
Submitted:
30 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
Post-operative pain is the unpleasant sensory and emotional experience after surgery, its origin being both the inflammatory reaction induced by the surgical trauma on the abdominal wall, and the splanchnic pain, induced by the activation of nociceptors of the viscera, which are highly sensi-tive to distension, ischemia and inflammation. Nowadays, it is well recognized that there is a close relationship between the gut microbiome and pain perception, and that microbiome is highly af-fected by both anaesthesia and surgical manipulation. Thus, efforts to restore the disturbed micro-biome by supplementation with beneficial bacteria, namely probiotics, seems to be effective. In this article the knowledge gained mainly from experimental research on this topic is analyzed, the con-cluding message being that each probiotic strain works in its own way towards pain relief.
Keywords:
probiotics
; post-operative pain
; microbiome
; visceral pain
; abdominal wall pain
; inflammation
; opioids
; cannabinoids
1. Introduction
The word “pain” is thought to have come from the word “ponos,” the modern Greek word for pain, which originally meant labour and hard work in the Homeric epics. This meaning may have originated from the personification of “ponos” (toil) and “algea” (sorrows) as children of the hard-hearted goddess “Eris” (Strife) in Greek mythology. [Ἔρις στυγερὴ τέκε μὲν Πόνον ἀλγινόεντα,Λήθην τε Λιμόν τε καὶ Ἄλγεα δακρυόεντα] [Hesiodus,Theogonia, 0020.001] [1]. Ponos is the name of the Greek goddess of revenge, sent to punish mortal fools who had angered the Olympian gods because they had accepted and used the fire stolen by Prometheus from Mount Olympus. A similar belief about pain as a revenge of the goddess was also common in many ancient cultures. Similarly, in The Bible, when Adam and Eva were banished from Eden, God’s punishment was for them to experience pain and for Eva to be subjected to the severe intermittent pain of birth [“odynes”].
Aulus Cornelius Celsus [25 BC-50 AD] in his famous medical work De Medicina was the first to record the cardinal signs of inflammation known as the "Celsus tetrad of inflammation": calor (warmth), dolor (pain), tumor (swelling) and rubor (redness and hyperaemia), while he also goes into great detail regarding the preparation of numerous ancient medicinal remedies including opioids, also greatly valued by Hippocrates [460-370 BC] for the treatment of pain, most frequently recognized as “algos” [άλγος], “algema” [άλγημα], “odyne” [οδύνη], and “ponos” [πóνοσ] [2].
The Greeks and Romans were the first to inspire the “theory of sensation”, that is the brain and nervous system intervene in the perception of pain. But it was not until the Middle Ages, and well into the Renaissance - the 1400s and 1500s - that evidence began to accumulate to support these theories. Leonardo da Vinci and his contemporaries came to believe that the brain was the central organ responsible for sensation and that the spinal cord transmits sensations to the brain [2].
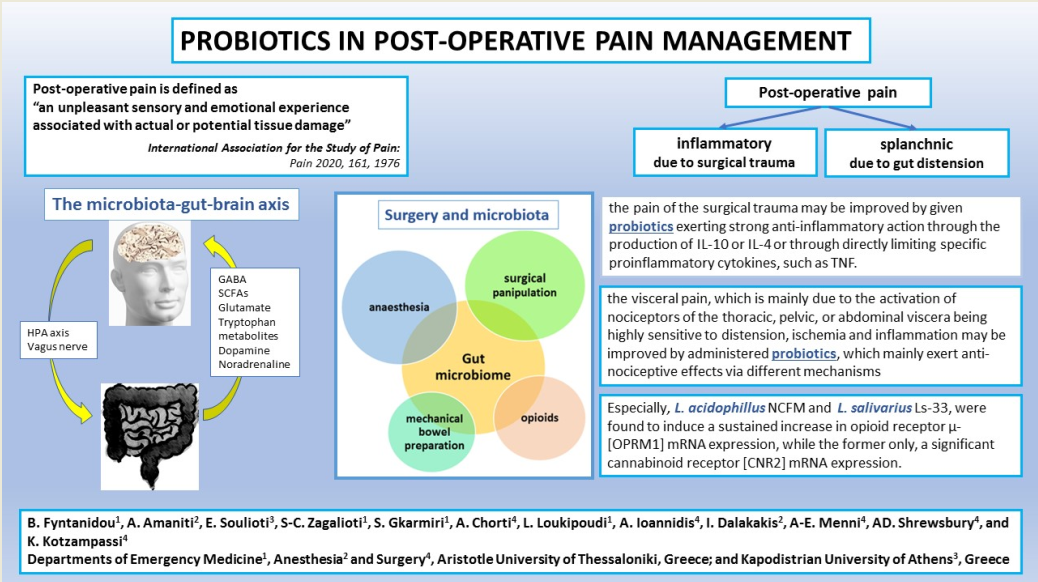
Today the International Association for the Study of Pain has updated the definition of pain to “an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage” [3]. However, in parallel with the ongoing knowledge of the interference by the body’s overall microbiome in almost any disease pathophysiology, there is an augmented literature on the implication of the gut microbiome in pain perception. The close association of the gut microbiome, and more specifically of the low-diversity microbiome or pathobiome, with functional bowel diseases in the form of “chicken and egg” has been well recognized for years. Only in recent decades has the cluster of beneficial bacteria belonging to the microbiome, i.e. probiotics, begun to be recognized as implicated in the manipulation of the damaged microbiome, to serve as an “alternative” treatment of high effectiveness.
The purpose of this narrated review is to present the current knowledge, mainly derived from animal experiments, in relation to postoperative pain management, by means of specific probiotic strains. For an overall understanding of the true implication of the microbiome in pain perception and visa-versa, we first look at the known association between the microbiome and pain and then to the alterations to the gut microbiome as a consequence of anaesthesia and surgery.
2. Pain and the Gut Microbiome
The gut microbiota typically hosts thousands of bacterial species, as well as human viruses, fungi, and bacteriophages [4,5,6]. This intricate ecosystem, which is continually reshaped by the host and its environment, and, at the same time, affecting its function and health, is now defined as “microbiome”, the term including both the microbiota within its well-defined habitat and all the bacterial and external structural parts, including their genes [7,8].
Microbiota generally live in a significant relationship, called homeostasis, with the host-gut, the interactions regulated by the integral gut barrier and the immune system [9,10]. On the other hand, the gut bidirectionally communicates with the central nervous system via immunological, hormonal, and neural pathways, the complex interaction being named the gut–brain axis [11]. Today, when everyone recognizes the microbiome as an integral part of the gut, both in the mucosa and the lumen, influencing the bidirectional signaling pathways between the central nervous system and the gut, this gut–brain axis is now called as the microbiome-gut-brain axis [12,13,14,15,16].
In this context, the gut microbiome can synthesize and secrete various neurotransmitters and neuro-modulatory bacterial metabolic end-products, including short chain fatty acids (SCFAs), tryptophan metabolites, gamma aminobutyric acid (GABA), glutamate, dopamine, and noradrenaline [17,18,19,20,21]. In physiological conditions, the blood–brain barrier permits the passage of tryptophan metabolites and SCFAs into the central-nervous system, but not allows the circulating neurotransmitters, excluding GABA [21,22,23,24]. Disruption of the blood–brain barrier, due to whatever pathology, permits an increased amount of the circulating neurotransmitters to enter the brain. Injection of butyrate, the main SCFA produced from certain microbial strains, in rat and mouse brains was revealed to stimulate the release of the brain-derived neurotrophic factor favoring neurogenesis. Additionally, butyrate is considered to exert regulatory effects on inflammation-induced visceral pain [24,25].
Bacteria such as Prevotella, Fusobacterium, Enterococcus casseliflavus, Escherichia, and Bacteroides were found to produce tryptophan, which, by passing the blood–brain barrier, allows serotoninergic neuro-transmission in the brain [26]. Glutamate, produced by certain microbial strains, is also a major neurotransmitter in the central nervous system, which also regulates gut sensory and motor functions in the enteral nervous system [27,28,29,30]. In cases of dysbiosis due to stress, the expression of glutamate receptor is altered, while, in dysbiosis due to antibiotics treatment, in mice, the decreased levels of hippocampal N-methyl-D-aspartate receptor and brain-derived neurotrophic factor were able to be restored by probiotic/prebiotic treatment [31,32,33,34].
Early studies have associated the altered structure of the gut microbiome [decrease in Bifidobacterium, Faecalibacterium and Faecalibacterium prausnitzii, and increase in Lactobacillaceae, Bacteroides, and Enterobacteriaceae] with human digestive tract diseases [35,36]. Nowadays, there is augmented evidence that gut microbiota is one of the most important pain modulators, working towards pain regulation in both the central and peripheral nervous systems. This evidence is much more pronounced in the cases of visceral or abdominal pain [37,38], such as the irritable bowel syndrome, for which we know not only that the diversity of the microbiome is directly related to the diagnosis, but also with its phenotypic subtypes, i.e. predominantly diarrhea or constipation, as well as with the severity syndrome and its flare-ups [39,40,41]. There are also significant indications for inflammatory pain, migraine, joint pain, neuropathic pain and opioid tolerance [15].
The fundamental role of the gut microbiota has been confirmed in mice, having undergone spared nerve injury, who then received fecal transplantation from pain-free donors or from patients suffering chronic postoperative pain. Mechanical thresholds were found significantly decreased after receiving feces from pain-suffering patients rather than from pain-free donors, and was accompanied by a significant reduction of the peroxisome proliferator-activated receptor γ [PPAR γ] levels, a key metabolism modulator implicated in pain and a trend toward an increase in activated microglia in the spinal cord [35,42,43,44].
In another set of experiments in germ-free mice subjected to systemic LPS or ischemia/reperfusion injury, Amaral et al found that the lack of microbiota was followed by inflammatory hypo-responsiveness, mediated by IL-10, via inhibition of the inflammatory cytokines release, and of the expression of COX-2, since IL-10 down-modulates hyper-nociception, that is a state of increased sensation to painful stimuli, or, in other words, a decrease in the behavioral nociceptive threshold [45]. Microbiota transplantation thereafter, was followed by loss of the ability to produce IL-10 and a re-gained ability to develop inflammation in response to diverse stimulation. All the above imply a critical role in the interaction between the host and the commensal bacteria, in favoring adaptation to environmental stresses, including those which release pain.
Microbiome analysis in fecal samples from patients suffering chronic pelvic pain syndrome and healthy controls was correlated with the NIH-Chronic Prostatitis Symptom Index. Patients have significantly decreased α-diversity and a wider clustering in relation to controls, who revealed tighter clustering in a space distinct from the patients. Additionally, compared to controls, 3 taxa were over-represented and 12 were under-represented, as occurs with Prevotella [46].
In a recent clinical study, gut microbiota diversity and abundance were analyzed in 20 patients undergoing surgical fixation of distal radius fracture under axillary brachial plexus block. The results then correlated with [i] the verbal pain rating scale; [ii] the level of pain deemed “acceptable” by the patient; [iii] the 24 h max self-reported pain score; and [iv] the 7-days analgesic consumption. The major finding was that the 7-days analgesic consumption was inversely correlated with the Shannon index of α-diversity, known to be decreased in certain pain conditions [47]. Main perception was found associated with the abundance of certain microbial genera, including Collinsella, previously recognized to influence the production of the pro-inflammatory cytokine IL-17A, to increase gut permeability and disease severity in experimental arthritis [48]. In patients whose pain was perceived as “not acceptable”, a greater abundance of Lachnospira and Alistipes was prominent, while the analgesic consumption was positively correlated with abundance of Dialister, previously shown to correlate with ankylosing spondylitis [49]. On the other hand, Porphyromonas was more abundant in those with acceptable levels of pain, its anti-nociceptive action having been shown previously in a preclinical model [46,50,51].
3. Surgical Operation and the Microbiome
For the needs of the present analysis, the term "surgical operation" includes all those manipulations that precede the main surgical time of a programmed laparotomy; i.e. the procedure of pre-operative mechanical bowel cleansing, when required, and the administration of anesthesia/ analgesia. However, in cases of emergency surgery, the medical event leading to the operating theater must also be taken into account - severe trauma, gut perforation, or gut obstruction, being the most common - as seriously damaging the body and its microbiome. The possibility of pre-operative food deprivation as well as the administration of antibiotics and, perhaps, opioids for pain management must not be overlooked [52,53,54,55,56,57].
Mechanical bowel preparation aimed at reducing fecal materials and bacterial count with the objective to decrease surgical site infection rate, including anastomosis leakage, has been a common practice for more than a century. However, cleansing preparations osmotically increase the volume of water within the gut, washing out the lumenal contents, including gut bacteria. Additionally, the rapid increase in bowel movements will flush out bacteria incapable of adhering to the gut mucosa, distorting the faecal microbiota composition [58,59,60,61].
By approaching the issue of bowel cleansing from the perspective of the gut microbiome, an early randomized controlled trial reports a significant decrease in the total bacteria number of the Clostridium coccoides group, the Clostridium leptum subgroup, Bifidobacteria, Lactobacillus and Enterobacteriaceae; but no effect on Enterococci and Staphylococci [62]. Jalanka et al, in feces samples collected immediately after bowel cleansing, reported a reduction in the number of bacteria in relation to baseline, and the number of methanogenic archaea per gram of feces to be decreased by 20-fold [58]. Specifically, they found a decrease in the members of Clostridium cluster IV; an increase in the Clostridium cluster XIVa and Proteobacteria, Fusobacteria and bacteria related to Dorea formicigenerans, all returning to baseline only after the 14- and 28-day. More recent studies have documented these findings: at the phylum level, mechanical cleansing resulted in a reduction in the relative abundance of Firmicutes, and increase of Proteobacteria, both restored one month later; and at class level, an increase of γ-Proteobacteria and Coriobacteria, and significantly reduced Clostridia. Finally, at family level, a significant reduction of Lactobacilli and a rise in Enterobacteriaceae were found to have persisted one month later [63,64].
Regarding the procedure of anaesthesia, it is now recognized that it could provoke perioperative neurocognitive dysfunction, attributable to modifications in the gut microbiome. The use of certain general anesthetics could result in a depletion of microbial α-diversity, and of neurologically relevant metabolite variations. Most of our knowledge on the association between anaesthetics and microbiota has been derived from studies testing different anaesthetics to inhibit infections after surgery [65], their findings supporting a wide range of effects, including phenotype modifications, the changes seeming depend mostly on the duration of anaesthesia [66,67]. Isoflurane and sevoflurane given in vitro, but not propofol, were found to increase motility and biofilm formation of pathogenic E. coli, S. aureus, E. faecalis, and P. aeruginosa, both changes being associated with increased bacterial virulence, but also being drug dependent [68]. Moreover, Maier et al. have evaluated the effect of different drugs, including propofol and etomidate, on the growth of the 40 most common bacteria in the human gut. Etomidate, but not Propofol, showed significant antibacterial activity against Ruminococcus bromii, Roseburia intestinalis, Ruminococcus torques, and Blautia obeum [69].
The association between anaesthetics and gut microbiota is now clarified by their shared affinity for neurotransmitters, which empower the anaesthetics to directly interact with prokaryotes. Additionally, acute variations in the levels of neuro-transmitters due to anaesthesia can lead to transitory changes in the gut mucosa, including the postoperative gastrointestinal tract dysfunctions, commonly observed in abdominal surgery patients [70,71,72]. The impact of anaesthesia on the gut milieu have also been considered an essential trigger of neuro-inflammation and development of cognitive dysfunction [73,74,75,76].
Recent experimental models have demonstrated a significant alteration in the gut microbiome after exposure to anaesthetics, even after a short-term exposure. Moreover, these studies suggest bidirectional interactions: alterations in the composition of the microbiota after anaesthesia, and of the altered microbiome on the postoperative outcomes, including postoperative pain and postoperative cognitive dysfunction; both being prominent in early studies in humans, showing a close relationship between microbiota composition and postoperative outcomes, including pain and delirium [35]. Specifically, Han et al. showed a strong perturbation in fecal microbiome following sevoflurane anaesthesia in mice, the changes lasting for more than 14 days: the genus Bacteroides, Alloprevotella, and Akkermansia remained significantly elevated in relation to baseline and did not return to their original levels [77]. Similarly, the genus Lactobacillus remained significantly reduced even after two weeks. In the same manner, the intravenously given propofol for 3 hours in rats induced gut microbiota changes: a reduction of Prevotella, Alloprevotella and Lactobacillus, showing a trend to recover two weeks later [78].
Furthermore, Liufu et al found in mice that a 1.4% isoflurane anesthesia of two hours duration, in conjunction with a ten-minute surgery [extended laparotomy and immediate closure] – which potentiates the anesthesia neurotoxicity – could cause changes in gut microbiota, being age-dependent [79,80,81,82]. Specifically, significant alterations in the composition of the gut microbiota were found in older [18 months], but not in 9 months mice. However, there was different microbiota diversity in the two age groups: the reduction rate in the abundance of Lactobacillus at genus and species levels being, generally, more pronounced in the older mice, and especially of Lactobacillus salivarius; which also had a longer recovery time of more that 11 days. However, it should be noticed that a 2.5% lidocaine plus 2.5% prilocaine local anaesthesia cream was applied to the incision site, 3-times daily up to euthanasia, to alleviate pain, and thus avoid the interference of pain in microbiome alterations [80,81,83].
Regarding the consequences of surgical manipulation on the gut microbiome, it has long been accepted that operational stress can induce alterations in the intestinal bacteria via corticotropin-mediated routes, which can activate systemic inflammatory responses [52,84,85,86]. Surgery represents a form of controlled trauma, which is an established source of tissue injury, and every injury is a key driver of inflammation [87]; the cellular damage triggering endogenous factors, the damage-associated molecular patterns, which activate immune cells to resolve the damage and restore homeostasis [88,89]. However, the physical trauma of surgery itself, in cooperation with the surgery-induced alterations in tissue oxygenation, perfusion and pH and augmented by the stress of anesthesia and the intraoperative pharmacological manipulations, have long been shown to cause gut microbial dysbiosis [90,91]. In colorectal surgery patients, when comparing pre- and post-operative stool samples a significant decrease in total bacterial counts and in the numbers in six groups of obligate anaerobes were prominent [92], while the populations of Enterobacteriaceae, Enterococcus, Staphylococcus and Pseudomonas were significantly increased [93].
A systematic review of 10 studies scoping the relationship between the gut microbiome and postoperative complications has suggested that complications might be influenced by the patients’ gut microbiota [94]. Surgical procedures of different types tend to result in an increase in potentially pathogenic bacteria such as Pseudomonas, Staphylococcus and Enterococci and a decrease in Lactobacilli and Bifidobacteria [95,96,97].
An early study by Shogan et al in rats subjected to colon resection and anastomosis, as is usually performed in colon cancer patients, revealed that colorectal surgery does not affect the bacteria of the lumenal contents (stool), but induces significant alterations in the composition of those being adherent to the gut epithelium [98]. Most of the changes were observed in Proteobacteria, Actinobacteria, and Firmicutes: among Proteobacteria, the relative abundance of Escherichia/Shigella increased 200-fold; in Actinobacteria, the relative abundance of an uncultured bacterium increased approximately 50-fold; and, non-uniform changes among the genera Firmicutes were also prominent. On the other hand, the relative abundance of Ruminococcaceae and Clostridia decreased 20-fold, of Allobaculum and Coprococcus increased 10-fold, and of Enterococcus 500-fold. Finally, a 20-fold decrease of Prevotellaceae clearly suggests that Bacteroidetes were also affected by anastomotic injury [98].
In a recent review, Agnes et al questioned “How surgery affects the gut microbiome?”, in order to analyze the different factors involved. Besides the operational stress and the disruption of tissue homeostasis, the authors considered that many species of gut microbiota, being either facultative or obligate anaerobes, when exposed to oxygen during colon surgery, could be subjected to a significant effect on their species; in other words, a reduction in “good” obligate anaerobes, such as some Bacteroides, and an increase in “bad” facultative anaerobes, such as Enterococcus could occur [86,98]. Another cause of decrease could also be tissue ischemia or hypoperfusion, due to permanent or temporary occlusion of gut vessels. In an ischemia/reperfusion model in rats, a 30min of gut ischemia followed by reperfusion resulted in changes in the gut microbiota at one hour of reperfusion, reaching the peak at 6h and then progressively recovering: E. coli and Prevotella fist and Lactobacillus later [99,100,101].
However, the extent of surgery-induced gut microbiome depletion is also related to or affected by different factors, such as the length of surgical line incision or the type of suture used [79,102]. Although there are no references as to the exact relationship between the gut microbiome and the degree of inflammatory reaction caused by surgery, it is clear that less abdominal wall trauma as occurs in laparoscopy versus laparotomy and/or less inflammation as occurs by using absorbable sutures or other implanted material should accelerate less aseptic inflammation and thus less pain and less catabolism [103]. The catabolic modifications that develop mainly in muscles to provide energy and essential aminoacids to improve repair of damaged tissues and wound healing and to maintain the function of the critical organs must not be overlooked [52,104,105]. However, catabolic hormones, such as catecholamines and cortisol are known to increase, proportional to the extent of trauma of any type and etiology. It is also known that at least noradrenaline can interact with Escherichia coli and the Pseudomonas aeruginosa quorum sensing receptors and activate the transcription of some microbial genes to virulent phenotypes [65,106]. The same cell-to-cell signaling mechanism that allows the release and detection of extracellular signals, called quorum sensing, has also been recognized after 5-HT, substance P, or epinephrine release, which all increase the motility of the virulent phenotype of Pseudomonas fluorescens [107,108].
Finally, of importance is the bidirectional interaction among opioids, opioid receptors, and the microbiome: opioids were found to modulate the diversity of the gut microbiome, and that, in turn, affects the host response to opioids. Morphine given to mice resulted in significant alterations of the gut microbiome, evident as early as 1-day after administration, but were reversible by the co-administration of the μ-receptor antagonist naltrexone [109]. More interestingly, the host response to morphine was dependent on its microbiome: in an experimental model of morphine tolerance, naive mice were more prone to developing tolerance than germ-free and antibiotic-treated mice [110]. Moreover, the tolerance was found related to the decrease of Bifidobacteria and Lactobacillaeae, which, when “re-enter” in the gut by means of fecal transplantation, re-established opioid tolerance in this model [35,65].
4. Probiotics in post-operative pain management
Based on the above, we may say that the effort to control or reduce postoperative pain, by any means, has two simultaneous approaches: one relating to the pain of the surgical trauma on the abdominal wall which is purely of an inflammatory etiology, and two, the visceral pain, which is due to overdistension of the intestine before its mobilization from the paralytic ileus, resulting from anaesthesia.
According to the current definition “probiotics are live microorganisms that, when administered in adequate amounts, confer a health effect on the host” [111]. Today, many in vitro and in vivo studies have shown that probiotics exert potential suppressive effects on various infectious, and inflammatory conditions, thus significantly shortening the inflammatory phase of the wound healing process [112,113,114]. At the same time, there are quite a lot of clinical studies referring to the reduction of postoperative bloating and the shortening of the time-duration up to the first flatus, all indirectly indicating the shortening of the post-operative paralytic ileus [115,116,117]. Both the above may not have a direct, suppressive, effect on the pain originating from the abdominal wall trauma, or from the distension of the gut wall, but there are many studies documenting the specific cytokines and other molecules changes, which point in that direction.
4.1. Probiotics in relation to the inflammation-induced pain of surgical trauma
Surgical incisional trauma causes inflammatory reaction as part of the healing process. Inflammation seems to contribute to the sensitization of peripheral nociceptors, leading to hyperalgesia or allodynia, when noxious or non-noxious stimuli cause pain [118]. Inflammation reduces the pain threshold of nociceptors and increases the individual pain response. Post-operative pain is exacerbated by the released pro-inflammatory cytokines, not only locally but in the spinal dorsal horn tissues, which are further involved in the pain mechanism, the magnitude of pain being proportionally dependent on the incision length and the surgical manipulations [102,119,120]. The released chemo- and cytokines trigger the activation of the intracellular downstream signal pathways and, subsequently, to phosphorylation of receptors and ion channels, in primary sensory neurons. This whole process leads to neuronal hyperexcitability and peripheral sensitization [15]. Furthermore, it is known that skin-deep incisions induce mechanical and heat hypersensitivity similar to incisions extended deeper than the skin; thus, cutaneous trauma drive the most of post-operative pain and, consequently, the most of the inflammatory process, or the opposite [121,122,123].
Probiotic species seem capable to modulate the gut microbiota to prevent or alleviate the inflammatory pain, the beneficial effects being the consequence of their multifaceted action: to control pro-inflammatory signaling via increased expression of anti-inflammatory cytokines, or by directly limiting the bioavailability of specific pro-inflammatory cytokines [112,124]. The anti-inflammatory IL-10, binds to the IL-10 receptor, with the IL-10R1 being the high affinity sub-unit necessary for signal transduction, expressed by astrocytes, microglia, endothelial cells, and trafficking leukocytes [125]. Its anti-inflammatory action is accelerated by blocking downstream signaling of TLR4, which normally promote a pro-inflammatory milieu, critical for the induction of neuropathic pain [126]. Additionally, IL-10 signaling also induces the production of the suppressors of cytokine signaling 1 and 3, thereby further reducing pro-inflammatory cytokine production by targeting the p65 NF-κB subunit [127]. This effect has also been confirmed in a clinical study on colorectal surgery patients treated with the four-probiotic regime L. plantarum, L. acidophilus LA-5, B. lactis BB-12 and S. boulardii [115].
Diabetic rats subjected to a dorsal wound and topically treated with Lactiplantibacillus plantarum presented a significantly increased expression of the anti-inflammatory mediators IL-10 and TGF1 to day 14, in parallel with a markedly decreased expression of the pro-inflammatory IL-1 and TNF from day 7 and thereafter, in comparison to control-treated rats. These findings were consistent to the speeding up of wound closure from day 3 and thereafter [128]. Staphylococcus aureus-infected excisional wounds treated with L. plantarum MTCC 2621 exhibited up-regulation of IL-10 in the later phase of the healing process in relation to controls, in parallel with earlier re-epithelization, accompanied by a reduction of leukocytes infiltration, and increased activity of fibroblasts and deposition of collagen [129]. Other probiotic bacteria, such as L. rhamnosus UBLR-58, L. acidophilus LA-5, L. fermentum SGL10, L. brevis GQ4237768, L. brevis SGL 12, L. paracasei SGL 04, and B. longum UBBL-64 also exert anti-inflammatory action by means of similar mechanisms, but to a significantly lesser degree, in relation to L. plantarum [130,131,132].
TNF is considered of particular value as a marker of inflammation, due to its close relationship to the prostaglandin release implicated in pain, swelling and fever [133]. In an ex-vivo human skin explant model, the administration of the postbiotic B. longum Reuter lysate led to an improvement in parameters relating to inflammation, but mainly it significantly inhibited capsaicin-induced CGRP release by neurons. Similarly, the same regime in humans led to increased skin resistance to physical and chemical aggression in relation to control [134].
Lactobacillus paracasei MSMC39 or placebo given as mouth wash in 30 patients who had had an impacted mandibular third molar removed, was found to significantly reduce the TNF level in gingival crevicular fluid, in relation to placebo. Pain, swelling and trismus, evaluated in a visual analog scale score, were also reduced, but not to a statistically significant degree [135]. Thirty-eight patients subjected to third molar surgery were randomized to Levilactobacillus brevis CECT7480 and Lactoplantibacillus plantarum CECT7481 or control regime for one week. Probiotics were found to significantly reduce pain and eating difficulty scores on the 5th post-operative day and thereafter. The infection rate and swelling values showed no significant difference between probiotics and placebo at any time point [136]. [Table 1].
A total of 283 patients with a single rib fracture were allocated to either Lactobacillus casei Shirota in skimmed milk or placebo daily for a month following the fracture. Pain relief was assessed during activities specifically to elicit pain, while sleep quality and maximal inspiration lung volumes were also examined. Probiotic treatment was found to be more effective in relieving pain during deep breathing, coughing and turning over, while patients also had increased inspiration volumes - sleep quality being unaffected [137]. Contrary to these positive findings, Lactobacillus reuteri LR06 or Bifidobacterium longum BL5b, given in the drinking water in a rat model of inflammatory pain by means of injection of complete Freund's adjuvant into the left hind paw, were found to have no antinociceptive effect. This was assessed by means of mechanical and thermal hyperalgesia, while Iba1 was used to verify the microglial inflammatory activation in relation to placebo [138].
There is also evidence that probiotic beneficial bacteria may affect inflammatory cytokine levels by acting on the NF-κB and mitogen-activated protein kinases [MAPK] signaling pathways [139]. L. rhamnosus GG ATCC 53103 was found to modulate signal transduction pathways by triggering MAPKs, while L. acidophilus LA-5 were found showing higher values in all MAPKs analyzed [140,141].
Finally, besides anti-inflammatory cytokines, microbiota also produce neuro-transmitters, which is able to alter pain signaling. Lactobacillus spp., B. dentium and Bifidobacterium spp. produce GABA, the most important inhibitory neuro-transmitter in the brain, through enzymatic decarboxylation of glutamate. GABA binds to its receptors on the surface of DRG neurons, resulting to their depolarization, thus inhibiting the nociceptive transmission [142].
4.2. Probiotics in relation to gut distension-induced visceral pain
Visceral or splanchnic pain results from the activation of nociceptors of the thoracic, pelvic, or abdominal organs. Splanchnic organs are highly sensitive to distension, ischemia and inflammation, but are relatively insensitive to other stimuli that normally evoke pain, such as cutting or burning [16,38]. Thus, the colorectal distention model – although scheduled for inflammatory bowel disease or irritable bowel syndrome simulation – could be considered a model characterized by visceral pain associated with altered transit time, as occurs in the post-operative situation of paralytic ileus, before the gut starts to move again.
One of the basic findings correlating probiotic strains with pain relief is that documented by Rousseaux et al. They initially hypothesize that some probiotics may stimulate the expression of receptors on epithelial cells that locally control the transmission of nociceptive stimuli to the gut nervous system. After testing different probiotic bacteria, they found that both L. acidophillus NCFM and L. salivarius Ls-33 induced a sustained increase in opioid receptor μ- [OPRM1] mRNA expression in human HT-29 epithelial cells, but only the former, either live or heat-killed, was able to also induce significant cannabinoid receptor [CNR2] mRNA expression. Then, they conducted the same experiment in mice and rats, orally administering live L. acidophillus NCFM strain for 15 days, and found the same receptors expressed in 25% to 60% of epithelial cells as against 0% to 20% in placebo treated animals. Additionally, in treated rats the mean colorectal distention required to induce pain was 20% more than the 50±2mm Hg of controls [143].
Similar results were obtained in humans. Twenty women experiencing mild to moderate abdominal pain were enrolled to receive either L. acidophilus NCFM or this in combination with Bifidobacterium lactis Bi-07 for 21 days. L. acidophilus alone, but not with Bifidobacterium, induced colonic opioid receptor μ- [MOR] mRNA and protein expression. In contrast, cannabinoid receptor [CNR2] mRNA expression was decreased, as seen in colonic biopsies obtained before and at the end of treatment. Both treatment groups trended towards improvement in symptoms, but not significantly [144].
In a colorectal distention model, the commercially available fermented dairy product containing Bifidobacterium lactis CNCM I-2494 and Lactcoccus lactis CNCM I-1631 and two common yogurt starters, Streptococcus thermophilus and Lactobacillus bulgaricus, was found to reduce stress-induced visceral hypersensitivity and pain. Of interest, the anti-nociceptive effect of fermented dairy products was dose-dependent and generally more effective than B. lactis CNCM I-2494 strain alone, at the same dose, findings supporting a synergistic interplay. In detail, the anti-nociceptive action of B. lactis CNCM I-2494, when given as a single dose per day, was clear at a dose of 1010 CFU while, when in co-administration with the fermented dairy product at the lower dose of 108 CFU [145].
In a model of colonic visceral hypersensitivity induced by infusion of 2,4,6-trinitro-benzene-sulfonic acid into the proximal colon, the 8-day probiotic Lactobacillus rhamnosus Lcr35 treatment produces an anti-hypersensitivity activity. As this model is known to involved an increase in IL-13 secretion, it may be that the action of probiotic treatment involves the regulation of the local IL-13/Th17 immune activation [146].
Colonic visceral hypersensitivity of inflammatory origin, occurring in rats after intracolonic instillation of zymosan, was found to be significantly attenuated by Lactobacillus rhamnosus GG ATCC53103, the viscero-motor response for grading colorectal distension being determined by measurement of the electro-myographic activity of the abdominal external oblique muscles. The levels of neuro-transmitters serotonin, noradrenaline, and dopamine and biogenic amines quantified in the frontal cortex, subcortex, brain stem, and cerebellum were also found significantly altered in L. rhamnosus-treated rats, suggesting that they could be involved in pain modulation [147,148].
Rats exposed to maternal separation, from weaning onwards, were given drinking water with or without supplementation of Lactobacillus rhamnosus GG soluble mediators. Maternal separation followed by restraint stress in adulthood led to increased splanchnic sensitivity and corticosterone plasma levels, as well as to alterations in β-diversity and abundance of specific bacteria, including parabacteroides - all these effects being ameliorated through L. rhamnosus GG soluble mediator supplementation [149].
Intrarectally given butyrate enemas effected reduction in pain perception and discomfort, when given to healthy volunteers [150]. Based on this knowledge, Roseburia hominis, a species of Lachnospiraceae family, well known to consume lactate and acetate to synthesize butyrate via different pathways, was administered orally in rats subjected to water avoidance stress [151]. Rats subjected to stress but having received placebo treatment exhibited a reduced representation of the pathways involved in the metabolism of butyrate and a reduced abundance of several operational taxonomic units associated with butyrate-producing bacteria, such as Lachnospiraceae. However, Roseburia hominis treatment led to alleviation of visceral pain perception and hypersensitivity, as well as to an increase in cecal butyrate concentration [152].
On the other hand, in a rat model of colonic hypersensitivity elicited by butyrate enemas, the hypersensitivity improved after administration of Lactobacillus acidophilus NCFM strain, since treatment increases the colorectal distention threshold by 44% compared to untreated rats. It is very important to underline the fact that the anti-nociceptive effect of L. acidophillus was similar to that elicited by the subcutaneous injection of 1mg/Kg body weight morphine, and it enhanced by 65% the suboptimal analgesic effects of 0.1mg/Kg morphine. Furthermore, L. acidophillus-induced analgesia was significantly inhibited by an intraperitoneally given dose of the cannabinoid receptor 2 [CB2] selective antagonist AM-630, but not by the opioid receptor antagonist naloxone methiodide, thus given indirect evidence for the physiological role of CB2 in the regulation of splanchnic pain [143].
The combination of Bifidobacterium longum and Lactobacillus helveticus, as pretreatment, in mice subjected to water avoidance stress, significantly reduced the chronic stress-induced visceral hypersensitivity, in comparison to a single one. Furthermore, the combination of L. helveticus and B. longum was superior in regulating glucocorticoid negative feedback on the HPA axis [153].
The same authors, having previously reported that Lactobacillus farciminis suppresses stress-induced hypersensitivity in response to colorectal distension, evaluated whether this anti-nociceptive result is related with changes in neuronal activation at spinal and supraspinal sites induced by L. farciminis strain. The neuronal activation was assessed by means of Fos protein expression, which is a marker of neuronal activation, rapidly expressed in neurons of the central nervous system in response to somato-cutaneous or splanchnic sensory stimuli. They then assessed whether restrain stress-induced splanchnic hyperalgesia in rats modifies Fos protein expression provoked by colorectal distention and whether this expression can be modulated by L. farciminis. After colorectal distention or restrain stress, Fos expression was found increased in the sacral spinal cord, in the nucleus tractus solitarius, the hypothalamus paraventricular nucleus, and in the medial nucleus of the amygdala. When both stimuli were applied, Fos was overexpressed in the sacral spinal cord section, in the paraventricular nucleus and the medial nucleus of the amygdala, but not in nucleus tractus solitarius. L. farciminis pretreatment largely reduced the Fos positive cells in all these areas, suggesting that its anti-nociceptive effect is the result of the reduction in the activation of sensory neurons in the spinal and supraspinal level due to stress [154,155].
The implication of probiota in the modulation of the gut–brain axis and the HPA axis was further documented after an oral administration of L. plantarum PS128 in rats for a 14-d period. L. plantarum PS128 was found to inhibit the 5-hydroxytryptophan-induced visceral hypersensitivity during colorectal distention, the effect followed by decreased serum corticosterone, decreased neurotransmitter protein [substance P, CGRP, BDNF, and NGF] in the spinal cord, and increase of the glucocorticoid receptor and decrease of mineralocorticoid receptor in the amygdala. These findings suggest that L. plantarum PS128 decreased splanchnic hypersensitivity through modulating the gut–brain axis and the HPA axis [156].
Faecalibacterium prausnitzii is an extremely oxygen-sensitive commensal butyrate-producer bacterium, populating the most anaerobic parts of GI tract of mammals [157]. In a neonatal-maternal-separation induced stress model, a decrease in Faecalibacterium prausnitzii has been confirmed. This is also evidence in several intestinal disorders, including that of colon anastomosis leakage, after colon surgery, the F. prausnitzii being generally recognized as a biomarker of intestinal health [157,158,159]. In two experimental models of chronic [neonatal-maternal-separation] and acute stress [partial restraint stress] colorectal distension was applied, for colonic hypersensitivity induction; then the F. prausnitzii A2-165 strain, or its supernatant were tested [157]. Both demonstrated anti-inflammatory properties, as also seen by others; however, only the F. prausnitzii decreased colonic sensitivity and exhibited curative anti-nociceptive properties in response to a colorectal distension [157,160,161]. The authors suggest that the F. prausnitzii-induced anti-nociceptive properties are possibly correlated to intestinal epithelial barrier enhancement, since others support the option that it can modulate tight junctions in animal models of low grade or of acute inflammation [157,162,163].
One of the fundamental receptors responsible for pain perception in the intestine is the transient receptor potential vanilloid 1 [TRPV1], which is a member of the vanilloid receptor family [164,165,166]. It is expressed in spinal and vagal primary afferent neurons and activated by capsaicin, noxious heat, acidosis, depolarization and endovanilloids [164,166,167]. In a distension-dependent gut pain model the effectiveness of Lactobacillus reuteri DSM 17938 was tested. In a dose-depended manner, L. reuteri DSM 17938 was found to reduce the jejunal spinal nerve firing evoked by distension; 80% of this reduction being inhibited by the TRPV1 channel antagonist, which mediates nociceptive signals [168]. In order to further confirm the effect of L. reuteri on TRPV1, the authors used a murine jejunal mesenteric nerve bundles model in which capsaicin was applied on serosa; capsaicin induces intracellular calcium in dorsal root ganglion [DRG] neurons but L. reuteri DSM 17938 inhibits it, as assessed by means of Ca2+ fluorescence intensity [168]. Furthermore, the L. reuteri DSM anti-nociceptive effects, when given by gavage, were tested on gastric distension in rats: L. reuteri DSM as pre-treatment was found to inhibit bradycardia induction after the painful gastric distension [168]. Finally, it has recently found that both isoflurane inhalation and ketamine intravenously affect acetylcholine-activated TRPC4 channels, which considerably inhibit the muscarinic cation current in ileal myocytes, even when G proteins are activated directly by intracellular GTPγS, i.e., by-passing muscarinic receptors, thus seriously implicated in anaesthesia-induced postoperative ileus [169,170].
The same strain of Lactobacillus reuteri DSM 17938, given orally in a dose of 108 CFU for 21 days, was reported to be effective in improving symptoms of infantile colic and thus reduce the crying time [171]. Previous research had revealed that the gut bacteria in colicky infants varies in relation to healthy ones, exhibiting a reduction in bacterial richness and specifically in Lactobacillus and Bifidobacterium genera, while Gram-negative bacteria are increased [172,173]. Newer research also reveals that 30 days Lactobacillus reuteri DSM 17938 treatment significantly alters the mRNA levels of the transcription factors retinoid-related orphan receptor-γ [RORγ] and forkhead box P3 [FOXP3] in the peripheral blood, which modulate T-cell responses to gut microbes: it increases the FOXP3 concentration, thus resulting in decreased RORγ/ FOXP3 ratio. In parallel, it increases the percentage of Lactobacillus in feces and decreases calprotectin [174].
Trichinella spiralis infection-induced muscle hypercontractility in mice was attenuated after Lactobacillus paracasei, but not after Lactobacillus johnsonii, Bifidobacterium lactis, or Bifidobacterium longum treatment from days 10 to 21, post infection. This finding was related to a decrease in the T-helper 2 response triggered by T. spiralis and in transforming growth factor-1, cyclooxygenase-2, and prostaglandin E2 levels in muscles. Based on this experiment, the authors conclude that muscle hypercontractility attenuation is a strain-dependent effect, which follows both immunologic response to infection and a direct effect of L. paracasei on muscles [175]. Similarly, acute ex-vivo exposure of colonic mucosa to Lactobacillus rhamnosus GG resulted in a significant [almost 70%] impairment in smooth muscle cell contraction, the effect being attributed to the reduced contractile response to Acetylcholine [176].
Using the same as previous colorectal distension-dependent gut pain model the effectiveness of Lactobacillus rhamnosus JB-1 was also tested; although its visceral anti-nociceptive activity was confirmed, the nociceptive signals were not found mediated by the specific TRPV1 channel antagonist [168]. It had been previously found in rats that L. rhamnosus JB-1, as well as L. reuteri ATCC23172, later recognized as being L. rhamnosus, and L. plantarum NCIMB 826 [WT] inhibited pain perception after even the maximum colorectal distension pressure of 80mmHg, by altering signaling in DRG fibers [177,178]. Similarly, L. reuteri treatment was found to inhibit the mechano-sensitive response to gastric distension, but not the L. plantarum NCIMB 826 treatment [178].
The probiotics Bifidobacterium infantis 35624, Lactobacillus salivarius UCC4331 or Bifidobacterium breve UCC2003 were given by gavage for a 14-day period, to two strains of visceral normo-sensitive and visceral hyper-sensitive rats, exposed to a novel stress and then to colorectal distension. Their nociceptive responses were analyzed by recording visceral pain behavior. B. infantis 35624 only was found to reduce the total number of pain behaviors in the open field assessment, while significantly increasing the threshold pressure of the first pain behavior [179]. There is also evidence in irritable bowel syndrome patients, where it was found to alleviates symptoms of pain/discomfort, bloating/distention, and bowel movement difficulty [180,181]. [Table 2]
5. Discussion - Conclusions
Pain, the unpleasant sensory and emotional experience associated with actual or potential tissue damage remains a crucial point in the treatment of the postoperative patient. And this is not only due to the stress and psychological state of the just-operated patient, but also to the double origin of this pain: the pain of the surgical trauma on the abdominal wall which is purely of an inflammatory etiology, and the visceral pain, which is due to over-distension of the intestine up to its mobilization from the paralytic ileus, resulting from anesthesia. Post-operative pain is exacerbated by the released pro-inflammatory cytokines, mainly IL-1β, IL-6 and TNF-α, not only locally but in the spinal dorsal horn tissues, which are further involved in the pain mechanism. It is documented that inflammation reduces the pain threshold of nociceptors and increases the individual pain response.
It would be of interest to identify in the near future [i] whether there is a more specific causal relationship between microbiota changes and post-operative pain; [ii] whether microbiota alterations vary, based on the nature or severity of pain; and [iii] whether pain further accelerate microbiota changes in the host.
Since pain is linked with alterations in the gut microbiome, and the overall process of surgery is known to alter this microbiome, there has been considerable research towards the treatment of postoperative pain with probiotics. Thus, today we know that:
- the pain of the surgical trauma on the abdominal wall which is of an inflammatory etiology may be improved by given probiotics exerting strong anti-inflammatory action through the production of IL-10 or IL-4 or through directly limiting specific pro-inflammatory cytokines, such as TNF. Such benefits have been recognized after treatments mainly with Lactiplantibacillus plantarum, and to a lesser extent with L. acidophilus LA-5, L. rhamnosus GG ATCC 53103 and UBLR-58, L. fermentum SGL10, L. brevis GQ4237768, SGL 12, and CECT7480, L. paracasei SGL04, and MSMC39, B. longum UBBL-64 and Reuter and L. casei Shirota. Additionally, Lactobacillus spp., B. dentium and Bifidobacterium spp. are able to modify pain signaling by producing GABA, the most important inhibitory neurotransmitter.
- the visceral pain, which is mainly due to the activation of nociceptors of the thoracic, pelvic, or abdominal organs being extremely sensitive to distension, tissue ischemia and inflammation may be improved by administered probiotics, which mainly exert anti-nociceptive effects via different mechanisms: L. plantarum PS128, L. acidophilus NCFM, L. rhamnosus GG ATCC53103, L. reuteri DSM 17938, L. paracasei, B. infantis 35624, B. longum and L. helveticus in combination, Bifidobacterium lactis CNCM I-2494 and Lactococcus lactis CNCM I-1631 in combination, and the less known L. farciminis, Roseburia hominis, a species of butyrate-producing Lachnospiraceae family, and Faecalibacterium prausnitzii.
- Finally, particular mention must be made of the extraordinary action of L. acidophillus NCFM and of L. salivarius Ls-33, which induce a sustained increase in opioid receptor μ- [OPRM1] mRNA expression, while the former only, also induce significant cannabinoid receptor [CNR2] mRNA expression.
Author Contributions
1. Study conception and design: B. Fyntanidou, K. Kotzampassi. 2. Data collection: S. Gkarmiri, S. Zagalioti, A. Ioannidis, I. Dalalakis, E. Soulioti, L. Loukipoudi 3. Analysis and interpretation of results: A. Chorti, A. Menni, A. Amaniti, 4. Draft manuscript preparation: B. Fyntanidou, A. Amaniti, A. Chorti, A. Shrewsbury 5. Revision of the manuscript: B. Fyntanidou, A. Shrewsbury, K. Kotzampassi. All authors reviewed the results and approved the final version of the manuscript.
Funding
There is no funding for this article.
Data Availability Statement
All data of this article will be available on reasonable demand.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Astyrakaki, E.; Papaioannou, A.; Askitopoulou, H. References to anesthesia, pain, and analgesia in the Hippocratic Collection. Anesth Analg 2010, 110, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.wikidoc.org/index.
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Sommer, F.; Backhed, F. The gut microbiota--masters of host development and physiology. Nat Rev Microbiol 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat Med 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Verges, M.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef]
- Mayer, E.A.; Nance, K.; Chen, S. The Gut-Brain Axis. Annu Rev Med 2022, 73, 439–453. [Google Scholar] [CrossRef]
- Cryan, J.F.; Mazmanian, S.K. Microbiota-brain axis: Context and causality. Science 2022, 376, 938–939. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Cristiano, C.; Avagliano, C.; De Caro, C.; La Rana, G.; Raso, G.M.; Canani, R.B.; Meli, R.; Calignano, A. Gut-brain Axis: Role of Lipids in the Regulation of Inflammation, Pain and CNS Diseases. Curr Med Chem 2018, 25, 3930–3952. [Google Scholar] [CrossRef]
- Felice, V.D.; O'Mahony, S.M. The microbiome and disorders of the central nervous system. Pharmacol Biochem Behav 2017, 160, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Chen, L.H.; Xing, C.; Liu, T. Pain regulation by gut microbiota: molecular mechanisms and therapeutic potential. Br J Anaesth 2019, 123, 637–654. [Google Scholar] [CrossRef]
- Alizadeh, N.; Naderi, G.; Kahrizi, M.S.; Haghgouei, T.; Mobed, A.; Shah-Abadi, M.E. Microbiota-Pain Association; Recent Discoveries and Research Progress. . Curr Microbiol. 2022, 80, 29. [Google Scholar] [CrossRef]
- Bistoletti, M.; Bosi, A.; Banfi, D.; Giaroni, C.; Baj, A. The microbiota-gut-brain axis: Focus on the fundamental communication pathways. Prog Mol Biol Transl Sci 2020, 176, 43–110. [Google Scholar] [CrossRef] [PubMed]
- Lozano, J.; Fernández-Ciganda, S.; González Revello, Á.; Hirigoyen, D.; Martínez, M.; Scorza, C.; Zunino, P. Probiotic potential of GABA-producing lactobacilli isolated from Uruguayan artisanal cheese starter cultures. J Appl Microbiol. 2022, 133, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Attar, N. Good for the gut, good for the brain. Nature Reviews Microbiology 2016, 14, 269–269. [Google Scholar] [CrossRef]
- Bosi, A.; Banfi, D.; Bistoletti, M.; Giaroni, C.; Baj, A. Tryptophan Metabolites Along the Microbiota-Gut-Brain Axis: An Interkingdom Communication System Influencing the Gut in Health and Disease. Int J Tryptophan Res 2020, 13, 1178646920928984. [Google Scholar] [CrossRef]
- Lagomarsino, V.N.; Kostic, A.D.; Chiu, I.M. Mechanisms of microbial-neuronal interactions in pain and nociception. Neurobiol Pain 2021, 9, 100056. [Google Scholar] [CrossRef]
- Eicher, T.P.; Mohajeri, M.H. Overlapping Mechanisms of Action of Brain-Active Bacteria and Bacterial Metabolites in the Pathogenesis of Common Brain Diseases. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, L.; Gao, J.; Zhu, M.; Zhang, Y.; Zhang, H.L. Gut Microbial Metabolites in Parkinson's Disease: Implications of Mitochondrial Dysfunction in the Pathogenesis and Treatment. Mol Neurobiol 2021, 58, 3745–3758. [Google Scholar] [CrossRef]
- Guo, C.; Huo, Y.J.; Li, Y.; Han, Y.; Zhou, D. Gut-brain axis: Focus on gut metabolites short-chain fatty acids. World J Clin Cases 2022, 10, 1754–1763. [Google Scholar] [CrossRef]
- Varela, R.B.; Valvassori, S.S.; Lopes-Borges, J.; Mariot, E.; Dal-Pont, G.C.; Amboni, R.T.; Bianchini, G.; Quevedo, J. Sodium butyrate and mood stabilizers block ouabain-induced hyperlocomotion and increase BDNF, NGF and GDNF levels in brain of Wistar rats. J Psychiatr Res 2015, 61, 114–121. [Google Scholar] [CrossRef]
- Ustianowska, K.; Ustianowski, L.; Machaj, F.; Goracy, A.; Rosik, J.; Szostak, B.; Szostak, J.; Pawlik, A. The Role of the Human Microbiome in the Pathogenesis of Pain. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F.; Jorge, J.M.P.; Perez-Garcia, F.; Sgobba, E. Updates on industrial production of amino acids using Corynebacterium glutamicum. World J Microbiol Biotechnol 2016, 32, 105. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Hashimoto, K.I.; Sawada, Y.; Sokabe, M.; Kawasaki, H.; Martinac, B. Corynebacterium glutamicum mechanosensitive channels: towards unpuzzling "glutamate efflux" for amino acid production. Biophys Rev 2018, 10, 1359–1369. [Google Scholar] [CrossRef]
- Zareian, M.; Ebrahimpour, A.; Bakar, F.A.; Mohamed, A.K.S.; Forghani, B.; Ab-Kadir, M.S.B.; Saari, N. A glutamic acid-producing lactic acid bacteria isolated from Malaysian fermented foods. Int J Mol Sci 2012, 13, 5482–5497. [Google Scholar] [CrossRef] [PubMed]
- Neunlist, M.; Michel, K.; Reiche, D.; Dobreva, G.; Huber, K.; Schemann, M. Glycine activates myenteric neurones in adult guinea-pigs. J Physiol. 2001, 536, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol 2004, 558, 263–275. [Google Scholar] [CrossRef]
- Caputi, V.; Marsilio, I.; Filpa, V.; Cerantola, S.; Orso, G.; Bistoletti, M.; Paccagnella, N.; De Martin, S.; Montopoli, M.; Dall'Acqua, S.; et al. Antibiotic-induced dysbiosis of the microbiota impairs gut neuromuscular function in juvenile mice. Br J Pharmacol 2017, 174, 3623–3639. [Google Scholar] [CrossRef] [PubMed]
- Gronier, B.; Savignac, H.M.; Di Miceli, M.; Idriss, S.M.; Tzortzis, G.; Anthony, D.; Burnet, P.W.J. Increased cortical neuronal responses to NMDA and improved attentional set-shifting performance in rats following prebiotic (B-GOS((R))) ingestion. Eur Neuropsychopharmacol 2018, 28, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Lin, W.J.; Salton, S.R. Role of a VGF/BDNF/TrkB Autoregulatory Feedback Loop in Rapid-Acting Antidepressant Efficacy. J Mol Neurosci 2019, 68, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Minerbi, A.; Shen, S. Gut Microbiome in Anesthesiology and Pain Medicine. Anesthesiology 2022, 137, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Shock, T.; Badang, L.; Ferguson, B.; Martinez-Guryn, K. The interplay between diet, gut microbes, and host epigenetics in health and disease. J Nutr Biochem 2021, 95, 108631. [Google Scholar] [CrossRef] [PubMed]
- Moloney, R.D.; Johnson, A.C.; O'Mahony, S.M.; Dinan, T.G.; Greenwood-Van Meerveld, B.; Cryan, J.F. Stress and the Microbiota-Gut-Brain Axis in Visceral Pain: Relevance to Irritable Bowel Syndrome. CNS Neurosci Ther 2016, 22, 102–117. [Google Scholar] [CrossRef] [PubMed]
- O' Mahony, S.M.; Dinan, T.G.; Cryan, J.F. The gut microbiota as a key regulator of visceral pain. Pain 2017, 158 Suppl 1, S19–S28. [Google Scholar] [CrossRef]
- Greenwood-Van Meerveld, B.; Moloney, R.D.; Johnson, A.C.; Vicario, M. Mechanisms of Stress-Induced Visceral Pain: Implications in Irritable Bowel Syndrome. J Neuroendocrinol 2016, 28. [Google Scholar] [CrossRef]
- Tap, J.; Derrien, M.; Tornblom, H.; Brazeilles, R.; Cools-Portier, S.; Dore, J.; Storsrud, S.; Le Neve, B.; Ohman, L.; Simren, M. Identification of an Intestinal Microbiota Signature Associated With Severity of Irritable Bowel Syndrome. Gastroenterology 2017, 152, 111–123. [Google Scholar] [CrossRef]
- Mars, R.A.T.; Yang, Y.; Ward, T.; Houtti, M.; Priya, S.; Lekatz, H.R.; Tang, X.; Sun, Z.; Kalari, K.R.; Korem, T.; et al. Longitudinal Multi-omics Reveals Subset-Specific Mechanisms Underlying Irritable Bowel Syndrome. Cell 2020, 182, 1460–1473. [Google Scholar] [CrossRef]
- Maeda, T.; Kishioka, S. PPAR and Pain. Int Rev Neurobiol 2009, 85, 165–177. [Google Scholar]
- Galiazzo, G.; De Silva, M.; Giancola, F.; Rinnovati, R.; Peli, A.; Chiocchetti, R. Cellular distribution of cannabinoid-related receptors TRPV1, PPAR-gamma, GPR55 and GPR3 in the equine cervical dorsal root ganglia. Equine Vet J 2021, 54, 788–798. [Google Scholar] [CrossRef] [PubMed]
- Ghaderi, F.; Sotoodehnejadnematalahi, F.; Hajebrahimi, Z.; Fateh, A.; Siadat, S.D. Effects of active, inactive, and derivatives of Akkermansia muciniphila on the expression of the endocannabinoid system and PPARs genes. Sci Rep 2022, 12, 10031. [Google Scholar] [CrossRef] [PubMed]
- Amaral, F.A.; Sachs, D.; Costa, V.V.; Fagundes, C.T.; Cisalpino, D.; Cunha, T.M.; Ferreira, S.H.; Cunha, F.Q.; Silva, T.A.; Nicoli, J.R.; et al. Commensal microbiota is fundamental for the development of inflammatory pain. Proc Natl Acad Sci U S A 2008, 105, 2193–2197. [Google Scholar] [CrossRef] [PubMed]
- Shoskes, D.A.; Wang, H.; Polackwich, A.S.; Tucky, B.; Altemus, J.; Eng, C. Analysis of Gut Microbiome Reveals Significant Differences between Men with Chronic Prostatitis/Chronic Pelvic Pain Syndrome and Controls. J Urol. 2016, 196, 435–441. [Google Scholar] [CrossRef]
- Brenner, D.; Cherry, P.; Switzer, T.; Butt, I.; Stanton, C.; Murphy, K.; McNamara, B.; Iohom, G.; O'Mahony, S.M.; Shorten, G. Pain after upper limb surgery under peripheral nerve block is associated with gut microbiome composition and diversity. Neurobiol Pain 2021, 10, 100072. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med 2016, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Tito, R.Y.; Cypers, H.; Joossens, M.; Varkas, G.; Van Praet, L.; Glorieus, E.; Van den Bosch, F.; De Vos, M.; Raes, J.; Elewaut, D. Brief Report: Dialister as a Microbial Marker of Disease Activity in Spondyloarthritis. . Arthritis Rheumatol 2017, 69, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Puchimada, B.; Kadouri, D.; Zusman, T.; Javed, F.; Eliav, E. The anti-nociceptive effects of Porphyromonas gingivalis lipopolysaccharide. . Arch Oral Biol. 2019, 102. [Google Scholar] [CrossRef]
- Cruz-Aguliar, R.M.; Wantia, N.; Clavel, T.; Vehreschild, M.J.G.T.; Buch, T.; Bajbouj, M.; Haller, D.; Busch, D.; Schmid, R.M.; Stein-Thoeringer, C.K. An Open-Labeled Study on Fecal Microbiota Transfer in Irritable Bowel Syndrome Patients Reveals Improvement in Abdominal Pain Associated with the Relative Abundance of Akkermansia Muciniphila. . Digestion. 2019, 100, 127–138. [Google Scholar] [CrossRef]
- Kotzampassi, K.; Kolios, G.; Manousou, P.; Kazamias, P.; Paramythiotis, D.; Papavramidis, T.; Heliadis, S.; Kouroumalis, E.; Eleftheriadis, E. Oxidative stress due to anesthesia and surgical trauma: importance of early enteral nutrition. . Mol Nutr Food Res. 2009, 53, 770–779. [Google Scholar] [CrossRef]
- Lange, K.; Buerger, M.; Stallmach, A.; Bruns, T. Effects of Antibiotics on Gut Microbiota. . Dig Dis. 2016, 34, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Tilg, H.; Gasbarrini, A. Antibiotics as deep modulators of gut microbiota: between good and evil. Gut 2016, 65, 1906–1915. [Google Scholar] [CrossRef]
- Bolton, L. Feed Abdominal Surgery Patients to Improve Outcomes. . Wounds. 2021, 33, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Sindberg, G.; Wang, F.; Meng, J.; Sharma, U.; Zhang, L.; Dauer, P.; Chen, C.; Dalluge, J.; Johnson, T.; et al. Opioid-induced gut microbial disruption and bile dysregulation leads to gut barrier compromise and sustained systemic inflammation. Mucosal Immunol 2016, 9, 1418–1428. [Google Scholar] [CrossRef] [PubMed]
- Williamson, A.J.; Alverdy, J.C. Influence of the Microbiome on Anastomotic Leak. Clin Colon Rectal Surg 2021, 34, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Jalanka, J.; Salonen, A.; Salojarvi, J.; Ritari, J.; Immonen, O.; Marciani, L.; Gowland, P.; Hoad, C.; Garsed, K.; Lam, C.; et al. Effects of bowel cleansing on the intestinal microbiota. Gut 2015, 64, 1562–1568. [Google Scholar] [CrossRef] [PubMed]
- Stavrou, G.; Kotzampassi, K. Gut microbiome, surgical complications and probiotics. Ann Gastroenterol 2017, 30, 45–53. [Google Scholar] [CrossRef]
- Gorkiewicz, G.; Thallinger, G.G.; Trajanoski, S.; Lackner, S.; Stocker, G.; Hinterleitner, T.; Gully, C.; Hogenauer, C. Alterations in the colonic microbiota in response to osmotic diarrhea. PLoS One 2013, 8, e55817. [Google Scholar] [CrossRef]
- Harrell, L.; Wang, Y.; Antonopoulos, D.; Young, V.; Lichtenstein, L.; Huang, Y.; Hanauer, S.; Chang, E. Standard colonic lavage alters the natural state of mucosal-associated microbiota in the human colon. PLoS One 2012, 7, e32545. [Google Scholar] [CrossRef]
- Watanabe, M.; Murakami, M.; Nakao, K.; Asahara, T.; Nomoto, K.; Tsunoda, A. Randomized clinical trial of the influence of mechanical bowel preparation on faecal microflora in patients undergoing colonic cancer resection. Br J Surg 2010, 97, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Toscano, M.; De Grandi, R.; Casini, V.; Pace, F. Persisting changes of intestinal microbiota after bowel lavage and colonoscopy. . Eur J Gastroenterol Hepatol 2016, 28, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Valentina, C.; Fabio, P. Gut microbiota, dysbiosis and colon lavage. Dig Liver Dis 2019, 51, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shang, L.; Jin, D.; Wu, X.; Long, B. General anesthesia bullies the gut: a toxic relationship with dysbiosis and cognitive dysfunction. Psychopharmacology (Berl). 2022, 239, 709–728. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Serrano, M.; Gerónimo-Pardo, M.; Martínez-Monsalve, A.; Crespo-Sánchez, M.D. Antibacterial effect of sevoflurane and isoflurane. Rev Esp Quimioter 2017, 30, 84–89. [Google Scholar] [PubMed]
- Koutsogiannaki, S.; Schaefers, M.M.; Okuno, T.; Ohba, M.; Yokomizo, T.; Priebe, G.P.; DiNardo, J.A.; Sulpicio, S.G.; Yuki, K. From the Cover: Prolonged Exposure to Volatile Anesthetic Isoflurane Worsens the Outcome of Polymicrobial Abdominal Sepsis. Toxicol Sci 2017, 156, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, M.; Koutsogiannaki, S.; Schaefers, M.; Babazada, H.; Liu, R.; Yuki, K. The Differential Effects of Anesthetics on Bacterial Behaviors. PLoS One 2017, 12, e0170089. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The enteric nervous system and neurogastroenterology. . Nat Rev Gastroenterol Hepatol 2012, 9, 286–294. [Google Scholar] [CrossRef]
- Raybould, H.E. Gut chemosensing: interactions between gut endocrine cells and visceral afferents. Auton Neurosci 2010, 153, 41–46. [Google Scholar] [CrossRef]
- Mazzotta, E.; Villalobos-Hernandez, E.C.; Fiorda-Diaz, J.; Harzman, A.; Christofi, F.L. Postoperative Ileus and Postoperative Gastrointestinal Tract Dysfunction: Pathogenic Mechanisms and Novel Treatment Strategies Beyond Colorectal Enhanced Recovery After Surgery Protocols. Front Pharmacol 2020, 11, 583422. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.; Yang, D.; Vella, M.; Chiu, I.M. The intestinal neuro-immune axis: crosstalk between neurons, immune cells, and microbes. Mucosal Immunol 2021, 14, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Li, Z.; Liu, T.; Yang, N.; Li, Y.; He, J.; Qian, M.; Kuang, Z.; Zhang, W.; Ni, C.; et al. Prebiotics Regulation of Intestinal Microbiota Attenuates Cognitive Dysfunction Induced by Surgery Stimulation in APP/PS1 Mice. Aging Dis 2020, 11, 1029–1045. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Ding, Y.; Wang, L.; Xiao, Y. Gut microbiome improves postoperative cognitive function by decreasing permeability of the blood-brain barrier in aged mice. Brain Res Bull 2020, 164, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, X.; Wu, H. Juvenile Rats Show Altered Gut Microbiota After Exposure to Isoflurane as Neonates. Neurochem Res. 2019, 44, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Zhang, Z.; Guo, N.; Li, X.; Yang, M.; Peng, Y.; Ma, X.; Yu, K.; Wang, C. Effects of Sevoflurane Inhalation Anesthesia on the Intestinal Microbiome in Mice. Front Cell Infect Microbiol 2021, 11, 633527. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Zhang, Z.; Han, C.; Chen, L.; Zheng, X.; Yu, K.; Zhang, Z.; Wang, C. Effects of continuous intravenous infusion of propofol on intestinal flora in rats. Biomed Pharmacother 2021, 134, 111080. [Google Scholar] [CrossRef] [PubMed]
- Liufu, N.; Liu, L.; Shen, S.; Jiang, Z.; Dong, Y.; Wang, Y.; Culley, D.; Crosby, G.; Cao, M.; Shen, Y.; et al. Anesthesia and surgery induce age-dependent changes in behaviors and microbiota. Aging (Albany NY) 2020, 12, 1965–1986. [Google Scholar] [CrossRef]
- Yang, S.; Gu, C.; Mandeville, E.T.; Dong, Y.; Esposito, E.; Zhang, Y.; Yang, G.; Shen, Y.; Fu, X.; Lo, E.H.; et al. Anesthesia and Surgery Impair Blood-Brain Barrier and Cognitive Function in Mice. Front Immunol 2017, 8, 902. [Google Scholar] [CrossRef]
- Ren, Q.; Peng, M.; Dong, Y.; Zhang, Y.; Chen, M.; Yin, N.; Marcantonio, E.R.; Xie, Z. Surgery plus anesthesia induces loss of attention in mice. Front Cell Neurosci 2015, 9, 346. [Google Scholar] [CrossRef]
- Miao, H.; Dong, Y.; Zhang, Y.; Zheng, H.; Shen, Y.; Crosby, G.; Culley, D.J.; Marcantonio, E.R.; Xie, Z. Anesthetic Isoflurane or Desflurane Plus Surgery Differently Affects Cognitive Function in Alzheimer's Disease Transgenic Mice. Mol Neurobiol 2018, 55, 5623–5638. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Zhang, C.; Dong, Y.; Zhang, Y.; Nakazawa, H.; Kaneki, M.; Zheng, H.; Shen, Y.; Marcantonio, E.R.; Xie, Z. Battery of behavioral tests in mice to study postoperative delirium. Sci Rep 2016, 6, 29874. [Google Scholar] [CrossRef]
- Rodino-Janeiro, B.K.; Alonso-Cotoner, C.; Pigrau, M.; Lobo, B.; Vicario, M.; Santos, J. Role of Corticotropin-releasing Factor in Gastrointestinal Permeability. J Neurogastroenterol Motil 2015, 21, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Matzaras, R.; Anagnostou, N.; Nikopoulou, A.; Tsiakas, I.; Christaki, E. The Role of Probiotics in Inflammation Associated with Major Surgery: A Narrative Review. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Agnes, A.; Puccioni, C.; D'Ugo, D.; Gasbarrini, A.; Biondi, A.; Persiani, R. The gut microbiota and colorectal surgery outcomes: facts or hype? A narrative review. BMC Surg 2021, 21, 83. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Velagapudi, R.; Terrando, N. Neuroinflammation after surgery: from mechanisms to therapeutic targets. Nat Immunol 2020, 21, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu Rev Pathol 2020, 15, 493–518. [Google Scholar] [CrossRef] [PubMed]
- Defaye, M.; Gervason, S.; Altier, C.; Berthon, J.Y.; Ardid, D.; Filaire, E.; Carvalho, F.A. Microbiota: a novel regulator of pain. J Neural Transm (Vienna) 2020, 127, 445–465. [Google Scholar] [CrossRef]
- Morowitz, M.J.; Babrowski, T.; Carlisle, E.M.; Olivas, A.; Romanowski, K.S.; Seal, J.B.; Liu, D.C.; Alverdy, J.C. The human microbiome and surgical disease. Ann Surg 2011, 253, 1094–1101. [Google Scholar] [CrossRef]
- Kotzampassi, K. What Surgeon Should Know about Probiotics. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Ohigashi, S.; Sudo, K.; Kobayashi, D.; Takahashi, T.; Nomoto, K.; Onodera, H. Significant changes in the intestinal environment after surgery in patients with colorectal cancer. J Gastrointest Surg. 2013, 17, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, N.; Dang, Q.; Liu, L.; Wang, L.; Li, H.; Han, X. Exploring the roles of intestinal flora in enhanced recovery after surgery. iScience 2023, 26, 105959. [Google Scholar] [CrossRef] [PubMed]
- Lederer, A.K.; Pisarski, P.; Kousoulas, L.; Fichtner-Feigl, S.; Hess, C.; Huber, R. Postoperative changes of the microbiome: are surgical complications related to the gut flora? A systematic review. BMC Surg 2017, 17, 125. [Google Scholar] [CrossRef]
- Usami, M.; Miyoshi, M.; Kanbara, Y.; Aoyama, M.; Sakaki, H.; Shuno, K.; Hirata, K.; Takahashi, M.; Ueno, K.; Tabata, S.; et al. Effects of perioperative synbiotic treatment on infectious complications, intestinal integrity, and fecal flora and organic acids in hepatic surgery with or without cirrhosis. JPEN J Parenter Enteral Nutr. 2011, 35, 317–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qin, H.; Yang, Z.; Xia, Y.; Liu, W.; Yang, J.; Jiang, Y.; Zhang, H.; Yang, Z.; Wang, Y.; et al. Randomised clinical trial: the effects of perioperative probiotic treatment on barrier function and post-operative infectious complications in colorectal cancer surgery - a double-blind study. Aliment Pharmacol Ther 2011, 33, 50–63. [Google Scholar] [CrossRef]
- Lu, H.; He, J.; Wu, Z.; Xu, W.; Zhang, H.; Ye, P.; Yang, J.; Zhen, S.; Li, L. Assessment of microbiome variation during the perioperative period in liver transplant patients: a retrospective analysis. Microb Ecol. 2013, 65, 781–791. [Google Scholar] [CrossRef]
- Shogan, B.D.; Smith, D.P.; Christley, S.; Gilbert, J.A.; Zaborina, O.; Alverdy, J.C. Intestinal anastomotic injury alters spatially defined microbiome composition and function. . Microbiome. 2014, 3, 35. [Google Scholar] [CrossRef]
- Wang, F.; Li, Q.; Wang, C.; Tang, C.; Li, J. Dynamic alteration of the colonic microbiota in intestinal ischemia-reperfusion injury. PLoS One 2012, 7, e42027. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriadis, E.; Kotzampassi, K.; Botsios, D.; Tzartinoglou, E.; Farmakis, H.; Dadoukis, J. Splanchnic ischemia during laparoscopic cholecystectomy. Surg Endosc. 1996, 10, 324–326. [Google Scholar] [CrossRef]
- Eleftheriadis, E.; Kotzampassi, K.; Papanotas, K.; Heliadis, N.; Sarris, K. Gut ischemia, oxidative stress, and bacterial translocation in elevated abdominal pressure in rats. . World J Surg. 1996, 20, 11–16. [Google Scholar] [CrossRef]
- Ioannidis, A.; Arvanitidis, K.; Filidou, E.; Valatas, V.; Stavrou, G.; Michalopoulos, A.; Kolios, G.; Kotzampassi, K. The Length of Surgical Skin Incision in Postoperative Inflammatory Reaction. JSLS 2018, 22. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, J.C.; Keller, D.S.; Baldini, G.; Bordeianou, L.; Weiss, E.; Lee, L.; Boutros, M.; McClane, J.; Feldman, L.S.; Steele, S.R. Clinical Practice Guidelines for Enhanced Recovery After Colon and Rectal Surgery From the American Society of Colon and Rectal Surgeons and Society of American Gastrointestinal and Endoscopic Surgeons. Dis Colon Rectum 2017, 60, 761–784. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.; Nicholson, G.; Hall, G. Endocrine and metabolic response to surgery. Continuing Education in Anaesthesia Critical Care & Pain 2004, 4, 144–147. [Google Scholar] [CrossRef]
- Schricker, T.; Lattermann, R. Perioperative catabolism. Can J Anaesth. 2015, 62, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.T.; Sperandio, V. Inter-kingdom signalling: communication between bacteria and their hosts. Nat Rev Microbiol 2008, 6, 111–120. [Google Scholar] [CrossRef]
- Kendall, M.M.; Sperandio, V. What a Dinner Party! Mechanisms and Functions of Interkingdom Signaling in Host-Pathogen Associations. mBio 2016, 7, e01748. [Google Scholar] [CrossRef] [PubMed]
- Biaggini, K.; Barbey, C.; Borrel, V.; Feuilloley, M.; Déchelotte, P.; Connil, N. The pathogenic potential of Pseudomonas fluorescens MFN1032 on enterocytes can be modulated by serotonin, substance P and epinephrine. . Arch Microbiol. 2015, 197, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Meng, J.; Zhang, L.; Johnson, T.; Chen, C.; Roy, S. Morphine induces changes in the gut microbiome and metabolome in a morphine dependence model. Sci Rep 2018, 8, 3596. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Meng, J.; Ban, Y.; Jalodia, R.; Chupikova, I.; Fernandez, I.; Brito, N.; Sharma, U.; Abreu, M.T.; Ramakrishnan, S.; et al. Morphine tolerance is attenuated in germfree mice and reversed by probiotics, implicating the role of gut microbiome. Proc Natl Acad Sci U S A 2019, 116, 13523–13532. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Menni, A.; Moysidis, M.; Tzikos, G.; Stavrou, G.; Tsetis, J.K.; Shrewsbury, A.D.; Filidou, E.; Kotzampassi, K. Looking for the Ideal Probiotic Healing Regime. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Xiang, Q.; Tang, X.; Zhang, Q.; Liu, X.; Zhao, J.; Cui, S.; Zhang, H. Lactobacillus reuteri CCFM1175 and Lactobacillus paracasei CCFM1176 Could Prevent Capsaicin-Induced Ileal and Colonic Injuries.. Probiotics Antimicrob Proteins. 2023, 15:797-812. [CrossRef]
- Meenakshi, S.; Santhanakumar, R. The role of probiotics as wound healers: an overall view. . J Wound Care 2023, 32, 318–328. [Google Scholar] [CrossRef]
- Kotzampassi, K.; Stavrou, G.; Damoraki, G.; Georgitsi, M.; Basdanis, G.; Tsaousi, G.; Giamarellos-Bourboulis, E.J. A Four-Probiotics Regimen Reduces Postoperative Complications After Colorectal Surgery: A Randomized, Double-Blind, Placebo-Controlled Study. . World J Surg. 2015, 39, 2776–2783. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, O.; Chatzakis, C.; Tirta, M.; Anestiadou, E.; Zapsalis, K.; Symeonidis, S.; Bitsianis, S.; Kotidis, E.; Pramateftakis, M.G.; Mantzoros, I.; et al. The Efficacy of Probiotics, Prebiotics, and Synbiotics in Patients Who Have Undergone Abdominal Operation, in Terms of Bowel Function Post-Operatively: A Network Meta-Analysis. J Clin Med 2023, 12. [Google Scholar] [CrossRef]
- Tang, G.; Huang, W.; Tao, J.; Wei, Z. Prophylactic effects of probiotics or synbiotics on postoperative ileus after gastrointestinal cancer surgery: A meta-analysis of randomized controlled trials. PLoS One 2022, 17, e0264759. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, B.; Li, J.; Feng, M.; Zhang, Y.; Yao, W.; Zhang, C.; Wan, L. Celastrol attenuates incision-induced inflammation and pain associated with inhibition of the NF-κB signalling pathway via SARM. . Life Sci. 2018, 205, 136–144. [Google Scholar] [CrossRef]
- Kim, M.K.; Kang, H.; Baek, C.W.; Jung, Y.H.; Woo, Y.C.; Choi, G.J.; Shin, H.Y.; Kim, K.S. Antinociceptive and anti-inflammatory effects of ginsenoside Rf in a rat model of incisional pain. J Ginseng Res 2018, 42, 183–191. [Google Scholar] [CrossRef]
- Sommer, C.; Kress, M. Recent findings on how proinflammatory cytokines cause pain: peripheral mechanisms in inflammatory and neuropathic hyperalgesia. . Neurosci Lett. 2004, 361, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Brennan, T.J. Comparison of skin incision vs. skin plus deep tissue incision on ongoing pain and spontaneous activity in dorsal horn neurons. Pain 2009, 144, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, S.; Takeuchi, H.; Fujii, K.; Shiraishi, N.; Adachi, Y.; Kitano, S. Length of laparotomy incision and surgical stress assessed by serum IL-6 level. Injury. 2006, 37, 247–251. [Google Scholar] [CrossRef]
- Ronchetti, S.; Migliorati, G.; Delfino, D.V. Association of inflammatory mediators with pain perception. . Biomed Pharmacother. 2017, 96, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Vanderwall, A.G.; Milligan, E.D. Cytokines in Pain: Harnessing Endogenous Anti-Inflammatory Signaling for Improved Pain Management. Front Immunol 2019, 10, 3009. [Google Scholar] [CrossRef] [PubMed]
- Milligan, E.D.; Penzkover, K.R.; Soderquist, R.G.; Mahoney, M.J. Spinal interleukin-10 therapy to treat peripheral neuropathic pain. Neuromodulation 2012, 15, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Tanga, F.Y.; Nutile-McMenemy, N.; DeLeo, J.A. The CNS role of Toll-like receptor 4 in innate neuroimmunity and painful neuropathy. . Proc Natl Acad Sci U S A. 2005, 102, 5856–5861. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Naka, T.; Kubo, M. SOCS proteins, cytokine signalling and immune regulation. Nat Rev Immunol. 2007, 7, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Mohtashami, M.; Mohamadi, M.; Azimi-Nezhad, M.; Saeidi, J.; Nia, F.F.; Ghasemi, A. Lactobacillus bulgaricus and Lactobacillus plantarum improve diabetic wound healing through modulating inflammatory factors. . Biotechnol Appl Biochem. 2021, 68, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.K.; Podia, M.; Priyanka; Raut, S. ; Singh, S.; Pinnaka, A.K.; Khatri, N. Insight Into the Beneficial Role of Lactiplantibacillus plantarum Supernatant Against Bacterial Infections, Oxidative Stress, and Wound Healing in A549 Cells and BALB/c Mice. Front Pharmacol 2021, 12, 728614. [Google Scholar] [CrossRef]
- Panagiotou, D.; Filidou, E.; Gaitanidou, M.; Tarapatzi, G.; Spathakis, M.; Kandilogiannakis, L.; Stavrou, G.; Arvanitidis, K.; Tsetis, J.K.; Gionga, P.; et al. Role of Lactiplantibacillus plantarum UBLP-40, Lactobacillus rhamnosus UBLR-58 and Bifidobacterium longum UBBL-64 in the Wound Healing Process of the Excisional Skin. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Tarapatzi, G.; Filidou, E.; Kandilogiannakis, L.; Spathakis, M.; Gaitanidou, M.; Arvanitidis, K.; Drygiannakis, I.; Valatas, V.; Kotzampassi, K.; Manolopoulos, V.G.; et al. The Probiotic Strains Bifidomicronbacterium lactis, Lactobacillus acidophilus, Lactiplantibacillus plantarum and Saccharomyces boulardii Regulate Wound Healing and Chemokine Responses in Human Intestinal Subepithelial Myofibroblasts. Pharmaceuticals (Basel) 2022, 15. [Google Scholar] [CrossRef]
- Brandi, J.; Cheri, S.; Manfredi, M.; Di Carlo, C.; Vita Vanella, V.; Federici, F.; Bombiero, E.; Bazaj, A.; Rizzi, E.; Manna, L.; et al. Exploring the wound healing, anti-inflammatory, anti-pathogenic and proteomic effects of lactic acid bacteria on keratinocytes. Sci Rep 2020, 10, 11572. [Google Scholar] [CrossRef]
- Chen, X.; Thibeault, S.L. Role of tumor necrosis factor-alpha in wound repair in human vocal fold fibroblasts. Laryngoscope 2010, 120, 1819–1825. [Google Scholar] [CrossRef] [PubMed]
- Gueniche, A.; Bastien, P.; Ovigne, J.M.; Kermici, M.; Courchay, G.; Chevalier, V.; Breton, L.; Castiel-Higounenc, I. Bifidobacterium longum lysate, a new ingredient for reactive skin. Exp Dermatol 2010, 19, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Banjonjit, S.; Taweechotipatr, M.; Rungsiyanont, S. Effect of probiotic Lactobacillus paracasei on tumor necrosis factor-alpha level in gingival crevicular fluid of patients undergoing impacted third molar removal. J Oral Sci 2022, 64, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Ferrés-Amat, E.; Espadaler-Mazo, J.; Calvo-Guirado, J.L.; Ferrés-Amat, E.; Mareque-Bueno, J.; Salavert, A.; Aguiló-García, M.; Moreno-Centeno, J.; Ferrés-Padró, E. Probiotics diminish the post-operatory pain following mandibular third molar extraction: a randomised double-blind controlled trial (pilot study). Benef Microbes. 2020, 11, 631–639. [Google Scholar] [CrossRef]
- Lei, M.; Guo, C.; Wang, Y.; Hua, L.; Xue, S.; Yu, D.; Zhang, C.; Wang, D. Oral administration of probiotic Lactobacillus casei Shirota relieves pain after single rib fracture: a randomized double-blind, placebo-controlled clinical trial. Asia Pac J Clin Nutr 2018, 27, 1252–1257. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, C.; Wang, J.; Guo, Q.; Zou, W. Oral Lactobacillus reuteri LR06 or Bifidobacterium BL5b supplement do not produce analgesic effects on neuropathic and inflammatory pain in rats. Brain Behav 2019, 9, e01260. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Versalovic, J. Probiotics-host communication: Modulation of signaling pathways in the intestine. . Gut Microbes. 2010, 1, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Drabik, K.A.; Waypa, T.S.; Musch, M.W.; Alverdy, J.C.; Schneewind, O.; Chang, E.B.; Petrof, E.O. Soluble factors from Lactobacillus GG activate MAPKs and induce cytoprotective heat shock proteins in intestinal epithelial cells. Am J Physiol Cell Physiol 2006, 290, C1018–C1030. [Google Scholar] [CrossRef]
- Vale, G.C.; Mota, B.I.S.; Ando-Suguimoto, E.S.; Mayer, M.P.A. Effect of Probiotics Lactobacillus acidophilus and Lacticaseibacillus rhamnosus on Antibacterial Response Gene Transcription of Human Peripheral Monocytes. Probiotics Antimicrob Proteins. 2023, 15, 264–274. [Google Scholar] [CrossRef]
- Du, X.; Hao, H.; Yang, Y.; Huang, S.; Wang, C.; Gigout, S.; Ramli, R.; Li, X.; Jaworska, E.; Edwards, I.; et al. Local GABAergic signaling within sensory ganglia controls peripheral nociceptive transmission. J Clin Invest 2017, 127, 1741–1756. [Google Scholar] [CrossRef]
- Rousseaux, C.; Thuru, X.; Gelot, A.; Barnich, N.; Neut, C.; Dubuquoy, L.; Dubuquoy, C.; Merour, E.; Geboes, K.; Chamaillard, M.; et al. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat Med. 2007, 13, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Ringel-Kulka, T.; Goldsmith, J.R.; Carroll, I.M.; Barros, S.P.; Palsson, O.; Jobin, C.; Ringel, Y. Lactobacillus acidophilus NCFM affects colonic mucosal opioid receptor expression in patients with functional abdominal pain - a randomised clinical study. Aliment Pharmacol Ther 2014, 40, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Agostini, S.; Goubern, M.; Tondereau, V.; Salvador-Cartier, C.; Bezirard, V.; Lévèque, M.; Keränen, H.; Theodorou, V.; Bourdu-Naturel, S.; Goupil-Feuillerat, N.; et al. A marketed fermented dairy product containing Bifidobacterium lactis CNCM I-2494 suppresses gut hypersensitivity and colonic barrier disruption induced by acute stress in rats. Neurogastroenterol Motil. 2012, 24, 376–e172. [Google Scholar] [CrossRef] [PubMed]
- Darbaky, Y.; Evrard, B.; Patrier, S.; Falenta, J.; Garcin, S.; Tridon, A.; Dapoigny, M.; Silberberg, C.; Nivoliez, A.; Diop, L. Oral probiotic treatment of Lactobacillus rhamnosus Lcr35® prevents visceral hypersensitivity to a colonic inflammation and an acute psychological stress. J Appl Microbiol. 2017, 122, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Kannampalli, P.; Pochiraju, S.; Chichlowski, M.; Berg, B.M.; Rudolph, C.; Bruckert, M.; Miranda, A.; Sengupta, J.N. Probiotic Lactobacillus rhamnosus GG (LGG) and prebiotic prevent neonatal inflammation-induced visceral hypersensitivity in adult rats. Neurogastroenterol Motil. 2014, 26, 1694–1704. [Google Scholar] [CrossRef]
- Roman, P.; Abalo, R.; Marco, E.M.; Cardona, D. Probiotics in digestive, emotional, and pain-related disorders. Behav Pharmacol. 2018, 29, 103–119. [Google Scholar] [CrossRef]
- McVey Neufeld, K.A.; Strain, C.R.; Pusceddu, M.M.; Waworuntu, R.V.; Manurung, S.; Gross, G.; Gerry, M.M.; Hoban, A.E.; Murphy, K.; Stanton, C.; et al. Lactobacillus rhamnosus GG soluble mediators ameliorate early life stress-induced visceral hypersensitivity and changes in spinal cord gene expression. Neuronal Signal 2020, 4, NS20200007. [Google Scholar] [CrossRef]
- Vanhoutvin, S.A.; Troost, F.J.; Kilkens, T.O.; Lindsey, P.J.; Hamer, H.M.; Jonkers, D.M.; Venema, K.; Brummer, R.J. The effects of butyrate enemas on visceral perception in healthy volunteers. Neurogastroenterol Motil. 2009, 21, 952–e976. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc Nutr Soc 2015, 74, 13–22. [Google Scholar] [CrossRef]
- Zhang, J.; Song, L.; Wang, Y.; Liu, C.; Zhang, L.; Zhu, S.; Liu, S.; Duan, L. Beneficial effect of butyrate-producing Lachnospiraceae on stress-induced visceral hypersensitivity in rats. J Gastroenterol Hepatol 2019, 34, 1368–1376. [Google Scholar] [CrossRef]
- Ait-Belgnaoui, A.; Payard, I.; Rolland, C.; Harkat, C.; Braniste, V.; Theodorou, V.; Tompkins, T.A. Bifidobacterium longum and Lactobacillus helveticus Synergistically Suppress Stress-related Visceral Hypersensitivity Through Hypothalamic-Pituitary-Adrenal Axis Modulation. J Neurogastroenterol Motil 2018, 24, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Ait-Belgnaoui, A.; Eutamene, H.; Houdeau, E.; Bueno, L.; Fioramonti, J.; Theodorou, V. Lactobacillus farciminis treatment attenuates stress-induced overexpression of Fos protein in spinal and supraspinal sites after colorectal distension in rats. Neurogastroenterol Motil. 2009, 21, 567–573. [Google Scholar] [CrossRef]
- Ait-Belgnaoui, A.; Han, W.; Lamine, F.; Eutamene, H.; Fioramonti, J.; Bueno, L.; Theodorou, V. Lactobacillus farciminis treatment suppresses stress induced visceral hypersensitivity: a possible action through interaction with epithelial cell cytoskeleton contraction. Gut 2006, 55, 1090–1094. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Wang, Y.P.; Yen, H.F.; Liu, P.Y.; Tzeng, W.J.; Tsai, C.F.; Lin, H.C.; Lee, F.Y.; Jeng, O.J.; Lu, C.L.; et al. Lactobacillus plantarum PS128 Ameliorated Visceral Hypersensitivity in Rats Through the Gut-Brain Axis. Probiotics Antimicrob Proteins. 2020, 12, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Martin, R.; Lashermes, A.; Gillet, M.; Meleine, M.; Gelot, A.; Eschalier, A.; Ardid, D.; Bermudez-Humaran, L.G.; Sokol, H.; et al. Anti-nociceptive effect of Faecalibacterium prausnitzii in non-inflammatory IBS-like models. Sci Rep 2016, 6, 19399. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr Opin Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- van Praagh, J.B.; de Goffau, M.C.; Bakker, I.S.; van Goor, H.; Harmsen, H.J.M.; Olinga, P.; Havenga, K. Mucus Microbiome of Anastomotic Tissue During Surgery Has Predictive Value for Colorectal Anastomotic Leakage. Ann Surg 2019, 269, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Leclerc, M.; Martin, R.; Chain, F.; Lenoir, M.; Raguideau, S.; Hudault, S.; Bridonneau, C.; Northen, T.; Bowen, B.; et al. Identification of metabolic signatures linked to anti-inflammatory effects of Faecalibacterium prausnitzii. mBio 2015, 6. [Google Scholar] [CrossRef]
- Martin, R.; Chain, F.; Miquel, S.; Lu, J.; Gratadoux, J.J.; Sokol, H.; Verdu, E.F.; Bercik, P.; Bermudez-Humaran, L.G.; Langella, P. The commensal bacterium Faecalibacterium prausnitzii is protective in DNBS-induced chronic moderate and severe colitis models. Inflamm Bowel Dis 2014, 20, 417–430. [Google Scholar] [CrossRef]
- Carlsson, A.H.; Yakymenko, O.; Olivier, I.; Håkansson, F.; Postma, E.; Keita, A.V.; Söderholm, J.D. Faecalibacterium prausnitzii supernatant improves intestinal barrier function in mice DSS colitis. Scand J Gastroenterol. 2013, 48, 1136–1144. [Google Scholar] [CrossRef]
- Laval, L.; Martin, R.; Natividad, J.N.; Chain, F.; Miquel, S.; Desclee de Maredsous, C.; Capronnier, S.; Sokol, H.; Verdu, E.F.; van Hylckama Vlieg, J.E.; et al. Lactobacillus rhamnosus CNCM I-3690 and the commensal bacterium Faecalibacterium prausnitzii A2-165 exhibit similar protective effects to induced barrier hyper-permeability in mice. Gut Microbes 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P. TRPV1: a new target for treatment of visceral pain in IBS? Gut 2008, 57, 882–884. [Google Scholar] [CrossRef] [PubMed]
- Winston, J.; Shenoy, M.; Medley, D.; Naniwadekar, A.; Pasricha, P.J. The vanilloid receptor initiates and maintains colonic hypersensitivity induced by neonatal colon irritation in rats. Gastroenterology 2007, 132, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Boesmans, W.; Owsianik, G.; Tack, J.; Voets, T.; Vanden Berghe, P. TRP channels in neurogastroenterology: opportunities for therapeutic intervention. Br J Pharmacol 2011, 162, 18–37. [Google Scholar] [CrossRef] [PubMed]
- Vay, L.; Gu, C.; McNaughton, P.A. The thermo-TRP ion channel family: properties and therapeutic implications. Br J Pharmacol 2012, 165, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Perez-Burgos, A.; Wang, L.; McVey Neufeld, K.A.; Mao, Y.K.; Ahmadzai, M.; Janssen, L.J.; Stanisz, A.M.; Bienenstock, J.; Kunze, W.A. The TRPV1 channel in rodents is a major target for antinociceptive effect of the probiotic Lactobacillus reuteri DSM 17938. J Physiol 2015, 593, 3943–3957. [Google Scholar] [CrossRef] [PubMed]
- Zholos, A.V.; Dryn, D.O.; Melnyk, M.I. General anaesthesia-related complications of gut motility with a focus on cholinergic mechanisms, TRP channels and visceral pain. Front Physiol 2023, 14, 1174655. [Google Scholar] [CrossRef]
- Melnyk, M.I.; Dryn, D.O.; Al Kury, L.T.; Dziuba, D.O.; Zholos, A.V. Suppression of mI(CAT) in Mouse Small Intestinal Myocytes by General Anaesthetic Ketamine and its Recovery by TRPC4 Agonist (-)-englerin A. Front Pharmacol 2020, 11, 594882. [Google Scholar] [CrossRef] [PubMed]
- Savino, F.; Cordisco, L.; Tarasco, V.; Palumeri, E.; Calabrese, R.; Oggero, R.; Roos, S.; Matteuzzi, D. Lactobacillus reuteri DSM 17938 in infantile colic: a randomized, double-blind, placebo-controlled trial. . Pediatrics. 2010, 126, e526–e533. [Google Scholar] [CrossRef]
- Savino, F.; Cresi, F.; Pautasso, S.; Palumeri, E.; Tullio, V.; Roana, J.; Silvestro, L.; Oggero, R. Intestinal microflora in breastfed colicky and non-colicky infants. Acta Paediatr. 2004, 93, 825–829. [Google Scholar] [CrossRef]
- de Weerth, C.; Fuentes, S.; de Vos, W.M. Crying in infants: on the possible role of intestinal microbiota in the development of colic. Gut Microbes 2013, 4, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Savino, F.; Garro, M.; Montanari, P.; Galliano, I.; Bergallo, M. Crying Time and RORgamma/FOXP3 Expression in Lactobacillus reuteri DSM17938-Treated Infants with Colic: A Randomized Trial. J Pediatr 2018, 192, 171–177. [Google Scholar] [CrossRef]
- Verdú, E.F.; Bercík, P.; Bergonzelli, G.E.; Huang, X.X.; Blennerhasset, P.; Rochat, F.; Fiaux, M.; Mansourian, R.; Corthésy-Theulaz, I.; Collins, S.M. Lactobacillus paracasei normalizes muscle hypercontractility in a murine model of postinfective gut dysfunction. Gastroenterology. 2004, 127, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Guarino, M.P.; Altomare, A.; Stasi, E.; Marignani, M.; Severi, C.; Alloni, R.; Dicuonzo, G.; Morelli, L.; Coppola, R.; Cicala, M. Effect of acute mucosal exposure to Lactobacillus rhamnosus GG on human colonic smooth muscle cells. J Clin Gastroenterol. 2008, 42, S185–S190. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Wang, L.; Forsythe, P.; Goettsche, G.; Mao, Y.; Wang, Y.; Tougas, G.; Bienenstock, J. Inhibitory effects of Lactobacillus reuteri on visceral pain induced by colorectal distension in Sprague-Dawley rats. Gut 2006, 55, 191–196. [Google Scholar] [CrossRef]
- Duncker, S.C.; Kamiya, T.; Wang, L.; Yang, P.; Bienenstock, J. Probiotic Lactobacillus reuteri alleviates the response to gastric distension in rats. J Nutr 2011, 141, 1813–1818. [Google Scholar] [CrossRef]
- McKernan, D.P.; Fitzgerald, P.; Dinan, T.G.; Cryan, J.F. The probiotic Bifidobacterium infantis 35624 displays visceral antinociceptive effects in the rat. Neurogastroenterol Motil. 2010, 22, 1029–1035. [Google Scholar] [CrossRef]
- Whorwell, P.J.; Altringer, L.; Morel, J.; Bond, Y.; Charbonneau, D.; O'Mahony, L.; Kiely, B.; Shanahan, F.; Quigley, E.M. Efficacy of an encapsulated probiotic Bifidobacterium infantis 35624 in women with irritable bowel syndrome. Am J Gastroenterol. 2006, 101, 1581–1590. [Google Scholar] [CrossRef]
- OMahony, L.; McCarthy, J.; Kelly, P.; Hurley, G.; Luo, F.; Chen, K.; O'Sullivan, G.C.; Kiely, B.; Collins, J.K.; Shanahan, F.; et al. Lactobacillus and bifidobacterium in irritable bowel syndrome: symptom responses and relationship to cytokine profiles. Gastroenterology. 2005, 128, 541–551. [Google Scholar] [CrossRef]

Table 1.
Probiotics in relation to the inflammation-induced pain of surgical trauma.
| Probiotics | Action | Type of operation | Type of study |
|---|---|---|---|
| L. plantarum (128,135) | ↑ IL-10 and TGF1 ↓ TNF |
1. dorsal wound 2. mandibular 3rd molar excision |
1. diabetic rats 2. humans |
| L. rhamnosus UBLR-58 (130-132) | ↑ IL-10 | Skin Wound healing | Rats and humans |
|
L. acidophilus LA-5 (130-132) |
↑ IL-10 and MAPKs | Skin Wound healing | Rats and humans |
| L. fermentum SGL10 (130-132) | ↑ IL-10 and TGF1 | Skin Wound healing | Rats and humans |
| L. brevis GQ4237768 (130-132, 135) | ↑ IL-10 ↓ TNF |
mandibular 3rd molar excision | Humans |
| L. brevis SGL 12 (130-132) | ↑ IL-10 | Skin Wound healing | Rats and humans |
| L. paracasei SGL 04 (130-132, 135) | ↑ Increase of IL-10 | Skin Wound healing | Rats and humans |
| B. longum UBBL-64 (130-132) | ↑ Increase of IL-10 ↓ TNF |
mandibular 3rd molar excision | Humans |
| L. casei Shirota (137) | NA | Rib fracture pain modulation | Humans |
| L. rhamnosus GG ATCC 53103 (140-141) | MAPKs | Skin Wound healing | Rats |
| Lactobacillus spp (142) | GABA | Skin Wound healing | Rats |
| Bifidobacterium spp (142) | GABA | Skin Wound healing | Rats |
| B. dentium (142) | GABA | Skin Wound healing | Rats |
Numbers in parenthesis represent reference number, NA=not applicable.
Table 2.
Probiotics in relation to gut distension-induced visceral pain.
| Probiotics | Action | Type of operation |
|---|---|---|
| Experimental studies | ||
| L. acidophillus NCFM (143) | Decrease in μ-opioid receptor and cannabinoid receptor R2 | Colorectal distention Reduction of stress-induced visceral hypersensitivity and pain |
| L. salivarius Ls-33 (143) | Decrease in opioid receptor μ | Colorectal distention |
| L. rhamnosus Lcr35 (145) | IL-13/Th 17 immune activation | Colorectal distention |
| L. rhamnosus GG ATCC53103 (145) | NA | Colorectal distention |
| Roseburia hominis (151-152) | Butyrate activation | Reduction of stress-induced visceral hypersensitivity and pain |
| B. longum (153) | regulating glucocorticoid negative feedback on the HPA axis | Reduction of stress-induced visceral hypersensitivity and pain |
| L. helveticus (153) | regulating glucocorticoid negative feedback on the HPA axis | Reduction of stress-induced visceral hypersensitivity and pain |
| L. farciminis (154-155) | Fos downregulation | Reduction of stress-induced visceral hypersensitivity and pain |
| L. plantarum PS128 (156) | 5-HTP regulation and altering signaling in DRG fibres | Reduction of stress-induced visceral hypersensitivity and pain |
| Faecalibacterium prausnitzii (157,160-163) | intestinal epithelial barrier enhancement/ modulation of tight junctions in inflammation models | Reduction of stress-induced visceral hypersensitivity and pain |
| L. reuteri DSM 17938 (168,171) | TRPV1 channel antagonist modulation | Reduction of stress-induced visceral hypersensitivity and pain |
|
L. johnsonii, B. lactis, or B. longum (175) |
GF-1, COX-2, and PGE2 levels transformation | Reduction of muscle hypercontractility |
| L. rhamnosus JB-1 (177) | altering signaling in DRG fibres | Reduction of stress-induced visceral hypersensitivity and pain |
| B. infantis 35624 (179) | NA | Reduction of stress-induced visceral hypersensitivity and pain |
| Clinical studies | ||
| B. lactis CNCM I-2494 (144) | NA | Reduction of stress-induced visceral hypersensitivity and pain |
| Lactococcus lactis CNCM I-1631 (144) | NA | Reduction of stress-induced visceral hypersensitivity and pain |
| L. reuteri DSM 17938 (168,171) | TRPV1 channel antagonist modulation | Reduction of stress-induced visceral hypersensitivity and pain |
Numbers in parenthesis represent reference number, NA=not applicable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



