
Submitted:
16 October 2023
Posted:
30 October 2023
You are already at the latest version
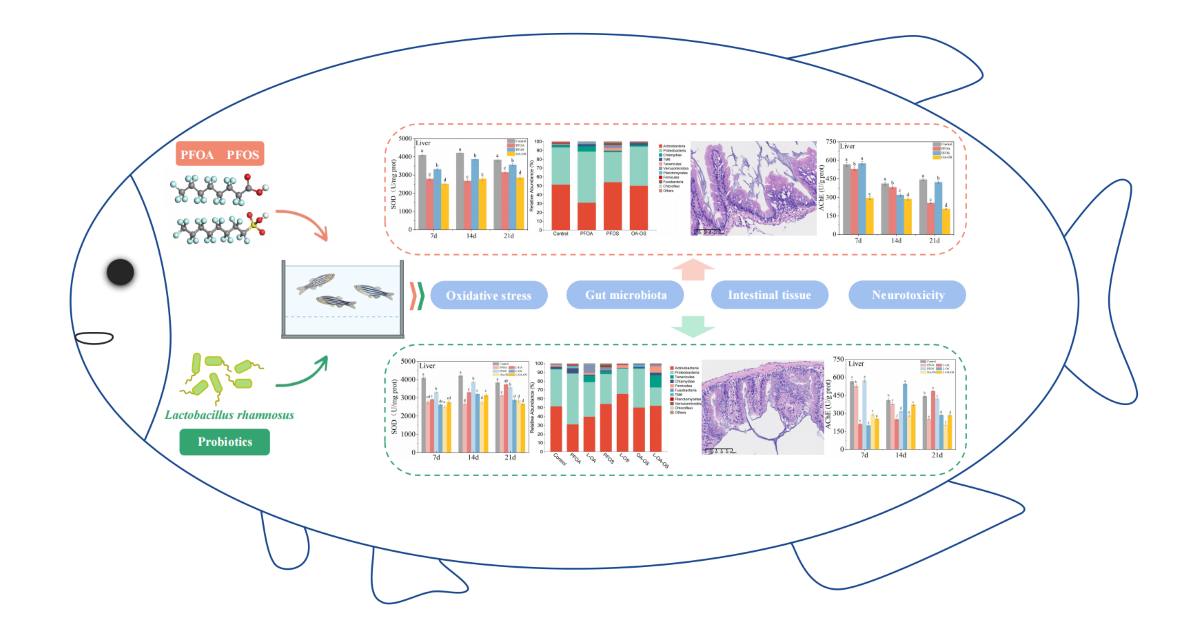
Abstract
The EPA has banned PFOA and PFOS as persistent organic pollutants, but their residues in environment should not be underestimated. Based on the demand for healthy aquaculture and safe aquatic products, it is necessary to explore alleviative methods for the toxicity of PFOA and PFOS to fish. This study is the first to investigate the protective effects of Lactobacillus rhamnosus against oxidative damage, neurotoxicity, and gut microbial dysbiosis induced by PFOA and PFOS in adult zebrafish. The results showed that L. rhamnosus effectively alleviated PFOA- and PFOS-induced oxidative stress by restoring superoxide dismutase (SOD), catalase (CAT), and lactate dehydrogenase (LDH) levels. Both PFOA and PFOS caused intestinal tissue damage, and the degree of damage is PFOA+PFOS>PFOS>PFOA. L. rhamnosus can restore damaged intestinal villi and lymphocyte numbers and also have a restorative effect on intestinal tissue damage. The up-regulated proportion of beneficial Firmicutes and Actinobacteria was also found after L. rhamnosus feeding. These findings provide new insights for L. rhamnosus to alleviate the toxicity of PFOA and PFOS to zebrafish and promote the progress of research on the successful use of probiotics in freshwater aquaculture.
Keywords:
PFOA
; PFOS
; Lactobacillus rhamnosus
; Chronic toxicity
; Oxidative stress
; Gut microbiota
1. Introduction
Perfluorinated and polyfluoroalkyl substances (PFAS) are organic compounds containing at least one perfluorinated carbon atom. Since the 1950s, polymers and surfactants containing PFAS have been widely used in pesticide synthesis, electroplating, firefighting, textiles, and polymer production industries [1]. However, the environmental persistence of PFAS, which is not easily photolyzed, hydrolyzed, oxidized and biodegraded, has caused widespread concern. Currently, PFAS are detected globally in air, water, sediment, soil, wildlife, and even polar ice sheets [2,3]. Compared to atmospheric sources (<5%), the aquatic environment is a significant sink for PFAS (>95%) [4].
PFAS are widespread in aquatic environments, transferred through the food chain, and accumulated in organisms, which resulting in higher PFAS concentrations in aquatic products than in aqueous environments [5]. Many studies have shown that PFAS can affect the growth, development, and reproduction of aquatic organisms and cause serious harm to the liver, immune system, reproductive system, etc. [6]. China is a big fishery country with a large variety of aquaculture and is the world's top exporter. Therefore, it is crucial to reduce the toxicity of PFAS to aquatic products and thus reduce the risk of human intake of PFAS.
Existing studies have shown that although members of PFAS can be removed from the aquaculture water environment by physical and chemical methods, the economics and biosafety need to be evaluated [7]. Functional probiotics can improve the resistance of aquatic products to contaminants and were thought to be effective biological control methods for PFAS pollutants and its toxicity [8]. Probiotics are living microorganisms that regulate the body's mucous membrane and immune system and improve intestinal nutrient absorption and microbial community function, thus playing important roles in mitigating the toxic effects of contaminants in aquaculture [9]. It is interesting to use zebrafish as a model for aquaculture species. Eighty percent of farmed fish are other carp, and therefore close relatives [10].
As the most studied probiotic, L. rhamnosus can alleviate intestinal microbial dysbiosis, reproductive developmental toxicity, and lipid metabolism disorders induced by PFBS in zebrafish [11,12,13]. However, the toxic effects and mechanisms of PFOA and PFOS mixtures in zebrafish are unknown. Therefore, this study aimed to assess the alleviating effect and mechanism of probiotic L. rhamnosus on PFOA/PFOS-induced ecotoxicity in zebrafish by evaluating intestinal histological changes, intestinal microbiota, and oxidative stress response.
2. Materials and methods
2.1. Chemicals and Zebrafish maintenance
Perfluorooctane sulfonates (PFOS, CAS No. 2795-39-3, purity: 98%) was purchased from Beijing Bailingway Technology Co., Ltd (Beijing, China). Perfluorooctanoic acid (PFOA, CAS No. 335-67-1, purity: 98%) was purchased from Baidisin Technology Co. Ltd (Beijing, China). Probiotic Lactobacillus rhamnosus ATCC53103 (108 CFU/g; Culturelle, Italy) was acquired from China General Microbiological Culture Collection Center (CGMCC, Beijing, China).
Adult wild-type zebrafish (AB, 3-4 months old, all males) were purchased from the China Zebrafish Resource Center (Wuhan, China). Zebrafish were 3.0±0.5 cm in body length and 0.4±0.05 g in weight. The male-to-female ratio was 1:1. Before entering the laboratory, and zebrafish were domesticated for 14 days. The culture conditions were 26±1°C, 12/12h light/dark cycle, oxygen saturation over 70%, and pH of 7.4 to 8.1. Zebrafish were regularly fed twice a day with fairy shrimp for apparent satiety.
2.2. Experimental design
The domesticated zebrafish were randomly assigned to seven experimental groups with three replicates per group. The experimental groups setting were Control (fully aerated tap water), PFOA, L-OA (PFOA and L. rhamnosus), PFOS, L-OS (PFOS and L. rhamnosus), OA-OS (PFOA and PFOS), and L-OA-OS (PFOA, PFOS, and L. rhamnosus). PFOA and PFOS exposure concentrations were 30 mg/L and 0.3 mg/L. Exposure concentrations of PFOA and PFOS were determined based on environmental concentrations [14,15] and LC50 of PFOA and PFOS obtained from acute experiments (Figure S1).
Each exposure group had three identical glass water tanks (n = 3), with 25 zebrafish in each tank. The exposure solution was changed every two days to maintain water quality and concentrations. The feeding conditions, such as light, temperature, and food, were consistent with the domestication period. The exposure lasted for 21 days. Stop feeding 24 hours before each sampling to prevent feces from contaminating the water and influencing the experimental results.
2.3. Sample collection
Adult zebrafish tissues were collected at 7, 14, and 21 days after exposure. Three zebrafish were randomly selected from each tank and dissected to obtain intestines, livers, brains, and gills. At the same time, one zebrafish was randomly selected to scrape the intestinal contents. Immediately freeze it in liquid nitrogen and store it at -80 °C for subsequent molecular analysis. One zebrafish was randomly selected at 21 days of exposure to obtain intestinal tissues for histological analyses. All procedures are approved by the Animal Experiment Ethics Committee of Beijing Technology and Business University and conducted according to the guidelines for the protection of animal welfare.
2.4. Intestinal histology
Dissected zebrafish intestinal tissues were quickly stored in 4% paraformaldehyde, fixed for 24 h, dehydrated, embedded, sectioned, and stained with hematoxylin and eosin (H & E). After the sections were made, microscopy was used to observe and record the histological changes.
2.5. 16S rRNA amplicon sequencing and bioinformatic analysis
To characterize the changes in gut microbial communities and metabolic activities in each exposure group, total DNA from gut samples was extracted using the MoBio PowerFecal™ Fecal DNA Ki. The V3-V4 hypervariable regions of bacterial 16S rRNA gene were amplified from extracted DNA using bar-coded primers (forward 338F: ACTCCTACGGGAGGCAGCA, reverse 806R: GGACTACHVGGGTWTCTAAT). After purification, all amplicons were pooled with equimolar amounts from each sample and sequenced using the Illumina platform (Novogene, Beijing, China).
High-throughput sequencing data were further analyzed using QIIME (V1.9.1). Closed operational taxonomic units (OTUs) packing and taxonomic information were annotated with a 97% sequence similarity threshold [16]. Alpha diversity was estimated by Chao1, Shannon, Simpson's diversity, and Good's coverage index to describe species composition in a given treatment and differences between treatments. PCoA analysis based on Bray-Curtis was performed to assess Beta diversity, revealing differences in community structure between samples. Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt2) was used to predict functional gene changes and metabolic activities in the gut microbial community of different treatment groups.
2.6. Measurement of biochemical indicators
Zebrafish gut, liver, brain, and gill tissue samples were accurately weighed, and a 10% tissue homogenate was made by adding nine times of normal saline. The samples were then centrifuged at 2,500 rpm for 10 minutes. After dilution, the supernatant was collected. Superoxide dismutase (SOD), catalase (CAT), glutathione (GSH), malondialdehyde (MDA), lactate dehydrogenase (LDH), and acetylcholinesterase (AChE) were tested using assay kits (Nanjing Jiancheng Institute of Biological Engineering, Jiangsu, China).
2.7. Statistical Analysis
Statistical analysis was performed using SPSS software (version 24.0). Data from each experimental group were tested for normality (Shapiro-Wilk) and chi-square (Leven). In order to analyze the variances of data results, one-way analysis of variance (ANOVA) was used to determine significant differences between groups, followed by a post hoc LSD test. Graphs were constructed using Origin 2021 (Microcal, Redmond, Washington, USA). Statistical significance was considered when P<0.05. All data are expressed as mean ± standard deviation.
3. Results
3.1. Effect of different exposure treatments on SOD activity of zebrafish tissues
Based on the results, liver SOD activity was the highest among the four sampled tissues (Figure 1). SOD activity in the intestine and liver decreased overall in all exposure groups compared to the control group. At the same time, it showed a more significant decrease in the combined exposure group than in the single exposure group (Figure 1a,b). The changes of SOD activity in the brain were insignificant (Figure 1c). In gills, SOD activity increased on day 14 in each exposure group (Figure 1d).
The effect of L. rhamnosus on PFOA and OA-OS groups were significant. Compared with the exposed groups, liver SOD activity was increased by 15.52% and 7.74% in the L-OA and L-OA-OS groups, respectively, with the most significant effect on day 14 (Figure 1b). Intestinal SOD activity was induced to increase in L-OA and L-OA-OS groups at 14 days (Figure 1a). At this point, L. rhamnosus inhibited the abnormally elevated SOD activity in the gills and returned it to control levels (Figure 1d).
3.2. Activities of CAT in zebrafish tissues of different treatments
Under the effect of PFOA and PFOS, CAT activity in the intestine increased and then decreased in the exposed groups, and the impact of L. rhamnosus was not significant (Figure 1e). Compared with the control group, CAT activity in the liver, brain, and gills decreased with prolonged exposure time in the exposed group. The induction of CAT activity by L. rhamnosus was increasingly significant (Figure 1f-h).
3.3. MDA content in zebrafish tissues of different treatments
In the liver, there was no significant change in MDA content of the PFOA group. MDA content increased in PFOS and OA-OS groups before returning to control levels (Figure 2b). Feeding of L. rhamnosus increased the level of MDA in L-OA group. Compared to the PFOA group, the mean MDA increased by 35.96%. The MDA content increased by 27.50% in the L-OS group at 7 days and 46% in L-OA-OS group at 14 days. In the intestine, brain, and gills, PFOA and PFOS decreased MDA content (Figure 2a,c,d). L. rhamnosus has restored MDA content in the intestine to some extent. Compared to the exposed groups, MDA content showed marked increases after 21 days of L. rhamnosus feeding by 102.70%, 13.32%, and 47.37%, respectively. However, the effect on MDA content in the gills was not significant. Notably, L. rhamnosus caused a further reduction of MDA in the brain at 7 and 14 days.
3.4. Changes of GSH content in zebrafish tissues of different treatments
Compared with the control group, GSH content in the intestine and liver were decreased in all exposed groups. The effects of L. rhamnosus on GSH in the intestine and liver were insignificant (Figure 2e,f). In the brain, GSH levels decreased first, then increased in the PFOA and OA-OS groups. GSH levels first remained unchanged and then increased in the PFOS group. At the end of exposure, GSH content was higher in all exposure groups than control. L. rhamnosus feeding suppressed the elevation of GSH in each exposure group and restored it to normal levels in the brain at 21 days. The GSH levels decreased by 50.46%, 43.73%, and 40.43% in OA, OS, and OA-OS groups, respectively (Figure 2g). In the gills, L. rhamnosus restored GSH to normal levels in the PFOA and OA-OS groups (Figure 2h).
3.5. Influence of various exposure treatments on LDH activity of zebrafish tissues
In the brain, LDH activity of PFOA and PFOS groups were first inhibited and then restored. However, the OA-OS group was consistently inhibited. Feeding L. rhamnosus inhibited LDH activity in the PFOA group and increased LDH activity in the OA-OS group (Figure 3c). In the gills, PFOA and PFOS had no significant effect on LDH activity, but L. rhamnosus induced increased LDH activity (Figure 3d). PFOA and PFOS inhibited LDH activity in the liver and intestine, and L. rhamnosus restored LDH levels in the control group (Figure 3a,b).
3.6. Differences in AChE activity of different treatment groups
In the brain, no significant difference in AChE activity was found in the PFOA and PFOS groups compared with the control group at 21 days. However, AChE activity was mainly inhibited in the OA-OS group. AChE was inhibited in the PFOA and PFOS groups under the feeding of L. rhamnosus (Figure 3g). In the gills, the trend of AChE activity in PFOS and OA-OS groups was elevated-recovered-lowered compared to control. The PFOA group remained unchanged and then increased. L. rhamnosus inhibited the increase in AChE activity, with the best effect at 7 days (Figure 3h).
3.7. Changes in zebrafish intestinal tissue morphology
Under the tissual staining and section analysis, it was observed that the control group had no apparent pathological damage. The intestinal wall was thick, the intestinal villi were morphologically intact, and their epithelial cells and epidermal structures were intact and undamaged (Figure 4a). The intestinal villi of the PFOA, PFOS, and OA-OS groups showed different degrees of damage, and the number of goblet cells decreased. The epithelial cells showed signs of shedding and vacuolization (Figure 4b,d,f). The degree of intestinal damage: PFOA+PFOS>PFOS>PFOA. These injuries were improved to some extent by adding the probiotic L. rhamnosus (Figure 4c,e,g).
3.8. Community composition and metabolic activities of gut microbiota
After sequencing and data quality control analysis, the sparsity curves showed that all samples reached the saturation plateau, indicating that the sampling depths were sufficient (Figure S2). In this study, a total of 13,396 OTUs were obtained with a 97% similarity. The total number of OTUs was 101 (Figure S3).
The Chao1 index, Shannon index, and Simpson index were used to determine the gut microbial alpha-diversity of zebrafish. Compared to the control group, PFOA and PFOS affected the Chao1, Shannon, and Simpson indexes (Figure 5a-c). After adding L. rhamnosus, the Chao1 index of the gut microbiota was higher than that of the control (P<0.05). PCoA results based on Bray-Curtis distance showed that the addition and non-addition of L. rhamnosus resulted in the formation of two independent clusters for the gut microbial community of zebrafish (P<0.05) (Figure 5d).
To further investigate the changes in the gut microbial community of zebrafish, the frequencies of bacterial phyla and genera were analyzed based on high-throughput sequencing of the metagenome (Figure 6). The results showed that the significant phyla were Actinobacteria, Proteobacteria, Firmicutes, and Tenericutes, with relative abundance of 46.78%, 43.19%, 1.58%, and 4.24%, respectively (Figure 6a). The main genera were Rhodobacter, Microbacterium, Leucobacter, Mycobacterium, Aeromonas, Gordonia (Figure 6b).
At the phylum level, the addition of L. rhamnosus increased the abundance of Firmicutes and Actinobacteria but decreased Proteobacteria. At the genus level, Leucobacter and Microbacterium were raised, but Rhodobacter, Gordonia, and Rhodococcus were decreased. These results showed that L. rhamnosus regulated adult zebrafish gut microbiota in the presence of PFOA and PFOS exposure interference.
Considering that the function of the gut microbial community determines the physiological properties of the host, we used PICRUSt2 to analyze and predict function variance. PICRUSt2 analysis was carried out on the MetaCyc database to identify differentially relevant metabolic passages, biochemical processes, enzymes, and gene alterations to anticipate microbial metabolic functions.
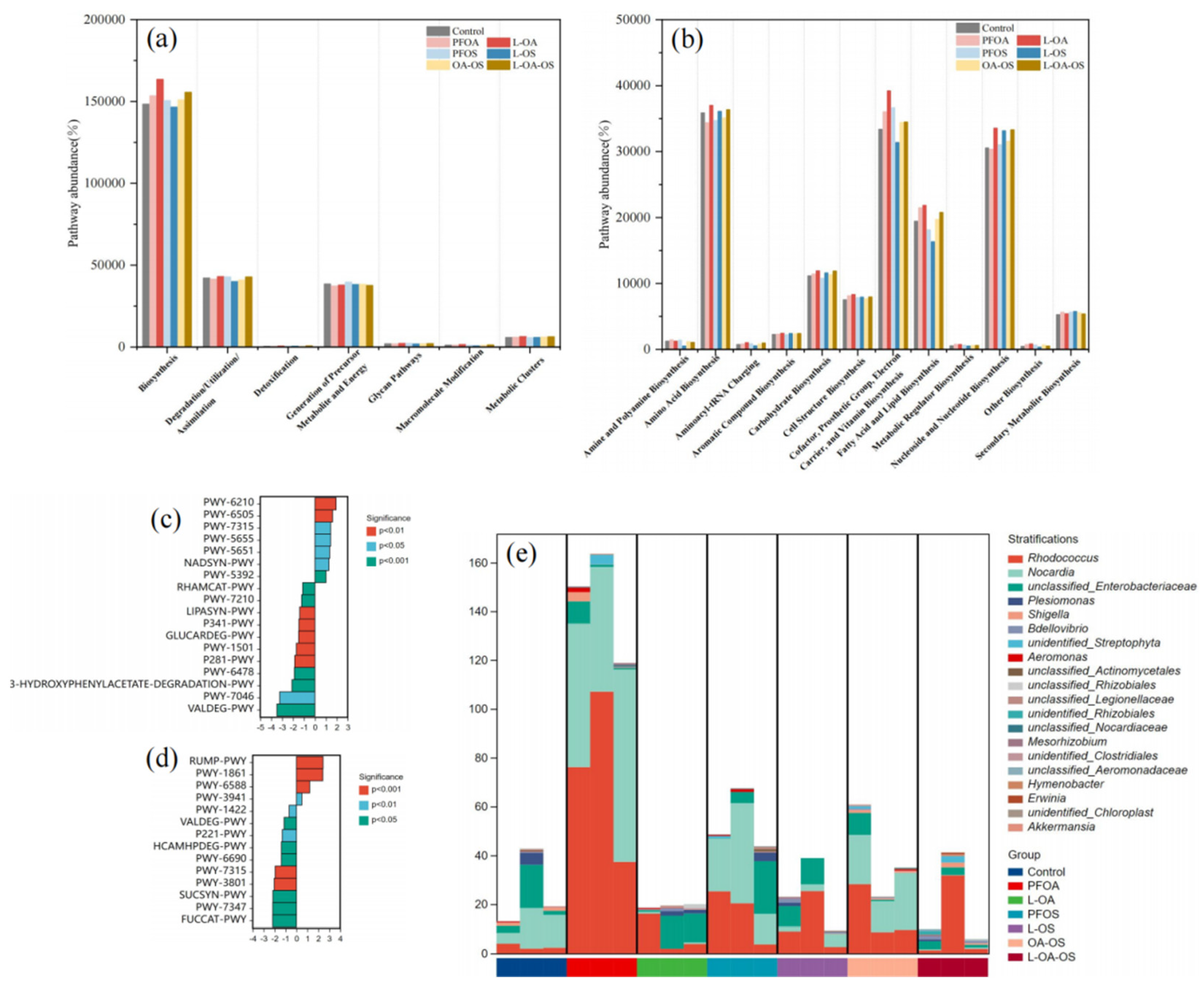
As shown in Figure 7a, each signaling pathway was affected to varying degrees. PFOA and PFOS increased the abundance of the biosynthesis pathway and decreased the abundance of the generation of precursor metabolite and energy and degradation/utilization/assimilation pathways. L. rhamnosus reduced the abundance of these pathways. The biosynthesis metabolic pathway had the greatest abundance, and L. rhamnosus increased the abundance of amino acid biosynthesis pathway in the secondary pathways (Figure 7b). By metabolic pathway difference analysis (Figure 7c,d), the PWY-7315 pathway was up-regulated in the PFOA group compared to the control group and downregulated by adding L. rhamnosus. Figure 7e showed the species composition of this metabolic pathway. The results showed that the exposure group resulted in a higher proportion of Rhodococcus in PWY-7315 pathway, while L. rhamnosus alleviated this trend.
4. Discussion
4.1. Probiotic L. rhamnosus relieves zebrafish tissue oxidative stress
Reactive oxygen species (ROS) include hydrogen peroxide (H2O2), hydroxyl radicals (·OH), and superoxide anion radicals (·O2-). These are produced by organisms under the action of normal cellular metabolism and environmental factors. Moreover, typical concentrations of ROS do not cause damage to organisms. However, when adverse environmental influences stress organisms, large amounts of ROS will be produced in cells, leading to an imbalance between oxidation and antioxidation, inducing oxidative stress. Excess ROS trigger lipid peroxidation, which disrupts the structure and permeability of cell membranes. Moreover, the lipid peroxidation product, MDA, further damages protein activity [17]. In addition, excessive ROS can damage DNA to produce genotoxicity in the host [18]. The probiotic L. rhamnosus was found to efficiently scavenge hydroxyl radicals (·OH) and superoxide anion radicals (·O2-) and improve the antioxidant capacity of H2O2-induced oxidative stress [12].
SOD can convert ·O2- into H2O2 and O2 in vivo, protect cells from damage, and maintain the oxidative and antioxidant balance of the body [19]. SOD can be used as a biomarker of environmental stress in aquatic organisms. When organisms are subjected to mild adversity, SOD activity increases to activate the immune system. If hosts are subjected to severe adversity stress, SOD activity decreases, and the accumulation of ·O2- in the body suppresses the immune system [20].
In this study, PFOA and PFOS inhibited SOD activity. It indicated that to resist the activity of toxic substances, SOD was reduced and did not eliminate ROS in time. Therefore, PFOA and PFOS caused significant oxidative damage to zebrafish. In addition, the production of large amounts of H2O2 by organisms also inhibits SOD activity [21]. In the gills, SOD activity was first increased and then inhibited in exposed groups. It might be due to pollutant stress in zebrafish, which produced large amounts of ROS, which in turn activated the immune mechanism. Excess free radicals were scavenged by accelerated SOD synthesis, but the immune system was impaired with increasing exposure time.
L. rhamnosus regulated PFOA and PFOS-induced abnormalities in SOD activity. The effect of L. rhamnosus on the restoration of liver SOD activity was more pronounced than that in the intestine. Notably, L. rhamnosus inhibited SOD activity with prolonged exposure time, which was possibly due to the need to maintain superoxide anion levels or through metal-catalyzed singlet oxygen (1O2) and/or ·OH interactions to enhance microbial killing by phagocytes [22].
CAT can break down H2O2 produced under the action of SOD into H2O and O2, effectively preventing the production of harmful hydroxyl groups, which can cause phospholipid dysfunction, biofilm damage, and DNA breakage [23]. This experiment showed that the intestinal CAT activity of zebrafish exhibited the trend of increasing first and then decreasing, and the liver CAT activity was inhibited. In the presence of L. rhamnosus, CAT activity was restored to some extent in all tissues.
The liver of zebrafish is most sensitive to the toxicity of PFOA and PFOS, a vital target tissue for PFAS toxicity that causes oxidative stress, endocrine disorders, and altered lipid metabolism in the host [24]. In the early stages of exposure to pollutants, the SOD and CAT activities were increased to protect the body against excess ·O2-. With prolonged exposure, large amounts of H2O2 inhibited enzyme activity. It indicated that the antioxidant system of zebrafish was disrupted.
ROS attacks polyunsaturated fatty acids in biological membranes leading to lipid peroxidation. MDA is a product of lipid peroxidation which can reflect the extent of cellular damage [25]. At the end of this experiment, MDA levels were equal to or lower than the control group in all exposure groups. Previous studies also showed that PFOS caused the increase of ROS but stable remaining MDA levels in tilapia liver [26]. A possible explanation might be that the pollutants did not cause severe lipid peroxidation and glutathione peroxidase (GPx) inhibition [27], and the decrease in MDA levels may also be related to GPx activity. L. rhamnosus can reduce MDA by activating the antioxidant enzyme activity system to protect zebrafish from damage caused by oxidative stress.
GSH is a non-enzymatic antioxidant that eliminates free radicals associated with selenium-dependent glutathione peroxidase and also regulates intracellular redox homeostasis [28]. In this study, PFOA and PFOS exposure decreased GSH content in the intestine and liver, indicating that GSH was heavily depleted to overcome oxidative stress. It has been shown that GSH content was reduced under PFOA and PFOS [26]. The increased GSH in the brain and gills may be an adaptation to oxidative stress or a failure of adaptation due to severe oxidative stress. Probiotic L. rhamnosus restored the GSH content to normal levels. Similar results demonstrated that probiotics can influence oxidative stress by producing or inducing the release of GSH and extracellular polysaccharide (EPS) from the intestine [29].
LDH is a cytoplasm oxidoreductase enzyme that facilitates the transfer of hydrogen ions. LDH can catalyze the conversion process of lactate to pyruvate to participate in oxidative reactions and cause oxidative stress in organisms [30]. It is a metabolic biomarker of oxidative stress and plays an important role in the glycolytic cycle [31]. In this study, PFOA and PFOS led to the reduction of LDH activity, while the addition of probiotic L. rhamnosus could induce and restore LDH activity. It has been shown that LDH decreased in the liver of Oreochromis mossambicus exposed to an organophosphorus insecticide [32]. The reduction in LDH activity reflects possible decreases in biosynthetic activity and anaerobic capacity. In contrast, the addition of probiotics L. rhamnosus induced LDH activity and may indicate an increase in the glycolytic capacity of the tissue.
The study showed that PFOA and PFOS significantly altered antioxidant enzyme activities, indicating oxidative stress in the exposed groups of zebrafish. The oxidative stress was the most pronounced in the liver, which may be related to the highest concentration of PFAS in the liver of fish [33]. SOD, CAT, GSH, and LDH activities were inhibited in zebrafish after PFOA/PFOS exposure alone and in combination. Probiotic L. rhamnosus reduced oxidative stress by inducing SOD, CAT, and LDH increases in zebrafish. The probiotic bacteria affected fish oxidative stress by other toxic substances for the same trend. Cadmium exposure inhibited SOD, CAT activities in carp, and Bacillus up-regulated SOD, CAT activities to alleviate oxidative stress [34].
4.2. Probiotic L. rhamnosus alleviate neurotoxicity induced by PFOA and PFOS
AChE, a landmark enzyme for evaluating the toxic effects of neurotoxic agents on aquatic organisms, is widely present in various animal tissues [35]. It catalyzes the hydrolysis reaction of the neurotransmitter acetylcholine (Ach) in the synaptic gap to maintain the normal transmission of nerve impulses. Changes in AChE activity affect the behavior of zebrafish, as evidenced by increased muscle activity, loss of locomotor activity, and anxiety-like behaviors [36].
In the present study, AChE activity was more affected by the combined exposure of PFOA and PFOS than single. The decrease in AChE activity with increasing exposure time may be caused by adaptation to a bad external environment. Similar results were shown in a previous study where zebrafish AChE decreased to adapt to the toxic deltamethrin [55]. The presumed reason for this is that zebrafish produce stress responses when exposed to external environmental stressors. Besides, acetylcholine in the synaptic gap decreases with exposure time, thus inhibiting excitation transmission. The effect of the probiotic L. rhamnosus on AChE activity in this experiment was complex. But it has been shown that the novel extracellular polysaccharide EPSRam12 produced by L. rhamnosus strain Ram12 protects against D-galactose-induced brain damage in mice [37].
4.3. Gut histomorphology and microbial community function regulation
Gut microbiota is crucial for host health, regulating host physiological and immune responses, but also the morphology and function of intestinal epithelium [38]. In this study, the results of intestinal histopathological sections showed that PFOA and PFOS caused lesions in zebrafish intestines, with the degree of damage in the following order: PFOA+PFOS>PFOS>PFOA. L. rhamnosus improved zebrafish intestinal health as evidenced by increased cupped cells, decreased vacuolization, and repairment of intestinal villi and basement membrane.
Probiotics can activate immune cells by recognizing damage related molecules released by stressed or damaged cells, thus altering the intestinal mucosa [39]. Feeding probiotics increased the intestinal cell count and microvillus density, thereby enhancing disease resistance by altering the gut microbiota composition in tilapia [40]. The increases in microvillus density and intestinal absorption surface area reduced the colonization rate of pathogenic bacteria, thus increasing the resistance of fish to pathogenic microorganisms [41]. Adding L. rhamnosus to the diet for 30 days showed increased proliferation of intraepithelial lymphocytes and eosinophils [42].
In terms of the number of microorganisms, the OTUs specific to each treatment group were higher, indicating that PFOA, PFOS, and L. rhamnosus all had greater impacts on the gut microbial community. The Chao1, Shannon, and Simpson index showed that L. rhamnosus altered the alpha-diversity of the intestinal microbial community, which could promote the colonization and replication of beneficial bacteria. A similar previous study found that probiotic L. rhamnosus could regulate PFBS-induced gut microbial dysbiosis in zebrafish [11]. PCoA plots could observe clearly separated clusters, suggesting that L. rhamnosus could alter the overall structure of the zebrafish gut microbiota. PICRUSt2 predicted that exposure to PFOA, PFOS, or/and L. rhamnosus led to the changes in PWY-7315 pathway, suggesting the link between L. rhamnosus and PFOA toxicity.
Gut microbes can produce large numbers of metabolites through metabolic pathways, such as volatile carboxylic acids, esters, neurotransmitters (e.g., dopamine), and fatty acids, some of which can affect brain physiology and behavior [43]. The results of this study suggested that exposure to PFOA and PFOS leads to dysbiosis of microbial communities in the intestine of zebrafish. The probiotic L. rhamnosus alleviated the toxic effects of pollutants on zebrafish by increasing beneficial bacteria and reducing pathogenic bacteria. Previous studies have shown that probiotics may reduce the disturbing effects on the gut microbial community under the stress of environmental pollutants [44]. This was consistent with the conclusion that pollutants and probiotics have antagonistic effects on the gut microbiota in the present study.
At the phylum level, the probiotic L. rhamnosus increased Firmicutes and Actinobacteria abundance and decreased Proteobacteria abundance. Studies have shown that Proteobacteria includes potentially pathogenic bacteria, and their over-representation is a potential marker of epithelial dysfunction, leading to ecological dysbiosis of the gut microbiota and causing small intestinal colitis [45,46]. L. rhamnosus belongs to Firmicutes, most of which are beneficial bacteria that improve fish growth, immunity, and disease resistance [47]. Actinobacteria not only removes organic pollutants to improve pond water quality but also helps to improve the immunity and growth of cultured fish [48]. At the genus level, the probiotic L. rhamnosus increased Leucobacter and Microbacterium and decreased Rhodobacter, Gordonia, and Rhodococcus. Gordonia is a pathogenic bacterium capable of producing certain secondary metabolites, which cause malformation and death in seahorses [49]. Some species of Rhodococcus cause nephritis and anophthalmia in salmon [50].
Various mechanisms have been proposed to explain the beneficial effects of probiotics. Examples include antagonism to pathogens, competition for adhesion sites and nutrients, enzymatic effects on digestion, improvement of water quality, and stimulation of host immune responses [51]. In the present study, the probiotic L. rhamnosus alleviated oxidative stress, neurotoxicity, and gut microbial community dysbiosis caused by PFOA and PFOS in zebrafish. Gut microbial dysbiosis causes metabolic disorders and immune damage in organisms but also affects neuronal signaling in the enteric nervous system by synthesizing neurotransmitters such as acetylcholine and serotonin [52]. L. rhamnosus can effectively shape intestinal microbes, regulate genes related to lipid metabolism and increase lipolytic metabolism for growth through β-oxidation [53]. It can also activate the interconversion of pentose and glucuronide in the liver [54], which eases the toxic effects of the organism.
5. Conclusion
In conclusion, this study demonstrated for the first time that L. rhamnosus alleviates the toxic effects of PFOA and PFOS on oxidative stress, neurotoxicity, gut microbial homeostasis, and enhancement of intestinal barrier function in zebrafish. L. rhamnosus alleviated the effects of PFOA and PFOS on SOD, CAT, and LDH levels, thus alleviating oxidative stress. L. rhamnosus regulated PFOA and PFOS-induced dysbiosis of intestinal flora in zebrafish. At the phylum level, Firmicutes and Actinobacteria abundance were increased, but Proteobacteria abundance was decreased in the zebrafish intestine. At the genus level, Leucobacter and Microbacterium increased, while deleterious genera such as Rhodobacter, Gordonia, and Rhodococcus decreased. Both PFOA and PFOS caused intestinal tissue damage. L. rhamnosus restored damaged intestinal villi and lymphocyte numbers and enhanced the intestinal barrier. These findings demonstrate that L. rhamnosus can alleviate the toxic effect of PFOA and PFOS in zebrafish, highlighting the potential application of probiotics in maintaining aquatic animal health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, G.L., S.C. and C.W.; methodology, G.L., S.C. and C.W.; software, G.L., S.C. and R.L.; validation, G.L., S.C. and M.W.; formal analysis, G.L., S.C. and C.W.; investigation, G.L., S.C., C.W. and J.L.; resources, S.C.; data curation, G.L. and S.C.; writing—original draft preparation, G.L. and S.C.; writing—review and editing, G.L., S.C. and C.W.; visualization, G.L., S.C. and M.Y.; supervision, G.L., S.C.,C.W. and Y.S.; project administration, G.L. and S.C.; funding acquisition, G.L. and S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Engineering Research Center of Solid-state Brewing, the National Natural Science Foundation of China (22278007), Research on countermeasures and key technologies for the governance and protection of the Yangtze River under the new situation (CX2021Z02) and the Opening Project of Key Laboratory of Environment Controlled Aquaculture (Dalian Ocean University) Ministry of Education (No. 202218).
Institutional Review Board Statement
All procedures are approved by the Animal Experiment Ethics Committee of Beijing Technology and Business University (Approval No. BTBU-2022-50) and conducted according to the guidelines for the protection of animal welfare.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within the article. The Supplementary material for this article can be found online at https://
Acknowledgments
We would like to thank the staff at our laboratory for their ongoing assistance.
Conflicts of Interest
The authors declare that they have no known competing financial interest or personal relationships that may have appeared to influence the work reported in this paper. The authors declare no conflict of interest.
References
- Melo, T.M.; Schauerte, M.; Bluhm, A.; Slany, M.; Paller, M.; Bolan, N.; Bosch, J.; Fritzsche, A.; Rinklebe, J. Ecotoxicological effects of per- and polyfluoroalkyl substances (PFAS) and of a new PFAS adsorbing organoclay to immobilize PFAS in soils on earthworms and plants. J. Hazard. Mater. 2022, 433, 128771. [Google Scholar] [CrossRef] [PubMed]
- Blum, A.; Balan, S.A.; Scheringer, M.; Trier, X.; Goldenman, G.; Cousins, I.T.; Diamond, M.; Fletcher, T.; Higgins, C.; Lindeman, A.E.; Peaslee, G.; de Voogt, P.; Wang, Z.; Weber, R. The Madrid Statement on Poly- and Perfluoroalkyl Substances (PFASs). Environ. Health. Persp. 2015, 123, A107–A111. [Google Scholar] [CrossRef] [PubMed]
- MacInnis, J.J.; French, K.; Muir, D.C.; Spencer, C.; Criscitiello, A.; De Silva, A.O.; Young, C.J. Emerging investigator series: a 14-year depositional ice record of perfluoroalkyl substances in the High Arctic. Environ. Sci-Proc. Imp. 2017, 19, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, L.; Bundschuh, M. Fate and Effects of Poly- and Perfluoroalkyl Substances in the Aquatic Environment: A Review. Environ. Toxicol. Chem. 2014, 33, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Campo, J.; Lorenzo, M.; Perez, F.; Pico, Y.; Farre, M.; Barcelo, D. Analysis of the presence of perfluoroalkyl substances in water, sediment and biota of the Jucar River (E Spain). Sources, partitioning and relationships with water physical characteristics. Environ. Res. 2016, 147, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Wu, P.; Wang, L.; Li, Q.; Li, X.; Luo, Y. Toxicity of per- and polyfluoroalkyl substances to aquatic vertebrates. Front. Env. Sci-Switz. 2023, 11, 37. [Google Scholar] [CrossRef]
- Vu, C.T.; Wu, T. Recent progress in adsorptive removal of per- and poly-fluoroalkyl substances (PFAS) from water/wastewater. Crit. Rev. Env. Sci. Tec. 2022, 52, 90–129. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S.; Abdel-Daim, M.M.; Van Doan, H. Probiotic application for sustainable aquaculture. Rev. Aquacult. 2019, 11, 907–924. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: a fish perspective. Fish. Shellfish. Immnol. 2010, 29, 2–14. [Google Scholar] [CrossRef]
- López Nadal, A.; Ikeda-Ohtsubo, W.; Sipkema, D.; Peggs, D.; McGurk, C.; Forlenza, M.; Wiegertjes, G.F.; Brugman, S. Feed, Microbiota, and Gut Immunity: Using the Zebrafish Model to Understand Fish Health. Front. Immunol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Chen, L.; Lam, J.C.W.; Tang, L.; Hu, C.; Liu, M.; Lam, P.K.S.; Zhou, B. Probiotic Modulation of Lipid Metabolism Disorders Caused by Perfluorobutanesulfonate Pollution in Zebrafish. Environ. Sci. Technol. 2020, 54, 7494–7503. [Google Scholar] [CrossRef]
- Sun, B.; Liu, M.; Tang, L.; Hu, C.; Huang, Z.; Chen, L. Probiotics inhibit the stunted growth defect of perfluorobutanesulfonate via stress and thyroid axes in zebrafish larvae. Environ. Pollut. 2021, 290, 118013. [Google Scholar] [CrossRef]
- Sun, B.; Liu, M.; Tang, L.; Hu, C.; Huang, Z.; Zhou, X.; Chen, L. Probiotic supplementation mitigates the developmental toxicity of perfluorobutanesulfonate in zebrafish larvae. Sci. Total. Environ. 2021, 799, 149458. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, X.; Ge, X.; Wang, D.; Wang, T.; Zhang, L.; Tanguay, R.L.; Simonich, M.; Huang, C.; Dong, Q. Chronic perfluorooctanesulphonic acid (PFOS) exposure produces estrogenic effects in zebrafish. Environ. Pollut. 2016, 218, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; He, M.Y.; Ling, Y.; Wang, X.J.; Zhang, F.; Feng, X.S.; Zhang, Y.; Xing, S.G.; Li, J.; Qiu, X.; Li, Y.R. Tissue distribution study of perfluorooctanoic acid in exposed zebrafish using MALDI mass spectrometry imaging. Environ. Pollut. 2022, 293, 118505. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2 ' s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Hematyar, N.; Rustad, T.; Sampels, S.; Dalsgaard, T.K. Relationship between lipid and protein oxidation in fish. Aquac. Res. 2019, 50, 1393–1403. [Google Scholar] [CrossRef]
- Liu, H.; Shi, L.; Giesy, J.P.; Yu, H. Polychlorinated diphenyl sulfides can induce ROS and genotoxicity via the AhR-CYP1A1 pathway. Chemosphere 2019, 223, 165–170. [Google Scholar] [CrossRef]
- Zheng, J.; Zhu, Q.; Wu, C.; Zhu, A.; Shen, B. Zinc acclimation mitigated high zinc induced oxidative stress by enhancing antioxidant defenses in large yellow croaker Pseudosciaena crocea. Aquat. Toxicol. 2016, 172, 21–29. [Google Scholar] [CrossRef]
- Huang, J.; Wang, Q.; Liu, S.; Lai, H.; Tu, W. Comparative chronic toxicities of PFOS and its novel alternatives on the immune system associated with intestinal microbiota dysbiosis in adult zebrafish. J. Hazard. Mater. 2022, 425, 127950. [Google Scholar] [CrossRef]
- Beauchamp, C.O.; Fridovich, I. Isozymes of superoxide dismutase from wheat germ. Biochim. Biophys. Acta. 1973, 317, 50–64. [Google Scholar]
- Son, V.M.; Chang, C.C.; Wu, M.C.; Guu, Y.K.; Chiu, C.H.; Cheng, W.T. Dietary administration of the probiotic, Lactobacillus plantarum, enhanced the growth, innate immune responses, and disease resistance of the grouper Epinephelus coioides. Fish. Shellfish. Immun. 2009, 26, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Miao, L.; Pan, W.; Huang, X.; Dengu, J.M.; Zhang, W.; Ge, X.; Liu, B.; Ren, M.; Zhou, Q.; Xie, J.; Pan, L.; Xi, B. Effect of nitrite exposure on the antioxidant enzymes and glutathione system in the liver of bighead carp, Aristichthys nobilis. Fish. Shellfish. Immun. 2018, 76, 126–132. [Google Scholar] [CrossRef]
- Dale, K.; Yadetie, F.; Muller, M.B.; Pampanin, D.M.; Gilabert, A.; Zhang, X.; Tairova, Z.; Haarr, A.; Lille-Langoy, R.; Lyche, J.L.; Porte, C.; Karlsen, O.A.; Goksoyr, A. Proteomics and lipidomics analyses reveal modulation of lipid metabolism by perfluoroalkyl substances in liver of Atlantic cod (Gadus morhua). Aquat. Toxicol. 2020, 227, 105590. [Google Scholar] [CrossRef] [PubMed]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Liu, C.; Yu, K.; Shi, X.; Wang, J.; Lam, P.K.; Wu, R.S.; Zhou, B. Induction of oxidative stress and apoptosis by PFOS and PFOA in primary cultured hepatocytes of freshwater tilapia (Oreochromis niloticus). Aquat. Toxicol. 2007, 82, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Budni, J.; Zomkowski, A.D.; Engel, D.; Santos, D.B.; dos Santos, A.A.; Moretti, M.; Valvassori, S.S.; Ornell, F.; Quevedo, J.; Farina, M.; Rodrigues, A.L. Folic acid prevents depressive-like behavior and hippocampal antioxidant imbalance induced by restraint stress in mice. Exp. Neurol. 2013, 240, 112–121. [Google Scholar] [CrossRef]
- Guedes, R.P.; Dal Bosco, L.; Araújo, A.S.; Belló-Klein, A.; Ribeiro, M.F.; Partata, W.A. Sciatic nerve transection increases gluthatione antioxidant system activity and neuronal nitric oxide synthase expression in the spinal cord. Brain. Res. Bull. 2009, 80, 422–427. [Google Scholar] [CrossRef]
- Kodali, V.P.; Sen, R. Antioxidant and free radical scavenging activities of an exopolysaccharide from a probiotic bacterium. Biotechnol J. 2008, 3, 245–251. [Google Scholar] [CrossRef]
- Serrano-Lorenzo, P.; Coya, O.N.; López-Jimenez, A.; Blázquez, A.; Delmiro, A.; Lucia, A.; Arenas, J.; Martín, M.A. Plasma LDH: A specific biomarker for lung affectation in COVID-19? Pract. Lab. Med. 2021, 25, e00226. [Google Scholar] [CrossRef]
- Fantin, V.R.; St-Pierre, J.; Leder, P. Attenuation of LDH-A expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell. 2006, 10, 172–172. [Google Scholar] [CrossRef]
- Venkateswara, R.J. Sublethal effects of an organophosphorus insecticide (RPR-II) on biochemical parameters of tilapia, Oreochromis mossambicus. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2006, 143, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Xia, X.; Zhou, D.; Wang, H.; Zhai, Y.; Lin, H.; Chen, J.; Hu, D. Bioconcentration and tissue distribution of shorter and longer chain perfluoroalkyl acids (PFAAs) in zebrafish (Danio rerio): Effects of perfluorinated carbon chain length and zebrafish protein content. Environ. Pollut. 2019, 249, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Kang, M.; Shen, Y.; Yun, L.; Yang, G.; Zhu, L.; Meng, X.; Zhang, J.; Su, X. Bacillus coagulans SCC-19 maintains intestinal health in cadmium-exposed common carp (Cyprinus carpio L.) by strengthening the gut barriers, relieving oxidative stress and modulating the intestinal microflora. Ecotox. Environ. Safe. 2021, 228, 112977. [Google Scholar] [CrossRef] [PubMed]
- Devi, M.; Fingerman, M. Inhibition of acetylcholinesterase activity in the central nervous system of the red swamp crayfish, Procambarus clarkii, by mercury, cadmium, and lead. Bull. Environ. Contam. Toxicol. 1995, 55, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Li, Z.; Yang, Y.; Jiao, Y.; Qu, J.; Wang, Y.; Zhang, Y. Effects of common environmental endocrine-disrupting chemicals on zebrafish behavior. Water. Res. 2022, 208, 117826. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Dasriya, V.L.; Nataraj, B.H.; Nagpal, R.; Behare, P.V. Lacticaseibacillus rhamnosus-Derived Exopolysaccharide Attenuates D-Galactose-Induced Oxidative Stress and Inflammatory Brain Injury and Modulates Gut Microbiota in a Mouse Model. Microorganisms 2022, 10, 2046. [Google Scholar] [CrossRef] [PubMed]
- Tarnecki, A.M.; Burgos, F.A.; Ray, C.L.; Arias, C.R. Fish intestinal microbiome: diversity and symbiosis unravelled by metagenomics. J. Appl. Microbiol. 2017, 123, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Anim. Nutr. 2020, 6, 69–79. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.; Tang, J.; Cai, J.; Yu, H.; Wang, Z.; Abarike, E.D.; Lu, Y.; Li, Y.; Afriyie, G. In vivo assessment of the probiotic potentials of three host -associated Bacillus species on growth performance, health status and disease resistance of Oreochromis niloticus against Streptococcus agalactiae. Aquaculture 2020, 527, 735440. [Google Scholar] [CrossRef]
- Pirarat, N.; Pinpimai, K.; Endo, M.; Katagiri, T.; Ponpornpisit, A.; Chansue, N.; Maita, M. Modulation of intestinal morphology and immunity in nile tilapia (Oreochromis niloticus) by Lactobacillus rhamnosus GG. Res. Vet. Sci. 2011, 91, E92–E97. [Google Scholar] [CrossRef] [PubMed]
- Roshchina, V.V. New trends and perspectives in the evolution of neurotransmitters in microbial, plant, and animal cells. Adv. Exp. Med. Biol. 2016, 874, 25–77. [Google Scholar] [PubMed]
- Tian, X.; Yu, Z.; Feng, P.; Ye, Z.; Li, R.; Liu, J.; Hu, J.; Kakade, A.; Liu, P.; Li, X. Lactobacillus plantarum TW1-1 Alleviates Diethylhexylphthalate-induced testicular damage in mice by modulating gut microbiota and decreasing inflammation. Front. Cell. Infect. Microbiol. 2019, 9, 221. [Google Scholar] [CrossRef]
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Baumler, A.J. Dysbiotic Proteobacteria expansion: a microbial signature of epithelial dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Ran, C.; Wang, Y.; Zhang, Z.; Ding, Q.; Yang, Y.; Olsen, R.E.; Ringo, E.; Bindelle, J.; Zhou, Z. Use of probiotics in aquaculture of China-a review of the past decade. Fish. Shellfish. Immun. 2019, 86, 734–755. [Google Scholar] [CrossRef] [PubMed]
- James, G.; Prasannan Geetha, P.; Thavarool Puthiyedathu, S.; Vattringal Jayadradhan, R.K. Applications of Actinobacteria in aquaculture: prospects and challenges. 3 Biotech. 2023, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.A.; Petty, B.D. Mixed Metazoan and Bacterial Infection of the Gas Bladder of the Lined Seahorse-A Case Report. J. Aquat. Anim. Health. 2013, 25, 42–52. [Google Scholar] [CrossRef]
- Zhan, M.; Huang, Z.; Cheng, G.; Yu, Y.; Su, J.; Xu, Z. Alterations of the Mucosal Immune Response and Microbial Community of the Skin upon Viral Infection in Rainbow Trout (Oncorhynchus mykiss). Int. J. Mol. Sci. 2022, 23, 14037. [Google Scholar] [CrossRef]
- Tinh, N.T.; Dierckens, K.; Sorgeloos, P.; Bossier, P. A review of the functionality of probiotics in the larviculture food chain. Mar. Biotechnol. 2008, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, P.; Kunze, W.A.; Bienenstock, J. On communication between gut microbes and the brain. Curr. Opin. Gastroen. 2012, 28, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Falcinelli, S.; Picchietti, S.; Rodiles, A.; Cossignani, L.; Merrifield, D.L.; Taddei, A.R.; Maradonna, F.; Olivotto, I.; Gioacchini, G.; Carnevali, O. Lactobacillus rhamnosus lowers zebrafish lipid content by changing gut microbiota and host transcription of genes involved in lipid metabolism. Sci. Rep. 2015, 5, 1–11. [Google Scholar]
- Sun, H.; Zhang, A.; Song, Q.; Fang, H.; Liu, X.; Su, J.; Yang, L.; Yu, M.; Wang, X. Functional metabolomics discover pentose and glucuronate interconversion pathways as promising targets for Yang Huang syndrome treatment with Yinchenhao Tang. Rsc. Adv. 2018, 8, 36831–36839. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, H.; Ren, Z.; Cui, Z. The toxic effects of deltamethrin on Danio rerio: the correlation among behavior response, physiological damage and AChE. Rsc. Adv. 2016, 6, 109826–109833. [Google Scholar] [CrossRef]
Figure 1.
SOD and CAT levels in each treatment group. (a), (e) for intestine; (b), (f) for liver; (c), (g) for brain; (d), (h) for gills. Data are shown as mean ± SD (n = 4). Columns with different letters indicate values with significant differences (P< 0.05).
Figure 1.
SOD and CAT levels in each treatment group. (a), (e) for intestine; (b), (f) for liver; (c), (g) for brain; (d), (h) for gills. Data are shown as mean ± SD (n = 4). Columns with different letters indicate values with significant differences (P< 0.05).
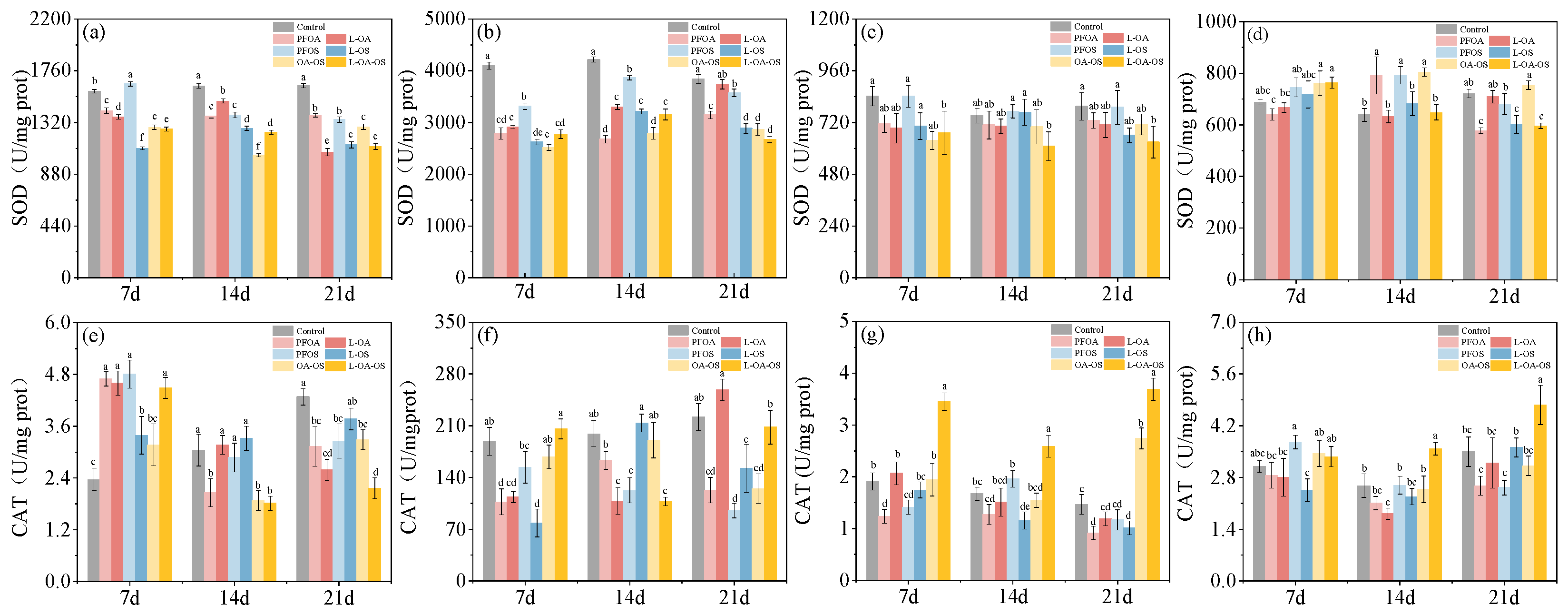
Figure 2.
MDA and GSH levels in each treatment group. (a), (e) for intestine; (b), (f) for liver; (c), (g) for brain; (d), (h) for gills. Data are shown as mean ± SD (n = 4). Columns with different letters indicate values with significant differences (P< 0.05).
Figure 2.
MDA and GSH levels in each treatment group. (a), (e) for intestine; (b), (f) for liver; (c), (g) for brain; (d), (h) for gills. Data are shown as mean ± SD (n = 4). Columns with different letters indicate values with significant differences (P< 0.05).
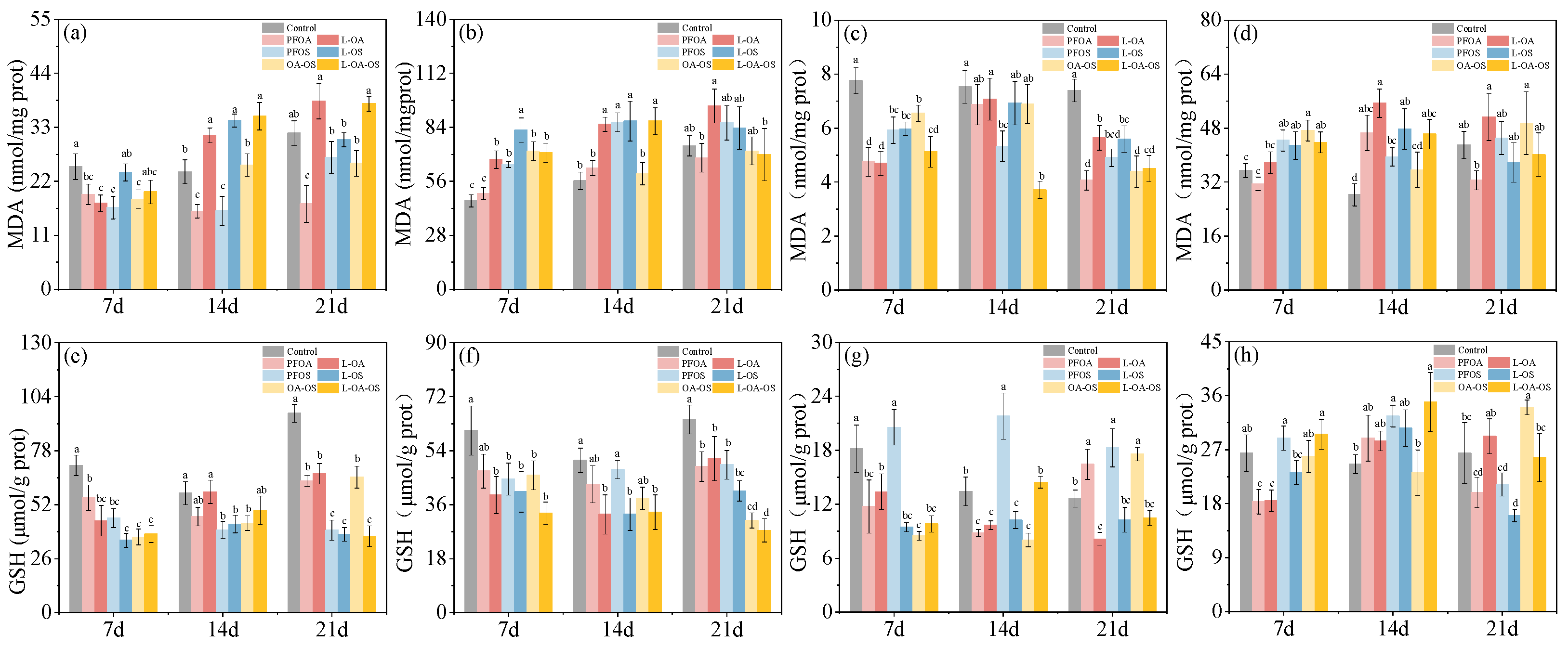
Figure 3.
LDH and AChE levels in each treatment group. (a), (e) for intestine; (b), (f) for liver; (c), (g) for brain; (d), (h) for gills. Data are shown as mean ± SD (n = 4). Columns with different letters indicate values with significant differences (P< 0.05).
Figure 3.
LDH and AChE levels in each treatment group. (a), (e) for intestine; (b), (f) for liver; (c), (g) for brain; (d), (h) for gills. Data are shown as mean ± SD (n = 4). Columns with different letters indicate values with significant differences (P< 0.05).
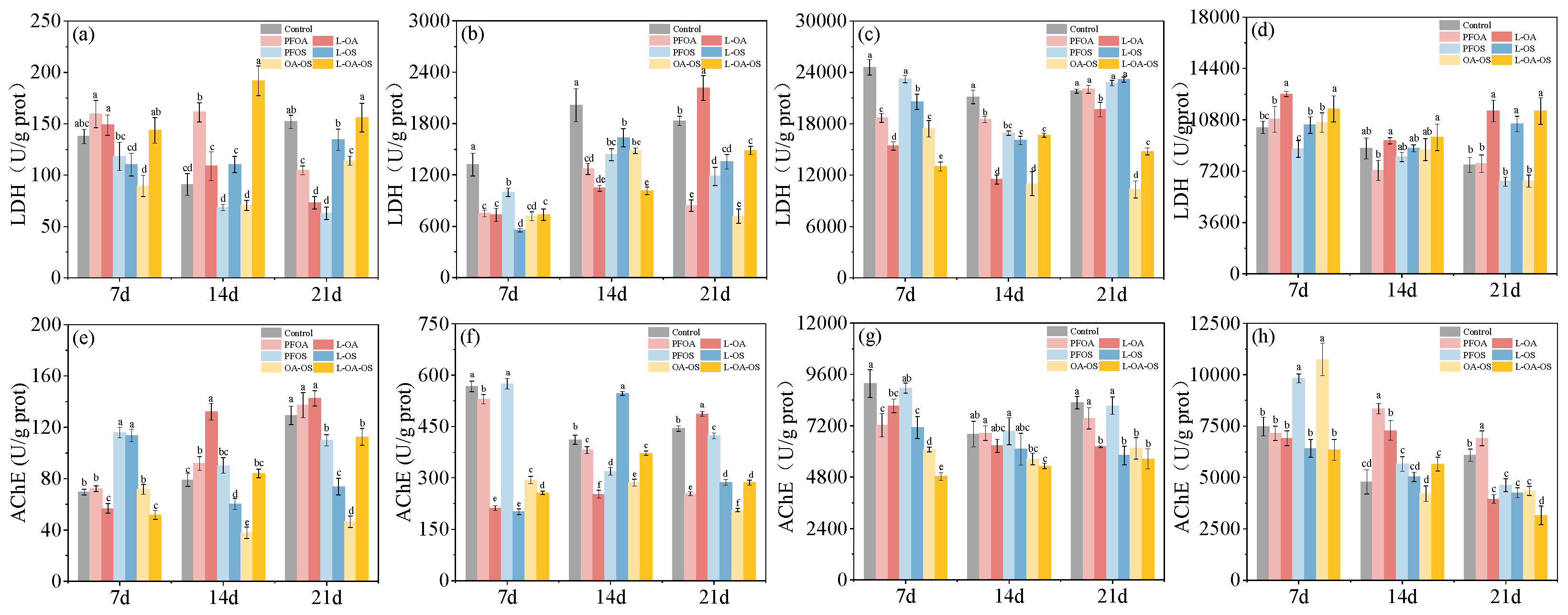
Figure 4.
21-day intestinal tissue sections of each treatment group. (a) Control group, (b) PFOA group, (c) L-OA group, (d) PFOS group, (e) L-OS group, (f) OA-OS group, (g) L-OA-OS group.
Figure 4.
21-day intestinal tissue sections of each treatment group. (a) Control group, (b) PFOA group, (c) L-OA group, (d) PFOS group, (e) L-OS group, (f) OA-OS group, (g) L-OA-OS group.
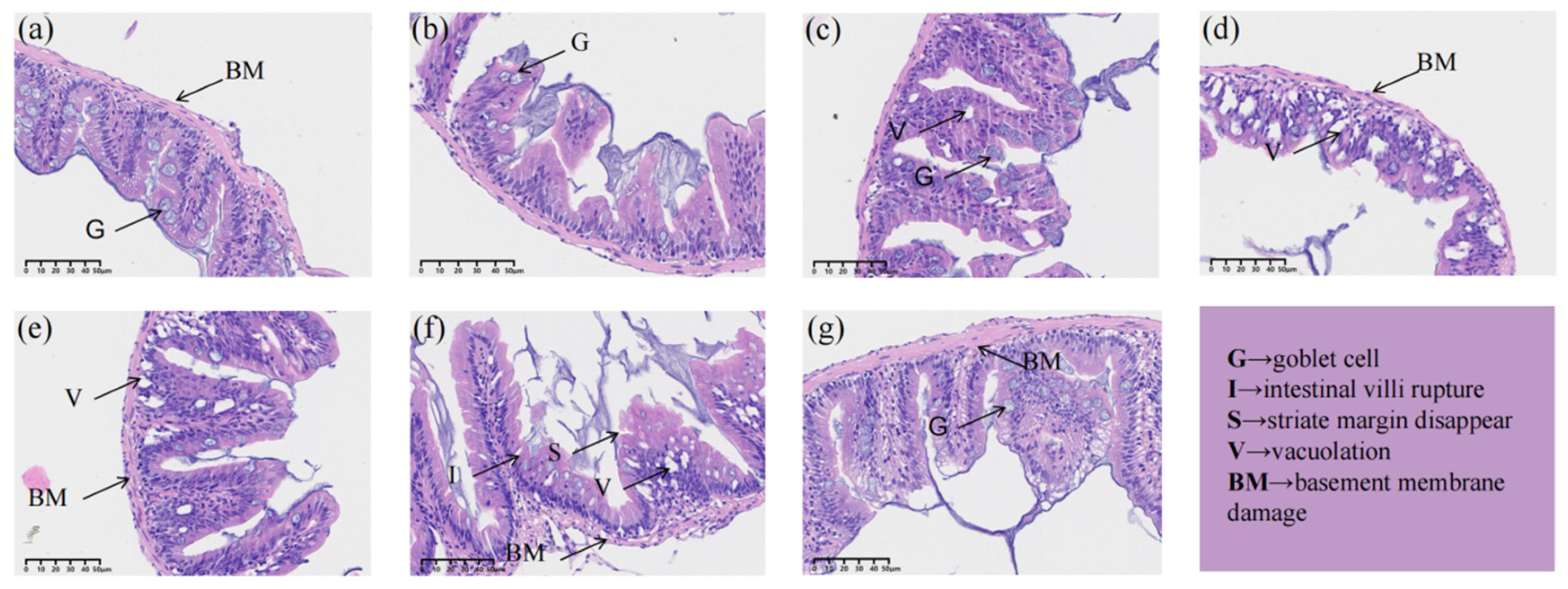
Figure 5.
Effect of different treatments on the community structure of the intestinal flora of zebrafish. (a) Chao1, (b) Shannon and (c) Simpson indices showing the diversity of intestinal flora under different treatments. (d) PCoA reveals the differences in community structure between treatments.
Figure 5.
Effect of different treatments on the community structure of the intestinal flora of zebrafish. (a) Chao1, (b) Shannon and (c) Simpson indices showing the diversity of intestinal flora under different treatments. (d) PCoA reveals the differences in community structure between treatments.
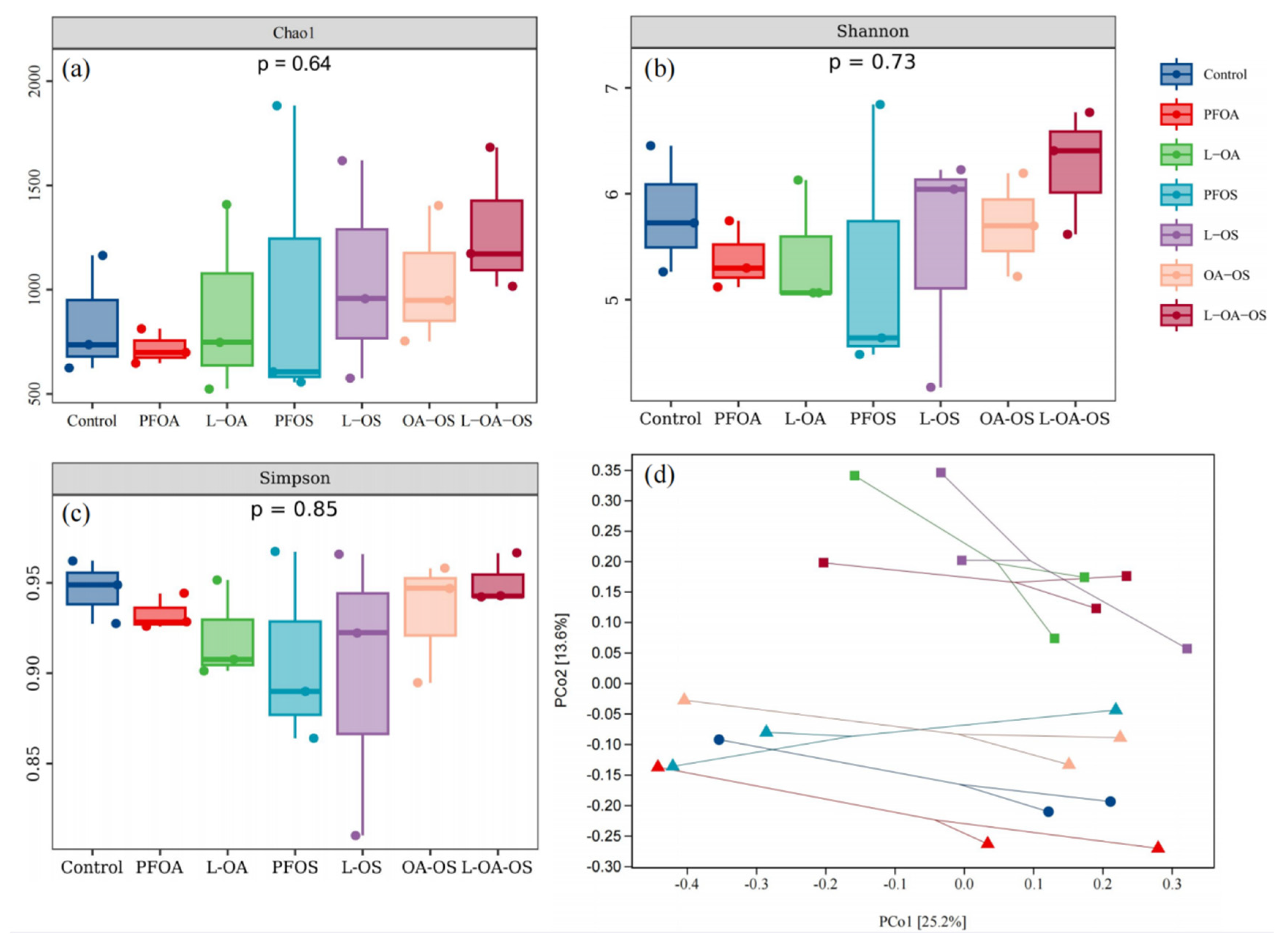
Figure 6.
Abundance of bacterial composition of different communities. (a) represents the phylum level, and (b) represents the genus level.
Figure 6.
Abundance of bacterial composition of different communities. (a) represents the phylum level, and (b) represents the genus level.
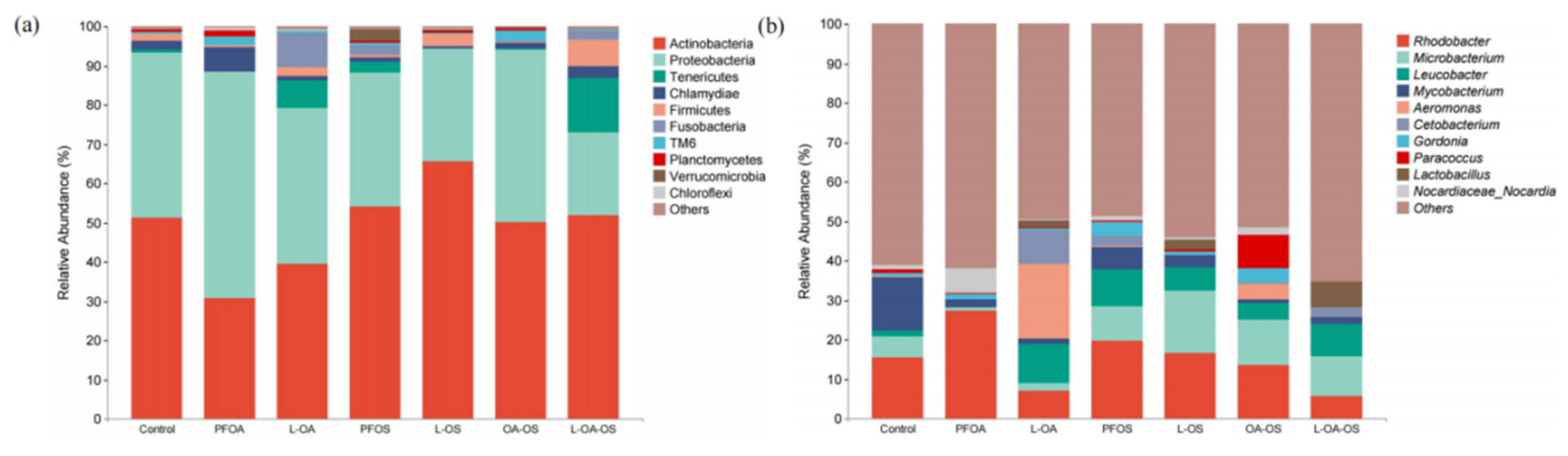
Figure 7.
MetaCyc pathway analysis. (a) Predicted changes in metabolic activity of the gut microbiota (Primary pathway) (b) Secondary pathways (c) control vs PFOA (d) PFOA vs L-OA (e) species composition of the PWY-7315 metabolic pathway.
Figure 7.
MetaCyc pathway analysis. (a) Predicted changes in metabolic activity of the gut microbiota (Primary pathway) (b) Secondary pathways (c) control vs PFOA (d) PFOA vs L-OA (e) species composition of the PWY-7315 metabolic pathway.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



