
Submitted:
26 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
The cryopreservation of gametes and embryos is an important element of biodiversity conservation. One species in need of conservation is the honey bee Apis mellifera L. Changing environmental factors, especially the anthropogenic factor, have led to a reduction in the numbers of this insect species. In this study, we provide an example of the creation of a biobank of honey bee drone sperm. For sperm cryopreservation, drones of the most common subspecies of honey bees common in Russia were selected. These were the dark forest bee, Apis mellifera mellifera from the Republic of Bashkortostan, with three subspecies (A. m. carnica, A. m. carpatica, and A. m. caucasica) from the southern regions of Russia, as well as two breeding stocks, the Far Eastern bee and Prioksky bee. For subspecies identification, morphometric and genetic methods were used. The subspecies of the studied samples were confirmed by the analysis of the tRNAleu-COII locus of mitochondrial DNA and nine microsatellite markers of nuclear DNA. It was shown that bees of the Prioksky breeding stock belong to the subspecies A. m. caucasica based on phylogenetic analysis, and the Far Eastern breeding stock is a stable hybrid, descending on the maternal line from the evolutionary lineage C or O. The results of the morphometric analysis are consistent with the results of the genetic analysis. For the cryopreservation of sperm, we used a honey diluent. As a result, the viability of frozen–thawed sperm decreased by 20.3% compared to fresh sperm, and overall motility decreased by 25-fold. The measurement of the sperm concentration in the spermatheca of artificially inseminated queens showed that it varied from 0.22 to 4.4 million/μL. Therefore, the use of honey in sperm cryopreservation has great potential.
Keywords:
Apis mellifera
; honey bee subspecies
; cryopreservation
; breeding stock
; honey bee selection
; tRNAleu-COII (COI-COII)
; microsatellite markers
; morphometry
1. Introduction
The cryopreservation of gametes and embryos is an important tool for preserving the biodiversity of both wild and farmed animals and plants [1-4]. Research related to the possibility of the long-term preservation of honey bee sperm in liquid nitrogen was actively carried out around the world at the end of the last century. One of the founders of the cryopreservation of drone sperm [5], who used 25% dimethyl sulfoxide (DMSO) as a cryoprotectant, obtained 8% bee brood from queens artificially inseminated with frozen–thawed sperm. The follow-up results on the artificial insemination of queens with frozen–thawed sperm were published several decades later. Hopkins et al. (2012) [6] pioneered the use of egg yolk as a cryoprotectant for drone sperm. According to their data, out of five inseminated queens, only two produced offspring with more than 50% worker bees. The remaining queens had mixed broods or only drone broods. In order to reduce the toxic effects of DMSO on the quality characteristics of sperm, researchers used a citrate–HEPES buffer containing trehalose [7], soy lecithin [8], the seminal plasma of sheep, and honey bee drone sperm [9], with royal jelly as an extender [10]. The first successful experiments on the cryopreservation of drone sperm in the hemolymph of a honey bee were carried out in the USSR [11]. And, only at the beginning of this century was a patent issued (No. 2173045 dated 10 September 2001; http://allpatents.ru/patent/2173045.html?ysclid=lmh7tuqzuy819222269, accessed on 1 September 2023) for the technology of the cryopreservation of drone sperm with the production of fertile queens inseminated with frozen–thawed sperm [12]. The cryopreservation of drone sperm was carried out on the basis of the C46 nutrient medium [12]. An assessment was made of the egg production of queen bees inseminated with frozen–thawed sperm after 25 years of storage in liquid nitrogen [13]. It has been shown that sperm viability is maintained at a fairly high level (93%) and does not depend on the duration of cryopreservation. However, the fertilizing ability of these doses of sperm was low; the amount of capped worker brood from queens inseminated with frozen–thawed sperm was less than 50%. But research results have demonstrated the possibility of preserving drone sperm in liquid nitrogen for 25 years [13]. During long-term storage in liquid nitrogen, the sperm is exposed to a number of factors that cause structural and functional changes in cells. Damage to the acrosome and changes in chromatin condensation, induced by the processes of freezing and thawing of the gamete, are clearly reflected in the morphology of sperm and its morphometric parameters [14]. The morphological parameters of sperm correlate more closely with the rate of fertilization than sperm concentration and motility [15,16]. Today, specialists from the Federal Beekeeping Research Centre (Russia, Ryazan region, Rybnoe) are developing a honey extender for the cryopreservation of drone sperm [17].
During the natural spread of the honey bee Apis mellifera L., it formed various subspecies and ecotypes adapted to specific climatic zones [18,19]. In addition to naturally occurring subspecies, work is being done to develop various breeding stocks and lines that differ in terms of their economically useful traits [20-23]. Due to human economic activity, the boundaries between subspecies began to blur, and some subspecies were absorbed by others [24,25]. To differentiate subspecies, morphometric and genetic methods are used. “Classical morphometry” includes the analysis of 36 morphometric features [18,26]. But, usually, no more than 10 features are used [27]. One of the most used genetic methods is the analysis of polymorphism of the mitochondrial locus tRNAleu-COII (or COI-COII) [28,29]. Using the analysis of this locus, it is possible to differentiate honey bees from the evolutionary lineages A, M, C, O, and Y [18,19]. Allelic variants P(Q)1-n are markers of the origin of bees from A. m. mellifera and A. m. iberiensis (lineage M), with allelic variant Q from subspecies from the evolutionary lineages C (A. m. carnica, A. m. ligustica) and O (A. m. caucasica, A. m. anatoliaca, A. m. remipes, A. m. macedonica) on the maternal line. The letters P and Q indicate repeats located between the tRNAleu and COII genes. Subspecies from the evolutionary lineages C and O lack the P repeat [29]. This mitochondrial marker allows us to establish the origin of bees only on the maternal line. To assess the drone background, microsatellite [30] and SNP [31-33] markers are used.
Consequently, beekeeping has the important role of preserving naturally occurring gene pools and gene pools obtained through selection for economically useful traits. And one of the tools for preserving the genetic diversity of honey bees is the cryopreservation of genetic material.
The aim of this study is to create a biobank of the sperm of Apis mellifera L. drones with a known origin on the basis of the Federal Beekeeping Research Centre (Rybnoye, Ryazan region, Russia, https://beecentr.ru/, accessed on 1 September 2023). To assess the origin of bees, morphometric (length of the proboscis, length and width of the forewing and third tergite, tarsal index, cubital index, length of the third sternite, length and width of the wax mirror, distance between wax mirrors) and genetic (analysis of the polymorphism of the microsatellite loci of nuclear DNA and the intergenic locus tRNAleu-COII of mitochondrial DNA) methods were used.
2. Materials and Methods
2.1. Sampling
Bees (drones and worker bees) were selected from 76 colonies (Table 1) belonging to four naturally occurring honey bee subspecies (A. m. caucasica, A. m. carnica, A. m. carpatica, and A. m. mellifera) and two breeding stocks (Far Eastern and Prioksky bees). Samples of the subspecies A. m. mellifera were selected from two districts of the Republic of Bashkortostan: from the Iglinsky district, in an apiary specializing in breeding A. m. mellifera, and from the Burzyansky district, an ancient reserve of the Burzyan population of A. m. mellifera [34]. The gray mountain Caucasian bee was collected at the Krasnopolyansk experimental beekeeping station in the Krasnodar Krai [35]. Samples of A. m. carnica and A. m. carpatica were selected in the Republic of Adygea. The Far Eastern bee was selected in the Primorsky Krai of the Far East, and Prioksky bees in the Ryazan region were obtained from the apiary of the Federal Beekeeping Research Centre.
2.2. Preparation of Honey Bee Drone Sperm
Sperm was collected from sexually mature drones at the age of 20–30 days by artificially stimulating endophallus eversion using SCHLEY-System model 1.04 equipment (A&G Wachholz, Espelkamp, Germany). One hundred microliters of sperm was collected from 110–125 drones from each bee colony. Freshly collected sperm samples were transported to the deposit site in glass capillaries (L = 90 ± 1.0 mm, d = 1.8 ± 0.2 mm) with a volume of 50 μL without the use of antibacterial contamination agents in a foam container with refrigerants within a temperature range of 2–8 °C.
Sperm quality was assessed by motility and membrane integrity by fluorescence microscopy using SYBR-14 and PI fluorochromes [36]. With the use of equipment for the instrumental insemination of queen bees, sperm with a volume of 50 μL was subjected to short-term storage at a temperature of 3 °C. After 2–3 months of storage in the refrigerator at 3 °C, cryopreservation of the prepared sperm samples was performed. The extender included the following components: 10% honey (50 mL), lactose (10 mg), sucrose (10 mg), egg yolk (2.5 mL), and DMSO (5 mL) (10% of the volume of the honey solution). To prepare a 10% solution, honey was preheated in a water bath at 40–45 °C for 30 minutes. The concentration of hydrogen ions in the finished extender was adjusted with 6 M NaOH to a pH value of 8–9.
Next, to prepare one sample, 80 µL of freshly prepared extender and 10 µL of cooled sperm were added to a 1.5-mL Nunc cryovial (for 1 part sperm, 8 parts extender). All components were mixed until homogeneous and placed in a refrigerator for 1 hour at 3 °C for equilibration.
The freezing of samples was carried out using a Bio Freeze BV-65 program freezer (Consarctic, Westerngrund, Germany). The protocol for freezing drone sperm at a rate of 3 °C/min was as follows:
- -
- start at 3 °C;
- -
- from 3 °C to –5 °C at a speed of 3 °C/min;
- -
- hold at –5 °C for 1 min;
- -
- from –5 to –12 °C at a speed of 1 °C/min;
- -
- hold at –12 °С for 9 min;
- -
- from –12 °С to –50 °С at a speed of 3 °C/min;
- -
- after –50 °C, drop the free temperature to –196 °С.
2.3. Instrumental Insemination of Queen Bees
The instrumental insemination of queen bees was carried out using SCHLEY-System model 1.04 equipment (A&G Wachholz). A single insemination was used with a volume of injected sperm of 10–12 μL. For insemination, virgin queens aged 7–8 days were used. The assessment of the reproductive values of artificially inseminated (AI) queens was replaced by an assessment of their physiological values—the concentration of sperm in the seminal receptacle and the presence of sperm residues in the paired oviducts [37].
The queens were dissected under an MBS-10 light microscope (Lytkarino Optical Glass Plant, Lytkarino, Russia). The presence or absence of sperm residues in the paired oviducts of the uterus was visually recorded. The seminal receptacle was freed from tissue and placed in a sterile 1.5 mL Eppendorf tube containing 250 μL of 10% honey extender. Then, the seminal receptacle was pierced with a needle, releasing the contents into the extender. After careful pipetting, a drop of the suspension was taken and Goryaev’s counting chamber was filled. Sperm counting and the determination of the sperm concentration were carried out according to a previously published method [13], taking into account that the volume of the seminal receptacle was 1 μL [38].
2.4. Morphometric Analysis
With the use of Altami Studio software(http://altamisoft.ru/, accessed on 1 September 2023), the following characteristics of bees were measured (Figure S1): proboscis length (Lx), fore wing length (FL) and fore wing width (FW), cubital index (CI, %), length of third tergite (Lt3) and width of third tergite (Wt3), length of third sternite (Ls3), length (Lwm) and width (Wwm) of the wax mirror, distance between wax mirrors (Lwmd), and the tarsal index (TI). Thirty worker bees from each colony were used.
2.5. Genetic Analysis
DNA was isolated from the thorax muscles of worker bees using the DNA-EXTRAN-2 kit (Syntol, LLC, Moscow, Russia). The quality and quantity of total DNA were analysed on an Implen N60 spectrophotometer (Implen GmbH, Munich, Germany).
To establish the maternal origin of bee colonies, analysis of the mtDNA intergenic locus tRNAleu-COII was performed [28] using primers (5’-TCTATACCACGACGTTATTC-3’) and (5’-GATCAATATCATTGATGACC-3’). Subspecies from the evolutionary lineage M have allelic variants P(Q)1-n. Subspecies from the evolutionary lineages C (A. m. carnica, A. m. carpatica) and O (A. m. caucasica) have allelic variant Q.
To establish the drone background, an analysis of the polymorphism of nine microsatellite loci of nuclear DNA (Ap243, 4a110, A24, A8, A43, A113, A88, Ap049, A28) was performed [30]. Samples of A. m. mellifera from the Burzyansky district and the Perm Krai (N = 136) were used as a reference group for the evolutionary lineage M. Samples from the Republic of Adygea, Krasnodar Krai, and Uzbekistan (N = 120) were used as representatives of the C and O evolutionary lineages.
PCRs were performed in a final volume of 20 μL:15 μL sterile deionized water, 2 μL of 10× PCR Buffer, 0.4 μL dNTP, 0.6 μL each primer (10 pmol/μL), 0.3 μL Taq DNA polymerase, and 2 μL DNA template. All PCR amplifications were carried out on a Bio-Rad T100 thermocycler (Bio-Rad, Hercules, CA, USA) with the following conditions: initial denaturation at 94 °C for 5 min, followed by 30 cycles of denaturation at 94 °C for 30 s, annealing at 50/55 °C (tRNAleu-COII/microsatellites) for 30 s, and elongation at 72 °C for 1 min with a final elongation at 72 °C for 10 min. All PCR products were examined on 8% polyacrylamide gels stained with ethidium bromide and observed under an ultraviolet transilluminator Gel Doc™ XR+ (BioRad, Hercules, CA, USA).
To determine the genetic structure of samples, the Structure 2.3.4 program was used with a given number of clusters from 1 to 10. The number of intended groups (K) was calculated in Structure Harvester. The analysis was performed using the Admixture model with Burn-in Period and MCMC equal to 10,000 and 100,000 repetitions, respectively. The results of the analysis were processed in CLUMPP 1.1.2 using the FullSearch algorithm. Genetic differentiation between populations was computed using unbiased estimates of FST values with GENEPOP.
3. Results
3.1. Assessment of Morphometric Parameters
The results of the morphometric analysis of the bees are presented in Table 2 and Table 3. It was established that, according to the length of tergite 3 and the length of the proboscis, all colonies of all subspecies and stocks corresponded to standard values. According to the cubital index, colonies of Prioksky bees corresponded not to the standard values of the breeding stock but to the subspecies A. m. caucasica. The cubital index for samples of A. m. mellifera was also below the standard value [39,40*]. The cubital index of the Burzyan sample (AmmB), despite the fact that it was lower than standard values, corresponded to the CI for the Burzyan population of A. m. mellifera [41].
There are no standard values for other characters. We calculated the mean, minimum, and maximum values for our seven samples (Table 3). The highest value of the tarsal index (TI) was found in the gray mountain Caucasian bee and in the Prioksky bees. The samples also differed in the length of the wax mirror (Lwm)—the highest values were found in samples of A. m. mellifera (AmmB and AmmI). For samples Pr, Amcau, Amcarp, and Amcarn, this indicator was ≤1.40. In the Far Eastern sample, Lwm had an intermediate value (1.44). The same can be said about the distance between wax mirrors (Lwmd).
Thus, morphometric analysis showed that Prioksky bees corresponded to the subspecies A. m. caucasica. The Far Eastern bee had intermediate values between the samples of A. m. mellifera and subspecies from lineage C or O. Samples of A. m. mellifera from the Republic of Bashkortostan (AmmB and AmmI) did not correspond to standard values according to the cubital index. In particular, the AmmI sample, according to CI, corresponded to the subspecies A. m. caucasica. Perhaps this population underwent hybridization. In the sample from the Burzyansky district, CI was 57%, corresponding to the standard of the Burzyansky population of A. m. mellifera.
3.2. Assessment of the Genetic Structure of the Apis Mellifera Samples
From the evolutionary lineage M on the maternal line came samples from the Iglinsky and Burzyansky districts of Bashkortostan, as well as one colony from the Krasnodar Krai, positioned as A. m. carnica. All other samples belonged to the C/O evolutionary lineages. Bees of the Prioksky breeding stock also descended on the maternal line from lineage C or O (with the markers used, we could not differentiate A. m. caucasica from lineage C subspecies).
The analysis of the polymorphism of microsatellite loci also showed that Prioksky bees, along with bees from the Krasnodar Krai and the Republic of Adygea, belonged to subspecies from the C/O lineages (Table 4, Figure S2). Samples from the Burzyansky and Iglinsky districts were confirmed to belong to A. m. mellifera. The level of introgression of the gene pool of the evolutionary lineages C/O in the AmmI sample was 18.4%, while in the sample AmmB it was 8%. All colonies from the Far Eastern sample originated on the maternal line from lineage C or O. The level of hybridization in this sample was 73.7%. Consequently, the gene pool of this sample was formed by subspecies from the evolutionary lineages M, C, and O, which is confirmed by the history of the settlement of this territory by bees of different subspecies.
We calculated pairwise Fst between the studied samples (Table 5). The greatest divergence was observed between samples of AmmI (dark forest bee A. m. mellifera, Iglinsky district) and Amcau (A. m. caucasica, Krasnopolyansk experimental station), i.e., 0.5736. The smallest divergence was between the samples AmmI and AmmB (A. m. mellifera from the Burzyansky district), i.e., 0.0133.
Phylogenetic analysis was also performed based on a polymorphism analysis of microsatellite loci using genetic distance estimation, according to Nei (1983).
Thus, both samples from the Republic of Bashkortostan corresponded to the declared subspecies A. m. mellifera based both on the results of the analysis of the mitochondrial marker tRNAleu-COII and on the analysis of microsatellite markers. However, in the AmmI sample, the level of introgression of the C/O gene pool was 18.4%. The set of microsatellite loci we used was not capable of differentiating subspecies from the evolutionary lineages C (samples of A. m. carnica and A. m. carpatica) and O (A. m. caucasica). However, a phylogenetic analysis showed (Figure 1) that these samples formed different clusters: samples of A. m. carnica and A. m. carpatica were included in one cluster; samples of A. m. caucasica and Prioksky bees formed another cluster. The Far Eastern sample occupied an intermediate position between the samples of A. m. mellifera and A. m. caucasica.
3.3. Evaluation of Cryopreserved Sperm
In order to evaluate the effectiveness of the cryopreservation method used, some of the cryopreserved sperm was thawed after four days. A comparative assessment of the fresh (n = 100) and frozen–thawed (27) sperm of drones of the Prioksky bees was performed (Table 6). The viability of frozen–thawed sperm (membrane integrity) in the test samples decreased by 20.3% compared to that of fresh sperm. Previously, we showed that honey exhibits cryoprotectant properties, maintaining sperm viability at a level of 37.2 ± 0.5%, and provides sufficiently high protection of the vital resource of the sperm of honey bee drones in combination with 10% DMSO (79.6 ± 1.2%). Of the six queen bees artificially inseminated with frozen–thawed sperm based on a 10% honey extender, two yielded offspring of 96.5–99.1% worker bees [17].
The overall motility of frozen–thawed sperm was reduced by 25-fold compared to that of fresh sperm. In order to determine the fertilizing ability of frozen–thawed sperm, we carried out artificial insemination of 10 virgin queens. During the assessment of the physiological parameters of AI queens, the presence of sperm in the spermatheca of four out of ten queens was revealed (Table 7).
An important physiological parameter of the queen bee, which determines its reproductive abilities, is the concentration of sperm in the seminal receptacle [42]. Previously [9,43], a positive correlation was identified between the concentration of sperm in the spermatheca of the queen and the number of fertilized eggs laid by the queen (r = 0.54 and r = 0.91). The concentration of sperm in the spermatheca of artificially inseminated queens was reported to vary from 1.8 million/μL [44] to 6 million/μL [45], and in naturally mated queens from 4 to 7 million/μL [42]. At the same time, sperm in the spermatheca of natural mating queens can be stored for several years. In our case, the concentration of sperm in the spermatheca of AI queens varied from 0.22 to 4.4 million/μL.
Thus, the reproductive period of the queen bee will depend on the number of sperm that entered its seminal receptacle. It is obvious that queen bees 1 and 3 will have a longer reproductive period (oviposition period) than queens 2 and 4. All four queen bees will produce more than 50% worker bee offspring.
4. Discussion
In this study, we provide an example of creating a biobank of the sperm of honeybee drones of certain subspecies. For subspecies identification, we used morphometric and genetic data. Genetic analysis showed that Prioksky bees, along with bees from the Krasnodar Krai and Adygea, belonged to subspecies from the C/O lineage. At the same time, phylogenetic analysis showed that the Prioksky bees formed one cluster with a sample of A. m. caucasica from the Krasnopolyansk experimental station. The Prioksky breeding stock was bred at the Federal Beekeeping Research Centre on the basis of crossing A. m. mellifera and A. m. caucasica [41,46]. The results of a genetic analysis showed that the Prioksky sample was more consistent with the subspecies A. m. caucasica and the level of the gene pool of A. m. mellifera in it was only 1.4%. Samples of A. m. carnica and A. m. carpatica also formed one cluster. Samples AmmB and AmmI were confirmed to belong to A. m. mellifera. The sample from the Far East was of hybrid origin. The Far Eastern breeding stock was created by crossing A. m. mellifera, A. m. carpatica, A. m. caucasica, A. m. remipes, and A. m. ligustica [39]. Thus, the genetic analysis confirmed the hybrid origin of this breeding stock.
Morphometric analysis also showed that Prioksky bees corresponded to the subspecies A. m. caucasica. The Far Eastern bees had intermediate values between the samples of A. m. mellifera and the samples A. m. carnica, A. m. caucasica, and A. m. carpatica. Samples of A. m. mellifera from the Republic of Bashkortostan did not correspond to standard values according to the cubital index. In particular, the AmmI sample, according to the cubital index, corresponded to the subspecies A. m. caucasica. Perhaps this population has undergone hybridization. According to the genetic analysis of the AmmI sample, the level of introgression of the C/O gene pool was 18.4%, which confirms hybridization. In the AmmB sample, the cubital index was 57%, which corresponds to the standard of the Burzyansky population of A. m. mellifera. The level of hybridization in this sample was only 8%. Previously, Oleksa and Tofilski (2015) [47] showed that morphometric and genetic methods yielded almost the same result—more than 90% of colonies were classified as one subspecies. These authors used 17 microsatellite loci and the mtDNA COI-COII locus as a genetic method, and as a morphometric method, they employed the geometric morphometry of the venation of the fore wing. In our study, the results of genetic and morphometric analyses were also consistent with each other.
An analysis of the viability and motility of cryopreserved sperm showed that the use of honey makes it possible to achieve significant progress in the preservation of reproductive gametes during low-temperature freezing. The main component of honey is carbohydrates (fructose, glucose, sucrose, maltose), dissolved in a small amount of water, as well as vitamins B1, B2, B6, E, K, C, carotene, and folic acid in small quantities. Honey has proven antimicrobial (antibacterial, antimycotic, antimycobacterial) properties, the interest in which has recently been growing [48]. The addition of honey to an extender for cryopreservation significantly improves sperm motility after thawing and the integrity of membranes and acrosomes and also reduces the number of abnormalities in sperm morphology in horses [49], bulls [50], buffalo [51], goats [52], sheep [53], rats and mice [54,55], fish [56,57], and humans [58].
When using honey as a cryoprotectant, it is necessary to take into account the fact that the physicochemical composition of honey also depends on its botanical origin. This circumstance may explain the different results obtained by other researchers when using honey as the main extender. For example, Malik et al. (2017) [50], experimenting with the replacement of glycerol with honey at a concentration of 8%, revealed a significant increase in abnormalities in sperm morphology (8.35 ± 0.16%), while the viability (82.19 ± 1.41%) and motility (76.63 ± 3.21%) did not differ significantly from those of fresh sperm. Shikh Maidina et al. (2018) and Fanni et al. (2018) [52,56], on the contrary, did not find major changes in sperm morphology but noted frequent changes in the head and tail of the flagellum. Other authors [53] reported a lower percentage of dead and abnormal sperm when using honey.
5. Conclusions
Using morphometric and genetic methods, we selected the sperm of the drone bees of different subspecies. A honey solution was used as a cryopreservative. It was shown that the main indicators of frozen–thawed sperm decreased compared to those of fresh sperm. However, the use of honey in sperm cryopreservation has great potential. With the help of cryopreservation, it will be possible to preserve endangered subspecies of Apis mellifera L.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: The studied morphometric parameters; Figure S2: Genetic structure of studied samples.
Author Contributions
Conceptualization, K.M.D. and G.A.N.; methodology, G.A.N., B.A.S., K.M.D.; formal analysis, G.A.N., B.A.S., L.E.O., K.M.D.; investigation, G.A.N., B.A.S., L.E.O., S.E.S., K.M.D.; writing—original draft preparation, K.M.D. and G.A.N.; writing—review and editing, B.A.S., L.E.O., S.E.S.; funding acquisition, G.A.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Ministry of Science and Higher Education of the Russian Federation, grant no. 075-15-2021-1037 (internal no. 15.BRК.21.0001).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Edesi, J.; Tolonen, J.; Ruotsalainen, A.L.; Aspi, J.; Häggman, H. Cryopreservation enables long-term conservation of critically endangered species Rubus humulifolius. Biodivers Conserv 2020, 29, 303–314. [Google Scholar] [CrossRef]
- Kaviani, B.; Kulus, D. Cryopreservation of Endangered Ornamental Plants and Fruit Crops from Tropical and Subtropical Regions. Biology 2022, 11, 847. [Google Scholar] [CrossRef]
- Leroy, G.; Boettcher, P.; Besbes, B.; Danchin-Burge, C.; Baumung, R.; Hiemstra, S.J. Cryoconservation of Animal Genetic Resources in Europe and Two African Countries: A Gap Analysis. Diversity 2019, 11, 240. [Google Scholar] [CrossRef]
- Paillard, M.; Rousseau, A.; Giovenazzo, P.; Bailey, J.L. Preservation of Domesticated Honey Bee (Hymenoptera: Apidae) Drone Semen. J Econ Entomol 2017, 110, 1412–1418. [Google Scholar] [CrossRef]
- Harbo, J.R. Survival of honey bee (Hymenoptera, Apidae) spermatozoa after 2 years in liquid-nitrogen (-196°C). Ann. Entomol. Soc. Am. 1983, 76, 890–891. [Google Scholar] [CrossRef]
- Hopkins, B. K.; Herr, C.; Sheppard, W.S. Sequential generations of honey bee (Apis mellifera) queens produced using cryopreserved semen. Reproduction, Fertility and Development 2012, 24, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Wegener, J.; May, T.; Kamp, G.; Bienefeld, K. New methods and mediafor the centrifugation of honey bee (Hymenoptera: Apidae) drone semen. Journal of Economic Entomology 2014, 107, 47–53. [Google Scholar] [CrossRef]
- Dadkhah, F.; Nehzati-Paghaleh, G.; Zhandi, M.; Hopkins, B.K. Preservation of honey bee spermatozoa using egg yolk and soybean lecithin-based semen extenders and a modified cryopreservation protocol. Journal of Apicultural Research 2016, 55, 279–283. [Google Scholar] [CrossRef]
- Gul, А.; Nuray, S.; Onal, A.G.; Hopkins, B.K.; Sheppard, W.S. Effects of diluents and plasma on honey bee (Apis mellifera L.) drone frozen-thawed semen fertility. Theriogenology 2017, 101, 109–113. [Google Scholar] [CrossRef]
- Alcay, S.; Cakmak, S.; Cakmak, I.; Mulkpinar, E.; Gokce, E.; Ustuner, B.; Sen, H.; Nur, Z. Successful cryopreservation of honey bee drone spermatozoa with royal jelly supplemented extenders. Cryobiology 2019, 87, 28–31. [Google Scholar] [CrossRef]
- Melnichenko, A.N.; Vavilov, Y.L. Long term storage of drone semen by freezing in liquid nitrogen. SP S Kennan, Soviet Agric. Sci 1976, 1, 34–36. (In Russian) [Google Scholar]
- Kakpakov, V.T. Center for instrumental (artificial) insemination of honey bees (CIIHB). Veterinary pathology, 2007; 1, 28–30. (In Russian) [Google Scholar]
- Gulov, A.N. Problems of honeybee conservation of genetic resources. Bee Journal 2018, 6, 22–25. (In Russian) [Google Scholar]
- Gulov, A.N.; Bragina, E.E. Cryopreservation effects on drone swarm Apis mellifera L. morphometric parameters and infrastructure. Veterinary medicine, animal science, biotechnology 2022, 1, 75–86. [Google Scholar] [CrossRef]
- Hinting, A.; Comhaire, F.; Vermeulen, L.; Dhort, M.; Vermeulen, A.; Vanderberhove, D. Value of sperm characteristics and the result of in vitro fertilization for predicting the outcome of assisted reproduction. Int J Androl. 1990, 13, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Duncan, W.W.; Glew, M.J.; Wang, X.; Flaherty, S.P.; Matthews, C.D. Prediction of fertilization rates from semen variables. Fertil Steril 1993, 59, 1233–8. [Google Scholar] [CrossRef]
- Gulov, A.N. , Laskin, A. S. Honey diluent for cryopreservation of honey bee drone sperm. Genetics and animal breeding 2020, 4, 27–36. (In Russian) [Google Scholar] [CrossRef]
- Ruttner, F. Biogeography and Taxonomy of Honeybees. Springer, Berlin, 1988; 291 p. [CrossRef]
- Cridland, J.M.; Tsutsui, N.D.; Ramírez, S.R. The complex demographic history and evolutionary origin of the western honey bee, Apis mellifera. Genome Biol. Evol. 2017, 9, 457–472. [Google Scholar] [CrossRef]
- Harbo, J.R.; Harris, J.W. Selecting honey bees for resistance to Varroa jacobsoni. Apidologie 1999, 183–196. [Google Scholar] [CrossRef]
- Hoppe, A.; Du, M.; Bernstein, R.; Tiesler, F.-K.; Kärcher, M.; Bienefeld, K. Substantial Genetic Progress in the International Apis mellifera carnica Population Since the Implementation of Genetic Evaluation. Insects 2020, 11, 768. [Google Scholar] [CrossRef]
- Maucourt, S.; Fortin, F.; Robert, C.; Giovenazzo, P. Genetic Progress Achieved during 10 Years of Selective Breeding for Honeybee Traits of Interest to the Beekeeping Industry. Agriculture 2021, 11, 535. [Google Scholar] [CrossRef]
- Sprau, L.; Traynor, K.; Rosenkranz, P. Honey bees (Apis mellifera) preselected for Varroa sensitive hygiene discriminate between live and dead Varroa destructor and inanimate objects. Sci Rep 2023, 13, 10340. [Google Scholar] [CrossRef]
- Jensen, A.B.; Palmer, K.A.; Boomsma, J.J.; Pedersen, B.V. Varying degrees of Apis mellifera ligustica introgression in protected populations of the black honeybee, Apis mellifera mellifera, in northwest Europe. Mol. Ecol. 2005, 14, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Oleksa, A.; Chybicki, I.; Tofilski, A.; Burczyk, J. Nuclear and mitochondrial patterns of introgression into native dark bees (Apis mellifera mellifera) in Poland. J Apicult Res. 2011, 50, 116–129. [Google Scholar] [CrossRef]
- Alpatov, W.W. Biometrical studies on variation and races of the honey bee (Apis mellifera L.). Quarterly Review of Biology 1929, 4, 1–58. [Google Scholar] [CrossRef]
- Meixner, M.D.; Pinto, M.A.; Bouga, M.; Kryger, P.; Ivanova, E.; Fuchs, S. Standard methods for characterising subspecies and ecotypes of Apis mellifera. Journal of Apicultural Research 2013, 52, 1–28. [Google Scholar] [CrossRef]
- Garnery, L.; Solignac, M.; Celebrano, G.; Cornuet, J.-M. A simple test using restricted PCR-amplified mitochondrial DNA to study the genetic structure of Apis mellifera L. Experientia 1993, 49, 1016–1021. [Google Scholar] [CrossRef]
- Bertrand, B.; Alburaki, M.; Legout, H.; Moulin, S.; Mougel, F.; Garnery, L. MtDNA COI-COII marker and drone congregation area: an efficient method to establish and monitor honeybee (Apis mellifera L.) conservation centers. Mol Ecol Resour. 2015, 15, 673–683. [Google Scholar] [CrossRef]
- Solignac, M.; Vautrin, D.; Loiseau, A.; Mougel, F.; Baudry, E.; Estoup, A.; Garnery, L.; Haberl, M. ; Cornuet, J-M. Five hundred and fifty microsatellite markers for the study of the honeybee (Apis mellifera L.) genome. Molecular Ecology Notes 2003, 3, 307–311. [Google Scholar] [CrossRef]
- Parejo, M.; Wragg, D.; Gauthier, L.; Vignal, A.; Neumann, P.; Neuditschko, M. Using Whole-Genome Sequence Information to Foster Conservation Efforts for the European Dark Honey Bee, Apis mellifera mellifera. Frontiers in Ecology and Evolution 2016, 4, 583. [Google Scholar] [CrossRef]
- Chapman, N.C.; Harpur, B.A.; Lim, J.; Rinderer, T.E.; Allsopp, M.H.; Zayed, A.; Oldroyd, B.P. A SNP test to identify Africanized honeybees via proportion of “African” ancestry. Mol. Ecol. Resour. 2015, 15, 1346–1355. [Google Scholar] [CrossRef] [PubMed]
- Momeni, J.; Parejo, M.; Nielsen, R.O.; Langa, J.; Montes, I.; Papoutsis, L.; Farajzadeh, L.; Bendixen, C.; Căuia, E.; et al. Authoritative subspecies diagnosis tool for European honey bees based on ancestry informative SNPs. BMC Genomics 2021, 22, 101. [Google Scholar] [CrossRef]
- Petrov, E. M. Bashkirskaya bortevaya pchela. Ufa, Bashkirskoe knizhnoe izdatel'stvo, 1983. 200 p. (In Russian).
- Krivcov, N.I.; Sokol'skij, S.S.; Lyubimov, E.M. Serye gornye kavkazskie pchyoly. Nauchnoe izdanie, Sochi, 2009; 192 p. (In Russian).
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Effect of in-hive miticides on drone honey bee survival and sperm viability. Journal of Apicultural Research 2013, 52, 88–95. [Google Scholar] [CrossRef]
- 37. Bienkowska, М; Panasiuk, B.; Loc, K. Influence of the age of honey bee queens and dose of semen on condition of instrumentally inseminated queens kept in cages with 25 worker bees in the colonies. Journal of Apicultural Science 2008, 52, 23–33.
- Ruttner, F. Instrumental Insemination of the Queen Bee. Bucharest, Apimondia, 1975; 127 p.
- Bilash, G. D., Krivcov N. I. Selekciya pchyol. Moscow, Agropromizdat, 1991; 304 p. (In Russian).
- Bouga, M.; Alaux, C.; Bienkowska, M.; Büchler, R.; Carreck, N.L.; Cauia, E.; Chlebo, R.; Dahle, B.; Dall'Olio, R.; De la Rúa, P.; Gregorc, A.; Ivanova, E.; Kence, A.; Kence, M.; Kezic, N.; Kiprijanovska, H.; Kozmus, P.; Kryger, P.; Le Conte, Y.; Lodesani, M.; Manuel, A.; Siceanu, A.; Soland, G.; Uzunov, A.; Wilde, J. A review of methods for discrimination of honey bee populations as applied to European beekeeping. Journal of Apicultural Research 2011, 50, 51–84. [Google Scholar] [CrossRef]
- Frunze, O.; Brandorf, A.; Kang, E.-J.; Choi, Y.-S. Beekeeping Genetic Resources and Retrieval of Honey Bee Apis mellifera L. Stock in the Russian Federation: A Review. Insects 2021, 12, 684. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S.W. Comparison studies of instrumentally inseminated and naturally mated honey bee queens and factors affecting their performance. Apidologie 2007, 38, 390–410. [Google Scholar] [CrossRef]
- Wegener, J.; Bienefeld, K. Toxicity of cryoprotectants to honey bee semen and queens. Theriogenology 2012, 77, 600–607. [Google Scholar] [CrossRef]
- Woyke, J.; Jasinski, Z. Influence of the number of attendant workers on the number of spermatozoa entering the spermatheca of instrumentally inseminated queens kept outdoors in mating nuclei. J. Apicultural Science 1982, 21, 129–133. [Google Scholar] [CrossRef]
- Mannapov, A.G.; Lyakhov, V.V.; Brovarsky, V.D. Evaluation of instrumental insemination technologies. Bee Journal 2013, 6, 21. (In Russian) [Google Scholar]
- Savushkina, L.N.; Borodachev, A.V. Biological signs of Prioksky bees. Bee Journal 2014, 10, 10–12. (In Russian) [Google Scholar]
- Oleksa, A.; Tofilski, A. Wing geometric morphometrics and microsatellite analysis provide similar discrimination of honey bee subspecies. Apidologie 2015, 46, 49–60. [Google Scholar] [CrossRef]
- Bicudo de Almeida-Muradian, L.; Barth, O.M.; Dietemann, V.; Eyer, M.; da Silva de Freitas, A.; Martel, A.-C.; Marcazzan, G.L.; Marchese, C.M.; Mucignat-Caretta, C.; Pascual-Maté, A.; Reybroeck, W.; Sancho, M.T.; Gasparotto Sattler, J.A. Standard methods for Apis mellifera honey research. Journal of Apicultural Research 2020, 59, 1–62. [Google Scholar] [CrossRef]
- El-Sheshtawy, R.I.; El-Badry, D.A. Natural honey as a cryoprotectant to improve Arab stallion post-thawing sperm parameters. Asian Pacific Journal of Reproduction 2016, 5, 331–334. [Google Scholar] [CrossRef]
- Malik, A.; Fauzi, R.; Zakir, M.I. Subtitusi Madu Asli Pengganti Gliserol dalam Pembekuan pada Kualitas Pasca-thawing Spermatozoa Sapi Bali. Acta veterinaria indonesiana 2017, 5, 98–104. [Google Scholar] [CrossRef]
- Kandiel, M.M.M.; El-Khawagah, A.R.M.; Hussein, M.N.A.; Caoet, X. Quantitative Ultrastructure Evaluation of Egyptian Buffalo Bull Frozen-Thawed Spermatozoa under the Effect of Honey. Scholars Journal of Agriculture and Veterinary Sciences 2019, 6, 92–98. [Google Scholar] [CrossRef]
- Shikh Maidin, M.; Padlan, M.H.; Azuan, S.A.N.; Jonit, R.; Mohammed, N.H.; Abdullah, R. Supplementation of Nigella sativa Oil and Honey Prolong the Survival Rate of Fresh and Post-Thawed Goat Sperms. Tropical Animal Science Journal 2018, 41, 94–99. [Google Scholar] [CrossRef]
- Zaghloul, A.A. Relevance of Honey Bee in Semen Extender on the Quality of Chilled-Stored Ram Semen. J. Animal and Poultry Prod. 2017, 8, 1–5. [Google Scholar] [CrossRef]
- Syazana, N.S.; Hashida, N.H.; Majid, A.M.; Durriyah Sharifah, H.A.; Kamaruddin, M.Y. Effects of Gelam Honey on Sperm Quality and Testis of Rat. Sains Malaysiana 2011, 40, 1243–1246. [Google Scholar]
- Zoheir, K.M.A.; Harisa, G.I.; Abo-Salem, O.M.; Ahmad, S.F. Honey bee is a potential antioxidant against cyclophosphamide induced genotoxicity in albino male mice. Pak. J. Pharm. Sci. 2015, 28, 973–981. [Google Scholar]
- Fanni, N.A.; Santanumurti, M.B.; Suprayogi, T.W.; Bendryman, S.S. Quality enhancement of cryopreserved spermatozoa of sutchi catfish (Pangasianodon hypophthalmus) with honey addition. Iraqi Journal of Veterinary Sciences 2018, 32, 231–236. [Google Scholar] [CrossRef]
- Ogretmen, F.; İnanan, B.E. Evaluation of cryoprotective effect of turkish pine honey on common carp (Cyprinus Carpio) spermatozoa. CryoLetters 2014, 35, 427–437. [Google Scholar] [PubMed]
- Fakhrildin, M.B.; Alsaadi, R.A. Honey Supplementation to Semen-Freezing Medium Improves Human Sperm Parameters Post-Thawing. Journal of Family and Reproductive Health 2014, 8, 27–31. [Google Scholar]
Figure 1.
Dendrogram of the genetic relatedness of the studied samples of honey bees (Nei, 1983). .
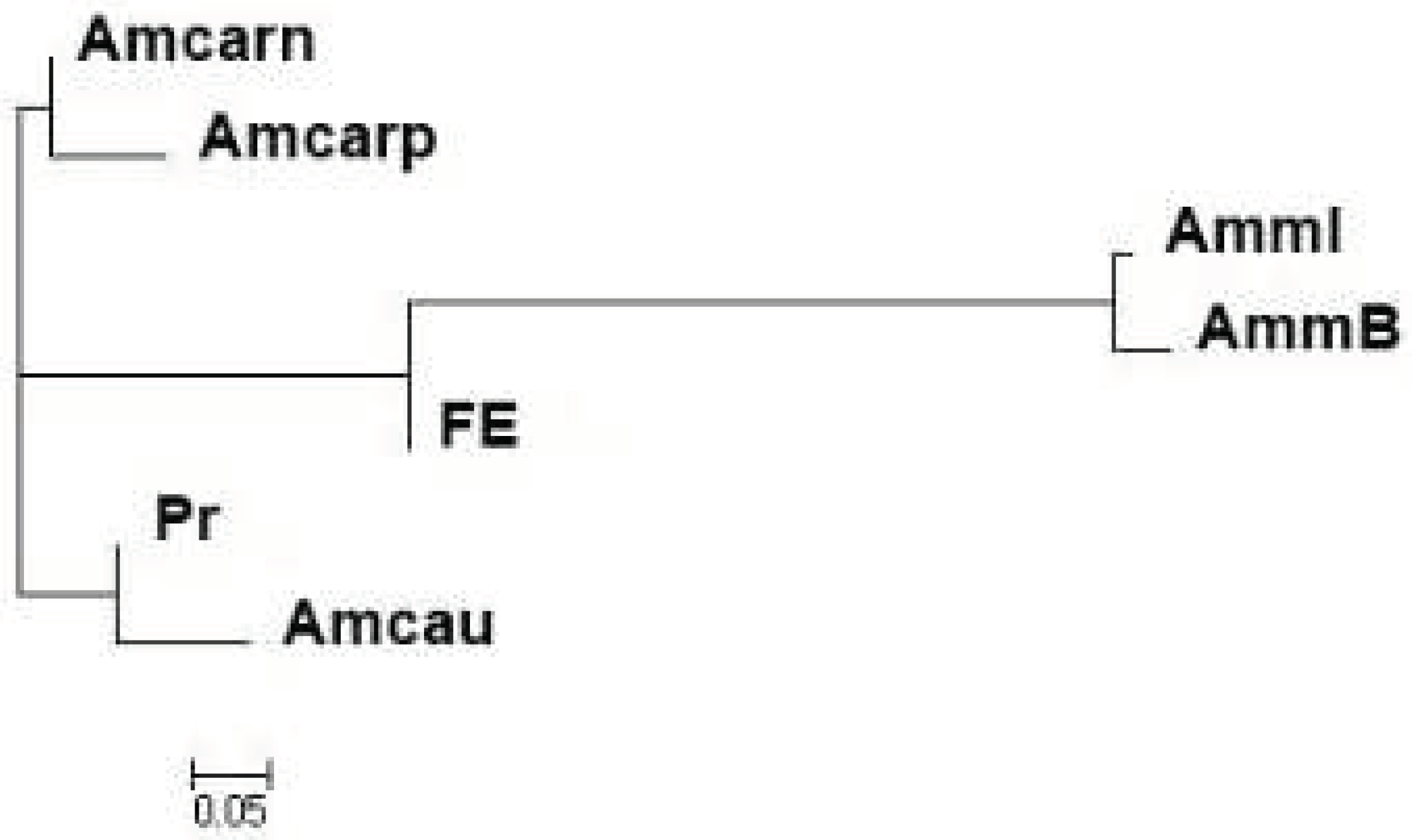

Table 1.
Characteristics of the studied samples.
| Sampling region | Sample name | N | Subspecies |
|---|---|---|---|
| The Republic of Bashkortostan, Iglinsky district | AmmI | 10 | A. m. mellifera |
| The Republic of Adygea, Maykop | Amcarn | 8 | A. m. carnica |
| The Republic of Adygea, Maykop | Amcarp | 10 | A. m. carpatica |
| Far East, Primorsky Krai (Kondratenovka and Tichoreshnoe villages) | FE | 10 | Hybrid of A. m. mellifera, A. m. carpatica, and A. m. caucasica |
| The Republic of Bashkortostan, Burzyansky district, Shulgan-Tash Nature Reserve | AmmB | 9 | A. m. mellifera |
| Ryazan region, Rybnoye, The Federal Beekeeping Research Centre apiary | Pr | 10 | Prioksky bees, hybrid of A. m. mellifera and A. m. caucasica |
| Krasnodar Krai, Adler, Krasnopolyansk experimental beekeeping station | Amcau | 19 | A. m. caucasica |
Table 2.
Morphometric characteristics of the study and reference samples.
| Sample | N | CI, % | Lx, mm | Wt3, mm |
| Pr | 10 | 50.5 | 6.88 | 4.84 |
| Standard values for Prioksky bees | 55–60 | 6.6–6.9 | 4.6–5.0 | |
| AmmI | 10 | 52.9 | 6.17 | 4.92 |
| AmmB | 9 | 57.0 | 6.21 | 4.91 |
| Standard values for A. m. mellifera | 60–65 | 6.0–6.4 | 4.8–5.2 | |
| Amcarp | 10 | 41.9 | 6.63 | 4.81 |
| Standard values for A. m. carpatica | 33–43 | 6.3–7.0 | 4.4–5.1 | |
| Amcarn | 8 | 39.4 | 6.71 | 4.90 |
| Standard values for A. m. carnica | <40.0 | 6.4–6.8 | 4.7–5.1 | |
| FE | 10 | 43.9 | 6.53 | 4.97 |
| Standard values for Far Eastern bees | 28–60 | 6.1–6.8 | 4.6–5.4 | |
| Amcau | 15 | 52.4 | 7.01 | 4.80 |
| Standard values for A. m. caucasica | 50–55 | 6.7–7.2 | 4.4–5.0 | |
| A. m. mellifera* | 61.4 | 6115 | - | |
| A. m. carnica* | 51.2 | 6458 | - | |
| A. m. caucasica* | 54.7 | 6976 | - |
Table 3.
Morphometric characteristics of the study samples.
| Sample | N | FL, mm |
FW, mm |
TI, % | Ls3, mm | Lwm, mm | Lwm, mm | Lwmd, mm | Lt3, mm |
| Pr | 10 | 9.43 | 3.16 | 56.5 | 2.86 | 1.39 | 2.48 | 0.29 | 2.29 |
| AmmI | 10 | 9.34 | 3.09 | 54.6 | 2.92 | 1.50 | 2.55 | 0.22 | 2.36 |
| AmmB | 9 | 9.37 | 3.09 | 55.6 | 2.91 | 1.50 | 2.53 | 0.21 | 2.33 |
| Amcarp | 10 | 9.30 | 3.12 | 55.7 | 2.82 | 1.39 | 2.46 | 0.29 | 2.25 |
| Amcarn | 8 | 9.36 | 3.14 | 55.1 | 2.85 | 1.40 | 2.50 | 0.29 | 2.27 |
| FE | 10 | 9.37 | 3.16 | 55.5 | 2.93 | 1.44 | 2.53 | 0.27 | 2.34 |
| Amcau | 15 | 9.40 | 3.13 | 56.8 | 2.85 | 1.39 | 2.47 | 0.31 | 2.27 |
| Mean | 9.37 | 3.13 | 55.7 | 2.88 | 1.43 | 2.50 | 0.27 | 2.30 | |
| Min | 9.30 | 3.09 | 54.6 | 2.82 | 1.39 | 2.46 | 0.21 | 2.25 | |
| Max | 9.43 | 3.16 | 56.8 | 2.93 | 1.50 | 2.55 | 0.31 | 2.36 |
Table 4.
Results of the genetic analysis of the studied samples of Apis mellifera.
| Sample | Allelic variant of tRNAleu-COII | Gene pool of С/О | Gene pool of М |
|---|---|---|---|
| M lineage | 98 PQQ, 38 PQQQ | 0.016 | 0.984 |
| C/O lineage | 120 Q | 0.993 | 0.007 |
| AmmI | 10 PQQ | 0.184 | 0.816 |
| Amcarn | 7 Q, 1 PQQ | 0.946 | 0.054 |
| Amcarp | 10 Q | 0.967 | 0.033 |
| FE | 10 Q | 0.737 | 0.263 |
| AmmB | 9 PQQ | 0.080 | 0.920 |
| Pr | 10 Q | 0.986 | 0.014 |
| Amcau | 19 Q | 0.988 | 0.012 |
Table 5.
Pairwise Fst between samples (* = P < 0.05, NS = not significant).
| AmmI | Amcarn | Amcarp | FE | AmmB | Pr | Amcau | |
|---|---|---|---|---|---|---|---|
| AmmI | 0.0000 | * | * | * | NS | * | * |
| Amcarn | 0.4174 | 0.0000 | NS | * | * | NS | * |
| Amcarp | 0.4602 | 0.0389 | 0.0000 | * | * | * | * |
| FE | 0.2546 | 0.1140 | 0.1959 | 0.0000 | * | * | * |
| AmmB | 0.0133 | 0.4682 | 0.5177 | 0.3030 | 0.0000 | * | * |
| Pr | 0.4414 | 0.0944 | 0.1606 | 0.2014 | 0.4677 | 0.0000 | * |
| Amcau | 0.5736 | 0.2466 | 0.3267 | 0.4059 | 0.5843 | 0.1311 | 0.0000 |
Table 6.
Main indicators of the sperm quality of Prioksky drones before and after four days of cryopreservation.
Table 6.
Main indicators of the sperm quality of Prioksky drones before and after four days of cryopreservation.
| Indicator | Frozen–thawed sperm (n = 27) | Fresh sperm (n = 100) | ||||
| M ± m (min – max) | σ | Cv, % | M ± m (min – max) | σ | Cv, % | |
| Motility, % | 2.2 ± 0.6 (0–11.5) | 3.1 | 141.04 | 55.0 ± 2.6 (0–99.8) | 26.5 | 48.3 |
| Viability, % | 64.0 ± 1.8 (41.5–83.7) | 9.6 | 14.87 | 84.3 ± 1.2 (40–99.9) | 12.2 | 14.5 |
Table 7.
Physiological parameters of Prioksky queen bees artificially inseminated with frozen–thawed sperm.
Table 7.
Physiological parameters of Prioksky queen bees artificially inseminated with frozen–thawed sperm.
| No. of AI queens | Concentration of sperm in the spermatic receptacle, millions/μL | Presence of sperm in paired oviducts |
| 1 | 2.4 ± 0.25 (2.2–2.7) | absent |
| 2 | 0.9 ± 0.3 (0.6–1.2) | absent |
| 3 | 4.4 ± 0.3 (4.1–4.7) | absent |
| 4 | 0.22 ± 0.02 (0.2–0.25) | sperm traces |
| 5 | 0 | large amount of sperm |
| 6 | 0 | large amount of sperm |
| 7 | 0 | large amount of sperm |
| 8 | 0 | absent |
| 9 | 0 | absent |
| 10 | 0 | absent |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



