
Submitted:
31 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
The emergence of antimicrobial resistance remains one of the greatest public health concerns. Biofilm formation has been postulated as a mechanism of microbial pathogens to resist antimicrobial agents. Lactic Acid Bacteria (LAB) and their metabolites have been proposed to combat bacterial biofilms, due to their antimicrobial activity. In this vein, the aim of the present study was to investigate the biofilm removal potential of cell-free supernatant (CFS) of five wild-type Lacticaseibacillus rhamnosus LAB strains, isolated from Greek natural products, in comparison to the commercial available L. rhamnosus GG strain, formed by common foodborne pathogens (Salmonella Enteritidis, Salmonella Typhimurium, Escherichia coli, Listeria monocytogenes, and Staphylococcus aureus). Biofilm removal activity of LAB was assessed on two-days mature biofilm using a microtiter plate-based procedure. Both non-neutralized and neutralized CFSs removed biofilms in a concentration-dependent manner. Biofilm removal activity of the non-neutralized CFSs was significantly higher compared to the neutralized CFSs, as expected, ranging 40 – 90% and 10 – 78%, respectively. Biofilm removal efficiency of L. rhamnosus OLXAL-3 was signifanctly higher among the wild-type L. rhamnosus strains tested (20 – 100% v/v). In conclusion, our results suggested the great potential of application of wild-type Lacticaseibacillus rhamnosus strains CFSs as effective natural agents against pathogenic bacterial biofilms.
Keywords:
Biofilm removal
; L. rhamonsus
; cell free supernatant
; foodborne pathogens
; probiotics
1. Introduction
The rising anti-microbial resistance of pathogens consists a worldwide risk to human health and thus efficient antimicrobial alternatives are required urgently [1]. Bacterial biofilms, formed by the polymeric metabolites secreted by microbes, are one of the main resistance mechanisms that bacteria utilize to survive against various stresses, including antibiotics, disinfectants, and host defenses. The use of Lactic Acid Bacteria (LAB) to combat bacterial biofilms is a rapidly growing trend. Among others, it is documented that LAB produce several bioactive molecules, such as organic acids, alcohols, carbon dioxide, diacetyl, hydrogen peroxide and bacteriocins [2,3], many of which exert powerful antimicrobial activity [2,4]. Generally, most of these compounds are secreted during cultivation in a broth medium following the proliferation of bacteria cells, known as a supernatant. Hence, it has been observed that the LAB culture supernatant acts efficiently against bacterial biofilms [5,6,7,8]. Thus, probiotics and the antimicrobial agents produced by probiotics have recently been proposed as potential candidates for controlling bacterial biofilm formation against foodborne pathogens [9,10,11].
Bacterial biofilm formation is a consequence of the accumulation and non-reversible attachment of bacterial cells on a biological or non-biological surface, as well as of a body of extracellular polymers (Extracellular Polymeric Substance – ESP) or glycocalyx that is secreted by the same microorganisms [12,13].
In the food industry, microbial biofilms have been found in dairy products, fish and poultry, as well as in the production lines, e.g. of ready-to-eat manufacturing plants [14]. Members of the species Salmonella have been reported to form biofilms on a variety of surfaces and equipment, while Bacillus cereus, Escherichia coli, Shigella sp. and Staphylococcus aureus have been isolated during the production process of dairy products [15]. Furthermore, biofilm formation by E. coli was observed and isolated from surfaces during production of cattle derived meat products [14]. Frequently observed species, related to biofilm formation, are staphylococci, Enterobacteriaceae and the foodborne pathogen Listeria monocytogenes. Therefore, addressing and controlling the formation of microbial biofilms from foodborne pathogens is a challenge for the food industry, where the need to produce safe products is of paramount importance.
Hence, the aim of the present study was to assess biofilm removal potential of cell-free supernatant (CFS) of 5 LAB strains belonging to Lacticaseibacillus rhamnosus species, isolated from traditional Greek foods, formed by common foodborne pathogens.
2. Materials and Methods
2.1. Microbial strains
Five wild-type lactic acid bacteria, whose techonological and functional properties have been described by a previous study [16], isolated from Greek natural products, were used as shown in Table 1. All strains were grown in de Man, Rogosa and Sharpe broth (MRS, Condalab, Spain) at 37oC for 24 h. The commercial Lacticaseibacillus rhamnosus GG served as a reference strain, since its activity against pathogenic biofilm formation is reported in literature [17].
Salmonella enterica subsp. enterica ser. Enteritidis FMCC B56 PT4 (kindly provided by Prof. Nychas G.J.E., Agricultural University of Athens, Athens, Greece), Salmonella enterica subsp. enterica ser. Τyphimurium DSMZ 554, Escherichia coli ATCC 25922, Listeria monocytogenes NCTC 10527 serotype 4b and Staphylococcus aureus ATCC 25923 were grown in Brain Heart Infusion (BHI) broth (Condalab, Spain) at 37oC for 24 h.
2.2. Preparation of LAB CFS (Cell Free Supernatants)
An overnight bacterial culture of each isolated wild-type LAB strain (109 cfu/mL) was centrifuged at 8500 rpm, 4oC for 15 minutes and the supernatant was collected. In order to study the neutralized CFS (pH 7) against bacterial biofilm removal, part of the collected CFS was neutralized using 5M NaOH solution using a pH-meter (WTW Ph 330i, WTW, Germany). All CFS were then sterilized through filtration (0.22μm, Merck, Darmstadt, Germany) (cell free supernatant-CFS).
2.3. Biofilm removal activity
Biofilm removal activity was determined using 24-well polystyrene microtiter plates, according to Koohestani et al. [18] with some modifications. In each well 1800μL Brain Heart Infusion broth and 200μL bacterial suspension after two ten-fold dilutions to obtain a final concentration of 106 logcfu/mL were poured. For the establishment of the bacterial biofilms, the microplates were incubated at 37oC for 48h. After incubation, the free (planktonic) cells were discarded and the wells were washed with ¼ Ringer’s (2.25 g/L NaCl, 0.105 g/L KCl, 0.12 g/L CaCl2H12O6 and 0.05 g/L NaHCO3) sterilized solution to remove any weakly-attached cells. In the next step, CFS (neutralized and non-neutralized) of all LAB strains were gently added to the wells at concentration 100%, 80%, 60%, 40% or 20% and left undisturbed for 1 h at ambient temperature. Then, the CFS were vigorously decanted and the wells were washed again with ¼ Ringer’s sterilized solution. The remaining attached bacterial biofilms were stabilized by the addition of 2000μL of methanol solution and allowed to dry for 5 min. Cell staining was performed using 1% w/v crystal violet (Sigma-Aldrich, St. Louis, USA) for 30 min followed by a final wash with distilled H2O, in order to remove any excess of the staining solution. The remaining crystal violet at the bottom of the wells was diluted in ethanol/acetone solution (Chem-Lab NV, Belgium) at a ratio of 80/20 for 15 min. The solutions were then transferred into clear bottom 96-well microplates to determine the optical absorbance at 540nm. Wells containing only BHI broth and bacterial suspension without CFSs were used as negative and positive controls, respectively.
The estimation of the reduction percentage of biofilms exposed to the different CFSs was calculated according to following equation:
where, C are the OD values of negative control wells, B are the OD values of positive control wells and T are the OD values of CFS treated wells at 540nm.
2.4. Statistical analysis
All experiments were performed at least in quadruple and the results were analyzed for statistical significance with analysis of variance (ANOVA). Duncan’s test was used to determine significant differences [Coefficients, ANOVA tables, and significance (p < 0.05) were computed using Statistica for Windows, v.12.5 (StatSoft, Tulsa, USA)].
3. Results and Discussion
The effect of neutralized and non-neutralized CFSs resulted in significant high removal percentages of biofilms formed by the bacterial species S. Enteritidis, S. Typhimurium, E. coli, L. monocytogenes and S. aureus (Figure 1 and Figure 2). The removal percentage decreased with decreasing CFSs concentrations (from 100 to 20%). The non-neutralized CFSs were significantly more effective at removing the bacterial biofilms (p < 0.05), than the neutralized CFSs.
The non-neutralized CFSs with the greatest activity against S. Enteritidis bacterial biofilms were those of L. rhamnosus OLXAL-3 and L. rhamnosus OLXAL-4 strains (88% and 89%, respectively, p < 0.05), while for S. Tyhpimurium biofilms the most effective CFS was of L. rhamnosus OLXAL-3 strain (88%, p < 0.05) (Figures 1a and 1b). Likewise, the neutralized CFSs with the greatest activity against S. Enteritidis bacterial biofilms were L. rhamnosus CHTH-2 (78%, p < 0.05) and for S. Tyhpimurium of L. rhamnosus OLXAL-3 (79%, p < 0.05) (Figures 2a and 2b).
The above results are in agreement with previous research by Divyashree et al. [19], in which the effect of non-neutralized CFS of Lactobacillus casei MYSRD 108 and Lactobacillus plantarum MYSRD 71 against Salmonella paratyphi bacterial biofilms at 15% v/v CFS concentration was studied, reporting a removal activity over 75% by the non-neutralized Lactobacillus casei MYSRD 108 CFS and 81% by the non-neutralized Lactobacillus plantarum MYSRD 71 CFS of S. paratyphi biofilms, respectively. As expected, neutralized CFSs had significantly lower activity. The greater biofilm removal activity of the non-neutralized CFSs of the two strains was due to the presence of organic acids, according to Divyashree et al. [19]. In another study [20], the non-neutralized CFS of the probiotic strain Weissella confusa WM36 resulted in the removal of 95.68% of Salmonella typhi biofilm at 20% v/v, while the non-neutralized CFS of Weissella viridescens WM33 removed 66.46% of Salmonella Typhimurium biofilms at 15% v/v. Similarly, Tazehabadi et al. [21] studied the activity of Bacillus subtilis KATMIRA 1933 and Bacillus amyloliquefaciens B-1895 CFS against Salmonella enterica subsp. enterica serovar Hadar, Salmonella enterica subsp. enterica serovar Enteritidis phage type 4 and Salmonella enterica subsp. enterica serovar Thompson biofilms. The CFS of Bacillus subtilis KATMIRA 1933 removed 51.1, 48.3 and 56.9% of the biofilms formed by the Salmonella species studied, while the corresponding removal percentages observed by Bacillus amyloliquefaciens B-1895 CFS were 30.4, 28.6 and 35.5%, respectively. The activity against biofilms was not associated with low pH and organic acids production, as the pH of CFS was approximately 5.85 for both strains, but with the production of subtilisin peptides.
The highest removal activity against E. coli bacterial biofilms was noted by non-neutralized L. rhamnosus OLXAL-3 CFS (87%, p < 0.05), while the CFSs of L. rhamnosus OLXAL-3 and L. rhamnosus OLXAL-4 strains were most effective against L. monocytogenes biofilms (84% and 82% removal percentage, respectively, p < 0.05) (Figures 1c and 1d). On the other hand, the most efficient removal of E. coli and L. monocytogenes biofilms was noted for L. rhamnosus GG neutralized CFS (67% and 69%, respectively, p < 0.05) (Figures 2c and 2d).
Abdelhamid et al. [22] studied the activity of non-neutralized CFSs of six probiotic bacteria of the genera Bifidobacterium and Lacticaseibacillus against biofilms of multi-resistant E. coli WW1 and IC2 strains. The CFSs of B. longum and L. plantarum removed 57.94% and 64.57% of E. coli IC2 and E. coli WW1 biofilms, respectively. Furthermore, the skim milk CFS fermented by L. helveticus or L. rhamnosus, separately, removed 31.52 and 17.68% of E. coli IC2 biofilms, respectively, while the corresponding removal percentages recorded by B. longum or L. helveticus CFS were 70.81 and 69.49%. In a similar study published by Apiwatsiri et al. [23], neutralized and non-neutralized CFSs of Lactobacillus plantarum 22F, 25F and Pediococcus acidilactici 72N strains showed significant removal activity against bacterial biofilm strains of E. coli resistant to the antibiotic colistin.
According to the study by Shao et al. [24], non-neutralized CFS of three Leuconostoc mesenteroides strains removed effectively L. monocytogenes biofilms. Leuconostoc species produce bacteriocins, such as mesentericin Y105 produced by Leuconostoc mesenteroides spp. mesenteroides, leucocin A-UAL 187, which is produced by Leuconostoc gelidum, carnosin 44A produced by Leuconostoc carnosum and leuconocin S, which is produced by Leuconostoc paramesenteroides [25]. Bacteriocins of Leuconostoc species possess significantly high removal activity against Listeria biofilms. Moreover, Moradi et al. [26] studied the effect of L. acidophilus LA5 and L. casei 431 CFS on L. monocytogenes biofilms. A greater biofilm removal activity was noted for L. acidophilus CFS and it was found that L. acidophilus CFS contains 1.8% lauric acid, which is considered a possible surfactant, yet the mechanism of action is not fully understood. According to Moradi et al. [26] and in agreement with our results, the action of LAB CFSs in the removal of bacterial biofilms was strain-specific and was due to the presence of compounds, such as exopolysaccharides, organic acids and surfactants.
Regarding the removal activity of non-neutralized CFSs against S. aureus bacterial biofilms, the highest removal percentages were observed by L. rhamnosus OLXAL-2 and L. rhamnosus OLXAL-3 strains (87% and 89%, respectively, p < 0.05) (Figure 1e). On the other hand, the neutralized L. rhamnosus CHTH-2 CFS was the most effective on removing S. aureus bacterial biofilms (70%, p < 0.05) (Figure 2e).
In a recent study, Koohestani et al. [18] explored the removal activity of L. acidophilus LA5 and L. casei 431 CFSs against S. aureus bacterial biofilms. In accordance to our findings, their results highlighted the significant removal activity of both CFSs tested, in a concentration-dependent manner. Of note, the subsequent neutralization of both Lactobacillus CFSs resulted in reduced activity, in agreement to our results, as well as to previous reports [27].
4. Conclusions
A strong and concentration-dependent removal activity of S. Enteritidis, S. Typhimurium, E. coli, L. monocytogenes and S. aureus biofilms was observed by the CFSs of the wild-type L. rhamnosus strains studied. However, their effectiveness was reduced after neutralization, but still remained significant. Thus, more research is still required to identify the presence of other microbial metabolites, such as bacteriocins, hydrogen peroxide, diacetyl, etc., and clarify their effect and mechanism of action in biofilm removal.
Since the food industry is susceptible to the risks associated by biofilm formation and biofilms not only have an impact on food manufacturing operations, but also pose a wide public health risk through the contamination of food products, a better understanding of the mechanisms of their formation in the food chain is essential for developing prevention and control measures.
Author Contributions
Conceptualization, Y.K.; methodology, G.M.; validation, G.M., V.K., G.N. and Y.K.; formal analysis, G.M. and V.K.; investigation, G.M., V.K.., G.N.; resources, Y.K.; data curation, G.M., V.K., G.N. and Y.K.; writing—original draft preparation, G.M., V.K., G.N.; writing—review and editing, Y.K.; visualization, G.M., V.K., G.N and Y.K.; supervision, Y.K.; project administration, Y.K.; funding acquisition, Y.K. All authors have read and agreed to the published version of the manuscript.
Funding
The research work was supported by the Hellenic Foundation for Research and Innovation (H.F.R.I.) under the “First Call for H.F.R.I. Research Projects to support Faculty members and Researchers and the procurement of high-cost research equipment grant” (Project number: HFRI-FM17-2496, acronym: iFUNcultures).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to restrictions of the funding authorities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clinical Microbiology Reviews 2020, 33, 10.1128/cmr.00181-19. [Google Scholar] [CrossRef] [PubMed]
- Šušković, J. , Kos, B., Beganović, J., Pavunc, A., Habjanič, K., & Matošić, S. Antimicrobial Activity – the Most Important Property of Probiotic and Starter Lactic Acid Bacteria. Food Technol Biotechnol 2010, 3, 296–307. [Google Scholar]
- Júnior, W.; Ferrari, Í.; Viana de Souza, J.; Silva, C.; Costa, M.; Dias, F. Characterization and Evaluation of Lactic Acid Bacteria Isolated from Goat Milk. Food Control 2015, 53. [Google Scholar] [CrossRef]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In Vitro Evaluation of the Antimicrobial Activity of a Range of Probiotics against Pathogens: Evidence for the Effects of Organic Acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Ait Ouali, F.; Al Kassaa, I.; Cudennec, B.; Abdallah, M.; Bendali, F.; Sadoun, D.; Chihib, N.-E.; Drider, D. Identification of Lactobacilli with Inhibitory Effect on Biofilm Formation by Pathogenic Bacteria on Stainless Steel Surfaces. Int J Food Microbiol 2014, 191, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Aminnezhad, S.; Kasra-Kermanshahi, R. Antibiofilm Activity of Cell-Free Supernatant from Lactobacillus Casei in Pseudomonas Aeruginosa. Feyz 2014. [Google Scholar]
- Khiralla, G.M.; Mohamed, E.A.H.; Farag, A.G.; Elhariry, H. Antibiofilm Effect of Lactobacillus Pentosus and Lactobacillus Plantarum Cell-Free Supernatants against Some Bacterial Pathogens. J Biotech Res 2015. [Google Scholar]
- Bulgasem, B.Y.; Lani, M.N.; Hassan, Z.; Wan Yusoff, W.M.; Fnaish, S.G. Antifungal Activity of Lactic Acid Bacteria Strains Isolated from Natural Honey against Pathogenic Candida Species. Mycobiology 2016, 44, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Stitt, G.; Son, L.; Enioutina, E.Y. Probiotics and Their Bioproducts: A Promising Approach for Targeting Methicillin-Resistant Staphylococcus Aureus and Vancomycin-Resistant Enterococcus. Microorganisms 2023, 11, 2393. [Google Scholar] [CrossRef]
- Xin, W.-G.; Li, X.-D.; Zhou, H.-Y.; Li, X.; Liu, W.-X.; Lin, L.-B.; Wang, F. Isolation, Antibacterial Characterization, and ATF-Based Preparation of Viable Cells of Lacticaseibacillus Paracasei XLK 401 and Its Potential Application in Milk Preservation. J Dairy Sci 2023, S0022-0302(23)00701-4. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Z.; Li, J.; Qin, G. New Strategies for Biocontrol of Bacterial Toxins and Virulence: Focusing on Quorum-Sensing Interference and Biofilm Inhibition. Toxins (Basel) 2023, 15, 570. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg Infect Dis 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Barzegari, A.; Kheyrolahzadeh, K.; Hosseiniyan Khatibi, S.M.; Sharifi, S.; Memar, M.Y.; Zununi Vahed, S. The Battle of Probiotics and Their Derivatives Against Biofilms. Infect Drug Resist 2020, 13, 659–672. [Google Scholar] [CrossRef]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm Formation in Food Industries: A Food Safety Concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Shi, X.; Zhu, X. Biofilm Formation and Food Safety in Food Industries. Trends in Food Science & Technology 2009, 20, 407–413. [Google Scholar] [CrossRef]
- Nelios, G.; Santarmaki, V.; Pavlatou, C.; Dimitrellou, D.; Kourkoutas, Y. New Wild-Type Lacticaseibacillus Rhamnosus Strains as Candidates to Manage Type 1 Diabetes. Microorganisms 2022, 10, 272. [Google Scholar] [CrossRef]
- Lebeer, S.; De Keersmaecker, S.C.J.; Verhoeven, T.L.A.; Fadda, A.A.; Marchal, K.; Vanderleyden, J. Functional Analysis of luxS in the Probiotic Strain Lactobacillus Rhamnosus GG Reveals a Central Metabolic Role Important for Growth and Biofilm Formation. J Bacteriol 2007, 189, 860–871. [Google Scholar] [CrossRef] [PubMed]
- Kohestani, M.; Moradi, M.; Tajik, H.; Badali, A. Effects of Cell-Free Supernatant of Lactobacillus Acidophilus LA5 and Lactobacillus Casei 431 against Planktonic Form and Biofilm of Staphylococcus Aureus. Vet Res Forum 2018. [Google Scholar] [CrossRef] [PubMed]
- Divyashree, S.; Anjali, P.G.; Somashekaraiah, R.; Sreenivasa, M.Y. Probiotic Properties of Lactobacillus Casei - MYSRD 108 and Lactobacillus Plantarum-MYSRD 71 with Potential Antimicrobial Activity against Salmonella Paratyphi. Biotechnol Rep (Amst) 2021, 32, e00672. [Google Scholar] [CrossRef]
- Pelyuntha, W.; Chaiyasut, C.; Kantachote, D.; Sirilun, S. Cell-Free Supernatants from Cultures of Lactic Acid Bacteria Isolated from Fermented Grape as Biocontrol against Salmonella Typhi and Salmonella Typhimurium Virulence via Autoinducer-2 and Biofilm Interference. PeerJ 2019, 7, e7555. [Google Scholar] [CrossRef]
- Tazehabadi, M.H.; Algburi, A.; Popov, I.V.; Ermakov, A.M.; Chistyakov, V.A.; Prazdnova, E.V.; Weeks, R.; Chikindas, M.L. Probiotic Bacilli Inhibit Salmonella Biofilm Formation Without Killing Planktonic Cells. Front Microbiol 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, A.G.; Esaam, A.; Hazaa, M.M. Cell Free Preparations of Probiotics Exerted Antibacterial and Antibiofilm Activities against Multidrug Resistant E. Coli. Saudi Pharm J 2018, 26, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Apiwatsiri, P.; Pupa, P.; Yindee, J.; Niyomtham, W.; Sirichokchatchawan, W.; Lugsomya, K.; Shah, A.A.; Prapasarakul, N. Anticonjugation and Antibiofilm Evaluation of Probiotic Strains Lactobacillus Plantarum 22F, 25F, and Pediococcus Acidilactici 72N Against Escherichia Coli Harboring Mcr-1 Gene. Front Vet Sci 2021, 8, 614439. [Google Scholar] [CrossRef]
- Shao, X.; Fang, K.; Medina, D.; Wan, J.; Lee, J.; Hong, S.H. The Probiotic, Leuconostoc mesenteroides, Inhibits Listeria monocytogenes Biofilm Formation. Journal of Food Safety 2020, 40, e12750. [Google Scholar] [CrossRef]
- Stiles, M.E. Bacteriocins Produced by Leuconostoc Species. J Dairy Sci 1994, 77, 2718–2724. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Mardani, K.; Tajik, H. Characterization and Application of Postbiotics of Lactobacillus Spp. on Listeria Monocytogenes in Vitro and in Food Models. LWT 2019, 111, 457–464. [Google Scholar] [CrossRef]
- Mahasneh, A.M.; Hamdan, S.; Mahasneh, S.A. Probiotic Properties of Lactobacillus Species Isolated from Local Traditional Fermented Products. JJBS 2015, 8, 81–87. [Google Scholar] [CrossRef]
Figure 1.
Biofilm removal activity of non-neutralized CFSs of five wild-type L. rhamnosus strains in comparison to L. rhamnosus GG against (a) S. Enteritidis, (b) S. Typhimurium, (c) E. coli, (d) L. monocytogenes and (e) S. aureus.
Figure 1.
Biofilm removal activity of non-neutralized CFSs of five wild-type L. rhamnosus strains in comparison to L. rhamnosus GG against (a) S. Enteritidis, (b) S. Typhimurium, (c) E. coli, (d) L. monocytogenes and (e) S. aureus.
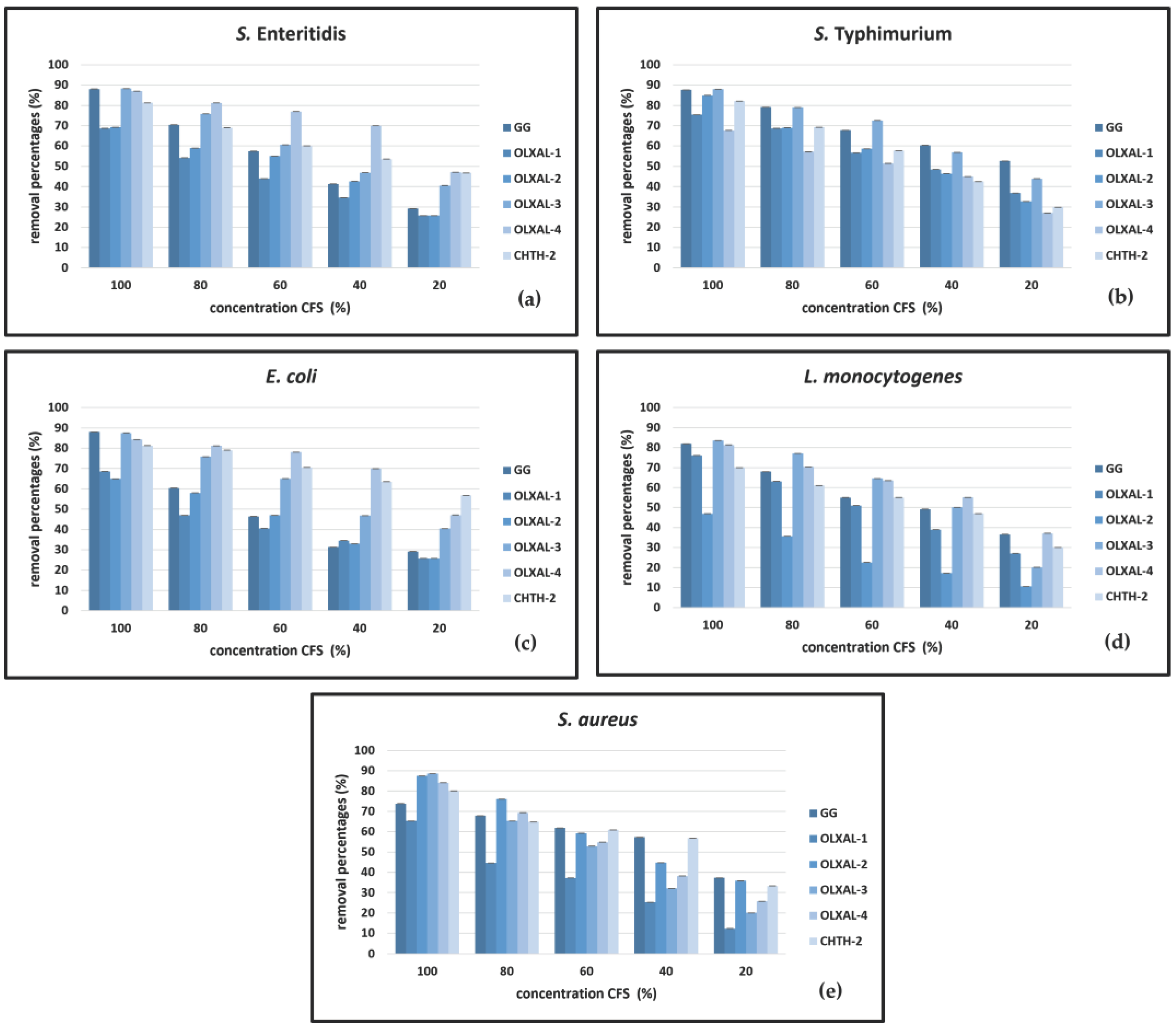
Figure 2.
Biofilm removal activity of neutralized (pH 7) CFSs of five wild-type L. rhamnosus strains in comparison to L. rhamnosus GG against (a) S. Enteritidis, (b) S. Typhimurium, (c) E. coli, (d) L. monocytogenes and (e) S. aureus.
Figure 2.
Biofilm removal activity of neutralized (pH 7) CFSs of five wild-type L. rhamnosus strains in comparison to L. rhamnosus GG against (a) S. Enteritidis, (b) S. Typhimurium, (c) E. coli, (d) L. monocytogenes and (e) S. aureus.
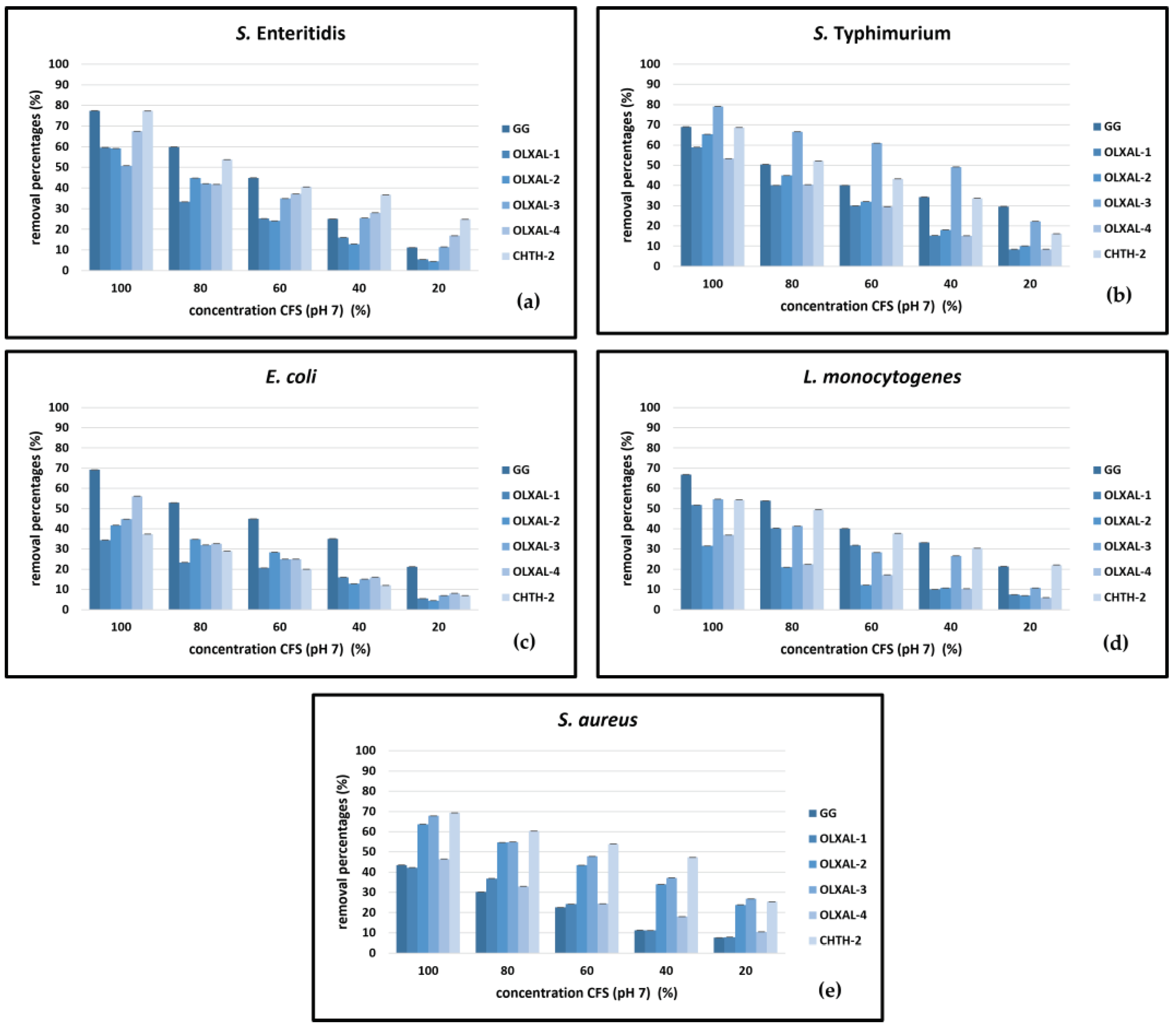

Table 1.
Wild-type Lactic Acid Bacteria (LAB) used in the present study.
| Isolate code | Bacterial Species | Source of isolation |
|---|---|---|
| GG (ATCC 53103) | Lacticaseibacillus rhamnosus | Human intestines |
| OLXAL-1 | Lacticaseibacillus rhamnosus | Olive (fruit) |
| OLXAL-2 | Lacticaseibacillus rhamnosus | Olive (fruit) |
| OLXAL-3 | Lacticaseibacillus rhamnosus | Olive (fruit) |
| OLXAL-4 | Lacticaseibacillus rhamnosus | Olive (fruit) |
| CHTH-2 | Lacticaseibacillus rhamnosus | Feta-type cheese |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



