
Submitted:
01 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
Keywords: Agrobacterium 1; Xenorhabdus 2; NR-AMP 3 T-DNA keyword 4; TI-plasmid 5; Intact/Cured/T-DNA Deleted 6 Sensitive/Resistant 7; EMA_PF2 8; HPLC 9
Keywords:
Agrobacterium
; Xenorhabdus
; NR-AMP
; T-DNA keyword
; TI-plasmid
; Intact/Cured/T-DNA Deleted
; Sensitive/Resistant
; EMA_PF2
; HPLC
1. Introduction
The emergence of antibiotic multi-resistance (MDR) in pathogenic bacteria has become alarming in recent decades, all over the world. Antibiotic poly-resistance (multidrug-, extreme-, and pan-drug resistance) is governed by adaptive evolution, [1] MDR has been invoking an enormous public concern, first of all, because of from humans human-clinical aspects. MGR strains of those bacterium species that have been put in the ESKAPE list [2], including (Enterococcus faecium, [3,4,5]; Staphylococcus aureus, (MRSA) [6,7,8]; Klebsiella pneumoniae [9,10]; Acinetobacter baumannii, [11,12], Pseudomonas aeruginosa, [13,14,15,16,17], and Enterobacter [2] species cannot be controlled by chemotherapy. From a veterinary aspect, the situation is also critical. A number of pathogenic bacterium species means serious challenge for veterinarians [18] (Gebreyes and Thakur, 2005),[19] Endimiani et al., 2011), [20] (Szmolka & Nagy, 2013), [21] (McManus al, 2015), [22] (Rzewuska et al., 2015), [23] (Marques et al., 2016). Furthermore, no antibiotics that are used in human clinical practice are permitted to be applied as veterinary drugs, but pets are frequent sources of human infections. In plant pathology, similar problems have been appearing, [24] Fodor et al., 2012). [25] (Załuga et al., 2014), [26] (Li et al.,), 2015). To overcome extended-spectrum beta-lactamase (ESBL)–caused resistance problems ˙[27] (Pitout, 2008), carbapenem antibiotics [28] (Papp-Wallace et al., 2011) were developed, but carbapenem-resistant (CRE) Enterobacteriaceae [29] (Temkin et al., 2014), and Klebsiella [30] (Gupta et al., 2011). appeared soon. Later, the rediscovered and rehabilitated colistin was considered a final trump [31] (Kádár et al., 2013) until colistin resistance was found in Gram-negative bacterium species, [32] (Otter et al., 2017). Antibiotics are also used in plant medicine [33] (Mc Manus et al., 2002; [34] Stockwell, [36] Sundin and Jones, 2002; [35] Aćimović et al., 2015), but the increasing number of streptomycin-resistant Erwinia amylovora isolates has been causing serious problems both in the USA [36] (Förster et al., 2015) and in Europe [37] (Gusberti et al., 2015). Environmentally friendly plant, - veterinary, - and human antibiotics of novel modes of action are imperatively needed. Antimicrobial peptides (AMP) have been hoped to provide perspectives. AMPs have been found in practically each of the known prokaryotic and eukaryotic organisms, (but no Archea) [38] (Jenssen, 2006; [39] Ötvös and Wade, 2014; [40] Mojsoska & Jenssen, 2015). produces them. AMPs are mostly of broad target spectra and strong antibiotic activity. The patented AMPs have been listed, [41], (Kosikowska and, Lesner 2016), [42] (Fosgerau and Hoffmann 2014), [43] (Sharma et al., 2023), and there is already a number of peptide-based drugs (>100 drugs in clinic). The options of finding novel natural have recently been revolutionized by tools provided by bioinformatics, allowing curation and comparative analysis of genomic and bioinformatics metabolic data of potential antibiotic-producing organisms [44] (Vallenet et al., 2013); especially since the discovery of the “On-Demand Production” of bioactive natural products, [45] (Bode et al., 2015). The symbiotic bacterial partners of the entomopathogenic nematode/bacteria (EPN/EPB) associations (Steinernema / Xenorhabdus and Heterorhabditis / Photorhabdus) produce anti-microbial [46](Akhurst, 1982; [47] Forst & Nealson, 1996) mainly AMPs [48] (Vivas & Goodrich-Blair,2001), [49] (Bode, 2009). The natural role of these antimicrobial compounds is to provide pathobiom conditions [50] (Ogier et al., 2020). or the respective EPN/ EPB) complex in polyxenic (insect cadaver; soil) environmental conditions. All known EPB-produced AMP compounds are non-ribosomal peptides (NRP), which means that they are synthesized enzymatically by multi-enzyme thiotemplate mechanisms using non-ribosomal peptide synthetases (NRPS), fatty acid synthase (FAS)-related polyketide synthases (PKS), or a hybrid biosynthesis thereof [51] (Reimer & Bode 2014). Some recently discovered AMPs are xenocoumacins [52] (Park et al., 2009); a novel new lysine-rich cyclolipopeptide family [53] (Gualtieri et al., 2009) from Xenorhabdus nematophila; and the cabanilasin from X. cabanillasii [54] (Houard et al., 2013).
Our main contributions to the field were the discovery and some pioneer studies on two EPB species, X. budapestensis, and X. szentirmaii [55]´ (Lengyel et al., 2005); [56], (Furgani et al., 2008), [57] Böszörményi et al., 2009; have recently been reviewed, [58] (Fodor et al., 2023). (The respected French team sequenced our EMC strain (protecting our “copyright” to our strains in such a way,˙[59] (Gualtieri et al., 2014). Our data concerning the antimicrobial potential of EMA and EMC [56] Furgani et al, 2008; [57] Böszörményi et al, 2009) were carefully re-evaluated and the research was further extended by Bode and his associates, who discovered several cationic AMPs in different Xenorhabdus species, sequenced EMA, but they did not publish, however, they let the sequence information publicly available. They discovered that the most powerful antimicrobial non-ribosomal peptide (NRP) compound produced by our EMA and EMC strains is the fabclavine, [60,61,62,63].
Meanwhile we ourselves went on to plant the direction of plant pathogenic applications [64,65,66,67] (Vozik et al., 2015). The most active antimicrobial component (the fabclavine) produced by X. budapestensis (EMA) has been discovered. EMA CFCM is condemned as being generally toxic, there are some we found resistant bacterium species and organisms are resistant, indicating that the EMA AMPs are not overall (“sulfuric-acid-like”) poisons and, therefore usable for resistance studies related to peptide-type antimicrobials in nature. At this point, we have become focused on the resistance/sensitivity problems rather than on the options of the immediate application.
The traditional resistance studies are based on working with a single molecule with antibacterial, (antifungal, anti-oomycete, or anti-protist) potential, and with a single (bacterium, fungus, oomycete, or protist) species that has sensitive and resistant variants (mutants), and the task is to discover a molecular mechanism of resistance. However, the defense mechanisms of EPB species providing a safe pathobiom condition in a polyxenic (soil, cadaver) milieu, [50] (Ogiert, 2020) lets us suppose that the existence of species resistant to CFCM of EMA and EMC may be an indirect evidence of the existence of complex defense mechanisms competitor organisms as well. If we managed to find sensitive variants within that competitor species we would have a chance to reveal the details of that complex resistance mechanism presenting the wild-type. When bio-assayed the CFCM, and the ambelite-adsorbed, and methanol-eluted peptide fraction (PF) on different plant-pathogenic bacteria [63], we found that the Agrobacterium tumefaciens bacterium species fulfills this criterion. We decided to try to benefit from the sophisticated genetic toolkit established by fellow researchers on A. tumefaciens as the number one tool of molecular plant biotechnology, (recently reviewed by [68] Nester, 2015).
Although this ambitious goal has only partly been achieved and we are only moderately satisfied, we gathered quite a lot of information about the profile of antimicrobials active against plant pathogen bacteria of the EMA CFCM peptides, what we feel necessary to share with the scientific community especially readers of Applied Microbiology.
This study aims to contribute the developing developing an amenable experimental system for studying resistance mechanisms toward natural individual and complex antimicrobial peptides in the future. For those fellow scientists who may not be quite familiar with the Agrobacterium system, let us summarize its advantages: A. tumefaciens DNA consists of the indispensable genome DNA or bacterial chromosome, (either C58 [69] (Wood et al., 2001); or Ach) [70] Henkel et al., 2014); and the dispensable plasmon DNA including a large circular tumor-inducing (Ti) [71] (Van Larebeke et al., 1974; [72] Currier & Nester,1976) , [73] (Shell and Montague, 1977), [74] (Hooykass, 2023) plasmid responsible for virulence and tumor-induction in infected plants. Most but not all plasmid genes are expressed in the bacterium living as vegetative in the rhizosphere. The vir genes, which are responsible for virulence, are inducible by chemicals (of phenolic, - and sugar compounds) released from wounded plant tissues through the virA-gene encoded membrane histidine kinase receptor. VirA protein then phosphorylates the transcription activator VirG, which binds to vir-box sequences, located in the promoter regions of vir genes (Koncz, personal communication).
The genes encoding for enzymes synthesizing tumor-specific compounds (including opines) are located in the transfer (T-DNA) region that is being inserted into the plant chromosomes, [75] (Chilton et al., 1977), and have all signals necessary for expression in plants during crown-gall tumor formation [76] (Koncz et al; 1983). The T-DNA located opine-synthase genes are responsible for the synthesis of respective (nopaline, - octopine, or agrpine –type) opines characteristic for a given Agrobacterium strain; while enzymes catabolizing (only the respective) opine are located outside of the T-DNA region. Agrobacterium strains are scored as nopaline (NOP), octopine (OCT), and agropine, as well as L, L,-succinamopine (AGR) opine-catabolizing ones [77] (Montoya et al., 1977; [78] (Guyon et al., 1980). [79] (Tremblay et al., 1987). (For more details, see Supplementary material (Suppl. Text 1; Table S1).
A given sensitivity/resistance (S/R) phenotype could be a consequence of more than one mechanism. Genes responsible for S/R phenotypes to EMA_PF2 may be located either on the chromosome; on the Ti plasmid; or on the second large cryptic plasmid, (in the case) of C58 strains, on pAtC58). If S/R phenotypes to EMA_PF2 were plasmid-related, genetic studies could be carried out by complementation analysis in Agrobacterium, [80] (Hoekema, 1983). A toolkit for genetic analysis may also include comparisons of S/R phenotypes of strains with different genetic backgrounds; such as of different opine types and of plasmid state. For the latter, wild-type, plasmid-cured, and helper-plasmid harboring strains producing/catabolizing the same opine are worthwhile to compare.
The mutant hunt and mutation analysis of candidate sequences is another way of genetic analysis and reproducible methodology has also been available in Agrobacterium [81], (Koekman et al., 1979; [82] Klapwijk & Schilperoort, 1979; [83] Ooms et al., 1980; [84] Ooms et al., 1981; [85] Ooms et al., 1982).
There are three more unique attributes provided by the Agrobacterium genetic analytical system. First, the Ti and RI plasmids of different origins are compatible and mutually exchangeable. Second, the “DNA content” of the T-DNA region flanked by border sequences [86] (Jen & Chilton, 1986) could “freely” be replaced by other sequences. Third, the existence and special function of (prokaryotic) vir genes that can mobilize and activate T-DNA cassettes.
These genes are coding for Vir proteins. The latter plays a key role in Type 4 secretion (conjugation of the T-DNA) and processing the T-DNA borders trans by using the virD1/2 relaxation complex, allowing whose function is to mobilize the T-DNA region, (whatever DNA sequences are inside), which cannot be imagined without severely influencing the cell membranes. The greatest advantage from our aspect is that they are capable of acting either from a cis or a trans position (Csaba Koncz, personal communication)
2. Material and Methods
2.1. Bacterium Strains
2.1.1. Bacterium strains: their origin
AMP-producing [63] Xenorhabdus EMA and EMC strains [55,58] are from the Fodor Laboratory in the Department of Genetics at Eötvös University, Budapest, Hungary. Agrobacterium tumefaciens (HP1836 – HP1843) strains were from the frozen stock-collection of coauthor F. O. HP1836 - HP1840 had been deposited there by B. Dudás; HP1841 by D. Silhavy; HP1842 by V. Tisza, and HP1843 by G. B. Kiss. A. tumefaciens SZL1SZL2, SZL3, SZL4, and SZL5 were provided by coauthor L. Sz., (BRC, Hungarian Academy of Sciences, Szeged, Hungary. Agrobacterium and Xenorhabdus strains were grown and cultured according to the respective routine protocols of [87] (Ausubel et al., 1999); [88] (Leclerc & Boemare,1991); [89] (Wise et al., 2006). All the in vitro bioassays were carried out in Luria Bertani broth and/or Luria Bertani Agar.
2.1.2. AMP-producing Xenorhabdus strains
Xenorhabdus budapestensis (EMA) isolated from Steinernema bicornutum was discovered and identified by us [55] (Lengyel et al., 2005). Samples were deposited in DSMZ (DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Inhoffenstr. 7B, - Braunschweig – Germany; (http://www.dsmz.de ) as DSM16342 in 2004. It has also been deposited in Hungary (asAF13); and also in the Laboratory of Prof. Heidi Goodrich-Blair (Department of Bacteriology, University of Wisconsin –Madison, Madison, WI, USA) as HGB033. A spontaneous rifampicin-resistant mutant strain was isolated from HGB033 by András Fodor and also deposited there as HGB2238. (Some comparative tests also used the antibiotic-producing X. szentirmaii HGB036, as well as the spontaneous rifampicin-resistant HGB2239 strain isolated from HGB036 by A. Fodor). All information concerning keeping, culturing, fermenting, and bio-assaying EMA has previously been reported [56] (Furgani et al., 2008); [57] (Böszörményi et al), 2009; [67] (Vozik et al., 2015).
2.1.3. Test organisms
Control organisms
Negative (EMA_PF2 sensitive) control) bacteria:
Gram-negative bacteria:
Multi-resistant E.coli strains were used: HGB 1333 /BW29427 (Dap-requiring, CmR) from H. Goodrich-Blair). This strain was also used as an antibiotic double-resistant (KmR; CmR) E. coli strain. ABC 0801 (harboring plasmid with KK88 antigen; KmR, CmR, SmR, TcR); ABC 1609 (with plasmid TcA1; KmR, ApR, / SmS, SpS, GmS); ABC 0156 (TG90nalR; R55 with integrated SG11 genomic island; CmR, KmR, SulR, SmR, ApR, RifR, EryR) from F. Olasz; ABC 0785 (hly+, sta, stb; plasmids: pTC, 18ac; TcR, from B. Nagy); ABC 1611 (Serotype: K12; pR16A; KmR, ApR, SmR, SpS, GmS) from P. Dublet, (personal com); ABC 1499 (Human clinical isolate, KmR, GmR, CmR, FloR, SmR, TcR ) from F. De la Cruz (personal com.; ABC 0280 (Human clinical isolate A3R; CmR, KmR, SulR, SmR, ApR, RifR, EryR) A. Cloeackert, personal com. Also Salmonella strains: S. Typhimurium ABC 0159 (Natural isolate, SG11 genomic island, CmR, ApR TcR, SmR, RifR); S. Typhimurium ABC 0208, (Natural isolate, SG11 genomic island; CmR, NalR, ApR SmR, TcR, RifR); S. enteritidis ABC 0741, (Natural isolate, pFOL1111; ApR); S. enteritis ABC 1844 (Serotype LT2; recA1; srl-202::Tn10 TcR rifR; TcR, RifR) and S. infantis ABC 1748 (Natural isolate RifR, SpR EryR, SuR sulfamethoxazole / SmR) all from F. Olasz.
Gram-positive bacteria:
Staphylococcus aureus (SA) JE commercial strain (J.C. Ensign, unpublished) from Dr. J.C. Ensign’s Lab was used.
Fungi:
Candida albicans (CA) JE strain (J.C. Ensign, unpublished) was used as a fungal target for testing each preparation for antimicrobial activity in Agar Diffusion Bioassays, which were carried out as described [67] (Vozik et al. 2015) with minor, actual modifications.
Xenorhabdus strains as test organisms.
HGB1795 was generously provided to Prof. Helge B. Bode via Prof. Heidi Goodrich-Blair as a transposon-induced insertion mutant of the XNC1_2022 gene (Gene ID: 9430524; Gene Page Link: NCBI UniProtKB; Locus Tag: XNC1_2022 see gene page for GenePage for the XNC1_2022 gene EcoGene-RefSeq) from Xenorhabdus nematophila (strain ATCC 19061 / DSM 3370 / LMG 1036 / NCIB 9965 / AN6). The reason why we involved this mutant in this study on EMA_PF resistance studies is that previously Bicornutin A was believed to be the active EMA antibiotic molecule, [57] (Böszörményi et al., 2009) and XNC1_2022 gene of X. nematophila was thought to be a homolog of Xenorhabdus budapestensis NrpS (nrpS) gene, (GenBank: Accession Number is JX424818.1; gene synonym="bicA) which is responsible for the biosynthesis of Bicornutin A, [60] (Fuchs et al., 2012). It turned out that it was not the case. However, some roles in the scenario related to antibiotic activity and self-resistance cannot be ruled out, since the coexistence of Bicornutin A and fabclavine in our peptide preparations. Later, however, we were informed by Prof. H. Bode (via H. Goodrich-Blair), that the strain is not as what was believed, or maybe contaminated. Therefore, we publish the exciting-looking data found in the Preprint version without comment in this article. The data are correct, but as we learnt the strain is dubious. (Please consider this statement as an “ERRATUM”).
The AMP-producing Xenorhabdus strains were used as positive controls. They were: X. budapestensis HGB033 and HGB2238 (rifR), X. szentirmaii HGB036, HGB2239 (rifR), . X. nematophila ATCC 19061 (from S. A. Forst), HGB081 (rifR), and HGB1789 (rifR).
Agrobacterium strains
To reveal the sensitivity (S) / resistance (R) phenotypes to the EMA: PF2 antimicrobial peptide complex and its subfractions, we chose Agrobacterium strains of different genotypes for in vitro liquid bio-assaying of EMA_PF2 on them first. We worked on strains of different opine types and those on different plasmid genotypes within the opine groups. Agrobacterium strains used in this study are listed in Table 1. From our point of view the most important subgroup is the four AGR) agropine (the most up-to-date name:L, L, - succinamopine, - catabolizing strains). The wild type strain: A281 [79] (Guyon et al., 1980); and each of the other here is disarmed (T-box free) derivative: EHA5 [87] (Hood et al., 1986); AGL1 [91] (Lazo, Stein and Ludwig, 1991); EHA105 [92] (Hood et al., 1993); and A4T [93] (White and Nester, 1980); [94] (Petit et al., 1982); [95] (Jouanin et al, 1986); [96] (Slater et al, 2009).
pAl4404; pTiA136Bo542; pEHA101; pEHA105 and A4T) see Textext and Supplementary material TextS1.
We worked on 4 agropine (L, L, - succinamopine, AGR) - catabolizing strains in this study: A281 [79] (Guyon et al., 1980); [761] ([80] (Tremblay, 1987), [90] (Hood et al., 1986); AGL1 [91] (Lazo, Stein and Ludwig, 1991);EHA105 [92] (Hood et al., 1993); and A4T [93] (White and Nester, 1980); [94] (Petit et al., 1982); [95] (Jouanin et al, 1986); [96] (Slater et al, 2009). All of them are C58RifRstrains. All but A4T have a C58 (“S”) chromosome - (the abbreviation indicates the geographic origin (Seattle) of strain A136 (C58 (Rif®), its chromosome also called “Seattle C58”); the sequence of which is slightly different from that of the previously discovered and sequenced “Gent/Leiden C58C” chromosome of nopaline catabolizing plasmid-cured strains Dr. Paul J.J. Hooykaas, personal communication).
A281 has a wild-type C58 (S) (Rif®)´ chromosome from one of its ancestors, (the nopaline-catabolizing A136); and an intact, virulent atropine-catabolizing pBo542 [T-DNA] (+) plasmid (from its other ancestor, Bo542). A281 is a hyper, - [90] (Hood et al, 1986); [97] (Hood et al, 1987); and also a super, -[98] (Jin et al., 1987) virulent strain. A known sequence of the pTiBo542 plasmid, outside the T-DNA box [90] (Hood et al, 1986); [99] (Komari, Halperin and Nester, 1986) is responsible for both hyper, - and super-virulence. The intact pTiBo542 plasmid has the T-DNA cassette, containing genes responsible for the synthesis of tumor openings L, L-SAP, LOP, and AGR. The disarmed-DNA deleted remainder sequence, called pEHA 101, contains genes coding for catabolizing enzymes of these opines.
AGL1 is a disarmed derivative of A281 with a mutated C58 (S) (Rif® chromosome with a deletion in the in the RecA gene; its exact genotype is (C58(S), RecA::bla; Rif® arb®, and is called AGL0; pEHA101.(The pEHA101= pTiBo542 DEL-T-DNA plasmid). The plasmid markers are NalR Mop (+) [91] (Lazo, Stein and Ludwig, 1991); see also DNA Cloning Service, www.dna-cloning.com).
EHA105 is an indirect derivative of the intact plasmid from A281 (pTiBo542). It was generated from pEHA101 through site-directed deletion of the kanamycin resistance gene on pEHA101 and by Gen® [92] (Hood et al., 1993). (Previously pEHA101 had similarly been created from the wild-type (pTiBo542) when the T-DNA was replaced by KmR, [90] (Hood et al., 1986).´The genotype is C58(S) Rif® (pTiBo542DT-DNA = pEHA105 / / pBIN-19 –intronGus100-Km®).
(See also: (http://www.springerlink.com//content/t02h1486p1862715/).
A4T is an agropine-catabolizing helper strain of the “Gent/Leiden C58C” chromosome; and harbors a T-DNA-deleted (disarmed, helper [T-DNA] (-)]. A4T plasmid originated from A. rhizogenes; and the binary vector pBIN19 intron (Gus Km®) [100] (Bevan, 1984). For more details on A. rhizogenes helper plasmid harboring strains and their agro-biotechnological importance, see review [101] (Taylor et al., 2006).
As for the NOP strains used in this study, we did not have a chance to the virulent wild-type ([T-DNS]) (+) strai5 of the 6 (HP1836 (C58C*-NOP1); HP1840 (C58C*-NOP2); HP1843 C58C*-NOP3; HP1841 (C58C1-NOP4); HP1842 (C58C1-NOP5) are plasmid-cured, [102] (Uraji, Suzuki, and Yoshida 2002); and only the SZL4 C58C1- pMP90 - NOP6 harbors the disarmed (helper, T-DNA deleted, pTiC58 [T-DNA [(-) called pMP90) plasmid, [103] (Koncz and Shell, 1986). Each of them has a C58 chromosome [69] (Wood et al, 2001). The SZL4 (C58C1-pMP90-NOP6 strain has (the original “Gent/Leiden”) C58C chromosome. [104] (Koncz and Schell, 1986), The C58 chromosomes of the other 5 are other (Hungarian) isolates that have not been sequenced yet. The genome-selective marker for HP1836 (C58C*-NOP1); HP1840 (C58C*-NOP2) and HP1843 (C58C*-NOP3) strains are nalidixic acid resistance (Nal®); while that for HP1841 (C58C1-NOP4); HP1842 C58C1-NOP5; and C58C1-pMP90-NOP6 are of rifampicin resistance (Rif®).
As for the OC)T strains used in this study, we did not have a chance to work either on the ancestor wild-type strain, harboring the virulent pTiAch5 [T-DNA] (+) plasmid; or on the first disarmed derivative of that plasmid is LBA4213 [85] (Ooms et al., 1982); or on any plasmid-cured OCT strains. Our work has been restricted to HP1837 (LBA4404/0-OCT1) and SZL2 (LBA4404/pBIN-OCT2) strains. They both have the Ach5 chromosome [70], (Henkel et al, 2014), and the chromosomal marker for them is Rif®. Each of the two strains, HP1837 (LBA4404/0-OCT1) and SZL2 (LBA4404/pBIN-OCT2), strains harbor the disarmed T-DNA deleted helper plasmid pAL 4404; (as known, encoding genes needed for both T-DNA transfer; and octopine degradation [82] (Klapwijk, & Schilperoort, 1979 ]; [104] (Dessaux et al., 1988). The plasmid marker is Sm.
All these are necessary to understand what are summarized in Table 1.
2.2. Antibiotics-active peptide fractions isolated from EMA CFCM.
Agar-diffusion bio-assays of all studied antimicrobial active peptide fractions isolated from EMA_CFCM on four sensitive test organisms were carried out. The isolated and bio-assayed fractions are enumerated (and together with the results of their antimicrobial activity data) given in Table 2 (Results section).
2.2.1. Isolation of Antimicrobial Active Peptide Fractions EMA_PF1, and EMA_PF2, were carried out by Amberlite adsorption followed by washing (purification)and eluted by acetone with methanol gradient and eluted by methanol [56] (Furgani,2008), [57] (Böszörményi et al., 2009). 2.2.2. Isolation of Antimicrobial Active Peptide Fraction (EMA30)
who allowed the publishing of the results but did not want to be coauthor in this paper and kept the methodical details unpublished. However we used the reversed-phase HPLC (RP-HPLC) technique which is one of the most important techniques for peptide separation, [106] (Josic and Kovac, 2010). We used RP-HPLC for separating biologically active peptide fractions. We used the respective protocols provided for cleaning, regenerating, and storing reversed-phase chromatography columns as es described, [106] (Josic and Kovac, 2010). Briefly, similarly as described by Ensign and his associates [107] (Bowen and Ensign 1998), ˙[108] (Bowen and Ensign 2001), [109] (Chiche et al., 2012), the proper volume of the CFCM or the diluted 10 mg/ml stock solution of (previously freeze-dried) EMA_PF2 were used. All buffers and stock solutions for column chromatography were filtered through 0.2-μm-pore-size filters and autoclaved before use. The Sigma protocol was modified by Professor J. Ensign (unpublished) and we used his modified method. Briefly, the column was eluted with a mixture of acetonitrile (AN), and CH3CN in 0.1% TFA (trifluoroacetic acid) at a flow rate of 0.4 ml/min at room temperature, that is 0, 10, 20, 30, 35 40, 50, and 70, V/V% of AN containing 0.1% TFA. RPCC fractions were named by the number of the concentration of AN, which eluted them from the column. The antimicrobial active peptides from EMA cell-free culture media were quantitatively eluted as one single faction by 30 V/V AN (containing 0.1% TFA) and called EMA30. It exerted strong anti-Gram-positive, anti-fungal, and anti-Gram/negative activity (data not shown) and was used for biochemical characterization.
2.2.2. Antimicrobial active HPLC fractions
Each HPLC sample was of a given volume of a distilled water-dissolved and diluted freeze-dried antimicrobial peptide-complex solution, and, depending upon the column, each respective volume was loaded, following the protocol. The HPLC protocols we used as described by [110] (Carr (2002). The eluent absorbance at 218 and 280 nm was routinely monitored. The peaks were detected at 168 -215 nm and 168-280 nm, respectively. Fractions were collected corresponding to the appearing peaks. Both EMAPF2 (the first HPLC sample was called af3, and the second run af6), and EMA30 (called AF103) were subjected to HPLC. As for af6, three HPLC peaks were detected, and 5 fractions from below the latest peak (called A2) exerted strong cytotoxic activities on both Gram-positive (SA) and Gram-negative (EC) targets; (see Results). Each experiment was repeated at least twice. Three peaks from below the main peak of AF103 (called AF103-40; AF103-43 and AF103-44)exerted strong anti-Gram-negative, anti-Gram-positive, and anti-Candida activity. These fractions were collected on the 40th, 43rd, and 44th minutes of the 60-minute HPLC run. None of the other fractions showed anti-Gram-negative activity. These fractions were then used in MALDI analysis.
2.3. Bioassays of Antimicrobial Peptide Fractions from EMA CFCM
2.3.1. Methodology of Liquid Bioassay of EMA AMP-Active Fractions on Agrobacterium strains.
In vitro Liquid - Culture Bioassays of EMA PF on Agrobacterium strains were carried out in sterile 96-well tissue culture plates.
Briefly, each culture had 200 µl in the final volume containing 100 µl of 2X LB (supplemented with the respective selective antibiotics) and 95 µl of a sterile water solution from the partially purified EMA PF2, and inoculation of 5 µl bacterial LB suspension from 100 µl; which contained 1 loop-size bacteria from a single, individual test bacterium colony grown on an LBA plate in 24-h. For the three replicates, 3 independent clones were used. Purified, freeze-dried, and re-dissolved preparations of EMA PF were used at 0, 30, 46, 6,0, and 75 µg/ml concentrations. We incubated the experimental plates for 24h at 28 0C and then determined the OD values spectrophotometrically. The growth of bacteria was quantified based on the optical densities (OD values) of the cultures by screening the plates spectrophotometrically. The lower OD values indicated the stronger antibacterial activity of the EMA PF and higher sensitivity of the Agrobacterium strain tested. Other technical details of the experimental conditions of Liquid Bioassays had been published earlier, [66] (Fodor et al., 2012), [67] (Vozik et al., 2015).
2.3.2. Quantitative evaluations
If we had worked with a single antimicrobial active compound, we should have determined the minimum inhibitory concentration (the MIC value, that is the lowest concentration of the tested compound at which bacterial growth is completely inhibited. We, however, have had to provide some quantitative parameters of the antimicrobial activity of a mixture of peptides of different antimicrobial activities. If we determined the quantitative amounts of peptides that exerted a complete inhibitory effect on the tested bacterium strain, these “MIC” values could not be considered as quantitative data referring to one active component, but still provide an option for comparing the activity of different EMA_PF2 peptide fractions in different strains. Therefore, we determined a value that the “gross MIC values” of freeze-dried as if EMA_PF2 solution. The “gross MIC” value is suitable for comparison of the activities of the EMA_PF2 on different targets, and this is the aim of this study because this is what is happening in nature. Technically the “gross MIC values” of the EMA_PF2 were determined similarly to the MIC value of a single AMP, following the standard protocol, [111] (Wiegand et al, 2008). We determined the lowest growth-inhibiting dose of the EMA_PF2 mixture (and separated fractions) on Agrobacterium and control (E. coli, Xenorhabdus, S. aureus) strains. We used LB broth for dilutions. Briefly, we worked in “SARSTEDT Multiple Well Plate 96-Well Round Bottom with Lid” culture plates, (Sarstedt, Inc., Newton, NC 28658, USA). Test bacteria were inoculated into a liquid growth medium containing different concentrations of EMA_PF2. Growth was determined based on the OD values of the liquid cultures, after incubation for 24h (at 28 0C, when the test targets were Agrobacterium and Xenorhabdus) and 12h (at 37 0C, when E. coli, S. aureus, and Candida were the test organisms). When the OD value of the culture did not differ significantly from that of the freshly inoculated LB culture of the same composition, we cautiously considered the applied MA PF concentration (given in µg/ml) as (gross) MIC90. In case of complete cytotoxicity, we kept the cultures for another two weeks on the bench top and considered the final result if no growth was detected during this period. The “MIC” /- /gross MIC) - values added in the Tables and Figures are the means of three replicates. In Agar Diffusion Bioassays we -pipetted 100 μl of samples into a hole in the center of a 1/cm thick LB agar plate. The respective plate was then overlaid by the suspension of the test organism, and diluted with soft agar as published earlier [67] (Vozik et al., 2015). The diameter of the inactivation zone was measured and the volume of agar media was calculated from that measurement. We considered these data as also informative but preliminary.
2.4. Statistics
The data analysis was performed using [SAS/STAT] software, Version [9.4] of the SAS System for [Windows X 64 Based Systems]; (Copyright © [2013 of copyright]; SAS Institute Inc. SAS, Cary, NC, USA. We used ANOVA and GLM Procedures alternatively following the requirements of the SAS 9.4 Software. The design of the experiment could be considered as a randomized complete block design with the number of the respective treatments, concentrations, and replicates. Data have been averaged to allow the analysis of variance (ANOVA). The significance of differences of the means (α=0.05) was determined by using t (Least Significant Difference, LSD) tests or Duncan’s Multiple Range Tests, depending on the experiment. (For more details, see Supplementary Material, S_Text_2).
3. Results
3.1. Antimicrobial Activity of EMA_PF2 on E. coli and Xenorhabdus strains. Results of Liquid Culture Bioassays.
The results of the essential control experiments, that is those of the liquid culture bioassays on closely related (belonging to the genus Xenorhabdus) and unrelated (E. coli, HGB1333. and HGB2226) Gram-negative bacteria EMA_PF2 are presented in Figure 1A,B. and demonstrating the strong anti-Gram-negative potential of the EMA_PF2 peptide fraction. The data show, that the AMPs in the EMA_PF2 fraction of X. budapestensis are very efficient against the non-related E. coli strains while the closely related Xenorhabdus stains (with the only exception of HGB1795) are resistant, as expected [55] (Fodor et al., 2022), [65] (Fodor et al., 2010). EMA_PF2 (and EMA_PF1, which is probably an aggregate of the same molecules) AMP prepare rations also tested on the X. nematophila mutant HGB1795 and X. nematophila clones (HGB081 and HGB1789) of its parental genotype (but different isolates), and these data are presented in Figure 1A and Figure 1B, in comparison with other different Xenorhabdus strains. HGB1795 has a mutation of a strange operon in X. nematophila. One of the mutants (X. nematophila (pCEP_kan_XNC1_1711) is an acaricidal bioactive strain [112] (Incedayi et al., 2021). The operon went through a detailed molecular analysis and was identified as a toxin-antitoxin module, [113] (Yadav and Rathore, 2020). We found it extremely surprising that all but HGB1795 proved resistant. The resistance of the wild-type Xenorhabdus strains indicates that the EMA_PF1 and EMA_PF2 fractions supposedly contained only antibiotic-active peptides, but not -but not R-type bacteriocin [114] (Morales-Soto and, Forst, (2011). But the sensitivity of the mutants is strange. We lost the train but wanted to repeat this experiment on an independent new culture of the stain, but our request for the sample was refused.
3.2. Antimicrobial Activity Profile EMA_PF2 and EMA 30 peptide fractions
Antimicrobial active fractions from EMA_CFCM could be separated either by amberlite adsorption or RFLP and could be purified by HPLC. Data on the antimicrobial activities of those preparations that proved antimicrobial active in each of the target organisms are presented in Table 2. As for the fractions, information concerning purification and description of different AMP preparations made during these experiments are listed in Supplementary Material Table S2.
As for PF1, it was found in very small quantities, it was not enough quantities for each test, but seems to act similarly to EMA_PF2. We suppose that the same AMPs are present in both fractions, but we cannot interpret the existence of EMA_PF1 at all, since the known AMPs in Xenorhabdus are of small molecular site.Although it was very potent in each target organism, we could know, whether it consists of spontaneously polymerized active smaller peptides, or large, originally inactive peptides that were simply “contaminated” with smaller antimicrobial active ones. As for EMA_PF2, the data of the antimicrobial activity (measured in two different experiments) on the examined Gram-positive and unrelated Gram-negative Gram-the proved strong, and concentration-dependent. The Statistical (ANOVA Procedure) Analysis of the data is present in Supplementary Material.
3.3. HPLC and MALDI Profile of EMA_PF2 and EMA 30 peptide fractions
The EMA_PF2 preparation, which was used in the liquid bioassays on Agrobacterium strains, could be separated into three sharp peaks after repeated HPLC runs (Figure 2).
Some but not all fractions collected from below the third (called A2) peak exerted both anti-Gram-positive and -Anti-Gram-negative activities when tested on S. aureus JE and E. coli (HGB2226) strains. Anti-Gram-Positive and Anti-Gram-Negative activities could not be separated in such a way (Table 3).

Table 3.
.
| HPLC run | Staphylococcus aureus J.E. | Escherichia coli HGB2226 J.K |
|---|---|---|
| OD Values at 600 nm for the fractions collected between 40-50 min (Mean ± SD, n=2 at each time) | ||
| 1. | 0.3577±0.0797 | 0.5380± 0.009 |
| 2. | 0.4404±0.0511 | 0.4214±0.0002 |
| 3. | 0.4273±0.0377 | 0.4335±0.0002 |
| 4. | 0.4588±0.0307 | 0.4625±0.001 |
| 5. | 0.4027±0.0285 | 0.48135±0.00063 |
| 6. | 0.3874±0.0510 | 0.4651±0.00198 |
| 7. | 0.4255±0.0571 | 0.4395±0.0004 |
| 8. | 0.0003±0.0247 | 0.00155000±0.0006 |
| 9. | 0.0003±0.0201 | 0.00020000±0.0002 |
| 10. | 0.0081±0.0547 | 0.001±0.0001 |
| 11 | 0.0040±0.0061 | 0.0015±0.0002 |
Footer to Table 3 Anti-Gram Positive and Anti-Gram-Negative Activities of HPLC (EMA_PF2) isolated from the Cell-Free Culture Medium (CFCM) of Xenorhabdus budapestensis (EMA) by Amberlite XAD1148® Adsorption, Methanol Elution, and Ultrafiltration in liquid culture bioassay.). Abbreviations: HPLC Sample: af3; Peak: A2. fractions collected (from below A2, between 40 – 47 min exerted complete bactericidal activities on both the Gram-positive and the Gram-negative test organisms. Fractions collected before and after this time interval were completely inactive. Each mean was calculated from 3 replicates. In repeated experiments, using different columns got similar results. (We were not interested in the growth rates of the bacterial cultures; only in the completely toxic fractions. This explains the layaways from the original protocol).
Thus, we figured that peptides between 1340 – 1366 m/Z (believed to involve, 1346 m/Z, fabclavine) were responsible for the antimicrobial activity on four different EMA-sensitive targets. The MALDI profiles of both the antimicrobial active (Figure 3 A,B) and inactive (not shown). HPLC fractions contained many peptide peaks in the range (about 946 m/Z), believed to be where Bicornutin A is located.
The text continues here (Figure 3
The antimicrobial activity could be increased when the most active fraction obtained by RFLP was further purified by HPLC, but the ant-Gram-positive, the anti-Gram-negative, and antifungal activities could not be separated in such a way, (see Figure 4).
3.4. Results of Liquid Culture Bioassays of EMA_PF2 on Agrobacterium Strains of Different Genotypes, Opine Types, and Plasmid Genotype
The MALDI profiles of two antimicrobial active preparations from the cell-free culture medium (CFCM) of X. budapestensis (EMA PF2 (HPLC) A2 and EMA RFLP AF30 AF103 Fr.43) were obtained from EMA cell-free culture media by different protocols. (A) MALDI profile of pooled antimicrobial fractions from below A2 peak of the HPLC profile of EMA_PF2. (B) MALDI profile of EMA 30, purified by RPLC. For more details, see the Footer to Figure 3. The distribution of OD values as a function of EMA_PF concentrations is presented in Figure 5. (As for the respective statistics, see Supplementary material, Figure S1A – H; Tables S3 and 4).
Of the twelve tested Agrobacterium strains, eight proved unambiguously resistant to each applied dose (at somewhat different degrees), that is, the gross MIC values could not be determined. One of them was the HP1838 (A281, of T-DNA (+) genotype). wild-type AGR strain. Each of the studied four T-DNA deleted strains proved extremely sensitive, (represented by low OD (<0.2) values even at the lowest applied EMA_PF2) dose, (Figure 5). However, these experiments should be repeated with these and several different strains of similar genotypes before concluding, considering that the Agrobacteria does not produce antimicrobials, therefore unexpected (as those found in HP1837 and HP1839 after the Preprint paper appeared) and unrevealed contamination with EMA-sensitive bacteria may bias the results. Although our statements about HP1837 and HP1839 aim to be an erratum, considering that neither in the case of the wild-type nor in that of the 7 and plasmid-cured Agrobacterium strains any doubt has never appeared concerning their EMA resistance, (it was always unambiguous) we should not simply forget about these data as erroneous ones but the tests should be repeated on many T-DNA-deleted Agrobacterium strains.
It is not a surprise that the wild-type A281 (HP1838) was resistant to the unpurified EMA CFCM, while its Δ –TDNA derivative, EHA105, a T-DNA deleted (∆ –T-DNA) was very sensitive (Figure 6A and B, respectively). When comparing the OD values of the four AGR strains, it a spectacular difference between the strain (HP1838) of T-DNA (+) and of the three strains (HP1839, A4T, and SZL3) of T-DNA (-) genotype.
The S/R profiles of three disarmed AGR strains hardly differed from each other (Figure A, B, C, and D). (As for the respective statistics related to the results of AGR strains, see Supplementary material, Table S5A, S5B). As for the octopine strains, the picture is not so clear. SZL2 is resistant, and HP1837 is sensitive, (Supplementary material Figure S3).
As for the studied NOP strains, we did not have disarmed ones, each of the studied ones proved resistant to EMA_PF2 in vitro liquid bioassay but none of them harbored T-DNA-del Ti- plasmid. Data on NOP strains are presented in Supplementary Material, Figure S3; Suppl Text_6; Supplementary Material, S_Text_6; Table S7.
4. Discussion
This study was inspired by our special interest in better understanding the mechanism and the intraspecific diversity in resistance (not simply to a simple AMP molecule but) to a branch of AMP molecules complex released to the environment together by our AMP-producing organism, X. budapestensis (EMA).
EMA is an EPB living as an obligate symbiont of eukaryotic nematode partner (the insect-parasitic Steinernema bicornutum ), [55] (Lengyel et al., 2005). [58] (Fodor et al., 2023) in an extreme polyxenic environment (insect cadaver in soil), and one of its “symbiotic job” is to produce an efficient set of (defensive) AMP molecules that provides safe living, and pathobionts conditions for itself and its eukaryotic (EPN) symbiont partner [50] (Ogiert et al., 2020) by overcoming all potential competitors. These AMP molecules accidentally inactivate (not all but many) harmful plant pathogen bacterium (like Clavibacter,[66] (Fodor et al., 2010) and Erwinia [54] (Böszörményi, et al., 2009), [67] (Vozik et al., 2015)) species), although they are not natural competitors of EMA, therefore not natural targets of these AMPs. We intend to benefit from that. This is the explanation for why a significant part of this publication provides data about the antimicrobial potential of different peptide fractions. This part of the work is scheduled to be continued in a practical direction and we planning to involve analytical chemists especially experts in evaluating the HPLC and MALDI profiles. We also intend to work on more test organisms, including plant and veterinary pathogens. We believe that this information is important and deserves to be published.
A challenging but most critical part of this study describes experiments aiming at getting closer to revealing the details of the genetic background of collective resistance to a set of different AMP molecules designed by nature to act together against a selected group of competitors of the AMP-producing organism. We are at the very beginning. We found the studied wild-type strain (A281) of A. tumefaciens is fully resistant, and several variants of this resistant species we found sensitive. This seems to provide an option for genetic analysis.
From a technical point of view, it is a critical weakness, that the Agrobacterium did not produce any antimicrobial compounds, therefore accidental unrevealed contamination during the 24-hour residence/sensitivity bioassay may happen and could bias the results if the contaminant was a sensitive organism that otherwise overgrows the culture. (It never happened that an Agrobacterium strain that was found as resistant changed later to sensitive, or vice versa, but HP1837 and HP1839 which were found sensitive, (see Preprint version) proved contaminated with E. coli years later, and we do not know when they became contaminated. Therefore we declare here those results as ERRATUM and do not discuss them here either).
The answer to the first arising question (whether ever existing such a type of “collective” resistance mechanism at all), is MDR-efflux pumps exist and efficiently work in Gram-negative pathogen bacteria, [15] (Nehme and Pole, 2005). In the RND-type multidrug efflux system, AcrABR, and its regulation mechanism have recently been described [117] (Nuonming et al., 2018). It was published that the AcrR is the transcriptional repressor of the acrABR operon in A. tumefaciens, [117] (Nuonming et al., 2018). The AcrAB efflux pump confers resistance to various toxic compounds of very different chemical structure, including several antibiotics like ciprofloxacin (CIP), nalidixic acid (NAL), novobiocin (NOV), and tetracycline (TET)], also two detergents (sodium dodecyl sulfate (SDS) and biocide triclosan (TRI)) [117] (Nuonming et al., 2018). There has been no published data concerning any AMP molecule available, but we suppose that the AcrABR multidrug efflux system may provide resistance against AMP- molecules as well. The studied (supposedly) sensitive A. tumefaciens variants are all laboratory strains, but none of them was designed to be sensitive to EMA-released AMP molecules. Their detected sensitivities need explanation, whether they might be considered as a spectacular example of pleiotropy, [118] (Watanabe et al., 2019), or simply experimental artifacts explained by unrevealed contamination. If our observations were confirmed by other labs, it might mean that these strains are not protected by those MDR-resistance mechanisms, which protect the wild-type A281.
The answer to the second arising question (whether these (supposedly) sensitive laboratory strains could be considered as real genetic variants) is that the well-defined genotype of each of them is different from that of the wild-type, as well as from those of each other. It should be discussed whether these genetic differences could logically explain the differences found in the EMA_PF2 sensitivities at the level of present knowledge. Our data prove that the (only available) virulent wild-type A281 (HP1838) A.tumefaciens strain of known genotype is extremely resistant to EMA_PF2.
We suppose that there must be a gene (let us call it REmaAmp) ) responsible for the EMA-PF2 AMP resistance of the wild-type. If this gene happened to be hit by a loss-of-function mutation, the EMA-PF2 resistant phenotype should be switched to a sensitive one. We suppose that REmaAmp is an essential [119] (Ward et al., 2023) gene of indispensable function. REmaAmp must be located in the chromosome, because (i) strains with both (known) chromosomes (C58 and Ach5 ) were found among both the EMA-PF2 sensitive, and the EMA-PF2 resistant strains; (ii) each of the (studied, nopaline-producing) plasmid-cured [102], (Uraji, 2002), [120] (Rodrigues et al., 2021) strains proved strongly resistant to the EMA-PF2 AMP-complex. These data also indicate that the presence of the Ti plasmid is not needed for the EMA-PF2-resistant phenotype either. As for the plasmid-cured strains, we should keep in mind that the “opine-type” as a phenotype in plasmid-free Agrobacterium cells simply does not exist at all, (no restriction concerning to transport to any opine-type plasmid to any plasmid-cured strain). but we may not know everything about the narure of plasmid incompatibility. (Ti plasmids of different origins might be incompatible with each other but not with the chromosome). The idea based on a simple model that there is a structure gene on the chromosome and there is a regulatory gene on the Ti-plasmid should not be taken into consideration. Other genetic interactions, however, cannot be ruled out. We expect mainly recessive epistatic interactions [121] (Elston et al., 2017) between chromosomal, and Ti-located genes. Our data suggest some communication between membrane-expressed genes of chromosomal and Ti-plasmid locations. We found reproducibly spectacular differences between the wild-type and the disarmed derivative strains (at least within the (originally named) agropine [78] (Guyon et al., 1980), (recently called) L, L,-succinamopine, [79] (Tremblay et al, 1987) opine group in their EMA-PF2 sensitivities are proven by the ANOVA-based statistical analyses, (Supplementary Material Table S3). We found each of the disarmed [122] (Kiyokawa et al., 2009) Agrobacterium strains EMA-PF2 sensitive. The question is whether it was just an accident or a general rule. Each of the disarmed plasmid has its original, complete, functioning vir-region, [123] (Stahel and Nester, 1986)], [124] (Palanichelvam et al., 2000), [125] (Hattori et al., 2001), essential for their for conjugation and gene transfer to another cell, [126] (Lacroix and Citovsky, 2016) but they lack their original transfer DNA (T-DNA) region, including not only the respective opine synthase genes but all signals necessary for expression for anything in the T-Box in plants after successful transformation, [76] (Koncz et al., 1983), but those of conserved part of the T-region which expresses four proteins in bacteria [127] (Schröder et al., 1983), (and might have been considered as potentially playing some role in EMA_PF resistance). If any plasmid-located gene might play a role in the EMA-PF2 sensitivity of the disarmed strains must be located in the vir region concerning the respective Ti (in our case, pTiBo542) plasmid.
The enormously significant differences between the fully resistant wild-type (that is the ([T-DNA] (+) pTiBo542 plasmid harboring) A281 strain, and the super-sensitive disarmed ([T-DNA] (-) pTiBo542 plasmid harboring) derivative strains, AGL1, [91] (Lazo et el., 1991),], EHA 105 [92] (Hood et al., 1993)) and A4T [95] (Jouanin et al., 1986) (see Figure 5) might be interpreted as the allelic difference of an unknown gene located in the Ti plasmid. This hypothetic gene must be present in the wild type and deleted from the disarmed strains, and its absence from the plasmid influences the AMP permeability of the cell, at least for one or more found in the EMA_PF2 peptide. (During conjugation the permeability of the cell membrane also alters, allowing macromolecular transfer, and this is under the control of genes located in the vir region).. Each was uniformly sensitive to EMA CFCM (Figure 6A) and purified EMA_PF2 peptide fraction, (Figure 7A – 7B).
We intend to use these two strains in designing our genetic analytic experimental system. If this observation were confirmed by another lab, it would mean that there would be a toolkit for carrying out classical Mendelian genetic analysis of an efficient multidrug efflux system providing resistance against AMPs (see Conclusions).
5. Conclusions and Perspectives
2-years after the Preprint version of this paper appeared, two of us (L.M., and J.K.) checked the purity of all the Agrobacterium strains used in this study by MALDI and then by sequencing. We, unfortunately, found that strains HP1837 and HP1839 were contaminated with E. coli which is extremely susceptible to EMA-CFCMs. Therefore although the data HP1837 and HP1839 were correct in the Results section of the published Preprint paper, (therefore we left them in this MS as well unchanged), we have withdrawn some comments and conclusions done in the Preprint version. Please consider this statement as an ERRATUM. We did not withdraw the supposition about the possible EMA_PF2 sensitivity of some disarmed AGR strains but put our previous statement in first conditional like that: if we had a chance to test more disarmed AGR strains (including independent isolates from the previously tested strains) for EMA_PF2 sensitivity and got similar results, we would conclude the discovery novel interactions between genes expressed in the chromosome and (probably in the vir region) of the Ti plasmid of Agrobacterium strains belonging to the AGR opine group, (the wild-type is A281), contributing to the resistance mechanisms to some AMP molecules produced by the entomopathogenic nematode-symbiont bacterium Xenorhabdus budapestensis DSM 16342 (EMA).
Suppose that the AMP resistance, (including or at least) AMP_PF2 resistance) as a phenotype is due to the efficient work of the AcrABR multidrug efflux system, in A. tumefaciens [117] (Nuonming et al., 2018) a question arises: whether the resistance mechanisms against completely different chemicals could be separated by the tools of classical Mendelian genetics. The availability of different phenotypes concerning resistance and sensitivity seems to provide an option to do so. It would be great, considering the biological and structural diversity of Type IV secretion systems, [128] (Li et al., 2019), used by A. tumefaciens. But if this option could not be justified by the similar results of similar experiments on other disarmed strains, it should be forgotten.
If we had a chance to carry on this project, we would go ahead with a Mendelian complementation analysis [129] (Ward et al., 1990) based on the option of producing a partial heterozygous constellation ˙[80] (Hoekema et al., 1983), [100] (Bevan, 1984), [103] (Koncz and Shell, 1986), and using the sophisticated molecular techniques [120] (Rodriguez et al), [130] (Bitrián et al., 2011), [131] (Wang et al, 2023) of the binary vector technique for Agrobacterium research. The target of such a genetic analysis must be the vir region consisting of approximately 10 operons (depending upon the Ti- or Ri-plasmid) that serve four major functions. Secreting T-DNA and Vir proteins from the bacterium via a type IV secretion system (virB operon and virD4). The Agrobacterium virB operon contains 11 genes, most of which form a pore through the bacterial membrane for the transfer of Vir proteins [132] (Christie, et al., 2004). Currently, we know of five such proteins that are secreted through this apparatus: VirD2 (unattached or attached to the T-strand), VirD5, VirE2, VirE3, and VirF [133] (Vergunst et al., 2000), [134] (Vergunst et al., 2005). VirD4 acts as a coupling factor to link the VirD2-T-strand to the type IV secretion apparatus [132] (Christie et al., 2004). These genes could easily be cloned into small T-DNA regions [135] (Koncz et al., into that kind of binary vector via E. coli.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization: A.F., F. O., and J. K., who designed the study. Methodology, A.F., L.M. and L.F. (bioassays); A.F. Software: M. A.B.A.G. (ANOVA), HPLC, MALDI). Validation: M.G.K. Formal analysis, M. A.B.A.G. Investigation, G.M.F., D.V., A.N.E., and A.F.. Resources: K.B.B., Bacterium strains: F.O., J.K., and L. Sz. Data curation: M. A.B.A.G. Writing:—original draft preparation: A.F.; writing—review and editing: F.O., and Supervision, J.K. Project administration: A.F.; funding acquisition: A.F. and K.B.B All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a Fulbright Grant Biological Science Grant (1214102) awarded to András Fodor to conduct research in the Laboratory of Heidi Goodrich-Blair at the Department of Bacteriology at the University of Wisconsin-Madison, USA. Valent BioSciences also contributed to the Bench Costs with a Special Grant, (provided by R&D Director, Dr. József Racskó).
Data Availability Statement
The following information was supplied regarding data availability: Experimental Data Analyzed by ANOVA Procedure: provided in the Supplemental File.
Acknowledgments
We would like to express our thanks and gratitude to those fellow scientists, who were contributors and coauthors in the earlier (Preprint) version of this paper: Erzsébet Böszörményi, who used to make the first observation about the different sensitivities of Arobacterium isolates to EMA CFCMs but did not participate in the revisions, and re-evaluations later on. We express our appreciation and sincerest thanks for the professional guidance of Heidi Goodrich-Blair in the field of bacteriology, supporting the work of A. Fodor (her Fulbright fellow). She was who kindly provided the strain HGB1975, and the needed direct and related sequence information, as well as many other E. coli and Xenorhabdus strains and equipment, lab infrastructure, and consumables we needed. Professor Jerald C. Ensign guided all work related to separation biotechnology, HPLC, RFLP, and MALDI. He provided software and many other things. We also thank people from their laboratories, especially Dr. Angel Casanova-Torres, Terra Maurer, Dr. Kristen Murfin; Mengy Cao, Neta Millet, Kai Hillman, and Daren Ginete for technical and intellectual help. For his abundant intellectual help in connection with the Agrobacterium part at finalizing the MS, we had to express our thanks to Csaba Koncz and Zsuzsanna Koncz (Max Planck Institute for Plant Breeding Research, Cologne, Germany).The experiments at the University of Pannonia, Keszthely, Hungary) were kindly supported by András Takács, Head of the Institute of Plant Protection. We feel obliged to express our thanks to fellow scientist Sándor Józsa (University of Pannonia, Keszthely, Hungary) for his help with statistics, and to Mrs. Andrea Máthé-Fodor her support in completing the Manuscript. We would like to express thanks and appreciation to the professional technical help to lab technicians in the Vet School for their invaluable technical help, especially Miss Éva Kolozsvári and Miss Teréz Halasi. We are thankful for the generous intellectual and technical help of the Head of the OARDC/OSU Library, Mrs. Gwen Short, and her associate Mrs. Laura Appelgate who introduced ZOTERO and made it possible to complete the Manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fodor A, Abate BA, Deák P, Fodor L, Gyenge E, Klein MG, Koncz Z, Muvevi J, Ötvös L, Székely G, Vozik D, Makrai L. Multidrug Resistance (MDR) and Collateral Sensitivity in Bacteria, with Special Attention to Genetic and Evolutionary Aspects and to the Perspectives of Antimicrobial Peptides-A Review. Pathogens. 2020 Jun 29;9(7):522. [CrossRef] [PubMed]
- Rice, L. B. 2008. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: no ESKAPE. Journal of Infectious Diseases, 197(8):1079-1081. [CrossRef]
- Williamson R, Calderwood SB, Moellering RC Jr, Tomasz A. 1983. Studies on the mechanism of intrinsic resistance to beta-lactam antibiotics in group D streptococci. Journal of General Microbiology 129:813-822.
- Gilmore MS, Lebreton F, van Schaik W. 2013. Genomic transition of enterococci from gut commensals to leading causes of multidrug-resistant hospital infection in the antibiotic era. Current Opinion in Microbiology 16: 10-16.
- Miller, W. R. , Munita, J. M. and Arias, C. A. 2014. Mechanisms of antibiotic resistance in enterococci Expert Review of Anti-infective Therapy 2014 Oct; 12:1221-1236. [CrossRef]
- Tomasz, A. 1998. Accelerated evolution: the emergence of multidrug-resistant gram-positive bacterial pathogens in the 1990's.Netherlands Journal of Medicine; 52:219-227.
- Tenover FC, Sinner SW, Segal RE, Huang V, Alexandre SS, McGowan JE Jr, Weinstein MP 2009. Characterization of a Staphylococcus aureus strain with progressive loss of susceptibility to vancomycin and daptomycin during therapy. International Journal of Antimicrobial Agents, Jun; 33(6):564-568. [CrossRef]
- Shi J, Mao NF, Wang L, Zhang HB, Chen Q, Liu H, Tang X, Jin T, Zhu CT, Li FB, Sun LH, 1109 Xu XM, Xu YQ 2014. Efficacy of combined vancomycin and fosfomycin against methicillin-resistant Staphylococcus aureus in biofilms in vivo. PLoS One 9:e113133. [CrossRef]
- Schechner V, Straus-Robinson K, Schwartz D, Pfeffer I, Tarabeia J, Moskovich R, Chmelnitsky I, Schwaber MJ, Carmeli Y, Navon-Venezia S. 2009. Evaluation of PCR-based testing for surveillance of KPC-producing carbapenem-resistant members of the Enterobacteriaceae family. Journal of Clinical Microbiology 47:3261–3265. [CrossRef]
- Schwaber MJ1, Lev B, Israeli A, Solter E, Smollan G, Rubinovitch B, Shalit I, Carmeli Y; 2011. Israel Carbapenem-Resistant Enterobacteriaceae Working Group Containment of a country-wide outbreak of carbapenem-resistant Klebsiella pneumoniae in Israeli hospitals via a nationally implemented intervention. Clinical Infectious Diseases 52:848-855. [CrossRef]
- Vila J, Martí S, Sanchez-Céspedes J. 2007. Porins, efflux pumps, and multidrug resistance in Acinetobacter baumanii. Journal of Antimicrobial Chemotherapy 59:1210–1215. Epub 2007 Feb 26.
- Antunes, L.C.S. , Visca, P., Towner, K.J., 2014. Acinetobacter baumannii: evolution of a global pathogen. Pathogens and Diseases 71:292-301. [CrossRef]
- Nordmann P, Ronco E, Naas T, Duport C, Michel-Briand, Y. Labia R. 1993. Characterization of 1041 a novel extended-spectrum beta-lactamase from Pseudomonas aeruginosa. Antimicrobial Agents 1042 and Chemotherapy 37: 962-969. PMID: 8517722 PMCID: PMC187863.
- Strateva T, Yordanov D. 2009. Pseudomonas aeruginosa – a phenomenon of bacterial resistance. Journal of Medical Microbiology 58:1133–1148. [CrossRef]
- Nehme D, Poole K. 2005. Interaction of the MexA and MexB components of the MexAB-Opr Multidrug efflux system of Pseudomonas aeruginosa: identification of MexA extragenic 103suppressors of a T578I mutation in MexB. Antimicrobial Agents and Chemotherapy 49:4375– 4378. [CrossRef]
- Mulcahy, L.R. , Isabella, V.M., and Lewis, K. 2014. Pseudomonas aeruginosa biofilms in disease.Microb Ecol. 68:1. [CrossRef]
- Jeukens J, Kukavica-Ibrulj I, Emond-Rheault JG, Freschi L, Levesque RC. 2017. Comparative genomics of a drug-resistant Pseudomonas aeruginosa panel and the challenges of antimicrobial resistance prediction from genomes. FEMS Microbiology Letters 364, (18), 2 October 2017). [CrossRef] [PubMed]
- Gebreyes, W. A. and Thakur, S. 2005. Multidrug-Resistant Salmonella enterica Serovar Muenchen from pigs and humans and potential interserovar transfer of antimicrobial resistance. Antimicrobial Agents and Chemotherapy. 49: 503–511. [CrossRef]
- Endimiani A1, Hujer KM, Hujer AM, Bertschy I, Rossano A, Koch C, Gerber V, Francey T, Bonomo RA, Perreten V.. 2011. Acinetobacter baumannii isolates from pets and horses in Switzerland: molecular characterization and clinical data 2011. Journal of Antimicrobial Chemotherapy. 66: 2248-2254. [CrossRef]
- Szmolka A, Nagy B, 2013. Multidrug resistant commensal Escherichia coli in animals and its 1130 impact for public health. Search Results Frontiers in Microbiology. 4: Article 258, 1-13. 1131 Published online 2013 Sep 3. [CrossRef]
- McManus, BA. , Coleman, DC., Deasy, EC., Brennan, GI., O’ Connell, B., Monecke, S., Ehricht, R., Leggett, Leonard, NB., Anna C. Shore, AC. 2015. Comparative Genotypes, Staphylococcal Cassette Chromosome mec (SCCmec) Genes and Antimicrobial Resistance amongst Staphylococcus epidermidis and Staphylococcus haemolyticus isolates from infections in humans and companion animals. PLoS One 10: e0138079. [CrossRef]
- Rzewuska M, Stefańska I, Kizerwetter-Świda MI, Chrobak M, Chimel D, Szczygielska P, Leśniak M, Binek M. 2015. Characterization of Extended-Spectrum-β-Lactamases Produced by Escherichia coli Strains Isolated from Dogs in Poland. Polish Journal of Microbiology 64: 285–288.
- Marques, C. , Gama, L. T., Belas, A., Bergström, K., Beurlet, S., Briend-Marchal, A., Broens, E.M., Costa, M., Criel, D., Damborg, P., van Dijk, M. A. M., van Dongen, A. M.,Dorsch, R., Espada, C.M., Gerber, B., Kritsepi-Konstantinou, M., Loncaric, I., Mion, D. Misic, D., Movilla, R., Overesch, G., Perreten,V., Roura, X., Steenbergen, J., Timofte, D.,Wolf, G., Zanoni, R. G. Schmitt, S., Guardabassi, L., Pomba, C. 2016. European multicenter study on antimicrobial resistance in bacteria isolated from companion animal urinary tract infections. BMC Veterinary Research 2016; 12:213. [CrossRef]
- Fodor A, Varga I, Hevesi M, Máthé-Fodor A, Racsko J, Hogan JA. 2012. Novel anti-microbial peptides of Xenorhabdus origin against multidrug-resistant plant pathogens, In Bobbarala, V. (Ed. Biochemistry, Genetics and Molecular Biology - A Search for Antibacterial Agents, 872 9.
- Załuga J, Stragier P, Baeyen S, Haegeman A, Van Vaerenbergh J, Maes M, De Vos P. 2014. Comparative genome analysis of pathogenic and non-pathogenic Clavibacter strains reveals adaptations to their lifestyle. BMC Genomics 15. [CrossRef]
- Li, X.-Z. , Plésiat, P., Hiroshi Nikaido, H. 2015. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clinical Microbiology Reviews 28: 337-418. [CrossRef] [PubMed]
- Pitout, JD. 2008. Multiresistant Enterobacteriaceae: new threat of an old problem. Expert Review of Antimicrobial Infection Therapy 6:657–669. [CrossRef]
- Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA. 2011. Carbapenems: past, present,and future. Antimicrobial Agents and Chemotherapy 55:4943-4960. [CrossRef]
- Temkin, E. , Adler, A., Lerner, A. Carmeli, Y. 2014. Carbapenem-resistant Enterobacteriaceae: biology, epidemiology, and management. New York Academy of Sciences 1323:22-42. [CrossRef]
- Gupta N, Limbago BM, Patel JB, Kallen AJ.2011. Carbapenem-resistant Enterobacteriaceae: epidemiology and prevention. Clinical Infectious Diseases, 53: 60-67.
- Kádár B, Kocsis B, Nagy, K, Szabó, D. 2013. The renaissance of polymyxins. Current Medicinal Chemistry Journal 20: 3759-3773. [PubMed]
- Otter, J. A. , Doumith, M., Davies, F., Mookerjee, S., Dyakova, E., Gilchrist, M., Brannigan, E. 1055 T., Bamford, K., Galletly, T., Donaldson, H., Aanensen, D. M., Ellington, M. J., Hill, R., 1056 Turton, J. F., Hopkins, K. L., Woodford, N. Holmes, A. 2017. Emergence and clonal spread 1057 of colistin resistance due to multiple mutational mechanisms in carbapenemase-producing Klebsiella pneumoniae in London. Scientific Reports 7:12711.
- McManus PS, Stockwell VO, Sundin GW, Jones AL. 2002. Antibiotic use in plant agriculture. Annual Review of Phytopathology 40:443–465. [CrossRef]
- Stockwell VO, Sundin GW. Jones AL. 2002. Antibiotic use in plant agriculture. Annual Review of Phytopathology 40:443–465.
- Aćimović SG, Zeng Q, McGhee GC, Sundin GW, Wise JC. 2015. Control of fire blight (Erwinia amylovora) on apple trees with trunk-injected plant resistance inducers and antibiotics, and 764 assessment of induction of pathogenesis-related protein genes. Frontiers of Plant Science 6:16. [CrossRef]
- Förster H, McGhee GC, Sundin GW, Adaskaveg JE. 2015. Characterization of streptomycin resistance in isolates of Erwinia amylovora in California. Phytopathology 105:1302–1310. [CrossRef]
- Gusberti M, Klemm U, Meier MS, Maurhofer M, Hunger-Glaser I. 2015. Fire blight control: The struggle goes on. A comparison of different fire blight control methods in Switzerland with respect to biosafety, efficacy and durability. International Journal of Environmental Research and Public Health 12:11422–11447. [CrossRef]
- Jenssen H, Hamill P, Hancock RE. 2006. Peptide antimicrobial agents. Clinical Microbiology Review 19:491–511. [CrossRef]
- Ötvös, L. Jr. D. Wade, J. D. 2014. Current challenges in peptide-based drug discovery. Specialty Grand Challenge Article Frontiers in Chemistry 2, 62. [CrossRef]
- Mojsoska B, Jenssen H. 2015. Peptides and Peptidomimetics for Antimicrobial Drug Design. Pharmaceuticals (Basel). 2015 Jul 13; 8:366-415. [CrossRef]
- Kosikowska P, Lesner A.2016. Antimicrobial peptides (AMPs) as drug candidates: a patent review(2003–2015) Expert Opinion on Therapeutic Patents, 26:689-702. [CrossRef]
- Fosgerau K, Hoffmann T. Peptide therapeutics: current status and future directions. Drug Discov Today. 2015 Jan;20(1):122-8. [CrossRef] [PubMed]
- Sharma, K.; Sharma, K.K.; Sharma, A.; Jain, R. Peptide-based drug discovery: Current status and recent advances. Drug Dis-cov. Today 2023, 28, 103464. [Google Scholar] [CrossRef]
- Vallenet D, Belda E, Calteau A, Cruveiller S, Engelen S, Lajus A, Le Fèvre F, Longin C, Mornico D, Roche D, Rouy Z, Salvignol G, Scarpelli C, Thil Smith AA, Weiman M, Médigue C. 2013. MicroScope—an integrated microbial resource for the curation and comparative analysis of genomic and metabolic data. Nucleic Acids Research 41:D636 –D647. [CrossRef]
- Bode E, Brachmann AO, Kegler C, Simsek R, Dauth C, Zhou Q, Kaiser M, Klemmt P, Bode HB. 2015a. Simple “on-demand” production of bioactive natural products. ChemBioChem 16:1115–1119.
- Akhurst, RJ. 1982. Antibiotic activity of Xenorhabdus spp., bacteria symbiotically associated with 767 insect pathogenic nematodes of the families Heterorhabditidae and Steinernematidae. J GenMicrobiol.128:3061-3065. [CrossRef] [PubMed]
- Forst S, Nealson K. Molecular biology of the symbiotic-pathogenic bacteria Xenorhabdus spp. and Photorhabdus spp. Microbiol Rev. 1996 Mar;60(1):21-43. [CrossRef] [PubMed]
- Vivas EI, Goodrich-Blair H. 2001. Xenorhabdus nematophilus as a model for host–bacterium interactions: rpoS is necessary for mutualism with nematodes. Journal of Bacteriology 183:4687–4693.
- Bode, HB. 2009. Entomopathogenic bacteria as a source of secondary metabolites. Current Opinions in Chemistry & Biology 13:224–230. doi.org/10.1016/j.cbpa.2009.02.037.
- Ogier JC, Pagès S, Frayssinet M, Gaudriault S. Entomopathogenic nematode-associated microbiota: from monoxenic paradigm to pathobiome. Microbiome. 2020 Feb 24;8(1):25. [CrossRef] [PubMed]
- Reimer D, Bode HB. 2014. A natural prodrug activation mechanism in the biosynthesis of non- ribosomal peptides. Natural Products Report 31:154-159. [PubMed]
- Park, D. , Ciezki K., van der Hoeven R., Singh S., Reimer D., Bode H.B., Forst S. 2009. Genetic analysis of xenocoumacin antibiotic production in the mutualistic bacterium Xenorhabdus nematophila. Molecular Microbiology 73:938–949. [CrossRef]
- Gualtieri M, Aumelas A, Thaler JO. 2009. Identification of a new antimicrobial lysine-rich cyclolipopeptide family from Xenorhabdus nematophila. Journal of Antibiotics 62:295–302.
- Houard J, Aumelas A, Noël T, Pages S, Givaudan A, Fitton-Ouhabi V, Villain-Guillot P, Gualtieri M. Cabanillasin, a new antifungal metabolite, produced by entomopathogenic Xenorhabdus cabanillasii JM26. J Antibiot (Tokyo). 2013 Oct;66(10):617-20. [CrossRef] [PubMed]
- Lengyel K, Lang E, Fodor A, Szállás E, Schumann P, Stackebrandt E. 2005. Description of four 978 novel species of Xenorhabdus, family Enterobacteriaceae: Xenorhabdus budapestensis sp. nov., 979 Xenorhabdus ehlersii sp. nov., Xenorhabdus innexi sp. nov., and Xenorhabdus szentirmaii sp. nov. Systematics of Applied Microbiology 28:115-122. Erratum in: Systematics of Applied Microbiology 30:83; also in March/April 2014 Volume 2 Issue 2 e00190-14 Genome Announcements genomea.asm.org. Xenorhabdus budapestensis sp. Nov., Xenorhabdus ehlersii sp. Nov., Xenorhabdus innexi sp. Nov., and Xenorhabdus szentirmaii sp. Nov. Systematics of Applied Microbiology 28:115–122. [CrossRef]
- Furgani G, Böszörményi E, Fodor A, Máthé-Fodor A, Forst S, Hogan JS, Katona Z, Klein,MG, Stackebrandt E, Szentirmai A, Sztaricskai F, Wolf SL. 2008. Xenorhabdus antibiotics: a 890 comparative analysis and potential utility for controlling mastitis caused by bacteria. Journal of 891 Applied Microbiology 104:745–758. [CrossRef]
- Böszörményi E, Érsek T, Fodor A, Fodor AM, Földes LS, Hevesi M, Hogan JS, Katona Z, 791 Klein MG, Kormány A, Pekár S, Szentirmai A, Sztaricskai F, Taylor RA. 2009. Isolation 792 and activity of Xenorhabdus antimicrobial compounds against the plant pathogens Erwinia amylovora and Phytophthora nicotianae. Journal of Applied Microbiology 107:746–759. [CrossRef]
- Fodor A, Gualtieri M, Zeller M, Tarasco E, Klein MG, Fodor AM, Haynes L, Lengyel K, Forst SA, Furgani GM, Karaffa L, Vellai T. Type Strains of Entomopathogenic Nematode-Symbiotic Bacterium Species, Xenorhabdus szentirmaii (EMC) and X. budapestensis (EMA), Are Exceptional Sources of Non-Ribosomal Templated, Large-Target-Spectral, Thermotolerant-Antimicrobial Peptides (by Both), and Iodinin (by EMC). Pathogens. 2022 Mar 11;11(3):342. [CrossRef] [PubMed]
- Gualtieri M, Ogier J-C, Pagès S, Givaudan A, Gaudriault S. 2014. Draft genome sequence and annotation of the entomopathogenic\ bacterium Xenorhabdus szentirmaii Strain DSM16338. Genome Announcements 2(2): e00190-14 genomea.asm.org 1).
- Fuchs, S.W.; Sachs, C.C.; Kegler, C.; Nollmann, F.I.; Karas, M.; Bode, H.B. Neutral loss fragmentation pattern based screening for arginine-rich natural products in Xenorhabdus and Photorhabdus. Anal. Chem. 2012, 84, 6948–6955 [CrossRef]. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.W. Grundmann, F.; Kurz, M.; Kaiser, M.; Bode, H.B. Fabclavines: Bioactive peptide-polyketide-polyamine hybrids from Xenorhabdus. Chembiochem 2014, 15, 512–516 [CrossRef]. [Google Scholar] [CrossRef] [PubMed]
- Wenski, S.L.; Kolbert, D.; Grammbitter, G.L.C.; Bode, H.B. Fabclavine biosynthesis in X. szentirmaii: Shortened derivatives and characterization of the thioester reductase FclG and the condensation domain-like protein FclL. J. Ind. Microbiol. Biotechnol. 2019, 46, 565–572 [CrossRef]. [Google Scholar] [CrossRef] [PubMed]
- Wenski, S.L.; Cimen, H.; Berghaus, N.; Fuchs, S.W.; Hazir, S.; Bode, H.B. Fabclavine diversity in Xenorhabdus bacteria. Fabclavine diversity in Xenorhabdus bacteria. Beilstein J. Org. Chem. 2020, 16, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Fodor A, Fodor AM, Forst S, Hogan J, Hevesi M, Klein MG, Stackebrandt E, Szentirmai A, Sztaricskai F. 2007. New aspects of Xenorhabdus research. In: Abstracts of the 11th Europea Meeting of IOBC.WPRS Working Group “Insect Pathogens and Insect Parasitic Nematodes” in association with COST 862 Bacterial toxins for insect control” Ales (Gard) France, June 03-07 2007.
- Fodor A, Fodor AM, Forst S, Hogan JS, Klein MG, Lehoczky É. 2010. Comparative analysis of antibacterial activities of Xenorhabdus species on related and non-related bacteria in vivo. Journal of Microbiology and Antimicrobials 2:30–35.
- Fodor A, Varga I, Hevesi M, Máthé-Fodor A, Racsko J, Hogan JA. 2012. Novel anti-microbial peptides of Xenorhabdus origin against multidrug resistant plant pathogens, In: Bobbarala, V. (Ed.): Biochemistry, Genetics and Molecular Biology - A Search for Antibacterial Agents, 9:147–196.
- Vozik D, Bélafi-Bakó K, Hevesi M, Böszörményi E, Fodor A. 2015. Effectiveness of a peptide-rich fraction from Xenorhabdus budapestensis culture against fire blight disease on apple blossoms. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 43:547-553. [CrossRef]
- Nester, EW. 2015. Agrobacterium: nature's genetic engineer. Front Plant Sci. 2015 Jan 6;5:730. [CrossRef]
- Wood DW, Setubal JC, Kaul R, Monks DE, Kitajima J, Vagner P, Okura K, Zhou Y, Chen L, Wood GE, Almeida NF Jr., Woo L, Chen Y, Paulsen IT, Eisen JA, Karp PD, Bovee D, Sr., Chapman P, Clendenning J, Deatherage G, Gillet W, Grant C, Kutyavin T, Levy R, Li M-J, McClelland E, Palmieri A, Raymond C, Rouse G, Saenphimmachak C, Wu Z, RomeroP, Gordon D, Zhang S, Yoo H, Tao Y, Biddle P, Jung M, Krespan W, Perry M, Gordon- Kamm B, Liao L, Kim S, Hendrick C, Zhao Z-Y, Dola M, Chumley F, Tingey SV, Tomb JF, Gordon MP, Olson MV, Nester EW. 2001. The genome of the natural genetic engineer Agrobacterium tumefaciensC58. Science 294:2317–2323. [CrossRef]
- Henkel CV, Dulk-Ras A, Zhang X, Hooykaas PJJ. 2014. Genome sequence of the octopine-type Agrobacterium tumefaciens Strain Ach5. Genome Announcements 2: e00225-14.
- Van Larebeke N, Engler G, Holsters M, Van den Elsacker S, Zaenen I, Schilperoort RA, Schell J. 1974. Large plasmid in Agrobacterium tumefaciens essential for crown gall-inducing ability. Nature 252:169–170. (08 November 1974). [CrossRef]
- Currier TC, Nester EW. 1976. Evidence for diverse types of large plasmids in tumor- inducing strains of Agrobacterium. Journal of Bacteriology 126:157–165.
- Schell J, Van Montagu M. The Ti-plasmid of Agrobacterium tumefaciens, a natural vector for the introduction of nif genes in plants? Basic Life Sci. 1977;9:159-79. [CrossRef] [PubMed]
- Hooykaas PJJ. The Ti Plasmid, Driver of Agrobacterium Pathogenesis. Phytopathology. 2023 Apr;113(4):594-604. [CrossRef] [PubMed]
- Chilton M-D, Drummond M, Merlo D, Sciaky D, Montoya A, Gordon M, Nester E. 1977. Stable incorporation of plasmid DNA into higher plant cells: the molecular basis of crown gall tumorigenesis. Cell 11:263–271.
- Koncz Cs, DeGreve H, Andre D, Deboeck F, Van Montagu M, Schell J. 1983. The opine synthase genes carried by Ti plasmids contain all signals necessary for expression in plants. EMBO Journal 2:1597–1603.
- Montoya A, Chilton M-D, Gordon MP, Sciaky D, Nester EW. 1977. Octopine and nopaline metabolism in Agrobacterium tumefaciens and crown gall tumor cells: role of plasmid genes. Journal of Bacteriology 129:101–107.
- Guyon P, Chilton M-D, Petit A, Tempo J. 1980. Agropine in "null-type" crown gall tumors: Evidence for generality of the opine concept. Proceedings of the National Academy of Science USA 77:2693–2697.
- Tremblay G, Gagliardo R, Chilton WS, Dion P. Diversity among Opine-Utilizing Bacteria: Identification of Coryneform Isolates. Appl Environ Microbiol. 1987 Jul;53(7):1519-24. [CrossRef] [PubMed]
- Hoekema A, Hirsch PR, Hooykaas PJJ, Schilperoort RA. 1983. A binary plant vector strategy based on separation of vir-and T-region of the Agrobacterium tumefaciens Ti-plasmid. Nature 926 303:179–180.
- Koekman BP, Ooms G, Klapwijk PM, Schilperoort RA. 1979. Genetic map of an octopine Ti-plasmid. Plasmid 2:346–357.
- Klapwijk PM, Schilperoort RA 1979. Negative control of octopine degradation and transfer genes of octopin Ti plasmids in Agrobacterium tumefaciens. Journal of Bacteriology 132:424-431.
- Ooms G, Klapwijk PM, Poulis JA, Schilperoort RA. 1980. Characterization of Tn904 insertions in octopine Ti-plasmid mutants of Agrobacterium tumefaciens. Journal of Bacteriology 144:82– 1045.
- Ooms G, Hooykaas PJJ, Moolenaar G, Schilperoort RA. 1981. Crown gall tumors of abnormal morphology, induced by Agrobacterium tumefaciens carrying mutated octopine Ti-plasmids: Analysis of T-DNA functions. Gene 14:33–50.
- Ooms G, Hooykaas PJJ, Van Veen RJM, Van Beelen P, Regensburg-Tuink AJG, Schilperoort RA. 1982. Octopine Ti plasmid deletion mutants of Agrobacterium tumefaciens with emphasis 1053 on the right side of the T-region. Plasmid 7:15-29.
- Jen GC, Chilton M-D. 1986. Activity of T-DNA borders in plant cell transformation by mini-Ti Plasmids. Journal of Bacteriology 166:491–499.
- Ausubel FM, Brent R, Kingston RE, Moor DD, Seidman JG, Smith JA, Struhl K. (Eds.) 1999. Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology. pp. 1-1 – 1-6; 5-1 – 6-30. John Wiley & Sons, New York.
- Leclerc MC, Boemare NE. (1991). Plasmids and phase variation in Xenorhabdus spp. Applied Environmental Microbiology 57:2597–3601.
- Wise AA, Liu Z, Binns AN. 2006. Culture and maintenance of Agrobacterium strains. Methods in Molecular Biology 343:3–14. Review.
- Hood EE, Helmer GL, Fraley RT, Chilton M.-D. 1986. The hypervirulence of Agrobacterium tumefaciens A281 is encoded in a region of pTiBo542 outside of T-DNA. Journal of Bacteriology 168:1291–1301.
- Lazo GR, Stein PA, Ludwig RA. 1991. A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. Biotechnology (N Y) 9: 963–967.
- Hood EE, Gelvin SB, Melchers LS, Hoekema A. 1993. New Agrobacterium helper plasmids for gene transfer to plants. Transgenic Research 2:208–218.
- White FF, Nester EW. Hairy root: plasmid encodes virulence traits in Agrobacterium rhizogenes. J Bacteriol. 1980 Mar;141(3):1134-41. [CrossRef] [PubMed]
- Petit A, David C, Dahl GA, Ellis JG, Guyon P, Casse-Delbart F, Tempe J. 1982. Further extension of the opine concept: Plasmids in Agrobacterium rhizogenes cooperate for opine degradation. Molecular and General Genetics 190:204–214.
- Jouanin L, Tourneur J, Tourneur C, Casse-Delbart F. 1986. Restriction maps and homologies of the three plasmids of Agrobacterium rhizogenes Strain A4. Plasmid 16:124–134.
- Slater SC, Goldman BS, Goodner B, Setubal JC, Farrand SK, Nester EW, Burr TJ, Banta L, Dickerman AW, Paulsen I, Otten L, Suen G, Welch R, Almeida NF, Arnold F, Burton OT, Du Z, Ewing A, Godsy E, Heisel S, Houmiel KL, Jhaveri J, Lu J, Miller NM, Norton S, Chen Q, Phoolcharoen W, Ohlin V, Ondrusek D, Pride N, Stricklin SL, Sun J, Wheeler C, Wilson L, Zhu, H, Wood DW. 2009. Genome sequences of three Agrobacterium biovars help elucidate the evolution of multichromosome genomes in bacteria. Journal of Bacteriology 191:2501–2511. [CrossRef]
- Hood EE, Helmer GL, Fraley RT, Chilton MD. 1987. Virulence of Agrobacterium tumefaciens strain A281 on legumes. Plant Physiology 83:529–534.
- Komari T, Halperin W, Nester EW. 1986. Physical and functional map of supervirulent Agrobacterium tumefaciens tumor-inducing plasmid pTiBoS42. Journal of Bacteriology 166:88–94.
- Jin S, Komari T, Milton P, Gordon MP, Nester EW. 1987. Genes Responsible for the supervirulence Phenotype of Agrobacterium tumefaciens A281. Journal of Bacteriology 169:4417–4425.
- Bevan M, 1984. Binary Agrobacterium vectors for plant transformation. Nucleic Acid Research 12:8711–8721.
- Taylor CG, Fuchs B, Collier R, Lutke WK. 2006. Generation of composite plants using Agrobacterium rhizogenes. Methods in Molecular Biology 343:155–167. Review.
- Uraji M, Suzuki K, Yoshida K. 2002. A novel plasmid curing method using incompatibility of plant pathogenic Ti plasmids in Agrobacterium tumefaciens. Genes and Genetic Systematics 77:1–9.
- Koncz C, Schell J. 1986. The promoter of TL-DNA gene 5 controls the tissue-specific expression of 960 chimaeric genes carried by a novel type of Agrobacterium binary vector. Molecular and General Genetics 204:383–396.
- Dessaux Y, Guyon P, Petit A, Tempe J, Demarez M, Legrain C, Tate ME, Farrand SK. 1988. 830 Opine utilization by Agrobacterium spp.: Octopine-type Ti plasmids encode two pathways for mannopinic acid degradation. Journal of Bacteriology 170:2939–2946.
- Jandera, P. Gradient elution in normal-phase high-performance liquid chromatographic systems. J Chromatogr A. 2002 Aug 2;965(1-2):239-61. [CrossRef] [PubMed]
- Josic D, Kovac S. Reversed-phase High Performance Liquid Chromatography of proteins. Curr Protoc Protein Sci. 2010 Aug;Chapter 8:8.7.1-8.7.22. [CrossRef] [PubMed]
- Bowen DJ, Ensign JC. 1998. Purification and characterization of a high-molecular-weight insecticidal protein complex produced by the entomopathogenic bacterium Photorhabdus luminescens. Applied Environmental Microbiology 64:3029–3035. [PubMed]
- Bowen DJ, Ensign JC. 2001. Isolation and characterization of intracellular protein inclusions produced by the entomopathogenic bacterium Photorhabdus luminescens. Applied Environmental Microbiology 67:4834–4841.
- Ciche TA, Blackburn M, Carney JR, Ensign JC. Photobactin: a catechol siderophore produced by Photorhabdus luminescens, an entomopathogen mutually associated with Heterorhabditis bacteriophora NC1 nematodes. Appl Environ Microbiol. 2003 Aug;69(8):4706-13. [CrossRef] [PubMed]
- Carr D 2002. The Handbook of Analysis and. Purification of Peptides and. Proteins by Reversed- Phase HPLC. Presented by Vydac (The Separations Group). 17434 Mojave Street. Hesperia CA 92345 USA.
- Wiegand, Hilpert & Hancock, 2008; Clinical and Laboratory Standards Institute (CLSI, 2012). M07: Methods for Dilution Antimicrobial Susceptibility Tests... Clinical & Laboratory Standards Institute. https://clsi.org media.
- Incedayi G, Cimen H, Ulug D, Touray M, Bode E, Bode HB, Orenlili Yaylagul E, Hazir S, Cakmak I. Relative potency of a novel acaricidal compound from Xenorhabdus, a bacterial genus mutualistically associated with entomopathogenic nematodes. Sci Rep. 2021 ;11(1):11253. 27 May. [CrossRef] [PubMed]
- Yadav M, Rathore JS. The hipBAXn operon from Xenorhabdus nematophila functions as a bonafide toxin-antitoxin module. Appl Microbiol Biotechnol. 2020 Apr;104(7):3081-3095. [CrossRef] [PubMed]
- Morales-Soto N, Forst SA. The xnp1 P2-like tail synthesis gene cluster encodes xenorhabdicin and is required for interspecies competition. J Bacteriol. 2011 Jul;193(14):3624-32. [CrossRef] [PubMed]
- Völgyi, A. , Fodor A, Forst S. 2000. Inactivation of a novel gene produces a phenotypic variant cell and affects the symbiotic behavior of Xenorhabdus nematophilus. Applied and Environmental Microbiology. 66:1622-1628.
- Cowles KN, Cowles CE, Richards GR, Martens EC, Goodrich-Blair H. 2007. The global regulator Lrp contributes to mutualism, pathogenesis, and phenotypic variation in the bacterium Xenorhabdus nematophila. Cellular Microbiology 9:1311-1323. 2007.
- Nuonming P, Khemthong S, Dokpikul T, Sukchawalit R, Mongkolsuk S. Characterization and regulation of AcrABR, a RND-type multidrug efflux system, in Agrobacterium tumefaciens C58. Microbiol Res. 2018 Sep;214:146-155. [CrossRef] [PubMed]
- Watanabe K, Stringer S, Frei O, Umićević Mirkov M, de Leeuw C, Polderman TJC, van der Sluis S, Andreassen OA, Neale BM, Posthuma D. A global overview of pleiotropy and genetic architecture in complex traits. Nat Genet. 2019 Sep;51(9):1339-1348. [CrossRef] [PubMed]
- Ward RD, Tran JS, Banta AB, Bacon EE, Rose WE, Peters JM. Essential Gene Knockdowns Reveal Genetic Vulnerabilities and Antibiotic Sensitivities in Acinetobacter baumannii. bioRxiv [Preprint]. 2023 Aug 2:2023.08.02.551708. [CrossRef] [PubMed]
- Rodrigues SD, Karimi M, Impens L, Van Lerberge E, Coussens G, Aesaert S, Rombaut D, Holtappels D, Ibrahim HMM, Van Montagu M, Wagemans J, Jacobs TB, De Coninck B, Pauwels L. Efficient CRISPR-mediated base editing in Agrobacterium spp. Proc Natl Acad Sci U S A. 2021 Jan 12;118(2):e2013338118. [CrossRef] [PubMed]
- Elston RC, Satagopan J, Sun S. Statistical Genetic Terminology. Methods Mol Biol. 2017;1666:1-9. [CrossRef] [PubMed]
- Kiyokawa K, Yamamoto S, Sakuma K, Tanaka K, Moriguchi K, Suzuki K. Construction of disarmed Ti plasmids transferable between Escherichia coli and Agrobacterium species. Appl Environ Microbiol. 2009 Apr;75(7):1845-51. [CrossRef] [PubMed]
- Stachel SE, Nester EW. The genetic and transcriptional organization of the vir region of the A6 Ti plasmid of Agrobacterium tumefaciens. EMBO J. 1986 Jul;5(7):1445-54. [CrossRef] [PubMed]
- Palanichelvam K, Oger P, Clough SJ, Cha C, Bent AF, Farrand SK. A second T-region of the soybean-supervirulent chrysopine-type Ti plasmid pTiChry5, and construction of a fully disarmed vir helper plasmid. Mol Plant Microbe Interact. 2000 Oct;13(10):1081-91. [CrossRef] [PubMed]
- Hattori Y, Iwata K, Suzuki K, Uraji M, Ohta N, Katoh A, Yoshida K. Sequence characterization of the vir region of a nopaline type Ti plasmid, pTi-SAKURA. Genes Genet Syst. 2001 Apr;76(2):121-30. [CrossRef] [PubMed]
- Lacroix B, Citovsky V. Transfer of DNA from Bacteria to Eukaryotes. mBio. 2016 Jul 12;7(4):e00863-16. [CrossRef] [PubMed]
- Schröder G, Klipp W, Hillebrand A, Ehring R, Koncz C, Schröder J. The conserved part of the T-region in Ti-plasmids expresses four proteins in bacteria. EMBO J. 1983;2(3):403-9. [CrossRef] [PubMed]
- Li YG, Hu B, Christie PJ. Biological and Structural Diversity of Type IV Secretion Systems. Microbiol Spectr. 2019 Mar;7(2):10.1128/microbiolspec.PSIB-0012-2018. [CrossRef] [PubMed]
- Ward JE Jr, Dale EM, Christie PJ, Nester EW, Binns AN. Complementation analysis of Agrobacterium tumefaciens Ti plasmid virB genes by use of a vir promoter expression vector: virB9, virB10, and virB11 are essential virulence genes. J Bacteriol. 1990 Sep;172(9):5187-99. [CrossRef] [PubMed]
- Bitrián M, Roodbarkelari F, Horváth M, Koncz C. BAC-recombineering for studying plant gene regulation: developmental control and cellular localization of SnRK1 kinase subunits. Plant J. 2011 Mar;65(5):829-42. [CrossRef] [PubMed]
- Wang X, Teng C, Wei H, Liu S, Xuan H, Peng W, Li Q, Hao H, Lyu Q, Lyu S, Fan Y. Development of a set of novel binary expression vectors for plant gene function analysis and genetic transformation. Front Plant Sci. 2023 Jan 12;13:1104905. [CrossRef] [PubMed]
- Christie, PJ. 2004. Type IV secretion: the Agrobacterium VirB/D4 and related conjugation systems. Biochimica et Biophysica Acta 1694:219–234. [CrossRef]
- Vergunst AC, Schrammeijer B, den Dulk-Ras A, de Vlaam CM, Regensburg-Tuïnk TJ, Hooykaas PJ. VirB/D4-dependent protein translocation from Agrobacterium into plant cells. Science. 2000 Nov 3;290(5493):979-82. [CrossRef] [PubMed]
- Vergunst AC, van Lier MC, den Dulk-Ras A, Stüve TA, Ouwehand A, Hooykaas PJ. Positive charge is an important feature of the C-terminal transport signal of the VirB/D4-translocated proteins of Agrobacterium. Proc Natl Acad Sci U S A. 2005 Jan 18;102(3):832-7. [CrossRef] [PubMed]
- Koncz C, Kreuzaler F, Kalman Z, Schell J. A simple method to transfer, integrate and study expression of foreign genes, such as chicken ovalbumin and alpha-actin in plant tumors. EMBO J. 1984 May;3(5):1029-37. [CrossRef] [PubMed]
Figure 1.
Antimicrobial Activity of EMA_PF2 on E. coli and Xenorhabdus strains. Results of Liquid Culture Bioassays. 1A The distribution of the OD values (Means +/- StDev) measured in 24-h cultures of Escherichia coli (strains HGB1333 and HGB2226, grown at 37 oC) and those of Xenorhabdus budapestensis (HG2338), X. szentirmaii (HGB2339) and X. nematophila HGB1789 wild-type strains (grown at 28 oC) demonstrate the strong anti-Gram-negative activity of the antimicrobial peptide (AMP) fraction EMA_PF2 isolated from the cell-free-culture of X. budapestensis (HGB033) against which not only the cells of the EMA-producing species, X. budapestensis, but those the wild-type strains of X. szentirmaii and X. nematophila cells are completely protected. (Not all data are given, but each tested X. nematophila strains, including X. nematophila Types Strain ATTC 19061 deposited in Madison as well, as HGB800; and var-1 [115] (Völgyi, Fodor and Forst, 2002); all tested lap mutants characteristically resembling secondary form strains [116] (Cowles et al., 2007) were unambiguously resistant to EMA_PF, [67] Vozik et al., 2015). Strain HGB1795, however, which had been isolated as a Tn insertion mutant from the spontaneous rifampicin-resistant X. nematophila strain HGB081 (of H. Goodrich – Blair, a spontaneous mutant a derivative of X. nematophila Types Strain ATTC 19061, see Materials and Methods) proved as sensitive as the examined E. coli strains. To confirm this interesting discovery, we repeated the test in an independent experiment, in which HGB1795 was treated with another preparation of EMA_PF2, in comparison with the original HGB081 deposited in the laboratory of Prof. H. Goodrich-Blair and that clone of this mutant from which H. Bode isolated a transposon-induced insertion mutant of the XNC1_2022 gene what was then deposited in the Laboratory of H. Goodrich- Blair in Madison, WI as HGB1795, see data on Figure 1 B. When we later asked for a new sample from this strain in 2019, he request was refused.
Figure 1.
Antimicrobial Activity of EMA_PF2 on E. coli and Xenorhabdus strains. Results of Liquid Culture Bioassays. 1A The distribution of the OD values (Means +/- StDev) measured in 24-h cultures of Escherichia coli (strains HGB1333 and HGB2226, grown at 37 oC) and those of Xenorhabdus budapestensis (HG2338), X. szentirmaii (HGB2339) and X. nematophila HGB1789 wild-type strains (grown at 28 oC) demonstrate the strong anti-Gram-negative activity of the antimicrobial peptide (AMP) fraction EMA_PF2 isolated from the cell-free-culture of X. budapestensis (HGB033) against which not only the cells of the EMA-producing species, X. budapestensis, but those the wild-type strains of X. szentirmaii and X. nematophila cells are completely protected. (Not all data are given, but each tested X. nematophila strains, including X. nematophila Types Strain ATTC 19061 deposited in Madison as well, as HGB800; and var-1 [115] (Völgyi, Fodor and Forst, 2002); all tested lap mutants characteristically resembling secondary form strains [116] (Cowles et al., 2007) were unambiguously resistant to EMA_PF, [67] Vozik et al., 2015). Strain HGB1795, however, which had been isolated as a Tn insertion mutant from the spontaneous rifampicin-resistant X. nematophila strain HGB081 (of H. Goodrich – Blair, a spontaneous mutant a derivative of X. nematophila Types Strain ATTC 19061, see Materials and Methods) proved as sensitive as the examined E. coli strains. To confirm this interesting discovery, we repeated the test in an independent experiment, in which HGB1795 was treated with another preparation of EMA_PF2, in comparison with the original HGB081 deposited in the laboratory of Prof. H. Goodrich-Blair and that clone of this mutant from which H. Bode isolated a transposon-induced insertion mutant of the XNC1_2022 gene what was then deposited in the Laboratory of H. Goodrich- Blair in Madison, WI as HGB1795, see data on Figure 1 B. When we later asked for a new sample from this strain in 2019, he request was refused.
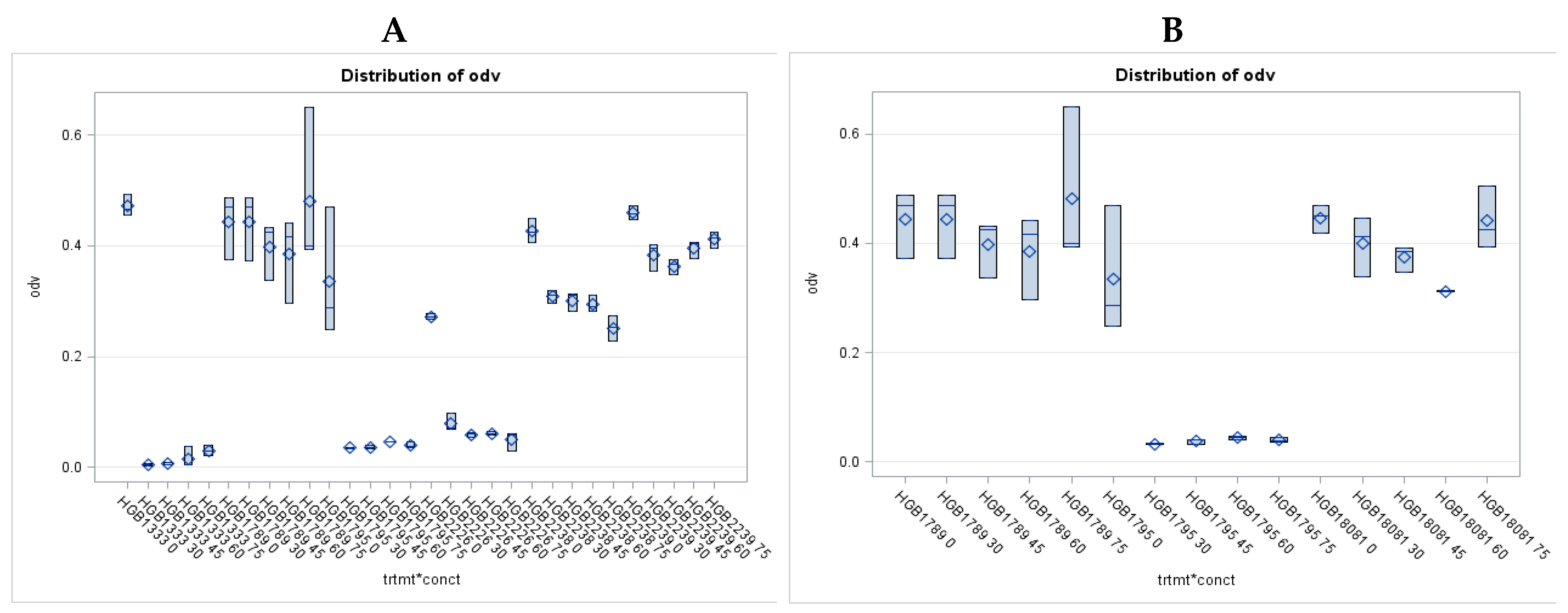
Figure 2.
EMA_PF2 is peptide- fraction (EMA_PF2), which had been isolated from cell-free culture media (CFCM) of X. budapestensis (EMA) by Amberlite XAD1148® adsorption; purified followed by gradient MeOH elution; and eluted by with 99V/V% of methanol, followed by sterile ultrafiltration (Vozik et al., 2015). HPLC peaks were detected at 168 -215 nm and 168-280nm, respectively. When tested in Agar Diffusion Bioassays Fractions collected from below the third (A2) peak (between 40-45 min intervals) exerted strong antimicrobial activities on both Gram-positive (Staphylococcus aureus, strain JE) and Gram-negative (Escherichia coli strain HGB2226) test organisms, (see Table 2) and also on Candida albicans (not shown). Fractions collected from below the peaks at the first B2, (5-10 min), and the second, B1. (18-30 min) intervals showed no antimicrobial activity on either target. This experiment was carried out in the Laboratory of Professor Emeritus Jerald C. Ensign using his equipment, solutions, and standard methods, Department of Bacteriology, University of Madison, Linden Drive, Madison WI, and the USA. MALDI analysis of the most active fraction pooled samples from below the A2 peak three peaks are presented in Figure 3A.
Figure 2.
EMA_PF2 is peptide- fraction (EMA_PF2), which had been isolated from cell-free culture media (CFCM) of X. budapestensis (EMA) by Amberlite XAD1148® adsorption; purified followed by gradient MeOH elution; and eluted by with 99V/V% of methanol, followed by sterile ultrafiltration (Vozik et al., 2015). HPLC peaks were detected at 168 -215 nm and 168-280nm, respectively. When tested in Agar Diffusion Bioassays Fractions collected from below the third (A2) peak (between 40-45 min intervals) exerted strong antimicrobial activities on both Gram-positive (Staphylococcus aureus, strain JE) and Gram-negative (Escherichia coli strain HGB2226) test organisms, (see Table 2) and also on Candida albicans (not shown). Fractions collected from below the peaks at the first B2, (5-10 min), and the second, B1. (18-30 min) intervals showed no antimicrobial activity on either target. This experiment was carried out in the Laboratory of Professor Emeritus Jerald C. Ensign using his equipment, solutions, and standard methods, Department of Bacteriology, University of Madison, Linden Drive, Madison WI, and the USA. MALDI analysis of the most active fraction pooled samples from below the A2 peak three peaks are presented in Figure 3A.
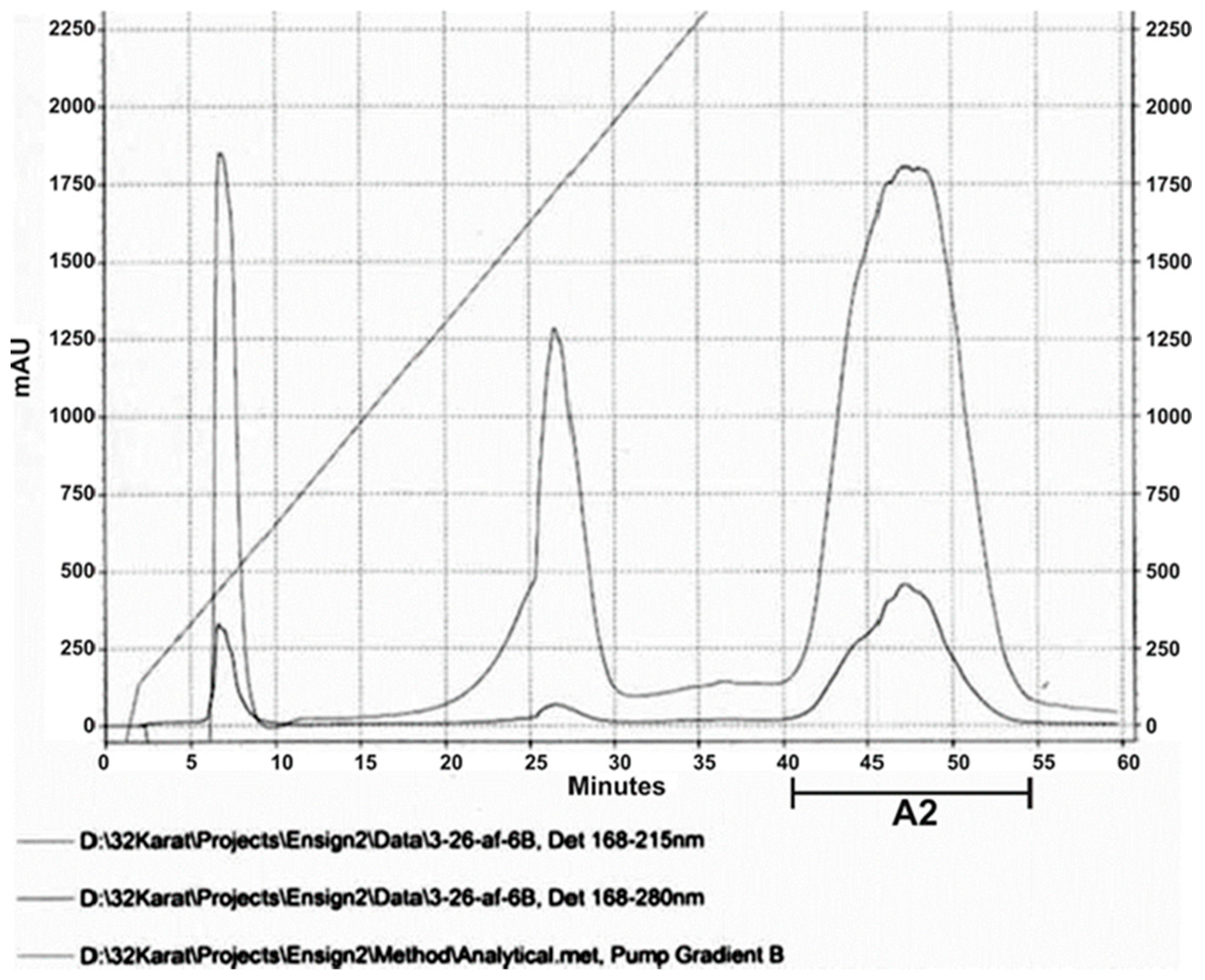
Figure 3.
Comparison of the MALDI profiles of two antimicrobial active preparations from the cell-free culture medium (CFCM) of Xenorhabdus budapestensis (EMA PF2 (HPLC) A2 and EMA RFLP AF30) AF103 Fr.43. The complete MALDI profile of the pooled fractions from below A2 peak of the HPLC chromatogram (Figure 2) of the Amberlite-Adsorbed, MeOH-purified EMA_PF2 at three magnifications (not shown). Each peptide peak is < 2,200 m/Z. There are many peaks in the m/Z range, where Bicornutin A is expected to be present (500-1000) and there are many in the m/Z range of 1,200-1500, (lowermost chromatogram); but the majority are located between 1,300-1,400 (uppermost diagram). Figure 3A shows this region in more detail. Two arrows indicate the most densely populated sub-regions. The HPLC profile of purified peptide preparation obtained by column chromatography (EMA30, (AF103), is very similar (not shown). The MALDI profile of the most active antimicrobial HPLC fraction, Fr43 of EMA30 (AF103) is rather similar (Figure 3B) but not identical to that of A2. The main difference is that only the left sub-region of the m/Z range in Figure 3C has peptide peaks, the right one, labeled with a red arrow is almost empty. The left molecular region is where fabclavine isomers [] (2012, [] Fuchs et al., 2014) are expected to be located, and this region is very similar to Figure 3B and 3C. The m/Z regions where Bicornutin A is expected to be localized are very similar in 3B and 3C. This observation may be considered as an indirect confirmation that the predominant active antimicrobial peptide component of EMA AMP complexes is the fabclavine, but Bicornutin A (or similar peptides in other Xenorhabdus species, with of unknown role) may also be present in antimicrobial active bio-preparations). .
Figure 3.
Comparison of the MALDI profiles of two antimicrobial active preparations from the cell-free culture medium (CFCM) of Xenorhabdus budapestensis (EMA PF2 (HPLC) A2 and EMA RFLP AF30) AF103 Fr.43. The complete MALDI profile of the pooled fractions from below A2 peak of the HPLC chromatogram (Figure 2) of the Amberlite-Adsorbed, MeOH-purified EMA_PF2 at three magnifications (not shown). Each peptide peak is < 2,200 m/Z. There are many peaks in the m/Z range, where Bicornutin A is expected to be present (500-1000) and there are many in the m/Z range of 1,200-1500, (lowermost chromatogram); but the majority are located between 1,300-1,400 (uppermost diagram). Figure 3A shows this region in more detail. Two arrows indicate the most densely populated sub-regions. The HPLC profile of purified peptide preparation obtained by column chromatography (EMA30, (AF103), is very similar (not shown). The MALDI profile of the most active antimicrobial HPLC fraction, Fr43 of EMA30 (AF103) is rather similar (Figure 3B) but not identical to that of A2. The main difference is that only the left sub-region of the m/Z range in Figure 3C has peptide peaks, the right one, labeled with a red arrow is almost empty. The left molecular region is where fabclavine isomers [] (2012, [] Fuchs et al., 2014) are expected to be located, and this region is very similar to Figure 3B and 3C. The m/Z regions where Bicornutin A is expected to be localized are very similar in 3B and 3C. This observation may be considered as an indirect confirmation that the predominant active antimicrobial peptide component of EMA AMP complexes is the fabclavine, but Bicornutin A (or similar peptides in other Xenorhabdus species, with of unknown role) may also be present in antimicrobial active bio-preparations). .
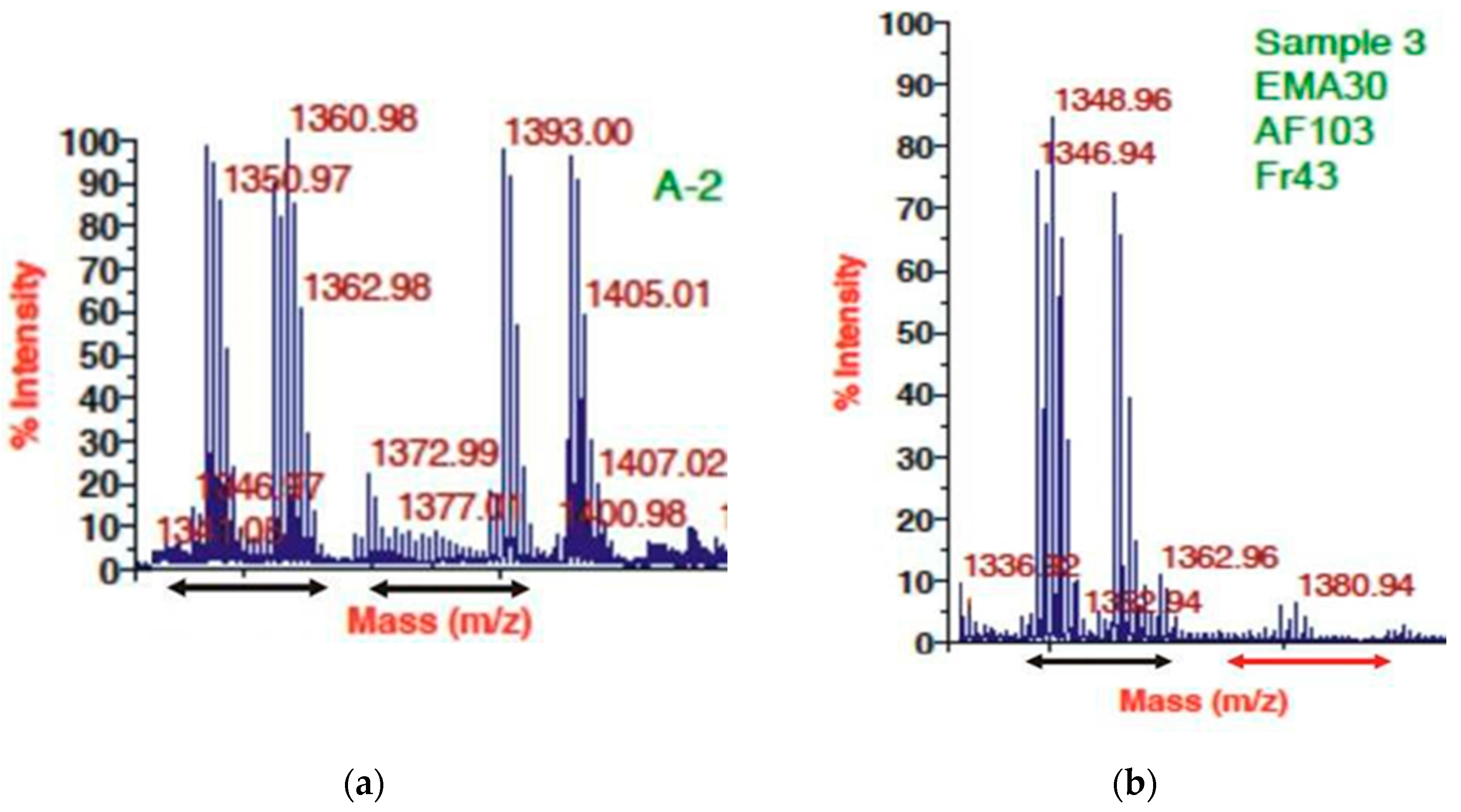
Figure 4.
Comparison of the antimicrobial activity of peptide fractions separated from HPLC from EMA(30) and tested on 4 different targets in Agar Diffusion Bioassays. Test organisms: SA = S. aureus JE; as a Gram-positive; EC= E. coli HGB2226; as a Gram-negative target; CA= Candida albicans JE, as a fungal target; HGB1795: an insertion mutant of X. nematophila with extreme sensitivity to Xenorhabdus antimicrobials. Columns of holes: the places into which 0.1 ml of samples were pipetted. from left to right: sample collected in the 19th, 21st, 23rd, 25th, 40th, 43rd, 44th, 45th, 49th and 57th min of HPLC run. Each hole had been filled with 100 µl volume of a sample, overlaid with the uniformly soft-agar diluted cell-suspension of the respective test organism. Samples (holes) from left to right Left plates: fractions collected at retention time 19, 21, 23, and 25 from below the first peak of HPLC run), Plates in the middle: fractions collected at the 40, 43, and 44 minutes retention times; from below the largest sharp peak of HPLC run, Right plates: fractions collected at the 45th, 47th, 53rd and 57th minutes from below the last large, not-very-sharp peak (at the end of the chromatogram). From top to bottom: Sensitive test organisms: CA =Candida albicans, SA. = Staphylococcus aureus, EC =Escherichia coli HGB2226, X. nematophila HGB 1795, two replicates of each.
Figure 4.
Comparison of the antimicrobial activity of peptide fractions separated from HPLC from EMA(30) and tested on 4 different targets in Agar Diffusion Bioassays. Test organisms: SA = S. aureus JE; as a Gram-positive; EC= E. coli HGB2226; as a Gram-negative target; CA= Candida albicans JE, as a fungal target; HGB1795: an insertion mutant of X. nematophila with extreme sensitivity to Xenorhabdus antimicrobials. Columns of holes: the places into which 0.1 ml of samples were pipetted. from left to right: sample collected in the 19th, 21st, 23rd, 25th, 40th, 43rd, 44th, 45th, 49th and 57th min of HPLC run. Each hole had been filled with 100 µl volume of a sample, overlaid with the uniformly soft-agar diluted cell-suspension of the respective test organism. Samples (holes) from left to right Left plates: fractions collected at retention time 19, 21, 23, and 25 from below the first peak of HPLC run), Plates in the middle: fractions collected at the 40, 43, and 44 minutes retention times; from below the largest sharp peak of HPLC run, Right plates: fractions collected at the 45th, 47th, 53rd and 57th minutes from below the last large, not-very-sharp peak (at the end of the chromatogram). From top to bottom: Sensitive test organisms: CA =Candida albicans, SA. = Staphylococcus aureus, EC =Escherichia coli HGB2226, X. nematophila HGB 1795, two replicates of each.

Figure 5.
Distribution of the OD values measured in in-vitro liquid bioassays of EMA_PF2 in Agrobacterium strains. Note that (the gross) MIC values could be determined only for 4 Agrobacterium strains, HP1837, H1839, (these should be repeated with on independent samples), A4T, and EHA105, not for the others, because they proved fully resistant. Consequently, the OD values detected at the different concentrations did not differ significantly from each other. Each of the sensitive strains was Δ –TDNA. All the others, which should be considered as resistant, seem resistant but to different degrees. For more details, see the Footer of Figure 5 (below). The mean values and standard deviations of OD values (odv) were obtained from 3 replicates. Distribution of the OD values measured in 24h liquid cultures of 0, 30, 45, 60, and 75 µg/ml dose of EMA_PF2. The odv of each strain (HP1836; HP1837; HP1838; HP1839; HP1840; HP1841; HP1842; HP1843; SZL1; SZL2; SZL3; SZL4) were determined at each concentration. Because of the space limit, not all data are noted on the abscissa. Data were analyzed by ANOVA procedure. Abbreviations: Odv = values of optical density (OD) determined spectroscopically, trmt*conct: ANOVA results of the strain (treatment) and dose (concentration) interaction (*); which is hardly different from those of the freshly inoculated LB culture control. These four data groups of OD values are those of the 30 and 60 µg/ml dose of EMA PF treated HP1837; HP1839; SZL1 and SZL3 cultures, respectively. These Agrobacterium strains are considered to be of S phenotype (sensitive to EMA PF), while the rest of the strains are R phenotype (resistant to EMA PF), even if they are not uniformly resistant. (For raw data see Supplementary material).
Figure 5.
Distribution of the OD values measured in in-vitro liquid bioassays of EMA_PF2 in Agrobacterium strains. Note that (the gross) MIC values could be determined only for 4 Agrobacterium strains, HP1837, H1839, (these should be repeated with on independent samples), A4T, and EHA105, not for the others, because they proved fully resistant. Consequently, the OD values detected at the different concentrations did not differ significantly from each other. Each of the sensitive strains was Δ –TDNA. All the others, which should be considered as resistant, seem resistant but to different degrees. For more details, see the Footer of Figure 5 (below). The mean values and standard deviations of OD values (odv) were obtained from 3 replicates. Distribution of the OD values measured in 24h liquid cultures of 0, 30, 45, 60, and 75 µg/ml dose of EMA_PF2. The odv of each strain (HP1836; HP1837; HP1838; HP1839; HP1840; HP1841; HP1842; HP1843; SZL1; SZL2; SZL3; SZL4) were determined at each concentration. Because of the space limit, not all data are noted on the abscissa. Data were analyzed by ANOVA procedure. Abbreviations: Odv = values of optical density (OD) determined spectroscopically, trmt*conct: ANOVA results of the strain (treatment) and dose (concentration) interaction (*); which is hardly different from those of the freshly inoculated LB culture control. These four data groups of OD values are those of the 30 and 60 µg/ml dose of EMA PF treated HP1837; HP1839; SZL1 and SZL3 cultures, respectively. These Agrobacterium strains are considered to be of S phenotype (sensitive to EMA PF), while the rest of the strains are R phenotype (resistant to EMA PF), even if they are not uniformly resistant. (For raw data see Supplementary material).
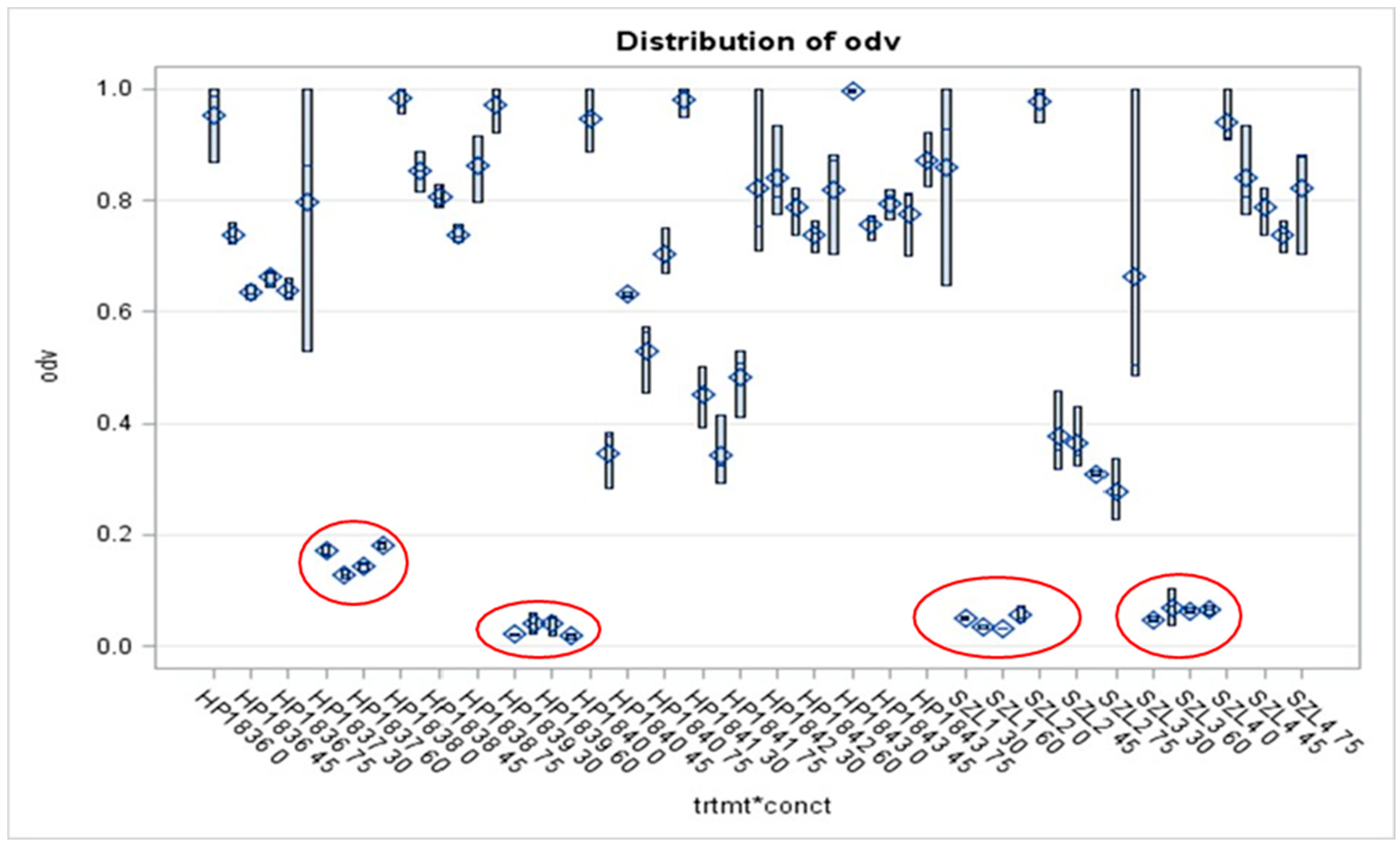
Figure 6.
Comparison of the sensitivities of the wild-type, virulent (A281) and a T-DNA deleted (Δ–TDNA) derivative (EHA105), detected in Agar Diffusion Bioassay. Both of them are L, L,- succinamopine, AGR) - catabolizing Agrobacterium strains. Agardfiffusion bioassay of sterile cell-free culture medium (EMA CFCM) of nematode-symbiotic bacterium Xenorhabdus budapestensis (EMA) on agropine—(L, L,-succinamoipine catabolizing (utilizing), AGR) Agrobacterium tumefaciens strains. Left (A) EHA105, a T-DNA deleted (∆ –T-DNA) strain. Right (B): A281, the virulent, wild-type (T-DNA (+) strain. Plasmid genotypes: EHA 105 (pTiBo542 (T-DNA(-) = (pEHA105); Nal Mop(+)). A281: (+) pT Bo542 (T-DNA(+) HP1838 (A281, right)) Agrobacterium strains. (Photo: Ahmed Nour El-Deen). .
Figure 6.
Comparison of the sensitivities of the wild-type, virulent (A281) and a T-DNA deleted (Δ–TDNA) derivative (EHA105), detected in Agar Diffusion Bioassay. Both of them are L, L,- succinamopine, AGR) - catabolizing Agrobacterium strains. Agardfiffusion bioassay of sterile cell-free culture medium (EMA CFCM) of nematode-symbiotic bacterium Xenorhabdus budapestensis (EMA) on agropine—(L, L,-succinamoipine catabolizing (utilizing), AGR) Agrobacterium tumefaciens strains. Left (A) EHA105, a T-DNA deleted (∆ –T-DNA) strain. Right (B): A281, the virulent, wild-type (T-DNA (+) strain. Plasmid genotypes: EHA 105 (pTiBo542 (T-DNA(-) = (pEHA105); Nal Mop(+)). A281: (+) pT Bo542 (T-DNA(+) HP1838 (A281, right)) Agrobacterium strains. (Photo: Ahmed Nour El-Deen). .

Figure 7.
OD values of the wild-type (a) (A281) wild-type and the disarmed Ti plasmid bearing (b) A4T) agropine (the L, L,-succinamopine) —catabolizing (AGR) Agrobacterium tumefaciens strains in of in- vitro liquid bioassays. OD values were measured at different concentrations of EMA_PF2 in liquid bioassays.
Figure 7.
OD values of the wild-type (a) (A281) wild-type and the disarmed Ti plasmid bearing (b) A4T) agropine (the L, L,-succinamopine) —catabolizing (AGR) Agrobacterium tumefaciens strains in of in- vitro liquid bioassays. OD values were measured at different concentrations of EMA_PF2 in liquid bioassays.
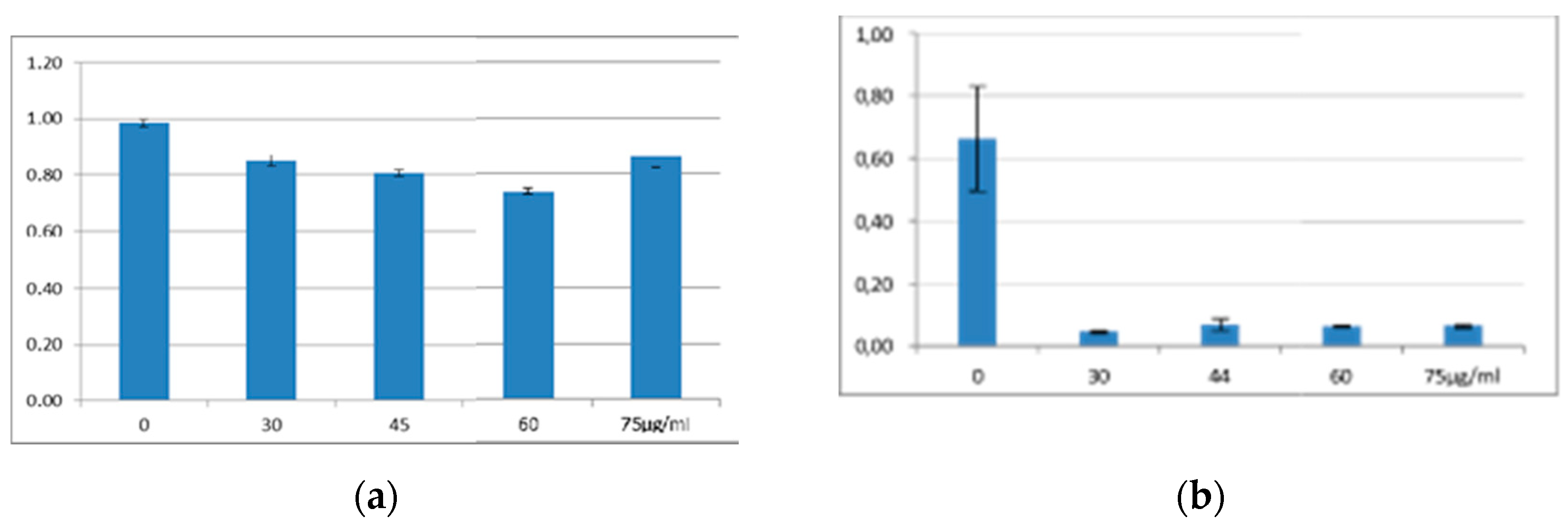
Table 1.
Agrobacterium strain.
| Name | Genotype | ||||||
|---|---|---|---|---|---|---|---|
|
LAB |
REF |
Genome | Plasmon | T-DNA | Opine | ||
| Chromo some |
Selective marker |
Ti plasmid | BIN | ||||
| HP1836 | C58C*NOP1 | C58C* | NalR | Cured | [T-DNA](-) | NOP | |
| HP1840 | C58C*NOP2 | C58C* | NalR | Cured | [T-DNA](-) | NOP | |
| HP1843 | C58C*NOP3 | C58C* | NalR | Cured | [T-DNA](-) | NOP | |
| HP1841 | C58C1NOP4 | C58C1* | RifR | Cured | [T-DNA](-) | NOP | |
| HP1842 | C58C1NOP5 | C58C1* | RifR | Cured | [T-DNA](-) | NOP | |
| SZL4 | C58C1/ pMP90NOP6 |
C58C1* | RifR | pMP90 GeR | pHP9- Gus101 |
Δ− [T-DNA] | NOP |
| HP1837 | LBA4404/0 OCT1 |
Ach5 | RifR | pAl4404 SmR | Δ− [T-DNA] | OCT | |
| SZL2 | LBA 4404/ pBIN-OCT2 |
Ach5 | RifR | pAl4404 SmR | pBIN | Δ− [T-DNA] | OCT |
| HP1838 | A281 | C58 | RifR | pTiA136 Bo542 |
[T-DNA](+) | AGR | |
| HP1839 | AGL1 | C58C* (AG0) |
CaR RecA(-) |
pEHA101NalR | Δ− [T-DNA] | AGR | |
| SZL1 | EHA 105 | C58C* | RifR | pEHA105NalR | Δ− [T-DNA] | AGR | |
| SZL3 | A4T | RifR | A4T | Δ− [T-DNA] | AGR | ||
* Footer to Table 1. Agrobacterium tumefaciens strains were used in this study. Genotype: includes genome that is the respective chromosome C58; Ach5) with genome-selective markers; and the Plasmon (including the respective Ti (Ri) plasmid with plasmid/selective markers and the respective binary vector (BIN) with binary vector selective markers). The binary vectors are engineered DNA constructions that are capable of replicating both Agrobacterium and E. coli cells and could be transmitted to plant cells as well because they. include T-DNA border sequences which could be recognized by the respective vir gene product. Abbreviations of the selective markers: Nal®, Rif®; Carb®; Ge®, and Sm®: resistant to nalidixic acid, rifampicin, carbenicillin, gentamycin, and streptomycin, respectively. T-DNA: a special segment (cassette) of the Ti (Ri) plasmid that could covalently be inserted into the infected plant chromosome. It carries genes expressed and regulated in the infected plant cell. T-DNA (+): genotype of the wild-type Ti plasmid having the intact T-DNA cassette; (Δ –TDNA) means [T-DNA] (-) genotype of the disarmed (non-virulent, helper) Ti (RI) plasmid from which the T-DNA cassette had been precisely excised. Opine: opines are strain-specific compounds, synthesized by the respective opine-synthase gene in plant tumors and can also be catabolized by decomposing enzymes encoded in the respective Ti (RI) plasmid, located outside of the T-DNA cassette. Abbreviations of opines: NOP =nopaline catabolizing (and synthetizing) Agrobacterium strain; OCT = octopine catabolizing (and synthetizing) Agrobacterium strain; AGR=agropine and L, L - succinamopine catabolizing (and synthetizing) Agrobacterium strain. Abbreviations of strains from which the complete Ti (RI) plasmid had been removed: The C58 strains cured for the pTiC58 plasmid general C58C* means C58 cured. The C58 strains cured especially for the pTiC58 (rifR) Ti plasmid are labeled as C58C1. C58C1 means that the cured stain carries a chromosomal rifampicin resistance mutation. C58C1RifR was alternatively designated as GV3101. C58C* = (in this study) the cured stain carries a chromosomal naladixic acid resistance mutation. We did not have a chance to work on plans-mid-cured OCT and AGR strains. For details on plasmids (pMP90; pAl4404; pTiA136Bo542; pEHA101; pEHA105 and A4T) see Textext and Supplementary Material TextS1.
Table 2.
Results of agar-diffusion assays - Inactivation zone in mm2 Gram-positive and fungal targets.
Table 2.
Results of agar-diffusion assays - Inactivation zone in mm2 Gram-positive and fungal targets.
| Antimicrobial peptide | . Inactivation zone in mm2 Gram-negative targets | |||
|---|---|---|---|---|
| HGB2226 | HGB1795* | |||
| N | Mean ± SD | N | Mean ± SD | |
| EMA_PF1 | 3 | 3820.00±690.22 | 3 | 4280.00 ±415.69 |
| EMA_PF2 | 4 | 3683.75±799.23 | 2 | 6602.50 456.08 |
| EMA_PF2*20 | 3 | 0 | 3 | 3370.00± 635.83 |
| EMA_PF2*30 | 3 | 0 | 3 | 3119.00 ±842.61 |
| EMA_PF2*40 | 3 | 0 | 3 | 4088.00± 678.70 |
| EMA_PF2*50 | 3 | 0 | 3 | 3821.67 ±214.20 |
| EMA_PF2*70 | 3 | 0 | 3 | 4640.00± 850.97 |
| EMA_PF2*TF | 3 | 0 | 3 | 4172.00±1502.02 |
| EMA_(RPLC)30 | 3 | 1452.50 ±95.45 | 3 | 1761.33± 173.78 |
| AF103_(EMA)_HPLC40 | 3 | 617.00±88.02 | 3 | 1135.33± 119.52 |
| AF103_(EMA)_HPLC43 | 3 | 1614.00± 81.41 | 3 | 2073.33 ±244.32 |
| AF103_(EMA)_HPLC44 | 3 | 1019.33 ±113.52 | 3 | 1385.00± 100.00 |
| Results of agar-diffusion assays - Inactivation zone in mm2 Gram-(+) and fungal targets. | ||||
| Antimicrobial peptide | ||||
| SA | CA | |||
| 3 | 8723.33 ±600.44 | 3 | 11746.67 ±704.37 | |
| EMA_PF1 | 3 | 5931.67± 453.22 | 3 | 6291.6667±627.58134 |
| EMA_PF2 | 3 | 0 | 1 | 530.00 ±0.00 |
| EMA_PF2*20 | 3 | 0 | 3 | 696.33± 279.69 |
| EMA_PF2*30 | 3 | 0 | 3 | 544.67 ±226.68 |
| EMA_PF2*40 | 3 | 0 | 3 | 0.00 |
| EMA_PF2*50 | 3 | 0 | 3 | 623.33 ±175.65 |
| EMA_PF2*70 | 3 | 0 | 558.33± 49.07 | |
| EMA_PF2*TF | 3 | 1656.67± 40.41 | 3 | 1526.00±233.83 |
| EMA_(RPLC)30 | 3 | 1614.00 ±81.41 | 3 | 2289.00±0.000 |
| AF103_(EMA)_HPLC40 | 3 | 1886.33± 66.97 | 3 | 2930.00± 287.51 |
| AF103_(EMA)_HPLC43 | 3 | 1613.67± 81.98 | 3 | 2834.33± 377.57 |
1 The footer of Table 2: Agar-Diffusion Bioassays of Antimicrobial Activities of Antimicrobial-active Peptide Fractions Isolated from EMA CFCM: Inactivation Zone Sizes (IZ values) Determined in four Targets Agar-Diffusion test Antimicrobial Activities of Peptide Fractions Isolated from EMA CFCM were determined in four targets in Agar-Diffusion test. Inactivation zones were calculated from the diameter of inactivation in 1-cm tick LBA plates and given in mm3 Abbreviations: AMP Fractions EMA_PF1: A peptide fraction isolated from the cell-free culture medium (CFCM) of Xenorhabdus budapestensis (EMA). EMA_PF1is a fraction supposed to be containing molecules of MW > 10,000 D. We suppose that the large peptides of PF1 this fraction adsorbed small reactive peptides and we detected the activity of the complex rather than antimicrobial activity of peptides of MW > 10,000 D EMA_PF2 (PF2) consists of peptides of MW < 10,000 D; EMA_PF2*20, EMA_PF2*30, EMA_PF2*40, EMA_PF2*50 and EMA_PF2*70: EMA_PF2 was further fractionated by, RPLC, and fractions eluted by 20. 30, 40, 50 and 70 V/V% of AN containing 0.1% TFA, freeze-dried, taken up by PBS and bio-assayed. TF: fraction which did not adsorb to the column. EMA_ (RPLC) 30 or (EMA30) also mentioned as AF103 (as HPLC sample) is a purified fraction of EMA PF by Reverse Phase Column Chromatography; eluted with 30 V/V% AN. (This was the only RPLC fraction active on Gram-negative targets). AF103_ (EMA) _HPLC40, AF103_ (EMA) _HPLC43 and AF103_ (EMA) _HPLC44 HPLC subfractions of EMA30 (AF103) collected in the 40th, 43rd, and 44th minute of the run of sample AF103. Test organisms: Gram-negative organisms: HGB2226 = Escherichia coli (E. coli) mutator strain of Km® Cm® genotype, constructed by L.K. (In this experiment it was used just as a Geram-negative est organism.); HGB1795 = was received as a transposon-induced insertion mutant of the XNC1_2022 gene (Gene ID: 9430524; Gene Page Link: NCBI UniProtKB; Locus Tag: XNC1_2022 see gene page for GenePage for the XNC1_2022 gene EcoGene-RefSeq) from Xenorhabdus nematophila (strain ATCC 19061 / DSM 3370 / LMG 1036 / NCIB 9965 / AN6), kindly provided by Prof. Helge Bode, via Prof. Heidi Goodrich-Blair, which behaves like an “immune-suppressed” X. nematophila, a Xenorhabdus-antibiotic-sensitive Xenorhabdus mutant. Later we did not get any information about this strain from the Bode lab. (In his experiment this mysterious strain was used just as a Gram-negative est organism.);) SA = Staphylococcus aureus (as a Gram-positive), CA = Candida albicans as a fungal target, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



