
Submitted:
30 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
The negative impact of drought on plant growth may be modified by different availability of mineral nutrients and by their adaptation to different local habitat conditions. In this study, we examine the impact of drought, fertilization with phosphorus and provenance, as well as their interactions, on the growth and allometric growth relationships between belowground and aboveground organs of common beech (Fagus sylvatica) and sessile oak (Quercus petraea). The research was conducted on saplings originating from two mature mixed stands (dry and wet provenances) dominated by these species. In the common garden experiment, saplings were exposed to regular watering and drought in interaction with moderate and high phosphorus concentrations in the growing substrate (achieved by phosphorus fertilization). Obtained results indicate negative impact of drought and phosphorus fertilization on the growth of both species. In common beech, phosphorus fertilization had a negative impact on its adaptive capacity to drought, because of the unfavourable ratios between fine root mass and mass of other organs. The sessile oak provenances under the impact of drought showed a different root collar diameter/stem height increment ratio, which indicates its different phenotypic plasticity as a consequence of adaptation to different frequencies of dry periods in their natural habitats.
Keywords:
Fagus sylvatica
; Quercus petraea
; Allometric growth relationship
; Adaptation to drought
; Luxury nutrition with phosphorus
1. Introduction
Common beech (Fagus Sylvatica L.) and sessile oak (Quercus Petraea (Matt.) Liebl.) represent two species of forest trees that form mixed forest stands around Europe, with very high biodiversity and economic, ecological and social value [1]. However, forest ecosystems across Europe have recently been increasingly exposed to the negative impact of climate changes, i.e., long-lasting drought periods [2,3,4] and severe drought stress [5,6,7]. Such events have a negative impact on vitality [8,9], carbon storage [10,11], biomass production [12,13] and natural regeneration of forest trees [14,15].
For this reason, scientists and forestry experts invest a lot of effort in finding appropriate solutions to mitigate the negative impact of drought on the growth of forest trees and the survival of entire forest ecosystems. One of the ways to mitigate the negative impact of drought on the growth and vitality of forest trees is the selection of more drought-tolerant genotypes and/or provenances [16]. Common beech and sessile oak provenances often show differentiation in physiological or morphological traits, which are conditioned by their adaptation to specific local habitat conditions [17,18,19]. Provenances of the mentioned species originating from dry compared to wet habitats under the impact of experimentally induced drought have better survival, higher stem diameter and height growth increment, leaf area and leaf thickness, take deeper roots, and invest more of their resources in the development of belowground (fine and coarse roots) at the expense of aboveground (leaves and stems) organs [18,20,21,22,23,24,25,26,27]. This indicates that provenances from dry compared to wet habitats are better adapted to drought [28,29]. Therefore, the use of forest reproductive material (seeds and saplings) originating from dry provenances for the establishment of new and regeneration of existing forest stands could mitigate the negative impact of drought on their growth and survival in the future [30,31].
An abundance of saplings used for the establishment of new and regeneration of existing forest stands are produced in forest nurseries. During nursery production, saplings are fertilized with mineral fertilizers which positively affects their quality [32,33,34,35] and capacity for drought tolerance [36,37,38,39]. Saplings with high quality are defined by a well-developed stem and root, as well as the optimal concentration of mineral nutrients in the leaves, stems and roots [40,41]. According to previous experience and knowledge, saplings with higher compared to lower quality have better survival and growth after transplanting into natural habitats affected by the drought [42,43].
The positive effect of fertilization with NPK (complex) mineral fertilizers or individual nitrogen (N) fertilizers on the growth of forest trees in the initial stages of their development (seedlings and saplings) is quite well documented [11,44,45,46]. However, the effect of fertilizing with phosphorus (P) mineral fertilizers on the growth of forest trees has rarely been investigated so far [47,48,49,50]. Current knowledge about the effect of P fertilization on plant growth and its physiological processes is mainly based on research conducted on annual plants, mainly on crops. The results of these studies indicate that P fertilization has a positive effect on their antioxidant metabolism [51,52], cell membrane stability [53,54], leaf conductance and nitrate reductase activity [55,56,57], water balance [58], photosynthesis and leaf area [54,59], root growth [60] and the ratio of belowground and aboveground growth [61]. However, there are reports about the negative effects of P fertilization on the growth of annual plants. For example, exposure of the model species Oryza sativa and Arabidopsis thaliana to high P concentrations in growing substrate negatively affects their photosynthesis and root morphological traits [62,63]. Sudden exposure to higher P availability of species adapted to relatively low P concentrations in the soils of their natural habitats has a negative effect on its growth [64]. Thus, the contradictory results of previous research indicate that the effect of P fertilization on plant growth and adaptation to drought is still quite unknown.
Natural forest soils in Europe are generally poorly supplied with P [65,66,67,68,69]. Earlier reports about the negative impact of sudden exposure of plants to high P availability on their growth [64], especially on root system [62,63] lead to the assumption that P fertilization could have a negative impact on the growth of the European forest trees and their adaptative capacity to drought. Primarily because the adaptation of forest trees to drought largely depends on the morphological traits of their root system [70]. The effect of P fertilization or different concentrations of P in the natural soil on the growth of common beech and sessile oak is currently poorly investigated [50,71]. Previous research has mainly focused on inter- and intra-provenance differences in the mechanisms of uptake and internal allocation of P during the growing season and leaf phenology, which was mainly investigated in common beech [50,65,72,73,74,75,76,77]. Previous research on sessile oak was focused on the effect of P fertilization on leaf P concentration and total biomass production [71], or leaf phenology [77]. Finally, the effect of drought in interaction with P fertilization on the growth of common beech and sessile oak, as well as other European forest trees, has not been investigated so far. However, results of recent research conducted on two Chinese forest tree species (Phoebe zhennan and Alnus cremastogyne) indicate that P fertilization in interaction with drought has a positive effect on the total root biomass (only for Phoebe zhennan), while it had no significant effect on growth of other aboveground (leaves and stem) and belowground (fine and coarse root) organs [48,49].
To explore the effects of drought and P fertilization on common beech and sessile oak growth, an experiment was conducted with saplings originating from two mixed stands (provenances). One originating from a wet and the other from a dry habitat, with low P concentration in the soil of both provenances. The aims of the research were to:
- examine the effects of drought, P fertilization and provenance on the growth of common beech and sessile oak.
- examine how fertilization with P affects the growth and allometric growth relationships between belowground and aboveground organs of common beech and sessile oak, i.e. their adaptation capacity to drought.
- examine the common beech and sessile oak provenance differentiation with regard to different local habitat conditions.
2. Results
2.1. Soil water conditions and saplings water balances
Under regular watering in the W/+P and W/-P treatments, the volumetric water content (VWC) in the substrate during the entire growing season was between 18.3 and 40.6 %. The water deprivation in the D/+P and D/-P treatments resulted in a continuous decrease in VWC in the substrate, which was reduced to 8.0 and 10.1 % until the end of August (at the end of the dry period), respectively (Figure 1). Regular watering had a significant and positive effect on the Ψ of both species (Table 1 and Figure 1). Accordingly, Ψ in regularly watered saplings of common beech and sessile oak was not lower than -0.4 MPa, neither in May nor at the end of August. However, Ψ in the drought-treated saplings of common beech and sessile oak until the end of August reached mean values of -2.5 and -3.0 MPa, respectively ( Figure 1).
2.2. Substrate P concentration and saplings nutrition with P
Fertilization with P had a significant and positive effect on the substrate and leaf P concentrations of both species (Table 1 and Figure 2). Accordingly, substrate P concentration in the W/+P and D/+P treatments was high, whilst in the W/-P and D/-P it was moderate (Figure 2a). Range of mean leaf P concentration in common beech saplings fertilized with P was between 1.79 – 1.97 mg P g-1 DW, indicating their nutrition with P between upper normal and surplus (Figure 2b), whilst in the sessile oak saplings fertilized with P it was between 1.92 – 1.99 mg P g-1 DW, indicating their upper normal nutrition with P (Figure 2c). Range of mean leaf P concentration in common beech saplings non-fertilized with P was between 1.49 – 1.69 mg P g-1 DW, indicating their central normal nutrition with P (Figure 2b), whilst in the sessile oak saplings non-fertilized with P it was between 1.41 – 1.55 mg P g-1 DW, indicating their lower normal nutrition with P (Figure 2c). Other chemical traits of the substrate and the concentration of other mineral nutrients in the leaves of common beech and sessile oak were similar in all treatments (Table A1 and Table A2).
2.3. Effect of drought on saplings growth
The drought had a negative impact on the growth of both species, indicated by lower mean values of all measured growth parameters in drought-treated compared to regularly watered saplings. Mean values of almost all allometric growth relationship parameters were higher in drought-treated than in regularly watered saplings, for both species. However, in common beech, drought significantly reduced only Hincrement by 33 %, and in sessile oak Dincrement, Hincrement, DSeptember, HSeptember, STmass, AGmass and Larea by 26, 28, 13, 18, 31, 26 and 26 %, respectively. However, in sessile oak drought significantly increased ratios of TRlength/HSeptember and CRmass/STmass (Table 2).
2.4. Effect of P fertilization on saplings growth
Fertilization with P did not have a positive effect on the growth of common beech and sessile oak saplings. Lower mean values of almost all measured growth parameters in the saplings fertilized with P compared to non-fertilized with P, for both species (Table 3) indicate negative effect of P fertilization on comon beech and sessile oak growth. Moreower, P fertilization significantly reduced TRlength in common beech by 16 %, and FRmass in sessile oak by 33 %. In addition, in common beech fertilization with P significantly decreased the ratio TRlength/HSeptember (Table 3).
2.5. Effect of provenance on saplings growth
Provenance significantly affected only Dincrenment in common beech and FRmass in sessile oak saplings. Common beech originating from KA provenance had a 25 % higher Dincrenment compared to SB provenance. Sessile oak from KA provenance had a 28 % higher FRmass compared to SB provenance. In addition, ratios D/HMarch and D/HSeptember in common beech, as well as ratios FRmass/Lmass, FRmass/STmass, FRmass/CRmass and FRmass/L+ST+CRmass in sessile oak were significantly higher in KA compared to SB provenance (Table 4).
2.6. Effect of drought, P fertilization and provenance interactions on saplings growth
Interactions of P fertilization x provenance, as well as drought x P fertilization x provenance, did not significantly affect any of the measured growth and/or allometric growth relationship parameters, for both species (Table A3 and Table A4). However, interaction drought x P fertilization significantly affected FRmass and the ratios FRmass/STmass, FRmass/CRmass and FRmass/L+ST+CRmass in common beech (Table A3). LSD post-hoc test revealed significantly lower FRmass in drought-treated saplings fertilized with P compared to other treatments (Figure 3a), as well as significantly lower ratios of FRmass/STmass, FRmass/CRmass and FRmass/L+ST+CRmass in drought-treated saplings fertilized with P in compared to drought-treated saplings non-fertilized with P (Figure 3b). Interaction drought x provenance significantly affected the ratio of Dincrement/Hincrement in sessile oak (Table A4). LSD post-hoc test revealed a significantly higher Dincrement/Hincrement ratio in drought-treated sessile oak saplings originating from KA compared to SB provenance, as well as compared to regularly watered sessile oak saplings originating from KA and SB provenance (Figure 4).
3. Discussion
The investigated saplings of common beech and sessile oak grew with the same dynamics in the period between transplanting from natural habitats (March 2021) and the beginning of the research (March 2022). This is confirmed by similar DMarch and HMarch at the beginning of the study (in March 2022) in regularly watered and drought-treated saplings (Table 2), in saplings fertilized and non-fertilized with P (Table 3) and in saplings originated from KA and SB provenance (Table 4). Such a result justifies the further interpretation of the obtained results during the growing season of 2022 when the investigation was conducted.
3.1. Effect of drought on saplings growth
According to previous research carried out on common beech and sessile oak, ψ (measured before dawn) higher than -0.4 MPa indicates their regular water supply and lower than -2.0 MPa indicates their exposure to severe drought stress [80,81,82,83,84,85]. Thus, in our case common beech and sessile oak saplings treated with drought were affected by severe drought stress (Figure 1 and Table 1).
The negative effect of drought on the growth of aboveground and belowground organs, as well as its positive effect on ratios between the growth of belowground and aboveground organs in both species (Table 2) are consistent with the results of earlier research in which common beech and sessile oak were experimentally treated by drought [19,20,22,70,86,87,88,89,90,91]. According to Brunner et al. [92], such response of plants to drought is related to optimizing water uptake and simultaneously minimizing water loss from transpiration.
However, our results indicate that the impact of drought was more negative for the growth of sessile oak than for common beech (Table 2). Under favourable environmental conditions after spring growth, the saplings of forest trees (including common beech and sessile oak) usually have a subsequent secondary and/or tertiary growth that occurs during the summer [93,94,95]. Their subsequent summer growth is accompanied by the production of new leaves and xylem vessels, which increase their leaf area and stem diameter [93,94,95,96,97,98,99,100,101,102,103]. Accordingly, the regularly watered (favourable environmental conditions) beech saplings had only secondary stem growth (which ended in mid-June), whilst sessile oak saplings had secondary and tertiary stem growth (which ended in mid-July). In drought-treated saplings, secondary and/or tertiary stem growth was almost completely absent for both species (unpublished data). This quite well explains why the negative (significant) impact of the drought was more pronounced in sessile oak than in common beech (Table 2).
Significantly higher ratios between growth of tap root and stem in the sessile oak treated with drought compared to regular watering (Table 2) confirm its very pronounced adaptation capacity to drought [104,105]. This adaptation is manifested by deeper rooting at the expense of the stem height growth and/or the investment of more dry mass in the coarse root than in the stem [19,20,22,91].
3.2. Effect of P fertilization and interaction drought x P fertilization on saplings growth
In both species, mean values of almost all measured growth parameters in saplings fertilized with P were lower compared to saplings non-fertilized with P (Table 3), indicating a negative effect of P fertilization on the growth of common beech and sessile oak saplings. Such results are contrary to earlier reports about the positive effect of P fertilization on the growth of some forest trees [42,49,106,107].
According to our results, the leaf P concentrations of both species fertilized with P were quite high (Figure 2), indicating their almost excessive (luxury) nutrition with P [79]. According to earlier reports, luxury nutrition of plants with P in natural conditions is a very rare phenomenon and occurs mainly in species adapted to habitats with a low soil P concentration when they are suddenly exposed to a higher soil P concentration, which negatively affects their growth [62,63,64,108,109,110,111,112]. The soils of both provenances are poorly supplied with P [19], which could explain the worse growth of common beech and sessile oak saplings fertilized with P compared to non-fertilized with P (Table 3). In addition, the significantly lower tap root length and the tap root length/stem height ratio in common beech, as well as fine root mass in sessile oak fertilized with P compared to non-fertilized with P (Table 3) agree very well with the results of an earlier study. This study was carried out on the model species Arabidopsis thaliana, which responds to high P concentration in the growth substrate with the production of shallow root system architecture and reduced primary root growth, root apical meristem size, and meristematic activity [62].
However, a few studies on common beech and/or sessile oak determined quite high leaf P concentrations which did not have a negative impact on their growth [65,71,113] and physiology [73,74,114]. Moreover, Newnham and Carlisle [71] point out that the sessile oak in experimental conditions achieves the best growth at a leaf P concentration of 2.2 mg/g DW, which roughly corresponds to our leaf P concentration of sessile oak fertilized with P (Figure 2). This suggests that in addition to excessive P nutrition, something else could negatively affect the growth of our common beech and sessile oak saplings. According to Güsewell [115], the N/P ratio ˂ 10 and ˃ 20 in plant biomass often corresponds to limited biomass production due to an unfavourable balance in plant nutrition with N or P, which most often occurs during artificial fertilization. In our case, the N/P ratio in the leaves of both species fertilized with P was 11, and in the non-fertilized with P was 15 (Table A2). This indicates that an unfavourable N/P ratio in the leaves could also be one of the reasons for the worse growth of the common beech and the sessile oak saplings fertilized with P compared to non-fertilized with P.
The drought in combination with P fertilization compared to other combinations of water and P fertilization treatments significantly reduced fine root mass in common beech (Figure 3a). Such a result is in accordance with the earlier reports where the drought had negatively affected fine root growth in common beech [70,116,117,118,119,120,121] and which indicate that high P concentrations in the growing substrate and/or excessive plant nutrition with P negatively affects the growth of their fine roots [62,63,64]. It is also in line with previous research, according to which common beech in its natural habitats produces lower fine root mass under high compared to low concentrations of soil mineral nutrients [75,122,123,124]. In addition, the ratios of fine root growth and growth of other aboveground organs in common beech, under the impact of drought and fertilization with P were also significantly lower compared to the impact of drought and non-fertilization with P (Figure 3b). Such results indicate that P fertilization could have a negative effect on the ability of common beech to adapt to drought, due to an unfavourable allometric growth relationship between the fine root (responsible for uptake of water and mineral nutrients) and the other organs (especially leaves and stems responsible for transpiration and/or assimilation).
3.3. Effect of provenance and interaction drought x provenance on saplings growth
Previous research indicates that saplings of forest trees with a higher root collar diameter/stem height ratio have greater mechanical stability, making them more resistant to storms and heavy snow drifts [20,125]. Higher maximum snow depths in months with snow cover in KA compared to SB provenance [19] could explain the significantly higher stem diameter increment, as well as stem diameter/stem height ratios in common beech saplings originating from KA compared to SB provenance (Table 4). Furthermore, soils with a lower compared to a higher proportion of silt and clay have a favourable effect on the growth of fine roots in forest trees [123,126,127]. A lower proportion of silt and clay, as well as a higher proportion of sand in the soil of KA provenance compared to SB provenance [19], could explain the significantly higher fine root mass and ratios between fine root mass and mass of the other organs in sessile oak saplings originating from KA compared to SB provenance (Table 4). Such results indicate genetic differentiation between common beech and/or sessile oak provenances as a consequence of adaptation to specific local habitat conditions.
European oaks from provenances with lower annual precipitation respond to experimentally induced drought with a higher root collar diameter/stem height ratio compared to provenances with higher annual precipitation, indicating a more pronounced strengthening of the root system at the expense of its stem height growth (with the aim of more efficient uptake of water from the dried soil) in dry compared to wet provenances [20,22]. However, our result was contrary. Despite lower annual precipitation in the period from 1949 to 2019 in SB than in KA provenance [19], drought-treated sessile oak saplings from SB provenance had a significantly lower stem diameter/stem height increment ratio compared to KA provenance (Figure 4). However, a similar stem diameter/stem height increment ratio in regularly watered saplings of both provenances compared to a significantly higher stem diameter/stem height increment ratio in drought-treated saplings from KA than from SB provenance indicates higher phenotypic plasticity of saplings from KA provenance (Figure 4). According to earlier reports, changes in plant phenotypic plasticity can be triggered by epigenetic imprinting, which helps them adapt to adverse environmental conditions, including drought [128,129]. In the period from 2016 to 2020 during the saplings' growth in their natural habitats, more dry months were recorded in KA provenance than in SB provenance [19]. This could have triggered epigenetic changes in the sessile oak from KA provenance and resulted in a higher stem diameter/stem height increment ratio under the impact of experimentally induced drought (Figure 4).
4. Materials and Methods
4.1. Plant material and provenance habitat conditions
Investigated four-year-old saplings of common beech and sessile oak originated from two mature mixed stands (provenances) dominated by these species, from the continental part of the Republic of Croatia (RH). One is 100 years old, from the north-western part of RH, near the city of Karlovac (KA provenance, 15.524041 N, 45.466135 E), and the other 105 years old, from the eastern part of RH, near the city of Slavonski Brod (SB provenance, 17.973173 N, 45.273451 E). In both provenances, during early March 2021, saplings were carefully excavated with minimal damage to their root system, under mature trees, at least 100 meters apart. Sever et al. [19] provide more details about the habitat conditions of the investigated provenances (phytosociological, geomorphological, climatological and meteorological traits, as well as mechanical, physical and chemical soil traits). However, here it is important to point out that the mean annual precipitation for the period from 1949 to 2019 in KA provenance (1111.8 mm) was higher than in SB provenance (770.3 mm). Despite this, in the period from 2016 to 2020 (during the growth of four-year-old saplings in their natural habitats) in KA provenance there were 17 moderately to extremely dry months recorded (nine during the growing seasons) compared to SB provenance in which there were nine moderately to extremely dry months (four during growing seasons). The mean maximum snow depths in the months with snow cover for the period from 1949 to 2019 were higher in the KA (32 cm) compared to the SB (18 cm) provenance. Most of the physical and/or chemical soil traits were similar in both provenances, including P concentration, which in KA provenance was 0.50 ± 0.32 mg P2O5 100 g-1 of soil, and in SB provenance was 0.64 ± 0.21 mg P2O5 100 g-1 of soil, indicating low P concentration in soils (on depth 0 – 30 cm) of both provenances. Soil mechanical composition (on depth 0 – 30 cm) described by the relative proportion of sand, silt and clay in KA provenances was 21.3, 59.6 and 19.1 %, respectively, and in SB provenance 2.4, 73.8 and 23.8 %, respectively.
4.2. Experimental design and growth conditions
The excavated saplings (in the previously mentioned stands) with an average height of 36.6 ± 8.01 cm, were transported to the garden of the Faculty of Forestry and Wood Technology, University of Zagreb, where the common garden experiment was established (45.82065 N, 16.02303 E). The establishment of the common garden experiment included sapling transplantation in four wooden boxes (dimensions 155 x 275 x 80 cm, with a volume of 3.41 m3) which were previously filled with Klasmann TS 3 substrate (3800 L in each box) in which concentration of P2O5 was 160 mg L-1 substrate. In the substrate of two boxes, 1182 g of triple superphosphate (Triplex) fertilizer was added, which contains 45 % of P2O5, to increase the concentration of P2O5 up to 300 mg L-1 substrate, which indicates high concentrations of easily accessible P. In the substrate of the other two (non-fertilized) boxes, the concentration of P2O5 was 160 mg L-1 substrate, which indicates moderate concentrations of easily accessible P.
After fertilization, in each box 25 common beech and 25 sessile oak saplings originating from the KA provenance were transplanted, and the same number of common beech and sessile oak saplings from the SB provenance (100 saplings in each box), according to a random arrangement at a distance of 20 x 18 cm. It means that 400 plants were planted in the whole experiment. During the growing season of 2021, all transplanted saplings were exposed to natural meteorological conditions with regular watering during the summer with the aim of their better acclimatization and survival. During the growing season of 2022, all boxes were covered with a transparent PVC roof to prevent natural precipitation. After that, two boxes (one fertilized with P and the other non-fertilized with P) were manually watered with 40 L water per box every four days during the entire growing season. The other two boxes (one fertilized with P and the other non-fertilized with P) were exposed to drought (from 15th May to 1st September 2022 when the drought period was interrupted by re-watering) and watered very rare with 20 L water per box, only at the moment of the appearance of wilting leaves, indicating drought stress. That happened only three times, at the end of July, the beginning of August and the middle of August. As mentioned above, during the growing season of 2022 investigated saplings were growing in four different treatments: regular watering and fertilization with P (W/+P treatment), regular watering and non-fertilization with P (W/-P treatment), drought and fertilization with P (D/+P treatment), drought and non-fertilization with P (D/-P treatment).
4.3. Soil water content and chemical traits
Seasonal water dynamics in the substrate of all treatments was controlled using a data logger and sensors for measurement of volumetric water content (VWC) in the soil (Spectrum Technologies, Inc., Aurora, USA). In each treatment, there were four sensors installed at a depth of 5 – 20 cm. Sampling of the substrate to determine its chemical traits was carried out in mid-September 2021 (six months after the P fertilization). In each box (treatment) one composite sample was collected, formed with nine sub-samples collected in a diagonal (X) arrangement. The samples were subjected to soluble component extraction by mixing with deionized water in a 1:2 ratio. The material was mixed for 1 h using a rotating mixer. Then, the suspension was filtered through filter paper and the clear filtrate was subjected to an analytical procedure aiming to determine the pH reaction, total nitrogen (N total), nitrate (NO3-), ammonium (NH4+), phosphorus (P3-), potassium (K+), calcium (Ca2+), magnesium (Mg2+), chloride (Cl-) and sodium (Na+) ions and salt, as well as electrical conductivity (E.C.) in the substrate of each treatment separately. The applied methodology for the chemical analyses of the substrate is described more in detail in Page et al. [78].
4.4. Leaf water potential, growth and dry mass production
The investigation was carried out on 192 saplings. After the establishment of the common garden experiment in the spring of 2021, in each treatment, 12 common beech and 12 sessile oak saplings originating from the KA provenance, and the same number of saplings from the SB provenance (48 saplings per treatment) were randomly selected and labelled. On three selected and/or labelled common beech and sessile oak saplings originating from KA and SB provenance per each treatment pre-down leaf water potential (Ψ) was measured, using a portable pressure chamber (PMS Instrument Company, Oregon, USA). The first measurements were done on the 26th of May (at the beginning of the drought period) and the second measurements on the 31st of August of 2022 (at the end of the drought period). The root collar diameter (D) in mm and the stem height (H) in cm were measured twice, for all labelled saplings. The first time was in March, before the drought treatment (DMarch and HMarch), and the second time was in mid-September after the drought treatment (DSeptember and HSeptember). Root collar diameter increment (Dincrement) and stem height increment (Hincrement) were calculated as differences between D or H measured in September and March of 2022. At the beginning of September (shortly after the drought period was interrupted by re-watering), the labelled saplings were carefully excavated from the substrate and their tap root length (TRlength) in cm and total leaf area (Larea) in m2 were measured. Leaf area was measured using the software package WinFOLIA 2005b (Regent Instruments, Quebec City, Canada). After drying at 105 °C for 48 hours (to constant mass), using an analytical balance with a precision of 0.01 g, the dry mass of leaves (Lmass), stem (STmass), coarse roots >2 mm (CRmass) and fine roots < 2 mm (FRmass) was determined for each sapling. The total dry mass of aboveground (AGmass = Lmass + STmass) and belowground (BGmass = CRmass + FRmass) organs was also calculated. Based on previously measured growth parameters, the following allometric growth relationship parameters were calculated: D/HMarch, Dincrement/Hincrement, D/HSeptember, TRlength/HSeptember, CRmass/STmass, FRmass/Lmass, FRmass/STmass, FRmass/CRmass, FRmass/L+ST+CRmass and BGmass/AGmass, which describe the growth of belowground (coarse and fine roots) compared to aboveground (leaves and stems) organs.
4.5. Leaf chemical traits
After determining the leaf dry mass and leaf area, dry leaves were grounded, homogenized and subjected to chemical analysis to determine the nutrition of the investigated plants with mineral nutrients. The concentration of N was determined using the Kjeldahl method. Digestion was performed with concentrated nitric acid (HNO3) and perchloric acid (HClO4). The concentration of P was determined spectrophotometrically. The concentration of K was determined by a flame photometer. The concentrations of Ca, Mg and Fe were determined by atomic absorption spectrophotometry with previous digestion with concentrated HNO3 and HClO4. The aforementioned analyses were performed according to standardized international protocols [130].
4.6. Statistical analysis
All statistical tests were performed using the SAS statistical software package (SAS Institute Inc., Cary, NC, USA). Assumptions of residual normality and variance homogeneity were tested by using the Shapiro–Wilk test and Levine’s test with the GLM and UNIVARIATE procedures in SAS. Residuals were plotted as a function of fitted values to test for variance homogeneity, and the distribution of residuals was also tested. Factorial ANOVA was performed to evaluate the fixed effects of drought, P fertilization and provenance, as well as the interaction of these effects on leaf water potential, concentrations of mineral nutrition in leaves, growth parameters and allometric growth relationship parameters, for each species separately. In all cases, an LSD post-hoc test was performed to determine the significance of differences (p < 0.05) between studied effect levels.
5. Conclusions
The drought had a negative impact on the growth of both species but stimulated the growth of their belowground compared to aboveground organs. Such effect of drought on the growth and allometric growth relationship between belowground and aboveground organs was stronger in the sessile oak than in the common beech. P fertilization did not have a positive effect on the growth of common beech and sessile oak. Moreover, in common beech, P fertilization had a negative effect on the tap root length and tap root length/stem height ratio, as well as on the fine root mass in sessile oak. In addition, in common beech, P fertilization in interaction with drought had a negative effect on the fine root mass and the ratios of fine root mass/mass of other aboveground organs, which could reduce its adaptive capacity to drought. Significantly different root collar diameter increments and root collar diameter/stem height ratios (in March and September) between common beech provenances could indicate its genetic differentiation as a consequence of adaptation to different snow depths during the winter months in their natural habitats. Significantly different fine root mass and the ratios of fine root mass/mass of other organs between sessile oak provenances could indicate its genetic differentiation as a consequence of adaptation to different soil mechanical compositions in their natural habitats. Significantly different root collar diameter/height growth increment ratio between sessile oak saplings treated by drought originating from KA and SB provenance could indicate its different phenotypic plasticity, as a consequence of its adaptation to the different frequency of dry periods in their natural habitats.
Author Contributions
conceptualization, A.V. and K.S.; methodology, A.V., K.S., S.B. and Ž.Š.; validation, K.S.; formal analysis, A.V and K.S.; investigation A.V., K.S., S.B., Ž.Š., D.K., I.K.B., T.K., M.B. and M.B.; resources, K.S.; data curation, A.V., K.S. and S.B.; writing—original draft preparation, A.V.; writing—review and editing, K.S., S.B., Ž.Š., D.K., I.K.B., T.K., M.B. and M.B.; visualization, A.V., K.S. and Ž.Š.; supervision, K.S.; project administration, K.S. and A.V.; funding acquisition, K.S. All authors have read and agreed to the published version of the manuscript.
Funding
this research was funded by the Croatian Science Foundation, grant number IP-2020-02-5204; project title: “Phenotypic response of common beech and sessile oak provenances to long-lasting drought in interaction with different phosphorus concentrations in the soil”.
Data Availability Statement
the data in this study are available from the corresponding author upon request.
Conflicts of Interest
the authors declare no conflict of interest.
Appendix A

Table A1.
pH reaction and concentration of water-extractable mineral nutrition in substrate sampled from regular watered and phosphorus fertilization treatment (W/+P), regular watered and non-phosphorus fertilization treatment (W/-P) drought and phosphorus fertilization treatment (D/+P) and drought and non-phosphorus fertilization treatment (D/-P). Description of water-extractable mineral nutrition concentration were taken from Page et al. [78].
Table A1.
pH reaction and concentration of water-extractable mineral nutrition in substrate sampled from regular watered and phosphorus fertilization treatment (W/+P), regular watered and non-phosphorus fertilization treatment (W/-P) drought and phosphorus fertilization treatment (D/+P) and drought and non-phosphorus fertilization treatment (D/-P). Description of water-extractable mineral nutrition concentration were taken from Page et al. [78].
| Parameter | W/+P | W/-P | D/+P | D/-P | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Values | Description | Values | Description | Values | Description | Values | Description | |||||
| pH (H2O) | 6.33 | Slightly acidic | 6.88 | Neutral | 6.81 | Neutral | 7.07 | Neutral | ||||
| P3- (mg/L) | 9.7 | High | 3.31 | Moderate | 8.4 | High | 2.66 | Moderate | ||||
| NH4+ (mg/L) | 12.13 | Optimal | 12.56 | Optimal | 13.24 | Optimal | 13.76 | Optimal | ||||
| NO3- (mg/L.) | 69.8 | Optimal | 50.3 | Optimal | 59.5 | Optimal | 60 | Optimal | ||||
| N total (mg/L) | 47.4 | Medium - normal | 46.57 | Medium - normal | 46.92 | Medium - normal | 49.69 | Medium - normal | ||||
| K+ (mg/L) | 59.9 | Medium - normal | 60.4 | Medium - normal | 63.0 | Medium - normal | 60.9 | Medium - normal | ||||
| Mg2+ (mg/L) | 51.68 | Moderate | 53.52 | Moderate | 61.38 | Moderate | 54.53 | Moderate | ||||
| Ca2+ (mg/L) | 196 | Low | 244 | Low | 204 | Low | 234 | Low | ||||
| Cl- (mg/L) | 57.3 | Medium -normal | 58.4 | Medium -normal | 54.8 | Medium -normal | 52.4 | Low | ||||
| Na+ (mg/L) | 35.49 | Moderate | 36.5 | Moderate | 34.79 | Moderate | 36.6 | Moderate | ||||
| E.C. (mS/cm) | 1.198 | Medium - normal | 1.171 | Medium - normal | 1.185 | Medium - normal | 1.156 | Medium - normal | ||||
| Salt (%) | 0.153 | Medium - normal | 0.149 | Medium - normal | 0.151 | Medium - normal | 0.147 | Medium - normal | ||||
Table A2.
Mean values ± SD of phosphorus (P), nitrogen (N), calcium (Ca), magnesium (Mg), potassium (K) and iron (Fe) concentration, as well as, N/P ratio in leaves of common beech and sessile oak saplings treated by regular watering (W), drought (D), fertilization with phosphorus (+P) and non-fertilization with phosphorus (-P), originated from Karlovac (KA) and Slavonski Brod (SB) provenances. Bolded values indicate significant differences between W and D, as well as, +P and -P saplings, or between saplings originated from KA and SB provenances, as calculated with factorial ANOVA, at p < 0.05. Critical foliar concentration was taken from Mellert and Göttlein (2012). .
Table A2.
Mean values ± SD of phosphorus (P), nitrogen (N), calcium (Ca), magnesium (Mg), potassium (K) and iron (Fe) concentration, as well as, N/P ratio in leaves of common beech and sessile oak saplings treated by regular watering (W), drought (D), fertilization with phosphorus (+P) and non-fertilization with phosphorus (-P), originated from Karlovac (KA) and Slavonski Brod (SB) provenances. Bolded values indicate significant differences between W and D, as well as, +P and -P saplings, or between saplings originated from KA and SB provenances, as calculated with factorial ANOVA, at p < 0.05. Critical foliar concentration was taken from Mellert and Göttlein (2012). .
| Species | Nutrients | Water treatments | Fertilization treatments | Provenances | Critical foliar concentration | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| W | D | +P | -P | KA | SB | Deficiency | Normal range | Surplus | ||
| Common beech | P (mg g-1) | 1.77 ± 0.34 | 1.70 ± 0.33 | 1.91 ± 0.29 | 1.56 ± 0.29 | 1.74 ± 0.29 | 1.73 ± 0.38 | ˂1.2 | 1.2 – 1.9 | >1.9 |
| N (mg g-1) | 22.68 ± 4.55 | 21.53 ± 3.85 | 21.25 ± 4.44 | 22.96 ± 3.87 | 22.48 ± 4.13 | 21.73 ± 4.35 | ˂18.7 | 18.7 – 23.2 | >23.2 | |
| Ca (mg g-1) | 6.27 ± 2.00 | 5.99 ± 1.99 | 6.51 ± 2.32 | 5.75 ± 1.53 | 6.03 ± 2.12 | 6.23 ± 1.88 | ˂6.7 | 6.7 – 14.0 | >14.0 | |
| Mg (mg g-1) | 2.14 ± 0.42 | 2.16 ± 0.38 | 2.21 ± 0.41 | 2.09 ± 0.37 | 2.12 ± 0.37 | 2.18 ± 0.42 | ˂1.1 | 1.1 – 2.3 | >2.3 | |
| K (mg g-1) | 7.33 ± 1.58 | 7.31 ± 1.78 | 7.13 ± 1.86 | 7.51 ± 1.46 | 7.49 ± 1.79 | 7.15 ± 1.55 | ˂6.1 | 6.1 – 9.7 | >9.7 | |
| Fe (mg g-1) | 0.11 ± 0.03 | 0.12 ± 0.05 | 0.11 ± 0.03 | 0.12 ± 0.06 | 0.11 ± 0.05 | 0.12 ± 0.04 | - | - | - | |
| N/P | 13.27 ± 3.62 | 13.35 ± 3.92 | 11.42 ± 3.07 | 15.20 ± 3.43 | 13.24 ± 3.12 | 13.38 ± 4.33 | - | - | - | |
| Sessile oak | P (mg g-1) | 1.72 ± 0.41 | 1.69 ± 0.40 | 1.96 ± 0.26 | 1.46 ± 0.37 | 1.73 ± 0.39 | 1.69 ± 0.42 | ˂1.4 | 1.4 – 2.1 | >2.1 |
| N (mg g-1) | 20.94 ± 3.04 | 20.21 ± 2.93 | 20.77 ± 3.22 | 20.38 ± 2.77 | 20.47 ± 2.88 | 20.68 ± 3.14 | ˂19.8 | 19.8 – 26.8 | >26.8 | |
| Ca (mg g-1) | 9.78 ± 3.17 | 10.00 ± 3.46 | 10.27 ± 3.85 | 9.5 ± 2.64 | 9.70 ± 3.25 | 10.08 ± 3.39 | ˂5.3 | 5.3 – 10.2 | >10.2 | |
| Mg (mg g-1) | 2.47 ± 0.43 | 2.39 ± 0.40 | 2.44 ± 0.47 | 2.42 ± 0.37 | 2.40 ± 0.44 | 2.45 ± 0.40 | ˂1.2 | 1.2 – 2.4 | - | |
| K (mg g-1) | 9.71 ± 1.98 | 9.43 ± 1.60 | 10.36 ± 1.91 | 8.78 ± 1.25 | 9.63 ± 1.96 | 9.51 ± 1.63 | ˂7.2 | 7.2 – 11.4 | >11.4 | |
| Fe (mg g-1) | 0.11 ± 0.05 | 0.12 ± 0.05 | 0.12 ± 0.06 | 0.11 ± 0.04 | 0.11 ± 0.04 | 0.12 ± 0.05 | - | - | - | |
| N/P | 13.13 ± 4.90 | 12.69 ± 3.84 | 10.82 ± 2.41 | 15.00 ± 4.91 | 12.68 ± 4.46 | 13.13 ± 4.34 | - | - | - | |
Table A3.
Main effects of drought (regularly watered vs. drought-treated saplings), phosphorus fertilization (saplings fetilized with phosphorus vs. non-fertilized with phosphorus), provenance (saplings originated from Karlovac vs. Slavonski Brod provenance) and its interactions on growth and allometric growth relationship parematers in common beech saplings, as calculated with factorial ANOVA, (F and p values). Bolded values indicate significant effects at p ˂ 0.05.
Table A3.
Main effects of drought (regularly watered vs. drought-treated saplings), phosphorus fertilization (saplings fetilized with phosphorus vs. non-fertilized with phosphorus), provenance (saplings originated from Karlovac vs. Slavonski Brod provenance) and its interactions on growth and allometric growth relationship parematers in common beech saplings, as calculated with factorial ANOVA, (F and p values). Bolded values indicate significant effects at p ˂ 0.05.
| Parameter | Drought (D) |
Fertilization (P) | Provenance (Pr) | D x P | D x Pr | P x Pr | D x P x Pr | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| Growth | DMarch (mm) | 0.029 | 0.866 | 0.502 | 0.480 | 1.019 | 0.315 | 1.370 | 0.245 | 0.248 | 0.620 | 0.323 | 0.571 | 0.384 | 0.537 |
| Dincrement (mm) | 0.602 | 0.440 | 2.463 | 0.120 | 4.957 | 0.029 | 0.003 | 0.955 | 0.095 | 0.759 | 0.470 | 0.495 | 1.610 | 0.208 | |
| DSeptember (mm) | 0.223 | 0.638 | 1.540 | 0.218 | 3.112 | 0.081 | 0.784 | 0.378 | 0.274 | 0.602 | 0.553 | 0.459 | 0.006 | 0.940 | |
| HMarch (cm) | 0.000 | 0.983 | 0.107 | 0.744 | 1.050 | 0.308 | 0.065 | 0.799 | 0.170 | 0.681 | 0.090 | 0.765 | 1.117 | 0.293 | |
| Hincrement (cm) | 10.433 | 0.002 | 0.033 | 0.857 | 1.125 | 0.292 | 1.284 | 0.260 | 0.001 | 0.973 | 0.032 | 0.859 | 0.692 | 0.408 | |
| HSeptember (cm) | 1.956 | 0.166 | 0.053 | 0.818 | 0.256 | 0.614 | 0.066 | 0.799 | 0.165 | 0.686 | 0.131 | 0.718 | 0.405 | 0.526 | |
| Larea (m2) | 0.757 | 0.387 | 1.566 | 0.214 | 0.321 | 0.573 | 2.766 | 0.100 | 0.189 | 0.665 | 0.182 | 0.671 | 0.103 | 0.749 | |
| TRlength (cm) | 1.259 | 0.265 | 5.585 | 0.020 | 0.004 | 0.951 | 0.603 | 0.439 | 0.449 | 0.505 | 0.336 | 0.564 | 0.047 | 0.828 | |
| Lmass (g) | 0.781 | 0.379 | 0.642 | 0.425 | 0.377 | 0.541 | 1.153 | 0.286 | 0.081 | 0.776 | 0.169 | 0.682 | 0.000 | 0.988 | |
| STmass (g) | 2.118 | 0.149 | 0.059 | 0.809 | 0.386 | 0.536 | 0.549 | 0.461 | 0.005 | 0.945 | 0.004 | 0.952 | 0.163 | 0.687 | |
| AGmass (g) | 1.783 | 0.185 | 0.158 | 0.692 | 0.402 | 0.528 | 0.718 | 0.399 | 0.001 | 0.981 | 0.024 | 0.877 | 0.091 | 0.764 | |
| FRmass (g) | 1.470 | 0.229 | 2.778 | 0.099 | 1.652 | 0.202 | 5.042 | 0.027 | 0.004 | 0.948 | 0.052 | 0.821 | 0.132 | 0.718 | |
| CRmass (g) | 3.261 | 0.074 | 0.459 | 0.500 | 1.341 | 0.250 | 0.089 | 0.766 | 0.232 | 0.631 | 0.367 | 0.546 | 0.016 | 0.901 | |
| BGmass (g) | 2.959 | 0.089 | 0.906 | 0.344 | 1.516 | 0.222 | 0.642 | 0.425 | 0.130 | 0.719 | 0.174 | 0.678 | 0.000 | 0.995 | |
Table A3.
cont.
| Parameter | Drought (D) |
Fertilization (P) | Provenance (Pr) | D x P | D x Pr | P x Pr | D x P x Pr | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| Allometric growth relationship | D/HMarch | 0.119 | 0.731 | 0.996 | 0.321 | 11.745 | 0.001 | 0.619 | 0.434 | 0.094 | 0.760 | 1.153 | 0.286 | 0.009 | 0.923 |
| Dincrement/Hincrement | 1.633 | 0.205 | 2.053 | 0.155 | 1.762 | 0.188 | 1.060 | 0.306 | 0.173 | 0.678 | 2.322 | 0.131 | 0.029 | 0.865 | |
| D/HSeptember | 1.946 | 0.167 | 2.553 | 0.114 | 9.133 | 0.003 | 2.352 | 0.129 | 0.011 | 0.915 | 1.010 | 0.318 | 0.167 | 0.683 | |
| TRlenght/HSeptember | 0.015 | 0.903 | 8.513 | 0.004 | 0.316 | 0.575 | 1.837 | 0.179 | 0.101 | 0.752 | 0.438 | 0.510 | 0.449 | 0.505 | |
| CRmass/STmass | 0.376 | 0.541 | 0.064 | 0.801 | 0.075 | 0.785 | 3.816 | 0.054 | 0.510 | 0.477 | 0.347 | 0.558 | 0.184 | 0.669 | |
| FRmass/Lmass | 0.011 | 0.917 | 0.154 | 0.696 | 0.164 | 0.687 | 3.231 | 0.076 | 0.047 | 0.829 | 0.048 | 0.827 | 0.251 | 0.617 | |
| FRmass/STmass | 0.139 | 0.710 | 0.533 | 0.467 | 0.018 | 0.894 | 4.278 | 0.042 | 0.215 | 0.644 | 0.123 | 0.727 | 0.518 | 0.474 | |
| FRmass/CRmass | 0.694 | 0.407 | 1.936 | 0.168 | 0.030 | 0.862 | 13.164 | 0.000 | 0.000 | 1.000 | 0.790 | 0.377 | 0.836 | 0.363 | |
| FRmass/L+ST+CRmass | 0.163 | 0.687 | 0.869 | 0.354 | 0.002 | 0.962 | 8.957 | 0.004 | 0.094 | 0.760 | 0.094 | 0.760 | 0.876 | 0.352 | |
| BG/AG | 0.017 | 0.896 | 0.025 | 0.876 | 0.056 | 0.814 | 0.861 | 0.356 | 0.175 | 0.677 | 0.048 | 0.827 | 0.025 | 0.874 | |
| Abbreviations: D – root collar diameter, Dincrement – root collar diameter increment, H – stem height, Hincrement – stem height increment, L – leaf, TR – tap root, ST – stem, AG – aboveground, FR – fine root, CR – coarse root, BG – beloweground, mass – dry biomass | |||||||||||||||
Table A4.
Main effects of drought (regularly watered vs. drought-treated saplings), phosphorus fertilization (saplings fetilized with phosphorus vs. non-fertilized with phosphorus), provenance (saplings originated from Karlovac vs. Slavonski Brod provenance) and its interactions on growth and allometric growth relationship parematers in sessile oak saplings, as calculated with factorial ANOVA, (F and p values). Bolded values indicate significant effects at p ˂ 0.05.
Table A4.
Main effects of drought (regularly watered vs. drought-treated saplings), phosphorus fertilization (saplings fetilized with phosphorus vs. non-fertilized with phosphorus), provenance (saplings originated from Karlovac vs. Slavonski Brod provenance) and its interactions on growth and allometric growth relationship parematers in sessile oak saplings, as calculated with factorial ANOVA, (F and p values). Bolded values indicate significant effects at p ˂ 0.05.
| Parameter | Drought (D) |
Fertilization (P) | Provenance (Pr) | D x P | D x Pr | P x Pr | D x P x Pr | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| Growth | DMarch (mm) | 0.707 | 0.403 | 2.874 | 0.094 | 0.282 | 0.597 | 0.673 | 0.414 | 0.946 | 0.333 | 0.039 | 0.844 | 0.013 | 0.910 |
| Dincrement (mm) | 6.663 | 0.011 | 0.295 | 0.589 | 0.211 | 0.647 | 0.518 | 0.474 | 0.221 | 0.639 | 0.560 | 0.456 | 0.000 | 0.990 | |
| DSeptember (mm) | 6.096 | 0.015 | 2.336 | 0.130 | 0.000 | 0.990 | 1.162 | 0.284 | 0.989 | 0.323 | 0.179 | 0.674 | 0.004 | 0.948 | |
| HMarch (cm) | 1.974 | 0.164 | 0.111 | 0.740 | 0.000 | 0.997 | 0.003 | 0.957 | 1.373 | 0.245 | 0.082 | 0.775 | 0.110 | 0.741 | |
| Hincrement (cm) | 10.623 | 0.002 | 0.094 | 0.759 | 2.407 | 0.124 | 0.109 | 0.742 | 0.000 | 0.982 | 0.003 | 0.959 | 0.198 | 0.657 | |
| HSeptember (cm) | 7.416 | 0.008 | 0.192 | 0.662 | 0.471 | 0.494 | 0.009 | 0.923 | 1.148 | 0.287 | 0.080 | 0.778 | 0.010 | 0.919 | |
| Larea (m2) | 4.404 | 0.039 | 3.847 | 0.053 | 0.565 | 0.454 | 0.193 | 0.661 | 0.603 | 0.439 | 0.010 | 0.919 | 0.771 | 0.382 | |
| TRlength (cm) | 0.209 | 0.649 | 0.468 | 0.496 | 1.591 | 0.210 | 0.133 | 0.716 | 0.147 | 0.702 | 1.950 | 0.166 | 0.096 | 0.757 | |
| Lmass (g) | 2.124 | 0.149 | 3.539 | 0.063 | 0.121 | 0.729 | 0.113 | 0.737 | 1.355 | 0.248 | 0.131 | 0.719 | 0.138 | 0.711 | |
| STmass (g) | 5.686 | 0.019 | 2.405 | 0.125 | 0.416 | 0.521 | 0.060 | 0.807 | 1.480 | 0.227 | 0.098 | 0.755 | 0.189 | 0.665 | |
| AGmass (g) | 4.442 | 0.038 | 2.930 | 0.090 | 0.306 | 0.582 | 0.081 | 0.776 | 1.510 | 0.222 | 0.115 | 0.735 | 0.179 | 0.673 | |
| FRmass (g) | 0.329 | 0.567 | 5.006 | 0.028 | 6.733 | 0.011 | 0.026 | 0.872 | 0.373 | 0.543 | 0.008 | 0.928 | 0.000 | 0.994 | |
| CRmass (g) | 3.618 | 0.060 | 1.949 | 0.166 | 0.002 | 0.965 | 0.010 | 0.919 | 0.023 | 0.879 | 0.798 | 0.374 | 0.038 | 0.846 | |
| BGmass (g) | 3.422 | 0.068 | 2.190 | 0.143 | 0.047 | 0.830 | 0.008 | 0.931 | 0.035 | 0.852 | 0.734 | 0.394 | 0.034 | 0.854 | |
Table A4.
cont.
| Parameter | Drought (D) |
Fertilization (P) | Provenance (Pr) | D x P | D x Pr | P x Pr | D x P x Pr | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| Allometric growth relationship | D/HMarch | 0.610 | 0.437 | 0.007 | 0.934 | 0.058 | 0.811 | 0.209 | 0.648 | 1.334 | 0.251 | 0.020 | 0.888 | 0.016 | 0.901 |
| Dincrement/Hincrement | 1.998 | 0.161 | 1.187 | 0.279 | 2.425 | 0.123 | 0.159 | 0.691 | 4.069 | 0.047 | 0.074 | 0.787 | 1.175 | 0.281 | |
| D/HSeptember | 0.498 | 0.482 | 0.275 | 0.602 | 0.375 | 0.542 | 0.000 | 0.994 | 0.184 | 0.669 | 0.338 | 0.562 | 0.002 | 0.969 | |
| TRlenght/HSeptember | 11.186 | 0.001 | 1.252 | 0.266 | 0.344 | 0.559 | 0.006 | 0.939 | 0.147 | 0.702 | 2.032 | 0.158 | 0.556 | 0.458 | |
| CRmass/STmass | 4.108 | 0.046 | 0.187 | 0.666 | 1.098 | 0.298 | 0.011 | 0.915 | 0.611 | 0.436 | 0.008 | 0.929 | 1.168 | 0.283 | |
| FRmass/Lmass | 0.026 | 0.873 | 0.269 | 0.605 | 7.636 | 0.007 | 0.704 | 0.404 | 0.025 | 0.875 | 0.506 | 0.479 | 0.068 | 0.795 | |
| FRmass/STmass | 2.046 | 0.156 | 0.110 | 0.741 | 13.779 | 0.000 | 0.108 | 0.743 | 0.138 | 0.711 | 1.560 | 0.215 | 0.704 | 0.404 | |
| FRmass/CRmass | 0.001 | 0.970 | 0.021 | 0.885 | 9.347 | 0.003 | 0.319 | 0.573 | 0.000 | 0.993 | 0.761 | 0.385 | 0.091 | 0.764 | |
| FRmass/L+ST+CRmass | 0.163 | 0.688 | 0.051 | 0.821 | 11.147 | 0.001 | 0.348 | 0.557 | 0.014 | 0.906 | 1.175 | 0.281 | 0.134 | 0.715 | |
| BG/AG | 0.980 | 0.325 | 0.494 | 0.484 | 1.231 | 0.270 | 0.336 | 0.564 | 0.791 | 0.376 | 0.005 | 0.943 | 0.487 | 0.487 | |
| Abbreviations: D – root collar diameter, Dincrement – root collar diameter increment, H – stem height, Hincrement – stem height increment, L – leaf, TR – tap root, ST – stem, AG – aboveground, FR – fine root, CR – coarse root, BG – beloweground, mass – dry biomass | |||||||||||||||
References
- Pretzsch, H.; Bielak, K.; Block, J.; Bruchwald, A.; Dieler, J.; Ehrhart, H.-P.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zasada, M.; et al. Productivity of Mixed versus Pure Stands of Oak (Quercus Petraea (Matt. ) Liebl. and Quercus Robur L.) and European Beech (Fagus Sylvatica L.) along an Ecological Gradient. Eur J Forest Res 2013, 132, 263–280. [Google Scholar] [CrossRef]
- Hajek, P.; Link, R.M.; Nock, C.A.; Bauhus, J.; Gebauer, T.; Gessler, A.; Kovach, K.; Messier, C.; Paquette, A.; Saurer, M.; et al. Mutually Inclusive Mechanisms of Drought-induced Tree Mortality. Global Change Biology 2022, 28, 3365–3378. [Google Scholar] [CrossRef] [PubMed]
- Kasper, J.; Leuschner, C.; Walentowski, H.; Petritan, A.M.; Weigel, R. Winners and Losers of Climate Warming: Declining Growth in Fagus and Tilia vs. Stable Growth in Three Quercus Species in the Natural Beech–Oak Forest Ecotone (Western Romania). Forest Ecology and Management 2022, 506, 119892. [Google Scholar] [CrossRef]
- Rukh, S.; Sanders, T.G.M.; Krüger, I.; Schad, T.; Bolte, A. Distinct Responses of European Beech (Fagus Sylvatica L. ) to Drought Intensity and Length—A Review of the Impacts of the 2003 and 2018–2019 Drought Events in Central Europe. Forests 2023, 14, 248. [Google Scholar] [CrossRef]
- Hanel, M.; Rakovec, O.; Markonis, Y.; Máca, P.; Samaniego, L.; Kyselý, J.; Kumar, R. Revisiting the Recent European Droughts from a Long-Term Perspective. Sci Rep 2018, 8, 9499. [Google Scholar] [CrossRef] [PubMed]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A First Assessment of the Impact of the Extreme 2018 Summer Drought on Central European Forests. Basic and Applied Ecology 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Meyer, P.; Spînu, A.P.; Mölder, A.; Bauhus, J. Management Alters Drought-induced Mortality Patterns in European Beech ( Fagus Sylvatica L. ) Forests. Plant Biol J 2022, 24, 1157–1170. [Google Scholar] [CrossRef] [PubMed]
- Beloiu, M.; Stahlmann, R.; Beierkuhnlein, C. High Recovery of Saplings after Severe Drought in Temperate Deciduous Forests. Forests 2020, 11, 546. [Google Scholar] [CrossRef]
- Rohner, B.; Kumar, S.; Liechti, K.; Gessler, A.; Ferretti, M. Tree Vitality Indicators Revealed a Rapid Response of Beech Forests to the 2018 Drought. Ecological Indicators 2021, 120, 106903. [Google Scholar] [CrossRef]
- Wiley, E. Do Carbon Reserves Increase Tree Survival during Stress and Following Disturbance? Curr Forestry Rep 2020, 6, 14–25. [Google Scholar] [CrossRef]
- Schönbeck, L.; Li, M.-H.; Lehmann, M.M.; Rigling, A.; Schaub, M.; Hoch, G.; Kahmen, A.; Gessler, A. Soil Nutrient Availability Alters Tree Carbon Allocation Dynamics during Drought. Tree Physiology 2021, 41, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Galle, A.; Esper, J.; Feller, U.; Ribas-Carbo, M.; Fonti, P. Responses of Wood Anatomy and Carbon Isotope Composition of Quercus Pubescens Saplings Subjected to Two Consecutive Years of Summer Drought. Ann. For. Sci. 2010, 67, 809–809. [Google Scholar] [CrossRef]
- Wilkinson, S.; Ogee, J.; Domec, J.-C.; Rayment, M.; Wingate, L. Biophysical Modelling of Intra-Ring Variations in Tracheid Features and Wood Density of Pinus Pinaster Trees Exposed to Seasonal Droughts. Tree Physiology 2015, 35, 305–318. [Google Scholar] [CrossRef]
- Redmond, M.D.; Weisberg, P.J.; Cobb, N.S.; Clifford, M.J. Woodland Resilience to Regional Drought: Dominant Controls on Tree Regeneration Following Overstorey Mortality. Journal of Ecology 2018, 106, 625–639. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Turcsán, A.; Erdélyi, É.; Beeckman, H. Drought Treated Seedlings of Quercus Petraea (Matt. ) Liebl., Q. Robur L. and Their Morphological Intermediates Show Differential Radial Growth and Wood Anatomical Traits. Forests 2020, 11, 250. [Google Scholar] [CrossRef]
- Petrík, P.; Grote, R.; Gömöry, D.; Kurjak, D.; Petek-Petrik, A.; Lamarque, L.J.; Sliacka Konôpková, A.; Mukarram, M.; Debta, H.; Fleischer, P. The Role of Provenance for the Projected Growth of Juvenile European Beech under Climate Change. Forests 2022, 14, 26. [Google Scholar] [CrossRef]
- Thiel, D.; Kreyling, J.; Backhaus, S.; Beierkuhnlein, C.; Buhk, C.; Egen, K.; Huber, G.; Konnert, M.; Nagy, L.; Jentsch, A. Different Reactions of Central and Marginal Provenances of Fagus Sylvatica to Experimental Drought. Eur J Forest Res 2014, 133, 247–260. [Google Scholar] [CrossRef]
- Wang, F.; Israel, D.; Ramírez-Valiente, J.-A.; Sánchez-Gómez, D.; Aranda, I.; Aphalo, P.J.; Robson, T.M. Seedlings from Marginal and Core Populations of European Beech (Fagus Sylvatica L. ) Respond Differently to Imposed Drought and Shade. Trees 2021, 35, 53–67. [Google Scholar] [CrossRef]
- Sever, K.; Vukmirović, A.; Hodak, L.; Bogdan, S.; Katičić Bogdan, I.; Krstonošić, D.; Karažija, T.; Franjić, J.; Škvorc, Ž. Funkcionalna Prilagodba Prirodnog Pomlatka Hrasta Kitnjaka i Obične Bukve na Različite Stanišne Prilike. Šumar. list (Online) 2022, 146, 293–307. [Google Scholar] [CrossRef]
- Arend, M.; Kuster, T.; Gunthardt-Goerg, M.S.; Dobbertin, M. Provenance-Specific Growth Responses to Drought and Air Warming in Three European Oak Species (Quercus Robur, Q. Petraea and Q. Pubescens). Tree Physiology 2011, 31, 287–297. [Google Scholar] [CrossRef]
- Günthardt-Goerg, M.S.; Kuster, T.M.; Arend, M.; Vollenweider, P. Foliage Response of Young Central European Oaks to Air Warming, Drought and Soil Type. Plant Biology 2013, 15, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Kuster, T.M.; Arend, M.; Günthardt-Goerg, M.S.; Schulin, R. Root Growth of Different Oak Provenances in Two Soils under Drought Stress and Air Warming Conditions. Plant Soil 2013, 369, 61–71. [Google Scholar] [CrossRef]
- Stojnić, S.; Orlović, S.; Miljković, D.; Galić, Z.; Kebert, M.; Von Wuehlisch, G. Provenance Plasticity of European Beech Leaf Traits under Differing Environmental Conditions at Two Serbian Common Garden Sites. Eur J Forest Res 2015, 134, 1109–1125. [Google Scholar] [CrossRef]
- Stojnic, S.; Orlovic, S.; Miljkovic, D.; Von, W. Intra- and Interprovenance Variations in Leaf Morphometric Traits in European Beech (Fagus Sylvatica L. ). Arch biol sci (Beogr) 2016, 68, 781–788. [Google Scholar] [CrossRef]
- Stojnić, S.; Suchocka, M.; Benito-Garzón, M.; Torres-Ruiz, J.M.; Cochard, H.; Bolte, A.; Cocozza, C.; Cvjetković, B.; De Luis, M.; Martinez-Vilalta, J.; et al. Variation in Xylem Vulnerability to Embolism in European Beech from Geographically Marginal Populations. Tree Physiology 2018, 38, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, P.; Menard, T.; Arend, M.; Kuster, T.M.; Günthardt-Goerg, M.S. Structural Changes Associated with Drought Stress Symptoms in Foliage of Central European Oaks. Trees 2016, 30, 883–900. [Google Scholar] [CrossRef]
- Rabarijaona, A.; Ponton, S.; Bert, D.; Ducousso, A.; Richard, B.; Levillain, J.; Brendel, O. Provenance Differences in Water-Use Efficiency Among Sessile Oak Populations Grown in a Mesic Common Garden. Front. For. Glob. Change 2022, 5, 914199. [Google Scholar] [CrossRef]
- Montwé, D.; Isaac-Renton, M.; Hamann, A.; Spiecker, H. Drought Tolerance and Growth in Populations of a Wide-ranging Tree Species Indicate Climate Change Risks for the Boreal North. Global Change Biology 2016, 22, 806–815. [Google Scholar] [CrossRef]
- Csilléry, K.; Buchmann, N.; Fady, B. Adaptation to Drought Is Coupled with Slow Growth, but Independent from Phenology in Marginal Silver Fir ( Abies Alba Mill. ) Populations. Evolutionary Applications 2020, 13, 2357–2376. [Google Scholar] [CrossRef]
- Eilmann, B.; de Vries, S.M.G.; den Ouden, J.; Mohren, G.M.J.; Sauren, P.; Sass-Klaassen, U. Origin Matters! Difference in Drought Tolerance and Productivity of Coastal Douglas-Fir (Pseudotsuga Menziesii (Mirb.)) Provenances. Forest Ecology and Management 2013, 302, 133–143. [Google Scholar] [CrossRef]
- Montwé, D.; Spiecker, H.; Hamann, A. Five Decades of Growth in a Genetic Field Trial of Douglas-Fir Reveal Trade-Offs between Productivity and Drought Tolerance. Tree Genetics & Genomes 2015, 11, 29. [Google Scholar] [CrossRef]
- Haase, D.L.; Rose, R.; Trobaugh, J. Field Performance of Three Stock Sizes of Douglas-Fir Container Seedlings Grown with Slow-Release Fertilizer in the Nursery Growing Medium. New Forest 2006, 31, 1–24. [Google Scholar] [CrossRef]
- Seletković, I.; Potočić, N.; Jazbec, A.; Ćosić, T.; Jakovljević, T. Utjecaj različitih sjetvenih supstrata i vrsta sporo topivih gnojiva na rast i fiziološke parametre sadnica obične bukve (Fagus Sylvatica L.) u rasadniku i nakon presadnje. Šum list 2009, 9–10: 469 – 481.
- Schmal, J.L.; Jacobs, D.F.; O’Reilly, C. Nitrogen Budgeting and Quality of Exponentially Fertilized Quercus Robur Seedlings in Ireland. Eur J Forest Res 2011, 130, 557–567. [Google Scholar] [CrossRef]
- Uscola, M.; Salifu, K.F.; Oliet, J.A.; Jacobs, D.F. An Exponential Fertilization Dose–Response Model to Promote Restoration of the Mediterranean Oak Quercus Ilex. New Forests 2015, 46, 795–812. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Puértolas, J.; Peñuelas, J.L.; Planelles, R. Effect of Nitrogen Fertilization in the Nursery on the Drought and Frost Resistance of Mediterranean Forest Species. Invest. Agrar.: Sist. Recur. For. 2005, 14, 408. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Peñuelas, J.L.; Nicolás-Peragón, J.L.; Benito, L.F.; Domínguez-Lerena, S. Is Nitrogen Fertilization in the Nursery a Suitable Tool for Enhancing the Performance of Mediterranean Oak Plantations? New Forests 2013, 44, 733–751. [Google Scholar] [CrossRef]
- Fini, A.; Ferrini, F.; Di Ferdinando, M.; Brunetti, C.; Giordano, C.; Gerini, F.; Tattini, M. Acclimation to Partial Shading or Full Sunlight Determines the Performance of Container-Grown Fraxinus Ornus to Subsequent Drought Stress. Urban Forestry & Urban Greening 2014, 13, 63–70. [Google Scholar] [CrossRef]
- Salto, C.S.; Sagadin, M.B.; Luna, C.M.; Oberschelp, G.P.J.; Harrand, L.; Cabello, M.N. Interactions between Mineral Fertilization and Arbuscular Mycorrhizal Fungi Improve Nursery Growth and Drought Tolerance of Prosopis Alba Seedlings. Agroforest Syst 2020, 94, 103–111. [Google Scholar] [CrossRef]
- Haase, D. L. Morphological and Physiological Evaluations of Seedling Quality. USDA Forest Service Proceedings RMRS P-50 2007, 3–8.
- Drvodelić, D.; Oršanić, M. Izbor Kvalitetne Šumske Sadnice Poljskog Jasena ( Fraxinus Angustifolia Vahl) Za Umjetnu Obnovu i Pošumljavanje. Šumar. list (Online) 2019, 143, 577–585. [Google Scholar] [CrossRef]
- Oliet, J.A.; Salazar, J.M.; Villar, R.; Robredo, E.; Valladares, F. Fall Fertilization of Holm Oak Affects N and P Dynamics, Root Growth Potential, and Post-Planting Phenology and Growth. Annals of Forest Science 2011, 68, 647–656. [Google Scholar] [CrossRef]
- Zadworny, M.; Mucha, J.; Jagodziński, A.M.; Kościelniak, P.; Łakomy, P.; Modrzejewski, M.; Ufnalski, K.; Żytkowiak, R.; Comas, L.H.; Rodríguez-Calcerrada, J. Seedling Regeneration Techniques Affect Root Systems and the Response of Quercus Robur Seedlings to Water Shortages. Forest Ecology and Management 2021, 479, 118552. [Google Scholar] [CrossRef]
- Catovsky, S.; Kobe, R.K.; Bazzaz, F.A. Nitrogen-Induced Changes In Seedling Regeneration And Dynamics Of Mixed Conifer-Broad-Leaved Forests. Ecological Applications 2002, 12, 1611–1625. [Google Scholar] [CrossRef]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.L.; Tanner, E.V.J.; Sayer, E.J.; Santiago, L.S.; Kaspari, M.; Hedin, L.O.; Harms, K.E.; et al. Potassium, Phosphorus, or Nitrogen Limit Root Allocation, Tree Growth, or Litter Production in a Lowland Tropical Forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yao, F.; Wu, J.; Zhang, P.; Xu, W. Effect of Nitrogen Levels on Photosynthetic Parameters, Morphological and Chemical Characters of Saplings and Trees in a Temperate Forest. J. For. Res. 2018, 29, 1481–1488. [Google Scholar] [CrossRef]
- Brown, K.R.; Van Den Driessche, R. Effects of Nitrogen and Phosphorus Fertilization on the Growth and Nutrition of Hybrid Poplars on Vancouver Island. New Forest 2005, 29, 89–104. [Google Scholar] [CrossRef]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Sun, X.; Song, D.; Chen, W.; Zhang, A.; et al. Phosphorous Application Improves Drought Tolerance of Phoebe Zhennan. Front. Plant Sci. 2017, 8, 1561. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Zhang, L.; Wu, X.; Chen, W.; Song, D.; et al. Phosphorous Fertilization Alleviates Drought Effects on Alnus Cremastogyne by Regulating Its Antioxidant and Osmotic Potential. Sci Rep 2018, 8, 5644. [Google Scholar] [CrossRef]
- Zavišić, A.; Yang, N.; Marhan, S.; Kandeler, E.; Polle, A. Forest Soil Phosphorus Resources and Fertilization Affect Ectomycorrhizal Community Composition, Beech P Uptake Efficiency, and Photosynthesis. Front. Plant Sci. 2018, 9, 463. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant Growth under Drought Stress: Significance of Mineral Nutrients. In Water Stress and Crop Plants; Ahmad, P., Ed.; Wiley, 2016; pp. 649–668 ISBN 978-1-119-05436-8. [CrossRef]
- Kayoumu, M.; Iqbal, A.; Muhammad, N.; Li, X.; Li, L.; Wang, X.; Gui, H.; Qi, Q.; Ruan, S.; Guo, R.; et al. Phosphorus Availability Affects the Photosynthesis and Antioxidant System of Contrasting Low-P-Tolerant Cotton Genotypes. Antioxidants 2023, 12, 466. [Google Scholar] [CrossRef]
- Sawwan, J.; Shibli, R.A.; Swaidat, I.; Tahat, M. Phosphorus Regulates Osmotic Potential and Growth of African Violet under in Vitro-induced Water Deficit. Journal of Plant Nutrition 2000, 23, 759–771. [Google Scholar] [CrossRef]
- Singh, V.; Pallaghy, C.K.; Singh, D. Phosphorus Nutrition and Tolerance of Cotton to Water Stress. Field Crops Research 2006, 96, 191–198. [Google Scholar] [CrossRef]
- Silveira, J.A.G.; Matos, J.C.S.; Cecatto, V.M.; Viegas, R.A.; Oliveira, J.T.A. Nitrate Reductase Activity, Distribution, and Response to Nitrate in Two Contrasting Phaseolus Species Inoculated with Rhizobium Spp. Environmental and Experimental Botany 2001, 46, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.; Khan, M.M.A. Phosphorus Ameliorates Crop Productivity, Photosynthesis, Nitrate Reductase Activity and Nutrient Accumulation in Coffee Senna ( Senna Occidentalis L. ) under Phosphorus-Deficient Soil. Journal of Plant Interactions 2009, 4, 145–153. [Google Scholar] [CrossRef]
- Oliveira, M.T.; Medeiros, C.D.; Frosi, G.; Santos, M.G. Different Mechanisms Drive the Performance of Native and Invasive Woody Species in Response to Leaf Phosphorus Supply during Periods of Drought Stress and Recovery. Plant Physiology and Biochemistry 2014, 82, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Faustino, L.I.; Bulfe, N.M.L.; Pinazo, M.A.; Monteoliva, S.E.; Graciano, C. Dry Weight Partitioning and Hydraulic Traits in Young Pinus Taeda Trees Fertilized with Nitrogen and Phosphorus in a Subtropical Area. Tree Physiology 2013, 33, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Badgujar, G.B.; Reddy, V.R.; Fleisher, D.H.; Timlin, D.J. Effect of Phosphorus Nutrition on Growth and Physiology of Cotton Under Ambient and Elevated Carbon Dioxide. J Agronomy Crop Science 2013, 199, 436–448. [Google Scholar] [CrossRef]
- Singh, D.K.; Sale, P.W.G. Phosphorus Supply and the Growth of Frequently Defoliated White Clover (Trifolium repens L. ) in Dry Soil. Plant and Soil 1998, 205, 155–162. [Google Scholar] [CrossRef]
- Kang, L.; Yue, S.; Li, S. Effects of Phosphorus Application in Different Soil Layers on Root Growth, Yield, and Water-Use Efficiency of Winter Wheat Grown Under Semi-Arid Conditions. Journal of Integrative Agriculture 2014, 13, 2028–2039. [Google Scholar] [CrossRef]
- Shukla, D.; Rinehart, C.A.; Sahi, S.V. Comprehensive Study of Excess Phosphate Response Reveals Ethylene Mediated Signaling That Negatively Regulates Plant Growth and Development. Sci Rep 2017, 7, 3074. [Google Scholar] [CrossRef]
- Takagi, D.; Miyagi, A.; Tazoe, Y.; Suganami, M.; Kawai-Yamada, M.; Ueda, A.; Suzuki, Y.; Noguchi, K.; Hirotsu, N.; Makino, A. Phosphorus Toxicity Disrupts Rubisco Activation and Reactive Oxygen Species Defence Systems by Phytic Acid Accumulation in Leaves. Plant Cell & Environment 2020, 43, 2033–2053. [Google Scholar] [CrossRef]
- Lambers, H. Phosphorus Acquisition and Utilization in Plants. Annu. Rev. Plant Biol. 2022, 73, 17–42. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zavišić, A.; Pena, R.; Polle, A. Phenology, Photosynthesis, and Phosphorus in European Beech ( Fagus Sylvatica L. ) in Two Forest Soils with Contrasting P Contents. J. Plant Nutr. Soil Sci. 2016, 179, 151–158. [Google Scholar] [CrossRef]
- Rieger, I.; Kowarik, I.; Ziche, D.; Wellbrock, N.; Cierjacks, A. Linkages between Phosphorus and Plant Diversity in Central European Forest Ecosystems—Complementarity or Competition? Forests 2019, 10, 1156. [Google Scholar] [CrossRef]
- Ognjenović, M. 2021. Nutritional Status and Defoliation of Common Beech (Fagus Sylvatica L.) in Changed Climate Conditions. Doctoral Thesis, University of Zagreb, Faculty of Forestry and Wood Technology, Croatia, 07. 09.2017. [Google Scholar]
- Prietzel, J.; Krüger, J.; Kaiser, K.; Amelung, W.; Bauke, S.L.; Dippold, M.A.; Kandeler, E.; Klysubun, W.; Lewandowski, H.; Löppmann, S.; et al. Soil Phosphorus Status and P Nutrition Strategies of European Beech Forests on Carbonate Compared to Silicate Parent Material. Biogeochemistry 2022, 158, 39–72. [Google Scholar] [CrossRef]
- Spohn, M.; Stendahl, J. Spatial Patterns of Nitrogen Isotope Ratios in Forest Soils Are Related to Latitude and Soil Phosphorus Concentration. Biogeochemistry 2023, 165, 43–56. [Google Scholar] [CrossRef]
- Zang, U.; Goisser, M.; Meyer, N.; Häberle, K.-H.; Borken, W. Chemical and Morphological Response of Beech Saplings (Fagus Sylvatica L. ) to an Experimental Soil Drought Gradient. Forest Ecology and Management 2021, 498, 119569. [Google Scholar] [CrossRef]
- Newnham, R.M.; Carlisle, A. The Nitrogen and Phosphorus Nutrition of Seedlings of Quercus Robur L. and Q. Petraea (Mattuschka) Liebl. The Journal of Ecology 1969, 57, 271. [Google Scholar] [CrossRef]
- Peuke, A.D.; Rennenberg, H. Carbon, Nitrogen, Phosphorus, and Sulphur Concentration and Partitioning in Beech Ecotypes (Fagus Sylvatica L. ): Phosphorus Most Affected by Drought. Trees 2004, 18, 639–648. [Google Scholar] [CrossRef]
- Netzer, F.; Herschbach, C.; Oikawa, A.; Okazaki, Y.; Dubbert, D.; Saito, K.; Rennenberg, H. Seasonal Alterations in Organic Phosphorus Metabolism Drive the Phosphorus Economy of Annual Growth in F. Sylvatica Trees on P-Impoverished Soil. Front. Plant Sci. 2018, 9, 723. [Google Scholar] [CrossRef]
- Zavišić, A.; Polle, A. Dynamics of Phosphorus Nutrition, Allocation and Growth of Young Beech (Fagus Sylvatica L. ) Trees in P-Rich and P-Poor Forest Soil. Tree Physiology 2018, 38, 37–51. [Google Scholar] [CrossRef]
- Meller, S.; Frossard, E.; Luster, J. Phosphorus Allocation to Leaves of Beech Saplings Reacts to Soil Phosphorus Availability. Front. Plant Sci. 2019, 10, 744. [Google Scholar] [CrossRef]
- Yang, F.; Magh, R.-K.; Ivanković, M.; Lanšćak, M.; Haberstroh, S.; Du, B.; Dannenmann, M.; Rennenberg, H.; Herschbach, C. Foliar P Nutrition of European Beech (Fagus Sylvatica L. ) Depends on the Season but Remains Unaffected by Co-Cultivation with Silver Fir (Abies Alba Mill.). Eur J Forest Res 2020, 139, 853–868. [Google Scholar] [CrossRef]
- Bačurin, M.; Bogdan, S.; Katičić Bogdan, I.; Sever, K. Leaf Phenological Responses of Juvenile Beech and Oak Provenances to Elevated Phosphorus. Forests 2023, 14, 834. [Google Scholar] [CrossRef]
- Page, A.L.; Miller, R.H.; Keeney, D.R. (1982) Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties. American Society of Agronomy 1982; In Soil Science Society of America, Vol. 1159.
- Mellert, K.H.; Göttlein, A. Comparison of New Foliar Nutrient Thresholds Derived from van Den Burg’s Literature Compilation with Established Central European References. Eur J Forest Res 2012, 131, 1461–1472. [Google Scholar] [CrossRef]
- Epron, D.; Dreyer, E. Long-term Effects of Drought on Photosynthesis of Adult Oak Trees [ Quercus Petraea (Matt. ) Liebl. and Quercus Robur L.] in a Natural Stand. New Phytologist 1993, 125, 381–389. [Google Scholar] [CrossRef]
- Arend, M.; Brem, A.; Kuster, T.M.; Günthardt-Goerg, M.S. Seasonal Photosynthetic Responses of European Oaks to Drought and Elevated Daytime Temperature. Plant Biology 2013, 15, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Arend, M.; Sever, K.; Pflug, E.; Gessler, A.; Schaub, M. Seasonal Photosynthetic Response of European Beech to Severe Summer Drought: Limitation, Recovery and Post-Drought Stimulation. Agricultural and Forest Meteorology 2016, 220, 83–89. [Google Scholar] [CrossRef]
- Cocozza, C.; De Miguel, M.; Pšidová, E.; Ditmarová, L.; Marino, S.; Maiuro, L.; Alvino, A.; Czajkowski, T.; Bolte, A.; Tognetti, R. Variation in Ecophysiological Traits and Drought Tolerance of Beech (Fagus Sylvatica L. ) Seedlings from Different Populations. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Pflug, E.E.; Buchmann, N.; Siegwolf, R.T.W.; Schaub, M.; Rigling, A.; Arend, M. Resilient Leaf Physiological Response of European Beech (Fagus Sylvatica L. ) to Summer Drought and Drought Release. Front. Plant Sci. 2018, 9, 187. [Google Scholar] [CrossRef]
- Walthert, L.; Ganthaler, A.; Mayr, S.; Saurer, M.; Waldner, P.; Walser, M.; Zweifel, R.; Von Arx, G. From the Comfort Zone to Crown Dieback: Sequence of Physiological Stress Thresholds in Mature European Beech Trees across Progressive Drought. Science of The Total Environment 2021, 753, 141792. [Google Scholar] [CrossRef]
- Van Hees, A. Growth and Morphology of Pedunculate Oak (Quercus Robur L) and Beech (Fagus Sylvatica L) Seedlings in Relation to Shading and Drought. Ann. For. Sci. 1997, 54, 9–18. [Google Scholar] [CrossRef]
- Thomas, F.M.; Gausling, T. Morphological and Physiological Responses of Oak Seedlings ( Quercus Petraea and Q. Robur ) to Moderate Drought. Ann. For. Sci. 2000, 57, 325–333. [Google Scholar] [CrossRef]
- Nielsen, C.N.; Jørgensen, F.V. Phenology and Diameter Increment in Seedlings of European Beech (Fagus Sylvatica L. ) as Affected by Different Soil Water Contents: Variation between and within Provenances. Forest Ecology and Management 2003, 174, 233–249. [Google Scholar] [CrossRef]
- Rose, L.; Leuschner, C.; Köckemann, B.; Buschmann, H. Are Marginal Beech (Fagus Sylvatica L. ) Provenances a Source for Drought Tolerant Ecotypes? Eur J Forest Res 2009, 128, 335–343. [Google Scholar] [CrossRef]
- Jensen, J.S.; Hansen, J.K. Genetic Variation in Responses to Different Soil Water Treatments in Quercus Robur L. Scandinavian Journal of Forest Research 2010, 25, 400–411. [Google Scholar] [CrossRef]
- Bruschi, P. Geographical Variation in Morphology of Quercus Petraea (Matt. ) Liebl. as Related to Drought Stress. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology 2010, 144, 298–307. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How Tree Roots Respond to Drought. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Le Hir, R.; Leduc, N.; Jeannette, E.; Viemont, J.-D.; Pelleschi-Travier, S. Variations in Sucrose and ABA Concentrations Are Concomitant with Heteroblastic Leaf Shape Changes in a Rhythmically Growing Species (Quercus Robur). Tree Physiology 2006, 26, 229–238. [Google Scholar] [CrossRef]
- Kuster, T.M.; Dobbertin, M.; Günthardt-Goerg, M.S.; Schaub, M.; Arend, M. A Phenological Timetable of Oak Growth under Experimental Drought and Air Warming. PLoS ONE 2014, 9, e89724. [Google Scholar] [CrossRef]
- Sever, K.; Bogdan, S.; Škvorc, Ž.; Sever, M.Z.O.; Franjić, J. Estimation of Leaf Nitrogen Concentrations in Quercus Robur L. Using the CCM-200 Portable Chlorophyll Meter for Different Patterns of Vegetative Growth and Acorn Production. New Forests 2016, 47, 513–527. [Google Scholar] [CrossRef]
- Lavarenne-Allary, S. Recherches Sur La Croissance Des Bourgeons de Chêne et de Quelques Autres Espèces Ligneuses. Ann. Sci. forest. 1965, 22, 7–203. [Google Scholar] [CrossRef]
- Harmer, R.; Baker, C. An Evaluation of Decapitation as a Method for Selecting Clonal Quercus Petraea (Matt) Liebl with Different Branching Intensities. Ann. For. Sci. 1995, 52, 89–102. [Google Scholar] [CrossRef]
- Collet, C.; Colin, F.; Bernier, F. Height Growth, Shoot Elongation and Branch Development of Young Quercus Petraea Grown under Different Levels of Resource Availability. Ann. For. Sci. 1997, 54, 65–81. [Google Scholar] [CrossRef]
- Mansour, A. ; de Faÿ, Elisabeth Rhythmic Growth Rings of Wood and Their Relationship with the Foliage in Oak Seedlings Grown in a Favourable Environment. Annals of Botany 1998, 82, 89–96. [Google Scholar] [CrossRef]
- Mikhalevskaya, O.B. Growth Rhythms at Different Stages of Shoot Morphogenesis in Woody Plants. Russ J Dev Biol 2008, 39, 65–72. [Google Scholar] [CrossRef]
- Spieß, N.; Oufir, M.; Matušíková, I.; Stierschneider, M.; Kopecky, D.; Homolka, A.; Burg, K.; Fluch, S.; Hausman, J.-F.; Wilhelm, E. Ecophysiological and Transcriptomic Responses of Oak (Quercus Robur) to Long-Term Drought Exposure and Rewatering. Environmental and Experimental Botany 2012, 77, 117–126. [Google Scholar] [CrossRef]
- Turcsán, A.; Steppe, K.; Sárközi, E.; Erdélyi, É.; Missoorten, M.; Mees, G.; Mijnsbrugge, K.V. Early Summer Drought Stress During the First Growing Year Stimulates Extra Shoot Growth in Oak Seedlings (Quercus Petraea). Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Gaytán, Á.; Moreira, X.; Castagneyrol, B.; Van Halder, I.; De Frenne, P.; Meeussen, C.; Timmermans, B.G.H.; Ten Hoopen, J.P.J.G.; Rasmussen, P.U.; Bos, N.; et al. The Co-existence of Multiple Oak Leaf Flushes Contributes to the Large Within-tree Variation in Chemistry, Insect Attack and Pathogen Infection. New Phytologist 2022, 235, 1615–1628. [Google Scholar] [CrossRef] [PubMed]
- Lloret, F.; Casanovas, C.; Peñuelas, J. Seedling Survival of Mediterranean Shrubland Species in Relation to Root:Shoot Ratio, Seed Size and Water and Nitrogen Use. Functional Ecology 1999, 13, 210–216. [Google Scholar] [CrossRef]
- Thomas, F.M. Growth and Water Relations of Four Deciduous Tree Species (Fagus Sylvatica L. , Quercus Petraea [Matt.] Liebl., Q. Pubescens Willd., Sorbus Aria [L.] Cr.) Occurring at Central-European Tree-Line Sites on Shallow Calcareous Soils: Physiological Reactions of Seedlings to Severe Drought. Flora 2000, 195, 104–115. [Google Scholar] [CrossRef]
- Brown, K.R.; Van Den Driessche, R. Effects of Nitrogen and Phosphorus Fertilization on the Growth and Nutrition of Hybrid Poplars on Vancouver Island. New Forest 2005, 29, 89–104. [Google Scholar] [CrossRef]
- Goswami, S.; Fisk, M.C.; Vadeboncoeur, M.A.; Garrison-Johnston, M.; Yanai, R.D.; Fahey, T.J. Phosphorus Limitation of Aboveground Production in Northern Hardwood Forests. Ecology 2018, 99, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Groves, R.; Keraitis, K. Survival and Growth of Seedlings of Three Sclerophyll Species at High Levels of Phosphorus and Nitrogen. Aust. J. Bot. 1976, 24, 681. [Google Scholar] [CrossRef]
- Nichols, D.G.; Beardsell, D.V. The Response of Phosphorus-Sensitive Plants to Slow-Release Fertilizers in Soil-Less Potting Mixtures. Scientia Horticulturae 1981, 15, 301–309. [Google Scholar] [CrossRef]
- Hawkins, H.-J.; Hettasch, H.; Mesjasz-Przybylowicz, J.; Przybylowicz, W.; Cramer, M.D. Phosphorus Toxicity in the Proteaceae: A Problem in Post-Agricultural Lands. Scientia Horticulturae 2008, 117, 357–365. [Google Scholar] [CrossRef]
- Lambers, H.; Raven, J.; Shaver, G.; Smith, S. Plant Nutrient-Acquisition Strategies Change with Soil Age. Trends in Ecology & Evolution 2008, 23, 95–103. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, D.; Xiong, Z.; Wang, Z.; Gao, M. Changes in Rhizosphere Phosphorus Fractions and Phosphate-Mineralizing Microbial Populations in Acid Soil as Influenced by Organic Acid Exudation. Soil and Tillage Research 2023, 225, 105543. [Google Scholar] [CrossRef]
- André, F.; Jonard, M.; Ponette, Q. Biomass and Nutrient Content of Sessile Oak (Quercus Petraea (Matt. ) Liebl.) and Beech (Fagus Sylvatica L.) Stem and Branches in a Mixed Stand in Southern Belgium. Science of The Total Environment 2010, 408, 2285–2294. [Google Scholar] [CrossRef]
- Hofmann, K.; Heuck, C.; Spohn, M. Phosphorus Resorption by Young Beech Trees and Soil Phosphatase Activity as Dependent on Phosphorus Availability. Oecologia 2016, 181, 369–379. [Google Scholar] [CrossRef]
- Güsewell, S. N : P Ratios in Terrestrial Plants: Variation and Functional Significance. New Phytologist 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Leuschner, C.; Backes, K.; Hertel, D.; Schipka, F.; Schmitt, U.; Terborg, O.; Runge, M. Drought Responses at Leaf, Stem and Fine Root Levels of Competitive Fagus Sylvatica L. and Quercus Petraea (Matt.) Liebl. Trees in Dry and Wet Years. Forest Ecology and Management 2001, 149, 33–46. [Google Scholar] [CrossRef]
- Mainiero, R.; Kazda, M. Depth-Related Fine Root Dynamics of Fagus Sylvatica during Exceptional Drought. Forest Ecology and Management 2006, 237, 135–142. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Leaf Size and Leaf Area Index in Fagus Sylvatica Forests: Competing Effects of Precipitation, Temperature, and Nitrogen Availability. Ecosystems 2008, 11, 655–669. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Belowground Drought Response of European Beech: Fine Root Biomass and Carbon Partitioning in 14 Mature Stands across a Precipitation Gradient: Belowground Drought Response of Beech. Global Change Biology 2008, 14, 2081–2095. [Google Scholar] [CrossRef]
- Zang, U.; Goisser, M.; Häberle, K.; Matyssek, R.; Matzner, E.; Borken, W. Effects of Drought Stress on Photosynthesis, Rhizosphere Respiration, and Fine-root Characteristics of Beech Saplings: A Rhizotron Field Study. Z. Pflanzenernähr. Bodenk. 2014, 177, 168–177. [Google Scholar] [CrossRef]
- Zwetsloot, M.J.; Bauerle, T.L. Repetitive Seasonal Drought Causes Substantial Species-specific Shifts in Fine-root Longevity and Spatio-temporal Production Patterns in Mature Temperate Forest Trees. New Phytologist 2021, 231, 974–986. [Google Scholar] [CrossRef] [PubMed]
- Calvaruso, C.; Kirchen, G.; Saint-André, L.; Redon, P.-O.; Turpault, M.-P. Relationship between Soil Nutritive Resources and the Growth and Mineral Nutrition of a Beech ( Fagus Sylvatica ) Stand along a Soil Sequence. CATENA 2017, 155, 156–169. [Google Scholar] [CrossRef]
- Weemstra, M.; Sterck, F.J.; Visser, E.J.W.; Kuyper, T.W.; Goudzwaard, L.; Mommer, L. Fine-Root Trait Plasticity of Beech (Fagus Sylvatica) and Spruce (Picea Abies) Forests on Two Contrasting Soils. Plant Soil 2017, 415, 175–188. [Google Scholar] [CrossRef]
- Likulunga, L.E.; Clausing, S.; Krüger, J.; Lang, F.; Polle, A. Fine Root Biomass of European Beech Trees in Different Soil Layers Show Different Responses to Season, Climate, and Soil Nutrients. Front. For. Glob. Change 2022, 5, 955327. [Google Scholar] [CrossRef]
- Moore, J.R.; Tombleson, J.D.; Turner, J.A.; Van Der Colff, M. Wind Effects on Juvenile Trees: A Review with Special Reference to Toppling of Radiata Pine Growing in New Zealand. Forestry 2008, 81, 377–387. [Google Scholar] [CrossRef]
- Bécel, C.; Vercambre, G.; Pagès, L. Soil Penetration Resistance, a Suitable Soil Property to Account for Variations in Root Elongation and Branching. Plant Soil 2012, 353, 169–180. [Google Scholar] [CrossRef]
- Ostrogović Sever, M.Z.; Alberti, G.; Delle Vedove, G.; Marjanović, H. Temporal Evolution of Carbon Stocks, Fluxes and Carbon Balance in Pedunculate Oak Chronosequence under Close-To-Nature Forest Management. Forests 2019, 10, 814. [Google Scholar] [CrossRef]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and Promises of Epigenetics for Forest Trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Miryeganeh, M. Plants’ Epigenetic Mechanisms and Abiotic Stress. Genes 2021, 12, 1106. [Google Scholar] [CrossRef] [PubMed]
- AOAC, Official Methods of Analysis, 2000, Association of Official Analytical Chemists, Washington, DC, USA, 17th edition.
- Sparks, D.L. , Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E. Methods of Soil Analysis: Part 3 Chemical Methods; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; ISBN 978-0-89118-866-7. [Google Scholar]
Figure 1.
Patterns of substrate volumetric water content (VWC) in regular watered and phosphorus fertilization treatment (W/+P), regular watered and non-phosphorus fertilization treatment (W/-P), drought and phosphorus fertilization treatment (D/+P) and drought and non-phosphorus fertilization treatment (D/-P) with mean values of pre-down leaf water potential (Ψ) in regular watered (W) and drought treated (D) saplings of common beech (C. beech) and sessile oak (S. oak), measured on 26th May and 31st August.
Figure 1.
Patterns of substrate volumetric water content (VWC) in regular watered and phosphorus fertilization treatment (W/+P), regular watered and non-phosphorus fertilization treatment (W/-P), drought and phosphorus fertilization treatment (D/+P) and drought and non-phosphorus fertilization treatment (D/-P) with mean values of pre-down leaf water potential (Ψ) in regular watered (W) and drought treated (D) saplings of common beech (C. beech) and sessile oak (S. oak), measured on 26th May and 31st August.
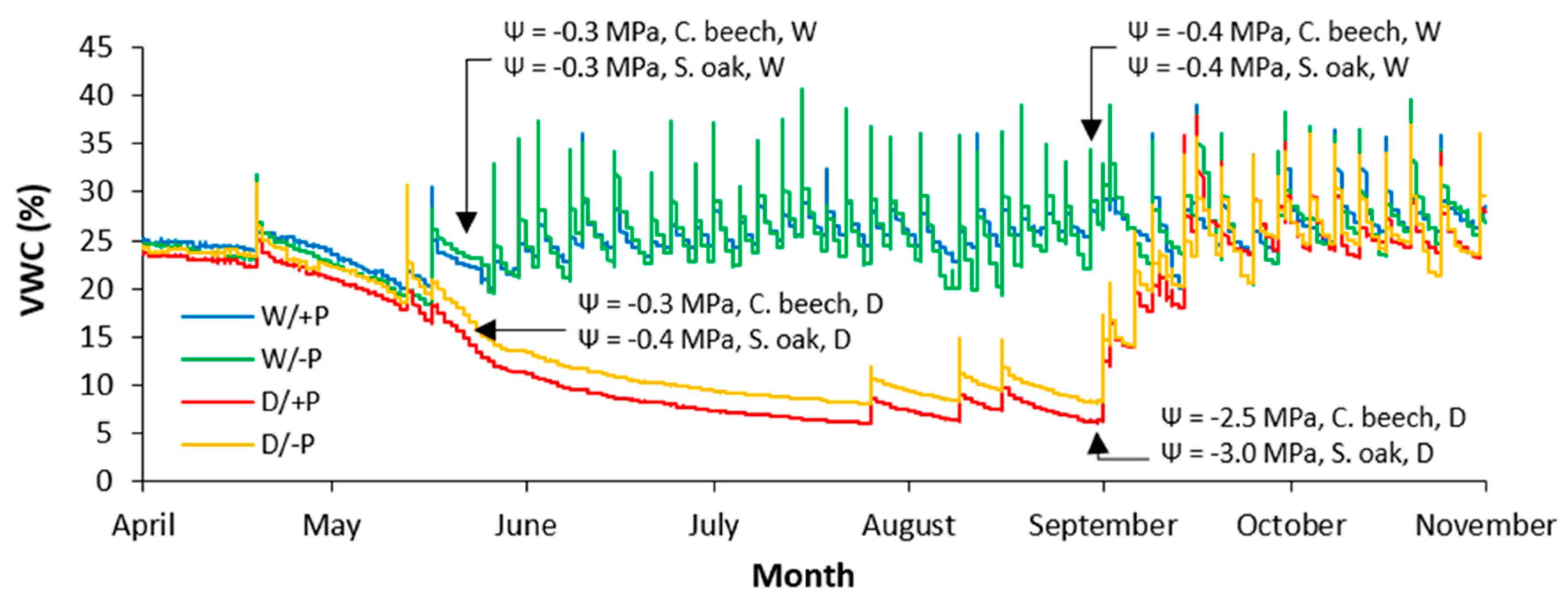
Figure 2.
Total phosphorus concentration in substrate (A) and leaves (means ± SD) of common beech (B) and sessile oak (C) saplings originated from Karlovac (KA) and Slavonski Brod (SB) provenances, fertlized with phosphorus (+P), non-fertilized with phosphorus (-P), regularly watered (W) and drought-treated (D). Red horizontal dashed lines indicate critical phosphorus concentration in substrate [78] and in leaves of common beech and sessile oak saplings [79].
Figure 2.
Total phosphorus concentration in substrate (A) and leaves (means ± SD) of common beech (B) and sessile oak (C) saplings originated from Karlovac (KA) and Slavonski Brod (SB) provenances, fertlized with phosphorus (+P), non-fertilized with phosphorus (-P), regularly watered (W) and drought-treated (D). Red horizontal dashed lines indicate critical phosphorus concentration in substrate [78] and in leaves of common beech and sessile oak saplings [79].
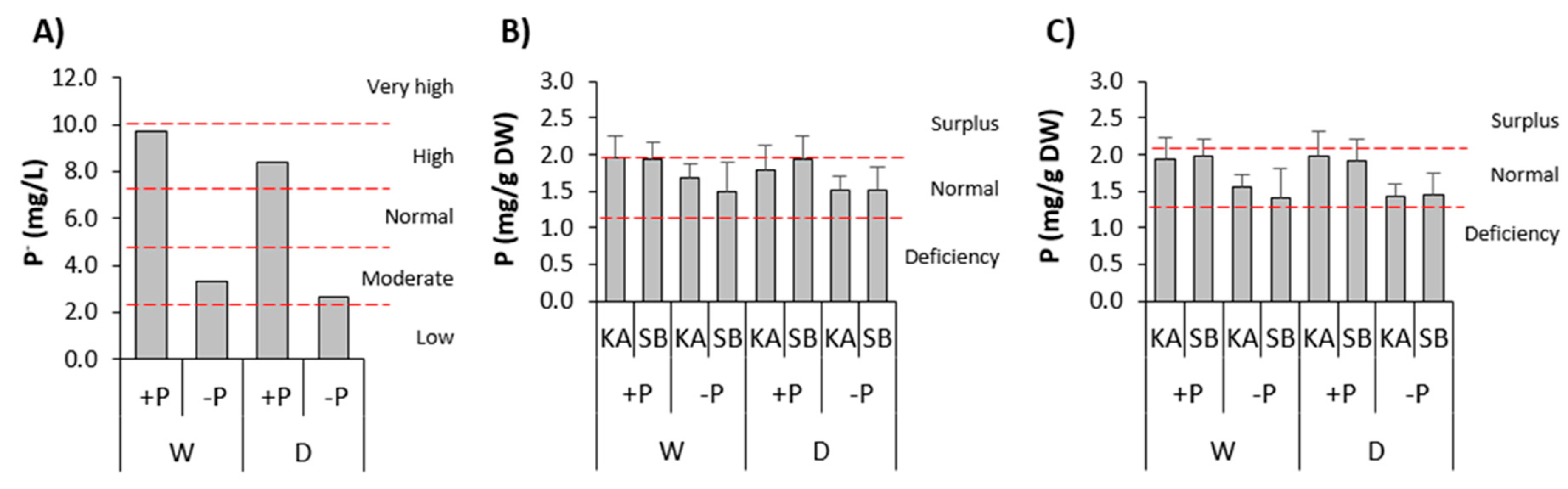
Figure 3.
Mean values ± SD of fine root dry mass (FRmass) in common beech saplings (A), as well as, mean ratios ± SD of fine root/stem dry mass (FRmass/STmass), fine root/coarse root dry mass (FRmass/CRmass) and fine root/total dry mass of leaves, stem and coarse root (FRmass/L+ST+CRmass) in common beech saplings (B), which was regularly waterd (W), drought-treated (D), fertilized with phospohorus (+P) and non-ferilized with phosphorous (-P). Different capital (for FRmass and FRmass/STmass), small (for FRmass/CRmass) and italic small (for FRmass/L+ST+CRmass) letters indicate significant differences among W/+P, W/-P, D/+P and D/-P saplings at p ˂ 0.05.
Figure 3.
Mean values ± SD of fine root dry mass (FRmass) in common beech saplings (A), as well as, mean ratios ± SD of fine root/stem dry mass (FRmass/STmass), fine root/coarse root dry mass (FRmass/CRmass) and fine root/total dry mass of leaves, stem and coarse root (FRmass/L+ST+CRmass) in common beech saplings (B), which was regularly waterd (W), drought-treated (D), fertilized with phospohorus (+P) and non-ferilized with phosphorous (-P). Different capital (for FRmass and FRmass/STmass), small (for FRmass/CRmass) and italic small (for FRmass/L+ST+CRmass) letters indicate significant differences among W/+P, W/-P, D/+P and D/-P saplings at p ˂ 0.05.
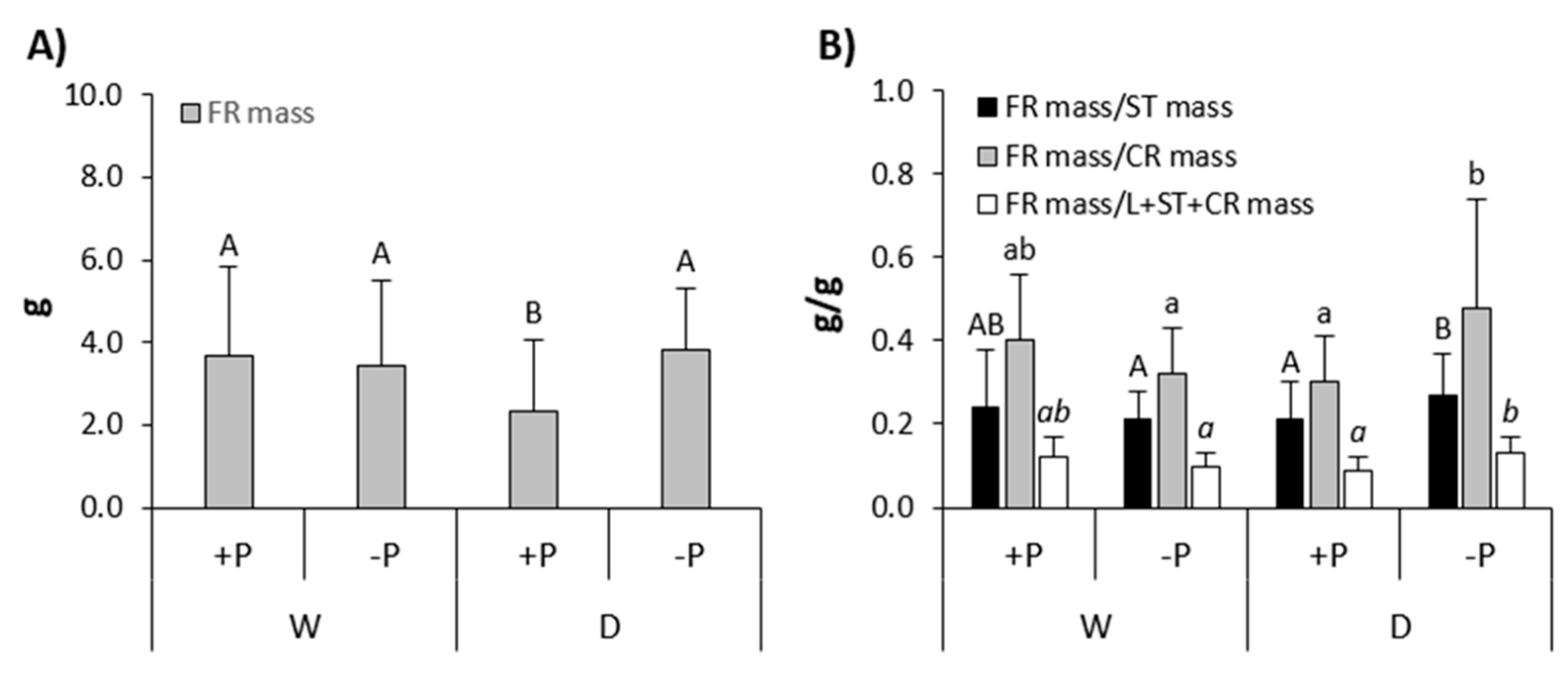
Figure 4.
Mean ratio ± SD of root colar diameter increment/stem height increment (Dincrement/Hincrement) of regularly watered (W) and drought-treated (D) sessile oak saplings originated from Karlovac (KA) and Slavonski Brod (SB) provenences. Different small letters indicate significant differences among W/KA, W/SB, D/KA and D/SB saplings at p ˂ 0.05.
Figure 4.
Mean ratio ± SD of root colar diameter increment/stem height increment (Dincrement/Hincrement) of regularly watered (W) and drought-treated (D) sessile oak saplings originated from Karlovac (KA) and Slavonski Brod (SB) provenences. Different small letters indicate significant differences among W/KA, W/SB, D/KA and D/SB saplings at p ˂ 0.05.
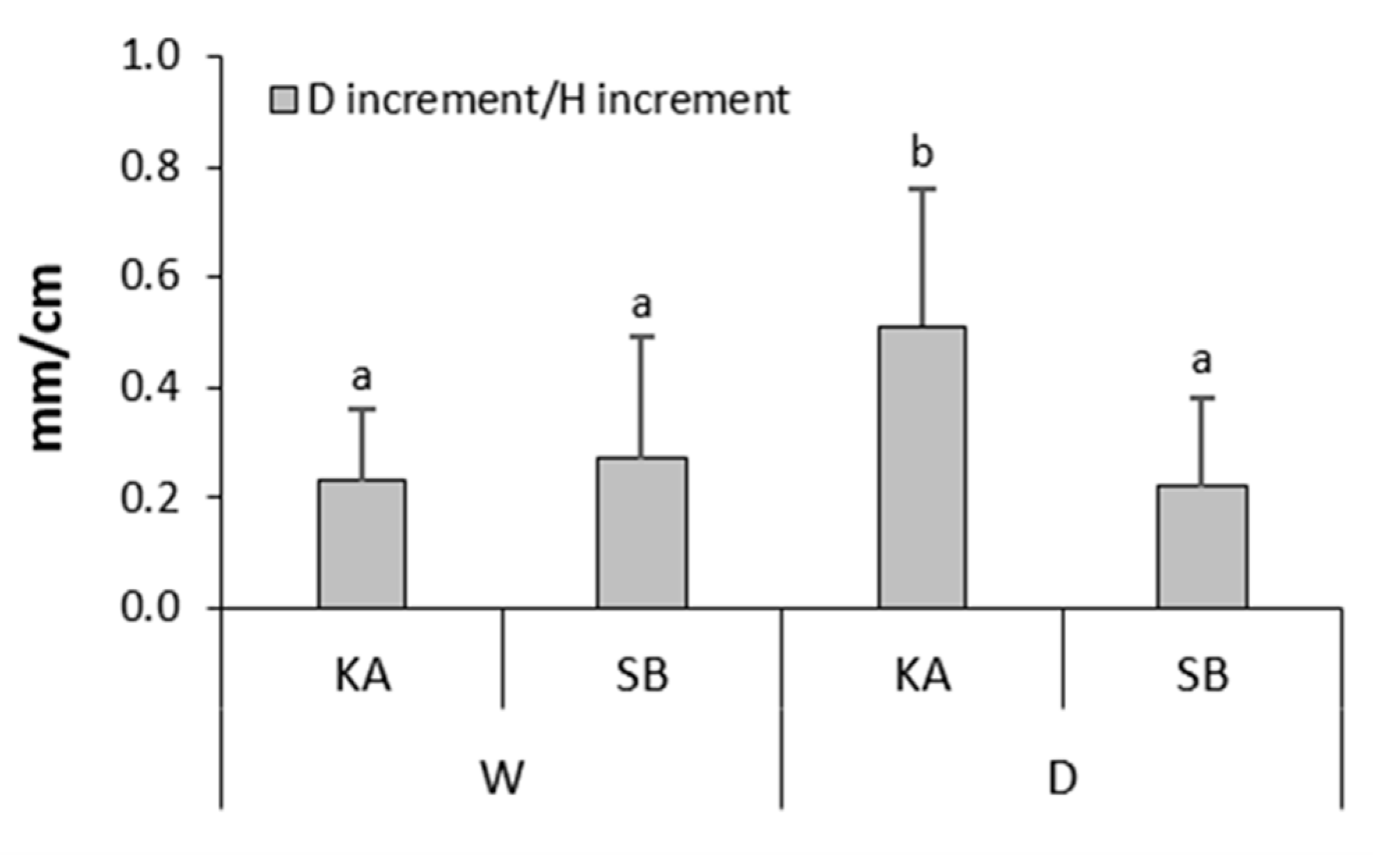
Table 1.
Main effects of drought (regularly watered vs. drought-treated saplings), fertilization (saplings fetilized with phosphorus vs. non-fertilized with phosphorus), provenance (saplings originated from Karlovac vs. Slavonski Brod provenance) and its interactions, on pre-down leaf water potential measured in May (ΨMay) and August (ΨAugust), as well as, on leaf phosphorus concentration (PLeaves) of common beech and sessile oak saplings, as calculated with factorial ANOVA.
Table 1.
Main effects of drought (regularly watered vs. drought-treated saplings), fertilization (saplings fetilized with phosphorus vs. non-fertilized with phosphorus), provenance (saplings originated from Karlovac vs. Slavonski Brod provenance) and its interactions, on pre-down leaf water potential measured in May (ΨMay) and August (ΨAugust), as well as, on leaf phosphorus concentration (PLeaves) of common beech and sessile oak saplings, as calculated with factorial ANOVA.
| Species | Parameter | Drought (D) | Fertilization (P) | Provenance (Pr) | D x P | D x Pr | P x Pr | D x P x Pr |
|---|---|---|---|---|---|---|---|---|
| Common beech | ΨMay | * | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| ΨAugust | *** | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | |
| PLeaves | n.s. | *** | n.s. | n.s. | n.s. | n.s. | n.s. | |
| Sessile oak |
ΨMay | * | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| ΨAugust | *** | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | |
| PLeaves | n.s. | *** | n.s. | n.s. | n.s. | n.s. | n.s. | |
| Levels of significance: *p < 0.05; ***p < 0.001; n.s., not significant. | ||||||||
Table 2.
Mean values ± SD of growth and allometric growth relationship parameters for regularly watered (W) and drought-treated (D) common beech and sessile oak saplings, with main effect of drought (D effect) as calculated with factorial ANOVA (W vs. D saplings). Red arrows (↓) indicate lower and green arrows (↑) indicate higher mean values of measured parameters in D compared to W saplings.
Table 2.
Mean values ± SD of growth and allometric growth relationship parameters for regularly watered (W) and drought-treated (D) common beech and sessile oak saplings, with main effect of drought (D effect) as calculated with factorial ANOVA (W vs. D saplings). Red arrows (↓) indicate lower and green arrows (↑) indicate higher mean values of measured parameters in D compared to W saplings.
| Parameters | Common beech | Sessile oak | |||||
|---|---|---|---|---|---|---|---|
| D effect | W | D | D effect | W | D | ||
| Growth | DMarch (mm) | n.s. | 7.78 ± 2.26 | 7.70 ± 1.91 ↓ | n.s. | 7.73 ± 2.37 | 7.34 ± 2.04 ↓ |
| Dincrement (mm) | n.s. | 1.97 ± 1.31 | 1.78 ± 1.13 ↓ | * | 5.03 ± 2.35 | 3.72 ± 2.49 ↓ | |
| DSeptember (mm) | n.s. | 9.74 ± 2.92 | 9.48 ± 2.51 ↓ | * | 12.75 ± 3.52 | 11.06 ± 3.12 ↓ | |
| HMarch (cm) | n.s. | 46.04 ± 14.88 | 46.20 ± 15.54 ↓ | n.s. | 48.83 ± 22.77 | 42.84 ± 17.71 ↓ | |
| Hincrement (cm) | ** | 14.47 ± 8.24 | 9.74 ± 5.66 ↓ | ** | 24.24 ± 10.42 | 17.42 ± 9.73 ↓ | |
| HSeptember (cm) | n.s. | 60.60 ± 16.67 | 55.94 ± 15.12 ↓ | ** | 73.07 ± 25.00 | 60.26 ± 19.78 ↓ | |
| Larea (m2) | n.s. | 0.25 ± 0.19 | 0.21 ± 0.18 ↓ | * | 0.35 ± 0.23 | 0.26 ± 0.16 ↓ | |
| TRlength (cm) | n.s. | 41.14 ± 12.63 | 38.29 ± 12.34 ↓ | n.s. | 63.15 ± 17.52 | 61.59 ± 15.38 ↓ | |
| Lmass (g) | n.s. | 7.44 ± 5.55 | 6.49 ± 4.80 ↓ | n.s. | 14.91 ± 10.15 | 12.30 ± 7.12 ↓ | |
| STmass (g) | n.s. | 19.33 ± 17.64 | 14.92 ± 10.36 ↓ | * | 24.90 ± 19.24 | 17.16 ± 11.31 ↓ | |
| AGmass (g) | n.s. | 26.77 ± 22.74 | 21.41 ± 14.83 ↓ | * | 39.82 ± 28.70 | 29.46 ± 18.01 ↓ | |
| FRmass (g) | n.s. | 3.55 ± 2.11 | 3.08 ± 1.77 ↓ | n.s. | 3.04 ± 1.91 | 2.82 ± 1.85 ↓ | |
| CRmass (g) | n.s. | 10.60 ± 6.86 | 8.42 ± 4.49 ↓ | n.s. | 40.28 ± 29.92 | 30.26 ± 19.77 ↓ | |
| BGmass (g) | n.s. | 14.15 ± 8.74 | 11.50 ± 5.87 ↓ | n.s. | 43.32 ± 31.30 | 33.09 ± 21.02 ↓ | |
| Allometric growth relationship | D/HMarch | n.s. | 0.17 ± 0.04 | 0.18 ± 0.04 ↑ | n.s. | 0.18 ± 0.07 | 0.19 ± 0.07 ↑ |
| Dincrement/Hincrement | n.s. | 0.20 ± 0.29 | 0.28 ± 0.33 ↑ | n.s. | 0.25 ± 0.19 | 0.37 ± 0.53 ↑ | |
| D/HSeptember | n.s. | 0.16 ± 0.04 | 0.17 ± 0.04 ↑ | n.s. | 0.19 ± 0.06 | 0.20 ± 0.08 ↑ | |
| TRlenght/HSeptember | n.s. | 0.70 ± 0.20 | 0.70 ± 0.22 | ** | 0.90 ± 0.21 | 1.10 ± 0.34 ↑ | |
| CRmass/STmass | n.s. | 0.65 ± 0.23 | 0.69 ± 0.36 ↑ | * | 1.64 ± 0.52 | 1.88 ± 0.60 ↑ | |
| FRmass/Lmass | n.s. | 0.55 ± 0.25 | 0.54 ± 0.22 ↓ | n.s. | 0.26 ± 0.18 | 0.26 ± 0.19 | |
| FRmass/STmass | n.s. | 0.23 ± 0.11 | 0.24 ± 0.10 ↑ | n.s. | 0.16 ± 0.11 | 0.19 ± 0.12 ↑ | |
| FRmass/CRmass | n.s. | 0.36 ± 0.14 | 0.39 ± 0.21 ↑ | n.s. | 0.11 ± 0.08 | 0.11 ± 0.06 | |
| FRmass/L+ST+CRmass | n.s. | 0.11 ± 0.04 | 0.11 ± 0.04 | n.s. | 0.05 ± 0.03 | 0.05 ± 0.03 | |
| BG/AG | n.s. | 0.61 ± 0.20 | 0.62 ± 0.20 ↑ | n.s. | 1.09 ± 0.34 | 1.17 ± 0.37 ↑ | |
| Levels of significance: *p < 0.05; **p < 0.01; n.s., not significant. Abbreviations: D – root collar diameter, Dincrement – root collar diameter increment, H – stem height, Hincrement – stem height increment, L – leaf, TR – tap root, ST – stem, AG – aboveground, FR – fine root, CR – coarse root, BG – beloweground, mass – dry biomass | |||||||
Table 3.
Mean values ± SD of growth and allometric growth relationship parameters for common beech and sessile oak saplings fertilized with phosphorus (+P) and non-fertilized with phosphorus (-P), with main effect of phosphorus fertilization (P effect) as calculated with factorial ANOVA (+P vs. -P saplings). Red arrows (↓) indicate lower and green arrows (↑) indicate higher mean values in +P compared to -P saplings.
Table 3.
Mean values ± SD of growth and allometric growth relationship parameters for common beech and sessile oak saplings fertilized with phosphorus (+P) and non-fertilized with phosphorus (-P), with main effect of phosphorus fertilization (P effect) as calculated with factorial ANOVA (+P vs. -P saplings). Red arrows (↓) indicate lower and green arrows (↑) indicate higher mean values in +P compared to -P saplings.
| Parameters | Common beech | Sessile oak | |||||
|---|---|---|---|---|---|---|---|
| P effect | +P | -P | P effect | +P | -P | ||
| Growth | DMarch (mm) | n.s. | 7.59 ± 2.18 ↓ | 7.89 ± 1.99 | n.s. | 7.15 ± 2.16 ↓ | 7.92 ± 2.21 |
| Dincrement (mm) | n.s. | 1.68 ± 1.20 ↓ | 2.06 ± 1.22 | n.s. | 4.23 ± 2.67 ↓ | 4.51 ± 2.33 | |
| DSeptember (mm) | n.s. | 9.27 ± 2.87 ↓ | 9.96 ± 2.53 | n.s. | 11.38 ± 3.18 ↓ | 12.43 ± 3.59 | |
| HMarch (cm) | n.s. | 45.65 ± 13.85 ↓ | 46.69 ± 16.45 | n.s. | 45.13 ± 21.28 ↓ | 46.54 ± 19.92 | |
| Hincrement (cm) | n.s. | 12.23 ± 6.65 ↑ | 11.97 ± 8.19 | n.s. | 20.51 ± 9.93 ↓ | 21.15 ± 11.32 | |
| HSeptember (cm) | n.s. | 57.89 ± 15.55 ↓ | 58.66 ± 16.60 | n.s. | 65.64 ± 23.08 ↓ | 67.70 ± 23.78 | |
| Larea (m2) | n.s. | 0.21 ± 0.19 ↓ | 0.25 ± 0.18 | n.s. | 0.27 ± 0.19 ↓ | 0.35 ± 0.21 | |
| TRlength (cm) | * | 36.72 ± 14.16 ↓ | 42.71 ± 9.85 | n.s. | 63.53 ± 16.43 ↑ | 61.21 ± 16.49 | |
| Lmass (g) | n.s. | 6.53 ± 5.96 ↓ | 7.40 ± 4.30 | n.s. | 11.92 ± 7.05 ↓ | 15.29 ± 10.08 | |
| STmass (g) | n.s. | 16.75 ± 16.14 ↓ | 17.49 ± 12.95 | n.s. | 18.52 ± 12.88 ↓ | 23.55 ± 18.71 | |
| AGmass (g) | n.s. | 23.29 ± 21.63 ↓ | 24.89 ± 16.81 | n.s. | 30.43 ± 19.52 ↓ | 38.84 ± 28.02 | |
| FRmass (g) | n.s. | 2.99 ± 2.08 ↓ | 3.64 ± 1.77 | * | 2.51 ± 1.47 ↓ | 3.35 ± 2.14 | |
| CRmass (g) | n.s. | 9.10 ± 6.80 ↓ | 9.92 ± 4.80 | n.s. | 31.60 ± 21.51 ↓ | 38.95 ± 29.10 | |
| BGmass (g) | n.s. | 12.09 ± 8.62 ↓ | 13.56 ± 6.24 | n.s. | 34.11 ± 22.28 ↓ | 42.29 ± 30.73 | |
| Allometric growth relationship | D/HMarch | n.s. | 0.17 ± 0.03 ↓ | 0.18 ± 0.05 | n.s. | 0.19 ± 0.08 | 0.19 ± 0.05 |
| Dincrement/Hincrement | n.s. | 0.20 ± 0.22 ↓ | 0.29 ± 0.38 | n.s. | 0.27 ± 0.28 ↓ | 0.35 ± 0.50 | |
| D/HSeptember | n.s. | 0.16 ± 0.04 ↓ | 0.17 ± 0.04 | n.s. | 0.19 ± 0.08 ↓ | 0.20 ± 0.07 | |
| TRlenght/HSeptember | ** | 0.64 ± 0.20 ↓ | 0.76 ± 0.20 | n.s. | 1.03 ± 0.32 ↑ | 0.97 ± 0.28 | |
| CRmass/STmass | n.s. | 0.68 ± 0.35 ↑ | 0.66 ± 0.25 | n.s. | 1.79 ± 0.57 ↑ | 1.74 ± 0.57 | |
| FRmass/Lmass | n.s. | 0.56 ± 0.27 ↑ | 0.54 ± 0.19 | n.s. | 0.27 ± 0.22 ↑ | 0.25 ± 0.15 | |
| FRmass/STmass | n.s. | 0.23 ± 0.12 ↓ | 0.24 ± 0.09 | n.s. | 0.18 ± 0.13 | 0.18 ± 0.09 | |
| FRmass/CRmass | n.s. | 0.35 ± 0.14 ↓ | 0.40 ± 0.21 | n.s. | 0.11 ± 0.08 | 0.11 ± 0.06 | |
| FRmass/L+ST+CRmass | n.s. | 0.11 ± 0.04 | 0.11 ± 0.04 | n.s. | 0.05 ± 0.04 | 0.05 ± 0.03 | |
| BG/AG | n.s. | 0.61 ± 0.20 ↓ | 0.62 ± 0.20 | n.s. | 1.16 ± 0.36 ↑ | 1.10 ± 0.35 | |
| Levels of significance: *p < 0.05; **p < 0.01; n.s., not significant. Abbreviations: D – root collar diameter, Dincrement – root collar diameter increment, H – stem height, Hincrement – stem height increment, L – leaf, TR – tap root, ST – stem, AG – aboveground, FR – fine root, CR – coarse root, BG – beloweground, mass – dry biomass | |||||||
Table 4.
Mean values ± SD of growth and allometric growth relationship parameters for common beech and sessile oak saplings originated from Karlovac (KA) and Slavonski Brod (SB) provenances with main effect of provenance (Pr effect) as calculated with factorial ANOVA (saplings from KA vs. SB provenance). Red arrows (↓) indicate lower and green arrows (↑) indicate higher mean values in saplings originated from SB compared to KA provenance.
Table 4.
Mean values ± SD of growth and allometric growth relationship parameters for common beech and sessile oak saplings originated from Karlovac (KA) and Slavonski Brod (SB) provenances with main effect of provenance (Pr effect) as calculated with factorial ANOVA (saplings from KA vs. SB provenance). Red arrows (↓) indicate lower and green arrows (↑) indicate higher mean values in saplings originated from SB compared to KA provenance.
| Parameters | Common beech | Sessile oak | |||||
|---|---|---|---|---|---|---|---|
| Pr effect | KA | SB | Pr effect | KA | SB | ||
| Growth | DMarch (mm) | n.s. | 7.96 ± 2.11 | 7.52 ± 2.05 ↓ | n.s. | 7.65 ± 2.39 | 7.41 ± 2.03 ↓ |
| Dincrement (mm) | * | 2.14 ± 1.21 | 1.60 ± 1.17 ↓ | n.s. | 4.26 ± 2.35 | 4.49 ± 2.66 ↑ | |
| DSeptember (mm) | n.s. | 10.10 ± 2.57 | 9.12 ± 2.79 ↓ | n.s. | 11.91 ± 3.19 | 11.90 ± 3.66 ↓ | |
| HMarch (cm) | n.s. | 44.55 ± 16.70 | 47.79 ± 13.37 ↑ | n.s. | 45.84 ± 19.40 | 45.83 ± 21.78 ↓ | |
| Hincrement (cm) | n.s. | 12.88 ± 7.71 | 11.33 ± 7.12 ↓ | n.s. | 19.21 ± 10.35 | 22.46 ± 10.70 ↑ | |
| HSeptember (cm) | n.s. | 57.43 ± 15.49 | 59.11 ± 16.62 ↑ | n.s. | 65.05 ± 22.41 | 68.28 ± 24.34 ↑ | |
| Larea (m2) | n.s. | 0.24 ± 0.17 | 0.22 ± 0.20 ↓ | n.s. | 0.29 ± 0.18 | 0.32 ± 0.22 ↑ | |
| TRlength (cm) | n.s. | 39.79 ± 12.24 | 39.64 ± 12.89 ↓ | n.s. | 60.23 ± 18.07 | 64.51 ± 14.44 ↑ | |
| Lmass (g) | n.s. | 7.30 ± 4.72 | 6.64 ± 5.64 ↓ | n.s. | 13.29 ± 7.57 | 13.92 ± 9.98 ↑ | |
| STmass (g) | n.s. | 18.06 ± 14.82 | 16.18 ± 14.39 ↓ | n.s. | 19.99 ± 13.90 | 22.08 ± 18.26 ↑ | |
| AGmass (g) | n.s. | 25.36 ± 19.12 | 22.81 ± 19.57 ↓ | n.s. | 33.28 ± 20.92 | 36.00 ± 27.59 ↑ | |
| FRmass (g) | n.s. | 3.56 ± 1.79 | 3.06 ± 2.08 ↓ | * | 3.41 ± 2.04 | 2.45 ± 1.56 ↓ | |
| CRmass (g) | n.s. | 10.21 ± 6.15 | 8.81 ± 5.56 ↓ | n.s. | 35.39 ± 25.51 | 35.16 ± 26.21 ↓ | |
| BGmass (g) | n.s. | 13.77 ± 7.73 | 11.88 ± 7.26 ↓ | n.s. | 38.80 ± 26.98 | 37.60 ± 27.32 ↓ | |
| Allometric growth relationship | D/HMarch | * | 0.19 ± 0.04 | 0.16 ± 0.04 ↓ | n.s. | 0.19 ± 0.07 | 0.18 ± 0.07 ↓ |
| Dincrement/Hincrement | n.s. | 0.29 ± 0.38 | 0.20 ± 0.21 ↓ | n.s. | 0.37 ± 0.53 | 0.25 ± 0.21 ↓ | |
| D/HSeptember | * | 0.18 ± 0.04 | 0.16 ± 0.04 ↓ | n.s. | 0.20 ± 0.07 | 0.19 ± 0.07 ↓ | |
| TRlenght/HSeptember | n.s. | 0.71 ± 0.22 | 0.69 ± 0.19 ↓ | n.s. | 0.98 ± 0.33 | 1.01 ± 0.26 ↑ | |
| CRmass/STmass | n.s. | 0.68 ± 0.34 | 0.66 ± 0.27 ↓ | n.s. | 1.82 ± 0.61 | 1.70 ± 0.52 ↓ | |
| FRmass/Lmass | n.s. | 0.54 ± 0.19 | 0.56 ± 0.27 ↑ | ** | 0.31 ± 0.21 | 0.21 ± 0.13 ↓ | |
| FRmass/STmass | n.s. | 0.23 ± 0.09 | 0.23 ± 0.11 | *** | 0.22 ± 0.13 | 0.14 ± 0.08 ↓ | |
| FRmass/CRmass | n.s. | 0.37 ± 0.14 | 0.38 ± 0.21 ↑ | ** | 0.13 ± 0.08 | 0.09 ± 0.05 ↓ | |
| FRmass/L+ST+CRmass | n.s. | 0.11 ± 0.04 | 0.11 ± 0.05 | ** | 0.06 ± 0.04 | 0.04 ± 0.02 ↓ | |
| BG/AG | n.s. | 0.61 ± 0.17 | 0.62 ± 0.22 ↑ | n.s. | 1.17 ± 0.38 | 1.09 ± 0.33 ↓ | |
| Levels of significance: *p < 0.05; **p < 0.01; n.s., not significant. Abbreviations: D – root collar diameter, Dincrement – root collar diameter increment, H – stem height, Hincrement – stem height increment, L – leaf, TR – tap root, ST – stem, AG – aboveground, FR – fine root, CR – coarse root, BG – beloweground, mass – dry biomass | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



