Submitted:
30 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
The new class of mRNA vaccines is studied extensively, leading to many novel prospects based on the pivotal success of vaccines against the SARS-Cov-2 virus. Many novel mechanisms of immune response discovered have led to the possibility of mutation-resistant vaccines by combining multiple conserved epitopes as antigens to manipulate the T-cell and B-cell responses; these antigens can also come from different organisms, providing protection against numerous diseases and of most tremendous significance is the utility of mRNA vaccines in preventing and treating more than 100 autoimmune disorders. This is a significant humanitarian breakthrough given the ease and faster and low cost of mRNA vaccines, being a chemical entity. This paper provides a prospective analysis of mRNA vaccines with much broader applications than anticipated when these vaccines entered treating SARS-Cov-2 infections.
Keywords:
T-cell-mediated immunity
; vaccines
; infectious diseases
; viral vectors
; mRNA vaccines
Introduction
Early vaccines, such as those developed by Edward Jenner in the 18th century, relied on inoculating live or inactivated pathogens to induce immune responses and confer immunity. Over time, vaccine technology evolved, introducing methods to subunit vaccines, viral vectors, and nucleic acid-based vaccines. The latter, including mRNA vaccines, represents a paradigm shift in vaccine development. The landscape of vaccine development has been profoundly transformed by advancements in technology, marking a departure from traditional approaches. Analytical tools, capable of dissecting the molecular intricacies of pathogens and host responses, provide unprecedented insights. Manufacturing technologies, including recombinant DNA techniques, enable the production of highly purified vaccine components.
mRNA vaccines, like those developed for COVID-19 by Pfizer-BioNTech and Moderna, have revolutionized the field of vaccinology, offering several distinct advantages over traditional platforms. First and foremost, the speed of development is unparalleled. Once the genetic sequence of a virus is available, an mRNA vaccine can be designed in just a few days—a process crucial for the rapid response to the COVID-19 pandemic [1]. Key observations include:
- Traditional vaccine development techniques are time-consuming; conversely, in-silico epitope-based vaccines are developed quickly with high precision. [2]
- Reverse vaccinology, a branch of bioinformatics, can find novel antigens that conventional approaches can miss yet are essential for the immunogenicity of next-generation vaccines.
- Furthermore, unlike traditional vaccines that might require adjuvants to bolster the immune response, mRNA vaccines have been shown to elicit strong immunity without them [3].
- Additionally, mRNA vaccines don't have the risk of vector immunity, a concern with viral vector vaccines where prior exposure to the vector might reduce vaccine efficacy [4].
- Another notable advantage is the scalability of mRNA vaccine production, as they can be synthesized without the need for cell cultures, facilitating consistent and large-scale production [5].
- Importantly, mRNA does not integrate into the host genome, alleviating concerns associated with insertional mutagenesis [6].
- The first-generation COVID-19 mRNA vaccines posed challenges with ultra-cold storage requirements that are now resolved with improved formulations. [7]
Moreover, because of the flexibility in designing mRNA sequences, there's considerable interest in creating vaccines that can simultaneously target multiple pathogens or strains of a single pathogen. This could be revolutionary for diseases with high mutation rates or for developing broad-spectrum vaccines [8].
However, as with all innovations, there are challenges to overcome. Although mostly mild and transient, the side effects associated with mRNA vaccines require continuous monitoring and study to ensure long-term safety [9]. Additionally, given that mRNA vaccines represent a relatively new technological frontier in human medicine, there's a need for continued research to understand their long-term effects and to address any potential concerns the public may have.
With the success of the COVID-19 mRNA vaccines, investments in this technology are expected to grow, leading to innovations that may address current limitations and open the door to new therapeutic possibilities [10].
mRNA technology represents a momentary innovation and paradigm shift in how we approach therapeutic design and vaccination. MRNA vaccines can be seen as a platform technology where the core delivery mechanism remains consistent, allowing for streamlined adaptations to various diseases or conditions [11].
As of the end of 2022, there were more than 50 vaccines approved globally (15) [12] , and according to the tracking by the World Health Organization, there are 199 SARS-Cov-2 vaccines in the preclinical stage and 183 in the clinical stage [13]. Globally, as of 18 October 2023, there have been 771,407,825 confirmed cases of COVID-19, including 6,972,152 deaths, reported to WHO [14]. A better metric of current vaccine development can be gained from the research on preventing SARS-CoV-2. Table 1 (Source WHO) lists the types of vaccines that are being developed. There are more than 200 vaccines in the testing phase and an equal number in early development. A review of clinicaltrials.gov [15] shows 761 clinical trials registered, of which 590 are interventional comprising: Early Phase 1 (10), Phase 1 (199), Phase 2 (234), Phase 3 (170), Phase 4 (55), Not applicable (49).
However, despite the clear advantages of mRNA vaccines, other vaccines continue to grow, as evidenced by the SARS-Cov-2 vaccines under development. [WHO data]
Interestingly, over a dozen mRNA vaccines were discontinued due to suboptimal efficacy, and all these vaccines were based on natural bases, not the modified ones used by the BioNtech and Moderna vaccines. The base modification for mRNA vaccines was the subject of the Nobel Prize in Medicine and Physiology in 2023 [17] to Katalin Karikó and Drew Weissman. This observation highlights the possibilities of further advancements in the efficacy and safety of mRNA vaccines.
The potential applications of mRNA vaccines extend beyond infectious diseases. Researchers are exploring their utility in developing cancer vaccines and therapies for genetic disorders, demonstrating the broad applicability of this technology [18]. For instance, mRNA therapies are being investigated for their potential in personalized cancer treatments, where a patient's unique tumor antigens could be targeted, ushering in a new era of precision medicine [19].
Patients with melanoma and pancreatic ductal adenocarcinoma have successfully induced tumor-specific T-cell responses using therapeutic mRNA vaccination combined with checkpoint inhibition [20]. The mRNA used in these experiments contained unmodified bases, but the poly(A) tail was modified [21] to increase mRNA stability and translational efficiency. Thus, alternative methods are being developed to generate effective mRNA-based vaccines and therapeutics. It is anticipated that the number of clinical trials employing various forms of mRNA to induce prophylactic or therapeutic responses in the fields of infection [22] and cancer [23] will increase in the coming years.
One metric of research interest can be derived from the research publications on the topic. At the same time, there were less than 50 publications before the first mRNA vaccine's approval; now, more than 4,000 papers are published on mRNA vaccines. [24]
The longevity of a vaccine is highly variable; for example, vaccination requirements for bacterial and viral infections are influenced by the specific characteristics of the pathogens and the vaccines themselves. Often, bacterial vaccines require booster shots to maintain immunity over time, such as the vaccines for tetanus and diphtheria with boosters every ten years. In contrast, viral vaccines, like those for measles, mumps, and rubella, confer long-lasting or lifelong immunity after the initial vaccinations. However, exceptions exist, such as the influenza vaccine, which requires annual administration due to the virus's rapid mutation and the emergence of new strains. Thus, the need for repeat vaccinations is not strictly tied to the nature of the infection (bacterial or viral) but depends on immunity duration, pathogen variability, and vaccine efficacy.
Scientific advancements in analytical and manufacturing tools, coupled with computational technologies supported by Artificial Intelligence (AI) and Machine Learning (ML), are facilitating the reimagining of vaccines for their prolonged activity. Moreover, applying these tools is paving the way for creating vaccines targeting multiple pathogens or their diverse antigens.
Artificial intelligence, which now offers the capabilities to decipher the enormous complexity of the human immune system and influence the future of human health, has revolutionized many sectors of business and society. [25] We can now assess host responses to infectious and non-communicable diseases, vaccines, and immunotherapies in detail thanks to the development of single-cell and high-throughput transcriptomics, metabolomics, proteomics, epigenetics assays, and mass cytometry and biosensors. We must comprehend human biology and immunological diversity better since most of the world's illness burden affects vulnerable groups with biologically diverse immune systems.
The complexity of the human immune system may now be understood using Deep learning, probabilistic, and hybrid models, as well as the use of supercomputing resources to undertake sophisticated biological system simulations. [26] For instance, "immune age" parameters—measures of inflammatory indicators strongly associated with multimorbidity, immunological senescence, frailty, and cardiovascular aging—have been identified using deep learning approaches. [27] Additionally, AlphaFold used deep learning to predict the 3D structures of 200 million known proteins. It will not be able to predict the structure of a mutation or a random sequence, indicating that there is still a lot of potential to challenge the AI-based protein structuring paradigm. [28]
The discovery of treatments, particularly vaccinations, for sudden outbreaks and unidentified infections has been dramatically accelerated by advancements in bioinformatics. [29] Developing AI models for the human immune system will be the next significant advance in biomedicine.
Reverse vaccinology (RV), which analyses the complete viral genome using computer software to detect pathogen antigens/epitopes, is the most practical use of bioinformatics in vaccine production that can best benefit the emergence of newer mRNA vaccines. [30,31]
Advances in immune profiling technologies, such as flow cytometry mass cytometry (CyTOF) and high-throughput and high-dimensional sequencing, have enabled in-depth analyses of immune responses at the single-cell level. [32] These technologies allow researchers to identify and quantify specific T-cell subsets, cytokine profiles, and immune activation markers associated with protection. Biomarkers from such analyses are valuable tools for evaluating vaccine efficacy and guiding vaccine design.
This paper presents the current scientific understanding of mRNA technology, which is fast growing, with advice and plans for developers to adopt this technology as the primary means of future disease prevention and treatment.
Immune System
Understanding the functionality of the immune system is pivotal in vaccine science. The system constitutes a sophisticated network involving organs, cells, and proteins, operating to safeguard the body against infections while ensuring the protection of its cells. (Figure 1)
At its core, the immune system comprises hematopoietic cells originating from the bone marrow through a process known as hematopoiesis. These cells further differentiate into two major lineages: myeloid and lymphoid. The myeloid lineage gives rise to various immune cells, including neutrophils, monocytes, macrophages, and dendritic cells. Neutrophils, often called first responders, are crucial for rapid defense against bacterial infections. Upon entering tissues, monocytes can mature into macrophages, contributing to immune surveillance and the clearance of cellular debris. Dendritic cells specialize in presenting antigens to initiate specific immune responses. [33,34]
On the other hand, the lymphoid lineage encompasses cells involved in adaptive immunity, which includes B cells, T cells, and natural killer (NK) cells. B cells produce antibodies, essential components of humoral immunity that target pathogens circulating in bodily fluids. T cells, categorized into helper (CD4+) and cytotoxic (CD8+) subsets, play pivotal roles in coordinating immune responses and directly eliminating infected or aberrant cells. NK cells contribute to the rapid recognition and elimination of virus-infected cells. This intricate division of labor within the immune system allows for a multifaceted and highly coordinated defense mechanism, highlighting the complexity and sophistication that underlie the body's ability to combat a diverse array of threats. [35]
A portion of the stimulated T and B cells differentiate into memory cells in the weeks that follow vaccination, but the bulk of these cells incur death. Importantly, immune memory cells play a crucial role in reducing the severity of infectious diseases. During the 2009 H1N1 pandemic, older adults with pre-existing cross-reactive CD4+ T cells prevented severe influenza symptoms. [36]
Cellular Immunity
Strategies to promote enduring T-cell memory hold the potential for sustained protection against infections and the prevention of cancer recurrence [37]. Cross-reactive T-cell-mediated immunity plays a pivotal role in enhancing immune responses against severe acute respiratory syndrome SARS-CoV-2 infection, wherein about 40–60% of individuals who had not been previously exposed to SARS-CoV-2 possessed virus-specific T-cell immunity. This pre-existing immunity is attributable to past exposure to common coronaviruses, and it may help specific patient populations have a less severe illness development. [38,39]
Memory T cells exist in various subpopulations, each serving distinct roles. Effector memory T cells (Tem) play a vital role in immunosurveillance by continuously recirculating between tissues and blood through the lymphatic system. T cells with central memory (Tcm) monitor secondary lymphoid organs. Without recirculation, memory T cells reside at infection-frontline sites like the skin, lungs, and gut, where tissue-resident memory T cells (Trm) play crucial roles in mounting a rapid local immune response to prevent pathogen invasion.
For vaccination protection to persist, memory cells must be maintained. While the germinal center (GC) response produces long-lasting memory B cells, interleukin-15 (IL-15) controls the lifetime and proliferation of memory T cells. [40]
T-cells recognize short peptides displayed on the cell surface in complex with human leukocyte antigen (HLA) class I or II molecules. Memory T-cells can rapidly develop effector functions to inhibit viral replication, limit infection, and prevent host-wide spread upon recognizing their cognate peptide presented by HLA molecules. CD8+ T-cells, for example, directly kill infected cells and produce antiviral cytokines and inflammatory molecules that recruit additional immune cells to infection sites. Some CD4+ T-cells possess direct antiviral properties like CD8+ T-cells. CD4+ T-cells contribute to various aspects of immunity, including supporting B cell responses in the GC; some CD4+ T-cells can also have direct antiviral properties like those of CD8+ T-cells. Memory T-cells can be maintained for decades in humans, and memory T-cells specific to SARS–CoV–2 can persist for decades. T-cells persist after vaccination or infection. [41]
MHC molecules are integral to the immune response, presenting antigens to T cells [42]. MHC class I molecules present antigens derived from intracellular pathogens. Proteins, including those from pathogens, are broken down into peptides by proteasomes in the cytoplasm. These peptides are then transported into the endoplasmic reticulum (ER) by Transporter Associated with Antigen Processing (TAP), where they bind to MHC class I molecules. The peptide-MHC class I complexes are presented on the cell surface for recognition by CD8+ T cells [43]. (Figure 2)
On the other hand, MHC class II molecules present antigens from extracellular pathogens. Pathogens are engulfed by endocytosis or phagocytosis and degraded into peptides within endosomes or lysosomes. These peptides bind to MHC class II molecules and are presented on the cell surface, where they can be recognized by CD4+ T cells [44]. The interaction between T cells and antigens presented by MHC molecules is crucial for adaptive immune responses, including cell-mediated immunity and the activation of B cells to produce antibodies. [45]
This highly specific recognition process allows T-cells to discriminate between foreign invaders and healthy host cells. [46] Upon successful antigen recognition, T-cells undergo activation, triggering a cascade of events leading to their differentiation into effector T-cells, which mediate immune responses, or memory T-cells, which provide long-lasting immunity. [47]
The fundamental aspects of T-cell immunity are a primary concern in the design of vaccines [48] , where the indispensable partnership between CD4+ T helper cells and B cells in generating protective humoral immunity proves pivotal. [49] Adaptive immune responses must be developed to control and eradicate harmful infections. Memory T-cells offer an extra line of defense since T-cells can only recognize invading viruses after they have entered the host cell; as a result, they cannot stop infection but may limit viral reproduction and spread after infection.
Immune checkpoint pathways, such as PD-1/PD-L1 and CTLA-4, play pivotal roles in modulating T-cell responses within the immune system. These pathways act as control mechanisms to balance the immune system's activity, preventing excessive responses that could lead to autoimmune reactions or tissue damage. PD-1 (Programmed Cell Death Protein 1) and CTLA-4 (Cytotoxic T-Lymphocyte-Associated Protein 4) are cell surface receptors found on T cells. When these receptors interact with their corresponding ligands—PD-L1 (Programmed Death-Ligand 1) for PD-1 and B7 molecules for CTLA-4—they transmit signals that inhibit T-cell activation. In normal immune function, these checkpoints are crucial for maintaining self-tolerance and preventing immune cells from attacking healthy tissues. However, in certain conditions like cancer, tumor cells can exploit these pathways to evade the immune response. Cancer cells may express PD-L1, engaging with PD-1 on T cells and suppressing their activity. Similarly, CTLA-4 can dampen the immune response by inhibiting the early stages of T-cell activation.
One of the foremost challenges in developing T-cell-focused vaccines is navigating the remarkable diversity of pathogens, particularly those with high mutation rates like HIV. These pathogens continuously evolve, giving rise to an ever-expanding repertoire of genetic variants. This diversity poses a significant hurdle in selecting antigens that elicit effective T-cell responses. Traditional approaches targeting conserved regions of the pathogen aim to overcome this challenge. [50] The creation of technological platforms, including elements like antigen design, adjuvants, and delivery methods, is essential to eliciting responses from vaccinations on both the cellular and humoral levels. Live-attenuated vaccines (LAVs) are traditional vaccinations that have been altered to include microorganisms in a weakened form. LAVs demonstrate significantly heightened immunogenicity, capable of inducing both humoral and cellular immune responses. [51]
Bacillus Calmette-Guérin (BCG), created in the 1920s as the first T-cell-inducing vaccine against TB, is one illustrative example of a live-attenuated vaccine. BCG promotes the development of specific memory T cells, providing defense against the intracellular parasite infection brought on by Mycobacterium TB. [52] Recently, the mechanism of mycobacterium tuberculosis bacteria in cords remains translationally active despite antibiotic exposure and regrows rapidly upon cessation of treatment. [53].(1)
Humoral Immunity
The humoral immune response is a vital arm of the immune system orchestrated by B lymphocytes (B cells) and their antibody-mediated defenses. When confronted with pathogens, B cells recognize specific antigens and transform into plasma cells, prolifically producing antibodies. These antibodies, or immunoglobulins, play multifaceted roles in combating infections, including neutralizing pathogens, marking them for phagocytosis, activating the complement system, and facilitating agglutination. Memory B cells formed during this process retain a memory of encountered antigens, enabling a swift and robust response upon subsequent infections. The diverse classes of antibodies, such as IgM and IgG, serve distinct functions, contributing to both immediate and long-term immunity. (Figure 3)
Antibody-mediated humoral immunity protects against extracellular pathogens. It is common knowledge that the passive transport of maternal antibodies across the placenta protects newborns from a broad spectrum of pathogens. [54] The magnitude of the humoral immune response is correlated with disease severity. [55]
Antigen-presenting cells recognize a pathogen's infiltration into a tissue as an exogenous antigen. The peptide antigen is subsequently presented to T lymphocytes by these activated antigen-presenting cells via MHC class II in the draining lymph node. As CD4+ T cells become activated, cytokine production is increased. T follicular helper (Tfh) cells, a subgroup of CD4+ T cells, have been shown to increase antibody affinity for the antigen by encouraging B cell proliferation and humoral response maturation. (Figure 3)
B cells produce two waves of antibody responses. Short-lived plasma cells often develop without passing through a germinal center (GC) and have significant titers of subpar antibodies very quickly. To make high-quality antibodies, a second set of B cells enters the GC and undergoes somatic hypermutation and affinity maturation. Long-lived plasma cells produce antibodies in the bone marrow. A pool of memory B cells, the biological repository for top-notch antibody sequences, is the other product of the GC. These cells may quickly differentiate into plasma cells upon reinfection (or booster vaccination) to create fresh antibodies and seed fresh GCs to resume antibody affinity maturation. Memory B cells are very long-lived and may work with memory T cells to guard against illness, even while serum antibody titers viral vaccinations quickly fall. [56,57]
Upon detecting peptide antigens with MHC on antigen-presenting cells (APCs), CD4+ T helper (Th) cells undergo differentiation into various effector cells. The three significant subsets—Th1, Th2, and Th17—are pivotal in enhancing and coordinating antipathogen immunity. Th1 cells are particularly adept at eradicating specific intracellular pathogens by producing substantial interferon-gamma (IFN-) and interleukin-12 (IL-12). Cellular immunity is facilitated by these Th1 cytokines, which also promote the activation of monocytes and macrophages and the development of cytotoxic T lymphocytes (CTLs). The reduction of infectious illness severity is correlated with the predominance of Th1-dominant antiviral immunity. [58]
Eosinophils and mast cells are controlled by the interleukins (ILs) that Th2 cells produce, including IL-4, IL-5, IL-6, IL-10, and IL-11. Among these, IL-4, a Th2 cytokine, boosts B cells' humoral immune response and promotes the generation of antibodies. Notably, T follicular helper (Tfh) cells—not Th2 cells—are essential for developing and operating B cell responses. Tfh cells in secondary lymphoid organs' germinal centers produce IL-21 (GC), and GC Tfh cells also produce IL-4, which is crucial for the best possible B cell activation. As a result, the immunogenicity of the vaccination and humoral immunity are related to the stimulation of Tfh cells. Additionally, Th17 cells serve crucial roles in eradicating cellular pathogens at mucosal locations by generating IL-17A, -17F, and -22. [59,60] Th17 cytokines regulate neutrophils, and IL-17 controls interferon (IFN) production, essential for promoting the Th1 response. Th17 cells are necessary for maintaining mucosal immunity, which controls the translocation of microbes. Notably, Th17 cells impact the prevention of AIDS and the management of chronic HIV-1 infection.
Regulatory T cells (Tregs) constitute a subpopulation essential for immune tolerance. Immune tolerance depends on a subset called regulatory T cells (Tregs). Interleukin-10 (IL-10) produced in large quantities by these cells prevents T-cell activation and differentiation. A rise in antigen-specific Tregs compromises the effectiveness of vaccine immunity and pathogen control.
The polarization of the Thf response and the efficacy of vaccinations are affected by formulations, adjuvants, and delivery methods. Numerous technical platforms for vaccines that induce T-cell-mediated immunity have been developed. [61] Vaccines strategically leverage the humoral immune response by priming the system to generate targeted antibodies and memory cells, fostering a heightened defense against potential threats. This intricate orchestration effectively protects against a spectrum of pathogens, particularly those in the extracellular domain. The humoral immune response works synergistically with cellular immunity to safeguard the host organism comprehensively.
Neutralizing antibody (NAb) responses have been the main research topic on SARS-Cov-2 vaccines, but cellular immunity has received less attention. Despite this, there is evidence that T-cell responses, especially against viral variants that partly elude NAb detection, play a critical role in vaccination protection against severe COVID-19 illness.
Vaccine Types
When developing vaccines, it is crucial to consider important factors like immunogenicity, genomic structure, pathogenicity, epidemiology, target antigens, correlated immune protection, outbreak forecasting, and characteristics of the target population's cellular and humoral immune responses. Despite substantial efforts from the scientific community, many aspects related to vaccines remain unknown. [62] For example, local reactions at the injection site and adverse systemic reactions are observed following the administration of vaccines. Adjuvants are necessary alongside antigenic epitopes to induce robust immune responses. Live attenuated vaccines face challenges due to the potential for defective inactivation causing antibody-dependent enhancement (ADE). Vector-based vaccines, while capable of inducing neutralizing antibodies and cellular immune responses, may encounter anti-vector immunity as a challenge. [63]
Moreover, there are instances where antigens are only generated during the disease, making laboratory production impossible. Additionally, these methods do not apply to non-cultivable microorganisms. [64] The development of oral or nasal vaccinations is recommended to overcome these restrictions since these immunization methods are thought to be the most successful at generating potent immune responses at the mucosal level. [65]
Pathogens and cancer cells have developed various mechanisms to evade the host immune system, posing significant challenges to developing effective T-cell-focused vaccines and immunotherapies. For example, the variants of SARS-CoV-2 have evolved convergent strategies to remodel the host by modulating viral RNA and protein levels, altering viral and host protein phosphorylation, and rewiring virus-host protein-protein interactions. [66]
Understanding and countering these evasion mechanisms is a critical aspect of vaccine research. Pathogens like HIV and influenza undergo rapid antigenic variation, allowing them to escape recognition by host immune cells. This diversity challenges the development of vaccines targeting specific antigens. Strategies to address antigenic variation include targeting conserved regions, employing mosaic antigens, or using polyvalent vaccines incorporating multiple antigens or strains. [67]
Attenuated cell
Attenuated bacterial vaccines use live bacteria that have been weakened (attenuated) in a laboratory setting to the point where they can stimulate an immune response without causing the disease they are intended to prevent. The attenuation process involves modifying the bacteria to lose their ability to cause disease in a healthy host while retaining their ability to induce an immune response. Oral Polio Vaccine (OPV) is a milestone in the global fight against polio. The oral poliovirus vaccine (OPV) administration triggers the production of antibodies and the activation of T cells, leading to the development of immunological memory. This memory provides swift and effective protection upon subsequent exposure to the wild-type poliovirus. A similar approach is employed with Bacille Calmette-Guerin (BCG), an attenuated live vaccine derived from the bacteria causing bovine tuberculosis, Mycobacterium bovis. When administered subcutaneously or intradermally to newborns, BCG protects them from severe and disseminated manifestations of human tuberculosis. Developed around 100 years ago to combat Mycobacterium tuberculosis infection, BCG is currently being repurposed to investigate its potential to reduce morbidity and mortality associated with SARS-CoV-2 infection.
While effective in generating robust and long-lasting immunity, live attenuated vaccines are unsuitable for all pathogens, especially those that can revert to virulence. [33]
Protein-based vaccines
Protein-based vaccines, including subunit and recombinant protein vaccines, offer precision in antigen delivery and have played essential roles in combatting infectious diseases. [48] Protein-based vaccines represent another approach to eliciting T-cell immunity with precision in antigen delivery. These vaccines contain purified proteins or protein subunits from the pathogen of interest. Protein-based vaccines can induce targeted T-cell responses by presenting specific antigens to the immune system. Examples include subunit vaccines, which contain select components of the pathogen, and recombinant protein vaccines, where proteins are produced using genetic engineering techniques [68]. Protein-based vaccines have played essential roles in combatting infectious diseases such as hepatitis B and human papillomavirus (HPV). [69] They are known for their safety and efficacy, making them valuable tools in vaccine development.
Viral vector
Viral vectors and nucleic acid-based vaccines are used extensively to combat SARS-CoV-2. Both platforms are based on genetic engineering, which allows for the induction of Th1-biased vaccine immunity via intracellular expression of genetically engineered antigens. [70] Viral vector vaccines can induce humoral and cellular immunity specific to an antigen. [71] These systems allow for the delivery of antigen to cells and subsequent antigen-presenting cell (APC) uptake. Viral vectors have been approved for treating HIV-1 and other retroviral infections because they may stimulate potent cytotoxic T lymphocytes (CTLs). [72] Inactivated vaccines may sometimes suffer from weak immunogenicity. Adjuvants and biomaterials together enhance the immunogenicity of vaccinations. Immunogenicity is also influenced by the delivery method. Increased immunogenicity and local immunity are brought about by tissue-specific dosing. For quick responses to antigen re-exposure, tissue-resident memory T-cells (Trm) must be induced. [73] Vaccine distribution necessitates optimized administration routes and novel medical devices. [74,75]
Viral antigen
Viral antigens are typically inactivated trivalent or quadrivalent hemagglutinin (HA) subunits in most commercially available seasonal influenza vaccines. Based on surveillance data, these vaccines select strains twice a year, and their effectiveness relies on predicting epidemic strains. [76,77] T-cell activation that targets conserved epitopes, such as those on non-structural (NS) proteins, may provide defense against various influenza strains throughout the year. Multimeric/M-001, which has nine conserved B-cell, CD4+, and CD8+ T-cell epitopes from HA, nucleo-protein, and matrix protein 1 (M1), is one prospective candidate recombinant protein with this capability. Another candidate vaccine, Flu-v, includes conserved peptides derived from M1, M2, and the nucleoprotein. However, achieving a definitive and robust induction of conserved, cell-specific epitopes has not been accomplished thus far.
Nucleic acid
Vaccines based on nucleic acids use DNA/mRNA that has been genetically modified to elicit an immune response against antigens. These vaccines benefit from inducing cellular and humoral responses against specific epitopes. SARS-CoV-2 is being extensively combated using viral vectors and vaccines based on nucleic acids. Both platforms are built on genetic engineering, which allows for the intracellular production of genetically modified antigens and the generation of Th1-biased vaccination immunity.
Dendritic cell
Dendritic cell vaccines are designed to harness the unique antigen-presenting capabilities of dendritic cells (DCs), which are critical players in initiating immune responses. Dendritic cell vaccines showcase the intricate orchestration of antigen presentation, instilling robust T-cell responses. [74] The development of dendritic cell vaccines involves manipulating these cells to enhance their ability to recognize and attack specific targets, such as cancer cells. In this approach, DCs are loaded with antigens from the pathogen and then administered to the patient. DCs are highly efficient at presenting T-cell antigens, stimulating potent T-cell responses [52]. Cancer vaccines have explored this strategy, where DCs are loaded with tumor antigens to activate T-cell-mediated anti-tumor responses. [69] Dendritic cell vaccines hold promise for infectious disease and cancer immunotherapy and are an exciting avenue for research and development.
Adjuvants
Adjuvants are traditionally used with protein-based vaccines to augment their efficacy by boosting the immune response to antigens. [78] Tailoring adjuvants for T-cell-focused responses often involves incorporating specific immune-stimulating molecules. For example, Toll-like receptor (TLR) agonists, such as CpG oligonucleotides, mimic pathogen-associated molecular patterns (PAMPs) and can stimulate APCs to induce T-cell responses. [79] Developing synthetic adjuvants that precisely target T-cell activation pathways also holds promise in vaccine design. [80] These approaches aim to optimize adjuvants to induce strong and durable T-cell immunity. While adjuvants enhance vaccine efficacy, their safety and compatibility with other vaccine components are paramount. Rigorous testing and monitoring ensure that adjuvanted vaccines meet stringent safety standards. Adverse events, such as local reactions or systemic symptoms, are closely monitored during clinical trials and post-marketing surveillance. [81] Ensuring the safety of adjuvanted vaccines is essential for public confidence and vaccine acceptance.
Future directions in adjuvant development for T-cell-focused vaccines involve a deeper understanding of immune pathways and the rational design of adjuvants to modulate these pathways precisely. Developing personalized adjuvant strategies considering an individual's immunological profile and genetic background is an emerging concept. Additionally, exploring adjuvant combinations and novel adjuvant platforms will continue to advance the field, offering new tools to enhance T-cell-mediated immune responses.
Adjuvants that specifically target immune evasion mechanisms are under investigation. For example, toll-like receptor agonists can enhance antigen presentation and promote T-cell responses, overcoming the immune evasion tactics employed by specific pathogens [159]. These adjuvants can be incorporated into vaccine formulations to boost their efficacy.
Furthermore, several useful biomaterials have been created to enhance the immunogenicity of vaccinations. Scaffold-based vaccines combine pore-forming polymer gel matrices with immune-modulating compounds, such as adjuvants and cytokines, to concentrate and excite immune cells at the injection site. [82] Additionally, several antigen-conjugated functional materials have shown effective lymph node delivery. [83]
Nucleoside Vaccines
Nucleic acid-based vaccines use genetically altered DNA or mRNA to activate the immune system against specific antigens. These vaccines can trigger cellular and humoral responses directed to epitopes, a significant benefit. Although safe, DNA and nanoparticle-based vaccinations have lesser immunogenicity than conventional vaccines; their effectiveness requires appropriate adjuvants, immunization types, and optimization of antigenic epitope sequences to induce heightened immune responses. Existing vaccines, such as live attenuated or inactivated vaccines, follow established protocols, but novel mRNA, DNA, and RNA vaccines require time-consuming design processes. [84] The surface proteins and the RBD domains of viruses are promising targets for subunit vaccines. They efficiently prevent the illness because they trigger a higher immune response than entire virus particles, such as inactivated or attenuated viral vaccines. [85]
DNA Vaccines
In DNA vaccines, the DNA encodes pathogen-specific antigens. This DNA is injected into the body, taken up by cells, and used to synthesize proteins based on the genetic code within the plasmid. These proteins are recognized as foreign due to their amino acid sequences typical of bacteria or viruses. This recognition triggers immune responses when the antigens are processed by host cells and displayed on their surface. Alternatively, DNA can be encapsulated in protein to facilitate cell entry. If the capsid protein is incorporated into the DNA, the resulting vaccine may possess the potency of a live vaccine without the associated risks of reversion.
DNA vaccines represent an innovative approach to immunization, utilizing genetic material to trigger an immune response. Diverging from traditional vaccines that use inactivated or attenuated pathogens, DNA vaccines directly introduce DNA sequences encoding specific antigens into the body. These vaccines include DNA encoding viral proteins or peptides that stimulate the immune system. DNA vaccines have been created for the SARS-CoV-2 virus after being tested for their effectiveness against HIV, rabies, hepatitis B and C, influenza, and hepatitis B and C. In the case of SARS-CoV-2, the vaccines include nucleotide sequences for the MHC-1 and MHC-2 domains to stabilize the MHC-1-peptide complex. The DNA encodes the most antigenic T-cell epitope in the N protein. A multi-epitope DNA vaccine that encodes B cell epitopes found in the S and M proteins is also available. It encodes the spike protein of SARS-CoV-2 with an additional arginine-glycine-aspartic acid (RGD) sequence. [86] It is under inquiry whether attenuated viruses, such as MVA, adenovirus, and measles, are safe to utilize as vectors to transfer nucleotide sequences encoding protein-coding genes into a benign vector virus (such as the adenovirus that causes the common cold). However, this kind of vaccination is limited due to worries about reversion and instability that might result in the creation of more virulent variants. [87,88]
DNA vaccines provide various advantages, including cheap mobility, heat resistance, and safety. [89,90] In the 1990s, Wolff et al. demonstrated the production of foreign antigens by the intramuscular delivery of plasmid DNA, introducing the idea of DNA vaccinations. This procedure involves injecting the body with plasmid DNA that encodes pathogen-specific antigens. When the DNA is ingested by cells, it is carried into the cell's nucleus, where proteins are produced following the genetic code contained in the plasmid. These proteins are identified as being foreign because they include portions of amino acid sequences that are typical of bacteria or viruses. The immune system is notified after being digested by host cells and shown on their surface, which triggers further immunological reactions. [91] Alternatively, DNA may be covered with a protein to make cell entrance easier. The resultant vaccine might have the potency of a live vaccination without the dangers of reversion if this capsid protein is integrated into the DNA.
DNA vaccines, while promising, often elicit less robust immune responses than other vaccine platforms due to limitations in vaccine delivery technology. As a result, they are frequently assessed using novel injection systems to enhance their efficacy. The first DNA vaccine approved in India for emergency use was administered intradermally using a jet injector. [92,93,94] DNA vaccination can induce high-specific antibody titers and cellular immune responses against rapidly mutating pathogens like HIV. In non-human primates infected with HIV, DNA vaccines targeting envelope proteins have demonstrated effectiveness with minimal DNA and quasi-homologous antigens. The co-polymer 704 is considered a potentially ideal synthetic vector for future DNA-based immunotherapeutic and infectious disease vaccines. [95] Furthermore, the delivery of mRNA-encoding DNA sensors as "adjuvants" holds promise for maximizing vaccine efficacy while minimizing side effects. Adjuvant development involving innovative formulations is anticipated to enhance T-cell responses while maintaining safety. [96] In vaccine evaluation and optimization, immune correlates of protection play a crucial role, serving as guiding beacons to enhance our understanding and refine vaccine strategies. [97,98]
]DNA-based vaccinations provide several benefits: cost, mobility, heat resistance, and safety. When Wolff et al. showed that plasmid DNA administered intramuscularly may cause the expression of foreign antigens, the idea of DNA vaccinations was first proposed. Transport into the cell's nucleus is needed for plasmid DNA antigens to manifest themselves.
Despite these benefits, DNA vaccines tend to produce less powerful immune responses due to vaccine delivery technology limitations than other vaccination platforms. DNA vaccines are often tested using cutting-edge injection techniques to overcome this. A jet injector was used to give the first DNA vaccination that India has authorized for use in an emergency intradermally.
mRNA Vaccines
mRNA vaccines have recently gained significant attention and success. These vaccines utilize a small genetic material—mRNA—to instruct cells to produce a specific antigen. mRNA vaccines offer several advantages, including their rapid development and adaptability to emerging pathogens. [72] The two SARS-CoV mRNA vaccines induced high levels of both neutralizing antibodies and Th1-biased SARS-CoV-2-specific T-cell responses, [99] in addition to an increase in Tfh cells in draining lymph nodes, which may provide long-lasting protective antibody responses, according to the results of representative mRNA vaccines and T-cell-mediated immunity. [100]
Modifications to mRNA can increase the stability of mRNA vaccines, ranging from capping to polyA tail, uridine replacement, and lipid nanoparticle formulations to improve the efficacy of mRNA vaccines. [101]
Various adjuvant types are employed to enhance T-cell-focused immune responses. One well-known adjuvant is alum, used in numerous vaccines, including hepatitis B and human papillomavirus. Alum primarily stimulates Th2-type antibody responses, making it less suitable for T-cell-focused vaccines. [102] In contrast, newer adjuvants like AS01 and MF59 have shown efficacy in enhancing both antibody and T-cell responses. AS01, used in the RTS, S malaria vaccine, contains a liposome-based adjuvant and a Toll-like receptor agonist, while MF59, used in seasonal influenza vaccines, is an oil-in-water emulsion. [103,104]
Longer-Term mRNA Vaccines
Longer-term vaccines aim to confer a more persistent defense than traditional vaccines; the key to successful vaccine development is the deliberate induction of T-cell-mediated immunity and humoral immunity, regularly assessed by serological tests as the seroconversion rate of antigen-specific antibodies after vaccination. In contrast, T-cell responses to antigen-stimulated peripheral blood mononuclear cells (PBMCs) are assessed. Standard methods for determining cytokine production from PBMCs include intracellular cytokine labeling and enzyme-linked immune spots (ELISPOT). Analyzing the production of cytokines by T cells in response to repeated antigen exposure is necessary to comprehend the antipathogenic effectiveness of vaccinations.
Specificity
Epitope-based vaccines constitute a specialized category designed to target specific antigenic regions, known as epitopes, on the surface of pathogens. Unlike traditional vaccines that may use weakened or inactivated forms of the entire pathogen, epitope-based vaccines focus on the fundamental molecular components that elicit an immune response. Epitope-based vaccines have shown promise in combating various infectious diseases, including viral infections and certain types of cancer. Their precision and adaptability make them valuable tools for developing effective and targeted vaccination strategies. Notably, CD4+ and CD8+ T cells, integral immune system components, recognize these epitopes.
To predict the epitopes, there are different databases and approaches. After the prediction of epitopes, Shannon entropy [105] , and TOPSIS [106] scoring are run and plotted through antigenicity and conservity successfully to identify the best epitopes with two factors. The subsequent and final step is to select the optimal epitopes by plotting to provide a road map for comprehending antigens and epitope modification in variants. Additionally, this could aid in selecting epitopes based on various criteria. This approach is an open-source, multi-criteria platform for selecting the top epitopes. The anticipated CTL epitopes from the N and S proteins, LSPRWYFYY and DLSPRWYFYY, as well as VVFLHVTYV, GVVFLHVTY, VRFPNITNL, and PYRVVVVLSF, are highly conserved and immunogenic, making them ideal candidates for vaccine development. One may create a vaccination with numerous epitopes using an AAY linker between CTL epitopes. Additionally, linkers (like GPGPG) may be used to connect HTL epitopes predicted by WPQIA-QFAPSASAFF, QIAQFAPSASAFFGM, and LBL epitopes predicted by AGLPYGANK to CTL epitopes. More vaccine design possibilities exist between anticipated epitopes and IEDB epitopes that can be added to this vaccine. [107]
These novel variations are characterized by several spike (S) protein alterations that improve replication efficiency, immunological evasion, and transmissibility. The spike sequence is, in essence, a "moving target" because of the continual high frequency of mutations in various locations. The second generation of vaccine candidates supports swapping or co-expressing the spike gene with the nucleocapsid (N) gene. The internal N gene is more stable and conserved than the external spike gene. Due to their significance to the viral life cycle, including RNA packaging, replication, and transcription, fewer mutations have occurred. The non-surface N protein is a possible vaccine target for cytotoxic CD8+ T lymphocytes because of its conserved sequence linked to efficient viral clearance and less severe illness. [108] Indeed, strong T-cell responses to nucleocapsid have been identified.
The T-cells recognize short, 8– to 15–amino acid linear peptides that are not restricted to the RBD and NTD domains of the spike, where most mutations occur. With >80% of T-cell epitopes conserved across variants, T-cell responses against variants such as Omicron remain largely intact). Moreover, suppose T-cell epitope escape occurs. In this case, differences in HLA-peptide presentation indicate that a mutation causing T-cell immunity evasion in one individual is unlikely to do so in another. In general, emerging viral variants substantially affect antibody neutralization but have negligible effect on T-cell responses. Table 1 lists the conserved epitopes of SARS-CoV-2 [109]
Reverse Vaccinology
Reverse Vaccinology (RV) is a cutting-edge approach that revolutionizes vaccine design by allowing the precise determination of antigens, focusing on epitopes as a critical parameter. The most restrictive step in conventional vaccinology is precisely determining antigen number. Computational vaccinology uses high-performance information analysis and machine-learning tools to elucidate essential antigens and antigenic potential for discovering optimal vaccines by observing the valuable components and units in pathogens and pathogenesis fundamental organic processes (Figure 4). [110,111]
Computational tools and algorithms analyze protein structures to predict potential epitopes, providing valuable insights into their abundance and distribution. Immuno-informatics is a subfield of Bioinformatics that develops algorithms for validating potential B cell epitopes (BCEs) and T cell epitopes (TCEs). Examining the affinity of antigenic peptides for binding to MHC particles can pose a formidable obstacle to predicting epitopes. Quantifying B/T cell epitopes contributes to a comprehensive understanding of the immune response, guiding the selection of antigenic components for vaccine development. By combining computational predictions with experimental validation, researchers can refine their knowledge of epitope characteristics and tailor vaccines to elicit targeted and robust immune responses. Integrating quantification methods enhances the precision and effectiveness of epitope-based vaccine strategies.
B-cell Epitopes
B cell epitopes are specific antigenic regions on proteins or other molecules that can be recognized by B cells and subsequently trigger the production of antibodies. These epitopes are essential for humoral immunity, where antibodies are central in neutralizing pathogens. B cell epitopes can be located on the surface of antigens and are often characterized by their ability to bind to the B cell receptor (BCR). For example, researchers have used computational methods such as the BepiPred algorithm [112] to predict potential B cell epitopes on viral proteins, aiding in developing vaccines against infectious diseases.
The nature and type of BCEs determine in-silico-BCEs prognostication since B cell lymphocytes are drawn to both continuous (linear) and discontinuous (conformational, CBCEs) BCE epitopes. [113,114] A few key guidelines for precise in-silico-BCEs mapping are as follows:
- disposal of short epitopes; [115]
- analysis of candidate antigens' fundamental properties (such as hydrophilicity, flexibility, surface exposure, and solvent availability); [116]
- use of multimethod BCEs prediction strategies; [117]
- comparison of web-based BCEs prediction tool results with results from molecular interaction strategies like molecular docking and molecular dynamics simulations. [118]
It is unknown how long antigenic peptides must be to bind to the paratope site of antibodies, even though nearly 90% of BCEs are CBCEs (mostly). This is often a CBCEs-related issue because it results in less accurate (e.g., false-negative and false-positive) predictions of CBCEs.
T-cell Epitopes
T Cell epitope prediction strategies fall into two types, like BCE: direct and indirect. While indirect approaches locate MHC binders, direct methods rely on the structural and sequential study of TCEs. Indirect procedures are often used for TCE prediction because of their high accuracy and specificity. The indirect methods are MHC-I covers (CD8+ TCEs for processing endogenous antigens) and MHC-II binders (CD4+ helper TCEs for processing external antigens). [119]
Several strategies have been employed to address antigen selection in the context of T-cell-focused vaccines. One approach is to target conserved regions of the pathogen's genome, regions less likely to undergo mutations without compromising the pathogen's viability. Another strategy is to focus on epitopes shared among different strains or serotypes of the pathogen. By selecting epitopes that are common to multiple variants, vaccines can induce broader and more cross-protective T-cell responses. [120] Advancements in immunoinformatic and computational biology have greatly facilitated antigen selection. These tools enable researchers to predict potential T-cell epitopes within pathogen genomes, streamlining the process of antigen design. [10] By identifying conserved epitopes likely to be recognized by a broad range of T-cells, immunoinformatic contributes to the rational design of T-cell-focused vaccines. Structural biology and bioinformatics tools also help visualize antigen-TCR interactions, providing insights into epitope-TCR binding affinities. [12]
Identification of antigenic epitopes is the most crucial step in vaccine development. For example, the spike protein (S) of SARS-CoV-2 has been a promising target for developing a vaccine. Still, the choice between the receptor-binding domain (RBD) and the entire protein remains challenging. [121,122] Concern has repeatedly been raised about the advent of virus types immune to current antiviral immunity or vaccinations. [123] Although SARS-CoV-2 vaccinations provide severe illness protection as a booster, the developing Omicron SARS-CoV-2 variant evades vaccination-induced anti-spike neutralizing antibodies. [124] This variation is also highly resistant to therapeutic monoclonal antibodies. The use of widely neutralizing antibodies during immunization is effective against all variations. [125] Immunity mediated by T cells is essential. Most current vaccines target the spike, M, N, and other nonstructural (NS) proteins, [126] including many immunodominant T-cell epitopes (Figure 4). [127]
Figure 1.
Viral proteins are subject to vaccination selection.
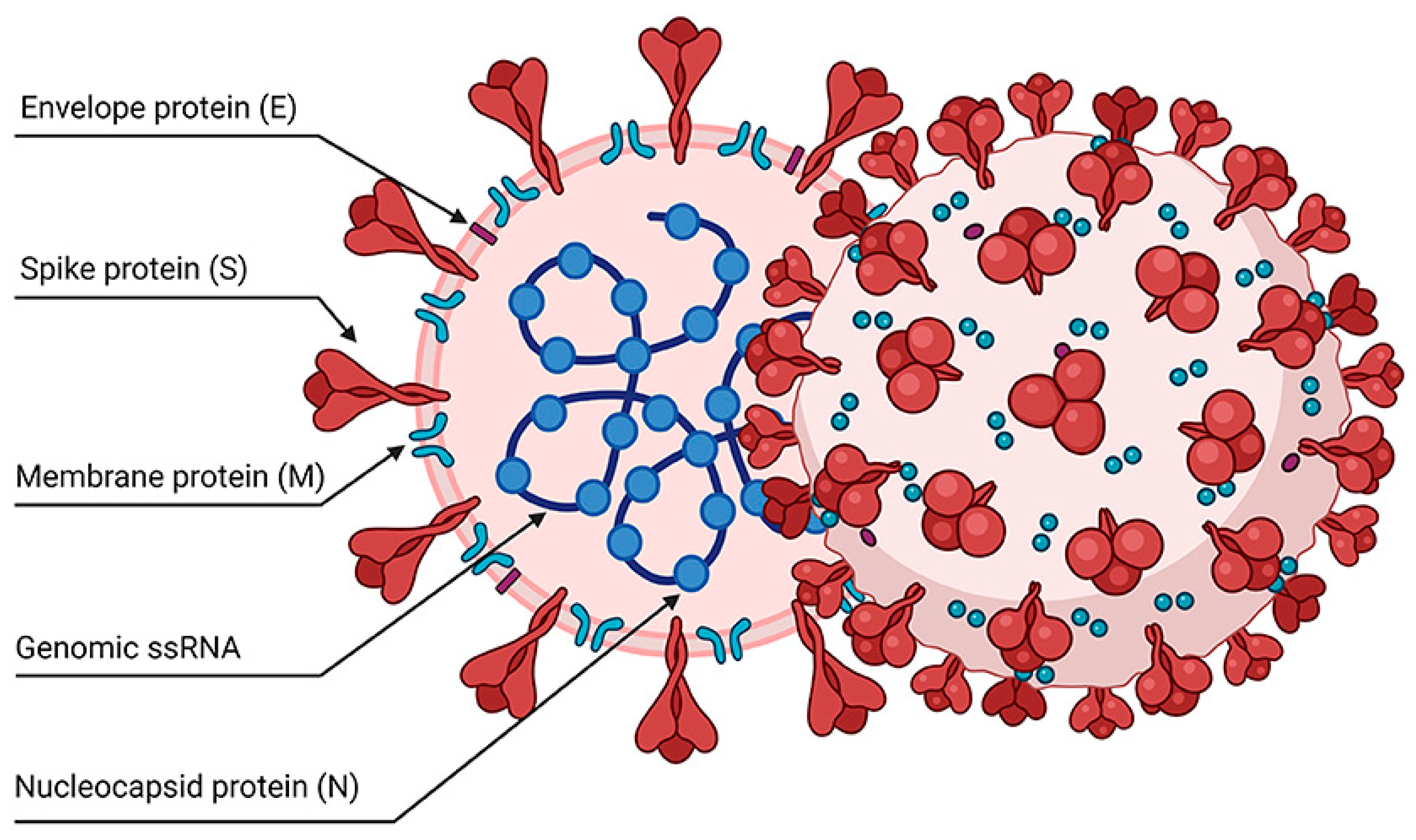
Targeting the immunodominant epitopes of pathogens is also essential for developing cross-reactive, immunogenic vaccines. For example, in COVID-19 patients, CD8+ T cells specific for conserved epitopes correlate with milder disease [128]. Bioinformatics and analyses of peptide HLA complexes have revealed that ORF1ab-derived epitopes have the potential to serve as optimal vaccine antigens [129]. These hypothesized epitopes are immunodominant and highly conserved.
T cell epitopes are specific peptide sequences within antigens that can be recognized by T cells, a critical component of the cellular immune response. T cell epitopes are presented by major histocompatibility complex (MHC) molecules, and two main types of T cells, CD4+ T cells and CD8+ T cells recognize these epitopes through their T cell receptors (TCRs). CD4+ T cells interact with epitopes presented on MHC class II molecules, helping coordinate immune responses, while CD8+ T cells target cells displaying epitopes on MHC class I molecules for destruction. Various tools, such as epitope prediction algorithms, have been developed to understand their role in immunity better, facilitating vaccine design and immunotherapy development (Table 1).
Another interesting aspect of the exceptionally complex SARS-CoV-2 virus is that it exhibits a composition of structural proteins (S, M, H, and E) and non-structural proteins (NSPs), both of which play pivotal roles in its infectivity and potential involvement in chronic ailments such as cancer and neurodegenerative disorders. [152] The interaction between SARS-CoV-2 proteins, particularly the spike protein, and the angiotensin-converting enzyme 2 (ACE2) receptor is identified as a critical factor in the infection process. Certain compounds and drugs, such as polyphenols and specific inhibitors, demonstrate inhibitory effects on these protein interactions. The critical epitopes of the SARS-CoV-2 virus encompass structural and non-structural proteins (NSPs), offering promising candidates for developing vaccines and treatments. Numerous studies have shown that the NSP3 protein is a promising target for developing an effective COVID-19 vaccine. This is attributed to its possessing epitopes for both MHC and T cells, along with linear B cell epitopes translated in specific locations and functional domains of the protein. [153,154]
Despite significant progress, challenges persist in antigen selection and design. Achieving broad and long-lasting T-cell responses remains an ongoing goal, particularly for pathogens with high mutation rates.
Integrating multi-omics data, such as genomics, transcriptomics, and proteomics, will likely enhance our ability to predict and select antigens that can effectively engage T-cell responses [155]. Moreover, advances in structural vaccinology, epitope-based vaccine design, and immunoinformatics will continue to drive innovation in T-cell-focused vaccines. These approaches offer promise in infectious disease control and personalized cancer treatment, marking a transformative era in vaccine development.
Numerous clinical outcome investigations on infectious diseases have shown that cellular and humoral immune responses trigger the most potent antipathogen immunity. The mechanism of action of effective vaccines is clarified by understanding the antipathogenic immune response. T-cell responses are often assessed during the first stages of therapeutic studies. The characteristics of T-cells produced because of immunization show the effectiveness of these vaccinations. Which kind of immunity is preferred—humoral or cell-mediated—is determined by Th-cell differentiation. The possibility of ADEs is decreased by cross-reactive CTLs. Memory cell performance relates to the length of vaccination protection. Preclinical and clinical vaccine studies should define T-cell function-based criteria.
Individual differences exist in vaccine effectiveness and immune response, with older persons showing delayed and decreased humoral and cellular responses compared to younger ones. Effector T cells with a limited lifetime are more likely to differentiate as we age than Tfh and memory cells. [156]
In summary, leveraging T-cell immunity for vaccines and immunotherapies has enormous promise for treating malignancies and infectious disorders. Researchers and healthcare professionals are at the forefront of improving T-cell-focused therapies by tackling antigenic diversity, using immune checkpoint modulation, utilizing adjuvant-based techniques, and adopting personalized approaches.
. While T-cell-focused vaccines hold great promise, they are not without their challenges. Antigen selection for T-cell vaccines requires careful consideration, as it involves identifying conserved epitopes shared among diverse strains of pathogens. The rapid mutation rates of certain viruses, such as HIV, pose a formidable obstacle, necessitating innovative strategies to elicit broad and durable T-cell responses. [72] Additionally, understanding the balance between protective and pathological immune responses, as excessive T-cell activation can lead to immunopathology.
Mutations
Since antibodies and T-cells recognize viruses and contribute to protection by different mechanisms, the impact of viral mutations on immune escape is distinct. NAbs identify conformational epitopes on viral proteins and typically mediate their effects by inhibiting the interaction between a viral coat protein and the host cell entry receptor. In the case of SARS-CoV-2, NAbs bind to the receptor binding domain (RBD) and N-terminal domain (NTD) of the spike protein, preventing its interaction with the host receptor, angiotensin-converting enzyme 2 (ACE-2). Mutations in the RBD and NTD of the spike can significantly affect antibody binding. Immune pressure from antibodies is likely driving the evolution of the spike protein, resulting in incomplete neutralization of new viral variants by neutralizing antibodies (NAbs) induced by vaccination or infection. Multiple therapeutic monoclonal antibodies have similarly lost efficacy against current SARSCoV-2 variants.
The evolving mutations may result in structural alterations in essential proteins involved in the pathogenesis and transmission of the virus. [157,158,159] This indicates that the vaccine target must be carefully selected concerning acceptable antigenicity and adequate conservatism, independent of the vaccine design technique. So far, several techniques have been investigated to detect different mutations and variations. [160,161,162] Only eight of the examined IDRs of the SARS-CoV-2 S, M, N, and E proteins demonstrate adequate conservatism, indicating that most IDRs have mutated. Significant alterations have been found in the S, M, and E proteins of SARS-CoV-2 and the HTL and T lymphocyte epitopes.
Finding mutation alone is insufficient to identify the best epitopes. Their conservation can be quantified using the TOPSIS method. Using this algorithm and a plot of their immunogenicity properties, the optimal epitopes are chosen for further laboratory analysis. Over time, almost all epitopes have changed, making the immune response to SARS-CoV-2 difficult. This may also account for the emergence of new, riskier varieties or volatile chemical compounds. Due to IDR mutation, the antibody neutralization has also altered. [163]
Targeted Vaccine Delivery
Targeted vaccine delivery is the precise and controlled administration of vaccines to specific cells, tissues, or organs in the body. The goal is to enhance the immune response and optimize vaccine efficacy while minimizing side effects. This approach involves the design of delivery systems that can selectively target immune cells or antigen-presenting cells, such as dendritic cells, which play a crucial role in initiating immune responses. Most licensed vaccines are given intramuscularly. The antigen delivery pathway affects immunogenicity because tissue-resident immune cells are present. This implies that a new delivery method may improve the immunological qualities of a vaccination. [164] Creating Trm cells (Tissue-resident memory) is crucial since it permits a quick reaction upon re-exposure to the antigen. As a result, the distribution method for vaccines and the control of local inflammation must be considered in innovative medical devices and biomaterials.
Intradermal (ID) administration of seasonal influenza vaccines increases immunogenicity and provides dose-sparing effects [165]. ID delivery activates local immune cells, quickly improving humoral and cellular responses. The ID approach is sometimes employed for nucleic acid-based DNA and mRNA vaccines with innovative medical devices. A significant antigen-specific Tfh cell response is induced by ID vaccination with mRNA-LNPs encoding several viral surface antigens associated with long-lasting/high-affinity neutralizing antibodies and persistent protection. [166] Local and systemic mucosal immunity may be induced with intranasal delivery. [167] The production of IgA on the mucosal surface by mucosal immunity stops respiratory infections from entering the body. The use of a non-invasive, needle-free nasal immunization method is also useful. Additionally, SARS-CoV-2 vaccinations are given orally or nasally, and several trials have shown encouraging outcomes.
Multi-target Vaccines
The development of combination mRNA vaccines is in its early stages, such as with the co-circulation of influenza and SARS-CoV-2, a combined respiratory vaccine targeting both COVID-19 and the flu is an excellent target as these vaccines enter phase 3 trials [168,169]. The most common influenza vaccine in the U.S. is quadrivalent; this means the combined influenza/COVID-19 vaccine would also likely need to be quadrivalent or trivalent, making these vaccines more complicated. Beyond infectious diseases, there's growing interest in therapeutic cancer vaccines, utilizing the specificity of mRNA to target an array of tumor-specific antigens [170].
One benefit of peptide-based vaccinations over conventional whole-protein vaccines is combining numerous peptides into a single (multivalent) vaccine. This allows for maximal coverage so that one or more epitopes will trigger an efficient immune response. Repeated changes perhaps hamper the development of efficient vaccinations in the viral genome. Therefore, the key to resolving this ubiquitous issue is the development of a broadly protective vaccination against the whole viral family. [171]
Moreover, specific serologic and clinical studies must be conducted to identify high-risk populations. [172]
Combination therapies that simultaneously target multiple evasion mechanisms hold promise. Combining immune checkpoint inhibitors with therapeutic vaccines or adoptive T-cell transfer therapies can enhance cancer immunotherapy's overall anti-tumor immune response. [173] Combinatorial approaches are being explored for infectious diseases as well.
Cancer Vaccines
Cancer vaccines represent a promising frontier in oncology, aiming to harness the body's immune system to recognize and eliminate cancer cells. Unlike traditional vaccines that prevent infectious diseases, cancer vaccines are designed to stimulate an immune response against specific tumor-associated antigens. These antigens are unique or overexpressed proteins found on cancer cells. There are two main types of cancer vaccines: preventive and therapeutic. Preventive cancer vaccines aim to prevent specific infections that can lead to cancer, such as the human papillomavirus (HPV) vaccine for cervical cancer. On the other hand, therapeutic cancer vaccines are developed to treat existing cancers by boosting the patient's immune response against tumor cells. [174,175]
Cancer vaccines function by introducing tumor-specific antigens into the body, triggering an immune response targeted at cancer cells. One of the advantages of using mRNA is its capacity to be tailored for individual patients, allowing for the targeting of unique tumor mutations. This has been applied to mRNA-based vaccines targeting prostate-specific antigens, showcasing their potential for triggering immune responses against prostate cancer [2,176]. The adaptability and specificity of mRNA make it an attractive platform for developing therapeutic cancer vaccines. As this field continues to evolve, mRNA cancer vaccines promise to revolutionize oncological treatment, providing personalized therapies tailored to each patient's unique tumor profile.
The advancements in mRNA cancer vaccine research, beyond their individualized potential, also encompass efforts to target commonly shared tumor antigens across a patient population. This would facilitate the creation of "off-the-shelf" mRNA vaccines that can be mass-produced and widely distributed, similar to vaccines for infectious diseases. A pivotal study wherein dendritic cells pulsed with tumor mRNA were shown to induce potent antitumor immune responses [177]. Such research establishes a foundation for developing broadly applicable mRNA cancer vaccines.
Moreover, the combination of mRNA vaccines with other immunotherapies is also being actively explored. For instance, integrating mRNA vaccines with checkpoint inhibitors might amplify the immune system's ability to recognize and destroy tumor cells [178]. This synergy could potentially overcome some of the challenges faced in cancer treatments, such as immune evasion by tumor cells.
Furthermore, the delivery method of mRNA cancer vaccines is vital to their efficacy. Nanoparticle carriers, lipid nanoparticles, and other novel delivery systems are under investigation to ensure optimal mRNA stability, cellular uptake, and translation [179]. Researchers aim to maximize the immune response against tumor antigens by improving delivery efficiency. [180]
It's also important to consider the challenges ahead. The tumor microenvironment is known for its immunosuppressive nature, which can limit the effectiveness of immunotherapies, including mRNA vaccines. Comprehensive understanding and strategies to counteract these suppressive mechanisms will be pivotal in successfully applying mRNA cancer vaccines.
As research continues to expand in mRNA cancer vaccines, the potential for combination therapies is another dimension under exploration. The rationale is to employ mRNA vaccines alongside other established cancer treatments, such as radiation, chemotherapy, or targeted therapies. By doing so, the goal is to capitalize on the strengths of each treatment modality while potentially mitigating their respective limitations.
For instance, radiation therapy induces immunogenic cell death, releasing tumor antigens and making the tumor environment more receptive to immune system attacks. This can be synergistically combined with mRNA vaccines to enhance antitumor immunity [181]. Similarly, certain chemotherapeutic agents can modify the tumor microenvironment, making it more amenable to immune cell infiltration and action. Pairing these agents with mRNA vaccines might thus bolster the overall immune response against cancer cells [182].
Beyond the immediate science of mRNA cancer vaccines, the infrastructure for manufacturing, distribution, and administration plays a crucial role in ensuring their successful deployment. As evidenced by the COVID-19 pandemic, the large-scale production of mRNA vaccines requires sophisticated facilities with stringent quality controls. This would need considerable investments and collaboration between public and private sectors, especially if mRNA cancer vaccines become a mainstay of oncological care [183].
Furthermore, with any innovative treatment modality, cost becomes a factor. Ensuring that mRNA cancer vaccines are accessible and affordable will be a significant challenge, requiring thoughtful policy decisions, possible subsidy programs, and innovative pricing models by pharmaceutical companies [184].
Various strategies are employed in cancer vaccine development. Peptide-based vaccines use short fragments of proteins (peptides) found in cancer cells to trigger an immune response. DNA and RNA vaccines introduce genetic material into cells to produce tumor-associated antigens, promoting an immune response. Whole-cell vaccines use inactivated cancer cells or parts of cells to stimulate immune recognition. Checkpoint inhibitors, a type of immunotherapy, complement cancer vaccines by blocking specific proteins that inhibit immune responses. The combination of cancer vaccines and immunotherapies is a promising avenue for enhancing the effectiveness of cancer treatments. [185]
In the context of cancer vaccines, neoantigens have gained prominence. Neoantigens are unique peptides generated from mutations in tumor cells. Because these mutations are absent in healthy tissues, neoantigens offer targets for T-cell-mediated immune responses. [186] Personalized cancer vaccines are designed to elicit T-cell responses against a patient's unique set of neoantigens, holding great potential for precision medicine in oncology. [187]
Additionally, strategies for selecting neoantigens in cancer vaccines must consider the heterogeneity of tumors and the dynamic nature of tumor evolution.
Other Autoimmune Disorder Vaccines
Perhaps the largest impact that the mRNA vaccine will make in the future is to prevent and treat autoimmune disorders. Studies from the SARS-COv-2 virus show that it triggers intracellular signaling pathways, including hypoxia-inducible factor-1 (HIF-1), neuropilin-1 (NRP-1), CD147, and Eph receptors, suggesting potential implications for diseases like Alzheimer's, epilepsy, various cancers, and cardiovascular conditions. [188]
Intellectual Property
The field of T-cell vaccines is characterized by a diverse range of patents covering various critical aspects:
1. Antigen Selection and Design: Patents are granted for identifying and designing novel antigens or epitopes that target T-cell-based vaccines. These antigens can be derived from infectious agents like viruses or bacteria, including cancer-specific antigens known as neoantigens.
2. Vaccine Formulations: Patents encompass the formulation of T-cell vaccines involving the integration of adjuvants, delivery systems, and other components to enhance T-cell responses. These formulations contribute to the effectiveness and safety of the vaccines.
3. Methods of Production: Patents protect innovative methods for producing T-cell vaccines. This includes traditional manufacturing processes and cutting-edge techniques, such as mRNA-based vaccine production, reflecting advancements in vaccine technology.
4. Adjuvants and Immunomodulators: Patents cover adjuvants and immunomodulators designed to augment T-cell responses when used with vaccines. These components enhance the immune system's ability to recognize and respond to antigens.
5. Therapeutic Applications: Patents extend to the therapeutic use of T-cell vaccines. These applications may involve T-cell vaccines to treat various conditions, including infectious diseases, cancer, and autoimmune disorders. The patents outline the specific therapeutic applications and methods for achieving therapeutic effects.
These patents advance T-cell vaccine technologies, fostering innovation in antigen discovery, formulation strategies, production methods, and therapeutic applications for addressing various health challenges.
10. Conclusions
Scientific history is full of transformation of early thoughts and theories leading to remarkable paradigm shifts; the mRNA technology, validated by the SARS-COv-2 virus, is one such example because of its simplicity, ingenuity, and the prospect of leaving the body take care of itself; instead of a bioreactor vessel, we are now getting ready to ride the ribosome trail. The era of perpetual vaccines is not far; neither is the possibility of combining multiple antigens in a single vaccine that will enhance this healthcare instrument's affordability. While we have understood the workings of the immune system well, it is now that we can exploit it to provide solutions to the problems it causes, such as autoimmune disorders, including cancer. I foresee investments rising in this field and anticipate the regulator agencies to create pathways for “biosimilar” mRNA therapies, though this option is unavailable. Extending the use of anti-infective vaccines towards resolving the dilemma of hundreds of untreatable diseases is the highlight of this breakthrough, made possible by the unfortunate incidence of COVID-19.
Funding
No funding was received for the preparation of the manuscript.
Conflicts of Interest
Authors declare no conflict of interest. The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Jackson, N.A.C., Kester, K.E., Casimiro, D., Gurunathan, S., & DeRosa, F. (2020). The promise of mRNA vaccines: a biotech and industrial perspective. NPJ Vaccines, 5(1), 11. [CrossRef]
- Vivona S, Gardy JL, Ramachandran S, Brinkman FS, Raghava GP, Flower DR, Filippini F. Computer-aided biotechnology: from immuno-informatics to reverse vaccinology. Trends Biotechnol. 2008 Apr;26(4):190-200. [CrossRef]
- Pardi, N., Hogan, M.J., Porter, F.W., & Weissman, D. (2018). mRNA vaccines—a new era in vaccinology. Nature Reviews Drug Discovery, 17(4), 261-279. [CrossRef]
- Ewer, K.J., Lambe, T., Rollier, C.S., Spencer, A.J., Hill, A.V., & Dorrell, L. (2016). Viral vectors as vaccine platforms: from immunogenicity to impact. Current Opinion in Immunology, 41, 47-54. [CrossRef]
- Crommelin, D.J.A., Anchordoquy, T.J., Volkin, D.B., Jiskoot, W., & Mastrobattista, E. (2021). Addressing the Cold Reality of mRNA Vaccine Stability. Journal of Pharmaceutical Sciences, 110(3), 997-1001. Authors. [CrossRef]
- Kowalski, P.S., Rudra, A., Miao, L., & Anderson, D.G. (2019). Delivering the messenger: advances in technologies for therapeutic mRNA delivery. Molecular Therapy, 27(4), 710-728. [CrossRef]
- Moderna. (2021). Moderna’s COVID-19 Vaccine Candidate Meets its Primary Efficacy Endpoint in the First Interim Analysis of the Phase 3 COVE Study. [Press release].
- Freyn, A.W., Ramos da Silva, J., Rosado, V.C., Bliss, C.M., Pine, M., Mui, B.L., Sahin, U. (2020). A multi-targeting, nucleoside-modified mRNA influenza virus vaccine provides broad protection in mice. Molecular Therapy, 28(7), 1569-1584. [CrossRef]
- Polack, F.P., Thomas, S.J., Kitchin, N., Absalon, J., Gurtman, A., Lockhart, S., Perez, J.L. (2020). Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. New England Journal of Medicine, 383(27), 2603-2615. [CrossRef]
- Dolgin, E. (2021). The tangled history of mRNA vaccines. Nature, 597(7874), 318-324. [CrossRef]
- Alameh, M.G., Weissman, D., & Pardi, N. (2021). Messenger RNA-Based Vaccines Against Infectious Diseases. Current Topics in Microbiology and Immunology, 429, 1-40. [CrossRef]
- Vaccine Tracker. https://covid19.trackvaccines.org (accessed on 1 September 2023).
- WHO Covid-19 Vaccine Landscape. https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 1 September 2023).
- WHO Covid Dashboard. https://covid19.who.int (accessed on 1 September 2023).
- https://clinicaltrials.gov/search?cond=SARS-CoV2&intr=Vaccine (accessed on 1 September 2023).
- https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 1 September 2023).
- Nobel Prize 2023 Medicine or Physiology. https://www.nobelprize.org/prizes/medicine/2023/press-release/ (accessed on 12 October 2023).
- Sahin, U., & Türeci, Ö. (2018). Personalized vaccines for cancer immunotherapy. Science, 359(6382), 1355-1360. [CrossRef]
- Pardi, N., Tuyishime, S., Muramatsu, H., Kariko, K., & Weissman, D. (2015). Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. Journal of Controlled Release, 217, 345-351. [CrossRef]
- Rojas LA, Sethna Z, Soares KC, Olcese C, Pang N, Patterson E, Lihm J, Ceglia N, Guasp P, Chu A, et al., Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature. 2023 Jun;618(7963):144-150. [CrossRef]
- Holtkamp S, Kreiter S, Selmi A, Simon P, Koslowski M, Huber C, Türeci O, Sahin U. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood. 2006 Dec 15;108(13):4009-17. [CrossRef]
- Barbier AJ, Jiang AY, Zhang P, Wooster R, Anderson DG. The clinical progress of mRNA vaccines and immunotherapies. Nat Biotechnol. 2022 Jun;40(6):840-854. [CrossRef]
- Liu C, Shi Q, Huang X, Koo S, Kong N, Tao W. mRNA-based cancer therapeutics. Nat Rev Cancer. 2023 Aug;23(8):526-543. [CrossRef]
- Pubmed Search. https://pubmed.ncbi.nlm.nih.gov/?term=mrna+vaccine&filter=datesearch.y_5&sort=pubdate (accessed on 1 September 2023).
- Rajpurkar P, Chen E, Banerjee O, Topol EJ. AI in health and medicine. Nat Med. 2022 Jan;28(1):31-38. [CrossRef]
- Brenden K. Petersen, Jiachen Yang, Will S. Grathwohl, Chase Cockrell, Claudio Santiago, Gary An, and Daniel M. Faissol, Deep Reinforcement Learning and Simulation as a Path Toward Precision Medicine Journal of Computational Biology 2019 26:6, 597-604. [CrossRef]
- Goudsmit, J., van den Biggelaar, A.H.J., Koudstaal, W. et al. Immune age and biological age as determinants of vaccine responsiveness among elderly populations: the Human Immunomics Initiative research program. Eur J Epidemiol 36, 753–762 (2021). [CrossRef]
- Jumper J, Evans R, Pritzel A, Green T, Figurnov M, Ronneberger O, Tunyasuvunakool K, Bates R, Žídek A, Potapenko A, et al.,. Highly accurate protein structure prediction with AlphaFold. Nature. 2021 Aug;596(7873):583-589. [CrossRef]
- Hie, B.L., Shanker, V.R., Xu, D. Efficient evolution of human antibodies from general protein language models. Nat Biotechnol (2023). [CrossRef]
- Ong, E., He, Y. (2022). Vaccine Design by Reverse Vaccinology and Machine Learning. Methods in molecular biology (Clifton, N.J.), 2414, 1–16. [CrossRef] [PubMed]
- Parihar, A., Malviya, S., Khan, R. (2022). Immunoinformatics and reverse vaccinomic approaches for effective design. Computational Approaches for Novel Therapeutic and Diagnostic Designing to Mitigate SARS-CoV-2 Infection, 357–378. [CrossRef]
- Newell EW, Sigal N, Bendall SC, Nolan GP, Davis MM. Cytometry by time-of-flight shows combinatorial cytokine expression and virus-specific cell niches within a continuum of CD8+ T cell phenotypes. Immunity. 2012 Jan 27;36(1):142-52.
- Chaplin, D.D. (2010). Overview of the immune response. The Journal of allergy and clinical immunology, 125(2 Suppl 2), S3–S23. [CrossRef]
- Kantari, C., Pederzoli-Ribeil, M., Witko-Sarsat, V. (2008). The role of neutrophils and monocytes in innate immunity. Contributions to microbiology, 15, 118–146. [CrossRef]
- Cano RLE, Lopera HDE. Introduction to T and B lymphocytes. In: Anaya JM, Shoenfeld Y, Rojas-Villarraga A, et al., editors. Autoimmunity: From Bench to Bedside Bogota (Colombia): El Rosario University Press; 2013 Jul 18. Chapter 5. (accessed on 10 July 2023).
- Wilkinson, T.M.; Li, C.K.F.; Chui, C.S.C.; Huang, A.K.Y.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.S.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef]
- Vita R, Mahajan S, Overton JA, Dhanda SK, Martini S, Cantrell JR, Wheeler DK, Sette A, Peters B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019 Jan 8;47(D1):D339-D343. [CrossRef]
- Loyal, L.; Braun, J.; Henze, L.; Kruse, B.; Dingeldey, M.; Reimer, U.; Kern, F.; Schwarz, T.; Mangold, M.; Unger, C.; et al. Cross-reactive CD4+ T cells enhance SARS-CoV-2 immune responses upon infection and vaccination. Science 2021, 374, eabh1823. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Garcia-Ibanez, L.; Ulbricht, C.; Lok, L.S.C.; Pike, J.A.; Mueller-Winkler, J.; Dennison, T.W.; Ferdinand, J.R.; Burnett, C.J.M.; Yam-Puc, J.C.; et al. Recycling of memory B cells between germinal center and lymph node subcapsular sinus supports affinity maturation to antigenic drift. Nat. Commun. 2022, 13, 2460. [Google Scholar] [CrossRef] [PubMed]
- Liu J, Chandrashekar A, Sellers D, Barrett J, Jacob-Dolan C, Lifton M, McMahan K, Sciacca M, VanWyk H, Wu C, Yu J, Collier AY, Barouch DH. Vaccines elicit highly conserved cellular immunity to SARS-CoV-2 Omicron. Nature. 2022 Mar;603(7901):493-496. [CrossRef]
- Janeway, C. A., Travers, P., Walport, M., & Shlomchik, M. J. (2001). Immunobiology: The Immune System in Health and Disease (5th ed.). Garland Science.
- Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., & Walter, P. (2002). Molecular Biology of the Cell (4th ed.). Garland Science.
- Murphy, K., Travers, P., Walport, M., & Janeway, C. (2012). Janeway's Immunobiology (8th ed.). Garland Science.
- Harty, J.T.; Tvinnereim, A.R.; White, D.W. CD8+ T cell effector mechanisms in resistance to infection. Annu. Rev. Immunol. 2000, 18, 275–308. [Google Scholar] [CrossRef]
- Marrack, P., McKee, A. S. (2017) MHC class II antigen processing in immune tolerance and inflammation. Annual Review of Immunology, 35, 13-27.
- GAVI: COVAX explained. Retrieved from. https://www.gavi.org/vaccineswork/covax-explained (accessed on 10 July 2023).
- Ura, T., Takeuchi, M., Kawagoe, T., Mizuki, N., Okuda, K., Shimada, M. (2022). Current Vaccine Platforms in Enhancing T-Cell Response. Vaccines, 10(8), 1367. [CrossRef]
- Crotty, S. (2011) Follicular helper CD4 T-cells (TFH). Annual Review of Immunology, 29, 621-663.
- Plotkin, S. A. (2008) Vaccines: correlates of vaccine-induced immunity. Clinical Infectious Diseases, 47(3), 401-409.
- Gilbert, S.C. T-cell-inducing vaccines—What’s the future. Immunology 2012, 135, 19–26. [Google Scholar] [CrossRef]
- Colditz, G.A.; Brewer, T.F.; Berkey, C.S.; Wilson, M.E.; Burdick, E.; Fineberg, H.V.; Mosteller, F. Efficacy of BCG vaccine in the prevention of tuberculosis. Meta-analysis of the published literature. JAMA 1994, 271, 698–702. [Google Scholar] [CrossRef]
- Mishra R, Hannebelle M, Patil VP, Dubois A, Garcia-Mouton C, Kirsch GM, Jan M, Sharma K, Guex N, Sordet-Dessimoz J, er al., Mechanopathology of biofilm-like Mycobacterium tuberculosis cords. Cell. 2023 Oct 11:S0092-8674(23)01037-1. [CrossRef]
- Prabhudas, M.; Bonney, E.; Caron, K.; Dey, S.; Erlebacher, A.; Fazleabas, A.; Fisher, S.; Golos, T.; Matzuk, M.; McCune, J.M.; et al. Immune mechanisms at the maternal-fetal interface: Perspectives and challenges. Nat. Immunol. 2015, 16, 328–334. [Google Scholar] [CrossRef]
- Guthmiller, J.J.; Stovicek, O.; Wang, J.; Changrob, S.; Li, L.; Halfmann, P.; Zheng, N.Y.; Utset, H.; Stamper, C.T.; Dugan, H.L.; et al. SARS-CoV-2 Infection Severity Is Linked to Superior Humoral Immunity against the Spike. MBio 2021, 12, e02940-20. [Google Scholar] [CrossRef]
- Turner JS, O'Halloran JA, Kalaidina E, Kim W, Schmitz AJ, Zhou JQ, Lei T, Thapa M, Chen RE, Case JB, er al., SARS-CoV-2 mRNA vaccines induce persistent human germinal centre responses. Nature. 2021 Aug;596(7870):109-113. [CrossRef]
- Goel RR, Painter MM, Apostolidis SA, Mathew D, Meng W, Rosenfeld AM, Lundgreen KA, Reynaldi A, Khoury DS, Pattekar A, Gouma S, Kuri-Cervantes L, Hicks P, Dysinger S, Hicks A, Sharma H, Herring S, Korte S, Baxter AE, Oldridge DA, Giles JR, Weirick ME, McAllister CM, Awofolaju M, Tanenbaum N, Drapeau EM, Dougherty J, Long S, D'Andrea K, Hamilton JT, McLaughlin M, Williams JC, Adamski S, Kuthuru O; UPenn COVID Processing Unit‡; Frank I, Betts MR, Vella LA, Grifoni A, Weiskopf D, Sette A, Hensley SE, Davenport MP, Bates P, Luning Prak ET, Greenplate AR, Wherry EJ. mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern. Science. 2021 Dec 3;374(6572):abm0829. [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Invest. 2020, 130, 2620–2629. [Google Scholar] [CrossRef]
- Leung, S.; Liu, X.; Fang, L.; Chen, X.; Guo, T.; Zhang, J. The cytokine milieu in the interplay of pathogenic Th1/Th17 cells and regulatory T cells in autoimmune disease. Cell Mol. Immunol. 2010, 7, 182–189. [Google Scholar] [CrossRef]
- Khader, S.A.; Gaffen, S.L.; Kolls, J.K. Th17 cells at the crossroads of innate and adaptive immunity against infectious diseases at the mucosa. Mucosal. Immunol. 2009, 2, 403–411. [Google Scholar] [CrossRef]
- Ndure, J.; Flanagan, K.L. Targeting regulatory T cells to improve vaccine immunogenicity in early life. Front. Microbiol. 2014, 5, 477. [Google Scholar] [CrossRef]
- Amanat F, Krammer F. SARS-CoV-2 Vaccines: Status Report. Immunity. 2020;52(4):583–9.
- Chen J, Ting N. Design Considerations for Vaccine Trials with a Special Focus on COVID-19 Vaccine Development. Data Sci J. 2021;18(3):550–80. [CrossRef]
- Borah P, Deb PK, Deka S, Venugopala KN, Singh V, Mailavaram RP, Kalia K, Tekade RK. Current Scenario and Future Prospect in the Management of COVID-19. Curr Med Chem. 2021;28(2):284-307. [CrossRef]
- Kim MH, Kim HJ, Chang J. Superior immune responses induced by intranasal immunization with recombinant adenovirus-based vaccine expressing full-length Spike protein of Middle East respiratory syndrome Coronavirus. PLoS One. 2019;14(7). e0220196. [CrossRef]
- Bouhaddou M, Reuschl A-K, Polacco BJ, Thorne LG, Ummadi MR, Ye C, et al. SARS-CoV-2 variants evolve convergent strategies to remodel the host response. Cell. 2023;186(21):4597-614.e26.
- Plotkin, S. A. (2010) Correlates of protection induced by vaccination. Clinical and Vaccine Immunology, 17(7), 1055-1065. [CrossRef]
- Palucka, K., Banchereau, J. (2012) Cancer immunotherapy via dendritic cells. Nature Reviews Cancer, 12(4), 265-277. [CrossRef]
- Zhen, A., Kamata, M. (2020) Viral vectors for the delivery of human genes. Journal of Cellular Physiology, 235(4), 3235-3249. [CrossRef]
- Sadoff, J.; Le Gars, M.; Shukarev, G.; Heerwegh, D.; Truyers, C.; de Groot, A.M.; Stoop, J.; Tete, S.; Damme, W.V.; Leroux-Roels, I.; et al. Interim Results of a Phase 1–2a Trial of Ad26.COV2.S COVID-19 Vaccine. N. Engl. J. Med. 2021, 384, 1824–1835. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Hokello, J.; Sharma, A.L.; Tyagi, M. An Update on the HIV DNA Vaccine Strategy. Vaccines 2021, 9, 605. [Google Scholar] [CrossRef]
- Schenkel, J.M.; Masopust, D. Tissue-resident memory T cells. Immunity 2014, 41, 886–897. [Google Scholar] [CrossRef]
- Yusuf, H.; Kett, V. Current prospects and future challenges for nasal vaccine delivery. Hum. Vaccin Immunother. 2017, 13, 34–45. [Google Scholar] [CrossRef]
- Menon, I.; Bagwe, P.; Gomes, K.; Bajaj, L.; Gala, R.; Uddin, M.; D’Souza, M.; Zughaier, S. Microneedles: A New Generation Vaccine Delivery System. Micromachines 2021, 12, 435. [Google Scholar] [CrossRef] [PubMed]
- Pleguezuelos, O.; James, E.; Fernandez, A.; Lopes, V.; Rosas, L.A.; Cervantes-Medina, A.; Cleath, J.; Edwards, K.; Neitzey, D.; Gu, W.; et al. Efficacy of FLU-v, a broad-spectrum influenza vaccine, in a randomized phase IIb human influenza challenge study. NPJ Vaccines 2020, 5, 22. [Google Scholar] [CrossRef]
- Atsmon, J.; Kate-Ilovitz, E.; Shaikevich, D.; Singer, Y.; Volokhov, I.; Haim, K.Y.; Ben-Yedidia, T. Safety and immunogenicity of multimeric-001—A novel universal influenza vaccine. J. Clin. Immunol. 2012, 32, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Coffman, R. L., Sher, A., Seder, R. A. (2010). Vaccine adjuvants: Putting innate immunity to work. Immunity, 33(4), 492-503. [CrossRef]
- Krieg, A. M. (2002) CpG motifs in bacterial DNA and their immune effects. Annual Review of Immunology, 20, 709-760. [CrossRef]
- Lynn GM, Laga R, Darrah PA, Ishizuka AS, Balaci AJ, Dulcey AE, Pechar M, Pola R, Gerner MY, Yamamoto A. In vivo characterization of the physicochemical properties of polymer-linked TLR agonists that enhance vaccine immunogenicity. Nat Biotechnol. 2015 Nov;33(11):1201-10. [CrossRef]
- Graham, B. S., Sullivan, N. J. (2018) Emerging viral diseases from a vaccinology perspective: preparing for the next pandemic. Nature Immunology, 19(1), 20-28. [CrossRef]
- Shah, N.J.; Najibi, A.J.; Shih, T.-Y.; Mao, A.S.; Sharda, A.; Scadden, D.T.; Mooney, D.J. A biomaterial-based vaccine eliciting durable tumour-specific responses against acute myeloid leukaemia. Nat. Biomed. Eng. 2020, 4, 40–51. [Google Scholar] [CrossRef]
- Liu, H.; Moynihan, K.; Zheng, Y.; Szeto, G.; Li, A.V.; Huang, B.; Van Egeren, D.; Park, C.; Irvine, D.J. Structure-based programming of lymph-node targeting in molecular vaccines. Nature 2014, 507, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Pollard C, De Koker S, Saelens X, Vanham G, Grooten J. Challenges and advances towards the rational design of mRNA vaccines. Trends Mol Med. 2013;19(12):705–13. [CrossRef]
- Kanampalliwar, AM. Reverse Vaccinology and Its Applications. Methods Mol Biol. 2020;2131:1–16. [CrossRef]
- Cheung YK, Cheng SC, Sin FW, Chan KT, Xie Y. Induction of T- cell response by a DNA vaccine encoding a novel HLA-A*0201 severe acute respiratory syndrome coronavirus epitope. Vaccine. 2007;25(32):6070–7. [CrossRef]
- Wang X, Xu W, Tong D, Ni J, Gao H, Wang Y, Chu Y, Li P, Yang X, Xiong S. A chimeric multi-epitope DNA vaccine elicited specific antibody response against severe acute respiratory syndrome-associated coronavirus which attenuated the virulence of SARS-CoV in vitro. Immunol Lett. 2008 Aug 15;119(1-2):71-7. [CrossRef]
- Voysey M, Clemens SAC, Madhi SA, Weckx LY, Folegatti PM, Aley PK, Angus B, Baillie VL, Barnabas SL, Bhorat QE,; Oxford COVID Vaccine Trial Group. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: an interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet. 2021 Jan 9;397(10269):99-111. [CrossRef]
- Alarcon JB, Waine GW, McManus DP (1999). "DNA Vaccines: Technology and Application as Anti-parasite and Anti-microbial Agents". Advances in Parasitology Volume 42. Vol. 42. pp. 343–410. [CrossRef]
- Robinson HL, Pertmer TM (2000). DNA vaccines for viral infections: basic studies and applications. Advances in Virus Research. Vol. 55. pp. 1–74. [CrossRef]
- Sheridan, C. First COVID-19 DNA vaccine approved, others in hot pursuit. Nat. Biotechnol. 2021, 39, 1479–1482. [Google Scholar] [CrossRef]
- India’s DNA COVID vaccine is a world first – more are coming. https://www.nature.com/articles/d41586-021-02385-x (accessed on 10 July 2023).
- Khobragade A, Bhate S, Ramaiah V, Deshpande S, Giri K, Phophle H, Supe P, Godara I, Revanna R, Nagarkar R, Sanmukhani J, Dey A, Rajanathan TMC, Kansagra K, Koradia P; ZyCoV-D phase 3 Study Investigator Group. Efficacy, safety, and immunogenicity of the DNA SARS-CoV-2 vaccine (ZyCoV-D): the interim efficacy results of a phase 3, randomised, double-blind, placebo-controlled study in India. Lancet. 2022 Apr 2;399(10332):1313-1321. [CrossRef]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef]
- Mothe, B., Brander, C. (2016) HIV T-cell vaccines. Human Vaccines Immunotherapeutics, 12(8), 2013-2016.
- Colombani T, Haudebourg T, Pitard B. 704/DNA vaccines leverage cytoplasmic DNA stimulation to promote anti-HIV neutralizing antibody production in mice and strong immune response against alpha-fetoprotein in non-human primates. Mol Ther Nucleic Acids. 2023 May 4;32:743-757. [CrossRef]
- Lim, W. A., June, C. H. (2017) The principles of engineering immune cells to treat cancer. Cell, 168(4), 724-740. [CrossRef] [PubMed]
- Twyman-Saint Victor C, Rech AJ, Maity A, Rengan R, Pauken KE, Stelekati E, Benci JL, Xu B, Dada H, Odorizzi PM, Herati RS, et al., Radiation and dual checkpoint blockade activate non-redundant immune mechanisms in cancer. Nature. 2015 Apr 16;520(7547):373-7. [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Buddy Creech, C.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Güler, A.; et al. BNT162b vaccines protect rhesus macaques from SARS-CoV-2. Nature 2021, 592, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Niazi, SK. RNA Therapeutics: A Healthcare Paradigm Shift. Biomedicines. 2023 Apr 25;11(5):1275. [CrossRef]
- Oleszycka, E., Lavelle, E. C. (2014) Immunomodulatory properties of the vaccine adjuvant alum. Current Opinion in Immunology, 28, 1-5. 28, 1–5. [CrossRef] [PubMed]
- Agnandji ST, Lell B, Fernandes JF, Abossolo BP, Methogo BG, Kabwende AL, Adegnika AA, Mordmüller B, Issifou S, Kremsner PG, A phase 3 trial of RTS,S/AS01 malaria vaccine in African infants. N Engl J Med. 2012 Dec 13;367(24):2284-95. [CrossRef]
- Khurana, S., Verma, N. (2021) MF59 adjuvant: The best insurance against influenza strain diversity. Expert Review of Vaccines, 20(8), 947-951. [CrossRef]
- Gustiananda M, Sulistyo BP, Agustriawan D, Andarini S. Immunoinformatics Analysis of SARS-CoV-2 ORF1ab Polyproteins to Identify Promiscuous and Highly Conserved T-Cell Epitopes to Formulate Vaccine for Indonesia and the World Population. Vaccines (Basel). 2021 Dec 9;9(12):1459. [CrossRef]
- Mitra R, Das J, Kamruzzaman M. Application of TOPSIS method for flood susceptibility mapping using Excel and GIS. MethodsX. 2023 Jun 15;11:102263. [CrossRef]
- Bagherzadeh MA, Izadi M, Baesi K, Jahromi MAM, Pirestani M. Considering epitopes conservity in targeting SARS-CoV-2 mutations in variants: a novel immunoinformatics approach to vaccine design. Sci Rep. 2022 Aug 18;12(1):14017. [CrossRef]
- Moss P. The T cell immune response against SARS-CoV-2. Nat Immunol. 2022;23:186–193. [CrossRef]
- Olukitibi TA, Ao Z, Warner B, Unat R, Kobasa D, Yao X. Significance of Conserved Regions in Coronavirus Spike Protein for Developing a Novel Vaccine against SARS-CoV-2 Infection. Vaccines (Basel). 2023 Feb 24;11(3):545. [CrossRef]
- Heinson, A. I., Woelk, C. H., Newell, M. L. (2015). The promise of reverse vaccinology. International health, 7(2), 85–89. [CrossRef] [PubMed]
- Pritam M, Singh G, Swaroop S, Singh AK, Singh SP. Exploitation of reverse vaccinology and immunoinformatics as promising platform for genome-wide screening of new effective vaccine candidates against Plasmodium falciparum. BMC Bioinformatics. 2019 Feb 4;19(Suppl 13):468. [CrossRef]
- Jespersen MC, Peters B, Nielsen M, Marcatili P. BepiPred-2.0: improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017 Jul 3;45(W1):W24-W29. [CrossRef]
- Larsen JE, Lund O, Nielsen M. Improved method for predicting linear B-cell epitopes. Immunome Res. 2006;2:2. [CrossRef]
- Saha S, Raghava GP. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins. 2006;65(1):40–8. [CrossRef]
- Rahman Kh S, Chowdhury EU, Sachse K, Kaltenboeck B. Inadequate Reference Datasets Biased toward Short Non-epitopes Confound B-cell Epitope Prediction. J Biol Chem. 2016;291(28):14585–99. [CrossRef]
- Ansari HR, Raghava GP. Identification of conformational B-cell Epitopes in an antigen from its primary sequence. Immunome Res. 2010;6:6. [CrossRef]
- Parvizpour S, Razmara J, Pourseif MM, Omidi Y. In silico design of a triple-negative breast cancer vaccine by targeting cancer testis antigens. Bioimpacts. 2019;9(1):45–56. [CrossRef]
- Potocnakova L, Bhide M, Pulzova LB. An Introduction to B-Cell Epitope Mapping and In Silico Epitope Prediction. J Immunol Res. 2016;2016:6760830. [CrossRef]
- Lundegaard C, Hoof I, Lund O, Nielsen M. State of the art and challenges in sequence based T-cell epitope prediction. Immunome Res. 2010;6 Suppl 2. S3. [CrossRef]
- Ott PA, Hu Z, Keskin DB, Shukla SA, Sun J, Bozym DJ, Zhang W, Luoma A, Giobbie-Hurder A, Peter L,. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature. 2017 Jul 13;547(7662):217-221. [CrossRef]
- Burgos RM, Badowski ME, Drwiega E, Ghassemi S, Griffith N, Herald F, Johnson M, Smith RO, Michienzi SM. The race to a COVID-19 vaccine: opportunities and challenges in development and distribution. Drugs Context. 2021 Feb 16;10:2020-12-2. [CrossRef]
- Mills MC, Salisbury D. The challenges of distributing COVID-19 vaccinations. EClinicalMedicine. 2021;31:100674. [CrossRef]
- Krause, P.R.; Fleming, T.R.; Longini, I.M.; Peto, R.; Briand, S.; Heymann, D.L.; Beral, V.; Snape, M.D.; Rees, H.; Ropero, A.M.; et al. SARS-CoV-2 Variants and Vaccines. N. Engl. J. Med. 2021, 385, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Accorsi, E.K.; Britton, A.; Fleming-Dutra, K.E.; Smith, Z.R.; Shang, N.; Derado, G.; Miller, J.; Schrag, S.J.; Verani, J.R. Association Between 3 Doses of mRNA COVID-19 Vaccine and Symptomatic Infection Caused by the SARS-CoV-2 Omicron and Delta Variants. JAMA 2022, 327, 639–651. [Google Scholar] [CrossRef]
- Hoffmann, M.; Krüger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.-S.; Winkler, M.S.; et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2021, 185, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Kidd, C.K.; Dan, J.M.; Ramirez, S.I.; Yu, E.D.; Mateus, J.; da Silva Antunes, R.; Moore, E.; Rubiro, P.; et al. Comprehensive analysis of T cell immunodominance and immunoprevalence of SARS-CoV-2 epitopes in COVID-19 cases. Cell Rep. Med. 2021, 2, 100204. [Google Scholar] [CrossRef]
- Swadling, L.; Diniz, M.O.; Schmidt, N.M.; Amin, O.E.; Chandran, A.; Shaw, E.; Pade, C.; Gibbons, J.M.; Le Bert, N.; Tan, A.T.; et al. Pre-existing polymerase-specific T cells expand in abortive seronegative SARS-CoV-2. Nature 2022, 601, 110–117. [Google Scholar] [CrossRef]
- Mallajosyula, V.; Ganjavi, C.; Chakraborty, S.; McSween, A.M.; Pavlovitch-Bedzyk, A.J.; Wilhelmy, J.; Nau, A.; Manohar, M.; Nadeau, K.C.; Davis, M.M. CD8+ T cells specific for conserved coronavirus epitopes correlate with milder disease in COVID-19 patients. Sci. Immunol. 2021, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Gangaev, A.; Ketelaars, S.L.C.; Isaeva, O.I.; Patiwael, S.; Dopler, A.; Hoefakker, K.; Biasi, S.D.; Gibellini, L.; Mussini, C.; Guaraldi, G.; et al. Identification and characterization of a SARS-CoV-2 specific CD8+ T cell response with immunodominant features. Nat. Commun. 2021, 12, 2593. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov I, Flower DR, Doytchinova I. AllerTOP--a server for in silico prediction of allergens. BMC Bioinformatics. 2013;14 Suppl 6(Suppl 6):S4. [CrossRef]
- Doytchinova IA, Flower DR. VaxiJen: a server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinformatics. 2007 Jan 5;8:4. [CrossRef]
- http://www.ddg-pharmfac.net/vaxijen/ (accessed on 10 July 2023).
- https:// webs.iiitd.edu.in/raghava/toxinpred/design.php (accessed on 10 July 2023).
- http://tools.iedb.org/ellipro/ (accessed on 10 July 2023).
- Bui HH, Sidney J, Dinh K, Southwood S, Newman MJ, Sette A. Predicting population coverage of T-cell epitope-based diagnostics and vaccines. BMC Bioinformatics. 2006 Mar 17;7:153. [CrossRef]
- Lamiable A, Thévenet P, Rey J, Vavrusa M, Derreumaux P, Tufféry P. PEP-FOLD3: faster de novo structure prediction for linear peptides in solution and in complex. Nucleic Acids Res. 2016 Jul 8;44(W1):W449-54. [CrossRef]
- Schneidman-Duhovny D, Inbar Y, Nussinov R, Wolfson HJ. PatchDock and SymmDock: servers for rigid and symmetric docking. Nucleic Acids Res. 2005 Jul 1;33(Web Server issue):W363-7. [CrossRef]
- Mashiach E, Schneidman-Duhovny D, Andrusier N, Nussinov R, Wolfson HJ. FireDock: a web server for fast interaction refinement in molecular docking. Nucleic Acids Res. 2008 Jul 1;36(Web Server issue):W229-32. [CrossRef]
- https:// web.expasy.org/protparam (accessed on 10 July 2023).
- Petersen TN, Brunak S, von Heijne G, Nielsen H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Methods. 2011 Sep 29;8(10):785-6. [CrossRef]
- https://zhanglab.ccmb.med.umich.edu/I-TASSER/ (accessed on 10 July 2023).
- https://zhanggroup.org/ModRefiner/ (accessed on 10 July 2023).
- Wiederstein M, Sippl MJ. ProSA-web: interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007 Jul;35(Web Server issue):W407-10. [CrossRef]
- https:// servicesn.mbi.ucla.edu/ERRAT/ (accessed on 10 July 2023).
- https:// servicesn.mbi.ucla.edu/PROCHECK/ (accessed on 10 July 2023).
- https://www.rcsb.org/ (accessed on 10 July 2023).
- https:// cluspro.bu.edu/login.php (accessed on 10 July 2023).
- https://pymol.org/2/ (accessed on 10 July 2023).
- https://kraken.iac.rm.cnr.it/CIMMSIM/ (accessed on 10 July 2023).
- http://apps.cytoscape.org/apps/cytohubb (accessed on 10 July 2023).
- https://services.healthtech.dtu.dk/services/NetMHCpan-4.1/ (accessed on 10 July 2023).
- Poonam, B., Gill, P. K. (2021). Coronavirus: History, genome structure and pathogenesis. Coronaviruses, 2(3), 325-338. [CrossRef]
- Kakavandi, S., Zare, I., Vaezjalali, M., Dadashi, M., Azarian, M., Akbari, A., Farani, M. R., Zalpoor, H., Hajikhani, B. (2023). Structural and non-structural proteins in SARS-CoV-2: potential aspects to COVID-19 treatment or prevention of progression of related diseases. Cell Communication and Signaling, 21(1). [CrossRef]
- Ong, E., Wong, M. U., Huffman, A., He, Y. (2020). COVID-19 coronavirus vaccine design using reverse vaccinology and machine learning. Frontiers in immunology, 11, 1581. [CrossRef]
- Zhang S, Gong C, Ruiz-Martinez A, Wang H, Davis-Marcisak E, Deshpande A, Popel AS, Fertig EJ. Integrating single cell sequencing with a spatial quantitative systems pharmacology model spQSP for personalized prediction of triple-negative breast cancer immunotherapy response. Immunoinformatics (Amst). 2021 Oct;1-2:100002. [CrossRef]
- Gustafson, C.E.; Kim, C.; Weyand, C.M.; Goronzy, J.J. Influence of immune aging on vaccine responses. J. Allergy Clin. Immunol. 2020, 145, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Allen H, Vusirikala A, Flannagan J, Twohig KA, Zaidi A, Chudasama D, Lamagni T, Groves N, Turner C, Rawlinson C, et al; COVID-19 Genomics UK (COG-UK Consortium). Household transmission of COVID-19 cases associated with SARS-CoV-2 delta variant (B.1.617.2): national case-control study. Lancet Reg Health Eur. 2022 Jan;12:100252. [CrossRef]
- Zhou B, Thao TTN, Hoffmann D, Taddeo A, Ebert N, Labroussaa F, Pohlmann A, King J, Steiner S, Kelly JN, Portmann J, SARS-CoV-2 spike D614G change enhances replication and transmission. Nature. 2021 Apr;592(7852):122-127.). [CrossRef]
- Jangra S, Ye C, Rathnasinghe R, Stadlbauer D; Personalized Virology Initiative study group; Krammer F, Simon V, Martinez-Sobrido L, García-Sastre A, Schotsaert M. SARS-CoV-2 spike E484K mutation reduces antibody neutralisation. Lancet Microbe. 2021 Jul;2(7):e283-e284. [CrossRef]
- Almubaid, Z. Al-Mubaid, H. Analysis and comparison of genetic variants and mutations of the novel coronavirus SARS-CoV-2. Gene Rep. 23, 101064 (2021). [CrossRef]
- Hassan, S. S., Choudhury, P. P., Basu, P. Jana, S. S. Molecular conservation and differential mutation on ORF3a gene in Indian SARS-CoV2 genomes. Genomics 112, 3226–3237 (2020). [CrossRef]
- Dai Y, Chen H, Zhuang S, Feng X, Fang Y, Tang H, Dai R, Tang L, Liu J, Ma T, Zhong G. Immunodominant regions prediction of nucleocapsid protein for SARS-CoV-2 early diagnosis: a bioinformatics and immunoinformatics study. Pathog Glob Health. 2020 Dec;114(8):463-470.). [CrossRef]
- Garrett ME, Galloway J, Chu HY, Itell HL, Stoddard CI, Wolf CR, Logue JK, McDonald D, Weight H, Matsen FA 4th, Overbaugh J. High-resolution profiling of pathways of escape for SARS-CoV-2 spike-binding antibodies. Cell. 2021 May 27;184(11):2927-2938.e11.).
- Zeng, C.; Zhang, C.; Walker, P.G.; Dong, Y. Formulation and Delivery Technologies for mRNA Vaccines. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2020.
- Kenney, R.T.; Frech, S.A.; Muenz, L.R.; Villar, C.P.; Glenn, G.M. Dose sparing with intradermal injection of influenza vaccine. N. Engl. J. Med. 2004, 351, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.; Naradikian, M.S.; Parkhouse, K.; Cain, D.W.; Jones, L.; Moody, M.A.; Verkerke, H.P.; Myles, A.; Willis, E.; et al. Nucleoside-modified mRNA vaccines induce potent T follicular helper and germinal center B cell responses. J. Exp. Med. 2018, 215, 1571–1588. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, S45–S53. [Google Scholar] [CrossRef]
- Xiong F, Zhang C, Shang B, Zheng M, Wang Q, Ding Y, Luo J, Li X. An mRNA-based broad-spectrum vaccine candidate confers cross-protection against heterosubtypic influenza A viruses. Emerg Microbes Infect. 2023 Dec;12(2):2256422. [CrossRef]
- https://investors.modernatx.com/news/news-details/2023/Moderna-Announces-First-Participant-Dosed-in-Phase-3-Study-of-mRNA-1083-a-Combination-Vaccine-Against-Influenza-and-COVID-19/default.aspx.
- Sahin, U., Derhovanessian, E., Miller, M., Kloke, B.P., Simon, P., Löwer, M., Omokoko, T. (2017). Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature, 547(7662), 222-226. [CrossRef]
- Berg MK, Yu Q, Salvador CE, Melani I, Kitayama S. Mandated Bacillus Calmette-Guerin (BCG) vaccination predicts flattened curves for the spread of COVID-19. Sci Adv. 2020;6(32):eabc1463. [CrossRef]
- Singh, S. BCG vaccines may not reduce COVID-19 mortality rates.; medRxiv 2020.04.11.20062232.
- June CH, O'Connor RS, Kawalekar OU, Ghassemi S, Milone MC. CAR T cell immunotherapy for human cancer. Science. 2018 Mar 23;359(6382):1361-1365. [CrossRef]
- Liu J, Fu M, Wang M, Wan D, Wei Y, Wei X. Cancer vaccines as promising immuno-therapeutics: platforms and current progress. J Hematol Oncol. 2022 Mar 18;15(1):28. [CrossRef]
- Apostolopoulos, V. (2019). Cancer Vaccines: Research and Applications. Cancers, 11(8), 1041. [CrossRef]
- Kübler, H., Scheel, B., Gnad-Vogt, U., Miller, K., Schultze-Seemann, W., Vom Dorp, F. Kallen, K.J. (2015). Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: a first-in-man phase I/IIa study. Journal of Immunotherapy of Cancer, 3(1), 26. [CrossRef]
- Boczkowski, D., Nair, S.K., Snyder, D., & Gilboa, E. (1996). Dendritic cells pulsed with RNA are potent antigen-presenting cells in vitro and in vivo. The Journal of Experimental Medicine, 184(2), 465-472. [CrossRef]
- Ribas, A., & Wolchok, J.D. (2018). Cancer immunotherapy using checkpoint blockade. Science, 359(6382), 1350-1355. [CrossRef] [PubMed]
- Reichmuth, A. M., Oberli, M. A., Jaklenec, A., Langer, R., & Blankschtein, D. (2016). mRNA vaccine delivery using lipid nanoparticles. Therapeutic Delivery, 7(5), 319-334. [CrossRef]
- Binnewies, M., Roberts, E. W., Kersten, K., Chan, V., Fearon, D. F., Merad, M., Krummel, M. F. (2018). Understanding the tumor immune microenvironment (TIME) for effective therapy. Nature Medicine, 24(5), 541-550. [CrossRef]
- Vanpouille-Box, C., Alard, A., Aryankalayil, M. J., Sarfraz, Y., Diamond, J. M., Schneider, R. J. Formenti, S. C. (2017). DNA exonuclease Trex1 regulates radiotherapy-induced tumour immunogenicity. Nature Communications, 8, 15618. [CrossRef]
- Galluzzi, L., Buqué, A., Kepp, O., Zitvogel, L., & Kroemer, G. (2017). Immunogenic cell death in cancer and infectious disease. Nature Reviews Immunology, 17(2), 97-111. [CrossRef]
- Slaoui, M., & Hepburn, M. (2020). Developing Safe and Effective COVID Vaccines—Operation Warp Speed's Strategy and Approach. The New England Journal of Medicine, 383(18), 1701-1703. [CrossRef]
- Bach, P. B. (2021). The Price of Progress—Cost of the COVID-19 Vaccine. JAMA Health Forum, 2(5), e210524-e210524. [CrossRef]
- Le, I., Dhandayuthapani, S., Chacon, J., Eiring, A. M., and Gadad, S. S. (2022). Harnessing the Immune System with Cancer Vaccines: From Prevention to Therapeutics. Vaccines, 10(5), 816. [CrossRef]
- Schumacher, T. N., Schreiber, R. D. (2015) Neoantigens in cancer immunotherapy. Science, 348(6230), 69-74. [CrossRef] [PubMed]
- Hilf N, Kuttruff-Coqui S, Frenzel K, Bukur V, Stevanović S, Gouttefangeas C, Platten M, Tabatabai G, Dutoit V, et al.,Actively personalized vaccination trial for newly diagnosed glioblastoma. Nature. 2019 Jan;565(7738):240-245. Erratum in: Nature. 2019 Feb;566(7745):E13. [CrossRef]
- Gorkhali, R., Koirala, P., Rijal, S., Mainali, A., Baral, A., Bhattarai, H. K. (2021). Structure and function of major SARS-CoV-2 and SARS-CoV proteins. Bioinformatics and biology insights, 15, 11779322211025876. [CrossRef]
Figure 1.
Body Immune System (Shutterstock).
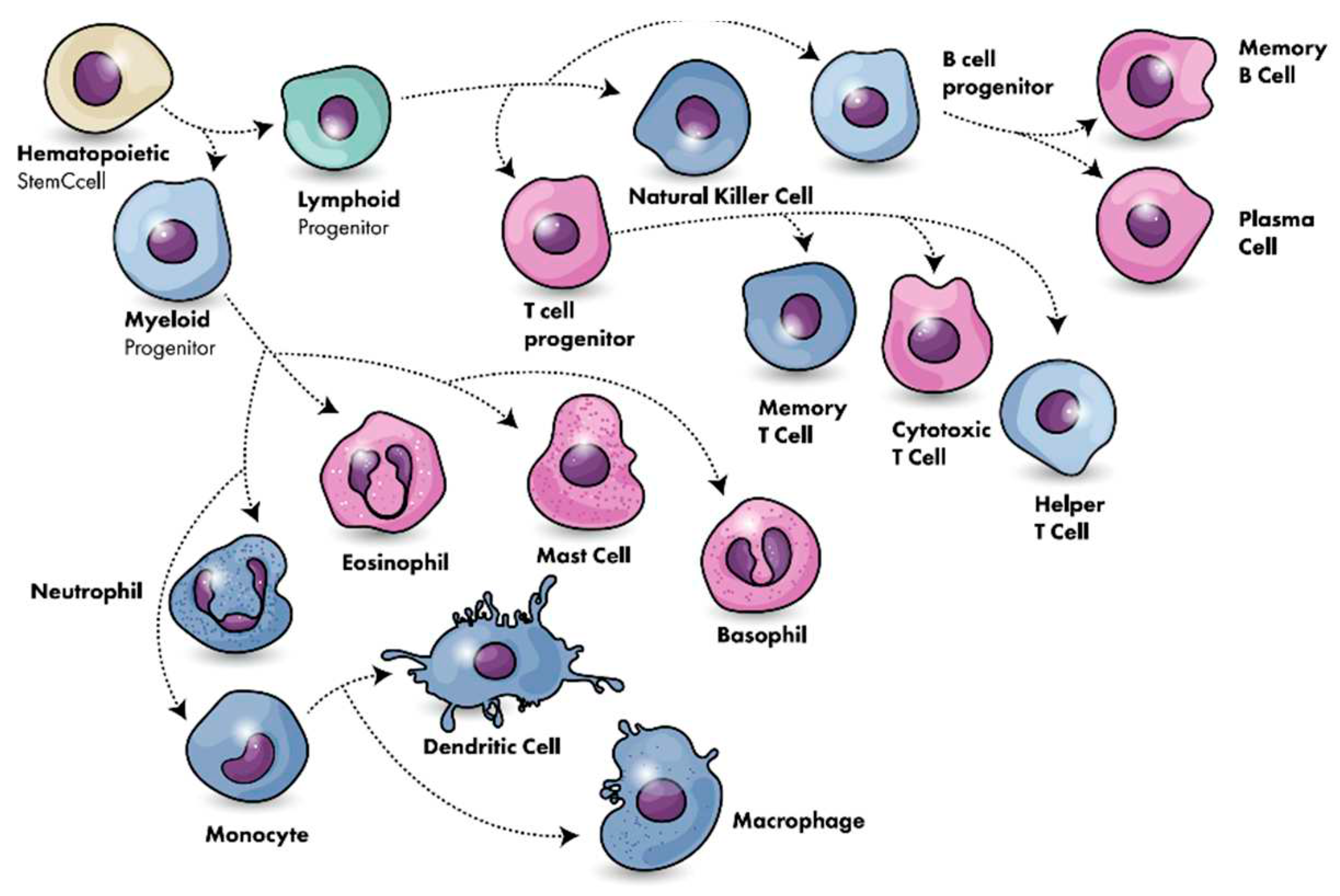
Figure 2.
T-cell activation and immune process. [Shutterstock].
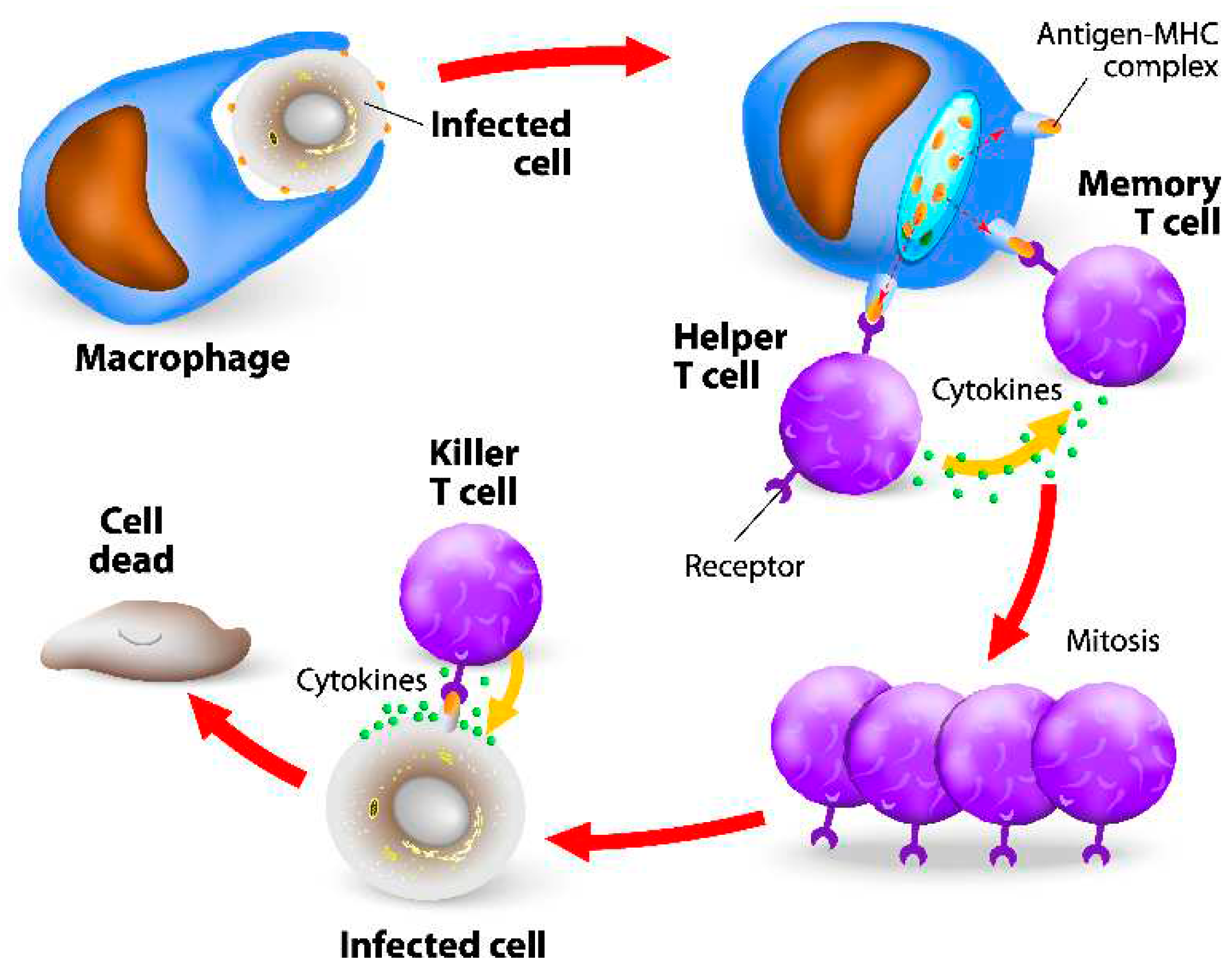
Figure 3.
Humoral immunity activation scheme [Shutterstock].
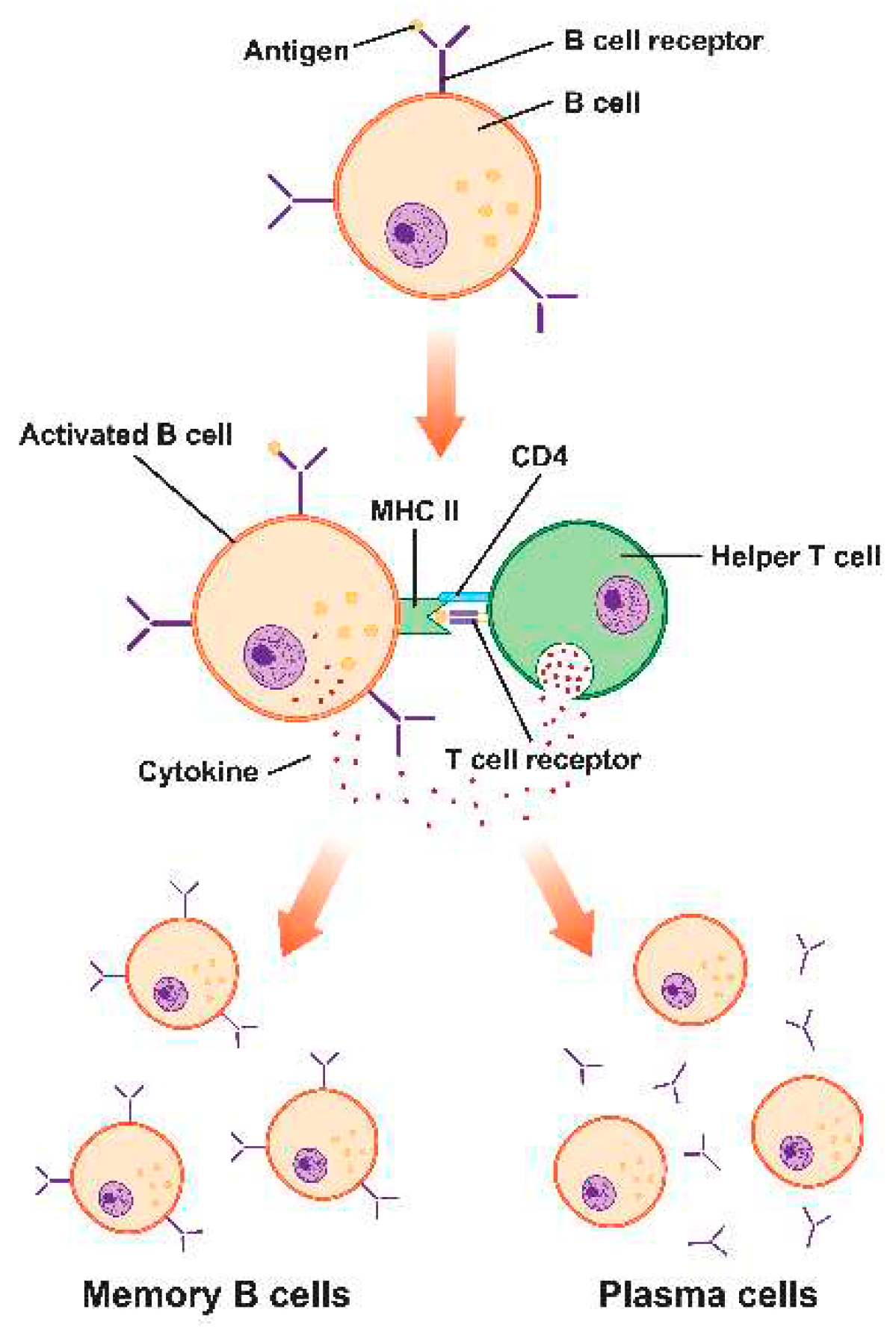
Figure 4.
Bioinformatics-driven pipeline for reverse vaccine analysis.
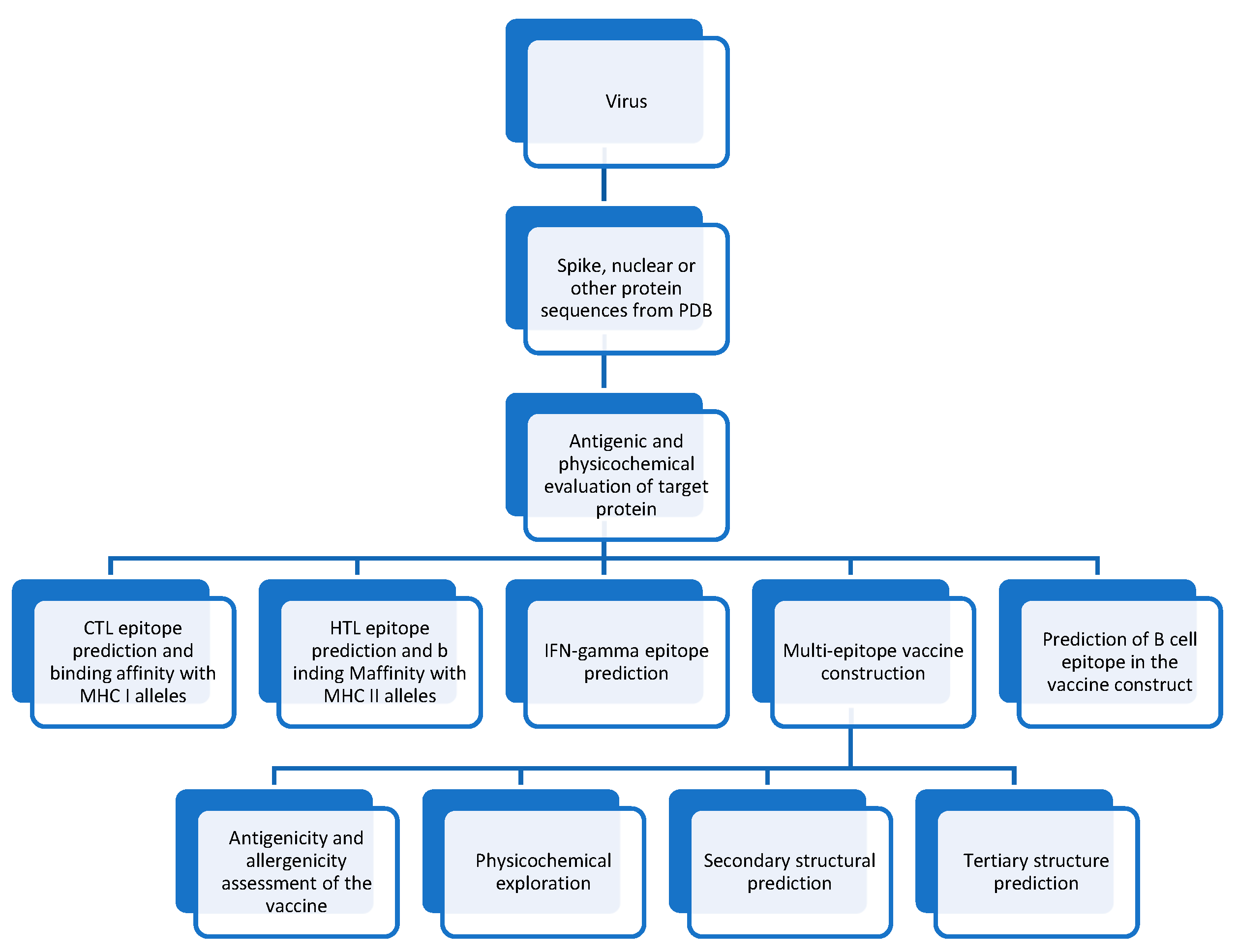

Table 1.
SARS-Cov-2 Vaccines under development [16].
Table 1.
SARS-Cov-2 Vaccines under development [16].
| Type Abb. | Vaccine Type | # |
| PS | Protein subunit | 59 |
| RNA | RNA | 43 |
| VVnr | Viral Vector (non-replicating) | 25 |
| IV | Inactivated Virus | 22 |
| DNA | DNA | 17 |
| VLP | Virus-Like Particle | 7 |
| VVr | Viral Vector (replicating) | 4 |
| VVr + APC | VVr + Antigen Presenting Cell | 2 |
| LAV | Live Attenuated Virus | 2 |
| VVnr + APC | VVnr + Antigen Presenting Cell | 1 |
| BacAg-SpV | Bacterial antigen-spore expression vector | 1 |
Table 1.
Projected conserved epitopes of SARS-Cov2.
| Conserved Epitopes | Position | Immune Response Induced | Type of Study | |
| B cells/Neutralizing antibodies | T cells | |||
| YLTPGDSSSGWTAGAAAYYV | 248–267 aa | Yes | Yes | Mathematically (in-house developed PERL scripts), in vivo |
| YYVGYLQPRTFLLKY | 264–278 aa | NT | Yes | Web-based analytic tools |
| VRFPNITNL | 327–335 aa | NT | Yes | Immunoinformatic, in vivo |
| FNATRFASVYAWNRK | 342–356 aa | Yes | Yes | In silico, T-cell epitope mapping, molecular dynamics simulations, immunoinformatic |
| TFKCYGVSPTKLNDL | 376–390 aa | Yes | Yes | Mathematically (in-house developed PERL scripts), bioinformatics, monoclonal antibody targeting |
| PYRVVVLSF | 507–515 aa | NT | Yes | Immunoinformatic |
| LPFQQFGRDIADT | 560–572 aa | Yes | Yes | PepSeq Analysis |
Table 2.
Epitope testing tools for vaccine development.
| Testing | Tools |
| Antigenic, non-toxic, and non-allergenic epitopes | AllerTop v2. [130] VaxiJen v.2.0 [131,132] and ToxinPred [133] |
| Linear and discontinuous B-cell epitope prediction | Ellipro [134] |
| Overlapping epitopes | IEDB. [135] |
| Conservancy of epitopes analysis | Infectious Disease Epidemiology Bureau conservancy tool. [135] |
| 3-D structure of screened-out best epitope sequences. | PEPFOLD 3l [136] |
| Epitope and allele interaction pattern | PatchDockr [137] |
| Complex Refining | FireDock [138] |
| Antigenicity anticipation | ANTIGENpro and VaxiJen v2.0 |
| Protein allergens based on auto and cross-covariance (ACC) transformation | AllerTOP v2.0 |
| Theoretical pI and molecular weight | ProtParam [139] |
| Existence of any signal peptide | SignalP4.1 [140] |
| Secondary structure. | SOPMA [44] |
| 3D structure prediction | I-TASSER [141] |
| Protein structure refining from Ca traces | ModRefiner [142] |
| 3D structure validation | ProSA-web, [143] The ERRAT server [144] |
| Ramachandran plot | ProCheck server [145] |
| TLR2 and TLR4 Crystal structures | PDB [146] |
| TLR2 and TLR4 molecular interactions | ClusPro 2.0 server [147] |
| Molecular interaction visualization | PyMOL [148] |
| Immunogenicity | C-ImmSim server [149] |
| Hub residues | CytoHubba [150] |
| Predict potential T cell epitopes | NetMHC [151] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



