
Submitted:
02 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
Increasing CO2 concentration ([CO2]) in atmosphere decreases mineral nutrients concentration in crops whereas increasing water use efficiency (WUE) especially in drought conditions. Partial root-zone irrigation (PRI) could stimulate soil nutrient mineralization and improve crop nutrient status. Yet the effect of PRI combined with elevated CO2 concentration (e[CO2]) on the element stoichiometry of tomato leaves remains unknown. This study was to investigate the responses of leaf mineral nutrients status and element stoichiometric ratio in tomato grown at PRI exposed to e[CO2]. Tomato plants (cv. Ailsa Craig) were grown in pots in climate-controlled growth chambers with ambient [CO2] (a[CO2], 400ppm) and elevated [CO2] (e[CO2] 800ppm), respectively. Three irrigation regimes (IR) i.e. full irrigation (FI), deficit irrigation (DI) and partial root-zone irrigation (PRI) were applied to tomato plants at flowering stage. The result showed that plants exposed to both reduced irrigations had a similar biomass, an enhanced root growth including greater root to shoot ratio (R/S), root length (RL), surface area (RS), volume (RV) and specific length (RSL), and an improved WUE under e[CO2]. Compared to a[CO2], e[CO2] growth environment resulted in a similar leaf [C], [K] and [Mg], leaf C/K and C/Mg, increased leaf [Ca], [S] and leaf C/N, C/P and N/P, but decreased leaf [N], [P], leaf C/Ca, C/S and N/K at PRI. Additionally, under e[CO2], PRI showed an increase in leaf [C], [N] and [S], no decrease in leaf [K], Ca], [Mg], [S] and [15N], but a decrease in leaf C/N and C/S as compared with FI. Conclusively, PRI had the ability to alleviate the negative effects on mineral nutrient concentrations and maintain or improve most leaf element stoichiometric ratios under e[CO2]. Therefore PRI would be a practicable irrigation for optimizing WUE and nutrient status in tomato leaves in a future freshwater-limited and higher CO2 environment.
Keywords:
Elevated CO2
; partial root-zone irrigation
; root morphological traits
; mineral nutrient
; elemental stoichiometric ratio
1. Introduction
The atmospheric [CO2] has been continuing to rise since past decades and is predicted to reach nearly 800 ppm at the end of this century, even if further actions are taken to reduce emissions[1]. This has aroused widely concern on the effect of rising atmospheric [CO2] on crop yield and quality. Positive responses of e[CO2] has been well observed in most crop plants and the results can be summarized as increasing net photosynthetic rate, increasing biomass and yield, decreasing stomatal conductance, enhancing WUE and C:N ratio [2,3,4,5,6]. Nonetheless, e[CO2] decreases transpiration, and hence soil nutrient mass flow which can reduce delivery of mobile mineral nutrients to the root and thereby limit plant mineral nutrient acquisition [7], which has been associated with significant decrease in the concentrations of important macro- and micro-nutrients elements in C3 crops [8,9], and this effect has been explained as growth dilution phenomenon [10]. Earlier studies have also shown that e[CO2] could reduce mineral concentration in plants at deferent level [9,11] and decrease N more than P and K [11,12], which can disturb the stoichiometry (the comparative ratio of chemical elements) in plants [13]. The nutrient imbalance in plants has serious impact on plant structure and metabolism [14]. Additionally, mineral nutrient content in plants can be influenced by the capacity of roots to absorb mineral nutrients from soil solution and plant root systems have morphological plasticity in response to e[CO2] [15]. Roots of plants grown under e[CO2] grow faster with more numerous, thicker and longer roots [16]. An increase in root length under e[CO2] may change the spatial patterns of exploitation of soil water and nutrients from different soil layers [15,17]. Hence the change of root morphological traits would affect water and mineral nutrient uptake in tomatoes grown under e[CO2] condition.
Alternate partial root-zone drying irrigation (PRI) and deficit irrigation (DI) are water-saving irrigation strategies [18]. And the DI method irrigates the entire root zone with less water than the potential evapotranspiration, while the mild water stress developed has minimal effects on plant growth [19]. Alternate PRI is a further improvement of DI [20]. Alternate PRI involves irrigating only part of the root zone and permitting the other part to a predetermined level before the next irrigation [21]. Due to spatial and temporal differences in soil water status under PRI, the plants have more root biomass and longer RL, lager root length density, and R/S but decrease leaf area [22,23,24,25], implying that PRI plants had greater capacity for deeper and more widespread water and nutrient exploration, less water loss than DI and FI plants [20,23,26]. It was reported that PRI could increase N concentration in tomato upper leaves, leading to a greater photosynthesis and WUE compared with DI [19]. Furthermore, soil drying and rewetting cycles can stimulate organic matter decomposition and mineralization of soil nutrients, hence resulting in enhancing nutrients availability and plant uptake [27,28,29]. Moreover, accumulated evidence have demonstrated that PRI can increase N [19,30] and P [31,32] accumulation in plants, enhance the concentration of anions and cations in xylem sap [29]. Therefore, it is suggested that PRI has the potential to improve nutrient status in tomatoes grown under e[CO2].
Recent studies have revealed that PRI can improve fruit quality and mineral nutrient status of tomatoes grown under e[CO2] [6,33,34]. However, the combined effects of e[CO2] and PRI on tomato root morphological traits and mineral stoichiometry in leaf have not been well studied. Accordingly, the objective of this study was to investigate the responses of leaf mineral nutrient status and leaf element stoichiometry of tomatoes grown at PRI exposed to e[CO2]. In this study, tomatoes (cv. Ailsa Craig) grown in two growth chambers with a[CO2] and e[CO2], respectively, were investigated and 15N isotope technique was used to trace N accumulation in tomato leaves grown under e[CO2] during the irrigation treatments period. The effects of irrigation regimes on plant growth, root morphological traits, leaf element concentrations and uptakes, leaf element stoichiometric ratios of tomatoes gown under e[CO2] were determined. we hypothesized that:1) e[CO2] would ameliorate the negative effects of PRI on tomato plant growth; 2) PRI would have positive effect on root morphological traits for mineral nutrient absorption, which would relieve the negative effect on leaf mineral nutrient concentrations, thereby improving element stoichiometric ratios of tomato leaves grown exposed to e[CO2].
2. Materials and Methods
The experiment was carried out from September to November 2020 in the climate-controlled green-house of Northwest A&F University ( N 34◦ 15′, E 108◦ 04′), Yangling, Shaanxi, China. The tomato seeds (cv. Ailsa Craig) were sown on 13th Sep. 2020 under either a[CO2] or e[CO2] respectively, at day/night air temperature of 25/16 ± 2 ℃, relative humidity of 55 ± 5 %, and under long-day (16 h of light provided by high pressure sodium lamps and metal halide lamps) conditions. The daily average temperature (Temp), relative humidity (RH) and vapour pressure deficit (VPD) are illustrated in Figure 1
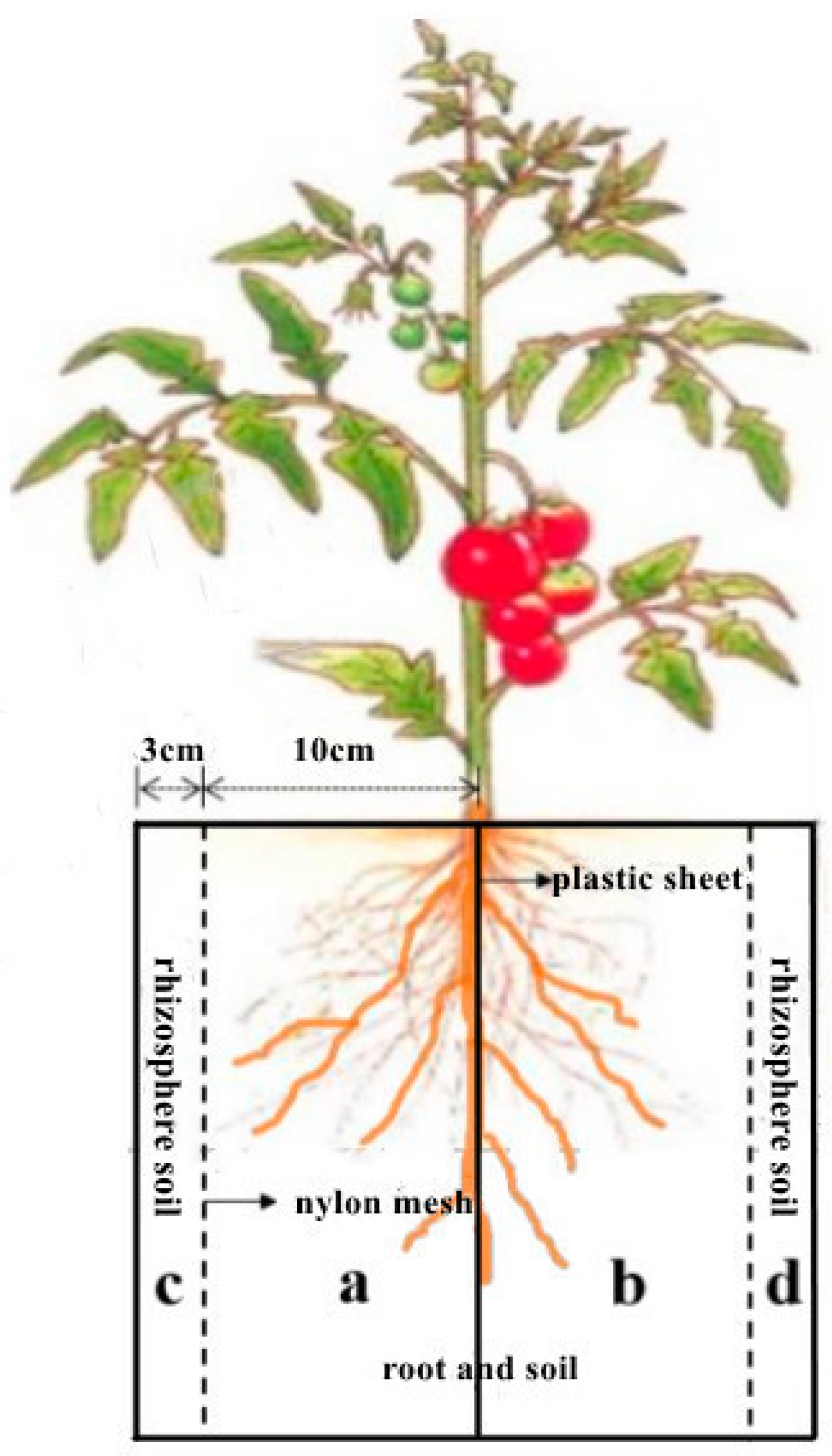
Four weeks after sowing, the seedlings were transplanted into pots with a volume of 16 liter (16 cm inner width, 26 cm length inner, and 40 cm height) at 4-leaf stage with one plant per pot. The pots were divided into two equally sized vertical compartments by plastic sheets to prevent water and nutrient exchange between the two compartments. A small piece of plastic (6×8 cm) was removed from the top-middle of the sheet where the tomato seedlings were transplanted. Nylon meshes with aperture of 150 μm were inserted 3cm away from both inner edge of the compartments along the length direction, and therefore the pot was divided into four sections (a, b, c, d) (Figure 2). The tomato roots were grown in the soil volume of a and b. The soils in compartments of c and d were considered as rhizosphere soil. Each pot was filled with 18.5 kg pot-1 naturally dried soil at a bulk density of 1.30 g cm-3. The soil was taken from topsoil of 25 cm at a conventional agricultural field in Yangling. The soils were air-dried and sieved through a 0.5 cm mesh. The soil had a pH of 7.8, EC of 0.36 ms/cm, total C of 17.79 g/kg, total N of 0.46 g/kg, total P of 0.60 g/kg, total K of 24.24g/kg. The pot water holding capacity (WHC, %) was 30%. Two weeks after transplanting, 2.0 g N (as urea), 2.0 g K (as KH2PO4 + K2SO4) and 1.6 g P (as KH2PO4) were applied to each pot with the irrigation water to meet the nutrient requirement for plant growth. To prevent soil evaporation, the soil surface was covered by a 2-3 cm layer pearlite. Each pot was watered daily to 90% WHC from 18th Oct. to 4th Nov. 2020.
2.2. Treatments
The tomato seedlings were grown either in growth chambers with a[CO2] or e[CO2], and in each of the growth chambers the pots were randomly arranged, comprising 3 treatments with 4 replicates each, namely 12 pots, additional 4 pots in each growth chamber for acquiring the basic growth indices of the plants before starting irrigation treatments. Each growth chamber was equipped with a CO2 monitor that controls the CO2 concentrations over the experimental period by connecting with an automated CO2 adjustable valve, installed on a special high-pressure gas cylinder.
All plants were well-watered for 18 days from one week after transplanting. To stimulate the root growth and activity, the soil water content was maintained at 60% of WHC during the first week after transplanting. Three days before the onset of the irrigation treatments, 0.10 g 15N urea (5% 15N abundance) was dissolved in 1 ml water and added into the c and d soil compartments with a syringe, respectively. The irrigation treatments included (I) full irrigation (FI) where the whole pot was watered daily to 90% of WHC; (II) deficit irrigation (DI) in which 70% volume of water used in FI was irrigated evenly to the entire pot; (III) partial root-zone drying irrigation (PRI) where the soil compartments of a+c were watered using the same volume of water for DI while the b+d were allowed to dry to a soil water content at 12% -14% , then the irrigation was switched. The irrigation treatments lasted 20 days, during which tomatoes had experienced each soil compartment two drying/wetting cycles. The average soil water content was determined by TDR (time-domain reflectometer, MINITRASE, Soil Moistures Equipment Crop., SEC, US) with probes (35 cm) inserted in both a and b soil compartments. The plants were irrigated manually with tap water daily between 16:30 and 18:00.
2.3. Measurements
2.3.1. Harvest
The plants were harvest two times during the experimental period. The first harvest was done at the onset the irrigation treatment, during which four pots of each tomato genotype from each growth chambers were harvested. The last harvest was conducted when tomato plants reached blossom stage, then all of the 24 pots were harvested. During each harvest, the plant samples were divided into three parts: leaf, stem, and root. The entire-root was collected by carefully washing with tap water after collecting the rhizosphere soil following the procedure by Bornø et al.[35], and then stored at 4℃ for further analysis. Leaves and stems were oven dried separately at 75℃ to constant weight.
2.3.2. Root morphological analyses
The stored root samples were thawed and washed with deionized water before measuring root traits. The procedure is described in detail by Liu et al. [36]. Roots were carefully washed on a 2 mm sieve and broken segments were also collected during flushing. Then a representative root subsample from each clean root samples was used for scanning on a 20 × 25 cm transparent trays with deionized water using WinRHIZO Pro (Modified Epson Expression 12000XL, Regent Instruments Inc., Québec, QC, Canada) at 400 dots per inch resolution. The obtained images were analyzed with WinRHIZO software (Regent Instruments Inc., Quebec, Canada) for root length (RL), root average diameter (RAD), root surface area (RS), root volume (RV). Afterwards, root samples were oven dried to constant weight at 75℃ to determine dry matter. Specific root length (SRL) was calculated as the root length per unit of root dry weight and root tissue density (RTD) as the ratio of root dry weight to root volume [36].
2.3.3. Plant biomass, water use, water use efficiency
The dry matter (DM) of each plant part was measured after oven drying to a constant weight. Total dry matter accumulation (ΔDM) during the irrigation treatment period was calculated as the difference of DM between the first harvest and the last harvest. Biomass partitioning were calculated as the ratio between DM of leaves, stems and roots to plant total DM, respectively. Plant water use (WU) was calculated based on the amount of irrigation and changes of soil water content in the pots from the initiation to the end of the irrigation treatments. Plant water use efficiency (WUE) was calculated as the ratio of ΔDM to WU. The oven-dried leaf sample was thoroughly grounded into powder and passed through a 0.2 mm sieve for further analysis.
2.3.4. Elemental analyses
The values of δ15N as well as the total C and N content in the leaves were measured using elemental analyzer (vario PYRO cube, Elementar Analysensysteme GmbH, Germany) coupled to an isotope-ratio mass spectrometer (Isoprime100, Elementar Analysensysteme GmbH, Germany).
Leaf P, K , S concentrations were determined, after digestion with 10 ml concentrated nitric acid + 2ml perchloric acid, using an Automatic Flow Elemental Analyzer ( Seal AA3, Germany). Leaf Ca and Mg concentrations were measured by Atomic Absorption Spectroscopy (PE-pinAAcle, 900 F), after digestion with 5ml concentrated nitric acid + 1ml perchloric acid . The obtained values of the leaf were used to calculate stoichiometric ratios of C/N, C/P, C/K, C/Ca, C/Mg, C/ S, N/P, N/K. The elements uptake contant were calculated as leaf elemental concentration multiplies by leaf DM.
2.4. Statistical analysis
The data was analyzed by two-way analysis of variance (ANOVA) using SPSS 20.0 (IBM Corporation, New York, USA) to evaluate the effect of [CO2] and irrigation regimes as well as their interactions on plants growth, WUE, root morphological traits, leaf element concentrations, leaf element uptakes, and leaf element stoichiometry. One-way ANOVA and Duncan’s multiple range test at the 5% confidence level was further conducted when there were significant interactions between the independent factors to test for significant differences among treatments. Principle component analysis (PCA) and Pearson correlation coefficient were performed in origin 2021 (OriginLab, Massachusetts, USA)
3. Results
3.1. Plant dry matter accumulation (ΔDM), Dry biomass allocation, shoot dry mass, Plant water use (WU)and plant water use efficiency (WUE
The ΔDM, SDW, LDW, biomass allocation to leaf (LMR), stem (SMR) and root (RMR), WU and WUE of tomato plants and the output of two-way ANOVA was shown in (Table 1). [CO2] significantly affected these parameters. e[CO2] had significantly 101.5%, 99.2%, 101.5%, 3.8%, 14.1%, 38.5%, 45.4%, respectively, more ΔDM, SDW, LDW, LMR, RMR, WU and WUE while had 13.3% lower SMR of tomato plants than a[CO2]. IR significantly influenced LMR, SMR, RMR, LDW, WU and WUE. Compared to FI, reduced irrigations (DI and PRI) increased SMR (+13.7% at DI, +16.2% at PRI ), RMR (+22.7% at DI, +18.5% at PRI) and WUE (+35.5% at DI, +38.7% at PRI), but reduced LMR (-5.5% at DI, 5.2% at PRI), LDW (-7.5% at DI, -9.0% at PRI) and WU (ca. 30% at both DI and PRI). C×IR had little effect on ΔDM, SDW, LDW, biomass allocation to leaf (LMR), stem (SMR) and root (RMR), WU and WUE of tomato plants.

Table 1.
The main effects of treatments and output of two-way ANOVA for dry biomass accumulation, dry biomass allocation, shoot dry mass, water use, water-use efficiency.
Table 1.
The main effects of treatments and output of two-way ANOVA for dry biomass accumulation, dry biomass allocation, shoot dry mass, water use, water-use efficiency.
| [CO2] | Irrigation regimes | ∆DM (g plant-1) | biomass allocation (%) | SDW (g plant-1) | LDW (g plant-1) | WU (L plant-1) | WUE (g L-1) | |||
| leaf | stem | root | ||||||||
| C400 | FI | 25.9 ± 1.6 | 73.0 ± 1.5 | 21.6 ± 1.4 | 5.4 ± 0.2 | 26.6 ± 1.5 | 20.5 ± 1.0 | 7.0 ± 0.3 | 3.7 ± 0.3 | |
| DI | 25.7 ± 0.5 | 68.5 ± 1.2 | 24.9 ± 1.2 | 6.7 ± 0.2 | 26.2 ± 1.8 | 19.2 ± 1.2 | 4.9 ± 0 | 5.2 ± 0.1 | ||
| PRI | 25.8 ± 2.0 | 67.6 ± 1.2 | 25.6 ± 1.2 | 6.8 ± 0.1 | 26.1 ± 0.5 | 18.9 ± 0.3 | 4.9 ± 0 | 5.3 ± 0.4 | ||
| C800 | FI | 53.8 ± 2.1 | 74.3 ± 1.3 | 19.2 ± 1.4 | 6.5 ± 0.2 | 52.6 ± 2.1 | 41.8 ± 1.9 | 9.7 ± 0.7 | 5.6 ± 0.3 | |
| DI | 50.3 ± 1.6 | 70.7 ± 1.1 | 21.5 ± 1.0 | 7.9 ± 0.4 | 50.2 ± 2.4 | 38.4 ± 1.3 | 6.8 ± 0 | 7.4 ± 0.2 | ||
| PRI | 52.1 ± 2.7 | 72.0 ± 0.7 | 21.8 ± 0.3 | 7.3 ± 0.4 | 48.8 ± 1.3 | 37.8 ± 0.8 | 6.8 ± 0 | 7.6 ± 0.4 | ||
| Output of two-way ANOVA | ||||||||||
| [CO2] (C) | *** | * | ** | ** | *** | *** | *** | *** | ||
| Irrigation regimes (IR) | ns | ** | * | *** | ns | ** | *** | *** | ||
| C×IR | ns | ns | ns | ns | ns | ns | ns | ns | ||
The table shows the significance results of two-way analysis of variance (ANOVA) of dry mass accumulation (ΔDM), the allocation of ΔDM to leaf (LMR), stem (SMR) and root (RMR), shoot dry weight (SDW), leaf dry weight (LDW), water use (WU) and water use efficiency (WUE) of tomato plants as affected by the CO2 concentration (400ppm and 800ppm) and irrigation regimes (IR) (FI, full irrigation; DI, deficit irrigation; and PRI, alternative partial root drying irrigation) and their interactions. Values are means ± standard error of the means (SE) (n = 4).Different letters after the means indicate significant differences among treatments determined by Duncan’ s multiple range test at p < 0.05. *, **, *** indicates significance level at p < 0.05, p < 0.01, and p < 0.001 respectively, ‘ns’ is non-significant.
3.2. Root growth and morphological traits
The RL, RS, RD, RV, SRL, RTD, R/S and RDW of tomatoes grown at different irrigation regimes under a[CO2] and e[CO2], respectively, and the output of the two-way ANOVA were shown in (Table 2). [CO2] significantly affected RL, RS, RV, R/S and RDW. e[CO2] significantly increased RL, RS, RV, R/S and RDW by 154.7%, 144.9%, 134.5%, 20.0% and 139.2%, respectively. IR significantly influenced RL, RS, RV, SRL,RTD and R/S. reduced irrigations had longer RL, greater RS and RV, larger SRL, RTD and R/S than FI. C×IR significantly affected RSL but had no influence on other indices. Compared to a[CO2], e[CO2] enhanced SRL by 121.2% and 19.4%, respectively, at FI and PRI whereas decrease SRL by 26.7% at DI.
Table 2.
The effects of treatments and output of two-way ANOVA for root traits of tomato plants.
| [CO2] | Irrigation regimes | RL (m) | RS (cm2) | RD (mm) | RV (cm3) | SRL (m g-1) | RTD (g cm-3) | R/S | RDW (g plant-1) | |
| C400 | FI | 12.9 ± 1.1 | 280.5 ± 28.6 | 0.54 ± 0.02 | 4.5 ± 0.6 | 9.9 ± 0.8c | 0.3 ± 0.02 | 0.06±0.003 | 1.5 ± 0.1 | |
| DI | 60.8 ± 4.6 | 718.0 ± 55.8 | 0.38 ± 0.01 | 7.1 ± 0.5 | 35.9 ± 2.7a | 0.2 ± 0.02 | 0.07±0.003 | 1.9 ± 0.2 | ||
| PRI | 43.1 ± 6.4 | 609.8 ± 62.7 | 0.45 ± 0.03 | 7.4 ± 0.6 | 25.8 ± 2.9ab | 0.2 ± 0.01 | 0.07±0.003 | 1.9 ± 0.0 | ||
| C800 | FI | 75.0 ± 19.7 | 1045.5 ± 132.7 | 0.47 ± 0.04 | 12.4 ± 0.6 | 21.9 ± 5.9b | 0.3 ± 0.01 | 0.07±0.003 | 3.6 ± 0.1 | |
| DI | 99.5 ± 24.5 | 1324.4 ± 240.7 | 0.47 ± 0.05 | 15.2 ± 1.6 | 26.3 ± 4.5ab | 0.2 ± 0.01 | 0.08±0.005 | 4.3 ± 0.4 | ||
| PRI | 123.0 ± 14.3 | 1569.4 ± 123.7 | 0.43 ± 0.03 | 17.0 ± 1.1 | 30.8 ± 4.6ab | 0.2 ± 0.01 | 0.09±0.005 | 3.8 ± 0.3 | ||
| Output of two-way ANOVA | ||||||||||
| [CO2] (C) | *** | *** | ns | *** | ns | ns | ** | *** | ||
| Irrigation regimes (IR) | * | ** | ns | ** | ** | ** | ** | ns | ||
| C×IR | ns | ns | ns | ns | * | ns | ns | ns | ||
The table reports the significance results of two-way analysis of variance (ANOVA) of root lengh (RL), root surfarea (RS), root average diameter (RD), root volume( RV), specific root length (SRL), root tissue density (RTD), root dry weight (RDW) and root to shoot ratio (R/S) of tomato plants as affected by the CO2 concentration (400ppm and 800ppm) and irrigation regimes (FI, full irrigation; DI, deficit irrigation; and PRI, alternative partial root drying irrigation) and their interactions. Values are means ± standard error of the means (SE) (n = 4). Different letters after the means indicate significant differences among treatments determined by Duncan’s multiple range test at p < 0.05. *, **, *** indicates significance level at p < 0.05, p < 0.01, and p < 0.001 respectively, ‘ns’ is non-significant.
3.3. Leaf element concentrations.
The leaf [C], [N], [P], [K], [Ca], [Mg], [S] and [15N] of tomatoes grown at different irrigation regimes under a[CO2] and e[CO2], respectively, and the output of the two-way ANOVA were presented in Table 3. [CO2] significantly affected leaf [N], [Ca], [Mg] and [S]. e[CO2] significantly lowered leaf [N] and [Mg], respectively, by 13.1% and 6.9% but increased leaf [Ca] and [S]leaf, respectively, by 8.2% and 20.2%. IR significantly affected leaf [C]leaf, [P]leaf, [Ca]leaf, [Mg]leaf and [15N]. Reduced irrigations significantly decreased leaf [P] (-10.7% at DI, -19.0% at PRI), [Ca] (- 2.5% at DI, -8.3% at PRI) and [Mg] (- 2.6% at DI, -10.3% at PRI), while increased leaf [C] (+ 1.2% at DI, +1.7% at PRI) and [15N] (+46.2% at DI, +11.5% at PRI). C×IR had little effect on leaf [C], [N], [P], [K], [Ca], [Mg], [S] and [15N] of tomatoes. Nevertheless, compared to a[CO2], e[CO2] decreased leaf [N] and [P], had equivalent leaf [C], [K], [Mg] and [15N], but increased leaf [Ca] and [S] (Table 3). Moreover, under e[CO2], PRI increased leaf [C], [N] and [S] and had no decrease in leaf [K], [Ca], [Mg] and [15N] in relative to FI (Table 3).
Table 3.
The effects of treatments and output of two-way ANOVA for leaf element concentrations.
| [CO2] | Irrigation regimes | [C] (mg g-1) | [N] (mg g-1) | [P] (mg g-1) | [K] (mg g-1) | [Ca] (mg g-1) | [Mg] (mg g-1) | [S] (mg g-1) | [15N] (mg g-1) | |
| C400 | FI | 412.2 ± 4.1 | 50.9 ± 0.9 | 4.2 ± 0.09 | 25.0 ± 1.9 | 51.2 ± 1.1 | 4.1 ± 0.2 | 7.2 ± 0.5 | 1.5 ± 0.1 | |
| DI | 421.4 ± 2.9 | 54.8 ± 1.1 | 3.8 ± 0.1 | 24.6 ± 2.0 | 49.5 ± 1.5 | 3.9 ± 0.2 | 5.7 ± 0.3 | 2.2 ± 0.3 | ||
| PRI | 420.4 ± 1.1 | 54.0 ± 1.8 | 3.8 ± 0.05 | 22.5 ± 1.4 | 47.1 ± 1.7 | 3.6 ± 0.1 | 5.9 ± 0.4 | 1.4 ± 0.2 | ||
| C800 | FI | 417.6 ± 1.4 | 45.4 ± 2.2 | 4.2 ± 0.16 | 25.0 ± 0.7 | 55.2 ± 1.7 | 3.7 ± 0.2 | 7.8 ± 0.1 | 1.1 ± 0.1 | |
| DI | 418.4 ± 3.1 | 43.7 ± 1.6 | 3.7 ± 0.27 | 24.5 ± 1.8 | 54.2 ± 1.9 | 3.7 ± 0.1 | 7.6 ± 0.4 | 1.6 ± 0.1 | ||
| PRI | 423.9 ± 1.7 | 49.6 ± 3.3 | 3.0 ± 0.39 | 22.7 ± 1.2 | 50.5 ± 1.6 | 3.4 ± 0.1 | 7.2 ± 0.7 | 1.5 ± 0.1 | ||
| Output of two-way ANOVA | ||||||||||
| [CO2] (C) | ns | *** | ns | ns | ** | * | ** | 0.052 | ||
| Irrigation regimes (IR) | * | ns | ** | ns | * | * | ns | ** | ||
| C×IR | ns | ns | ns | ns | ns | ns | ns | ns | ||
The table shows the significance results of two-way analysis of variance (ANOVA) of leaf C concentration ([C]), leaf N concenrationt ([N]), leaf Pconcentration ([P]), leaf K concentration ([K]), leaf Ca concentration ([Ca]), leaf Mg concentration ([Mg]), leaf S concentration ([S]) and leaf [15N] of tomato plants as affected by the CO2 concentration (400ppm and 800ppm) and irrigation regimes (FI, full irrigation; DI, deficit irrigation; and PRI, alternative partial root drying irrigation) and their interactions. Values are means ± standard error of the means (SE) (n = 4). Different letters after the means indicate significant differences among treatments determined by Duncan’s multiple range test at p < 0.05. *, **, *** indicates significance level at p < 0.05, p < 0.01, and p < 0.001 respectively, ‘ns’ is non-significant.
3.4. Leaf element uptakes
The leaf C, N, P, K, Ca, Mg, S an 15N uptakes of tomatoes grown at different irrigation regimes under a[CO2] and e[CO2], respectively, and the output of the two-way ANOVA were depicted in Table 4. [CO2] significantly affected leaf C, N, P, K, Ca, Mg, S and 15N uptakes. e[CO2] significantly enhanced these indices. IR significantly influenced leaf P, K, Ca, Mg, S and 15N uptakes. Reduced irrigations significantly lowered leaf P, K, Ca, Mg, S uptakes but increased leaf 15N uptakes. C×IR significantly affected leaf P uptake. Compared to a[CO2], e[CO2] increased more leaf P uptake at FI than reduced irrigations.
Table 4.
The effects of treatments and output of two-way ANOVA for leaf element uptakes.
| [CO2] | Irrigation regimes | C (g plant-1) | N (g plant-1) | P (mg plant-1) | K (mg plant-1) | Ca (g plant-1) | Mg (mg plant-1) | S (mg plant-1) | 15N (mg plant-1) |
| C400 | FI | 8.5 ± 0.5 | 1.0 ± 0.05 | 86.4 ± 5.1d | 506.8 ± 17.2 | 1.1 ± 0.1 | 82.5 ± 3.1 | 147.7 ± 13.2 | 31.0 ± 2.2 |
| DI | 8 ± 0.1 | 1.0 ± 0.02 | 71.2 ± 1.5d | 463.6 ± 31.8 | 0.9 ± 0.0 | 74.3 ± 2.4 | 107.5 ± 6.6 | 41.0 ± 5.2 | |
| PRI | 8.1 ± 0.5 | 1.0 ± 0.06 | 73.2 ± 3.9d | 431.9 ± 41.5 | 0.9 ± 0.1 | 68.7 ± 5.5 | 112.4 ± 8.2 | 26.9 ± 4.7 | |
| C800 | FI | 17.5 ± 0.8 | 1.9 ± 0.05 | 175.8 ± 11.2a | 1041.3 ± 43.5 | 2.3 ± 0.1 | 155.2 ± 7.9 | 328.0 ±17.6 | 45.7 ± 5.8 |
| DI | 15.8 ± 0.4 | 1.7 ± 0.04 | 141.4 ± 13.3b | 929.5 ± 89.3 | 2.0 ± 0.0 | 138.7 ± 6.3 | 287.9 ± 21.9 | 61.7 ± 3.5 | |
| PRI | 16.3 ± 0.6 | 1.9 ± 0.11 | 113.3 ± 11.1c | 867.4 ± 18.5 | 1.9 ± 0.1 | 129.5 ± 5.8 | 273.9 ± 21.1 | 58.2 ± 4.2 | |
| Output of two-way ANOVA | |||||||||
| [CO2] (C) | *** | *** | *** | *** | *** | *** | *** | *** | |
| Irrigation regimes (IR) | ns | ns | ** | * | ** | ** | * | * | |
| C×IR | ns | ns | * | ns | ns | ns | ns | ns | |
The table reports the significance results of three-way analysis of variance (ANOVA) of leaf C accumulation, N, P, K, Ca, Mg and S uptakes of tomato plants as affected by the CO2 concentration (400ppm and 800ppm) and irrigation regimes (FI, full irrigation; DI, deficit irrigation; and PRI, alternative partial root drying irrigation) and their interactions. Values are means ± standard error of the means (SE) (n = 4). Different letters after the means indicate significant differences among treatments determined by Duncan’s multiple range test at p < 0.05. *, **, *** indicates significance level at p < 0.05, p < 0.01, and p < 0.001 respectively, ‘ns’ is non-significant.
3.5. C/N, C/P, C/K, C/Ca, C/Mg, C/S. N/P and N/K in tomato leaves
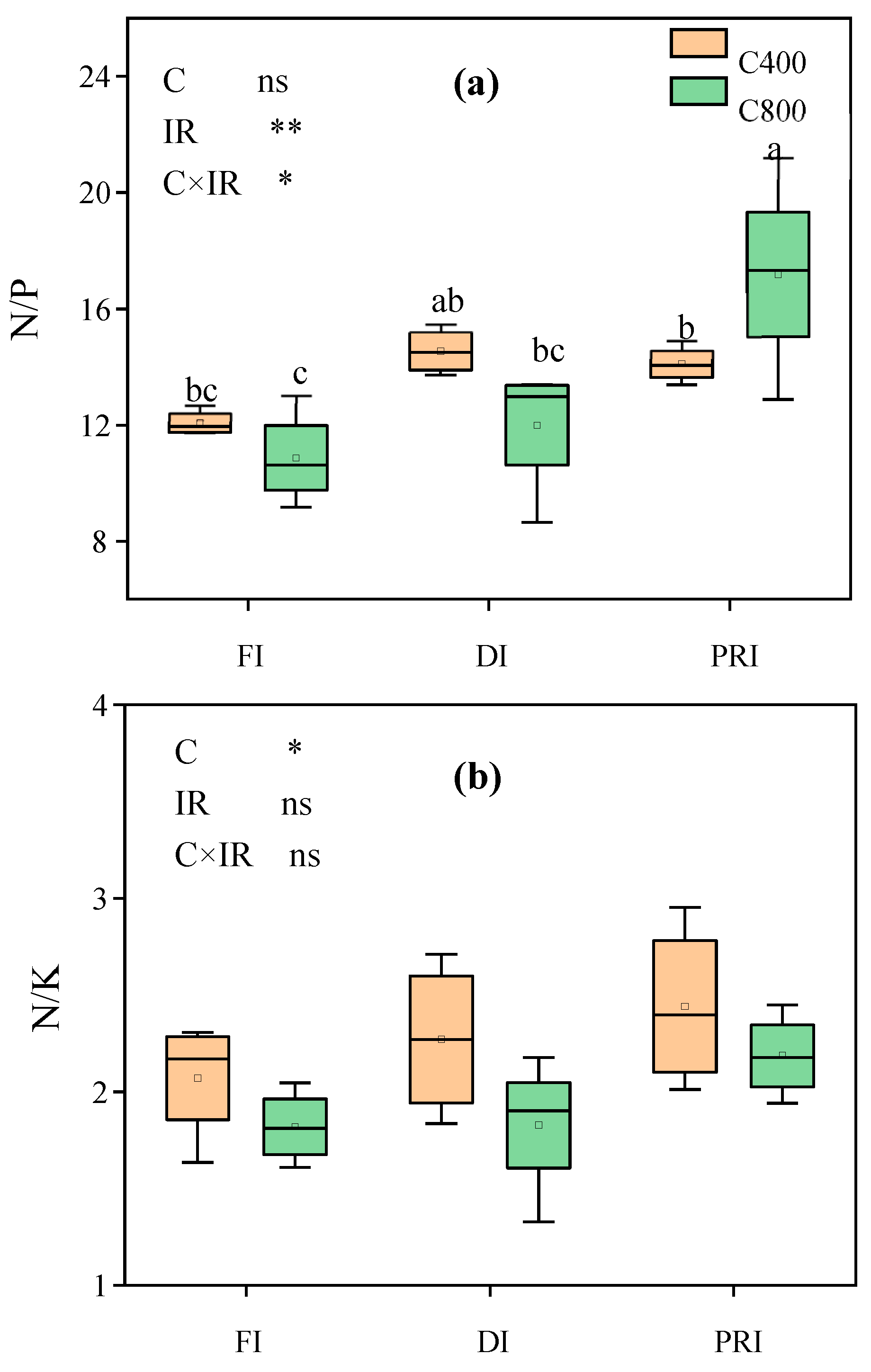
Leaf C/N, C/P, C/K, C/Ca, C/Mg, C/S, N/P, N/K and P/K of tomatoes grown at different irrigation regimes under a[CO2] and e[CO2], respectively, and the output of the two-way ANOVA were shown in Figure 3 and 4. [CO2] significantly affected leaf C/N, C/Ca, C/Mg, C/S and N/K. e[CO2] significantly increased leaf C/N and C/Mg, respectively by 16.6% and 7.6%, while decrease leaf C/Ca, C/S and N/K, respectively, by 7.0%, 17.2% and 14.0%. IR significantly affected leaf C/P, C/Ca, C/Mg, C/S and N/P. Reduced irrigations significantly increased leaf C/P, C/Ca, C/Mg, C/S and N/P. C×IR significantly affected leaf N/P. Compared to a[CO2], e[CO2] decreased leaf N/P at FI (-10.0%) and DI (-17.5%) but increased it by 21.8% at PRI. Nevertheless, compared to a[CO2], e[CO2] had similar leaf C/K and C/Mg, increased leaf C/N, C/P and N/P, but decreased leaf C/Ca, C/S and N/K at PRI. Additionally, under e[CO2], PRI showed an increase in leaf C/N and C/S as compared with FI.
Figure 3.
Leaf C:N ratio ([C:N]leaf, a), leaf C:P ratio ([C:P]leaf, b), leaf C:K ratio ([C:K]leaf, c), leaf C:Ca ratio ([C:Ca]leaf, d), leaf C:Mg ([C:Mg]leaf, e ), as well as leaf C:S ratio ([C:S]leaf, f) of tomato plants grown at three irrigation regimes ([IR]) (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRD) in either ambient (400 ppm, C400) or elevated (800 ppm, C800) atmospheric CO2 concentrations. Bars show the maximum (top edge) and minimum (lower edge) percentiles, whereas boxes represent the 25% and 75% percentiles across tomato plants (n=4). The median (50%) percentile is represented by the horizontal line within the box. Symbols indicate extreme values. *, ** and *** indicate significant levels at P < 0.05,P < 0.01, P < 0.001, respectively; “ns” denotes non-significant differences.
Figure 3.
Leaf C:N ratio ([C:N]leaf, a), leaf C:P ratio ([C:P]leaf, b), leaf C:K ratio ([C:K]leaf, c), leaf C:Ca ratio ([C:Ca]leaf, d), leaf C:Mg ([C:Mg]leaf, e ), as well as leaf C:S ratio ([C:S]leaf, f) of tomato plants grown at three irrigation regimes ([IR]) (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRD) in either ambient (400 ppm, C400) or elevated (800 ppm, C800) atmospheric CO2 concentrations. Bars show the maximum (top edge) and minimum (lower edge) percentiles, whereas boxes represent the 25% and 75% percentiles across tomato plants (n=4). The median (50%) percentile is represented by the horizontal line within the box. Symbols indicate extreme values. *, ** and *** indicate significant levels at P < 0.05,P < 0.01, P < 0.001, respectively; “ns” denotes non-significant differences.
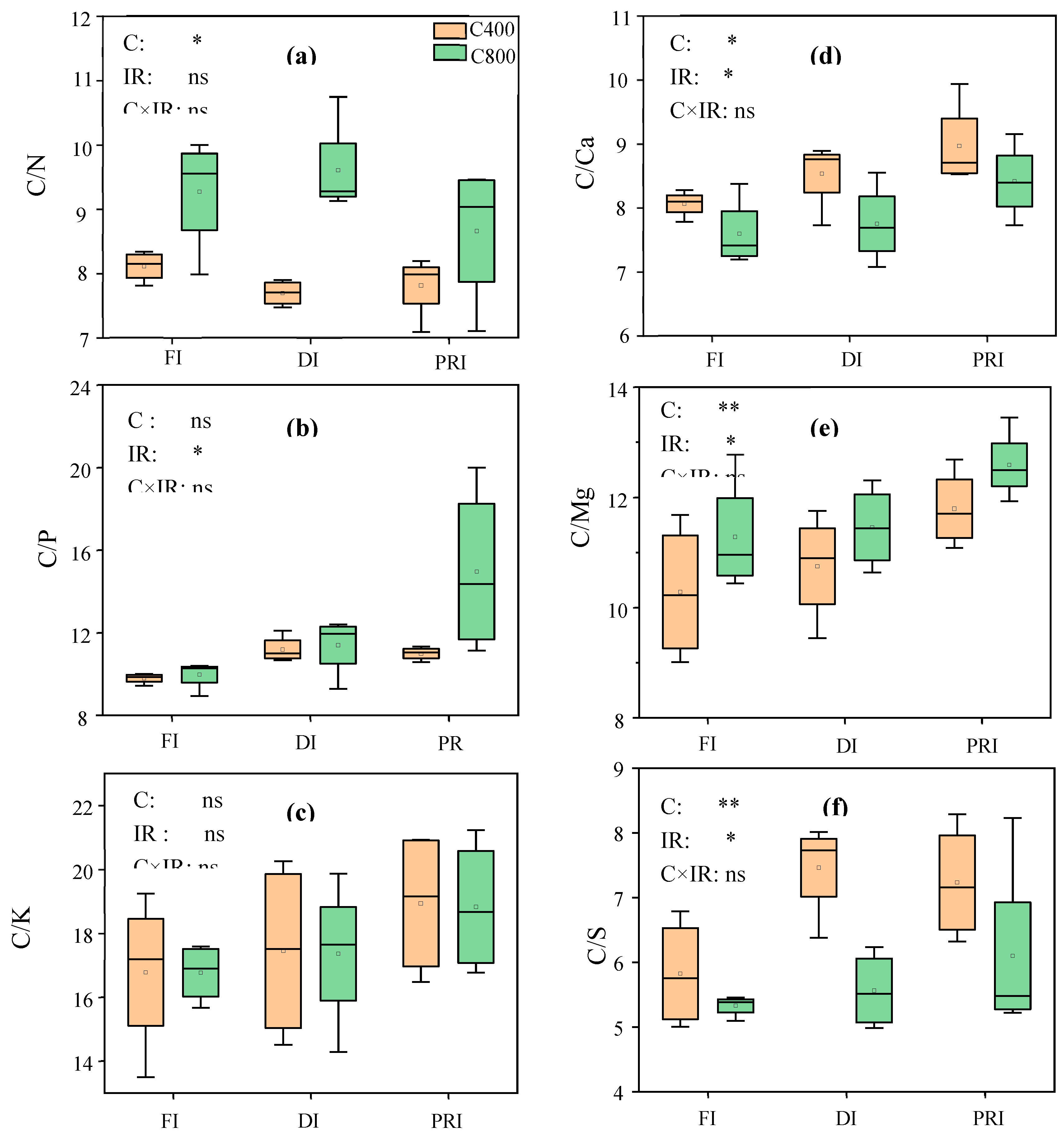
Figure 4.
Leaf N:P ratio ([N:P]leaf, a) and leaf N:K ratio ([N:K]leaf, b) of tomato plants grown at three irrigation regimes ([IR]) (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRD) in either ambient (400 ppm, C400) or elevated (800 ppm, C800) atmospheric CO2 concentrations. Bars show the maximum (top edge) and minimum (lower edge) percentiles, whereas boxes represent the 25% and 75% percentiles across tomato plants (n=4). The median (50%) percentile is represented by the horizontal line within the box. Symbols indicate extreme values. Different letters indicate significant difference determined by Duncan’s multiple range test at p < 0.05. *, ** and *** indicate significant levels at P < 0.05, P < 0.01, P < 0.001, respectively; “ns” denotes non-significant differences.
Figure 4.
Leaf N:P ratio ([N:P]leaf, a) and leaf N:K ratio ([N:K]leaf, b) of tomato plants grown at three irrigation regimes ([IR]) (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRD) in either ambient (400 ppm, C400) or elevated (800 ppm, C800) atmospheric CO2 concentrations. Bars show the maximum (top edge) and minimum (lower edge) percentiles, whereas boxes represent the 25% and 75% percentiles across tomato plants (n=4). The median (50%) percentile is represented by the horizontal line within the box. Symbols indicate extreme values. Different letters indicate significant difference determined by Duncan’s multiple range test at p < 0.05. *, ** and *** indicate significant levels at P < 0.05, P < 0.01, P < 0.001, respectively; “ns” denotes non-significant differences.
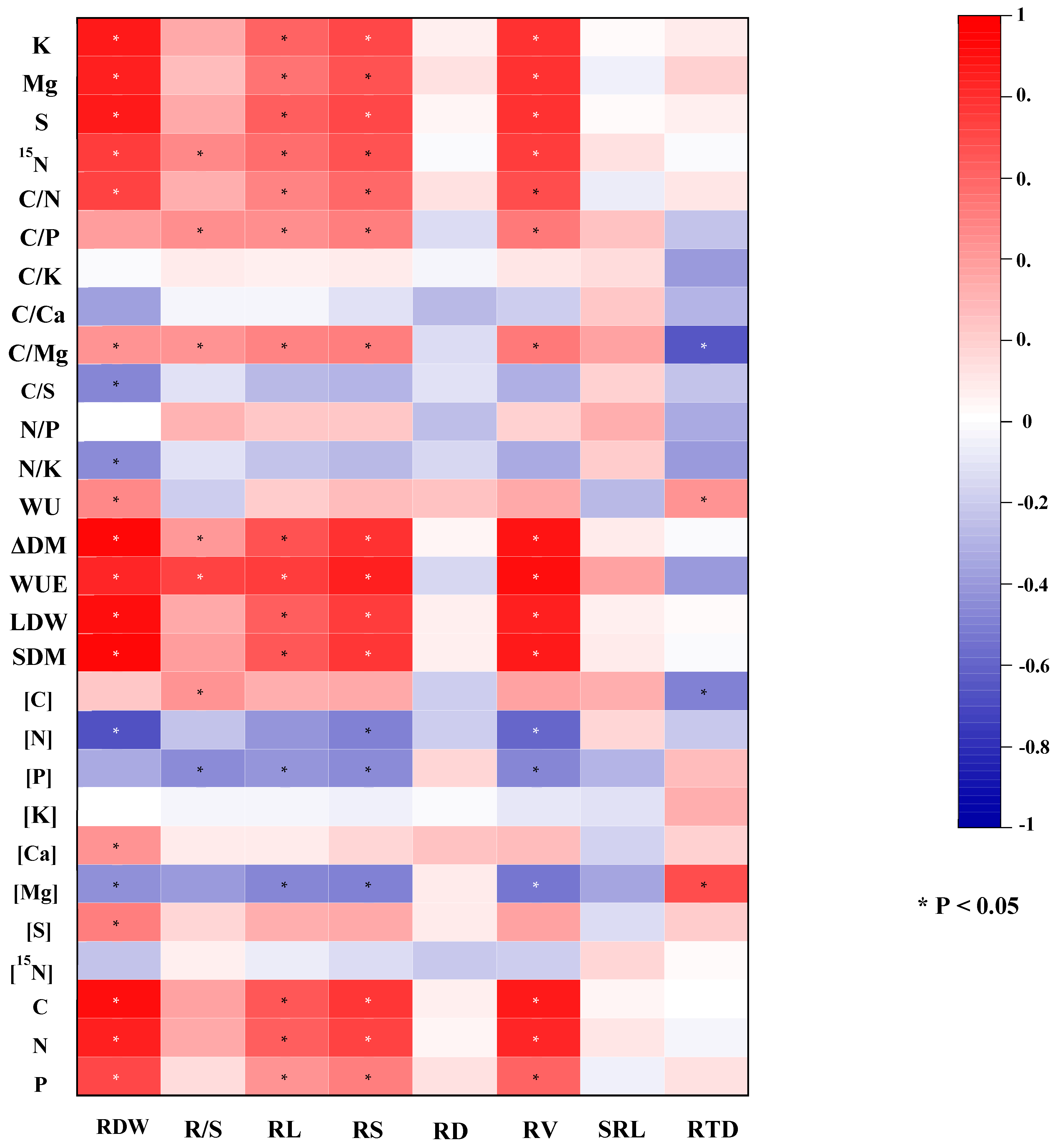
3.6. The Pearson correlation analysis between root morphology traits and tomato growth, element concentrations, element uptakes and stoichiometric ratios
The pearson correlation analysis indicated that ΔDM , LDM, WU, WUE, SDW, leaf element uptakes, leaf C/N, C/Mg correlated significantly positively, whereas leaf [Mg] correlated significantly negatively with RV, RS, RL, RDW (Figure 5). ΔDM, WUE, leaf 15N uptake, Leaf [C], leaf C/Mg and C/P correlated significantly positively, whereas leaf [P] correlated significantly negatively with R/S. RD was significant positive correlation with WU and leaf [Mg], but significant negative with leaf [C] and leaf C/Mg.
Figure 5.
Heat map of Pearson correlation coefficient between root morphological traits and plant dry mass and WUE, leaf element concentration, leaf element uptake and leaf nutrition stoichiometry of tomato plants. The color gradient is proportional to the Pearson's correlation coefficient. Red and blue colors denote positive and negative relationships, respectively. * indicates significant at P < 0.05.
Figure 5.
Heat map of Pearson correlation coefficient between root morphological traits and plant dry mass and WUE, leaf element concentration, leaf element uptake and leaf nutrition stoichiometry of tomato plants. The color gradient is proportional to the Pearson's correlation coefficient. Red and blue colors denote positive and negative relationships, respectively. * indicates significant at P < 0.05.
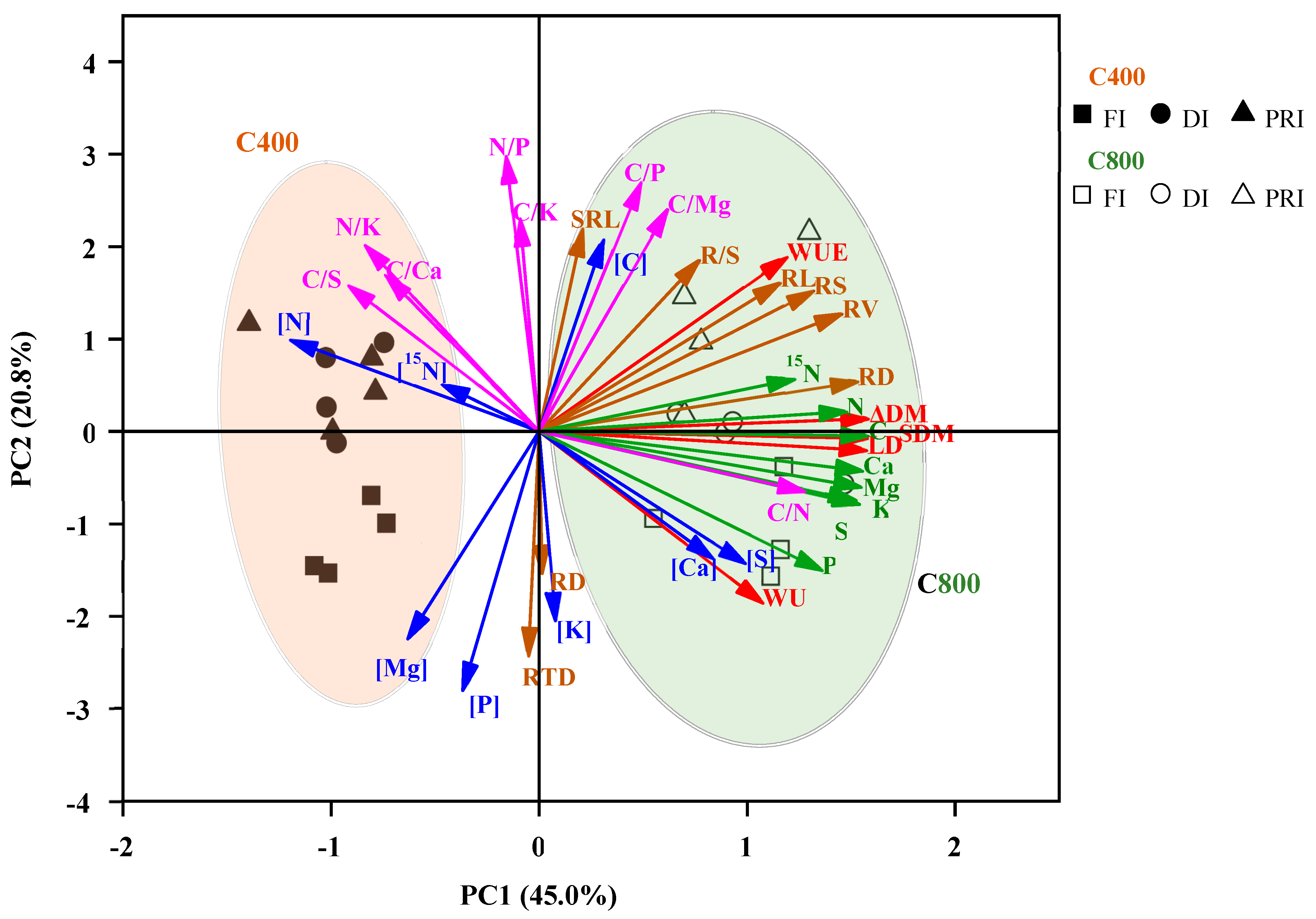
3.7. PCA analysis of tomato parameters
PCA plots of tomato attributes for tomato plants including plant growth parameters, plant root morphology parameters, leaf element concentrations, leaf element uptakes, leaf nutrition stoichiometry ratios were shown in Figure 6. PC1 and PC2 explained 45.0% and 20.8% of the variation of tomato attributes, respectively. The clusters for tomato plants were distinct, since a[CO2] and e[CO2] treatments were distributed on the left and right sides of the PCA plot, respectively. a[CO2] was clustered in the same direction as the vectors of [N], [15N], [Mg], [P], leaf C/S, leaf N/K and C/Ca, indicating tomatoes grown under a[CO2] had higher of these parameters. [C400, FI] clustered to the same direction as leaf [Mg] and [P], indicating it had higher of the parameters. [ C400, DI] and [C400, PRI] clustered to the higher [N], [15N], leaf C/S, N/K and C/Ca. e[CO2] clustered towards the higher ΔDM, SDW, RDW, LDW, WU, WUE, RL, RS, RV, R/S, SRL, mineral nutrient uptakes, leaf [C], [Ca], [S], leaf C/N, C/P and C/Mg. [C800, PRI] clustered to the upper right of the PCA plot, possessing higher RL, RS, RV, RDW, SRL, R/S, WUE, leaf C/P, C/Mg and [C]. [C800, FI] clustered to the bottom right of the PCA plot, having higher leaf C, Ca, S, Mg, P, K, [Ca], [S] and plant WU, but having lower leaf [N], [15N], leaf C/Ca, C/S and N/K.
Figure 6.
PCA plots for different parameters of tomato plants grown at three irrigation regimes ([IR]) (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRD) in either ambient (400 ppm, C400) or elevated (800 ppm, C800) atmospheric CO2 concentrations. Brown vectors are related to plant root traits (RV, RL, RS, RDW, RD, RTD, SRL, root to shoot), red vectors are related to dry mass and WUE (mass, LDM, SDW, WU and WUE ), pink vectors are related to leaf nutrition stoiochiometry (leaf C/N, leaf C/P, leaf C/K, leaf C/Ca, leaf C/Mg, leaf C/S, leaf N/P, leaf N/K), green vectors are related to leaf nutrition uptake (C, N, P, K, Ca, Mg, S,15N), blue vectors are related to leaf nutrition concentration (leaf [C],leaf [N], leaf [P], leaf [K], leaf [Ca], leaf [Mg], leaf [S], leaf [15N]).
Figure 6.
PCA plots for different parameters of tomato plants grown at three irrigation regimes ([IR]) (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRD) in either ambient (400 ppm, C400) or elevated (800 ppm, C800) atmospheric CO2 concentrations. Brown vectors are related to plant root traits (RV, RL, RS, RDW, RD, RTD, SRL, root to shoot), red vectors are related to dry mass and WUE (mass, LDM, SDW, WU and WUE ), pink vectors are related to leaf nutrition stoiochiometry (leaf C/N, leaf C/P, leaf C/K, leaf C/Ca, leaf C/Mg, leaf C/S, leaf N/P, leaf N/K), green vectors are related to leaf nutrition uptake (C, N, P, K, Ca, Mg, S,15N), blue vectors are related to leaf nutrition concentration (leaf [C],leaf [N], leaf [P], leaf [K], leaf [Ca], leaf [Mg], leaf [S], leaf [15N]).
4. Discussion
Root morphological traits mediate the responses of shoot to environmental change, which would be altered by PRI and e[CO2] scenario. Nevertheless, most studies concentrated on effects of PRI combined with e[CO2] on the aboveground tissues [5,33,37], leading to roots less being understood. In the present study, the responses of plant growth, root morphological traits, leaf element concentration and uptakes and leaf element stoichiometry of tomato plants to different irrigation regimes under e[CO2] were studied. Our results not only confirmed e[CO2] could ameliorate the negative effects of PRI on tomato plant growth but also revealed PRI had positive effect on root morphological traits for mineral nutrient uptakes, relieving the negative effect on leaf mineral nutrient concentrations and optimizing leaf stoichiometrical ratios caused by e[CO2]. More importantly, studying root morphological traits would deepen our understanding the effect of PRI on nutrient status and element balance in plants under e[CO2].
4.1. The effects of PRI on Plant biomass, biomass allocation and water use efficiency under e[CO2]
In accordance with the findings of Pazzagli et al. [38] and Wei et al.[5], the ΔDM, SDM, LDM and RDM of tomatoes were greater at e[CO2] than a[CO2] (Table 1and 2, Figure 6). In addition, in the present study, despite the 30% water reduction in relative to the FI plants, reduced irrigation had similar ΔDM with FI in tomato plants grown under either [CO2], which coincided with accumulated evidences [18,19,24,39], and they believed that both DI and PRI could consume 25-50% less water without significant reduction in biomass compared to FI. Therefore, reduced irrigation had higher WUE than FI under either [CO2], since WUE was calculated as ΔDM/WU. e[CO2] has been suggested to result in decreasing plant WU attributed to the decline in leaf stomatal conductance [4]. Nonetheless, decreased stomatal conductance could increase leaf temperature and accordingly enhance the rates of leaf transpiration [40]. On the other hand, Plants grown at high [CO2] usually have larger leaf area, RS and RV, which increases transpiration of whole plants and water acquisition [40,41]. Although leaf area was not measured in this study, e[CO2] had larger LDM (+ 101.5%), RS and RV, RDW than a[CO2] (Table 1 and 2, Figure 6), and WU was positively associated RDW (Figure 5). The greater LDM reflects a larger leaf area, combined with a larger RDW, which could interpret the response of an increase in WU to CO2 enrichment. Therefore, plants grown at e[CO2] might totally offset the effect of lowered stomatal conductance and ultimately bring about an increase in transpiration and WU at plant level. Tomato plants exposed to e[CO2] obtained 45.3% higher WUE than those exposed to a[CO2] (Table 1, Figure 6). We ascribed this result to 102.6% more increase in ΔDM and 38.5% more increase in WU (Table 1).
Furthermore, biomass allocation to different plant tissues is affected by growth environment [42], and changes in biomass allocation patterns would provide a explaination for physiological deference in plants grown under various environments [43]. A theory of functional equilibrium is that plants shift their biomass allocation towards the part that is short of resources [44], which means that under e[CO2], photosynthesis rate per unit leaf mass is increased and nutrient demand may also be increased due to accelerated plant growth, but transpiration rate and mineral nutrients mass flow are decreased, which increases the need to allocate more biomass to roots [42]. Many previous studies have indicated that root biomass of most horticultural crops is enhanced under e[CO2], often stimulated more than shoot biomass, and the increases in R/S occurred more frequently [15,45,46,47]. In agreement with these findings, our results showed that compared to a[CO2], e[CO2] increased more biomass allocation to leaves and roots from stems, enhanced RDW by 139.2% and R/S by 20% (Table 1 and 2).
Similarly, according to the functional equilibrium theory [42], the water uptake is decreased per unit root mass and probably a decline in nutrient uptake due to a decrease in mass flow, which is expected to increase in biomass allocation to roots under drought condition[42]. Moreover, many studies also showed that PRI could decline leaf area, leaf and stem biomass [24,38] but promote roots growth and increase biomass allocation to root from shoot organs [48] and increase R/S [22,49]. These findings were partly consistent with our results that reduced irrigations increased RDM, R/S ratio and biomass allocation to roots from leaves (Table 1). Hence the less LDW and lower LMR at reduced irrigations were due to more ΔDM allocated to stems and roots at reduced irrigations than those at FI (Table 1). The reasons for the enhanced SMR in this study are still unknown, needing to be further studied. Our findings suggested that more biomass allocation to root and higher R/S, to some extent, could expect that the tomato plants have a stronger ability to capture water and nutrients.
4.2. The effects of PRI on root morphological traits of tomato leaves under e[CO2]
Generally, roots of high plants are the unique tissues responsible for mineral nutrient uptake, hence the growth and development of roots greatly affects the ability of plants to acquire mineral nutrients and water [50]. The growth of roots is close coordination with the supply of photosynthesis from the shoot [51], in turn, mineral nutrients uptake by roots have strong effect on the shoot growth and development [52]. Consistent with those results, our results showed that RDW, RL, RS, RV positively correlated with ΔDM, SDW and LDW (Figure 5). In addition, root growth is highly sensitive to soil mineral nutrient and water condition [50,53]. Plants could have a fast relative growth rate, a deeper resources exploration and a rapid water and nutrients acquisition by increasing RL, RS, RV and SRL, decreasing RD and RTD to expend root system with a low investment on dry matter under limited resources condition [47,54,55,56]. This has been considered as an important indicator of plant resource use strategy [54]. Likewise, in this study, reduced irrigation under either [CO2] enlarged root system size, enhanced RL, RS, RV, and SRL, but decreased RTD (Table 2). These results may be due to upregulation of genes expression controlling root RL, RS, RV and elongation of seminal roots under reduced irrigations [53]. Furthermore, ABA induced by reduced irrigations promotes seminal roots elongation, lateral roots development and branching [57] by upregulation the expression of auxin-related genes [58], and inhibition of the synthesis of ethylene [53,59]. Additionly, a decline in RD and RTD is considered to a function trait for drought tolerance, and RD and RTD regulates SRL and RS per root biomass [60]. In the present study, the increasesed SRL at reduced irrigations (particularly at PRI exposed to e[CO2]) were due to reduced irrigations decreased RTD while they had no influence on RD as compared with FI, implying that tomato plants grown at reduced irrigations had higher efficiency of water and nutrients acquisition by permitting the exploration of more soil volume per unit dry matter investment in root length [61] than those grown at FI. But the no decline in RD at reduced irrigations needs further study.
In addition, many studies have demonstrated that root traits are sensitive to e[CO2] [15,50]. e[CO2] stimulate root growth much more than that of shoot, increasing RL, RD, RV, RS, RTD, R/S and RDW [4762,63], whereas decreasing SRL of absorptive root [64], decreasing the efficiency of direct water and nutrients acquisition [65,66]. In this study, we found e[CO2] increased RL, RS, RV, R/S and RDW as compared to a[CO2] (Table 2, Figure 6) These results may be attributed to e[CO2] allocating more biomass allocated to roots but having little effect on RD, RTD and SRL in tomato plants (Table 2, Figure 6), since an increase in root biomass can reflect an increase in total RL or average RD or RTD [67]. The equivalent SRL indicated that tomato plants had similar efficiency of direct water and nutrients acquisition under the two [CO2]. Additionally, rapid growth caused by e[CO2] is related to the synthesis of auxin in shoots, which is transported to root and stimulates primary roots elongation and lateral roots development [50], resulting in the enhanced RL, RS, RV of tomato plants grown under e[CO2].
There have been an interaction between [CO2] and IR on SRL (Table 2). Compared to a[CO2], e[CO2] increased SRL at FI and at PRI, while decreased it at DI. This might be partly due to e[CO2] showing similar RTD at three irrigations, a slight decrease in RD at FI and PRI, but a tiny increase in RD at DI, in relative to a[CO2]. Since RD and RTD regulates SRL and RS [60].
Nutrient uptake in roots is achieved in three ways, namely root interception, mass flow and diffusion [50,68]. Root interception denpends on the content of nutrients in soil where root grow and the quantity of nutrients intercepted by roots equals the amount of nutrients in a volume of soil identical to the root volume [50], therefore the enhanced RL, RS, RV can increase root nutrients interception. Mass flow and diffusion are the transport of nutrients from bulk soil to root surface [68,69]. The further enlarged RS, RL, RV and SRL in tomato plants grown at PRI under e[CO2] (Table 2, Figure 6) would be more beneficial for acquisition of diffusion-limited nutrients such as phosphorus and potassium, whereas the co-optimised traits optimising water acquisition would also better assist in the acquisition of soluble mobile resources such as nitrate, calcium, magnesium, sulfur [61]. In good agreement with these, RL, RS, RV and RDW were significantly positive correlation with element uptakes.
4.3. The effects of PRI on leaf nutrient concentration, nutrient uptake, nutrient stoiochiometry of tomato leaves under e[CO2]
Plants nutrient uptake not only depends on morphological traits of roots, but also on plant physiological properties and nutrient availability in soil [50]. It is reported that mass flow of a nutrient driven by transpiration drawing water form roots and releasing it to the atmosphere, which can be calculated as nutrient concentration multiplied by the amount of water by transpired, and the mass flow of water is equal to the volume of water transpired [69]. In this study, e[CO2] enhanced WU (amount of water by transpirated) of tomato plants as compared with a[CO2], with the same amount of mineral nutrients supply to the plants, which indicated that the mineral nutrients mass flow were enhanced or not lowered. In addition, RL, RS, RV and RDW were significantly positive correlation with element uptakes (Figure 5). These might be the reasons for the enhanced mineral nutrient uptakes, leaf [Ca] and [S] and similar leaf [C], [P]leaf , [K] and [15N] of tomato plants grown under e[CO2] (Table 3 and 4).
Furthermore, compared with a[CO2], e[CO2] increased LDM , leaf N and Mg uptakes, respectively, by 101.46%, 75.16%, and 87.8% which led to decreasing leaf [N] and leaf [Mg], respectively, by 13.1% and 6.9% (Table 1, 3 and 4). Additionally, e[CO2] enhanced leaf carbon uptake by 102.37%, leading to increased leaf C/N and C/Mg. Therefore the decrease in leaf [N], [Mg], higher leaf C/N and C/Mg were mainly due to the dilution effect [4].
Plant 15N values are related with acquisition, assimilation and allocation of N in plants [70]. It is reported that 15N uptake under a[CO2] was greater than that under e[CO2] in Arabidopsis [71]. And in leaves, 15N concentration was positive correlation with N concentration [30,72]. However, we found that e[CO2] enhanced 15N uptake ,while had little effect on leaf [15N] and decreased leaf [N] as compared with a[CO2] (Table 3 and 4, Figure 6). These results might be attributed to the N of leaves transport to stems and roots and the fractionation of 15N during reallocation of N, leaving 15N in leaves, in the faster growing period [73].
Stoichiometry is the comparative ratio of nutrients [13]. Recent studies stated that leaf C/N, C/P, C/K, C/Mg, C/Ca and C/S could indicate variations in balance between leaf structural (i.e. cell wall construction) and metabolic functional (i.e. photosynthesis and synthesis of biomolecules) investments [14,74]. And leaf stoichiometry can also demonstrate leaf structural and metabolic strategies, such as leaf C and Ca primarily linked with cell wall construction, P and K mostly regulating leaf cell protoplasm metabolic status, N, Mg and S associated with both leaf structural function and metabolic activity [14,74]. Furthermore, the nutrient ratios can reflect the relative availability of nutrient availability and can be used to determine the type of nutrient limitation [75]. When plant N/P ratio is < 14, N is limiting, plant N/P ratio is > 16, P is limiting, whereas within this range along with N/K ratio = 2.1, the effects of fertilization are not unequivocally related to N/P and N/K ratio [76,77]. But when N/K ratio is < 2.1, N is limiting, N/K ratio is > 2.1, K is limiting [77]. In this study, compared to a[CO2], e[CO2] increased leaf C/N and C/Mg, while decreased C/Ca, C/S and N/K, had no influence on leaf C/P , C/K and N/P , with N/P < 14, and N/K < 2.1 (Figure 3 and 4). These results indicated that N limited tomatoes growth under e[CO2], and e[CO2] positively affected cell wall construction but negatively influenced leaf cell protoplasm metabolism.
It has been reported that the drying and wetting cycles of the soil in PRI enhance root growth [48], improve leaf mineral nutrient concentrations [19,30,78]. In accordance with these, we found reduced irrigations enhanced RL, RS, RV, SRL and R/S, particularly in PRI (Table 2). And RDW, RL,RS and RV correlated positively with element uptakes, LDW, leaf C/N and C/Mg, whereas correlated negatively with leaf [N], [P] and [Mg](Figure 6), might due to the greater LDW diluted the leaf N, P and Mg concentrations in tomato leaves exposed to e[CO2]. And in part agreement with previous studies [19,30,79], reduced irrigation plants had equivalent leaf [N], [K] and [S], leaf C and N uptakes and leaf C/N, C/K and N/K, increased leaf [C] and [15N], leaf C/P, C/Ca, C/Mg, C/S and N/P while decreased leaf [P], [Ca] and [Mg], leaf P, K, Ca, Mg and S, compared to FI (Table 4, Figure 3 and 4). The decrease in mineral nutrient uptakes were mostly due to the decreased LDW at reduced irrigations (particularly at PRI). Since element uptakes were computed as LDW multiplied by element concentration. Moreover, the decreased leaf [P], [Ca] and [Mg] at reduced irrigations were due to the decreased diffusion of soil P and mass flow of soil Ca and Mg at reduced irrigations [80,81]. The similar leaf [N] ,[K] and [S] with FI might be due to the positive effect on bioavailability on mineral nutrients at reduced irrigations [28]. The equivalent leaf C/N, C/K and N/K were due to the similar level of increase in leaf [C], [N] and [K], while the increase leaf C/P, C/Ca, C/Mg, C/S and N/P were due to the increased leaf [C], the decreased leaf [P], [Ca] and [Mg] and equivalent leaf [N] at reduced irrigations as compared with those at FI. Additionally, the results of stoichiometric ratio suggested that more C but less Ca investment to leaf cell wall construction, less P investment to leaf cell protoplasm metabolic ability, and less Mg investment to leaf structural and metabolic activity [14].
Reduced irrigations enhanced leaf [15N] but had the similar leaf [N] in relative to FI, attributed to leaf N transported to stems and roots with higher 15N left in leaves due to 15N discrimination during leaf N reallocation [73]. Since more dry mass was allocated to stems and roots at reduced irrigations (Table 1), implying that these tissues needed more N.
There has been an interaction between [CO2] and IR on leaf P uptake and leaf N/P (Table 4 and Figure 4). Compared to a[CO2], e[CO2] increased leaf P uptake by 103.5%, 98.6% and 54.8% respectively at FI, DI and PRI, while decreased leaf N/P by 10.0% and 15.56%, respectively at FI and DI, but enhanced leaf N/P by 21.8% at PRI. A less increase in leaf P uptake at PRI under e[CO2] was due to e[CO2] decreasing leaf [P] at PRI in comparison with a[CO2] (Table 3). e[CO2] decreased leaf N/P at FI and DI whereas increased it at PRI, which was attributed to e[CO2] decreased more leaf [N] but had similar leaf [P] at FI and DI, and it slightly lowered leaf [N] but significantly decreased leaf [P], in relative to a[CO2].
Moreover, compared to a[CO2], e[CO2] had similar leaf [C], [K] and [Mg], leaf C/K and C/Mg, decreased leaf [N], [P], leaf C/Ca, C/S and N/K, increased leaf [Ca], [S] and leaf C/N, C/P and N/P at PRI. These results indicated that PRI could relieve the negative effect on leaf [Mg], maintain or improve most leaf mineral nutrient concentration and element stoichiometric ratios in tomatoes grown under e[CO2]. The decreased leaf [N] and [P] were due to dilution effect. Since compared to compared to a[CO2], e[CO2] increased LDW by 100% but enhanced leaf N and P uptake, respectively by 90% and 54.8%, at PRI. The similar leaf [K] and [Mg] and enhanced leaf [Ca] and [S] might be attributed to the enlagerd RL, RS, RV (Table 2) and the higher transpiration (WU) (Table 1) of tomato plants grown at PRI under e[CO2] condition, implying that PRI enhanced the ability to acquire mineral nutrients an increased mineral nutrient diffusion and mass flow under e[CO2]. The similar leaf C/K and C/Mg at PRI were due to the similar leaf [C], [K] and [Mg] at PRI under the two [CO2] condition. The decreased leaf C/Ca, C/S and N/K were due to e[CO2] having similar leaf [C] and [K], but enhancing leaf [Ca] and [S]and the decreasing leaf [N] at PRI in relative to a[CO2]. The increased leaf C/N, C/P, N/P were attributed to e[CO2] having similar leaf [C] but less decrease in leaf [N] than leaf [P] at PRI as compared with a[CO2].
Furthermore, under e[CO2], PRI showed an increase in leaf [C], no decrease in leaf [N], [K], Ca], [Mg], [S] and [15N], even tiny increase in leaf [N] and [15N] but decrease in leaf C/N and leaf C/S as compared with FI. The similar in mineral nutrient concentrations at PRI might be ascribed to the enlagerd RL, RS, RV and SRL (Table 2, Figure 6) and the improved bioavailability of mineral nutrients induced by PRI [30]. The results suggested that PRI had the ability to alleviate the negative effects on N concentration and maintain most other mineral nutrient concentration and improve leaf C, N and S balance under e[CO2]. The decreased leaf C/N was due to the higher leaf [N] at PRI than that at FI under e[CO2]. Simultaneously, under e[CO2], leaf N/P was > 16 and leaf N/K was about 2.1 at PRI (Figure 4), but leaf N/P was < 14 and leaf N/K was < 2.1 at FI and DI, indicating that the growth of tomato plants grown at PRI was limited by P, while that at FI and DI both limited by N, under e[CO2]. This result was consistent with our previous findings that PRI could alleviate the negative effect on N concentration caused by e[CO2] [6,33,34].
5. Conclusions
Collectively, reduced irrigations had no effect on ΔDM and SDW, stimulated root growth, increased R/S ratio, enlargerd RL, RS, RV and SRL and improved WUE of tomato plants. and e[CO2] could further stimulated root growth, enlarged root-system size, enhanced R/S ratio and WUE, indicating that reduced irrigations had higher acquisition ability of water and mineral nutrients under e[CO2] environment. e[CO2] had negative effect on leaf [N] and [Mg]. Nevertheless, compared with a[CO2], e[CO2] had similar leaf [C], [K] and [Mg], leaf C/K and C/Mg, decreased leaf [N], [P], leaf C/Ca, C/S and N/K, but increased leaf [Ca], [S] and leaf C/N, C/P and N/P at PRI. These results indicated that PRI could alleviate the negative effects on leaf [Mg] and maintain or improve most leaf mineral nutrient concentration and element stoichiometric ratios in tomatoes grown under e[CO2]. Additionally, under e[CO2], PRI showed an increase in leaf [C], no decrease in leaf [N], [K], [Ca], [Mg], [S] and [15N], but a decrease in leaf C/N and C/S as compared with FI. These results suggested that PRI had the ability to alleviate the negative effects on N concentration and maintain most other mineral nutrient concentration and improve leaf C, N and S balance under e[CO2]. Therefore, PRI would be a practicable IR for optimizing WUE and nutrient status in tomato leaves in a future water-deficit and higher CO2 environment.
Author Contributions
Conceptualization, F.L. and G.W.; methodology, F.L., G.W., X.X., B.C. and M.Z. ; software, G.W., X.X.; formal analysis, G.W.; investigation, G.W., X.X, B.C., M.Z., J.L., and Z.W; data curation, G.W.; writing—original draft preparation, G.W.; writing—review and editing, F.L.; supervision, Z.W.,funding acquisition, Z.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (51909220).
Data Availability Statement
The data related to this study was shown in the paper. Any additional data may be acquired from the first corresponding author upon request.
Acknowledgments
We were grateful to college of Natural Resources and Environment for analyzing element concentrations and division of laboratory safety and services for making plant growth chambers for use.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IPCC. Climate change 2014: synthesis report. . In CoreWriting Team, Pachauri, R. K., Meyer, L.A. (Eds.), Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, Geneva., 2014.
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol 2005, 165, 351–371. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Glob Chang Biol 2021, 27, 27–49. [Google Scholar] [CrossRef]
- Leakey, A.D.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: six important lessons from FACE. J Exp Bot 2009, 60, 2859–2876. [Google Scholar] [CrossRef]
- Wei, Z.; Du, T.; Li, X.; Fang, L.; Liu, F. Interactive effects of CO2 concentration elevation and nitrogen fertilization on water and nitrogen use efficiency of tomato grown under reduced irrigation regimes. Agricultural Water Management 2018, 202, 174–182. [Google Scholar] [CrossRef]
- Yang, X.; Bornø, M.L.; Wei, Z.; Liu, F. Combined effect of partial root drying and elevated atmospheric CO2 on the physiology and fruit quality of two genotypes of tomato plants with contrasting endogenous ABA levels. Agricultural Water Management 2021, 254. [Google Scholar] [CrossRef]
- McDonald, E.P.; Erickson, J.E.; Kruger, E.L. Can decreased transpiration limit plant nitrogen acquisition in elevated CO2? Functional Plant Biology 2002, 22, 1115–1120. [Google Scholar] [CrossRef]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; Holbrook, N.M.; Nelson, R.L.; Ottman, M.J.; Raboy, V.; Sakai, H.; Sartor, K.A.; Schwartz, J.; Seneweera, S.; Tausz, M.; Usui, Y. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139–142. [Google Scholar] [CrossRef]
- Loladze, I. Hidden shift of the ionome of plants exposed to elevated CO(2)depletes minerals at the base of human nutrition. Elife 2014, 3, e02245. [Google Scholar] [CrossRef]
- Müller, C.; Elliott, J.; Levermann, A. Fertilizing hidden hunger. Nature Climate Change 2014, 4, 540–541. [Google Scholar] [CrossRef]
- Loladze, I. Rising atmospheric CO2 and human nutrition: toward globally imbalanced plant stoichiometry? Trends in Ecology & Evolution 2002, 17, 457–461. [Google Scholar] [CrossRef]
- Du, C.; Wang, X.; Zhang, M.; Jing, J.; Gao, Y. Effects of elevated CO2 on plant C-N-P stoichiometry in terrestrial ecosystems: A meta-analysis. Sci Total Environ 2019, 650, 697–708. [Google Scholar] [CrossRef]
- Kundu, P.; Goel, K.; Zinta, G. Nutritional imbalance in plants under rising atmospheric CO2. In Plant Nutrition and Food Security in the Era of Climate Change, 2022; pp 513-536.
- Xing, K.; Zhao, M.; Niinemets, U.; Niu, S.; Tian, J.; Jiang, Y.; Chen, H.Y.H.; White, P.J.; Guo, D.; Ma, Z. Relationships Between Leaf Carbon and Macronutrients Across Woody Species and Forest Ecosystems Highlight How Carbon Is Allocated to Leaf Structural Function. Front Plant Sci 2021, 12, 674932. [Google Scholar] [CrossRef]
- Benlloch-Gonzalez, M.; Bochicchio, R.; Berger, J.; Bramley, H.; Palta, J.A. High temperature reduces the positive effect of elevated CO2 on wheat root system growth. Field Crops Research 2014, 165, 71–79. [Google Scholar] [CrossRef]
- Chaudhuri, U.N.; Kirkham, M.B.; Kanemasu, E.T. Root Growth of Winter Wheat under Elevated Carbon Dioxide and Drought. Crop Sci 1990, 30, 853–857. [Google Scholar] [CrossRef]
- Uddin, S.; Low, M.; Parvin, S.; Fitzgerald, G.J.; Tausz-Posch, S.; Armstrong, R.; O'Leary, G.; Tausz, M. Elevated [CO2] mitigates the effect of surface drought by stimulating root growth to access sub-soil water. PLoS One 2018, 13, e0198928. [Google Scholar] [CrossRef]
- Kang, S.; Zhang, J. Controlled alternate partial root-zone irrigation: its physiological consequences and impact on water use efficiency. J Exp Bot 2004, 55, 2437–2446. [Google Scholar] [CrossRef]
- Wang, Y.S.; Liu, F.L.; Andersen, M.N.; Jensen, C.R. Improved plant nitrogen nutrition contributes to higher water use efficiency in tomatoes under alternate partial root-zone irrigation. Functional Plant Biology 2010, 37, 175–182. [Google Scholar] [CrossRef]
- Dodd, I.C. Soil moisture heterogeneity during deficit irrigation alters root-to-shoot signalling of abscisic acid. Funct Plant Biol 2007, 34, 439–448. [Google Scholar] [CrossRef]
- Wang, Y.S.; Liu, F.L.; Jensen, C.R. Comparative effects of partial root-zone irrigation and deficit irrigation on phosphorus uptake in tomato plants. Journal of Horticultural Science & Biotechnology 2012, 87, 600–604. [Google Scholar] [CrossRef]
- Kang, S.Z.; Liang, Z.S.; Hu, W.; Zhang, J.H. Water use efficiency of controlled alternate irrigation on root-divided maize plants. Agricultural Water Management 1998, 38, 69–76. [Google Scholar] [CrossRef]
- Kang, S.Z.; Shi, W.J.; Cao, H.X.; Zhang, J.H. Alternate watering in soil vertical profile improved water use effciency of maize (Zea mays). Field Crops Research 2002, 77, 31–41. [Google Scholar] [CrossRef]
- Liu, F.; Shahnazari, A.; Andersen, M.N.; Jacobsen, S.-E.; Jensen, C.R. Effects of deficit irrigation (DI) and partial root drying (PRD) on gas exchange, biomass partitioning, and water use efficiency in potato. Scientia Horticulturae 2006, 109, 113–117. [Google Scholar] [CrossRef]
- Wang, L.; de Kroon, H.; Bogemann, G.M.; Smits, A.J.M. Partial root drying effects on biomass production in Brassica napus and the significance of root responses. Plant and Soil 2005, 276, 313–326. [Google Scholar] [CrossRef]
- Dodd, I.C. Rhizosphere manipulations to maximize 'crop per drop' during deficit irrigation. J Exp Bot 2009, 60, 2454–2459. [Google Scholar] [CrossRef]
- Birch, H.F. The Effec tOf Soi lDrying On Humus Decomposition and Nitrogen availability. Plant and Soil 1958, 1, 9–31. [Google Scholar] [CrossRef]
- Wang, Y.S.; Jensen, C.R.; Liu, F.L. Nutritional responses to soil drying and rewetting cycles under partial root-zone drying irrigation. Agricultural Water Management 2017, 179, 254–259. [Google Scholar] [CrossRef]
- Wang, Y.S.; Liu, F.L.; Jensen, C.R. Comparative effects of deficit irrigation and alternate partial root-zone irrigation on xylem pH, ABA and ionic concentrations in tomatoes. J Exp Bot 2012, 63, 1907–1917. [Google Scholar] [CrossRef]
- Wang, Y.S.; Liu, F.L.; de Neergaard, A.; Jensen, L.S.; Luxhøi, J.; Jensen, C.R. Alternate partial root-zone irrigation induced dry/wet cycles of soils stimulate N mineralization and improve N nutrition in tomatoes. Plant and Soil 2010, 337, 167–177. [Google Scholar] [CrossRef]
- Sun, Y.Q.; Cui, X.Y.; Liu, F.L. Effect of irrigation regimes and phosphorus rates on water and phosphorus use efficiencies in potato. Scientia Horticulturae 2015, 190, 64–69. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Keller, B.; Hoop, D.; Jud, K.; Boivin, P.; Frossard, E. Increased availability of phosphorus after drying and rewetting of a grassland soil: processes and plant use. Plant and Soil 2013, 370, 511–526. [Google Scholar] [CrossRef]
- Wei, G.; Zhang, M.; Cui, B.; Wei, Z.; Liu, F. Ammonium nitrogen combined with partial root-zone drying enhanced fruit quality of tomato under elevated atmospheric CO2. Scientia Horticulturae 2024, 323. [Google Scholar] [CrossRef]
- Wei, Z.; Du, T.; Li, X.; Fang, L.; Liu, F. Interactive Effects of Elevated CO2 and N Fertilization on Yield and Quality of Tomato Grown Under Reduced Irrigation Regimes. Front Plant Sci 2018, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Bornø, M.L.; Eduah, J.O.; Müller-Stöver, D.S.; Liu, F. Effect of different biochars on phosphorus (P) dynamics in the rhizosphere of Zea mays L. (maize). Plant and Soil 2018, 431, 257–272. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Y.; Manevski, K.; Andersen, M.N.; Li, Y.; Wei, Z.; Liu, F. Biochar and alternate wetting-drying cycles improving rhizosphere soil nutrients availability and tobacco growth by altering root growth strategy in Ferralsol and Anthrosol. Sci Total Environ 2021, 806, 150513. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, P.; Wei, Z.; Liu, J.; Hu, X.; Liu, F. Effects of elevated CO2 and nitrogen supply on leaf gas exchange, plant water relations and nutrient uptake of tomato plants exposed to progressive soil drying. Scientia Horticulturae 2022, 292. [Google Scholar] [CrossRef]
- Pazzagli, P.T.; Weiner, J.; Liu, F. Effects of CO2 elevation and irrigation regimes on leaf gas exchange, plant water relations, and water use efficiency of two tomato cultivars. Agricultural Water Management 2016, 169, 26–33. [Google Scholar] [CrossRef]
- Elhani, S.; Haddadi, M.; Csákvári, E.; Zantar, S.; Hamim, A.; Villányi, V.; Douaik, A.; Bánfalvi, Z. Effects of partial root-zone drying and deficit irrigation on yield, irrigation water-use efficiency and some potato (Solanum tuberosum L.) quality traits under glasshouse conditions. Agricultural Water Management 2019, 224. [Google Scholar] [CrossRef]
- Reddy, A.R.; Rasineni, G.K.; Raghavendra, A.S. The impact of global elevated CO2 concentration on photosynthesis and plant productivity. Current Science 2010, 99, 46–57. [Google Scholar]
- Fleisher, D.H.; Timlin, D.J.; Reddy, V.R. Interactive Effects of Carbon Dioxide and Water Stress on Potato Canopy Growth and Development. Agronomy Journal 2008, 100, 711–719. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Aust. J. Plant Physiol 2000, 27, 595–607. [Google Scholar] [CrossRef]
- Kizildeniz, T.; Pascual, I.; Irigoyen, J.J.; Morales, F. Future CO2, warming and water deficit impact white and red Tempranillo grapevine: Photosynthetic acclimation to elevated CO2 and biomass allocation. Physiol Plant 2021, 172, 1779–1794. [Google Scholar] [CrossRef]
- Brouwer, R. Distribution of dry matter in the plant. Netherlands Journal of Agricultural Science 1962, 10, 361–376. [Google Scholar] [CrossRef]
- Benlloch-Gonzalez, M.; Berger, J.; Bramley, H.; Rebetzke, G.; Palta, J.A. The plasticity of the growth and proliferation of wheat root system under elevated CO2. Plant and Soil 2013, 374, 963–976. [Google Scholar] [CrossRef]
- Madhu, M.; Hatfield, J.L. Dynamics of Plant Root Growth under Increased Atmospheric Carbon Dioxide. Agronomy Journal 2013, 105, 657–669. [Google Scholar] [CrossRef]
- Rogers, H.H.; Peterson, C.M.; Mccrimmon, J.N.; CURE, J.D. Response of plant roots to elevated atmospheric carbondioxide. Plant, Celt and Environment 1992, 15, 749–752. [Google Scholar] [CrossRef]
- Mingo, D.M.; Theobald, J.C.; Bacon, M.A.; Davies, W.J.; Dodd, I.C. Biomass allocation in tomato (Lycopersicon esculentum) plants grown under partial rootzone drying: enhancement of root growth. Funct Plant Biol 2004, 31, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xu, F.; Zhu, J.R.; Wang, R.F.; Xu, Z.H.; Shu, L.Z.; Xu, W.W. Nitrogen forms affect root growth, photosynthesis, and yield of tomato under alternate partial root-zone irrigation. Journal of Plant Nutrition and Soil Science 2015, 179, 104–112. [Google Scholar] [CrossRef]
- Wang, H.; Inukai, Y.; Yamauchi, A. Root Development and Nutrient Uptake. Critical Reviews in Plant Sciences 2007, 25, 279–301. [Google Scholar] [CrossRef]
- Ogawa, A.; Kawashima, C.; Yamauchi, A. Sugar Accumulation along the Seminal Root Axis, as Affected by Osmotic Stress in Maize: A Possible Physiological Basis for Plastic Lateral Root Development. Plant Production Science 2015, 8, 173–180. [Google Scholar] [CrossRef]
- Ristova, D.; Busch, W. Natural variation of root traits: from development to nutrient uptake. Plant Physiol 2014, 166, 518–527. [Google Scholar] [CrossRef]
- Ranjan, A.; Sinha, R.; Singla-Pareek, S.L.; Pareek, A.; Singh, A.K. Shaping the root system architecture in plants for adaptation to drought stress. Physiol Plant 2022, 174, e13651. [Google Scholar] [CrossRef] [PubMed]
- Birouste, M.; Zamora-Ledezma, E.; Bossard, C.; Pérez-Ramos, I.M.; Roumet, C. Measurement of fine root tissue density: a comparison of three methods reveals the potential of root dry matter content. Plant and Soil 2013, 374, 299–313. [Google Scholar] [CrossRef]
- Kong, D.; Wang, J.; Wu, H.; Valverde-Barrantes, O.J.; Wang, R.; Zeng, H.; Kardol, P.; Zhang, H.; Feng, Y. Nonlinearity of root trait relationships and the root economics spectrum. Nat Commun 2019, 10, 2203. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, C.M.; Lindahl, B.; Wardle, D.A.; Sundqvist, M.K.; Gundale, M.J.; Fanin, N.; Kardol, P. Root trait-microbial relationships across tundra plant species. New Phytol 2021, 229, 1508–1520. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, C.; Zhang, Y.; Wang, B.; Ran, Q.; Zhang, J. The bHLH family member ZmPTF1 regulates drought tolerance in maize by promoting root development and abscisic acid synthesis. J Exp Bot 2019, 70, 5471–5486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yuan, W.; Wang, Q.; Cao, Y.; Xu, F.; Dodd, I.C.; Xu, W. ABA regulation of root growth during soil drying and recovery can involve auxin response. Plant Cell Environ 2022, 45, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Rowe, J.H.; Topping, J.F.; Liu, J.; Lindsey, K. Abscisic acid regulates root growth under osmotic stress conditions via an interacting hormonal network with cytokinin, ethylene and auxin. New Phytol 2016, 211, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Wasaya, A.; Zhang, X.; Fang, Q.; Yan, Z. Root Phenotyping for Drought Tolerance: A Review. Agronomy 2018, 8. [Google Scholar] [CrossRef]
- Ho, M.D.; Rosas, J.C.; Brown, K.M.; Lynch, J.P. Root architectural tradeoffs for water and phosphorus acquisition. Funct Plant Biol 2005, 32, 737–748. [Google Scholar] [CrossRef]
- Nie, M.; Lu, M.; Bell, J.; Raut, S.; Pendall, E. Altered root traits due to elevated CO2: a meta-analysis. Global Ecology and Biogeography 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Pokorný, R.; Tomášková, I.; Marek, M.V. Response of Norway spruce root system to elevated atmospheric CO2 concentration. Acta Physiologiae Plantarum 2013, 35, 1807–1816. [Google Scholar] [CrossRef]
- Wang, N.; Gao, G.; Wang, Y.; Wang, D.; Wang, Z.; Gu, J. Coordinated responses of leaf and absorptive root traits under elevated CO2 concentration in temperate woody and herbaceous species. Environmental and Experimental Botany 2020, 179. [Google Scholar] [CrossRef]
- Li, X.; Jiang, D.; Liu, F. Soil warming enhances the hidden shift of elemental stoichiometry by elevated CO2 in wheat. Scientific Reports 2016, 6. [Google Scholar] [CrossRef]
- Taub, D.R.; Wang, X. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J Integr Plant Biol 2008, 50, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.E.; LeCain, D.R.; McCormack, M.L.; Pendall, E.; Carlson, M.; Blumenthal, D.M.; Lamb, E. Root responses to elevated CO 2, warming and irrigation in a semi-arid grassland: Integrating biomass, length and life span in a 5-year field experiment. Journal of Ecology 2018, 106, 2176–2189. [Google Scholar] [CrossRef]
- Jungk, A. Root hairs and the acquisition of plant nutrients from soil. Journal of Plant Nutrition and Soil Science 2001, 164, 121–129. [Google Scholar] [CrossRef]
- McGrath, J.M.; Lobell, D.B. Reduction of transpiration and altered nutrient allocation contribute to nutrient decline of crops grown in elevated CO(2) concentrations. Plant Cell Environ 2013, 36, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, T.; Ito, O.; Engelaar, W.M.H.G. Uptake, metabolism and distribution of nitrogen in crop plants traced by enriched and natural 15N: Progress over the last 30 years. Phytochemistry Reviews 2003, 2. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Rubio Asensio, J.S.; Cousins, A.B. Carbon dioxide enrichment inhibits nitrate assimilation in wheat and Arabidopsis. Science 2010, 328, 899–903. [Google Scholar] [CrossRef]
- Ullah, S.; Zhao, Q.; Wu, K.; Ali, I.; Liang, H.; Iqbal, A.; Wei, S.; Cheng, F.; Ahmad, S.; Jiang, L.; Gillani, S.W.; Amanullah, *!!! REPLACE !!!*; Anwar, S. ; Khan, Z. Biochar application to rice with (15)N-labelled fertilizers, enhanced leaf nitrogen concentration and assimilation by improving morpho-physiological traits and soil quality. Saudi J Biol Sci 2021, 28, 3399–3413. [Google Scholar] [CrossRef]
- Zhou, W.; Hu, C.-S.; Li, J.; Christie, P.; He, X.-H.; Ju, X.-T. Natural 15N Abundance in Winter Wheat Amended with Urea and Compost: A Long-Term Experiment. Pedosphere 2013, 23, 835–843. [Google Scholar] [CrossRef]
- Fratte, M.D.; Pierce, S.; Zanzottera, M.; Cerabolini, B.E.L. The association of leaf sulfur content with the leaf economics spectrum and plant adaptive strategies. Funct Plant Biol 2021, 48, 924–935. [Google Scholar] [CrossRef]
- Gusewell, S.; Koerselman, W. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspectives in Plant Ecology Evolution and Systematics 2002, 5, 37–61. [Google Scholar] [CrossRef]
- Craine, J.M.; Morrow, C.; Stock, W.D. Nutrient concentration ratios and co-limitation in South African grasslands. New Phytol 2008, 179, 829–836. [Google Scholar] [CrossRef]
- Olde Venterink, H.; Wassen, M.J.; Verkroost, A.W.M.; De Ruiter, P.C. Species Richness–Productivity Patterns Differ between N-, P-, and K-Limited Wetlands. Ecology 2003, 84, 2191–2199. [Google Scholar] [CrossRef]
- Sun, Y.; Holm, P.E.; Liu, F. Alternate partial root-zone drying irrigation improves fruit quality in tomatoes. Horticultural Science 2014, 41, 185–191. [Google Scholar] [CrossRef]
- Sun, Y.; Yan, F.; Cui, X.; Liu, F. Plasticity in stomatal size and density of potato leaves under different irrigation and phosphorus regimes. J Plant Physiol 2014, 171, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Rubæk, G.H.; Liu, F.; Andersen, M.N. Effect of partial root zone drying and deficit irrigation on nitrogen and phosphorus uptake in potato. Agricultural Water Management 2015, 159, 66–76. [Google Scholar] [CrossRef]
- Oliveira, E.M.M.; Ruiz, H.A.; Alvarez V, V.H.; Ferreira, P.A.; Costa, F.O.; Almeida, I.C.C. Nutrient supply by mass flow and diffusion to maize plants in response to soil aggregate size and water potential. Revista Brasileira de Ciência do Solo 2010, 34, 317–328. [Google Scholar] [CrossRef]
Figure 1.
The average daily climate data inside the climate-controlled growth chambers during the treatment period.
Figure 1.
The average daily climate data inside the climate-controlled growth chambers during the treatment period.
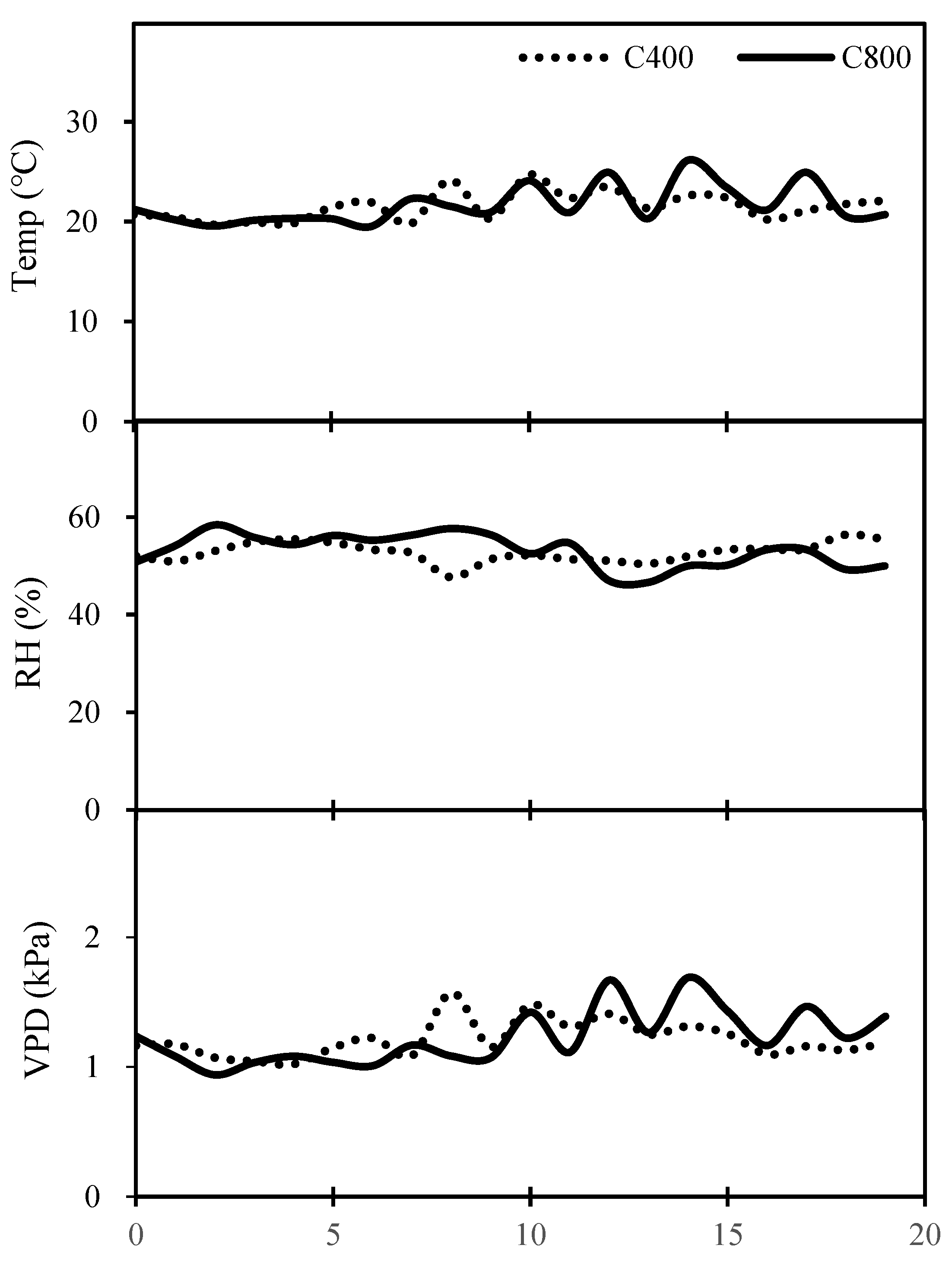
Figure 2.
The profile of tomato growth pot.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



