
Submitted:
27 October 2023
Posted:
30 October 2023
You are already at the latest version
Abstract
Influenza is an acute respiratory illness caused by influenza virus infection, which is managed using vaccines and antiviral drugs. Recently, the antiviral effects of plants and foods have gained attention. This review summarizes the therapeutic properties of Euglena against influenza from a functional food viewpoint. While it has been reported that Euglena contributes to suppressing blood sugar levels and ameliorates symptoms caused by stress by acting on the autonomic nervous system, the immunostimulatory and antiviral activities of Euglena have also been reported. In this review, I focused on the immunostimulation of antiviral activity via the intestinal environment and the suppression of viral replication in infected cells. The functions of specific components of Euglena, which also serve as a source of a wide range of nutrients such as vitamins, minerals, amino acids, unsaturated fatty acids, and β-1,3-glucan (paramylon), are also reviewed. Euglena has animal and plant properties and natural compounds with a wide range of functions, providing crucial information for improved antiviral strategies.
Keywords:
Euglena
; immunostimulation
; antiviral activity
; b-1
; 3-glucan
; paramylon
; minerals
; influenza virus
1. Introduction
Sudden outbreaks of viruses, such as the recent outbreak of SARS-CoV2, have occurred regularly. Particularly, influenza, a respiratory tract infection caused by influenza viruses, is a disease far more serious than the common cold syndrome. Although diverse vaccines and anti-influenza drugs are clinically used yearly, adapting the annual vaccines to the circulating influenza viruses prevalent during the influenza season is challenging. In addition, with the emergence of drug-resistant viruses, new approaches to treat influenza must be considered. Moreover, the risk of spread of infection through droplet and airborne transmission increases during disasters, especially in evacuation centers. Under these circumstances, it is reasonable that daily food ingredients can be incorporated into our daily lives to improve our physical condition, prevent diseases, and alleviate certain symptoms.
Furthermore, carbohydrates, proteins, and fats consumed daily are digested to produce cellular energy. Vitamins and minerals are indispensable coenzymes for this process; however, it is challenging for most people to consume sufficient amounts of these vitamins and minerals in their diets. To fundamentally reduce the gap between a healthy life span and expectancy, it is essential to treat the symptoms and disorders symptomatically and maintain homeostasis in the body and mind for they not to occur.
According to the Dietary Intake Standards for Japanese (2020 edition) [1], the target daily intake of dietary fiber is 21 g or more for men aged 18 and older (20 g or more for those aged 65 and older) and 18 g or more for women (17 g or more for those aged 65 and older). However, the dietary fiber intake is below the target for men and women across all age groups [1]. One source of dietary fiber that is attracting attention is β-glucan, a non-starch polysaccharide composed of D-glucose linked by beta-glycosidic bonds. The cell walls of various organisms are made of β-glucan. Therefore, we unintentionally consume β-glucan daily. Typical organisms from which β-glucan is derived include cereals (especially oats and barley), mushrooms, seaweeds, and yeast, with the structure differing depending on the source. In addition, the intake of β-glucan is not only a dietary fiber supplement but also has various functions. The functions of β-glucan have been recognized for more than half a century, and the effects of its intake on the human body include immunostimulatory [2,3,4,5], antiviral [6], regulation of intestinal microflora [7], lowering of cholesterol levels [2,8,9], prevention of diabetes [10], antioxidant effects [2,8,11,12,13], and anti-tumor [2,3,14,15,16,17,18]. The differences in the physiological functions of β-glucan may depend on its structure, including the composition of the glucan backbone, type and frequency of side chains, molecular size, solubility, and steric structure.
Euglena is a source of β-glucan that has gained attention recently. Euglena is a microalga of approximately 0.05 mm in length and is characterized by its chlorophyll, photosynthesis, and use of flagella to perform cell deformation movements called swimming and Euglena movements. Because of its plant and animal characteristics, it contains a wide range of nutrients such as vitamins, minerals, amino acids, and unsaturated fatty acids (Table 1) [19,20,21,22]. It also contains nutrients such as paramylon [19,21,23], GABA (gamma-aminobutyric acid) [23], chlorophyll [24], lutein [25], zeaxanthin [25], spermidine [26], and putrescine [26]. Euglena utilizes a unique β-1,3-glucan called paramylon as a storage polysaccharide, making it one of the main components of Euglena as a food since it represents 30 to 80% of its dry weight, depending on the environment and culture method. As mentioned above, there are various types of β-glucans; however, paramylon is characterized by its linear and non-branched β-1,3-glucan structure [27,28], high crystal structure with a triple-helical structure [29,30], unique sugar composition (glucose only) [31], and insolubility in water.
The intake of Euglena may contribute to the suppression of high blood glucose levels [32] and the alleviation of stress-related symptoms [33] via autonomic nervous system balance; nonetheless, the effect of Euglena on the immunological and antiviral activities has not been studied in detail. Moreover, the active ingredients of Euglena, which contains a wide range of nutrients, have not been thoroughly analyzed. Unlike β-glucans from other sources, Euglena also serves as a broad source of vitamins, minerals, amino acids, unsaturated fatty acids, and other nutrients, and thus may exhibit a wide range of mechanisms of action. In this review, I focused on the immunostimulation of antiviral activity and the possible contribution from the intestinal environment, which is one of the mechanisms of immunostimulation. Moreover, the effects of Euglena's intake, focusing on its components and effects, are also described.
2. Effects of Euglena intake on allergic diseases
Allergic rhinitis (AR) is the most common allergic disease and represents a health problem worldwide. In Japan, the number of patients with AR has increased. Recently, the number of patients with pollinosis, particularly Japanese cedar pollinosis (JCP), has markedly increased beyond that associated with house dust mites (HDM) or pollinosis other than JCP. An epidemiological study revealed a marked increase (approximately 10%) in the prevalence of AR between 1998 and 2008 [34]. JCP is an immediate-type (type I) allergic disease that causes allergic symptoms due to a specific reaction with IgE antibodies. Cry j1 and Cry j2 in the pollen have been identified as the major antigens (allergens) causing JCP [35,36]. Although immunotherapy and drug therapy are available as treatments for JCP, there is still no effective cure. Recently, there have been reports that functional foods suppress allergic symptoms [37,38,39], raising the expectations for functional foods.
Helper T cells (Th), which are classified into Th1 and Th2 according to their cytokine production patterns, are involved in the pathogenesis of allergy [40]. Th1 participates in cellular immunity and secretes cytokines such as IL-2 and IFN-γ. Th2 is involved in humoral immunity and secretes cytokines such as IL-4 and IL-5. When Th1 and Th2 are imbalanced, allergies are thought to be triggered. For example, Th1 predominates in delayed (type IV) allergic development and Th2 in type I allergic development. Therefore, pollinosis is thought to develop when Th2 predominates.
Euglena is attracting attention as a new functional food, and the paramylon (β-1,3-D-glucan) contained in Euglena has immunostimulatory effects involving cytokines [41], hepatoprotective effects on acute liver injury [42], and anti-human immunodeficiency virus (HIV) [43], and antibacterial effects [44]. Furthermore, Sugiyama et al. [45] indicate that paramylon treatment could provide an effective alternative therapy for the management of atopic dermatitis (AD). Oral administration of paramylon was suggested to suppress the development of atopic dermatitis in NC/Nga mice, which spontaneously develop an eczematous AD-like skin lesion when kept under conventional care but not under specific pathogen-free (SPF) conditions [46,47], by inhibiting Th1 and Th2 responses [45]. The effect of Euglena and palamylon were observed in the early stages of A4gnt KO mice, which are mutant models that spontaneously develop gastric cancer through hyperplasia-dysplasia-adenocarcinoma mechanisms in the antrum of the stomach. The results suggest that the administration of Euglena and paramylon may ameliorate the early involvement of A4gnt mice through inflammatory modulation in the gastric mucosa [48]. Amorphous paramylon had a greater effect on intestinal immunity than that of paramylon, inhibiting colon cancer [49].
The production of allergen-specific IgE was significantly suppressed, and the production of IL-12 and IFN-γ increased when low-molecular-weight β-glucan was administered to mice [50]. It was also found that low-molecular-weight lentinan-suppressed allergic symptoms, such as seasonal and perennial rhinorrhea, sneezing, nasal obstruction, itching, tearing in humans, and allergen-specific IgE and total IgE levels [51], indicating that the β-1,3-1,6-D-glucan found in mushrooms and yeast has immunomodulatory and allergy-suppressing effects. Although a decrease in serum IgE concentration was reported in atopic dermatitis-induced mice after the oral administration of paramylon, no decrease in IgE was observed in a pollinosis model mouse created by inoculation with Cry j1, because the hypersensitivity response to externally introduced specific antigens is biased toward Th2 dominance [52]. Furthermore, Euglena intake may directly reduce pollinosis symptoms, suggesting that components other than paramylon also relieve pollinosis [52].
3. Effects of Euglena's intake on the intestinal microbiota and defecation
The intestinal tract hosts the gut microbiota, a complex bacterial community. The gut microbiota interacts with the host and strongly influences homeostasis and immunity in the host. Therefore, the gut microbiota is essential for maintaining the health of the host [53,54,55]. There is growing interest in optimizing the composition of the gut microbiota through dietary therapy using functional foods containing probiotics [56] or prebiotics [57].
For example, β-glucans in cereals are fermented by microorganisms living in the large intestine, and they are converted into short-chain fatty acids (SCFA), such as acetic, propionic, and butyric acids [58]. SCFAs produced in the colon exert various effects, including immunomodulation [59], mediating apoptosis of colon cancer cells [60], and preventing obesity [61]. For example, the prevention of obesity is achieved by regulating energy metabolism via the SCFA receptor GPR43 and preventing the accumulation of excess lipids in adipose tissue [61]. In addition, SCFAs inhibit the growth of harmful bacteria such as Clostridium spp. and pathogenic Escherichia coli, thereby maintaining a healthy intestinal microflora [62].
The modulatory effects of diet on the gut microbiota are often investigated through in vivo studies in humans [63]. Information on the composition of the colon microbiota comes primarily from the analysis of fecal samples in human dietary intervention studies. However, this method has the experimental limitation that the production of certain metabolites, such as short-chain fatty acids (SCFA), cannot be measured in situ (at the intestinal tract). A model culture system was developed to rigorously reproduce the microbial components of human fecal collections in vitro [64]. This in vitro human colon microbiota model was used to detect the decreased butyrate production in patients with ulcerative colitis [64]. Thus, combining in vitro human colon microbiota models with in vivo studies can help interpret changes in the human gut microbiota.
Since the effects of Euglena’s ingestion on the human gut microbiota are not yet clear, this section evaluates the effects of Euglena on the colon microbiota of healthy human subjects. Furthermore, by analyzing the effects of adding Euglena or paramylon to an in vitro human colon microbiota model, verifying the effects of Euglena on the gut microbiota is feasible.
Studies on the effects of Euglena’s intake on gut microbiota and defecation showed that the occupancy of the genus Faecalibacterium was increased by Euglena in vitro and in vivo [65]. However, this effect may not be due to the paramylon contained in Euglena [65]. The only known species in the genus Faecalibacterium, F. prausnitzii, is one of the butyrate-producing bacteria with the highest occupancy in the intestinal tract [66]. Other butyrate-acid-producing bacteria, such as Roseburia, showed no differences in vitro and in vivo. Thus, the changes in the gut microbiota induced by Euglena may be specific to Faecalibacterium. Previous studies reported that butyrate improves insulin resistance [67], and that increasing the amount of butyrate-producing bacteria may be useful to treat type 2 diabetes [68]. Such results of increased butyrate production induced by Euglena are discussed with the results of a study showing that Euglena consumption lowered blood glucose levels in a rat model of type 2 diabetes [69], indicating that butyrate production by the gut microbiota may be one of the mechanisms by which blood glucose levels are reduced. Euglena is also a source of vitamins, minerals, and unsaturated fatty acids [70,71]. These components promote the growth of Faecalibacterium. Our previous study showed that ingestion of Euglena increased bifidobacteria in the intestinal microflora of mice [72]. However, in another experiment, there was no significant increase in the relative occupancy of bifidobacteria in vitro or in vivo. Therefore, Euglena consumption may promote acetic acid production by Bifidobacterium, which is then consumed by Faecalibacterium, promoting Bifidobacterium growth and butyrate production. In Bifidobacterium adolescentis and F. prausnitzii, a cross-feeding process has been previously reported using fructooligosaccharides as a carbon source [73].
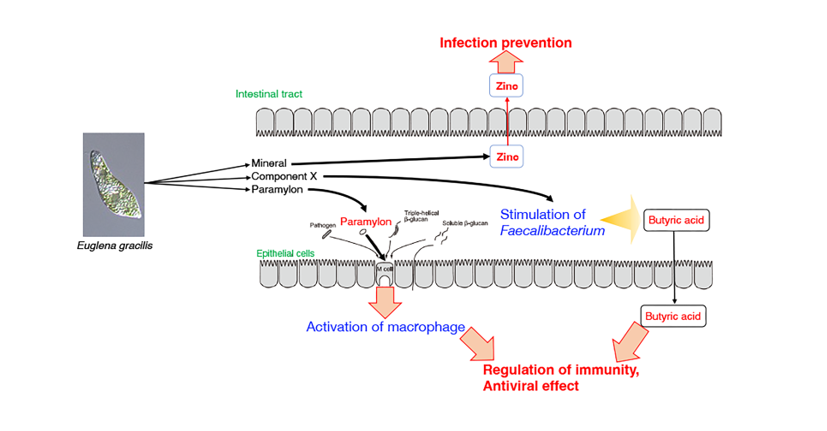
Furthermore, stool volume was increased by the ingestion of Euglena [65], as shown by Asayama et al. [74]. These results are consistent with the findings of Kawano et al. that rats fed a diet containing cholesterol and Euglena had a shorter cholesterol retention time in the intestine than rats fed a diet containing only cholesterol [75]. Dietary fiber intake may increase stool frequency in patients with constipation [76]. The paramylon in Euglena is insoluble and is neither digested nor absorbed. Therefore, it is believed to exert the same effect as a dietary fiber. Moreover, the consumption of Euglena enhanced butyrate production by F. prausnitzii and is consistent with the results of a previous study showing that butyrate supplementation may alleviate defecation disorders [77]. Thus, ingesting Euglena may have beneficial effects on constipation, such as reducing pain during defecation. In addition, butyrate produced in the intestine increases the number of specific CD8+ cells that eliminate influenza viruses and control the infection [78]. These findings indicate Euglena may contribute to immunomodulation and antiviral activity (Figure 1).
Overall, these findings suggest that Euglena increases the occupancy of Faecalibacterium, which in turn promotes butyrate production (Figure 1). This could be the mechanism by which Euglena improves stool volume. One limitation of these studies is the small sample size for the in vivo analysis. Therefore, an increase in sample size is warranted in subsequent studies to validate the results. Further experiments are also needed to identify the effect of components other than paramylon in Euglena. Thus, Euglena has great potential as a novel prebiotic.
4. Effect of Euglena intake on symptoms of influenza virus infection
Influenza is an acute respiratory illness caused by the influenza virus [79,80]. Influenza increases in winter and is a serious social and economic problem in many countries. Influenza virus infection causes multiple systemic symptoms, including fever over 38 °C, headache, arthralgia, myalgia, and fatigue. Typically, healthy adults recover spontaneously without antiviral treatment through self-healing mechanisms involving the immune response. However, in the elderly, infants, or patients with an underlying respiratory disease, impaired immune function can exacerbate symptoms due to viral infection and, in the worst case, pneumonia or encephalitis, which can be fatal. Because influenza viruses are segmental-stranded RNA viruses, gene replication and reassortment are frequent. Therefore, vaccination does not provide adequate protection against influenza virus infection.
In the previous section, it was shown that Euglena consumption contributes to the regulation of butyrate-producing bacteria occupancy in the gut and may contribute to antiviral activity by stimulating butyrate production; however, the effect of Euglena or paramylon on influenza virus infection is unknown.
his section reviews the alleviating effects of Euglena and paramylon on influenza virus infection symptoms in mice based on survival, lung virus titer, and cytokine production.
Euglena or paramylon administration prevented a decrease in survival after influenza virus infection. High lung virus titers and/or abnormal production of inflammatory cytokines frequently occur with the progression of influenza virus infection, which are associated with the severity of the morbidity [81,82,83,84]. Therefore, lung homogenates from infected mice were used to measure lung viral titers and cytokine production. On day 1 post-infection, pulmonary viral titers were similar in the Euglena and paramylon-fed groups. However, the paramylon-treated group showed lower viral titers on day 2 compared to those of the Euglena-treated group. The production of IL-1β, IFN-β, IFN-γ, and TNF-α was higher in the Euglena group than those in the control group. Significant increases in IL-1β, IL-6, IL-12, IL-10, IFN-γ, and TNF-α were detected in the paramylon-treated group, and increased production of IFN-β was observed. These results suggest that paramylon is one of the active substances that alleviate influenza virus infection symptoms. Paramylon is a β-1,3-glucan, and the Euglena used in this experiment contained approximately 30% paramylon.
Recent immunological studies have shown that influenza virus infection induces the production of type I interferons such as INF-β, leading to acute inflammation of the lungs, after which INF-β contributes to viral elimination via induction of NK and CD8+ T cells [85,86]. It has also been reported that β-1,3-glucan enhances NK cell activity in influenza virus-infected mice, increases IL-1β, TNF-α, and IFN-γ levels in the lungs [87,88], activates type I interferon, and induces CD8+ T cells via Dectin-1 in dendritic cells [89]. Furthermore, the proliferation, deviation, and activation of NK cells are induced by the IFN, TNF-α, and IL-1 secreted by dendritic cells and macrophages [90,91,92].
Generally, Dectin-1, a C-type lectin-like pattern recognition receptor on the surface of leukocytes, is the primary receptor for β-glucan [93]. The triple-helix structure formed by the backbone of β-1,3-glucan is specifically recognized by Dectin-1. As mentioned above, the crystal structure of paramylon consists of a triple helix [94,95], which is recognized by the β-glucan receptor Dectin-1, and it is presumed that paramylon activates Dectin-1. Moreover, the size of paramylon is typically 2-3 μm, which is comparable to the size of pathogenic bacteria. This suggests paramylons may pass through epithelial cells using the same mechanism as pathogenic bacteria. Indeed, consistent with the above data, paramylon has been shown to bind directly to the recombinant Dectin-1 [96] and upregulate inflammatory factors such as NO, TNF-α, IL-6, and COX-2 [97]. Studies have shown that the number and size of β-1,6 branches on the β-1,3 skeleton are crucial for the function of β-glucans [98,99]; however, despite its limited number of branches, paramylon shows functional diversity. In the results of the study by Nakashima et al. [100], mice were fed the same amount of Euglena and paramylon; however, the survival rate and viral titer in the lungs results tended to be better in the paramylon-fed than in the Euglena-fed group. However, the Euglena-fed group did not show the same results as those of the paramylon-fed group. In particular, the Euglena-fed group showed a different pattern of cytokine production, suggesting the involvement of components other than paramylon in Euglena treatment. These results indicate that Euglena may prevent influenza virus infection not only through the actions of components other than paramylon in the intestinal microflora but also through systemic immune regulation, mainly through the contribution of paramylon in Euglena.
Oral intake of Euglena and paramylon significantly reduced pulmonary viral titers and increased survival. In addition, paramylon induced significant increases in cytokine levels in the lungs, including those of IL-1β, IL-6, IL-12, IL-10, IFN-γ, and TNF-α (Figure 2). These results are consistent with those reported in the literature on the effects of β-glucan in mice infected with influenza. However, the pattern of cytokine production in the Euglena-fed group did not completely match that of the paramylon-fed group, suggesting the involvement of components other than the paramylon contained in Euglena. The oral intake of Euglena and paramylon can eliminate influenza viruses, mainly through the activity of β-1,3-glucan on dendritic cells and induction of CD8+ T and/or NK cells (Figure 2). We believe that further studies are needed to better understand the detailed functions of Euglena and paramylon and to enable influenza prevention, possibly through their direct effects on the viral replication cycle.
5. Effect of Euglena on the cellular infection with influenza virus
Influenza is an infectious respiratory tract disease caused by A, B, or C influenza viruses. Type A and C are the most and least common, respectively. Influenza A and B viruses, the most prevalent types, comprise two surface glycoproteins, hemagglutinin (HA) and neuraminidase (NA), which are antigens the host targets for protective immunity. The antigenic properties of these viruses differ depending on the combination of HA and NA subtypes. Viruses with different combinations of these glycoproteins are widely distributed in humans and other hosts such as pigs and birds. Even for the same subtype, the antigenic properties of HA and NA gradually change due to the accumulation of viral gene mutations. Influenza viruses are endemic every year because of frequent antigenic mutations. Generally, after an incubation period of 1-3 days after influenza virus infection, symptoms such as fever (usually higher than 38 °C), headache, general malaise, and muscle and joint pain suddenly occur. This is followed by upper respiratory tract inflammatory symptoms such as cough and nasal discharge, and symptoms usually abate after one week. Particularly, the elderly, patients with chronic diseases of the respiratory, cardiovascular, and renal systems, with metabolic diseases such as diabetes, and patients of any age with compromised immune function are more susceptible to secondary bacterial infections of the respiratory system, which can worsen the original disease and increase the risk of hospitalization or death. In children, influenza can also cause otitis media, febrile convulsions, and bronchial asthma. Currently, vaccines are used to prevent influenza virus infection, and antiviral drugs are used for treatment. However, there are concerns about the side effects of adjuvants contained in existing influenza vaccines, the inability to respond rapidly to new viruses, and the fact that these vaccines are less effective in prevention than vaccines against other viruses. Furthermore, the side effects of antiviral drugs are equally problematic, as is the emergence of drug-resistant bacteria [101,102,103,104]. Thus, there is a need to develop new preventive and therapeutic methods to overcome these issues. In light of this, we have focused on the functional properties of foods and investigated the mechanisms of action of components with antiviral activity. The previous section mentioned that mice fed a diet containing Euglena powder for 2 weeks and subsequently infected with the A/Puerto Rico/8/34 (H1N1) strain of influenza virus showed improved survival and reduced virus counts in the lungs [100]. Based on the pattern of cytokine production in these mice, it was speculated that paramylon, which is mainly contained in Euglena, may have contributed to the elimination of the virus by mobilizing the systemic immune system. However, the antiviral effect of Euglena may have mechanisms other than systemic immunity. In fact, the cytokine production pattern of the Euglena-fed group in the in vivo experiment described in the previous section did not completely match that of the paramylon-fed group, suggesting that components other than the paramylon contained in Euglena may be involved. The direct mechanism by which Euglena suppresses viruses in cells has not yet been elucidated. In vitro experiments are necessary to examine the direct effect; however, the paramylon contained in Euglena is an insoluble β-glucan that is difficult to add to the culture medium, making it difficult to conduct in vitro experiments. Therefore, this section aims to clarify the antiproliferative effect of Euglena hot water extract on the influenza virus and its mechanism.
The anti-influenza effect of Euglena hydrothermal extract was evaluated using MDCK cells infected with various strains of influenza virus, and it was confirmed that Euglena hydrothermal extract inhibited the growth of all virus strains. Particularly, the results showed that the Euglena hot water extract was effective against oseltamivir-resistant virus strains such as A/Osaka/2024/2009 and A/Osaka/71/2011, suggesting that it inhibits the growth of influenza viruses by a mechanism different from that of oseltamivir [105]. In addition, no significant difference in IC50 values was observed among the virus strains examined. Therefore, the virus replication inhibitory activity of the Euglena hot water extract does not show virus specificity [105]. This differs from amantadine, which is effective against type A influenza virus strains but ineffective against type B strains [106].
Once the virus attaches to the cell membrane, it is endocytosed, and RNA is released from the viral particles into the cytoplasm, where it is transferred to the nucleus for replication and transcription, followed by the synthesis of viral proteins and the viral genome. Once the virus components are in place, the budding phase of the viral growth cycle occurs when the viral particles aggregate near the cell membrane and are released via neuraminidase activity. One growth cycle lasts approximately eight hours, and studying the inhibition of the viral process is possible. "Relenza" and "Tamiflu," which are the mainstream anti-influenza drugs, inhibit the budding phase of the virus by blocking neuraminidase activity [107], whereas the recently launched "Xofluza" inhibits viral RNA replication [108]. Although the mechanism of inhibition by Euglena hydrothermal extract has not yet been predicted, Euglena hydrothermal extract reduced the viral titer by pretreatment or prolonged treatment of infected cells. The antiviral effects of the Euglena hydrothermal extract remained potent even after 16 h of pretreatment followed by infection [105]. These results suggest that the extract activates host cell defense mechanisms and does not act directly on the influenza virus.
In the mouse infection experiments described in the previous section, oral ingestion of Euglena alleviated the symptoms of influenza virus infection, mainly through the contribution of paramylon. The ingestion of paramylon by mice reduced the amount of interferon β in their blood, which the virus infection had increased, on day 3 [100]. After insoluble paramylon is recognized by Dectin-1 (a major β-glucan receptor expressed on intestinal immune cells such as dendritic cells and macrophages) and endocytosed, the activation of tyrosine-protein kinase SYK and transcription factor nuclear factor-kappa B promote cytokine secretion [109,110]. The hydrothermal extract of Euglena obtained by hot water extraction of Euglena powder, consists of water-soluble components, from which insoluble paramylons are thought to be excluded. The concentration of carbohydrates in the hydrothermal extract of Euglena was 0.53% using the phenol sulfate method, whereas the concentration of carbohydrates in the Euglena powder was approximately 30-40%. However, to discard the possibility that a small amount of glucan was involved, the hydrothermal extract of Euglena was treated with β-glucanase to verify if it affected its antiviral activity, and it was found to have no effect. This suggests that β-glucan is not involved in the in vitro influenza virus growth suppression observed with the hydrothermal extract of Euglena.
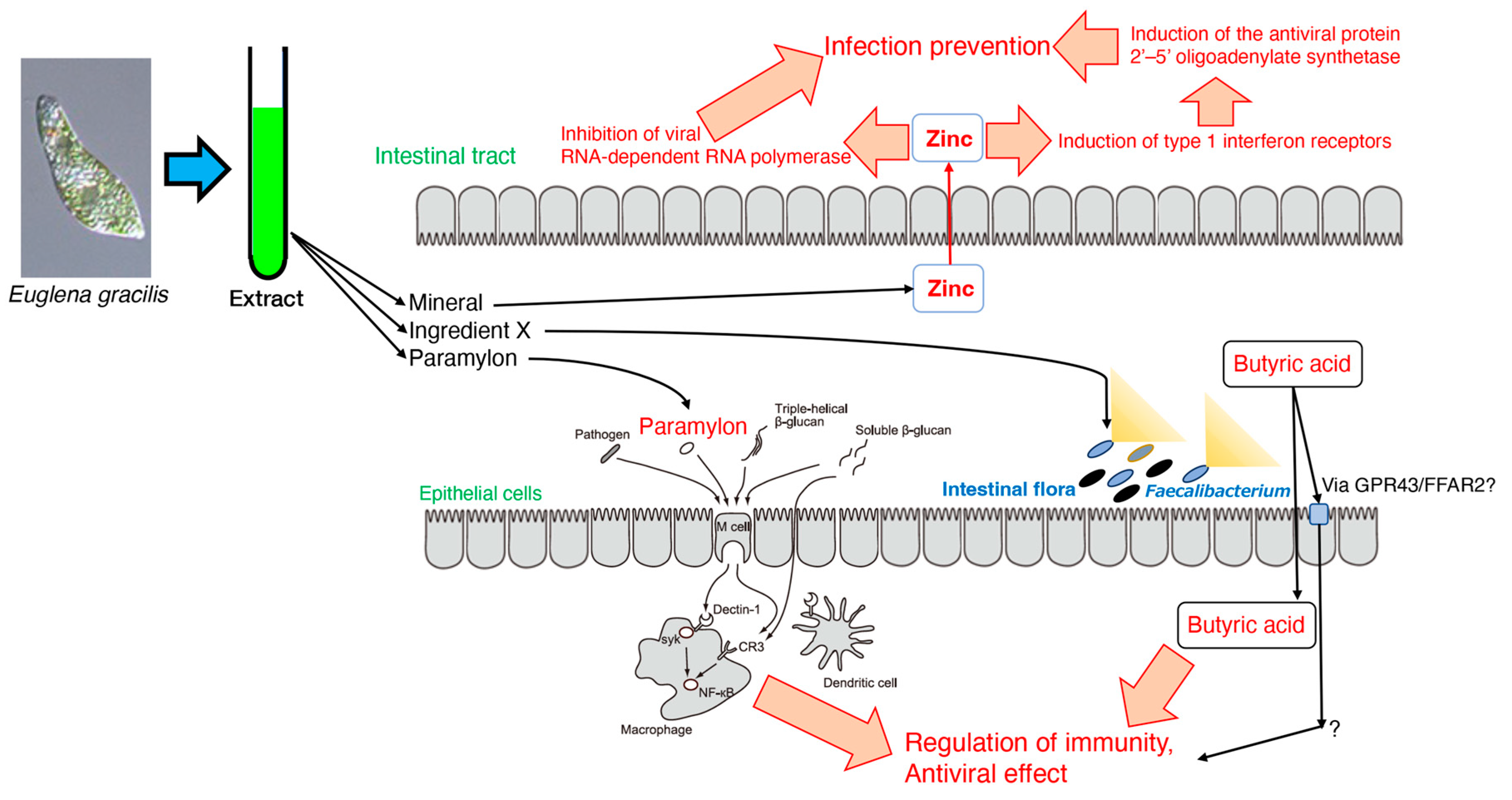
Recently, it has been confirmed that the hydrothermal extract of Euglena suppresses lung cancer symptoms in mice by stimulating host immunity [111]. Therefore, components other than paramylon may be involved in such immune mechanisms. Generally, polyphenols exhibit antiviral effects [112]. However, our experiments revealed that polyphenols were not involved in the antiviral activity of the hydrothermal extract of Euglena. As β-glucan and polyphenols showed no activity, the minerals were analyzed. Zinc is required for the growth of Euglena and accumulates in the cells. In addition, compared to other metals, zinc has antiviral activity by promoting the induction of type 1 interferon receptors, thereby inducing the production of the antiviral protein 2’–5’ oligoadenylate synthetases [113]. In the case of SARS-CoV and equine arteritis virus (EAV), the RNA synthesis is catalyzed by an RNA-dependent RNA polymerase, which is directly inhibited by zinc [114]. Specifically, zinc blocks the initiation step of EAV RNA synthesis, whereas the RNA-dependent RNA polymerase elongation was inhibited and template binding reduced in the case of the SARS-CoV [114]. In our study [105], the addition of zinc acetate inhibited influenza virus growth in a concentration-dependent manner (Figure 3). Furthermore, the addition of zinc acetate to the demineralized Euglena extract restored its influenza virus inhibitory activity; at 0.031 mg/mL of Euglena extract, the zinc concentration was 111.6 nM. At 125 nM with only zinc acetate, its anti-influenza virus activity was comparable to that of the 0.031 mg/mL Euglena extract. Thus, it can be inferred that zinc is responsible for this antiviral activity. Conversely, when 2 mg/mL of membrane-treated Euglena extract was added, the zinc concentration was 44.06 nM. The addition of zinc acetate alone (31 nM) did not inhibit influenza virus growth compared to CER (cation exchange resin)-treated Euglena extract, which has an equivalent zinc concentration. Thus, the CRE-treated Euglena extract, which has an equivalent zinc concentration, has higher anti-influenza virus inhibitory activity than that of zinc alone [105]. Ionophores are required for zinc uptake into cells [114]. Euglena may contain substances that act similarly to ionophores, whereas the hydrothermal extract of Euglena may contain higher concentrations. The ionophores may capture zinc, ensuring some anti-influenza virus activity of the membrane-treated extract at high concentrations. Therefore, the effect of zinc acetate was retained in the normal Euglena extract because of the presence of free zinc, but in CER-treated Euglena, the removal of free zinc by membrane treatment allowed the ionophores to exert the effect of the zinc captured by the ionophores. This suggests that substances other than zinc are also involved in the anti-influenza virus activity of Euglena.
This indicate that not only the paramylon in Euglena eliminates viruses by activating the systemic immunity, but also by the action of other components of Euglena, including zinc, which may act directly on cellular defense mechanisms. A limitation is that the MDCK cells used in the aforementioned study are regularly used in viral research, whereas cells of other origins should also be investigated.
In conclusion, hydrothermal extracts of Euglena reduced the viral titer of various influenza viruses in vitro, and its inhibitory activity was more potent when host cells were pre-exposed to hydrothermal extracts of Euglena, suggesting that Euglena components may provide signals to enhance host cell defense mechanisms. Furthermore, the Euglena extract inhibit the growth of influenza viruses by a nonspecific mechanism different from those of existing drugs. Therefore, this extract may be a promising treatment for infections caused by newly mutated influenza virus strains.
6. Conclusions
In this review, I focused on the immunostimulation of antiviral activities and its potential contribution to the intestinal environment. While reviewing whether the diverse components of Euglena are involved in the antiviral activity, the beneficial effects of Euglena’s intake are clear.
In section 2, Euglena extract and its components, such as paramylon, were shown to be effective in alleviating allergies such as hay fever. In section 3, it was shown that Euglena components other than paramylon may affect Faecalibacterium occupancy and promote defecation by increasing butyrate production [65]. Previous studies have shown that butyrate production from the intestinal flora increases the number of specific CD8+ cells that eliminate influenza viruses and control infection, and these findings indicate that Euglena may be involved in immunomodulation and antiviral activity. In anticipation of its involvement in immunity by improving the intestinal environment, it was confirmed the suppression of influenza virus infection symptoms by Euglena’s intake in vivo [100]. Thus, influenza virus infection could be prevented not only through the action of the intestinal microflora mediated by Euglena components, but also through the systemic immune regulation mediated by the contribution of paramylon in Euglena. In section 5, the direct action of Euglena on influenza virus infection was confirmed [105]. Since this review focused on in vitro experiments, and assumed the involvement of water-soluble components in Euglena, studies with the hydrothermal extracts of Euglena, excluding the insoluble paramylon, were reviewed. These studies showed that the hydrothermal extract of Euglena strongly inhibited the infection of all influenza virus strains, including strains resistant to the anti-influenza drugs oseltamivir and amantadine, indicating that the hydrothermal extract of Euglena may activate the host cell defense mechanism rather than acting directly on the influenza virus. Furthermore, it became clear that the minerals contained in the hydrothermal extract of Euglena, especially zinc, were involved in the infection-inhibitory action of the extract. These studies indicate that not only the paramylon in Euglena eliminates viruses by systemic immunity, but also other components of Euglena, including zinc, may exert antiviral activity by directly acting on cellular defense mechanisms (Figure 4).
Thus, Euglena is effective in the prevention and treatment of influenza through the improvement of intestinal microflora, as well as through the regulation of systemic immunity and the enhancement of cellular defense mechanisms. These functions were associated with one of the major components, paramylon, and the combined contribution of other components in Euglena. This suggests the value of eating whole Euglena as a source of β-glucan (Figure 4).
I propose the ingestion of Euglena as a strong candidate for daily health management through food, prevention against viral infections, and alleviation of symptoms of influenza.
Figure 4.
Model of the anti-influenza virus effect by Euglena extract.
Author Contributions
Conceptualization, original draft preparation, and writing—review and editing, Y.I.
Funding
This work was partially supported by a Grant-in-Aid for Scientific Research (C) (Grant number 22K11792) from the Japan Society for the Promotion of Science (KAKENHI) and by the JST-OPERA Program (Grant Number JPMJOP1832), Japan.
Data Availability Statement
Data will be made available on request.
Acknowledgments
The author would like to thank to Drs. Ayaka Nakashiima and Yoshihisa Nakano for providing detailed information on Euglena. We thank Editage (www.editage.com) for their support with language editing.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ministry of Health, Labour and Welfare, Japan. 2020. The national health and nutrition survey in Japan, 2018. https://www.mhlw.go.jp/content/00106684.
- Daou, C.; Zhang, H. Oat beta-glucan: Its role in health promotion and prevention of diseases. Compr. Rev. Food Sci. Food Saf. 2012, 11, 355–365. [Google Scholar] [CrossRef]
- Chan, G.C.-F.; Chan, W.K.; Sze, D.M.-Y. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Tada, R.; Ikeda, F.; Aoki, K.; Yoshikawa, M.; Kato, Y.; Adachi, Y.; Tanioka, A.; Ishibashi, K.; Tsubaki, K.; Ohno, N. Barley-derived beta-D-glucan induces immunostimulation via a dectin-1-mediated pathway. Immunol. Lett. 2009, 123, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, D.; Iwai, A.; Aoki, S.; Uchiyama, H.; Kawata, K.; Nakayama, Y.; Nikawa, Y.; Kusano, K.; Okabe, M.; Miyazaki, T. β-Glucan derived from Aureobasidium pullulans is effective for the prevention of influenza in mice. PLoS ONE. 2012, 7, e41399. [Google Scholar] [CrossRef]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In vitro fermentation of oat and barley derived beta-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef]
- Gupta, M.; Abu-Ghannam, N.; Gallaghar, E. Barley for brewing: Characteristic changes during malting, brewing and applications of its by-products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 318–328. [Google Scholar] [CrossRef]
- Reyna-Villasmil, N.; Bermúdez-Pirela, V.; Mengual-Moreno, E.; Arias, N.; Cano-Ponce, C.; Leal-Gonzalez, E.; Souki, A.; Inglett, G.E.; Israili, Z.H.; Hernández-Hernández, R.; Valasco, M.; Arraiz, N. Oat-derived beta-glucan significantly improves HDLC and diminishes LDLC and non-HDL cholesterol in overweight individuals with mild hypercholesterolemia. Am. J. Ther. 2007, 14, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Battilana, P.; Ornstein, K.; Minehira, K.; Schwarz, J.M.; Acheson, K.; Schneiter, P.; Burri, J.; Jéquier, E.; Tappy, L. Mechanisms of action of beta-glucan in postprandial glucose metabolism in healthy men. Eur. J. Clin. Nutr. 2001, 55, 327–333. [Google Scholar] [CrossRef]
- Kofuji, K.; Aoki, A.; Tsubaki, K.; Konishi, M.; Isobe, T.; Murata, Y. Antioxidant activity of β-glucan. ISRN Pharm. 2012, 2012, 125864. [Google Scholar] [CrossRef]
- Bayrak, O.; Turgut, F.; Karatas, OF.; Cimentepe, E.; Bayrak, R.; Catal, F.; Atis, O.; Akcay, A.; Unal, D. Oral beta-glucan protects kidney against ischemia/reperfusion injury in rats. Am. J. Nephrol. 2008, 28, 190–196. [Google Scholar] [CrossRef]
- Sener, G.; Toklu, H.; Ercan, F.; Erkanli, G. Protective effect of beta-glucan against oxidative organ injury in a rat model of sepsis. Int. Immunopharmacol. 2005, 5, 1387–1396. [Google Scholar] [CrossRef]
- Taguchi, T.; Furue, H.; Kimura, T.; Kondo, T.; Hattori, T.; Ogawa, N. Clinical efficacy of lentinan on neoplastic diseases. Adv. Exp. Med. Biol. 1983, 166, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Orita, K.; Kimura, T.; Kondo, T.; Taguchi, T.; Yoshida, K.; Ogawa, N.; Furue, H. Clinical evaluation of SPG (schizophyllan) as a therapeutic adjuvant after surgery of gastric cancer--controlled study by an envelope method. Gan To Kagaku Ryoho. 1983, 10, 1135–1145. [Google Scholar] [PubMed]
- Isoda, N.; Eguchi, Y.; Nukaya, H.; Hosho, K.; Suga, Y.; Suga, T.; Nakazawa, S.; Sugano, K. Clinical efficacy of superfine dispersed lentinan (beta-1,3-glucan) in patients with hepatocellular carcinoma. Hepatogastroenterology. 2009, 56, 437–441. [Google Scholar] [PubMed]
- Shimizu, K.; Watanabe, S.; Watanabe, S.; Matsuda, K.; Suga, T.; Nakazawa, S.; Shiratori, K. Efficacy of oral administered superfine dispersed lentinan for advanced pancreatic cancer. Hepatogastroenterology. 2009, 56, 240–244. [Google Scholar] [PubMed]
- Fedorov, S.N.; Ermakova, S.P.; Zvyagintseva, T.N.; Stonik, V.A. Anticancer and cancer preventive properties of marine polysaccharides: Some results and prospects. Mar. Drugs. 2013, 11, 4876–4901. [Google Scholar] [CrossRef]
- Matsumoto, T.; Inui, H.; Miyatake, K.; Nakano, Y.; Murakami, K. Comparison of nutrients in Euglena with those in other respresentative food sources. Eco-Engineering. 2009, 21, 81–86. [Google Scholar]
- Kott, Y.; Wachs, A.M. Amino acid composition of bulk Protein of Euglena growth in wast water. Appl. Microbiol. 1964, 12, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Seppänen-Laakso, T.; Rischer, H.; Wiebe, M.G. Euglena gracilis growth and cell composition under different temperature, light and trophic conditions. PLoS ONE. 2017, 13, e0195329. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Cirpus, P.; Ott, C.; Schlecker, R.; Zähringer, U.; Heinz, E. Biosynthesis of docosahexaenoic acid in Euglena gracilis: Biochemical and Molecular evidence for the involvement of a ∆4-fatty acyl group desaturase. Biochemistry. 2003, 42, 9779–9788. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wan, X.; Zhao, Z.; Liu, H.; Wu, W.; Ge, X.; Zhao, C. Metabolomic analysis and pathway profiling of paramylon production in Euglena gracilis grown on different carbon sources. Int. J. Biol. Macromol. 2023, 246, 125661. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.X.; Schiff, J.A. Chlorophyll-protein complexes from Euglena gracilis and mutants deficient in chlorophyll b: I. Pigment composition. Plant Physiol. 1986, 80, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Yso, R.; Fu, W.; Du, M.; Chen, Z.-X.; Lei, A.-P.; Wang, J.-X. Carotinoids biosynthesis, accumulation, and applications of a model microalga Euglena gracilis. Marine Drugs. 2022, 20, 496. [Google Scholar] [CrossRef]
- Aleksijevic, A .; Grove, J.; Schuber, F. Studies on polyamine biosynthesis in Euglena gracilis. Biochim. Biophys. Acta. 1979, 565, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Deslandes, Y.; Marchessault, R.H.; Sarko, A. Triple-helical structure of (1->3)-β-D-glucan. Macromolecules. 1980, 13, 1466–1471. [Google Scholar] [CrossRef]
- Clarke, A.E.; Stone, B.A. Structure of the paramylon from Euglena gracilis. Biochim. Biophys. Acta. 1960, 44, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.Z.; Roberts, E.M.; Brown, R.M.; Triemer, R.E. X-ray and dissolution studies of paramylon storage granules from Euglena. Protoplasma. 1988, 146, 150–156. [Google Scholar] [CrossRef]
- Chuah, C.T.; Sarko, A.; Deslandes, Y.; Marchessault, R.H. Packing analysis of carbohydrates and polysaccharides. Part 14. Triple-helical crystalline structure of curdlan and paramylon hydrates. Macromolecules. 1983, 16, 1375–1382. [Google Scholar] [CrossRef]
- Barsanti, L.; Passarelli, V.; Evangelista, V.; Frassanito, A.M.; Gualtieri, P. Chemistry, physico-chemistry and applications linked to biological activities of β-glucans. Nat. Prod. Rep. 2011, 28, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Shimada R, Fujita M, Yuasa M, Sawamura H, Watanabe T, Nakashima A, Suzuki K. 2016. Oral administration of green algae, Euglena gracilis, inhibits hyperglycemia in OLETF rats, a model of spontaneous type 2 diabetes. Food Funct. 2016, 7, 4655–4659. [CrossRef] [PubMed]
- Nakashima, A.; Yasuda, K.; Murata, A.; Suzuki, K.; Miura, N. Effects of Euglena gracilis Intake on Mood and Autonomic Activity under Mental Workload, and Subjective Sleep Quality: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients. 2020, 12, 3243. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Kurono, Y.; Ichimura, K.; Enomoto, T.; Okamoto, Y.; Kawauchi, H.; Suzuki, H.; Fujieda, S.; Masuyama, K. The Japanese Society of Allergology. 2017. Japanese guidelines for allergic rhinitis. Allergol. Int. 2017, 66, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Yasueda, H.; Yui, Y.; Shimizu, T. Isolation and partial characterization of the major allergen from Japanese cedar (Cryptomeria japonica) polln. J. Allergy Clin. Immunol. 1983, 71, 77. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Inouye, S.; Taniai, M.; Ando, S.; Usui, M.; Matuhasi, T. Identification of the second major allergen of Japanese cedar pollen. Allergy. 1990, 45, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, D.; Wakabayashi, H.; Watanabe, H.; Nishida, S.; Iino, H. A double-blind trial of Lactobacillus paracasei strain KW3110 administration for immunomodulation in patients with pollen allergy. Allergol. Int. 2005, 54, 143–149. [Google Scholar] [CrossRef]
- Maeda-Yamamoto, M.; Ema, K.; Monobe, M.; Shibuichi, I.; Shinoda, Y.; Yamamoto, T.; Fujisawa, T. The efficancy of early treatment of seasonal allergic rhinitis with benifuuki green tea containing O-methylated catechin before pollen exposure: an open randomized study. Allergol. Int. 2009, 58, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Akiyama, H.; Sugamura, H.; Watanabe, T.; Hamano-Nagaoka, M.; Inakuma, T.; Goda, Y.; Maitani, T. The feeding of β-carotene down-regulates serum IgE levels and inhibits the type I allergic response in mice. Biol. Pharm. Bull. 2004, 27, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Kopf, M.; Le Gros, G.; Bachmann, M.; Lamers, M.C.; Bluethmann, H.; Köhler, G. Disruption of the murine IL-4gene blocks Th2 cytokine respnses. Nature. 1993, 362, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Kato, A.; Hojo, H.; Nozoe, S.; Takeuchi, M.; Ochi, K. Cytokine-related immunopotentiating activities of paramylon, a β-(1–>3)-D-glucan from Euglena gracilis. J. Pharmacobio.-Dyn. 1992, 15, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, A.; Suzuki, K.; Mitra, S.; Arashida, R.; Yoshida, E.; Nakano, R.; Yabuta, Y.; Takeuchi, T. Hepatoprotective effects of pramylon, a β-1,3-D-glucan isolated from Euglena gracilis Z, on acute liver injury induced by carbon tetrachloride in rats. J. Vet. Med. Sci. 2009, 71, 885–890. [Google Scholar] [CrossRef]
- Koizumi, N.; Sakagami, H.; Utsumi, A.; Fujinaga, S.; Takada, M.; Asano, K.; Sugawara, I.; Ichikawa, S.; Kondo, H.; Mori, S.; Miyatake, K.; Nakano, Y.; Nakashima, H.; Murakami, T.; Miyano, N.; Yamamoto, N. Anti-HIV (human immunodeficiency virus) activity of sulfated paramylon. Antiviral Res. 1993, 21, 1–14. [Google Scholar] [CrossRef]
- Sakagami, H.; Kikuchi, K.; Takeda, M.; Sato, T.; Ichikawa, S.; Fujimaki, M.; Wada, C.; Komatsu, N. Macrophage stimulation activity of antimicrobial N, N-dimethylaminoethyl paramylon. In Vivo. 1991, 5, 101–105. [Google Scholar] [PubMed]
- Sugiyama, A.; Hata, S.; Suzuki, K.; Yoshida, E.; Nakano, R.; Mitra, S.; Arashida, R.; Asayama, Y.; Yabuta, Y.; Takeuchi, T. Oral administration of paramylon, a β-1,3-glucan isolated from Euglena gracilis Z inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice. J. Vet. Med. Sci. 2010, 72, 755–763. [Google Scholar] [CrossRef]
- Matsuda, H.; Watanabe, N.; Geba, G.P.; Sperl, J.; Tsudzuki, M.; Hiroi, J.; Matsumoto, M.; Ushio, H.; Saito, S.; Askenase, P.W.; Ra, C. Development of atopic dermatitis-like skin lesion with IgE hyperproduction in NC/Nga mice. Int. Immunol. 1997, 9, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Suto, H.; Matsuda, H.; Mitsuishi, K.; Hira, K.; Uchida, T.; Unno, T.; Ogawa, H.; Ra, C. NC/Nga mice: a mouse model for atopic dermatitis. Int. Arch. Allergy Immunol. 1999, 120, 70–75. [Google Scholar] [CrossRef]
- Iida, M.; Desamero, M.J.; Yasuda, K.; Nakashima, A.; Suzuki, K.; Chambers, J.K.; Uchida, K.; Ogawa, R.; Hachimura, S.; Nakayama, J.; Kyuwa, S.; Miura, K.; Kakuta, S.; Hirayama, K. Effect of orally administered Euglena gracilis and its reserve polysaccharide, paramylon, on gastric dysplasia in A4gnt knockout mice. Sci. Rep. 2021, 11, 13640. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Shimada, R.; Matsuyama, A.; Yuasa, M.; Sawamura, H.; Yoshida, E.; Suzuki, K. Antitumor activity of the β-glucan paramylon from Euglena against preneoplastic colonic aberrant crypt foci in mice. Food Funct. 2013, 4, 1685–1690. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Sumiyoshi, M.; Suzuki, T.; Sakanaka, M. Inhibitory effects of water-soluble low-molecular-weight beta-(1,3-1,6) d-glucan purified from Aureobasidium pullulans GM-NH-1A1 strain on food allergic reactions in mice. Int. Immunopharmacol. 2007, 7, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Yamada, J.; Hamuro, J.; Hatanaka, H.; Hamabata, K.; Kinoshita, S. Alleviation of seasonal allergic symptoms with super fine beta-1,3-glucan: a randomized study. J. Allergy Clin. Immunol. 2007, 119, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, M.; Yoshii, Y.; Yuasa, M.; Sawamura, H.; Watanabe, T.; Yoshida, E.; Suzuki, K. Effect of euglena on cedar pollen allergies in cry j1-sensitized mice. J. Integrated Study Dietary Habits. 2013, 24, 171–176. [Google Scholar] [CrossRef]
- Lazar, V.; Ditu, L.-M.; Pircalabioru, G.G.; Gheorghe, I.; Curutiu, C.; Holban, A.M.; Picu, A.; Petcu, L.; Chifiriuc, M.C. Aspects of gut microbiota and immune system interactions in infectious disease, immunopathology, and cancer. Front. Immunol. 2018, 9, 1830. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; Mende, D.R.; Li, J.; Xu, J.; Li, S.; Li, D.; Cao, J.; Wang, B.; Liang, H.; Zheng, H.; Xie, Y.; Tap, J.; Lepage, P.; Bertalan, M.; Batto, J.-M.; Hansen, T.; Le Paslier, D.; Linneberg, A.; Nielsen, H.B.; Pelletier, E.; Renault, P.; Sicheritz-Ponten, T.; Turner, K.; Zhu, H.; Yu, C.; Li, S.; Jian, M.; Zhou, Y.; Li, Y.; Zhang, X.; Li, S.; Qin, N.; Yang, H.; Wang, J.; Brunak, S.; Doré, J.; Guarner, F.; Kristiansen, K.; Pedersen, O.; Parkhill, J.; Weissenbach, J.; Consortium, M.; Bork, P.; Ehrlich, S.D.; Wang, J. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Blaser, M.J. The human microbiome: at the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res. 2010, 61, 219–225. [Google Scholar] [CrossRef]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In vitro fermentation of oat and barley derived β-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Meijer. K.; de Vos, P.; Priebe, M.G. Butyrate and other short-chain fatty acids as modulators of immunity: what relevance for health? Curr. Opin. Clin. Nutr. Metab. Care. 2010, 13, 715–21. [Google Scholar] [CrossRef] [PubMed]
- Hague. A.; Elder, D.J.; Hicks, D.J.; Paraskeva, C. Apoptosis in colorectal tumour cells: induction by the short chain fatty acids butyrate, propionate and acetate and by the bile salt deoxycholate. Int. J. cancer. 1995, 60, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; Takahashi, T.; Miyauchi, S.; Shioi, G.; Inoue, H.; Tsujimoto, G. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef]
- Schneider, S.M.; Le Gall, P.; Girard-Pipau, F.; Piche, T.; Pompei, A.; Nano, J.L.; Hébuterne, X.; Rampal, P. Total artificial nutrition is associated with major changes in the fecal flora. Eur. J. Nutr. 2000, 39, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health. Dis. 2015, 26, 26164. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Inoue, J.; Sasaki, D.; Hoshi, N.; Shirai, T.; Fukuda, I.; Azuma, T.; Kondo, A.; Osawa, R. Construction of a model culture system of human colonic microbiota to detect decreased Lachnospiraceae abundance and butyrogenesis in the feces of ulcerative colitis patients. Biotechnol. J. 2019, 14, e1800555. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Sasaki, K.; Sasaki, D.; Yasuda, K.; Suzuko, K.; Kondo, A. The alga Euglena gracilis stimulates Faecalibacterium in the gut and contributes to increased defecation. Sci. Rep. 2021, 11, 1074. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Martin, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Li, D.; Feng, N.; Shamoon, M.; Sun, Z.; Ding, L.; Zhang, H.; Chen, W.; Sun, J.; Chen, Y.Q. Anti-diabetic effects of Clostridium butyricum CGMCC0313.1 through promoting the growth of gut butyrate-producing bacteria in type 2 diabetic mice. Sci. Rep 2017, 7, 7046. [CrossRef]
- Shimada, R.; Fujita, M.; Yuasa, M.; Sawamura, H.; Watanabe, T.; Nakashima, A.; Suzuki, K. Oral administration of green algae, Euglena gracilis, inhibits hyperglycemia in OLETF rats, a model of spontaneous type 2 diabetes. Food Funct. 2016, 7, 4655–4659. [Google Scholar] [CrossRef] [PubMed]
- Gissibl, A.; Sun, A.; Care, A.; Nevalainen, H.; Sunna, A. Bioproducts from Euglena gracilis: synthesis and applications. Front. Bioeng. Biotechnol. 2019, 7, 108. [Google Scholar] [CrossRef]
- Sugimoto, R.; Ishibashi-Ohgo, N.; Atsuji, K.; Miwa, Y.; Iwata, O.; Nakashima, A.; Suzuki, K. Euglena extract suppresses adipocyte-differentiation in human adipose-derived stem cells. PLoS ONE. 2018, 13, e0192404. [Google Scholar] [CrossRef] [PubMed]
- Okouchi, R.; E, S.; Yamamoto, K.; Ota, T.; Seki, K.; Imai, M.; Ota, R.; Asayama, Y.; Nakashima, A.; Suzuki, K.; Tsuduki, T. Simultaneous intake of Euglena gracilis and vegetables exerts synergistic anti-obesity and anti-inflammatory effects by modulating the gut microbiota in diet-induced obese mice. Nutrients 2019, 11, 204. [Google Scholar] [CrossRef] [PubMed]
- Rios-Covian, D.; Gueimonde, M.; Duncan, S.H.; Flint, H.J.; de los Reyes-Gavilan, C.G. Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS. Microbiol. Lett. 2015, 362, fnv176. [Google Scholar] [CrossRef] [PubMed]
- Asayama, Y.; Suzuki, K.; Nakashima, A.; Shioya, N.; Sugimura, H. The study of the effects of food containing Euglena gracilis on improvement in constipation. Jpn. Pharmacol. Ther. 2017, 45, 1359–1364. [Google Scholar]
- Kawano, Y.; Nakano, Y.; Kitaoka, S.; Katou, K.; Shigeoka, S.; Ohnishi, T. Effects of Euglena cells on the absorption and tissue distribution of dietary cholesterol in rats. J. Jpn. Soc. Nutr. Food. Sci. 1987, 40, 193–198. [Google Scholar] [CrossRef]
- Yang, J.; Wang, H.-P.; Zhou, L.; Xu, C.-F. Effect of dietary fiber on constipation: a meta analysis. World J. Gastroenterol. 2012, 18, 7378–7383. [Google Scholar] [CrossRef] [PubMed]
- Pituch, A.; Walkowiak, J.; Banaszkiewicz, A. Butyric acid in functional constipation. Prz. Gastroenterol. 2013, 8, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary fiber confers protection against flu by shaping Ly6c– patrolling monocyte hematopoiesis and CD8+ T cell metabolism. Immunity 2018, 48, 992–1005.e8. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Morens, D.M. Influenza: the once and future pandemic. Public Health Rep. 2010, 125, 16–26. [Google Scholar] [CrossRef] [PubMed]
- McCaughey, C. Influenza: a virus of our times. Ulster Med. J. 2010, 79, 46–51. [Google Scholar] [PubMed]
- Van Reeth, K.; Nauwynck, H.; Pensaert, M. Bronchoalveolar interferon-α, tumornecrosis factor-α, interleukin-1, and inflammation during acute influenza inpigs: a possible model for humans. J. Infect. Dis. 1998, 177, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Julkunen, I.; Sareneva, T.; Pirhonen, J.; Ronni, T.; Melén, K.; Matikaine, S. Molecular pathogenesis of influenzaA virus infection and virus-induced regulation of cytokine gene expression. Cytokine Growth Factor Rev. 2001, 12, 171–180. [Google Scholar] [CrossRef]
- Kaiser, L.; Fritz, R.S.; Straus, S.E.; Gubareva, L.; Hayden, F.G. Symptom pathogenesis during acuteinfluenza: interleukin-6 and other cytokine responses. J. Med. Virol. 2001, 64, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Mok, K.P.; Wong, C.H.K.; Cheung, C.Y.; Chan, M.C.; Lee, S.M.Y.; Nicholls, J.M.; Peiris, J.S.M. Viral genetic determinants of H5N1 influenza viruses that contribute to cytokine dysregulation. J. Infect. Dis. 2009, 200, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Arimori, Y.; Nakamura, R.; Yamada, H.; Shibata, K.; Maeda, N.; Kase, T.; Yoshikai, Y. Type I interferon limits influenza virus-induced acute lung injury by regulation of excessive inflammation in mice. Antiviral Res. 2013, 99, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Arimori, Y.; Nakamura, R.; Yamada, H.; Shibata, K.; Maeda, N.; Kase, T.; Yoshikai, Y. Type I interferon plays opposing roles in cytotoxicity and interferon-γ production by natural killer and CD8 T cells after influenza A virus infection in mice. J. Innate Immun. 2014, 6, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vetvickova, J. Glucan supplementation enhances the immune response against an influenza challenge in mice. Ann. Transl. Med. 2015, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, D.; Iwai, A.; Aoki, S.; Uchiyama, H.; Kawata, K.; Nakayama, Y.; Nikawa, Y.; Kusano, K.; Okabe, M.; Miyazaki, T. β-Glucan derived from Aureobasidium pullulans is effective for the prevention of influenza in mice. PLoS ONE. 2012, 7, e41399. [Google Scholar] [CrossRef] [PubMed]
- Hassanzadeh-Kiabi, N.; Yáñez, A.; Dang, I.; Martins, G.A.; Underhill, D.M.; Goodridge, H.S. Autocrine type I IFN signaling in dendritic cells stimulated with fungal β-glucans or lipopolysaccharides promotes CD8 T cell activation. J. Immunol. 2017, 198, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Gibbert, K.; Schlaak, J.F.; Yang, D.; Ditmer, U. IFN-α subtypes: distinct biological activities in anti-viral therapy, Br. J. Pharmacol. 2013, 168, 1048–1058. [Google Scholar] [CrossRef]
- Hunter, C.A.; Chizzonite, R.; Remington, J.S. IL-1 beta is required for IL-12 to induce production of IFN-gamma by NK cells. A role for IL-1 beta in the T cell-independent mechanism of resistance against intracellular pathogens, J. Immunol. 1995, 155, 4347–4354. [Google Scholar] [CrossRef]
- Sabel, M.S.; Arora, A.; Su, G.; Mathiowitz, E.; Reineke, J.J.; Chang, A.E. Synergistic effect of intratumoral IL-12 and TNF-alpha microspheres: systemic anti-tumor immunity is mediated by both CD8+ CTL and NK cells, Surgery. 2007, 142, 749–760. [CrossRef]
- Brown, G.D.; Gordon, S. Immune recognition. A new receptor for beta-glucans. Nature. 2001, 413, 36–37. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.Z.; Roberts, E.M.; Brown, R.M.; Triemer, R.E. X-ray and dissolution studies of paramylon storage granules from Euglena. Protoplasma. 1988, 146, 150–56. [Google Scholar] [CrossRef]
- Chuah, C.T.; Sarko, A.; Deslandes, Y.; Marchessault, R.H. Packing analysis of carbohydrates and polysaccharides. Part 14. Triple-helical crystalline structure of curdlan and paramylon hydrates. Macromolecules. 1983, 16, 1375–1382. [Google Scholar] [CrossRef]
- Ujita, M.; Nagayama, H.; Kanie, S.; Koike, S.; Ikeyama, Y.; Ozaki, T.; Okumura, H. Carbohydrate binding specificity of recombinant human macrophage beta-glucan receptor dectin-1. Biosci. Biotechnol. Biochem. 2009, 73, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Barsanti, L.; Evangelista, V.; Frassanito, A.M.; Longo, V.; Pucci, L.; Penno, G.; Gualtieri, P. Euglena gracilis paramylon activates human lymphocytes by upregulating pro-inflammatory factors. Food Sci. Nutr. 2016, 5, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Legentil L, Paris F, Ballet C, Trouvelot S, Daire X, Vetvicka V, Ferrières V. Molecular interactions of β-(1->3)-glucans with their receptors. Molecules 2015, 20, 9745–9766. [CrossRef] [PubMed]
- Bohn, J.A.; BeMiller, J.N. (1->3)- β-D-Glucans as biological response modifiers: a review of structure-functional activity relationships. Carbohydr. Polym. 1995, 28, 3–14. [Google Scholar] [CrossRef]
- Nakashima, A.; Suzuki, K.; Asayama, Y.; Konno, M.; Saito, K.; Yamazaki, N.; Takimoto, H. Oral administration of Euglena gracilis Z and its carbohydrate storage substance provides survival protection against influenza virus infection in mice. Biochem. Biophys. Res. Commun. 2017, 494, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.B.; Burk, K.; Newman, F.; Cerruti, R.L.; Sim, I.S. Resistance of influenza A virus to amantadine and rimantadine: results of one decade of surveillance. J. Infect. Dis. 1989, 159, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Bloom, J.D.; Gong, L.I.; Baltimore, D. Permissive secondary mutations enable the evolution of influenza oseltamivir resistance. Science 2010, 328, 1272–1275. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.M.; Kiso, M.; Someya, K.; Sakai, Y.T; Nguyen, T.H; Ngyuen, K.H; Pham, N.D; Ngyen, H.H.; Yamada, S.; Muramoto, Y.; Horimoto, T.; Takada, A.; Goto, H.; Suzuki, T.; Suzuki, Y.; Kawaoka, Y. Avian flu: isolation of drug resistant H5N1 influenza A virus. Nature. 2005, 437, 1108. [Google Scholar] [CrossRef] [PubMed]
- Seibert, C.W.; Rahmat, S.; Krammer, F.; Palese, P.; Bouvier, N.M. Efficient transmission of pandemic H1N1 influenza viruses with high-level oseltamivir resistance. J. Virol. 2012, 86, 5386–5389. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Horio, Y.; Suzuki, K.; Isegawa, Y. Antiviral activity and underlying action mechanism of Euglena extract against influenza virus. Nutrients. 2021, 13, 3911. [Google Scholar] [CrossRef] [PubMed]
- Barr, I.G.; Hurt, A.C.; Iannello, P.; Tomasov, C.; Deed, N.; Komadina, N. Increased adamantane resistance in influenza A(H3) viruses in Australia and neighbouring countries in 2005. Antiviral Res. 2007, 73, 112–117. [Google Scholar] [CrossRef]
- Nitsch-Osuch, A.; Brydak, L.B. Influenza viruses resistant to neuraminidase inhibitors. Acta Biochim. Pol. 2014, 61, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Todd, B.; Tchesnokov, E.P.; Götte, M. The active form of the influenza cap-snatching endonuclease inhibitor baloxavir marboxil is a tight binding inhibitor. J. Biol. Chem. 2021, 296, 100486. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Yamada, K.; Iwata, O.; Sugimoto, R.; Atsuji, K.; Ogawa, T.; Ishibashi-Ohgo, N.; Suzuki, K. β-Glucan in foods and its physiological functions. J. Nutr. Sci. Vitaminol. (Tokyo) 2018, 64, 8–17. [Google Scholar] [CrossRef]
- Ujita, M.; Nagayama, H.; Kanie, S.; Koike, S.; Ikeyama, Y.; Ozaki, T.; Okumura, H. Carbohydrate binding specificity of recombinant human macrophage β-glucan receptor dectin-1. Biosci. Biotechnol. Biochem. 2009, 73, 237–240. [Google Scholar] [CrossRef]
- Ishiguro, S.; Upreti, D.; Robben, N.; Burghart, R.; Loyd, M.; Ogun, D.; Le, T.; Delzeit, J.; Nakashima, A.; Thakkar, R.; Nakashima, A.; Suzuki, K.; Comer, J.; Tamura, M. Water extract from Euglena gracilis prevents lung carcinoma growth in mice by attenuation of the my-eloid-derived cell population. Biomed. Pharmacother. 2020, 127, 110166. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-F.; Bai, L.-P.; Huang, W.-B.; Li, X.-Z.; Zhao, S.-S.; Zhong, N.-S.; Jiang, Z.-H. Comparison of in vitro antiviral activity of tea polyphenols against influenza A and B viruses and structure-activity relationship analysis. Fitoterapia. 2014, 93, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T.; Nakajima, K.; Takada, H.; Sekine, Y.; Suzuki, K. Induction of type 1 interferon receptor by zinc in U937 cells. Cytokine. 2009, 46, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Te Velthuis, A.J.; van den Worm, S.H.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn(2+) inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef]
Figure 1.
Model of the activation of Faecalibacterium, butyrate production, and CD8+ cells by component(s) of Euglena.
Figure 1.
Model of the activation of Faecalibacterium, butyrate production, and CD8+ cells by component(s) of Euglena.
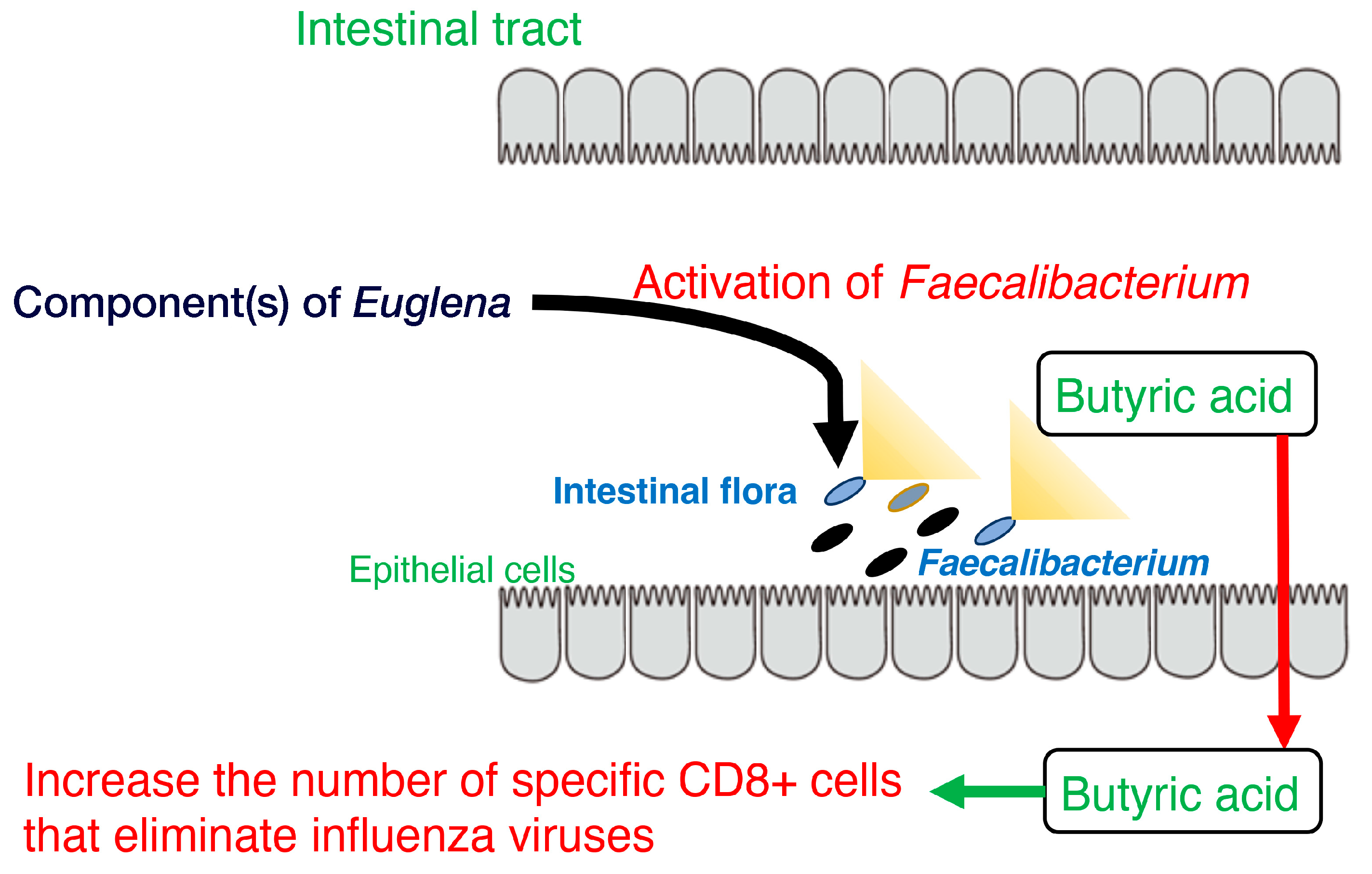
Figure 2.
Immune regulation by paramylon.
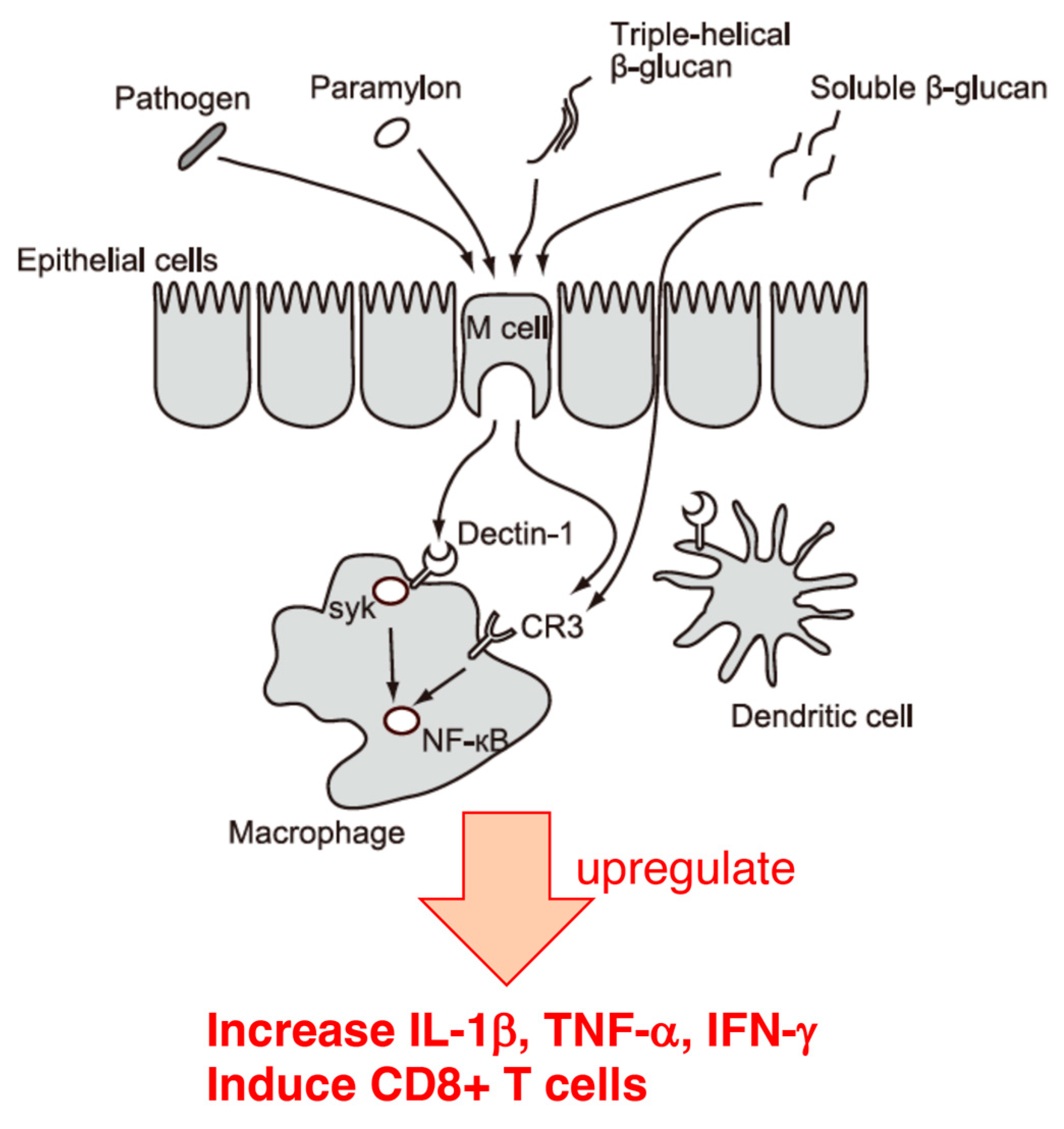
Figure 3.
Model of the inhibition of influenza virus replication by zinc. INF: type 1 interferon; INFR: type 1 interferon receptor; vRNP: viral ribonucleoprotein.
Figure 3.
Model of the inhibition of influenza virus replication by zinc. INF: type 1 interferon; INFR: type 1 interferon receptor; vRNP: viral ribonucleoprotein.
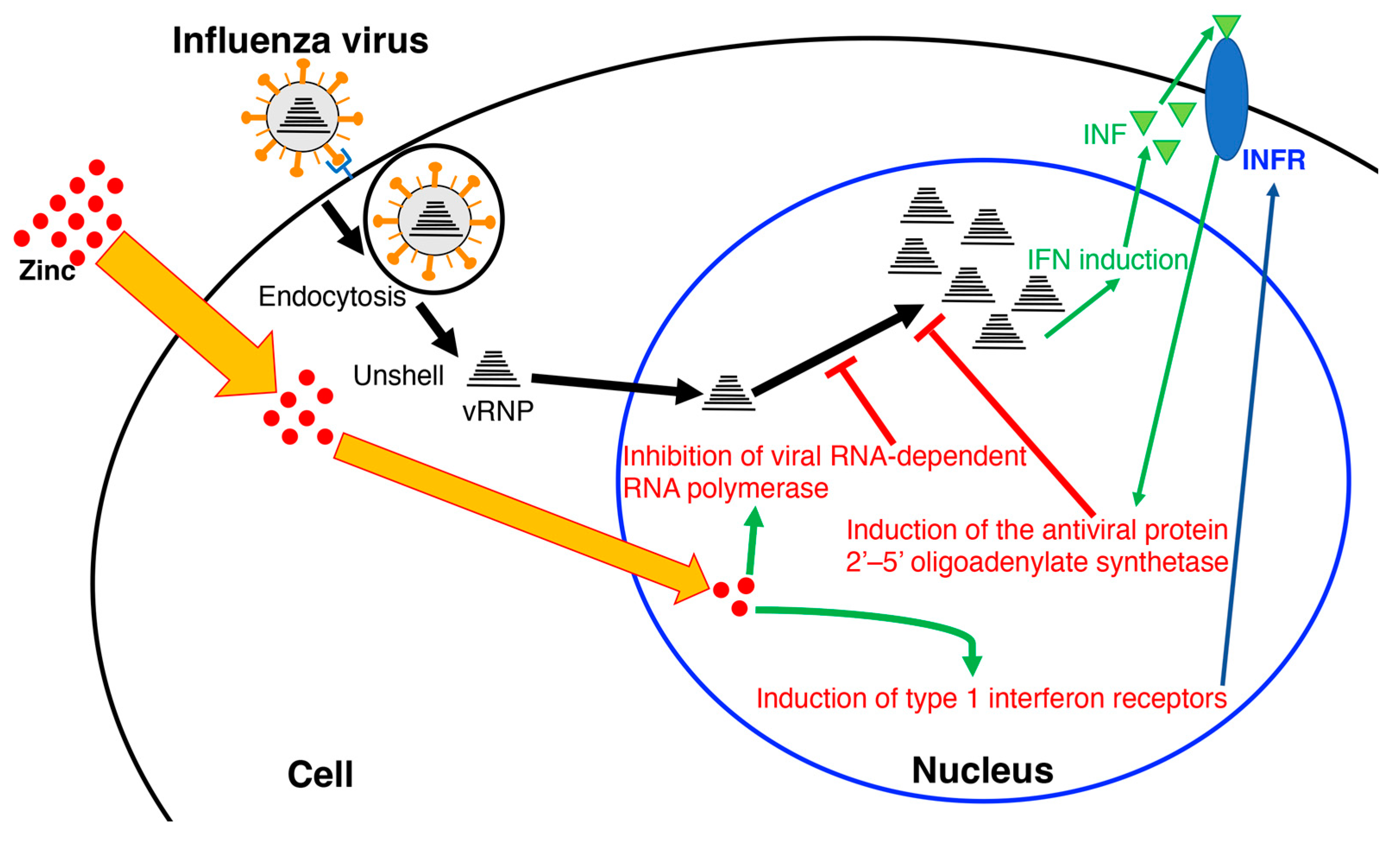

Table 1.
Nutrients contained in Euglena.
| Vitamins | Minerals | Amino acids | Unsaturated fatty acids | ||||||
| Content | Ref. | Content | Ref. | Ref. | Ref. | ||||
| µg/106 cells) | µg/106 cells) | ||||||||
| Vitamin B1 | 1.4 ± 0.1 | [19] | Zn2+ | 0.795 ± 0.104 | [19] | Alanine | [20] | Docosahexaenoic acid | [21,22] |
| Vitamin B2 | 3.5 ± 0.2 | [19] | Mg2+ | 0.165 ± 0.076 | [19] | Arginine | [20] | Eicosapentaenoic acid | [21] |
| Vitamin B6 | 7.5 ± 0.6 | [19] | Fe2+ | 0.153 ± 0.092 | [19] | Aspertic acid | [20] | α-Linolenic acid | [19,21] |
| Vitamin B12 | 0.05 ± 0.01 | [19] | Mn2+ | 0.240 ± 0.056 | [19] | Cystine | [20] | Arachidonic acid | [19,21] |
| Folic acid | 1.7 ± 0.5 | [19] | Ca2+ | 0.046 ± 0.070 | [19] | Glycine | [20] | Palmitoleic acid | [21] |
| Pantotheric acid | 18.2 ± 0.4 | [19] | Cu2+ | 0.004 ± 0.0023 | [19] | Glutamic acid | [20] | Oleic acid | [19,21] |
| Biotin | 4.6 ± 0.3 | [19] | Ni2+ | 0.006 ± 0.0014 | [19] | Histidine | [20] | Linoleic acid | [19,21] |
| Vitamin C | 27.2 ± 0.4 | [19] | Isoleucine | [19,20] | Eicosadienoic acid | [21] | |||
| Tocopherol | 517.5 ± 10.2 | [19] | Leucine | [19,20] | Docosatetraenoic Acid | [21] | |||
| b-Carotene | 0.71 ± 0.15 | [19] | Lysine | [19,20] | Docosapentaenoic acid | [21,22] | |||
| Phenylalanine | [20] | Dihomo-γ-linolenic acid | [21] | ||||||
| Preoline | [20] | ||||||||
| Serine | [20] | ||||||||
| Threonine | [19,20] | ||||||||
| Tryptophan | [19,20] | ||||||||
| Tyrosine | [20] | ||||||||
| Varine | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



