
Submitted:
26 October 2023
Posted:
27 October 2023
You are already at the latest version
Abstract
A key response to glucose stress is an increased production of unsaturated fatty acids to balance the increase of saturated fatty acids in the membrane. The C. elegans homolog of stearoyl-CoA desaturase, FAT-7, introduces the first double bond into saturated C18 fatty acids yielding oleic acid and is a critical regulatory point for surviving cold and glucose stress. Here, we incorporated 13C stable isotopes into the diet of nematodes and quantified the 13C-labelled fatty acid by GC-MS and HPLC/MS-MS to track the metabolic response to various concentrations of glucose. Previous work has analyzed membrane composition of C. elegans when responding to mild glucose stress and showed few alterations in overall fatty acid composition in the membrane. Here, in nematodes exposed to higher concentrations of glucose, a specific reduction of oleic acid and linoleic acid was observed. Using time courses and stable isotope tracing, the response of fatty acid me-tabolism to increasing levels of glucose stress has been characterized, revealing a funneling of monounsaturated fatty acids to preserve polyunsaturated fatty acid abundance. Taken together, higher levels of glucose unveil a specific reduction of oleic and linolenic acid in the metabolic re-wiring required to survive glucose stress.
Keywords:
oleic acid
; glucose
; phospholipids
; mass spectrometry
; stable isotopes
; lipidomics
; fatty acid metabolism
1. Introduction
As animals encounter a perturbation in their environment or their diet, there is often a rewiring of metabolic pathways that tunes or adjusts membrane composition to the new conditions [1,2]. In certain circumstances, like temperature changes, a different phospholipid content is established that promotes membrane function with the altered requirements for the membrane [1,3]. There are also situations such as with the addition of moderate glucose stress to the diet where the membrane composition remains stable; however, this stability requires a shift of the metabolic pathways to alter the flux of certain lipid species to the membrane. The PAQR-2 response network positively regulates enzymes including FAT-7 that is essential in the production of unsaturated fatty acids (UFAs) and ELO-5 that is responsible for monomethyl-branched chain fatty acids (mmBCFAs) synthesis. PAQR-2 is a membrane sensor homologous to the mammalian protein AdipoR2 which plays a similar role regulating membrane fluidity in the membrane of HEK 293 cells [4,5,6]. The upregulation of both FAT-7 and ELO-5 is needed for the survival in glucose stress, highlighting the importance of fatty acid metabolism in this response [7,8,9,10].
To promote the correct balance of saturated fatty acids (SFAs) and unsaturated fatty acids (UFAs), fatty acids incorporated from the diet or produced by de novo synthesis must be processed and incorporated into the membrane in the correct ratios [11,12]. In C. elegans, FAT-7 introduces the first double bond to stearic acid (C18:0) producing oleic acid (C18:1n9), which is further elongated and desaturated to produce PUFAs (polyunsaturated fatty acids) including linoleic acid (C18:2n6) and the C20 PUFAs including eicosapentaenoic acid (C20:5n3) [13]. Here, in addition to their common names, a nomenclature of CX:YnZ, where X stands for the number of carbons, Y indicates the number of double bonds and Z defines the location of the first double bond, is used for simplicity. It is important to note that the FA composition of E. coli (OP50), the standard laboratory diet used to feed Caenorhabditis elegans (C. elegans) nematodes, consists primarily of saturated FAs and cyclopropyl FAs [7]. Therefore, to produce and incorporate polyunsaturated fatty acids (PUFAS) in the membrane, FAT-7 is essential for the conversion of C18:0 to C18:1n9 and thus the production of the PUFAs in the nematode [9,14]. Additionally, quantification of new fatty acid incorporation in C. elegans showed that overall lipid production is compromised after FAT-7 RNAi knockdown implicating this enzymatic step in overall fatty acid metabolism12. Furthermore, fat-7 is upregulated upon cold and glucose exposure [7,15]. Together, both findings reinforce the importance of the C18:0 to C18:1n9 conversion in membrane homeostasis.
To promote membrane adaptation, mechanisms to adjust the production and the allocation of different lipid molecules must be present. Techniques using 13C or 15N stable isotopes have been used to evaluate and quantify the dynamics of lipids in the phospholipid (PL) membrane. The evaluation of 13C stable isotope labeling in FAs showed that membrane rejuvenation is rapid in nematodes with most membrane lipids being replaced within 24 hours in young adults [11,12]. Recently, stable isotope studies showed that although the overall composition of the membrane is nearly unchanged with mild (15 mM) glucose stress, there are significant changes to the dynamics of fatty acids like C16:0 and 15-methyl hexadecanoic acid (C17iso) [7]. The altered labeling patterns led to the investigation of the role of monomethyl-branched chain fatty acids in glucose response where it was shown that the production of these fatty acids is essential to survival in glucose, highlighting the utility of probing dynamics along with overall abundance of fatty acids.
Glucose supplementation to the diet reduces the lifespan of nematodes and reduces their resistance to stress. Research has also shown that the gradual elevation in glucose stress has an inverse proportion to the nematode's survival [16,17]. The incorporation of glucose was previously shown to alter overall FA composition in the bacteria, increasing the levels of C16:0 8. We recently found that E. coli growing in high growth (HG) media enriched with glucose had lower levels of vaccenic acid (C18:1n7), suggesting a direct influence in the membrane composition assessed in nematodes [7]. Moreover, fat-7 expression in nematodes was shown to induce a dose dependent response to increased levels of glucose stress [18]. The content of the diet also impacts the regulation and adaptation of membrane metabolism pathways [19]. Many fatty acids present in nematodes are partially obtained intact from the diet; therefore, modification in bacterial composition also influences phospholipid metabolism in nematodes. It is not fully understood the role of bacterial metabolism in the processing of glucose, but studies have found that living bacteria is necessary to see the longevity and healthspan effects on the nematode [20].
Much of what we know about the response to glucose is from studies that compromise the response to glucose via reduction of PAQR-2, FAT-7 or ELO-5. Because of the relationship between glucose stress and membrane adaptation, we hypothesize that higher levels of glucose would reveal other mechanisms not seen previously in wildtype nematodes. Here, we assessed the membrane response to higher concentrations of glucose using GC-MS and HPLC-MS/MS to completely evaluate the response. In doing so, we revealed specific alterations in oleic acid metabolism and describe the requirement for a recovery period to correct the membrane composition.
2. Materials and Methods
2.1. Strains maintenance and population synchronization
All experiments were conducted using wildtype N2 nematodes obtained from the C. elegans Genetics Center (CGC, MN, USA). To synchronize each population of animals, gravid adults were treated with 20% bleach solution, and the recovered eggs were washed at least 3 times and left rotating overnight at 20ºC in M9 solution. OP50 bacteria on HG plates was used in this study unless otherwise stated.
2.2. Stress conditions, labeling strategy, and time-course experiments
The preparation of glucose stress plates (+gluc) followed the protocol that we published previously in Vieira et al., 2022. Briefly, the +gluc plates were made to a final concentration of 15 mM, 100 mM, or 200 mM of glucose by adding a filtered glucose solution to cooled autoclaved HG media. All plates were seeded using regular OP50 bacteria and the +gluc plates were seeded at least 4 days before plating the worms. To start the stress, synchronized L4 stage worms growing on HG plates were transferred to +gluc plates and fed for 12, 24, 48, and 72 hours.
Our labeling strategy followed the protocols established in Perez & Van Gilst, 2008. Briefly, Isogro media (13C) and LB media (12C) were inoculated with OP50 colonies to allow the growth of bacteria for 16 hours at 37ºC. Next, the bacteria were harvested and resuspended in M9 in the concentration of 0.15 g/mL. A mixture containing enriched bacteria 13C:12C (60%:40%) was transferred to agarose plates and allowed to dry. Nematodes from +gluc plates were collected, washed three times using M9, and plated onto stable isotope labeling plates containing 800 uL of bacteria mixture for 6 hours. Labeled worms were removed from the labeling plates, washed, and stored at -80°C until lipid extraction and analysis by mass spectrometry.
Labeling with stable isotopes (13C) allows for the analysis of the percentage of all newly incorporated FA species simultaneously. Briefly, to calculate the MPE the isotopomers were normalized and corrected to the incorporation of natural isotopes. The % Newly Incorporated Fatty Acids considers all newly modified fat independent of its source (de novo synthesized, elongated, or directly absorbed) as described in Dancy et al., 2015 [12]. Error bars of the 13C labeling show the standard error of the mean, and t-tests were used to identify significant differences between the fatty acids. The t-tests were calculated using the GraphPad Prism software version 9.4.1 from July 18, 2022. Samples were considered significantly different if P <0.05 when running an unpaired t-test, using F-test to compare variances.
2.3. Conditions with recovery period after glucose stress
The glucose plates (+gluc) were made as described above to a final concentration of 100 mM of glucose. Synchronized L4 stage worms were transferred to +gluc plates and followed the stress as described. For 12 hours (No Recovery), nematodes spent 12 hours on glucose plates and were collected and washed three times before immediately snap-freezing for further GC/MS analysis. For 12 hours (Recovery), nematodes spent 12 hours on glucose plates and 6 hours of “recovery period” on agarose plates seeded with concentrated OP50 (0.15 mg/mL) to mimic the conditions of the stable isotope labeling plates. After recovery, animals were collected and frozen for further analysis by GC-MS. In 18 hours (No Recovery), nematodes spent 18 hours on glucose plates and were collected and washed three times before immediately snap-freezing for further GC/MS analysis. Statistical analysis used GraphPad Prism software and samples were considered significantly different if P<0.05, marked by the * sign, or if P<0.1, marked by the # sign. The levels of decrease and increase shown in Figure 2C was indicated based on the levels of “P”, where light blue and orange had P values in between 0.05 - 0.005, salmon pink and middle blue had P values between 0.005 - 0.002, and dark blue and red had P values between 0.002 - 0.001.
2.4. Lifespan analysis
To quantify survival on +gluc plates, L4440 bacteria were seeded onto 3 cm NGM +CI plates, and L1 worms were grown for 48 hours. Approximately 50 L4 stage nematodes were then transferred to fresh NGM +CI +gluc plates each day, and the number of dead animals was determined by gently prodding with a pick.
2.5. Heat-killed bacteria tests
Concentrated OP50 E. coli stocks were generated from OP50 stock plates by inoculating 2-3 colonies in each of 4 flasks of 50 mL LB media. These flasks were left to shake overnight (16-18 hours) at 37ºC. Bacteria was pelleted from each flask and resuspended in M9 according to the pellet’s mass at a concentration of 0.15 g/mL. Two of the four resuspended pellets were heat-killed at 65ºC for 20 minutes (HK and HK gluc). All four tubes of living and dead bacteria were centrifuged at 3900 rpm for 10 minutes to re-pellet. The volume of media in each HG plate was considered 30 mL and glucose was added to 100 mM for the +gluc conditions. The two remaining tubes of living and dead bacteria pellets were resuspended in 1X M9 (OP50 and HK OP50).
The heat-killed tubes of 1X M9 and 1X M9 glucose resuspensions were streaked on LB plates and left overnight in a 37ºC incubator to confirm death. All tubes were kept at room temperature for about 12-14 hours until seeded. Before plates were seeded, the tubes holding heat-killed bacteria were placed again in a 65ºC water bath for 20 minutes to ensure no survival of bacteria. Nematodes in the L4 stage were transferred to each plate condition (OP50, OP50 gluc, HK, and HK gluc) right after plates were completely dry and allowed to feed for 12 hours before they were frozen and saved for analysis.
2.6. Lipid Extraction and GC Analysis
Total lipid was extracted using a chloroform:methanol solution (2:1 mixture) and the PL population was separated using chromatography before GC-MS analysis [11,12]. Lipid standards, 1,2-diundecanoyl-sn-glycero-3-phosphocholine (Avanti Polar Lipids) and tritridecanoin (Nu-Chek Prep), were added to each sample, and used as internal control to confirm separation of lipid classes. Dried total lipids resuspended in 1mL of chloroform were loaded onto HyperSep Silica SPE columns (100 mg capacity, Thermo Scientific), and after a sequence of chloroform (3 x 1 mL), acetone:methanol (9:1; 5 x 1 mL) and methanol (3 x 1 mL) was used to each lipid class (i.e., phospholipids, glycolipids, neutral lipids) from the column. Purified PLs were dried and resuspended in 1 mL of 2.5% H2SO4 in methanol, then incubated for 1 hour at 80°C to create fatty acid methyl esters (FAMEs) to run on GC-MS (Thermo Trace 1310, ISQ LT).
The relative % of FAs in each sample was calculated by using the integrated area under the peaks seen in the gas chromatograph and quantified with the Thermo Fisher software Chromeleon version 7.2.10 ES. Error bars of the lipid composition show the standard error of the mean, and t-tests were used to find significant differences between the fatty acids. The t-tests were calculated using the GraphPad Prism software version 9.4.1 from July 18, 2022. Samples were considered significantly different if P <0.05 when running unpaired t-test, also using F-test to compare variances.
2.7. Quantification the synthesis of fatty acids
To determine the percentage of synthesized fatty acids, we followed the protocol published [11,12]. Briefly, using the variation on mass in the parent ion (m+1, m+2, etc.) analyzed by GC-MS, we excluded peaks directly incorporated from the diet and elongated from dietary content. More specifically, the molecular weight of C16:0 can vary from 270 to 286 depending on the number of stable isotopes (13C) that were incorporated into the molecule. Based on the examination of bacterial data and the normalization for the natural incorporation of 13C, peaks in between 270-272 and 284-286 were considered directly incorporated or elongated from the diet. Therefore, to calculate the synthesis of FA, peaks in between 273-283, after normalization, were selected. The percentage calculation takes into consideration the sum of 273-283 peaks over the sum of total peaks 270-286 multiplied by 100%, as represented in the equation: Synthesis % of FA = (Σ273-283)/(Σ270-286) ×100
2.8. Phospholipid extraction and analysis using HPLC-MS/MS
The phospholipid analysis was conducted based on previous studies [12]. Briefly, total phospholipids were extracted from frozen nematodes based on the Folch procedure using 2:1 chloroform: methanol. Extracted lipids were resuspended in 200 μL of acetonitrile/2-propanol/water (65:30:5 v/v/v) and 10 μL was injected into the Dionex UHPLC UltiMate 3000. The phospholipids were separated on a reverse-phase LC column (C18 Hypersil Gold 2.1 x 50 mm, 1.9μm column) at a flow rate of 300 µL/min. The phospholipids were eluted using gradient solvents A and B containing 10 mM ammonium formate (NH4COOH) and 0.1% formic acid (FA). Solvent A was composed of 60/40, water/acetonitrile and solvent B composed of 90/10, isopropyl alcohol/acetonitrile. The schedule for the gradient was 32% B over 0-1.5 min; 32-45% B from 1.5-4 min; 45-52% B from 4-5 min; 52-58% B from 5-8 min; 58-66% B from 8-11min; 66-70% B from 11-14 min; 70-75% B from 14-18 min; 75-97% B from 18-21 min; 97% B up to 25 min; 97-32% B from 25-26 min; 32% B is maintained until 30 min for column equilibration.
Mass spectrometry of PLs was performed on a Q Exactive mass spectrometer (Themo Fisher Scientific). The analysis was performed in the negative ion mode and Full scan data dependent MS2 (ddMS2) mode. The scan range for MS analysis was 300-1200 m/z with maximum injection time of 100 ms and the AGC target 106. The capillary spray voltage was set at 3.2 kV and the capillary temperature was set at 325°C. The sheath gas flow rate was at 45 units and the auxiliary gas flow at 10 units. For MS1 profiling, scans were run at a resolution of 70k. MS2 analyses were performed at NCE of 35 using 6 scan events, with the top five ions chosen from an initial MS1 scan.
Analysis of the LC-MS/MS data was conducted using the software Lipid Data Analyzer (LDA) Version 2.8.1. A 0.1% relative peak cutoff value was applied to the RAW files to focus on the major phospholipid species [21].
3. Results
3.1. High dietary glucose alters the allocation of oleic acid to the membrane.
In nematodes supplemented with low levels of glucose (15mM), the dynamics of specific fatty acids are altered as assayed by stable isotope labeling; however, most fatty acid species do not show significant changes [7]. To further probe the membrane adaptations needed with glucose supplementation, we fed nematodes increasing concentrations of glucose (referred to as +gluc) from 15 mM (mild stress) to 200 mM (high stress) and profiled the dynamics of the fatty acid populations by stable isotope feeding and GC-MS (Figure 1B). Fatty acids were considered new to the membrane if the molecular weight of intact FA was increased in at least one mass unit (MW+1, MW+2, MW+3, etc.) after correction for natural abundance of stable isotopes in the environment and presence of 12C in the labeled OP50 (Isogro media, Sigma Aldrich, St Louis, MO).
At the lowest concentration (15 mM), there was a significant reduction in 13C-incorporation into C16:0 and a significant increase in the 13C-incorporation in C17iso consistent with our previous reports [7]. As the amount of glucose was increased, the levels of newly incorporated C16:0 were reduced from 9.5% ± 0.5 in controls to 7.3% ± 0.1 and 6.6% ± 0.05 in 100 mM and 200 mM, respectively (Figure 1B). There was no further decrease in C16:0 abundance with increasing glucose concentrations. Interestingly, the increase in C17iso was seen only at the two lower concentrations of 15 mM and 100 mM but not at 200 mM (Figure 1B) suggesting that the mmBCFA response may only be needed at lower concentrations of glucose. The stable isotope labeling patterns clearly reveal distinct responses to different concentrations of glucose calling for further investigation of other fatty acid species.
The high concentrations of glucose revealed significant decreases in multiple species that are not compromised with lower glucose supplementation including C18:0, C18:1n9, and C18:2n6. Specifically, C18:0 decreases from 8.8 ± 0.3% in control worms to 8.0 ± 0.1% in 100 mM and 7.7 ± 0.1% in 200 mM; C18:1n9 decreases from 15.0 ± 1.2% in control worms to 12.8 ± 0.6% in 100 mM and 12.8 ± 0.4% in 200 mM; and C18:2n6 decreases from 12.3 ± 0.9% in control worms to 10.2 ± 0.6% in 100 mM and 9.4 ± 0.6% in 200 mM (Figure 1B). The reduction in new C18:1n9 and C18:2n6 suggests that these fatty acid species are not being produced at adequate levels or that any newly synthesized molecules are specifically funneled to maintain highly polyunsaturated species production. Here, the fatty acids are detected using an electron impact (EI) source for the mass spectrometry, and, therefore, the only other PUFA that has sufficient detectable parent ions for analysis is C20:3n6. The amount of newly incorporated C20:3n6 does not change significantly from controls suggesting that C18:1n9 and C18:2n6 are being consumed to funnel resources towards C20 PUFA populations (Figure 1B).
To further support the funneling hypothesis, the total relative abundance of the fatty acid species involved in C20 PUFA production was quantified using GC-MS. The fatty acid profiles confirmed the increase in palmitate (C16:0) as previously seen at all concentrations with this increase being the only significant change seen in the 15mM treated animals in the species measured (Figure 1C). There was a similar increase in C16:0 at all concentrations demonstrating that the increase in C16:0 does not continue to rise with higher concentrations of glucose (Figure 1C). The C16:0 pool is elongated to C18:0 and the levels of C18:0 remain constant at all glucose concentrations showing a specific impact on C16:0 accumulation. C18:0 is converted to C18:1n9 by the FAT-7 desaturase which is implicated in the response to glucose previously [7,8]. Here, there is a significant reduction in overall C18:1n9 levels in only the higher +gluc plates (100 mM and 200 mM). Because the precursors to C18:1n9 are elevated or maintained, we suspected that the C18:1n9 was being consumed to produce the C20 PUFAs needed in the nematode. In fact, there is maintenance in the abundance of all measured PUFAs except for C18:2n6 where we find a trend to reduced levels in high +gluc plates but significance was reached only on 100 mM plates (Figure 1C). Taken together, the stable isotope and GC-MS analysis shows no significant changes in the C20 PUFA population at any glucose concentration suggesting that new dietary carbon is being funneled to maintain these populations.
Elevated glucose in the diet is associated with decreased longevity, and therefore we sought to test whether increasing glucose concentrations would lead to further impacts on lifespan [17]. For each glucose condition, the mean lifespan of the nematode’s is significantly reduced by approximately 30% (Figure 1D). There was no further lifespan reduction as the glucose concentration rose to 100 mM and 200 mM glucose. This is consistent with the fatty acid abundance and dynamics data where we see an impact at lower concentrations but not a correlation with glucose concentration for most fatty acid species. We hypothesize that the metabolic rewiring that occurs at 15 mM is largely sufficient to accommodate the increased glucose levels, but this observation dictates more careful probing of fatty acid dynamics.
3.2. A recovery period is needed to drive the shift in C18:1n9 abundance.
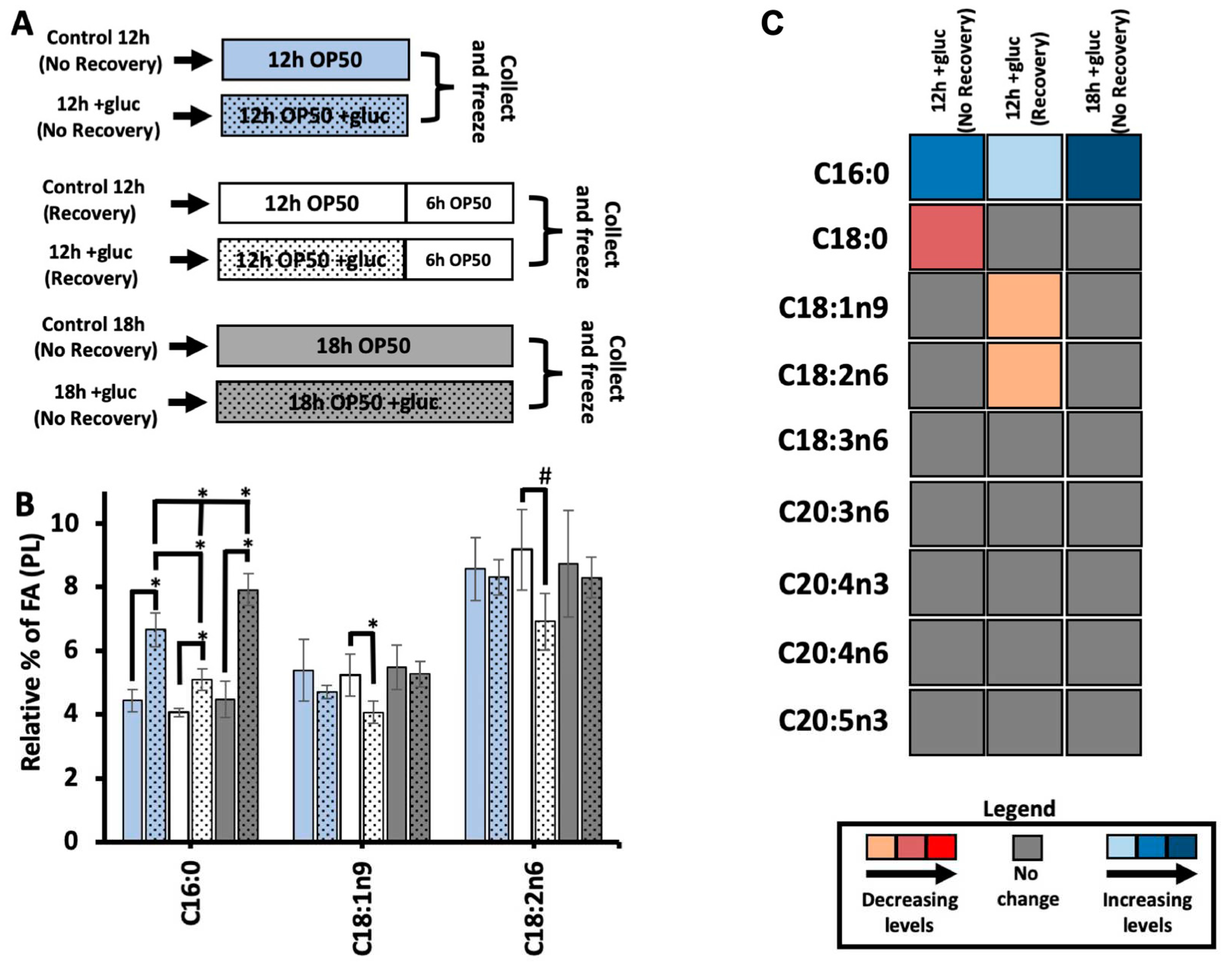
The higher concentrations of glucose revealed altered oleic acid dynamics consistent with the role of the FAT-7 desaturase in surviving glucose stress. Therefore, we further probed the kinetics of fatty acid metabolism with a focus on C18:1n9. First, the stable isotope labeling technique used here introduces 13C-OP50 on agarose media, which is free of nutrients to prevent the labeled bacteria from incorporating 12C from the unlabeled nutrients in the plate. A consequence of this protocol is that the nematodes are given to a “recovery period” of 6 hours on the labeling plates when there is no glucose stress. Because it was previously shown that removing nematodes from a short glucose stress allows for recovery of development in PAQR-2 mutants, we examined the impact of this recovery period of the fatty acids of the membrane [9]. To do so, we used GC-MS to quantify the abundance of saturated and unsaturated fatty acids in nematodes stressed for 12 hours and frozen immediately following the stress (+gluc 12h - No Recovery); and nematodes stressed for 12 hours followed by 6 hours of “recovery period” (+gluc 12h - Recovery). We also included nematodes that were stressed for 18 hours and frozen immediately after stress (+gluc 18h – No Recovery) which would allow us to identify changes that were occurring not due to recovery but to the extra hours on the plates (Figure 2A). To induce the glucose stress, we selected 100 mM of glucose which caused the most significant alterations in membrane dynamics and composition.
We first quantified the abundance of C16:0 in the three conditions and found a significant increase in all glucose treatment groups regardless of the timing and recovery period. The increase in C16:0 with the recovery period was significantly less than in either of the treatment groups without a recovery period (Figure 2A). This trend suggests that the input of C16:0 to the membrane is higher in glucose conditions and that higher input is reduced once glucose is removed from the plate. The amount of C16:0 does continue to significantly rise between 12 hours and 18 hours of glucose feeding supporting the hypothesis that the input of C16:0 is linked to glucose supplementation (Figure 2A). Next, we analyze the relative levels of C18:1n9 in the PL membrane with and without a recovery. If the nematodes are not given to a recovery period, oleic acid levels were not significantly different in 12-hour glucose-stressed animals versus controls (Figure 2B). When given a recovery period, +gluc 12h (Recovery) showed a significant decrease when compared to controls where the level of C18:1n9 decreased from 5.2% ± 0.6 to 4.0% ± 0.3 (Figure 2B). C18:1n9 is converted to C18:2n6 via FAT-2, and we quantified similar trends in this species with a change from 9.1% ± 1.2 to 6.9% ± 0.8 after the recovery period (Figure 2B). This reduction was not a result of the longer time period as these fatty acid species after 18 hours of glucose stress had no change in overall levels (Figure 2B).
The reduction in both C18:1n9 and C18:2n6 abundance after a recovery may show an overall reduction in PUFA production or a specific focus on the production of C20 PUFAs. Therefore, we quantified the relative fatty acid abundances for all major species in the nematode in the three glucose treatment groups versus their respective controls (Figure 2C, Supplementary Figure 1A). We found that there are no significant modifications in the relative abundance of any C20 PUFAs in any stress conditions (Figure 2C). The fatty acids that had significant changes in our previous results remained consistent: increased levels of C16:0 in all treatment groups, decreased C18:1n9 abundance in the 12h plus recovery and decreased C18:2n6 abundance in the 12h plus recovery treatment group (Figure 2C). In addition, there was a small but significant decrease in C18:0 in the 12 h group without recovery. Taken together, our data suggests that the reduction in C18:1n9 and C18:2n6 occurs as these fatty acids are converted to C20 PUFAs in the period following glucose exposure.
Figure 2.
Recovery from glucose is required to observe the shift in C18:1n9. (A) For each comparison, the addition of glucose is represented by the inclusion of black dots vs control with no dots. The blue bars represent nematodes collected immediately after 12 hours of stress (12h; No Recovery), the white bars are nematodes allowed to recover for 6 hours in agarose plates seeded 0.15mg/mL of OP50 (12h: Recovery), and the gray bars are nematodes collected immediately after 18 hours of stress (18h; No Recovery). (B) C16:0 SFA showed significant increase in all glucose stressed animals relative to its controls, and nematodes stressed for a longer period (18h) showed significantly larger accumulation of C16:0 compared to all other two stress conditions. Both C18:1n9 and C18:2n6 maintained stable levels in “No Recovery” animals but showed significant decrease in +gluc 12h (Recovery) compared to its respective control. C18:1n9 decreased from 5.2% ± 0.6 in control 12h (Recovery) to 4% ± 0.3 in +gluc 12h (Recovery), and C18:2n6 decreased from 9.1% ± 1.2 in control 12h (Recovery) to 6.9% ± 0.8 in +gluc 12h (Recovery). (C) A heat map shows the alteration in FA levels comparing stressed animals to controls (+gluc/controls). Significant decreases are shown in light orange (little), dark orange (intermediate), and red (substantial). Significant increases are shown in light blue (little), median blue (intermediate), and dark blue (substantial). Gray bars indicate FAs that did not have significant alteration. For all GC-MS analysis, values represent means ± SEM of at least 9 replicates. Statistical significance, P<0.05 indicated by * and P<0.1 is indicated by #, was calculated using unpaired T tests and F tests to compare variances. .
Figure 2.
Recovery from glucose is required to observe the shift in C18:1n9. (A) For each comparison, the addition of glucose is represented by the inclusion of black dots vs control with no dots. The blue bars represent nematodes collected immediately after 12 hours of stress (12h; No Recovery), the white bars are nematodes allowed to recover for 6 hours in agarose plates seeded 0.15mg/mL of OP50 (12h: Recovery), and the gray bars are nematodes collected immediately after 18 hours of stress (18h; No Recovery). (B) C16:0 SFA showed significant increase in all glucose stressed animals relative to its controls, and nematodes stressed for a longer period (18h) showed significantly larger accumulation of C16:0 compared to all other two stress conditions. Both C18:1n9 and C18:2n6 maintained stable levels in “No Recovery” animals but showed significant decrease in +gluc 12h (Recovery) compared to its respective control. C18:1n9 decreased from 5.2% ± 0.6 in control 12h (Recovery) to 4% ± 0.3 in +gluc 12h (Recovery), and C18:2n6 decreased from 9.1% ± 1.2 in control 12h (Recovery) to 6.9% ± 0.8 in +gluc 12h (Recovery). (C) A heat map shows the alteration in FA levels comparing stressed animals to controls (+gluc/controls). Significant decreases are shown in light orange (little), dark orange (intermediate), and red (substantial). Significant increases are shown in light blue (little), median blue (intermediate), and dark blue (substantial). Gray bars indicate FAs that did not have significant alteration. For all GC-MS analysis, values represent means ± SEM of at least 9 replicates. Statistical significance, P<0.05 indicated by * and P<0.1 is indicated by #, was calculated using unpaired T tests and F tests to compare variances. .
3.3. The abundance of oleate and linoleate stabilizes with longer glucose exposure.
Once we found that the recovery period was essential to see the alterations in C18:1n9 metabolism, we next tested the impact of longer durations of glucose stress. Nematodes were subjected to 100 mM glucose for 12 hours, 24 hours, 48 hours and 72 hours, and all treatment groups had a 6-hour recovery period to elicit the reduction in C18:1n9 and in C18:2n6 as well as to allow for stable isotope labeling. First, C16:0 abundance was considered, and, with all durations of glucose stress, there was a significant increase in C16:0 (Figure 3A). Interestingly, the longer periods of glucose stress did not lead to further increases in C16:0 abundance. Because we saw an increase in C16:0 with 18 hours compared to 12 hours, we believe that the recovery period along with metabolic rewiring can compensate for the longer exposure to glucose.
We next quantified the abundance of C18:1n9 with different lengths of 100 mM exposure. Here, we found a decrease of C18:1n9 from 4.4% ± 0.1 in control 12h to 3.9 ± 0.1 in +gluc 12h, and a greater decrease from 5.6% ± 0.6 in control 24h to 4.3 ± 0.2 in +gluc 24h. However, the 48h and the 72h treatment did not lead to significant changes in C18:1n9 (Figure 3A). To interpret this data, we considered the oleic acid populations in the control populations which revealed that the baseline C18:1n9 levels increased in the 24h and the 48h controls (Figure 3A). The longer glucose exposure dictates that the lipid populations are being examined in older animals, and, although 24 h is a brief period, the first three days of adulthood are the peak reproductive period within the nematodes and associated with glucose-independent metabolic changes [22,23]. Despite the impact of aging, this data is consistent with an upregulation of FAT-7 that requires time to stabilize the higher levels of the enzyme.
To further understand the impact of the altered C18:1n9 levels on fatty acid elongation and desaturation pathway, we next examined C18:2n6, the immediate product of C18:1n9 desaturation (Figure 1A). The trends in C18:2n6 were similar to C18:1n9 with significantly decreased abundance from 10.8 ± 1.0% in control 24h to 7.5 ± 0.3 in +gluc 24h (Figure 3A). Like C18:1n9, there is no significant change with 48 hours of glucose feeding. However, the 72-hour analysis revealed significantly higher levels of C18:2n6 in control animals compared to the young 12 h stressed controls and a significant decrease in C18:2n6 with 72h glucose exposure (Figure 3A). Next, we examined if the reduction in C18:2n6 and C18:1n9 affected the abundance of the C20 PUFAs downstream. To do so, we examined the levels of the major C20 PUFAs with 24 hours glucose stress as that treatment had the greatest impact on the precursor populations (see Supplementary Figure 1B for all treatment durations). For C20:3n6, C20:4n3 and C20:5n3, there was some small but significant change in these pools; however, the extent of these changes was relatively minor. In fact, the C20:4n3 levels increased following 24 hours of glucose (Figure 3B), supporting the hypothesis that the C18:1n9 produced by FAT-7 upregulation is funneled to preserve C20 PUFAs (Figure 3B).
In addition to monitoring the overall abundance of these fatty acid pools, we implemented a stable isotope labeling strategy to determine the flux in these populations. Consistent, with the 12-hour data, the amount of newly incorporated or isotopically labeled C16:0 was reduced with all durations of 100 mM glucose exposure (Figure 3C). In both C18:1n9 and C18:2n6, there was a significant reduction in the number of 13C-fatty acids at 12 hours as seen previously. There are no significant changes in the labeling of C18:1n9 at 24 or 48 hours despite reducing C18:1n9 abundance at 24 hours. This data suggests that FAT-7 levels reach their peak within 24 hours and the turnover of the fatty acids requires 48 hours for the fatty acid pool to stabilize. A similar trend is seen with C18:2n6 but there are significant differences at 48 hours after labeling. We hypothesize that the upregulation of FAT-3, the enzyme that drives the conversion between C18:2n6 and C18:3n6 has slower kinetics.
Finally, each of the fatty acid pools examined had a dramatic reduction of labeling at 72 hours in the control nematodes. This reduction likely indicates an alteration in phospholipid metabolism as the animals leave the reproductive period, but it also uncovers the relative production of these fatty acids. New C16:0 production continues to be reduced (Figure 3C), consistent with the fact that the overall levels of C16:0 are still elevated at 72 hours (Figure 3A). Both C18:1n9 and C18:2n6 have significant increases in new fatty acids compared to the unstressed controls (Figure 3C). The increases in these populations likely reflect the stabilization or activation of FAT-7 and FAT-3.
The changes in fatty acid abundance can ultimately reflect increased production, increased dietary absorption or decreased consumption. The stable isotope strategy we used here lets us define the origin of the oleate pool as altered with glucose supplementation. Under basal conditions, 11 ± 0.3% C18:1n9 is derived from de novo fatty acid synthesis consistent with past reports11. The contribution of synthesis to C18:1n9 production falls to 9.2 ± 0.2% in +gluc 12h (Figure 3D). The amount of synthesis is not statistically different in any of the longer treatment periods; however, it is interesting to note that the amount of synthesized fatty acids increases in 24 hours and 48 hours compared to 12 hours in the controls (Figure 3D). The trends in synthesized C18:1n9 closely mirror the overall amount of stable isotope incorporation suggesting that the processing of synthesized fatty acids is the driver of those dynamics. This is consistent with the composition of the bacterial diet which has few overall changes in fatty acids [7].
Because C16:0 is the main product of de novo fatty acid synthesis, we quantified the amount of synthesized palmitate in the time course. Similar to C18:1n9, there is a decrease in synthesized C16:0 at 12 hours and no significant changes at other timepoints (Figure 3D). We believe that this quantification shows that synthesis is initially reduced to prevent further production of C16:0. This also shows that the elevated levels of C16:0 are not derived from using the excess glucose to drive fatty acid synthesis in the nematode. Furthermore, there is an increase in synthesized C16:0 in 24 hour and 48 hours populations reinforcing the altered fatty acid metabolism over the first few days of adulthood. Unlike the C18:1n9, the pattern of synthesized C16:0 does not match the overall labeling implicating fatty acid absorption as a key contributor to palmitate metabolism.
3.4. Living bacteria is needed for the impact of glucose stress on the membrane.
The addition to glucose to the diet may induce excess saturated fatty acid production or alter membrane metabolism through another mechanism. For instance, it has been seen that glucose can cause oxidative stress through increased production of advanced glycation agents[20]. Because the bacterial food source (OP50) is present during glucose exposure, the bacteria could theoretically metabolize glucose and contribute to the effects of glucose stress. To test the impact of bacteria processing glucose on the membrane composition, the OP50 bacteria was grown in LB broth media, and then bacteria were separated into two groups: living and heat killed bacteria (scheme shown in Figure 4A). Living bacteria was resuspended into fresh LB (OP50) for the control treatment and LB media containing 100 mM glucose (OP50 +gluc) for the stress. In the other treatment group, bacteria were heat killed at 65 °C for 20 mins and resuspended into fresh LB media (OP50 HK (Heat Killed)) and LB media containing 100 mM glucose (OP50 HK + gluc).
To determine the impact of heat killing on the bacteria food source, we analyzed the fatty acid composition of the bacteria using GC-MS. Notably, we found that killing the bacteria significantly increased the level of saturated fatty acids, C12:0, C14:0 and C16:0, even in the absence of glucose (Figure 4B). Specifically, the level of C16:0, the most abundant saturated fatty acid in OP50, increased from 33.7 ± 1.0% of the dietary fatty acids to 52 ± 4.5% in heat killed OP50. Additionally, there was a corresponding decrease in the level of unsaturated fatty acids particularly C18:1n7 which decreased from 28.4 ± 4.5% in OP50 to 6.7 ± 2.1% in killed OP50. We next examined the impact of glucose in both populations and found that glucose did not affect the composition of living bacteria. There was a small but not significant increase in C16:0 which suggests that the accumulation of C16:0 in the nematodes is not a result of increased dietary C16:0. There was a small but significant decrease in C14:0 glucose in the killed OP50 with glucose, but the GC-MS analysis confirmed that the glucose did not have a major impact on the fatty acid composition of the killed bacterial populations (Figure 4B).
To assess the impact of bacteria processing glucose on the lipid composition of the nematodes, the phospholipid composition was analyzed using HPLC/MS-MS in animals fed living or killed bacteria. Here, we focused on the most abundant phospholipids of the membrane, phosphatidylcholine (PC) and phosphatidylethanolamine (PE) (see supplementary table 1 for full list). First, we considered the total number of double bonds present in the two fatty acid tails (i.e., 0-1, 2-3, 4-5, 6-7 and 8 or more) (Figure 4C) and the chain length (Figure 4D) of the PC populations. Feeding the animals with live OP50 +gluc led to a significant decrease in the level of species with 2-3 double bonds and an increase in species with 4-5 double bonds (Figure 4C). The decrease in lipid species with 2-3 double bonds is consistent with GC-MS data as the fatty acids C18:1n9 and C18:2n6 are mostly present in this population. Animals that were fed a heat killed OP50 +gluc did not have any significant change in their degree of unsaturation, suggesting that the living bacteria is needed for processing glucose. Except for a reduction in the populations with 1 double bond, there was no significant difference between the PC profiles in living versus killed bacteria without glucose treatment (Figure 4C). The phospholipid was also binned according to their chain length (total number of carbon present) as the length of phospholipids can influence the biophysical property of the membrane such as thickness. However, there were no significant changes in the chain length of PC lipids when the worms were fed either living bacteria or heat killed bacteria (Figure 4D).
In analyzing the double bond distribution of the PE population, the animals were fed OP50 + gluc diet only had a significant increase in phospholipids with 4-5 double bonds (Figure 4E) and no changes in the distribution of chain lengths (Figure 4F). Again, there were no significant alterations in the level of unsaturation or chain length when the nematodes were fed a HK OP50 +gluc diet versus the killed diet without glucose supplementation (Figure 4E-F). Notably, there were significant changes in the chain length distribution of OP50 compared with OP50 HK which makes it difficult to disentangle the impact of the different dietary source from the requirement for living bacteria to process the glucose. However, these results indicate that living bacteria may have to process glucose to drive the changes in the membrane lipid composition.
4. Discussion
Here, we used increasing concentrations of glucose along with different treatment windows to map the metabolic responses to glucose stress in wildtype C. elegans. In doing so, we confirmed that a main feature to glucose stress response is in regulating the saturation balance in the PL membrane [8,9]. Although regulation of membrane saturation is largely influenced by the activity of FAT-7, most of the data testing membrane composition used mild concentrations (15 to 20 mM) of glucose [7,9]. These concentrations were enough to show accumulation of SFA in healthy nematodes, but species like C18:1n9 and C18:2n6 were maintained stable levels [8]. At the higher concentrations used here, the abundance and the dynamics of C18:1n9 and C18:2n6 were compromised. However, there was no reduction in the fatty acids downstream of C18:2n6 indicating that C20 PUFA synthesis is prioritized. Each double bond is responsible to create “kinks” in the FA structure disrupting the tight packing of more linear molecules such as saturated fat. Previous fluorescence studies in cell culture have shown that the incorporation of fatty acids with four or more double bonds such as C20:4n6, C20:5n3, and C22:6n3 increases membrane fluidity more than MUFAs [24]. We hypothesize that the decreased levels in all precursor FAs and the strict maintenance of PUFAs is a funneling process towards the maintenance of highly unsaturated fat to control membrane fluidity.
Higher concentrations of glucose have been used to stress nematodes and evaluate the role of glucose in health span, stress response and lifespan. In fact, here, we have defined 100 mM and 200 mM as high glucose condiftions, but other studies have used even higher concentrations [25,26]. However, it is important to consider that the concentrations here are supraphysiological and may not be reasonable models of glucose stress. The utility of these higher concentrations is to tax the nematode’s glucose response network including PAQR-2 to reveal the changes in dynamics that may be occurring at lower concentrations particularly in certain cell types or in certain organelles. It is striking to see how little the FA and the PL composition change with substantial amounts of exogenous glucose. The limited alterations in the membrane highlight the flexibility of the metabolic pathways to process a variety of diets and to support the composition of the lipid pool regardless of the input in wild-type animals.
Although we can map the response to elevated glucose in the diet, it is not clear how the excess glucose is driving those changes. Here, we find that this response requires living bacteria as we do not see the same changes with heat-killed bacteria. The role of living bacteria may be in ensuring that the glucose is taken up by the nematodes or in processing the glucose to a metabolite. Although others have hypothesized that the bacteria convert the glucose to SFA, we see a very modest increase in C16:0 with our experimental conditions. Because we do see an impact on the dynamics and the longevity of animals in our study, we believe that the extra synthesis of C16:0 by the bacteria does not account for the entirety of the increased C16:0 in the nematode. Furthermore, we find that there is a decrease in de novo fatty acid synthesis within the worm. Taken together, we hypothesize that elevated levels of glucose stress may have an added component. Several groups have associated glucose accumulation with the increase of oxidative stress ultimately affecting cell migration and promoting inflammation in human tissues[27,28,29,30,31]. In the membrane, the FAs form a major oxidation target, resulting in the production of peroxidation products [32]. This happens because the double bonds are particularly susceptible to reactive oxygen species attack allowing for hydrogen abstraction from the carbon atoms [14]. In our experiments, we substantially increased glucose concentration, which could have raised the levels of oxidative damage in the PUFAs. Thus, it is possible that the maintenance of PUFAs is an effort to correctly replace damaged lipids avoiding membrane dysregulation and consequently apoptosis. In support of this model, the decrease in lifespan seen in nematodes exposed to living bacteria fed glucose, does not happen in dead bacteria supplemented with glucose [26].
To better understand the importance of C18:1n9 and C18:2n6 in the overall response to elevated levels of glucose, we proposed to test the effectiveness of recovery mechanisms in restoring membrane composition after removal from stress source. It was previously shown that removing PAQR-2 mutants from glucose stress allows for development to adulthood if the stress does not last longer than 12 hours [9]. However, little is known about the influence of these mechanisms in the membrane adaptation to the recovery. In our experiments, the labeling techniques give the nematodes a 6-hour recovery period after stress, which becomes an opportunity to further understand the process of membrane recovery. When no recovery period is present, C16:0 continues to accumulate suggesting an increased input of SFA to the animal that overwhelms the fatty acid elongation and desaturation pathway. When glucose is removed, the C16:0 input is reduced allowing the downstream enzymes to catch up on processing these fatty acids and the decrease seen in C18:1n9 and C18:2n6 appears. We hypothesize that the reduced processing may be a result of overall slowed membrane turnover when there is an imbalance of SFA:UFA in the animal.
5. Conclusions
In conclusion, our findings indicate that the regulation of oleic and linoleic fatty acids is essential to respond against high concentrations of glucose consistent with the key role of FAT-7 in this response. In wild-type nematodes, upregulation of fatty acid processing enzymes likely through PAQR-2 eventually eliminates the lag in oleic acid processing. We believe that this study highlights the utility of stable isotope labeling in C. elegans. In fact, the labeling time course also revealed altered fatty acid synthesis and turnover as the control nematodes with no glucose exposure went through the first four days of adulthood. This observation shows the need for stable isotope studies over the entire C. elegans lifespan.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Fatty Acid composition from recovery from glucose stress; Table S1: Phospholipidomic profile of C. elegans fed OP50 with or without 100 mM glucose.
References
- Chen, Y.L.; Tao, J.; Zhao, P.J.; Tang, W.; Xu, J.P.; Zhang, K.Q.; Zou, C.G. Adiponectin Receptor PAQR-2 Signaling Senses Low Temperature to Promote C. Elegans Longevity by Regulating Autophagy. Nature Communications 2019 10:1 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Svensk, E.; Ståhlman, M.; Andersson, C.H.; Johansson, M.; Borén, J.; Pilon, M. PAQR-2 Regulates Fatty Acid Desaturation during Cold Adaptation in C. Elegans. PLoS Genet 2013, 9. [Google Scholar] [CrossRef]
- Farkas, T.; Fodor, E.; Kitajka, K.; Halver, J.E. Response of Fish Membranes to Environmental Temperature. Aquac Res 2001, 32, 645–655. [Google Scholar] [CrossRef]
- Ruiz, M.; Ståhlman, M.; Borén, J.; Pilon, M. AdipoR1 and AdipoR2 Maintain Membrane Fluidity in Most Human Cell Types and Independently of Adiponectin. J Lipid Res 2019, 60, 995–1004. [Google Scholar] [CrossRef]
- Pilon, M. Paradigm Shift: The Primary Function of the “Adiponectin Receptors” Is to Regulate Cell Membrane Composition. Lipids in Health and Disease 2021 20:1 2021, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sheng, W.; Sun, G.Y.; Lee, J.C.M. Effects of Fatty Acid Unsaturation Numbers on Membrane Fluidity and α-Secretase-Dependent Amyloid Precursor Protein Processing. Neurochem Int 2011, 58, 321–329. [Google Scholar] [CrossRef]
- Vieira, A.F.C.; Xatse, M.A.; Tifeki, H.; Diot, C.; Walhout, A.J.M.; Olsen, C.P. Monomethyl Branched-Chain Fatty Acids Are Critical for Caenorhabitis Elegans Survival in Elevated Glucose Conditions. Journal of Biological Chemistry 2022, 298, 101444. [Google Scholar] [CrossRef]
- Svensk, E.; Devkota, R.; Ståhlman, M.; Ranji, P.; Rauthan, M.; Magnusson, F.; Hammarsten, S.; Johansson, M.; Borén, J.; Pilon, M. Caenorhabditis Elegans PAQR-2 and IGLR-2 Protect against Glucose Toxicity by Modulating Membrane Lipid Composition. PLoS Genet 2016, 12, 1–24. [Google Scholar] [CrossRef]
- Devkota, R.; Svensk, E.; Ruiz, M.; Ståhlman, M.; Borén, J.; Pilon, M. The Adiponectin Receptor AdipoR2 and Its Caenorhabditis Elegans Homolog PAQR-2 Prevent Membrane Rigidification by Exogenous Saturated Fatty Acids. PLoS Genet 2017, 13, 1–19. [Google Scholar] [CrossRef]
- Zhu, H.; Shen, H.; Sewell, A.K.; Kniazeva, M.; Han, M. A Novel Sphingolipid-TORC1 Pathway Critically Promotes Postembryonic Development in Caenorhabditis Elegans. 2013, 1–19. [CrossRef]
- Perez, C.L.; Van Gilst, M.R. A 13C Isotope Labeling Strategy Reveals the Influence of Insulin Signaling on Lipogenesis in C. Elegans. Cell Metab 2008, 8, 266–274. [Google Scholar] [CrossRef]
- Dancy, B.C.R.; Chen, S.W.; Drechsler, R.; Gafken, P.R.; Olsen, C.P. 13C- and 15N-Labeling Strategies Combined with Mass Spectrometry Comprehensively Quantify Phospholipid Dynamics in C. Elegans. PLoS One 2015, 10, 1–23. [Google Scholar] [CrossRef]
- Watts, J.L. Using Caenorhabditis Elegans to Uncover Conserved Functions of Omega-3 and Omega-6 Fatty Acids. J Clin Med 2016, 5. [Google Scholar] [CrossRef]
- Watts, J.L.; Browse, J. Genetic Dissection of Polyunsaturated Fatty Acid Synthesis in Caenorhabditis Elegans. Proc Natl Acad Sci U S A 2002, 99, 5854–5859. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.J.; Browse, J.; Watts, J.L. Fatty Acid Desaturation and the Regulation of Adiposity in Caenorhabditis Elegans. Genetics 2007, 176, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Alcántar-Fernández, J.; Navarro, R.E.; Salazar-Martínez, A.M.; Pérez-Andrade, M.E.; Miranda-Ríos, J. Caenorhabditis Elegans Respond to High-Glucose Diets through a Network of Stress-Responsive Transcription Factors. PLoS One 2018, 13, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Jeong, D.E.; Son, H.G.; Yamaoka, Y.; Kim, H.; Seo, K.; Khan, A.A.; Roh, T.Y.; Moon, D.W.; Lee, Y.; et al. SREBP and MDT-15 Protect C. Elegans from Glucose-Induced Accelerated Aging by Preventing Accumulation of Saturated Fat. Genes Dev 2015, 29, 2490–2503. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Horikawa, M.; Shimamura, S.; Hashimoto, T.; Sakamoto, K. Fat Accumulation in Caenorhabditis Elegans Is Mediated by SREBP Homolog SBP-1. Genes Nutr 2010, 5, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Diot, C.; García-González, A.P.; Vieira, A.F.; Walker, M.; Honeywell, M.; Doyle, H.; Ponomarova, O.; Rivera, Y.; Na, H.; Zhang, H.; et al. Bacterial Diet Modulates Tamoxifen-Induced Death via Host Fatty Acid Metabolism. Nature Communications 2022 13:1 2022, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, S.F.; Seo, Y.; Allen, C.; Ghanta, K.S.; Finkel, S.; Tissenbaum, H.A. Bacterial Processing of Glucose Modulates C. Elegans Lifespan and Healthspan. Sci Rep 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Hartler, J.; Triebl, A.; Ziegl, A.; Trötzmüller, M.; Rechberger, G.N.; Zeleznik, O.A.; Zierler, K.A.; Torta, F.; Cazenave-Gassiot, A.; Wenk, M.R.; et al. Deciphering Lipid Structures Based on Platform-Independent Decision Rules. Nature Methods 2017 14:12 2017, 14, 1171–1174. [Google Scholar] [CrossRef]
- Byerly, L.; Cassada, R.C.; Russell, R.L. The Life Cycle of the Nematode Caenorhabditis Elegans: I. Wild-Type Growth and Reproduction. Dev Biol 1976, 51, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Muschiol, D.; Schroeder, F.; Traunspurger, W. Life Cycle and Population Growth Rate of Caenorhabditis Elegans Studied by a New Method. BMC Ecol 2009, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bodhicharla, R.; Devkota, R.; Ruiz, M.; Pilon, M. Membrane Fluidity Is Regulated Cell Nonautonomously by Caenorhabditis Elegans PAQR-2 and Its Mammalian Homolog AdipoR2. Genetics 2018, 210, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Gao, C.; Wang, M.; Tran, P.; Mai, N.; Finley, J.W.; Heymsfield, S.B.; Greenway, F.L.; Li, Z.; Heber, D.; et al. Lower Doses of Fructose Extend Lifespan in Caenorhabditis Elegans. https://doi.org/10.1080/19390211.2016.1212959 2016, 14, 264–277. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, L.; Zhang, L.; Wang, W.; Wei, S.; Wang, J.; Che, H.; Zhang, Y. Effects of Excess Sugars and Lipids on the Growth and Development of Caenorhabditis Elegans. Genes Nutr 2020, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular Death, Reactive Oxygen Species (ROS) and Diabetic Complications. Cell Death & Disease 2018 9:2 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Wu, N.; Shen, H.; Liu, H.; Wang, Y.; Bai, Y.; Han, P. Acute Blood Glucose Fluctuation Enhances Rat Aorta Endothelial Cell Apoptosis, Oxidative Stress and pro-Inflammatory Cytokine Expression in Vivo. Cardiovasc Diabetol 2016, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Oguntibeju, O.O. Type 2 Diabetes Mellitus, Oxidative Stress and Inflammation: Examining the Links. Int J Physiol Pathophysiol Pharmacol 2019, 11, 45. [Google Scholar] [PubMed]
- Peng, C.; Ma, J.; Gao, X.; Tian, P.; Li, W.; Zhang, L. High Glucose Induced Oxidative Stress and Apoptosis in Cardiac Microvascular Endothelial Cells Are Regulated by FoxO3a. PLoS One 2013, 8, e79739. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid Med Cell Longev 2014, 2014. [Google Scholar] [CrossRef]
- Assies, J.; Mocking, R.J.T.; Lok, A.; Ruhé, H.G.; Pouwer, F.; Schene, A.H. Effects of Oxidative Stress on Fatty Acid- and One-Carbon-Metabolism in Psychiatric and Cardiovascular Disease Comorbidity. Acta Psychiatr Scand 2014, 130, 163–180. [Google Scholar] [CrossRef]
Figure 1.
High concentrations of glucose reveal a compromised flux of oleic and linoleic acids. (A) The fate of oleic acid (C18:1n9) in the elongase and desaturase pathway is shown with the desaturases (“fat” genes) listed. (B) L1 nematodes were grown for 48 hours followed by 12 hours of glucose stress at 15mM (black dashed), 100 mM (black dots), and 200 mM (black hatch). After glucose treatment, nematodes were labeled by feeding13C:12C (60:40) for 6 hours. Multiple fatty acids species showed significant differences in the % of newly incorporated fatty acid when compared to the control populations (white bars). (C) The relative % of the fatty acids upstream and downstream of oleic acid were determined in the same populations of animals described above. (D) Lifespans showed significantly shorter mean and maximum lifespan in animals fed glucose at all concentrations at L4. All GC/MS analysis values represent means ± SEM of at least 9 biological replicates. Statistical significance (P<0.05) is indicated by * and was calculated using unpaired T tests and F tests to compare variances. Survival curves are presented as means of at least three independent replicates with n = 50 nematodes/condition. Statistical analysis was performed by Log-rank (Mantel-Cox) test. P≥0.05 is represented by *. .
Figure 1.
High concentrations of glucose reveal a compromised flux of oleic and linoleic acids. (A) The fate of oleic acid (C18:1n9) in the elongase and desaturase pathway is shown with the desaturases (“fat” genes) listed. (B) L1 nematodes were grown for 48 hours followed by 12 hours of glucose stress at 15mM (black dashed), 100 mM (black dots), and 200 mM (black hatch). After glucose treatment, nematodes were labeled by feeding13C:12C (60:40) for 6 hours. Multiple fatty acids species showed significant differences in the % of newly incorporated fatty acid when compared to the control populations (white bars). (C) The relative % of the fatty acids upstream and downstream of oleic acid were determined in the same populations of animals described above. (D) Lifespans showed significantly shorter mean and maximum lifespan in animals fed glucose at all concentrations at L4. All GC/MS analysis values represent means ± SEM of at least 9 biological replicates. Statistical significance (P<0.05) is indicated by * and was calculated using unpaired T tests and F tests to compare variances. Survival curves are presented as means of at least three independent replicates with n = 50 nematodes/condition. Statistical analysis was performed by Log-rank (Mantel-Cox) test. P≥0.05 is represented by *. .
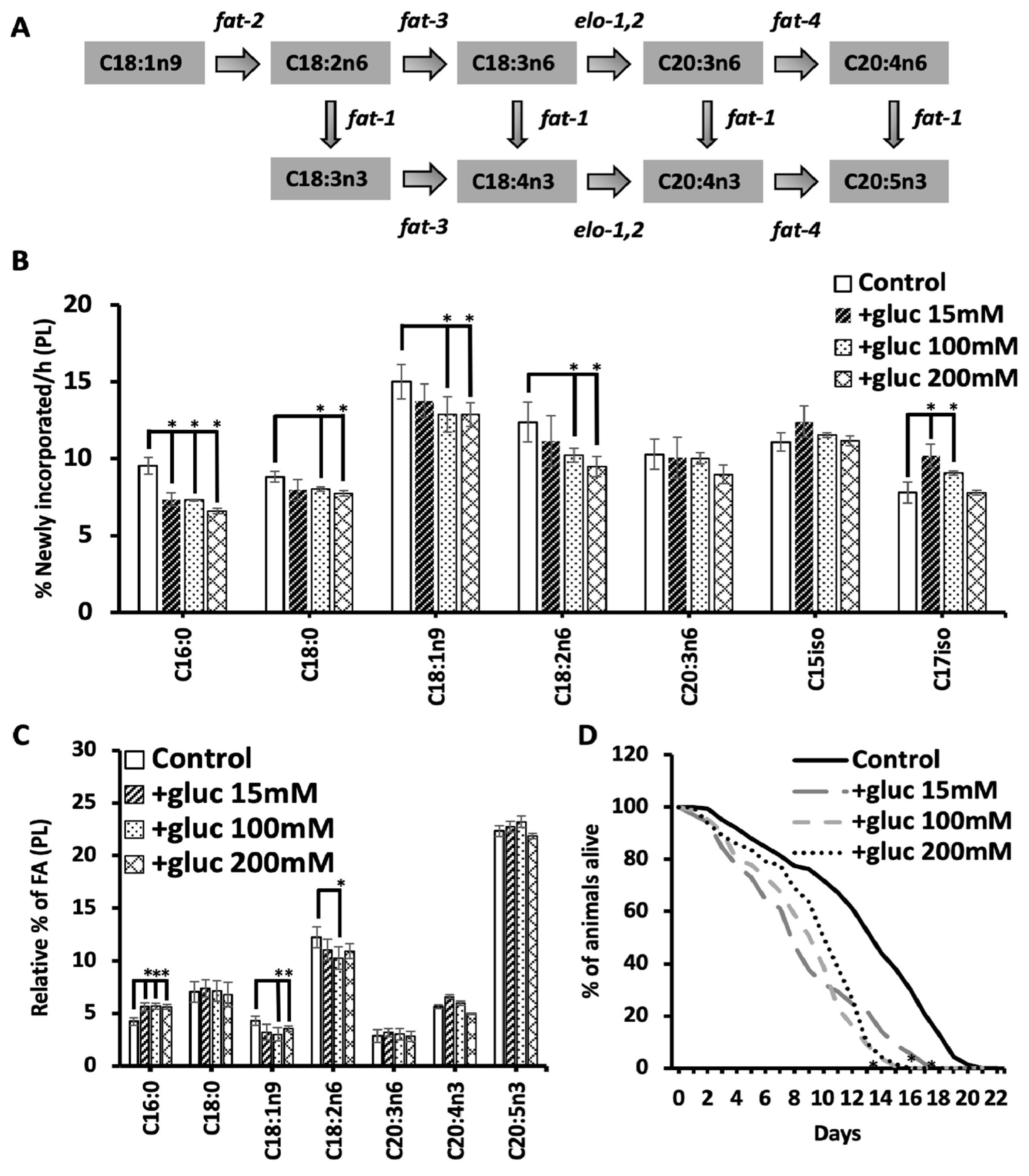
Figure 3.
Longer stress periods stabilize oleic acid and linoleic acid levels. (A) The relative abundance of C16:0, C18:1n9 and C18:2n6 were found by GC-MS in the following treatment groups: +gluc 12h (black dot), 24h +gluc (blue horizontal brick), +gluc 48h (orange hatch) and +gluc 72h (gold diagonal stripes). The fatty acid abundances were compared to age-matched controls without glucose exposure: 12h (black bars), 24h (blue), 48h (orange) and 72h (gold). (B) The relative % of C20:3n6, C20:4n3 and C20:5n3 were assessed in 12h and 24h nematodes with and without glucose treatment. The small but significant differences are noted with a *. (C) The % of newly incorporated FA was quantified after incorporating dietary stable isotopes in the same treatment groups as defined above. (D) FA synthesis was calculated based on the abundance of isotopomers characteristic of synthesis (i.e., MW 272-284 for C16:0 and MW 301-313 for C18:1n9) after correcting for natural abundance of isotopes and corrected by a normalization factor the accounts for differences in the distribution of isotopes in the diet. For all GC-MS analysis, values are means ± SEM of at least 4 replicates. Statistical significance (P<0.05) is shown by * and was calculated using unpaired T tests and F tests to compare variances. .
Figure 3.
Longer stress periods stabilize oleic acid and linoleic acid levels. (A) The relative abundance of C16:0, C18:1n9 and C18:2n6 were found by GC-MS in the following treatment groups: +gluc 12h (black dot), 24h +gluc (blue horizontal brick), +gluc 48h (orange hatch) and +gluc 72h (gold diagonal stripes). The fatty acid abundances were compared to age-matched controls without glucose exposure: 12h (black bars), 24h (blue), 48h (orange) and 72h (gold). (B) The relative % of C20:3n6, C20:4n3 and C20:5n3 were assessed in 12h and 24h nematodes with and without glucose treatment. The small but significant differences are noted with a *. (C) The % of newly incorporated FA was quantified after incorporating dietary stable isotopes in the same treatment groups as defined above. (D) FA synthesis was calculated based on the abundance of isotopomers characteristic of synthesis (i.e., MW 272-284 for C16:0 and MW 301-313 for C18:1n9) after correcting for natural abundance of isotopes and corrected by a normalization factor the accounts for differences in the distribution of isotopes in the diet. For all GC-MS analysis, values are means ± SEM of at least 4 replicates. Statistical significance (P<0.05) is shown by * and was calculated using unpaired T tests and F tests to compare variances. .
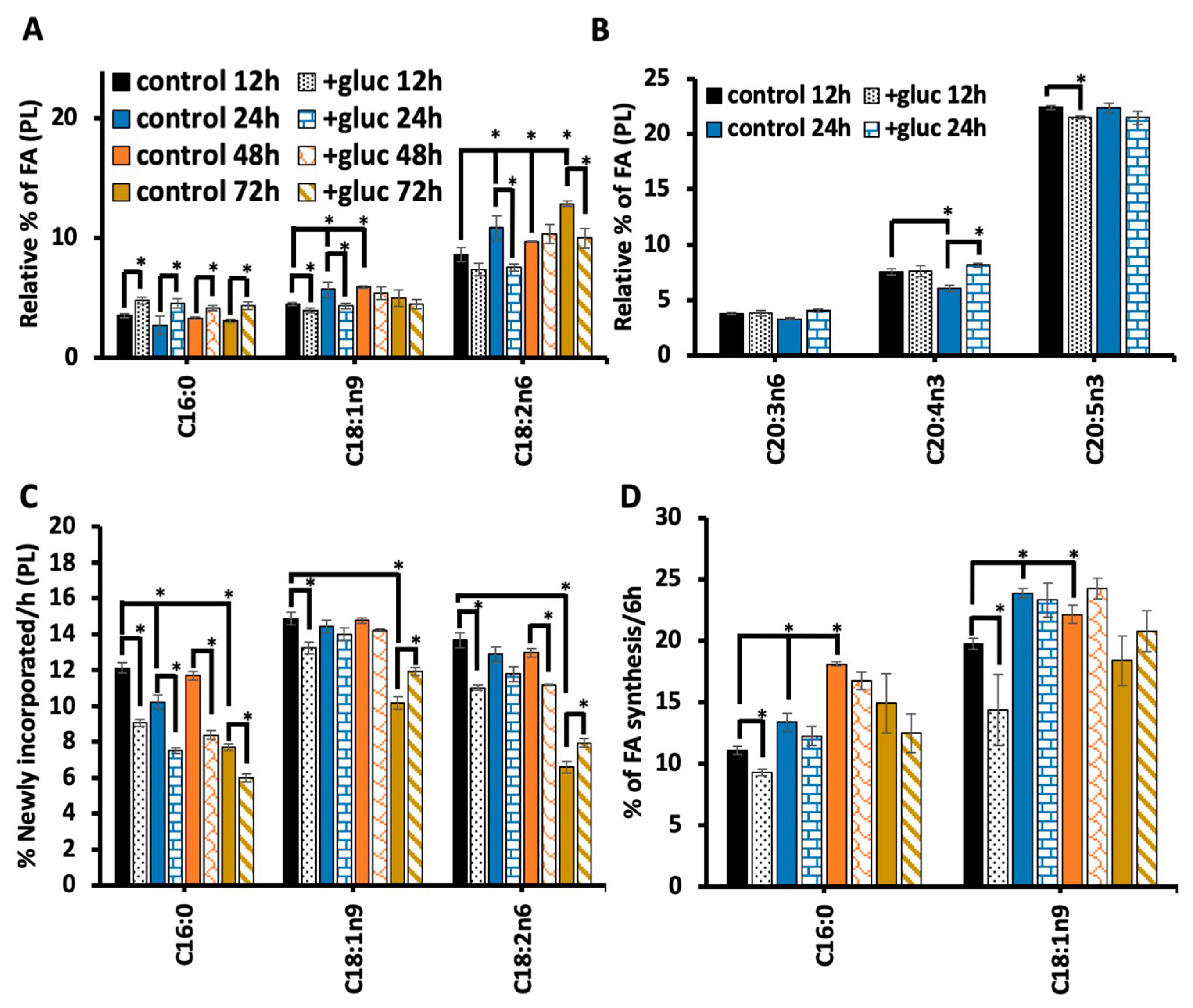
Figure 4.
Glucose stress phenotypes require living bacteria. (A) Representation of the experimental design to test the impact of the bacteria processing glucose. Nematodes were fed four different diets of living bacteria (OP50; white), living bacteria resuspended in glucose (OP50 +gluc; white with black dots), heat killed bacteria (OP50 HK; black), and heat killed bacteria resuspended in glucose (OP50 HK +gluc; black with white dote). (B) The relative % of FA in bacteria was assessed by GC-MS and did not show statistical significance when comparing OP50 to OP50 +gluc and OP50 HK to OP50 HK +gluc. Comparing the killed bacteria to living bacteria showed a significant increase in all SFA (C12:0, C14:0, and C16:0) and significant decrease in all UFA (unsaturated fatty acids) (C16:1n7 and C18:1n7). (C) The PCs in nematodes fed the different diets were evaluated by HPLC-MS/MS. A binned analysis of number of double bonds in the PC populations is shown with statistically significant changes denoted. (D) There was no significant alteration in the relative % of chain length of PC species in any condition. (E) PE species binned based on the double bond distribution (E) and chain length (F) are shown with statistically significant (P<0.05) changes marked by an *. .
Figure 4.
Glucose stress phenotypes require living bacteria. (A) Representation of the experimental design to test the impact of the bacteria processing glucose. Nematodes were fed four different diets of living bacteria (OP50; white), living bacteria resuspended in glucose (OP50 +gluc; white with black dots), heat killed bacteria (OP50 HK; black), and heat killed bacteria resuspended in glucose (OP50 HK +gluc; black with white dote). (B) The relative % of FA in bacteria was assessed by GC-MS and did not show statistical significance when comparing OP50 to OP50 +gluc and OP50 HK to OP50 HK +gluc. Comparing the killed bacteria to living bacteria showed a significant increase in all SFA (C12:0, C14:0, and C16:0) and significant decrease in all UFA (unsaturated fatty acids) (C16:1n7 and C18:1n7). (C) The PCs in nematodes fed the different diets were evaluated by HPLC-MS/MS. A binned analysis of number of double bonds in the PC populations is shown with statistically significant changes denoted. (D) There was no significant alteration in the relative % of chain length of PC species in any condition. (E) PE species binned based on the double bond distribution (E) and chain length (F) are shown with statistically significant (P<0.05) changes marked by an *. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



