Submitted:
25 October 2023
Posted:
26 October 2023
Read the latest preprint version here
Abstract
It was of great practical significance to repair the toxicity of salt stress environment to soybean seedlings and maintain the balance between agricultural production and ecological environment in the Yellow River valley of China. ‘Xudou 20’ was used as the experimental material, a large-scale planting variety in the old Yellow River basin. Endophytes 131 and 137 isolated from soybean root nodule were taken as the research objects, and the soybean plants were cultivated by pot culture in intelligent artificial incubator. Salt stress group, bacteria suspension group and control group were set up to study the effects of inoculation treatments on the activities of superoxide dismutase (SOD) and catalase (CAT) of soybean seedlings under different NaCl stress. The results showed that when inoculated with endophyte 131 bacterial suspension and cultured for 14 days, the SOD activity was the highest (14.89 U•g−1•FW) at the salt concentration 50 mmol•L-1. When inoculated with 131 bacterial suspension (2:1) and cultured for 28 days, the SOD activity reached the highest (0.94 ug-1FW) at the salt concentration of 50 mmol•L-1, and the CAT activity amounted to the highest (1910 U•g−1•min−1). The 16S rDNA sequencing and phylogenetic analysis showed that the most similar strain of strain 137 was Bacillus cereus, and the most similar strain of strain 131 was Enterobacter ludwigii. Therefore, the activities of SOD and CAT in soybean seedlings inoculated with endophytes 131 and 137 increased at different degrees under salt stress, which could effectively alleviate the damage of salt stress environment to plants, and thus improve the stress resistance of plants.
Keywords:
endophytic bacteria
; soybean seedlings
; salt stress
; SOD
; CAT
; resistance
; repair capability
1. Introduction
Soybean (Glycine max L.) belongs to the leguminous family, with a protein content of about 40% [1]. During its growth process, salt stress is a serious abiotic environmental hazard that affects the growth and development of soybeans, leading to a decrease in their yield and quality [2]. In general, the reactive oxygen system in soybean seedlings maintains a Dynamic equilibrium state. When it is under salt stress, the production and elimination mechanism of reactive oxygen in the body is inhibited, which destroys the Dynamic equilibrium of the system [3], affects its normal nutritional growth, and causes plant damage. Among them, the antioxidant enzymes that eliminate reactive oxygen species in soybean seedlings mainly include Catalase (CAT), Superoxide dismutase (SOD), etc.[4]. Under salt stress, it can induce the accumulation of reactive oxygen species such as superoxide free radicals (O2-) and hydrogen peroxide (H2O2) in plants[5], causing membrane lipid peroxidation, damaging cell membranes, exacerbating intracellular electrolyte leakage, and causing imbalance in cellular reactive oxygen metabolism[6]. At this time, the plant can accelerate the removal of active oxygen, repair the damage and effectively maintain the Dynamic equilibrium of the active oxygen system in the plant by increasing the activities of antioxidant enzymes[7,8], such as CAT and SOD, to reduce the damage of salt stress environment to the plant. In 2022, the planting area of soybean seedlings in China was 140 million acres, a decrease of 5.4% compared to the previous year. Some scientists adopt breeding methods to improve salt tolerance, but the cycle is long and the cost is high. Therefore, finding a green and environmentally friendly measure to enhance the salt tolerance of soybean seedlings and improve plant repair ability is of great practical significance for expanding China’s arable land area, especially for the high-quality development of agricultural production and ecological environment protection in the Yellow River Basin.
Endophytes refer to microbial groups that live between or within the host plant tissues during a certain stage or entire process of their life cycle and do not cause obvious harmful symptoms in the host plant[9]. They mainly include bacteria, fungi, or actinomycetes. Endophytic bacteria have strong colonization ability and can produce various antibacterial active substances in the host, resisting the invasion of plant pathogenic fungi[10,11]. Endophytes can not only promote the growth and development of the host, but also provide host plants with metabolites such as alkaloids, antibiotics, Hydrolase and other defense against external adverse environmental stimuli[12]. They evolve with the host, have the specific function of changing their species, and are important factors in repairing salt stress stress[13]. Research by Wen Caiyi et al.[14] has shown that endophytic bacteria form a symbiotic nitrogen fixation system with leguminous plants, which has the function of improving soil salinization toxicity, promoting a virtuous cycle between crops and the environment, and enhancing the ability of crops to resist adversity. The plant microorganism interaction alleviates the stress response of plants and helps the host recover after stress through various mechanisms. In particular, the root microbiome provides a variety of benefits for host plants, such as promoting plant growth, supporting nutrient absorption, improving tolerance to Abiotic stress, protecting plant hosts from pathogens, and regulating the plant immune system to induce resistance[15]. The previous research of the research group showed that endophyte could cooperate with soybean plants in salt stress environment to maintain the balance of plant Osmotic pressure, improve the salt tolerance of soybean plants, and promote the growth and development of soybean seedlings. Based on this, this study intends to use two endophytic bacteria 131 and 137, using Xudou 20 as experimental materials, to study the effects of endophytic bacteria 131 and 137 on SOD and CAT activities in soybean seedlings under salt stress. This study has important practical significance in repairing the toxicity of salt stress on soybean seedlings, maintaining agricultural production, ecological environment balance, and developing arable land.
2. Results
2.1. Sequence Determination and Phylogenetic Status of 16S rRNA Gene in Endophytic Bacteria
The 16S rRNA gene sequence size of two strains of bacteria is 1500 bp, and the most similar type strains were searched in the NCBI database to construct a phylogenetic tree (Figure 1). The entire phylogenetic tree has two main branches and one marginal branch (Coprococcus catus VPI-C6-61T). The main branch includes two genera: Bacillus and Enterobacter. Among them, strain 137 and type strain Bacillus cereus IAM 12605T (D16266) are located in the main branch I, with a maximum similarity rate of 99.8%. Strain 131 and type strain Enterobacter ludwigii EN-119T(AJ853891) are located in another main branch II, with a maximum similarity rate of 99.7%. Therefore, the most similar strain of 137 is Bacillus cereus, and the most similar strain of 131 is Enterobacter ludwigii.
2.2. SOD Activity Analysis
2.2.1. SOD Activity Analysis at 14 Days
As shown in Figure 2, after 14 days of cultivation of soybean seedlings, the overall SOD activity in the salt stress group was relatively stable. After inoculation with endophytic bacterium 131, as the salt concentration gradient continued to increase, SOD activity decreased, indicating that the antioxidant capacity of soybean seedlings under salt stress was limited by the high or low salt concentration. Inoculated with a 1:1 bacterial suspension, the highest SOD activity was 14.89 U·g-1 FW at a salt concentration of 50 mmol·L-1, which was significantly higher than the CK group, salt stress group, and other inoculation treatment groups. This indicates that the antioxidant capacity of soybean seedlings treated with inoculation under salt stress. After inoculation with a 1:2 bacterial suspension, the SOD activity was the highest at a salt concentration of 50 mmol·L-1, at 7.71 U·g-1 FW. The trend of SOD activity change was the same as that of the 1:1 bacterial suspension treatment group, but lower than that of the 1:1 bacterial suspension treatment group. After inoculation with 2:1 bacterial suspension, the activity was the highest at a salt concentration of 50 mmol·L-1, reaching 13.56 U·g-1FW, indicating that the 2:1 bacterial suspension has a significant repair effect on soybean seedlings. From this, it can be seen that when soybean seedlings were cultured for 14 days, the three bacterial suspensions of 131 endophytic bacteria were inoculated under salt stress, and all had repair effects at a salt concentration of 50 mmol·L-1, but the 1:1 bacterial suspension had better repair effects.
Figure 3 shows that the SOD activity of soybean seedlings in the salt stress group and the inoculation treatment group increased first and then decreased after 14 days of culture. After inoculation of endophytic bacteria 137, the SOD activity increased gradually when the salt concentration was 100-250 mmol·L-1, and the SOD activity was the highest when the salt concentration was 250 mmol·L-1 (0.84 u·g-1FW). When the salt concentration was between 250-300 mmol·L-1, the SOD activity gradually decreased, but was higher than that of all experimental groups. When inoculated with 1:1 bacterial suspension, the salt concentration was from 50 to 300 mmol·L-1, the SOD activity gradually increased and then decreased, and the active SOD activity was the highest when the salt concentration was 200 mmol·L-1. After inoculation with 2:1 bacterial suspension, the activity of SOD was the highest at the salt concentration of 200 mmol·L-1, which was 0.45 u·g-1FW, indicating that the inoculation with 2:1 bacterial suspension had good repair ability. Therefore, when soybean seedlings were cultured for 14 days, inoculation of 137 endophytes at salt concentration greater than 100 mmol·L-1 could significantly (P<0.05) improve the SOD activity of soybean seedlings and reduce the toxicity to soybean seedlings. Among them, the repair effect of 137 endophytic bacteria 1:2 suspension was the most obvious.
2.2.2. SOD Activity Analysis at 28 Days
When the cultivation time of soybean seedlings was 28 days, the overall change in SOD activity in the salt stress group showed a “decrease-increase-decrease” pattern (Figure 4). After inoculation with 2:1 bacterial suspension, the activity was the highest at a salt concentration of 50 mmol·L-1, at 0.94 U·g-1FW, significantly higher than the SOD activity in the salt stress group, and gradually rebounded under other salt concentrations. After inoculation with 1:1 bacterial suspension, the highest SOD activity was observed at a salt concentration of 100 mmol·L-1, at 0.54 U·g-1FW, which was higher than that of the salt stress group. Overall, the SOD activity was complementary to the salt stress group. After inoculation with a 1:2 bacterial suspension, the activity was the highest at a salt concentration of 100 mmol·L-1, at 0.72 U·g-1FW. At salt concentrations of 100 mmol·L-1, 150 mmol·L-1, and 300 mmol·L-1, the SOD activity was higher than that of the salt stress group and other inoculation treatment groups. Explanation: The 1:2 of bacterial suspension has a significant repair effect at a certain salt concentration. From this, it can be seen that when the cultivation time of soybean seedlings is 28 days, the inoculation of 131 endophytic bacteria under salt stress has relatively better repair ability than the control group.
When the cultivation time of soybean seedlings was 28 days, the overall SOD activity in the salt stress group showed an upward trend (Figure 5). After inoculation with 1:1 bacterial suspension, the highest activity (0.41 U·g-1FW) was observed at a salt concentration of 300 mmol·L-1, which was lower than the salt stress group. This indicates that inoculation with 1:1 bacterial suspension under salt stress did not have a repair effect. After inoculation with a 1:2 bacterial suspension, the activity was highest at a salt concentration of 100 mmol·L-1, at 0.58 U·g-1FW. Except for salt concentrations of 50 and 100 mmol·L-1, SOD activity was lower in all other salt concentrations than in the salt stress group. After inoculation with 2:1 bacterial suspension, with the increase of salt concentration, the SOD activity was significantly lower than that of the salt stress group. From this, it can be seen that soybean seedlings grow for 28 days and are inoculated with 137 endophytic bacteria in a 1:2 bacterial suspension. When the salt concentration is 50 and 100 mmol·L-1, the repair effect is better than that of the salt stress group.
2.3. CAT Activity Analysis
2.3.1. CAT Activity Analysis at 14 d
From Figure 6, it can be seen that under salt stress, when soybean seedlings grew for 14 days and were inoculated with 1:1 bacterial suspension, the CAT activity was the highest at a salt concentration of 50 mmol·L-1, at 1230 U·g-1·min-1, and lowest at a salt concentration of 200 mmol·L-1, at 870 U·g-1·min-1, indicating that the best repair effect was achieved at 50 mmol·L-1. After inoculation with a 1:2 bacterial suspension, the highest CAT activity was 1180 U·g−1·min−1 at a salt concentration of 50 mmol·L-1. As the salt concentration gradient increased, CAT activity decreased. At a salt concentration of 250 mmol·L-1, the lowest activity was 850 U·g−1·min−1, indicating that the 1:2 bacterial suspension had a relatively small repair effect on soybean seedlings. After inoculation with 2:1 bacterial suspension, the CAT activity showed a continuous increasing trend with the increase of salt concentration gradient. The activity reached its highest at a salt concentration of 300 mmol ·L-1, reaching 1380 U·g −1·min−1. The CAT activity was the lowest at 100 mmol·L-1, reaching 1030 U·g−1·min−1. At salt concentrations of 250 and 300 mmol·L-1, the activity was higher than the salt treatment group, while at other concentrations, it was lower than the salt treatment group. From this, it can be seen that when soybean seedlings grow for 14 days, the inoculation of endophytic bacteria 131 suspension in a 2:1 ratio has the strongest repair ability for soybean seedlings when the salt concentration is greater than 300 mmol·L-1, effectively alleviating the persecution of soybean seedlings and improving their salt tolerance.
As shown in Figure 7, when soybean seedlings grew for 14 days under salt stress, the salt stress group had the highest activity at a salt concentration of 50 mmol·L-1, which was 1650 U·g−1·min−1. CAT activity was higher than the control group at salt concentrations of 50 and 100 mmol·L-1, indicating that the antioxidant capacity of soybean seedlings under salt stress was limited by the high or low salt concentration. After inoculation with a 1:1 suspension of endophytic bacteria in a ratio of 137, the CAT activity showed an “increase decrease” trend with the increase of salt concentration. At a salt concentration of 50 mmol·L-1, the activity was the highest, at 1530 U·g−1·min−1, and was lower than that of the salt stress group, indicating that the 1:1 suspension had a smaller repair effect. After inoculation with a 1:2 bacterial suspension, the CAT activity showed an “increase decrease” trend, with the highest activity at 100 mmol·L-1, at 1840 U·g − 1·min −1. The activity was higher than that of the salt stress group at salt concentrations of 50, 100, 150, and 200 mmol·L-1, indicating that a 1:2 bacterial suspension has a good repair effect. After inoculation with 2:1 bacterial suspension, the activity was highest at a salt concentration of 100 mmol·L-1, at 1580 U·g-1·min-1, and lowest at a salt concentration of 250 mmol·L-1, at 1230 U·g-1 ·min-1. Except for the enzyme activity at 150 mmol·L-1, which was higher than the salt stress group, CAT activity was lower at each other salt concentration than the salt stress group, indicating that the repair ability of 2:1 bacterial suspension was different at each salt concentration, And the repair ability is best only when the salt concentration is 100 mmol·L-1. It can be seen that when soybean seedlings grow for 14 days, inoculating endophytic bacteria 137 and inoculating 1:2 bacterial suspension at 100 mmol·L-1 has the most significant repair effect (P<0.05), which can effectively alleviate salt stress and improve the salt tolerance of soybean seedlings.
2.3.2. CAT Activity Analysis at 28 Days
It can be seen from Figure 8 that when the soybean seedlings grow to 28 days, the CAT enzyme activity is the highest when the salt concentration is 300 mmol·L-1, which is 1950 u·g−1·min −1. The cat activity of the salt stress group is higher than that of the control group, indicating that the soybean seedlings have strong antioxidant ability under natural conditions. After inoculation with 1:1 endophytic bacteria suspension, the activity was the highest at 50 mmol·L-1 salt concentration, which was 1910 u·g−1·min−1, and the cat activity was the lowest at 250 mmol·L-1 salt concentration, which was 1280 u·g−1·min−1, indicating that 1:1 endophytic bacteria could increase the enzyme activity of soybean seedlings at 50 and 100 mmol·L-1 salt concentration. After inoculation with 1:2 endophytic bacteria suspension, cat enzyme activity increased first and then decreased, and the highest activity was 1590 u·g−1 ·min−1 when the salt concentration was 200 mmol·L-1, indicating that the repair effect of 1:2 endophytic bacteria suspension was limited when the salt concentration was increasing. The highest activity (1740 u·g−1·min−1) was observed when the concentration of salt was 300 mmol·L-1 after inoculation of 2:1 bacterial suspension, but the enzyme activities were lower than those in the salt stress group, indicating that the repair ability of 2:1 bacterial suspension was weak. In conclusion, when the soybean seedlings grew for 28 days, the effect of 1:1 endophyte inoculation was the best when the salt concentration was 50 mmol·L-1, which could effectively alleviate the damage of salt stress on soybean seedlings.
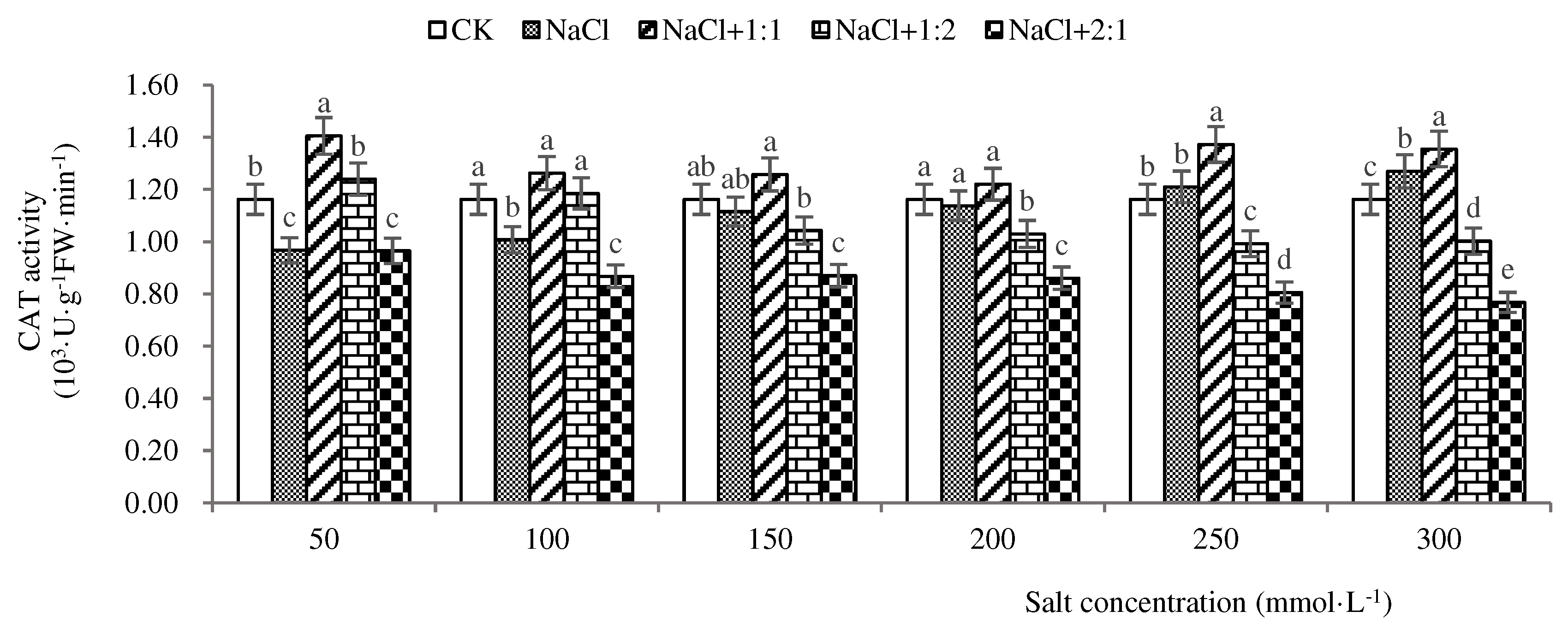
From Figure 9, it can be seen that when soybean seedlings grow to 28 days, the CAT activity is highest at a salt concentration of 300 mmol·L-1, which is 1270 U·g-1·min-1. At a salt concentration of 50 mmol·L-1, the CAT activity is lowest, which is 970 U·g-1·min-1. This indicates that the antioxidant capacity of soybean seedlings under salt stress is limited by salt concentration. After inoculation with 1:1 bacterial suspension, CAT activity showed a “decrease increase” trend with increasing salt concentration. The activity was highest at a salt concentration of 50 mmol·L-1, at 1410 U·g-1·min-1, and lowest at a salt concentration of 200 mmol·L-1, at 1220 U·g-1 ·min-1, indicating that the remediation effect of 1:1 bacterial suspension was weak. After inoculation with a 1:2 bacterial suspension, the overall CAT activity showed a decreasing trend. The enzyme activity was highest at a salt concentration of 50 mmol·L-1, at 1240 U·g−1·min−1. At 50 and 100 mmol·L-1, the CAT activity was higher than that of the control group and salt stress group, indicating that a 1:2 bacterial suspension has limited repair ability for soybean seedlings. After inoculation with 2:1 bacterial suspension, the enzyme activity was highest at a salt concentration of 50 mmol·L-1, reaching 970 U·g −1·min−1, and lowest at a salt concentration of 300 mmol·L-1, reaching 770 U·g−1·min−1. This indicates that inoculation with 2:1 bacterial suspension has no effect on soybean seedling repair. From this, it can be seen that when soybean seedlings grow to 28 days, inoculation with endophytic bacteria 137 can effectively alleviate the antioxidant capacity of natural soybean seedlings under salt stress and improve their salt tolerance when exposed to a 1:1 bacterial suspension.
3. Discussion
3.1. Effect of Endophytic Bacteria on SOD Activity of Soybean Seedlings under Salt Stress
SOD is the main enzyme related to the homeostasis of reactive oxygen species in the aging process of organisms[16], which has the function of clearing reactive oxygen species formed during metabolic processes[17,18]. When the accumulation of reactive oxygen species in plants exceeds a certain amount, it will disrupt the redox balance of cells. Plants improve the damage of reactive oxygen species through different antioxidant mechanisms, maintaining a basically non-toxic level of reactive oxygen species [19,20]. When plants are subjected to stress, exogenous substances are applied to regulate the activity of antioxidant enzymes to a certain extent, enhancing the plant’s stress resistance. The research results of Waqas[21] indicate that under salt stress, with the increase of salt concentration and the extension of cultivation time, SOD activity shows a trend of first increasing and then decreasing. It may be that when the salt concentration is low, the accumulation of reactive oxygen species in soybean seedlings is low[22], and the amount of SOD that clears reactive oxygen species is relatively low, causing a temporary increase in SOD activity. As the salt concentration increases, the accumulation of reactive oxygen species increases, and SOD increases to eliminate excess reactive oxygen species in the body to reduce the damage of salt stress on soybean seedlings [23], which is beneficial for the growth of soybean seedlings under salt stress. Under salt stress, the SOD activity in soybean seedlings significantly increased after inoculation with endophytic bacteria. Among them, at 28 days, the SOD activity in the suspension treatment group of strain 137 was higher than that in the salt stress group and CK group at different concentrations of salt. This indicates that endophytic bacterium 137 has a significant (P<0.05) repair effect on soybean seedlings under salt stress. Han Kun[24] have shown that endophytic bacteria with ACC deaminase activity promote plant nutritional growth and improve salt tolerance. Zhang[25] believe that different concentrations of salicylic acid (SA) induced (inoculated) have a significant impact on SOD activity in double petal rose leaves. The above research results strongly support the results of this experiment. Khan[26] found that inoculating endophytic bacteria LK5 can greatly improve SOD activity in the body under the same stress conditions. Li et al.[27] believe that after inoculation, the SOD activity of highly resistant varieties is rapidly activated and increased. With the prolongation of stress time, the accumulation of reactive oxygen species intensifies the damage to the protective enzyme system, and SOD activity begins to decrease, thereby improving plant salt tolerance. The experimental results showed that after inoculation with strain 137, soybean seedlings showed significant repair ability under salt stress, while strain 131 had limited repair ability. The research results are basically consistent with those of Khan and Li, but the impact of different environmental conditions on the quality and yield of salt stressed soybean seedlings under strain 131 and 137 still needs further experimental verification in the field.
3.2. Effect of Endophytic Bacteria on CAT Activity of Soybean Seedlings under Salt Stress
CAT is an antioxidant enzyme in plants that plays an important role in the process of plant stress resistance [28,29]. When plants encounter harsh environments such as drought and salinity during their growth and development, a complex set of coping mechanisms is formed within the plant body [30]. The results showed that the overall CAT activity in soybean seedlings showed a decreasing trend under salt stress. Zhao[8] enhanced CAT activity in plants by inoculating endophytic bacteria 252 and 254, thereby demonstrating a repair effect. Yang et al. [31] showed that after inoculation with bacterial agents, they enter the interior of the plant, and the antioxidant capacity of the plant, represented by CAT activity and total phenol content, increases. These research results provide strong support for this experiment. Saravanakumar[32] found that endophytic bacteria in plants can increase the vitality index, fresh and dry weight of soybean seedlings, induce the production of water stress related proteins, enhance the activity of antioxidant enzymes such as CAT, and enhance their drought resistance. In this experiment, the CAT activity in soybean seedlings showed an overall upward trend after inoculation with endophytic bacteria under salt stress. At 28 days of soybean seedling growth, inoculation with 1:1 endophytic bacteria at a salt concentration of 50 mmol·L-1 had the best repair effect, which can effectively alleviate the damage of salt stress to soybean seedlings. The experimental results are basically consistent with the conclusions of Zhao, Yang and Saravanakumar. However, this experiment only studied the effect of endophytic bacteria on antioxidant enzymes in soybean seedlings under salt stress, and further research is needed on the molecular mechanism and other physicochemical indicators of salt tolerance in soybean seedlings.
3.3. Discussion on Salt Tolerance of Endophytic Bacteria Species
The 16S rDNA sequencing results and phylogenetic analysis showed that the most similar strain of strain 137 was Bacillus cereus, and the most similar strain of strain 131 was Enterobacter ludwigii. Up to now, there have been reports both domestically and internationally that inoculation with Enterobacter Ludwigii and Bacillus cereus enhances the biological activity of host plants, mainly focusing on cucumber, corn, tomato, and wheat, but there are very few reports on soybean as the host. Using soybeans as experimental materials, the effects of inoculation with Enterobacter Ludwig and Bacillus cereus on SOD and CAT activities in soybean seedlings under salt stress were studied, expanding the research field of inoculation with endophytic bacteria to repair host plants under salt stress. After inoculation with endophytic bacteria under salt stress, the overall CAT activity in soybean seedlings showed an upward trend. At 28 days of soybean seedling growth, inoculation with 1:1 endophytic bacteria at a salt concentration of 50 mmol·L-1 had the best repair effect (P<0.05), which can effectively alleviate the damage of salt stress to soybean seedlings. Patricio[33]showed that inoculation of Enterobacter, Serratia, Achromobacter and Pseudomonas could significantly improve the emergence, growth, biomass and CAT activity of wheat seedlings under salt stress. Rajnish[34] believed that strain SBP-6 (Enterobacter sp.) with ACC deaminase activity significantly improved the growth traits of wheat under salt stress, including plant height, root length, aboveground and underground biomass, and CAT activity. These research results provide strong support for the results of this experiment, but the assistance of strains in improving salt resistance of host plants and promoting plant growth and development is still affected by environmental conditions, and further verification is still needed through field experiments. Upadhyay[35] showed that Bacillus could improve the dry weight of wheat root, the number of sprouts and the activity of SOD in the plant under salt stress to enhance its drought resistance. Lan[36] showed that after diluted 600 times with clean water, Bacillus mycoides could effectively reduce the cracking rate of pomelo, increase the average single fruit weight of pomelo, and improve the tolerance of pomelo to Alkali soil. Wang[37] showed that under salt stress, inoculation of YM6 (Bacillus amyloliquefaciens) strain increased the activities of SOD, POD and CAT in maize plants, which was more conducive to eliminating free radicals in maize plants, reducing the accumulation of oxides, and alleviating salt stress damage. Soybean seedlings grew for 28 days and were inoculated with 137 endophytic bacteria in a 1:2 bacterial suspension. When the salt concentration was 50 and 100 mmol·L-1, the repair effect was better than that of the salt stress group, indicating that endophytic bacteria 137 had a significant (P<0.05) repair effect on soybean seedlings under salt stress. The experimental results are basically consistent with the conclusions of Upadhyay, Lan, and Wang. However, this experiment only studied the effect of endophytic bacteria on antioxidant enzymes in soybean seedlings under salt stress, and further research is needed on the molecular mechanism and other physicochemical indicators of salt tolerance in soybean seedlings.
4. Materials and Methods
4.1. Test Materials and Culture
Soybean variety: Xudou 20 (National audit number 2014012), was developed by Xuzhou Institute of Agricultural Sciences of Jiangsu province, it belongs to common summer soybean varieties and planted in central and old Yellow River basin in eastern Henan Province, southern Shandong Province, north of Huaihe River in Jiangsu Province and north of Huaihe River in Anhui Province. This variety was conformed to the national soybean variety certification standard, purchased from Seed Co., Ltd.in Shangqiu Suiyang District.
Endophytic bacteria 131 and 137 were isolated from soybean root nodules planted in eastern Henan Province of China by the research group, and the strains were stored at 4℃ for a short period in a slanted test tube. Beef-protein medium[38] (g/L): 3 g of beef extract, 10 g of tryptone, 5 g of NaCl, 1000 mL of water, pH 7.0-7.2. LB Culture[39] (g/L): yeast extract 4g, tryptone 10g, NaCl 10g, water 1000mL, pH 7.0-7.2.
4.2. Sequencing and Phylogenetic Analysis
The total genomic DNA was extracted from the culture of nodule isolates by using the previous method[40]. The 16S rRNA gene was amplified from the genomic DNA by PCR with the universal forward primer P1 (5-CGGGAT CCA GAG TTT GAT CCT GGC TCA GAA CGA ACG CT-3) and reverse primer P6 (5-CGGGAT CCT ACGGCT ACC TTG TTA CGA CTT CACCCC-3), respectively, which corresponded to the positions of 8-37 bp and 1479-1506 bp in Escherichia coli 16S rRNA gene[40]. An aliquot of PCR product of isolates was directly sequenced by Sangon Biotech (Shanghai) Co., Ltd. using the same primers mentioned above. Acquired and related sequences were matched with ClustalX1.81 software, imported into Bioedit 4.8.4, and manually corrected. A phylogenetic tree was constructed using the Jukes-Cantor model and the neighbor joining method in TREECON package (version 1.3b). The similarity of each tested strain was computed by using the DNAMAN application (version 6.0.3.40, Lynnon Corporation). The acquired 16S rRNA gene sequences were submitted to NCBI GenBank (http://www.ncbi.nlm.nih.gov/). The GenBank accession numbers of the sequences obtained in this study are KR822280 and KR822263.
4.3. Activation of Endophytic Bacteria and Preparation of Bacterial Suspension
Endophytic bacteria were activated twice on beef extract peptone medium, inoculated into LB liquid medium, and placed in a constant temperature oscillating culture shaker at a speed of 135 r/min at 30℃for 3-4 days. Bacterial suspension was prepared by follow the procedure below. Pour LB liquid culture medium was injected into a centrifuge tube, centrifuged at a speed of 10000 r/min for 10 minutes, collected sediment, and used a spectrophotometer to adjust the OD≈1 of the mother solution at a wavelength of 600 nm (× 106 CFU/mL), dilute the bacterial suspension with sterile water in proportion to 1:2, 1:1, and 2:1.
4.4. Pretreatment and Experimental Treatment of Soybean Seedling Seeds
Select soybean seeds with no crack in seed coat, no mildew, and full seeds, and sub package them in Petri dish. Each dish contains 45 soybean seeds. Wash them with sterile water for 3-4 times. Pipette absorbs 30 mL of sterile water and places them in Petri dish. Incubate them in a constant temperature incubator for 2 hours. Pour out sterile water from Petri dish. Add sterile water and bacterial suspension respectively in CK group and treatment group, and soak them for 1 hour. When soybean sprouts are exposed, they are sown in a pot filled with sterile vermiculite (caliber Φ = 21 cm, 14.9 cm high container), 40 capsules per pot are placed in an artificial intelligence climate incubator under conditions of 25℃ for 10 hours during the day and 20℃ for 14 hours at night.
The experiment was set up with 1 blank control group and 6 salt treatment groups, with 3 replicates for each treatment group. The salt treatment concentrations were 50, 100, 150, 200, 250, and 300 mmol·L-1, respectively. Inoculation and stress treatment: On the first day, 30 mL of sterile water was poured into the CK group, bacterial suspension group, and salt stress group; On the second day, 30 mL of sterile water was poured into the CK group, while 30 mL of NaCl solution was poured into the bacterial suspension group and salt stress group; On the third day, 30 mL of sterile water was poured into the CK group and salt stress group, while 30 mL of bacterial suspension was poured into the bacterial suspension group. This lasted for 3 days for a cycle.
4.5. SOD Activity Detection
The SOD activity was measured by NBT photoreduction method[18]. On the 14th and 28th day of growth, 8 soybean seedlings were randomly selected, and 2g of stems were weighed and added to the SOD reaction solution. They were ground to a homogenate and placed in a centrifuge tube. 150 mL SOD reaction solution was prepared with 0.05 mol·L-1 mixed phosphoric acid buffer solution at pH7.8 as the base solution, containing 5 mL Riboflavin solution, 135 mL Methionine solution, 5 mL EDTA solution, and 5 mL nitrotetrazolium chloride (NBT) solution. The SOD activity was measured at a wavelength of 560 nm. The formula is SOD activity (U·g-1FW)=[(ACK-AE) × V] /(1/2 × ACK × W × Vt), the total activity of SOD is expressed in enzyme units per gram of fresh sample mass (U/g), ACK is the absorbance of the illumination control tube, AE is the absorbance of the sample degree, V is the total volume of the sample solution, Vt is the amount of sample used for measurement, and W is the fresh mass of the leaves.
4.6. CAT Activity Detection
CAT activity was detected by UV absorption method[41]. On the 14th and 28th day of growth, 8 soybean seedlings were randomly selected and 1.5 mL of 0.05 mol·L-1 mixed phosphate buffer with a pH value of 7.8 was added to the leaves. The mixture was ground to a homogenous state, placed in a centrifuge tube, and centrifuged at 12000 r/min for 20 minutes. The supernatant was used as the sample extract. Measure the OD value at a wavelength of 240 nm to calculate CAT activity, and replace it with distilled water. The formula is CAT activity(U·g−1·min−1)= (U·g−1·min−1)=(ΔA240×Vt)/(W×VS×0.1×t) , ΔA240 represents the change in absorbance during the reaction time, W represents the fresh weight of the leaves, t represents the reaction time, and Vt represents the total volume of the extracted enzyme solution.
4.7. Data Processing
Use Excel 2010 to make a Line chart, use SPSS 22.0 to carry out one-way ANOVA, Duncan’s new multiple range method to carry out difference test analysis and least significant difference (LSD) analysis, draw a histogram after data processing, and set the significance level to α= 0.05. Measure the SOD and CAT activities of endophytic bacteria 137 and 131 in soybean seedlings under different salt concentrations of stress.
5. Conclusions
In summary, inoculation of soybean seedlings with endophytic bacteria 131 and 137 under salt stress resulted in varying degrees of changes in SOD and CAT activities, effectively repairing the damage caused by salt stress environment to plants and improving their stress resistance. As the salt concentration increases and the growth cycle prolongs, the antioxidant enzymes in soybean seedlings show an upward trend, and the repair effects of inoculating endophytic bacteria 131 and 137 at each salt concentration are different. Overall, the repair effect of strain 131 is better than that of strain 137.
Author Contributions
Conceptualization and project administration, L. Z.; Data curation, Writing-original draft,Y. X. and X.H.; Formal analysis, Investigation,L.X. ; Writing - review & editing, W.S.; Software (analyzed data), J. Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (U1204301), Natural Science Foundation of Henan Province (182300410069) and Key Specialized Research and Development Program of Henan Province (212102310223).
Data Availability Statement
The sequence data obtained in this study are openly available in NCBI GenBank and the accession numbers are stated in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jiang, Y.; Xue, E.Y.; Lu, W.C.; Cui, G.W.; Li, Y.M.; Han, T.F.; Wang, S.D. Breeding and feeding quality analysis of a new soybean strain deficient in Kunitz tryps in inhibitor. Acta Prataculturae Sinica 2020, 29, 91–98. [Google Scholar]
- De Carvalho Oliveira, R.A.; De Andrade, A.S.; Imparato, D.O.; Lima, J. Dalmolin R. Analysis of Arabidopsis thaliana redox gene network indicates evolutionary expansion of class iii peroxidase in plants. Sci. Rep. 2019, 9, 1–9. [Google Scholar]
- Cha, T.; Zhong, X.B.; Zhou, Q.Z.; He, M.D.; Wang, G.F.; You, J.H.; Wang, Z.Q.; Tang, G.X. Development status of China’s soybean industry and strategies of revitalizing. Soybean Sci. 2018, 37, 458–463. [Google Scholar]
- Khan, A.L.; Hamayun, M.; Kang, S.M.; Kim, Y.H.; Jung, H.Y.; Lee, I.J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: an example of Paecilomyces formosus LHL10. BMC microbiol. 2012, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Li, Y.T.; An, D.D.; Zhang, R. Research progress on effects of endophytes on plant drought resistance. Biotechnol.Bull. 2015, 31, 23–29. [Google Scholar]
- Zarea, M.J.; Karimi, N.; Goltapeh, E.M.; Ghalavand, A. Effect of cropping systems and Arbuscular mycorrhizal fungi on soil microbial activity and root nodule nitrogenase. J. Saudi Soc. Agric. Sci. 2011, 10, 109–120. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar, A.S.; Grover, M. Drought-tolerant plant growth promoting Bacillus spp.: effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Zhao, L.F.; Xv, Y.J.; Lai, X.H.; Chang, J.L.; Ou, Q.F.; Meng, J.Q.; Peng, D.H. Effects of endophytic bacteria 252 and 254 on peroxidase (POD) and catalase (CAT) activities of wheat seedlings under salt stress. Chin. J. Appl. Ecol. 2017, 28, 2984–2992. [Google Scholar]
- Yang, R.P.; Bao, Z.X.; Chen, G.R.; Li, Y. Research Progress on Drought Resistance of soybean seedlings. Crop J. 2018, 28, 8–12. [Google Scholar]
- Qin, Y.; Druzhinina, I.S.; Pan, X.; Yuan, Z. Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef]
- Diao, Q.; Song, Y.; Qi, H. Exogenous spermidine enhances chilling tolerance of tomato (Solanum lycopersicum L.) seedlings via involvement in polyamines metabolism and physiological parameter levels. Acta Physiol. Plant. 2015, 37, 1–15. [Google Scholar] [CrossRef]
- Jiang, X.W.; Li, H.Q.; Wang, J.H. Physiological response of Scutellaria baicalensis seed germination and seedling to exogenous ascorbic acid under salt stress. Plant Physiol. J. 2015, 51, 166–170. [Google Scholar]
- Dinneny, J.R. Traversing organizational scales in plant salt-stress responses. Curr. Opin. plant biol. 2015, 23, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.Y.; Wu, Y.H.; Tian, X.L. Recent advances and issues on the endophyte. Chin. J. Ecol. 2019, 2, 86–91. [Google Scholar]
- Elisa, K.; Hassan, M.; Asaph, A. Plant–microbe interactions in the rhizosphere via a circular metabolic economy. The Plant Cell 2022, 34, 3168–3182. [Google Scholar]
- Farrar, K.; Bryant, D.; Cope-Selby, N. Understanding and engineering beneficial plant-microbe interactions: plant growth promotion in energy crops. Plant biotechnol. J. 2014, 12, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.H.; Feng, R.Z.; Shi, S.L. Mitigation effect of NO on growth inhibition and oxidative damage of alfalfa roots under salt stress. Acta Ecol. Sinica 2015, 35, 3606–3614. [Google Scholar]
- Radhakrishnan, R.; Khan, A.L.; Lee, I.J. Endophytic fungal pre-treatments of seeds alleviates salinity stress effects in soybean plants. J. Microbiol. 2013, 51, 850–857. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Ahmad, N.; Waqas, M.; Kang, S.M.; Kim, Y.H.; Lee, I.J. Exophiala sp. LHL08 reprograms Cucumis sativus to higher growth under abiotic stresses. Physiol. plant. 2011, 143, 329–343. [Google Scholar] [CrossRef]
- Elsey-Quirk, T.; Middleton, B.A.; Proffitt, C.E. Seed flotation and germination of salt marsh plants: the effects of stratification, salinity, and/or inundation regime. Aquat. Bot. 2009, 91, 40–46. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Breusegem, F.V. Reactive oxygen gene network of plants. Trends plant sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Lee, I.J. Endophytic fungi produce gibberellins and indoleacetic acid and promotes host-plant growth during stress. Molecules, 2012, 17, 10754–10773. [Google Scholar] [CrossRef] [PubMed]
- Pereg, L.; McMillan, M. Scoping the potential uses of beneficial microorganisms for increasing productivity in cotton cropping systems. Soil Biol. Biochem. 2015, 80, 349–358. [Google Scholar] [CrossRef]
- Wu, Q.; Feng, H.Q.; Li, H.Y.; et al. Effect of Puccinia striiformis west infection on cyanide-resistant respiration and metabolism of reactive oxygen in wheat, Acta Phytopathol. Sinica 2006, 36, 49–56. [Google Scholar]
- Han, K.; Tian, Z.Y.; Liu, K.; Zhang, J.Y.; Chang, Y.Y.; Guo, Y.Q. Effect of endophytic bacteria with ACC deaminase activity in Kosteletzkya pentacarpos on wheat salt tolerance. Plant Physiol. J. 2015, 51, 212–220. [Google Scholar]
- Zhang, Y.Q. Study on induced resistance of R. rugosa ‘Plena’. Dissertation for the Degree of Master. HaErbin: Northeast Forestry University,2013.
- Khan, A.L.; Waqas, M.; Khan, A.R.; Hussain, J.; Kang, S.M.; Gilani, S.A.; Hamayun, M.; Shin, J.H.; Kamran, M.; Al-Harrasi, A.; Yun, B.W.; Adnan, M.; Lee, I.J. Fungal endophyte Penicillium janthinellum LK5 improves growth of ABA-deficient tomato under salinity. World J. Microbiol. Biotechnol. 2013, 29, 2133–2144. [Google Scholar] [CrossRef]
- Li, F.L.; Guan, L.L.; Hu, F.R. Physiological Response of Different Resistant Cultivars of Lilium Orietal Hybrid after Inoculation with Botrytis cinerea. J. Northeast Forest. Univ. 2020, 48, 107–113. [Google Scholar]
- Zhang, R.; Lv, J.; Mi, Q.S.; Wang, S.G. Effects of salicylic acid on antioxidant enzymes in rice seedlings under chilling stress. J. Southwest Agric. Univ. (Natural Sci.) 2016, 01, 29–32. [Google Scholar]
- Pi, E.; Xu, J.; Li, H.H.; Fan, W.; Zhu, C.M.; Zhang, T.Y.; Jiang, J.C.; He, L.T.; Lu, H.F.; Wang, H.Z.; Poovaiah, B.W.; Du, L.Q. Enhanced salt tolerance of rhizobia-inoculated soybean correlates with decreased phosphorylation of the transcription factor GmMYB183 and altered flavonoid biosynthesis. Mol. Cell.proteomics 2019, 18, 2225–2243. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.R.; Mao, S.X.; Yan, S.Z.; Lu, C.M. Effects of endophytes DLJ1 (Pseudomonas fluorescens)and SZ5(Bacillus cereus)on the resistance, yield and quality of pepper plants under the stress of Meloidogyne incognita. Plant Protection 2020, 46, 96–102. [Google Scholar]
- Saravanakumar, D.; Kavino, M.; Raguchander, T.; Subbian, P.; Samiyappan, R. Plant growth promoting bacteria enhance water stress resistance in green gram plants. Acta physiol. Plant. 2011, 33, 203–209. [Google Scholar] [CrossRef]
- Patricio, J.B.; Nitza, G.I.; Jacquelinne, J.A.; Mora, M.L.; Crowley, D.E.; Jorquera, M.A. Formulation of bacterial consortia from avocado ( Persea americana Mill.) and their effect on growth, biomass and superoxide dismutase activity of wheat seedlings under salt stress. Appl. Soil Ecol. 2016, 102, 80–91. [Google Scholar]
- Singh, R.P.; Jha, P.N. Mitigation of salt stress in wheat plant (Triticum aestivum) by ACC deaminase bacterium Enterobacter sp. SBP-6 isolated from Sorghum bicolor. Acta physiol. Plant. 2016, 38, 110–116. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, D.P. Effect of salt-tolerant plant growth-promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol. 2015, 17, 288–293. [Google Scholar] [CrossRef]
- Lan, Y.Y.; Chen, Y.Y.; Wang, S.F.; Xin, Y.R. Effects of different concentrations of Bacillus mycoides on fruit cracking, yield and quality of Honey pomelo. Mod. Agric. 2017, 11, 30–31. [Google Scholar]
- Wang, H.X.; Liu, H.; Yang, G.P.; Zhang, X.; Li, Z.; Zhang, Y. Effects of Bacillus amyloliquefaciens YM6 on Physiological and Biochemical Characteristics of Maize under Salt Stress. Acta Agric. Boreali-occidentalis Sinica, 2020, 29, 436–443. [Google Scholar]
- Sun, J.G.; Hu, H.Y.; Liu, J.; Chen, Q.; Gao, M.; Xu, J.; Zhou, Y.Q. Growth Promotion Potential and Distribution Features of Nitrogen-Fixing Bacteria in Field Environments. Scientia Agric. Sinica 2012, 45, 1532–1544. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree generated by the Neighbor-Joining method based on 16S rRNA gene sequences of endophytic bacteria.
Figure 1.
Phylogenetic tree generated by the Neighbor-Joining method based on 16S rRNA gene sequences of endophytic bacteria.
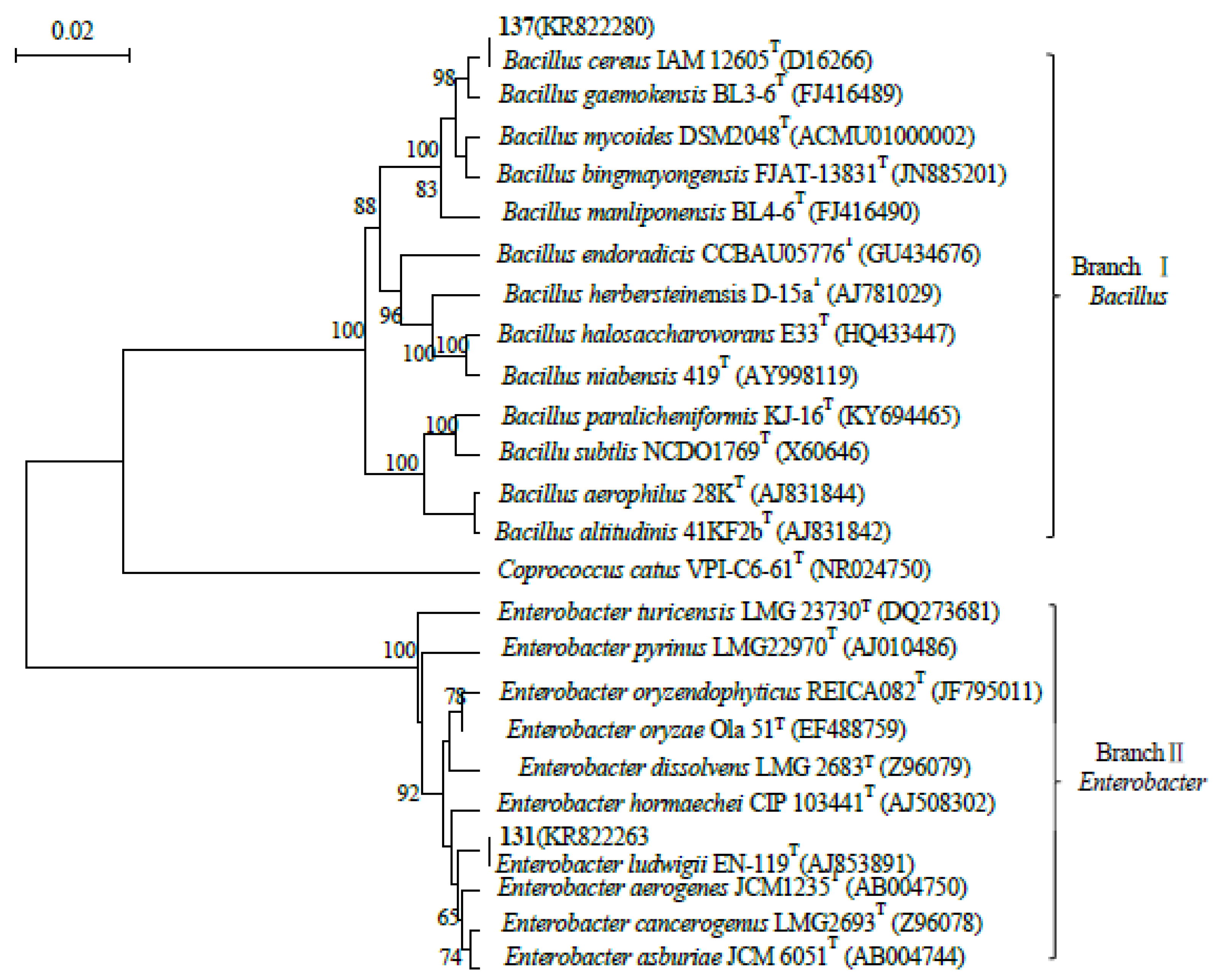
Figure 2.
SOD activities of soybean seedlings inoculated with endophyte 131 for 14 days.
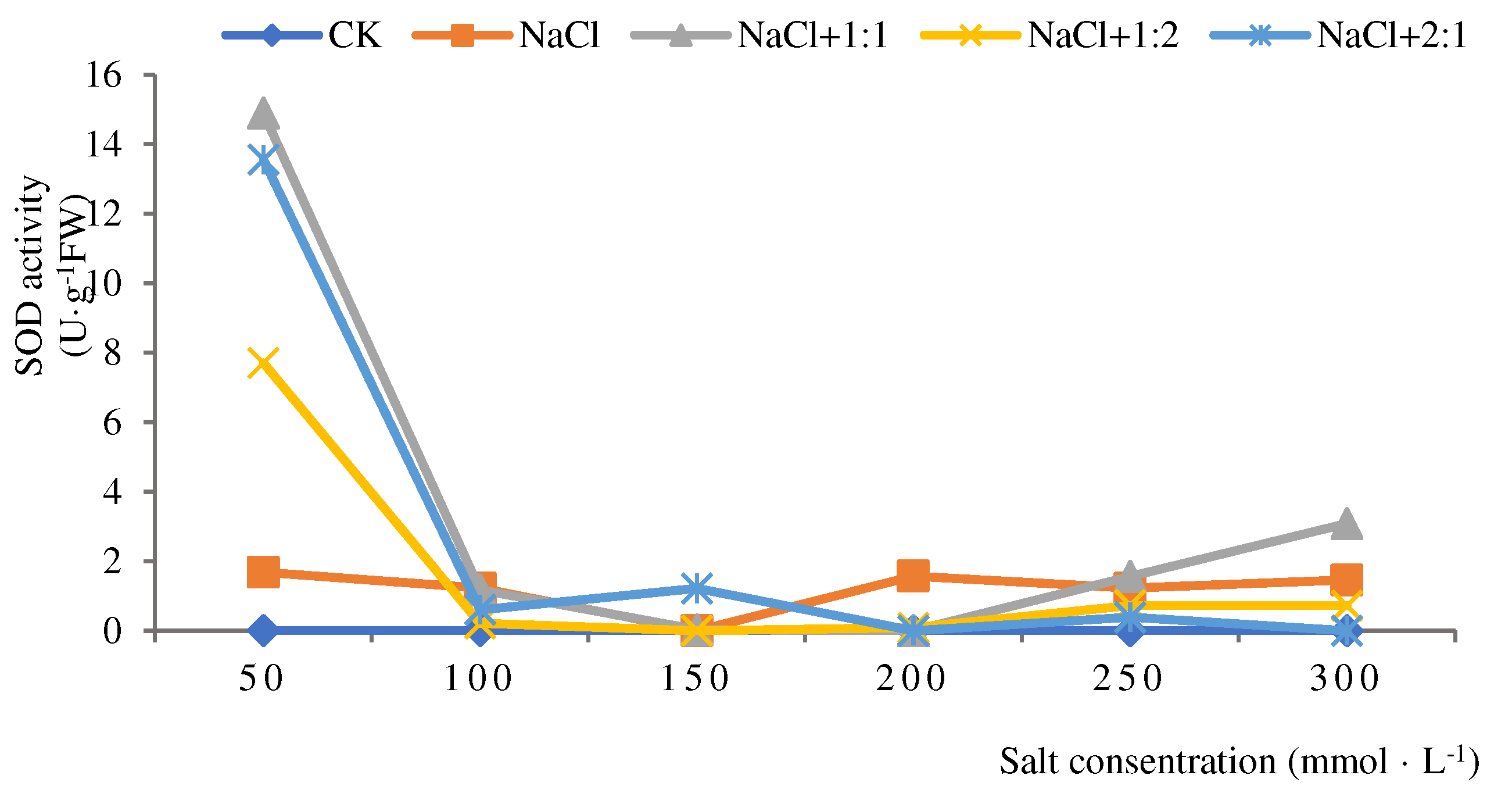
Figure 3.
SOD activities of soybean seedlings inoculated with endophyte 137 for 14 days.
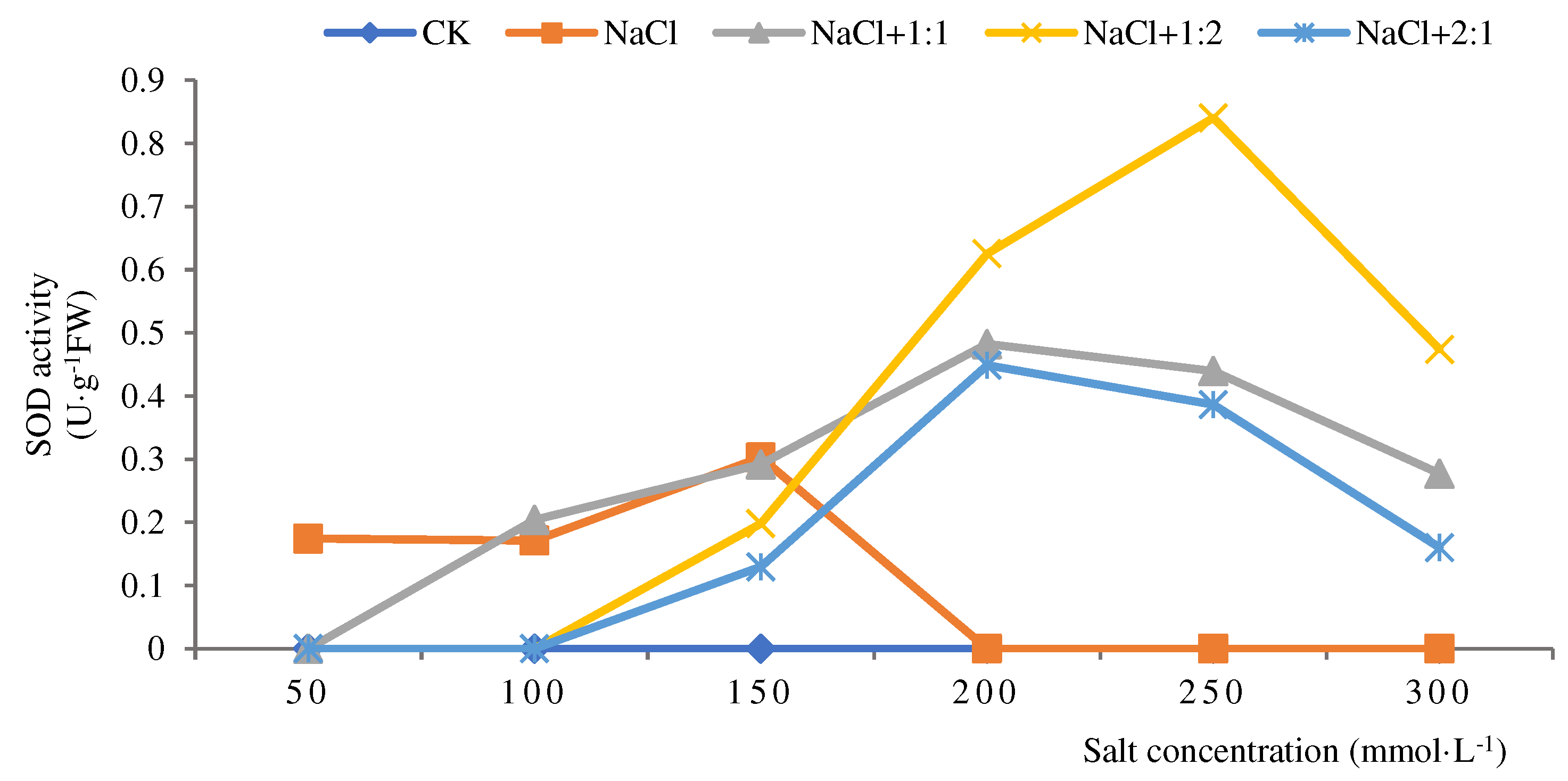
Figure 4.
SOD activities of soybean seedlings inoculated with endophyte 131 for 28 days.
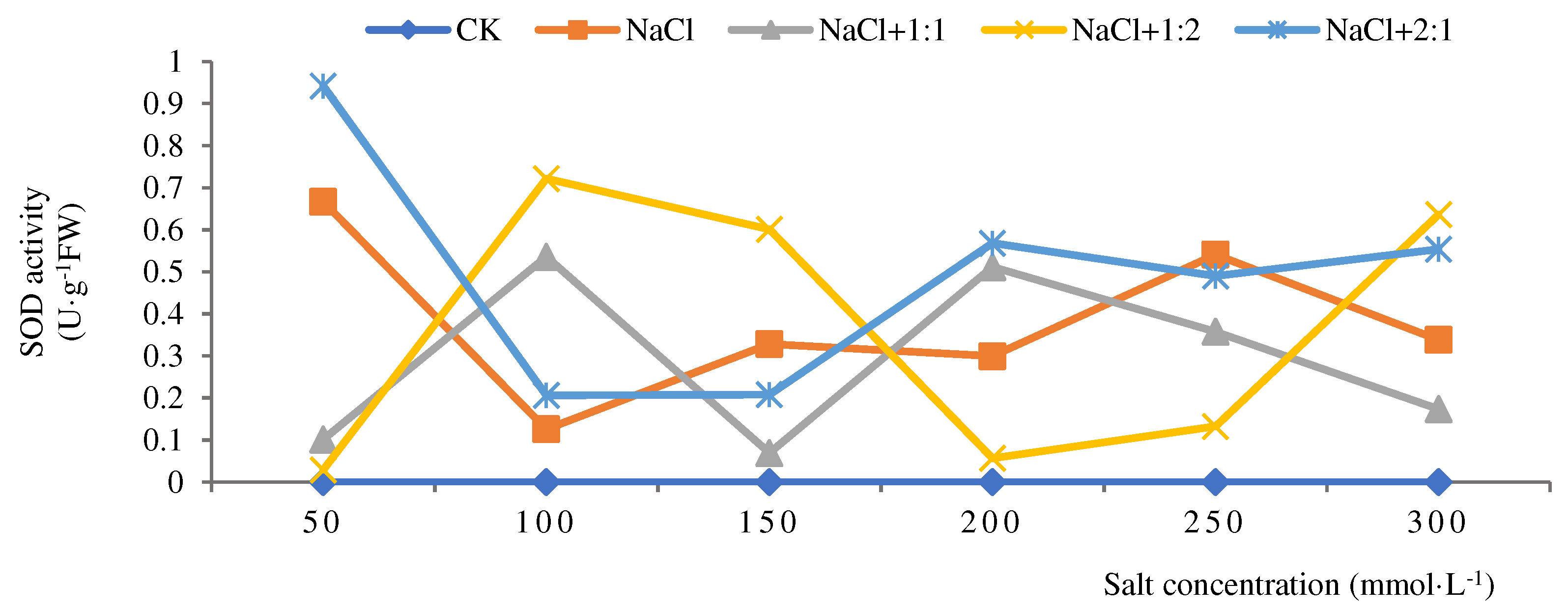
Figure 5.
SOD activities of soybean seedlings inoculated with endophyte 137 for 28 days.
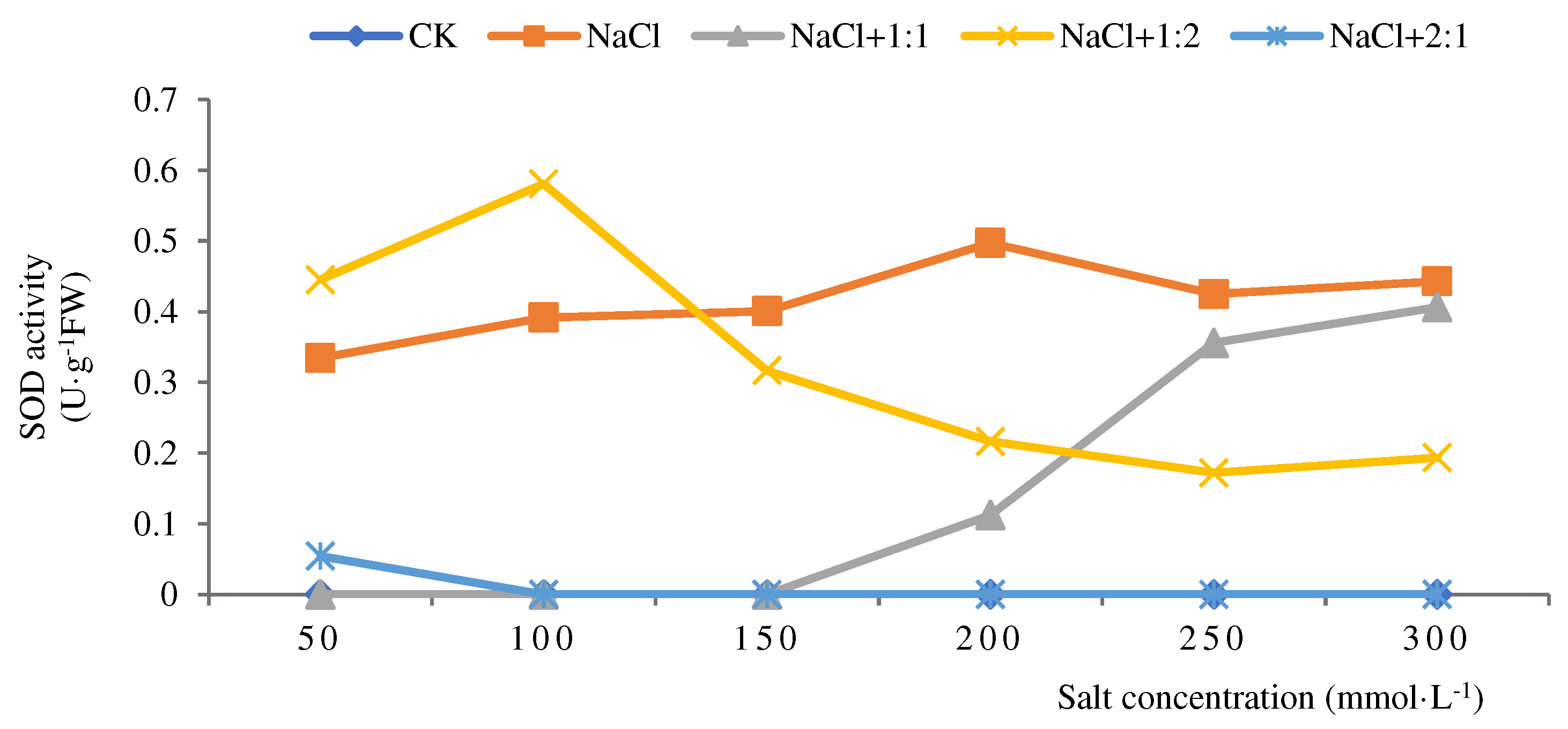
Figure 6.
CAT activities of soybean seedlings inoculated with endophyte 131 for 14 days.
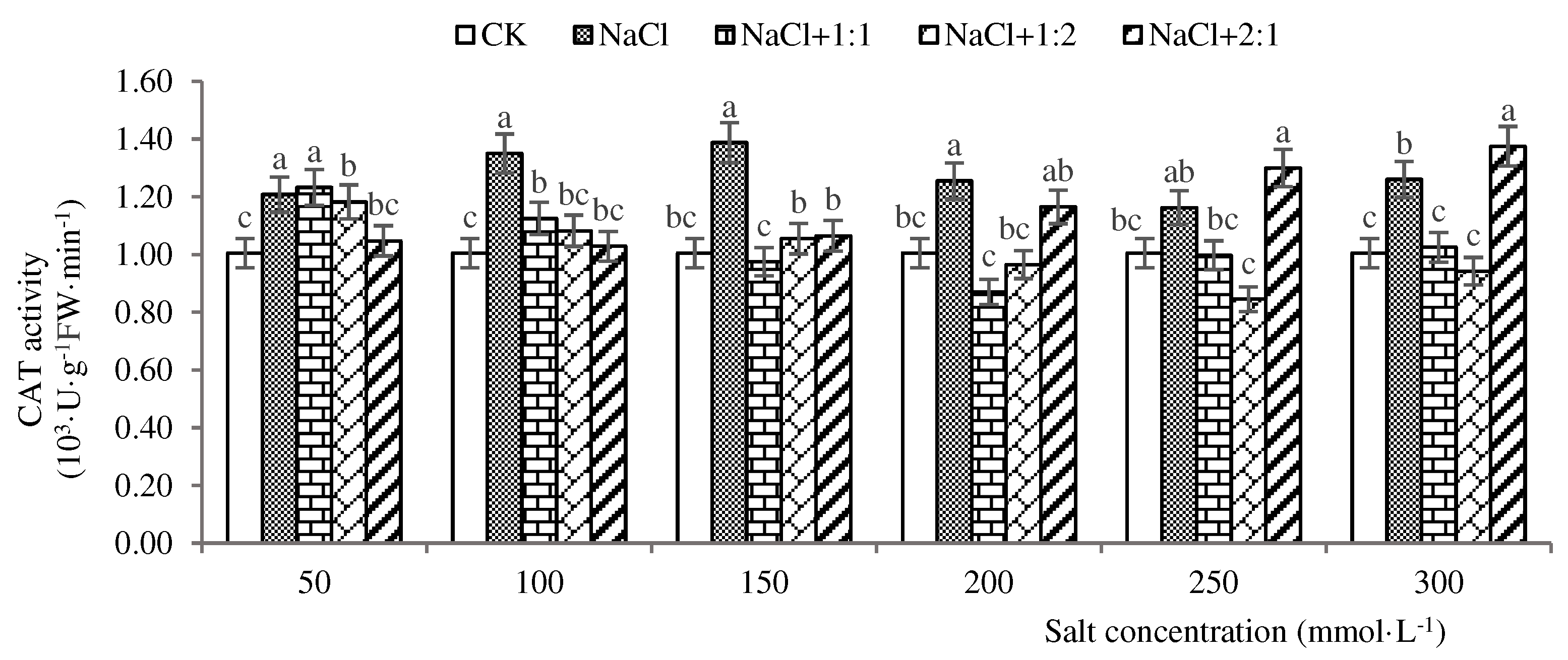
Figure 7.
CAT activities of soybean seedlings inoculated with endophyte 137 for 14 days.

Figure 8.
CAT activities of soybean seedlings inoculated with endophyte 131for 28 days.
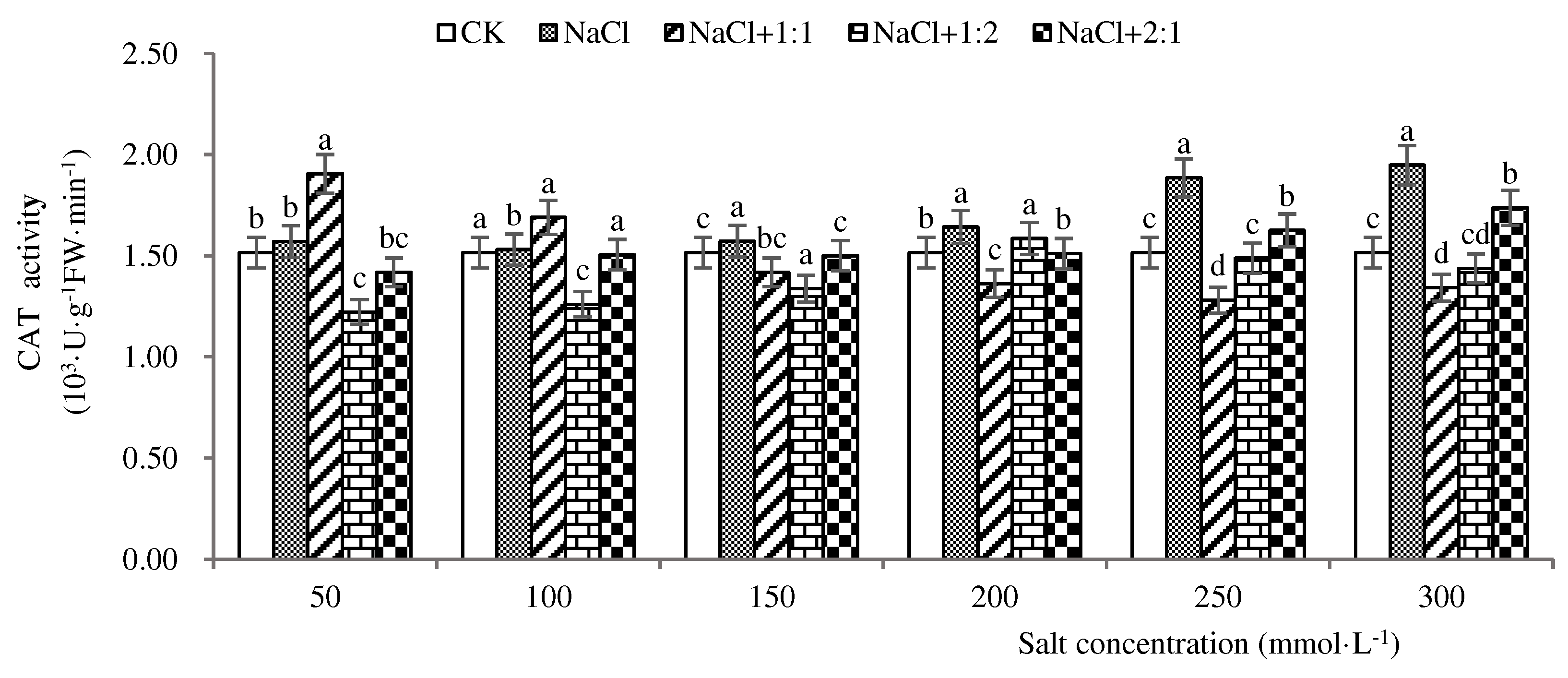
Figure 9.
CAT activities of soybean seedlings inoculated with endophyte 137 for 28 days.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



