
Submitted:
02 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
This experiment was conducted to study the effects of Cyfluthrin (Cy) exposure on neurobehaviour, hippocampal tissue and synaptic plasticity in Wistar rats. First, high-dose Cy exposure can cause nerve injury with symptoms such as deficits in learning and memory ability, spatial exploration and autonomic motor ability. Moreover, it was found that medium- and high-dose Cy exposure can cause disordered energy metabolism in hippocampal cells and an abnormal release of the neurotransmitter Glu. Second, pathological brain sections showed that the middle and high doses of Cy caused deformation, reduction and disorder of hippocampal pyramidal cells, a decrease in the number of Nissl bodies, pyknosis of the hippocampal cell nuclear membrane and serious damage to organelles, indicating that exposure to these doses of Cy may cause hippocampal tissue damage in rats. Third, with the increase in exposure dose, the morphological changes in hippocampal synapses were more obvious, including blurred synaptic spaces, decreased synaptic vesicles and a decreased number of synapses. Moreover, the expression levels of the key synaptic proteins PSD-95 and SYP also decreased in a dose-dependent manner, indicating obvious synaptic damage. Finally, the study found that medium and high doses of Cy could upregulate the expression of A2AR in the hippocampus, and the expression levels of inflammatory factors and apoptosis-related proteins increased in a dose-dependent manner. Moreover, the expression of A2AR mRNA was correlated with neurobehavioural indicators, inflammatory factors, synaptic plasticity factors and apoptosis factors, suggesting that Cy may cause nerve damage in rats and has a close relationship with A2AR.
Keywords:
Cyhalothrin
; Nerve injury
; Synaptic plasticity
; A2AR
1. Background
Although pesticides bring great economic benefits to human beings, they also pose a potential threat to human health. The wide use of pesticides is an important risk factor affecting human health. Cyfluthrin (Cy) is an important type II pyrethroid insecticide [1]. Because of its low toxicity, high efficiency, low residue and wide range of pest control, it has been widely selected and used by the agriculture, forestry, animal husbandry and fishery industries since its development in 1984 [2]. With the increasing use of Cy, the potential health risks of Cy residues on soil organisms and human beings have become a popular issue [3]. The accumulation of Cy in the body causes disorders in the nervous system, digestive system, reproductive system, cardiovascular system, and immune system [4,5,6,7]. Moreover, a previous research group also found that 56.70% of the investigated vegetable greenhouse growers used dangerous concentrations of Cy, which were higher than those of other pyrethroid insecticides [8].
Cy has a certain fat solubility. After entering the body, Cy may break through the blood‒brain barrier, accumulate in brain tissue, and damage the human nervous system to varying degrees [9]. Some studies have shown that this damage may be a trigger for the development of neurodegenerative diseases. A large number of epidemiological investigations have also shown that long-term exposure to Cy and other pyrethroid pesticides can lead to a variety of nervous system disorders and even the precursor symptoms of neurodegenerative diseases [10,11]. Therefore, this study intends to establish a model of nerve injury induced by Cy exposure in rats to explore the relationship between Cy exposure and neurobehaviour, synaptic plasticity, the inflammatory response and adenosine A2AR expression in rats and to confirm the damaging effect of Cy exposure on the nervous system of Wistar rats.
2. Material Method
2.1. Experimental Reagent and Dose Selection
Cy (c22h18cl2fno3, molecular weight 434.4, CAS NO.68359-37-5) was purchased from Dr. Company in the United Kingdom, and the purity was 99.52%. The solvent was a non-GMO corn oil. According to the relevant literature, international pesticide use threshold standards and the results of previous experiments, the solvent control group (corn oil), low-dose exposure group (6.25 mg/kg) (1/60 LD50), medium-dose exposure group (12.5 mg/kg) (1/30 LD50) and high-dose exposure group (25 mg/kg) (1/15 LD50) were finally established.
2.2. Animals
Forty male SPF grade Wistar rats weighing approximately 270 g were purchased from Liaoning Changsheng Biotechnology Co., Ltd. (licence No.: scxk (x) 20150001) and raised in the animal experiment centre of Ningxia Medical University. First, the purchased rats were disinfected and transferred to the Animal Center for adaptive feeding for one week. The housing conditions included a room temperature of approximately 22 ℃, normal day and night cycle lighting, and a relative humidity of approximately 60%. During the feeding period, they had free access to water and chow. The bedding was changed every three days. After one week of adaptive feeding, 40 Wistar rats were randomly divided into four groups with 10 rats in each group by the tail labelling method. Neurobehavioural tests were conducted before and after the next day’s exposure. The exposure time of each time point was guaranteed to be between 8:00 a.m. and 10:00 a.m. After the test, the rats were anaesthetized and sacrificed for follow-up experiments.
2.3. Tissue Sampling
Four rats in each group were randomly selected to be perfused and fixed with paraformaldehyde (Seville Company in China Wuhan), and then the brain tissue was quickly stripped and fully fixed in paraformaldehyde. The hippocampus of other rats was removed and used for subsequent electron microscopy and frozen at -80 ℃ for subsequent detection. All animal treatment methods strictly complied with the relevant provisions of the People’s Republic of China on the operation of experimental animals, and the experimental methods adopted were also approved by the ethics committee of Ningxia Medical University (No.: iacuc-nylac-2019-076).
2.4. Neurobehavioural Tests
2.4.1. Open Field Experiment
The open field test is a method to evaluate the autonomous activities; exploratory behaviour and anxiety-like behaviour in the new environment. The experimental device is composed of a square plastic box with a bottom area of 100*100 cm2 and a height of 60 cm. The open box is grey black. The bottom of the open field is divided into 16 square grids by black solid lines; and the middle 4 grids are defined as the central area. The open field lighting was artificial lighting; and the experiment was carried out in a quiet environment. Each rat was placed in the centre of the open field in turn and allowed to explore freely for five minutes in the open field area; and relevant parameters were recorded. Specific operations; such as Qin X; were carried out as previously described by Godin et al. [12].
2.4.2. New Object Recognition Experiment
The new object recognition experiment is a learning and memory test method based on the principle that rats have the innate tendency to explore new objects. This method is a highly validated method for memory recognition that can approximately simulate human learning and memory ability under free activities and evaluate the formation of long-term or short-term memory mechanisms of test animals by identifying the shape and size changes of objects. Specific operations, such as Ennaceur, were carried out as previously described by Godin et al. [13].
2.4.3. High Plus Maze Experiment
The elevated plus maze is widely used behavioural test for anxiety behaviour and curiosity about new and different environments to detect the anxiety and exploration state of animals. The elevated plus maze consisted of a pair of relatively open arms (50 cm long × L0 cm wide) and a pair of relatively closed arms (50 cm long × L0 cm wide × 40 cm high) and a central area (10 cm × L0 cm). The whole body of the device was grey black, the material was made of a high-grade medical organic plate, and there was no reflective surface. Under normal conditions, rats have a preference for dark, small spaces. However, they can exhibit exploratory behaviours and explore the open arms. A decrease in open arm activity represents anxiety behavior. This method is widely used in neurobiology, psychology and other fields. Specific operations, such as Knight, were carried out as previously described by Godin et al. [14].
2.4.4. Morris Water Maze Test
The Morris water maze test is an experiment in which rats are forced to swim and learn to find a platform hidden in the water. As a classical experimental method for testing spatial learning ability and spatial memory ability, it is widely used in research on learning and memory, neurodegenerative diseases, hippocampal/extrahippocampal, toxicology, preventive medicine, neuroethology and other disciplines. It is also the preferred classical experimental method for behavioural research in medical colleges. The Morris water maze is mainly used to test the ability of experimental animals to sense spatial position and direction (spatial positioning). This part of the experiment consists of two experimental parts: positioning navigation and space exploration. The experiment lasts for seven days and is divided into three stages. Specific operations, such as Goudarzi, were carried out as previously described by Godin et al. [15].
2.5. HE/Nissl Staining
The preparation of tissue sections for staining included the following steps: sampling, fixation, washing and dehydration, transparency, sectioning, staining, and mounting.
HE staining: First, the sections were dewaxed and hydrated by soaking in xylene I for 15 min, xylene II for 10 min, absolute ethanol I for 5 min, absolute ethanol II for 5 min, 95%, 80% and 70% alcohol for 5 min, and rinsed with distilled water 3 times. Then the sections were immersed in haematoxylin solution for 5 min, rinsed with running water, differentiated with hydrochloric acid and alcohol for 1 s, and then returned to a blue colour with distilled water. Next, the sections were immersed in eosin solution for 3 min and rinsed with running water. Finally, dehydration and sealing were performed as follows: 75%, 85%, 95%, soaking in absolute ethanol I and II for 2 min for gradient dehydration, and soaking in xylene I and II for 4 min, respectively. Neutral gum was used to mount the slides, and light microscopy was used to observe the tissue and capture images. The basic steps of Nissl staining are the same as those of HE staining.
2.6. Immunohistochemistry
The main steps included the following: sectioning, dewaxing and hydration; antigen repair; drawing a circle around the tissue with an immunohistochemical pen to identify region of interest; incubation with 3% hydrogen peroxide at 37 °C for 15 min, and rinsing with PBS; blocking; rinsing with PBS 3 times for 5 min each time, adding a horseradish peroxidase-labelled goat anti-rabbit IgG, and incubating in a 37 °C oven for 20 min, rinsing with PBS 3 times for 5 min each time; adding haematoxylin dye solution to stain for 5 min, rinsing with tap water, rising with hydrochloric acid and alcohol, and rinsing with distilled water for blue staining; dehydration, sealing and drawing acquisition.
2.7. Transmission Electron Microscopy
The main steps include sample fixation in glutaraldehyde; osmic acid fixation; dehydration; paraffin-embedding. sectioning and antigen retrieval; staining; observation and image capture.
2.8. WB
Protein expression was assayed following the method of Jamal et al. [16]. Briefly, hippocampal tissue was removed from the -80 °C ultralow temperature refrigerator, accurately weighed, and the weight of the tissue was recorded. The tissue was placed in a low-temperature grinding tube. The lysis reagent was prepared with PMSF solution at a ratio of 100:1, and the tissue was ground, placed on ice for 30 min, and centrifuged at 12000 r/min for homogenization. The sample was centrifuged at 4 °C for 10 min, and the supernatant protein sample was collected. The total protein concentration was quantified using a BCA protein kit (Biyuntian) according to the manufacturer’s instructions. The protein samples, PBS and buffer were added to the EP tube at 10 times the amount, heated at 100 °C for 10 min, denatured and stored at -20 °C for future use. The prepared samples (15 µl protein/lane) were electrophoresed on 12% SDS‒PAGE, electroblotted onto PVDF membranes (Thermo Company, U.S.A.) and incubated with 3% BSA (Biopped Biotechnology Company, China) on a shaking table for 1 hour. After subsequent washing, the blots were incubated with primary antibody (Table 1) for 24 h at 4 °C followed by incubation with horseradish peroxidase-linked secondary antibody (anti-rabbit IgG, 1:2000) at room temperature for 60 min. After incubation, the secondary antibody was recovered according to the recommended procedure, and an appropriate amount of TBST (Shanghai double helix Biotechnology Co., Ltd., China) was added for elution.
2.9. Q-PCR
Refer to the specific steps of Hong-qiang Chen [17] for experimental operation. In short, total RNA was extracted using TRIzol reagent. RNA (2 µg) was reverse transcribed with the GoScript™ Reverse Transcription System (Promega, USA) according to the manufacturer’s instructions. Real-time qPCR was performed on a fluorescent quantitative PCR instrument (Bio-Rad, USA) using a fluorescent quantitative kit (Promega, USA). The sequences of primers used for RT‒PCR are shown in Table 2. The data were analysed with respect to a calibrator sample using the 2-△△Ct method. All assays were conducted at least in triplicate.
2.10. ATP/Glu Test
The instructions of the ATP/Glu test kit were followed.
2.11. Statistical Analysis
SPSS 26.0 software was used to sort and analyse the data. The data were expressed as ± SD. ANOVA was used for comparisons among multiple samples, repeated measurement ANOVA was used for repeated measurement data, the Kruskal‒Wallis test was used for nonparametric data, and Pearson correlation analysis was used for correlation tests. Significance level was p≤0.05. The grey values of the WB bands were analysed by ImageJ, the fluorescence intensity was analysed by Image-Pro Plus, and the graphics software was GraphPad Prism 8.0.1.
3. Results
3.1. Learning and Memory Ability and Hippocampal Neuron Damage Induced by Cy Exposure in Rats
3.1.1. General Changes in Rats Exposed to Cy
During the whole experimental process, the growth and development of rats in each group were not significantly abnormal, the body shape was round, the hair colour was smooth and bright, the nutritional status was stable, and the diet, water consumption and urine output were all normal. Moreover locomotor ability was suitable, and the activity levels were normal. However, the resting state of rats in the middle- and high-dose exposure groups was higher than that of control rats, and they were more prone to irritability when subjected to intragastric administration and stimulation. With the increase in exposure days, the body weight of rats in each group showed a growing trend. The repeated measurement test results showed that there were differences in the body weight growth of rats on different exposure days (F=103.128, p<0.001). Due to the effect of Cy exposure, the body weight growth of the medium- and high-dose groups was hindered in the middle period of exposure, and the body weight growth slowed down significantly, but there was no significant difference in body weight growth among the groups (f=0.805, p>0.05) (Figure 1 and Figure 2).
3.2. Changes in Learning and Memory Ability in Cy-Exposed Rats
3.2.1. Open Field Experiment
The results showed that in the open field experiment of rats, before Cy exposure, there was no significant difference in the central grid residence time, the number of grid crossings, the time of grooming and the number of standing in each exposure group (p>0.05). After Cy exposure, the central grid residence time in the middle- and high-dose Cy exposure groups significantly increased compared with that in the control group (p<0.05, p<0.001), while the number of grid crossings in the middle- and high-dose Cy exposure groups significantly decreased compared with that in the control group (p<0.05, p<0.05). the time of grooming and the number of standing showed a gradual downwards trend in each exposure group compared with the control group, and the difference was statistically significant (p<0.05). See Figure 3 for details.
3.2.2. New Object Recognition Experiment
The results showed that in the new object recognition experiment, before Cy exposure, there was no significant difference in the new object recognition index (DI) between the rats in each treatment group and the control group (p>0.05). The DI values were significantly decreased in the low, medium and high Cy exposure dose groups compared with the control group, (p<0.05, p<0.05, p<0.001). See Figure 4 for the specific results.
3.2.3. Elevated Plus Maze Test
The results showed that in the elevated cross maze test, there was no significant difference in the percentage of open arm retention time or the number of open arm entries between the groups before Cy exposure (p>0.05). After Cy exposure, the percentage of open arm retention time and the number of open arm entries of rats in the middle and high Cy exposure dose groups significantly decreased compared with those of the rats in the control group, (p<0.001, p<0.001, p<0.05, p<0.001). See Figure 5 for details.
3.2.4. Morris Water Maze Test
The Morris water maze experiment is one of the most commonly used research tools in behavioural neuroscience. It is often used to verify rodent neurocognitive disease models and evaluate the efficacy of neurocognitive drugs.
- (1)
- Directional navigation experiment
The escape latency was used to evaluate the spatial learning and memory ability of rats. The results of the repeated-measures ANOVA showed that in the directional navigation experiment, with the extension of training time, the escape latency of rats in each group was shortened, and the learning ability increased. The difference was statistically significant (p<0.001), indicating that the extension of training time could improve the spatial learning and memory ability of rats in each group. Compared with that of rats in the control group, the escape latency of rats in the exposure groups was significantly prolonged in a dose-dependent manner (p<0.001), indicating that Cy exposure may reduce the spatial learning and memory ability of rats. However, there was no interaction between Cy exposure dose and training time (p>0.05), indicating that training time had no effect on the change in escape latency and the change in the learning and memory ability of rats induced by Cy exposure. See Figure 6 for details.
- (2)
- Space exploration experiment.
The spatial exploration memory ability of rats was expressed by the spatial exploration index in the water maze. The experimental results of spatial exploration ability showed that there was no significant difference in the target quadrant residence time or the number of target platform crossings in the low-dose Cy exposure group compared with the control group (p>0.05), indicating that the effect of low-dose Cy exposure on the spatial exploration ability of rats was not significant. However, the target quadrant residence time and target platform crossing times significantly decreased in the middle- and high-dose Cy exposure groups compared with the control group (p>0.05), indicating that the increase in Cy exposure dose had a certain reducing effect on the spatial exploration ability of rats. See Figure 7 for details.
3.3. Injury of Hippocampal Neurons in Rats Exposed to Cy
3.3.1. HE Staining
After the water maze, whole brain pathological sections fixed in paraformaldehyde, paraffin-embedded and stained with HE to verify whether Cy exposure caused damage to neurons in the hippocampus of rats. The results showed that the cytoplasm of the hippocampal CA1 neurons was intensely stained in the different dose exposure groups, especially in the medium- and high-dose Cy exposure groups compared with the control group. Moreover, the inclusions were smaller and degraded, the arrangement of neurons was loose and disordered, the structure of neurons was disordered, the inclusions swelled, pyknosis was present in the nuclei, and vacuolization was present (Figure 8A). Moreover, the number of neurons in the Cy-exposed groups decreased significantly compared to that in the control group (p<0.001) (Figure 8B), and the nuclei of some cells completely degraded, forming ghost cells. In conclusion, it can be seen that with increasing Cy exposure dose, the degree of neuronal damage in the rat hippocampus is progressively exacerbated, which preliminarily verifies the morphological damage effect of Cy exposure on the rat nervous system.
3.3.2. Nissl Staining
After the water maze test, the whole brain was fixed with paraformaldehyde, embedded in paraffin and sectioned for Nissl staining. Nissl staining detects the Nissl bodies of nerve cells, which could further reveal the damage caused by Cy exposure to the morphology and number of neurons in the hippocampus. The results showed that, compared with the control group, the low-dose Cy-exposed group had only a small number of hyperchromatic nuclei and a decreased number of Nissl bodies, but the arrangement was relatively neat, and Nissl bodies were still visible in most of the neuronal envelopes. Compared with those in the control group, the hippocampal neurons in the middle- and high-dose Cy-exposed groups were mostly absent, disorderly and loosely arranged, while the plasma was intensely stained and the number of Nissl bodies was significantly reduced. Microscopically, Nissl corpuscles were dark blue in shape; the nuclei were pale blue, and the background is basically colourless (Figure 8C). By counting the number of intact Nissl bodies in rat hippocampal neurons, it was found that the number of Nissl bodies decreased significantly with increasing exposure dose (p<0.05) (Figure 8D), which further showed that Cy exposure had a damaging effect on rat hippocampal neurons.
3.3.3. Electron Microscopy
To further observe the damage of hippocampal neurons in rats exposed to Cy, scanning electron microscope pictures were captured to observe the ultrastructural changes of hippocampal neurons. The electron microscopy results showed that the cell inclusions and nuclei of hippocampal neurons in the control group were full and round, the double-layer nuclear membrane was clearly visible, and each organelle was clear and complete, in good condition, without any abnormalities. However, with increasing Cy exposure dose, hippocampal neurons showed different degrees of damage, especially in the medium- and high-dose groups. The cell membrane and nuclear membrane of hippocampal neurons were severely shrunk and deformed, the double-layer nuclear membrane was blurred, the number of organelles were seriously reduced and the morphology was abnormal, and mitochondrial ridge breakage and lysosome phagocytosis of lipid droplet vesicles was visible (Figure 8E). These findings further show that the morphological damage of hippocampal neurons in Cy-exposed rats also causes ultrastructural changes that can then affect energy metabolism, information transmission and other cellular functions.
3.4. Damage to the Synaptic Plasticity of Hippocampal Neurons Induced by Cy Exposure in Rats
3.4.1. Ultrastructural Changes in the Synapses of Hippocampal Cells in Cy-Exposed Rats
The electron microscopy results showed that in the control group, the number of synapses in hippocampal neurons was high, the synaptic structure was clear and complete, the presynaptic membrane, synaptic vesicles and postsynaptic modules were clearly visible, and the synaptic gap was moderate. With an increase in exposure dose, the number of synapses in hippocampal neurons gradually decreased, and the normal synaptic structure changed, mainly manifested in the disappearance of synaptosomes and the degradation of presynaptic and postsynaptic membranes. The synaptic space became narrower, the synaptic vesicles decreased or disappeared, and the overall synaptic morphology showed ground-glass opacity. The area indicated by the arrow is the number and morphological changes of synaptic vesicles at the presynaptic membrane terminals (Figure 9A).
Western blot detection of the synaptic marker protein PSD-95 and synaptic vesicular protein SYP showed that, compared with that in the control group, the expression of PSD-95 in the middle- and high-dose Cy-exposed groups was significantly lower (p<0.05), while the expression of the synaptic vesicular protein SYP in the hippocampus of rats in all exposed groups showed a downwards trend compared with that in the control group. The difference was statistically significant (p<0.05). The changes in PSD-95 and SYP gene mRNA expression were consistent with those in their protein expression (Figure 9E,F). The detection results of ATP and the neurotransmitter Glu in the rat hippocampus showed that compared with that in the control group, the ATPase level in the hippocampus decreased with increasing Cy exposure dose (p<0.05) (Figure 9H). Compared with that in the control group, the level of the neurotransmitter Glu in the hippocampus increased with increasing exposure dose, and the difference was statistically significant (p<0.05) (Figure 9G).
In conclusion, Cy exposure can cause certain damage to the synaptic plasticity of hippocampal neurons from the aspects of morphology and the expression of key synaptic proteinsn and has a negative impact on the neurotransmitter release function of synapses.
3.5. Changes in Inflammatory Factors and A2AR Factors in the Neurons of Rats Exposed to Cy
3.5.1. Expression of Inflammatory Factors in the Hippocampus of Cy-Exposed Rats
The protein expression of inflammatory-related factors in the hippocampus was detected to explore the further damaging effect of Cy exposure on the hippocampus. The Western blot results showed that compared with the control treatment, medium and high doses of Cy exposure upregulated the inflammatory factor IL-6, and the difference was statistically significant (p<0.05). At the same time, compared with the control treatment, low, medium and high doses of Cy upregulated the inflammatory factor TNF-α. The difference was statistically significant (p<0.05) (Figure 10B).
At the same time, the mRNA expression corresponding to inflammatory factors was measured by Q-PCR. The results showed that compared with the control treatment, medium and high doses of Cy exposure could upregulate the inflammatory factors IL-6 and TNF-α. The difference in mRNA expression was statistically significant (p<0.05) (Figure 10B).
3.5.2. Expression of the A2AR Factor in the Hippocampus of Cy-Exposed Rats
The correlation between the damage of hippocampal tissue caused by Cy exposure and the expression of A2AR was explored, and the immunohistochemical analysis of hippocampal tissue of rats in each group showed that positive A2AR immunohistochemical staining was found in the cytoplasm of neurons (Figure 10A). Compared with that in the control group, the number of A2AR-positive cells was significantly increased in the middle- and high-dose Cy exposure groups (p<0.001) (Figure 10B). At the same time, the Q-PCR results showed that compared with that in the control group, the expression of A2AR mRNA in the medium- and high-dose Cy exposure groups was downregulated, and the difference was statistically significant (p<0.05) (Figure 10B,C).
In conclusion, Cy exposure can affect the expression of A2AR in the rat hippocampus and cause changes in energy metabolism and abnormal release of neurotransmitters.
3.6. Correlation Analysis of A2AR with Neurobehavioural Indices, Inflammatory Factors and Synaptic Plasticity Factors
The results of the Pearson correlation analysis showed that the expression of A2AR mRNA was positively correlated with the central lattice residence time in the open field test and the average latency in the water maze test (p<0.001) and negatively correlated with other neurobehavioural indices (p<0.05). At the same time, it can be seen that the expression of A2AR mRNA has a strong correlation with the corresponding indicators of the open field experiment, new object recognition experiment and water maze test. These three experiments mainly represent the recognition memory ability, spatial memory ability and spatial exploration ability of rats. In addition, the expression of A2AR mRNA was positively correlated with the expression of inflammatory factors (p<0.05) and negatively correlated with the expression of synaptic plasticity factors (p<0.05).

Table 3.
Correlation analysis between A2AR mRNA expression and related indicators.
| Experimental category | Indicators | Pearson correlation coefficient r | P |
|---|---|---|---|
| Open field test | |||
| Central grid dwell time | 0.7154 | <0.001 | |
| The time of grooming | -0.6354 | 0.001 | |
| Number of standing | -0.4381 | 0.032 | |
| Number of grid crossings | -0.5682 | 0.004 | |
| New object recognition experiment | |||
| Discrimination index | -0.6896 | <0.001 | |
| Plus maze experiment | |||
| Open arm entry times | -0.375 | 0.071 | |
| Open arm entry time | -0.4633 | 0.023 | |
| Water maze test | |||
| Average latency | 0.7000 | <0.001 | |
| Target Quadrant Dwell Time | -0.6044 | 0.002 | |
| Target platform traversal times | -0.6364 | 0.001 | |
| Inflammation related factors | |||
| IL-6 | 0.8612 | <0.001 | |
| TNF | 0.8433 | 0.001 | |
| Synaptic plasticity related factors | |||
| PSD-95 | -0.6377 | 0.026 | |
| SYP | -0.7935 | 0.002 | |
4. Discussion
Cy is an important type II pyrethroid pesticide. Because its molecular formula contains a benzene ring, Cy has a certain fat solubility, and some Cy entering the body can pass through the blood‒brain barrier and accumulate in brain tissue, which has certain neurotoxicity [9]. In this study, there was no significant difference in the weight and development status of rats in each group. However, compared with the control group, the rats in the medium- and high-dose exposure groups were more sluggish and were more prone to irritability during intragastric administration and stimulation.
4.1. Cy Exposure Can Cause Neurobehavioural Changes in Rats
Normal behavioural function is the concrete embodiment of the complete morphological structure and physiological function of the nervous system. Damage to the nervous system can be directly reflected in abnormalities in neurobehavioural function [18]. Therefore, the most direct and apparent method to evaluate the damaging effect of neurotoxins on the nervous system to perform animal neurobehaviour tests. Studies have also shown that neurobehavioural abnormalities caused by neurotoxins generally occur earlier than neurobiochemical and pathological indicators, so they are also more sensitive [19]. In this study, four neurobehavioural experiments were conducted to detect the degree of neurobehavioural damage in rats exposed to Cy.
The open field experiment is a behavioural experiment that reflects the motor exploration ability and spatial cognitive ability of experimental animals. It is a method to evaluate the exploratory behaviour and anxiety level of rats in unfamiliar environments [20]. The results showed that there was no significant difference in each index of the open field experiment of rats in each group before exposure, indicating that the randomization was consistent. After exposure, the number of grid crossings, the time of grooming and the number of standing decreased to varying degrees in the Cy-exposed group, while the central grid residence time was significantly prolonged. These changes were dose-dependent, indicating that with increasing exposure dose, the exploration ability of rats in the new environment became weaker. At the same time, the excitement and exploration ability of their independent activities decreased. Moreover, some studies have also shown that when the learning and memory ability of rats is abnormal, their corresponding exploration and activity ability decreases [21].
The new object recognition experiment tests the ability of rats to explore and recognize novel objects [13]. The results showed that there was no difference in the discrimination index of rats in each group before exposure, but with the increase in exposure dose and exposure days, especially in the high-dose Cy exposure group, the recognition of new objects decreased, indicating that Cy exposure reduced the recognition ability of rats to new objects and damaged their memory. This finding is consistent with the results of Hughes, MF [22].
The elevated plus maze is a behavioural test that reflects the exploratory ability and anxiety state of rats [14,23]. The results of this study found that there was no difference in the number and time of rats entering the open arm before exposure, but after exposure, the number of times that the rats entered the open arm in the medium- and high-dose groups was significantly shortened, indicating that the exploration ability of rats decreased and the degree of anxiety increased. Moreover, some studies have shown that rats with reduced times and time of entering the open arm of the cross maze may be related to brain cognitive damage and abnormal neurotransmitter release [23].
The Morris water maze test is a classic behavioural experiment to test the spatial learning ability and spatial memory ability of rats. It is one of the most commonly used methods to evaluate the impairment of learning and memory ability of rats [15] and is also known as the gold standard of learning and memory behavioural testing [24]. It is a practical, reliable and easy-to-implement method designed by Morris in 1981 [25]. In this experiment, the rats were trained for 5 days after exposure. The results showed that the escape latency of rats in each group could be shortened through continuous intensive training, while the longitudinal comparison of the escape latency of rats in different groups showed that the escape latency also increased with increasing exposure dose, with a dose-dependent relationship, indicating that Cy exposure reduced to the spatial learning ability of rats. On the sixth day of training, the water maze space exploration test was carried out. The results showed that the times of crossing the original platform and the residence time in the target quadrant in the medium- and high-dose exposure groups were significantly shorter than those in the control group, indicating that Cy exposure not only damaged the spatial learning ability but also the short-term spatial memory ability of the rats, with certain neurotoxicity. This finding is consistent with previous Syed f studies [26]. At the same time, since the hippocampus mainly controls the long-term and short-term memory and learning ability of the body [27], it can be speculated that Cy exposure may damage the hippocampus of rats, thus damaging the learning and memory ability and causing nervous system diseases.
4.2. Cy Exposure Can Cause Morphological Changes in Rat Hippocampus
Changes in function and behaviour may be accompanied by changes in tissue morphology. HE staining can intuitively evaluate the damage degree of toxic substances to tissue morphology. Therefore, HE staining was used to observe the changes in hippocampal tissue morphology after Cy exposure in this study. The results of HE staining showed that with increasing exposure dose, especially in the high-dose exposure group, the cells in the hippocampal CA1 area exhibited serious vacuolization, pyknosis, disordered arrangement and loss, which further indicated that Cy exposure caused serious damage to the morphology of rat hippocampal tissue. Many studies have shown that many neurodegenerative diseases such as Alzheimer’s disease have irregular arrangement and loss of neurons [28,29]. We should be more alert and suspect that Cy exposure may also be one of the environmental triggers of neurodegenerative diseases. To further observe neuronal damage, Nissl staining was used to observe the distribution and morphological characteristics of Nissl bodies [30]. The Nissl body is the main substance for protein synthesis, which can balance a variety of neurotransmitters in the brain and maintain the stability of neuronal excitability. Nissl corpuscles are very sensitive. When neurons are damaged, the nucleus begins to dissipate and dissolve. At the same time, Nissl corpuscles develop around the neurons and finally dissolve and dissipate in a vacuole shape. Therefore, Nissl bodies can be regarded as one of the important signs of neuronal damage [31]. The Nissl staining results showed that compared with those in the control group, the hippocampal neurons in the medium- and high-dose exposure groups were lost and disorderly and loosely arranged, while the plasma was intensely stained and the number of Nissl bodies was significantly reduced. It has been confirmed that Cy exposure can cause dissolution and vacuolation of Nissl bodies in hippocampal neurons and damage to neurons. In addition, the ultrastructure of rat hippocampal tissue was observed by transmission electron microscopy. With increasing exposure dose, the cell membrane and nuclear membrane of hippocampal neurons were severely shrunk and deformed, the double-layer nuclear membrane was blurred, and the morphology and number of organelles were seriously damaged. Moreover, mitochondrial ridge fragmentation and lysosome phagocytosis of lipid droplet vesicles was observed. As the energy factory of cells, mitochondria are responsible for energy transformation and transmission. Damaged mitochondria seriously affect the energy metabolism and normal information transmission of cells [32], while nuclear pyknosis and nuclear membrane structure damage further affect the replication of DNA and the normal operation of cells, resulting in cellular dysfunction, inflammation, apoptosis and other consequences.
In conclusion, whether observing the function of organelles and nuclei or the pathological morphology of cells, Cy exposure causes irreversible damage, which may seriously affect the normal function of cells.
Cy exposure can cause abnormal synaptic plasticity, energy transmission and neurotransmitter release in the rat hippocampus. Brain tissue is a high energy-consuming tissue, and its energy metabolism is of great importance for the normal operation of brain tissue, the transmission of information, and learning and memory [33]. In the ultrastructure, it was observed that the mitochondria of hippocampal neurons were seriously damaged, and the decrease in ATP levels, as the “energy currency” of intracellular energy transfer, could directly explain the disorder of cellular energy metabolism. Therefore, we determined the levels of ATPase in hippocampal tissue. The results showed that the level of ATPase in hippocampal tissue decreased in a dose-dependent manner, further indicating that Cy exposure had an impact on the morphological structure of hippocampal neurons. Changes in ultrastructure will damage the energy metabolism of cells. Moreover, the changes in the structure and function of the hippocampus are related to the morphology of dendrites and dendritic spines. Dendritic morphology is an important basis for establishing neural circuits. Changes in dendritic morphology are closely related to synaptic plasticity [34]. Dendritic morphology is also closely related to learning and memory [35]. Qingqing Li also found that the synaptic plasticity of hippocampal neurons is closely related to the learning and memory ability of rats [36]. Synaptic plasticity represents one of the most basic and important functions of the brain. It represents the ability to perceive, evaluate and store complex information. At the same time, it can make adaptive responses to related stimuli, such as energy metabolism disorders, inflammation and apoptosis. Studies have shown that changes in the number and morphology of synapses can directly affect the organizational structure and function of the hippocampus and have a profound impact on learning, memory and cognitive function [37]. Changes in synaptic plasticity include changes in structural plasticity and functional plasticity, that is, changes in synaptic transmission efficiency and changes in synaptic morphological structure [38]. This paper mainly discusses the plasticity of synaptic structure. The transmission electron microscopy results showed that, compared with the control treatment, with the increase in exposure dose, the number of synapses in hippocampal neurons gradually decreased, and the normal synaptic structure changed, mainly manifested in the disappearance of synaptosomes, the blurring of presynaptic and postsynaptic membranes, the narrowing of synaptic spaces, the reduction or disappearance of synaptic vesicles, and the overall synaptic morphology showed ground-glass opacity. This finding further confirms the theory that changes in neuronal cell morphology are bound to be accompanied by changes in neuronal synaptic plasticity morphology and function [39]. Then, we determined the protein and mRNA expression of key proteins of synaptic plasticity: postsynaptic density PSD-95 and synaptophysin SYP. The results showed that the expression of postsynaptic density PSD-95 was significantly downregulated in the middle- and high-dose Cy exposure groups compared with the control group, while the expression of synaptophysin SYP protein in the hippocampus of rats in each exposure group showed a downwards trend compared with the control group. The results of Q-PCR were consistent with those of Western blot. PSD-95 is an important postsynaptic compactor protein. The transmission of information between synapses, synaptic plasticity, the formation of synapses and the initiation of excitatory synapses all play key regulatory roles [40,41]. The decrease in the expression level of PSD-95 directly reflects the state of synaptic plasticity and the normal function of synapses. Synaptophysin SYP, a vesicular membrane protein located at the end of axons, reflects the number and distribution of synapses [42]. PSD-95 and synaptophysin SYP are not only representatives of synaptic membranes but also of synaptic plasticity, which is related to the formation and maturation of synapses. The loss of synapses may be the reason for the decreased secretion of the postsynaptic densification protein PSD-95 and synaptic vesicular peptide SYP [43]. After the structure and number of synapses are damaged, the transmission of neurotransmitters, one of the important functions of synapses, is affected. Abnormal release of neurotransmitters may affect subsequent intercellular dysfunction. Previous studies also found that Cy exposure can cause the abnormal release of neurotransmitters in rats [44]. In this study, it was found that compared with the control group, the release of Glu in the medium- and high-dose groups showed a significant upwards trend. A large amount of Glu release can cause overexcitation of neurons and eventually cause damage [45]. Injury causes a large amount of Na+ and Cl- to flow into the cells, causing swelling and necrosis of neurons. Moreover, Glu can act on the glutamate receptor on the postsynaptic membrane, resulting in the opening of receptor-dependent Ca2+ channels, a large amount of Ca2+ influx, a large amount of Ca2+ release from the endoplasmic reticulum, and a serious overload of intracellular Ca2+ leading to neuronal necrosis. In conclusion, Cy exposure can cause changes in the morphology and structure of hippocampal neurons, causing changes in energy metabolism, damaging the synaptic plasticity of neuronal cells, affecting the release of neurotransmitters, and changing cell morphology and function.
4.3. Cy Exposure Can Cause Abnormal Changes in the Inflammatory Response and Adenosine A2AR Expression in the Rat Hippocampus
A large number of studies have shown that damage to hippocampal neurons can be accompanied by the occurrence of hippocampal inflammation [46,47,48]. TNF-α is the earliest and most important inflammatory factor in the inflammatory response. It can induce nerve injury by mediating neuroinflammation and promoting T cells to produce various inflammatory factors [49]. IL-6 has a wide range of biological activities, which can regulate the immune response and promote the precursor of B cells to become antibody-producing cells. In coordination with colony stimulating factor, it can promote the growth and differentiation of primitive bone marrow-derived cells and enhance the lysis function of natural killer cells, which is of great importance for the reduction of inflammation, neuronal degeneration and necrosis [50]. In this study, the expression of two important inflammatory factors, TNF-α and IL-6, in the hippocampus was determined. Compared with the control group, TNF-α and IL-6 protein and mRNA were significantly upregulated, suggesting that Cy is likely to induce a neuroinflammatory reaction in the rat hippocampus, which is basically consistent with the previous results of Shang Jiaqi [51]. Studies have reported that A2AR can mediate a variety of physiological and pathological processes, including apoptosis, inflammation and synaptic plasticity [52,53]. Chen Jiangfan found that a large number of A2AR-positive neurons can be seen in brain tissue after cerebral ischaemia, which confirmed that A2AR is closely related to cerebral ischaemic injury [54] At the same time, some scholars also found that inhibiting the expression of A2AR can alleviate brain tissue damage, and this protective effect is accompanied by the reduction of TNF-α levels in brain tissue. The downregulation of inflammatory factors, including IL-6, can reduce the damage of neurons in brain tissue [55]. The results also showed that the protein and mRNA expression of A2AR in Cy-exposed rats, especially in the middle- and high-dose groups, was significantly higher than that in control rats. At the same time, in the Pearson correlation analysis, it was found that there was a close relationship between the expression of A2AR mRNA and various indicators of neurobehavioural experiments, and the expression of A2AR mRNA was positively correlated with the levels of inflammatory factors and apoptosis factors. However, it was negatively correlated with the levels of synaptic plasticity factors, which further suggests that A2AR may be involved in the process of neuronal injury in the hippocampus induced by Cy exposure, but the specific mechanism needs to be further explored.
5. Conclusions
The morphological and physical signs of Cy-exposed rats were not significantly abnormal compared with those of the control group; however, the neurobehavioural tests showed that the learning and memory ability, motor exploration ability decreased and anxiety-like behaviours increased with increasing exposure dose.
The pathological sections showed that the middle and high doses of Cy exposure caused deformation, reduction and disorder of hippocampal pyramidal cells, a decrease in Nissl body numbers, pyknosis of the hippocampal cell nuclear membrane and serious damage to organelles, indicating that Cy exposure at this dose may cause hippocampal tissue damage in rats.
In this study, with increasing exposure dose, the morphological changes in hippocampal synapses were more obvious, including blurred synaptic space, decreased synaptic vesicles and a decreased number of synapses. At the same time, the dose-dependent expression of the key synaptic proteins PSD-95 and SYP also decreased, indicating that synaptic damage was obvious.
In this study, medium and high doses of Cy exposure upregulated the expression of A2AR in hippocampal tissue. At the same time, the expression levels of inflammatory factors and apoptosis factor-related proteins increased in a dose-dependent manner, and there is a certain correlation between the expression of A2AR mRNA and neurobehavioural indicators, inflammatory factors and synaptic plasticity factors, suggesting that Cy exposure may cause nerve damage in rats and has a close relationship with A2AR.
Author Contributions
Conceptualization: H.-F.Y. and Y.-X.X. Data curation: Y.-X.X., J.Z. Formal analysis: J.S., Y.-X.X. and X.-Y.L. Funding acquisition: H.-F.Y. and Y.-X.X. Methodology: H.-F.Y., J.Z. and Y.-X.X. Writing—Original draft: Y.-X.X. Review and Editing: H.-F.Y. and Y.-X.X. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Natural Science Foundation of the Ningxia Hui Autonomous Region (grant number 2023AAC03153), and the National Natural Science Foundation of China (grant number 81960584).
Institutional Review Board Statement
The animal study protocol was approved by the Laboratory Animal Ethical and Welfare Committee of the Laboratory Animal Center, Ningxia Medical University (Approval No.: IACUC-NYLAC-2021-101). This study was approved by the Ethics Committee of Ningxia Medical University(No. 2021-N093).
Informed Consent Statement
No conflict of interest exists in the submission of this manuscript, and manuscript is approved by all authors for publication.
Data Availability Statement
All data included in this study are available upon request by contact with the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- YC, L.; WZ, L.; CC, K.; LJ, H.; CT, C.; CR, J. Action of the insecticide cyfluthrin on Ca signal transduction and cytotoxicity in human osteosarcoma cells. Hum Exp Toxicol 2020, 39, 1268-1276. [CrossRef]
- FAO; Specifications, R.I.P.O.; Standards, R.A.; AGP. Manual on the Development and Use of FAO Specifications for Plant Protection Products. 1999.
- F, H.; T, K. Pesticide residues and health risk appraisal of tomato cultivated in greenhouse from the Mediterranean region of Turkey. Environmental science and pollution research international 2021, 28, 22551-22562. [CrossRef]
- MK, M. Dietary Pyrethroid Exposures and Intake Doses for 188 Duplicate-Single Solid Food Items Consumed by North Carolina Adults. Toxics 2020, 8. [CrossRef]
- W, N.; H, G.; B, W.; J, Z.; J, S.; Y, S.; Y, S.; H, Y. Gestational Exposure to Cyfluthrin through Endoplasmic Reticulum (ER) Stress-Mediated PERK Signaling Pathway Impairs Placental Development. Toxics 2022, 10, 733. [CrossRef]
- J, Z.; LY, M.; YX, X.; LQ, Z.; WS, N.; R, W.; YN, S.; XY, L.; HF, Y. The role of stimulator of interferon genes-mediated AMPK/mTOR/P70S6K autophagy pathway in cyfluthrin-induced testicular injury. Environ Toxicol 2022, 38, 727-742. [CrossRef]
- L, M.; E, Z.; M, I. β-Cyfluthrin-Mediated Cytotoxicity of Cultured Rat Primary Hepatocytes Ameliorated by Cotreatment with Luteolin. Evidence-based complementary and alternative medicine: eCAM 2022, 2022, 3647988. [CrossRef]
- Xiaoyu, Z.; Bing, W.; Lijun, D.; Ji, Z.; Huifang, Y. Analysis on the current status and influencing factors of neurological subhealth among vegetable greenhouse growers in the suburbs of Yinchuan City. Journal of Ningxia Medical University 2017, 039, 293-298. [CrossRef]
- D, D.; S, K.; N, B.; R, E.; M, C.; RP, S.; M, S.; JO, G.; J, B.; V, K. Chlorpyrifos, permethrin and cyfluthrin effect on cell survival, permeability, and tight junction in an in-vitro model of the human blood-brain barrier (BBB). Neurotoxicology 2022, 93, 152-162. [CrossRef]
- M, V.; P, Q.; K, A.; C, M.; P, G.; C, B.; A, C. Aggregate and cumulative chronic risk assessment for pyrethroids in the French adult population. Food and chemical toxicology: an international journal published for the British Industrial Biological Research Association 2020, 143, 111519. [CrossRef]
- W, Z.; R, F.; S, L.; Y, L.; Y, J.; Y, L.; B, L.; Y, C.; L, J.; X, Y. Combined effects of chlorpyrifos and cyfluthrin on neurobehavior and neurotransmitter levels in larval zebrafish. Journal of applied toxicology: JAT 2022, 42, 1662-1670. [CrossRef]
- X, Q.; L, L.; X, N.; Q, N. Effects of Chronic Aluminum Lactate Exposure on Neuronal Apoptosis and Hippocampal Synaptic Plasticity in Rats. Biol Trace Elem Res 2020, 197, 571-579. [CrossRef]
- Ennaceur, A.; Delacour, J.A. A new one-trial test for neurobiological studies of memory in rats. I. Behavioral data. Behav Brain Res 1988, 31, 47-59. [CrossRef]
- P, K.; R, C.; R, W.; A, B.; S, P.; AW, B. Sex differences in the elevated plus-maze test and large open field test in adult Wistar rats. Pharmacology, biochemistry, and behavior 2021, 204, 173168. [CrossRef]
- N, G.; S, M.V.; A, N.; M, M.; M, M.; S, S.; M, G.; S, V.; M, H. Pharmacological Evidences for Curcumin Neuroprotective Effects against Lead-Induced Neurodegeneration: Possible Role of Akt/GSK3 Signaling Pathway. Iranian journal of pharmaceutical research: IJPR 2020, 19, 494-508. [CrossRef]
- M, J.; K, A.; S, A.; J, M.; W, W.; M, K.; U, I.; T, M.; I, I. Changes in cholinergic function in the frontal cortex and hippocampus of rat exposed to ethanol and acetaldehyde. Neuroscience 2007, 144, 232-238. [CrossRef]
- A, H.Q.C.; B, J.Z.A.; C, Y.L.A.; D, L.X.H.; D, Y.J.H.; D, W.Q.S.; A, J.C.; A, W.B.L.; A, J.Y.L. Gene expression network regulated by DNA methylation and microRNA during microcystin-leucine arginine induced malignant transformation in human hepatocyte L02 cells. Toxicol Lett 2018, 289, 42-53. [CrossRef]
- IA, A.; SE, O.; AK, O.; EO, F. Neuroprotective role of gallic acid in aflatoxin B -induced behavioral abnormalities in rats. J Biochem Mol Toxic 2021, 35, e22684. [CrossRef]
- ED, L.; E, U.; RE, B. Neurobehavioral toxicology of halothane in rats. Neurotoxicol Teratol 1991, 13, 461-470. [CrossRef]
- Z, L.; Z, H.; J, L.; H, L.; L, M.; Y, L.; J, Z.; Y, X.; Y, L. Regulation of CRMP2 by Cdk5 and GSK-3β participates in sevoflurane-induced dendritic development abnormalities and cognitive dysfunction in developing rats. Toxicol Lett 2021, 341, 68-79. [CrossRef]
- FDS, C.; de Souza Oliveira Tavares C; BHS, A.; F, M.; RÁB, L.; S, G.D.S. Improved Spatial Memory And Neuroinflammatory Profile Changes in Aged Rats Submitted to Photobiomodulation Therapy. Cell Mol Neurobiol 2022, 42, 1875-1886. [CrossRef]
- Hughes, M.F.; Ross, D.G.; Starr, J.M.; Scollon, E.J.; Wolansky, M.J.; Crofton, K.M.; Devito, M.J. Environmentally relevant pyrethroid mixtures: A study on the correlation of blood and brain concentrations of a mixture of pyrethroid insecticides to motor activity in the rat. Toxicology 2016, 359, 19-28. [CrossRef]
- ES, C. Dopamine Release in the Midbrain Promotes Anxiety. Biol Psychiat 2020, 88, 815-817. [CrossRef]
- S, C.; J, D.; K, C.; D, L.; K, C.; M, C.; L, K.; D, F.; C, T. NavWell: A simplified virtual-reality platform for spatial navigation and memory experiments. Behav Res Methods 2020, 52, 1189-1207. [CrossRef]
- K, B.; Y, D.; W, S. Morris water maze test for learning and memory deficits in Alzheimer’s disease model mice. Journal of visualized experiments: JoVE 2011, 20, 2920. [CrossRef]
- F, S.; LP, C.; VK, K.; I, S. Beta-cyfluthrin induced neurobehavioral impairments in adult rats. Chem-Biol Interact 2016, 243, 19-28. [CrossRef]
- D, L.; L, Y.; M, L.; Q, Z.; F, T.; W, C. Behavioral disorders caused by nonylphenol and strategies for protection. Chemosphere 2021, 275, 129973. [CrossRef]
- J, W.; B, L.; Y, X.; M, Y.; C, W.; M, S.; J, L.; W, W.; J, Y.; F, S., et al. Activation of CREB-mediated autophagy by thioperamide ameliorates β-amyloid pathology and cognition in Alzheimer’s disease. Aging Cell 2021, 20, e13333. [CrossRef]
- M, G.; DM, H. Targeting pre-synaptic tau accumulation: a new strategy to counteract tau-mediated synaptic loss and memory deficits. Neuron 2021, 109, 741-743. [CrossRef]
- G, P.; V, S.; A, V.; N, K.; N, N.; M, G.; T, B.; A, Y.; I, M.; S, K. A combined use of silver pretreatment and impregnation with consequent Nissl staining for cortex and striatum architectonics study. Front Neuroanat 2022, 16, 940993. [CrossRef]
- Y, Y.; H, G.; W, L.; X, L.; X, J.; X, L.; Q, W.; Z, X.; Q, Z. Arctium lappa L. roots ameliorates cerebral ischemia through inhibiting neuronal apoptosis and suppressing AMPK/mTOR-mediated autophagy. Phytomedicine: international journal of phytotherapy and phytopharmacology 2021, 85, 153526. [CrossRef]
- Y, Y.; SW, Z.; SQ, W.; QP, Z.; L, W.; H, Z.; JH, G.; XZ, L.; JC, B.; ZP, L. Alpha-Lipoic Acid Attenuates Cadmium- and Lead-Induced Neurotoxicity by Inhibiting Both Endoplasmic-Reticulum Stress and Activation of Fas/FasL and Mitochondrial Apoptotic Pathways in Rat Cerebral Cortex. Neurotox Res 2021, 39, 1103-1115. [CrossRef]
- Z, C.; Z, Y.; S, Y.; Y, Z.; M, X.; J, Z.; L, L. Brain Energy Metabolism: Astrocytes in Neurodegenerative Diseases. Cns Neurosci Ther 2023, 29, 24-36. [CrossRef]
- I, G.; T, T.; AL, S.; F, Y.; J, L.; D, V.; MS, H.; J, N.; AU, B. Systematic expression analysis of plasticity-related genes in mouse brain development brings PRG4 into play. Developmental dynamics: an official publication of the American Association of Anatomists 2022, 251, 714-728. [CrossRef]
- De la Torre-Iturbe S; RA, V.; De la Cruz-López F; G, F.; L, G. Dendritic and behavioral changes in rats neonatally treated with homocysteine; A proposal as an animal model to study the attention deficit hyperactivity disorder. J Chem Neuroanat 2022, 119, 102057. [CrossRef]
- QQ, L.; J, C.; P, H.; M, J.; JH, S.; HY, F.; FC, Q.; YY, Z.; YY, S.; G, C., et al. Enhancing GluN2A-type NMDA receptors impairs long-term synaptic plasticity and learning and memory. Mol Psychiatr 2022, 27, 3468-3478. [CrossRef]
- HL, T.; SL, C.; Q, Z.; RL, H. GRIP1 regulates synaptic plasticity and learning and memory. P Natl Acad Sci Usa 2020, 117, 25085-25091. [CrossRef]
- SD, S.; L, F.; M, Z.; P, G. Synaptic Plasticity, Dementia and Alzheimer Disease. CNS & neurological disorders drug targets 2017, 16, 220-233. [CrossRef]
- A, T.; MI, Á.; A, T.; JJ, M.; JJ, R. Brain local and regional neuroglial alterations in Alzheimer’s Disease: cell types, responses and implications. Curr Alzheimer Res 2016, 13, 321-342. [CrossRef]
- T, X.; J, L.; XR, L.; Y, Y.; X, L.; X, Z.; Y, C.; PQ, Y.; Y, L. The mTOR/NF-κB Pathway Mediates Neuroinflammation and Synaptic Plasticity in Diabetic Encephalopathy. Mol Neurobiol 2021, 58, 3848-3862. [CrossRef]
- D, D.; Y, C.; S, G.; Z, X.; S, C.; K, C.; S, W.; G, S.; L, Y.; S, B., et al. Sinisan alleviates depression-like behaviors by regulating mitochondrial function and synaptic plasticity in maternal separation rats. Phytomedicine: international journal of phytotherapy and phytopharmacology 2022, 106, 154395. [CrossRef]
- MA, C. Synaptophysin-dependent synaptobrevin-2 trafficking at the presynapse-Mechanism and function. J Neurochem 2021, 159, 78-89. [CrossRef]
- J, Y.; J, G.; KQ, S.; XD, F. Electroacupuncture improves learning and memory impairment and enhances hippocampal synaptic plasticity through BDNF/TRKB/CREB signaling pathway in cerebral ischemia-reperfusion injury rats. Zhen ci yan jiu = Acupuncture research 2023, 48, 843-851. [CrossRef]
- JL, R.; I, A.; V, C.; M, M.; MR, M.; A, A.; MA, M. Effects of exposure to pyrethroid cyfluthrin on serotonin and dopamine levels in brain regions of male rats. Environ Res 2016, 146, 388-394. [CrossRef]
- Y, G.; N, L.; J, C.; P, Z.; J, N.; S, T.; X, P.; J, W.; J, Y.; L, M. Neuropharmacological insight into preventive intervention in posttraumatic epilepsy based on regulating glutamate homeostasis. Cns Neurosci Ther 2023, 29, 2430-2444. [CrossRef]
- M, W.; H, Z.; J, L.; J, H.; N, C. Exercise suppresses neuroinflammation for alleviating Alzheimer’s disease. J Neuroinflamm 2023, 20, 76. [CrossRef]
- KM, L.; Y, H.; PP, W.; YH, L.; N, Z.; JJ, L.; CY, Y.; JJ, C.; L, Z.; C, Z., et al. Ursolic Acid Protects Neurons in Temporal Lobe Epilepsy and Cognitive Impairment by Repressing Inflammation and Oxidation. Front Pharmacol 2022, 13, 877898. [CrossRef]
- S, S.; A, Z.; P, B.; F, Y.; N, A.; E, N. The methanolic extract of Cinnamomum zeylanicum bark improves formaldehyde-induced neurotoxicity through reduction of phospho-tau (Thr231), inflammation, and apoptosis. Excli J 2020, 19, 671-686.
- Y, Z.; Q, F.; J, C.; Y, W.; X, L.; X, Z.; Y, S.; H, Z.; X, Z.; M, Y., et al. Melatonin Improves Mitochondrial Dysfunction and Attenuates Neuropathic Pain by Regulating SIRT1 in Dorsal Root Ganglions. Neuroscience 2023, S0306-4522, 454-455. [CrossRef]
- CA, M.; CN, F.; CMG, L.; VG, F.; AC, C.; ACR, O.; de Souza LC; EPF, R.; KC, C.; HC, G., et al. Leptin, hsCRP, TNF-α and IL-6 levels from normal aging to dementia: Relationship with cognitive and functional status. Journal of clinical neuroscience: official journal of the Neurosurgical Society of Australasia 2018, 56, 150-155. [CrossRef]
- Jia-Qi, S.; Qi, W.; Chao, T.; Ru-Nan, Z.; Shuai, Z.; Xiao-Rong, Z. Study on effect of beta-cypermethrin on expression of TH and TNF-α in rat brain tissue. Occupation and Health 2019, 35, 2187-2190.
- M, T.; DG, F.; VL, B.; I, M.; JE, C.; P, P.; R, G.; A, P.; S, C.; PM, C., et al. Age-related shift in LTD is dependent on neuronal adenosine A receptors interplay with mGluR5 and NMDA receptors. Mol Psychiatr 2020, 25, 1876-1900. [CrossRef]
- Silvia, V.D.S.; Haberl, M.G.; Zhang, P.; Bethge, P.; Lemos, C.; Gon Alves, N.; Gorlewicz, A.; Malezieux, M.; Gon Alves, F.Q.; Grosjean, N.L. Early synaptic deficits in the APP/PS1 mouse model of Alzheimer’s disease involve neuronal adenosine A2A receptors. Nat Commun 2016, 7, 11915. [CrossRef]
- Chen, J.F.; Pedata, F. Modulation of ischemic brain injury and neuroinflammation by adenosine A2A receptors. Curr Pharm Design 2008, 14, -. [CrossRef]
- S, H.; H, D.; H, Z.; S, W.; L, H.; S, C.; J, Z.; L, X. Noninvasive limb remote ischemic preconditioning contributes neuroprotective effects via activation of adenosine A1 receptor and redox status after transient focal cerebral ischemia in rats. Brain Res 2012, 1459, 81-90. [CrossRef]
Figure 1.
Morphological changes in rats exposed to different Cy doses.
Figure 2.
The weight changes of rats in each group (* indicates that compared with the control group, p<0.05** indicates that compared with the control group, p<0.001).
Figure 2.
The weight changes of rats in each group (* indicates that compared with the control group, p<0.05** indicates that compared with the control group, p<0.001).
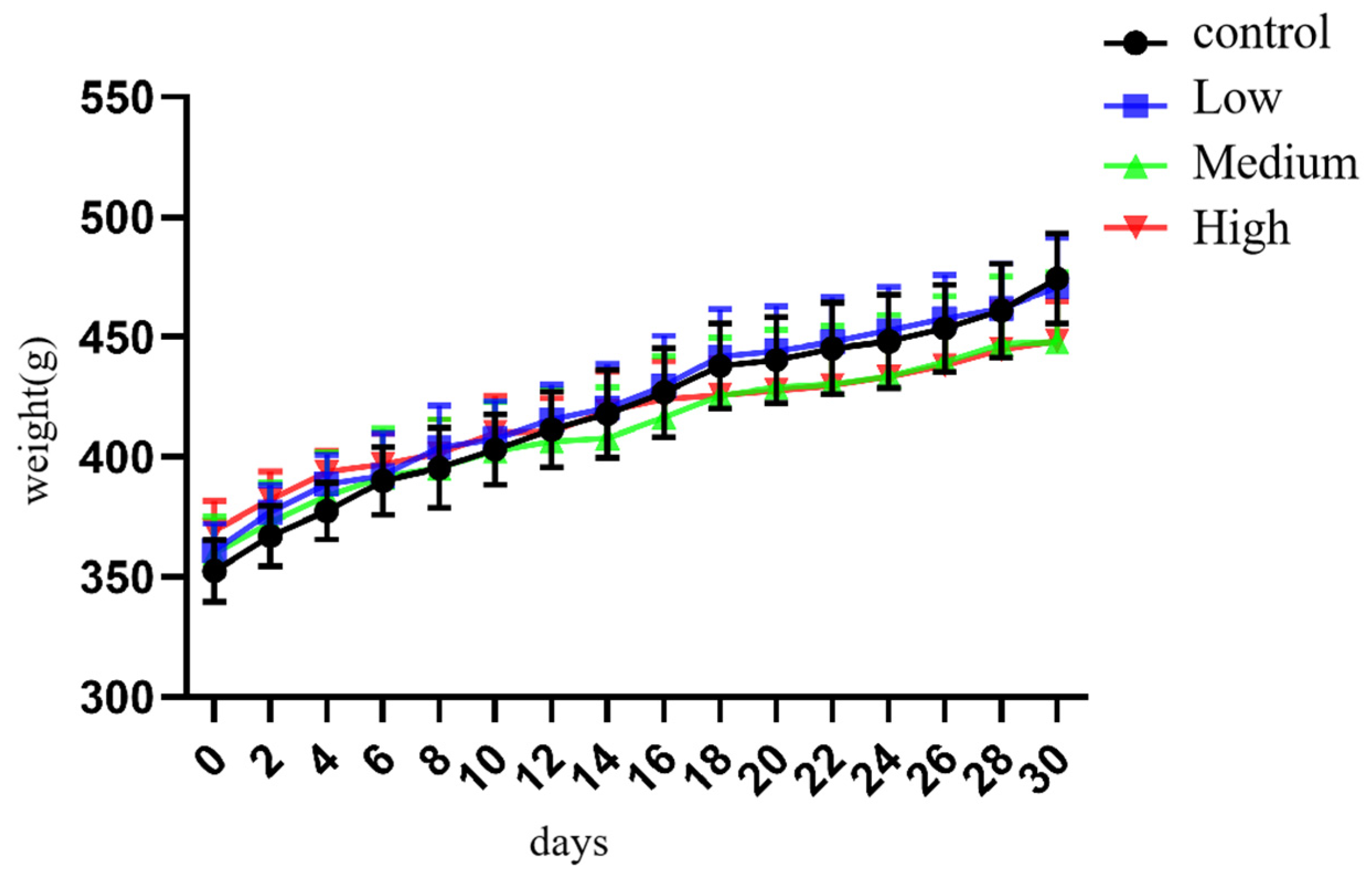
Figure 3.
Changes in the open field experiment of rats before and after Cy exposure. Note: Figure A shows the residence time in the central grid of rats in each group before and after exposure; Figure B shows the number of grid crossings before and after exposure; Figure C shows the time of grooming before and after exposure; Figure D shows the number of standing before and after exposure; * shows the comparison with the control group, p<0.05, * * shows the comparison with the control group, p<0.001.
Figure 3.
Changes in the open field experiment of rats before and after Cy exposure. Note: Figure A shows the residence time in the central grid of rats in each group before and after exposure; Figure B shows the number of grid crossings before and after exposure; Figure C shows the time of grooming before and after exposure; Figure D shows the number of standing before and after exposure; * shows the comparison with the control group, p<0.05, * * shows the comparison with the control group, p<0.001.
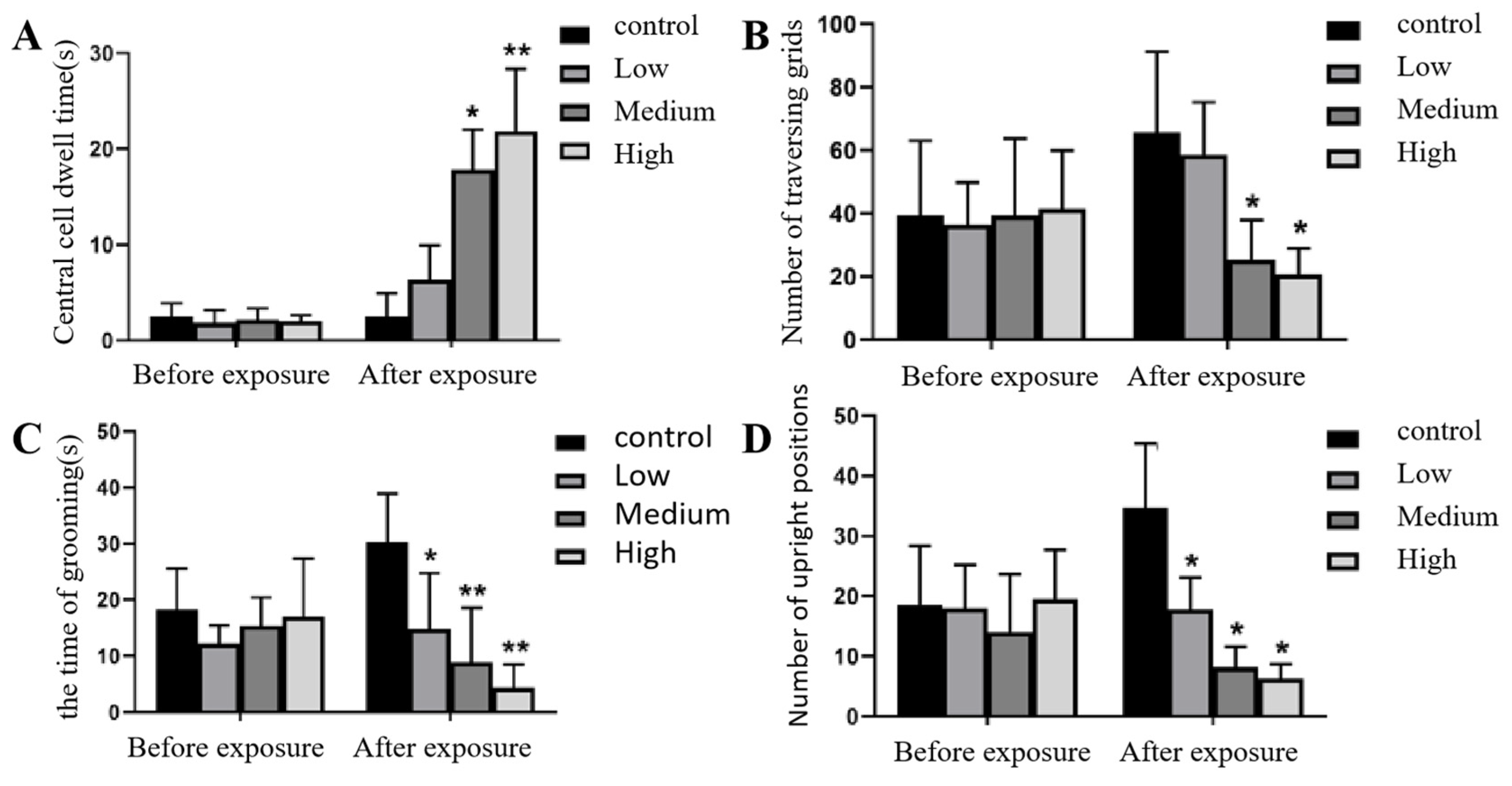
Figure 4.
Changes in the DI index of new object recognition in rats before and after Cy exposure Note: * indicates comparison with the control group, p<0.05, * * indicates comparison with the control group, p<0.001.
Figure 4.
Changes in the DI index of new object recognition in rats before and after Cy exposure Note: * indicates comparison with the control group, p<0.05, * * indicates comparison with the control group, p<0.001.
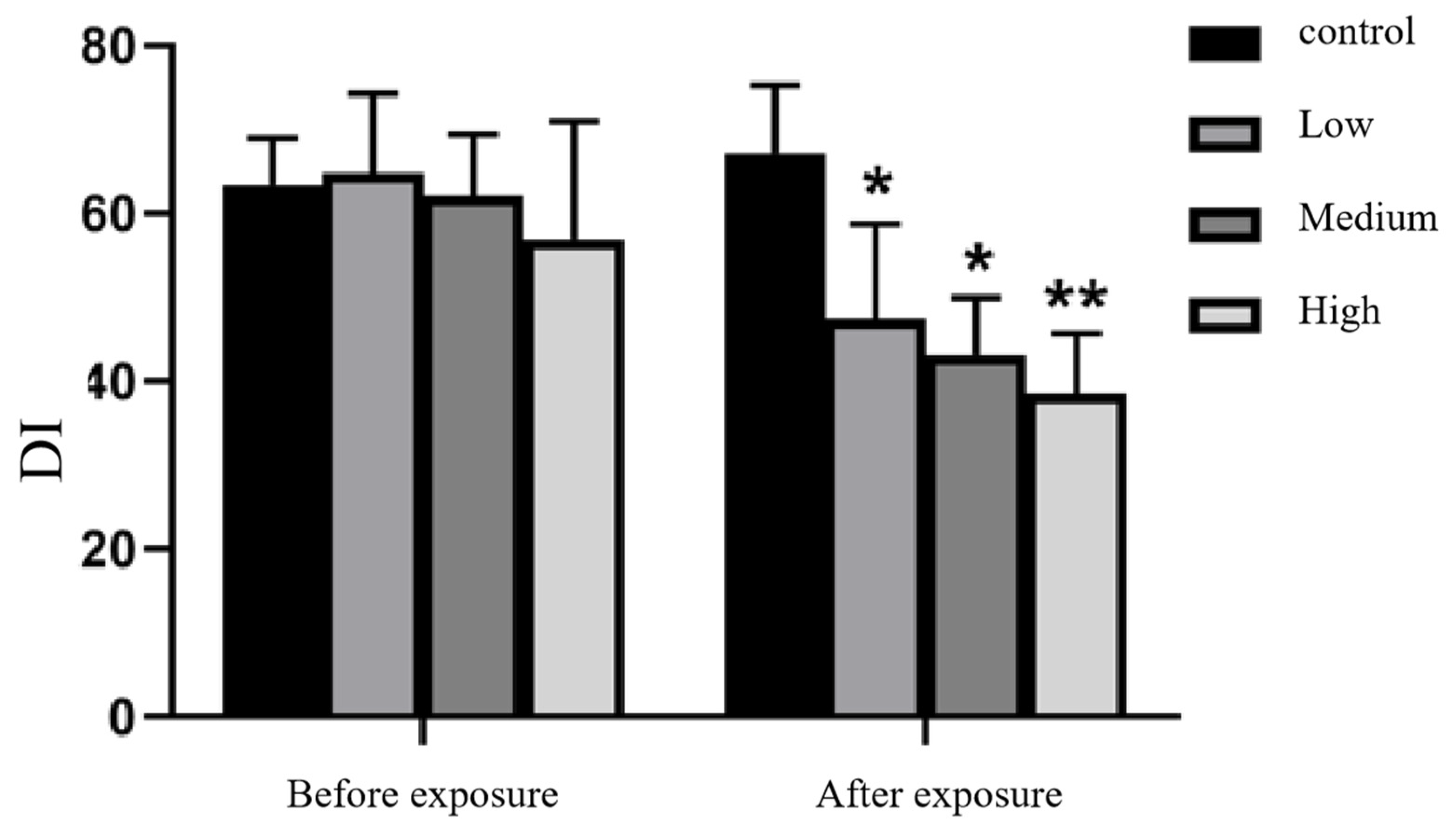
Figure 5.
Changes in the elevated cross maze metrics before and after Cy exposure. Note: Figure a shows the percentage of open arm retention time; Figure B shows the percentage of open arm entry times; * represents the comparison with the control group, p<0.05; * * represents the comparison with the control group, p<0.001.
Figure 5.
Changes in the elevated cross maze metrics before and after Cy exposure. Note: Figure a shows the percentage of open arm retention time; Figure B shows the percentage of open arm entry times; * represents the comparison with the control group, p<0.05; * * represents the comparison with the control group, p<0.001.
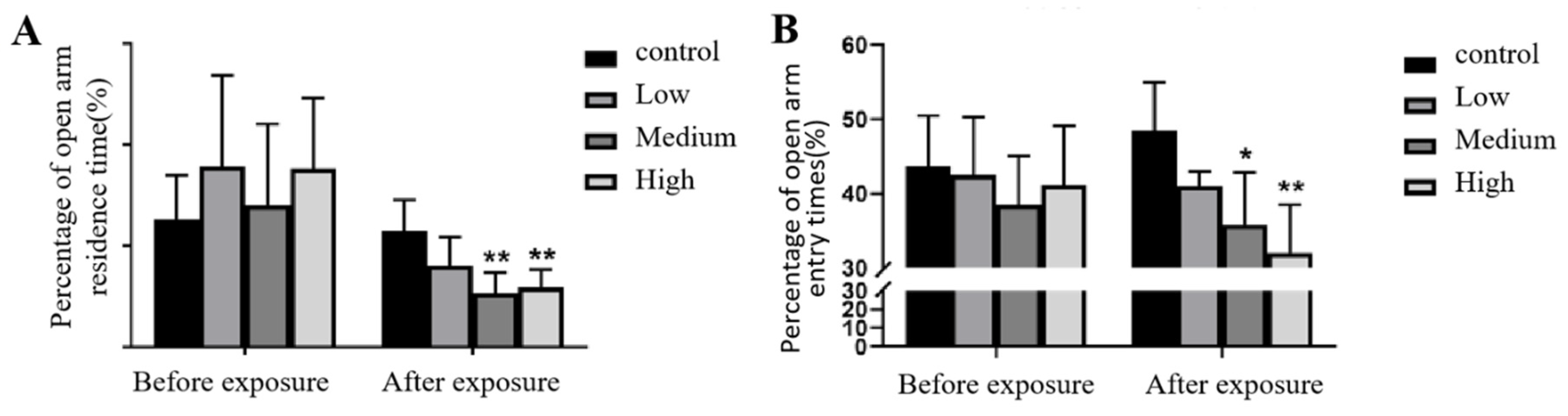
Figure 6.
Comparison of spatial exploration and escape latency.
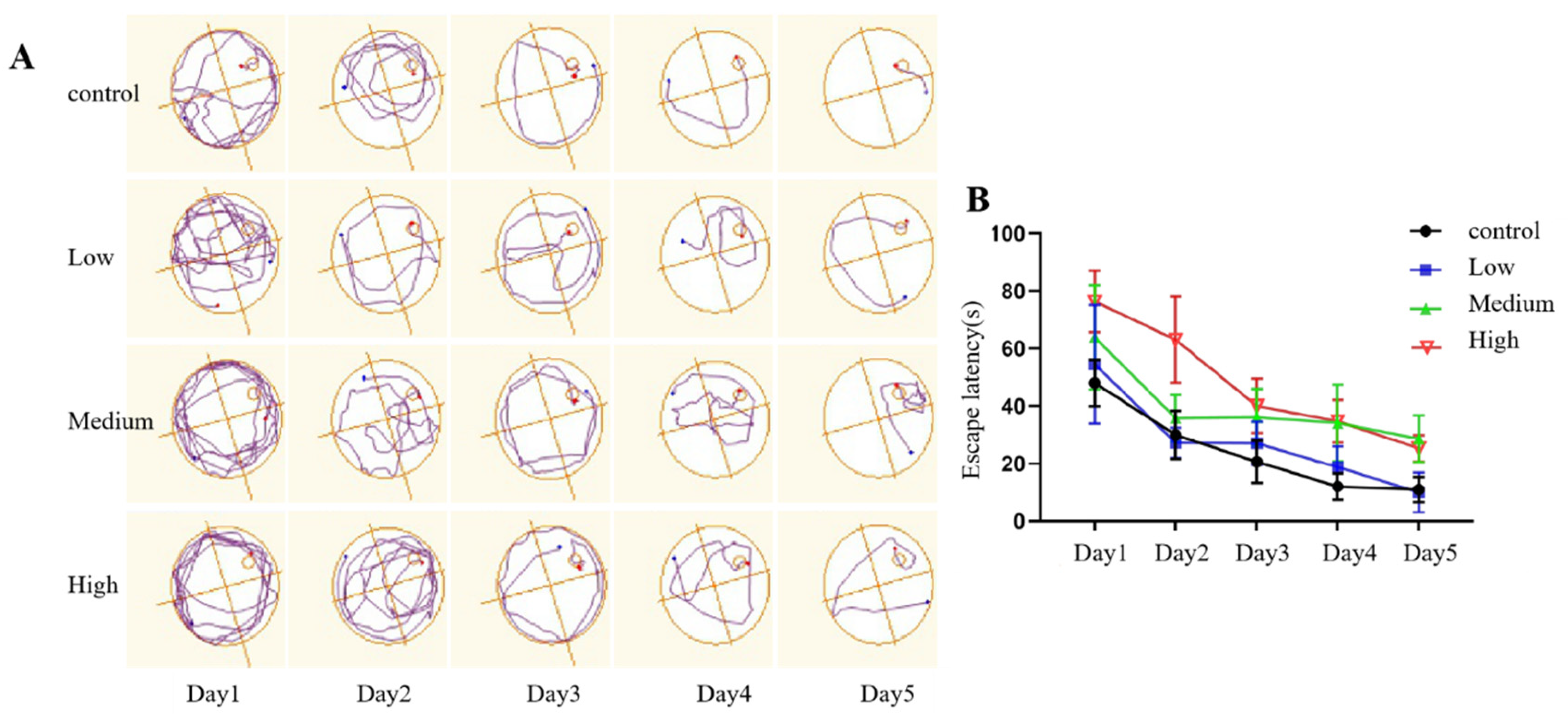
Figure 7.
Comparison of the spatial exploration ability of rats exposed to Cy in the Morris water maze test. Note: Figure A shows the residence time of the target quadrant of rats; Figure B shows the crossing times of the target platform of rats; Figure C shows the trajectory of the spatial exploration action of rats; * represents the comparison with the control group, p<0.05; * * represents the comparison with the control group, p<0.001.
Figure 7.
Comparison of the spatial exploration ability of rats exposed to Cy in the Morris water maze test. Note: Figure A shows the residence time of the target quadrant of rats; Figure B shows the crossing times of the target platform of rats; Figure C shows the trajectory of the spatial exploration action of rats; * represents the comparison with the control group, p<0.05; * * represents the comparison with the control group, p<0.001.
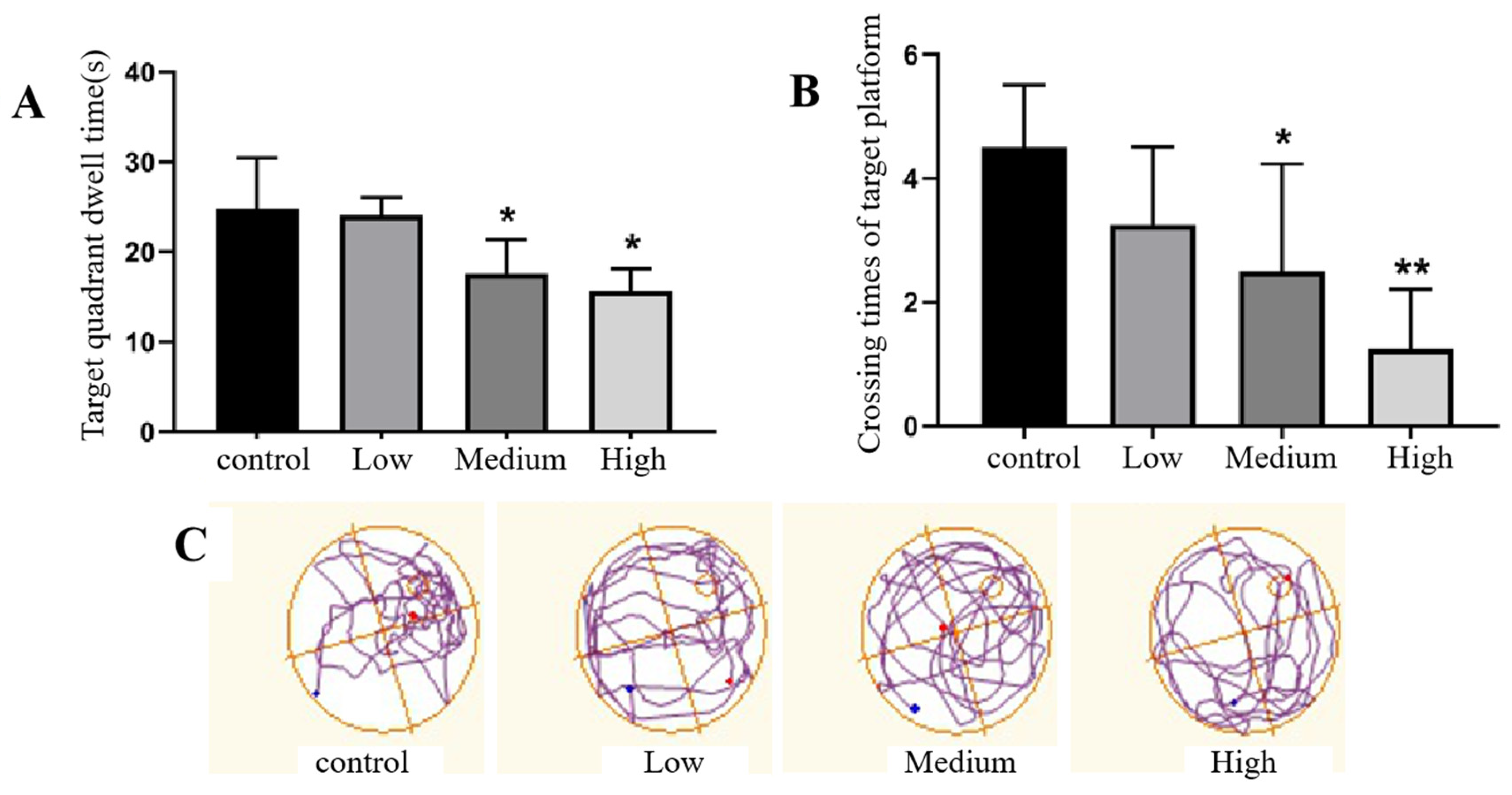
Figure 8.
Injury of hippocampal neurons in rats exposed to Cy. A: Comparison of HE-stained tissues (bar=20 μm, 400 ×); B: Statistics of the number of vertebral cells; C: Comparison of Nissl bodies in Nissl-stained neurons (bar=20 μm, 400 ×); D: Statistics of the number of Nissl corpuscles; E: Morphological changes in hippocampal cells under an electron microscope. Bar=5.0 μm, 1200×, Bar=2.0 μm, 8000×.
Figure 8.
Injury of hippocampal neurons in rats exposed to Cy. A: Comparison of HE-stained tissues (bar=20 μm, 400 ×); B: Statistics of the number of vertebral cells; C: Comparison of Nissl bodies in Nissl-stained neurons (bar=20 μm, 400 ×); D: Statistics of the number of Nissl corpuscles; E: Morphological changes in hippocampal cells under an electron microscope. Bar=5.0 μm, 1200×, Bar=2.0 μm, 8000×.
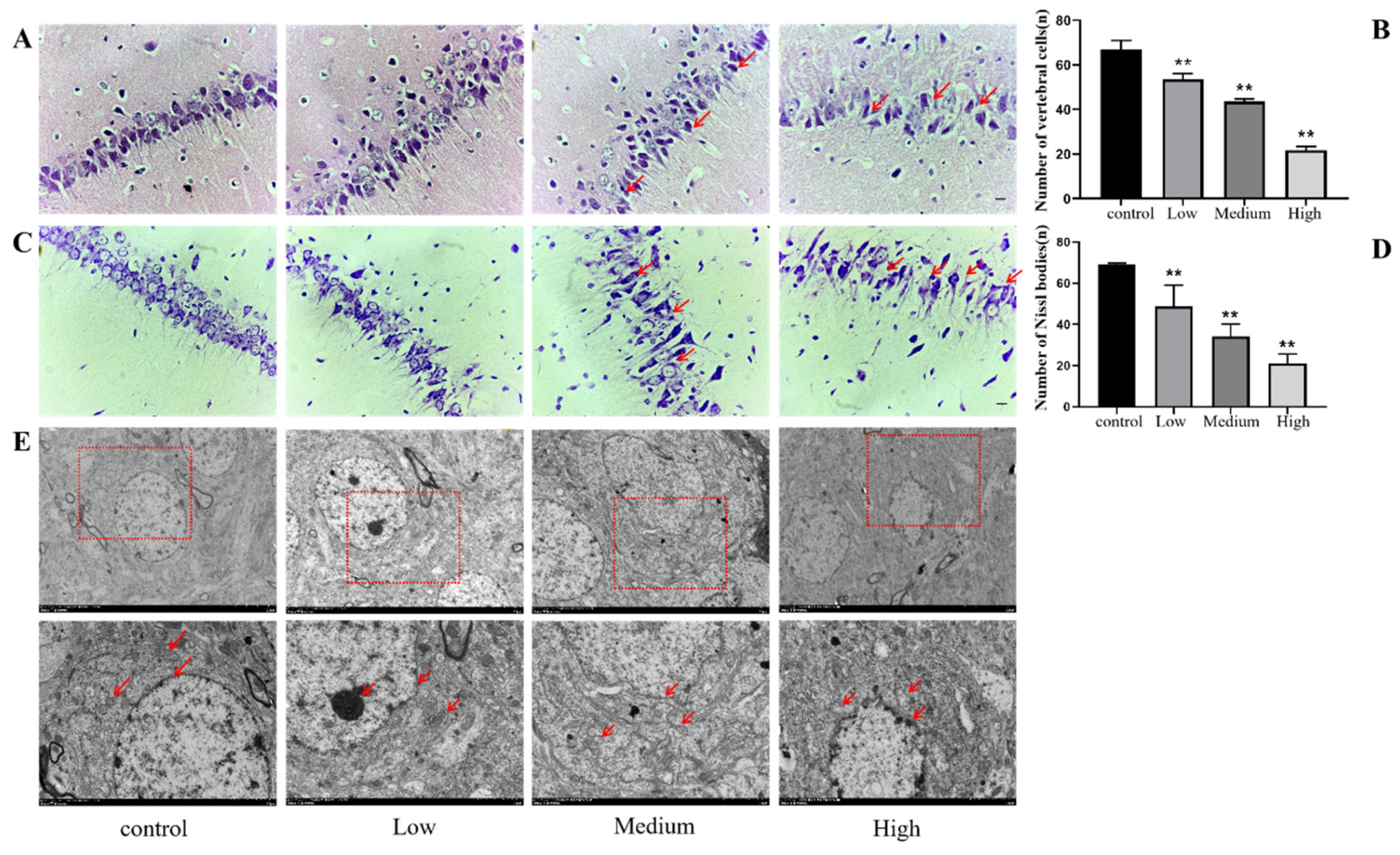
Figure 9.
Comparison of synaptic Ji and PSD-95 and SYP protein and mRNA expression in hippocampal neurons of Cy-exposed rats. A: Electron microscopy results. Bar=1.0 μm, 12000×; B: Western blot results; C, D: Statistical results of PSD-95 and SYP protein expression; E, F: Statistical results of mRNA expression of PSD-95 and SYP; G, H: Levels of glutamate and ATPase; *compared with the control group, p<0.05 **compared with the control group, p<0.001.
Figure 9.
Comparison of synaptic Ji and PSD-95 and SYP protein and mRNA expression in hippocampal neurons of Cy-exposed rats. A: Electron microscopy results. Bar=1.0 μm, 12000×; B: Western blot results; C, D: Statistical results of PSD-95 and SYP protein expression; E, F: Statistical results of mRNA expression of PSD-95 and SYP; G, H: Levels of glutamate and ATPase; *compared with the control group, p<0.05 **compared with the control group, p<0.001.
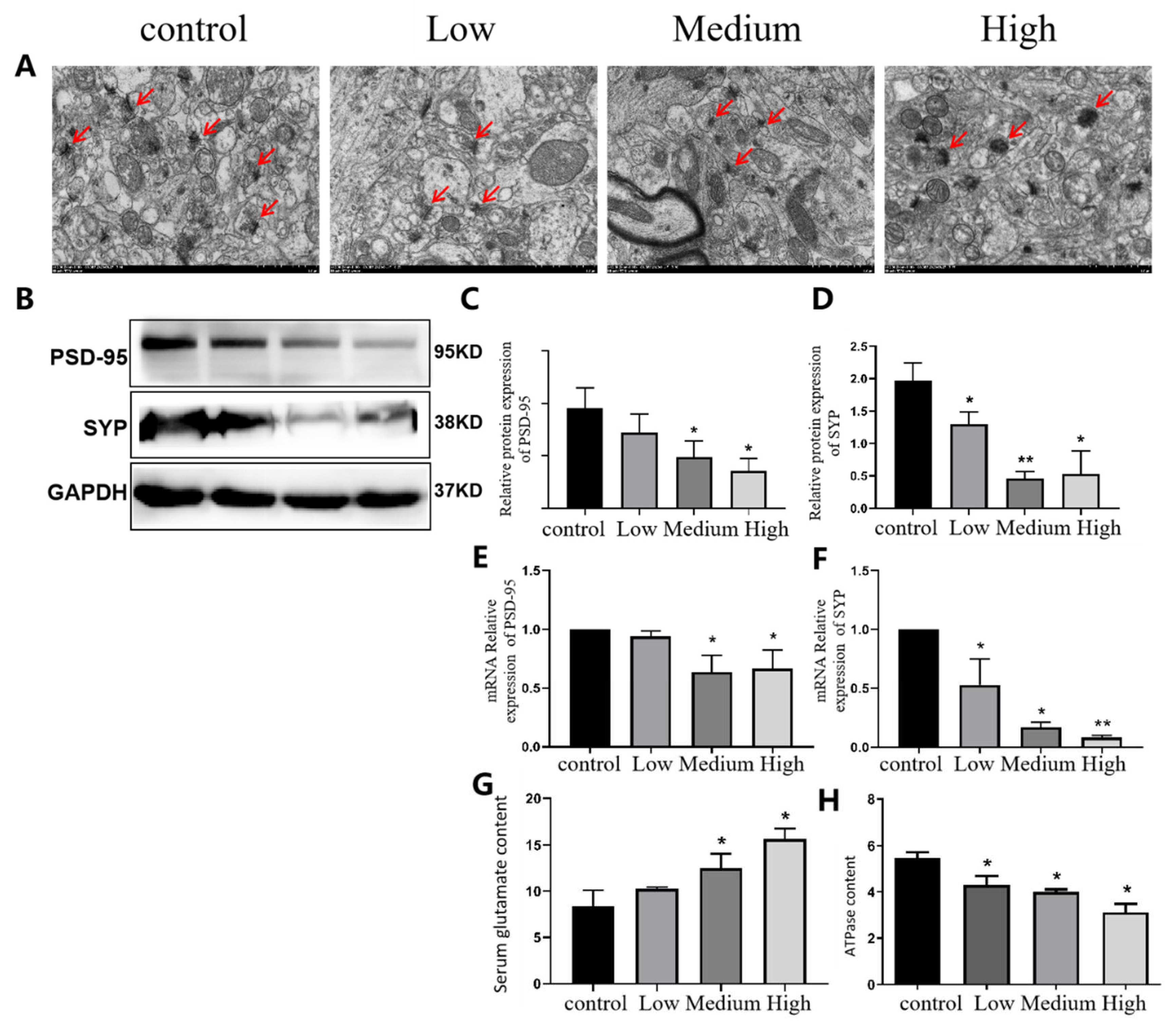
Figure 10.
Comparison of expression levels and immunohistochemistry results of IL-6, THF-α and A2AR proteins and mRNA levels in Cy-exposed rats. (A: Protein blotting results. Bar=20 μm, 400×; B: Statistical results of IL-6, THF-α, A2AR protein and mRNA expression; C, D: Immunohistochemistry results *compared to the control group, p<0.05 **compared to the control group, p<0.001).
Figure 10.
Comparison of expression levels and immunohistochemistry results of IL-6, THF-α and A2AR proteins and mRNA levels in Cy-exposed rats. (A: Protein blotting results. Bar=20 μm, 400×; B: Statistical results of IL-6, THF-α, A2AR protein and mRNA expression; C, D: Immunohistochemistry results *compared to the control group, p<0.05 **compared to the control group, p<0.001).
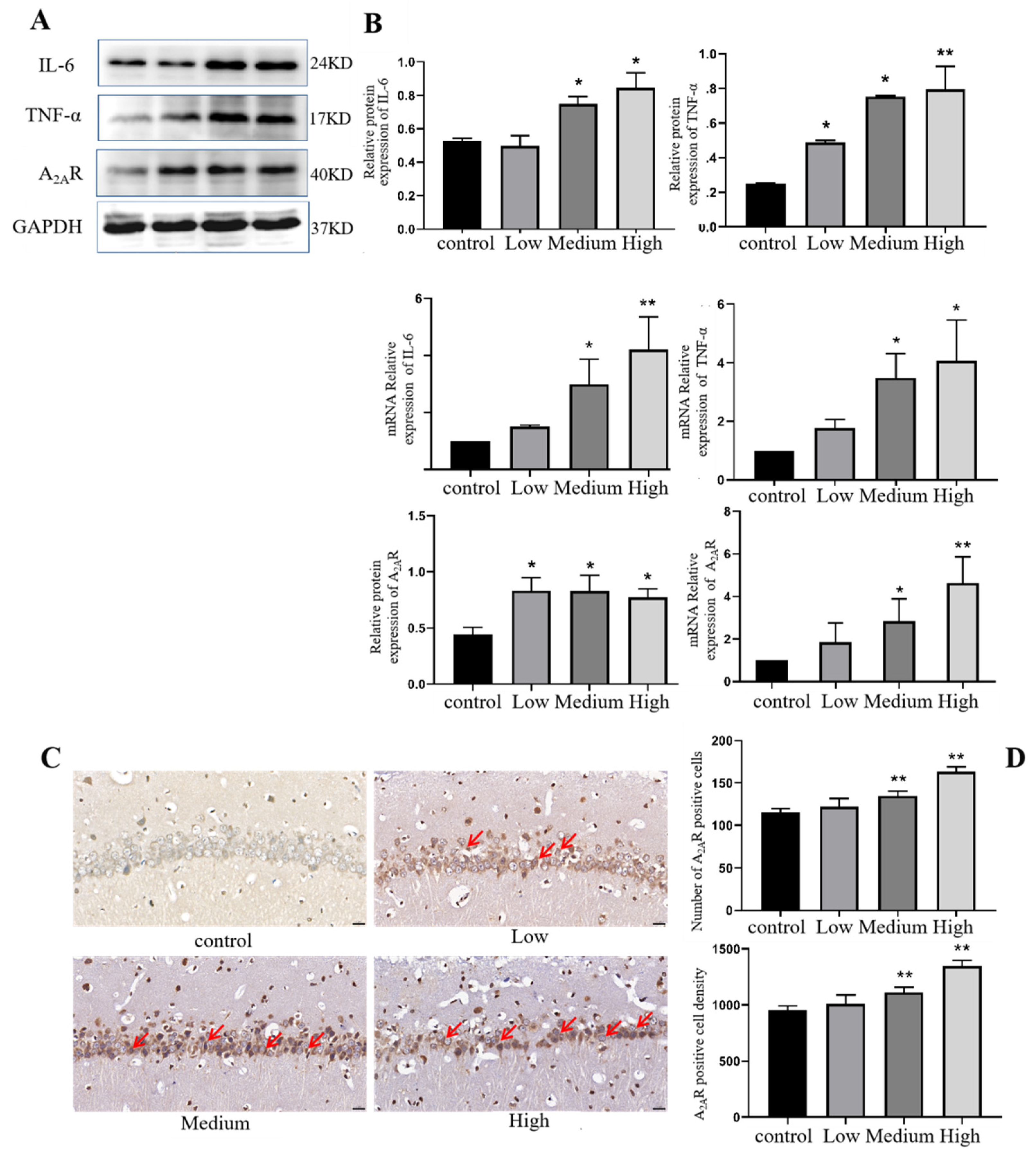
Table 1.
Primary Antibodies.
| Antibody name | Company | Country |
|---|---|---|
| Rabbit Anti-A2AR (ab3461) | Abcam | U.S.A |
| Rabbit Anti-PSD-95 (ab76115) | Abcam | U.S.A |
| Rabbit Anti-SYP (ab32127) | Abcam | U.S.A |
| Rabbit Anti-IL-6 | Affinity | China |
| Rabbit Anti-TNF-α | Affinity | China |
| Rabbit Anti-GAPDH | Bioss | China |
Table 2.
Primer sequences.
| Forward primer | Reverse primer | Species | |
|---|---|---|---|
| A2AR | GAAAGACGGGAACTCCACGAAGAC | GGCAGTAACACGAACGCAAAGAAG | rat |
| PSD-95 | TCCAGTCTGTGCGAGAGGTAGC | GGACGGATGAAGATGGCGATGG | rat |
| SYP | GCTGTGTTTGCCTTCCTCTACTC | TGATAATGTTCTCTGGGTCCGTG | rat |
| IL-6 | ACTTCCAGCCAGTTGCCTTCTTG | TGGTCTGTTGTGGGTGGTATCCTC | rat |
| TNF-α | AAAGGACACCATGAGCACGGAAAG | CGCCACGAGCAGGAATGAGAAG | rat |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



