovation distinguishing angiosperms from their peer gymnosperms is their fuller ovule-protection (Figure 10). Compared to the above gymnosperms with partial ovule/seed protection/enclosure (Figure 9), ovule enclosure in angiosperms is fulfilled before pollination (with rare exceptions). This feature, angio-ovuly (metamegasporangia enclosed by accessories) is frequently taken as a feature idiosyncratic to angiosperms. This feature ensures a congenial and favorable developmental environment for the ovules, giving edges to angiosperms in their competition against gymnospermous peers, including pre-zygotic selection of pollen grains, self-incompatibility, outcrossing-enhancing, etc[11,12,13,14,15]. The success in reproduction of these novel mechanisms is exemplified by the great diversity of angiosperms (over 300,000 species) in the current world, which enables the origin and survival of the humans.
In many textbooks, angiosperms seem to culminate the evolution of plants, and angio-ovuly seems to be the ultimate step of plant evolution, as if plants would not evolve thence after. However, this misleading impression apparently does not reflect the truth of plant evolution. A sound theoretical extrapolation is that, beyond angio-ovuly, fruits of angiosperms may be further enclosed and protected by adjacent accessories (Figure 11). This is not a plain hypothesis: the occurrences of hypanthodium (Figure 11a-c)[16] and inflated calyx syndrome (ICS, frequently seen in Solanaceae)[17] implement extra protection for the fruits in some angiosperms. Hypanthodium uses an expanded axis tip to protect the fruits, namely, it is derived through expansion and invagination of the flower axis tip. These processes not only have been seen in the basalmost angiosperm Amborella[18], but also may account for the inferior ovary in a Jurassic flower Nanjinganthus[19,20,21]. Such a combination of expansion and invagination of the flower axis tip is a phenomenon so far restricted to angiosperms. As a consequence of such a morphological innovation, a mutually beneficial relationship is established between Moraceae and agaonid wasps (Figure 11a): Moracea invite and trap otherwise free agaonid wasps in the figs for successful pollination, fruit-protecting, and increased seed setting, while agaonid wasps develop various features adapting to their living in host plants[22]. This may be termed tertiary metamegasporangium complex I (= angiocarpy, enclosed fruit). Tertiary metamegasporangium complex I has been seen not only in other extant angiosperms (Siparuna, Figure 11b), but also in an Early Cretaceous fossil angiosperm, Chaoyangia liangii (Figure 11c)[5].
Other than angio-ovuly and angiocarpy, many angiosperms aggregate multiple flowers into inflorescences of various geometries (Figure 12). Such an aggregation makes flowers more conspicuous and attractive especially during anthesis. Flowers in a single inflorescence differentiate spatially and temporally in morphology and development (especially anthesis timing) according to their positions in the inflorescence, and thus are conducive to successful pollination and fruitful reproduction. Thus inflorescence may be termed tertiary metamegasporangium complex II, as it integrates more flowers and accessories into a function unit.
As stated above, one of the major advantages of endomegasporangium is the extension of nutritional bond between mother plants and offspring. This extension seems to culminate in angiosperms that have enclosed ovules/seeds. However, this nutritional bond can be further extended in vivipary. Vivipary in plants designates that seeds germinate in ovaries/fruits. Apparently, not only ovules and seeds obtain their nutrition from their diploid parent plants, but their derivatives (seedlings) also do so, giving the offspring extra advantages through temporally extended nutritional bond between parent and offspring, namely, mother plants not only supply nutrition to ovules and seeds, but also to newer sporophytes (Figure 13)[16]. Such a nutritional bond spanning three generations (including sporophyte, gametophyte, and next generation sporophyte) culminates in Rhizophoraceae[16], and apparently benefits the survival of their offspring in the unique habitat. This may be termed tertiary metamegasporangium complex III.
These three types of tertiary metamegasporangium complexes are additional modifications added to SRC of equivalent level, namely, after secondary metamegasporangium complex, therefore they all are termed tertiary metamegasporangium complexes and distinguished by numbers.
3. The Underlying Trends
The above stairs of plant reproduction evolution are morphological reflections of plant evolution, which follows several underlying trends, including ODC, D&A, internalization, E&I, increasing dominance of sporophyte.
ODC (Offspring Development Conditioning) designates that, following the evolution path of a specific plant phylogeny, plants gain increasing control over the developmental environment of their offspring. ODC is implemented either as a strengthened and extended nutritional bond, enhanced physical protection, or their combinations in various lineages of plants[23].
The organization of plant reproductive organs becomes increasingly complicated and canalized during the evolution. A logical and natural consequence of such evolution is D&A (differentiation & aggregations), during which former equivalent parts are differentiated into the core part and the peripheral protective accessory parts, and both of which become indispensable parts of a more complicated organ. This is well demonstrated in the deriving of ovules and flowers.
Unavoidably in this process, the core reproductive part is shifted from a previously outer and more exposed position into an inner and more sheltered position. Such a morphological change can be termed as internalization. It is intriguing that internalization not only accounts for the evolution of plant reproductive organs, but also accounts for the evolution of the reproductive organs in vertebrates as well as the humans[23].
A previously largely ignored morphological change idiosyncratic of angiosperms is E&I (Expansion & Invagination) of flower axis. This trend has been demonstrated in Amborella [18], and it may account for the derivation of inferior ovary in the currently earliest fossil flower, Nanjinganthus [19,20,21] (Figure 14) and hypanthodia in some extant angiosperms (Figure 11a)[16,25,26,27].
Among vascular plants, a general evolution trend is increasing dominance and longer time of sporophyte generation in a SRC. Gametophyte plays an increasingly reduced and subdued role in a SRC, and gametophyte generation occupies an increasingly shorter time in SRC. There is little information on the evolution of gametophyte-dominating plants (bryophytes) yet.
A phylogeny of plants may be taken as a link composed of serial connected SRC of different individuals. As a link tends to break at its weakest point and the weakest point in a SRC is its initial vulnerable and weak period, naturally, whoever protects and nurtures its offspring better wins in evolution. This is the logic underlying ODC and other trends demonstrated in plant reproduction evolution. Decent applying of these rules contributes to the success of various land plants and accounts for their diversity in the current world.
Acknowledgments
I appreciate Dr. Shunong Bai at the Peking University and Dr. Mengxiang Sun at the Wuhan University for their careful reading and constructive suggestions. This research was supported by the National Natural Science Foundation of China (42288201, 41688103, 91514302) and the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (Grant No. XDB26000000).
References
- Bai, S.N. The concept of the sexual reproduction cycle and its evolutionary significance. Frontiers in Plant Science 2015, 6, 11. [Google Scholar] [CrossRef]
- Niklas, K.J. The evolutionary-developmental origins of multicellularity. American Journal of Botany 2014, 101, 6–25. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J.; Cobb, E.D.; Crawford, D.R. The evo-dev of multinucleate cells, issues, and organisms, and an alternative route to multicelluarity. Evolution and Development 2013, 15, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xiao, S.; Pang, K.; Zhou, C.; Yuan, X. Cell differentiation and germ-soma separation in Ediacaran animal embryo-like fossils. Nature 2014, 516, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. The Dawn Angiosperms, 2nd ed.; Springer: Cham, Switzerland, 2018; p. 407. [Google Scholar]
- Edwards, D.; Morris, J.L.; Richardson, J.B.; Kenrick, P. Cryptospores and cryptophytes reveal hidden diversity in early land floras. New Phytologist 2014, 202, 50–78. [Google Scholar] [CrossRef] [PubMed]
- Gonez, P.; Gerrienne, P. A new definition and a lectotypification of the genus Cooksonia Lang 1937. International Journal of Plant Sciences 2010, 171, 199–215. [Google Scholar] [CrossRef]
- Herr, J.M.J. The origin of the ovule. American Journal of Botany 1995, 82, 547–564. [Google Scholar] [CrossRef]
- Crane, P.R.; Kenrick, P. Diverted development of reproductive organs: A source of morphological innovation in land plants. Plant Systematics and Evolution 1997, 206, 161–174. [Google Scholar] [CrossRef]
- Taylor, T.N.; Taylor, E.L.; Krings, M. Paleobotany: the biology and evolution of fossil plants, 2nd ed.; Elsevier: Amsterdam, 2009; p. 1230. [Google Scholar]
- Williams, J.H. Amborella trichopoda (Amborellaceae) and the evolutionary developmental origins of the angiosperm progamic phase. American Journal of Botany 2009, 96, 144–165. [Google Scholar] [CrossRef]
- Rabe, A.J.; Soltis, D.E. Pollen tube growth and self-incompatibility in Heuchera micrantha var. diversifolia (Saxifragaceae). International Journal of Plant Sciences 1999, 160, 1157–1162. [Google Scholar] [CrossRef]
- Sage, T.L.; Hristova-Sarkovski, K.; Koehl, V.; Lyew, J.; Pontieri, V.; Bernhardt, P.; Weston, P.; Bagha, S.; Chiu, G. Transmitting tissue architecture in basal-relictual angiosperms: implications for transmitting tissue origins. American Journal of Botany 2009, 96, 183–206. [Google Scholar] [CrossRef]
- Thien, L.B.; Bernhardt, P.; Devall, M.S.; Chen, Z.D.; Luo, Y.B.; Fan, J.H.; Yuan, L.C.; Williams, J.H. Pollination biology of basal angiosperms (ANITA grade). American Journal of Botany 2009, 96, 166–182. [Google Scholar] [CrossRef]
- Wetzstein, H.Y.; Porter, J.A.; Janick, J.; Ferreira, J.F.S. Flower morphology and floral sequence in Artemisia annua (Asteraceae)1. American Journal of Botany 2014, 101, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Judd, W.S.; Campbell, S.C.; Kellogg, E.A.; Stevens, P.F. Plant systematics: a phylogenetic approach; Sinauer Associate Inc.: Sunderland, MA, 1999. [Google Scholar]
- He, C.Y.; Saedler, H. Molecular evolution of a morphological novelty in Solanaceae, the Inflated-Calyx-Syndrome (ICS) in Physalis. Acta Horticulturae 2007, 745, 171–182. [Google Scholar] [CrossRef]
- Posluszny, U.; Tomlinson, P.B. Aspects of inflorescence and floral development in the putative basal angiosperm Amborella trichopoda (Amborellaceae). Canadian Journal of Botany 2003, 81, 28–39. [Google Scholar] [CrossRef]
- Fu, Q.; Diez, J.B.; Pole, M.; Garcia-Avila, M.; Liu, Z.-J.; Chu, H.; Hou, Y.; Yin, P.; Zhang, G.-Q.; Du, K.; et al. An unexpected noncarpellate epigynous flower from the Jurassic of China. eLife 2018, 7, e38827. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Diez, J.B.; Pole, M.; García-Ávila, M.; Wang, X. Nanjinganthus is an angiosperm, isn't it? China Geology 2020, 3, 359–361. [Google Scholar] [CrossRef]
- Fu, Q.; Hou, Y.; Yin, P.; Diez, J.B.; Pole, M.; García-Ávila, M.; Wang, X. Micro-CT results exhibit ovules enclosed in the ovaries of Nanjinganthus. Scientific Reports 2023, 13, 426. [Google Scholar] [CrossRef]
- Elias, L.G.; Kjellberg, F.; Farache, F.H.A.; Almeida, E.A.B.; Rasplus, J.-Y.; Cruaud, A.; Peng, Y.-Q.; Yang, D.-R.; Pereira, R.A.S. Ovipositor morphology correlates with life history evolution in agaonid fig wasps. Acta Oecologica 2018, 90, 109–116. [Google Scholar] [CrossRef]
- Fu, Q.; Liu, J.; Wang, X. Offspring development conditioning (ODC): A universal evolutionary trend in sexual reproduction of organisms. Journal of Northwest University (Natural Science Edition) 2021, 51, 163–172. [Google Scholar] [CrossRef]
- Wang, X. Elite of plants; Phoenix Science Press: Nanjing, 2023; p. 168. [Google Scholar]
- Lorence, D.H. A monograph of the Monimiaceae (Laurales) in the Malagasy region (Southwest Indian Ocean). Annals Missouri Botanical Garden 1985, 72, 1–165. [Google Scholar] [CrossRef]
- Endress, P.K. Ontogeny, function and evolution of extreme floral construction in the Monimiaceae. Plant Systematics and Evolution 1980, 134, 79–120. [Google Scholar] [CrossRef]
- Renner, S.S.; Hausner, G. Siparunaceae, Monimiaceae. In Flora of Ecuador; Harling, G., Anderson, L., Eds.; Berlings Arloev: Sweden, 1997; Volume 59, pp. 1–125. [Google Scholar]
Figure 1.
Sexual reproductive cycle in its simple and primitive mode. Note that there is no visible difference between two gametes, and both diploid and haploid are unicellular. This cycle can be repeated infinitely, forming a phylogeny. Different generations are marked with background of black or white color.
Figure 1.
Sexual reproductive cycle in its simple and primitive mode. Note that there is no visible difference between two gametes, and both diploid and haploid are unicellular. This cycle can be repeated infinitely, forming a phylogeny. Different generations are marked with background of black or white color.
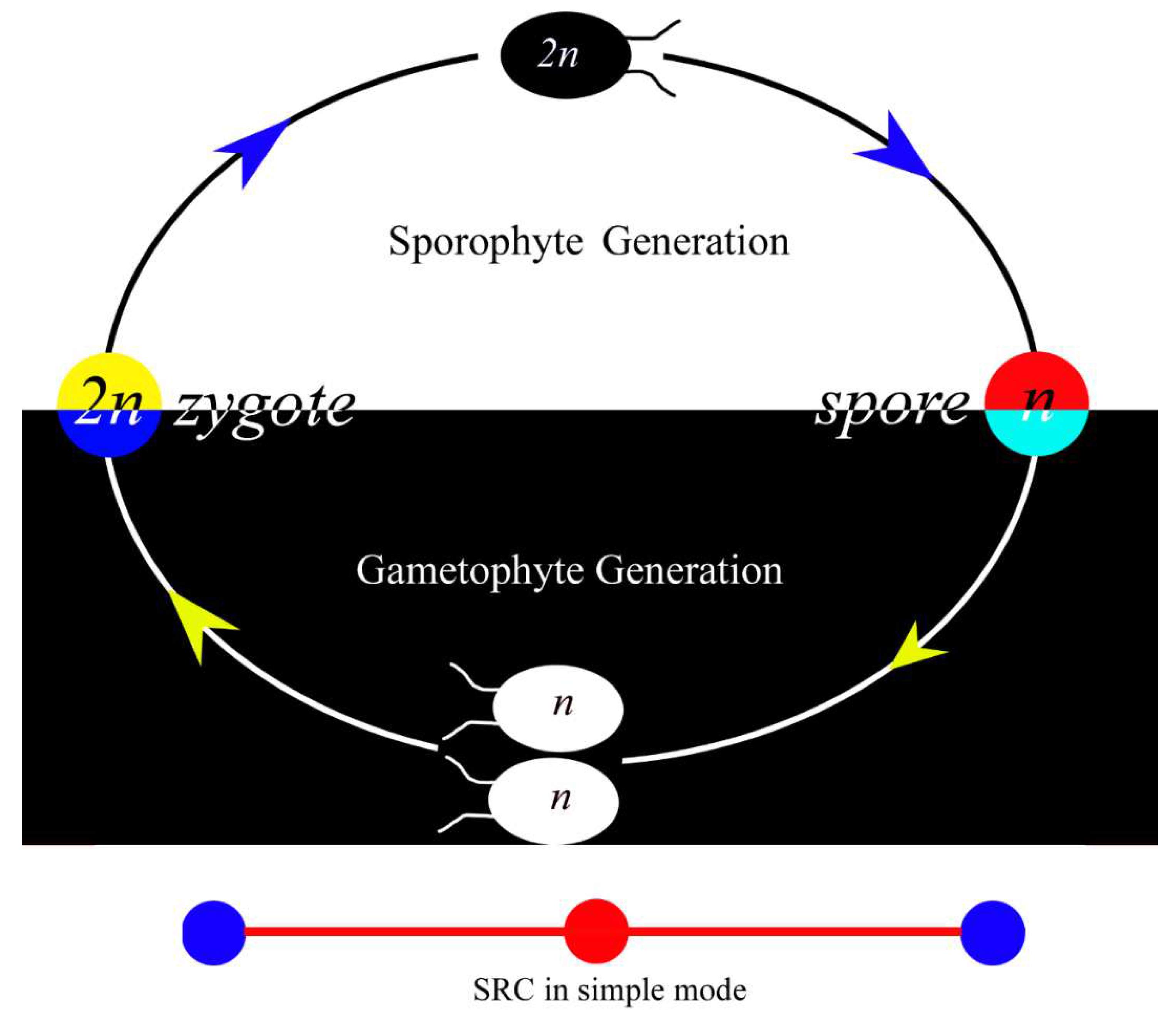
Figure 2.
Differentiation of gametes, from isomorphic to heteromorphic.
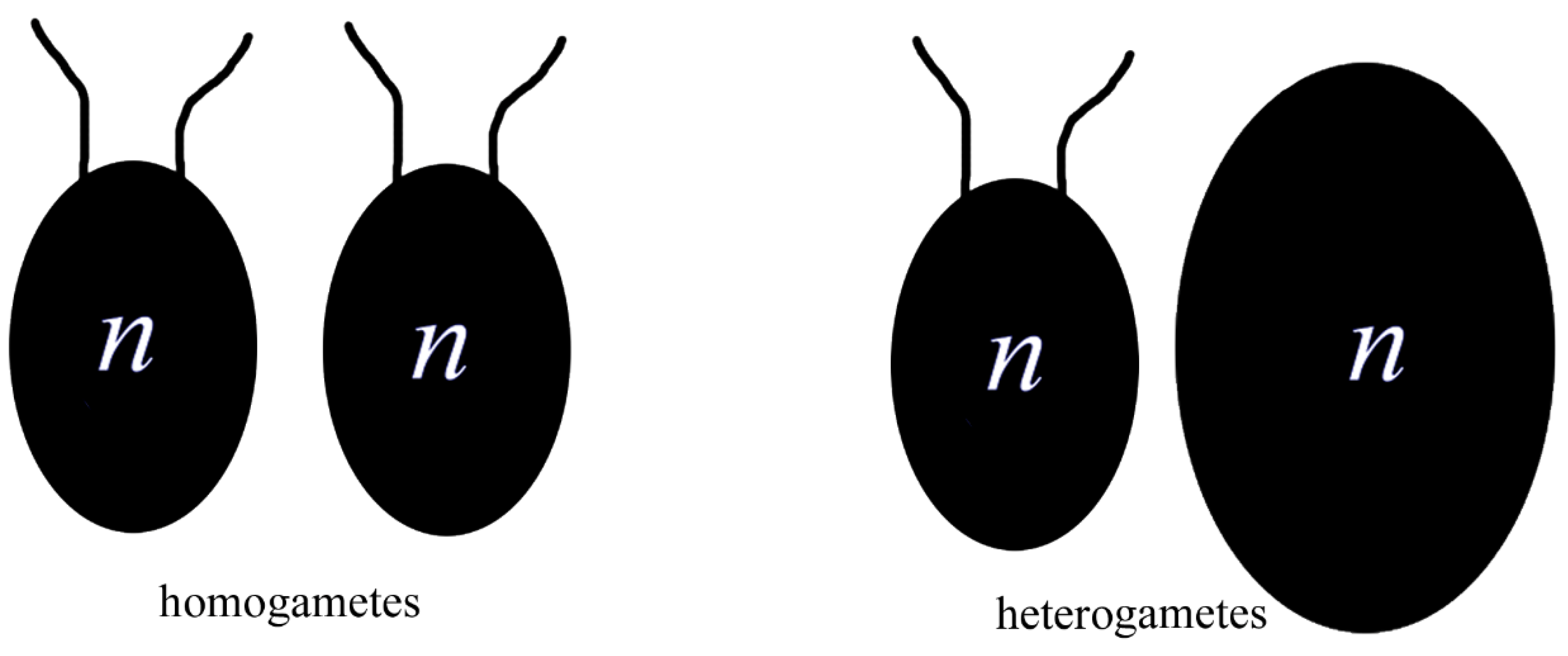
Figure 3.
From unicellularity to multicellularity, which enables complex morphology.
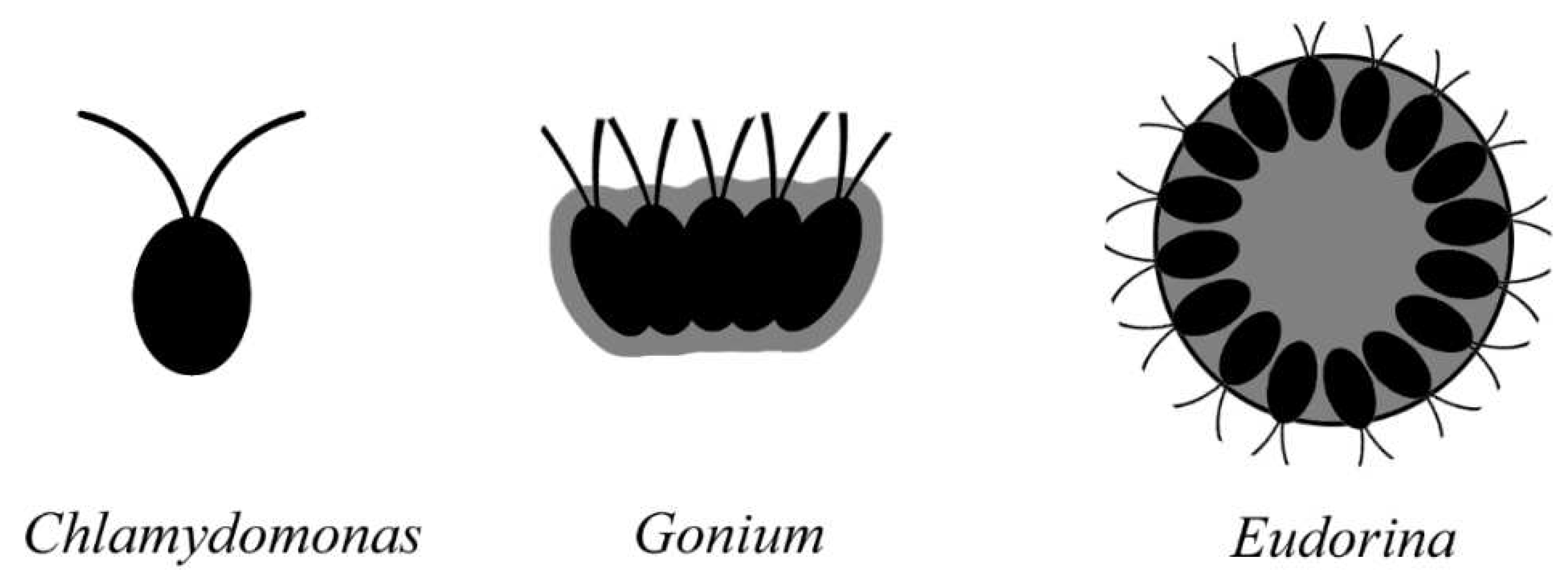
Figure 4.
Differentiation between somatic and reproductive parts. Reproduced from Fig. 8.3 of Wang (2018)[5]. Courtesy of Dr. Lei Chen.
Figure 4.
Differentiation between somatic and reproductive parts. Reproduced from Fig. 8.3 of Wang (2018)[5]. Courtesy of Dr. Lei Chen.

Figure 7.
Differentiation between microspore and megaspore, as well as between microsporangium and megasporangium.
Figure 7.
Differentiation between microspore and megaspore, as well as between microsporangium and megasporangium.
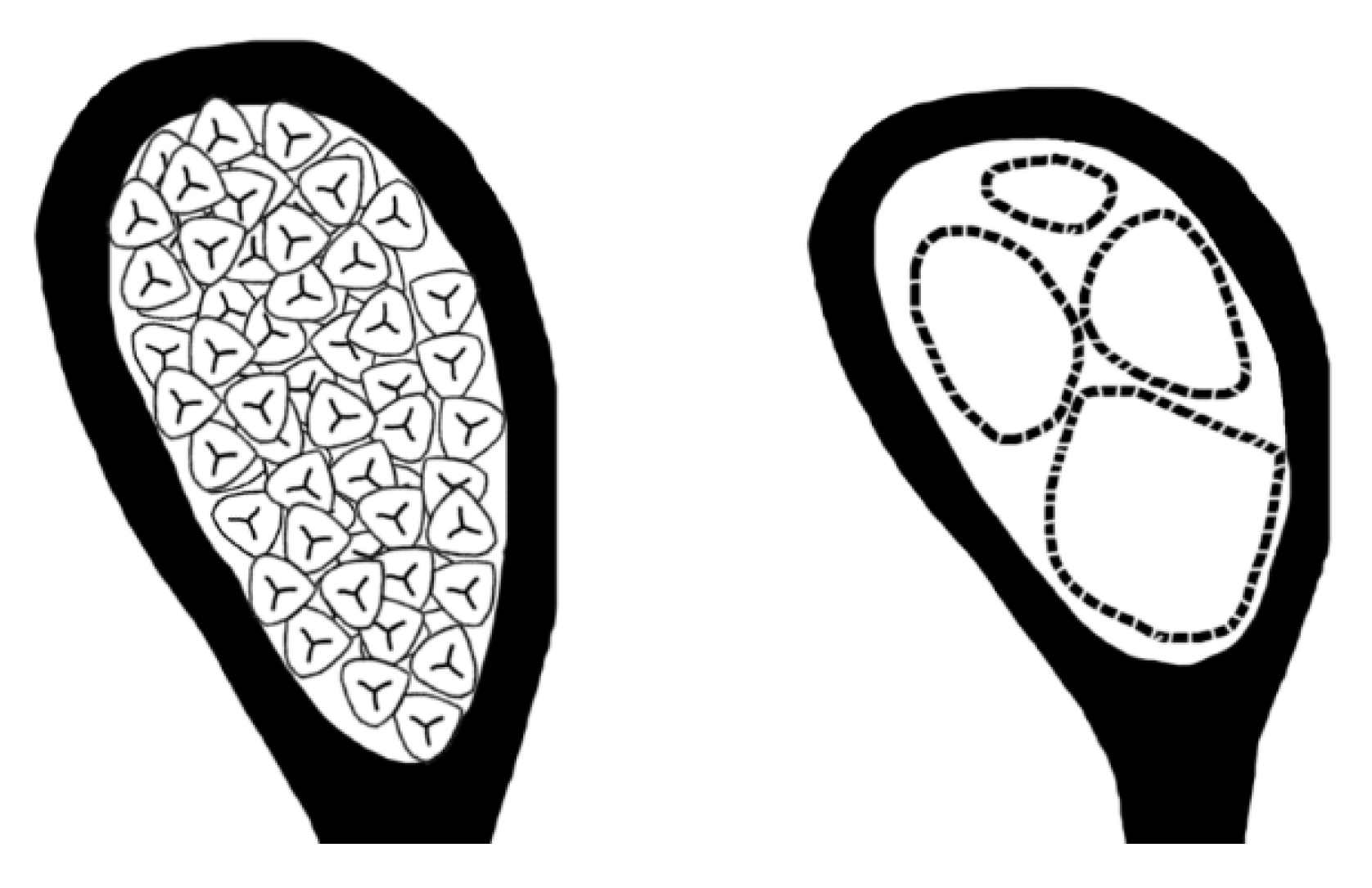
Figure 8.
Metamegasporangium (= ovule), developmental precursor of seed, and more complicated SRC. Note the nucellus (gray in color) has an archegonium-bearing gametophyte inside and is protected by integument, which is derived from sterilized sporangia and their pedicels. Compared to the SRC mode in Figure 6, heterospory and endospory are added to the SRC.
Figure 8.
Metamegasporangium (= ovule), developmental precursor of seed, and more complicated SRC. Note the nucellus (gray in color) has an archegonium-bearing gametophyte inside and is protected by integument, which is derived from sterilized sporangia and their pedicels. Compared to the SRC mode in Figure 6, heterospory and endospory are added to the SRC.
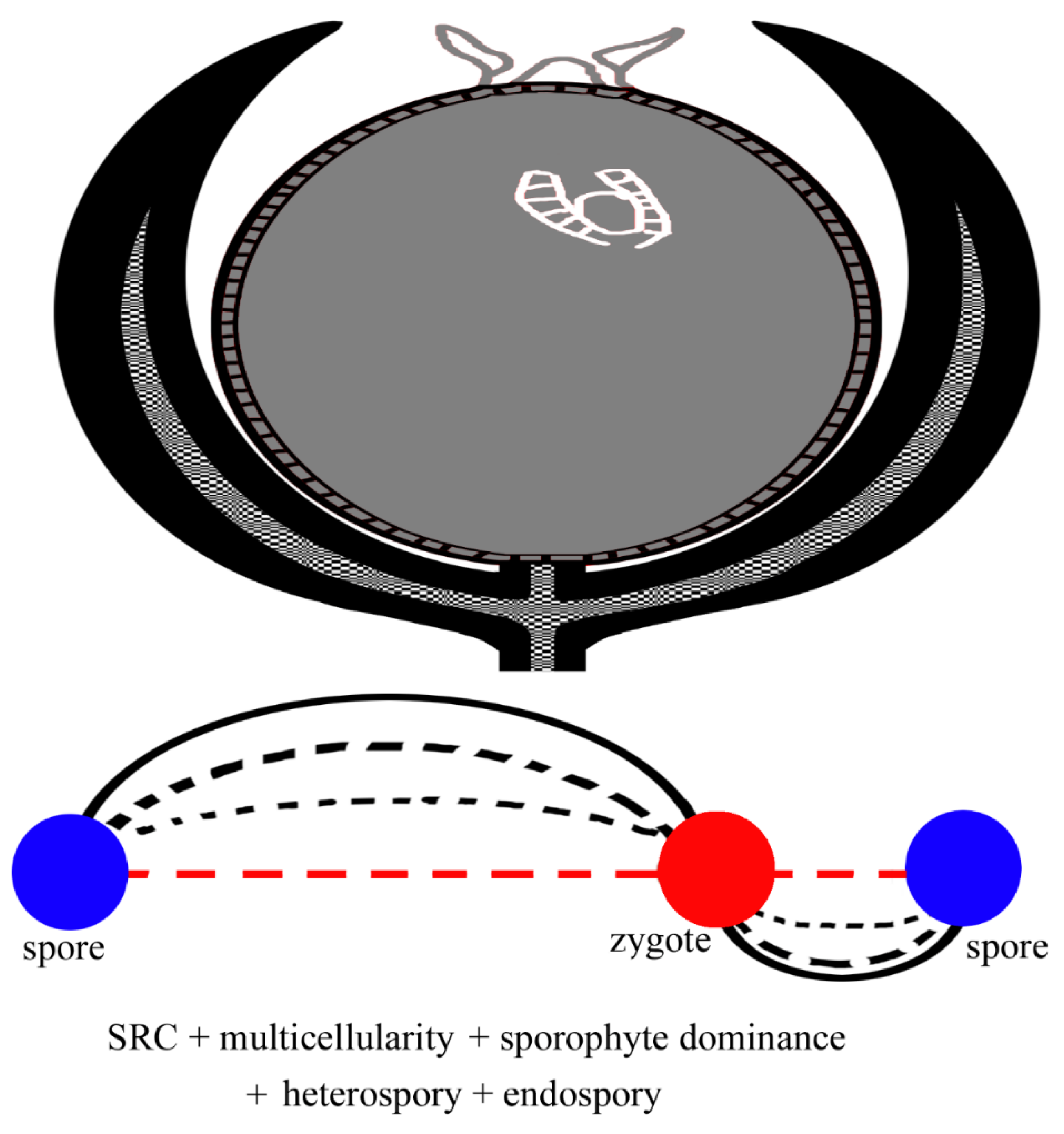
Figure 9.
Primary metamegasporangium complexes, with various ovule-protecting mechanisms, in different gymnosperms. Ovules are gray in color. a, Cycas. b, Zamia. c, Conifers. d, Caytonia.
Figure 9.
Primary metamegasporangium complexes, with various ovule-protecting mechanisms, in different gymnosperms. Ovules are gray in color. a, Cycas. b, Zamia. c, Conifers. d, Caytonia.
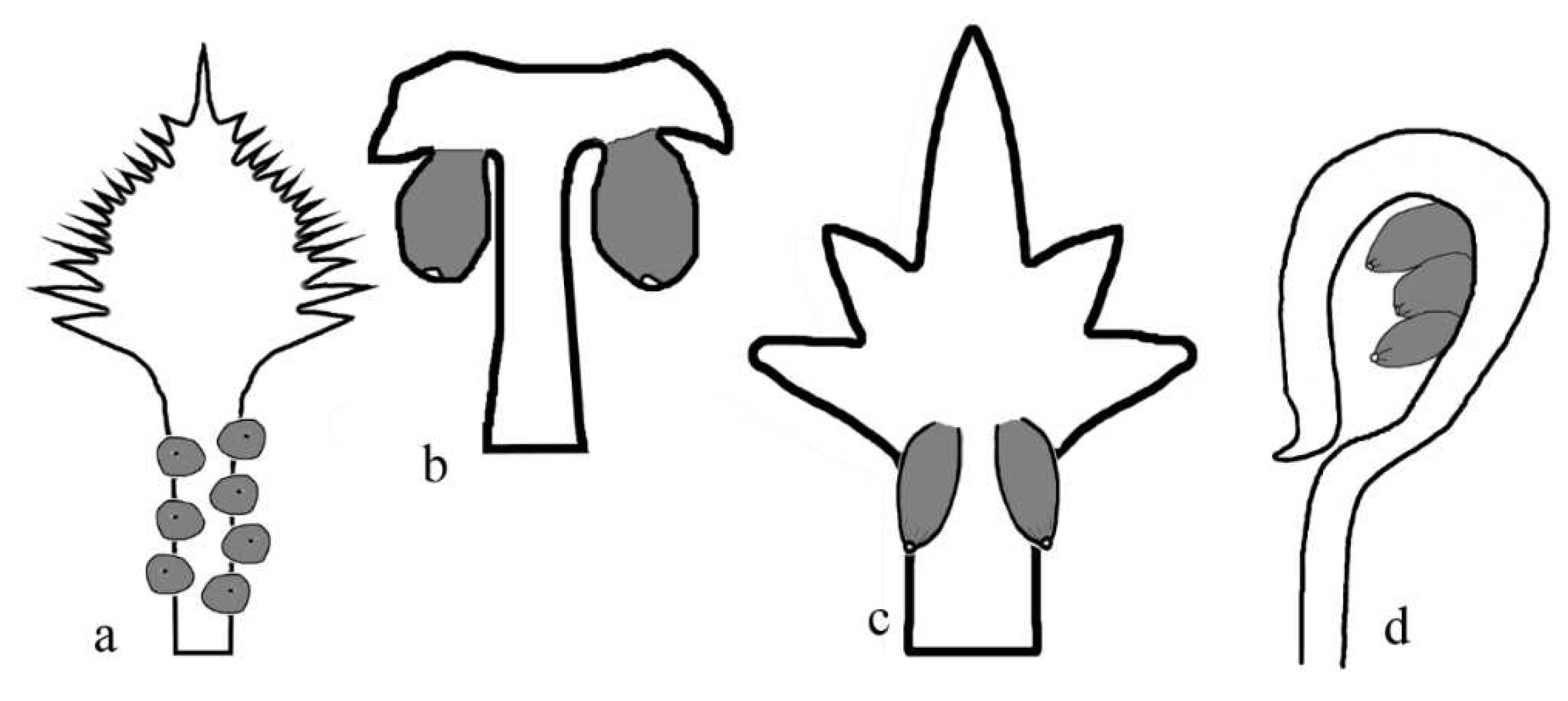
Figure 10.
Secondary metamegasporangium complexes, with various ovule-protecting structures. Ovules are gray in color. a, Amborellla. b, a Jurassic fossil plant. c, Cactaceae.
Figure 10.
Secondary metamegasporangium complexes, with various ovule-protecting structures. Ovules are gray in color. a, Amborellla. b, a Jurassic fossil plant. c, Cactaceae.
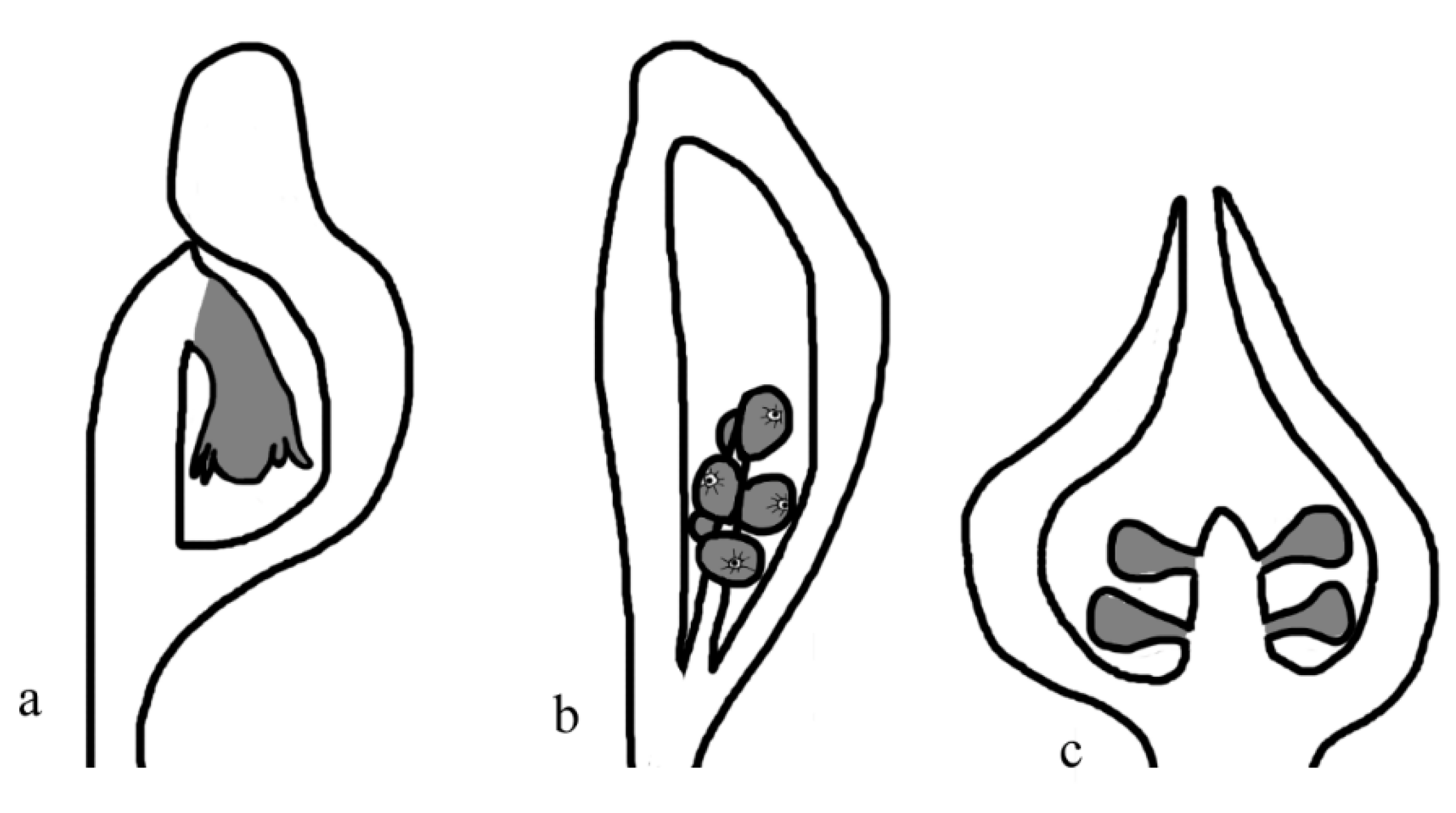
Figure 11.
Tertiary metamegasporangium complex I, with various fruit-protecting structures. a, Ficus. b, Siparuna. c, Chaoyangia liangii, a Cretaceous fossil plant. Fruits are gray in color.
Figure 11.
Tertiary metamegasporangium complex I, with various fruit-protecting structures. a, Ficus. b, Siparuna. c, Chaoyangia liangii, a Cretaceous fossil plant. Fruits are gray in color.
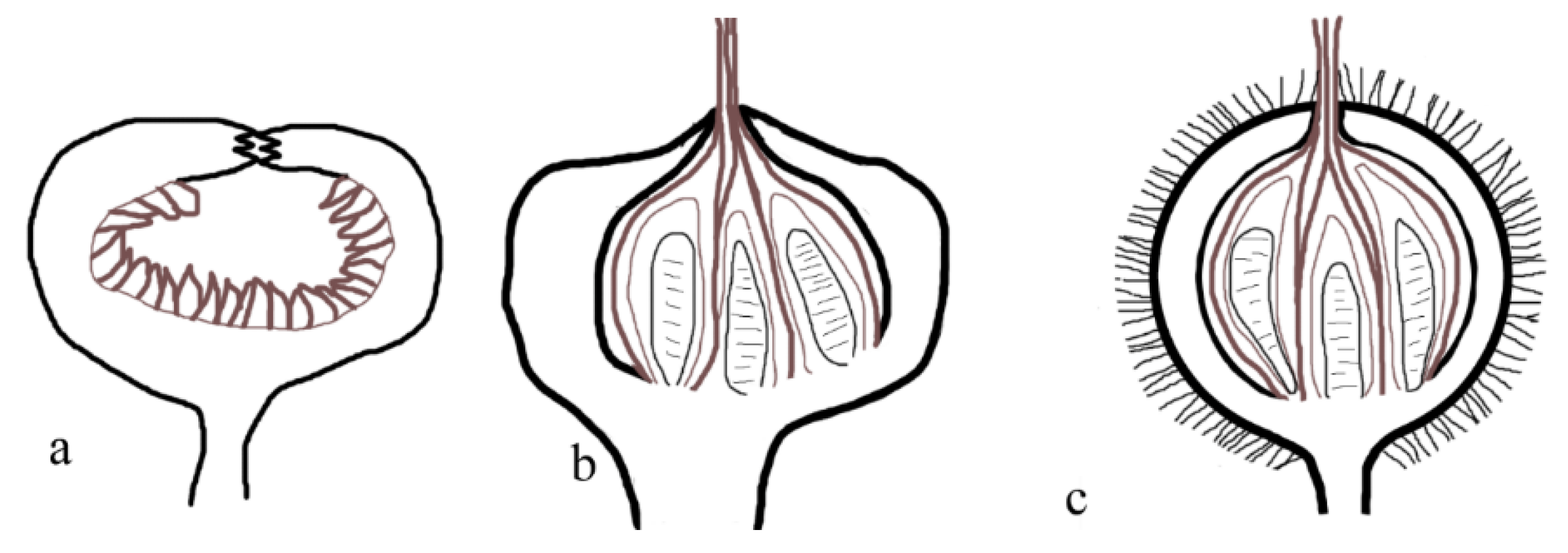
Figure 12.
Tertiary metamegasporangium complex II, aggregating flowers into an inflorescence.

Figure 13.
Tertiary metamegasporangium complex III, extended nutritional bond between two generations of sporophytes. Reproduced from Wang [24].
Figure 13.
Tertiary metamegasporangium complex III, extended nutritional bond between two generations of sporophytes. Reproduced from Wang [24].

Figure 14.
Inferior ovary in a Jurassic angiosperm, Nanjinganthus, may be derived through expansion and invagination of a flower axis tip. Reproduced from Fu et al. (2018, 2023)[19,21].

Figure 15.
Stairs of land plant reproduction evolution. Tracing a specific phylogeny of plants, the novelties of plant evolution, if existing, occurred sequentially, probably following this order.
Figure 15.
Stairs of land plant reproduction evolution. Tracing a specific phylogeny of plants, the novelties of plant evolution, if existing, occurred sequentially, probably following this order.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



