
Submitted:
31 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
In tropical areas, simultaneous transmission of several vector-borne diseases is common due to ecological factors that are shared by arthropod vectors. Malaria and dengue virus, transmitted by Anopheles and Aedes mosquitoes, respectively, are among the top vector-borne diseases that cause significant morbidity and mortality in endemic areas. Notably, tropical areas often have suitable conditions for the co-existence of these mosquito species, highlighting the importance of identifying markers that accurately indicate the risk of acquiring each specific disease entity. Aedes are daytime-biting mosquitoes, while Anopheles preferentially bite during the night. These biting patterns raises the possibility of concurrent exposure to bites from both species. This pattern is important since mosquito saliva, deposited in the skin during blood feeding, induces immune responses that modulate pathogen establishment and infection. Previous studies have focused on characterizing such effects on the vector-pathogen interface for an individual pathogen and its mosquito vector. In this study, we evaluated associations between immune responses to salivary proteins from non-dengue and non-malaria vector mosquito species with clinical characteristics of malaria and dengue, respectively. Surprisingly, antibody responses against Anopheles antigens in dengue patients correlated with red blood cell count and hematocrit, while antibody responses against Aedes proteins were associated with platelet count in malaria patients. Our data indicate that concurrent exposure to multiple disease-carrying mosquito vectors and their salivary proteins with differing immunomodulatory properties could influence the transmission, pathogenesis, and clinical presentation of malaria, dengue fever, and other vector-borne illnesses.
Keywords:
Aedes
; dengue
; Anopheles
; biomarkers.
; mosquito saliva
1. Introduction
Malaria and dengue fever are two of the most important vector-borne diseases of public health concern in tropical areas around the globe[1]. Dengue virus (DENV) has periodic outbreaks in Central and South America[2], with occasional outbreaks in the US, particularly in Texas and Florida[3,4]. Importantly, several recent reports have indicated the occurrence of local malaria transmission in the continental US, while this disease remains endemic in Latin America[5,6].
Mosquito-borne pathogens are usually deposited in the skin of the vertebrate host along with arthropod saliva during blood-feeding[7]. Compelling evidence suggests that mosquito saliva induces profound changes in immune responses locally (at the bite site) and systemically, eventually providing a vehicle for pathogen transmission[8,9]. Furthermore, salivary secretions are well known to induce significant antibody responses in the vertebrate host. These antibody responses may vary depending on factors such as age, seasonality, and vector abundance[10,11]. Several studies suggest that IgG responses to mosquito salivary proteins may serve as surrogate biomarkers for exposure to mosquito bites and an indirect marker for disease risk in travelers and individuals living in endemic areas[12-14].
We and others have previously shown that not all proteins in mosquito saliva are associated with disease risk[13], and that only a few proteins are suitable as markers for vector-bite exposure[15,16]. However, a small number of proteins may be useful in determining susceptibility to infection. For instance, we recently identified a D7 Long (D7L) (AAEL006424) salivary protein from Ae. aegypti that was capable of physically binding virions and inhibiting DENV infection in vitro and in vivo [17]. Interestingly, measurement of IgG antibodies against this D7L protein revealed higher anti-D7L antibody levels in people with active DENV infection compared to febrile DENV uninfected individuals and healthy participants[17]. In contrast, antibodies against AgBR1 and NeST1, two immunomodulatory mosquito salivary proteins, were higher in healthy individuals compared to DENV-infected individuals[18]. These data suggest that targeting the correct salivary proteins in vaccine formulations to induce specific antibodies may prevent or modify the course of mosquito-borne infections.
Immune responses to mosquito saliva can vary according to epidemiological factors such as age, sex, seasonality, use of vector control, etc. Interestingly, previous studies found higher IgG antibody levels after mosquito exposure in males compared to females[19], highlighting a potential sex-dependent incidence of mosquito-transmitted diseases. Such findings have been partially explained by physiological factors, host genetics, and gender-related social determinants, resulting in differences in exposure[20,21]. With the increase in global mosquito-borne infections, a better understanding of sex-dependent host responses may be critical to mitigating the negative consequences of vector-pathogen transfer and hormone-related antigen responses.
Currently, most arthropod-borne disease control relies heavily on decreasing human-vector contact through physical devices (i.e., bed nets) or insecticide treatment[22,23]. Effective vaccines or drugs against arthropod-borne diseases are scarce[24,25]. Importantly, the increase in insecticide and drug resistance calls for the design and implementation of new tools for disease control, and new protocols to measure exposure to arthropod vectors and estimate the potential risk of acquisition of vector-borne diseases, which will guide public health policy. Recently, an Ae. aegypti peptide, Nterm-34kDa, was evaluated as a quantitative measure of exposure to Aedes bites, and a positive correlation was observed between intensity of exposure, mosquito abundance, and anti-Nterm-34kDa IgG antibody levels[26] [27]. Also, the gSG6-P1 peptide, identified from the An. gambiae SG6 protein[28] has been extensively validated as a biomarker of exposure against Anopheles mosquitoes from the subgenus Cellia and Anopheles[29]. Both peptides have been used successfully to determine the level of exposure to mosquito bites associated with mosquito control interventions or risk of disease[15,30,31].
In tropical areas, people can be exposed to hundreds of mosquito bites per day, and an increase in mosquito abundance is often associated with an increase in vector-borne pathogen transmission[32]. Importantly, mosquito species are very particular in their feeding behavior. For instance, Anopheles mosquitoes are preferential nocturnal feeders, while Aedes mosquitoes are diurnal biters. Therefore, it is possible that an individual may sustain Aedes mosquito bites during the day, followed by Anopheles mosquito bites at night. To our knowledge, there are currently no studies that describe the effect of exposure to bites from different mosquito species on skin immune responses and pathogen replication. However, prior work suggests that saliva induces skin responses associated with the potential for pathogen establishment [33]. Also, recent studies suggest that saliva from different mosquito species may have variable impacts on the same pathogen[34]. Thus, there is a critical need to better understand how contact with salivary proteins from different mosquito species due to sequential exposure to diurnal and nocturnal biters may impact human arbovirus acquisition and anti-viral immune responses.
In this study, our primary goal was to evaluate the levels of antibodies against salivary antigens of different arthropod vectors of human disease in people with either malaria or dengue to assess whether responses against vector saliva are associated with blood parameters leading to severe clinical presentation. Since it is rare that only a single species of mosquito is found in a specific area, and several arthropod-borne diseases are usual in the tropics, we hypothesized that exposure to salivary proteins with different immunomodulatory properties would impact infection and progression to disease. Malaria and dengue fever transmission often co-occur in tropical areas, mainly because of the overlap in the ecological niches preferred by the main vectors of these infections[35-37]. We therefore leveraged our ongoing malaria and dengue surveillance study in Norte de Santander to evaluate exposure to Anopheles and Aedes mosquito saliva and compare these data with blood parameters at diagnosis. To our knowledge, this is the first study exploring a potential correlation between exposure to saliva of non-vectors in clinical presentation of disease and severity.
2. Results
2.1. P. vivax Malaria and Exposure to the Non-Malaria vector Aedes Aegypti
A total of 49 participants with current P. vivax malaria infection, confirmed by microscopy and rapid diagnostic test (RDT), from the areas of Tarra and Tibu in Norte de Santander, Colombia, were included in the study from 2018 to 2019 (Table 1). Mean parasite count was 6,665 parasites/µL (from 420 to 26,480 parasites/µL). We found that antibody levels were not correlated with parasite count or expression of the Pvs25 gene (gametes, ookinetes); however, IgG antibodies against An. albimanus SGE (Spearman correlation r=-0.6099, p=0.0269), Trans-1 (ρ=-0.7510, p=0.0031), gsG6-P1 (ρ=-0.6648, p=0.0132) and Nterm24kDa (Spearman correlation r=-0.7253, p=0.0050) showed a significant negative correlated with the level of expression of Pvs230 (gametocytes) (Table 2).

Table 1.
Description of the study population age in the malaria endemic areas (Tibu and Tarra, Norte de Santander – Colombia), dengue endemic area (Los Patios and Cucuta, Norte de Santander – Colombia); and healthy individuals from Los Patios (Colombia) and Manhattan – Kansas (USA).
Table 1.
Description of the study population age in the malaria endemic areas (Tibu and Tarra, Norte de Santander – Colombia), dengue endemic area (Los Patios and Cucuta, Norte de Santander – Colombia); and healthy individuals from Los Patios (Colombia) and Manhattan – Kansas (USA).
| Infection Status | All age years (range) | Female (range) | Male (Range) |
|---|---|---|---|
| Malaria | 33.4 (1 – 67), n=49 | 36.0 (16 – 53), n=14 | 32.4 (1 – 67), n=35 |
| Dengue | 15.3 (1 – 76), n=124 | 17.6 (1 – 76), n=70 | 12.4 (1 – 69), n=54 |
| Healthy | 28.9 (2 – 79), n=103 | 27.0 (2 – 79), n=65 | 32.3 (2 – 72), n=38 |
Table 2.
Correlation analysis between IgG antibody responses against each peptide and parasite count/gametocytemia by gender in Plasmodium positive volunteers. Data is presented in Spearman correlation r.
Table 2.
Correlation analysis between IgG antibody responses against each peptide and parasite count/gametocytemia by gender in Plasmodium positive volunteers. Data is presented in Spearman correlation r.
| Peptide | Pvs25 | Pvs230 | Parasite count |
|---|---|---|---|
| All | |||
| Peroxi-P1 | 0.0121 (p=0.9415) |
-0.2842 (p=0.0795) |
-0.0989 (p=0.5492) |
| Trans-1 | 0.0715 (p=0.6655) |
-0.1355 (p=0.4107) |
-0.1987 (p=0.2254) |
| Trans-2 | -0.1937 (p=0.2373) |
-0.0644 (p=0.6970) |
-0.0984 (p=0.5511) |
| An. albimanus SGE | -0.0497 (p=0.7638) |
-0.3072 (p=0.0571) |
-0.0538 (p=0.7451) |
| gSG6-P1 | -0.0901 (p=0.5855) |
-0.2913 (p=0.0720) |
-0.1123 (p=0.4960) |
| Nterm-34kDa | -0.0109 (p=0.9473) |
-0.1553 (p=0.3453) |
-0.0314 (p=0.8495) |
| Females | |||
| Peroxi-P1 | -0.0824 (p=0.7890) |
-0.5440 (p=0.0546) |
-0.2418 (p=0.4262) |
| Trans-1 | -0.3989 (p=0.1770) |
-0.7510 (p=0.0031) |
-0.1761 (p=0.5650) |
| Trans-2 | 0.1978 (p=0.5171) |
-0.3187 (p=0.2886) |
-0.0165 (p=0.9574) |
| An. albimanus SGE | -0.2802 (p=0.3538) |
-0.6099 (p=0.0269) |
-0.1593 (p=0.6031) |
| gSG6-P1 | -0.3132 (p=0.2974) |
-0.6648 (p=0.0132) |
-0.1593 (p=0.6031) |
| Nterm-34kDa | -0.5000 (p=0.0819) |
-0.7253 (p=0.0050) |
-0.3077 (p=0.3064) |
| Males | |||
| Peroxi-P1 | 0.0510 (p=0.8047) |
-0.1772 (p=0.3866) |
0.0395 (p=0.8480) |
| Trans-1 | 0.2113 (p=0.3001) |
0.1217 (p=0.5356) |
-0.1498 (p=0.4651) |
| Trans-2 | 0.2205 (p=0.2790) |
0.1022 (p=0.6193) |
-0.0728 (p=0.7236) |
| An. albimanus SGE | 0.0168 (p=0.9353) |
-0.1733 (p=0.3971) |
-0.0147 (p=0.9432) |
| gSG6-P1 | -0.0544 (p=0.7919) |
-0.1433 (p=0.4850) |
-0.0332 (p=0.8722) |
| Nterm-34kDa | 0.1829 (p=0.3711) |
0.0715 (p=0.7287) |
0.2151 (p=0.2913) |
We next tested the correlation between IgG antibody levels against the salivary proteins with blood parameters such as red blood cell (RBC) count, white blood cell (WBC) count, platelet count, hemoglobin, hematocrit, and parasite count (Table 3). Interestingly, we found that levels of antibodies against the Aedes peptide Nterm34kDa (non-malaria vector) were significantly negatively correlated with RBC count in males, while females presented a significant positive correlation between anti-Nterm34kDa antibodies and WBC count. We also compared antibody levels with age and observed a positive correlation with the level of antibodies against whole An. albimanus SGE, gSG6-P1, and Trans1 in females but not males (Table 4). When measuring odds ratios, we found that malaria patients with normal leucocyte counts were 3.3 times more likely to present high IgG antibodies against the gSG6-P1 peptide (Figure 1A), and people with low parasitemia were 3.3 times more likely to have low antibodies against Trasferrin 2 (Figure 1B).
Figure 1.
Odd ratios comparing levels of IgG antibodies against gSG6-P1 and Transferrin 2 peptide in people with and without leukemia (A) and low and high parasitemia levels (B).
Figure 1.
Odd ratios comparing levels of IgG antibodies against gSG6-P1 and Transferrin 2 peptide in people with and without leukemia (A) and low and high parasitemia levels (B).
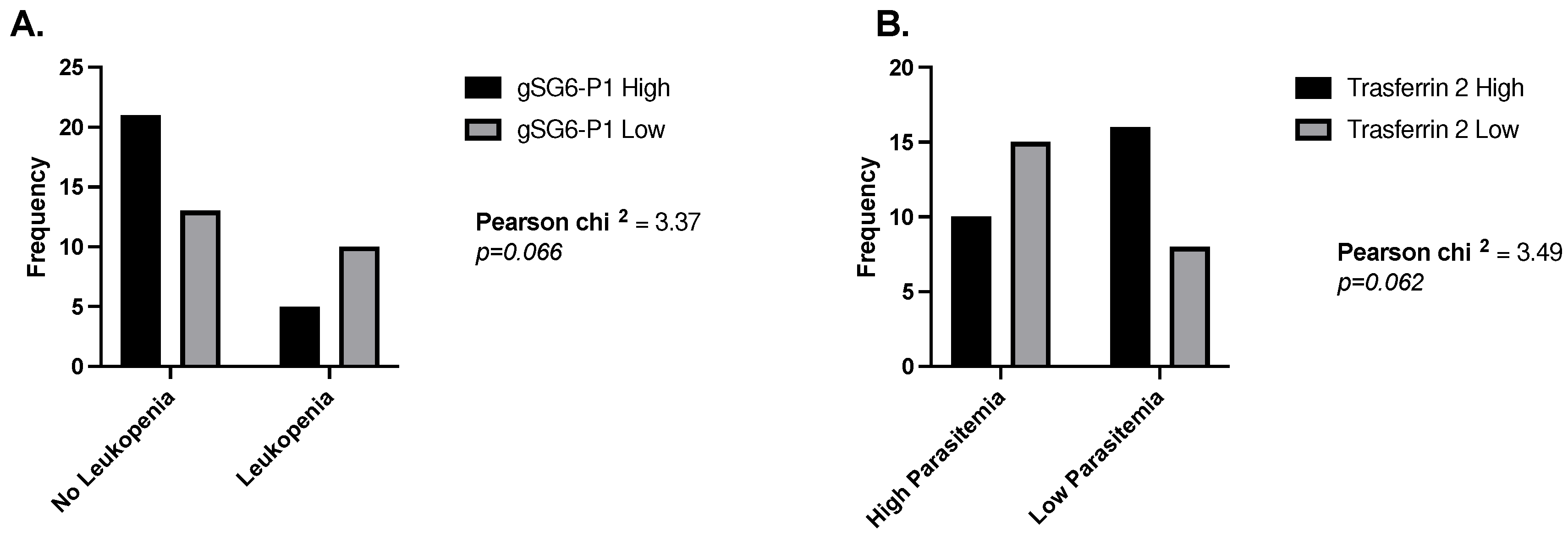
Table 3.
Correlation analysis between IgG antibody responses against each peptide and blood parameters by gender in Plasmodium positive volunteers. Data is presented in Spearman correlation r.
Table 3.
Correlation analysis between IgG antibody responses against each peptide and blood parameters by gender in Plasmodium positive volunteers. Data is presented in Spearman correlation r.
| Peptide | Red blood cell count | White blood cell count | Platelet count | Haemoglobin | Haematocrit |
|---|---|---|---|---|---|
| All | |||||
| Peroxi-P1 | -0.2660 (p=0.0847) |
-0.0269 (p=0.8642) |
-0.3167 (p=0.0385) |
-0.1691 (p=0.2782) |
-0.2145 (p=0.1671) |
| Trans-1 | -0.2462 (p=0.1115) |
0.2950 (p=0.0548) |
-0.3288 (p=0.0314) |
-0.1691 (p=0.2782) |
-0.2445 (p=0.1141) |
| Trans-2 | -0.2651 (p=0.0858) |
0.1180 (p=0.4510) |
-0.3651 (p=0.0161) |
-0.2651 (p=0.0974) |
-0.3740 (p=0.0135) |
| An. albimanus SGE | -0.1076 (p=0.4923) |
0.1589 (p=0.3088) |
-0.311 (p=0.0420) |
-0.1158 (p=0.4597) |
-0.1609 (p=0.3028) |
| gSG6-P1 | -2769 (p=0.0723) |
0.3205 (p=0.0361) |
-0.2636 (p=0.0877) |
-0.1189 (p=0.4477) |
-0.1300 (p=0.4061) |
| Nterm-34kDa |
-0.3288 (p=0.0313) |
0.2363 (p=0.1271) |
-0.2078 (p=0.1812) |
-0.1402 (p=0.3699) |
-0.1374 (p=0.3797) |
| Females | |||||
| Peroxi-P1 | -0.3714 (p=0.1910) |
0.6497 (p=0.0119) |
-0.4330 (p=1220) |
-0.1454 (p=0.62000 |
-0.2571 (p=0. 3748) |
| Trans-1 | -0.1958 (p=0.5023) |
0.7522 (p=0.0019) |
-0.5391 (p=0.0467) |
0.1235 (p=0.6741) |
-0.0726 (p=0.8052) |
| Trans-2 | -0.3099 (p=0.2809) |
0.6144 (p=0.0194) |
-0.2308 (p=0.4273) |
-0.1410 (p=0.6307) |
-0.3099 (p=0.2809) |
| An. albimanus SGE | -0.0330 (p=0.9109) |
0.6763 (p=0.0079) |
-0.4857 (p=0.0783) |
0.1828 (p=0.5316) |
0.0857 (p=0.7708) |
| gSG6-P1 | -0.0857 (p=0.7708) |
0.6188 (p=0.0153) |
-0.3626 (p=0.2026) |
0.1564 (p=0.5834) |
0.1121 (p=0.7028) |
| Nterm-34kDa | -01560 (p=0.5942) |
0.5923 (p=0.0256) |
-0.5560 (p=0.0389) |
-0.0683 (p=0.8166) |
-0.0989 (p=0.7366) |
| Males | |||||
| Peroxi-P1 | -0.3224 (p=0.0880) |
-0.2376 (p=0.2145) |
-0.2266 (p=0.2371) |
-0.2902 (p=0.1268) |
-0.2572 (p=0.1780) |
| Trans-1 | -0.2645 (p=0.1656) |
0.1199 (p=0.5356) |
-0.02303 (p=0.2294) |
-0.2740 (p=0.1504) |
-0.3069 (p=0.1053) |
| Trans-2 | -0.2334 (p=0.2230) |
-0.0712 (p=0.7135) |
-0.3548 (p=0.0589) |
-0.2930 9p=0.1229) |
-0.3972 (p=0.0329) |
| An. albimanus SGE | -0.1429 (p=0.4595) |
0.0094 (p=0.9615) |
-0.1947 (p=0.3116) |
-0.2169 (p=0.2584) |
-0.2341 (p=0.2216) |
| gSG6-P1 | -0.3300 (p=0.0804) |
0.2192 (p=0.2534) |
-0.1062 (p=0.5834) |
-0.1468 (p=0.4443) |
-0.1417 (p=0.4634) |
| Nterm-34kDa |
-0.4171 (p=0.0244) |
0.0561 (p=0.7727) |
-0.0017 (p=0.9929) |
-0.1594 (p=0.4090) |
-0.1264 (p=0.5135) |
Table 4.
Correlation analysis between IgG antibody responses against each peptide and age by gender in in P. vivax infected patients. Data is presented in Spearman correlation r.
Table 4.
Correlation analysis between IgG antibody responses against each peptide and age by gender in in P. vivax infected patients. Data is presented in Spearman correlation r.
| Peptide | Age |
|---|---|
| All | |
| Peroxi-P1 | -0.0207 (p=0.8880) |
| Trans-1 |
0.3270 (p=0.0218) |
| Trans-2 | 0.1596 (p=2733) |
| An. albimanus SGE | 0.0471 (p=0.07478) |
| gSG6-P1 | 0.2064 (p=0.1547) |
| Nterm-34kDa | 0.0828 (p=0.5719) |
| Females | |
| Peroxi-P1 | 0.3645 (p=0.2001) |
| Trans-1 |
0.6196 (p=0.0181) |
| Trans-2 | 0.3934 (p=0.1641) |
| An. albimanus SGE |
0.5934 (p=0.0253) |
| gSG6-P1 |
0.5334 (p=0.0495) |
| Nterm-34kDa | 0.2356 (p=0.4175) |
| Males | |
| Peroxi-P1 | -0.1172 (p=0.5027) |
| Trans-1 | 0.2603 (p=0.1310) |
| Trans-2 | 0.0793 (p=0.6507) |
| An. albimanus SGE | -0.0771 (p=0.6599) |
| gSG6-P1 | 0.1113 (p=0.5243) |
| Nterm-34kDa | 0.0417 (p=0.8121) |
2.2. DENV and Exposure to the Non-DENV vector Anopheles Albimanus
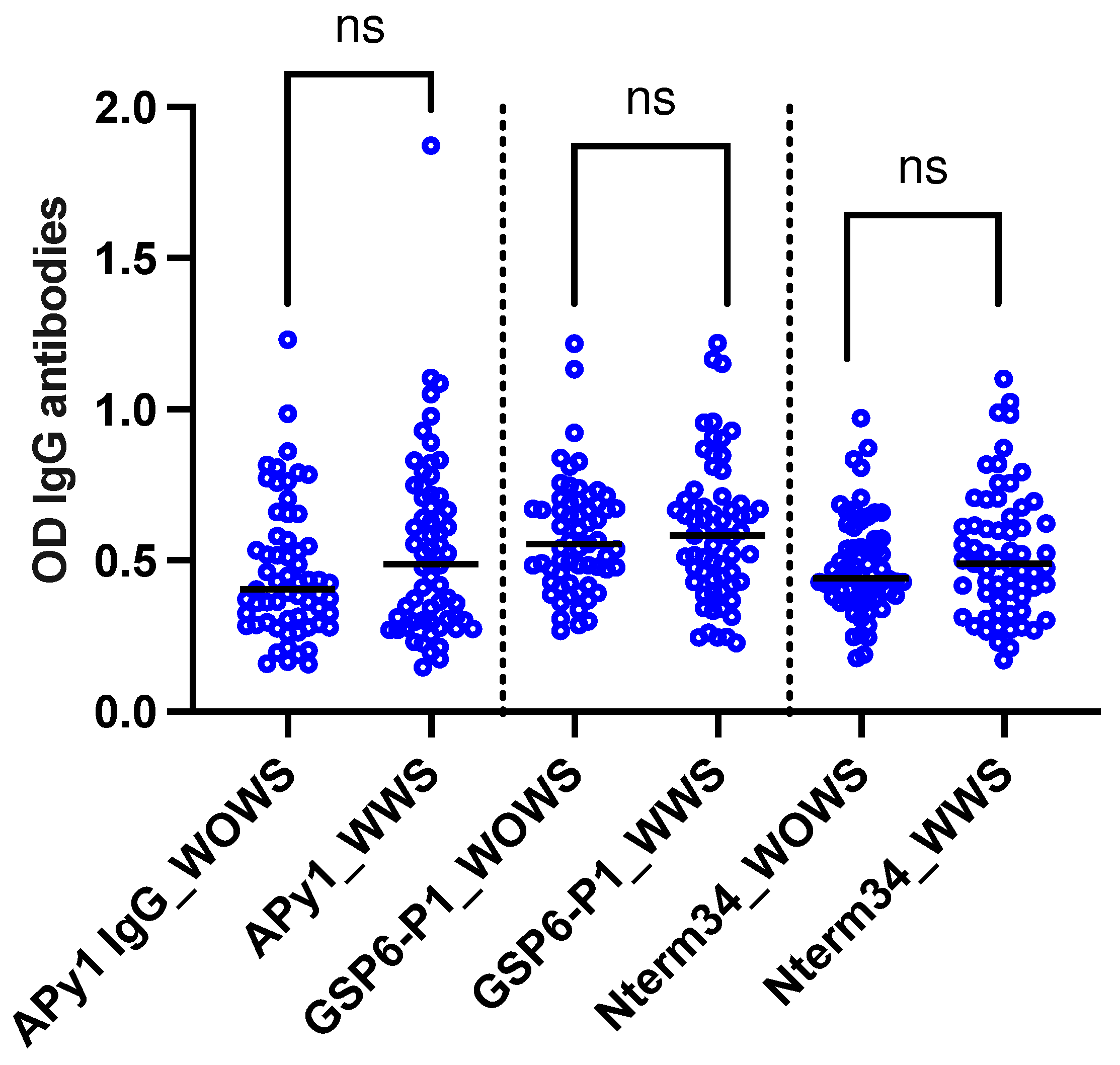
A total of 124 DENV-positive volunteers living in Los Patios (n=75), Cucuta (n=24), and Ocana (n=25) between October 2018 and September 2020 were included in this study. Following DENV classification according to WHO guidelines, we included 63 DENV patients with warning signs and 61 DENV patients without warning signs. No significant differences were observed in the level of IgG antibody levels against Nterm34kDa, AnDarApy1, and gSG6-P1 peptides when comparing DENV groups Mann-Whitney test, p>0.05) (Figure 2). However, we observed a significant positive correlation between RBC count and IgG antibody levels against Nterm34kDa (Spearman correlation sp=-0.2107, p=0.0193) and gSG6-P1 (Sρ=-0.1807, p=0.0455). Significant associations between the levels of antibodies against salivary peptides and blood parameters were observed in males but not females. All comparisons are found in Table 5.
Figure 2.
Scatterplot of IgG antibodies against the mosquito salivary peptides Nterm34kDa, AnDarApy1 and gSG6-P1 in people with dengue fever with warning signs (WWS) and dengue without warning signs. Mann-Whitney test, p>0.05.
Figure 2.
Scatterplot of IgG antibodies against the mosquito salivary peptides Nterm34kDa, AnDarApy1 and gSG6-P1 in people with dengue fever with warning signs (WWS) and dengue without warning signs. Mann-Whitney test, p>0.05.
2.3. Healthy Individuals from Endemic and Nonendemic Areas
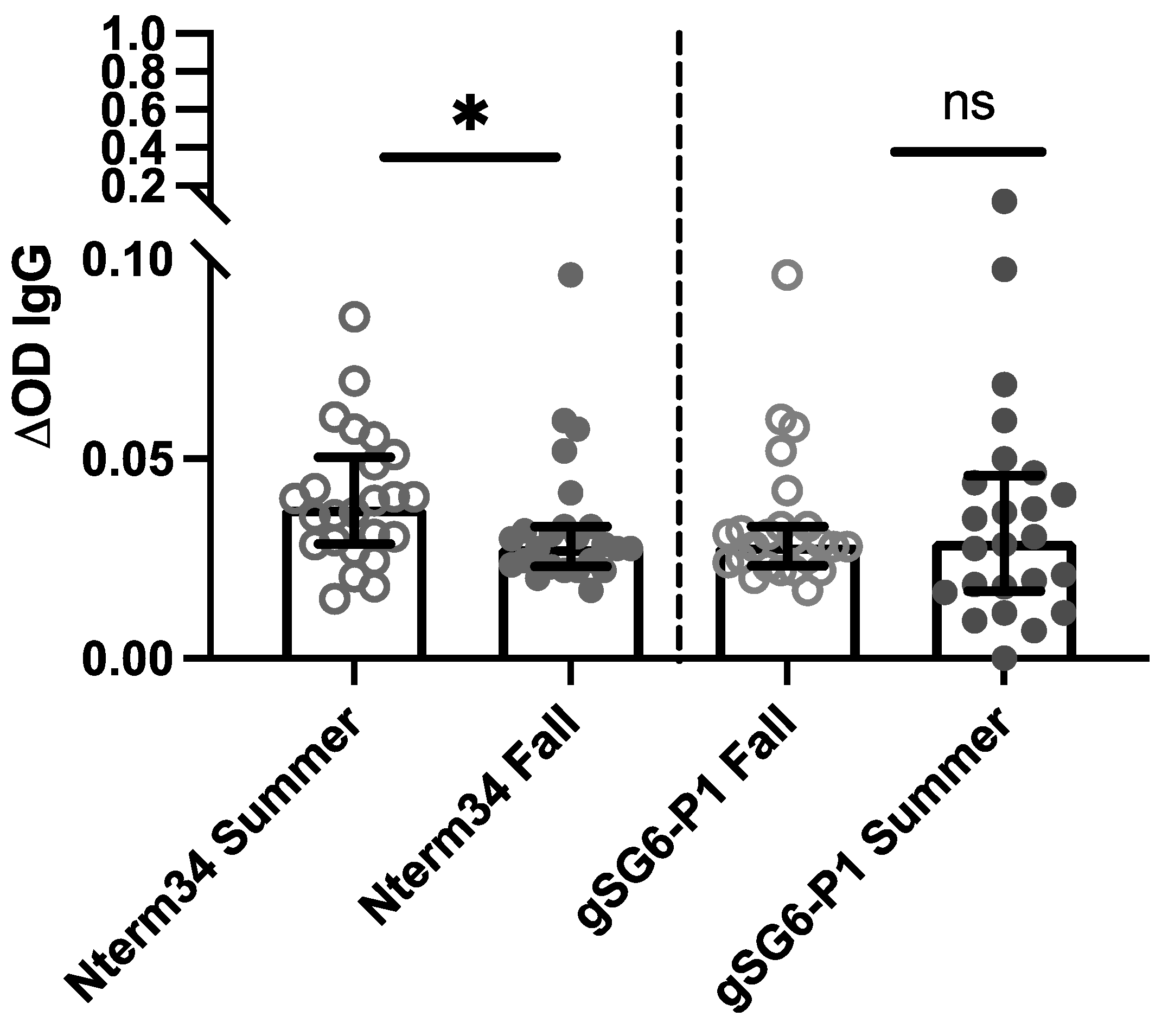
In 2018, we recruited participants in Manhattan, Kansas for a study focused on evaluating exposure to blood-sucking arthropods. A total of 27 participants donated blood samples in the summer (June – August) and fall (September – October). Our analyses revealed a significant reduction in antibody levels against the Nterm-34kDa, but not for the gSG6-P1 in the Fall (Figure 3), suggesting an association between the intensity of exposure to mosquito bites and IgG antibody levels against mosquito salivary peptides. Next, we compared antibody levels in healthy US participants with levels in healthy individuals living in areas with endemic DENV and malaria. In 2018, samples were collected from 54 healthy volunteers living in houses where a DENV case was reported in Los Patios – Norte de Santander (Colombia). We observed a significant negative correlation between age and IgG antibodies against Nterm34kDa (ρ=0.4182, p=0.000) and gSG6-P1 (ρ=-0.3553, p=0.0003). This significant negative correlation between age and anti-Nterm34kDa remained even when stratifying the data by gender (Table 5). When stratifying data by location, we observed that people from the US demonstrated significant negative correlations between antibody levels against both peptides and age, while healthy individuals from Colombia showed a positive correlation between age and gSG6-P1, but not Nterm-34kDa (Table 6).
Table 5.
Correlation analysis between IgG antibody responses against each peptide and blood parameters by gender. In dengue fever patients. Data is presented in Spearman correlation r.
Table 5.
Correlation analysis between IgG antibody responses against each peptide and blood parameters by gender. In dengue fever patients. Data is presented in Spearman correlation r.
| Peptide | Red blood cell count | White blood cell count | Platelet count | Haemoglobin | Haematocrit |
|---|---|---|---|---|---|
| All | |||||
| AnDarApy-1 | 0.0955 (p=0.2934) |
0.0277 (p=0.7607) |
-0.0831 (p=0.3609) |
0.0999 (p=0.2715) |
0.1092 (p=0.2294) |
| gSG6-P1 |
0.1807 (p=0.0455) |
-0.0829 (p=0.3617) |
-0.0484 (p=0.5949) |
0.0921 (p=0.3112) |
-0.1097 (p=0.2269) |
| Nterm-34kDa |
0.2107 (p=0.0193) |
-0.0429 (p=0.6375) |
-0.0649 (p=0.4754) |
0.1543 (p=0.0885) |
0.1587 (p=0.0797) |
| Females | |||||
| AnDarApy-1 | -0.0571 (p=0.6390) |
-0.0641 (p=0.5980) |
0.0739 (p=0.5434) |
0.0847 (p=0.4858) |
0.0520 (p=0. 6689) |
| gSG6-P1 | 0.0216 (p=0.8591) |
-0.1233 (p=0.3092) |
0.0338 (p=0.7815) |
0.0015 (p=0.9899) |
-0.0156 (p=0.8981) |
| Nterm-34kDa | 0.0741 (p=0.5422) |
0.0480 (p=0.6932) |
-0.0153 (p=0.9001) |
0.1431 (p=0.2373) |
0.01078 (p=0.3474) |
| Males | |||||
| AnDarApy-1 |
0.2797 (p=0.0425) |
0.2063 (p=0.1382) |
-0.3052 (p=0.0263) |
0.1138 (p=0.4173) |
0.1681 (p=0.2290) |
| gSG6-P1 |
0.4099 (p=0.0023) |
0.0559 (p=0.6908) |
-0.2038 (p=0.1433) |
0.2074 (p=0.1362) |
0.2408 (p=0.0824) |
| Nterm-34kDa |
0.3904 (p=0.0039) |
0.0304 (p=0.8289) |
-0.1611 (p=0.2493) |
0.1922 (p=0.1680) |
0.2288 (p=0.0994) |
Table 6.
Correlation analysis between IgG antibody responses against each peptide and age. Data is presented in Spearman correlation r.
Table 6.
Correlation analysis between IgG antibody responses against each peptide and age. Data is presented in Spearman correlation r.
| Peptide | Age |
|---|---|
| All | |
| gSG6-P1 |
-0.3533 (p=0.0003) |
| Nterm-34kDa |
-0.4182 (p=0.0000) |
| Correlations by gender | |
| Females | |
| gSG6-P1 |
-0.3575 (p=0.0035) |
| Nterm-34kDa |
-0.4087 (p=0.0007) |
| Males | |
| gSG6-P1 |
-0.3702 (p=0.0370) |
| Nterm-34kDa |
-0.4105 (p=0.0196) |
| Correlations by location | |
| Colombia | |
| gSG6-P1 |
0.2702 (p=0.0481) |
| Nterm-34kDa | 0.0796 (p=0.5675) |
| US | |
| gSG6-P1 |
-0.4772 (p=0.0005) |
| Nterm-34kDa |
-0.4049 (p=0.0039) |
Figure 3.
IgG antibody levels against the mosquito salivary peptides in healthy participants residents of Kansas (U.S.) in summer 2018 compared to levels in the IgG antibody levels found in volunteers followed during the Fall of 2018 (n=25). Significance was measured by the Mann-Whitney test p<0.05 (*=0.0, **=0.00, ***=0.000 and ns= not significant).
Figure 3.
IgG antibody levels against the mosquito salivary peptides in healthy participants residents of Kansas (U.S.) in summer 2018 compared to levels in the IgG antibody levels found in volunteers followed during the Fall of 2018 (n=25). Significance was measured by the Mann-Whitney test p<0.05 (*=0.0, **=0.00, ***=0.000 and ns= not significant).
3. Discussion
Mosquito saliva is composed of a plethora of molecules that are injected into the skin to counteract host responses and facilitate blood uptake. This saliva also induces production of host antibodies that correlate with the intensity of mosquito exposure. Notably, levels of IgG antibodies against whole or specific salivary proteins have been categorized as a reliable tool to measure exposure to mosquito bites and disease transmission intensity[16,38]. In malaria, Plasmodium gametocytes in vertebrate blood are the infectious stages for the mosquito vector. Interestingly, previous studies showed that the carriage of specific parasite stages influenced host attractiveness to mosquitoes, with people that harbored gametocytes in their blood being more attractive to mosquitoes than those presenting parasitemia with only asexual stages[39-42]. Here, we evaluated gametocyte carriage by measuring the expression of Plasmodium vivax Pvs25, while Pvs230 was used to determine gametocyte carriage[43,44]. Upon analysis of the level of antibodies against mosquito saliva, we observed that expression of Pvs230 was significantly negatively correlated with IgG antibody levels of all mosquito antigens tested in this study, including the non-malaria vector Aedes peptide Nterm34kDa. However, this association was not observed with Pvs25, suggesting that expression of Pvs230 may be associated with exposure to mosquito bites from both Anopheles and Aedes mosquitoes. Previous entomological data from this region indicate that Anopheles and Aedes mosquitoes are prevalent in the area[45,46]. However, a limitation of this study is that the gSG6-P1 and the Nterm34kDa peptides we evaluated as markers of mosquito exposure have not been extensively validated for their correlation with mosquito abundance[47-49], and we did not collect specific mosquito data from the sites in our study which could be used to confirm exposure to these mosquito species.
One of the objectives of this study was to determine if antibody levels against mosquito saliva were correlated with blood parameters. In the case of the malaria cohort, we observed that the non-malaria vector Aedes peptide Nterm34kDa was negatively associated with RBC count. This finding is notable as one of the signs of severe malaria is anemia, yet we did not observe correlations between any of the Anopheles salivary antigens and RBC count. The observed negative correlation between IgG anti-Nter34kDa and RBC count suggests that individuals with the lowest RBC counts may have the highest exposure to non-malaria vector Aedes mosquitoes or may produce more antibodies against their saliva. Interestingly, a previous study reported that anemia accelerates blood intake by Ae. aegypti, although it may negatively impact egg production[50]. Also, prior work has demonstrated that DENV acquisition by Ae. aegypti was inversely correlated with iron concentration in human serum[51]. Thus, it is possible that DENV benefits from the Aedes preference to feed on anemic individuals. Indeed, among our DENV-infected participants, we observed a significant positive correlation between RBC and both Nterm34kDa and gSG6-P1 peptides, suggesting increased exposure to mosquito bites while infected. Moreover, a recent study suggested that infection with DENV increased attractiveness to Aedes mosquitoes[52], similar to observations made with Anopheles mosquitoes and Plasmodium gametocyte carriers[39,42].
Mosquito abundance is associated with temperature, humidity, and other environmental factors[53-55]. In temperate and subtropical regions, mosquito abundance drastically changes in the summer vs. fall seasons, with higher mosquito populations in the former[56,57]. Previously, we observed a significant decrease in anti-tick antibodies during the fall months[58]. Interestingly, we found a significant decrease in IgG antibody levels against the Nter34kDa peptide in the fall compared to summer in people living in Kansas. Our results are in agreement with previous studies, which suggest a significant decrease in antibodies against Ae. aegypti mosquitoes after cessation of exposure[13]. However, we did not observe a decrease in antibodies against gSG6P1, although prior reports have indicated that antibodies against the gSG6 protein are short-lived[16], suggesting a potential continued exposure. In Kansas, An. quadrimaculatus and An. pseudopunctipennis are abundantly found in the summer, with An. quadrimaculatus populations receding in late fall[59]. Studies conducted in Northern Argentina indicate that An. pseudopunctipennis presents two peaks of abundance, one in spring and another in fall/autumn[60]. Together, these studies provide a potential explanation for the persistence of antibody levels in samples collected in early fall. Endemicity also appears to affect antibody levels against mosquito antigens in healthy individuals. Specifically, we observed that healthy volunteers living in a non-endemic area for either malaria or DENV (Kansas) presented negative correlations between age and anti-saliva antibodies, while healthy people living in Norte de Santander showed a positive correlation. We found a negative correlation between age and antibodies against Ae. aegypti whole salivary gland extract in healthy individuals living in Norte de Santander in 2015[10] that could be associated with the development of tolerance as observed in other studies[19,38]. However, in our recent study in a population with DENV, we did not observe any associations with age[61], as shown in other studies[62], suggesting that several factors may contribute to these discrepancies. Further and larger studies are needed to establish the relationship between age and response to arthropod salivary antigens.
In addition to previous studies showing sex-associated differences in DENV incidence, geographical area also appears to strongly influence the rates of association and occurrence. For instance, a study including DENV cases from at least six Asian countries showed a higher incidence of dengue fever in males[63], while other studies in Central and South America demonstrated a higher incidence in females[64,65]. Herein, we observed that males and females respond differently to mosquito salivary antigens. These results may have implications for pathogen transmission and clinical disease presentation. A major implication of our study is that characteristics, including age, seasonality, and vector control must be considered when assessing serum levels of IgG against Ae. aegypti salivary proteins as a surrogate of risk of human exposure to mosquito bites and pathogen transmission. Such information is needed in epidemiological studies aimed at control and prevention of mosquito-borne diseases. Furthermore, data on biological sex at the population level could potentially be used to inform calculations of total disease burden in regions where vector-borne diseases have the highest impact.
In conclusion, our data indicate that individuals living in malaria- and dengue-endemic areas are exposed to several mosquito species in a given area. Moreover, exposure to malaria and dengue mosquito vectors may be associated with clinical and immune responses to infection with the reciprocal pathogens transmitted by these specific species, as well as other mosquito species that may not be implicated as vectors of human disease. Our findings suggest that concurrent exposure to multiple disease-carrying mosquito vectors and their salivary proteins with differing immunomodulatory properties could influence the transmission and pathogenesis of malaria, dengue fever, and other vector-borne illnesses. A better understanding of the molecular mechanisms underlying how exposure to multiple and sequential bites from numerous mosquito species influences immune responses and pathogen transmission will advance the development of immune-targeting interventions to reduce disease spread. Moreover, including data on antibody responses against the main mosquito species found in specific areas will provide new knowledge on the interplay of such species, pathogen transmission, and disease severity in humans, which will help to inform the development of more effective vector control and disease prevention efforts.
4. Materials and Methods
4.1. Human Sample Collection and Diagnosis
All protocols involving human subjects were reviewed and approved by Universidad de Pamplona and by the IRB of Kansas State University (IRB#1206). Written informed consent was obtained from all subjects, and blood samples were collected from each subject living in two areas with different endemicity levels for malaria and DENV in the department of Norte de Santander Colombia. The sample size and ages of volunteers included in these studies are described in Table 1. Malaria diagnosis was done by thick blood smear evaluated by at least two experienced microscopists. Gametocyte carriage was performed by qRT-PCR on the Pvs25 and Pvs230 genes of P. vivax parasites following the methods described elsewhere[43,44]. DENV diagnosis was performed using the Rapid Diagnostic Test, Cassette Dengue AG (Xerion) and confirmed by qRT-PCR[66]. We also tested 48 human samples collected from healthy volunteers living in Kansas to measure antibody levels against the peptides in non-endemic areas.
4.2. Salivary Peptides
The previously reported salivary peptides Nterm-34kDa (Ae. aegypti) and gSG6-P1 (An. gambiae) were used to evaluate exposure to mosquito bites[27]. Peptides were synthesized by Genscript (Piscataway, NJ), dissolved in ultrapure water and frozen at -80°C until used as antigens in ELISA assays.
4.3. Human IgG Antibody Detection by ELISA
The level of human IgG antibodies against mosquito salivary proteins was determined by an indirect ELISA following the methods published by Londono-Renteria et al.[67]. Briefly, 96-well ELISA plates (Nunc-MaxiSorp, Nalgene Nunc International, Rochester, NY, USA) were coated with 50 μl/well of Ae. aegypti salivary protein in a final concentration of 2 µg/ml prepared in coating solution (1X PBS). Serum samples were tested in duplicate in a 1/100 dilution. After washes, plates were incubated with horseradish peroxidase-conjugated goat anti-human IgG (1: 1,000) (Abcam, Ab81202) and colorimetric development was obtained using tetra-methyl-benzidine (one-solution micro-well, Gene-Script, Piscataway, NJ, USA). The reaction was terminated with 1 M phosphoric acid and absorbance was measured at 450 nm. Two controls were included on each plate: (1) control blank: two wells with antigen and without sample as a control for nonspecific induction of color for any of the reagents used in the test; and (2) positive control: 1 control per plate to test plate variation and normalize OD (optical density) values. IgG antibody levels are reported as ΔOD = Average patient OD value (duplicate) less the Blank OD.
4.4. Data Analysis
The median OD value was selected for all IgG levels against salivary antigens to determine high (above the median) or low (below the median) antibody levels. Differences between two independent groups were tested using the nonparametric Mann-Whitney U test. Correlation analysis between age and antigens was performed using the Spearman correlation test. All differences were considered significant at p <0.05. All statistical tests were performed using Prism version 10 (Graph Pad Software Inc., La Jolla, CA).
5. Conclusions
Antibodies against salivary proteins are reliable markers for exposure intensity to bites from insect vectors. The data we have presented here support the use of IgG antibodies against salivary proteins as useful biomarkers of the intensity of exposure to mosquito bites and further indicate that these measurements can provide critical knowledge on how exposure to mosquito salivary proteins influences transmission of vector-borne diseases. Moreover, including the potential effects of sequential exposure to saliva from different mosquito species will promote a better understanding of the pathogenesis of diseases borne by the vectors themselves, as well as in the context of simultaneous exposure to non-vector mosquitos.
Author Contributions
conceptualization, BLR, JAM; methodology, BLR; sample collection, JCC, CAP, MUG, LYG-S and LAJ-V; sample testing, MMH and OO; formal analysis, MMH, OO and BLR; investigation, BLR, JAM; resources, BLR and JAM; writing—original draft preparation, BLR, JAM, JGS and BW; draft revisions BLR, JAM, JGS and BW, project administration, BLR and JAM; funding acquisition, BLR and JAM.
Funding
Tulane Center of Excellence in Emergent Infectious Diseases (CEEIRD) and A Studio In The Woods Program.
Acknowledgments
The authors thank the residents of Los Patios, Cucuta, Tarra and Tibu (Colombia); and Manhattan – Kansas (USA) for their participation in this study.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Rosenberg, R.; Lindsey, N.P.; Fischer, M.; Gregory, C.J.; Hinckley, A.F.; Mead, P.S.; Paz-Bailey, G.; Waterman, S.H.; Drexler, N.A.; Kersh, G.J.; et al. Vital Signs: Trends in Reported Vectorborne Disease Cases - United States and Territories, 2004-2016. MMWR Morb Mortal Wkly Rep 2018, 67, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; Adams, L.E.; Durbin, A.P.; Muñoz-Jordán, J.L.; Poehling, K.A.; Sánchez-González, L.M.; Volkman, H.R.; Paz-Bailey, G. Dengue: A Growing Problem With New Interventions. Pediatrics 2022, 149. [Google Scholar] [CrossRef] [PubMed]
- Brathwaite Dick, O.; San Martín, J.L.; Montoya, R.H.; del Diego, J.; Zambrano, B.; Dayan, G.H. The history of dengue outbreaks in the Americas. Am J Trop Med Hyg 2012, 87, 584–593. [Google Scholar] [CrossRef]
- Hernandez-Romieu, A.C.; Adams, L.E.; Paz-Bailey, G. Opportunities for Improved Dengue Control in the US Territories. Jama 2023, 330, 19–20. [Google Scholar] [CrossRef] [PubMed]
- Bagcchi, S. Locally acquired malaria cases in the USA. Lancet Infect Dis 2023, 23, e401. [Google Scholar] [CrossRef] [PubMed]
- Agudelo Higuita, N.I.; Franco-Paredes, C.; Henao-Martínez, A.F.; Mendez Rojas, B.; Suarez, J.A.; Naranjo, L.; Alger, J. Migrants in transit across Central America and the potential spread of chloroquine resistant malaria-a call for action. Lancet Reg Health Am 2023, 22, 100505. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Abreu, I.V.; Guimaraes-Costa, A.B.; Valenzuela, J.G. Impact of insect salivary proteins in blood feeding, host immunity, disease, and in the development of biomarkers for vector exposure. Curr Opin Insect Sci 2015, 10, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Arca, B.; Ribeiro, J.M. Saliva of hematophagous insects: a multifaceted toolkit. Curr Opin Insect Sci 2018, 29, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Spencer Clinton, J.L.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl Trop Dis 2018, 12, e0006439. [Google Scholar] [CrossRef]
- Londoño-Rentería, B.; Cárdenas, J.C.; Giovanni, J.E.; Cárdenas, L.; Villamizar, P.; Rolón, J.; Chisenhall, D.M.; Christofferson, R.C.; Carvajal, D.J.; Pérez, O.G.; et al. Aedes aegypti anti-salivary gland antibody concentration and dengue virus exposure history in healthy individuals living in an endemic area in Colombia. Biomedica 2015, 35, 572–581. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Shakeri, H.; Rozo-Lopez, P.; Conway, M.J.; Duggan, N.; Jaberi-Douraki, M.; Colpitts, T.M. Serosurvey of Human Antibodies Recognizing Aedes aegypti D7 Salivary Proteins in Colombia. Front Public Health 2018, 6, 111. [Google Scholar] [CrossRef] [PubMed]
- Londono-Renteria, B.; Drame, P.M.; Weitzel, T.; Rosas, R.; Gripping, C.; Cardenas, J.C.; Alvares, M.; Wesson, D.M.; Poinsignon, A.; Remoue, F.; et al. gambiae gSG6-P1 evaluation as a proxy for human-vector contact in the Americas: a pilot study. Parasit Vectors 2015, 8, 533. [Google Scholar] [CrossRef] [PubMed]
- Londono-Renteria, B.; Cardenas, J.C.; Cardenas, L.D.; Christofferson, R.C.; Chisenhall, D.M.; Wesson, D.M.; McCracken, M.K.; Carvajal, D.; Mores, C.N. Use of anti-Aedes aegypti salivary extract antibody concentration to correlate risk of vector exposure and dengue transmission risk in Colombia. PLoS One 2013, 8, e81211. [Google Scholar] [CrossRef] [PubMed]
- Traore, D.F.; Sagna, A.B.; Adja, A.M.; Zoh, D.D.; Adou, K.A.; Lingue, K.N.; Coulibaly, I.; Tchiekoi, N.B.; Assi, S.B.; Poinsignon, A.; et al. Exploring the heterogeneity of human exposure to malaria vectors in an urban setting, Bouake, Cote d'Ivoire, using an immuno-epidemiological biomarker. Malar J 2019, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Ndille, E.E.; Dubot-Pérès, A.; Doucoure, S.; Mouchet, F.; Cornelie, S.; Sidavong, B.; Fournet, F.; Remoue, F. Human IgG antibody response to Aedes aegypti Nterm-34 kDa salivary peptide as an indicator to identify areas at high risk for dengue transmission: a retrospective study in urban settings of Vientiane city, Lao PDR. Trop Med Int Health 2014, 19, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, C.; Lombardo, F.; Ronca, R.; Mangano, V.; Sirima, S.B.; Nèbiè, I.; Fiorentino, G.; Modiano, D.; Arcà, B. Differential antibody response to the Anopheles gambiae gSG6 and cE5 salivary proteins in individuals naturally exposed to bites of malaria vectors. Parasit Vectors 2014, 7, 549. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.J.; Londono-Renteria, B.; Troupin, A.; Watson, A.M.; Klimstra, W.B.; Fikrig, E.; Colpitts, T.M. Aedes aegypti D7 Saliva Protein Inhibits Dengue Virus Infection. PLoS Negl Trop Dis 2016, 10, e0004941. [Google Scholar] [CrossRef]
- Olajiga, O.M.; Marin-Lopez, A.; Cardenas, J.C.; Gutierrez-Silva, L.Y.; Gonzalez-Pabon, M.U.; Maldonado-Ruiz, L.P.; Worges, M.; Fikrig, E.; Park, Y.; Londono-Renteria, B. Aedes aegypti anti-salivary proteins IgG levels in a cohort of DENV-like symptoms subjects from a dengue-endemic region in Colombia. Frontiers in Epidemiology 2022, 2. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Ho, M.K.; Li, C.; Simons, F.E. Evidence for natural desensitization to mosquito salivary allergens: mosquito saliva specific IgE and IgG levels in children. Ann Allergy Asthma Immunol 2004, 93, 553–556. [Google Scholar] [CrossRef]
- Guerra-Silveira, F.; Abad-Franch, F. Sex bias in infectious disease epidemiology: patterns and processes. PLoS One 2013, 8, e62390. [Google Scholar] [CrossRef]
- Pathak, S.; Rege, M.; Gogtay, N.J.; Aigal, U.; Sharma, S.K.; Valecha, N.; Bhanot, G.; Kshirsagar, N.A.; Sharma, S. Age-dependent sex bias in clinical malarial disease in hypoendemic regions. PLoS One 2012, 7, e35592. [Google Scholar] [CrossRef]
- Bardach, A.E.; Garcia-Perdomo, H.A.; Alcaraz, A.; Lopez, E.T.; Gandara, R.A.R.; Ruvinsky, S.; Ciapponi, A. Interventions for the control of Aedes aegypti in Latin America and the Caribbean: Systematic Review and Meta-Analysis. Trop Med Int Health, 1111. [Google Scholar] [CrossRef]
- Dhiman, S. Are malaria elimination efforts on right track? An analysis of gains achieved and challenges ahead. Infect Dis Poverty 2019, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhang, Y.; Li, H.; Zhu, J.; Song, W.; Chen, K.; Zhang, Y.; Lou, Y. Vaccine development for mosquito-borne viral diseases. Front Immunol 2023, 14, 1161149. [Google Scholar] [CrossRef]
- Qian, X.; Qi, Z. Mosquito-Borne Flaviviruses and Current Therapeutic Advances. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Elanga Ndille, E.; Doucoure, S.; Poinsignon, A.; Mouchet, F.; Cornelie, S.; D'Ortenzio, E.; DeHecq, J.S.; Remoue, F. Human IgG Antibody Response to Aedes Nterm-34kDa Salivary Peptide, an Epidemiological Tool to Assess Vector Control in Chikungunya and Dengue Transmission Area. PLoS Negl Trop Dis 2016, 10, e0005109. [Google Scholar] [CrossRef]
- Elanga Ndille, E.; Doucoure, S.; Damien, G.; Mouchet, F.; Drame, P.M.; Cornelie, S.; Noukpo, H.; Yamadjako, S.; Djenontin, A.; Moiroux, N.; et al. First attempt to validate human IgG antibody response to Nterm-34kDa salivary peptide as biomarker for evaluating exposure to Aedes aegypti bites. PLoS Negl Trop Dis 2012, 6, e1905. [Google Scholar] [CrossRef]
- Poinsignon, A.; Cornelie, S.; Mestres-Simon, M.; Lanfrancotti, A.; Rossignol, M.; Boulanger, D.; Cisse, B.; Sokhna, C.; Arcà, B.; Simondon, F.; et al. Novel peptide marker corresponding to salivary protein gSG6 potentially identifies exposure to Anopheles bites. PLoS One 2008, 3, e2472. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, F.; Ronca, R.; Rizzo, C.; Mestres-Simòn, M.; Lanfrancotti, A.; Currà, C.; Fiorentino, G.; Bourgouin, C.; Ribeiro, J.M.; Petrarca, V.; et al. The Anopheles gambiae salivary protein gSG6: an anopheline-specific protein with a blood-feeding role. Insect Biochem Mol Biol 2009, 39, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Drame, P.M.; Poinsignon, A.; Besnard, P.; Cornelie, S.; Le Mire, J.; Toto, J.C.; Foumane, V.; Dos-Santos, M.A.; Sembène, M.; Fortes, F.; et al. Human antibody responses to the Anopheles salivary gSG6-P1 peptide: a novel tool for evaluating the efficacy of ITNs in malaria vector control. PLoS One 2010, 5, e15596. [Google Scholar] [CrossRef]
- Sagna, A.B.; Yobo, M.C.; Elanga Ndille, E.; Remoue, F. New Immuno-Epidemiological Biomarker of Human Exposure to Aedes Vector Bites: From Concept to Applications. Trop Med Infect Dis 2018, 3. [Google Scholar] [CrossRef]
- Bellone, R.; Failloux, A.B. The Role of Temperature in Shaping Mosquito-Borne Viruses Transmission. Front Microbiol 2020, 11, 584846. [Google Scholar] [CrossRef] [PubMed]
- Maharaj, P.D.; Widen, S.G.; Huang, J.; Wood, T.G.; Thangamani, S. Discovery of mosquito saliva microRNAs during CHIKV infection. PLoS Negl Trop Dis 2015, 9, e0003386. [Google Scholar] [CrossRef] [PubMed]
- Visser, I.; Koenraadt, C.J.M.; Koopmans, M.P.G.; Rockx, B. The significance of mosquito saliva in arbovirus transmission and pathogenesis in the vertebrate host. One Health 2023, 16, 100506. [Google Scholar] [CrossRef] [PubMed]
- Sulesco, T.M.; Toderas, L.G.; Uspenskaia, I.G.; Toderas, I.K. Larval Habitats Diversity and Distribution of the Mosquito (Diptera: Culicidae) Species in the Republic of Moldova. J Med Entomol 2015, 52, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Tazeen, A.; Abdullah, M.; Hisamuddin, M.; Ali, S.; Naqvi, I.H.; Verma, H.N.; Ahmed, A.; Parveen, S. Concurrent Infection with Plasmodium vivax and the Dengue and Chikungunya Viruses in a Paediatric Patient from New Delhi, India in 2016. Intervirology 2017, 60, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Eder, M.; Cortes, F.; Teixeira de Siqueira Filha, N.; Araujo de Franca, G.V.; Degroote, S.; Braga, C.; Ridde, V.; Turchi Martelli, C.M. Scoping review on vector-borne diseases in urban areas: transmission dynamics, vectorial capacity and co-infection. Infect Dis Poverty 2018, 7, 90. [Google Scholar] [CrossRef] [PubMed]
- Doucoure, S.; Mouchet, F.; Cournil, A.; Le Goff, G.; Cornelie, S.; Roca, Y.; Giraldez, M.G.; Simon, Z.B.; Loayza, R.; Misse, D.; et al. Human antibody response to Aedes aegypti saliva in an urban population in Bolivia: a new biomarker of exposure to Dengue vector bites. Am J Trop Med Hyg 2012, 87, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Debebe, Y.; Hill, S.R.; Birgersson, G.; Tekie, H.; Ignell, R. Plasmodium falciparum gametocyte-induced volatiles enhance attraction of Anopheles mosquitoes in the field. Malar J 2020, 19, 327. [Google Scholar] [CrossRef]
- Lacroix, R.; Mukabana, W.R.; Gouagna, L.C.; Koella, J.C. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol 2005, 3, e298. [Google Scholar] [CrossRef]
- Robinson, A.; Busula, A.O.; Voets, M.A.; Beshir, K.B.; Caulfield, J.C.; Powers, S.J.; Verhulst, N.O.; Winskill, P.; Muwanguzi, J.; Birkett, M.A.; et al. Plasmodium-associated changes in human odor attract mosquitoes. Proc Natl Acad Sci U S A 2018, 115, E4209–e4218. [Google Scholar] [CrossRef]
- Busula, A.O.; Bousema, T.; Mweresa, C.K.; Masiga, D.; Logan, J.G.; Sauerwein, R.W.; Verhulst, N.O.; Takken, W.; de Boer, J.G. Gametocytemia and Attractiveness of Plasmodium falciparum-Infected Kenyan Children to Anopheles gambiae Mosquitoes. J Infect Dis 2017, 216, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Wampfler, R.; Mwingira, F.; Javati, S.; Robinson, L.; Betuela, I.; Siba, P.; Beck, H.P.; Mueller, I.; Felger, I. Strategies for detection of Plasmodium species gametocytes. PLoS One 2013, 8, e76316. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, C.; Roobsoong, W.; Kangwanrangsan, N.; Bardelli, M.; Rawlinson, T.A.; Dambrauskas, N.; Trakhimets, O.; Parthiban, C.; Goswami, D.; Reynolds, L.M.; et al. A Humanized Mouse Model for Plasmodium vivax to Test Interventions that Block Liver Stage to Blood Stage Transition and Blood Stage Infection. iScience 2020, 23, 101381. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, M.A.; Cardenas, R.; Yañez, J.; Petzold, M.; Kroeger, A. Risk of dengue, Zika, and chikungunya transmission in the metropolitan area of Cucuta, Colombia: cross-sectional analysis, baseline for a cluster-randomised controlled trial of a novel vector tool for water containers. BMC Public Health 2023, 23, 1000. [Google Scholar] [CrossRef] [PubMed]
- Montoya-Lerma, J.; Solarte, Y.A.; Giraldo-Calderón, G.I.; Quiñones, M.L.; Ruiz-López, F.; Wilkerson, R.C.; González, R. Malaria vector species in Colombia: a review. Mem Inst Oswaldo Cruz 2011, 106 Suppl 1, 223–238. [Google Scholar] [CrossRef]
- Poinsignon, A.; Cornelie, S.; Ba, F.; Boulanger, D.; Sow, C.; Rossignol, M.; Sokhna, C.; Cisse, B.; Simondon, F.; Remoue, F. Human IgG response to a salivary peptide, gSG6-P1, as a new immuno-epidemiological tool for evaluating low-level exposure to Anopheles bites. Malaria Journal 2009, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Sagna, A.B.; Sarr, J.B.; Gaayeb, L.; Drame, P.M.; Ndiath, M.O.; Senghor, S.; Sow, C.S.; Poinsignon, A.; Seck, M.; Hermann, E.; et al. gSG6-P1 salivary biomarker discriminates micro-geographical heterogeneity of human exposure to Anopheles bites in low and seasonal malaria areas. Parasit Vectors 2013, 6, 68. [Google Scholar] [CrossRef] [PubMed]
- Ndo, C.; Elanga-Ndille, E.; Cheteug, G.; Metitsi, R.D.; Wanji, S.; Moukoko, C.E.E. IgG antibody responses to Anopheles gambiae gSG6-P1 salivary peptide are induced in human populations exposed to secondary malaria vectors in forest areas in Cameroon. PLoS One 2022, 17, e0276991. [Google Scholar] [CrossRef]
- Shieh, J.N.; Rossingnol, P.A. Opposite influences of host anaemia on blood feeding rate and fecundity of mosquitoes. Parasitology 1992, 105 ( Pt 2) Pt 2, 159–163. [Google Scholar] [CrossRef]
- Zhu, Y.; Tong, L.; Nie, K.; Wiwatanaratanabutr, I.; Sun, P.; Li, Q.; Yu, X.; Wu, P.; Wu, T.; Yu, C.; et al. Host serum iron modulates dengue virus acquisition by mosquitoes. Nat Microbiol 2019, 4, 2405–2415. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, Y.; Liu, Z.; Peng, Y.; Peng, W.; Tong, L.; Wang, J.; Liu, Q.; Wang, P.; Cheng, G. A volatile from the skin microbiota of flavivirus-infected hosts promotes mosquito attractiveness. Cell 2022, 185, 2510–2522. [Google Scholar] [CrossRef] [PubMed]
- Minakawa, N.; Sonye, G.; Mogi, M.; Githeko, A.; Yan, G. The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J Med Entomol 2002, 39, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Asgarian, T.S.; Moosa-Kazemi, S.H.; Sedaghat, M.M. Impact of meteorological parameters on mosquito population abundance and distribution in a former malaria endemic area, central Iran. Heliyon 2021, 7, e08477. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.J.; Kim, H.C.; Klein, T.A.; Chong, S.T.; Sim, K.; Chung, Y.; Cheong, H.K. Comparison of climatic factors on mosquito abundance at US Army Garrison Humphreys, Republic of Korea. PLoS One 2020, 15, e0240363. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Cayan, D.; Tyree, M.; Barker, C.M.; Eldridge, B.; Dettinger, M. Impact of climate variation on mosquito abundance in California. J Vector Ecol 2008, 33, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.W.; Ionides, E.L.; Knepper, R.G.; Stanuszek, W.W.; Walker, E.D.; Wilson, M.L. Cross-correlation map analyses show weather variation influences on mosquito abundance patterns in Saginaw County, Michigan, 1989-2005. J Med Entomol 2012, 49, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Ruiz, L.P.; Montenegro-Cadena, L.; Blattner, B.; Menghwar, S.; Zurek, L.; Londono-Renteria, B. Differential Tick Salivary Protein Profiles and Human Immune Responses to Lone Star Ticks (Amblyomma americanum) From the Wild vs. a Laboratory Colony. Front Immunol 2019, 10, 1996. [Google Scholar] [CrossRef] [PubMed]
- Robertson, L.C.; Prior, S.; Apperson, C.S.; Irby, W.S. Bionomics of Anopheles quadrimaculatus and Culex erraticus (Diptera: Culicidae) in the Falls Lake basin, North Carolina: seasonal changes in abundance and gonotrophic status, and host-feeding patterns. J Med Entomol 1993, 30, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Dantur Juri, M.J.; Claps, G.L.; Santana, M.; Zaidenberg, M.; Almirón, W.R. Abundance patterns of Anopheles pseudopunctipennis and Anopheles argyritarsis in northwestern Argentina. Acta Trop 2010, 115, 234–241. [Google Scholar] [CrossRef]
- Olajiga, O.M. , Cardenas, J. C., Paulina, L., Worges, M., Fikrig, E., & Park, Y. Aedes aegypti anti-salivary proteins IgG levels in a cohort of DENV-like symptoms subjects from a dengue-endemic region in Colombia. Frontiers in Immunology.
- Peng, Z.; Rasic, N.; Liu, Y.; Simons, F.E. Mosquito saliva-specific IgE and IgG antibodies in 1059 blood donors. J Allergy Clin Immunol 2002, 110, 816–817. [Google Scholar] [CrossRef]
- Anker, M.; Arima, Y. Male-female differences in the number of reported incident dengue fever cases in six Asian countries. Western Pac Surveill Response J 2011, 2, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.E.; Eliason, D.A.; Moore, M.; Sather, G.E.; Schonberger, L.B.; Cabrera-Coello, L.; Fernandez de Castro, J. Epidemiologic investigations of dengue infection in Mexico, 1980. Am J Epidemiol 1983, 117, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Trravassos da Rosa, A.P.; Vasconcelos, P.F.; Travassos Da Rosa, E.S.; Rodrigues, S.G.; Mondet, B.; Cruz, A.C.; Sousa, M.R.; Travassos Da Rosa, J.F. Dengue epidemic in Belém, Pará, Brazil, 1996-97. Emerg Infect Dis 2000, 6, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, J.C.; Giraldo-Parra, S.Y.; Gonzalez, M.U.; Gutierrez-Silva, L.Y.; Jaimes-Villamizar, L.; Roa-Parra, A.L.; Carvajal, D.J.; Valdivia, H.O.; Sanchez, J.F.; Colpitts, T.M.; et al. Laboratory Findings in Patients with Probable Dengue Diagnosis from an Endemic Area in Colombia in 2018. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Londono-Renteria, B.L.; Shakeri, H.; Rozo-Lopez, P.; Conway, M.J.; Duggan, N.; Jaberi-Douraki, M.; Colpitts, T.M. Serosurvey of Human Antibodies Recognizing. Front Public Health 2018, 6, 111. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



