
Submitted:
27 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
Superficial discolored spots on Atlantic salmon (Salmo salar) fillets are a serious quality problem for commercial seafood farming. Previous reports proposed that the black spots (called melanized focal changes (MFC)) may be melanin, but no convincing evidence has been reported. In this study, we performed chemical characterization of MFC, and also of red pigment (called red focal changes (RFC)), of salmon fillets using alkaline hydrogen peroxide oxidation and hydroiodic acid hydrolysis. This revealed that the MFC contain 3,4-dihydroxyphenylalanine (DOPA)-derived eumelanin whereas the RFC only contain trace amounts of eumelanin. Therefore, probably, the black color of MFC can be explained by the presence of eumelanin from accumulated melanomacrophages. For the red pigment, we could not find a significant signature of either eumelanin or pheomelanin, and the red color is probably predominantly hemorrhagic in nature. However, we found that in RFC the level of pigmentation did increase together with some melanogenic metabolites. Comparison with a “mimicking experiment,” in which a mixture of a salmon homogenate + DOPA was oxidized with tyrosinase, suggested that the RFC include conjugations of DOPAquinone and/or DOPAchrome with salmon muscle tissue proteins. In short, the results suggest that melanogenic metabolites in MFC and RFC derive from different chemical pathways, which would agree with the two different colorations deriving from distinct cellular origins, namely melanomacrophages and red blood cells, respectively.
Keywords:
Atlantic salmon
; Melanized focal changes
; Red focal changes
; Eumelanin
; Pheomelanin
; PTCA
; 4-AHP
; AHPO
; Melanomacrophages
; Red blood cells
1. Introduction
Atlantic salmon (Salmo salar) is a major commercial species in the salmonid family, produced in large numbers in Norway, Chile, the UK, and Canada. Norway’s long shoreline and cold, clean seawater provide excellent conditions for its aquaculture. However, superficial hyperpigmented areas on salmon fillets, called “black spots (BS),” represent a major problem for the commercial aquaculture of Atlantic salmon. BS in white skeletal muscle of farmed Atlantic salmon can appear initially as red spots (RS) characterized by hemorrhages and acute inflammation and progress into BS characterized by chronic inflammation and accumulating pigmented immune cells called “melanomacrophages” [1]. RS and BS are also known as “red focal changes (RFC)” and “melanized focal changes (MFC)” in the fillet, respectively [2] (Figure 1).
These discolorations cause large economic losses since fillets with pigmented abnormalities are commercially downgraded because of reduced attractiveness. A study found that in the production period of 59 weeks after the transfer of juvenile salmon (smolts) to sea, MFC first appeared with low frequency but then became more frequent, whereas the RFC maintained a constant low prevalence of about 4% [3]. Over time, the prevalence of MFC on fillets of Norwegian sea-reared salmon has increased substantially, from 7% in 2003 to 20% in 2015 [4]. Hence, the seafood farming industry is paying great attention to this problem and is seeking to understand the underlying causes of the dark discolorations on salmon fillets.
MFC can occur mainly in three locations: internal organs, the peritoneum, and the muscle fillet. Most MFC are up to 3 cm in width and a few mm thick, and are located in the front part of the fillet (Figure 1 and see below). Interestingly, although MFC are common in farmed harvest-sized Atlantic salmon all along the Norwegian coast, there have not been reports that such spots are very common in wild fish [1].
Melanomacrophage centers in teleost fish are distinctive groupings of phagocytic, melanin pigment-containing, immune cells that are generally found inside the stroma of tissues with immune functions, but these centers can also develop in association with inflammation elsewhere in the fish body [6,7]. Melanomacrophages are particularly concentrated in teleost fish spleen, kidney, and liver, and can also be found in spleen and liver of amphibians and reptiles [8-15]. In salmon, in some cases of granulomatous inflammation of the muscle, areas with pigmented melanomacrophages penetrate the peritoneum from the muscles, leading to externally visible pigmentation [5]. In case of Piscine orthoreovirus 1 (PRV-1) infection, the appearance of melanomacrophages was coincident with—and provided a likely explanation for—the transformation from the late phase of RFC into MFC [1,2,4,16]. However, melanomacrophage accumulations have also been found in cases of chronic inflammation initiated by trauma without specific causative agents [17]. It has been proposed that melanomacrophages may contain several pigments, such as melanin, lipofuscin, and hemosiderin [7,18], and the presence of melanin pigment in salmon melanomacrophases was suggested through Fontana Masson staining [19]. Gallone et al. [20] described that the melanin produced in the liver-pigmented macrophages of the frog Rana esculenta L. was a 5,6-dihydroxyindole (DHI)-rich eumelanin (EM) similar to Sepia melanin. However, to the best of our knowledge, there have not been reports regarding a biochemical and chemical analysis of melanin in melanomacrophages in fish. Gene expression studies, including the finding of tyrosinase expression in a cell line believed to represent melanomacrophages, support that melanin synthesis is the source of the dark pigment in salmon melanomacrophages and MFC [5,17,21,22]. However, although MFC in fillets have been associated with accumulations of melanomacrophages, they have not yet been examined by detailed chemical methods as to whether the focal changes do involve melanin and the nature of that melanin.
Melanin pigments, composed of black to dark brown EM and yellow to reddish-brown pheomelanin (PM), are widely distributed in vertebrates [23]. While much is now understood about the nature of melanin synthesized in higher vertebrates, little is known about melanin produced by aquatic animals including aquacultured fishes. Red seabream (Pagrus major), which in Japan is one of the commercially most valuable fish species, is usually found at around 20-m depth of seawater, and its skin is bright scarlet. However, their skin turns dark when bred in net cages–fish farmers call these seabreams “suntanned”–and this darkening severely reduces their market value. Adachi et al. [24] demonstrated that the cause of this suntanning is the production of melanin, and they were the first to chemically quantify melanin in fish. They reported that the skin of suntanned seabream accumulated five times higher levels of EM than the corresponding parts of shaded and wild red seabream, while PM was below detection limits in these fish [24].
Both EM and PM are derived from a common precursor, 3,4-dihydroxyphenylalanine-quinone (DOPAquinone), that is produced from tyrosine by the action of the melanogenic enzyme tyrosinase [25-28]. In the absence of thiol compounds, DOPAquinone undergoes an intramolecular cyclization of its amino group [29] to produce DOPAchrome, which is then spontaneously and gradually converted to DHI or 5,6-dihydroxyindole-2-carboxylic acid (DHICA) by tyrosinase-related protein 2 [30-32] or by copper ions [33]. DHI and DHICA are then further oxidized and polymerized to produce EM. Sulfhydryl compounds such as cysteine, if present at sufficient levels (> 0.13 µM), may enter a route different from the normal EM pathways to give thiol adducts of DOPA, that is, 5-S-cysteinyldopa (5SCD) along with a minor isomer 2-S-cysteinyldopa [26]. Further oxidation of these thiol adducts leads to the formation of benzothiazine and benzothiazole intermediates, which then are converted to PM. In vertebrates, melanin synthesis, called “mixed melanogenesis” because it produces both EM and PM, is biochemically controlled by tyrosinase activity and cysteine concentration. The casing model of mixed melanogenesis implies that PM is always produced first, after which EM is deposited on the preformed PM [27]. To characterize melanins and melanogenesis, in the early 1980s, we developed a microanalytical method to analyze EM and PM [34] based on the chemical degradation of melanin pigments followed by analysis of the degradation products using high-performance liquid chromatography (HPLC). Later, we established a novel, more convenient method for the simultaneous measurement of EM and PM using alkaline hydrogen peroxide oxidation (AHPO) and hydroiodic acid (HI) hydrolysis [28,35,36] (Figure 2).
The AHPO generates the specific markers pyrrole-2,3,5-tricarboxylic acid (PTCA), pyrrole-2,3-dicarboxylic acid (PDCA), thiazole-2,4,5-tricarboxylic acid (TTCA), and thiazole-4,5-dicarboxylic acid (TDCA). PTCA is a specific biomarker of DHICA units or 2-substituted DHI units in EM, whereas PDCA is a specific biomarker for DHI-derived units in EM, while TTCA and TDCA are specific biomarkers for benzothiazole-derived moieties in PM. Analysis of benzothiazine-derived moieties in PM is performed using HI hydrolysis to yield 4-amino-3-hydroxyphenylalanine (4-AHP) and its isomer 3-amino-4-hydroxyphenylalanine (3-AHP) [28,34,35].
As mentioned above, previous reports have suggested that the black pigments of MFC in salmon fillets may be melanin [2,6,19], but this assumption has been lacking chemical analysis other than histological staining [19], which, amongst others, cannot distinguish between eumelanin and pheomelanin. Therefore, for the present study, we performed the chemical analysis of MFC and RFC in salmon fillets using the chemical degradation methods for melanin pigments to help understand the character of MFC and RFC. The results revealed that the MFC are derived from EM while RFC contain little or no EM or PM. The results of analyzing RFC suggested that they include melanogenic metabolites derived from oxidized proteins produced by DOPAquinone and/or DOPAchrome binding to salmon proteins. We could find support for this hypothesis by performing tyrosinase oxidation of DOPA in the presence of salmon fillet proteins.
In short, the present study provides novel insights into the biochemistry and origin of MFC and RFC, which form a serious quality problem in the production of Atlantic salmon.
2. Results
We used the AHPO and HI hydrolysis assays that we previously developed to investigate whether the pigments in MFC and RFC in salmon are produced through the pathways of melanogenesis, which begins with tyrosinase oxidation of tyrosine, and determined the melanogenic components of these pigments.
2.1. Chemical characterization by AHPO and HI hydrolysis of MFC and RFC on salmon fillets
Salmon fillet samples prepared as described for Materials #1 and #2 from The Norwegian Institute of Food, Fisheries and Aquaculture Research (Nofima), Norway, were received in March, 2020, and analyzed. The pigmented areas had various degrees of pigmentation and were divided into three groups (small, medium, and large) depending on the size of the pigmented area: controls, MFC (BS-small, BS-medium, BS-large, and RFC (RS-small, RS-medium and RS-large) (Figure 3). Materials #1 are fillets from salmon cultivated in open cages in seawater (The LetSea Research Station, Dønna, Norway), and three pooled samples of each type of spot were analyzed. Materials #2 are fillets from salmon raised in large commercial cages hanging in sea (Lerøy Midt AS, Gjemnes, Norway), with various degrees of pigmentation (2 controls, 3 BS-small, 17 BS-medium, 7 BS-large, 1 RS-small and 2 RS-medium).
HPLC chromatograms of the AHPO mixtures obtained for BS and RS in Materials #1 are shown in Figure 4a and 4b, respectively. The PTCA (a specific biomarker for DHICA-derived units in EM) values observed for MFC were much higher than observed for RFC, where they were barely detectable. PDCA (a specific biomarker for DHI-derived units in EM) was only detected at trace levels in both MFC and RFC, while TTCA and TDCA, markers of benzothiazole-derived moieties in PM, were not detected. These AHPO analysis results conclude that MFC contain DHICA-rich EM and that RFC contain only a trace amount of EM. HPLC chromatograms of HI hydrolysates of both MFC and RFC in Materials #1 show the PM markers 4- and 3-AHP which are markers of benzothiazine-derived moieties in PM, and DOPA (Figure 4c, 4d). The 4-AHP value was larger in RFC than in MFC. Although 4-AHP was detected in RFC, the PM content in RFC was much smaller than in typical pheomelanic samples [28].
The VIS absorption spectrum of MFC in Soluene-350 of Materials #2 was similar in pattern to that of the control solution but with increased absorbance throughout the VIS region (Figure 5a). On the other hand, the spectrum of RFC in Soluene-350 had a large and increasing absorption below 550 nm with an absorbance 5-fold greater than the control at 400 nm. The difference spectra of MFC and RFC solutions subtracted from the control spectrum are shown in Figure 5b. These difference spectra show a small and flat absorption between 400 and 650 nm in MFC and a large and increasing absorption below 550 nm in RFC.
The results of spectrophotometric and chemical degradation analyses of MFC and RFC in Material #1 fillets are summarized in Figure 6. A500 values (total melanin values) obtained by spectrophotometry, after dissolving the samples in Soluene-350, progressively increased from the control (0.0065) to BS-small (0.0073), BS-medium (0.0097), and BS-large (0.0147). The A650/A500 ratio is an indicator of whether melanin is eumelanic or pheomelanic [37]. The A650/A500 ratio of 0.22 in BS-large suggests that this melanin is eumelanic, given that there is a background absorption from the control. The increasing trend was more obvious for PTCA values with 0.7 in the control, 3.4 in BS-small, 7.0 in BS-medium, and 19.4 ng/mg in BS-large (Supplemental Table 1). This clearly supports that the black color of MFC is attributable to DOPA-derived EM. PDCA values were only at trace levels. The low PDCA/PTCA ratio in the pigment of MFC indicates that this pigment is derived from DOPA but not from DA, because DA melanin should give comparable yields of PDCA and PTCA [38]. 4-AHP and 3-AHP were detected only at trace levels in MFC, irrespective of size, indicating that MFC contain little or no PM.
The identification of PTCA in MFC was confirmed by isolating some PTCA in a preparative scale AHPO experiment. The VIS and MS spectra are shown in Supplemental Figure 1, confirming the identification of PTCA.
RFC afforded a quite different pattern of degradation products compared with MFC (Figure 6). A500 values progressively increased from the control (0.0065), to RS-small (0.0148), RS-medium (0.0196), and RS-large (0.0208). However, the A650/A500 ratio remained similar and low (0.05 to 0.07), suggesting that RFC contain PM or another, not yet identified melanic pigment. PTCA values were only trace (<2.0 ng/mg), irrespective of size, indicating that this pigment is not a typical EM. 4-AHP values in RFC were many-fold higher than in MFC. Nevertheless, the low absolute values of 4-AHP (0.5 – 0.7 ng/mg) in RFC indicate that PM does not constitute a major portion of RFC. To characterize the pigment of MFC, we also analyzed DOPA after HI hydrolysis (named HI-DOPA). HI-DOPA can be produced from free DOPA, from free cysteinyldopa (CD) isomers, or from protein-bound DOPA and CD isomers [39,40]. HI-DOPA values (2.4 – 4.7 ng/mg) in MFC did not differ from the control (4.7 ng/mg). In contrast, HI-DOPA values progressively increased in RFC from the control (4.7), to RS-small (10.3), RS-medium (15.3), and RS-large (17.0 ng/mg). A similar trend was found for protein-bound (PB) 5-S-cysteinyldopa (5SCD), a metabolite of DOPA, that arises from the conjugation of DOPAquinone to sulfhydryl groups of proteins [39,40] (Figure 6). While PB-5SCD values in MFC remained the same (2.1 – 2.6 ng/mg) as the control (2.3 ng/mg), those in RFC were 2-fold higher (4.3 – 5.2 ng/mg) than the control. The above results indicate that the degree of pigmentation of RFC correlates well with the amount of melanin metabolites (HI-DOPA, PB-5SCD).
Analyses of Materials #1 led to the conclusion that the pigment of MFC is eumelanic while the pigment of RFC is neither eumelanic nor pheomelanic. We, therefore, wished to confirm these results using a greater number of MFC (a total of 27) from another set of materials (Materials #2) obtained from salmon raised in large cages in the sea.
Figure 7 summarizes the results of those analyses. The results show that pigments in MFC (n = 27) from Materials #2 had a similar pattern of melanin markers, e.g., A500, PTCA, 4-AHP, and HI-DOPA, as those in Materials #1. Only the high value of HI-DOPA in the control is puzzling (Figure 7, Supplemental Table 2). Taking advantage of analyzing a large number of MFC, we could assess the correlation between A500 and PTCA values. As shown in Supplemental Figure 2, the PTCA values correlated well (R2 = 0.754) with the A500 values with a background value of 0.006/mg, indicating that the black color is mostly due to DOPA-derived EM. RFC in Material #2 were only available from three fillets, but the melanin marker values in these RFC were similar to those in Materials #1.
Regarding the chemical characterization of RFC, it is noteworthy that the average 4-AHP/3-AHP ratio in three RFC (Materials #1) was 0.77 (Supplemental Table 1), which suggests that a melanic pigment was produced by interactions between DOPAquinone and NH2 groups in salmon proteins (Figure 8). This interaction would lead to the formation of a Schiff’s base between one of the two carbonyl groups in DOPAquinone and the protein amino group [41]. Reductive acid hydrolysis of such proteins with HI would give rise to the production of nearly equal amounts of 4-AHP and 3-AHP, as 4-AHP/3-AHP ratio should be much greater than 1 in PM [35]. Thus, we hypothesized that the pigment of RFC includes a form of oxidized proteins produced by DOPAquinone and/or DOPAchrome binding to proteins in salmon fillets. To confirm this hypothesis, we then performed an experiment mimicking the production of RFC in salmon fillets.
2.2. RFC are suggested to include melanogenic metabolites derived from oxidized proteins produced by DOPAquinone and/or DOPAchrome binding to salmon proteins
It is known that during a 4 h oxidation of DOPA by tyrosinase, all DOPA is oxidized to DOPAquinone, which is rapidly converted to the red pigment DOPAchrome that gives rise to EM via DHI and DHICA [25,26]. A minor amount of DOPAquinone may undergo conjugation with sulfhydryl or amino groups in proteins [41].
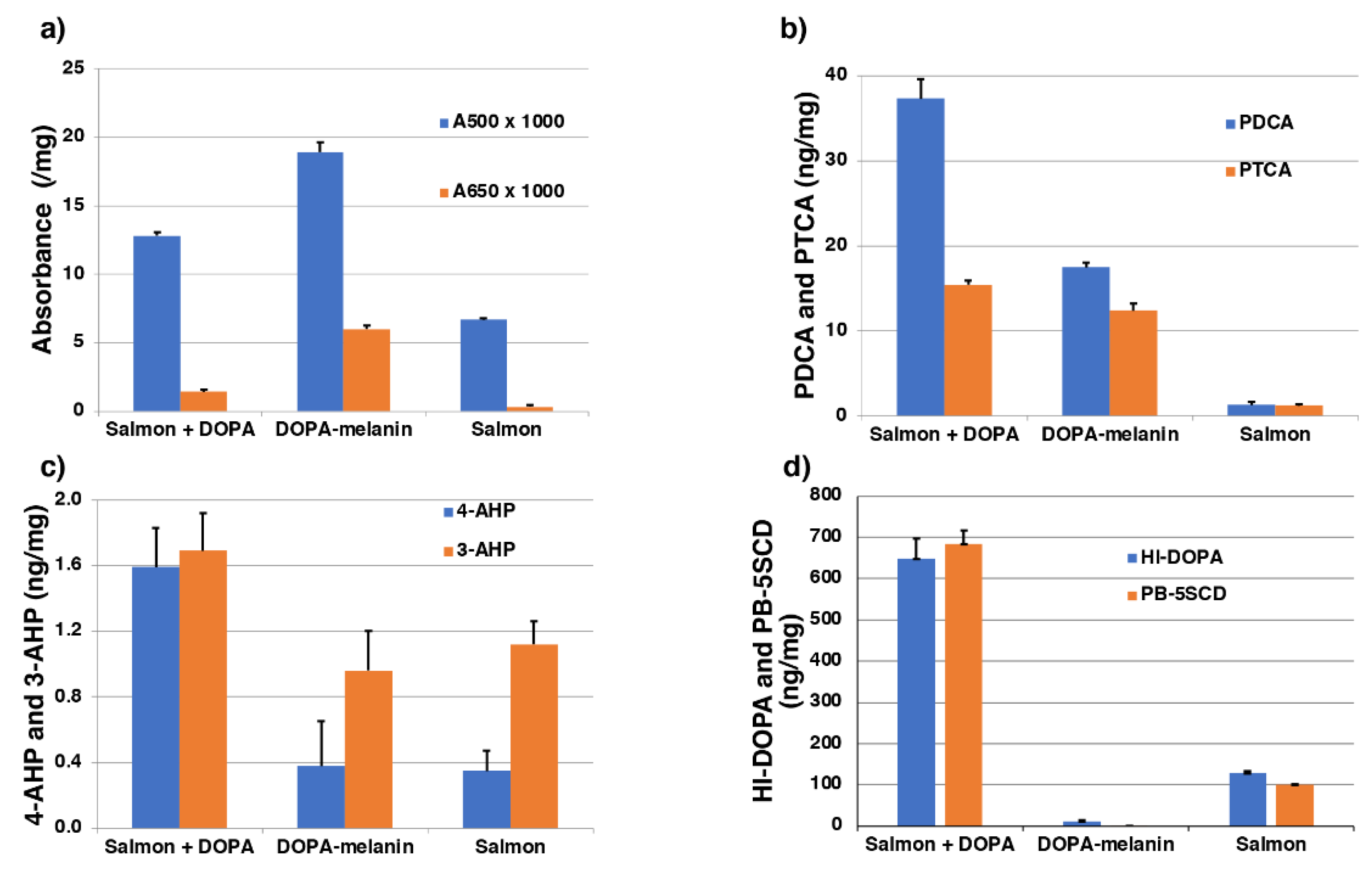
To replicate this, mushroom tyrosinase was added to a mixture of a salmon fillet homogenate (20 mg/mL) + DOPA (1 mM), DOPA (1 mM) alone, and salmon fillet homogenate (20 mg/mL) alone, and the reaction mixtures were incubated at pH 7.4 and 37°C for 4 h followed by measuring the melanin markers. The results of this mimicking experiment are summarized in Figure 9. The A500 value of 0.0128/mg after tyrosinase oxidation of the salmon + DOPA group was intermediate between the DOPA alone (DOPA-melanin) group (0.0189/mg) and the salmon alone group (0.0067/mg) (Figure 9a). The A650/A500 ratio in the salmon + DOPA group (0.110) was closer to the salmon alone group (0.050) than the DOPA alone group (0.317). These results indicate that the oxidation of DOPA to DOPA-melanin was suppressed in the salmon + DOPA group. This can be explained by interactions between salmon proteins and DOPA oxidation products. The PDCA value (37.3 ng/mg) in the salmon + DOPA group was higher than that of DOPA alone (17.5 ng/mg), suggesting that formation of DHI-protein conjugates derived from the conjugation of DOPAquinone or DOPAchrome and salmon proteins (Figure 9b). This suggests the presence of DHI-protein conjugates or related adducts in the salmon + DOPA group (Figure 8).
The 4-AHP value (1.6 ng/mg) in the salmon + DOPA group was very low, indicating that the contribution of PM can be ignored (Figure 9c). The 3-AHP value (1.7 ng/mg) in the salmon + DOPA group was also low, and the 4-AHP/3-AHP ratio of 0.9 had the same tendency as that of RFC in salmon (Figure 6). The salmon + DOPA group gave a higher yield of 4-AHP in comparison to the DOPA alone and the salmon alone groups. This suggests that 4-AHP is derived from interactions between DOPAquinone and the sulfhydryl and/or amino group in salmon proteins, since the tyrosinase oxidation of DOPA (no cysteine available) yielded no level of 4-AHP [26,41]. The DOPA value (648 ng/mg, 33% yield) following HI hydrolysis (HI-DOPA) in the salmon + DOPA group was much higher than in the DOPA alone group (12 ng/mg) or the salmon alone group (128 ng/mg) (Figure 9d). Similarly, the PB-5SCD value (685 ng/mg, 22% yield) in the salmon + DOPA group was much higher than in the DOPA alone group (0.5 ng/mg) or the salmon alone group (100 ng/mg) (Figure 9d). The high values of HI-DOPA and PB-5SCD in the salmon alone group may be ascribed to the oxidation of tyrosine residues in salmon proteins by tyrosinase. We have previously shown that tyrosine residues in proteins can be oxidized by tyrosinase to DOPA residues and then to CD residues [42]. These results indicate that a considerable amount (33%) of the DOPA oxidized (1,970 ng/mg) was consumed by binding with proteins but was not oxidized to DOPA melanin. As free DOPA (unoxidized DOPA) was at a negligible level (data not shown), DOPA produced by HI hydrolysis should be derived from protein-bound DOPA and CD [39,40]. This is consistent with the 22% yield of PB-5SCD from the salmon + DOPA group. This indicates that PB-5SCD can be considered as a marker of DOPA oxidation (to DOPAquinone). The binding of DOPA with proteins through sulfhydryl groups leads to a lower production of DOPA melanin, which is consistent with the lower A500 value in the salmon + DOPA group compared with the DOPA alone group. Thus, the above experimental results are consistent with the suggestion that RFC include oxidized proteins produced by DOPAquinone and/or DOPAchrome binding to salmon proteins.
3. Discussion
The results of the present study clearly indicate that the black pigment of the MFC is EM derived from DOPA. To the best of our knowledge, this is the first time that this is clearly shown by detailed chemical analytical methods. This EM is probably derived from melanomacrophages which are immune cells that may have entered the muscle for a number of possible reasons. We are not aware that there has been a proper biochemical characterization of the black pigment of fish melanomacrophages yet. Although we did not investigate isolated melanomacrophages directly, our study suggests that their pigment is EM because they are believed to give the black color to MFC.
In contrast, the origin of pigment of the RFC was less obvious. The levels of 4-AHP, HI-DOPA and PB-5SCD in RFC were higher than in MFC and were proportional to the degree of coloration (Figure 6). Based on these results, the red pigment appeared to include conjugations of DOPAquinone and/or DOPAchrome with salmon fillet proteins. To prove this hypothesis, we performed an experiment mimicking the process of RFC production. The exposure of salmon fillet proteins to DOPAquinone (or to DOPAchrome) resulted in the suppression of melanin production and the production of pigment producing PDCA, 4-AHP, HI-DOPA and PB-5SCD. Thus, we observed the characteristic features of RFC in the mimicking experiment except for the PDCA production, which remains puzzling. The production of 4-AHP (and 3-AHP) may be explained by Schiff’s base formation between DOPAquinone and the amino group in lysine residues in salmon proteins [41]. PM was detected only at trace levels in MFC, irrespective of size, indicating that MFC contain little or no PM. This may be because salmon muscle tissue has little cysteine levels compared to other amino acids [43,44], so the availability of cysteine in muscle tissue is limited. Since cysteine is required for the production of pheomelanin, low cysteine levels in salmon muscle are thought to lead to decreased PM production. This is consistent with the previous results that PM in red seabream was below the detection limits [24].
The absolute values of HI-DOPA and PB-5SCD were 40–100 times higher in the salmon + DOPA group than in RFC (Figure 9d). These differences may be ascribed to the fact that DOPA is rapidly oxidized by tyrosinase in the mimicking experiment, whereas in RFC in salmon, the oxidation (production of pigment of RFC) only gradually progresses over a long period of time. The level of PDCA in the salmon + DOPA group in the mimicking experiment was 2-fold higher than from the DOPA alone and the salmon alone groups combined (Figure 9b). This PDCA may arise from DHI-protein conjugates formed via cyclization of protein-bound CD. The reason why PTCA and PDCA values in RFC are similar to those of the control is unknown, but this may also be due to secondary changes of DHI-protein conjugates giving a reddish coloration of RFC (Figure 8).
The spectrum of RFC in Soluene-350 showed a significant increase in absorption below 550 nm, and the absorbance at 400 nm was 5-fold greater than the control at 400 nm (Figure 5). As Soluene-350 has a large absorption below 400 nm, the maximum of absorption spectrum could not be measured, but the large absorption of RFC at 400 nm indicates the existence of quinone with an absorbance maximum at 400 nm or DHI-protein conjugates with nearly 400 nm.
Knowing that MFC pigment consists of melanin may help to reduce MFC numbers by reducing melanin synthesis. For example, from studies in mammals it is known that unsaturated fatty acid decrease melanin synthesis and tyrosinase activity whereas saturated fatty acids increase those activities [45,46]. In a recent study, salmons fed a diet with increased n-3 long-chain polyunsaturated fatty acid, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), had a lower occurence of black spots [47,48].
The etiology of the focal melanization in the white muscle is complex and probably can include a variety of reasons that induce tissue damage (which can cause intramuscular bleeding) and/or immune responses (which can cause melanomacrophages to infiltrate the muscle). The aquaculture industry has also suggested a correlation between RFC (also known as “bleedings”) and MFC, and transient forms have been observed [4]. Our experimental results, however, do not support a possibility that the melanin pigment of MFC is a final product of the pigment found in RFC, which agrees with the common idea that RFC and MFC are derived from different cell types, namely erythrocytes and melanomacrophages. Our current hypothesis is that, in regard to the biochemistry of their melanogenic metabolites, MFC and RFC branch into two chemical pathways in a common biosynthetic pathways of melanogenesis that leads to the production of EM in MFC and melanogenic metabolites derived from protein-bound DOPAquinone and/or DOPAchrome in RFC (shown schematically in Figure 8).
Lastly, we will consider the involvement and role of ascorbic acid and red blood cell (RBC) oxidation on living organisms. To reduce lipid oxidation, researchers have explored adding antioxidants like vitamin E to fish diets or injecting mixtures of antioxidants (ascorbic acid, citric acid, and selenium) into fish fillets. Interestingly, the reaction of ascorbic acid and protein has been reported to produce a red color under aerobic conditions, which was shown to result from an amino-carbonyl reaction of oxidized ascorbic acid (dehydroascorbic acid) [49]. Iron in hemoglobin may contribute to changes in pigment intensity in RFC. It is known that RBC are exposed to reactive oxygen and nitrogen species, which can lead to the production of oxidants [50]. Oxyhemoglobin in RBC can undergo autoxidation to form active oxygen (superoxide, hydrogen peroxide, and so on). Free hemoglobin is particularly toxic and this is evident in several RBC diseases [51]. This oxidation process can also cause the oxidation of tyrosine residues, leading to the formation of dityrosine, dopamine, dopamine quinone, DHI, and other related products [52]. When considering the production of RFC, it is important to note that the above further research and experiments may be necessary to fully understand and confirm these reactions and their implications.
4. Materials and Methods
4.1. Fish material
The fish examined were clinically healthy farmed Atlantic salmon from two different populations. From both populations (Material #1 and #2), fillets were selected from the processing line, specifically focusing on focal discoloured spots located in the cranio-ventral part of the fillets. Stained tissue was carefully excised, and adjacent unstained muscle tissue located 3 cm posterior to the discoloured area was also cut out and utilized as a control. Samples within the same category were combined, frozen at -80°C and shipped to Fujita Health University in Japan in March, 2020, for chemical characterization.
Materials #1: Atlantic salmon with known pedigree, were obtained from the breeding nucleus of Benchmark Genetics, Norway (average body weight 4.2 kg). Out of the 2,005 registered fish at Austevoll Laksepakkeri, Norway, 32 % had focal dark spots on at least one of their fillets. In total, dark stained tissue samples were collected from 636 fillets.
The pigmented areas had various degrees of pigmentation. According to the color, black and red spots were separated. Pigmented spots with the same macroscopic appearance were divided into three groups (small, medium, and large) depending on the pigment intensity and pooled: controls, MFC (BS-small, BS-medium, BS-large), RFC (RS-small, RS-medium and RS-large) (Figure 6). Three pellets of pigmented areas from each group of MFC and RFC were dried in a desiccator (dry weights, 90 to 160 mg) and were homogenized at a concentration of 20 mg/mL in water using a Ten-Broeck glass homogenizer. Aliquots of 200 µL (4 mg) were subjected to AHPO to measure PTCA and PDCA [36], to HI hydrolysis to measure 4-AHP, 3-AHP, and HI-DOPA [35,39] and to solubilization in Soluene-350 to measure A500 and A650 [37]. PB-5SCD was analyzed after precipitation of proteins with 0.4 M HClO4 followed by HCl hydrolysis, as described in [40].
Materials #2: Atlantic salmon (average weight 4 kg) farmed in open commercial-sized sea-cages (Lerøy Midt AS, Gjemnes, Norway) [53]. The fish were filleted at the Lerøy processing plant on Hitra island, Norway. Salmon fillets, dissected from melanized or RFC (31 spotted samples and 2 controls; 250 to 350 mg) were dried in a desiccator (dry weights, 90 to 160 mg) and homogenized at a concentration of 20 mg dry weight/mL in water using a Ten-Broeck glass homogenizer. Aliquots of 100 µL (2 mg) were analyzed as described for Materials #1.
Mushroom tyrosinase (1,715 U/mg) and L-DOPA were purchased from Sigma-Aldrich (St Louis, MO, USA). Soluene-350 was purchased from PerkinElmer (Waltham, MA, USA). Other chemicals are of the highest purity commercially available.
4.2. HPLC conditions
A HPLC system consisting of an analytical UV/VIS detector, a JASCO pump (JASCO Co., Tokyo, Japan), a C18 column (Capcell Pak MG; 4.6 x 250 mm; 5 µm particle size, Osaka Soda, Osaka, Japan) and a JASCO UV-visible detector (JASCO Co., Tokyo, Japan) was used to measure PTCA, PDCA, and TTCA [36]. The mobile phase was 1 mM tetra-n-butylammonium bromide in 0.1 M potassium phosphate buffer (pH 2.1): methanol, 83:17 (v/v) [54]. Analyses were performed at 40°C at a flow rate of 0.7 ml/min. UV-visible spectra were measured using a JASCO V-630 UV-VIS spectrophotometer (JASCO Co., Tokyo, Japan). For the assay of 4-AHP, 3-AHP, and DOPA (HI-DOPA) in the HI hydrolysate, a Catecholpak C18 column was used with 4-AHP buffer – methanol, 98:2 (vol ⁄ vol) at 35°C, with an electrochemical detector set at +500 mV versus an Ag/AgCl electrode. The AHP buffer consisted of 0.1 M sodium citrate buffer, pH 3.0, containing 1 mM sodium octanesulfonate and 2% EDTA.2Na [35]. PB-5SCD was analyzed as described for serum 5SCD [40]. High-resolution MS spectra were obtained using a 6220 TOF mass spectrometer (mode: electrospray ionization-time-of-flight, negative; ESI (-)-TOF)(Agilent Technologies, Santa Clara, CA, USA).
4.3. Preparative isolation of PTCA from MFC
Preparative scale AHPO was performed. MFC from Materials #2 (wet weight 2.0 g) were combined and homogenized in 50 mL 1 M K2CO3, to which 10 mL 30% H2O2 was added. After stirring for 20 h at 25°C, 1.5 g Na2SO3 and 23 mL 6 M HCl were added to stop the oxidation and acidify the mixture to pH <2. After removal of proteins by filtration, the oxidation mixture was extracted 3 times with 100 mL ethyl acetate. The ethyl acetate was removed in vacuo and the residue was dissolved in 0.4 M HCOOH:MeOH, 90:10, and injected into a C18 column (Capcell Pak MG; 20 x 250 mm; 5 µm particle size, Osaka Soda, Osaka, Japan). HPLC separation was performed with a mobile phase of 0.4 M HCOOH:MeOH, 90:10 at a column temperature of 45°C and a flow rate of 7 mL/min. Fractions containing PTCA were recovered in a total yield of 18 µg. A PTCA standard (30 µg) was also injected into the HPLC and the fraction containing PTCA was recovered for comparison of UV spectra and high-resolution MS spectra. High-resolution MS 198.0036 [M-H]−, calc’d for C7H4O6N1, [M-H]−, 198.1044.
4.4. Experiment mimicking the production of RFC in salmon fillets
Solutions containing: 1) 0.2 mM L-DOPA and 20 mg salmon fillet homogenate, 2) 0.2 mM L-DOPA alone, or 3) 20 mg salmon fillet homogenate alone in 1 mL 50 mM sodium phosphate buffer, pH 7.4, were oxidized by mushroom tyrosinase (50 U) at 37°C. After vigorous mixing for 4 h, aliquots of each oxidation mixture were analyzed for melanin markers. Aliquots of 200 µL were mixed with 800 µL Soluene-350 and were then analyzed for A500 and A650 [37]. Aliquots of 100 µL were subjected to AHPO to analyze PDCA and PTCA, to HI hydrolysis to measure 4-AHP, 3-AHP, and HI-DOPA [35], and to HClO4 precipitation followed by HCl hydrolysis to measure PB-5SCD [39,40]. Melanin marker values were normalized per mg salmon protein.
5. Conclusions
In this study we firstly elucidated the melanogenic features of the pigments in MFC and RFC by detailed chemical analytical methods. The analytical results suggest that the pigment of MFC is DOPA-derived EM, and that the pigment of RFC, which are pigments produced by intramuscular hemorrhage, includes melanogenic metabolites derived from the oxidized proteins produced by DOPAquinone and/or DOPAchrome binding to salmon proteins. Although RFC have been reported to change into MFC [1,3], our data show that the pathways of melanogenesis in the production of MFC and RFC are distinct, agreeing with a different biochemical and cellular origin of MFC and RFC pigments.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: The identification of PTCA obtained by isolating PTCA in a preparative scale AHPO in MFC.; Figure S2: The correlation of PTCA with A500 values; Table S1: Average values of spectrophotometric and chemical degradation analyses of control, MFC (BS-small, BS-medium, BS-large) and RFC (RS-small, RS-medium, RS-large) in Materials #1. ; Table S2: Average values of spectrophotometric and chemical degradation analyses of control (n = 2), MFC (BS-small, n = 3; BS-medium, n = 17; BS-large, n = 7) and RFC (n = 3) in Materials #2.
Author Contributions
K.W. and S.I. were involved in the conceptualization and design of the study; S.I. performed the chemical analyses; T.M. was responsible for collecting the biological material (project leader); K.W. contributed to writing the original draft; K.W., J.M.D., T.M. and S.I. contributed to review and editing; All authors have read and agreed to the current version of the manuscript.
Funding
This research was supported in part by The Norwegian Seafood Research Fund (FHF 901487) to K.W and T.M.
Acknowledgments
The authors thank Thomas Larsson, Nofima for assistance in tissue preparation and shipment, and appreciate the mass measurements by Makoto Ojika, Graduate School of Bioagricultural Sciences, Nagoya University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Malik, M. S.; Bjørgen, H.; Nyman, I. B.; Wessel, Ø.; Koppang, E. O.; Dahle, M. K.; Rimstad, E. PRV-1 infected macrophages in melanized focal changes in white muscle of Atlantic salmon (Salmo salar) correlates with a pro-inflammatory environment. Front. Immunol. 2021, 12, 664624. [Google Scholar] [CrossRef] [PubMed]
- Bjørgen, H.; Wessel, Ø.; Fjelldal, P.G.; Hansen, T.; Sveier, H.; Sæbø, H.R.; Enger, K.B.; Monsen, E.; Kvellestad, A.; Rimstad, E.; Koppang, E.O. Piscine orthoreovirus (PRV) in red and melanised foci in white muscle of Atlantic salmon (Salmo salar). Vet. Res. 2015, 46, 89. [Google Scholar] [CrossRef] [PubMed]
- Mørkøre, T.; Larsson, T.; Kvellestad, A.S.; Koppang, E.O.; Åsli, M.; Krasnov, A.; Gannestad, K.H. Mørke flekker i laksefilet. Kunnskapsstatus og tiltak for å begrense omfanget (Melanised changes in salmon fillet. Knowledge status and efforts to limit their occurrence). NOFIMA report. 2015, Retrieved from https://brage.bibsys.no/xmlui/handle/11250/2368359.
- Bjørgen, H.; Haldorsen, R.; Oaland, Ø.; Kvellestad, A.; Kannimuthu, D.; Rimstad, E.; Koppang, E.O. Melanized focal changes in skeletal muscle in farmed Atlantic salmon after natural infection with Piscine orthoreovirus (PRV). J. Fish Dis. 2019, 42, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Larsen, H.A.; Austbø, L.; Mørkøre, T.; Thorsen, J.; Hordvik, I.; Fischer, U.; Jirilo, E.; Rimstad, E.; Koppang, E.O. Pigment-producing granulomatous myopathy in Atlantic salmon: a novel inflammatory response. Fish Shellfish Immunol. 2012, 33, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Macchi, G. J.; Romanol, A.; Christiansen, H.E. Melanomacrophage centres in white-mouth croaker Micropogonias fuerney, as biological indicators of environmental changes. J. Fish Biol. 1992, 40, 971–973. [Google Scholar] [CrossRef]
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Barni, S. Mechanisms of changes to the liver pigmentary component during the annual cycle (activity and hibernation) of Rana esculenta. L. J. Ant. 2002, 200, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Henninger, J.M.; Beresford, W.A. Is it coincidence that iron and melanin coexist in hepatic and other melanomacrophages? Histol. Histopathol. 1990, 5, 457–459. [Google Scholar] [PubMed]
- Ribeiro, H. J.; Procópio, M.S.; Gomes, J.M.; Vieira, F.O.; Russo, R.C.; Balzuweit, K.; Chiarini-Garcia, H.; Castro, A.C.S.; Rizzo, E.; Corrêa, J. D. Jr. Functional dissimilarity of melanomacrophage centres in the liver and spleen from females of the teleost fish Prochilodus argenteus. Cell Tissue Res. 2001, 346, 417–425. [Google Scholar] [CrossRef]
- Dubey, S.; Roulin, A. Evaluation and biomedical consequences of internal melanins. Pigment Cell Melanoma Res. 2014, 27, 327–338. [Google Scholar] [CrossRef]
- Guida, G.; Gallone, A.; Maida, I. , Boffoli, D.; Cicero, R. Tyrosinase gene expression in the Kupffer cells of Rana esculenta L. Pigment Cell Res. 2000, 13, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Purrello, M.; Scalia, M.; Corsaro, C.; Di Pietro, C.; Piro, S.; Sichel, G. Melanosynthesis, differentiation, and apoptosis in Kupffer cells from Rana esculenta. Pigment Cell Res. 2001, 14, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Scalia, M.; Di Pietro, C.; Poma, M.; Ragusa, M.; Sichel, G.; Corsaro, C. The spleen pigment cells in some Amphibia. Pigment Cell Res. 2004, 17, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Sichel, G.; Scalia, M.; Corsaro, C. Amphibia Kupffer cells. Microsc. Res. Tech. 2002, 57, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Bjørgen, H.; Kumar, S.; Gunnes, G.; Press, C.M.; Rimstad, E.; Koppang, E. O. Immunopathological characterization of red focal changes in Atlantic salmon (Salmo salar) white muscle. Vet. Immunol. Immunopathol. 2020, 222, 110035. [Google Scholar] [CrossRef]
- Krasnov, A.; Moghadam, H.; Larsson, T.; Afanasyev, S.; Mørkøre, T. Gene expression profiling in melanised sites of Atlantic salmon fillets. Fish Shellfish Immunol. 2016, 55, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Agius, C. The melano-macrophage centres of fish: A review. In M.J. Manning & M.F. Tatner (Eds.), Fish Immunology, 1985, 85-105. Academic Press. [CrossRef]
- Brimsholm, M.; Fjelldal, P.G.; Hansen, T.; Fraser, T.K.W.; Solberg, M.; Glover, K.; Koppang, E.O.; Bjørgen, H. Red and melanized focal changes in white skeltal muscle in Atlantic salmon (Salmo sala): Comaprative analysis of farmed, wild and hybrid reared under identical conditions. J. Fish Dis. 2023. [Google Scholar] [CrossRef]
- Gallone, A.; Sagliano, A.; Guida, G.; Ito, S.; Wakamatsu, K.; Capozzi, V.; Perna, G.; Zanna, P.; Cicero, R. The melanogenic system of the liver pigmented macrophages of Rana esculenta L. – Tyrosinase activity. Histol. Histopathol. 2007, 22, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Thorsen, J.; Høyheim, B.; Koppang, E.O. Isolation of the Atlantic salmon tyrosinase gene family reveals heterogenous transcripts in a leukocyte cell line. Pigment Cell Res. 2006, 19, 327–336. [Google Scholar] [CrossRef]
- Mackintosh, J.A. The antimicrobial properties of melanocytes, melanosomes and melanin and the evolution of black skin. J. Theor. Biol. 2001, 211, 101–113. [Google Scholar] [CrossRef]
- McNamara, M.E.; Rossi, V.; Slater, T.S.; Rogers, C.S.; Ducrest, A.-L.; Dubey, S.; Roulin, A. Decoding the evolution of melanin in vertebrates. Trends Ecol. Evol. 2021, 36, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Kato, K.; Wakamatsu, K.; Ito, S.; Ishimaru, K.; Hirata, T.; Murata, O.; Kumai. H. The histological analysis, colorimetric evaluation, and chemical quantification of melanin content in ‘suntanned’ fish. Pigment Cell Res. 2005, 18, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Sugumaran, M. Reactivities of quinone methides versus o-quinones in catecholamine metabolism and eumelanin biosynthsis. Int. J. Mol. Sci. 2016, 17, 1576. [Google Scholar] [CrossRef]
- d’Ischia, M.; Wakamatsu, K.; Napolitano, A.; Briganti, S.; Garcia-Borron, J-C. ; Kovacs, D.; Meredith, P.; Pezzella, A.; Picardo, M.; Sarna, T.; Simon, J.D.; Ito, S. Melanins and melanogenesis: methods, standards, protocols. Pigment Cell Melanoma Res. 2013, 26, 616–633. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, M. L.; Panzella, L. The multifaceted opportunities provides by the pheomelanin-inspired 1,4-benzothiazine chromophore: a still-undervalued issue. Molecules, 2023, 28, 6237. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Ito, S. Melanins in Vertebrates. 2021, pp 45-89, In: Hashimoto H., Goda M., Futahashi R., Kelsh R., Akiyama T. (eds). Pigments, Pigment Cells and Pigment Patterns. Springer, Singapore. [CrossRef]
- Land, E.J.; Ramsden, C.A.; Riley, P.A. Tyrosinase autoactivation and the chemistry of ortho-quinone amines. Acc. Chem. Res. 2003, 36, 300–308. [Google Scholar] [CrossRef]
- Pawelek, J.M.; Körner, A.M.; Bergstrom, A.; Bologna, J. New regulators of melanin biosynthesis and the autodestruction of melanoma cells. Nature, 1980, 286, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V.J. A second tyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519–526. [Google Scholar] [CrossRef] [PubMed Central]
- Kroumpouzos, G.; Urabe, K.; Kobayashi, T.; Sakai, C.; Hearing, V.J. Functional analysis of the slaty gene product (TRP2) as dopachrome tautomerase and the effect of a point mutation on its catalytic function. Biochem. Biophys. Res. Commun. 1994, 202, 1060–1068. [Google Scholar] [CrossRef]
- Ito, S.; Suzuki, N.; Takebayashi, S.; Commo, S.; Wakamatsu, K. Neutral PH and copper ions promote eumelanogenesis after the dopachrome stage. Pigment Cell Melanoma Res. 2013, 26, 817–825. [Google Scholar] [CrossRef]
- Guo, L.; Li, W.; Gu, Z.; Wang, L.; Guo, L.; Ma, S.; Li, C.; Sun, J.; Han, B.; Chang, J. Recent advances and progress on melanin: from source to application. Int. J. Mol. Sci. 2023, 24, 4360. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Ito, S.; Rees, J.L. The usefulness of 4-amino-3-hydroxyphenylalanine as a specific marker of peheomelanin. Pigment Cell Res. 2002, 15, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Nakanishi, Y.; Valenzuela, R.K.; Brilliant, M.H.; Kolbe, L.; Wakamatsu, K. Usefulness of alkaline peroxde oxidation to analyze eumelanin and pheomelanin in various tissue samples: application to chemical analysis of human hair melanin. Pigment Cell Res. 2011, 24, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Ozeki, H.; Ito, S.; Wakamatsu, K.; Thody, A.J. Spectrophotometric characterization of eumelanin and pheomelanin in hair. Pigment Cell Res. 1996, 9, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Fujikawa, K.; Zucca, F.A.; Zecca, L.; Ito, S. The structure of neuromelanin as studied by chemical degradative methods. J. Neurochem. 2003, 86, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Kato, T.; Maruta, K.; Jimbow, K.; Fujita, K. “Total”acidic metabolites of catecholamines in urine as determined by hydrolysis with hydriodoc acid and liquid chromatograohy: application to patients with neuroblastoma and melanoma. Clin. Chem. 1985, 31, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Kato, T.; Fujita, K. Covalent binding of catechols to proteins through the sulphydryl group. Biochem. Pharmacol. 1988, 37, 1707–1710. [Google Scholar] [CrossRef]
- Ito, S.; Sugumaran, M.; Wakamatsu, K. Chemical reactivities of ortho-quinones produced in living organisms: Fate of quinoid products formed tyrosinase and phenoloxidae action on phenols and catechols. Int. J. Mol. Sci. 2020, 21, 6080. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Kato, T.; Shinpo, K.; Fujia, K. Oxidation of tyrosine residues in proteins by tyrosinase. Formation of protein-bonded 3,4-dihydroxyphenylalanine and 5-S-cysteinyl-3,4-dihydroxyphenylalanine. Biochem. J. 1984, 222, 407–411. [Google Scholar] [CrossRef]
- Sathival, S.; Smiley, S.; Prinyawiwatkul, W.; Bechtel, P.J. Functional and nutritional properties of red salmon (Oncorhynchus nerka) enzymatic hydrolysates. J. Food Sci. 2005, 70, C401–C406. [Google Scholar] [CrossRef]
- Razzaq, W.; Masood, Z.; Hassan, H.U.; Benzar, S.; Nadeem, K.; Arai, T. An investigation on protein and amino acid contents in scales and muscles of pomfret Parastromateus niger (Bloch, 1795) and Pampus argenteus (Eupharasen, 1788). Brazilian J. Biol. 2024, 84, e258880. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Ryu, A.; Hashimoto, A.; Oka, M.; Ichihashi, M. Linoleic acid and alpha-linolenic acid lightens ultraviolet induced hyperpigmentation of the skin. Arch. Dermatol. Res. 1998, 290, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Shono, S.; Toda, K. Phenotypic expression in pigment cell. In Pigment Cell, M. Seiji, ed. (Tokyo, Japan: University of Tokyo Press), 1981, pp. 263-268.
- Sissener, N.H.; Waagbø, R.; Rosenlund, G.; Tvenning, L.; Susort, S.; Lea, T.B.; Oalamd, Ø.; Chen, L.; Breck, O. Reduced n-3 long chain fatty acid levels in feed for Atlantic salmon (Salmo salar L.) do not reduced growth, robustness or product quality through an entire full scale commercial production cycle in seawater. Aquac. 2016, 464, 236–245. [Google Scholar] [CrossRef]
- Lutfi, E.; Berge, G.M.; Bæverfjord, G.; Sigholt, T.; Bou, M.; Larsson, T.; Mørkøre, T.; Evensen, Ø.; Sissener, N.H.; Rosenlund, G.; Sveen, L.; Østbye, T-K. ; ruyter, B. Increasing dietary levels of the n-3 long-chain PUFA, EPA and DHA, improves the growth, welfare, rubustness and fillet quality of Atlantic salmon in sea cages. Br. J. Nut. 2023, 129, 10–28. [Google Scholar] [CrossRef] [PubMed]
- Balcos, M.C.; Kim, S.Y.; Jeong, H-S. ; Yun, H-Y.; Baek, K.J.; Kwon, N.S.; Park, K-C.; Kim, D-S. Docosahexaenoic acid inhibits melanin synthesis in murine melanoma cells in vitro through increasing tyrosinase degradation. Acta Pharmacol. Sin. 2014, 35, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Terao, A.; Ueda, S.; Namiki, M. Red pigment formation by the reaction of oxidized adcorbic acid and protein in a food model system of low moisture content. Agric. Biol. Chem. 1985, 49, 3139–3144. [Google Scholar] [CrossRef]
- Alayash, A.I.; Patel, R.P.; Cashon, R.E. Redox reactions of hemoglobin and myoglobin: Biological and toxicological implications. Antioxid. Redox Signal. 2001, 3, 313–327. [Google Scholar] [CrossRef]
- Schaer, D.J.; Buehler, P.W.; Alayash, A.I.; Belcher, J.D.; Vercellotti, G.M. Hemolysis and free hemoglobin revisited: Exploring hemoglobin and hemin scavengers as a novel class of therapeutic proteins. Blood 2013, 121, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Guerrero, R.; Baeverfjord, G.; Evensen, Ø.; Hamre, K.; Larsson, T.; Dessen, J-E. ; Gannestad, K-H.; Mørkøre, T. Rib abnormalities and their association withfocal dark spots in Atlantic salmon fillets. Aquac. 2022, 561, 738697. [Google Scholar] [CrossRef]
- Ito, S.; Del Bino, S.; Hirobe, T.; Wakamatsu, K. Improved HPLC conditions to determine eumelanin and pheomelanin in biological samples using an ion pair reagent. Int. J. Mol. Sci. 2020, 21, 5134. [Google Scholar] [CrossRef]
Figure 1.
The typical position of (A) red spots (red focal change (RFC)) and (B) black spots (melanized focal change (MFC)) on Atlantic salmon fillets.
Figure 1.
The typical position of (A) red spots (red focal change (RFC)) and (B) black spots (melanized focal change (MFC)) on Atlantic salmon fillets.

Figure 2.
Summary of chemical degradation by AHPO and HI hydrolysis of EM and PM. EM consists of DHI and DHICA (with a carboxyl group) units. For details, see the Introduction.
Figure 2.
Summary of chemical degradation by AHPO and HI hydrolysis of EM and PM. EM consists of DHI and DHICA (with a carboxyl group) units. For details, see the Introduction.
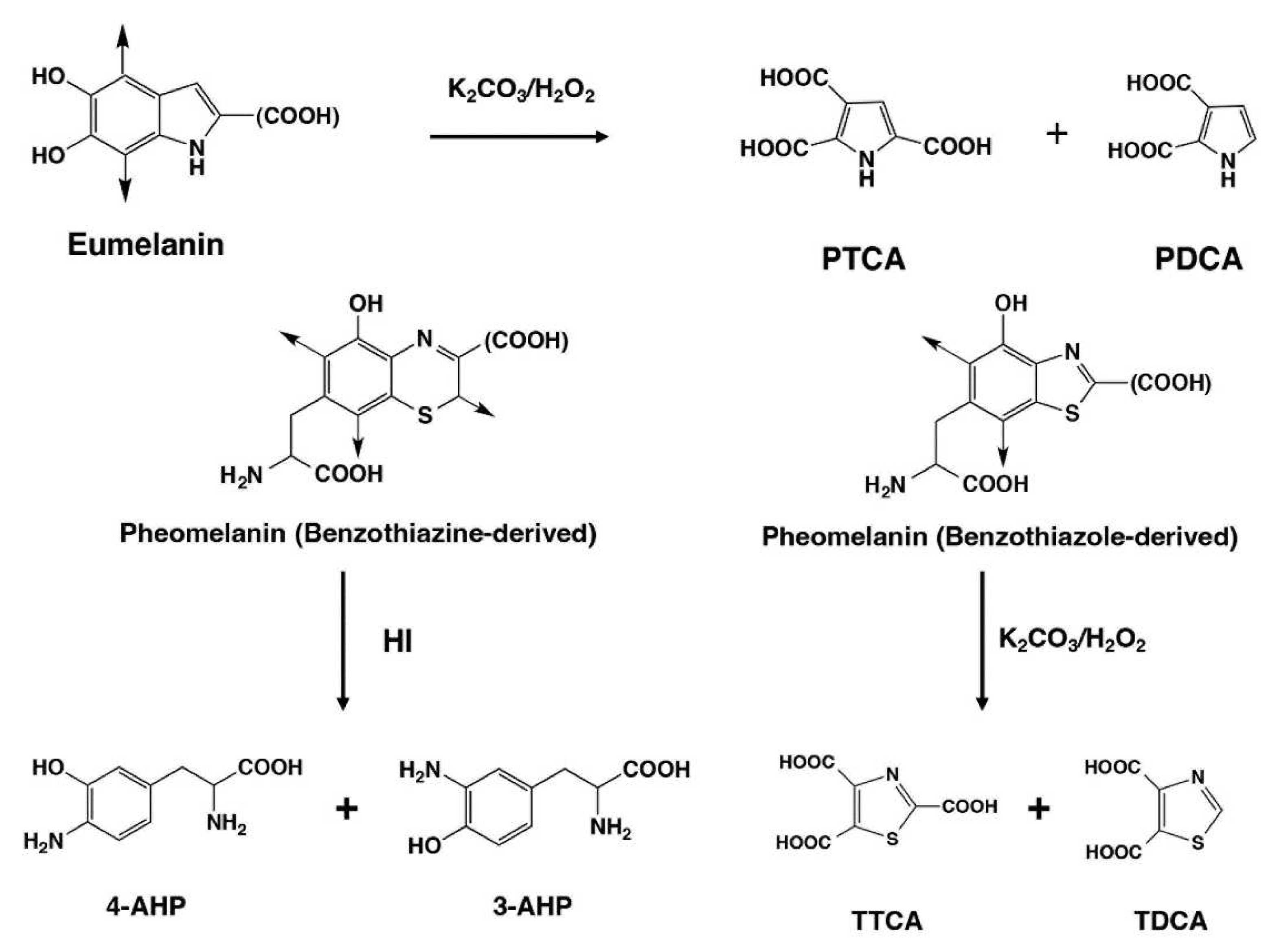
Figure 3.
Macroscopic grading of focal black spots (BS) (a-c) and red spots (RS) (d-f) of the cranioventral section of Atlantic salmon fillets (Material #1). The spots were classified into three categories based on their size (diameter) and pigmentation: small/weak stained spots (<3 cm), medium-sized spots (3 cm) with clear discolouration, and large spots (3-6 cm) with distinct discolouration.
Figure 3.
Macroscopic grading of focal black spots (BS) (a-c) and red spots (RS) (d-f) of the cranioventral section of Atlantic salmon fillets (Material #1). The spots were classified into three categories based on their size (diameter) and pigmentation: small/weak stained spots (<3 cm), medium-sized spots (3 cm) with clear discolouration, and large spots (3-6 cm) with distinct discolouration.

Figure 4.
HPLC chromatograms of AHPO (a, b) and HI hydrolysis (c, d) of BS-large (a, c) and RS-large (b, d), respectively.
Figure 4.
HPLC chromatograms of AHPO (a, b) and HI hydrolysis (c, d) of BS-large (a, c) and RS-large (b, d), respectively.
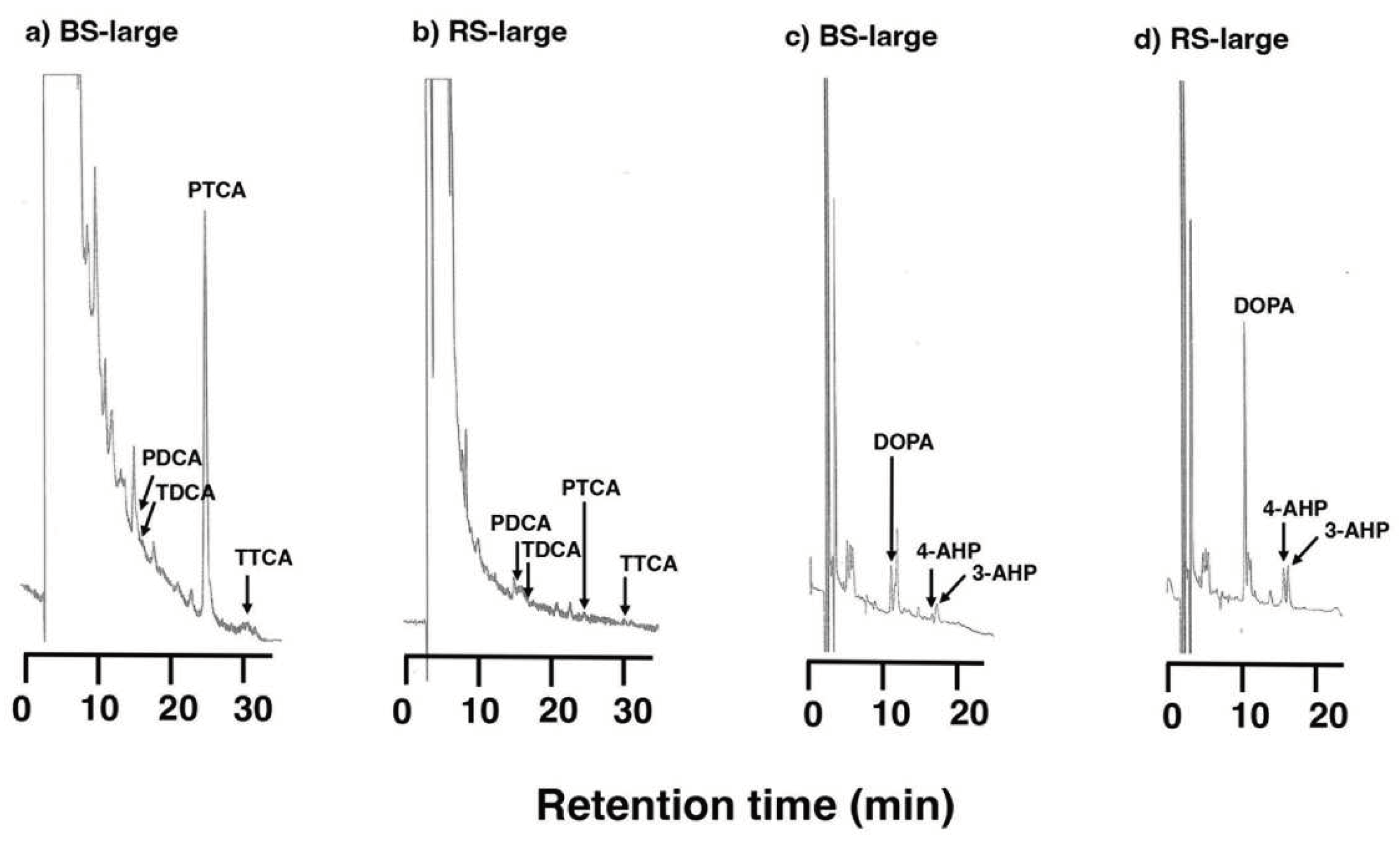
Figure 5.
(a) UV-VIS absorption spectra of MFC and RFC solution in Soluene-350. (b) Difference spectra of MFC and RFC solution that are subtracted from the control spectrum, respectively.
Figure 5.
(a) UV-VIS absorption spectra of MFC and RFC solution in Soluene-350. (b) Difference spectra of MFC and RFC solution that are subtracted from the control spectrum, respectively.
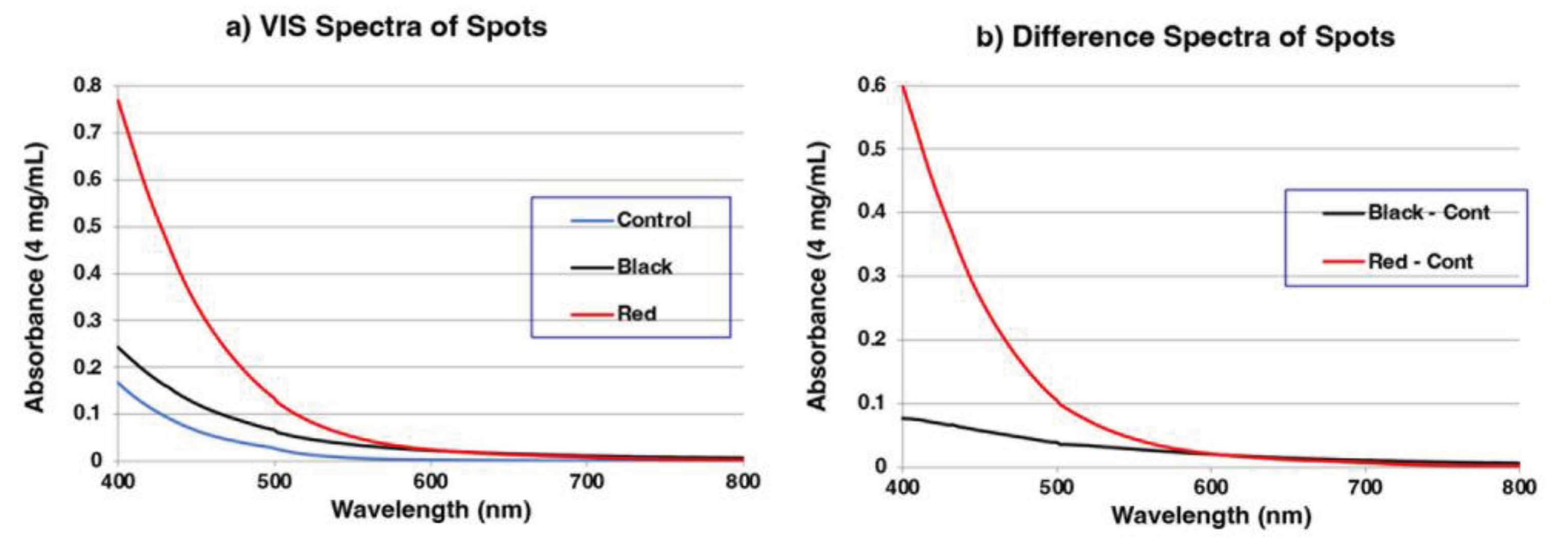
Figure 6.
Summary of spectrophotometric and chemical degradation analyses of control, MFC (BS-small, BS-medium, BS-large) and RFC (RS-small, RS-medium, RS-large) in Materials #1. The number of samples in each group = 3. Since A500, A650/A500 ratios, PDCA and 4-AHP were small values, these values were multiplied by 1000, 100, 2, and 10, respectively. Error bars represent one standard error from the mean.
Figure 6.
Summary of spectrophotometric and chemical degradation analyses of control, MFC (BS-small, BS-medium, BS-large) and RFC (RS-small, RS-medium, RS-large) in Materials #1. The number of samples in each group = 3. Since A500, A650/A500 ratios, PDCA and 4-AHP were small values, these values were multiplied by 1000, 100, 2, and 10, respectively. Error bars represent one standard error from the mean.
Figure 7.
Summary of spectrophotometric and chemical degradation analyses of control (n = 2), MFC (BS-small, n = 3; BS-medium, n = 17; BS-large, n = 7) and RFC (1 RS-small, 2 RS-medium, n = 3) in Materials #2. As A500, A650/A500 ratios, PDCA and 4-AHP showed small values, these values were multiplied by 1000, 100, 2, and 10, respectively. Error bars represent one standard error from the mean.
Figure 7.
Summary of spectrophotometric and chemical degradation analyses of control (n = 2), MFC (BS-small, n = 3; BS-medium, n = 17; BS-large, n = 7) and RFC (1 RS-small, 2 RS-medium, n = 3) in Materials #2. As A500, A650/A500 ratios, PDCA and 4-AHP showed small values, these values were multiplied by 1000, 100, 2, and 10, respectively. Error bars represent one standard error from the mean.
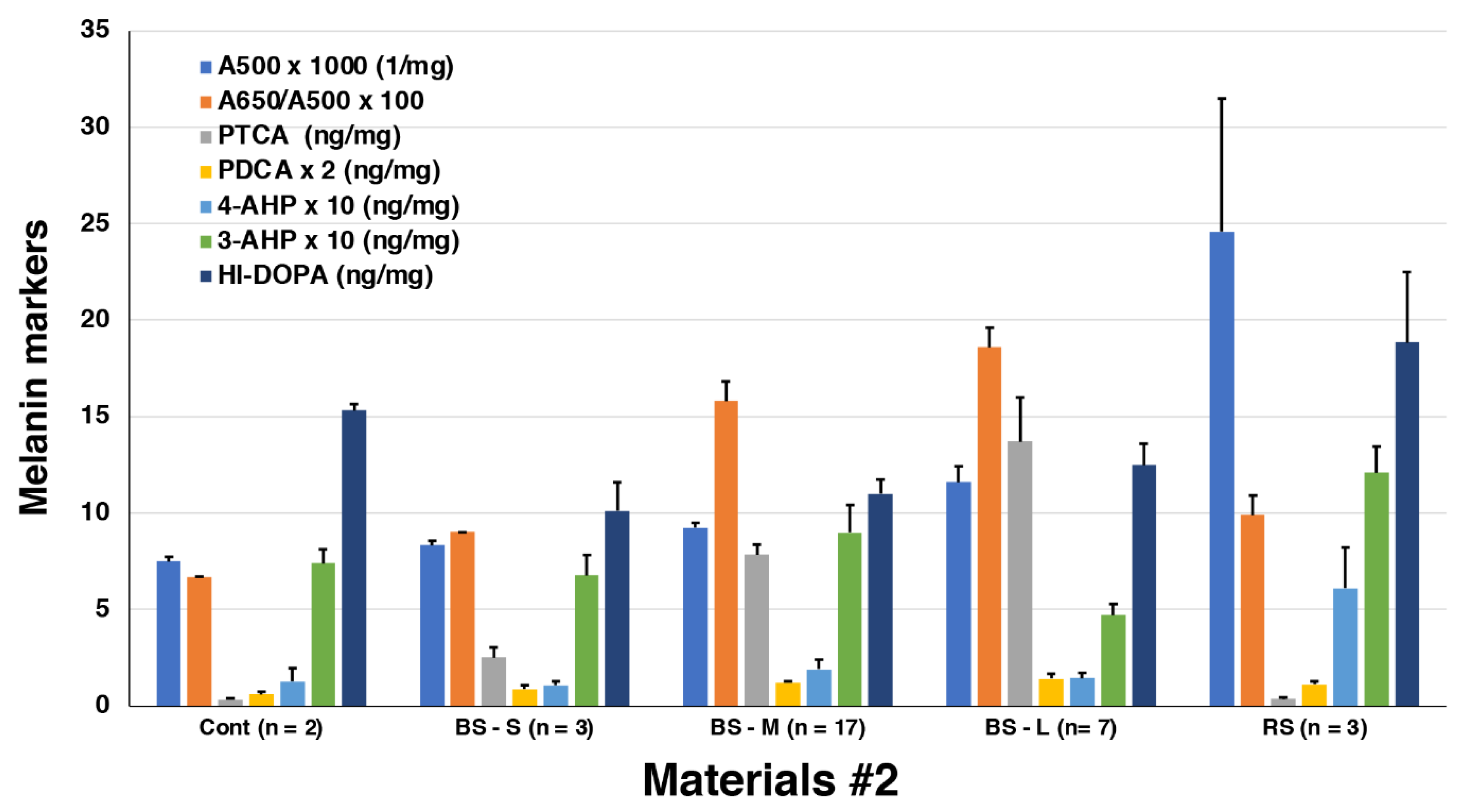
Figure 8.
Our current hypothesis of the biosynthetic pathway of the melanogenic components of MFC and RFC. The oxidation of DHI and DHICA via DOPAchrome gives MFC (EM). On the other hand, the protein-bound DOPAquinone and DOPAchrome formed by the reaction of DOPAquinone and DOPAchrome with salmon proteins produce protein-bound DOPA and protein-bound DHI included in RFC.
Figure 8.
Our current hypothesis of the biosynthetic pathway of the melanogenic components of MFC and RFC. The oxidation of DHI and DHICA via DOPAchrome gives MFC (EM). On the other hand, the protein-bound DOPAquinone and DOPAchrome formed by the reaction of DOPAquinone and DOPAchrome with salmon proteins produce protein-bound DOPA and protein-bound DHI included in RFC.
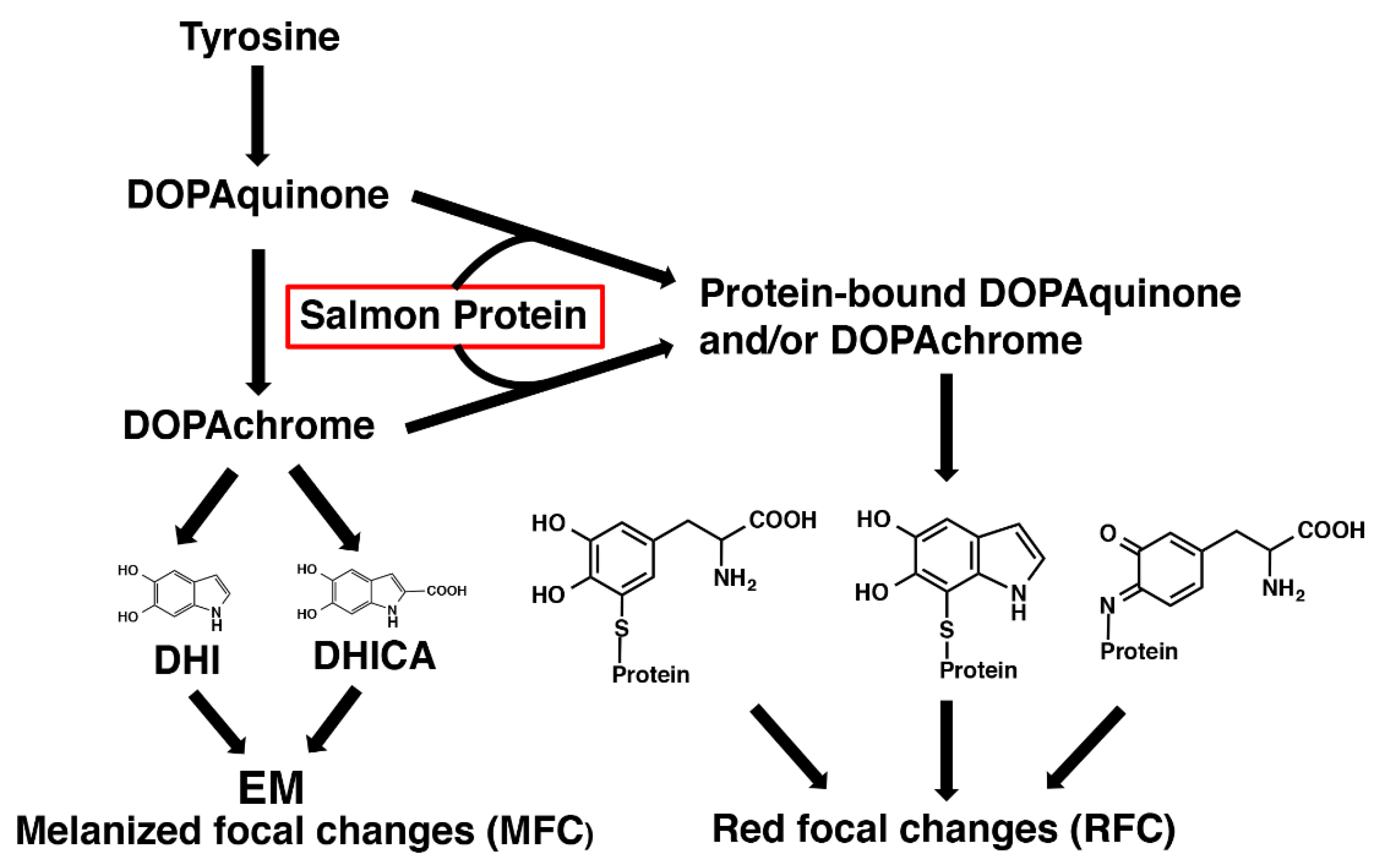
Figure 9.
Summary of the mimicking experiment. Mushroom tyrosinase was added to mixtures of salmon fillet homogenate (20 mg/mL) + DOPA (1 mM), DOPA (1 mM) alone, and salmon fillet (20 mg/mL) alone followed by measuring the melanin markers. Error bars represent one standard error from the mean.
Figure 9.
Summary of the mimicking experiment. Mushroom tyrosinase was added to mixtures of salmon fillet homogenate (20 mg/mL) + DOPA (1 mM), DOPA (1 mM) alone, and salmon fillet (20 mg/mL) alone followed by measuring the melanin markers. Error bars represent one standard error from the mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



