
Submitted:
02 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
Pericytes, as perivascular cells, are present in all vascularized organs and tissues, and they actively interact with endothelial cells in capillaries and microvessels. Their involvement includes functions like blood pressure regulation, tissue regeneration, and scarring.
Studies have confirmed that pericytes play a crucial role in bone tissue regeneration through direct osteodifferentiation processes, paracrine actions, and vascularization. Recent pre-clinical and clinical experiments have shown that combining perivascular cells with osteogenic factors and tissue-engineered scaffolds can be therapeutically effective in restoring bone defects. This approach holds promise for addressing bone-related medical conditions.
In this review, we have emphasized the characteristics of pericytes and their involvement in angiogenesis and osteogenesis. Furthermore, we have explored recent advancements in the use of pericytes in preclinical and clinical investigations, indicating their potential as a therapeutic resource in clinical applications.
Keywords:
pericyte
; osteogenesis
; bone defect
; regeneration
1. Introduction
The regeneration of massive bone defects remains a challenging and not fully resolved problem in traumatology and orthopedics. After the bone tumor, surgical removal or severe trauma the massive bone defects do not recover spontaneously and require a significant amount of bone grafting [1,2]. Currently, for the treatment of massive bone defects, various methods of osteoplasty using autologous and allogeneic osteografts, as well as bone substitutes are applied [3–5]. The application of bone autograft transplantation is considered the gold standard in bone defect treatment. However, this method has some drawbacks and limitations: 1) harvesting of bone from donor sites is a painful procedure and takes a long time to recover; 2) the lack of the required amount of donor bone tissue. In cases where autologous transplantation is difficult, allogeneic transplantation is the most common alternative. The cadaveric bones are used for this application, so additional operations are not required. However, there are some serious disadvantages, which include: reduced bone strength during sterilization, graft rejection, and the risk of contracting hepatitis or acquired immunodeficiency syndrome (AIDS). Use of the bone substitutes does not always lead to the complete restoration of massive bone defects. At present time, great hopes for massive bone defects’ regeneration are reasonably associated with the use of the tissue-engineered biomaterials. For the effective restoration of structural and functional characteristics of the damaged bones use of stem cells, growth factors, and biopolymers or scaffolds are suggested. Generally, searching for tissue-engineered methods for the development of an appropriate biomaterial is faced with several issues: the choice of an optimal cell source, the choice of factors stimulating regeneration, and the choice of an optimal biocompatible carrier or matrix. A promising cell source for tissue engineering is adipose tissue (AT), which, in comparison with other tissue sources, contains not only a large number of mesenchymal stem cells but also perivascular stem cells (PSCs) or pericytes [6]. This article highlights studying the pericytes and the therapeutic potential of pericytes combined with a different biological carrier and stimulating factors to accelerate the regeneration of damaged bone tissue.
2. Morphological characteristics of pericytes
According to classical histology, pericytes (“rouge cells”, or “mural cells”) are a population of outgrown cells of connective tissue that surrounds small blood vessels. The site of the localization of pericytes is the outer wall of the microvessels, blood capillaries, and small venules (postcapillaries) that form a basal membrane with endothelial cells. Pericytes are contractile cells that are similar to the smooth muscle cells (SMCs) of an arteriolar wall. In contrast to SMCs, they are completely embedded in the basal membrane of capillaries and are directly involved in the growth and functional development of a microcirculatory bed. In addition to the structural regulation of the capillary wall, pericytes are involved in the dynamic modulation of microvascular tone and diameter. Researchers identified the unique functions of pericytes that were associated with the regulation of the capillary blood flow, angiogenesis, phagocytosis, and other physiological processes of a microvasculature [7–9] (Table 1).
Depending on the localization of the microvascular bed and the degree of differentiation, pericytes exhibit different morphology, ranging from a typical flat, stellate shape (CNS) to a more rounded, shape (in the kidney). On average, cells are ~20µm long and ~0.5 µm thick. In general, pericytes have an elongated shape, a characteristic convex nucleus, and numerous processes. The body of the pericytes is always elongated along the direction of the blood flow. Long primary processes are located along the long axis of the capillary. Short secondary processes encircle the capillaries and form tight junctions with the endotheliocytes. The irregular distribution of the content that includes actins and myosins in the pericytes’ populations that depend on their localization causes notable differences in their contractility [8,9,31–35].
Pericytes regulate the formation of the basal membrane through the secretion of extracellular matrix proteins: collagen IV, laminin, and fibronectin. The interaction between pericytes and endotheliocytes occurs through dense intercellular contact and various signaling pathways. The slotted intercellular junctions (nexus) allow the interaction of pericytes’ cytoplasm with endotheliocytes to exchange substances with the help of connexin proteins by ion transport, and diffusion of nutrients and metabolites. Adhesion junctions ensure the transmission of mechanical contractile forces from pericytes to endothelial cells and the involvement of N-cadherin/catenin complexes with fibronectin-based cellular adhesion molecules. On the cytoplasm side, the glycoprotein complexes are bound to the actin filament bundles, which is important for pericytes attachment and movement. The pericyte-endothelial interdigitations provide strong mechanical binding, metabolism, and increase in the area of the intercellular interactions. The intercellular contacts between pericytes and endotheliocytes influence their mitotic activity, and gene expression and consequently, change each other's phenotype [7,8,32–37]. The density of the pericytes on the surface of the microvessels varies between the different organs. The vascular bed of the nervous system is considered to be the most covered by pericytes (endotheliocyte to pericyte ratio is 1:1-3:1). The much lower endothelial to pericyte ratio (100:1) was described for human skeletal muscle tissue. Pericytes’ coverage of the vascular surface ranges from 70% to 10%, which is related to the structure of the haematoparenchymal barrier, endotheliocyte proliferation, and vascular diameter. Reduced pericytes density and circumference of the microvessels are responsible for neurodegenerative diseases and uncontrolled angiogenesis [32,33,38,39].
3. Markers of pericytes
It is known that pericytes as multipotent cells are present in all vascularized tissues of the body. The role of the pericytes is not limited to the regulation and maintenance of the microvascular bed; they can also differentiate into phagocytes, chondrocytes, adipocytes, myocytes, and osteoblasts. Numerous studies have established that perivascular pericytes from various human organs and tissues have the differentiation potential and expression profile inherent to mesenchymal stem cells (MSCs) [40–43]. Pericytes as MSCs are associated with the microvasculature and express major phenotypical markers that include CD44, CD73, CD90, and CD105. Pericytes which carry the marker antigens of the MSCs are most effectively detected in blood vessels of the AT and dental pulp. It was determined that pericytes can differentiate into immune cells such as dendritic cells and macrophage-like cells, which play an important role in inflammation and under pathological conditions. Despite similarities in the localization and expression of surface immunomarkers, the ability to differentiate in vitro into osteoblasts, chondrocytes, adipocytes, myocytes, and smooth muscle cells, and form foci of ectopic osteogenesis in vivo, the biological equivalence of pericytes with MSCs remains open.
Currently, there is no single molecular marker that can be used to unambiguously identify the pericytes. Difficulties in the identification of them are associated with the fact that all known pericyte markers are used to phenotype other types of human cells. The most known markers of pericytes are smooth muscle alpha-actin (α-SMA), platelet-derived growth factor receptor beta (PDGFR-β), nerve/glial antigen 2 (NG-2), and CD146 [8,9,32–34,39–45].
The α-SMA (alpha-smooth muscle actin) marker is used to identify pericytes because they are identified as microvascular analogs of smooth muscle cells of the blood vessels. The α-SMA marker is the highly conserved contractile protein of smooth muscle cells that involves vasoconstriction and vasodilation of the vessels. The expression of α-SMA by capillary pericytes is significantly lower than in arterial smooth muscle cells, which was related to the rate of the regulation of blood pressure for vascular wall contraction. A mid-capillary pericytes with thin and spindle-shaped cytoplasmic processes do not express α-SMA, whereas pre- and post-capillary pericytes, with thick and stellate processes are rich in a-SMA expression [9,31–33,39–41].
NG2 (neural/glial antigen 2, chondroitin sulfate proteoglycan 4 - CSPG4) is an integral membrane proteoglycan that participates in the transmembrane signal transduction, cell adhesion, intercellular communication, migration, and proliferation. This marker is actively expressed by pericytes of arterioles and capillaries but is absent in pericytes of the venules. NG2 maintains the vascular network homeostasis, and its absence in venous vessels contributes to the regulation of arterial/venous anastomoses. However, NG2 expression is also characteristic of actively proliferating cells: activated macrophages, chondroblasts, osteoblasts, keratinocytes, fibroblasts, and some tumor cell types (gliomas and melanomas) [8,9,31–33,37,40–43].
For accurate phenotyping of NG2-expressing pericytes, an expression assay for the PDGFRβ marker is used. The PDGFRb (Platelet-derived growth factor receptor beta) - PDGFRb is a tyrosine kinase receptor that enables the differentiation of pericytic precursor cells. It is the most detectable marker of microvascular pericytes that are functionally involved in the recruitment of pericytes for normal blood vessel development and stabilization during angiogenesis. Dysregulation of the PDGFRβ kinase activity is responsible for the development of cardiovascular diseases [31–33,36,38,39].
CD146 (melanoma cell adhesion molecule, MCAM) Mel-CAM, MUC18, S-endo1) is a transmembrane glycoprotein with a molecular weight of approximately 110 kDa. Its function as a Ca2+-independent cell adhesion molecule which connected with intercellular interactions. It was reported that CD146 can promote pericytes recruitment with endotheliocytes through the direct regulation of the PDGF-B/PDGFRβ signaling pathway. Thus, this marker is presented as a universal marker of the brain pericytes, bone marrow, myocardium, and skeletal muscle. Authors have established an important role for CD146 positive pericytes in the control of the blood-brain barrier [9,32,33,38–45].
The aforementioned markers are notably expressed in pericytes at different stages. The NG2 and α-SMA markers are predominantly expressed by mature pericytes, whereas PDGFR-β is expressed by pericytic progenitor cells. Expression of the NG2 and α-SMA correlates with the type of vessels they surrounded. The pericytes enveloping capillaries are NG2+/α-SMA-, pericytes of the venules are NG2-/α-SMA+, pericytes of the arterioles are NG2+/α-SMA+, whereas PDGFR- and CD146 are ubiquitously expressed in all types of pericytes. Such evidence is related to stages of the pericytes differentiation, specific tissue structure, pathological condition, vascular hierarchy and stages of development [9,31,32,36,41–45].
The other markers that are used to identify pericytes are specific within the organ or tissue. For example, CD13 (aminopeptidase N), a type II membrane metalloprotease, is a marker of cerebral pericytes and is associated with the blood-brain barrier. ALP (alkaline phosphatase), an enzyme involved in metabolism, is actively expressed by skeletal pericytes. The regulator of G-protein-5 signaling (RGS5) is a protein that is expressed by activated pericytes during vascular remodeling and active tumor development. CD34 is a transmembrane phosphoglycoprotein that participates in cells adhesion as well as in the regulation of differentiation and proliferation. The CD34 marker is presented as a marker of the adventitial pericytes subpopulation. The expression of a nestin (cytoskeleton intermediate filament protein) marker by pericytes was found to be of two types, NG2+/Nestin- and NG2+/Nestin+, which differ in their angiogenic ability. The NG2+/Nestin+-positive pericytes that promote angiogenesis are targeted for anti-angiogenic therapy [7–9,36,37,42,43].
Unlike endothelial cells, pericytes do not express the next markers CD31, CD34, CD144, vW (von Willebrand factor).
4. A role of pericytes in angiogenesis
The localization of pericytes in the vascular space indicates that the endothelium is an important component of the perivascular niche capable of regulating the functional activity of pericytes. Pericytes play a key role in angiogenesis and are involved in the regulation of blood flow, vessel formation, remodeling, and stabilization. Determining the mechanisms that regulate these processes is crucial for understanding the pathogenesis of vascular complications.
Many soluble factors and signalling pathways are known to be fundamental for mutual intercellular interaction and communication between pericytes and endothelial cells (EC) at the molecular level. Pericytes are involved in the formation and sprouting of new blood vessels by signaling molecules such as PDGFR-b, transforming growth factor beta (TGF-b), vascular endothelial growth factor (VEGF), angioprotein 1 (Ang-1) and sphingosine-1-phosphate (S1P) [7–9,31–34,36,38–40].
Under hypoxic conditions, pericytes release VEGF-A to activate and transform endothelial cells (tip cells) and stimulate their migration during vascular sprouting. For pericytes’ adhesion (recruitment) on new vessels, PDGF-B secreted by sprouting endothelial cells binds to the pericyte-specific receptor PDGFRβ. The importance of PDGF/PDGFRβ signaling for capillary stabilization has been demonstrated in endothelio-specific PDGFB knockout mice resulting in endothelial hyperplasia, an aberrant vasculature and microaneurysms. Stabilization of the newly formed blood vessels is further regulated by the Ang-1/Tie-2 signaling pathway, in which Ang-1, a ligand produced by pericytes, binds to the endothelium-specific receptor Tie-2. Ang-1 is a natural inhibitor of vascular permeability. The Ang-1/Tie-2 binding promotes the association of pericytes and endothelium to form a tight vascular barrier.
The activation of the angiopoietin-1 antagonist, angiopoietin 2 (Ang-2), that binds to the Ang-1 receptor (Tie-2), contributes to vascular instability. The blocking of Ang-1-Tie-2 signaling leads to weakened contact between endothelial cells and pericytes, which increases their permeability and leads to vascular regression. Thus, the balance of the Angpt1/Angpt2 ratio determines vascular homeostasis and is essential for the regulation of new vessels formation and maturation [9,31–34,36,37,39,40,46–49].
The connection of pericytes with endothelial cells activates transforming growth factor beta (TGFβ), which inhibits endothelial cells (EC) proliferation and migration but stimulates the differentiation of pericytes and hence promotes blood vessels maturation. The importance of TGFβ is supported by studies in which the absence of TGFβ in experimental mice led to abnormal vascular network development which was characterized by a lack of pericytes coverage, vascular instability and rupture [7–9,31,32,34,40,47].
In addition, the interaction between ECs and pericytes is regulated by the S1P signaling molecule, a sphingolipid, that is involved in cell communication. The S1P maintains the vascular endothelial barrier by promoting the secretion of extracellular matrix components, enhancing the interaction of the connective proteins, N-cadherin and VE-cadherin, and inhibiting vascular destabilization factors such as Angpt2 and VEGF-A [9,32,40,47].
Thus, pericytes are important regulators of the angiogenesis process within definite signaling pathways that form the normal vascular bed.
This review of the current scientific data shows that researchers in the field of regenerative medicine have considered the pericytes as a promising therapeutic target. However, there are still many challenges, requiring the careful study of the identification of pericytes, the mechanism of pericytes differentiation, the determination of phenotypic differences between pericytes in angiogenesis and mature vessels, and the detailed determination of signaling pathways. Also, it is necessary to understand the functions of pericytes in the microcirculatory stream and the possibilities of their use for regenerative medicine (Figure 1).
5. Pericytes in osteogenesis
The potential of pericytes in osteogenesis was studied in an experimental animal models of muscle pocket, calvaria, spine fusion, and non-union fracture [36]. As reported, the significant potential for bone regeneration was found for CD146+ positive pericytes/progenitors and the optimal cell source was the subcutaneous adipose tissue (AT). The most meaningful effect in ossification for ectopic bone formation showed the combination of human AT-derived pericytes with osteoinductive demineralized bone matrix (DBM) with increased VEGF [36]. Therefore, the cell therapy by pericytes for spinal fusion in combination with the DBM scaffold resulted in notable osteodifferentiation and complete bone regeneration due to the paracrine mechanism. For the treatment of non-union fractures, the CD146+ AT pericytes showed a prominent bone healing however the complete regeneration depended on how the microenvironment of the injured site is attractable for pericytes shifting [36]. The potential mechanism of the effect of pericytes for bone healing was formulated: close ossification of transplanted cells, activation of the shifting and proliferation processes in osteoprogenitor cells, and paracrine and immunomodulatory effect on osteogenic and endothelial cells [49]. Authors reported that pericytes from human AT showed significant regeneration of the damaged bone in rats’ model of osteoporosis in the next combinations: within the bone chips and sodium hyaluronate-based scaffold, joined with the NELL-1 protein or with the gel foam-alginate 3D scaffold [50]. Also, for calvarial injuries, pericytes within the scaffold from poly(lactic-co-glycolic-acid) (PLGA) scaffold, from alginate gel, from collagen sponge or hydroxyapatite-based polymeric scaffold demonstrated the effect of regeneration [50] (Table 2). Thus, reported studies confirm the high osteogenic potential of pericytes in the acceleration of bone defects’ healing due to paracrine factors, immunomodulation, and vascularization.
6. Recent pre-clinical and clinical application of pericytes
A long-practiced approach widely used in clinics to facilitate fracture healing is demineralized bone matrix (DBM) implantation [57]. Among DMB implantation methods iliac crest autografts and fibula allografts are widely used in clinical practice to repair various fractures. However, in 10–25% of cases, these approaches are not effective in repairing delayed union and nonunion in the elderly patients [58]. Moreover, this procedures are known to have a number of adverse effects, not only during the initial surgery but also years later due to issues like scars, numbness, and pain that can impact on quality of life [59]. As an alternative strategy the cellular therapy with pericytes can be proposed [60]. Bone is an organ with an extensive vascular system that ensures efficient blood flow and nutrient delivery, therefore the use of mural cells as pericytes for bone tissue repair has long been posited as these cells are essential in bone development and regeneration. Experimental evidence has established that pericytes are the endogenous precursors of mesenchymal stem cells [61]. Pericytes can be isolated from AT, which, compared to other tissue sources, contains also a large number of mesenchymal stem cells and can relatively easy promote the acquirement of substantial quantities of the cells [6,62].
Recent animal studies reported that smooth muscle actin (SMA) after a fracture also participates in the formation of bone cells from endogenous mural cells [63]. SMA is a non-specific marker that is additionally expressed on smooth muscle and fibroblasts/myofibroblasts, therefore it cannot directly confirm this phenomenon. Studies have focused on identifying populations of pericytes which possess the highest potential for osteoblast formation. One of the study results demonstrate that CD146+ pericytes, obtained from skeletal tissue have the greatest potential for the repair and formation of bone both in vitro and in vivo compared with ones obtained from adipose or dermal tissue [64]. However, subsequent clinical studies confirmed the ability of AT pericytes to regenerate bones [53]. For successful isolation of pericytes from many tissues, the CD146+CD34-CD31-CD45-CD56- immunophenotype can be used [65]. Among soft tissue pericytes, cells expressing CXCR4 have a high osteoblastic potential, which are classified as non-adipocytic progenitor cells [64]. NG2, CD146 and PDGFRβ-positive pericytes in lineage-tracing experiments using NG2-Cre or tamoxifen-induced NG2-CreER mouse strain revealed that they are able to differentiate into osteogenic cells in mice [66] Pericytes labeled with tdTomato expression form callus in fractured femurs and have a positive expression of Runx2 and type I collagen corresponding to bone tissue in a bone fracture model [66]. Thus, pericytes isolated from AT demonstrate significant bone-forming potential in all autologous and xenogenic experiments in vivo. Additionally, recent studies have demonstrated that pericytes derived from human white AT with positive expression of alkaline phosphatase and bone matrix have bone formation potential in an intramuscular mouse model [45].
Another issue in cellular based therapy is the choice of a method for delivering cells into a defect area. Cell delivery can be done in a single or multiple dose, or using scaffolds, which are then implanted into a bone defect.
Polymers find extensive application in tissue engineering as scaffolds for cell cultures. These particular polymers are referred to as “cell-laden scaffolds”. The application of such polymers is based on the ability to create a three-dimensional tissue structure, followed by the incorporation of cells and the introduction of growth factors, cytokines, or chemokines into the scaffold [67]. The scaffolds should not only serve as a matrix for cells, but should promote cell proliferation and differentiation. Additionally, these polymers must possess specific properties, such as non-toxicity, biodegradability, biocompatibility and non-immunogenicity [68]. Demineralized bone matrix grafts, synthetic and natural polymers, bioactive ceramics, cross-linked hydrophilic hydrogels, metals, and composite scaffolds are often used as cell-laden scaffolds to retain cells and control the release of bioactive factors (Figure 2.) [63].
PSCs have been the subject of investigation in the context of scaffolds in various preclinical models to explore their osteogenic potential using DBM [62,69–71], collagen sponge [51], hydrogel [72,73], bone matrix [53,55,56], or polymer coated hydroxyapatite [6].
Several animal models in ectopic bone formation confirmed the potential of implanted human pericytes for osteogenic differentiation in vivo. The results of the study demonstrated the formation of inconspicuous bone after AT pericytes implantation with a carrier of a collagen sponge in a muscle [51]. In another work, AT pericytes have been implanted using an osteoinductive DBM carrier, which resulted in strong bone formation in muscle pocket osteogenesis [51]. Other researchers have reported similar findings, that CD146+ AT-derived cells did not showed significant bone formation when implanted in a subcutaneous model. In contrast to CD146+ precursor cells from the bone marrow or periosteum which exhibited strong bone tissue formation [74]. Comparisons of AT-derived pericytes, adventitial cells, and an unsorted/uncultured stromal population (termed stromal vascular fraction, SVF) showed that adventitial cells have a similar bone-forming potential and SVF are not effective enough [45,51]. SVF or pericytes in combination with adventitial cells (PSC) from the same patient on a hydroxyapatite-coated polymeric scaffold in a mouse calvarial defect model revealed that PSC led to a significant increase in bone regeneration [51]. Additionally, AT-derived human PSC using DBM carrier in a rat spinal fusion model resulted in paracrine-mediated bone formation [55].
A number of studies by James et al. confirmed the effectiveness of pericytes application in bone disease treatment [36,45,49,62,75]. In particular, the research group proved that human AT pericytes incorporated in PLGA scaffold accelerated the healing of critical size skull defects in mice within 2 weeks [6].
Another interesting research conducted by Zhang and colleagues suggested a novel pre-vascularization strategy using tri-culture of human induced pluripotent stem cell-derived mesenchymal stem cells (hiPS-MSCs), human umbilical vein endothelial cells (HUVECs) and pericytes on calcium phosphate cement (CPC). CPC scaffolds were designed to possess 100–300μm macropores. Six groups were tested in in vitro and in vivo research. According to the obtained results, bi-culture and tri-culture groups exhibited the formation of vessel-like structures in vitro, and in vivo study showed that the tri-cultured group exhibited a significantly larger quantity of new bone than bi-culture, mono-culture, and CPC control [76,77]. Thus, as shown by a multitude preclinical studies, pericytes may show promising results in clinical applications in the future [49].
Concerning modern clinical practice, the therapies in bone healing are based on local delivery of cells to the fracture site in combination with growth factors and scaffolds (Table 3).
Potential osteoinductive growth and differentiation factors in combination with pericytes are used for the optimal bone formation. Among the growth factors, the recombinant bone morphogenetic protein-2 (BMP-2), is known to be used in fusion facilitation [78]. Another factor, recombinant human platelet growth factor (rhPDGF) is used to accelerate the process of bone defects filling [79]. Additionally, a number of studies demonstrated the application of (BMP-2) [51], NELL-1 [51,52,74,80], the short isoform of NELL-1 [81] or WNT16 [82] which are used as osteoinductive factors for bone tissue restoration. Although all of these factors are osteoinductive in nature, the mechanism of action is different. For example, NELL-1 promotes both bone and cartilage formation, while WNT16 or the short isoform of NELL-1 have combined mitogenic/pro-osteogenic effects [79].
Recombinant BMP-2 is clinically available, and its combination with PSC induces synergistic effect in bone formation [49,83]. Mumcuoglu and colleagues developed an injectable hydrogel based on collagen microspheres and alginate to deliver BMP-2 into bone defects [84]. Other researchers, taking into account the heparin-binding ability of BMPs, mixed heparin with carriers such as chitosan, poly-L-lactic acid, and DBM [85]. Similar studies were conducted by Yang et al. who developed an injectable fibrin hydrogel for long-term delivery of BMP-2 through covalent conjugation of heparin to fibrinogen and showed its effectiveness in bone defect regeneration [86].
Jing Bai and colleagues have developed an organ-specific microfuidic platform recapitulating the in vivo angiogenic microenvironment by co-culturing mouse primary brain endothelial cells with brain pericytes in 3D collagen scaffold [87]. Morrison K. et al. fabricated prevascularized scaffold containing a hierarchical vascular network. Human pericytes and fibroblasts were incapsulated into the polydimethylsiloxane (PDMS) mold containing the 3D Pluronic F127 microfiber and macrofiber network. After 28 days of cultivation, investigators revealed that neoangiogenic sprouts formed in the collagen protodermis and pericytes self-assembled around both fabricated vessels and neoangiogenic sprouts [88]. Other researchers designed a PDMS device with a reservoir for a 3D fibrinogen gel with pericytes. It allows the interaction of endothelial cells (ECs) with pericytes and the extracellular matrix (ECM) in full bio-matrix encased 3D vessel structures (neovessels) [89].
Currently, clinical studies have been conducted for the restoration of bone defects using adipose-derived mesenchymal stem cells (Ad-MSC) and a preparation based on autologous microfragmented adipose tissue (Table 4), which have shown promising results. The use of an enriched fraction of pericytes may potentially yield better outcomes than merely halting the progression of the disease, as demonstrated in the clinical use of Ad-MSC and autologous microfragmented adipose tissue.
Thus, studies on pericytes application with growth factors in a cell-laden scaffolds have shown that the use of scaffolds or any other matrices for pericytes has great potential in restoring damaged bone tissue. The study of other properties of pericytes, such as pro-osteogenic, pro-vasculogenic and immunoregulatory functions, which have not yet been studied, may give further impetus to preclinical and clinical studies.
7. Conclusions
Human pericytes is a unique population of perivascular cells with mesenchymal stem cell properties present in all vascularized tissues. Pericytes have a high ability for multilineage differentiation, especially in the osteogenic direction. Given the enormous role of pericytes in the stabilization and formation of microvessels and angiogenesis, pericytes are of particular interest in the regeneration and engineering of vascularized bone tissue.
Efficient biomaterials and appropriate manufacturing techniques play a critical role in the development of injectable hydrogels that function as scaffolds for bone tissue engineering. Over the past 10 years, a variety of injectable hydrogels have been developed. These biomaterials include gelatin, alginate, collagen, chitosan, poly-L-lactic acid, hyaluronic acid, fibrin, heparin, and polyethylene glycol [102].
The application of various in vivo bone formation models based on injectable hydrogels and polymer scaffolds with pericytes, which include ectopic muscle pocket osteogenesis and calvarial replenishment in mice, spinal fusion in rats and dogs, and fracture healing in rats, confirmed the therapeutic efficacy of perivascular cells for accelerated osteoregeneration.
Thus, although scientists made progress in cellular-based approach for bone regeneration, there is still a necessity to advance the development of improved injectable hydrogel using stem cells and/or osteoinductive factors for efficient regeneration of damaged bone tissue.
Author Contributions
Conceptualization: S.E. (Saule Eskendirova) and V.O. (Vyacheslav Ogay); Data curation: A.I. (Assel Issabekova), S.E., G.K. (Gulshakhar Kudaibergen), A.S.(Aliya Sekenova), A.D. (Aidar Dairov); Formal analysis: M..S. (Madina Sarsenova) and V.O; Project administration: V.O.; Writing—original draft: S.E., A.I., G.K., A.S., A.D.; Writing—review and editing: M.S. and A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Ministry of Education and Science of the Republic of Kazakhstan: 102 “Grant funding for scientific research” program, No. AP09259357 Development of an injectable hydrogel for effective regeneration of massive bone defects project.
Institutional Review Board Statement
The study was conducted in accordance with the European Convention for the Protection of Vertebrate Animals (ETS No. 123) used for experimental and other scientific purposes. It was also approved by the Local Ethics Commission of the Republican State Enterprise “National Center for Biotechnology” CS MES RK (order No. 4, dated 8 September 2020) (Astana, Kazakhstan).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
A. Saparov (Nazarbayev University School of Medicine, Kazakhstan) for advices and assistance. M. Shakhatbayev (Stem cell laboratory, NCB, Kazakhstan) for technical support.
Conflicts of Interest
The authors have no conflict of interest.
References
- Roddy, E.; De Baun, M.R.; Daoud-Gray, A.; Yang, Y.P.; Gardner, M.J. Treatment of critical-sized bone defects: clinical and tissue engineering perspectives. Eur. J. Orthop. Surg. Traumatol. 2018, 28, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Nishida, J.; Shimamura, T. Methods of reconstruction for bone defect after tumor excision: a review of alternatives. Med. Sci. Monit. 2008, 14, 107–113. [Google Scholar]
- Goulet, J.A.; Senunas, L.E.; De Silva, G.L.; Greenfield, M.L.V.H. Autogenous iliac crest bone graft: complications and functional assessment. Clin. Orthop. Relat. Res. 1997, 339, 76–81. [Google Scholar] [CrossRef]
- Rabitsch, K.; Maurer-Ertl, W.; Pirker-Frühauf, U.; Wibmer, C.; Leithner, A. Intercalary reconstructions with vascularised fibula and allograft after tumour resection in the lower limb. Sarcoma 2013, 2013, 160295. [Google Scholar] [CrossRef]
- Tamai, N.; Myoui, A.; Tomita, T.; Nakase, T.; Tanaka, J.; Ochi, T.; Yoshikawa, H. Novel hydroxyapatite ceramics with an interconnective porous structure exhibit superior osteoconduction in vivo. J. Biomed. Mater. Res. 2002, 59, 110–117. [Google Scholar] [CrossRef] [PubMed]
- James, A.W.; Zara, J.N.; Corselli, M.; Askarinam, A.; Zhou, A.M.; Hourfar, A.; Nguyen, A.; Megerdichian, S.; Asatrian, G.; Pang, S.; et al. An abundant perivascular source of stem cells for bone tissue engineering. Stem Cells Transl. Med. 2012, 1, 673–684. [Google Scholar] [CrossRef]
- Zhao, H.; Chappell, J.C. Microvascular bioengineering: a focus on pericytes. J. Biol. Eng. 2019, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Meijer, E.M.; van Dijk, C.G.M.; Kramann, R.; Verhaar, M.C.; Cheng, C. Implementation of pericytes in vascular regeneration strategies. Tissue Eng. - Part B Rev. 2022, 28, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.S.; Zhou, H.N.; He, S.S.; Xue, M.Y.; Li, T.; Liu, L.M. Research advances in pericyte function and their roles in diseases. Chinese J. Traumatol. - English Ed. 2020, 23, 89–95. [Google Scholar] [CrossRef]
- Herndon, J.M. Chapter 9 – Development and maintenance of the blood-brain barrier. In Primer on cerebrovascular diseases.Second edition. Ed. Caplan, L.R. Publisher: Academic Press, 2017, 51-56. [CrossRef]
- Attwell, D.; Mishra, A.; Hall, C.N.; O'Farrell, F.M.; Dalkara, T. What is a pericyte? J. Cereb. Blood Flow Metab. 2016, 36, 451–455. [Google Scholar] [CrossRef]
- Brown, L.S.; Foster, C.G.; Courtney, J.-M.; King, N.E.; Howells, D.W.; Surtherland, B.A. Pericytes and neurovascular function in the healthy and diseased brain. Front. Cell.Neurosci. 2019, 13, 282. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Gao, C.; Gao, D.; Sun, R.; Li, W.; Wang, F.; Wang, Y.; Cao, H.; Zhou, G.; Zhang, J.; Shang, J. Reduction in pericyte coverage leads to blood-brain barrier dysfunction via endothelial transcytosis following chronic cerebral hypoperfusion. Fluids Barriers CNS. 2021, 18, 21. [Google Scholar] [CrossRef]
- Heymans, M.; Figueiredo, R.; Dehouck, L.; Francisco, D.; Sano, Y.; Shimizu, F.; Kanda, T.; Bruggmann, R.; Engelhardt, B.; Winter, P.; Gosselet, F.; Culot, M. Contribution of brain pericytes in blood–brain barrier formation and maintenance: a transcriptomic study of cocultured human endothelial cells derived from hematopoietic stem cells. Fluids Barriers CNS. 2020, 17, 48. [Google Scholar] [CrossRef] [PubMed]
- Ferland-McCollough, D.; Slater, S.; Richard, J.; Reni, C.; Mangialardi, G. Pericytes, an overlooked player in vascular pathobiology. Pharmacol.Ther. 2017, 171, 30–42. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhang, Z.; Peng, X.; Wu, Y.; Zhu, Y.; Wang, L.; Zhou, H.; Li, T.; Liu, L. The protective effect of pericytes on vascular permeability after hemorrhagic shock and their relationship with Cx43. Front. Physiol. 2022, 13, 1–16. [Google Scholar] [CrossRef]
- Welberg, L. Pericytes set the tone. Nature Rev. Neorosci. 2014, 15, 283. [Google Scholar] [CrossRef]
- Randall Harrell, C.; SimovicMarkovic, B.; Fellabaum, C.; Arsenijevic, A.; Djonov, V.; Volarevic, V. Molecular mechanisms underlying therapeutic potential of pericytes. J. Biomed. Sci. 2018, 25, 21. [Google Scholar] [CrossRef]
- Wu, Y.; Fu, J.; Huang, Y.; Duan, R.; Zhang, W.; Wang, C.; Wang, S.; Hu, X.; Zhao, H.; Wang, L.; Liu, J.; Gao, G.; Yuan, P. Biology and function of pericytes in the vascular microcirculation. Animal Model Exp. Med. 2023, 6, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, T.; Kubo, S.; Nakano-Doi, A.; Sakuma, R.; Lu, S.; Narita, A.; Kawahara, M.; Taguchi, A.; Matsuyama, T. Brain vascular pericytes following ischemia have multipotential stem cell activity to differentiate into neural and vascular lineage cells. Stem Cells. 2015, 33, 1962–1974. [Google Scholar] [CrossRef]
- Ahmed, T.A.; El-Badri, N. Pericytes: the role of multipotent stem cells in vascular maintenance and regenerative medicine. Adv. Exp. Med. Biol. 2018, 1079, 69–86. [Google Scholar] [CrossRef]
- Cheng, J.; Korte, N.; Nortley, R.; Sethi, H.; Tang, Y.; Attwell, D. Targeting pericytes for therapeutic approaches to neurological disorders. ActaNeuropathologica. 2018, 136, 507–523. [Google Scholar] [CrossRef]
- Courtney, J.-M.; Sutherland, B.A. Harnessing the stem cell properties of pericytes to repair the brain. Neural.Regen. Res. 2020, 15, 1021–1022. [Google Scholar] [CrossRef]
- Garrison, A.T.; Bignold, R.E.; Wu, X.; Johnson, J.R. Pericytes: the lung-forgotten cell type. Front. Physiol. 2023, 14, 1–16. [Google Scholar] [CrossRef]
- Sagare, A.P.; Bell, R.D.; Zhao, Z.; Ma, Q.; Winkler, E.A.; Ramanathan, A.; Zlokovic, B.V. Pericyte loss influences Alzheimer-like neurodegeneration in mice. Nature Communications. 2013, 4, 2932. [Google Scholar] [CrossRef]
- HosseinGeranmayeh, M.; Rahbarghazi, R.; Farhoudi, M. Targeting pericytes for neurovascular regeneration. Cell Commun.Signalling. 2019, 17, 26. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhou, J.; Li, L.; Liao, S.; He, J.; Zhou, S.; Zhou, Y. Pericytes in the tumor microenvironment. Cancer Lett. 2023, 556, 216074. [Google Scholar] [CrossRef]
- Sun, R.; Kong, X.; Qiu, X.; Huang, C.; Wong, P.-P. The emerging roles of pericytes in modulating tumor microenvironment. Front. Cell Dev. Biol. 2021, 9, 1–10. [Google Scholar] [CrossRef]
- Picoli, C.C.; Gonçalves, B.Ô.P.; Santos, G.S.P.; Rocha, B.G.S.; Costa, A.C.; Resende, R.R.; Birbrair, A. Pericytes cross-talks within the tumor microenvironment. BiochemicaetBiophysicaActa (BBA) – Reviews on Cancer. 2021, 1876, 188608. [Google Scholar] [CrossRef]
- Warmke, N.; Griffin, K.J.; Cubbon, R.M. Pericytes in diabetes-associated vascular disease. J. Diabetes Complications. 2016, 30, 1643–1650. [Google Scholar] [CrossRef]
- Harrell, C.R.; Simovic Markovic, B.; Fellabaum, C.; Arsenijevic, A.; Djonov, V.; Volarevic, V. Molecular mechanisms underlying therapeutic potential of pericytes. J. Biomed. Sci. 2018, 25, 1–12. [Google Scholar] [CrossRef]
- Mills, S.J.; Cowin, A.J.; Kaur, P. Pericytes, mesenchymal stem cells and the wound healing process. Cells 2013, 2, 621–634. [Google Scholar] [CrossRef]
- Cathery, W.; Faulkner, A.; Maselli, D.; Madeddu, P. Concise review: the regenerative journey of pericytes toward clinical translation. Stem Cells 2018, 36, 1295–1310. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef]
- Chen WC, Saparov A, Corselli M, Crisan M, Zheng B, Péault B, Huard J. Isolation of blood-vessel-derived multipotent precursors from human skeletal muscle. J Vis Exp. 2014 Aug 21;(90):e51195. PMID: 25177794; PMCID: PMC4762055. [CrossRef]
- Meyers, C.A.; Casamitjana, J.; Chang, L.; Zhang, L.; James, A.W.; Péault, B. Pericytes for therapeutic bone repair. In Pericyte Biology – Novel Concepts. Advances in Experimental Medicine and Biology; Birbrair, A., Ed.; Publisher: Springer, Cham, 2018; Volume 1109, pp. 21–32. [Google Scholar] [CrossRef]
- Blocki, A.; Beyer, S.; Jung, F.; Raghunath, M. The controversial origin of pericytes during angiogenesis - Implications for cell-based therapeutic angiogenesis and cell-based therapies. Clin. Hemorheol. Microcirc. 2018, 69, 215–232. [Google Scholar] [CrossRef]
- Gökçinar-Yagci, B.; Uçkan-Çetinkaya, D.; Çelebi-Saltik, B. Pericytes: properties, functions and applications in tissue engineering. Stem Cell Rev. Rep. 2015, 11, 549–559. [Google Scholar] [CrossRef]
- Wong, S.P.; Rowley, J.E.; Redpath, A.N.; Tilman, J.D.; Fellous, T.G.; Johnson, J.R. Pericytes, mesenchymal stem cells and their contributions to tissue repair. Pharmacol. Ther. 2015, 151, 107–120. [Google Scholar] [CrossRef]
- Caporarello, N.; D'Angeli, F.; Cambria, M.T.; Candido, S.; Giallongo, C.; Salmeri, M.; Lombardo, C.; Longo, A.; Giurdanella, G.; Anfuso, C.D.; et al. Pericytes in microvessels: from ‘mural’ function to brain and retina regeneration. Int. J. Mol. Sci. 2019, 20, 6351. [Google Scholar] [CrossRef]
- Yamazaki, T.; Mukouyama, Y.S. Tissue specific origin, development, and pathological perspectives of pericytes. Front. Cardiovasc. Med. 2018, 5, 1–6. [Google Scholar] [CrossRef]
- Yianni, V.; Sharpe, P.T. Perivascular-derived mesenchymal stem cells. J. Dent. Res. 2019, 98, 1066–1072. [Google Scholar] [CrossRef]
- Esteves, C.L.; Donadeu, F.X. Pericytes and their potential in regenerative medicine across species. Cytom. Part A 2018, 93, 50–59. [Google Scholar] [CrossRef]
- Grant, R.I.; Hartmann, D.A.; Underly, R.G.; Berthiaume, A.A.; Bhat, N.R.; Shih, A.Y. Organizational hierarchy and structural diversity of microvascular pericytes in adult mouse cortex. J. Cereb. Blood Flow Metab. 2019, 39, 411–425. [Google Scholar] [CrossRef]
- James, A.W.; Hindle, P.; Murray, I.R.; West, C.C.; Tawonsawatruk, T.; Shen, J.; Asatrian, G.; Zhang, X.; Nguyen, V.; Simpson, H.; et al. Pericytes for the treatment of orthopedic conditions. Pharmacol. Ther. 2017, 171, 93–103. [Google Scholar] [CrossRef]
- Lee, L.L.; Khakoo, A.Y.; Chintalgattu, V. Cardiac pericytes function as key vasoactive cells to regulate homeostasis and disease. FEBS Open Bio 2021, 11, 207–225. [Google Scholar] [CrossRef]
- Chiaverina, G.; di Blasio, L.; Monica, V.; Accardo, M.; Palmeiro, M.; Peracino, B.; Vara-Messler, M.; Puliafito, A.; Primo, L. Dynamic interplay between pericytes and endothelial cells during sprouting angiogenesis. Cells 2019, 8, 1–13. [Google Scholar] [CrossRef]
- Laredo, F.; Plebanski, J.; Tedeschi, A. Pericytes: problems and promises for CNS repair. Front. Cell. Neurosci. 2019, 13, 1–15. [Google Scholar] [CrossRef]
- James, A.W.; Péault, B. Perivascular mesenchymal progenitors for bone regeneration. J. Orthop. Res. 2019, 37, 1221–1228. [Google Scholar] [CrossRef]
- Avolio, E.; Alvino, V.V.; Ghorbel, M.T.; Campagnolo, P. Perivascular cells and tissue engineering: current applications and untapped potential. Pharmacol. Ther. 2017, 171, 83–92. [Google Scholar] [CrossRef]
- James, A.W.; Zara, J.N.; Zhang, X.; Askarinam, A.; Goyal, R.; Chiang, M.; Yuan, W.; Chang, L.; Corselli, M.; Shen, J.; et al. Perivascular stem cells: a prospectively purified mesenchymal stem cell population for bone tissue engineering. Stem Cells Transl. Med. 2012, 1, 510–519. [Google Scholar] [CrossRef]
- Askarinam, A.; James, A.W.; Zara, J.N.; Goyal, R.; Corselli, M.; Pan, A.; Liang, P.; Chang, L.; Rackohn, T.; Stoker, D.; et al. Human perivascular stem cells show enhanced osteogenesis and vasculogenesis with Nel-like molecule I protein. Tissue Eng. - Part A 2013, 19, 1386–1397. [Google Scholar] [CrossRef]
- Meyers, C.A.; Xu, J.; Zhang, L.; Asatrian, G.; Ding, C.; Yan, N.; Broderick, K.; Sacks, J.; Goyal, R.; Zhang, X.; et al. Early immunomodulatory effects of implanted human perivascular stromal cells during bone formation. Tissue Eng. - Part A 2018, 24, 448–457. [Google Scholar] [CrossRef]
- Tawonsawatruk, T. , West, C., Murray, I.; et al. Adipose derived pericytes rescue fractures from a failure of healing – non-union. Sci Rep 2016, 6, 22779 (2016). [Google Scholar] [CrossRef] [PubMed]
- Chung, C.G.; James, A.W.; Asatrian, G.; Chang, L.; Nguyen, A.; Le, K.; Bayani, G.; Lee, R.; Stoker, D.; Pang, S.; et al. Human perivascular stem cell-based bone graft substitute induces rat spinal fusion. Stem Cells Transl. Med. 2015, 4, 538. [Google Scholar] [CrossRef]
- Lee, S.; Zhang, X.; Shen, J.; James, A.W.; Chung, C.G.; Hardy, R.; Li, C.; Girgius, C.; Zhang, Y.; Stoker, D.; et al. Brief report: Human perivascular stem cells and Nel-Like Protein-1 synergistically enhance spinal fusion in osteoporotic rats. Stem Cells. 2015, 33, 3158–3163. [Google Scholar] [CrossRef]
- Gruskin, E.; Doll, B.A.; Futrell, F.W.; Schmitz, J.P.; Hollinger, J.O. Demineralized bone matrix in bone repair: history and use. Adv. Drug Deliv. Rev. 2012, 64, 1063–1077. [Google Scholar] [CrossRef]
- Lin, S.S.; Yeranosian, M.G. The role of orthobiologics in fracture healing and arthrodesis. Foot Ankle Clin. 2016, 21, 727–737. [Google Scholar] [CrossRef]
- Dodwad, S.N.M.; Mroz, T.E.; Hsu, W.K. Biologics in spine fusion surgery. Benzel’s Spine Surg. Tech. Complicat. Avoid. Manag. Vol. 1-2, Fourth Ed. 2017, 1–2, 280–284. [CrossRef]
- Zhu, S.; Chen, M.; Ying, Y.; Wu, Q.; Huang, Z.; Ni, W.; Wang, X.; Xu, H.; Bennett, S.; Xiao, J.; et al. Versatile subtypes of pericytes and their roles in spinal cord injury repair, bone development and repair. Bone Res. 2022, 10, 30. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef]
- Asatrian, G.; Pham, D.; Hardy, W.R.; James, A.W.; Peault, B. Stem cell technology for bone regeneration: current status and potential applications. Stem Cells Cloning Adv. Appl. 2015, 8, 39–48. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-renewing osteoprogenitors in bone marrow sinusoids can organize a hematopoietic microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef]
- Xu, J.; Li, D.; Hsu, C.-Y.; Tian, Y.; Zhang, L.; Wang, Y.; Tower, R.J.; Chang, L.; Meyers, C.A.; Gao, Y.; et al. Comparison of skeletal and soft tissue pericytes identifies CXCR4+ bone forming mural cells in human tissues. Bone Res. 2020, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Birbrair, A. Pericyte biology in disease. In Adv. Exp, Med. Biol. Publisher: Springer, Chem., Switzerland, 2019; Volume 1147. [CrossRef]
- Supakul, S.; Yao, K.; Ochi, H.; Shimada, T.; Hashimoto, K.; Sunamura, S.; Mabuchi, Y.; Tanaka, M.; Akazawa, C.; Nakamura, T.; et al. Pericytes as a source of osteogenic cells in bone fracture healing. Int. J. Mol. Sci. 2019, 20, 1079. [Google Scholar] [CrossRef] [PubMed]
- Çelebi-Saltik, B. Pericytes in tissue engineering. In Pericyte Biology - Novel Concepts Adv. Exp. Med. Biol.; Birbrair, A.; Publisher: Springer, Chem., 2018; Volume 1109, 125–137. [CrossRef]
- Annabi, N.; Tamayol, A.; Uquillas, J.A.; Akbari, M.; Bertassoni, L.E.; Cha, C.; Camci-Unal, G.; Dokmeci, M.R.; Peppas, N.A.; et al. Rational design and applications of hydrogels in regenerative medicine. Adv. Mater. 2014, 26, 85–124. [Google Scholar] [CrossRef] [PubMed]
- Asatrian, G.C.C.; James, A.W.; Liang, P.; et al. Human perivascular mesenchymal stem cells promote lumbar spinal fusion via induction of osteogenesis and vasculogenesis. Seattle, WA: International Association for Dental Research; 2013.
- Chung, C.G.K.J.; Velasco, O.; Asatrian, G.; et al. Perivascular stem cells with NELL-1 protein induce robust spinal fusion. Charlotte, NC: American Association of Dental Research; 2014.
- James, A.W.; Zara, J.N.; Corselli, M.; Chiang, M.; Yuan, W.; Nguyen, V.; Askarinam, A.; Goyal, R.; Siu, R.K.; Scott, V.; et al. Use of human perivascular stem cells for bone regeneration. J. Vis. Exp. 2012, 63, e2952. [Google Scholar] [CrossRef]
- Alakpa, E.V.; Jayawarna, V.; Burgess, K.E.V.; West, C.C.; Péault, B.; Ulijn, R.V.; Dalby, M.J. Improving cartilage phenotype from differentiated pericytes in tunable peptide hydrogels. Sci. Rep. 2017, 7, 6895. [Google Scholar] [CrossRef] [PubMed]
- Alakpa, E.V.; Jayawarna, V.; Lampel, A.; Burgess, K.V.; West, C.C.; Bakker, S.C.J.; Roy, S.; Javid, N.; Fleming, S.; Lamprou, D.A.; et al. Tunable supramolecular hydrogels for selection of lineage-guiding metabolites in stem cell cultures. Chem 2016, 1, 298–319. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Remoli, C.; Giannicola, G.; Kogler, G.; Liedtke, S.; Cossu, G.; Serafini, M.; Sampaolesi, M.; Tagliafico, E.; et al. No identical ‘mesenchymal stem cells’ at different times and sites: Human committed progenitors of distinct origin and differentiation potential are incorporated as adventitial cells in microvessels. Stem Cell Reports 2016, 6, 897–913. [Google Scholar] [CrossRef]
- Chang, L.; Nguyen, V.; Nguyen, A.; Scott, M.A.; James, A.W. Pericytes in sarcomas of bone. Med. Oncol. 2015, 32, 202. [Google Scholar] [CrossRef]
- Zhang, C.; Hu, K.; Liu, X.; Reynolds, M.A.; Bao, C.; Wang, P.; Zhao, L.; Xu, H.H.K. Novel hiPSC-based tri-culture for pre-vascularization of calcium phosphate scaffold to enhance bone and vessel formation. Mater. Sci. Eng. C 2017, 79, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huang, S.; Zou, R.; Gao, X.; Ruan, J.; Weir, M.D.; Reynolds, M.A.; Qin, W.; Chang, X.; Fu, H.; et al. Calcium phosphate cement scaffold with stem cell co-culture and prevascularization for dental and craniofacial bone tissue engineering. Dent. Mater. 2019, 35, 1031–1041. [Google Scholar] [CrossRef]
- Schroeder, G.D.; Hsu, W.K.; Kepler, C.K.; Kurd, M.F.; Vaccaro, A.R.; Patel, A.A.; Savage, J.W. Use of recombinant human bone morphogenetic protein-2 in the treatment of degenerative spondylolisthesis. Spine (Phila. Pa. 1976) 2016, 41, 445–449. [Google Scholar] [CrossRef]
- Dhote, R.; Charde, P.; Bhongade, M.; Rao, J. Stem cells cultured on beta tricalcium phosphate (β-TCP) in combination with recombinant human platelet-derived growth factor - BB (RH-PDGF-BB) for the treatment of human infrabony defects. J. Stem Cells 2015, 10, 243–254. [Google Scholar] [PubMed]
- Zhang, X.; Péault, P.; Chen, W.; Li, W.; Corselli, M.; James, A.W.; Lee, M.; Siu, R.K.; Shen, P.; Zheng, Z.; et al. The nell-1 growth factor stimulates bone formation by purified human perivascular cells. Tissue Eng. - Part A 2011, 17, 2497–2509. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Shen, J.; Liu, Y.; Chen, F.; Zheng, Z.; James, A.W.; Hsu, C.-Y.; Zhang, H.; Lee, K.S.; Wang, C.; et al. Proliferation and osteogenic differentiation of mesenchymal stem cells induced by a short isoform of NELL-1. Stem Cells 2015, 33, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Chen, X.; Jia, H.; Meyers, C.A.; Shrestha, S.; Asatrian, G.; Ding, C.; Tsuei, R.; Zhang, X.; Peault, B.; et al. Effects of WNT3A and WNT16 on the osteogenic and adipogenic differentiation of perivascular stem/stromal cells. Tissue Eng. - Part A 2018, 24, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Granjeiro, J.M.; Oliveira, R.C.; Bustos-Valenzuela, J.C.; Sogayar, M.C.; Taga, R. Bone morphogenetic proteins: from structure to clinical use. Brazilian J. Med. Biol. Res. 2005, 38, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Mumcuoglu, D.; Fahmy-Garcia, S.; Ridwan, Y.; Nicke, J.; Farrell, E.; Kluijtmans, S.G.; van Osch, G.J. Injectable BMP-2 delivery system based on collagen-derived microspheres and alginate induced bone formation in a time-and dose-dependent manner. Eur. Cells Mater. 2018, 35, 242–254. [Google Scholar] [CrossRef]
- Engstrand, T.; Veltheim, R.; Arnander, C.; Docherty-Skogh, A.-C.; Westermark, A.; Ohlsson, C.; Adolfsson, L.; Larm, O. A novel biodegradable delivery system for bone morphogenetic protein-2. Plast. Reconstr. Surg. 2008, 121, 1920–1928. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.S.; La, W.-G.; Bhang, S.H.; Jeon, J.-Y.; Lee, J.H.; Kim, B.-S. Heparin-conjugated fibrin as an injectable system for sustained delivery of bone morphogenic protein-2. Tissue Eng. Part A 2010, 16, 1225–1233. [Google Scholar] [CrossRef]
- Bai, J.; Khajavi, M.; Sui, L.; Fu, H.; Krishnaji, S.T.; Birsner, A.E.; Bazinet, L.; Kamm, R.D.; D'Amato, R.J. Angiogenic responses in a 3D micro-engineered environment of primary endothelial cells and pericytes. Angiogenesis 2021, 24, 111–127. [Google Scholar] [CrossRef]
- Morrison, K.A.; Weinreb, R.H.; Dong, X.; Toyoda, Y.; Jin, J.L.; Bender, R.; Mukherjee, S.; Spector, J.A. Facilitated self-assembly of a prevascularized dermal/epidermal collagen scaffold. Regen. Med. 2020, 15, 2273–2283. [Google Scholar] [CrossRef]
- van Dijk, C.G.M.; Brandt, M.M.; Poulis, N.; Anten, J.; van der Moolen, M.; Kramer, L.; Homburg, E.F.G.A.; Louzao-Martinez, L.; Pei, J.; Krebber, M.M.; et al. A new microfluidic model that allows monitoring of complex vascular structures and cell interactions in a 3D biological matrix. Lab Chip 2020, 20, 1827–1844. [Google Scholar] [CrossRef]
- Lu, L.; Dai, C.; Du, H.; Li, S.; Ye, P.; Zhang, L.; Wang, X.; Song, Y.; Togashi, R.; Vangsness, T.; et al. Intra-articular injections of allogeneic human adipose-derived mesenchymal progenitor cells in patients with symptomatic bilateral knee osteoarthritis: a phase I pilot study. Regen. Med. 2020, 15, 1625–1636. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Dai, C.; Zhang, Z.; Du, H.; Li, S.; Ye, P.; Fu, Q.; Zhang, L.; Wu, X.; Dong, Y.; et al. Treatment of knee osteoarthritis with intra-articular injection of autologous adipose-derived mesenchymal progenitor cells: a prospective, randomized, double-blind, active-controlled, phase IIb clinical trial. Stem Cell Res. Ther. 2019, 10, 143. [Google Scholar] [CrossRef]
- Lee, W.-S.; Kim, H.J.; Kim, K.I.; Kim, G.B.; Jin, W. Intra-articular injection of autologous adipose tissue-derived mesenchymal stem cells for the treatment of knee osteoarthritis: a phase IIb, randomized, placebo-controlled clinical trial. Stem Cells Transl. Med. 2019, 8, 504–511. [Google Scholar] [CrossRef]
- Freitag, J.; Bates, D.; Wickham, J.; Shah, K.; Huguenin, L.; Tenen, A.; Paterson, K.; Boyd, R. Adipose-derived mesenchymal stem cell therapy in the treatment of knee osteoarthritis: a randomized controlled trial. Regen. Med. 2019, 14, 213–230. [Google Scholar] [CrossRef]
- Kuah, D.; Sivell, S.; Longworth, T.; James, K.; Guermazi, A.; Cicuttini, F.; Wang, Y.; Craig, S.; Comin, G.; Robinson, D.; et al. Safety, tolerability and efficacy of intra-articular Progenza in knee osteoarthritis: a randomized double-blind placebo-controlled single ascending dose study. J. Transl. Med. 2018, 16, 49. [Google Scholar] [CrossRef] [PubMed]
- Jones, I.A., Wilson, M.; Togashi, R.; Han, B.; Mircheff, A.K.; Vangsness, T.J.R. A randomized, controlled study to evaluate the efficacy of intra-articular, autologous adipose tissue injections for the treatment of mild-to-moderate knee osteoarthritis compared to hyaluronic acid: a study protocol. BMC Mus. Dis. 2018, 19, 383. [CrossRef]
- Mikkelsen, R.K.; Blønd, L.; Hölmich, L.R.; Mølgaard, C.; Troelsen, A.; Hölmich, P.; Barfod, K.W. Treatment of osteoarthritis with autologous, micro-fragmented adipose tissue: a study protocol for a randomized controlled trial. Trials 2021, 22, 748. [Google Scholar] [CrossRef] [PubMed]
- Polancec, D.; Zenic, L.; Hudetz, D.; Boric, I.; Jelec, Z.; Rod, E.; Vrdoljak, T.; Skelin, A.; Plecko, M.; Turkalj, M.; et al. Immunophenotyping of a stromal vascular fraction from microfragmented lipoaspirate used in osteoarthritis cartilage treatment and its lipoaspirate counterpart. Genes 2019, 10, 474. [Google Scholar] [CrossRef]
- Borić, I.; Hudetz, D.; Rod, E.; Jeleč, Ž.; Vrdoljak, T.; Skelin, A.; Polašek, O.; Plečko, M.; Trbojević-Akmačić, I.; Lauc, G.; et al. A 24-month follow-up study of the effect of intra-articular injection of autologous microfragmented fat tissue on proteoglycan synthesis in patients with knee osteoarthritis. Genes 2019, 10, 1051. [Google Scholar] [CrossRef]
- Hudetz, D.; Borić, I.; Rod, E.; Jeleč, Ž.; Kunovac, B.; Polašek, O.; Vrdoljak, T.; Plečko, M.; Skelin, A.; Polančec, D.; et al. Early reasults of intra-articular micro-fragmented lipoaspirate treatment in patients with late stages knee osteoarthritis: a prospective study. Croat. Med. J. 2019, 60, 227–236. [Google Scholar] [CrossRef]
- Panchal, J.; Malanga, G.; Sheinkop, M. Safety and efficacy of percutaneous injection of lipogems micro-fractured adipose tissue for osteoarthritic knees. Am. J. Orthop. 2018, 47. [Google Scholar]
- Russo, A.; Condello, V.; Madonna, V.; Guerriero, V.; Zorzi, C. Autologous and micro-fragmented adipose tissue for the treatment of diffuse degenerative knee osteoarthritis. J. Exp. Orthop. 2017, 4, 33–10. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Takahashi, Y.; Tabata, Y. Controlled release by biodegradable hydrogels enhances the ectopic bone formation of bone morphogenetic protein. Biomaterials 2003, 24, 4375–4383. [Google Scholar] [CrossRef]
Figure 1.
Cell signaling pathways between pericytes and endothelial cells which involved in process of angiogenesis. (1) VEGF, secreted from pericytes, is implicated in propagation and differentiation processes; (2) Proliferation and migration of pericytes occur by PDGF-B, obtained from endothelial cells; (3) Ang-2/Tie-2 signaling ensures stabilization of new vessels and vascular homeostasis; (4) TGF-β that produced by both pericytes and endothelial cells, mediate the processes of maturation and stability of vessels; (5) S1P from endothelial cells maintain the vascular endothelial barrier. N-cadherin preserves cell communication between pericytes and endothelial cells.
Figure 1.
Cell signaling pathways between pericytes and endothelial cells which involved in process of angiogenesis. (1) VEGF, secreted from pericytes, is implicated in propagation and differentiation processes; (2) Proliferation and migration of pericytes occur by PDGF-B, obtained from endothelial cells; (3) Ang-2/Tie-2 signaling ensures stabilization of new vessels and vascular homeostasis; (4) TGF-β that produced by both pericytes and endothelial cells, mediate the processes of maturation and stability of vessels; (5) S1P from endothelial cells maintain the vascular endothelial barrier. N-cadherin preserves cell communication between pericytes and endothelial cells.
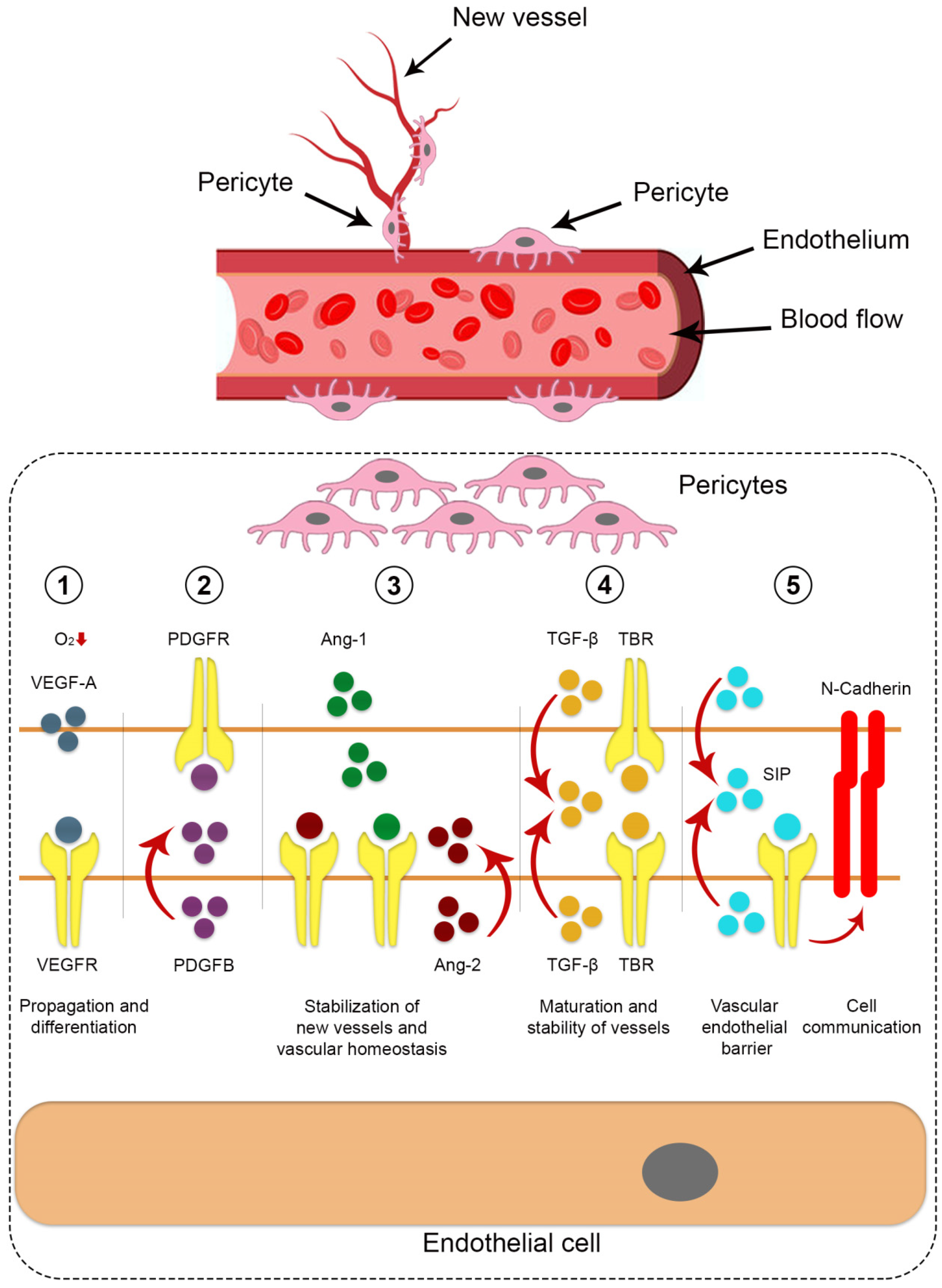
Figure 2.
Pericytes and growth factors based scaffold for bone regeneration.
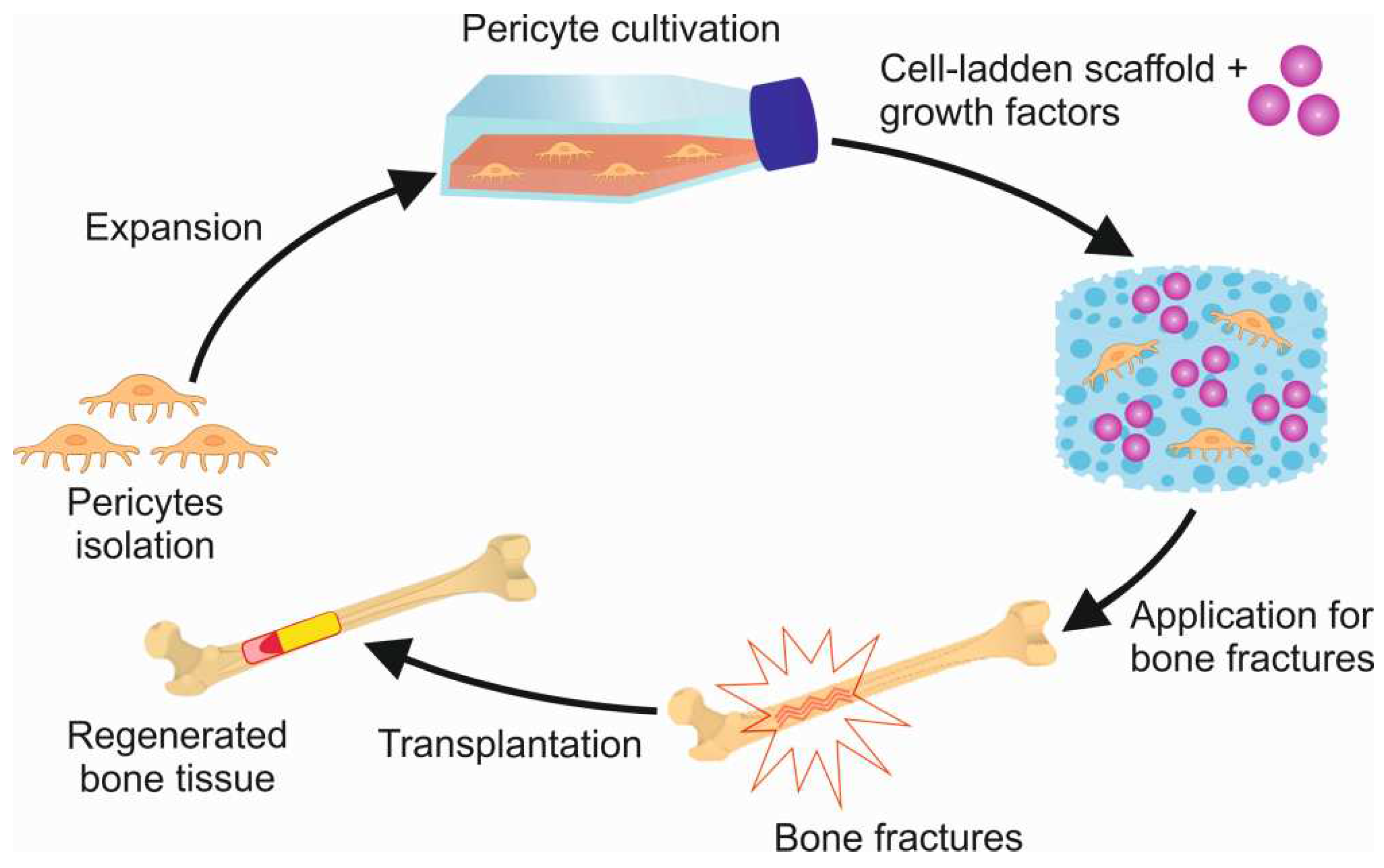

Table 1.
Physiological and pathological functions of pericytes.
| Function | Comments/reference |
|---|---|
| Physiological | |
| Blood brain barrier (BBB) | BBB is a specialized vascular structure that restricts the passage of most molecules from the systemic circulation into the central nervous system (CNS). It is crucial for proper neuronal function and is maintained by various cell types, collectively known as the neurovascular unit [10]. Pericytes, found along capillary walls, play a vital role in BBB maintenance, immune cell regulation, and brain blood flow control within the CNS ([11]. They are part of the neurovascular unit, which manages interactions between neurons and cerebral blood vessels to meet the brain's energy needs [12]. Loss of pericyte coverage can lead to BBB dysfunction and the accumulation of neurotoxic molecules, impacting white matter lesions [13]. The interaction between endothelial cells and brain pericytes can induce BBB characteristics during embryogenesis and is used in in vitro BBB models [14]. |
| Vascular permeability |
Increased pericyte coverage on tumor vasculature reduces vessel permeability, limiting the entry of pro-inflammatory and pro-tumorigenic cells [15]. In the context of hemorrhagic shock, pericytes act as protective cells situated on the basolateral side of endothelium, playing a crucial role in maintaining vascular barrier function in pulmonary and peripheral vessels [15]. |
| Vaso- constriction |
Pericytes play a key role in regulating blood flow in microvessels. They exhibit contractility through proteins like α-SMA, desmin, vimentin, and C-GMP, which affect actin filament bundles near endothelial cells. Pericytes respond to vasoconstrictors (e.g., angiotensin-II, serotonin) and vasodilators (e.g., nitric oxide, cholinergic agonists, adenosine) by changing the collagen lattice's surface area in vitro [9]. In rat cortex slices, ischemia causes vasoconstriction near pericytes, followed by pericyte death, suggesting their involvement in blood flow regulation and a potential role in reperfusion after ischemia [17]. |
| Angiogenesis | Pericytes, found in microvessels like capillaries and venules, play a vital role in maintaining structural integrity, regulating blood flow, promoting angiogenesis, stabilizing vasculature, and controlling permeability. In the central nervous system and retina, they form barriers protecting cells from harmful blood factors [18]. Pericytes are crucial for angiogenesis, influencing vessel stability by interacting with sprouting endothelial cells. They can also adjust capillary diameters, similar to smooth muscle cells, regulating microvessel blood flow [19]. |
| Impact on immune function |
Pericytes facilitate immune cell migration by releasing molecules, recruiting various immune cells, and promoting M2-like macrophages. They regulate the immune system in the central nervous system. Reduced CD4+ T-cells lead to decreased pericyte coverage, and retinal pericytes inhibit CD4+ T-cell activation [19]. |
| Stem cell | Pericytes maintain blood vessel integrity, prevent issues like vessel dilation and hemorrhaging, and exhibit stem cell-like qualities [20]. In the brain, vascular pericytes are crucial for the blood-brain barrier and demonstrate stem cell capabilities, particularly after brain damage from conditions like ischemia and hypoxia [21,22]. After ischemia, they can transform into various cell types, including neurons, microglia, and vascular cells, and assist in clearing damaged areas by becoming glial cells [23]. |
| Pathological | |
| Fibrosis |
Pericytes contribute to lung diseases like pulmonary arterial hypertension (PAH) and allergic asthma by transforming into scar-forming myofibroblasts, which leads to tissue fibrosis through collagen deposition and matrix remodeling [24]. |
| Neuro- degeneration |
Pericytes in the blood-brain barrier degenerate in Alzheimer's disease (AD), associated with neurovascular dysfunction, Aβ elevation, tau pathology, and neuronal loss [25]. Pericyte loss in neurological disorders increases blood-brain barrier permeability and may lead to vascular dementia [26]. |
| Cancer | Pericytes (PCs) in the tumor microenvironment have diverse roles, including forming the pre-metastatic niche, promoting cancer cell growth and drug resistance, and influencing M2 macrophage polarization [27]. In carcinogenesis, disrupted interaction between PCs and endothelial cells leads to dysfunctional tumor vasculature [28]. Recent studies employing advanced technologies confirm pericytes' communication with cancer cells [29]. |
| Diabetic retinopathy |
Pericyte loss is an early hallmark of diabetes-related microvascular diseases, including retinopathy and nephropathy. Pericytes actively contribute to vascular dysfunction by secreting pro-angiogenic factors, initiating neovascularization, thickening the basement membrane, and causing vasoconstriction [30]. |
Table 2.
Preclinical studies for bone healing therapy using adipose pericytes.
| Model | Type of cells and tissue origin | Animal type | Dosage | Method of investigation | Therapeutic outcome | Reference l |
|---|---|---|---|---|---|---|
| Intramuscular ectopic bone model | Adipose human perivascular stem cells (hPSCs) and adventitial cells derived from the same patient | severe combined immunodeficient (SCID) mice | 2.5 × 105 cells, sponge size 2.0 × 1.0 × 0.5 cm | Micro-CT imaging (bone mineral density and bone volume) | Both perivascular populations had a similar baseline osteogenic potential | [45] |
| Intramuscular ectopic bone model | Adipose hPSCs, SVF from the same patient | SCID mice | 2.5 × 105 cells, sponge size 2.0 × 1.0 × 0.5 cm | High resolution radiographic/3-Dimensional Micro-CT imaging | Larger particles of bone formation, increase in vascularity of the implant site, osteocalcin and bone sialoprotein expression | [51] |
| 3 mm non-healing calvarial defect centered in the parietal bone | Adipose hPSCs or SVF from the same patient | SCID mice | Radiographic imaging and histological examination | Significant bone defect healing overtime | [6] | |
| Intramuscular ectopic bone model | Adipose hPSCs | SCID mice | The demineralized bone matrix (DBX), with NELL-1 (3 μg/μL), hPSC (2.5×105 cells), or hPSC+NELL-1 | Micro-CT imaging, histological examination, immunohistochemical staining over 4 weeks | The additive effect of hPSC+NELL-1 on bone formation and vasculogenesis | [52] |
| Intramuscular ectopic bone model in the hindlimb | Adipose hPSCs or SVF from the same patient | SCID mice | DBX with SVF or hPSC | Histological examination, immunohistochemical staining | Significantly greater neutrophilic and macrophage infiltrates within and around SVF in comparison to PSC-laden implants, robust immunomodulatory effect | [53] |
| Tibial model | Adipose hPSCs, bone marrow MSC | Wistar rats | 5 × 106 cells were percutaneously injected into the fracture gap | Radiographic, micro-CT imaging and immunohistochemical staining | At eight weeks, 80% of animals in the cell treatment groups showed evidence of bone healing compared to only 14% of those in the control group | [54] |
| Posterolateral lumbar spinal fusion model | Adipose hPSCs | Athymic rats | DBX, 0,15Х106 hPSCs, 0,50x106 hPSCs, 1.5x106 hPSCs | Micro-CT imaging, immunohistochemical staining | Regulate bone formation via direct and paracrine mechanisms | [55] |
| Osteoporotic spinal fusion mode | Adipose hPSCs or NELL-1, BMP-2 | Athymic rats | 0.25 × 106 cells per milliliter of hPSCs or 33.3 μg/ml of NELL-1, 0.75 × 106 cells per milliliter of hPSCs or 66.6 μg/ml of NELL-1 | Micro-CT imaging, histological examination, immunohistochemical staining | The hPSC combined with NELL-1 synergistically enhances spinal fusion in osteoporotic rats | [56] |
Table 3.
FDA approved therapies for bone healing.
| Systemic | Local |
|---|---|
| Bisphosphonates | INFUSE (rhBMP-2) |
| Recombinant parathyroid hormone | Regranex (rhPDGF-BB) |
| RANKL inhibitors | rhBMP-7* |
| SOST inhibitors (pending) | Healos (GDF-5) (pending) |
| Demineralized bone matrix | |
| Fibula allograft | |
| Iliac crest autograft |
* FDA humanitarian device exemption in 2003, failed to pass FDA approval in 2009 (https://www.fda.gov/).
Table 4.
Features of clinical studies of stem cells from fat tissue.
| Dose (cell) (×106), donor, type of study | Age, Kellgren-Lawrence Grade, sample (M:F), follow up | Outcome, measures, effect | Study (year) |
|---|---|---|---|
| 1 × 10, 2 × 10, 5 × 10 x × 106, Autologous adipose tissue-MSC (Ad-MSC) In this single-site, randomized, double-blind, dose-ranging, phase I study |
|
WOMAC, VAS, WORMS, MRI, others MRI assessments showed slight improvements in the low-dose group |
Lu, L.; et al. (2020) [90] |
| 50 × 2 x × 106, Autologous Ad-MSC Randomized double-blind phase IIb clinical trial |
|
WOMAC, VAS, MRI, others 50% improvement of WOMAC 70% improvement rate in Re-Join® group after 12 months |
Lu, L.; et al. (2019) [91] |
| 100 × 106, autologous Ad-MSC Double-blinded, randomized controlled phase IIb clinical trial |
|
KOOS, WOMAC, VAS, MRI, others Improvement of WOMAC score at 6 months No significant change of cartilage defect in MSC group |
Lee, W.S.; et al. (2019) [92] |
| 100, 100 × 2 x × 106 at baseline and 6 months, autologous Ad-MSC Randomized controlled trial (RCT) |
|
KOOS, NPRS, WOMAC, others Significant pain and functional improvement in both treatment groups |
Freitag, J.; et al. (2019) [93] |
| 3.9 (Progenza 3.9M, n = 8) or placebo (n = 2) and 6.9 (Progenza 6.7M, n = 8) or placebo (n = 2), allogeneic Ad-MSC Double-blinded RCT |
|
|
Kuah, D.; et al. (2018) [94] |
| 6 ml, intra-articular injection, autologous adipose tissue (AT) or hyaluronic acid Prospective, single-center, parallel-group RCT |
|
WOMAC, WOMAC-A, PROMIS, force plate analysis, others |
Jones, I.; et al. (2018) [95] |
| 10 ml intra-articular injection, autologous, micro-fragmented AT or isotonic saline (placebo) Blinded RCT |
|
KOOS4, Tegner activity score, work status, others |
Mikkelsen, R.; et al. (2021) [96] |
| Intra-articular knee injection of autologous microfragmented lipoaspirate (MLA) |
|
Stromal vascular fraction isolation, flow cytometry | Polanec, D.; et al. (2019) [97] |
| 4-15 ml intra- articular injection, autologous microfragmented AT containing Ad-MSCs Prospective, non-randomized, interventional, single-center, open label clinical trial |
|
dGEMRIC, VAS A single intra-articular injection of autologous microfragmented AT improves glycosaminoglycans (GAG) content on a significant scale |
Borić, I.; et al. (2019) [98] |
| Intra-articular injection, autologous MLA Prospective, non-randomized study |
|
VAS, WOMAC, KOOS KOOS score improved from 46 to 176% when compared with baseline WOMAC decreased from 40 to 45% VAS rating decreased from 54% to 82% |
Hudetz, D.; et al. (2019) [99] |
| Intra-articular injection, autologous microfragmented AT |
|
NPRS, 100-point KSS with FXN and LEAS KSS score improved from 74 to 82 FXN score improved from 65 to 76 LEAS score improved from 36 to 47 |
Panchal, J.; et al. 2018 [100] |
| Intra-articular injection, autologous and microfragmented AT Retrospective study |
|
KOOS, IKDC, Tegner Lysholm knee, VAS IKDC-subjective and total KOOS improved by 20 points VAS-point and Tegner Lysholm knee improved by 24 and 31 points, respectively |
Russo, A.; et al. (2017) [101] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



