
Submitted:
31 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
The combined effects of the thermal processing (i.e., sterilisation treatment) and the prior frozen storage time (3 or 6 months at –18 ºC) on the quality loss in canned Atlantic horse mackerel (Trachurus trachurus) were determined. Thus, the sterilisation step led to a remarkable (p<0.05) formation in canned fish muscle of fluorescent compounds, free fatty acids (FFAs) and trimethylamine and to an increase of the L* and b* colour parameters; meantime, a decrease (p<0.05) of the total ω3 FA/total ω6 FA ratio and the a* colour value were detected. The prior frozen storage period led to an increased (p<0.05) lipid oxidation (peroxide, thiobarbituric acid reactive substance and fluorescent compound formation) and hydrolysis (FFA formation) development and to increased L* and b* colour values in the corresponding canned samples; additionally, a lower (p<0.05) polyene index and phospholipid content were observed in canned fish previously subjected to frozen storage. In most indices, physico-chemical changes related to quality loss were found higher if the previous storage period was increased. According to the marked effects of the sterilisation step and the prior frozen storage period, optimisation of such processing conditions is recommended to maximise the quality of canned horse mackerel.
Keywords:
fish
; canning
; cold storage
; lipid oxidation
; lipid hydrolysis
; fatty acid profile
; colour changes
; trimethylamine
; degradation
1. Introduction
Canning procedure can preserve marine species by the combination of sealing in a hermetic container and heating to destroy spoilage and pathogenic microorganisms and inactivate enzymes [1,2]. Provided duration and temperature during the heating and cooling cycle of the thermal process are properly carried out, canned seafood would be preserved for a long time. However, long-term high temperature may cause losses of quality in the nutritional value (destruction of vitamins and protein components) and loss in the sensory quality such as the soft texture, the separation of jelly and fat, discolouration, and undesirable taste [3,4]. Therefore, the search for optimised canning conditions [5] and the employment of advanced preserving technologies [6] has been found necessary.
According to the short shelf-life time of refrigerated marine species, excess raw material is frozen prior to canning. Freezing followed by frozen storage is considered a valuable alternative for relatively long preservation and has been used increasingly [7,8]. In most cases, marine species can be preserved at times of glut and stored under frozen conditions until supplied to canneries. However, measurements of sensory, chemical and physical changes have shown that deterioration of fish quality continues to some extent during the frozen storage, since undesirable changes associated with lipids and proteins are produced [9,10]. As a result, most concerns with canned fish have shown to be related to the quality of the raw material employed. Thus, previous studies have shown a relevant effect of time [11,12] and temperature [13] of prior cold storage on the quality of the resulting canned product. However, previous research regarding the combined effect of prior frozen storage and subsequent canning process can be considered scarce.
Atlantic horse mackerel (Trachurus trachurus), also known as European horse mackerel or common scad, is a pelagic fish species found in the eastern Atlantic Ocean of Europa and Africa and in the south-eastern Indian Ocean [14,15]. Previous research regarding quality changes of this species during processing has been focused on fresh and frozen products [16,17]. Thus, different preserving strategies have been developed for quality enhancement and for the shelf-life time increase under storage conditions such as addition of natural preserving extracts [18,19], biopreservation [20], irradiation [21], and high-pressure [22]. Although recent research has addressed the quality enhancement of brine-packed horse mackerel by addition of an antioxidant extract to the packing medium [23], it can be concluded that previous research on canned horse mackerel is very scarce.
The present study focused on the quality of canned horse mackerel (T. trachurus). Its basic objective was to analyse the combined effect of the canning process and a prior frozen storage time on the physico-chemical changes related to quality loss of the canned product. For it, complementary quality indices such as lipid oxidation and hydrolysis development, changes in fatty acid (FA) composition and colour parameters, and trimethylamine (TMA) formation were determined in raw and canned samples.
2. Materials and Methods
2.1. Raw fish and fish processing
Horse mackerel (thirty-two fish) (weight range: 165−185 g; length range: 27−31 cm) were obtained in winter 2022 at Vigo harbour (North-Western Spain) and transported on ice to the laboratory. Upon arrival at the laboratory, eight individuals were selected, divided into four batches (two individuals per batch) and considered as raw fish. For it, the fish individuals were beheaded, eviscerated, filleted and the dark muscle discarded. Within each batch, the white muscle was pooled together, minced and analysed independently (n=4).
On the same day, a second group of eight fish individuals was taken, divided into four batches (two individuals per batch), beheaded, eviscerated, filleted and subjected directly to the canning process according to the conditions expressed later on. Such canned samples were labelled as not previously frozen (0-month samples).
The remaining fish individuals (sixteen) were placed in individual flexible polyethylene bags, kept at –40 ºC for 48 h and then stored at –18 ºC for 3 or 6 months, respectively. At each storage period, eight individuals were thawed overnight (4 ºC), separated into four batches (two individuals per batch), beheaded, eviscerated, filleted and subjected to the canning procedure according to conditions expressed later on. The resulting samples were labelled as canned samples with prior 3- or 6-month frozen storage (3-month and 6-month samples), respectively.
At each canning time, 45–g portions of horse mackerel fillets (from one fish) were introduced in small flat rectangular cans (105 × 60 × 25 mm; 150 mL). Then, cans were filled with 90 mL of distilled water, vacuum–sealed (SOMME 222, Ezquerra, San Adrián, Navarra, Spain) and sterilised (115 ºC, 45 min; Fo = 7 min) (CIFP Coroso, Ribeira, A Coruña, Spain) in a steam retort (Presoclave II 75L, JP Selecta, Barcelona, Spain). When the heating time was accomplished, the steam was cut off, the remaining steam was flushed away by the use of air, and water at reduced pressure was employed for cooling the cans.
The cans were opened after 3 months of storage at room temperature (20 ºC). Then, the liquid part was carefully drained off gravimetrically and filtered through a filter paper and the dark muscle was discarded. The white muscle of horse mackerel was wrapped in filter paper and used for analysis. For each sample, the fish muscle corresponding to two cans was pooled together and considered for carrying out the different physico-chemical analyses. Four different batches of canned samples were analysed independently (n = 4). Additionally, each physico-chemical analysis was carried out in triplicate in each single sample.
In agreement with common practice employed in canneries, a 3-month storage was employed. Commonly, manufacturers indicate that a minimum of a 2-3-month storage is required to optimise the acceptability of commercial canned fish [24].
All chemical reagents and solvents employed were of reagent grade (Merck, Darmstadt, Germany).
2.2. Lipid oxidation assessment
The extraction of the lipid fraction was carried out on the fish white muscle by employing the Bligh and Dyer [25] procedure. This method based on a single-phase solubilisation of the lipids with a chloroform-methanol (1:1) mixture. The results were expressed as g lipid·kg-1 muscle.
Peroxide value (PV) was assessed by spectrophotometric analysis (Beckman Coulter, DU 640; London, UK) on the lipid extract. For it, peroxide reduction with ferric thiocyanate was employed, following the Chapman and McKay [26] method. The results were calculated as meq active oxygen·kg-1 lipids.
Thiobarbituric acid index (TBA-i) was assessed in agreement with the Vyncke [27] procedure. For it, reaction between thiobarbituric acid reactive substances (TBARS) present in a trichloroacetic acid extract of the fish muscle and thiobarbituric acid was carried out. Determination of TBARS content was accomplished spectrophotometrically at 532 nm and calculated from a standard curve prepared with 1,1,3,3-tetraethoxy-propane (TEP). Results were calculated as mg malondialdehyde·kg-1 muscle.
2.3. Assessment of fluorescent compounds
Formation of interaction compounds between oxidised lipids and nucleophilic compounds (i.e., protein-like molecules) were assessed by fluorescence spectroscopy (Fluorimeter LS 45; Perkin Elmer España; Tres Cantos, Madrid, Spain). In agreement with previous research [11,28], fluorescence was measured at excitation/emission of 393/463 and 327/415 nm in the lipid extract of the fish muscle. The relative fluorescence (RF) was calculated as follows: RF = F/Fst, where F is the fluorescence measured at each excitation/emission wavelength pair and Fst is the fluorescence intensity of a quinine sulphate solution (1 µg·mL-1 in 0.05 M H2SO4) at the corresponding wavelength pair. The fluorescence ratio (FR) was calculated as the ratio between the two RF values: FR = RF393/463 nm/RF327/415 nm.
2.4. Determination of free fatty acid (FFA) and phospholipid (PL) content
FFA content was assessed in the lipid extract of horse mackerel muscle according to the Lowry and Tinsley [29] procedure. This method implies a complex formation of FFA with cupric acetate-pyridine, followed by spectrophotometric (715 nm) determination. For quantitative purposes, oleic acid was employed as standard. Results were calculated as g FFAs·kg-1 lipids.
PL content was determined by measuring the organic phosphorus in the total lipid extracts in agreement with the Raheja et al. [30] procedure. This method implies a complex formation of organic phosphorus with ammonium molybdate. For quantitative purposes, 1,2-dipalmitoyl-rac-glycero-3-phosphocholine was employed as standard. Results were expressed as g PLs·kg-1 muscle and as g PL·kg-1 total lipids.
2.5. Analysis of FA composition
Fatty acid methyl esters (FAMEs) from lipid extracts were obtained by employing acetyl chloride in methanol. Then, analysis of FAMEs was carried out by gas-liquid chromatography (Perkin-Elmer 8700 chromatograph; Madrid, Spain), in agreement with an established procedure [31]. Resulting FAME peaks were identified by comparing their retention times with those of standard mixtures (Qualmix Fish, Larodan, Malmo, Sweden; FAME mix, Supelco, Inc., Bellefonte, PA, USA). For quantitative purposes, peak areas were automatically integrated and C19:0 FA was used as internal standard. The content of each FA was expressed as g·100 g-1 total FAs.
Results regarding total ω3 FAs, ω3/ω6 ratio and polyene index (PI) (C20:5ω3 + C22:6ω3/C16:0 ratio) were calculated taking into account the values of the corresponding individual FAs.
2.6. Other physico-chemical determinations
Colour parameters (L*, a* and b*) were determined in the surface of the raw and canned fish muscle. For it, instrumental colour analysis (CIE 1976 Lab), performed with a tristimulus Hunter Labscan 2.0/45 colorimeter, was carried out. For each sample analysis, colour scores were averaged over four determinations, which were taken by rotating the measuring head 90° between triplicate measurements per position.
TMA content was determined using the picrate colorimetric (Beckman Coulter, DU 640; London, UK) method, as previously described by Tozawa et al. [32]. This method involves the preparation of a 5% trichloroacetic acid extract of fish muscle (10 g/25 mL). Results were expressed as mg TMA-N·kg-1 muscle.
2.7. Statistical analysis
As expressed above, four replicates (n = 4) were considered in this study. Data obtained were evaluated by analysis of variance (ANOVA) to explore differences resulting from the effect of the sterilisation step and the prior frozen storage time. For carrying out mean values comparison, the least-squares difference (LSD) procedure was developed. A confidence interval at the 95% level (p < 0.05) was considered in order to establish significant differences among batches in all instances. For it, the PASW Statistics 18 software for Windows (SPSS Inc., Chicago, IL, USA) was employed. Correlation values among the different quality indices were determined by the Pearson test.
3. Results and discussion
3.1. Lipid oxidation development
Lipid content of raw horse mackerel was 14.17±2.41 g lipid·kg-1 muscle. With the aim of obtaining an accurate analysis of the lipid oxidation development in the current fish processing study, different and complementary methods, i.e. peroxide, TBARS and fluorescent compound determinations, were carried out.
Comparison of peroxide levels in raw fish and canned fish not previously frozen showed no effect (p > 0.05) of the sterilisation process on this kind of lipid oxidation compounds (Table 1). However, a marked increase (p < 0.05) was recorded in canned fish frozen stored for 3 or 6 months proved by including a prior frozen storage time. Notably, no differences (p > 0.05) were detected by comparison of canned samples corresponding to 3 and 6 months of prior frozen storage.
A similar result was observed for the evolution of the TBARS content (Table 1). Thus, although an increased average TBARS value was proved after the heating step, differences were not significant (p > 0.05). Additionally, canned fish corresponding to prior frozen storage (i.e., 3 and 6 months) showed higher (p < 0.05) levels of TBARS than the raw fish and the canned fish that was not subjected to frozen storage. As for the peroxide determination, no effect (p > 0.05) on the TBA-i was recorded by increasing the prior frozen storage from 3 to 6 months.
Contrary to peroxide and TBARS values, the FR showed a marked increase (p < 0.05) as a result of the sterilisation step (Table 1). Additionally, FR values increased (p < 0.05) if a prior frozen storage period (i.e., 3 or 6 months at –18 ºC) was included. As for peroxide and TBARS determinations, the highest average values were detected in canned fish previously subjected to the longest storage period; however, no significant differences (p > 0.05) with samples corresponding to a 3-month storage were detected.
Oxidation of the lipid fraction in seafood is considered a complex deteriorative mechanism. Thus, it involves the formation of a great diversity of molecules, most of them unstable, and therefore, susceptible to breakdown and lead to the formation of lower-weight compounds, which in turn can react with nucleophilic-type molecules (peptides, free amino acids, proteins, etc.) present in the fish muscle [3,33]. In the present study, the heat treatment (i.e., sterilisation) favoured the formation of interaction compounds with fluorescent properties, according to the development of two damage mechanisms (i.e., non-enzymatic lipid oxidation and non-enzymatic browning) [16]. Additionally, formation of lipid oxidation compounds ought to be also produced during the frozen storage period as a result of the endogenous enzyme (i.e., lipoxygenases, peroxidases, etc.) activity and autolysis [9,10]; this oxidant effect should increase with the storage duration and temperature. Values obtained in the present research were in all cases included in the 1.36-5.91 (PV) and 0.04-0.32 (TBA-i) ranges; therefore, a relevant value of primary and secondary lipid oxidation compounds was not obtained and could be explained on the basis of being relative unstable molecules [33,34].
The assessment of the fluorescent compounds produced as a result of the interaction between lipid oxidation compounds and nucleophilic-type molecules present in the fish muscle has shown to be a highly valuable tool in the present study to describe changes occurring as a result of the sterilisation step and the previous frozen storage. Fluorescence spectroscopy has already proved to be a valuable tool for the discrimination of fresh and frozen fish [35], for fish authenticity [36], and for the assessment of lipid damage during processing [11,28].
A low peroxide content has been observed in different canned fish species such as olive oil-packed bluefin tuna (Thunnus thynnus) and tomato sauce-packed sardine (Sardina pilchardus) [37], sunflower oil- and olive oil-packed eel (Anguilla anguilla) [38], and brine-packed [12] and water-packed [39] Atlantic Chub mackerel (Scomber colias). In such studies, the low primary oxidation compound content was explained on the basis that thermal treatment would partially destroy such kind of molecules and lead to the formation of low-molecular-weight molecules (i.e., carbonyl compounds). Additionally, interaction of lipid oxidation compounds with nucleophilic molecules present in the muscle may lead to the formation of fluorescent compounds [11,40]. Contrary to the present results, an increase of the peroxide presence in canned fish as a result of the sterilisation process was observed in brine-canned mackerel (S. colias) [41].
Previous investigation has concerned a great number of studies focused on the effect of canning on the TBARS content of canned fish. In most cases, an increase of the TBARS content has been described in canned fish when an oil-packing medium was employed. Contrary, aqueous-packing media have led either to non-different values or to a decrease of the TBARS level in canned fish. The fact of not detecting a TBARS content increase has been explained on the basis of TBARS leaching into the aqueous packing medium. Thus, an increased formation was observed in canned sardine (S. pilchardus) by employing brine- and olive oil-packing [11], in sunflower-, groundnut- and coconut-packed yellowfin tuna (Thunnus albacares) [42], in sunflower oil-, olive oil-, soybean oil and brine-packed silver carp (Hypophthalmichthys molitrix) [43] and in sunflower oil- and olive oil-packed eel (A. anguilla) [38]. Contrary, a marked decrease of the TBARS value was described for tomato sauce-packed sardine (S. pilchardus) [37] and brine-packed Atlantic Chub mackerel (S. colias) [12]. In agreement with the present results, no effect on the TBARS content was observed in water-packed mackerel (S. colias) [44].
Previous seafood research has shown a marked increase of the FR as a result of the sterilisation process. In fact, this analytical measurement was proposed as a lipid quality index for canned fish [11]. In agreement with the current results, an increased fluorescent compound formation was detected in varied canned fish products such as sunflower oil-, olive oil-, soybean oil- and brine-packed silver carp (H. molitrix) [43], brine-packed Atlantic Chub mackerel (S. colias) [12,41], water-packed Atlantic mackerel (S. scombrus) [45], and water-packed Atlantic Chub mackerel (S. colias) [39].
Regarding the effect of the prior frozen storage on the lipid oxidation development in canned seafood, a general increase of the FR value has been reported in canned fish with an increased prior frozen period. These results include olive oil-packed sardine (S. pilchardus) (prior 0-12-month period at –18 ºC) [11], brine-packed Atlantic Chub mackerel (S. colias) (0-15-month period at –18 ºC) [12], and brine-canned horse mackerel (T. trachurus) (0-6-month period at –18 ºC) [23].
3.2. Lipid hydrolysis development
Comparison of the FFA content in raw fish and canned fish without prior frozen period showed a strong formation of this kind of molecules resulting from the heating process (Figure 1). Furthermore, a subsequent increase (p < 0.05) of the FFA presence was proved by including a prior frozen storage, this increase being more important (p < 0.05) if the storage period was increased.
Contrary to FFA results, an average decrease could be observed for the PL content as a result of the sterilisation step (Figure 2); however, this decrease was not found significant (p > 0.05). If a prior frozen storage period was included (i.e., 3 or 6 months at –18 ºC), canned samples depicted lower (p < 0.05) PL values than those reported for the raw fish. The evolution of the PL content in the lipid fraction showed a high inverse correlation with the FFA presence (r = –0.924, logarithmic fitting; p < 0.05).
FFA are considered to be the result of the hydrolysis of high-molecular-weight lipid compounds such as triacylglycerols (TAGs) and PLs. In the present study, FFA content can be considered the result of different factors. First, the sterilisation process can lead to hydrolysis of lipid classes such as TAGs and PLs as a result of thermal breakdown (i.e., non-enzymatic lipid hydrolysis) [11,44]. On the other side, FFA formation is reported to be produced during the frozen storage by the action of endogenous enzymes (phospholipases and lipases in general) present in the fish muscle; this effect showed to increase with the storage duration and temperature [9,10]. Remarkably, FFA accumulation has been accepted as not having nutritional significance. Nevertheless, it has been recognised as being involved in several deteriorative mechanisms during seafood processing as leading to off-taste and off-odour development, muscle texture changes, and acceleration of the formation of lipid oxidation compounds [10,33].
Regarding PL compounds, this group of lipids has attracted a great attention as showing a high bioavailability and preserving effect on several diseases and as delivery systems [46,47]. Based on industrial requirements (food production and pharmaceutical industries), important efforts are being addressed to the retention of PL constituents from seafood and their corresponding by-products as presenting high PUFA levels [48,49].
Previous research has reported on the effect of canning on the FFA content of canned fish muscle. Thus, and in agreement with the present results, sunflower oil- or brine-packed sprat (Clupeonella cultriventris) [50], water-canned Chub mackerel (S. colias) [39], and brine-canned Chub mackerel (S. colias) [41] revealed a marked FFA formation when compared to the starting raw material. Notably, an important effect of the filling medium on the FFA value has been reported in previous research. Thus, Naseri et al. [43] showed a FFA content increase in canned silver carp (Hypophthalmichthys molitrix) including olive oil, sunflower oil, soybean oil or brine as packing medium. Additionally, the employment of sunflower oil increased the preservative effect of processed yellowfin tuna (T. albacares) against FFA formation when compared to groundnut and coconut oils as packing media [42].
Regarding the effect of the sterilisation process on the PL value in canned seafood, previous studies can be considered scarce. In agreement with the current results, a marked loss of PL compounds could be observed in brine-canned Chub mackerel (S. colias) muscle [44]. This loss could be explained on the basis of different mechanisms. First, as showing a high PUFA level [48,49], PLs could be damaged easily by the heating treatment. Additionally, PL classes may be partially hydrolysed during the prior frozen storage period by endogenous enzyme activity [9,10]. Finally, as being a group including relatively polar lipid compounds, PLs could be lost partly by leaching from the muscle into the packing medium [44].
Previous studies regarding the influence of a prior frozen storage period on the FFA and PL contents in canned seafood can be considered scarce. According to the current results, Prego et al. [12] proved an increase of the FFA content in brine-canned Atlantic Chub mackerel (S. colias) when an increased prior frozen period (0-15 months at -18 ºC). Additionally, Méndez et al. [23] observed a FFA value increase and a PL presence decrease in brine-canned horse mackerel (T. trachurus) if frozen period (0-6-month period) was previously employed.
3.3. Fatty acid analysis
The FA analysis of the lipid fraction corresponding to the raw fish indicated the following profile (g·100 g-1 total FAs): 4.24±0.76 (C14:0), 0.54±0.03 (C15:0), 22.15±0.10 (C16:0), 4.42±0.29 (C16:1ω7), 1.22±0.09 (C17:0), 7.52±0.16 (C18:0), 15.88±1.68 (C18:1ω9), 3.07±0.17 (C18:1ω7), 1.46±0.06 (C18:2ω6), 1.70±0.30 (C20:1ω9), 0.32±0.03 (C20:2ω6), 1.45±0.11 (C20:4ω6), 0.25±0.03 (C22:1ω9), 7.67±0.04 (C20:5ω3), 0.43±0.04 (C22:4ω6), 0.75±0.08 (C24:1ω9), 2.89±0.11 (C22:5ω3), and 23.78±2.82 (C22:6ω3).
In agreement with the profile obtained, DHA, C16:0, C18:1ω9, EPA, and C18:0 showed to be the major FAs in the present fish species. In recent decades, a great interest has been attributed to seafood according to their beneficial health effects. In agreement with recent reports [51,52], the total value of ω3 unsaturated FAs is considered as a highly valuable index. According to clinical and epidemiological research, EPA consumption has been associated to an inhibitory effect on the development of inflammatory, coronary, and circulatory diseases [53]. Regarding DHA, it has been associated with foetal development, prevention of neurodegenerative diseases, and appropriate functioning of the nervous system and visual organs in the foetus [54]. In this context, a great attention has been accorded to the ω3/ω6 ratio of foods included in the human diet [55,56]. With the aim of preventing relevant health concerns, values included in the 0.25-1.00 range have been recommended for the ω3/ω6 ratio [57]. According to such nutritional and healthy properties, discussion on FA results obtained in the present study will now be focused on the content of ω3 FAs and PUFAs in the lipid fraction. Evolution of their content during the thermal treatment and the previous frozen storage period would be closely related to the above-mentioned development of lipid oxidation and hydrolysis in the fish muscle.
Small differences (p > 0.05) could be observed for the EPA content among the different samples considered in the current study (Table 2). Thus, an increased average value was depicted resulting from the sterilisation step; however, differences between raw fish and canned samples without prior storage period were not found significant (p > 0.05). Canned samples corresponding to a 3-month storage period showed the highest average value, which was found significantly higher (p < 0.05) than the one reported for the raw fish; however, comparison to other canned samples (without prior storage period and with a 6-month period) did not show significant differences (p > 0.05).
Regarding the DHA value, the raw fish showed higher average values than any of the canned samples considered in the present study (Table 2). Remarkably, differences were found significant (p < 0.05) when compared to canned fish that had been kept under frozen condition, but not (p > 0.05) when compared to canned samples not previously stored. Additionally, the lowest average DHA content was found in samples corresponding to the longer prior storage time.
In a recent study, Prego et al. [31] analysed the effect of packing media (water, brine, sunflower oil, refined olive oil, and virgin olive oil) on canned Atlantic mackerel (S. scombrus). As a result, EPA content showed to increase when employing water and brine as packing media, DHA presence increased in canned mackerel packed in refined olive oil and virgin olive oil, the total ω3 FA value increased in canned fish when water, sunflower oil, refined olive oil and virgin olive oil were used as filling media, and the PUFA/STFA ratio value increased under all packing condition. Contrary, no influence on the ω3/ω6 ratio value was obtained under any of the packing conditions tested.
Previous studies regarding the influence of prior frozen holding on the FA composition of canned marine species are scarce. No remarkable influence was observed in the PI of olive oil-canned sardine (S. pilchardus) muscle with increased prior storage time (12 months at –18 ºC) [11]. Contrary, a PI decrease in brine-canned Chub mackerel (S. colias) was observed with by increasing the prior frozen period (0-15-month period) [12]. Additionally, a similar decreasing tendency for the PI was reported by Méndez et al. [23] in brine-canned horse mackerel (T. trachurus) previously stored for a 0-6-month period under frozen conditions. Recently, Prego et al. [31] studied the influence of a prior frozen storage (0-6-month period at –18 ºC) in canned Atlantic mackerel (S. scombrus) that was packed under different media. As a result, no effect on the EPA value and the ω3/ω6 ratio was observed by increasing the prior storage period; however, a decrease in DHA (sunflower oil packing) and total ω3 FA (sunflower oil packing) values was detected by increasing the prior storage time, which was also observed in the current study.
3.4. Determination of colour changes
A relevant increase (p < 0.05) of the L* value was observed in canned fish as a result of the sterilisation step (Table 3). An additional L* value increase (p < 0.05) was observed in canned fish that was previously stored for 6 months under frozen conditions. This additional increase may be justified according to an increased protein damage in fish corresponding to such holding condition (i.e., extended denaturation and interaction with oxidised lipids).
The analysis of the a* value revealed a strong decrease (p < 0.05) as a result of the sterilisation process (Table 3). The prior frozen storage led to higher a* values in canned horse mackerel; however, no differences (p > 0.05) could be observed when compared to canned fish without prior storage period. Remarkably, an a* value decrease has been reported to be correlated with haemoglobin-mediated lipid oxidation in fish and to show an inverse relationship with the secondary lipid oxidation compounds level [61,62]. Accordingly, an inverse correlation (r = –0.732, quadratic fitting; p > 0.05) between this colour index and the TBARS content has been obtained in the current study.
Regarding the b* value (Table 3), a marked increase was observed after the sterilisation step. Higher average values were observed in canned samples corresponding to batches previously subjected to frozen storage; remarkably, canned fish corresponding to the 6-month storage showed significantly higher (p < 0.05) values than their counterpart canned samples that were not subjected to prior storage. This b* increase has been explained on the basis that formation of interaction compounds between oxidised lipids and protein-type molecules and yellowish colour development are favoured by the heating step [63,64]. According to the direct relationship between the b* parameter and the lipid oxidation development, a good correlation value was obtained in the present research between this colour index and the FR (r = 0.942, linear fitting; p < 0.05); additionally, fair correlation values were observed with the PV (r = 0.792, quadratic fitting; p > 0.05) and the TBA-i (r = 0.861, quadratic fitting; p > 0.05).
According to its great effect on appearance and acceptability of seafood in general, assessment of colour changes has deserved a great attention. Therefore, previous research has addressed the changes produced in colour parameters as a result of the canning process. As a general behaviour, the sterilisation process has led to increased L* and b* values and to a* value decreases and the results obtained in this study are in agreement with those previous findings. Thus, an increase of L* and b* values and a decrease of a* value in yellowfin tuna (T. albacares) that was packed with baby corn, green pea, and broccoli [65] and with groundnut and coconut [42] have been reported. An L* value increase in sunflower oil-packed Atlantic mackerel (S. scombrus) [45], and an L* value increase and an a* value decrease in brine-canned Chub mackerel (S. colias) [41] have been described. Recently, Gómez-Limia et al. [66] reported an L* and b* value increase in sunflower oil- and olive oil-canned eel (A. anguilla); additionally, an a* decrease was observed in sunflower oil-canned product.
Regarding the effect of prior cold storage on colour changes in canned seafood, increasing the prior storage temperature and time led to an increase of the L* value and a decrease of the a* value in canned skipjack tuna (Katsuwonus pelamis) [13]. Recently, Méndez et al. [23] obtained an increase of L* and b* values by increasing the prior holding time (0–6 months at –18 ºC) in brine-canned horse mackerel (T. trachurus); however, and contrary to the present research, no effect on the a* value could be inferred.
3.5. TMA content
A strong effect (p < 0.05) of the sterilisation step on the TMA content was obtained (Figure 3). Thus, all kinds of canned fish revealed higher (p < 0.05) levels of this deteriorative molecule than the raw fish. Notably, no differences (p > 0.05) on the TMA value were detected as a result of including a prior frozen storage.
TMA is considered an important deteriorative molecule whose level can indicate the degree of quality loss in seafood in general [34]. Since TMA formation as a result of microbial activity is not likely to occur during the present study, formation of TMA should be explained from the breakdown of protein-like compounds [2,6]. Thus, previous research accounts for a relevant TMA formation as a result of the sterilisation step in canned seafood. These results have concerned different kinds of fish species and packing media such as olive oil-packed tuna (T. thynnus) and tomato sauce-packed sardine (S. pilchardus) [37], sunflower-, groundnut- and coconut-packed yellowfin tuna (T. albacares) [42], and brine-canned Chub mackerel (S. colias) [41].
Concerning the effect of the prior frozen storage, previous information can be considered scarce. A marked increase of the TMA value was observed in brine-canned Atlantic Chub mackerel (S. colias) by increasing the prior holding period (0–15 months at –18 ºC) [12].
4. Conclusions
The combined effect of the sterilisation treatment and the previous frozen storage period on the quality of the canned products was analysed. A marked effect of the thermal treatment and the prior frozen storage period (3 or 6 months at –18 ºC) was proved in water-packed canned horse mackerel. Thus, the sterilisation step led to a significant (p < 0.05) formation in canned fish muscle of fluorescent compounds, FFAs and TMA and to an increase of the L* and b* colour parameters; meantime, a decrease (p < 0.05) of the total ω3 FA/total ω6 FA ratio and the a* colour value were detected. The prior holding period led to an increased (p < 0.05) lipid oxidation (PV, TBA-i, and FR) and hydrolysis (FFA formation) development and to increased L* and b* colour values in the corresponding canned samples; furthermore, a lower (p < 0.05) PI and PL content were observed in canned horse mackerel previously subjected to the holding period. In most indices, quality changes were found higher with increased previous holding period.
In agreement with the marked effect of the sterilisation and the prior frozen storage time on the canning process of this pelagic fish species, optimisation of such processing conditions is recommended to maximise the quality of the corresponding canned product. Regarding the thermal process, optimisation of time and temperature of sterilisation ought to be carried out, taking into account the characteristics of the raw fish composition, the packing medium, and the geometry of the canning support. On the other side, and according to the canneries need of holding the raw material to be employed, it is recommended to carry out the freezing step as fast as possible and maintain the storage temperature at –18 ºC or even lower.
In order to better elucidate the influence of the previous frozen holding period on the canned fish quality, further research including physico-chemical analyses on fish samples at the frozen-thawed step is recommended. Additionally, the effect of processing conditions on horse mackerel muscle corresponding to individuals of different characteristics (size, lipid content, catching time, etc.) than in the present study ought to be addressed. To guarantee a high-quality raw material to be employed for the canning process of horse mackerel, the use of traditional and complementary strategies such as glazing, addition of natural antioxidants, and the use of impermeable packaging could constitute suitable alternatives.
Author Contributions
Conceptualization, B.M. and S.P.A.; methodology, E.V., B.M. and M.T.; data curation, E.V. and M.T.; writing—original draft preparation, S.P.A.; writing—review and editing, S.P.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Horner, W. Canning fish and fish products. In Fish Processing Technology; Hall, G., Ed.; 2nd edition. Blackie Academic and Professional, Chapman and Hall: London, UK, 1997; pp. 119–159. [Google Scholar]
- Veiga, A.; Martínez, E.; Ojea, G.; Caride, A. Principles of thermal processing in canned seafood. In Quality Parameters in Canned Seafoods; Cabado, A.G., Vieites, J.M., Eds.; Nova Science Publishers, Inc., New York, USA, 2008; pp. 83–103.
- Lukoshkina, M.; Odoeva, G. Kinetics of chemical reactions for prediction of quality of canned fish during storage. App. Biochem. Microb. 2003, 39, 321–327. [Google Scholar] [CrossRef]
- Ling, B.; Tang, J.; Kong, F.; Mitcham, E.J.; Wang, S. Kinetics of food quality changes during thermal processing: A review. Food Bioprocess Technol. 2015, 8, 343–358. [Google Scholar] [CrossRef]
- Pitarch, J.L.; Vilas, C.; de Prada, C.; Palacín, C.G.; Alonso, A.A. Optimal operation of thermal processing of canned tuna under product variability. J. Food Eng. 2021, 304, 110594. [Google Scholar] [CrossRef]
- Tokur, B.; Korkmaz, K. Novel thermal sterilisation technologies in seafood processing. In Innovative Technologies in Seafood Processing; Özoğul, Y., Ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 303–322. [Google Scholar]
- James, C.; Purnell, G.; James, S.J. A review of novel and innovative food freezing technologies. Food Bioprocess Technol. 2015, 8, 1616–1634. [Google Scholar] [CrossRef]
- Wu, X.F.; Zhang, M.; Adhikari, B.; Sun, J.C. Recent developments in novel freezing and thawing technologies applied to foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 3620–3631. [Google Scholar] [CrossRef] [PubMed]
- Sista, R.; Erickson, M.; Shewfelt, R. Quality deterioration in frozen foods associated with hydrolytic enzyme activities. In Quality in Frozen Food; Erickson, M., Hung, Y.C., Eds.; Chapman and Hall: New York, NY, USA, 1997; pp. 101–110. [Google Scholar]
- Sikorski, Z.; Kolakowski, E. Endogenous enzyme activity and seafood quality: Influence of chilling, freezing, and other environmental factors. In Seafood Enzymes; Haard, N., Simpson, B., Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 451–487. [Google Scholar]
- Aubourg, S.P.; Medina, I. Quality differences assessment in canned sardine (Sardina pilchardus) by fluorescence detection. J. Agric. Food Chem. 1997, 45, 3617–3621. [Google Scholar] [CrossRef]
- Prego, R.; Fidalgo, L.G.; Saraiva, J.A.; Vázquez, M.; Aubourg, S.P. Impact of prior high-pressure processing on lipid damage and volatile amines formation in mackerel muscle subjected to frozen storage and canning. LWT-Food Sci. Technol. 2021, 135, 109957. [Google Scholar] [CrossRef]
- Little, A. Effect of pre- and post-mortem handling on reflectance characteristics of skipjack tuna. J. Food Sci. 1972, 37, 502. [Google Scholar] [CrossRef]
- Bandarra, N.M.; Batista, I.; Nunes, M.L.; Empis, J.M. Seasonal variation in the chemical composition of horse mackerel (Trachurus trachurus). Eur. Food Res. Technol. 2001, 212, 535–539. [Google Scholar] [CrossRef]
- El Mghazli, H.; Znari, M.; Mounir, A. Stock discrimination in the horse mackerel Trachurus trachurus (Telostei: Carangidae) off the Moroccan Atlantic Coastal Waters using a morphometric-meristic analysis. Thalassas: An Int. J. Mar. Sci. 2022, 38, 171–181. [Google Scholar] [CrossRef]
- Aubourg, S.P. Damage detection in horse mackerel (Trachurus trachurus) during chilled storage. J. Amer. Oil Chem. Soc. 2001, 78, 857–862. [Google Scholar] [CrossRef]
- Aubourg, S.P.; Piñeiro, C.; González, M.J. Quality loss related to rancidity development during frozen storage of horse mackerel (Trachurus trachurus). J. Amer. Oil Chem. Soc. 2004, 81, 671–678. [Google Scholar] [CrossRef]
- Farvin, K.H.S.; Grejsen, H.D.; Jacobsen, C. Potato peel extract as a natural antioxidant in chilled storage of minced horse mackerel (Trachurus trachurus): Effect on lipid and protein oxidation. Food Chem. 2012, 131, 843–851. [Google Scholar] [CrossRef]
- Albertos, I.; Martín-Diana, A.B.; Jaime, I.; Avena-Bustillos, R.J.; McHugh, T.H.; Takeoka, G.R.; Dao, L.; Rico, D. Antioxidant effect of olive leaf powder on fresh Atlantic horse mackerel (Trachurus trachurus) minced muscle. J. Food Process Presev. 2018, 42, e13397. [Google Scholar] [CrossRef]
- Zarandona, I.; López-Caballero, M.E.; Montero, M.P.; Guerrero, P.; de la Caba, K.; Gómez-Guillén, M.C. Horse mackerel (Trachurus trachurus) fillets biopreservation by using gallic acid and chiotosan coatings. Food Cont. 2021, 120, 107511. [Google Scholar] [CrossRef]
- Alves Silva, H.; Mendes, R.; Nunes, M.L.; Empis, J. Protein changes after irradiation and ice storage of horse mackerel (Trachurus trachurus). Eur. Food Res. Technol. 2006, 224, 83–90. [Google Scholar] [CrossRef]
- Erkan, N.; Üretener, G.; Alpas, H.; Selçuk, A.; Özden, Ö.; Buzrul, S. Effect of high hydrostatic pressure (HHP) treatment on physicochemical properties of horse mackerel (Trachurus trachurus). Food Bioprocess Technol. 2011, 4, 1322–1329. [Google Scholar] [CrossRef]
- Méndez, L.; Trigo, M.; Zhang, B.; Aubourg, S.P. Antioxidant effect of octopus by-products in canned horse mackerel (Trachurus trachurus) previously subjected to different frozen storage times. Antioxidants 2022, 11, 2091. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Roso, B.; Cuesta, I.; Pérez, M.; Borrego, E.; Pérez-Olleros, L.; Varela, G. Lipid composition and palatability of canned sardines. Influence of the canning process and storage in olive oil for five years. J. Sci. Food Agric. 1998, 77, 244–250. [Google Scholar] [CrossRef]
- Bligh, E.; Dyer, W. A rapid method of total extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Chapman, R.; McKay, J. The estimation of peroxides in fats and oils by the ferric thiocyanate method. J. Am. Oil Chem. Soc. 1949, 26, 360–363. [Google Scholar] [CrossRef]
- Vyncke, W. Direct determination of the thiobarbituric acid value in trichloracetic acid extracts of fish as a measure of oxidative rancidity. Fette, Seifen, Anstrichm. 1970, 72, 1084–1087. [Google Scholar] [CrossRef]
- Aubourg, S.P.; Medina, I.; Pérez-Martín, R.I. A comparison between conventional and fluorescence detection methods of cooking-induced damage to tuna fish lipids. Z. Lebensm. Unters. Forsch. 1995, 200, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Lowry, R.; Tinsley, I. Rapid colorimetric determination of free fatty acids. J. Am. Oil Chem. Soc. 1976, 53, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Raheja, R.; Kaur, C.; Singh, A.; Bhatia, A. New colorimetric method for the quantitative determination of phospholipids without acid digestion. J. Lipid Res. 1973, 14, 695–697. [Google Scholar] [CrossRef] [PubMed]
- Prego, R.; Trigo, M.; Martínez, B.; Aubourg, S.P. Effect of previous frozen storage, packing medium and sterilisation on fatty acid composition of canned mackerel (Scomber scombrus). Marine Drugs 2022, 20, 666. [Google Scholar] [CrossRef] [PubMed]
- Tozawa, H.; Erokibara, K.; Amano, K. Proposed modification of Dyer’s method for trimethylamine determination in codfish. In Fish Inspection and Quality Control; Kreuzer, R., Ed.; Fishing News Books Ltd: London, UK, 1971; pp. 187–190. [Google Scholar]
- Rustad, T. Lipid oxidation. In Handbook of Seafood and Seafood Products Analysis; Nollet, L.M., Toldrá, F., Eds.; CRC Press, Francis and Taylor Group: Boca Raton, FL, USA, 2010; pp. 87–95. [Google Scholar]
- Özoğul, Y. Methods for freshness quality and deterioration. In Handbook of Seafood and Seafood Products Analysis; Nollet, L.M., Toldrá, F., Eds.; CRC Press, Francis and Taylor Group: Boca Raton, FL, USA, 2010; pp. 189–214. [Google Scholar]
- Hassoun, A. Exploring the Potential of Fluorescence Spectroscopy for the Discrimination between Fresh and Frozen-Thawed Muscle Foods. Photochem. 2021, 1, 247–263. [Google Scholar] [CrossRef]
- Hassoun, A.; Sahar, A.; Lakhal, L.; Aït-Kaddour, A. Fluorescence spectroscopy as a rapid and non-destructive method for monitoring quality and authenticity of fish and meat products: Impact of different preservation conditions. LWT-Food Sci. Technol. 2019, 103, 279–292. [Google Scholar] [CrossRef]
- Selmi, S.; Monser, L.; Sadok, S. The influence of local canning process and storage on pelagic fish from Tunisia: Fatty acids profile and quality indicators. J. Food Proc. Preserv. 2008, 32, 443–457. [Google Scholar] [CrossRef]
- Gómez-Limia, L.; Sanmartín, N.M.; Carballo, J.; Domínguez, R.; Lorenzo, J.M.; Martínez, S. Oxidative stability and antioxidant activity in canned eels: Effect of processing and filling medium. Foods 2021, 10, 790. [Google Scholar] [CrossRef] [PubMed]
- Malga, J.M.; Trigo, M.; Martínez, B.; Aubourg, S.P. Preservative effect on canned mackerel (Scomber colias) lipids by addition of octopus (Octopus vulgaris) cooking liquor in the packaging medium. Molecules 2022, 27, 739. [Google Scholar] [CrossRef] [PubMed]
- Tironi, V.; Tomás, M.; Añón, M.C. Structural and functional changes in myofibrillar proteins of sea salmon (Pseudopercis semifasciata) by interaction with malondialdehyde (RI). J. Food Sci. 2002, 67, 930–935. [Google Scholar] [CrossRef]
- Aubourg, S.P.; Trigo, M.; Martínez, B.; Rodríguez, A. Effect of prior chilling period and alga-extract packaging on the quality of a canned underutilised fish species. Foods 2020, 9, 1333. [Google Scholar] [CrossRef] [PubMed]
- Mohan, C.O.; Remya, S.; Murthy, L.N.; Ravishankar, C.N.; Kumar, K.A. Effect of filling medium on cooking time and quality of canned yellowfin tuna (Thunnus albacares). Food Cont. 2015, 50, 320–327. [Google Scholar] [CrossRef]
- Naseri, M.; Rezaei, M.; Moieni, S.; Hosseini, H.; Eskandari, S. Effects of different filling media on the oxidation and lipid quality of canned silver carp (Hypophthalmichthys molitrix). Int. J. Food Sci. 2011, 46, 1149–1156. [Google Scholar] [CrossRef]
- Barbosa, R.G.; Trigo, M.; Campos, C.A.; Aubourg, S.P. Preservative effect of algae extracts on lipid composition and rancidity development in brine–canned Atlantic chub mackerel (Scomber colias). Eur. J. Lipid Sci. Technol. 2019, 121, 1900129. [Google Scholar] [CrossRef]
- Barbosa, R.G.; Trigo, M.; Fett, R.; Aubourg, S.P. Impact of a packing medium with alga Bifurcaria bifurcata extract on canned Atlantic mackerel (Scomber scombrus) quality. J. Sci. Food Agric. 2018, 98, 3462–3467. [Google Scholar] [CrossRef] [PubMed]
- Küllenberg, D.; Taylor, L.A.; Schneider, M.; Massing, U. Health effects of dietary phospholipids. Lipids Health Dis. 2012, 11, Article 3. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Zhang, T.; Huang, Z.; Luo, X.; Deng, Y. A review on phospholipids and their main applications in drug delivery systems. Asian J. Pharm. Sci. 2015, 10, 81–98. [Google Scholar] [CrossRef]
- Aubourg, S.P.; Medina, I.; Pérez-Martín, R. Polyunsaturated fatty acids in tuna phospholipids: Distribution in the sn-2 location and changes during cooking. J. Agric. Food Chem. 1996, 44, 585–589. [Google Scholar] [CrossRef]
- Takahashi, K.; Inoue, Y. Marine by-product phospholipids as booster of medicinal compounds. Adv. Food Nutr. Res. 2012, 65, 31–46. [Google Scholar]
- Naseri, M.; Rezaei, M. Lipid changes during long-term storage of canned sprat. J. Aquat. Food Prod. Technol. 2012, 21, 48–58. [Google Scholar] [CrossRef]
- Magalhães, J.P.; Müller, M.; Rainger, G.; Steegenga, W. Fish oil supplements, longevity and aging. Aging 2016, 8, 1578–1582. [Google Scholar] [CrossRef] [PubMed]
- Devassy, J.G.; Leng, S.; Gabbs, M.; Monirujjaman, M.; Aukema, H.M. Omega-3 polyunsaturated fatty acids and oxylipins in neuroinflammation and management of Alzheimer disease. Adv. Nutr. 2016, 7, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Swanson, S.; Block, R.; Mousa, S. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ofosu, F.K.; Daliri, E.B.M.; Lee, B.H.; Yu, X. Current trends and future perspectives on omega-3 fatty acids. Res. J. Biol. 2017, 5, 11–20. [Google Scholar]
- Uauy, R.; Valenzuela, A. Marine oils: The health benefits of n-3 fatty acids. Nutrition 2000, 16, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Komprda, T. Eicosapentaenoic and docosahexaenoic acids as inflammation-modulating and lipid homeostasis influencing nutraceuticals: A review. J. Funct. Foods 2012, 4, 25–38. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharm. 2002, 56, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Limia, L.; Cobas, N.; Franco, I.; Martínez-Suárez, S. Fatty acid profiles and lipid quality indices in canned European eels: Effects of processing steps, filling medium and storage. Food Res. Int. 2020, 136, 109601. [Google Scholar] [CrossRef] [PubMed]
- Domiszewski, Z. Effect of sterilization on true retention rate of eicosapentaenoic and docosahexaenoic acid content in mackerel (Scomber scombrus), herring (Clupea harengus), and sprat (Sprattus sprattus) canned products. J. Food Proc. Preserv. 2021, 45, e15461. [Google Scholar] [CrossRef]
- Targueta Barreira, C.F.; Sales de Oliveira, V.; Hidalgo Chávez, D.W.; Domingues Gamallo, O.; Castro, R.N.; Côrrea Damasceno Júnior, P.; Frankland Sawaya, A.C.H.; da Silva Ferreira, M.; Rodrigues Sampaio, G.; Ferraz da Silva Torres, E.A.; Saldanha, T. The impacts of pink pepper (Schinus terebinthifolius Raddi) on fatty acids and cholesterol oxides formation in canned sardines during thermal processing. Food Chem. 2023, 403, 134347. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Joo, S.; Alderton, A.; Hill, D.; Faustman, C. Oxymioglobin and lipid oxidation in yellowfin tuna (Thunnus albacares) loins. J. Food Sci. 2003, 68, 1664–1668. [Google Scholar] [CrossRef]
- Wetterskog, D.; Undeland, I. Loss of redness (a*) as a tool to follow hemoglobin-mediated lipid oxidation in washed cod mince. J. Agric. Food Chem. 2004, 52, 7214–7221. [Google Scholar] [CrossRef] [PubMed]
- Undeland, I.; Hultin, H.; Richards, M. Aqueous extracts from some muscles inhibit hemoglobin-mediated oxidation of cod muscle membrane lipids. J. Agric. Food Chem. 2003, 51, 3111–3119. [Google Scholar] [CrossRef] [PubMed]
- Schubring, R. Comparative study of the DSC pattern, color, texture and water-binding capacity of rainbow trout muscle during heating. J. Food Process. Preserv. 2008, 32, 190–218. [Google Scholar] [CrossRef]
- Mohan, C.O.; Remya, S.; Ravishankar, C.N.; Vijayan, P.K.; Srinivasa Gopal, T.K. Effect of filling ingredient on the quality of canned yellowfin tuna (Thunnus albacares). Int. J. Food Sci. Technol. 2014, 49, 1557–1564. [Google Scholar] [CrossRef]
- Gómez-Limia, L.; Carballo, J.; Rodríguez-González, M.; Martínez, S. Impact of the filling medium on the colour and sensory characteristics of canned European eels (Anguilla anguilla L.). Foods, 2022, 11, 1115. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Free fatty acid (FFA) content of raw and canned horse mackerel previously subjected to different frozen storage times. Mean values of four (n = 4) replicates. Standard deviations are denoted by bars. Different letters (a-d) denote significant differences (p < 0.05).
Figure 1.
Free fatty acid (FFA) content of raw and canned horse mackerel previously subjected to different frozen storage times. Mean values of four (n = 4) replicates. Standard deviations are denoted by bars. Different letters (a-d) denote significant differences (p < 0.05).
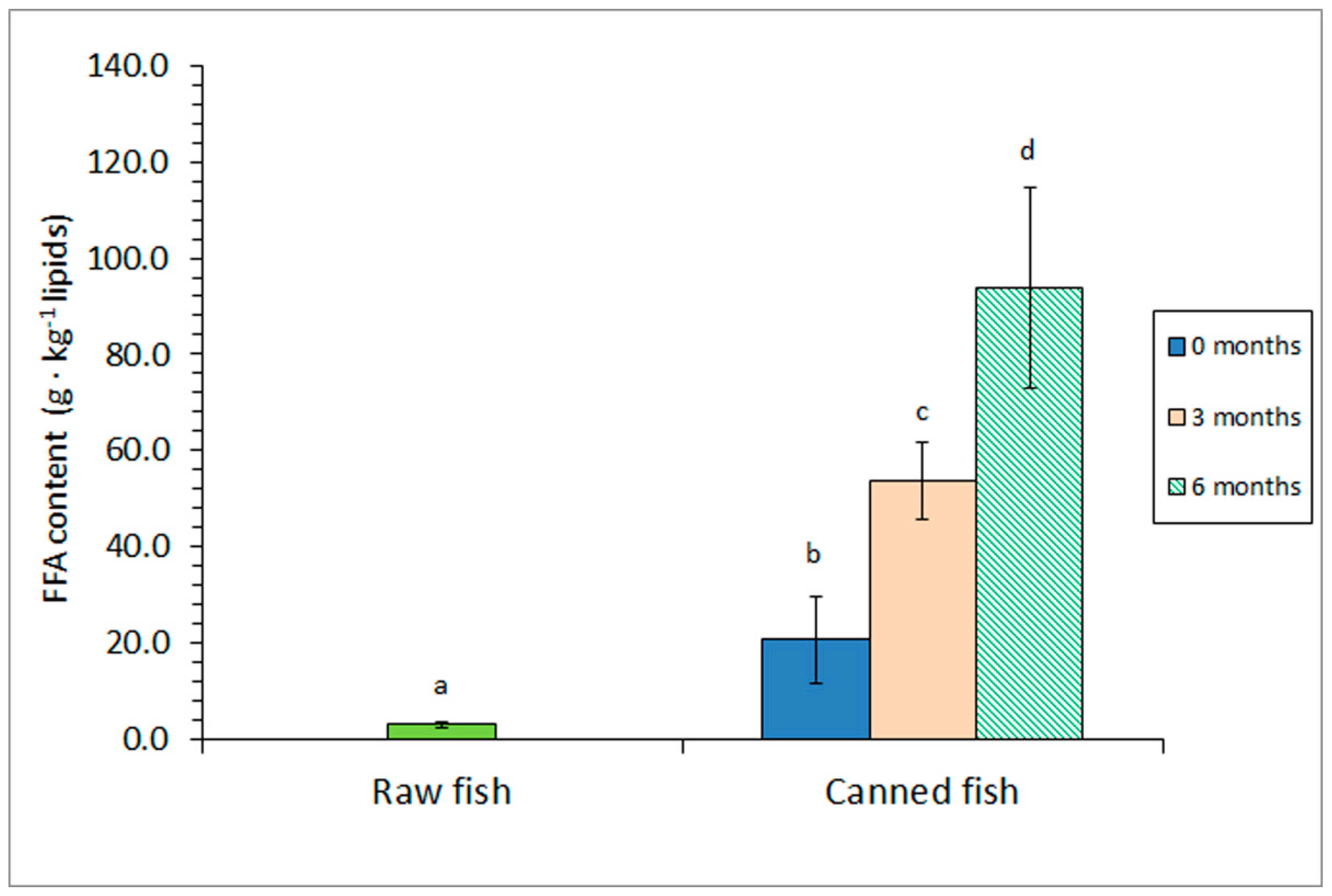
Figure 2.
Phospholipid (PL) content of raw and canned horse mackerel previously subjected to different frozen storage times. Mean values of four (n = 4) replicates. Standard deviations are denoted by bars. Different letters (a,b) denote significant differences (p < 0.05).
Figure 2.
Phospholipid (PL) content of raw and canned horse mackerel previously subjected to different frozen storage times. Mean values of four (n = 4) replicates. Standard deviations are denoted by bars. Different letters (a,b) denote significant differences (p < 0.05).
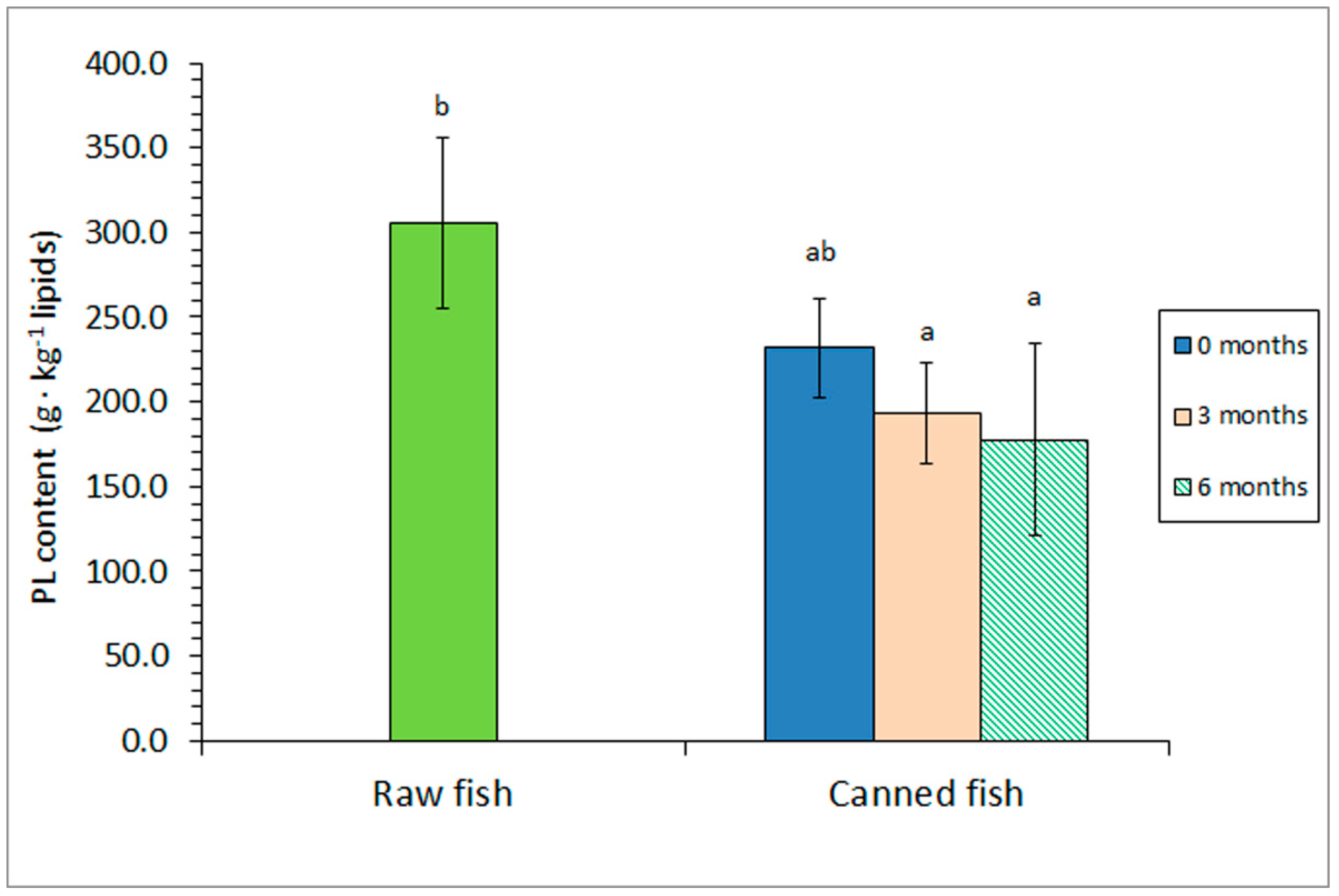
Figure 3.
Trimethylamine (TMA) content of raw and canned horse mackerel previously subjected to different frozen storage times. Average values of four (n = 4) replicates. Standard deviations are expressed by bars. Different letters (a,b) denote significant differences (p < 0.05).
Figure 3.
Trimethylamine (TMA) content of raw and canned horse mackerel previously subjected to different frozen storage times. Average values of four (n = 4) replicates. Standard deviations are expressed by bars. Different letters (a,b) denote significant differences (p < 0.05).
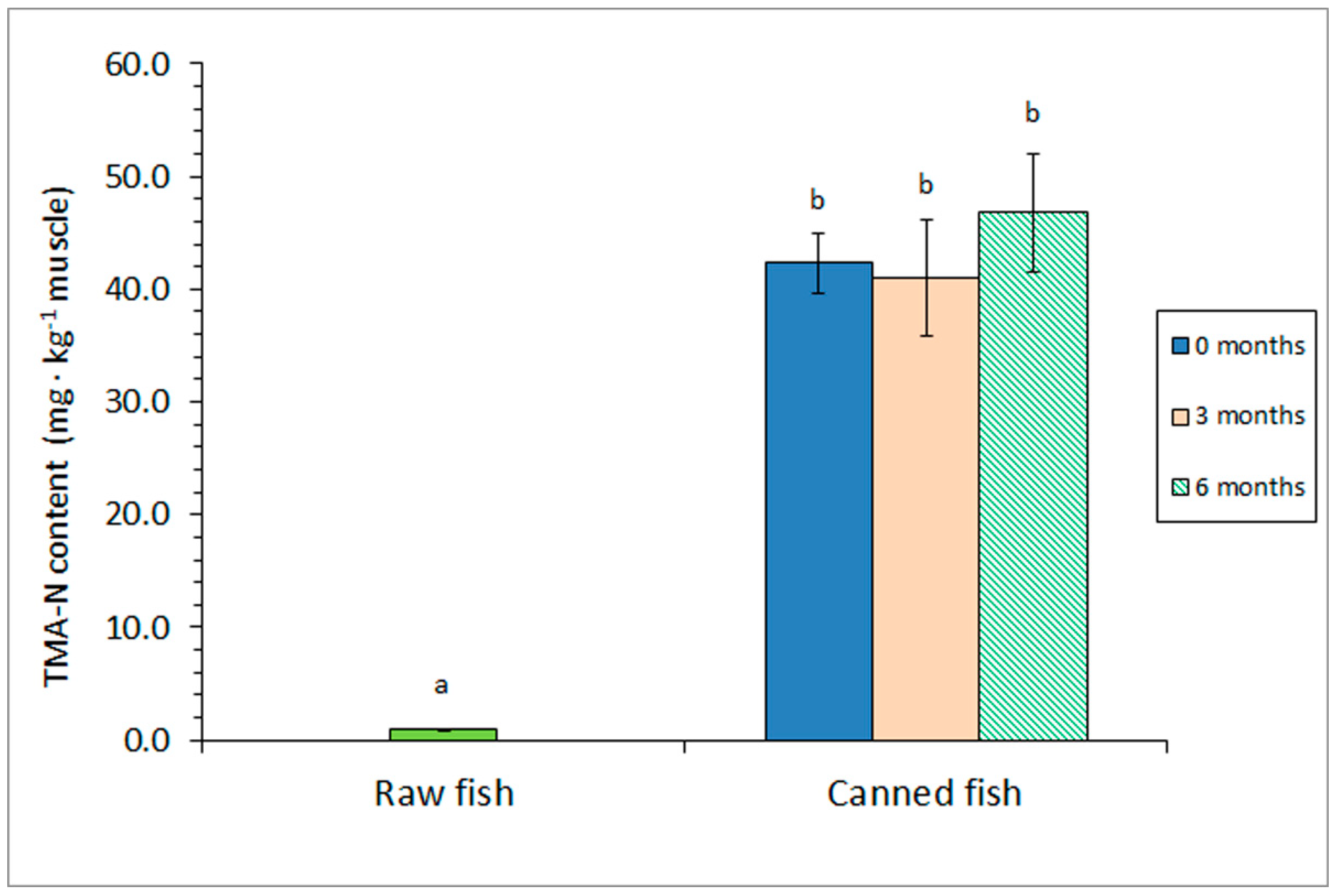

Table 1.
Lipid oxidation of raw and canned horse mackerel previously subjected to different frozen storage times§.
Table 1.
Lipid oxidation of raw and canned horse mackerel previously subjected to different frozen storage times§.
| Lipid quality index | Raw fish | Canned fish | ||
|---|---|---|---|---|
| Prior frozen storage time (months) | ||||
| 0 | 3 | 6 | ||
| Peroxide value (meq active oxygen·kg-1 lipids) | 1.36 a (0.51) |
1.45 a (0.32) |
4.21 b (1.07) |
5.91 b (1.23) |
| Thiobarbituric acid index (mg malondialdehyde·kg-1 muscle) | 0.04 a (0.02) |
0.12 a (0.07) |
0.30 b (0.02) |
0.32 b (0.03) |
| Fluorescence ratio | 1.31 a (0.40) |
3.43 b (0.54) |
4.32 c (0.17) |
4.89 c (0.81) |
§ Mean values of four (n = 4) replicates. Standard deviations are expressed in brackets. Different letters in row (a-c) indicate significant differences (p < 0.05).
Table 2.
Fatty acid (FA) parameters of raw and canned horse mackerel previously subjected to different frozen storage times§.
Table 2.
Fatty acid (FA) parameters of raw and canned horse mackerel previously subjected to different frozen storage times§.
| FA parameter§§ | Raw fish | Canned fish | ||
|---|---|---|---|---|
| Prior frozen storage time (months) | ||||
| 0 | 3 | 6 | ||
| EPA (g·100 g-1 total FAs) | 7.67 a (0.04) |
8.26 ab (1.29) |
8.79 b (0.41) |
8.03 ab (0.40) |
| DHA (g·100 g-1 total FAs) | 23.89 b (2.82) |
19.39 ab (2.87) |
17.04 a (1.71) |
15.91 a (3.13) |
| ω3 (g·100 g-1 total FAs) | 34.45 b (2.87) |
30.50 ab (2.64) |
28.74 a (1.51) |
26.32 a (2.71) |
| ω3/ω6 Ratio | 9.38 b (0.70) |
7.67 a (0.79) |
8.08 ab (0.71) |
7.55 a (0.52) |
| Polyene index | 1.42 c (0.12) |
1.21 bc (0.09) |
1.06 ab (0.07) |
0.92 a (0.11) |
§ Mean values of four (n = 4) replicates. Standard deviations are expressed in brackets. Different letters in row (a-c) denote significant differences (p < 0.05). §§ Abbreviations: EPA (eicosapentaenoic acid) and DHA (docosahexaenoic acid).
Table 3.
Colour parameters of raw and canned horse mackerel previously subjected to different frozen storage times §.
Table 3.
Colour parameters of raw and canned horse mackerel previously subjected to different frozen storage times §.
| Colour parameter | Raw fish | Canned fish | ||
|---|---|---|---|---|
| Prior frozen storage time (months) | ||||
| 0 | 3 | 6 | ||
| L* | 42.14 a (3.19) |
64.86 b (1.15) |
64.03 b (0.86) |
67.43 c (0.62) |
| a* | 4.90 b (0.73) |
0.77 a (0.34) |
1.46 a (0.51) |
0.93 a (0.25) |
| b* | 3.13 a (0.07) |
12.81 b (1.63) |
14.25 bc (2.33) |
16.47 c (1.32) |
§ Mean values of four (n = 4) replicates. Standard deviations are expressed in brackets. Different letters in row (a-c) denote significant differences (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



