
Submitted:
12 March 2024
Posted:
13 March 2024
You are already at the latest version
Abstract
In the brain, astrocytes regulate shape and functions of the synaptic and vascular compartments through a variety of released factors and membrane-bound proteins. An imbalanced astrocyte activity can therefore have drastic negative impacts on brain development, leading to the onset of severe pathologies. Clinical and pre-clinical studies show alterations in astrocyte cell number, morphology, molecular makeup and astrocyte-dependent processes in different affected brain regions in neurodevelopmental (ND) and neuropsychiatric (NP) disorders. Astrocytes proliferate, differentiate and mature during the critical period of early postnatal brain development, a time window of elevated glia-dependent regulation of a proper balance between synapse formation/elimination, which is pivotal in refining synaptic connectivity. Therefore, any intrinsic and/or extrinsic factors altering these processes during the critical period may result in an aberrant synaptic remodelling and onset of mental disorders. The peculiar bridging position of astrocytes between synaptic and vascular compartments further allows them to “compute” the brain state and consequently secrete factors in the bloodstream, which may serve as diagnostic biomarkers of distinct healthy or disease conditions.Here, we collect recent advancements regarding astrogenesis and astrocyte-mediated regulation of neuronal network remodelling during early postnatal critical periods of brain development, focusing on synapse elimination. We then propose alternative hypotheses for an involvement of aberrancies in these processes in the onset of ND and NP disorders. In light of the well-known differential prevalence of certain brain disorders between males and females, we also discuss putative sex-dependent influences on these neurodevelopmental events.From a translational perspective, understanding age- and sex-dependent astrocyte-specific molecular and functional changes may help to identify biomarkers of distinct cellular (dys)functions in health and disease, favouring the development of diagnostic tools or the selection of tailored treatment options for male/female patients.
Keywords:
astrocytes
; critical period
; synapse phagocytosis
; sex differences
; neuropsychiatric and neurodevelopmental disorders
Main Text
Astrocytes are the most abundant subtype of glial cells populating the brain and spinal cord [1]. Microscopically, they show a typical star-shaped morphology with few major processes extending from the soma, which further ramify into numerous fine branches and leaflets at more distal locations [2,3]. In the past years, several studies and excellent reviews have offered historical overviews about the discovery of astrocytes and covered their numerous physiological roles in the mammalian brain, ranging from their support of the formation/function of neuronal synapses to the development of a properly operating blood-brain barrier (BBB) [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18].
Here, we collect recent findings on astrocyte-dependent contributions to brain development during early postnatal critical periods, focusing on synapse elimination (phagocytosis) and its putative link to the onset of neurodevelopmental (ND) and neuropsychiatric (NP) disorders. Finally, considering the asymmetry in the prevalence of mental disorders between males and females, we discuss the impact of sex differences on astrocyte properties during critical periods and propose their hypothetical causal relation to sex-skewed brain pathologies.
During embryogenesis, in both human and rodent brains, astrocytes are generated from radial glia (RG), which self-renew and differentiate into neurons and macroglia cells, e.g. astrocytes, oligodendrocytes and Schwann cells [19,20,21,22,23,24,25]. Cell divisions of RGs are predominantly neurogenic at early/mid-gestation and turn into gliogenic at late-gestation/early postnatal developmental stages [23,26,27] (Figure 1). The initial steps of gliogenesis produce astrocyte precursors, which then locally proliferate in the different brain areas to increase their numbers and give rise to mature astrocytes, the majority of which consist of protoplasmic and fibrous astrocytes in the grey matter and white matter, respectively [17,20,23,28]. Several studies highlight the heterogeneity of astrocyte morphologies and functions in various regions of the central nervous system (CNS). The most specialized subtypes display very distinctive structural and functional properties such as e.g. the Bergmann glia and velate astrocytes in the cerebellum, the Müller glia cells in the retina or the pituicytes in the neurohypophysis [17,29,30,31,32]. In addition to these general features common to all mammalian brains, hominid primates exhibit evolutionary unique subtypes of astrocytes, i.e. the interlaminar and varicose projection astrocytes [15,18] (Figure 1). Interlaminar astrocytes were first observed in Golgi stained samples of the human cortex by Martinotti, Andriezen and Retzius already by the end of the 19th century and described as cells with small somata residing in the upper cortical layer and long processes extending through deeper layers [33,34,35]. Only later, Colombo and colleagues used glial fibrillary acidic protein (GFAP) to examine these long interlaminar astrocyte processes and proposed that they were primate-specific and putatively involved in favouring a radially acting, long distance intercellular communication across cortical layers [36,37,38,39]. A more recent study has, however, challenged the idea of interlaminar astrocytes being primate-specific, and showed their presence in rodent brains, although they appear morphologically more rudimentary than their primate counterpart [40]. Varicose projection astrocytes have been first identified by Oberheim and colleagues in 2009, after examining GFAP-labelled human cortical brain slices [15]. Located in layers 5 and 6, they morphologically resemble protoplasmic astrocytes, but possess less branched short processes and few [1,2,3,4,5] long projecting ones bearing varicosities and extending within single cortical layers, suggesting a tangential range of action [15,41,42]. The peculiar enrichment of interlaminar and varicose projection astrocytes in human and higher-order primates, when compared to other lower-order primates, suggests their relevance for evolutionary more refined CNS functions, but this has not been definitively proven yet. However, engraftment experiments of human glial progenitors into mouse forebrains at postnatal day 1 have shown increased synaptic plasticity and learning capabilities of the chimeric adult mice, further strengthening the hypothesis of enhanced supportive properties of human astrocytes for specialized CNS abilities [43]. In addition to these morphological differences, human astrocytes show a remarkable faster propagation of calcium waves when compared to rodents’ astrocytes, in contrast to more evolutionary conserved characteristics, such as an elevated intracellular calcium level in response to ATP and glutamate [15,43]. For more detailed comparisons between human and rodent astrocytes, we recommend the excellent reviews from Vasile and colleagues [14] and Verkhraktsy and Nedergaard [17].
Beside these differences in subtypes of astrocytes, in various species from worms and insects to rodents and humans, the number of glia cells and the ratio glia-to-neurons are additional factors worth of consideration, because they both increase with the complexity of the CNS [44,45], suggesting a tight link between the growing number of glial cells with increasingly advanced CNS functions. For instance, the development and evolution of mammalian higher cognitive abilities correlates with the process of gyrification in the cerebral cortex during early postnatal developmental stages [46,47]. This was shown to be associated with a fibroblast growth factor (FGF)-dependent localized expansion of astrocyte numbers in the ferret cortex, with FGF1 and FGF2 being the predominant ligands involved [48,49]. Using a targeted genetic manipulation, Shinmyo and colleagues demonstrated that inhibition of astrocyte proliferation in the ferret cortex prevented the formation of gyri, whereas an FGF-dependent overproduction of astrocytes in the lissencephalic mouse cortex was sufficient to induce gyrus-like protrusions [48].
In general, astrocytes show a great range of heterogeneity in the acquisition of various competencies selectively matching the function(s) of neighbouring cells [2,50,51,52]. To gain such a high degree of intra- and inter-regional diversity, astrocytes must undergo positionally and temporally regulated developmental programmes, strictly modulated by interactions between intrinsic (cell-autonomous) and extrinsic (non cell-autonomous) factors, to ultimately guarantee their specification most aligned to the requirements of the local environment (reviewed in [26]). Among intrinsic programmes, three pathways emerged as crucial players for astrogenesis: the Notch, the BMP, and the JAK-STAT signalling pathways [51,53,54,55]. These pathways, alone or in combination with exogenously secreted molecules, e.g. leukemia inhibitory factor (LIF), bone morphogenetic protein (BMP), sonic hedgehog (Shh), ciliary neurotrophic factor (CNTF) or cardiotrophin-1 (CT-1), induce chromatin changes, promoting astrocyte generation and differentiation from RGs [20,54]. For example, the coordinated activity of Notch and JAK/STAT pathways induces an astrocytic fate by demethylating and thereby activating astrocytic genes, such as GFAP or S100β, with the intermediate activation of the transcription factor STAT3 and the DNA methyltransferase 1 (DNMT1) ([20,56] and reviewed in [54]). On the contrary, the activity of pro-neurogenic factors such as Neurogenin1 (Ngn1), Ngn2, NeuroD, Dlx1/2, Nkx2.1 and Mesh1 inhibits astrogenesis by directly or indirectly blocking the JAK/STAT pathway, favouring the production of excitatory and inhibitory neurons both in vitro and in vivo [23,54,57,58,59,60,61,62,63,64,65,66,67,68,69]. Pro-neurogenic factors promote a temporally simultaneous generation of excitatory and inhibitory neurons during embryonic development, but exert different positional influences: excitatory neurons are produced in the dorsal pallium by the radial migration of locally born neuronal progenitors into their targeted cortical layers, whereas inhibitory neurons are generated in the ventral brain and tangentially migrate to dorsal locations, thus infiltrating the developing cortical plate and dorsal areas [70]. These differences influence each other’s developmental trajectories and astrocyte differentiation, ultimately affecting neuronal circuit formation. Imbalances in this finely tuned ratio may contribute to the cognitive dysfunction and neurological abnormalities of mental disorders, such as autism spectrum disorders (ASD), Alzheimer´s disease and schizophrenia (SCZ) [71,72,73].
Beyond intrinsic factors, evidence that also extrinsic signals guide astrogenesis initially came from co-culture experiments of mouse embryonic RG with cortical slices, which produced neurons when co-cultured on embryonic slices but shifted to a glial fate when co-cultured on postnatal slices [27]. Interestingly, the postnatal release of cytokines such as the gliogenic CT-1 from young neurons promote astrocyte differentiation and their depletion can severely impair astrogenesis [26,27,74]. Similarly, in the cerebellum, neuronal Shh diversifies molecular and functional features of Bergmann glia and velate astrocytes [75]. More generally, a coordinated expression of cell-type-specific ligands and receptors is required to favour selected cell-cell interactions and support the appropriate activation of intracellular pathways to drive an astrocyte heterogeneous specification. Very recently, Voss and colleagues demonstrated that the synergistic and combinatorial activity of five ligand–receptor pairs, driven by the ligands TGFβ2, NLGN1, TSLP, DKK1 and BMP4, guides astrogenesis in human cortical organoids and primary fetal tissue. In this work, they additionally identified a time frame of effective responsivity to gliogenic signals, which corresponds to the initial postnatal developmental stages [76] (Figure 1).
In summary, a tight regulation and coordination of all intracellular and extracellular stimuli along with cell-cell interactions is implicated in controlling astrocyte proliferation and maturation during perinatal and early postnatal developmental periods. This goes hand in hand with neuronal development and is critical for establishing the appropriate subtypes and numbers of astrocytes in any distinct brain region. The balanced relative number of all different cell types, their appropriate functional diversification and the correct formation of elaborated neuronal networks guide the specification of high-order morphological modifications of cortical areas, e.g. gyrification of the cerebral cortex [48,53,77]. These are in turn critical for the proper acquisition of increasingly more complex CNS functions, such as human mental and cognitive abilities [1,44].
Any disruption in these delicate processes might affect the formation and functions of astrocytic and neuronal networks, leading to the onset of severe brain disorders.
Astrocytes and the Critical Period of Early Postnatal Brain Development in Health and Mental Disorders
All phases of astrogenesis (generation/proliferation, differentiation and maturation) take place with distinct time courses in various areas of the CNS during the so-called “critical periods” of postnatal brain development. These periods may vary both in length and cell types involved in different species, depending on the functions that must be refined [78,79,80,81]. Critical periods are also sometimes referred to as “sensitive” periods, although it is still debated whether critical and sensitive periods temporally overlap or should be distinguished [81]. These are time frames when brain plasticity and differentiation/maturation processes are strongly dependent on experience and environmental cues to customize neural circuits and connectivity to the needs of each individual. Therefore, any interaction between intrinsic molecular/biochemical programmes with external factors becomes crucial during critical periods in shaping neuronal circuits to respond with the most adapted behaviours in juvenile, adolescent and adult life [82,83]. As described by Knudsen [81], during these developmental phases structural changes such as axon elaboration and synaptic modifications are instructed by life experiences across brain areas in various species (reviewed in [84]) (Figure 1).
In general, identifying the beginning and end of critical periods, as well as isolating the main factors shaping their opening and closing, might support investigations of time frame(s) and mechanisms relevant for an optimal acquisition of selected cognitive abilities and general mental competences, ultimately contributing to build an individual’s personality. From a translational perspective, a better knowledge of the neurobiological, genetic and environmental determinants affecting the opening/closing of critical periods and the reactions of specific cell types upon exposure to selected physiological or detrimental triggers might be essential to identify time windows, when therapeutic strategies are more effective in re-directing aberrant brain developmental trajectories by directly acting on affected cellular mechanisms.
In rodents, astrogenesis occurs during late-gestation and proceeds over the first three weeks of postnatal CNS development, concomitantly with synaptogenesis, both being key processes for the generation of fully functional neuronal networks [84] (Figure 1). Many astrocyte-derived molecules have been identified so far, playing crucial roles for the formation/functions of excitatory/inhibitory synapses during these developmental time frames. Tan and colleagues have recently reviewed the current knowledge about the cellular and molecular mechanisms by which astrocytes instruct synapse formation/functions through various proteins such as e.g glypicans, TNFα, thrombospondins, Hevin/SPARC ([85,86,87,88,89,90,91,92,93] further reviewed in [5,6,13,94,95]). Additionally, a growing body of evidence is featuring the central role of astrocytes in selectively eliminating synapses during developmental windows [96].
In humans, it is still debated whether critical periods are closed after the juvenile developmental stage (approximately corresponding to the first decade of life) or they further extend into adolescence and early adulthood [79,97]. The initial idea was that the closure of the critical period at the end of puberty marks the end for astrogenesis and synaptic spine formation/elimination in the human cortex. However, this hypothesis mostly relies on the first publications of Huttenlocher and colleagues [98,99], which contained only one brain specimen to support the claim. In contrast, more recent brain scan studies suggest that several dynamic changes in cortical gray matter density and remodelling extend into the third decade of life [79,97,100,101]. Insofar as dendritic growth and branching are mainly limited to early childhood stages, it has been postulated that this supplementary reorganization of neuronal circuitries is centred on synaptic modifications to favour the acquisition of higher brain functions, such as modulation of emotions, cognitive flexibility, socialization skills and others. In this line, investigating how a proper excitatory/inhibitory (E/I) balance instructs the opening/closing of critical periods may support our understanding of changes in plasticity related to synaptic modifications. Intriguingly, however, in disease states an E/I imbalance might seem to emerge more as a homeostatic compensatory mechanism to abnormal circuit activity, rather than a primary defective mechanism caused by e.g an inherited genetic background [102]. The E/I balance is in fact also heavily affected by neighbouring cells, including astrocytes. For example, the deletion of glutamate transporters GLAST and GLT1 in mice results in pathological repetitive behaviours commonly observed in ASD, obsessive-compulsive disorder, and Tourette´s syndrome [103]. Moreover, one study showed that astrocyte reactivity reduces inhibitory currents as a consequence of diminished glutamine and GABA availability [104], although astrocyte reactivity may not always homogeneously respond to disease conditions [105].
Besides these evidences, other theories to explain late-onset neurological or NP disorders suggest defective pruning mechanisms of the initially supernumerary spine synapses among the causes of disease conditions [106,107,108,109]. An over-abundance of unpruned weak synaptic inputs appears to negatively affects the synchronized development of E/I inputs, ultimately disrupting the connectivity of brain circuits in the limbic system [13,110,111].
However, due to the limitations of human neuroimaging tools and the ethical issues associated with studies involving children and adolescents, understanding the cellular mechanisms behind these developmental periods and how perceived adverse events may become neurobiologically embedded in brain circuits and result in ND and NP disorders, requires the continuous development of experimental models recapitulating the human conditions.
The use of animal models has been pivotal in revealing essential physiological underpinnings of critical periods and unravelling molecular pathways that respond to the simultaneous application of pharmacological treatments together with behavioural interventions, to ultimately rewire altered neuronal circuits [112,113,114]. Few seminal studies examined changes in the plasticity of brain circuits in various brain regions, after the application of effectors aimed at reopening critical period-like states. Specifically in two of them, the chronic administration of the antidepressant fluoxetine showed remarkable effects on the reopening of critical period-like neuronal responses, further associated with phenotypical alterations. Interestingly, in a first study, fluoxetine reinstated an ocular dominance plasticity in adult mice, promoting the recovery of visual capabilities in amblyopic animals, accompanied by a reduction of intracortical inhibition [113]. A later work showed how fluoxetine treatment combined with behavioural extinction training induced the erasure of amygdala-dependent fear memories, after reactivating a juvenile-like circuit plasticity associated with a disruption of extracellular matrix structures called perineuronal nets (PNNs) around parvalbumin-positive interneurons [112].
Although both studies evidenced a “rejuvenalization” of brain circuits with changes in the inhibitory system concurrent to the reopening of a critical period-like plasticity, neither of them proposed a cellular or molecular mechanisms possibly involved in the observed disruption of PNNs. Only very recently, the study of Ribot, Breton and colleagues has highlighted the role of astrocyte to close the critical period for visual plasticity in the mouse, favouring a remodelling of the extracellular matrix and associated maturation of the inhibitory system in the visual cortex, although the excitatory system was also, even if minorly, modulated [114]. Based on the knowledge that in rodents astrogenesis occurs during the early postnatal developmental stages and that it is for the most part completed by the end of the critical period around postnatal day (P) 28, the authors postulated that the maturation of astrocytes and its molecular regulators might be relevant for the closure of this period. In line with this, a previous study showed that transplanting immature astrocytes in the visual cortex of adult cats reactivated a period of high brain flexibility, with associated re-induction of ocular-dominance (OD) plasticity in adult animals, similar to what observed upon fluoxetine treatment [115]. In their study, Ribot, Breton and colleagues demonstrated that a molecular switch from proliferative to mature astrocytes corresponded to the closure of the critical period and the astrocyte-specific gap-junction channel subunit connexin 30 (Cx30), an important modulator of hippocampal astrocyte maturation [116], was a pivotal player in these events (Figure 1). Its lower expression in immature astrocytes and higher expression in mature astrocytes correlated with different stages of neuronal network maturation and plasticity in the visual cortex. Furthermore, the astrocyte-specific knockdown of Cx30 favoured a hightened ocular dominance plasticity until P50, confirming the astrocytic Cx30 as a factor necessary to mark the closing of the critical period [114]. It would be intriguing to investigate whether and how immature astrocytes might characterize ND or NP disorders and potentially identify underlying molecular determinants, which might trigger the onset of astrocyte-dependent brain disorders. In humans, early-life adversity (ELA) has been associated with the appearance of severe neurological and mental symptoms later in life, as well as with aberrant astrocyte functions ([82,117] and reviewed in [84]). Besides ELA other chronic stress paradigms are accompanied with reductions in both the number of GFAP-positive astrocytes and the length of GFAP-labeled processes in the prefrontal cortex (PFC), hippocampus or other brain regions, besides changes in the expression levels of various astrocyte-specific molecular markers, suggesting a negative impact of stress on proliferation/maturation processes of astrocytes [118,119,120,121,122,123,124]. For example, three weeks of mild chronic unpredictable stress reduce the number of Sox9/S100ß double positive cells in different subfields of the hippocampus [125]. Moreover, chronic social defeat stress (CSDS) hampers the expression of glucocorticoid receptors (GR) asymmetrically in astrocytes and neurons of the medial PFC. The reduced GR content in astrocytes sequentially affects Ca2+ oscillations and the release of adenosine triphosphate (ATP), which in turn modulates synaptic communication [126]. Additionally CSDS decreases the expression of Cx30 and Cx43, correlating with an impaired frequency of spontaneous EPSCs in slices from the mPFC and hippocampus and suggesting a reversal of the system to a functionally immature phenotype [127]. Furthermore, early social isolation in rodents induces an astrocytic GR-dependent activation of MERTK pathway, ultimately leading to an excessive phagocytosis of excitatory synapses and consequent unbalanced neuronal firing patterns [128]. Strikingly, a selectively lower expression of the astrocyte-specific protein Cx30 characterizes postmortem brain tissues of patients with major depressive disorder (MDD) [129], supporting a link between the expression of cell-type specific markers reminiscent of a critical period-like brain state and CNS pathologies. Furthermore, worth to mention is the association of several mental disorders with a reduced gyrification index (GI) in cortical areas, measured by magnetic resonance imaging (MRI) in patients, also suggestive of hypothetical deficits in astrocyte proliferation affecting cortical formation being behind certain brain diseases ([130,131,132] and reviewed in [133,134]).
Several studies have revealed astrocyte deficiencies in other brain disorders besides MDD, such as SCZ, bipolar disorder (BD), ASD and epilepsy [135,136,137,138,139,140,141,142,143,144]. In young MDD patients, postmortem brains present a profound reduction in the number and morphology of GFAP- and S100β-positive astrocytes in the prefrontal and anterior cingulate corteces and in the hippocampus, all regions heavily affected in MDD [135,138,145,146,147,148]. To validate this, it was also shown that antidepressants target astrocytes and reverse disease symptoms [136,149,150,151]. It remains to be clarified, however, which subtype(s) of astrocytes are affected, as far as currently available markers cannot discriminate among interlaminar, varicose projection or protoplasmic astrocytes. Conversely, in SCZ or BD, changes in astrocytes are less consistent, with different studies showing both their increased and decreased numbers in postmortem tissue of SCZ patients [140,141]. The most reproduced findings show a hypofunction of NMDA receptors (NMDAR) in SCZ. Astrocytes enhance NMDAR activity through the release of gliotransmitters, such as D-serine, which acts as co-agonist of glutamate [152]. The binding of D-serine to synaptic NMDARs additionally modulate the induction of long-term potentiation (LTP), the latter being the cellular correlate of memory formation [153]. Memory deficits are among the most common symptoms of various ND and NP disorders, thereby supporting an involvement of gliotransmission in their etiopathogenesis. In line with this hypothesis, a disrupted NMDAR function may also rely on a imbalanced postsynaptic AMPA receptor composition or missed AMPA receptor maturation due to an altered astrocyte-dependent glypican secretion or chordin-like secretion, respectively [90,154]. Related to this, the work of Caldwell and colleagues recently showed how an aberrant secretion of astrocyte-derived proteins is associated with severe ND disorders accompanied by an altered neuronal development [155]. Indeed, recent clinical work has proposed D-serine as a promising therapeutic option for the treatment of ND disorders, most notably SCZ [156,157]. Moreover, it has further been shown that mutations in Disrupted-In-Schizophrenia 1 (Disc1) reduce the stability of the D-serine synthesizing enzyme serine racemase, resulting in SCZ-like behavior [158]. Thus, in SCZ, not the number but rather the functionality of astrocyte seems to be mostly affected and influence disease onset and/or progression, principally driven by genetic factors. This might explain the inconsistent results from postmortem brain tissues, which did not show reproducible deficiencies in astrocyte counts [140,141]. Despite SCZ having been considered a disorder with a high genetic component, in reality the concordance rate among identical twins is only about 50% [159], with a substantial proportion of SCZ being of idiopathic origin. More interestingly, cortisol levels, an index of stress, measured in adolescents at-risk for SCZ were significantly elevated in subjects who transitioned to psychosis later in life, further supporting the negative impact of early life environmental factors such as stress for worsening NP disorders when encountered during critical periods of postnatal development [160,161]. Although a defective neurogenesis during the critical period has been long described in SCZ and associated with its etiopathogenesis [162], recent work has elucidated the putative role of astrocytes and their aberrant maturation paths impacting synapse formation during critical periods as a possible cause of disrupted neuronal circuit formation and disease onset [142,163]. In ASD, astrocyte involvement in its etiology has been recently investigated using patient-derived iPSCs [144,164]. Even in ASD, the essential role of the critical period has been proposed, with a shortened time frame of neuroplasticity indicated as a common cause of disease among various patients, despite their highly divergent genetic backgrounds. In particular, the language and social skill deficits observed in ASD have been led back to a premature closure of the critical period for their development, which should typically extend into adolescence. The hypothesis is that a shorter critical period may force the brain to rely on underdeveloped neuronal networks for learning language and social skills, among other mental abilities, leading to the typical symptoms observed in ASD patients [165]. However, as of today, no work has thoroughly investigated the possibility that an earlier closure of critical periods, marked by the premature differentiation of astrocytes, might be among the neurobiological underpinnings in the etiopathogenesis of ASD. In the same line, no work has ever considered a failure in the closure of this period among possible causes for other mental disorders such as depression (Figure 2 and Figure 3).
Astrocytes and the Synaptic Compartments: The Relevance of an Astrocyte-Mediated Phagocytosis for Neuronal Circuit Refinement in Health and Disease
In the CNS, the peculiar position of astrocytes between synaptic and vascular compartments supports the formation of a functional unit (neurovascular, NVU) “sensing” the brain state and secreting factors in the bloodstream as a reflection of this state. In pathological conditions, such factors may serve as biomarkers of cellular (dys)functions and help to improve diagnostic/treatment options specifically adapted to the needs of individual patients. Several excellent reviews have already described secreted astrocyte-derived molecules relevant for synapse and BBB properties in health and disease [5,6,10,12,13,166,167,168,169]. We also validated and characterized astrocyte and BBB deficits in an animal model of MDD [168,170] and identified the astrocyte-derived factor GDF15 as an effector of fluoxetine to simultaneously restore the disrupted astrocyte processes and loosened tight junctions between endothelial cells [171]. Ideally, identifying cell-type specific molecular underpinnings of disease may support the discovery of equally specific diagnostic biomarkers. However, ND and NP disorders often share overlapping symptoms and underlying neurobiological alterations, making it difficult to isolate disease-specific biomarkers for personalized medicine. Therefore, only a combination of efforts and methodological approaches may offer valid guidance to reach this ambitious goal. Among less explored functions of astrocytes, we focus here on astrocyte-mediated synaptic phagocytosis during critical periods of brain development and its putative involvement in NP and ND disorders.
Astrocytes derive from the embryonic ectodermal sheet and therefore share some developmental, genetic and functional similarities with other ectodermally-derived cells, such as neurons and oligodendrocytes, but less with other brain cells of mesodermal origin, such as microglia cells [172]. However, they might adopt analogous functions to microglia cells to cooperatively converge their efforts in the regulation of neuronal networks formation [96,173].
Together with microglia, astrocytes contribute to the formation/function of excitatory and inhibitory synapses and to the elimination of weak synapses (pruning) through released factors or membrane-bound molecules [51,169,173,174].
Early studies in the fly Drosophila melanogaster opened the field of glia-mediated phagocytosis, showing how this process is highly conserved across species, even when species-specific cellular and molecular mechanisms diverged along evolution to adapt to the growing complexity of selected functions [44]. In Drosophila, it was shown that, during its larvae-to-pupa metamorphosis, a high degree of tissue digestion occurs to reshape its body structures and nervous system, allowing the subsequent extension of adult, mature, neuronal projections [175]. The sequence of events leading to such drastic changes follows a developmentally regulated pattern, initiated by complex interactions between intrinsic and extrinsic factors, such as the neuronal hormone ecdysone. Specifically, the gene products of Draper and CED-6 were first identified to mediate the engulfment of pruned axons by glia cells during metamorphosis [176]. Accordingly, mutations in these two genes and glia cell-targeted knockdown experiments, suppressed glia-mediated axon pruning during metamorphosis. Genetic studies in Caenorhabditis elegans initially isolated the genes encoding for CED-1 and CED-6 as also essential mediators for the clearance of apoptotic cells [177].
In the mouse brain, the seminal work of Stevens [178] and Chung [174] has been key to underscore novel roles for the classical complement cascade and the MEGF10/MERTK proteins, respectively, for the refinement of neuronal synaptic networks during postnatal developmental periods. Initially, microglia were described as key players to refine synapses by engulfing presynaptic inputs during the peak retinogeniculate pruning via the microglial CR3/C3 phagocytic signalling pathway and under the regulatory influence of neuronal activity [179]. The involvement of microglia in synaptic pruning was simultaneously uncovered in the hippocampus and juvenile visual cortex, where the synaptic remodelling occurs through the fractalkine/CXC3CR1 signalling pathway [180]. Afterwards, numerous studies and comprehensive reviews have extensively examined the crucial role of microglia cells in the process of synapse elimination and in disease states [181,182,183,184,185,186,187,188,189]. Later, an additional involvement of the astrocytic MEGF10 and MERTK was highlighted for the synaptic remodelling during postnatal brain development [174] and the maintenance of hippocampal homeostasis in the adult brain [190]. Microglia and astrocytes collaborate through a synergistic and precisely orchestrated spatiotemporal coordination, ensuring efficient homeostatic phagocytosis of apoptotic cells and synaptic pruning, while respecting each other´s territorial boundaries and competences: microglia exhibit an engulfing preference for large cell bodies, whereas astrocytes favour small dendritic apoptotic bodies [191]. Moreover, Lee and colleagues demonstrated how astrocytes and microglia constantly engage in phagocytosis of both excitatory and inhibitory synaptic elements within the hippocampal CA1, preferring excitatory rather than inhibitory terminals [190]. Similarly, Dejanovic and colleagues observed that astrocytes preferentially engulf excitatory synapses, while microglia tend to target inhibitory ones, revealing a divergence between astrocytes and microglia during (patho)physiological processes ([181] and reviewed in [169]). A recent study has identified a unique GABA-receptive microglia subtype that selectively remodels inhibitory synapses during mouse postnatal cortical development [182]. Furthermore, the MERTK-mediated microglial phagocytosis is crucial for eliminating inhibitory post-synapses in the juvenile brain [183]. Elucidating the complex interplay between microglia and astrocyte regulatory roles in synapse elimination is the current crucial focus of many research labs. Regarding the molecular mechanisms driving these events, the current view postulates that a balanced distribution of “eat-me signals” and “don’t eat me signals” is key to determine which cells (or parts of them) should be excised/phagocytosed [51]. Among “eat-me signals”, the C1q has been described multiple times as a common tagging system shared by microglia cells and astrocytes to identify their targets. For instance, MEGF10 can mediate its phagocytic functions on apoptotic material via the recognition of C1q bound to phosphatidylserine (PS) exposed on dying cells [109]. This suggests that regulatory mechanisms controlling C1q expression/tagging and/or its binding partners may be responsible for sorting out which cell type between microglia cells and astrocytes should be recruited to execute the job. Because of the commonly shared target(s), it has also been proposed that the selective mechanism determining which cell type acts first may rely on the specific set of receptors localized on their membranes, possibly activated alone or in combination with other molecules “on demand” [96].
From the literature, the current hypothesis is that astrocytes stimulate neurons to produce C1q, which then triggers the downstream activation of the complement cascade and C3b deposition, finally promoting synapse elimination through its microglia-specific binding partner C3 receptor (CR3) [178,179] (Figure 3). Another extensively studied “eat-me signal” in the microglia-mediated synaptic elimination is represented by PS exposure on neuronal membranes. The PS presentation on weak synapses promotes microglia-mediated synaptic pruning via the TREM2 receptor and C1q in the developing hippocampus and retinogeniculate system [184]. Beside these positive mediators of microglial engulfment, “don’t eat me signals” also modulate synaptic pruning. These signals counterbalance the effects of “eat me signals”, preventing the engulfment of viable synapses by aberrant microglial removal. Among them, CD47 was first characterized as “don’t eat me signal” because it permitted glioma cells to infiltrate the healthy brain parenchyma, after blocking their macrophage-mediated engulfment [192]. Its alternative function in synaptic pruning was uncovered later, with CD47 expressed on neuronal membranes and interacting with the microglial receptor SIRPα being responsible for suppressing synaptic phagocytosis. Mice lacking CD47 exhibit excess synaptic pruning and decrease synaptic connectivity [193], whereas its increased expression was found in iPSCs derived from autistic patients with enlarged brains accompanied by reduced cellular phagocytosis [194]. However, in contrast to the extensive literature describing the mechanisms of a microglia-mediated synaptic pruning in health and disease [178,179,195,196], those instructing the astrocyte-mediated synaptic elimination are still less known [96,128,173,174,190] (Figure 3).
A reduced proliferation of astrocytes and/or disruption of astrocytic activity characteristic of ND and NP disorders may affect the appropriate formation of synaptic circuits in different ways. Their impaired activity in pathological conditions may for example either weaken mature synapses or prevent their maturation, thereby potentially enhancing their susceptibility to an excessive pruning. Alternatively, an aberrant astrocyte activity may negatively influence the synaptic tagging, leading to either an excessive or abrupted elimination of unwanted synapses. These possibilities further sustain the importance of the critical developmental periods, when astrocyte proliferation/maturation and synaptogenesis take place simultaneously, in sculpting brain circuits in health and disease.
Sex-Dependent Differences in Astrogenesis and in Time Frames of Vulnerability to Perturbations and Disease Onset
Sex differences have long been recognized as a variable affecting many brain disorders in terms of predisposition, rates of incidence, age of onset, symptomatology and outcome, with skewed prevalence towards one sex in different pathologies [111,119,197,198,199]. However, research on sex-dependent disorders and their neurobiological molecular and cellular causes is still scarce, thereby limiting the development of sex-specific diagnoses and treatments.
In the past, many human studies have often lacked sufficient sample availability to stratify results based on sex differences for any given disease category. Nowadays, the access to huge genomic, transcriptomic and proteomic datasets help to discriminate the impact of sex variables on various biological parameters [200,201,202,203]. However, our understanding of whether and how sex might affect astrogenesis, synaptogenesis and synapse elimination in humans and to which extent this contributes to the onset of brain diseases is still limited [204]. Currently, the overall idea is that mental conditions with early-onset neurodevelopmental origin, such as ASD, attention deficit/hyperactivity disorder and SCZ show a higher male prevalence. On the other hand, disorders with a higher emotional component, such as depression, anxiety disorder, and eating disorders, which usually start during puberty or later in life, show a higher female incidence [205,206]. This asymmetric development of psychopathologies has been analyzed across studies with the goal of elucidating how and which sex-specific developmental maturation trajectories might influence it. Interestingly, regional differences in volume and tissue density were found between sexes in areas implicated in sex-biased NP conditions, suggesting a few candidate regions susceptible to sex differences in the developing brain [207]. Among those regions, the cortex has been implicated as one of the brain areas with a high sex-dependent diversification in cellular and synaptic densities [99,111,119,208]. Most of these differences have been attributed to remodelling processes occurring at the synaptic level, with an aberrant pruning of weaker synapses being recently regarded as a mechanism relevant for the onset of brain disorders [110,111]. As previously mentioned, most of these remodelling events take place during the critical periods of brain development, when astrogenesis in parallel with synaptogenesis contribute to the formation and refinement of neuronal circuits [54,78,79,209]. Mechanisms controlling the opening and closure of these developmental time windows might be essential in building healthy neuronal networks and establishing appropriate behaviours in adult life. Recent work has highlighted sex differences in the maturation processes of astrocytes during early postnatal developmental stages, showing that astrocytes in male mice reach a mature state earlier than in female mice [210]. Sex-dependent maturational trajectories are influenced by the perinatal surge in testosterone aimed at masculinizing the brain after birth and establishing sexually dimorphic brain circuitries responsible for sex-differentiated behaviours and reproductive processes [211]. Astroglia respond to circulating gonadal hormones [212], which influence their relative sex-dependent maturation rates [210]. These may in turn correlate with sex-dependent closure times for temporal windows of high plasticity necessary for astrocyte-mediated modulation of neuronal circuit development. It would be highly interesting to investigate whether in ND diseases, for example, a further earlier closure of the critical period, accompanied by a putative premature differentiation of astrocytes, may account for the observed prevalence of disorders such as ASD in male individuals. Although this hypothesis has not yet been tested, we may speculate that a shorter time window for astrocyte maturation in males might make them more vulnerable to diseases affecting the astrocyte-dependent early-onset synaptic changes. This would explain some phenotypic effects seen in ASD. In such a case, there would be a narrower time frame for male individuals carrying those affected cells to recover from (or adapt to) detrimental environmental challenges occurring during this period and to rescue impaired neuronal circuits (Figure 4). Correspondingly, slower astrocyte maturation time frames in females might trigger the onset of female-biased disorders eventually characterized by an inappropriately longer immature brain state. Similar considerations can be true for other pathologies for which a sexual dimorphism has been already recognized, opening alternative lines for their investigations [213] (Figure 4).
Conclusions
In conclusion, we propose astrocytes as critical novel targets for the development of efficacious medical treatments for NP and ND disorders. These disorders could potentially result from aberrant opening/closure of critical developmental periods, accompanied by defective astrocyte proliferation/maturation processes either affecting synapse formation and gyrification and/or destabilizing neuronal circuits with E/I imbalances and/or impairing an astrocyte-dependent synapse elimination. These hypothesis point out the urge of intensifying research efforts to identify the molecular drivers of such processes and propose astrocyte-based therapeutic approaches to reverse disease phenotypes. Moreover, specific attention to early postnatal brain developmental periods, in combination with the study of sex-specific differential maturation patterns, may improve our knowledge regarding physiological and pathological processes taking place during these time spans and develop treatments personalized to the needs of female and male patients.
Author Contributions
This work was supported by intramural funding from the Department of Psychiatry and Psychotherapy of the University of Regensburg and by the German Research Council (DFG-GRK2174 “Neurobiology of Emotion Dysfunction” (P1)) to BDB. The sponsors did not have any role in writing the report and in the decision to submit the article for publication. We would like to thank Dr. Nicholaus Meyers for help with proofreading the manuscript. We further express our gratitude to the reviewers of our manuscript for their time and insightful comments, which surely supported us to improve its quality. We apologize to those colleagues whose work could not be cited because of space limitations.
Conflicts of Interest
The authors declare no conflict of interest in relation to the work described.
References
- Bandeira, F.; Lent, R.; Herculano-Houzel, S. Changing numbers of neuronal and non-neuronal cells underlie postnatal brain growth in the rat. Proc Natl Acad Sci. 2009, 106, 14108–14113. [Google Scholar] [CrossRef]
- Khakh, B.S.; Sofroniew, M.V. Diversity of astrocyte functions and phenotypes in neural circuits. Nat Neurosci. 2015, 18, 942–952. [Google Scholar] [CrossRef]
- Semyanov, A.; Verkhratsky, A. Astrocytic processes: from tripartite synapses to the active milieu. Trends Neurosci. 2021, 44, 781–792. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: from metabolic end-product to signalling molecule. Nat Rev Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron. 2017, 96, 697–708. [Google Scholar] [CrossRef]
- Bosworth, A.P.; Allen, N.J. The diverse actions of astrocytes during synaptic development. Curr Opin Neurobiol. 2017, 47, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.R.; Herrmann, J.E.; Woo, M.J.; Tansey, K.E.; Doan, N.B.; Sofroniew, M.V. Reactive Astrocytes Protect Tissue and Preserve Function after Spinal Cord Injury. J Neurosci. 2004, 24, 2143–2155. [Google Scholar] [CrossRef]
- Iram, T.; Frenkel, D. Targeting the Role of Astrocytes in the Progression of Alzheimers Disease. Curr Signal Transduct Ther. 2012, 7, 20–27. [Google Scholar] [CrossRef]
- Molofsky, A.V.; Kelley, K.W.; Tsai, H.H.; Redmond, S.A.; Chang, S.M.; Madireddy, L.; et al. Astrocyte-encoded positional cues maintain sensorimotor circuit integrity. Nature. 2014, 509, 189–194. [Google Scholar] [CrossRef]
- Cabezas, R.; Ávila, M.; Gonzalez, J.; El-Bachá, R.S.; Báez, E.; García-Segura, L.M.; et al. Astrocytic modulation of blood brain barrier: perspectives on Parkinson´s disease. Front Cell Neurosci [Internet]. 2014 Aug 4 [cited 2022 Jan 12];8. Available from: http://journal.frontiersin.org/article/10.3389/fncel.2014.00211/abstract. [CrossRef]
- Boulay, A.C.; Saubaméa, B.; Adam, N.; Chasseigneaux, S.; Mazaré, N.; Gilbert, A.; et al. Translation in astrocyte distal processes sets molecular heterogeneity at the gliovascular interface. Cell Discov. 2017, 3, 17005. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.I.; Katayama, T.; Prat, A. Glial influence on the blood brain barrier. Glia. 2013, 61, 1939–1958. [Google Scholar] [CrossRef]
- Eroglu, C.; Barres, B.A. Regulation of synaptic connectivity by glia. Nature. 2010, 468, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Vasile, F.; Dossi, E.; Rouach, N. Human astrocytes: structure and functions in the healthy brain. Brain Struct Funct. 2017, 222, 2017–2029. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.; Wang, F.; et al. Uniquely Hominid Features of Adult Human Astrocytes. J Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef]
- Mohn, T.C.; Koob, A.O. Adult Astrogenesis and the Etiology of Cortical Neurodegeneration. J Exp Neurosci. 2015 Jan;9s2:JEN.S25520. [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Bush, N.; Nedergaard, M.; Butt, A. The Special Case of Human Astrocytes. Neuroglia. 2018, 1, 21–29. [Google Scholar] [CrossRef]
- Götz, M.; Huttner, W.B. The cell biology of neurogenesis. Nat Rev Mol Cell Biol. 2005, 6, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Takouda, J.; Katada, S.; Nakashima, K. Emerging mechanisms underlying astrogenesis in the developing mammalian brain. Proc Jpn Acad Ser, B. 2017, 93, 386–398. [Google Scholar] [CrossRef]
- Ever, L.; Gaiano, N. Radial ‘glial’ progenitors: neurogenesis and signaling. Curr Opin Neurobiol. 2005, 15, 29–33. [Google Scholar] [CrossRef]
- Zhou, C.J.; Zhao, C.; Pleasure, S.J. Wnt Signaling Mutants Have Decreased Dentate Granule Cell Production and Radial Glial Scaffolding Abnormalities. J Neurosci. 2004, 24, 121–126. [Google Scholar] [CrossRef]
- Rowitch, D.H.; Kriegstein, A.R. Developmental genetics of vertebrate glial–cell specification. Nature. 2010, 468, 214–222. [Google Scholar] [CrossRef]
- Eze, U.C.; Bhaduri, A.; Haeussler, M.; Nowakowski, T.J.; Kriegstein, A.R. Single-cell atlas of early human brain development highlights heterogeneity of human neuroepithelial cells and early radial glia. Nat Neurosci. 2021, 24, 584–594. [Google Scholar] [CrossRef]
- Pollen, A.A.; Nowakowski, T.J.; Chen, J.; Retallack, H.; Sandoval-Espinosa, C.; Nicholas, C.R.; et al. Molecular Identity of Human Outer Radial Glia during Cortical Development. Cell. 2015, 163, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.D.; Gauthier, A.S. Timing is everything: making neurons versus glia in the developing cortex. Neuron. 2007, 54, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Morrow, T.; Song, M.R.; Ghosh, A. Sequential specification of neurons and glia by developmentally regulated extracellular factors. Dev Camb Engl. 2001, 128, 3585–3594. [Google Scholar] [CrossRef]
- Ge, W.P.; Miyawaki, A.; Gage, F.H.; Jan, Y.N.; Jan, L.Y. Local generation of glia is a major astrocyte source in postnatal cortex. Nature. 2012, 484, 376–380. [Google Scholar] [CrossRef]
- De Zeeuw, C.I.; Hoogland, T.M. Reappraisal of Bergmann glial cells as modulators of cerebellar circuit function. Front Cell Neurosci [Internet]. 2015 Jul 2 [cited 2023 Jun 6];9. Available from: http://journal.frontiersin.org/Article/10.3389/fncel.2015.00246/abstract. [CrossRef]
- Güngör Kobat, S. Importance of Müller Cells. Beyoglu Eye J [Internet]. 2020 [cited 2023 Jun 6]; Available from: http://beyoglueye.com/jvi.aspx?un=BEJ-28290&volume=.
- Choi, B.H.; Lapham, L.W. Evolution of Bergman glia in developing human fetal cerebellum: A Golgi, electron microscopic and immunofluorescent study. Brain Res. 1980, 190, 369–383. [Google Scholar] [CrossRef]
- Reichenbach, A.; Bringmann, A. Glia of the human retina. Glia. 2020, 68, 768–796. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, F. Contributo allo studio della corteccia cerebrale, ed all ́origine centrale dei nervi. Fratelli Bocca; 1889.
- Andriezen, W.L. The Neuroglia Elements in the Human Brain. BMJ. 1893, 2, 227–230. [Google Scholar] [CrossRef]
- Retzius, G. Die neuroglia des Gehirns beim Menschen und bei Saeugethieren. Jena: Chemie; 1894.
- Colombo, J.A.; Yáñez, A.; Puissant, V.; Lipina, S. Long, interlaminar astroglial cell processes in the cortex of adult monkeys. J Neurosci Res. 1995, 40, 551–556. [Google Scholar] [CrossRef]
- Colombo, J.A. Interlaminar Astroglial Processes in the Cerebral Cortex of Adult Monkeys But Not of Adult Rats. Cells Tissues Organs. 1996, 155, 57–62. [Google Scholar] [CrossRef]
- Colombo, J.A.; Yáñez, A.; Lipina, S.J. Interlaminar astroglial processes in the cerebral cortex of non human primates: response to injury. J Hirnforsch. 1997, 38, 503–512. [Google Scholar]
- Colombo, J.A.; Reisin, H.D. Interlaminar astroglia of the cerebral cortex: a marker of the primate brain. Brain Res. 2004, 1006, 126–131. [Google Scholar] [CrossRef]
- Falcone, C.; Penna, E.; Hong, T.; Tarantal, A.F.; Hof, P.R.; Hopkins, W.D.; et al. Cortical Interlaminar Astrocytes Are Generated Prenatally, Mature Postnatally, and Express Unique Markers in Human and Nonhuman Primates. Cereb Cortex. 2021, 31, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Falcone, C.; Martínez-Cerdeño, V. Astrocyte evolution and human specificity. Neural Regen Res. 2023, 18, 131. [Google Scholar] [PubMed]
- Falcone, C.; McBride, E.L.; Hopkins, W.D.; Hof, P.R.; Manger, P.R.; Sherwood, C.C.; et al. Redefining varicose projection astrocytes in primates. Glia. 2022, 70, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Chen, M.; Wang, F.; Windrem, M.; Wang, S.; Shanz, S.; et al. Forebrain Engraftment by Human Glial Progenitor Cells Enhances Synaptic Plasticity and Learning in Adult Mice. Cell Stem Cell. 2013, 12, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, F.A.C.; Carvalho, L.R.B.; Grinberg, L.T.; Farfel, J.M.; Ferretti, R.E.L.; Leite, R.E.P.; et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J Comp Neurol. 2009, 513, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Falcone, C. Evolution of astrocytes: From invertebrates to vertebrates. Front Cell Dev Biol. 2022, 10, 931311. [Google Scholar] [CrossRef] [PubMed]
- Degl’Innocenti, E.; Dell’Anno, M.T. Human and mouse cortical astrocytes: a comparative view from development to morphological and functional characterization. Front Neuroanat. 2023, 17, 1130729. [Google Scholar] [CrossRef]
- Namba, T.; Huttner, W.B. Neural progenitor cells and their role in the development and evolutionary expansion of the neocortex. WIREs Dev Biol. 2017, 6, e256. [Google Scholar] [CrossRef] [PubMed]
- Shinmyo, Y.; Saito, K.; Hamabe-Horiike, T.; Kameya, N.; Ando, A.; Kawasaki, K.; et al. Localized astrogenesis regulates gyrification of the cerebral cortex. Sci Adv. 2022, 8, eabi5209. [Google Scholar] [CrossRef] [PubMed]
- Zilles, K.; Palomero-Gallagher, N.; Amunts, K. Development of cortical folding during evolution and ontogeny. Trends Neurosci. 2013, 36, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Farmer, W.T.; Murai, K. Resolving Astrocyte Heterogeneity in the CNS. Front Cell Neurosci. 2017 Sep 27;11:300. [CrossRef]
- Holt, M.G. Astrocyte heterogeneity and interactions with local neural circuits. Bolaños, J.P.; editor. Essays Biochem. 2023, 67, 93–106. [Google Scholar] [PubMed]
- Miller, S.J. Astrocyte Heterogeneity in the Adult Central Nervous System. Front Cell Neurosci. 2018 Nov 15;12:401. [CrossRef]
- Oliveria, J.P.; Li, Z.J. critical role of astrogenesis and neurodevelopment in Fragile X Syndrome and Rett Syndrome. McMaster Univ Med J [Internet]. 2020 Dec 26 [cited 2023 May 31];17(1). Available from: https://journals.mcmaster.ca/mumj/article/view/2338. [CrossRef]
- Kanski, R.; Van Strien, M.E.; Van Tijn, P.; Hol, E.M. A star is born: new insights into the mechanism of astrogenesis. Cell Mol Life Sci. 2014, 71, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.; Nery, S.; Rutlin, M.L.; Radtke, F.; Fishell, G.; Gaiano, N. Fibroblast Growth Factor Receptor Signaling Promotes Radial Glial Identity and Interacts with Notch1 Signaling in Telencephalic Progenitors. J Neurosci. 2004, 24, 9497–9506. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Martinowich, K.; Chin, M.H.; He, F.; Fouse, S.D.; Hutnick, L.; et al. DNA methylation controls the timing of astrogliogenesis through regulation of JAK-STAT signaling. Development. 2005, 132, 3345–3356. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Nadal-Vicens, M.; Misono, S.; Lin, M.Z.; Zubiaga, A.; Hua, X.; et al. Neurogenin Promotes Neurogenesis and Inhibits Glial Differentiation by Independent Mechanisms. Cell. 2001, 104, 365–376. [Google Scholar] [CrossRef]
- Bonni, A.; Sun, Y.; Nadal-Vicens, M.; Bhatt, A.; Frank, D.A.; Rozovsky, I.; et al. Regulation of Gliogenesis in the Central Nervous System by the JAK-STAT Signaling Pathway. Science. 1997, 278, 477–483. [Google Scholar] [CrossRef]
- Nieto, M.; Schuurmans, C.; Britz, O.; Guillemot, F. Neural bHLH Genes Control the Neuronal versus Glial Fate Decision in Cortical Progenitors. Neuron. 2001, 29, 401–413. [Google Scholar] [CrossRef]
- Zhang, Y.; Pak, C.; Han, Y.; Ahlenius, H.; Zhang, Z.; Chanda, S.; et al. Rapid Single-Step Induction of Functional Neurons from Human Pluripotent Stem Cells. Neuron. 2013, 78, 785–798. [Google Scholar] [CrossRef]
- Yang, N.; Chanda, S.; Marro, S.; Ng, Y.H.; Janas, J.A.; Haag, D.; et al. Generation of pure GABAergic neurons by transcription factor programming. Nat Methods. 2017, 14, 621–628. [Google Scholar] [CrossRef]
- Vierbuchen, T.; Ostermeier, A.; Pang, Z.P.; Kokubu, Y.; Südhof, T.C.; Wernig, M. Direct conversion of fibroblasts to functional neurons by defined factors. Nature. 2010, 463, 1035–1041. [Google Scholar] [CrossRef]
- Hatakeyama, J.; Bessho, Y.; Katoh, K.; Ookawara, S.; Fujioka, M.; Guillemot, F.; et al. Hes genes regulate size, shape and histogenesis of the nervous system by control of the timing of neural stem cell differentiation. Development. 2004, 131, 5539–5550. [Google Scholar] [CrossRef] [PubMed]
- Mizutani K ichi, Saito, T. Progenitors resume generating neurons after temporary inhibition of neurogenesis by Notch activation in the mammalian cerebral cortex. Development. 2005, 132, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Hoch, R.V.; Rubenstein, J.L.R.; Pleasure, S. Genes and signaling events that establish regional patterning of the mammalian forebrain. Semin Cell Dev Biol. 2009, 20, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Leone, D.P.; Srinivasan, K.; Chen, B.; Alcamo, E.; McConnell, S.K. The determination of projection neuron identity in the developing cerebral cortex. Curr Opin Neurobiol. 2008, 18, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, F. Cellular and molecular control of neurogenesis in the mammalian telencephalon. Curr Opin Cell Biol. 2005, 17, 639–647. [Google Scholar] [CrossRef]
- He, F.; Ge, W.; Martinowich, K.; Becker-Catania, S.; Coskun, V.; Zhu, W.; et al. A positive autoregulatory loop of Jak-STAT signaling controls the onset of astrogliogenesis. Nat Neurosci. 2005, 8, 616–625. [Google Scholar] [CrossRef]
- Tomita, K. Mammalian achaete-scute and atonal homologs regulate neuronal versus glial fate determination in the central nervous system. EMBO J. 2000, 19, 5460–5472. [Google Scholar] [CrossRef]
- Llorca, A.; Deogracias, R. Origin, Development, and Synaptogenesis of Cortical Interneurons. Front Neurosci. 2022, 16, 929469. [Google Scholar] [CrossRef]
- Styr, B.; Slutsky, I. Imbalance between firing homeostasis and synaptic plasticity drives early-phase Alzheimer’s disease. Nat Neurosci. 2018, 21, 463–473. [Google Scholar] [CrossRef]
- Rubenstein, J.L.R.; Merzenich, M.M. Model of autism: increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef]
- Foss-Feig, J.H.; Adkinson, B.D.; Ji, J.L.; Yang, G.; Srihari, V.H.; McPartland, J.C.; et al. Searching for Cross-Diagnostic Convergence: Neural Mechanisms Governing Excitation and Inhibition Balance in Schizophrenia and Autism Spectrum Disorders. Biol Psychiatry. 2017, 81, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Barnabé-Heider, F.; Wasylnka, J.A.; Fernandes, K.J.L.; Porsche, C.; Sendtner, M.; Kaplan, D.R.; et al. Evidence that Embryonic Neurons Regulate the Onset of Cortical Gliogenesis via Cardiotrophin-1. Neuron. 2005, 48, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Farmer, W.T.; Abrahamsson, T.; Chierzi, S.; Lui, C.; Zaelzer, C.; Jones, E.V.; et al. Neurons diversify astrocytes in the adult brain through sonic hedgehog signaling. Science. 2016, 351, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Voss, A.J.; Lanjewar, S.N.; Sampson, M.M.; King, A.; Hill, E.J.; Sing, A.; et al. Identification of ligand-receptor pairs that drive human astrocyte development. Nat Neurosci. 2023, 26, 1339–1351. [Google Scholar] [CrossRef]
- Bayraktar, O.A.; Bartels, T.; Holmqvist, S.; Kleshchevnikov, V.; Martirosyan, A.; Polioudakis, D.; et al. Astrocyte layers in the mammalian cerebral cortex revealed by a single-cell in situ transcriptomic map. Nat Neurosci. 2020, 23, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Berardi, N.; Pizzorusso, T.; Maffei, L. Critical periods during sensory development. Curr Opin Neurobiol. 2000, 10, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Chugani, H.T. A Critical Period of Brain Development: Studies of Cerebral Glucose Utilization with PET. Prev Med. 1998, 27, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Sengpiel, F. The critical period. Curr Biol. 2007, 17, R742–3. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, E.I. Sensitive Periods in the Development of the Brain and Behavior. J Cogn Neurosci. 2004, 16, 1412–1425. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Gabard-Durnam, L.J. Early Adversity and Critical Periods: Neurodevelopmental Consequences of Violating the Expectable Environment. Trends Neurosci. 2020, 43, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Virolainen, S.J.; VonHandorf, A.; Viel KCMF, Weirauch, M. T.; Kottyan, L.C. Gene–environment interactions and their impact on human health. Genes Immun. 2022, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Milbocker, K.A.; Campbell, T.S.; Collins, N.; Kim, S.; Smith, I.F.; Roth, T.L.; et al. Glia-Driven Brain Circuit Refinement Is Altered by Early-Life Adversity: Behavioral Outcomes. Front Behav Neurosci. 2021 Dec 2;15:786234. [CrossRef]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.A.; Mullowney, C.E.; Hell, J.W.; Agah, A.; et al. Thrombospondins Are Astrocyte-Secreted Proteins that Promote CNS Synaptogenesis. Cell. 2005, 120, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Pfrieger, F.W.; Barres, B.A. Synaptic Efficacy Enhanced by Glial Cells in Vitro. Science. 1997, 277, 1684–1687. [Google Scholar] [CrossRef] [PubMed]
- Mauch, D.H.; Nägler, K.; Schumacher, S.; Göritz, C.; Müller, E.C.; Otto, A.; et al. CNS Synaptogenesis Promoted by Glia-Derived Cholesterol. Science. 2001, 294, 1354–1357. [Google Scholar] [CrossRef]
- Eroglu, Ç.; Allen, N.J.; Susman, M.W.; O’Rourke, N.A.; Park, C.Y.; Özkan, E.; et al. Gabapentin Receptor α2δ-1 Is a Neuronal Thrombospondin Receptor Responsible for Excitatory CNS Synaptogenesis. Cell. 2009, 139, 380–392. [Google Scholar] [CrossRef]
- Fossati, G.; Pozzi, D.; Canzi, A.; Mirabella, F.; Valentino, S.; Morini, R.; et al. Pentraxin 3 regulates synaptic function by inducing AMPA receptor clustering via ECM remodeling and β1-integrin. EMBO J. 2019, 38, e99529. [Google Scholar] [CrossRef]
- Allen, N.J.; Bennett, M.L.; Foo, L.C.; Wang, G.X.; Chakraborty, C.; Smith, S.J.; et al. Astrocyte glypicans 4 and 6 promote formation of excitatory synapses via GluA1 AMPA receptors. Nature. 2012, 486, 410–414. [Google Scholar] [CrossRef]
- Diniz, L.P.; Almeida, J.C.; Tortelli, V.; Vargas Lopes, C.; Setti-Perdigão, P.; Stipursky, J.; et al. Astrocyte-induced Synaptogenesis Is Mediated by Transforming Growth Factor β Signaling through Modulation of d-Serine Levels in Cerebral Cortex Neurons. J Biol Chem. 2012, 287, 41432–41445. [Google Scholar] [CrossRef] [PubMed]
- Diniz, L.P.; Tortelli, V.; Garcia, M.N.; Araújo, A.P.B.; Melo, H.M.; Seixas Da Silva, G.S.; et al. Astrocyte transforming growth factor beta 1 promotes inhibitory synapse formation via CaM kinase II signaling. Glia. 2014, 62, 1917–1931. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Casati, M.E.; Murtie, J.C.; Rio, C.; Stankovic, K.; Liberman, M.C.; Corfas, G. Nonneuronal cells regulate synapse formation in the vestibular sensory epithelium via erbB-dependent BDNF expression. Proc Natl Acad Sci. 2010, 107, 17005–17010. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Zhang, T.; Fan, K.; Cai, W.; Liu, H. Astrocyte-Neuron Signaling in Synaptogenesis. Front Cell Dev Biol. 2021, 9, 680301. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.X.; Burrus Lane, C.J.; Eroglu, C. Role of astrocytes in synapse formation and maturation. In: Current Topics in Developmental Biology [Internet]. Elsevier; 2021 [cited 2024 Jan 20]. p. 371–407. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0070215320301435.
- Konishi, H.; Koizumi, S.; Kiyama, H. Phagocytic astrocytes: Emerging from the shadows of microglia. Glia. 2022, 70, 1009–1026. [Google Scholar] [CrossRef] [PubMed]
- Juraska, J.M.; Willing, J. Pubertal onset as a critical transition for neural development and cognition. Brain Res. 2017, 1654, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Huttenlocher, P.R.; Dabholkar, A.S. Regional differences in synaptogenesis in human cerebral cortex. J Comp Neurol. 1997, 387, 167–178. [Google Scholar] [CrossRef]
- Peter, R. H. Synaptic density in human frontal cortex — Developmental changes and effects of aging. Brain Res. 1979, 163, 195–205. [Google Scholar] [CrossRef]
- Dosenbach, N.U.F.; Nardos, B.; Cohen, A.L.; Fair, D.A.; Power, J.D.; Church, J.A.; et al. Prediction of Individual Brain Maturity Using fMRI. Science. 2010, 329, 1358–1361. [Google Scholar] [CrossRef]
- Petanjek, Z.; Judaš, M.; Šimić, G.; Rašin, M.R.; Uylings, H.B.M.; Rakic, P.; et al. Extraordinary neoteny of synaptic spines in the human prefrontal cortex. Proc Natl Acad Sci. 2011, 108, 13281–13286. [Google Scholar] [CrossRef]
- Antoine, M.W.; Langberg, T.; Schnepel, P.; Feldman, D.E. Increased Excitation-Inhibition Ratio Stabilizes Synapse and Circuit Excitability in Four Autism Mouse Models. Neuron. 2019, 101, 648–661. [Google Scholar] [CrossRef] [PubMed]
- Aida, T.; Yoshida, J.; Nomura, M.; Tanimura, A.; Iino, Y.; Soma, M.; et al. Astroglial Glutamate Transporter Deficiency Increases Synaptic Excitability and Leads to Pathological Repetitive Behaviors in Mice. Neuropsychopharmacology. 2015, 40, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Ortinski, P.I.; Dong, J.; Mungenast, A.; Yue, C.; Takano, H.; Watson, D.J.; et al. Selective induction of astrocytic gliosis generates deficits in neuronal inhibition. Nat Neurosci. 2010, 13, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Eltokhi, A.; Janmaat, I.E.; Genedi, M.; Haarman, B.C.M.; Sommer, I.E.C. Dysregulation of synaptic pruning as a possible link between intestinal microbiota dysbiosis and neuropsychiatric disorders. J Neurosci Res. 2020, 98, 1335–1369. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, P.L.; De Lima, I.B.Q.; Maciel, E.M.A.; Silva, N.C.; Dobransky, T.; Ribeiro, F.M. Synaptic Elimination in Neurological Disorders. Curr Neuropharmacol. 2019, 17, 1071–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Xu, X.; Li, H.; Niu, F.; Yang, M.; Ge, Q.; et al. Megf10-related engulfment of excitatory postsynapses by astrocytes following severe brain injury. CNS Neurosci Ther. 2023 Apr 20;cns.14223. [CrossRef]
- Iram, T.; Ramirez-Ortiz, Z.; Byrne, M.H.; Coleman, U.A.; Kingery, N.D.; Means, T.K.; et al. Megf10 Is a Receptor for C1Q That Mediates Clearance of Apoptotic Cells by Astrocytes. J Neurosci. 2016, 36, 5185–5192. [Google Scholar] [CrossRef]
- Pattwell, S.S.; Liston, C.; Jing, D.; Ninan, I.; Yang, R.R.; Witztum, J.; et al. Dynamic changes in neural circuitry during adolescence are associated with persistent attenuation of fear memories. Nat Commun. 2016, 7, 11475. [Google Scholar] [CrossRef]
- Honeycutt, J.A.; Demaestri, C.; Peterzell, S.; Silveri, M.M.; Cai, X.; Kulkarni, P.; et al. Altered corticolimbic connectivity reveals sex-specific adolescent outcomes in a rat model of early life adversity. eLife. 2020, 9, e52651. [Google Scholar] [CrossRef]
- Karpova, N.N.; Pickenhagen, A.; Lindholm, J.; Tiraboschi, E.; Kulesskaya, N.; Ágústsdóttir, A.; et al. Fear Erasure in Mice Requires Synergy Between Antidepressant Drugs and Extinction Training. Science. 2011, 334, 1731–1734. [Google Scholar] [CrossRef]
- Vetencourt, J.F.M.; Sale, A.; Viegi, A.; Baroncelli, L.; De Pasquale, R.; F. O’Leary, O.; et al. The Antidepressant Fluoxetine Restores Plasticity in the Adult Visual Cortex. Science. 2008, 320, 385–388. [Google Scholar] [CrossRef]
- Ribot, J.; Breton, R.; Calvo, C.F.; Moulard, J.; Ezan, P.; Zapata, J.; et al. Astrocytes close the mouse critical period for visual plasticity. Science. 2021, 373, 77–81. [Google Scholar] [CrossRef]
- Müller, C.M.; Best, J. Ocular dominance plasticity in adult cat visual cortex after transplantation of cultured astrocytes. Nature. 1989, 342, 427–430. [Google Scholar] [CrossRef]
- Ghézali, G.; Calvo, C.F.; Pillet, L.E.; Llense, F.; Ezan, P.; Pannasch, U.; et al. Connexin 30 controls astroglial polarization during postnatal brain development. Development. 2018, 145, dev155275. [Google Scholar] [CrossRef]
- Abbink, M.R.; Deijk, A.F.; Heine, V.M.; Verheijen, M.H.; Korosi, A. The involvement of astrocytes in early-life adversity induced programming of the brain. Glia. 2019 Apr 30;glia.23625. [CrossRef]
- Codeluppi, S.A.; Chatterjee, D.; Prevot, T.D.; Bansal, Y.; Misquitta, K.A.; Sibille, E.; et al. Chronic Stress Alters Astrocyte Morphology in Mouse Prefrontal Cortex. Int J Neuropsychopharmacol. 2021, 24, 842–853. [Google Scholar] [CrossRef]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. Synaptic and behavioral effects of chronic stress are linked to dynamic and sex-specific changes in microglia function and astrocyte dystrophy. Neurobiol Stress. 2021, 14, 100312. [Google Scholar] [CrossRef] [PubMed]
- Dolotov, O.V.; Inozemtseva, L.S.; Myasoedov, N.F.; Grivennikov, I.A. Stress-Induced Depression and Alzheimer’s Disease: Focus on Astrocytes. Int J Mol Sci. 2022, 23, 4999. [Google Scholar] [CrossRef] [PubMed]
- Naskar, S.; Chattarji, S. Stress Elicits Contrasting Effects on the Structure and Number of Astrocytes in the Amygdala versus Hippocampus. eNeuro. 2019, 6, ENEURO.0338–182019. [Google Scholar] [CrossRef] [PubMed]
- Murphy-Royal, C.; Gordon, G.R.; Bains, J.S. Stress-induced structural and functional modifications of astrocytes—Further implicating glia in the central response to stress. Glia. 2019, 67, 1806–1820. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, K.; Oda, Y.; Kimura, M.; Kimura, H.; Nangaku, M.; Shirayama, Y.; et al. The alterations of glutamate transporter 1 and glutamine synthetase in the rat brain of a learned helplessness model of depression. Psychopharmacology (Berl). 2020, 237, 2547–2553. [Google Scholar] [CrossRef] [PubMed]
- Tynan, R.J.; Beynon, S.B.; Hinwood, M.; Johnson, S.J.; Nilsson, M.; Woods, J.J.; et al. Chronic stress-induced disruption of the astrocyte network is driven by structural atrophy and not loss of astrocytes. Acta Neuropathol (Berl). 2013, 126, 75–91. [Google Scholar] [CrossRef]
- Virmani, G.; D’almeida, P.; Nandi, A.; Marathe, S. Subfield-specific effects of chronic mild unpredictable stress on hippocampal astrocytes. Hughes, E.; editor. Eur J Neurosci. 2021, 54, 5730–5746. [Google Scholar] [CrossRef]
- Lu, C.L.; Ren, J.; Mo, J.W.; Fan, J.; Guo, F.; Chen, L.Y.; et al. Glucocorticoid Receptor–Dependent Astrocytes Mediate Stress Vulnerability. Biol Psychiatry. 2022, 92, 204–215. [Google Scholar] [CrossRef]
- Huang, D.; Li, C.; Zhang, W.; Qin, J.; Jiang, W.; Hu, C. Dysfunction of astrocytic connexins 30 and 43 in the medial prefrontal cortex and hippocampus mediates depressive-like behaviours. Behav Brain Res. 2019, 372, 111950. [Google Scholar] [CrossRef] [PubMed]
- Byun, Y.G.; Kim, N.S.; Kim, G.; Jeon, Y.S.; Choi, J.B.; Park, C.W.; et al. Stress induces behavioral abnormalities by increasing expression of phagocytic receptor MERTK in astrocytes to promote synapse phagocytosis. Immunity. 2023, 56, 2105–2120. [Google Scholar] [CrossRef]
- Miguel-Hidalgo, J.J.; Moulana, M.; Deloach, P.H.; Rajkowska, G. Chronic Unpredictable Stress Reduces Immunostaining for Connexins 43 and 30 and Myelin Basic Protein in the Rat Prelimbic and Orbitofrontal Cortices. Chronic Stress. 2018, 2, 247054701881418. [Google Scholar] [CrossRef]
- Kang, Y.; Kang, W.; Kim, A.; Tae, W.S.; Ham, B.J.; Han, K.M. Decreased cortical gyrification in major depressive disorder. Psychol Med. 2023, 53, 7512–7524. [Google Scholar] [CrossRef] [PubMed]
- Ning, M.; Li, C.; Gao, L.; Fan, J. Core-Symptom-Defined Cortical Gyrification Differences in Autism Spectrum Disorder. Front Psychiatry. 2021, 12, 619367. [Google Scholar] [CrossRef]
- Takayanagi, Y.; Sasabayashi, D.; Takahashi, T.; Komori, Y.; Furuichi, A.; Kido, M.; et al. Altered brain gyrification in deficit and non-deficit schizophrenia. Psychol Med. 2019, 49, 573–580. [Google Scholar] [CrossRef]
- Cao, B.; Mwangi, B.; Passos, I.C.; Wu, M.J.; Keser, Z.; Zunta-Soares, G.B.; et al. Lifespan Gyrification Trajectories of Human Brain in Healthy Individuals and Patients with Major Psychiatric Disorders. Sci Rep. 2017, 7, 511. [Google Scholar] [CrossRef] [PubMed]
- Sasabayashi, D.; Takahashi, T.; Takayanagi, Y.; Suzuki, M. Anomalous brain gyrification patterns in major psychiatric disorders: a systematic review and transdiagnostic integration. Transl Psychiatry. 2021, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Rajkowska, G.; Miguel-Hidalgo, J.J. Glial Pathology in Major Depressive Disorder: An Approach to Investigate the Coverage of Blood Vessels by Astrocyte Endfeet in Human Postmortem Brain. In: Di Benedetto, B.; editor. Astrocytes [Internet]. New York, NY: Springer New York; 2019 [cited 2023 ]. p. 247–54. (Methods in Molecular Biology; vol. 1938). Available from: http://link.springer.com/10. 9 May 1007. [Google Scholar]
- Di Benedetto, B.; Rupprecht, R. Targeting Glia Cells: Novel Perspectives for the Treatment of Neuropsychiatric Diseases. Curr Neuropharmacol. 2013, 11, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Roman, C.; Egert, L.; Di Benedetto, B. Astrocytic-neuronal crosstalk gets jammed: Alternative perspectives on the onset of neuropsychiatric disorders. Eur J Neurosci. 2021, 54, 5717–5729. [Google Scholar] [CrossRef] [PubMed]
- Nagy, C.; Suderman, M.; Yang, J.; Szyf, M.; Mechawar, N.; Ernst, C.; et al. Astrocytic abnormalities and global DNA methylation patterns in depression and suicide. Mol Psychiatry. 2015, 20, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Martins-Macedo, J.; Salgado, A.J.; Gomes, E.D.; Pinto, L. Adult brain cytogenesis in the context of mood disorders: From neurogenesis to the emergent role of gliogenesis. Neurosci Biobehav Rev. 2021, 131, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Feresten, A.H.; Barakauskas, V.; Ypsilanti, A.; Barr, A.M.; Beasley, C.L. Increased expression of glial fibrillary acidic protein in prefrontal cortex in psychotic illness. Schizophr Res. 2013, 150, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, V.V.; Svistunov, A.A.; Chubarev, V.N.; Sologova, S.S.; Mukhortova, P.; Levushkin, D.; et al. Alterations of Astrocytes in the Context of Schizophrenic Dementia. Front Pharmacol. 2020, 10, 1612. [Google Scholar] [CrossRef] [PubMed]
- Notter, T. Astrocytes in schizophrenia. Brain Neurosci Adv. 2021, 5, 239821282110091. [Google Scholar] [CrossRef]
- Vakilzadeh, G.; Martinez-Cerdeño, V. Pathology and Astrocytes in Autism. Neuropsychiatr Dis Treat. 2023, 19, 841–850. [Google Scholar] [CrossRef]
- Allen, M.; Huang, B.S.; Notaras, M.J.; Lodhi, A.; Barrio-Alonso, E.; Lituma, P.J.; et al. Astrocytes derived from ASD individuals alter behavior and destabilize neuronal activity through aberrant Ca2+ signaling. Mol Psychiatry. 2022, 27, 2470–2484. [Google Scholar] [CrossRef]
- Rajkowska, G.; Miguel-Hidalgo, J. Gliogenesis and Glial Pathology in Depression. CNS Neurol Disord - Drug Targets. 2007, 6, 219–233. [Google Scholar] [CrossRef]
- Belleau, E.L.; Treadway, M.T.; Pizzagalli, D.A. The Impact of Stress and Major Depressive Disorder on Hippocampal and Medial Prefrontal Cortex Morphology. Biol Psychiatry. 2019, 85, 443–453. [Google Scholar] [CrossRef]
- Malykhin, N.V.; Carter, R.; Seres, P.; Coupland, N.J. Structural changes in the hippocampus in major depressive disorder: contributions of disease and treatment. J Psychiatry Neurosci. 2010, 35, 337–343. [Google Scholar] [CrossRef]
- Geng, R.; Huang, X. Identification of major depressive disorder disease-related genes and functional pathways based on system dynamic changes of network connectivity. BMC Med Genomics. 2021, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Czéh, B.; Simon, M.; Schmelting, B.; Hiemke, C.; Fuchs, E. Astroglial Plasticity in the Hippocampus is Affected by Chronic Psychosocial Stress and Concomitant Fluoxetine Treatment. Neuropsychopharmacology. 2006, 31, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Czéh, B.; Di Benedetto, B. Antidepressants act directly on astrocytes: Evidences and functional consequences. Eur Neuropsychopharmacol. 2013, 23, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Czéh, B.; Nagy, S.A. Clinical Findings Documenting Cellular and Molecular Abnormalities of Glia in Depressive Disorders. Front Mol Neurosci. 2018, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Papouin, T.; Ladépêche, L.; Ruel, J.; Sacchi, S.; Labasque, M.; Hanini, M.; et al. Synaptic and Extrasynaptic NMDA Receptors Are Gated by Different Endogenous Coagonists. Cell. 2012, 150, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Henneberger, C.; Papouin, T.; Oliet, S.H.R.; Rusakov, D.A. Long-term potentiation depends on release of d-serine from astrocytes. Nature. 2010, 463, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Suarez, E.; Liu, T.F.; Kopelevich, A.; Allen, N.J. Astrocyte-Secreted Chordin-like 1 Drives Synapse Maturation and Limits Plasticity by Increasing Synaptic GluA2 AMPA Receptors. Neuron. 2018, 100, 1116–1132. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, A.L.M.; Sancho, L.; Deng, J.; Bosworth, A.; Miglietta, A.; Diedrich, J.K.; et al. Aberrant astrocyte protein secretion contributes to altered neuronal development in multiple models of neurodevelopmental disorders. Nat Neurosci. 2022, 25, 1163–1178. [Google Scholar] [CrossRef]
- Heresco-Levy, U.; Javitt, D.C.; Ebstein, R.; Vass, A.; Lichtenberg, P.; Bar, G.; et al. D-serine efficacy as add-on pharmacotherapy to risperidone and olanzapine for treatment-refractory schizophrenia. Biol Psychiatry. 2005, 57, 577–585. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Malhotra, A.K.; Cornblatt, B.; Silipo, G.; Balla, A.; Suckow, R.F.; et al. High dose D-serine in the treatment of schizophrenia. Schizophr Res. 2010, 121, 125–130. [Google Scholar] [CrossRef]
- Ma, T.M.; Abazyan, S.; Abazyan, B.; Nomura, J.; Yang, C.; Seshadri, S.; et al. Pathogenic disruption of DISC1-serine racemase binding elicits schizophrenia-like behavior via D-serine depletion. Mol Psychiatry. 2013, 18, 557–567. [Google Scholar] [CrossRef]
- Cardno, A.G.; Gottesman, I.I. Twin studies of schizophrenia: From bow-and-arrow concordances to Star Wars Mx and functional genomics. Am J Med Genet. 2000, 97, 12–17. [Google Scholar] [CrossRef]
- Walker, E.F.; Trotman, H.D.; Pearce, B.D.; Addington, J.; Cadenhead, K.S.; Cornblatt, B.A.; et al. Cortisol Levels and Risk for Psychosis: Initial Findings from the North American Prodrome Longitudinal Study. Biol Psychiatry. 2013, 74, 410–417. [Google Scholar] [CrossRef]
- Selemon, L.D.; Zecevic, N. Schizophrenia: a tale of two critical periods for prefrontal cortical development. Transl Psychiatry. 2015, 5, e623–e623. [Google Scholar] [CrossRef]
- Sheu, J.R.; Hsieh, C.Y.; Jayakumar, T.; Tseng, M.F.; Lee, H.N.; Huang, S.W.; et al. A Critical Period for the Development of Schizophrenia-Like Pathology by Aberrant Postnatal Neurogenesis. Front Neurosci. 2019, 13, 635. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Figueiredo, E.C.; Calì, C.; Petrelli, F.; Bezzi, P. Emerging evidence for astrocyte dysfunction in schizophrenia. Glia. 2022, 70, 1585–1604. [Google Scholar] [CrossRef] [PubMed]
- Russo, F.B.; Freitas, B.C.; Pignatari, G.C.; Fernandes, I.R.; Sebat, J.; Muotri, A.R.; et al. Modeling the Interplay Between Neurons and Astrocytes in Autism Using Human Induced Pluripotent Stem Cells. Biol Psychiatry. 2018, 83, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.M.; Rohn, T.T.; Oxford, J.T. Autism as the Early Closure of a Neuroplastic Critical Period Normally Seen in Adolescence. Biol Syst Open Access [Internet]. 2012 [cited 2023 Jul 23];02(03). Available from: https://www.omicsgroup.org/journals/autism-as-the-early-closure-of-a-neuroplastic-critical-period-normally-seen-in-adolescence-2329-6577-1000118.php?aid=43859.
- Hashimoto, Y.; Greene, C.; Munnich, A.; Campbell, M. The CLDN5 gene at the blood-brain barrier in health and disease. Fluids Barriers CNS. 2023, 20, 22. [Google Scholar] [CrossRef]
- Igarashi, Y.; Utsumi, H.; Chiba, H.; Yamada-Sasamori, Y.; Tobioka, H.; Kamimura, Y.; et al. Glial Cell Line-Derived Neurotrophic Factor Induces Barrier Function of Endothelial Cells Forming the Blood–Brain Barrier. Biochem Biophys Res Commun. 1999, 261, 108–112. [Google Scholar] [CrossRef]
- Rajkowska, G.; Hughes, J.; Stockmeier, C.A.; Javier Miguel-Hidalgo, J.; Maciag, D. Coverage of Blood Vessels by Astrocytic Endfeet Is Reduced in Major Depressive Disorder. Biol Psychiatry. 2013, 73, 613–621. [Google Scholar] [CrossRef]
- Lee, E.; Chung, W.S. Glial Control of Synapse Number in Healthy and Diseased Brain. Front Cell Neurosci. 2019, 13, 42. [Google Scholar] [CrossRef]
- Di Benedetto, B.; Malik, V.A.; Begum, S.; Jablonowski, L.; Gómez-González, G.B.; Neumann, I.D.; et al. Fluoxetine Requires the Endfeet Protein Aquaporin-4 to Enhance Plasticity of Astrocyte Processes. Front Cell Neurosci [Internet]. 2016 Feb 2 [cited 2022 Jan 13];10. Available from: http://journal.frontiersin.org/Article/10.3389/fncel.2016.00008/abstract.
- Malik, V.A.; Zajicek, F.; Mittmann, L.A.; Klaus, J.; Unterseer, S.; Rajkumar, S.; et al. GDF15 promotes simultaneous astrocyte remodeling and tight junction strengthening at the blood–brain barrier. J Neurosci Res. 2020, 98, 1433–1456. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.P.; Price, J. Evidence for multiple precursor cell types in the embryonic rat cerebral cortex. Neuron. 1995, 14, 1181–1188. [Google Scholar] [CrossRef]
- Chung, W.S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb Perspect Biol. 2015, 7, a020370. [Google Scholar] [CrossRef]
- Chung, W.S.; Clarke, L.E.; Wang, G.X.; Stafford, B.K.; Sher, A.; Chakraborty, C.; et al. Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature. 2013, 504, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Logan, M.A.; Freeman, M.R. The scoop on the fly brain: glial engulfment functions in Drosophila. Neuron Glia Biol. 2007, 3, 63–74. [Google Scholar] [CrossRef]
- Freeman, M.R.; Delrow, J.; Kim, J.; Johnson, E.; Doe, C.Q. Unwrapping Glial Biology. Neuron. 2003, 38, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Reddien, P.W.; Horvitz, H.R. THE ENGULFMENT PROCESS OF PROGRAMMED CELL DEATH IN CAENORHABDITIS ELEGANS. Annu Rev Cell Dev Biol. 2004, 20, 193–221. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.S.; Nouri, N.; et al. The Classical Complement Cascade Mediates CNS Synapse Elimination. Cell. 2007, 131, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Schafer, D.P.; Lehrman, E.K.; Kautzman, A.G.; Koyama, R.; Mardinly, A.R.; Yamasaki, R.; et al. Microglia Sculpt Postnatal Neural Circuits in an Activity and Complement-Dependent Manner. Neuron. 2012, 74, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; et al. Synaptic Pruning by Microglia Is Necessary for Normal Brain Development. Science. 2011, 333, 1456–1458. [Google Scholar] [CrossRef] [PubMed]
- Dejanovic, B.; Wu, T.; Tsai, M.C.; Graykowski, D.; Gandham, V.D.; Rose, C.M.; et al. Complement C1q-dependent excitatory and inhibitory synapse elimination by astrocytes and microglia in Alzheimer’s disease mouse models. Nat Aging. 2022, 2, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Favuzzi, E.; Huang, S.; Saldi, G.A.; Binan, L.; Ibrahim, L.A.; Fernández-Otero, M.; et al. GABA-receptive microglia selectively sculpt developing inhibitory circuits. Cell. 2021, 184, 4048–4063. [Google Scholar] [CrossRef]
- Park, J.; Choi, Y.; Jung, E.; Lee, S.; Sohn, J.; Chung, W. Microglial MERTK eliminates phosphatidylserine-displaying inhibitory post-synapses. EMBO J. 2021, 40, e107121. [Google Scholar] [CrossRef]
- Scott-Hewitt, N.; Perrucci, F.; Morini, R.; Erreni, M.; Mahoney, M.; Witkowska, A.; et al. Local externalization of phosphatidylserine mediates developmental synaptic pruning by microglia. EMBO J. 2020, 39, e105380. [Google Scholar] [CrossRef]
- Schmidtner, A.K.; Slattery, D.A.; Gläsner, J.; Hiergeist, A.; Gryksa, K.; Malik, V.A.; et al. Minocycline alters behavior, microglia and the gut microbiome in a trait-anxiety-dependent manner. Transl Psychiatry. 2019, 9, 223. [Google Scholar] [CrossRef]
- Cullheim, S.; Thams, S. The microglial networks of the brain and their role in neuronal network plasticity after lesion. Brain Res Rev. 2007, 55, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Datta, D.; Leslie, S.N.; Morozov, Y.M.; Duque, A.; Rakic, P.; Van Dyck, C.H.; et al. Classical complement cascade initiating C1q protein within neurons in the aged rhesus macaque dorsolateral prefrontal cortex. J Neuroinflammation. 2020, 17, 8. [Google Scholar] [CrossRef] [PubMed]
- Geloso, M.C.; D’Ambrosi, N. Microglial Pruning: Relevance for Synaptic Dysfunction in Multiple Sclerosis and Related Experimental Models. Cells. 2021, 10, 686. [Google Scholar] [CrossRef] [PubMed]
- Hammond, T.R.; Robinton, D.; Stevens, B. Microglia and the Brain: Complementary Partners in Development and Disease. Annu Rev Cell Dev Biol. 2018, 34, 523–544. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim J young, Noh, S. ; Lee, H.; Lee, S.Y.; Mun, J.Y.; et al. Astrocytes phagocytose adult hippocampal synapses for circuit homeostasis. Nature. 2021, 590, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Damisah, E.C.; Hill, R.A.; Rai, A.; Chen, F.; Rothlin, C.V.; Ghosh, S.; et al. Astrocytes and microglia play orchestrated roles and respect phagocytic territories during neuronal corpse removal in vivo. Sci Adv. 2020, 6, eaba3239. [Google Scholar] [CrossRef] [PubMed]
- Eladl, E.; Tremblay-LeMay, R.; Rastgoo, N.; Musani, R.; Chen, W.; Liu, A.; et al. Role of CD47 in Hematological Malignancies. J Hematol OncolJ Hematol Oncol. 2020, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Lehrman, E.K.; Wilton, D.K.; Litvina, E.Y.; Welsh, C.A.; Chang, S.T.; Frouin, A.; et al. CD47 Protects Synapses from Excess Microglia-Mediated Pruning during Development. Neuron. 2018, 100, 120–134. [Google Scholar] [CrossRef]
- Li, J.; Brickler, T.; Banuelos, A.; Marjon, K.; Shcherbina, A.; Banerjee, S.; et al. Overexpression of CD47 is associated with brain overgrowth and 16p11.2 deletion syndrome. Proc Natl Acad Sci. 2021, 118, e2005483118. [Google Scholar] [CrossRef]
- Chu, Y.; Jin, X.; Parada, I.; Pesic, A.; Stevens, B.; Barres, B.; et al. Enhanced synaptic connectivity and epilepsy in C1q knockout mice. Proc Natl Acad Sci. 2010, 107, 7975–7980. [Google Scholar] [CrossRef]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science. 2016, 352, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Dion-Albert, L.; Bandeira Binder, L.; Daigle, B.; Hong-Minh, A.; Lebel, M.; Menard, C. Sex differences in the blood-brain barrier: implications for mental health. Front Neuroendocrinol. 2022 Mar;100989. [CrossRef]
- Blokland, G.A.M.; Grove, J.; Chen, C.Y.; Cotsapas, C.; Tobet, S.; Handa, R.; et al. Sex-Dependent Shared and Nonshared Genetic Architecture Across Mood and Psychotic Disorders. Biol Psychiatry. 2022, 91, 102–117. [Google Scholar] [CrossRef]
- Riecher-Rössler, A. Sex and gender differences in mental disorders. Lancet Psychiatry. 2017, 4, 8–9. [Google Scholar] [CrossRef]
- Ramiro, L.; Faura, J.; Simats, A.; García-Rodríguez, P.; Ma, F.; Martín, L.; et al. Influence of sex, age and diabetes on brain transcriptome and proteome modifications following cerebral ischemia. BMC Neurosci. 2023, 24, 7. [Google Scholar] [CrossRef]
- Iturria-Medina, Y.; Adewale, Q.; Khan, A.F.; Ducharme, S.; Rosa-Neto, P.; O’Donnell, K.; et al. Unified epigenomic, transcriptomic, proteomic, and metabolomic taxonomy of Alzheimer’s disease progression and heterogeneity. Sci Adv. 2022, 8, eabo6764. [Google Scholar] [CrossRef]
- López-Cerdán, A.; Andreu, Z.; Hidalgo, M.R.; Grillo-Risco, R.; Català-Senent, J.F.; Soler-Sáez, I.; et al. Unveiling sex-based differences in Parkinson’s disease: a comprehensive meta-analysis of transcriptomic studies. Biol Sex Differ. 2022, 13, 68. [Google Scholar] [CrossRef]
- Maitra, M.; Mitsuhashi, H.; Rahimian, R.; Chawla, A.; Yang, J.; Fiori, L.M.; et al. Cell type specific transcriptomic differences in depression show similar patterns between males and females but implicate distinct cell types and genes. Nat Commun. 2023, 14, 2912. [Google Scholar] [CrossRef]
- Hyer, M.M.; Phillips, L.L.; Neigh, G.N. Sex Differences in Synaptic Plasticity: Hormones and Beyond. Front Mol Neurosci. 2018, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Rutter, M.; Caspi, A.; Moffitt, T.E. Using sex differences in psychopathology to study causal mechanisms: unifying issues and research strategies: Using sex differences in psychopathology to study causal mechanisms. J Child Psychol Psychiatry. 2003, 44, 1092–1115. [Google Scholar] [CrossRef] [PubMed]
- Ziemka-Nalecz, M.; Pawelec, P.; Ziabska, K.; Zalewska, T. Sex Differences in Brain Disorders. Int J Mol Sci. 2023, 24, 14571. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, A.N.V.; Salimi-Khorshidi, G.; Lai, M.C.; Baron-Cohen, S.; Lombardo, M.V.; Tait, R.J.; et al. A meta-analysis of sex differences in human brain structure. Neurosci Biobehav Rev. 2014, 39, 34–50. [Google Scholar] [CrossRef]
- Knickmeyer, R.C.; Styner, M.; Short, S.J.; Lubach, G.R.; Kang, C.; Hamer, R.; et al. Maturational Trajectories of Cortical Brain Development through the Pubertal Transition: Unique Species and Sex Differences in the Monkey Revealed through Structural Magnetic Resonance Imaging. Cereb Cortex. 2010, 20, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Dehorter, N.; Del Pino, I. Shifting Developmental Trajectories During Critical Periods of Brain Formation. Front Cell Neurosci. 2020, 14, 283. [Google Scholar] [CrossRef] [PubMed]
- Rurak, G.M.; Simard, S.; Freitas-Andrade, M.; Lacoste, B.; Charih, F.; Van Geel, A.; et al. Sex differences in developmental patterns of neocortical astroglia: A mouse translatome database. Cell Rep. 2022, 38, 110310. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, J.; Herbison, A.E. Hypothalamic control of the male neonatal testosterone surge. Philos Trans R Soc Lond B Biol Sci. 2016, 371, 20150115. [Google Scholar] [CrossRef]
- Acaz-Fonseca, E.; Avila-Rodriguez, M.; Garcia-Segura, L.M.; Barreto, G.E. Regulation of astroglia by gonadal steroid hormones under physiological and pathological conditions. Prog Neurobiol. 2016, 144, 5–26. [Google Scholar] [CrossRef]
- Rurak, G.M.; Woodside, B.; Aguilar-Valles, A.; Salmaso, N. Astroglial cells as neuroendocrine targets in forebrain development: Implications for sex differences in psychiatric disease. Front Neuroendocrinol. 2021, 60, 100897. [Google Scholar] [CrossRef]
Figure 1.
The critical period for astrogenesis and synaptic circuit formation during postnatal brain development in rodents and humans. The timelimes show the major processes occurring from pre- and postnatal early stages through adulthood in rodents and humans to develop properly functional astrocyic and neuronal synaptic networks. A few examples of intracellular signalling molecules which determine the switch between neurogenic to gliogenic cell fates are depicted, as well as selected proteins relevant for astrocyte-specific differentiation/maturation processes (e.g. Cx30 and TGFβ/NLGN1/TLSP/DKK1/BMP4 ligands). We refer the reader to the text for details regarding experimental evidences supporting our current knowledge about each event displayed in the Figure. Cx30, connexin 30. Figure created with BioRender.com.
Figure 1.
The critical period for astrogenesis and synaptic circuit formation during postnatal brain development in rodents and humans. The timelimes show the major processes occurring from pre- and postnatal early stages through adulthood in rodents and humans to develop properly functional astrocyic and neuronal synaptic networks. A few examples of intracellular signalling molecules which determine the switch between neurogenic to gliogenic cell fates are depicted, as well as selected proteins relevant for astrocyte-specific differentiation/maturation processes (e.g. Cx30 and TGFβ/NLGN1/TLSP/DKK1/BMP4 ligands). We refer the reader to the text for details regarding experimental evidences supporting our current knowledge about each event displayed in the Figure. Cx30, connexin 30. Figure created with BioRender.com.
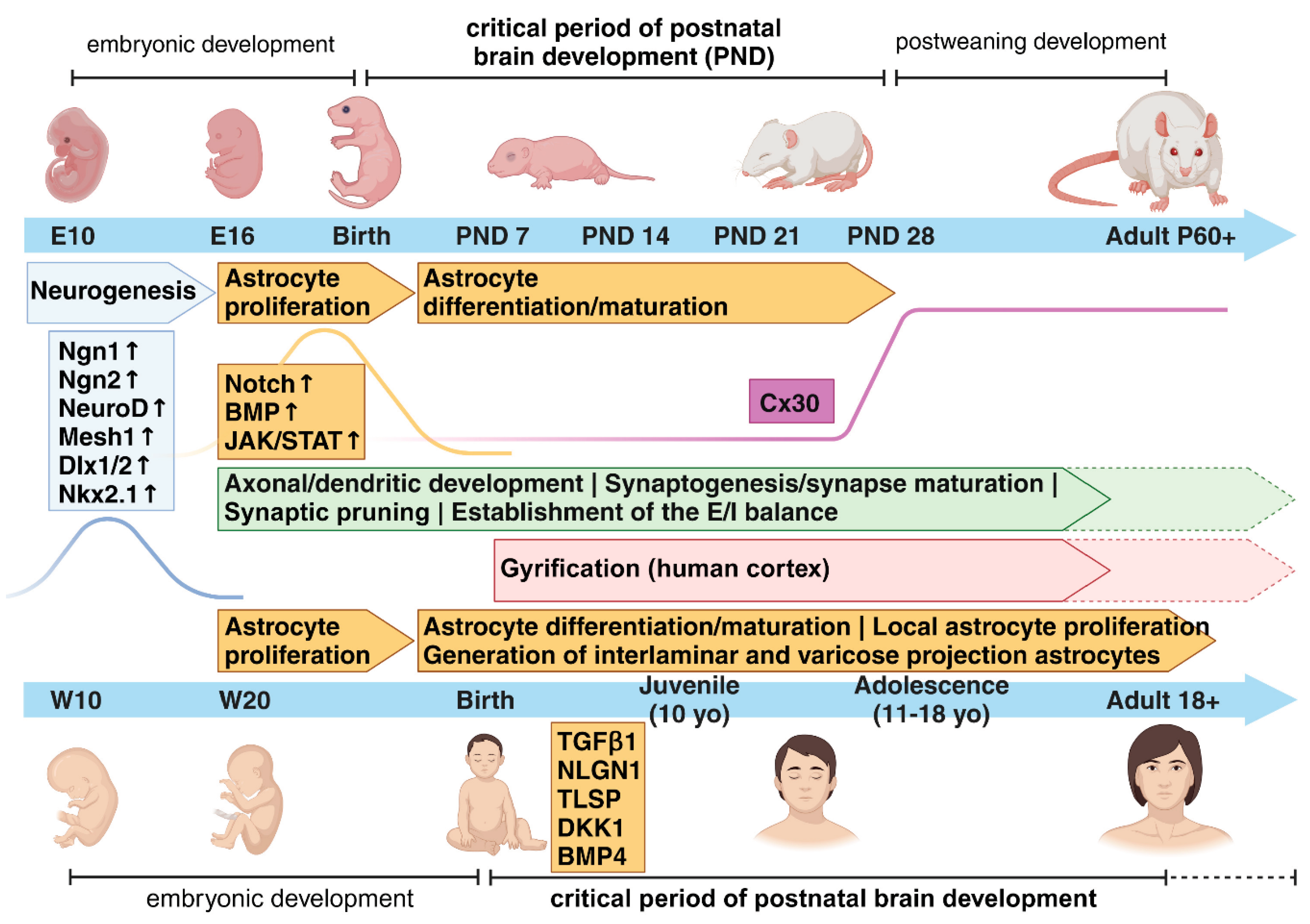
Figure 2.
The role of critical period for the onset of neurodevelopmental and neuropsychiatric disorders. Intrinsic molecular programmes and environmental factors may interact with each other during the critical period of brain development to drive the proper formation of neuronal circuits. Any alterations in the sequences of events occurring during these time frames might lead to the onset of neurodevelopmental (ND) disorders such as Autism Spectrum Disorder (ASD) or neuropsychiatric (NP) disorders such as Schizophrenia (SCZ) or Major depressive disorder (MDD). Stressful triggers, such as early-life adversity (ELA), chronic social defeat stress or early social isolation, experienced during the critical period show a higher impact on the development of ND and NP disorders later in life. Molecules like Cx30 may play a role in the closure of the critical period of astrocyte development, which is necessary for the proper formation of brain circuits. Figure created with BioRender.com.
Figure 2.
The role of critical period for the onset of neurodevelopmental and neuropsychiatric disorders. Intrinsic molecular programmes and environmental factors may interact with each other during the critical period of brain development to drive the proper formation of neuronal circuits. Any alterations in the sequences of events occurring during these time frames might lead to the onset of neurodevelopmental (ND) disorders such as Autism Spectrum Disorder (ASD) or neuropsychiatric (NP) disorders such as Schizophrenia (SCZ) or Major depressive disorder (MDD). Stressful triggers, such as early-life adversity (ELA), chronic social defeat stress or early social isolation, experienced during the critical period show a higher impact on the development of ND and NP disorders later in life. Molecules like Cx30 may play a role in the closure of the critical period of astrocyte development, which is necessary for the proper formation of brain circuits. Figure created with BioRender.com.
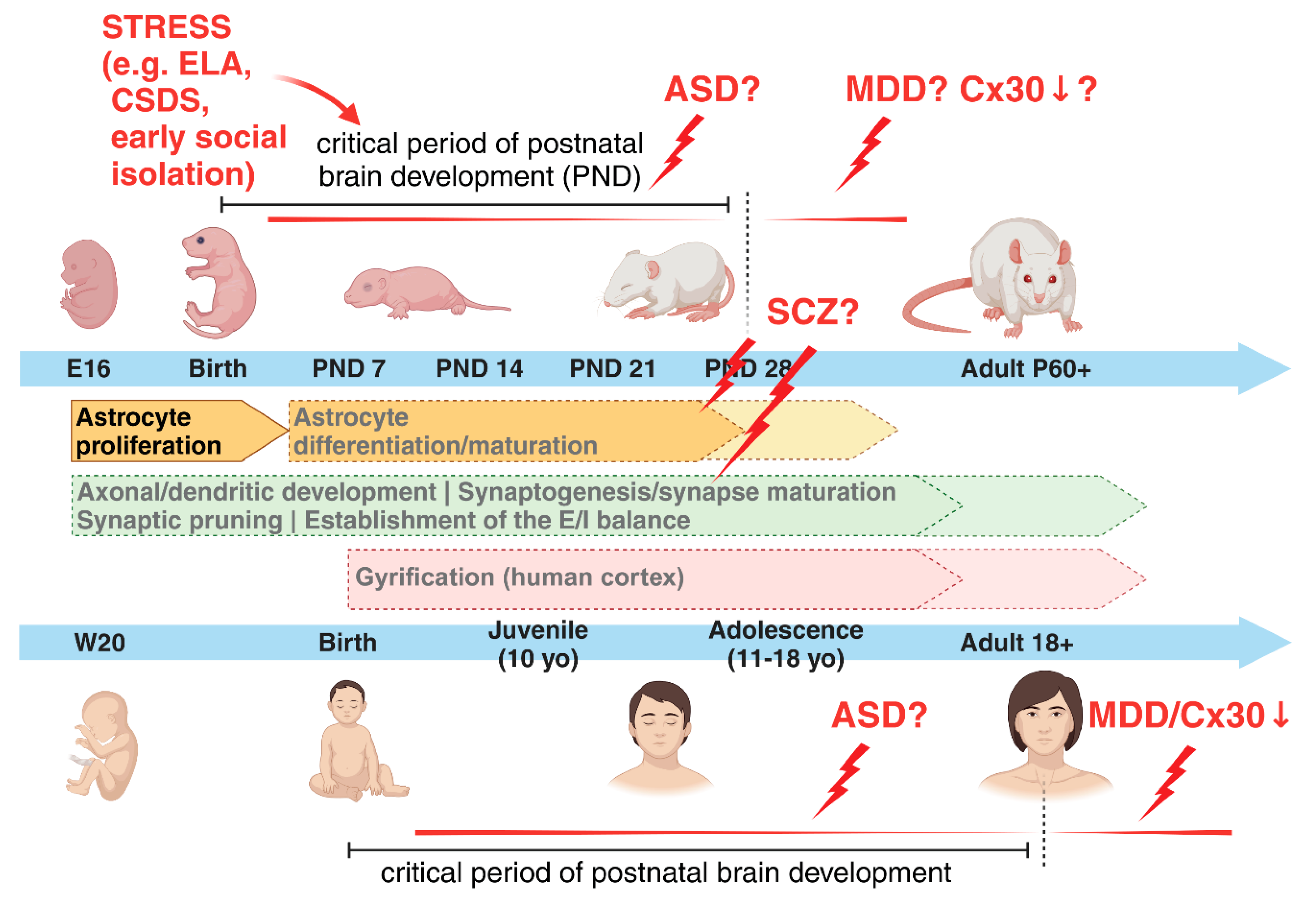
Figure 3.
Glia-mediated synaptic pruning. Astrocytes and microglia cells both regulate the refinement of synaptic neuronal networks through the elimination of weak synapses during early postnatal developmental stages. Signalling molecules called “eat me” signals, such as C1q and phosphatidylserine (PS), recruited during these events have been identified, as well as binding partners located on the respective cells, like MEGF10 receptor on astrocytes and the receptors of the complement cascade on microglia cells. It remains unclear whether PS and C1q are directly implicated in the astrocyte-mediated synaptic elimination as well. Figure created with BioRender.com.
Figure 3.
Glia-mediated synaptic pruning. Astrocytes and microglia cells both regulate the refinement of synaptic neuronal networks through the elimination of weak synapses during early postnatal developmental stages. Signalling molecules called “eat me” signals, such as C1q and phosphatidylserine (PS), recruited during these events have been identified, as well as binding partners located on the respective cells, like MEGF10 receptor on astrocytes and the receptors of the complement cascade on microglia cells. It remains unclear whether PS and C1q are directly implicated in the astrocyte-mediated synaptic elimination as well. Figure created with BioRender.com.
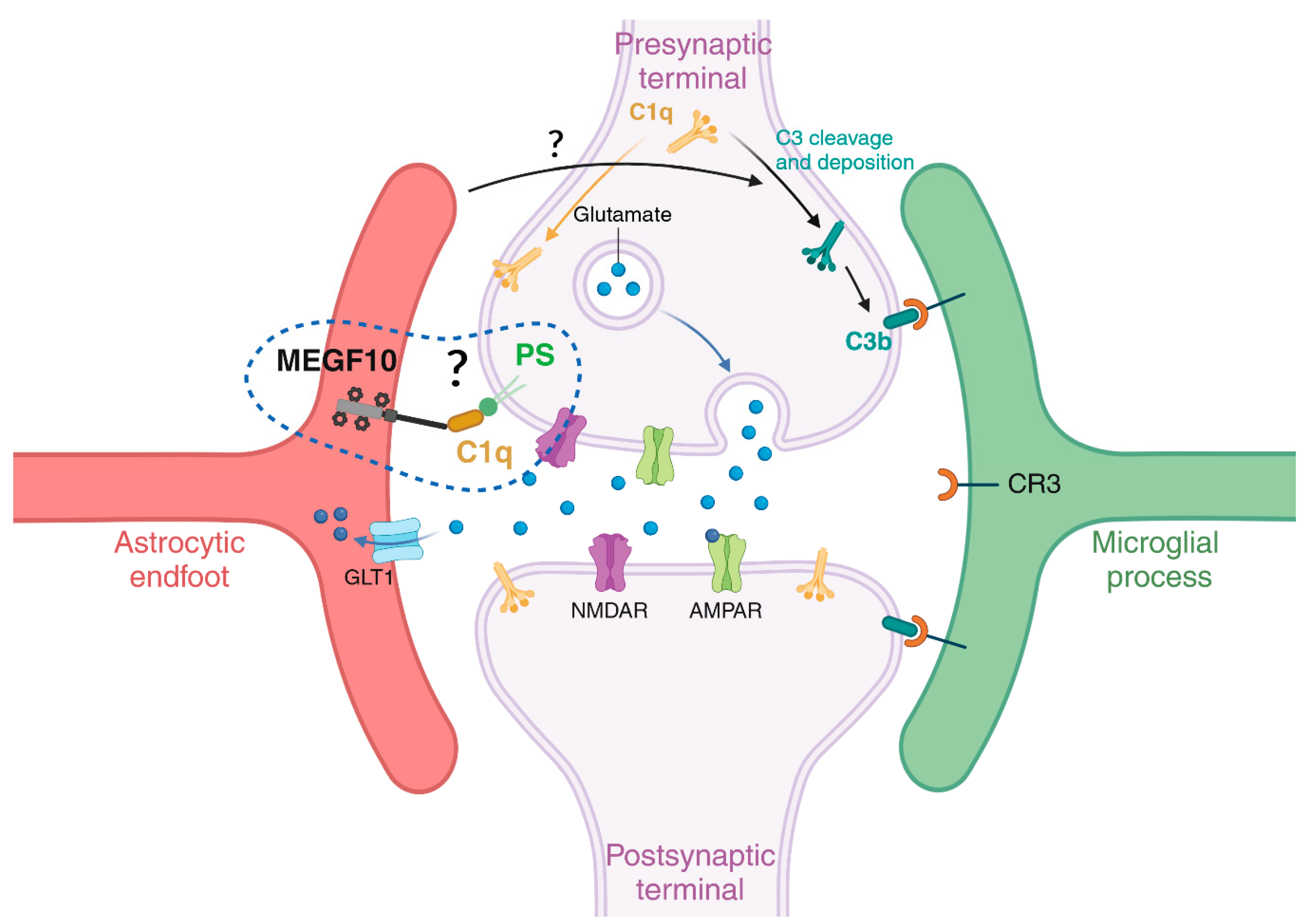
Figure 4.
Sex-dependent differences in astrogenesis and vulnerability to disease onset. A sex-dependent predisposition to brain disorders has been long recognized for many brain pathologies. Differential time-shifted maturation trajectories observed in astrocytes and astrocyte-mediated processes between males and females may account for such sex-dependent biased windows of vulnerability to disease onset. Figure created with BioRender.com.
Figure 4.
Sex-dependent differences in astrogenesis and vulnerability to disease onset. A sex-dependent predisposition to brain disorders has been long recognized for many brain pathologies. Differential time-shifted maturation trajectories observed in astrocytes and astrocyte-mediated processes between males and females may account for such sex-dependent biased windows of vulnerability to disease onset. Figure created with BioRender.com.
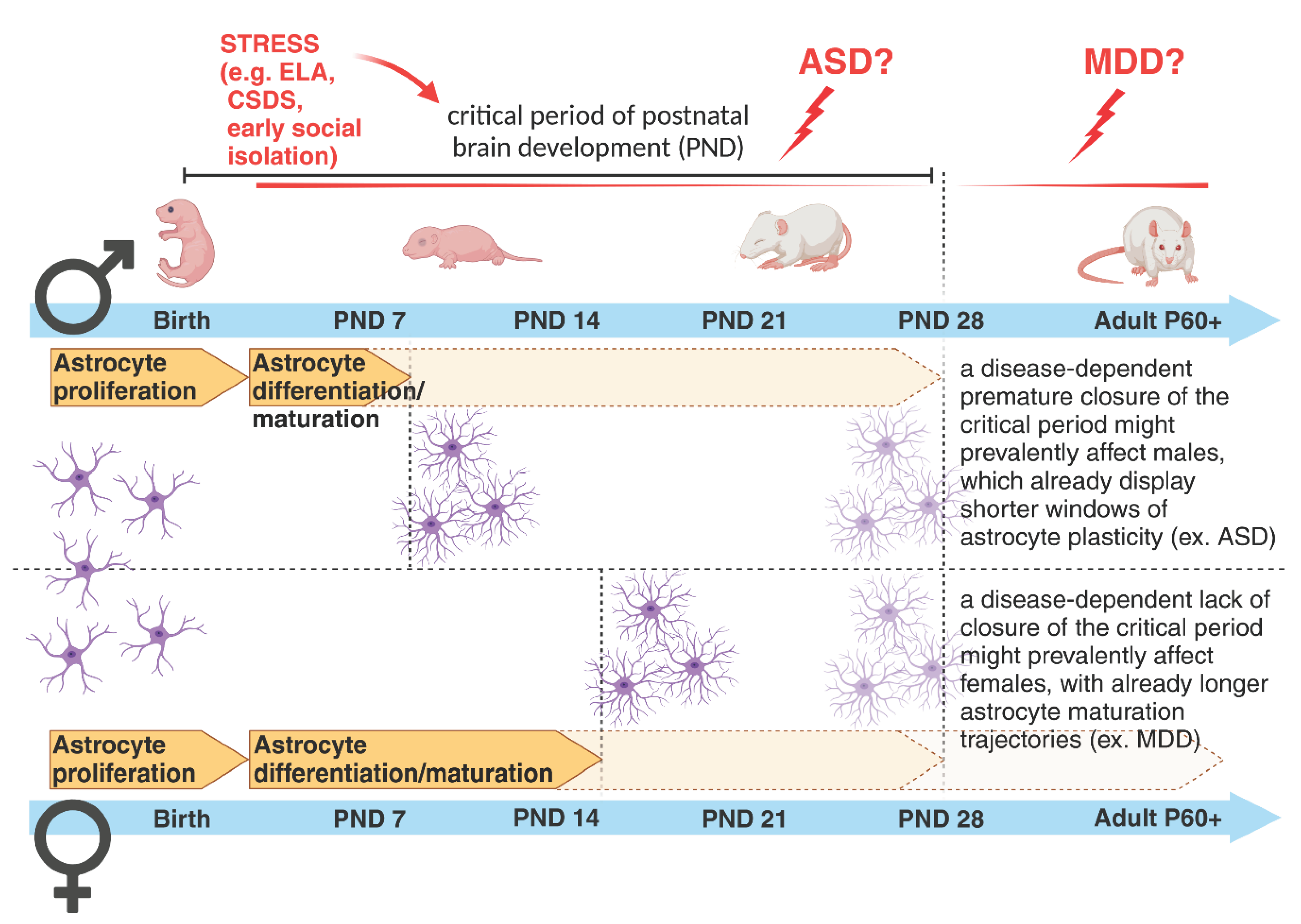
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



